Область применения
Изобретение относится к иммунологии и биотехнологии, в частности к методам получения иммуногенных препаратов и вакцин, которые могут быть использованы для профилактики гриппа.
Актуальность
Грипп является одним из наиболее распространенных инфекционных заболеваний человека и животных. Гриппозная инфекция часто протекает тяжело и иногда приводит к смертельному исходу. Некоторые штаммы вируса, например «испанка» в 1918-1920 годах, способны вызывать пандемии в мировом масштабе, сопровождающиеся высокой смертностью. Высокая изменчивость поверхностных белков вируса, гемаглютинина и нейраминидазы, приводит к возникновению нового эпидемического штамма каждые 1-2 года, с такой же частотой требуется изготовление «стандартных» штамм-специфических вакцин. Используемые в настоящее время противогриппозные вакцины основаны на получаемом в куриных эмбрионах вирусе гриппа или его компонентах [Nicholson K., Webster R., Hay A. (1998) Textbook of Influenza // Oxford, Blackwell Science]. Высокая изменчивость поверхностных белков вируса, гемаглютинина и нейраминидазы приводит к возникновению нового эпидемического штамма каждые 1-2 года [Webster et al., (1992), Microbiol. Rev.56. P.152-179], с такой же частотой требуется изготовление «стандартных» штамм-специфических вакцин.
Альтернативой традиционным вакцинам являются рекомбинантные вакцины, основанные на отдельных белках вируса гриппа, которые могут быть получены в стандартных организмах-продуцентах. Использование рекомбинантных вакцин не только снимает зависимость производства от куриных эмбрионов и решает проблемы безопасности, общие для вакцин, основанных на цельном патогене, но и открывает возможности создания «универсальных» вакцин при использовании консервативных белков вируса. Более того, такой подход позволяет, с одной стороны, подготовить вакцину очень быстро или создавать вакцины, которые «перекрывают» антигенные свойства большинства пандемических вирусов.
Поэтому задача создания рекомбинантной вакцины против гриппа продолжает являться актуальной.
Уровень техники
Современные противогриппозные вакцины
В таблице представлен перечень и характеристики противогриппозных вакцин, зарегистрированных и использующихся в России.
Все вакцины могут быть разделены на следующие типы:
1. Живые.
2. Инактивированные,
- цельновирионные,
- расщепленные,
- субъединичные.
Из этого перечня видно, что для таких актуальных вакцин, как противогриппозные, вообще не разрабатывались рекомбинантные эпитопные вакцины. Однако преимущество генно-инженерных технологий в решении проблем эффективности и безопасности гриппозных вакцин очевидны, и их можно перечислить в следующем порядке по степени важности:
1. Гарантированно полное отсутствие яичного белка (овальбумина) - основного аллергена в современных вакцинах.
2. Индивидуальность избранных эпиотопов - гарантии отсутствия аутоиммунных реакций и осложнений. Эта опасность особенно велика для живых и цельновирионных вакцин и не исключается для субъединичных.
3. Относительная простота производства, отсутствие зависимости от природного сырья, например куриных эмбрионов, и полное соответствие требования безопасности (отсутствие вирусных компонентов). Хорошие экономические показатели производства.
4. Возможность быстрого развертывания масштабного производства на случай роста потребностей в вакцине в предпандемический и пандемический периоды.
Биологически-активные наноструктуры на основе капсидов вирусов животных
Способность биологических макромолекул к самосборке и самоорганизации является одной из отличительных черт живых систем и предоставляет поистине неисчерпаемые возможности для использования биомолекул в качестве «строительных» блоков «молекулярного конструктора» для направленного создания новых наноархитектур, с заданными пространственными и функциональными свойствами. Одним из наиболее ярких примеров таких структур, обладающих четкой симметрией, обширными возможностями направленной модификации современными методами генной инженерии являются вирусные частицы.
Высокая стабильность рекомбинантных вирусноподобных частиц (VLP), их симметричная пространственная организация, легкость получения и очистки VLP, их безопасность, возможности направленной модификации структурных белков вирусов методами генной инженерии делают эти частицы идеальными векторами-носителями для представления чужеродных функционально-значимых пептидов и целых белков. Представление антигенных детерминант-эпитопов на поверхности VLP обеспечивает их высокую иммуногенность за счет использования вириона в качестве адьюванта в сочетании с высоким выходом, легкостью очистки и стабильностью при хранении. Эффективность такого подхода иллюстрируется многочисленными примерами создания VLP для борьбы с такими инфекционными заболеваниями, как малярия, бешенство, вирусные гепатиты В, С и Е папилломавирусная инфекция (рак шейки матки), СПИД и т.д.
Одним из наиболее эффективных носителей антигенных детерминант является нуклеокапсидный белок вируса гепатита В человека, а также близкородственные вирусы животных, в дальнейшем коллективно именуемые ВГБ. Мономеры этого белка, состоящие из 183-185 аминокислот, в инфицированных клетках собираются в стабильные агрегаты (вирусоподобные частицы), называемые НВс-частицы. Образуются два типа частиц - частицы диаметром около 30 нм, состоящие из 180 мономеров, и в большем количестве частицы диаметром 34 нм, состоящие из 240 субъединиц.
Известно, что НВс-частицы могут быть использованы в качестве высокоиммуногенного носителя чужеродных пептидов, стимулирующего Т-клеточный иммунный ответ. Два района НВс могут быть использованы для презентации чужеродных пептидов на поверхности НВс-частиц, - N - конец белка и так называемая иммунодоминантная петля, расположенная между 75 и 85 аминокислотными остатками белка. Было показано, что С - концевая аргинин-богатая область белка (150-183 аминокислотные остатки) может быть удалена без нарушения самосборки частиц, более того, в отличие от образованных полноразмерным белком, такие частицы при сборке не включают нуклеиновые кислоты клетки-хозяина.
Противогриппозные вакцины на основе рекомбинантных НВс-частиц
Использование рекомбинантных НВс-частиц для получения противогриппозных вакцин описано в работе [Neirynck et al., (1999) Nature Med., 5(10):1157-1163] и последующих работах этой группы. Их авторами получена серия рекомбинантных НВс-частиц, содержащих внеклеточный домен М2 (М2Е) белка вируса гриппа человека (М2еНВс частицы) и показана их высокая иммуногенность и протективность в отношении инфекции человеческим вирусом гриппа в опытах на лабораторных животных. В этих работах была использована М2Е последовательность S L L T E V E T P I R N E W G C R C N D S S D, присутствующая в практически неизменном виде во всех человеческих штаммах вируса, начиная с изолированного в 1933 г. штамма A/WSN/33.
Белок М2 вирусов гриппа типа А принципиально важен для конструирования универсальных вакцин в связи с рядом недооцененных его свойств:
1. Локализация и экспонирование на поверхности вириона.
2. Критическая важность в процессах фьюжина и почкования (ранняя и поздняя стадии инфекции).
3. Высокий консерватизм среди всех подтипов вируса вируса гриппа А.
Выбор высококонсервативного антигена предусматривал индукцию антител, способных к подавлению репродукции к большинству подтипов А вирусов гриппа. Такая вакцина получила название «универсальной» - предполагающей защиту фактически от всех вирусов гриппа типа А.
У вирусов гриппа животного происхождения последовательность М2Е отличается от приведенной выше. Например, последовательности S L L T E V E T P T R N G W E C K C S D S S D и S L L T E V E T P T R N E W E C R C S D S S D встречаются в различных штаммах «птичьего» гриппа (A/Duck/ Potsdam 1402-6/198 6 и A/Chicken/Kurgan/05/2005 соответственно), а М2е вируса «свиного» гриппа A/California/04/2009(H1N1) имеет последовательность S L L T E V E T P T R S E W E C R C S D S S D (подчеркнуты отличия от консенсусной последовательности "человеческих" штаммов вируса гриппа). Разработка рекомбинантных противогриппозных вакцин на основе НВс частиц, представляющих на своей поверхности М2Е пептиды вирусов гриппа животного происхождения, описана нами ранее [Равин Н.В., Киселев О.И., Скрябин К.Г. (2009) Универсальная вакцина против вируса гриппа птиц. Патент РФ №2358981 от 20.06.2009 г.] и [Котляров Р.Ю., Куприянов В.В., Мигунов А.И., Степанова Л.А., Цыбалова Л.М., Киселев О.И., Равин Н.В., Скрябин К.Г. (2010). Разработка рекомбинантной вакцины против гриппа A(H1N1) 2009 на основе вирусоподобных наночастиц - носителей внеклеточного домена М2 белка. Acta Naturae, т.2 №2(5), с.75-80]. Во всех описанных работах препараты вакцинных белков и рекомбинантных НВс-частиц, несущих М2е пептиды, были получены в клетках бактерий Escherichia coli.
Методы продукции рекомбинантных белков
Одним из наиболее ярких и убедительных достижений современной биотехнологии стало создание и бурное развитие в последние десятилетия новой области мировой экономики - биофармацевтической промышленности и промышленности биоматериалов, направленной на производство принципиально нового класса лекарств - рекомбинантных белков медицинского назначения. Различные вакцинные белки, интерфероны, эритпропоэтины, факторы роста, антитела, доступные ранее лишь в аналитических количествах, входят теперь в перечень жизненно-важных медицинских препаратов, производятся в объемах до нескольких тонн в год и прочно вошли в повседневную практику современной медицины, пищевой и фармацевтической промышленности. В подавляющем большинстве случаев источником получения этих продуктов служат клетки бактерий, животных или дрожжей.
Использование растений как «биофабрик» для продукции чужеродных белков имеет ряд преимуществ по сравнению с клетками бактерий, дрожжей и млекопитающих:
(1) Наработка целевых белков в растениях не требует применения дорогостоящей аппаратуры (ферментеры), культуральных сред и системы стерильности Для роста растений требуются только почва, вода и солнце, поэтому стоимость их выращивания несравнимо ниже стоимости культивирования клеток бактерий, дрожжей или животных. Вследствие этого стоимость белков, получаемых в растениях, уже сегодня в 10-30 раз ниже стоимости аналогичных белков, получаемых из бактерий [Giddings, G., Allison, G., Brooks, D., Carter, A (2000) Transgenic plants as factories for biopharmaceuticals. Nat. Biotechnol. 18 (11), 1151-1155].
(2) Клетки растений биологически безопасны, поскольку растения и человек не имеют общих патогенов. Поэтому получаемые в растениях продукты не содержат опасных для человека вирусов и прионов.
(3) В отличие от бактерий и дрожжей, растения и животные имеют сходную систему посттрансляционных модификаций белков. Поэтому в растительных клетках могут быть получены те белки человека и животных, которые не могут быть корректно экспрессированы в микроорганизмах.
Применяемые в настоящее время технологии экспрессии рекомбинантных белков в растениях, как правило, предполагают получение трансгенного растения - продуцента. Уже сегодня имеется ряд примеров коммерчески успешного производства белков в растениях таким способом. Например, получаемые в трансгенных растениях кукурузы трипсин, авидин и глюкуронидаза уже сейчас производятся и продаются фирмой Сигма-Алдрич. Близки к выходу на рынок апротинин (ProdiGene), коллаген (ProdiGene, Medicago, Meristem Therapeautics), липаза (Meristem Therapeautics), лактоферрин (Ventria, Meristem Therapeutics), лизоцим (Ventria), вакцина против вируса гастроэнтерита TGEV (ProdiGene), моноклональные антитела против кариеса и вируса герпеса (Planet Biotechnology, Epicyte Pharmaceutical). Первая полученная в растениях вакцина от болезни Ньюкасла, разработанная компанией Dow Agroscience успешно прошла регистрацию в США в 2006 году, более 20 полученных в растениях биофармацевтиков в настоящее время находятся на различных стадиях клинических испытаний (http://www.molecularfarmmg.com/news2.html).
Следует отметить, что получение трансгенных растений не является оптимальным способом создания растительных «биофабрик», так как уровень продукции целевых белков трансгенными растениями, как правило, весьма низок (около 0,1% общего белка). Низкий уровень экспрессии определяет высокую стоимость очистки продукта и, в конечном итоге, конкурентоспособность растительной системы экспрессии. Кроме того, культивирование трансгенных растений связано с рядом формальных ограничений.
Повышение экономической эффективности использования растений в качестве «биофабрик» - продуцентов белков - требует разработки новых технологий продукции в нетрансгенных растениях на высоком уровне целевых белков. Такие технологии позволят получать в растениях недорогие и безопасные белки и могут стать конкурентоспособной альтернативой традиционным методам, основанным на использовании бактерий, дрожжей или клеток животных.
Альтернативой трансгенным растениям как продуцентам рекомбинантных белков могут являться системы экспрессии, основанной на рекомбинантных вирусах растений (вирусы-векторы). Многие растительные вирусы, например вирус табачной мозаики и X-вирус картофеля, при заражении растения размножаются в больших количествах, а уровень продукции собственных белков вируса достигает 70% белков растительной клетки. Смысл этого метода состоит в интеграции гена, кодирующего необходимый чужеродный белок, в геном вируса и заражении модифицированным вирусом растительных клеток. При инфекции синтезируются не только собственные белки вируса, но и целевой белок, ген которого был специально вставлен в геном вируса. Уровень продукции рекомбинантного белка в этом случае может составлять до 20% общего белка [Marillonnet, S., Thoeringer, С., Kandzia, R., Klimyuk, V., Gleba, Y. (2005) Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nat. Biotechnol., 23, 718-723], что соответствует 1 грамму белка на 1 килограмм растительной биомассы, что в десятки-сотни раз выше уровней продукции, обычно достигаемых в трансгенных растениях.
До настоящего времени для получения М2еНВс частиц использовали только системы экспрессии в клетках бактерий Е. coli. Нам не известны опубликованные данные по продукции в растениях (как в трансгенных растениях, так и с помощью рекомбинантных вирусов-векторов) рекомбинантных НВс-частиц, несущих М2е пептиды. Поэтому актуальным остается создание альтернативных методов продукции этих перспективных противогриппозных вакцинных препаратов, одним из наиболее перспективных из которых являются растительные системы экспрессии.
Раскрытие изобретения
В настоящем изобретении ставилась задача создания вирусного вектора, который может обеспечивать продукцию в растениях рекомбинантных НВс-частиц, содержащих внеклеточный домен М2 белка вируса гриппа штамма A/Duck/Potsdam/4024/26 (далее -М2ерНВс-частицы), и соответствующей системы экспрессии этого продукта в клетках растений Nicotiana benthamiana.
Фактически задача была решена путем:
а) дизайна и синтеза оптимизированного для экспрессии в растениях гена, кодирующего гибридный белок, включающий М2Е пептид и НВс антиген вируса гепатита В.
б) клонирования синтезированного гена в вирусе-векторе на основе генома Х вируса картофеля,
в) разработки протоколов инфицирования рекомбинантным вирусом-вектором растений Nicotiana benthamiana и последующей продукции в растениях рекомбинантного белка М2ерНВс.
Первый аспект настоящего изобретения связан с дизайном гибридного белка М2ерНВс, включающего М2Е пептид вируса гриппа птиц штамма A/Duck/Potsdam/4024/26 и НВс антиген вируса гепатита В, и конструированием синтетического гена, кодирующего этот белок.
Вторым аспектом изобретения является получение экспрессионного вируса-вектора, обеспечивающего продукцию в растениях белка М2ерНВс.Для этого упомянутый выше синтетический ген был клонирован в вирусный вектор pA7248amvT.
Соответственно, третий аспект изобретения относится к оптимизации условий инфицирования растений Nicotiana benthamiana рекомбинантным вирусом-вектором и последующей продукции в растениях рекомбинантного белка М2еНВс.
Четвертый аспект изобретения связан с подтверждением факта продукции в растениях белка М2ерНВс в результате Вестерн-блот анализа белковых препаратов, выделенных из растений-продуцентов. При этом было показано, что рекомбинантный белок М2ерНВс действительно синтезируются в листьях растений-продуцентов. С помощью электронно-микроскопического анализа показано, что этот белок спонтанно собирается в вирусоподобные частицы размером около 30 нанометров, сходные с М2ерНВс-частицами, образуемыми этим белком при экспрессии в клетках бактерий Escherichia coli.
Краткое описание чертежей
Фиг.1 - Структура гибридного белка М2ерНВс, включающего последовательности М2Е пептида вируса гриппа штамма A/Duck/Potsdam/4024/26 и НВс антигена вируса гепатита В.
Приведена аминокислотная последовательность М2ерНВс белка. Последовательность М2е пептида выделена серым.
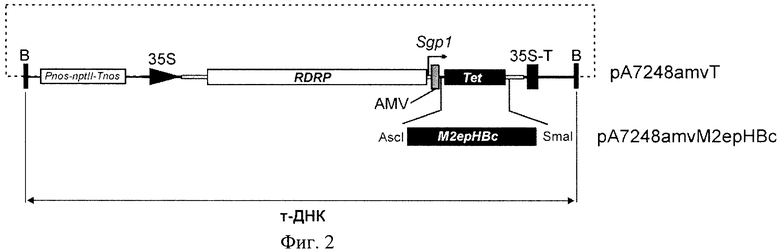
Фиг.2 - Структура вирусного вектора pA7248amvM2epHBc, обеспечивающего продукцию белка М2ерНВс в растениях.
Показаны структуры Т-ДНК областей вирусного вектора pA7248amvM2epHBc и исходного вектора pA7248amvT, участки за пределами области Т-ДНК показаны пунктирной линией. Границы Т-ДНК (бордеры, «В») отмечены жирными вертикальными линиями. 35S, - промотор, 35S-T - терминатор 35S РНК вируса мозаики цветной капусты. RDRP-ген полимеразы, Sgp1 - первый промотор субгеномной РНК Х вируса картофеля. Tet, - ген устойчивости к тетрациклину. AMV, - ДНК копия лидерной последовательности РНК вируса мозаики люцерны.
Фиг.3 - Вестерн-блот анализ белковых препаратов, выделенных из растений-продуцентов М2ерНВс.Детекция моноклональными антителами на М2е.
На гель нанесены белковые препараты:
1. Маркер молекулярного веса, размеры полос указаны в килодальтонах.
2. Растворимые белки из неинокулированных листьев N. benthamiana (контроль).
3. Растворимые белки из листьев N. benthamiana, инокулированных вирусом-вектором pA7248amvM2epHBc
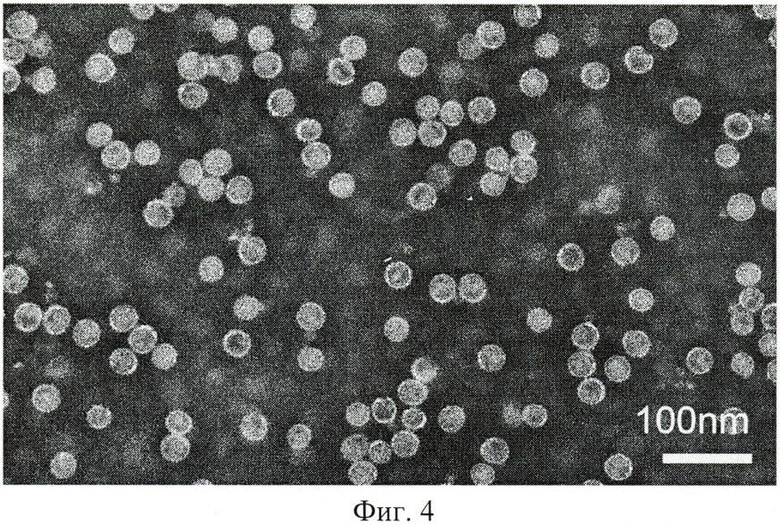
Фиг.4 - Результаты электронно-микроскопического анализа М2ерНВс частиц, выделенных из растений-продуцентов.
Осуществление изобретения.
Пример 1. Рекомбинантная белковая молекула М2ерНВс и кодирующая ее рекомбинантная нуклеиновая кислота.
Для получения иммуногенных вирусоподобных частиц использовали рекомбинантный белок, включающий внеклеточный домен М2 белка (М2Е) вируса гриппа штамма A/Duck/Potsdam/4024/26 и НВс антиген вируса гепатита В. Рекомбинантная белковая молекула, М2ерНВс включала последовательно следующие элементы: N-концевой метионин, последовательность аминокислот, соответствующую М2Е пептиду из штамма A/Duck/Potsdam/4024/26 вируса гриппа, аминокислотную последовательность НВс [Bichko, V., Pushko, P., Dreilina, D., Pumpen, P. and Gren, E. (1985) FEBS Lett. 185 (1), 208-212]. Для улучшения растворимости белка два цистеина в положениях 17 и 19 М2Е пептида, наличие которых может способствовать образованию дисульфидных связей, заменяли на серины, что не влияет на иммунологические характеристики М2Е. Аминокислотная последовательность рекомбинантной белковой молекулы М2ерНВс представлена в SEQ ID NO:1 и показана на Фиг.1.
Нуклеотидная последовательность синтетического гена, кодирующего М2ерНВс, оптимизирована для экспрессии в растениях и содержит последовательности сайтов рестрикции AscI и SmaI для клонирования в вирусный вектор pA7248amvTet. Последовательность гена М2еНВс была синтезирована in vitro. Нуклеотидная последовательность искусственного гена М2ерНВс представлена в SEQ ID NO:2.
Пример 2. Конструирование экспрессионного вируса-вектора, обеспечивающего продукцию в растениях рекомбинантного белка М2ерНВс.
В качестве основы для создания вектора был использован описанный в работе [Марданова Е.С., Котляров Р.Ю., Равин Н.В. (2009) Повышение эффективности продукции рекомбинантных белков в растениях за счет оптимизации трансляции РНК вируса-вектора. Молекулярная биология, т.43, №3, с.568-571] вирусный вектор pA7248amvT, основанный на геноме Х-вируса картофеля штамма UK3 [нуклеотидная последовательность генома этого вируса приведена в GenBank под номером М95516]. Этот вектор включает 5'-нетранслирумый участок генома ХВК, ген полимеоазы. первый промотор субгеномной РНК, трансляционный энхансер - нуклеотидную последовательность, соответствующую лидерной последовательности РНК вируса мозаики люцерны (AMV), ген устойчивости к тетрациклину (Tet), фланкированный уникальными сайтами рестрикции AscI и SmaI, последние 60 нуклеотидов гена белка оболочки и 3'-нетранслируемый участок генома Х-вируса картофеля. Вся эта конструкция помещена между 35S промотором и 35S терминатором и клонирована в бинарном векторе pBIN19. При доставке в клетки с помощью агроинфильтрации листьев происходит заражение большей части клеток листа в инфицированной области, репликация вирусного вектора в отдельных клетках и синтез продукта на высоком уровне.
Для создания вирусного вектора-продуцента М2ерНВс, ДНК вектора pA7248amvT обрабатывали рестриктазами AscI и SmaI и дотировали с обработанным этими рестриктазами фрагментом ДНК, содержащим искусственный ген М2ерНВс.Полученный рекомбинантные вектор, содержащий ген М2ерНВс на месте гена Tet, который был отобран по результатам рестрикционного анализа, обозначали pA7248amv М2ерНВс.
Структура вирусного вектора pA7248amv М2ерНВс, обеспечивающего продукцию в растениях рекомбинантного белка М2ерНВс, показана на Фиг.2.
Пример 3. Инфицирование растений Nicotiana benthamiana рекомбинантным вирусом-вектором и продукция в растениях белка М2ерНВс.
Для экспериментов по экспрессии белка М2ерНВс в растениях N. benthamiana вирусный вектор pA7248amvM2epHBc вводили в штамм Agrobaterizim tumefaciens GV3101, рекомбинантные агробактерии были использованы для инфильтрации листьев N. benthamiana. Агробактерии, содержащие рекомбинантный бинарный вектор, выращивали в течение 12 ч на шейкере при 30°С. Клетки (1,5 мл) осаждали центрифугированием (4000g, 5 мин), осадок ресуспендировали в 1,5 мл буфера, содержащего 10 мМ MES (рН 5.5) и 10 мМ MgCl2, оптическую плотность доводили до OD600=0,2. Листья растений N. benthamiana инъецировали суспензией агробактерии при помощи шприца без иглы. Инфильтрированные листья оставляли на растущих растениях.
Известно, что одним из основных факторов, ограничивающих вирусную инфекцию, в частности экспрессию целевых белков в вирусных системах экспрессии, является развитие посттранскипционного умолкания генов (PTGS), активированного двунитевыми репликативными формами вирусных РНК [Marillonnet, S., Thoeringer, С., Kandzia, R., Klimyuk, V., Gleba, Y. (2005) Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nat. Biotechnol., 23, 718-723]. Для предотвращения этого явления одновременно с агробактериями, содержащими вирус-вектор pA7248amv М2ерНВс, в листья растений инфильтрировали агробактерии, содержащие вектор-продуцент белка Р19 вируса кустистой карликовости томатов, известного суппрессора посттранскипционного умолкания генов.
После инфильтрации листьев N. benthamiana агробактериями с вирусом-вектором происходит заражение большей части клеток листа в инфицированной области, перенос Т-ДНК в ядро, транскрипция копии вирусной РНК с 35S промотора [Комарова Т.В., Скулачев М.В., Зверева А.С., Шварц A.M., Дорохов Ю.Л., Атабеков И.Г. (2006) Новый вирус-вектор для эффективной продукции целевых белков в растениях. Биохимия, 71(8), 1043-1049]. На последующей стадии происходит репликация вирусного вектора и синтез продукта на высоком уровне. Максимум синтеза продукта достигается на 6-10 сутки после инфильтрации.
Пример 4. Детекция продуцированного в растениях белка М2ерНВс методом Вестерн-блот анализа.
Наличие целевого белка в листьях растений-продуцентов регистрировали с помощью Вестерн-блот анализа. На 6-10 день после заражения фрагмент листа растирали в буфере 0.4М сахароза, 50 мМ Трис рН 8.0, 10 mM KCI, 5mM MgCl2, 10% глицерин, 10 мМ β-меркаптотанол. Полученный экстракт центрифугировали 10 минут при 14000 g, супернатант анализировали методом Вестерн-блота с использованием антител, полученных к М2е пептиду. Результаты анализа представлены на Фиг.3.
Как видно из результатов, представленных на Фиг.3, в клетках растения N. benthamiana, инфильтрированных вирусом-вектором pA7248amvM2epHBc, экспрессируется белок М2ерНВс, причем это белок является растворимым. Выход целевого продукта составляет 1-2% от растворимой фракции белков.
Пример 5. Образование продуцированным в растениях белком М2ерНВс вирусоподобных частиц.
На следующем этапе вирусоподобные частицы, образованные М2ерНВс, очищали ранее разработанным нами методом осаждения белков сульфатом аммония [Равин Н.В., Киселев О.И., Скрябин К.Г. (2009) Универсальная вакцина против вируса гриппа птиц. Патент РФ №2358981 от 20.06.2009 г.] и ультрацентрифугированием в градиенте сахарозы и хлористого цезия. Как видно из результатов, представленных на Фиг.4, экспрессированный в клетках растения гибридный белок М2ерНВс образует вирусоподобные наноразмерные частицы.
ЛИТЕРАТУРА
1. Giddings, G., Allison, G., Brooks, D., Carter, A. (2000) Transgenic plants as factories for biopharmaceuticals. Nat. Biotechnol. 18 (11), 1151-1155.
2. MacDiarmid, R. (2005) RNA silencing in productive virus infections. Annu. Rev. Phytopathol., 43, 523-544
3. Marillonnet, S., Thoeringer, C, Kandzia, R., Klimyuk, V., Gleba, Y. (2005) Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nat. Biotechnol., 23, 718-723.
4. Neirynck S., Deroo Т., Saelens X., Vanlandschoot P., Jou W.M., Fiers W. (1999) A universal influenza A vaccine based on the extracellular domain of the M2 protein. Nat Med, 5, 1157-1163.
5. Nicholson К., Webster R., Hay A. (1998) Textbook of Influenza. Oxford, Blackwell Science.
6. Webster R., Bean W., Gorman O., Chambers Т., Kawaoka Y. (1992) Microbiol. Rev., 56, 152-179.
7. Комарова Т.В., Скулачев М.В., Зверева А.С., Шварц A.M., Дорохов Ю.Л., Атабеков И.Г. (2006) Новый вирус-вектор для эффективной продукции целевых белков в растениях. Биохимия, 71(8), 1043-1049.
8. Котляров Р.Ю., Куприянов В.В., Мигунов А.И., Степанова Л.А., Цыбалова Л.М., Киселев О.И., Равин Н.В., Скрябин К.Г. (2010) Разработка рекомбинантной вакцины против гриппа A(H1N1) 2009 на основе вирусоподобных наночастиц - носителей внеклеточного домена M2 белка. Acta Naturae, 2(5), 75-80.
9. Марданова B.C., Котляров Р.Ю., Равин Н.В. (2009) Повышение эффективности продукции рекомбинантных белков в растениях за счет оптимизации трансляции РНК вируса-вектора. Молекулярная биология, 43(3), 568-571.
10. Равин Н.В., Киселев О.И., Скрябин К.Г. (2009) Универсальная вакцина против вируса гриппа птиц. Патент РФ №2358981 от 20.06.2009 г.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА "СВИНОГО" ГРИППА H1N1 И СПСОБ ЕЕ ПОЛУЧЕНИЯ | 2010 |
|
RU2451027C2 |
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| КРОСС-РЕАКТИВНАЯ РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА А ЧЕЛОВЕКА | 2020 |
|
RU2742336C1 |
| ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ, СОДЕРЖАЩИЕ БЕЛКИ-МИШЕНИ, СЛИТЫЕ С БЕЛКАМИ ОБОЛОЧКИ РАСТИТЕЛЬНЫХ ВИРУСОВ | 2010 |
|
RU2571223C2 |
| Антиген для универсальной рекомбинантной вакцины против вируса гриппа птиц и человека | 2021 |
|
RU2839428C2 |
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| РЕКОМБИНАНТНЫЙ ВИРУСНЫЙ ВЕКТОР ДЛЯ ПРОДУКЦИИ В РАСТЕНИЯХ БЕЛКА Е1 ВИРУСА КРАСНУХИ (ВАРИАНТЫ) И СИСТЕМА ЭКСПРЕССИИ БЕЛКА Е1 ВИРУСА КРАСНУХИ В КЛЕТКАХ РАСТЕНИЯ (ВАРИАНТЫ) | 2008 |
|
RU2390563C1 |
| ИММУНОГЕННЫЙ ЭПИТОП ВИРУСА ГРИППА | 2009 |
|
RU2546872C2 |
| АНТИГЕН ДЛЯ РЕКОМБИНАНТНОЙ ВАКЦИНЫ ПРОТИВ ВИРУСА КРАСНУХИ | 2019 |
|
RU2709328C1 |
| MDCK клетка-продуцент белков вируса гриппа (варианты) | 2018 |
|
RU2681482C1 |




Изобретение относится к области биотехнологии. Сконструирован оптимизированный для экспрессии в клетках растений синтетический ген, кодирующий гибридный белок М2ерНВс, включающий внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита В. Для продукции этого белка в растениях сконструирован рекомбинантный вирус-вектор на основе генома X-вируса картофеля. Фрагмент генома X вируса картофеля, в который была помещена последовательность гена, кодирующего М2ерНВс, был клонирован под контролем активного в клетках растений промотора, вся конструкция клонирована в пределах т-ДНК области бинарного агробактериального вектора. Вирусный вектор и система экспрессии могут быть использованы для недорогой и эффективной продукции в растениях вирусоподобных частиц на основе ядерного антигена вируса гепатита В, представляющих на своей поверхности полипептиды внеклеточного домена М2 белка вируса гриппа, которые могут являться основой для создания рекомбинантной противогриппозной вакцины. 2 н. и 5 з.п. ф-лы, 4 ил., 1 табл., 5 пр.
1. Рекомбинантный вирусный вектор для продукции в растениях гибридного белка, содержащего показанную на фиг.1 аминокислотную последовательность SEQ ID NO:1 и включающего внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита В, содержащий следующие последовательно расположенные элементы: функционально активный в клетках растения промотор, 5'-нетранслирумый участок генома Х-вируса картофеля, ген полимеразы X-вируса картофеля, первый промотор субгеномной РНК Х-вируса картофеля, нуклеотидная последовательность, кодирующая гибридный белок, включающий внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита В, 3'-нетранслируемый участок генома Х-вируса картофеля и терминатор транскрипции.
2. Рекомбинантный вирусный вектор по п.1, в который между первым промотором субгеномной РНК Х-вируса картофеля и нуклеотидной последовательностью, кодирующей гибридный белок, включающий внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита В, введена нуклеотидная последовательность, соответствующая последовательности 5'-нетранслируемого участка РНК вируса мозаики люцерны.
3. Рекомбинантный вирусный вектор по п.2, в котором промотор представляет собой промотор гена 35S РНК вируса мозаики цветной капусты.
4. Рекомбинантный вирусный вектор по п.3, в котором терминатор транскрипции представлен терминатором гена 35S РНК вируса мозаики цветной капусты.
5. Рекомбинантный вирусный вектор по п.4, представляющий собой плазмиду pA7248amvM2epHBc, в которой элементы объединены между собой как показано на фиг.2, а нуклеотидная последовательность, кодирующая гибридный белок, включающий внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита В, представляет собой нуклеотидную последовательность SEQ ID NO:2.
6. Система экспрессии гибридного белка, включающего внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита, в клетках растения, представляющего собой растение Nicotiana benthamiana, в клетки которого введен фрагмент рекомбинантного вирусного вектора по п.1, включающий функционально активный в клетках растений промотор, 5'-нетранслирумый участок генома Х-вируса картофеля, ген полимеразы Х-вируса картофеля, первый промотор субгеномной РНК Х-вируса картофеля, нуклеотидную последовательность, кодирующую гибридный белок, включающий внеклеточный домен белка М2 вируса гриппа, присоединенный к ядерному антигену вируса гепатита В, 3'-нетранслируемый участок генома Х-вируса картофеля и терминатор транскрипции.
7. Система экспрессии по п.6, представляющая собой растение Nicotiana benthamiana, в клетки которого с использованием бактерии Agrobacterium tumefaciens введен фрагмент рекомбинантного вирусного вектора pA7248amvM2epHBc, показанный как т-ДНК на фиг.2.
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| US 2007036826 A1, 15.02.2007 | |||
| NEIRYNCK S | |||
| et al., "A universal influenza A vaccine based on the extracellular domain of the M2 protein", Nat Med | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |

