Область техники
Настоящее изобретение относится к области иммунологии и биотехнологии, а именно к методам получения препаратов, формула которых в перспективе может быть использована как основа для разработки универсальной вакцины против вируса гриппа А. Изобретение может быть использовано в медицине, ветеринарии и биотехнологии.
Уровень техники
Грипп - высоко контагиозная острая респираторная вирусная инфекция. Согласно данным Всемирной организации здравоохранения (ВОЗ) ежегодно вирус гриппа заражает около 1 млрд человек во всем мире, при этом около 3-5 млн человек переносят болезнь тяжело (Krammer et al., 2018). По оценкам, полученным в рамках проекта «Глобальная смертность от гриппа» (Global Influenza Mortality project, GLaMOR), от 294000 до 518000 смертей ежегодно обусловлено вирусом гриппа (Paget et al., 2019).
Симптомы, связанные с инфицированием вирусом гриппа, варьируют от легких, типичных для респираторных заболеваний проявлений, до тяжелых осложнений в результате вторичных бактериальных инфекций нижних дыхательных путей, в том числе пневмоний с летальными исходами (Krammer et al., 2018). Кроме того, инфекция вирусом гриппа может привести и к широкому спектру нереспираторных осложнений, среди которых кардиологические, почечные и неврологические нарушения (Chow et al., 2020). Наиболее уязвимой к постинфекционным осложнениям группой населения являются люди с хроническими заболеваниями, пожилые, дети до двух лет, беременные женщины и лица с ослабленным иммунитетом (Egorov, 2016).
Ежегодные эпидемии гриппа, которые связаны, в том числе, и с низкой эффективностью традиционно применяемых вакцин, являются причиной существенных экономических издержек, что вызвано снижением продуктивности труда сотрудников компаний на фоне заболевания и последующих осложнений (Sah et al., 2019).
Грипп ассоциирован с инфицированием одним из четырех вирусов гриппа (А, В, С, D), при этом только вирусы гриппа А и В являются причиной сезонных эпидемий, и исключительно вирус гриппа А - глобальных пандемий (Krammer et al., 2018). Вирус гриппа А относится к семейству Orthomyxoviridae и имеет сегментированный РНК-геном отрицательной полярности. 8 сегментов вирусной РНК кодируют 17 белков, среди которых поверхностные гемагглютинин (hemagglutinin, НА), нейраминидаза (neuraminidase, NA), белок, образующий ионные каналы М2 (Chen et al., 2018). НА и NA - два основных поверхностных белка вириона. На данный момент известно 16 типов гемагглютинина (НА 1-16) и 9 типов нейраминидазы (NA 1-9) (Jang & Вае, 2018), помимо этого известны 2 варианта НА (17, 18) и 2 варианта NA (10, 11), специфические для летучих мышей.
Сегментированный геном ортомиксовирусов обуславливает возможность реассортации при инфицировании одной и той же клетки различными штаммами. Вирус гриппа А при этом способен заражать не только человека, но и сельскохозяйственных животных, а также диких водоплавающих птиц. Именно реассортация вируса гриппа А в организме диких птиц является одной из причин его высокой антигенной изменчивости и, таким образом, развития пандемий зоонозного происхождения. Помимо механизма реассортации существенный вклад в изменчивость антигенных свойств вируса гриппа вносит высокий темп накопления точечных мутаций (антигенный дрейф) (Egorov, 2016).
На основании разнообразия поверхностных гликопротеинов вируса гриппа А теоретически возможно существование 198 штаммов, однако на данный момент известно, что только 7% потенциально возможных штаммов могут быть опасны для человека (Jang & Вае, 2018). Тем не менее, все комбинации на основе НА (1-16) и NA (1-9) были обнаружены у диких водоплавающих птиц (de Vries et al., 2020).
НА играет ключевую роль в выработке нейтрализующих антител после болезни или вакцинации. Антитела, специфичные к NA не способны ингибировать вирусную инфекцию, но способствуют подавлению репликации и снижению тяжести заболевания (Marcelin et al., 2012). Антитела, вырабатывающиеся к вирусу гриппа А с определенным типом НА не обеспечивают протективность по отношению к штаммам с другим типом НА. Традиционные вакцины эффективны исключительно в случае совпадения антигенных вариантов циркулирующих и вакцинных штаммов вирусов, которые прогнозируются службой по эпидемиологическому надзору ВОЗ. При этом существует непреодолимый временной разрыв (от 6 до 9 месяцев) между этапом идентификации пандемического штамма и началом применения вакцины (Krammer & Palese, 2015). В связи с перечисленными факторами эпидемиологическая эффективность существующих традиционных вакцин может быть существенно ниже предполагаемой.
Согласно оценкам, сделанным Мао с соавторами (Мао et al., 2012), экономические последствия массовых заболеваний гриппом могут составлять от 13,9 тыс. до 957,5 млн долларов (в среднем около 2,47 млн долларов) в различных регионах США в зависимости от демографической ситуации, экономического благополучия и уровня развития системы здравоохранения. В исследовании Sah с соавторами (2019) для США было спрогнозировано, что замена всего лишь 10% используемых на данный момент сезонных вакцин на универсальную вакцину с 75% эффективностью сможет предотвратить около 5,3 миллиона случаев заболевания, 81 тыс. случаев госпитализации и 6,3 тыс.смертей от гриппа в год. Такое снижение заболеваемости поможет сэкономить более 1,1 млрд долларов прямых затрат на здравоохранение (в сравнении со средними затратами за период с 2010-11 по 2018-19 годы). При полной замене существующих сезонных вакцин универсальными вакцинами возможно предотвратить до 17 миллионов случаев заболеваний, 251 тыс. случаев госпитализаций, 19,5 тыс.смертей и 3,5 млрд долларов прямых затрат на здравоохранение. Ожидается, что страны с высокими медицинскими расходами и большой долей пожилых жителей получат максимальную экономическую выгоду от использования универсальных противогриппозных вакцин, так что замена даже части сезонных вакцин универсальными оправдывает значительную стоимость разработки универсальной вакцины (Sah et al., 2019).
Одной из перспективных мишеней для разработки универсальной вакцины против вируса гриппа А является эктодомен М2е минорного поверхностного белка М2 (Egorov, 2016). Белок М2 содержит 97 аминокислотных остатков (а.о.) (Lamb & Choppin, 1981), организованных в 3 структурных домена: короткий N-концевой (24 а.о.) домен М2е, экспонированный на поверхности вириона, трансмембранный (19 а.о.) и С-концевой участки (54 а.о.). Тетрамеры М2 формируют ионные каналы (Holsinger at al., 1994). Число молекул М2, образующих ионные каналы, составляет не более 2% от количества молекул НА, экспонированного на поверхности вирусной частицы (Feng et al., 2006).
Впервые М2 был идентифицирован в 1981 г. Lamb и Choppin. Вскоре после выявления М2-белка в ходе вирусной инфекции впервые были получены моноклональные антитела 14С2 (Zebedee & Lamb, 1988). Было установлено, что несмотря на невозможность 14С2 ингибировать адсорбцию вируса и его последующее проникновение в клетку, скорость роста бляшек в клеточных культурах в случае присутствия М2е-специфических антител оказывалась ниже для некоторых из штаммов вируса. Уменьшение скорости репликации вируса в данном эксперименте, по-видимому, обусловлено снижением уровня экспрессии М2, что косвенно негативно влияет на уровень продукции вирусного потомства (Hughey et al., 1995). При этом нечувствительность некоторых штаммов вируса к 14С2 обусловлена мутационной изменчивостью в области С-концевого домена белка М2 (Zebedee & Lamb, 1990). По-видимому, мутации в С-концевом участке, взаимодействующем с матриксным белком Ml, влияют на конформацию N-концевого домена, что обуславливает отсутствие ограничивающего рост бляшек эффекта 14С2 (Zebedee & Lamb, 1990).
В ходе естественной инфекции гриппа выявляемый титр антител к М2е-эпитопу оказывается низким, антитела присутствуют в сыворотке крови непродолжительное время (Feng et al., 2006) и не обладают нейтрализующей активностью. Тем не менее, несмотря на отсутствие нейтрализующих свойств, антитела к М2е играют существенную роль в формировании устойчивого иммунного ответа (Mezhenskaya et al., 2019).
В опытах in vitro показано подавление репликации вируса при взаимодействии с сывороткой, полученной путем иммунизации мышей препаратом на основе М2е-эпитопа (Liu et al., 2003), слитым с белком-носителем. Иммунизация мышей антигеном М2НВс, представляющем собой эпитоп М2е, слитый с носителем - НВс-антигеном вируса гепатита С, с 90-100% эффективностью приводит к снижению летальных случаев при заражении адаптированным к мышам штаммом Х47 (H3N2), 5LD50 (Neirynck et al., 1999). При иммунизации мышей синтетическим пептидом на основе М2е-эпитопа с белковым носителем был показан in vivo протективный эффект и снижение уровня летальных осложнений при заражении вирусами гриппа штаммов H1N1 и H3N1, LD90. Существенно, что аналогичный эффект был достигнут и при пассивном переносе сыворотки, содержащей высокие титры М2е специфических антител (Fan et al., 2004).
На данный момент механизмы обеспечения протективного эффекта М2е-специфическими антителами все еще не до конца ясны в полной мере и не могут быть обобщены в общую последовательную схему формирования устойчивости к инфицированию вирусом гриппа.
В работе Bakkouri с соавторами (2011) на мышах было продемонстрировано, что протективный эффект М2е специфических антител в значительной степени обусловлен активацией Fcγ-рецепторов. Кроме того, известно, что на поверхности клеточной мембраны инфицированных клеток М2 присутствует в количестве, превышающем таковое на поверхности вирионов и составляет до 50% от числа молекул НА, встроенного в клеточную мембрану (Feng et al., 2006). В связи с этим, по-видимому, антитела к М2е могут обеспечивать протективный эффект за счет связывания с поверхностью зараженных клеток (Bakkouri et al., 2011, Zebedee & Lamb, 1989) и активации системы комплимента или запуска антителозависимого цитотоксического иммунного ответа (Mozdzanowska et al., 1999). Участие С3-компонента комплемента в снижении титра вируса, обнаруживаемого в тканях легких, было показано на мышиной модели в работе Wang с соавторами (2008). Таким образом, М2е-специфические антитела способны активировать различные ключевые компоненты иммунного ответа, что в результате обуславливает их протективный эффект.
Концепция применения М2е-эпитопа в качестве антигена при создании вакцин кардинально отличается от традиционного подхода, который предполагает формирование гуморального ответа на НА и NA. Вирус гриппа А за счет механизмов антигенного дрейфа и антигенного шифта быстро развивает устойчивость и нечувствительность к НА- и NA-специфическим антителам. При этом в случае использования М2е-эпитопа скорость мутаций будет значительно ниже. Мутантные ускользающие от иммунного ответа штаммы, имеющие 1 аминокислотную замену в позиции 10, были выявлены при введении М2е-специфических моноклональных антител мышам с иммунодефицитом (Zharikova et al., 2005). Однако при пассажах мутантных штаммов на иммунокомпетентных клетках животных, иммунизированных антигеном на основе М2е, подобных ускользающих мутантов выявлено не было (Gerhard et al., 2006), что позволяет ожидать стабильный во времени протективный эффект после вакцинации препаратами на основе М2е.
Аминокислотная последовательность эктодомена М2е вируса гриппа А практически инвариантна для всех штаммов (Liu et al., 2003). Первые 9 аминокислотных остатков MSLLTEVET являются высоко консервативными и совпадают для штаммов птиц и человека (Bakkouri et al., 2011), в остальных позициях уровень мутационной изменчивости варьирует для вирусов, выделенных из различных организмов-хозяев (человек, птицы, свиньи) (Mezhenskaya et al., 2019), что позволяет выделить консенсусные последовательности М2е на основании специфичности к организму-хозяину. При этом существенно, что при иммунизации консенсусной для человека последовательностью М2е, протективный эффект в отношении свинных штаммов отсутствует (Heinen et al., 2002). В то же время моноклональные антитела 8С6, узнающие человеческий консенсус «EVETPIRN», не взаимодействуют с последовательностью птичьего консенсуса «EVETPTRN» (Liu et al., 2004), что свидетельствует о необходимости использования нескольких консенсусов для создания универсальной вакцины.
Прохождение межвидового барьера и инфицирование человека штаммами птичьего гриппа приводит к тяжелому течению болезни с высоко вероятным летальным исходом (Liu et al., 2005). В связи с этим целью изобретения являлась разработка подхода к созданию бивалентного антигена для вакцины, обеспечивающей эффективную защиту как против штаммов вируса гриппа А, специфических для человека, так и для штаммов вирусов гриппа птиц, представляющих особую опасность для людей в предпандемический период. Вакцина на основе предложенного антигена, с одной стороны, может быть применена для вакцинации домашней птицы, что поможет предотвратить распространение птичьего гриппа в птицеводческих хозяйствах и уменьшить как экономические убытки, так и риски заражения человека штаммами вируса гриппа птиц. С другой стороны, такая вакцина может обеспечить защиту человека как от сезонных, так и от новых штаммов вируса гриппа, образованных вследствие антигенного шифта между штаммами вируса гриппа птиц и человека.
В качестве прототипа изобретения может быть рассмотрен патент Равина Н.В. с соавторами (RU 2358981 С2, 20.06.2009), в котором предложен основной компонент для универсальной вакцины против вируса гриппа птиц, представляющий собой вирусоподобные частицы на основе НВс-антигена вируса гепатита В, несущие на своей поверхности М2е-эпитопы вируса гриппа птиц (штаммы H5N1, H5N2). Ключевым преимуществом предложенной разработки в сравнении с приведенным прототипом является возможность ее применения для создания бивалентной вакцины, обеспечивающей иммунопротективность к штаммам вируса гриппа А, специфических не только для птиц, но и для человека. Кроме того, антигены, использованные в указанном прототипе, содержат М2е-эпитопы только 2 штаммов вируса гриппа птиц: A/Duck/Potsdam 1402-6/1986 (H5N2), A /Chicken/Kurgan/05/2005 (H5N1), в то время как в состав предложенного в разработке антигена включена консенсусная аминокислотная последовательность М2е-эпитопа, выведенная на основании более широкого спектра штаммов: H5N1, H5N3, H5N4, H5N5, H5N6, H5N8, H5N9, H7N3, H7N7, H7N9.
Раскрытие сущности изобретения
Задачей данного изобретения является создание антигена, универсального в отношении специфических для птиц и человека штаммов вируса гриппа А. Данный антиген - ТМ2е - представляет собой рекомбинантный белок на основе высоко консервативного эктодомена М2е белка М2, образующего ионные каналы на поверхности вириона.
Сформулированная задача решается путем создания генетической конструкции для экспрессии в бактериальной культуре клеток Escherichia coli рекомбинантного белка ТМ2е с последовательностью, которая состоит, начиная с N-конца, из остатка метионина, гексагистидинового повтора, необходимого для выделения и очистки рекомбинантного белка, трехкратных повторов двух последовательностей аминокислот, консенсусных для М2е-эпитопа (2-24 а.о.) вирусов гриппа А птиц и человека, а также линкерных глицин-обогащенных участков, разделяющих повторы консенсусов (Фигура 2Б). В качестве консенсуса М2е, специфического для штаммов вируса гриппа А человека, в соответствии с литературными данными (Mezhenskaya et al., 2019) использована последовательность SLLTEVETPIRNEWGCRCNDSSD. В качестве консенсуса М2е птичьих штаммов выбрана последовательность SLLTEVETPTRNEWECRCSD. Консенсусная последовательность птичьего М2е-эпитопа отличается от такой для человека аминокислотами в положении 11, 16, 20 (Фигура 2А).
Синтетический фрагмент ДНК, содержащий ген описанного рекомбинантного антигена, был клонирован в экспрессионный вектор pQE30, содержащий Т5-промотор, репрессируемый LacI и индуцируемый изопропил-β-D-1-тиогалактопиранозидом (ИПТГ). Клетки E. coli (штамм SG13009) были трансформированы полученной конструкцией рТМ. Было показано, что целевой белок экспрессируется в растворимой форме.
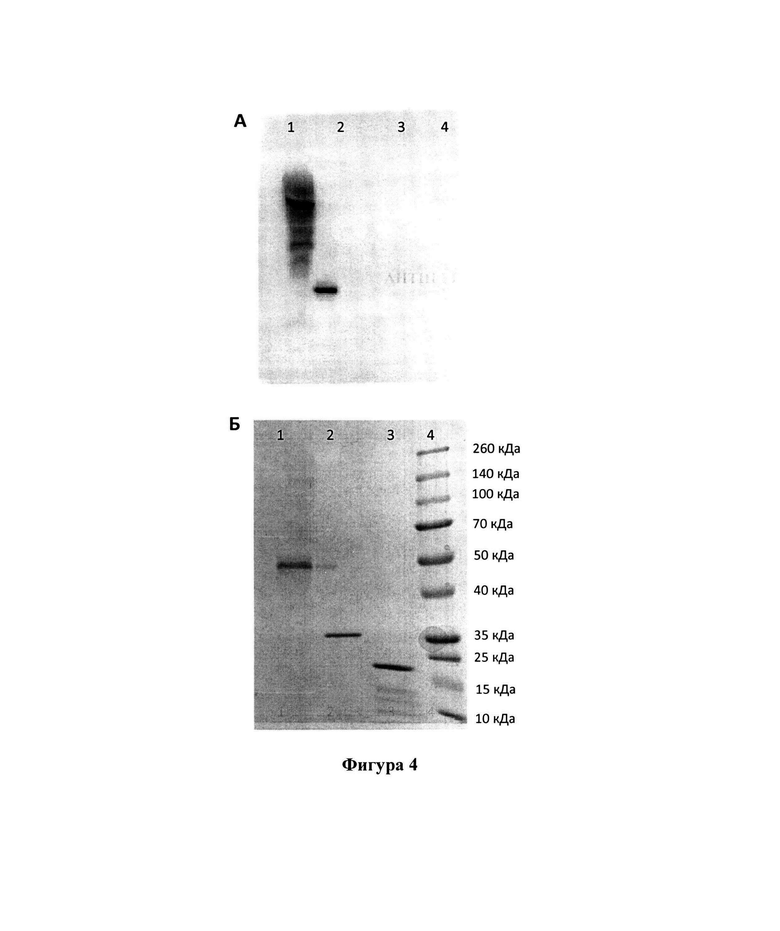
Накопленный рекомбинантный антиген был выделен и очищен методом афинной хроматографии. Получение целевого рекомбинантного антигена подтверждается методом электрофоретического анализа в денатурирующих условиях (8-20% ДСН-ПААГ-ЭФ). Большая часть рекомбинатного белка-антигена находится в форме олигомеров (Фигура 4А, расчетная молекулярная масса белка составляет 24 кДа), образующихся за счет дисульфильдных связей между остатками цистеинов (в последовательности присутствует 12 а.о. цистеина), которыми обогащена аминокислотная последовательность. Антигенная специфичность выделенного и очищенного белка подтверждается иммуноблот анализом с поликлональной мышиной антисывороткой, полученной при иммунизации неродственным рекомбинантным белком, представляющем собой М2е-эпитоп, конъюгированный с дегидрофолат редуктазой (DHFR-M2e) (Karpova et al., 2012) (Фигура 4A).
Техническим результатом предлагаемого изобретения является способ получения антигена для рекомбинантной вакцины для профилактики штаммов вируса гриппа А, специфических для птиц и человека.
Краткое описание чертежей
Фигура 1 представляет собой нуклеотидную последовательность рекомбинантного антигена ТМ2е в контексте плазмидного вектора pQE30. Стартовый кодон ATG и терминирующий кодон ТАА выделены жирным шрифтом и подчеркнуты.
Последовательность, кодирующая шесть гистидинов (His6), выделена жирным шрифтом. Сайты разрезания эндонуклеазами рестрикции BamHI и HindIII подчеркнуты и подписаны. Квадратными скобками и жирным шрифтом выделены последовательности, кодирующие М2е-эпитоп: последовательность, специфическая для штаммов, инфицирующих человека, дополнительно выделена серым.
Фигура 2 представляет собой таблицу, демонстрирующую различия в аминокислотных последовательностях консенсусов М2е-эпитопа для вируса гриппа А, специфических для птиц и человека (А), а также аминокислотную последовательность рекомбинантного белка ТМ2е (Б). Последовательность М2е, специфическая для человека, выделена жирным шрифтом на сером фоне, последовательность М2е, специфическая для птичьих штаммов, выделена исключительно жирным шрифтом. Шесть гистидинов (His6) выделены курсивом. Стартовый метионин подчеркнут.
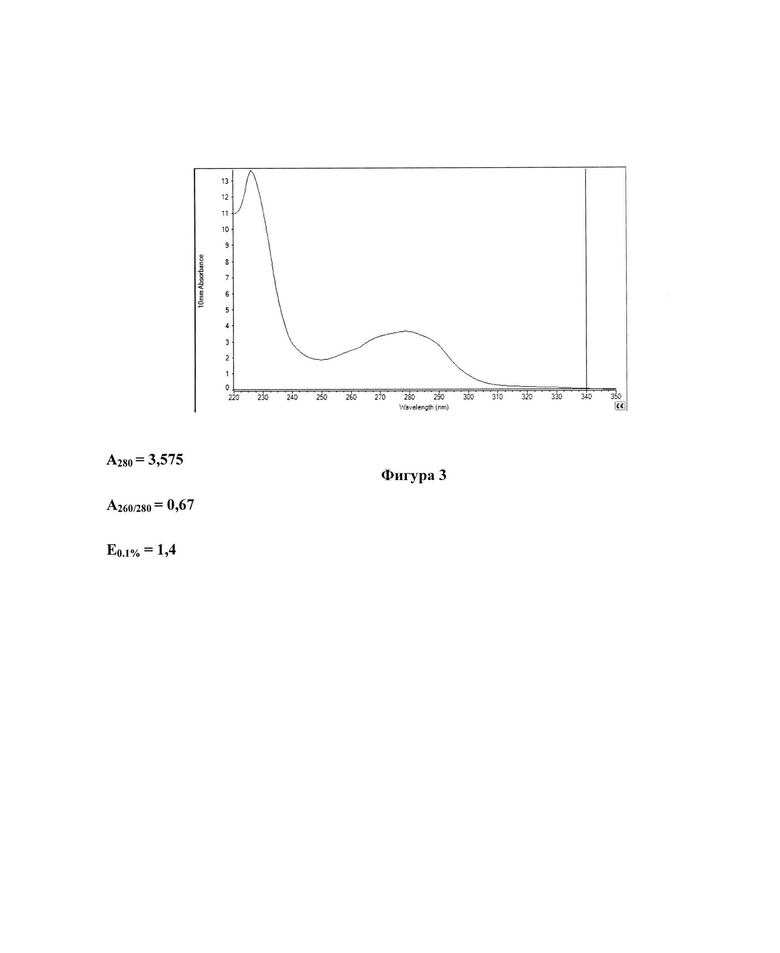
Фигура 3 представляет собой спектр поглощения водного раствора рекомбинантного белка ТМ2е, определенный с помощью спектрофотометра NanoDrop 2000 (Thermo Fisher Scientific, США). Указаны значения поглощения при длине волны 280 нм, соотношение значений поглощения при 260 и 280 нм, а также E0.1% (коэффициент поглощения белка ТМ2е с концентрацией 1 мг/мл при длине оптического пути 1 см). Е0.1% для белка ТМ2е был рассчитан с помощью сервиса https://web.expasy.org/protparam/.
Фигура 4 представляет иммуноблот анализ эффективности взаимодействия рекомбинантного белка ТМ2е с поликлональной мышиной антисывороткой к DHFR-M2e (Karpova et al., 2012) (разведение 1:5000). Описание дорожек: 1. - ТМ2е. 2. - DHFR-M2e. 3. - БО ВТМ (отрицательный контроль). 4. - маркеры молекулярной массы. Белки разделяли в 8-20% ДСН-ПААГ-ЭФ, после чего переносили на мембрану Hybond-P (GE Healthcare Life Sciences, Великобритания). Для проявления результатов иммуноблот анализа использовали вторичные противомышиные антитела, конъюгированные с пероксидазой хрена (разведение 1:20000) (Jackson ImmunoResearch, Великобритания). Визуализация результатов анализа проводилась с помощью ECL системы («Advansta», США). Б. Параллельный электрофоретический анализ - 8-20% ДСН-ПААГ-ЭФ, окрашенный Кумасси G-250. 1. - ТМ2е. 2. - DHFR-M2e. 3. - БО ВТМ (отрицательный контроль). 4. - маркеры молекулярной массы.
Осуществление изобретения
В соответствии с настоящим изобретением антиген для профилактической вакцины против вируса гриппа А представляет собой рекомбинантный белок (ТМ2е), состоящий из трех повторов двух аминокислотных мотивов М2е-эпитопа (2-24 а.о.), консенсусных для штаммов вируса гриппа А птиц и человека (Фигура 2Б). Выбор консенсуса, специфического для штаммов человека, был осуществлен в соответствии с полученными раннее данными анализа штаммов, выделенных от 27253 инфицированных людей (Mezhenskaya et al., 2019). Выбор консенсуса, специфического для штаммов птиц, был произведен на основерезультатов выравнивания с помощью программного обеспечения JalView 2.11.1.3. В качестве основы для выведения консенсусной последовательности М2е были выбраны аминокислотные мотивы следующих штаммов вируса гриппа A: H5N1, H5N3, H5N4, H5N5, H5N6, H5N8, H5N9, H7N3, H7N7, H7N9. Выбор штаммов был обусловлен тем, что в настоящий момент преобладающими штаммами вируса гриппа А в популяции человека являются штаммы, относящиеся к субтипам H1, Н2, Н3 (Liu et al., 2005), в связи с чем устойчивость в другим субтипам остается по-прежнему на низком уровне. На основе ежегодного мониторинга ВОЗ циркулирующих штаммов вируса гриппа А наиболее часто выявляемые у человека субтипы птичьего гриппа - Н5, Н7, Н9 (World Health Organization, 2021).
Приведенные примеры конкретного осуществления изобретения приведены для предоставления специалистам в данной области техники полного описания проведения и применения анализа по изобретению, и подразумевают, что приведенные примеры не ограничивают предполагаемый авторами изобретения объем изобретения.
Изобретение осуществляется следующим способом.
Плазмида рТМ получена путем клонирования нуклеотидных последовательностей, кодирующих М2е-эпитопы штаммов вируса гриппа А, специфических для птиц и человека, в вектор pQE30. Рекомбинантный белок ТМ2е с молекулярной массой 24 кДа, включающий в себя три повтора двух консенсусных последовательной М2е-домена вируса гриппа А птиц и человека, накоплен в бактериальной системе клеток Е. coli штамма SG13009 путем трансформации этих клеток плазмидой рТМ, содержащей нуклеотидную последовательность Seq ID 02, кодирующую белок ТМ2е. Экспрессированный белок выделен и очищен из бактериальных клеток методом металл-хелатной хроматографии и переведен в водный раствор посредством диализа. Анализ полученного белка, а также его специфических свойств осуществляется с использованием методик электрофореза в денатурирующих условиях в полиакриламидном геле, спектрофотометрии и иммуноблот-анализа.
Сущность изобретения поясняют следующие примеры, не ограничивая ее.
Пример 1
Продукция рекомбинантного антигена ТМ2е, содержащего М2е-эпитопы вируса гриппа А, специфического для штаммов птиц и человека, в бактериальной культуре Escherichia coli
На Фигуре 2Б представлена аминокислотная последовательность рекомбинантного антигена ТМ2е (Seq ID 01). Для накопления целевого белка были использованы клетки Е. coli штамма SG13009, содержащие плазмиду pRep4. Бактериальные клетки были трансформированы плазмидой рТМ, содержащей последовательность Seq ID 02, кодирующую целевой белок ТМ2е и использовались в дальнейшем в качестве основного продуцента.
На первом этапе продуцент был высеян в объеме 3 мл культуральной среды 2YT с добавлением селективных антибиотиков - ампициллина (итоговая концентрация 100 мкг/мл) и канамицина (итоговая концентрация 50 мкг/мл). Клетки были проинкубированы на качалке с24 Incubator Shaker (New Brunswick Scientific, США) в течение 14-16 часов при +37°С (180 об/мин). Полученная ночная культура была перенесена в больший объем питательной среды (200 мл 2YT с добавлением ампициллина и канамицина до конечной концентрации 100 мкг/мл и 50 мкг/мл соответственно) и культивировались на качалке при +37°С до оптической плотности 0,7-0,9 при длине волны λ=600 нм. При достижении заданной плотности для индукции экспрессии целевого белка к культуре клеток был добавлен ИПТГ до конечной концентрации 2 мМ. Индуцированная культура выращивалась в течение 4 часов на качалке при +37°С. Далее клеточная культура была отцентрифугирована при 5000 об/мин в течение 10 минут (центрифуга Beckman J2-21, ротор JA-14) при +4°С. Полученные осадки использовали для выделения рекомбинантного белка.
Осадки суспендировали в 0,05 М Трис-HCl pH 7,8 (10 мл буфера на 1 г осадка), содержащем фенилметилсульфонил фторид в концентрации 0,001 М в качестве ингибитора сериновых протеаз. Полученную суспензию клеток подвергали ультразвуковой дезинтеграции на ультразвуковом гомогенизаторе Q500 (QSonica, США), параметры: 150 циклов с продолжительностью работы 2 сек и интервалами 6 сек. Лизат клеток осветляли низкоскоростным центрифугированием 20 минут при 12 000 об/мин (центрифуга Beckman J2-21, ротор JA-14) при +4°С. Согласно результатам 8-20% ДСН-ПААГ-ЭФ целевой белок ТМ2е содержится преимущественно в супернатанте, образующемся после низкоскоростного центрифугирования, что свидетельствует о том, что целевой белок экспрессируется в растворимой форме. Для последующих стадий выделения и очистки был использован полученный описанным способом супернатант с добавлением NaCl до итоговой концентрации 0,3 М.
Для выделения и очистки ТМ2е из супернатанта проводили металл-хелатную хроматографию с применением Ni2+- нитрилотриацетатной агарозы (Ni-HTA) в качестве сорбента. Супернатант был нанесен на колонку, содержащую 0,8 мл сорбента, предварительно уравновешенного стартовым буфером (0,05 М Трис-HCl, рН 7,8; 0,3 М NaCl). Сорбцию белка на Ni-HTA агарозе осуществляли не менее часа. Далее промывали агарозу стартовым буфером (объемом не менее десятикратного объема сорбента), затем буфером, содержащем 0,03 М имидазол (0,05 М Трис-HCl, рН 8,0; 0,3 М NaCl; 0,03 М имидазол) объемом не менее пятикратного объема сорбента. Элюцию проводили буфером, содержащем 0,25 М имидазол (0,05 М Трис-HCl, рН 8,0; 0,3 М NaCl; 0,25 М имидазол), в объеме 1-1,5 объемов сорбента. Инкубировали Ni-HTA агарозу с элюирующим буфером в течение 10 минут, затем собирали целевую фракцию. Наиболее прочно связанные с сорбентом молекулы белка были элюированы буфером с 0,5 М имидазолом (0,05 М Трис-HCl, рН 8,0; 0,3 М NaCl; 0,5 М имидазол) в объеме двух объемов сорбента.
Очищенный белок ТМ2е переводили в водный раствор путем диализа. Для диализа использовали диализные мешки Servapor (Германия) диаметром 6 мм и порами, пропускающими молекулы с массой 12-14 кДа. Диализ проводили против MilliQ (соотношение раствор белка/вода 1:250) в течение 4 часов, при этом смена воды осуществлялась каждый час. Отдиализованный белок был осветлен низкоскоростным центрифугированием при +4°С (13,2 тыс. об/мин, 15 мин; центрифуга 5415D).
Полученный белок ТМ2е был проанализирован с помощью 8-20% ДСН-ПААГ-ЭФ. Согласно результатам, представленным на Фигуре 3, ТМ2е был успешно выделен и переведен в водный раствор. ТМ2е представлен преимущественно димерной формой (Фигура 4Б, дорожка 1), образующейся, по-видимому, благодаря насыщенности белка аминокислотными остатками цистеина (12 а.о.). Расчетная Mr димера ТМ2е составляет 48кДа (в соответствии с молекулярной массой мономера, полученной с использованием программы ProtParam сервиса https://web.expasy.org/protparam/), что согласуется с его расположением на электрофореграмме между маркерами молекулярной массы 40 кДа и 50 кДа (Фигура 4Б, дорожка 4).
Пример 2
Иммуноблот анализ антигенной специфичности рекомбинантного белка ТМ2е, содержащего М2е-эпитопы вируса гриппа А, специфического для штаммов птиц и человека
Анализ антигенной специфичности полученного рекомбинантного белка ТМ2е был проведен с помощью метода иммуноблот анализа с поликлональной антисывороткой к рекомбинантному белку DHFR-M2e (Karpova et al., 2012). В качестве положительного контроля был выбран DHFR-M2e. В качестве отрицательного контроля был использован белок оболочки вируса табачной мозаики (ВТМ).
После разделения белков в 8-20% ДСН-ПААГ методом электрофореза белки из геля были перенесены на поливинилиденфторидную мембрану Hybond-P (GE Healthcare Life Sciences, Великобритания) методом электропереноса (прибор Mini Trans-Blot cell, Bio-Rad, режим переноса: 16 часов, 30 V). После переноса мембрана была проинкубирована в 5% растворе сухого молока в буфере tTBS (0,01 М Трис-HCl рН 7,4; 0,15 М NaCl; 2% твин-20) в течение 1 часа на качалке при комнатной температуре. Далее была проведена отмывка мембраны 3 раза по 10 минут в буфере tTBS. На следующем этапе мембрану инкубировали с первичными антителами (мышиные поликлональные, полученные к DHFR-M2e) в разведении 1:5000 в 5% растворе сухого молока на tTBS в течение 1 часа на качалке при комнатной температуре, далее мембрана была отмыта 3 раза по 10 минут в буфере tTBS и проинкубирована со вторичными противомышиными антителами, конъюгированными с пероксидазой хрена («Jackson ImmunoResearch», разведение 1:20000) в 5% молоке на tTBS 1 час при комнатной температуре на качалке. Мембрана была отмыта 2 раза по 10 минут в буфере tTBS. Последняя отмывка проводилась в течение 10 минут в TBS без добавления твина-20. Проявка мембраны осуществлялась с помощью ECL системы «Advansta» (США). Хемилюминесцентный сигнал детектировали с помощью гельдокументирующей системы Chemi Doc XRS+ с программным обеспечением Image Lab Software (Bio Rad, США).
Из Фигуры 4А видно, что ТМ2е (Фигура 4А, дорожка 1) так же, как и DHFR-M2e (Фигура 4А, дорожка 2), взаимодействует с поликлональной антисывороткой к DHFR-M2e. Отрицательный контроль БО ВТМ (Фигура 4А, дорожка 3), маркеры молекулярной массы (Фигура 4А, дорожка 4) с данной антисывороткой не взаимодействуют. Взаимодействие ТМ2е с поликлональной антисывороткой к DHFR-M2e подтверждает возможность использования рекомбинантного белка в качестве антигена при создании вакцины против вируса гриппа. Параллельный электрофоретический анализ - 8-20% ДСН-ПААГ-ЭФ, окрашенный Кумасси G-250, представлен на Фигуре 4Б.
Литература
Патенты
1. Равин Н.В., Киселев О.И., Скрябин К.Г. Универсальная вакцина против вируса гриппа птиц // Патент RU 2358981 C2, 20.06.2009.
Непатентная литература
1. Bakkouri K, Descamps F, De Filette М et al. Universal vaccine based on ectodomain of matrix protein 2 of influenza A: Fc receptors and alveolar macrophages mediate protection. J Immunol. 2011;186(2): 1022-31.
2. Chen X, Liu S, Goraya MU et al. Host immune response to influenza A virus infection. Front Immunol. 2018;9:320.
3. Chow EJ, Rolfes MA, O'Halloran A et al. Respiratory and nonrespiratory diagnoses associated with influenza in hospitalized adults. JAMA Netw Open. 2020;3(3):e201323.
4. de Vries E, Du W, Guo H et al. Influenza A virus hemagglutinin-neuraminidase-receptor balance: preserving virus motility. Trends Microbiol. 2020;28(1):57-67.
5. Egorov A. The challenges of creating a universal influenza vaccine. MIR J, 2016;3(1),31-41.
6. Fan J, Liang X, Horton MS et al. Preclinical study of influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine. 2004;22(23-24):2993-3003.
7. Feng J, Zhang M, Mozdzanowska K et al. Influenza A virus infection engenders a poor antibody response against the ectodomain of matrix protein 2. Virol J 3, 2006;3:102.
8. Gerhard W, Mozdzanowska K, Zharikova D. Prospects for universal influenza virus vaccine. Emerg. Infect. Dis 2006;12(4):569-574.
9. Heinen P, Rijsewijk F, de Boer-Luijtze E et al Vaccination of pigs with a DNA construct expressing an influenza virus M2-nucleoprotein fusion protein exacerbates disease after challenge with influenza A virus. J Gen Virol. 2002;83(Pt 8):1851-1859.
10. Holsinger L, Nichani D, Pinto L et al. Influenza A virus M2 ion channel protein: a structure-function analysis. J Virol. 1994;68(3): 1551-63.
11. Hughey P, Roberts P, Holsinger L et al. Effects of antibody to the influenza a virus M2 protein on M2 surface expression and virus assembly. Virology. 1995;212(2):411-21.
12. Jang J, Вае SE. Comparative Co-Evolution Analysis Between the HA and NA Genes of Influenza A Virus. Virology (Auckl). 2018;9:1178122X18788328.
13. Karpova O, Nikitin N, Chirkov S et al. Immunogenic compositions assembled from tobacco mosaic virus-generated spherical particle platforms and foreign antigens. J Gen Virol. 2012;93:400-407.
14. Krammer F, Palese P. Advances in the development of influenza virus vaccines. Nat Rev Drug Discov. 2015; 14(3): 167-82.
15. Krammer F, Smith G, Fouchier R et al. Influenza. Nat Rev Dis Primers. 2018;28;4(1):3.
16. Lamb R, Choppin P. Identification of a second protein (M2) encoded by RNA segment 7 of influenza virus. Virology. 1981;112:729-737.
17. Liu W, Li H, Chen Y. N-terminus of M2 protein could induce antibodies with inhibitory activity against influenza virus replication. FEMS Immunol Med Microbiol. 2003;35(2): 141-6.
18. Liu W, Zou P, Chen YH. Monoclonal antibodies recognizing EVETPIRN epitope of influenza A virus M2 protein could protect mice from lethal influenza A virus challenge. Immunol Lett. 2004;93(2-3):131-6.
19. Liu W, Zou P, Ding J et al. Sequence comparison between the extracellular domain of M2 protein human and avian influenza A virus provides new information for bivalent influenza vaccine design. Microbes Infect. 2005;7(2):171-7.
20. Mao L, Yang Y, Qiu Y et al. Annual economic impacts of seasonal influenza on US counties: spatial heterogeneity and patterns. Int J Health Geogr. 2012; 11:16.
21. Marcelin G, Sandbulte M, Webby R. Contribution of antibody production against neuraminidase to the protection afforded by influenza vaccines. Rev Med Virol. 2012;22(4):267-279.
22. Mezhenskaya D, Isakova-Sivak I, Rudenko L. M2e-based universal influenza vaccines: a historical overview and new approaches to development. J Biomed Sci. 2019;26(1):76.
23. Mozdzanowska K, Maiese K, Furchner M et al. Treatment of influenza virus-infected SCID mice with nonneutralizing antibodies specific for the transmembrane proteins matrix 2 and neuraminidase reduces the pulmonary virus titer but fails to clear the infection. Virology. 1999;254(1): 138-46.
24. Neirynck S, Deroo T, Saelens X et al. A universal influenza A vaccine based on the extracellular domain of the M2 protein. Nat Med. 1999 Oct;5(10):1157-63.
25. Paget J, Spreeuwenberg P, Charu V et al. Global mortality associated with seasonal influenza epidemics: New burden estimates and predictors from the GLaMOR Project. J Glob Health. 2019;9(2):020421.
26. Pratha Sah, Jorge A. Alfaro-Murillo et al Future epidemiological and economic impacts of universal influenza vaccines. Proc Natl Acad Sci USA. 2019:116 (41):20786-20792.
27. World Health Organization. (2021). Antigenic and genetic characteristics of zoonotic influenza A viruses and development of candidate vaccine viruses for pandemic preparedness. https://www.who.int/influenza/vaccines/virus/202103_zoonotic_vaccinevir usupdate.pdf?ua=1
28. Zebedee S. & Lamb R. Growth restriction of influenza A virus by M2 protein antibody is genetically linked to the Ml protein. Proc Natl Acad Sci USA. 1989;86(3): 1061-1065.
29. Zharikova D, Mozdzanowska K, Feng J et al Influenza type A virus escape mutants emerge in vivo in the presence of antibodies to the ectodomain of matrix protein 2. J Virol. 2005;79(11):6644-6654.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Манухова, Татьяна Ивановна
<120> Антиген для универсальной рекомбинантной вакцины
против вируса гриппа птиц и человека
<130> 030441
<140> 2021114322
<141> 2021-05-20
<160> 2
<170> PatentIn version 3.5
<210> 1
<211> 224
<212> PRT
<213> Artificial Sequence
<220>
<223> Рекомбинантная белковая молекула (Seq ID 01), включающая
трехкратный повтор двух последовательностей аминокислот,
консенсусных для М2е-эпитопа вирусов гриппа А птиц и человека.
<400> 1
Met Arg Gly Ser His His His His His His Gly Ser Ala Gly Ser Thr
1 5 10 15
Gly Gly Gly Ser Leu Leu Thr Glu Val Glu Thr Pro Ile Arg Asn Glu
20 25 30
Trp Gly Cys Arg Cys Asn Asp Ser Ser Asp Gly Ser Ala Gly Ser Ala
35 40 45
Ala Gly Ser Gly Glu Gly Gly Ser Leu Leu Thr Glu Val Glu Thr Pro
50 55 60
Thr Arg Asn Glu Trp Glu Cys Arg Cys Ser Asp Ser Ser Asp Gly Ser
65 70 75 80
Ala Gly Ser Thr Gly Gly Gly Ser Leu Leu Thr Glu Val Glu Thr Pro
85 90 95
Ile Arg Asn Glu Trp Gly Cys Arg Cys Asn Asp Ser Ser Asp Gly Ser
100 105 110
Ala Gly Ser Ala Ala Gly Ser Gly Glu Gly Gly Ser Leu Leu Thr Glu
115 120 125
Val Glu Thr Pro Thr Arg Asn Glu Trp Glu Cys Arg Cys Ser Asp Ser
130 135 140
Ser Asp Gly Ser Ala Gly Ser Thr Gly Gly Gly Ser Leu Leu Thr Glu
145 150 155 160
Val Glu Thr Pro Ile Arg Asn Glu Trp Gly Cys Arg Cys Asn Asp Ser
165 170 175
Ser Asp Gly Ser Ala Gly Ser Ala Ala Gly Ser Gly Glu Gly Gly Ser
180 185 190
Leu Leu Thr Glu Val Glu Thr Pro Thr Arg Asn Glu Trp Glu Cys Arg
195 200 205
Cys Ser Asp Ser Ser Asp Gly Ser Ala Gly Ser Ser Gly Gly Gln Ala
210 215 220
<210> 2
<211> 675
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность (Seq ID 02), кодирующая
рекомбинантную белковую молекулу, включающую трехкратный повтор
двух последовательностей аминокислот, консенсусных для
М2е-эпитопа вирусов гриппа А птиц и человека.
<400> 2
atgagaggat cgcatcacca tcaccatcac ggatccgccg ggtcgactgg cgggggttcg 60
ctgctgaccg aagtcgagac tccgattcgc aatgaatggg gatgtcgctg caacgactct 120
tcggatgggt ccgctggatc tgcggcgggc tccggagagg gcggttcgct tctgacagag 180
gtcgagaccc ctactcgcaa tgaatgggag tgtcgctgta gcgattcgtc agatgggagc 240
gcaggctcga ctggcggggg ttcgctgctg accgaagtcg agactccgat tcgcaatgaa 300
tggggatgtc gctgcaacga ctcttcggat gggtccgctg gatctgcggc gggctccgga 360
gagggcggtt cgcttctgac agaggtcgag acccctactc gcaatgaatg ggagtgtcgc 420
tgtagcgatt cgtcagatgg gagcgcaggc tcgactggcg ggggttcgct gctgaccgaa 480
gtcgagactc cgattcgcaa tgaatgggga tgtcgctgca acgactcttc ggatgggtcc 540
gctggatctg cggcgggctc cggagagggc ggttcgcttc tgacagaggt cgagacccct 600
actcgcaatg aatgggagtg tcgctgtagc gattcgtcag atgggagcgc aggctcgagc 660
ggcggacaag cttaa 675
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| Рекомбинантный штамм вируса гриппа A/PR8-NS124-TB10.4-2A-HspX и способ специфической профилактики туберкулеза легких с использованием вакцины мукозального применения на его основе | 2019 |
|
RU2726106C1 |
| Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов H1, H2, H5 для создания противогриппозных иммуногенных препаратов, способ получения гена | 2018 |
|
RU2713722C1 |
| ХИМЕРНЫЕ ВАКЦИНЫ ПРОТИВ ГРИППА | 2021 |
|
RU2834969C1 |
| Рекомбинантный вакцинный штамм для живой интраназальной вакцины, обеспечивающей сочетанную профилактику гриппозной и коронавирусной инфекций | 2022 |
|
RU2782531C1 |
| Однодоменное антитело для нейтрализации вирусов и его модификации, и способ их применения для экстренной профилактики заболеваний, вызываемых вирусом гриппа А | 2021 |
|
RU2777073C1 |
| КОМПОЗИЦИИ ГЕМАГГЛЮТИНИНА ГРИППА С ГЕТЕРОЛОГИЧНЫМИ ЭПИТОПАМИ И/ИЛИ ИЗМЕНЕННЫМИ САЙТАМИ РАСЩЕПЛЕНИЯ ПРИ СОЗРЕВАНИИ | 2017 |
|
RU2769406C2 |
| Рекомбинантный штамм вируса гриппа A/PR8-NS124-Luc и способ оценки поствакцинальных нейтрализующих антител с использованием биолюминесцентной детекции | 2019 |
|
RU2759054C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ (VLP) С ИСПОЛЬЗОВАНИЕМ ПРОТЕИНА ГРУППОВОГО АНТИГЕНА (GAG) ВИРУСА БЫЧЬЕГО ИММУНОДЕФИЦИТА | 2016 |
|
RU2734118C2 |
| КРОСС-РЕАКТИВНАЯ РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА А ЧЕЛОВЕКА | 2020 |
|
RU2742336C1 |


Изобретение относится к области биотехнологии, конкретно к рекомбинантному получению пептидных антигенов, и может быть использовано в медицине для создания универсальной профилактической вакцины против вируса гриппа А птиц и человека. Предложен рекомбинантный антиген ТМ2е, представляющий собой рекомбинантный белок с молекулярной массой 24 кДа и содержащий в своем составе консенсусные для вирусов гриппа А птиц и человека последовательности М2е-эпитопа, соответствующие фрагменту со 2 по 24 аминокислотный остаток указанного эпитопа. Изобретение обеспечивает получение антигенного компонента для создания универсальной бивалентной вакцины против вируса гриппа А птиц и человека. 2 з.п. ф-лы, 4 ил., 2 пр.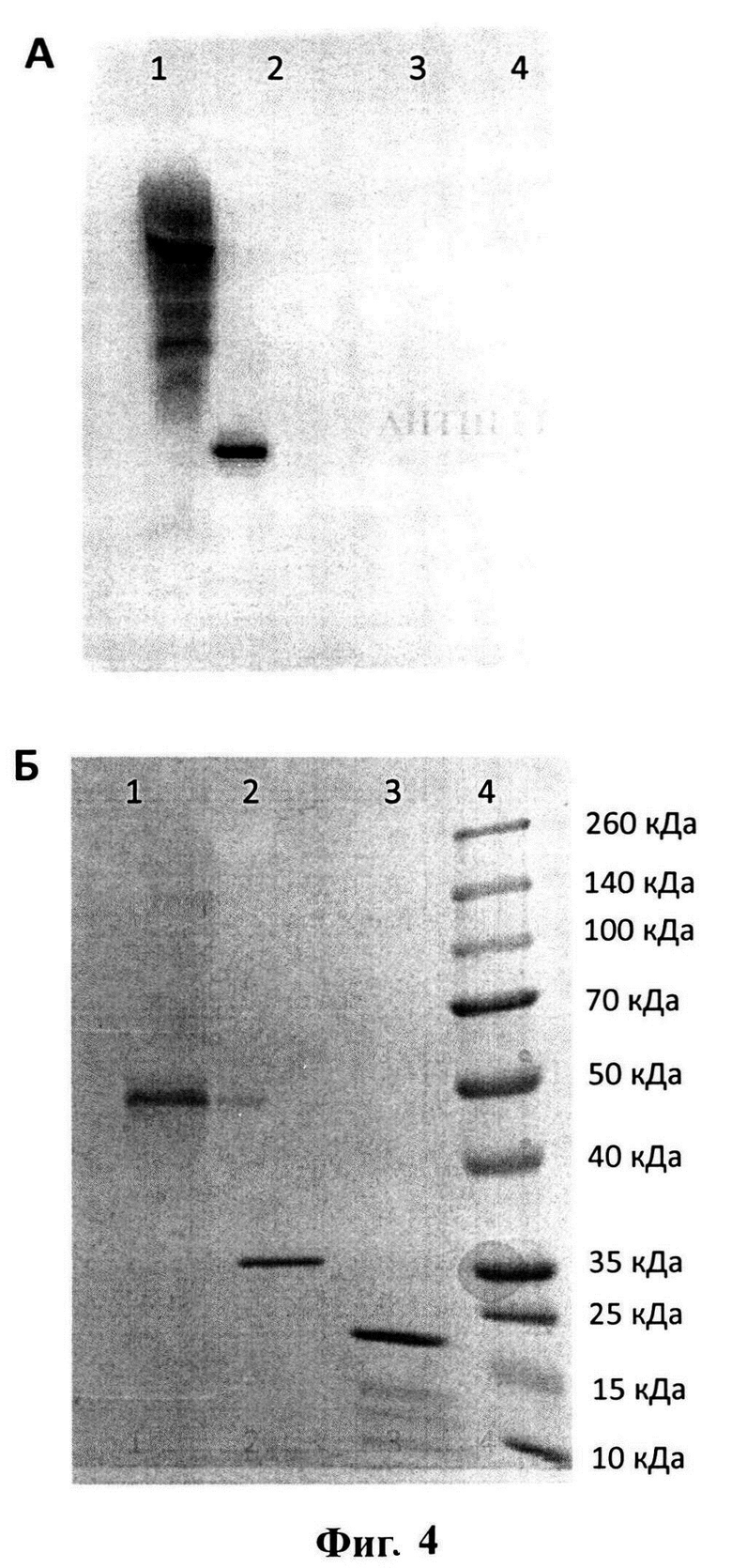
1. Рекомбинантный антиген ТМ2е для универсальной профилактической вакцины против вируса гриппа А птиц и человека, представляющий собой белок с последовательностью аминокислот SEQ ID NO: 1, которая включает консенсусные для вирусов гриппа А птиц и человека последовательности, соответствующие фрагменту с 2 по 24 аминокислотные остатки М2е-эпитопа вируса гриппа.
2. Рекомбинантный антиген по п. 1, кодируемый нуклеотидной последовательностью SEQ ID NO: 2.
3. Рекомбинантный антиген по п. 1, экспрессируемый в клетках Escherichia coli с помощью генно-инженерной конструкции рТМ, содержащей нуклеотидную последовательность SEQ ID NO: 2, связанную с регуляторными элементами, обеспечивающими ее экспрессию.
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| WO 2008054540 A2, 08.05.2008 | |||
| KIM MIN-CHUL et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| RU 2017144741 A, 19.06.2019 | |||
| СИНГЕР М | |||
| и др | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

