Область изобретения
Настоящее изобретение относится к области иммунологии и белковой инженерии и обеспечивает противогриппозную вакцину широкого спектра защиты («универсальную»), эффективную для борьбы с пандемией и эпидемией гриппа.
Уровень техники
Вакцинация населения является наиболее эффективным и доступным способом снижения ущерба, наносимого эпидемиями гриппа. Современные инактивированные противогриппозные вакцины обеспечивают профилактический эффект, в основном, за счет индукции иммунного ответа организма на поверхностные белки вируса гриппа: гемагглютинин (HA) и нейраминидазу (NA). Постоянный мутационный дрейф вирусов гриппа требует практически ежегодного обновления штаммового состава вакцин. Несмотря на тщательный глобальный мониторинг антигенной изменчивости вирусов гриппа, во многие эпидемические сезоны один из вирусных компонентов вакцин не совпадал с циркулирующим штаммом, что снижало эпидемиологическую эффективность традиционных вакцин. Кроме того, традиционные противогриппозные вакцины, производящиеся на куриных эмбрионах, имеют ряд противопоказаний, в первую очередь, наличие аллергических реакций.
В России к настоящему времени производится и используется в практике только штаммоспецифические противогриппозные вакцины (живые и инактивированные).
Живая вакцина Ультравак - вакцина гриппозная аллантоисная живая сухая интраназальная. Ультравак содержит аттенуированные эпидемически актуальные штаммы вируса гриппа типов А (H1N1, H3N2) и В, полученные из вируссодержащей аллантоисной жидкости куриных эмбрионов.
Инактивированные отечественные вакцины: Гриппол, Гриппол плюс, Совигрип, Флю-М, Флю-М квадро, Ультрикс. Полимер-субъединичные вакцины Гриппол и Совигрип представляют собой раствор поверхностных вирусных белков гемагглютинина и нейраминидазы в комплексе с водорастворимым высокомолекулярным иммуностимулятором, соответственно, Полиоксидонием и Совидоном. Гриппол Плюс - усовершенствованный аналог вакцины Гриппол без добавления консерванта тиомерсала, что снижает частоту поствакцинальных реакций; Гриппол Нео - культуральная полимер-субъединичная противогриппозная вакцина (в последние годы не выпускается). Флю-М, Флю-М квадро, Ультрикс - это расщепленные вакцины, содержащие, помимо поверхностных, внутренние вирусные белки.
Одним из основных направлений современной вакцинологии является создание рекомбинантных вакцин, нацеленных на широкий круг вирусов гриппа и основанных на использовании иммунодоминантных пептидов, инвариантных у различных штаммов вируса гриппа.
Примером консервативных иммунодоминантных пептидов для создания рекомбинантных «универсальных» вакцин является трансмембранный белок М2, последовательность внеклеточного домена (М2е) которого практически идентична для всех вирусов гриппа типа А, циркулировавших в человеческой популяции до появления вируса A(H1N1)pdm09, включая пандемические вирусы А/Сингапур/1/57 и А/Гонконг/1/68, вызвавшие пандемии, соответственно, в 1957 и 1968 годах. При этом, вирус гриппа А/Сингапур/1/57 (H2N2) рассматривается как один из самых вероятных возбудителей будущей пандемии, так как с момента исчезновения его из циркуляции прошло около 50 лет и появилось 2 поколения людей, не имеющих к нему анамнестического иммунитета. Небольшой по размеру (23 аминокислоты) высоко консервативный внеклеточный домен (эктодомен) белка М2 является объектом пристального внимания разработчиков вакцин. Одна из существенных характеристик пептида М2е - его обильная экспрессия на инфицированных клетках и доступность для эффекторов иммунной системы макроорганизма [Lamb, 1985, Holsinger, 1991; DeFillete, 2005]. Как показано в 1990-2000 годы, пассивный перенос моноклональных анти-М2е антител приводит к ограничению вирусной репликации и защите экспериментальных животных от заражения [Zebedee, 1988; Treanor, 1990; Fan, 2004; Zharikova, 2005; Liu, 2005]. Вместе с тем, после гриппозной инфекции и вакцинации антитела к низкоиммуногенному М2е образуются в малом количестве, но существует много способов увеличения иммуногенности этого пептида. В качестве вакцинных препаратов предлагаются разнообразные конструкции, включающие пептид М2е [Tompkins, 2007; Park, 2011; Bessa, 2008; Denis, 2008; Neirynck, 1999; DeFilette, 2005, 2006; Hulleatt, 2008, Mozdanovska, 2003; Kim, 2013]. Две универсальные вакцины Multimeric-001 (производство BiondVax) и Flu-v (производство компании SEEK) зарегистрированы, несколько находятся на стадии клинических исследований. [Scorza B.F., 2016; https://cordis.europa.eu/result/rcn/240188_es.html].Актуальность создания эффективных универсальных вакцин против гриппа обсуждается в обзорной статье EGOROV Andrej Y. «The challenges of creating a universal influenza vaccine». Microbiology Independent Research Journal, Volume 3, Number 1, 2016 p. 31-41.
Универсальные вакцины против гриппа A, способные обеспечить широкую перекрестную защиту и содержащие слитый белок, включающий гетерологичные домены M2e (внеклеточный домен М2 белка вируса гриппа) для усиления антигенности и перекрестной защиты также раскрыты в международных заявках WO 2016109656, WO 2014070848 и WO 2019070955, согласно которым слитый белок может быть включен в вирусоподобные частицы (VLP).
В статье SUN X. et al. (2015). «High copy numbers and N terminal insertion position of influenza A M2E fused with hepatitis B core antigen enhanced immunogenicity» BioScience Trends, 9(4), 221-227, отмечается, что M2e является перспективным кандидатом для разработки рекомбинантной вакцины против гриппа А широкого спектра действия. Однако иммуногенность M2e должна быть улучшена. Количество копий M2e и его экспрессия слияния с различными белками-носителями может влиять на его иммунопотентность.
В работе Ravin NV, Blokhina EA, Kuprianov VV, Stepanova LA, Shaldjan AA, Kovaleva AA, Tsybalova LM, Skryabin KG. (2015) Development of a candidate influenza vaccine based on virus-like particles displaying influenza M2e peptide into the immunodominant loop region of hepatitis B core antigen: Insertion of multiple copies of M2e increases immunogenicity and protective efficiency. Vaccine. 33(29):3392-3397, описаны сконструированные варианты рекомбинантных белков с одной, двумя и четырьмя копиями M2e, встроенными в иммунодоминантную петлю HBc антигена, а также отмечены преимуществами HBc, как носителя для слабоиммуногенного антигена.
Также проведена оценка влияния числа копий M2e в рекомбинантных белках на их иммуногенность. Показана тенденция усиления иммунного ответа при увеличении числа копий M2e в иммунодоминантной петле HBc. На основании полученных данных был выбран вариант рекомбинантного белка с четырьмя копиями M2e в иммунодоминантной петле HBc.
Одной из самых актуальных на сегодняшний день проблем здравоохранения является необходимость разработки отечественной универсальной вакцины против гриппа, обладающей повышенной стабильностью и иммуногенностью, по сравнению с известными на сегодняшний день разработками, указанными в описанном уровне техники, недостаточность стабильности и иммуногенности которых так и не позволила им использоваться в целях борьбы с гриппом.
При этом также необходимо отметить, что помимо аспекта применения разработанной вакцины для всего населения в качестве «баррикадной» при возникновении новой пандемии, имеются еще категории населения, для которых актуальность предлагаемой вакцины бесспорна. Во-первых, лица, нуждающиеся в щадящих низкоаллергенных вакцинных препаратах: беременные женщины, дети младшего возраста и лица с соматическими и хроническими инфекционными заболеваниями. Для этих категорий рекомбинантные гриппозные вакцины будут вакцинами выбора. Во-вторых, дети, праймирование которых рекомбинантной «универсальной» вакциной предотвратит развитие вакцинозависимости, возникающей при ежегодной иммунизации субъединичными вакцинами.
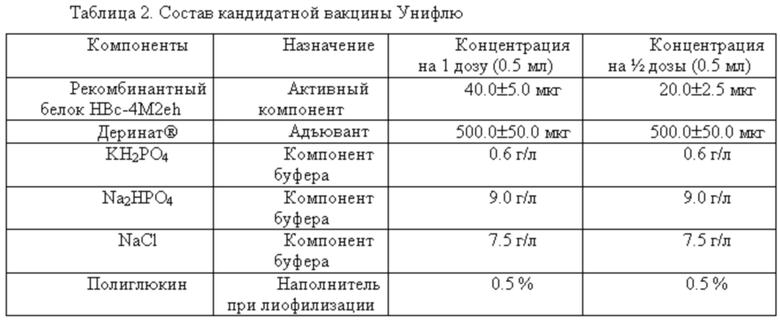
Таким образом, авторами настоящего изобретения была разработана новая универсальная вакцина против гриппа, содержащая в качестве активных ингредиентов рекомбинатный белок HBc-4M2eh, который имеет аминокислотную последовательность SEQ ID NO 6, в количестве от 20 до 90 мкг и адъювант дезоксирибонуклеат натрия в количестве от 1000 до 10000 мкг, которая обладает улучшенной стабильностью по сравнению с известными на сегодняшний день универсальными вакцинами против гриппа на основе рекомбинантных белков.
Сущность изобретения
Одним объектом настоящего изобретения является кросс-реактивная рекомбинантная вакцина против вируса гриппа А человека, содержащая рекомбинантный белок HBc-4M2eh, который имеет аминокислотную последовательность SEQ ID NO 6, в количестве от 20 до 90 мкг и адъювант дезоксирибонуклеат натрия в количестве от 1000 до 10000 мкг, а также фосфатно-буферный раствор (ФБР), рН 7,2-7,4, обладающая неожиданно увеличенной стабильностью по сравнению с вакцинами уровня техники. Данная вакцина также обладает улучшенной иммуногенностью.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 20 мкг рекомбинантного белка HBc-4M2eh.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 40 мкг рекомбинантного белка HBc-4M2eh.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 60 мкг рекомбинантного белка HBc-4M2eh.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 90 мкг рекомбинантного белка HBc-4M2eh.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит от 1000 до 3000 мкг адъюванта дезоксирибонуклеата натрия.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит от 1000 до 5000 мкг адъюванта дезоксирибонуклеата натрия.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 1000 мкг адъюванта дезоксирибонуклеата натрия.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 3000 мкг адъюванта дезоксирибонуклеата натрия.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 5000 мкг адъюванта дезоксирибонуклеата натрия.
В одном варианте выполнения настоящего изобретения вакцина согласно настоящему изобретению содержит 10000 мкг адъюванта дезоксирибонуклеата натрия.
Другим объектом настоящего изобретения является применение вакцины согласно настоящему изобретению для профилактики гриппа А человека.
Краткое описание чертежей
Фиг. 1 - Структура рекомбинантных белков HBc/M2ek(C-S), HBc/M2ek, HBc/2M2ek, HBc/2M2eh, HBc/4M2eh и HBc/2M2ek2M2eh.
Фиг. 2 - Доклинические исследования иммуногенности вакцины Унифлю. M2e - специфический гуморальный ответ у мышей (а, б, в) и хорьков (г). а - СГТ анти-M2e и анти-НВс IgG после 3-й иммунизации мышей одной дозой вакцины и плацебо; б - соотношение IgG1 и IgG2a в сыворотках крови иммунизированных мышей; в - СГТ анти-M2e IgG после 3-й иммунизации мышей в БАЛ; г - СГТ анти-M2e IgG.
Фиг. 3 - Доклинические исследования протективности вакцины Унифлю на модели летальной гриппозной инфекции (5 LD50) у мышей. а - выживаемость мышей после заражения вирусом А/PR/8/34 (H1N1); б - выживаемость мышей после заражения вирусом A/Aichi/2/68 (H3N2); в - выживаемость после заражения вирусом. A/Japan305/57 (H2N2); г - выживаемость после заражения вирусом A/California/7/2009 (H1N1)pdm09; * - достоверные результаты между вакциной и плацебо; ** - достоверные результаты между вакциной и ФБР.
Фиг. 4 - Экспрессия рекомбинантных белков HBc/4M2eh и HBc/2M2ek2M2eh в клетках E. coli. (A) SDS-PAGE анализ белковых препаратов. Дорожка 1, маркеры молекулярного веса (размер указан в килодальтонах; дорожка 2, препарат белков из штамма E. coli, не содержащего экспрессионный вектор; дорожка 3, препарат белков из штамма - продуцента HBc/4M2eh после индукции; дорожка 4, препарат белков из штамма - продуцента HBc/2M2ek2M2eh после индукции. (В) Вестерн-блот анализ белковых препаратов с использованием антител к M2ek. Дорожка 1, препарат белков из штамма - продуцента HBc/4M2eh после индукции; дорожка 2, препарат белков из штамма - продуцента HBc/2M2ek2M2eh после индукции.
Фиг. 5 - Аминокислотные последовательности SEQ ID NO 1-6.
Подробное описание изобретения
Одним из наиболее эффективных носителей антигенных эпитопов является нуклеокапсидный белок вируса гепатита В человека (НВс антиген). Мономеры этого белка, состоящие из 183-185 аминокислот, в инфицированных клетках собираются в стабильные агрегаты (вирусоподобные частицы), называемые HBc - частицы. Образуются два типа частиц, - частицы диаметром около 30 нм, состоящие из 180 мономеров, и в большем количестве частицы диаметром 34 нм, состоящие из 240 субъединиц. Аналогичные по размеру и структуре частицы могут образовываться в результате самосборки мономеров НВс антигена, экспрессированных в клетках бактерий, дрожжей, растений и животных.
НВс белок обладает рядом преимуществ как носитель антигенов небольших размеров:
• НВс частицы высоко иммуногенны, стимулируют как Т-независимый так и Т-зависимый иммунный ответ [Milich, 1986].
• три района HBc антигена могут быть использованы для вставки чужеродных пептидов для их презентации на поверхности HBc - частиц: N и С концы HBc антигена и, так называемая, иммунодоминантная петля, расположенная между 75 и 85 аминокислотными остатками белка, на вершине «шпильки» - поверхностного выпячивания на НВс частице [Pumpens, 2001].
• вставки на N- и С-конец вызывают иммунный ответ против встроенного эпитопа, однако, более сильный иммунный ответ на встроенную последовательность возникает при их локализации в области иммунодоминантной петли, поскольку основной иммунный ответ против НВс частиц направлен именно на выступающие на поверхности частицы «шпильки», образованные иммунодоминантными петлями [Bachmann, 1993; Fiers, 2009].
• рекомбинантный белок может накапливаться в клетках бактерий Escherichia coli и спонтанно собираться в вирусоподобные частицы (ВПЧ), а химерные HBc частицы, экспонирующие на своей поверхности чужеродные эпитопы, способны индуцировать высокий иммунный ответ на встроенные пептиды [DeFillete, 2008].
• С-концевая аргинин-богатая область HBc антигена (150-183 аминокислотные остатки) может быть удалена без нарушения самосборки частиц, более того, в отличие от образованных полноразмерным белком, такие частицы при сборке не включают нуклеиновые кислоты клетки-хозяина.
Химерные HBc частицы часто имеют менее упорядоченную структуру, чем частицы, не содержащие эпитопы [Schodel, 1994a]. Во многих случаях вставка эпитопа вызывает настолько сильную дестабилизацию, что гибридные частицы либо вообще не могут быть получены либо оказываются настолько нестабильными, что не могут быть использованы в качестве компонента вакцин [Schodel, 1994b; Karpenko, 2000]. Вставки в иммунодоминантную петлю особенно чувствительны к структуре чужеродного пептида, поскольку концевые участки встраиваемой последовательности должны соответствовать конформации акцепторного сайта. Поэтому конструирование химерных НВс частиц является нетривиальной задачей, решение которой требует применения современных методов белковой инженерии.
Авторы настоящего изобретения сначала получили рекомбинантную белковую молекулу HBc/M2ek(C-S), которая включала последовательно следующие элементы: N-концевой метионин, аминокислотную последовательность HBc антигена [Bichko, 1985], начиная с аспарагина в положении +4 до аспарагина в положении +78 (относительно первого метионина в нативном HBc), последовательность аминокислот SLLTEVETPTRNEWESRSSDSSD (далее обозначаемую M2ek), соответствующую М2е пептиду вируса гриппа птиц штамма A/Chicken/Kurgan/05/2005 (последовательность SLLTEVETPTRNEWECRCSDSSD в которой два цистеина в положениях +16 и +18 заменены на серины), аминокислотную последовательность HBc антигена, начиная с серина в положении +81 до валина в положении +149 (относительно первого метионина в нативном HBc), и цистеин на С-конце (Фиг. 1). Введение С-концевого цистеина имело целью повысить стабильность рекомбинантных М2Е-НВс частиц, но не обязательно для получения высокоактивных вирусоподобных частиц. Замена двух цистеинов в последовательности М2е пептида на серины была обусловлена тем, что, что цистеины за счет формирования дисульфидных связей могут приводить к агрегации белка; а их замена на серины предотвращает образование дисульфидных связей но не влияет на иммуногенные свойства М2е [De Filette, 2005]. Рекомбинантная белковая молекула HBc/M2ek(C-S) имеет аминокислотную последовательность SEQ ID NO 1.
Для получения описанной выше рекомбинантной белковой молекулы HBc/M2ek(C-S) была создана кодирующая ее рекомбинантная нуклеиновая кислота (ген). В качестве основы для ее создания был использован ранее созданный нами рекомбинантный вектор экспрессии pQE60-HBc [Blokhina, 2013]. В этом векторе рекомбинантная нуклеиновая кислота, кодирующая укороченный НВс антиген (N-концевой метионин, аминокислотную последовательность HBc антигена начиная с аспарагина в положении +4 до валина в положении +149 и цистеин на С-конце), оперативно связана с промотором PT5-lac и терминатором транскрипции Т0, что обеспечивает ее экспрессию в клетке бактерии Escherichia coli [Blokhina, 2013]. Нуклеотидную последовательность рекомбинантного вектора экспрессии pQE60-HBc модифицировали путем введения дополнительных рестрикционных сайтов BamHI (GGATCC) и ApaI (GGGCCC) между триплетами, кодирующими аспарагин в положении +78 и серин в положении +81 (с одновременным удалением триплетов, кодирующих аминокислоты в положении +79 и +80 относительно первого метионина в нативном HBc). Затем по сайтам BamHI и ApaI вводили полученную с помощью ПЦР нуклеиновую кислоту, кодирующую аминокислотную последовательность M2ek (SLLTEVETPTRNEWESRSSDSSD). Полученный вектор обозначали pQE60-HBc/M2ek(C-S) и использовали в качестве базового для дальнейших модификаций.
Рекомбинантная белковая молекула HBc/M2ek включала последовательно следующие элементы: N-концевой метионин, аминокислотную последовательность HBc антигена начиная с аспарагина в положении +4 до аспарагина в положении +78, последовательность глицин-серинового линкера GTSGSSGSGSGGSGSGGGG, последовательность M2ek пептида, последовательность глицин-серинового линкера GTSGSSGSGSGGSGSGGGG, аминокислотную последовательность HBc антигена, начиная с серина в положении +81 до валина в положении +149, и цистеин на С-конце. Введение гибких глицин-сериновых линкеров длиной по 19 аминокислот в места соединения последовательностей M2ek и НВс имело целью облегчить фолдинг гибридного белка. Для получения рекомбинантной белковой молекулы HBc/M2ek была создана кодирующая ее рекомбинантная нуклеиновая кислота (ген). В качестве основы для ее создания был использован рекомбинантный вектор экспрессии pQE60-HBc/M2ek(C-S). Чтобы фланкировать M2ek вставку два синтетических олигонуклеотидных фрагмента, кодирующих последовательность (GTSGSSGSGSGGSGSGGGG, были введены в вектор pQE60-HBc/M2ek(C-S) по сайтам BamHI и ApaI. Полученный рекомбинантный вектор экспрессии pQE60-HBc/M2ek позволяет получать белковую молекулу HBc/M2ek.
Введение в pQE60-HBc/M2ek дополнительных фрагментов ДНК, кодирующих M2ek, между участками, кодирующими глицин-сериновые линкеры, позволило получить рекомбинантные векторы экспрессии pQE60-HBc/2M2ek и pQE60-HBc/4M2ek, обеспечивающие экспрессию рекомбинантных белковых молекул HBc/2M2ek и HBc/4M2ek, включающих, соответственно, 2 и 4 копии M2ek в районе иммунодоминантной петли НВс антигена (Фиг. 1). Рекомбинантные белковые молекулы HBc/M2ek, HBc/2M2ek и HBc/4M2ek имеют аминокислотные последовательности SEQ ID NO 2, SEQ ID NO 3 и SEQ ID NO 4, соответственно.
Для получения штаммов-продуцентов рекомбинантных белков HBc/M2ek(C-S), HBc/M2ek, HBc/2M2ek и HBc/4M2ek рекомбинантные векторы экспрессии pQE60-HBc/M2ek(C-S), pQE60-HBc/M2ek, pQE60-HBc/2M2ek и pQE60-HBc/4M2ek, соответственно, вводили в клетки E. coli штамма DLT1270 с помощью трансформации. Штамм DLT1270 являющийся производным штамма DH10B [Grant, 1990], содержит ген репрессора лактозного оперона lacI, интегрированный в хромосому. Культуры трансформированных клеток DLT1270/ pQE60-HBc/M2ek(C-S), DLT1270/pQE60-HBc/M2ek, DLT1270/ pQE60-HBc/2M2ek и DLT1270/pQE60-HBc/4M2ek выращивали в стандартных условиях, подходящих для экспрессии рекомбинантного белка, например, в LB-бульоне до середины логарифмической фазы роста (OD600=0,5) при 37°C, добавляли IPTG до концентрации 0.5 мМ (достаточной для полной индукции промотора) и выращивали клетки еще в течение 48 часов при 20-30°C. Пробы для определения белка отбирали в начальный момент времени, а также через 16 часов после индукции. Определение уровня экспрессии рекомбинантных белков проводили путем анализа суммарных белковых препаратов, выделенных из бактериальных культур с помощью SDS-PAGE.
Рекомбинантный белок HBc/M2ek(C-S) успешно экспрессировали в клетках E. coli, однако он оказался нерастворимым, в связи с чем дальнейшая работа с ним не проводилась. Вероятно, вставка M2ek в район иммунодоминантной петли НВс антигена отрицательно повлияла на фолдинг гибридного белка, что привело к его нерастворимости.
Данную проблему удалось преодолеть за счет введения гибких глицин-сериновых линкеров в места соединения последовательностей M2ek и НВс антигена. Анализ белковых препаратов, выделенных из штаммов DLT1270/pQE60-HBc/M2ek, DLT1270/pQE60-HBc/2M2ek и DLT1270/pQE60-HBc/4M2ek после индукции синтеза целевых продуктов показал, что рекомбинантные белки HBc/M2ek, HBc/2M2ek и HBc/4M2ek успешно экспрессируются и являются растворимыми. Рекомбинантные белки также выявлялись при иммуноблотинге с анти-M2ek моноклональными антителами.
Для выделения вирусоподобных частиц клетки штаммов-продуцентов после индукции собирали с помощью центрифугирования и ресуспендировали в буфере TEN (10 мМ Tris-HCl pH 8.0; 100 мМ NaCl; 1 мМ EDTA). Клеточную суспензию обрабатывали лизоцимом 1 мг/мл в течение 30 минут при +4С, после чего взвесь замораживали при -20С в течение ночи. Затем суспензию размораживали и осаждали центрифугированием при 30,000 × g в течение 1 часа. Отбирали супернатант, белки осаждали с помощью 40% сульфата аммония. Дальнейшую очистку проводили с использованием колонок Sephacryl S-100HR для эксклюзионной хроматографии по размеру, после чего проводили очистку на колонках Q-Sepharose FF (GE Healthcare) для анионо-обменной хроматографии. Фракции, содержащие НВс-частицы были собраны, объединены и хранились при 4C. Чистота полученного препарата составляла не менее 95%. Указанные условия выделения и очистки подбирались экспериментальным путем и могут варьироваться в известных среднему специалисту в этой области значениях.
Факт образования рекомбинантными белками HBc/M2ek, HBc/2M2ek и HBc/4M2ek вирусоподобных частиц был показан с помощью электронной микроскопии выделенных препаратов, в качестве контроля был использован препарат HBc - частиц, не содержащих M2e пептида, а в остальном эквивалентный настоящим частицам. Электронно-микроскопический анализ подтвердил факт самосборки рекомбинантных белковых молекул HBc/M2ek, HBc/2M2ek и HBc/4M2ek в вирусоподобные частицы размером 30-35 нанометров, сходных с частицами, образуемыми HBc антигеном, не содержащим M2e.
Так как предназначенная для человека противогриппозная вакцина должна быть, в первую очередь, направлена против “человеческих” штаммов вируса гриппа и, следовательно, должна содержать М2е последовательность “человеческих” штаммов, авторами настоящего изобретения было сконструировано два типа рекомбинантных вирусоподобных частиц - носителей М2е пептида. В рекомбинантной белковой молекуле HBc/4M2eh в состав иммунодоминантной петли НВс антигена включено 4 копии последовательности SLLTEVETPIRNEWGSRSNDSSD (далее - M2eh), соответствующей консенсусной последовательности М2е пептида вируса гриппа человека SLLTEVETPIRNEWGCRCNDSSD, в которой два цистеина в положениях +16 и +18 заменены на серины. Для создания кодирующей эту рекомбинантную белковую молекулу рекомбинантной нуклеиновой кислоты заменили кодирующий M2ek фрагмент в векторе pQE60-HBc/M2ek на синтетическую нуклеотидную последовательность, кодирующую M2eh. Путем введения дополнительных копий M2eh получили вектор pQE60-HBc/4M2eh, кодирующий белок HBc/4M2eh, несущий 4 копии M2eh (Фиг. 1).
Поскольку последовательности М2е пептида вирусов гриппа человека и птиц различаются, для создания «универсальной» вакцины, которая была бы эффективна как против «человеческого», так и «птичьего» гриппа, сконструировали вторую рекомбинантную белковую молекулу, HBc/2M2ek2M2eh (Фиг. 1), содержащую 2 копии «человеческого» M2eh и 2 копии «птичьего» M2ek пептида (в очередности M2ek-M2eh-M2ek-M2eh). Для создания кодирующей эту рекомбинантную белковую молекулу рекомбинантной нуклеиновой кислоты в вектор pQE60-HBc/M2ek между последовательностями, кодирующими M2ek и глицин-сериновый линкер, был введен синтетический олигонуклеотидный фрагмент, кодирующий пептид M2eh. Затем копия вставки M2ek-M2eh была клонирована в тот же вектор правее первоначальной вставки. В результате был получен рекомбинантный вектор экспрессии pQE60-HBc/2M2ek2M2eh. Рекомбинантные белковые молекулы HBc/2M2ek2M2eh и HBc/4M2eh имеют аминокислотные последовательности SEQ ID NO 5 и SEQ ID NO 6, соответственно.
Для получения штаммов-продуцентов рекомбинантных белков HBc/4M2eh и HBc/2M2ek2M2eh рекомбинантные векторы экспрессии pQE60-HBc/4M2eh и pQE60-HBc/2M2ek2M2eh, соответственно, вводили в клетки E. coli штамма DLT1270 с помощью трансформации. Культуры трансформированных клеток DLT1270/pQE60-HBc/4M2eh и DLT1270/pQE60-HBc/2M2ek2M2eh выращивали в стандартных условиях, подходящих для экспрессии рекомбинантного белка, например, в LB-бульоне до середины логарифмической фазы роста (OD600=0,5) при 37°C, добавляли IPTG до концентрации 0.5 mM (достаточной для полной индукции промотора) и выращивали клетки еще в течение 48 часов при 20-30°C.
Анализ суммарных белковых препаратов, выделенных из бактериальных культур с помощью SDS-PAGE показал, что оба рекомбинантных белка продуцируются в достаточном количестве (Фиг. 4). В отличие от НВс-частиц, содержащих M2ek пептиды, оба рекомбинантных белка оказались нерастворимы при стандартных условиях экспрессии после индукции (37°C). Однако, экспрессия при пониженных температурах (28°C для HBc/4M2eh и 20°C для HBc/2M2ek2M2eh) позволила повысить содержание продукта в растворимой фракции приблизительно до 50%. Вестерн-блот анализ с использованием антител на «птичий» M2ek пептид выявил только HBc/2M2ek2M2eh, а белок HBc/4M2eh не взаимодействовал с этими антителами (Фиг. 4). Вероятно, разница в 3-х аминокислотах в последовательностях M2eh и M2ek критична для распознавания. Выделение вирусоподобных частиц из клеток штаммов-продуцентов проводили так же, как для ВПЧ, содержащих M2ek.
Авторы настоящего изобретения в диссертации БЛОХИНА Е. А. «Конструирование вирусоподобных частиц на основе корового белка вируса гепатита В и M2 белка вируса гриппа как основы новых противогриппозных вакцин», ДИССЕРТАЦИЯ, Москва, 2013, которая включена в настоящую заявку в полном объеме посредством ссылки, описали разработанный ими рекомбинантный белок HBc/4M2eh, содержащий четыре копии М2е пептида вируса гриппа человека и рекомбинантные коровые частицы вируса гепатита В (HBc). Представлена оценка иммунологических характеристик и протективного действия вирусоподобных частиц, образованных химерными белками HBc с вставкой четырех копий М2е пептида в район иммунодоминантной петли, которые позволяют рассматривать рекомбинантный белок HBc/4M2eh в качестве превосходного кандидата для получения универсальных противогриппозных вакцин. Показано, что число встроенных М2е пептидов положительно коррелировало с иммуногенностью и протективным действием химерных НВс-частиц. Описано получение рекомбинантного белка HBc/4M2eh.
В данной диссертации также раскрыты результаты исследований иммуногенности и протективного действия рекомбинантных белков HBc/M2ek-M2eh-M2ek-M2eh и HBc/4M2eh, которые были проведены в ФГБУ «НИИгриппа» Минздрава России. Группы опытных мышей (линия Balb/c, по 5 мышей в каждой группе) трехкратно иммунизировали препаратами очищенных химерных вирусоподобных частиц в дозе 50 мкг на мышь с адъювантом Деринат® (действующее вещество (МНН): натрия дезоксирибонуклеат) (100мкг/мышь). Контрольной группе мышей трехкратно вводили препарат очищенных HBc частиц в такой же дозе. Иммуногенность тестируемых препаратов оценивали по способности специфичных сывороточных антител распознавать линейные и нативные эпитопы М2е пептида. Протективность рекомбинантных белков оценивали на модели летальной гриппозной инфекции при заражении мышей вирусами гриппа штаммов A/Chicken/Kurgan/05/05RG (H5N1), A/PR/8/34 (H1N1) и A/Aichi (H3N2).
Определение титра специфических антител к линейным эпитопам М2е в индивидуальных сыворотках мышей проводили методом ИФА с использованием синтетических пептидов G-11 (SLLTEVETPTRNEWECRCSDSSD) и G-37 (SLLTEVETPIRNEWGCRCNDSSD). Пептид G-11 соответствует M2e последовательности вируса гриппа птиц A/Chicken/Kurgan/05/2005 (H5N1), а G-37 соответствует консенсусной последовательности 2е вируса гриппа человека. Сыворотки отбирали через две недели после 3-ей иммунизации. В качестве дополнительного контроля использовали сыворотки не иммунизированных животных.
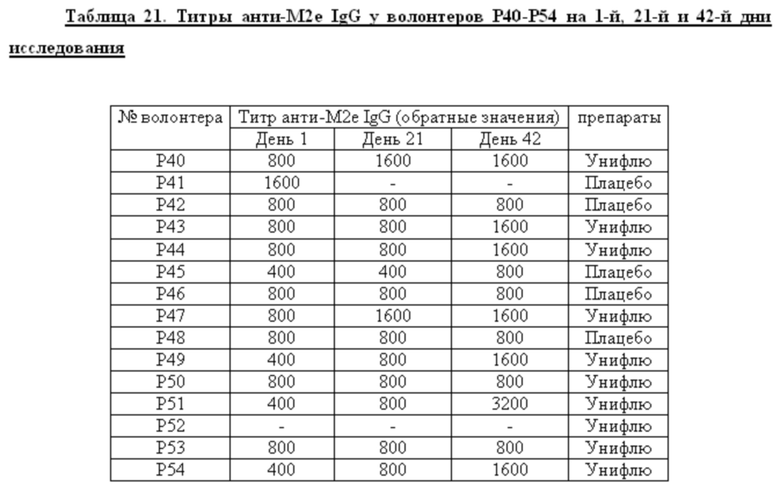
Исследования показали (Таблица 1), что иммунизация мышей препаратом HBc/M2ek-M2eh-M2ek M2eh инициирует образование иммуноглобулинов IgG, специфичных как к М2е последовательности вируса гриппа птиц A/Chicken/Kurgan/05/2005 (H5N1), так и к консенсусной последовательности М2е вируса гриппа человека. Обе эти М2е последовательности были включены в состав рекомбинантного белка исследуемого препарата. Образование меньшего титра антител к М2е последовательности вируса гриппа птиц (A/Chicken/Kurgan/05/2005) можно объяснить разной конформационной доступностью соответствующих копий М2е в составе химерных ВПЧ. В сыворотках мышей иммунизированных препаратом HBc/4M2eh, не содержащим в составе М2е последовательности штамма A/Chicken/Kurgan/05/2005, титр специфичных антител, связывающих пептид G-11 («птичий» М2е), был значительно ниже, чем титры в сыворотках мышей, иммунизированных препаратом HBc/M2ek-M2eh-M2ek-M2eh. Эти результаты говорят о важном влиянии аминокислотных замен на специфичность индуцируемых иммуноглобулинов.
Сравнение титров анти-HBc иммуноглобулинов в опытных и контрольной (мыши, иммунизированные HBc антигеном) группах показало, что включение в состав HBc антигена дополнительных последовательностей (четырехкратные повторы М2е) более чем в два раза снижает иммуногенность самого носителя. Кроме того, иммунизация как HBc/M2ek-M2eh-M2ek-M2eh, так и HBc/4M2eh препаратом индуцирует образование суммарного титра антител против М2е выше, чем титр антител против самого HBc антигена, что соотносится с данными, полученными De Filette et al., 2005.
Таблица 1. Титры IgG антител и средние геометрические титры (СГТ) в сыворотке к синтетическим пептидам G-11-1, G-37 и HBc белку после третьей иммунизации мышей препаратами вирусоподобных частиц HBc/M2ek-M2eh-M2ek-M2eh и HBc/4M2eh.
+ Дерината® 100 мкг, в/м
491 520
* p<0.05 между опытом и контролем
Для определения специфических сывороточных антител изотипа IgG к «нативному» М2е проводили ИФА с использованием клеток MDCK, инфицированных вирусами A/PR/8/34(H1N1) и A/Chicken/Kurgan/05/05RG. После заражения белок М2 экспонируется на поверхности инфицированных клеток MDCK. Связывание сывороток иммунизированных мышей с этими клетками характеризует взаимодействие антител с М2е в его «естественной» конформации. Через 2 недели после третьей иммунизации сыворотки мышей были тестированы на способность к связыванию in vitro с клетками MDCK, неинфицированными и инфицированными вирусами A/PR/8/34 (H1N1) или A/Kurgan/05/05RG (H5N1).
Определение уровня антител, способных связываться с нативным М2 белком в инфицированных клетках MDCK показало сходные результаты с данными по титрам специфичных IgG к синтетическим М2е пептидам (Таблица 1). Иммунизация опытных животных как HBc/M2ek-M2eh-M2ek-M2eh, так и HBc/4M2eh вызывает образование высокого титра антител к нативным М2е пептидам вируса гриппа человека.
При этом наибольший титр антител при инфицировании вирусом гриппа человека наблюдается для рекомбинантного белка HBc/4M2eh.
Протективное действие препаратов HBc/M2ek-M2eh-M2ek-M2eh и HBc/4M2eh оценивали по динамике гибели иммунизированных мышей после заражения адаптированными к мышам вирусами гриппа птиц A/Kurgan/05/05RG (H5N1), и человека A/PR/8/34 (H1N1) и A/Aichi (H3N2). Последовательность М2е штамма A/Aichi (H3N2) совпадает с консенсусной «человеческой» М2е последовательностью. Результаты этих исследований показывают, что трехкратная иммунизация мышей препаратом вирусоподобных частиц HBc/M2ek-M2eh-M2ek-M2eh, содержащих в составе рекомбинантного белка М2е пептиды вируса гриппа птиц и человека, обеспечивает 83% защиту при заражения вирусом гриппа человека A/PR/8/34, а также 25% защиту при заражении вирусом гриппа птиц A/Kurgan/05/05RG (H5N1). Эти результаты согласуются с данными определения уровня специфичных антител. Более низкие титры антител специфичных к М2е пептиду и нативному М2 белку вируса гриппа птиц A/Kurgan/05/2005 (H5N1) обеспечивают слабую защиту при заражении вирусом.
При заражении вирусом гриппа человека A/PR/8/34 мышей, иммунизированных препаратом вирусоподобных частиц HBc/4M2eh, наблюдали 92% защиту. Высокий уровень защиты (87%) наблюдали и при заражении вирусом A/Aichi (H3N2), последовательность М2е пептида которого совпадает с M2eh.
На основании проведенных разработок, исследований и полученных результатов авторами настоящего изобретения была разработана и описана рекомбинантная гриппозная вакцина Унифлю, которая является лиофилизированным препаратом, содержащим в 1 дозе (0,5 мл) высокоочищенный рекомбинантный белок НВс4М2е (40,0 ± 5,0 мкг) с консенсусной последовательностью М2е вирусов гриппа А человека (A/H1N1, A/H3N2, A/H2N2), адъювант - Деринат® (500 ± 50 мкг), фосфатно-буферный раствор (ФБР), рН 7,2-7,4. Особенностью рекомбинантной гриппозной вакцины Унифлю является ее широкое протективное действие против вирусов гриппа А человека различных субтипов (A/H1N1, A/H2N2, A/H3N2) как сезонных, так и вероятных пандемических.
Основой вакцины является рекомбинантный белок HBc-4M2eh, в котором целевой антиген (4 копии консенсусной последовательности эктодомена белка М2 - М2е - вирусов гриппа человека А субтипов H1N1, H2N2, H3N2) встроены в иммунодоминантную петлю корового антигена вируса гепатита В (HBc). Штамм E.coli DLT1270 pQE60 HBc/4M2eh, несущий плазмиду pQE60 HBc/4M2eh, является продуцентом гибридного белка.
Данная вакцина описана авторами настоящего изобретения в статье ЦЫБАЛОВА Л.М. И ДР. в «Кросс-протективные свойства противогриппозной вакцины на основе рекомбинантного белка HBc4M2e». Вопросы вирусологии. Апрель 2018; 63(2): 68-76, которая включена в настоящую заявку в полном объеме посредством ссылки. Доклинические исследования иммуногенности вакцины показаны на Фиг.2.
Результаты доклинического изучения протективности рекомбинантной вакцины Унифлю свидетельствуют о широком спектре её защитного действия. Оно проявлялось в отношении не только вирусов гриппа А, имеющих идентичную аминокислотную последовательность пептида М2е (субтипы вируса гриппа А/Н3, А/Н2, А/Н1), но и вируса А(H1N1)pdm09, у которого аминокислотная последовательность М2е пептида отличается по 4 позициям. С большой долей вероятности можно предположить, что такой же защитный эффект (около 80%) вакцина Унифлю будет иметь в отношении вирусов субтипа А/Н5, включая высокопатогенный вирус А/H5N1, так как пептид М2е вирусов А(H1N1)pdm09 и А/H5N1 отличается всего на одну аминокислоту в позиции 12, не критичной для связывания специфических антител. Протективность вакцины по отношению к другим потенциально пандемическим вирусам гриппа птичьего происхождения требует дополнительных исследований.
Исследование протективности вакцины на мышах показало, что вакцинация уменьшает тяжесть инфекции. Так, максимальная потеря массы тела в опытных группах при заражении разными вирусами, идентичными вакцине по M2e-пептиду, составляла 16,3-17,7%, тогда как в группах плацебо - 20,1-35,3%. Вакцина Унифлю обеспечивала 90-100% защиту животных от летального заражения (5 LD50) вирусами гриппа А человека различных субтипов: A/PR/8/34, A/Japan/305/57, A/Aichi/2/68 (Фиг. 3).
Таким образом, описанная кандидатная вакцина, которая является лиофилизированным препаратом, содержащим в 1 дозе (0,5 мл) высокоочищенный рекомбинантный белок НВс4М2е (40,0 ± 5,0 мкг) с консенсусной последовательностью М2е вирусов гриппа А человека (A/H1N1, A/H3N2, A/H2N2), адъювант - Деринат® (500 ± 50 мкг), фосфатно-буферный раствор (ФБР), рН 7,2-7,4, является хорошим кандидатом для применения в качестве универсальной противогриппозной вакцины на основе рекомбинантного белка с точки зрения ее иммуногенности и протективности.
Однако для крупномасштабного производства противогриппозных вакцин с целью защиты наибольшего процента населения от вируса гриппа А человека одним из важнейших показателей вакцины является ее стабильность. Повышение стабильности, которая является основным из важнейших показателей вакцины с точки зрения ее производства, применения и хранения, необходимо для обеспечения ее производства и возможности проведения вакцинации населения. Поэтому авторы настоящего изобретения решали задачу получения вакцин, обладающих наилучшей стабильностью.
Наиболее показательной оценкой стабильности вакцины является измерение дзета-потенциала, так как дзета-потенциал (ZP) - важный физико-химический показатель стабильности частиц. На границе между частицей и окружающей жидкостью возникает потенциал, который вызывает возникновение электростатических сил отталкивания, что обеспечивает стабильность раствора и препятствует агрегированию частиц. Чем выше значение дзета-потенциала, тем более стабильной можно считать систему. Абсолютные значения ZP выше 30 мВ обеспечивают хорошую стабильность, а выше 60 мВ - прекрасную стабильность. Около 20 мВ - только кратковременную стабильность, от -5 мВ до +5 мВ - показывают быструю агрегацию частиц. Согласно статье Soheyla Honary and Foruhe Zahir Effect of Zeta Potential on the Properties of Nano-Drug Delivery Systems - A Review (Part 2) .Tropical Journal of Pharmaceutical Research April 2013; 12 (2): 265-273, которая включена в настоящую заявку в полном объеме посредством ссылки, высокий ZP (положительный или отрицательный), обычно более 30 мВ, поддерживает стабильность системы. Кроме того, известно, что наночастицы с высоким поверхностным зарядом фагоцитируются более эффективно мышиными макрофагами (в 1,3 раза). Негативно заряженные сферические частицы потенциально могут связываться с доступными катионными сайтами на поверхности макрофагов и распознаваться scavenger рецепторами.
Авторы настоящего изобретения в статье TSYBALOVA L.M. et al. «Development of a candidate influenza vaccine based on virus-like particles displaying influenza M2e peptide into the immunodominant region of hepatitis B core antigen: Broad protective efficacy of particles carrying four copies of M2e. Vaccine. 2015; 33(29): 3398-406, которая включена в настоящую заявку в полном объеме посредством ссылки, раскрыли на Фиг. 2 сведения по иммунногенности вакцины, содержащей 50 мкг белка HBc/4M2eh и 100 мг Деринат®. В данной статье указано, что Деринат® оказывает влияние на иммунногенность рекомбинантного белка HBc/4M2eh, что также подтверждено данными, представленными на Фиг. 2. Однако данные по стабильности вакцины не приводятся.
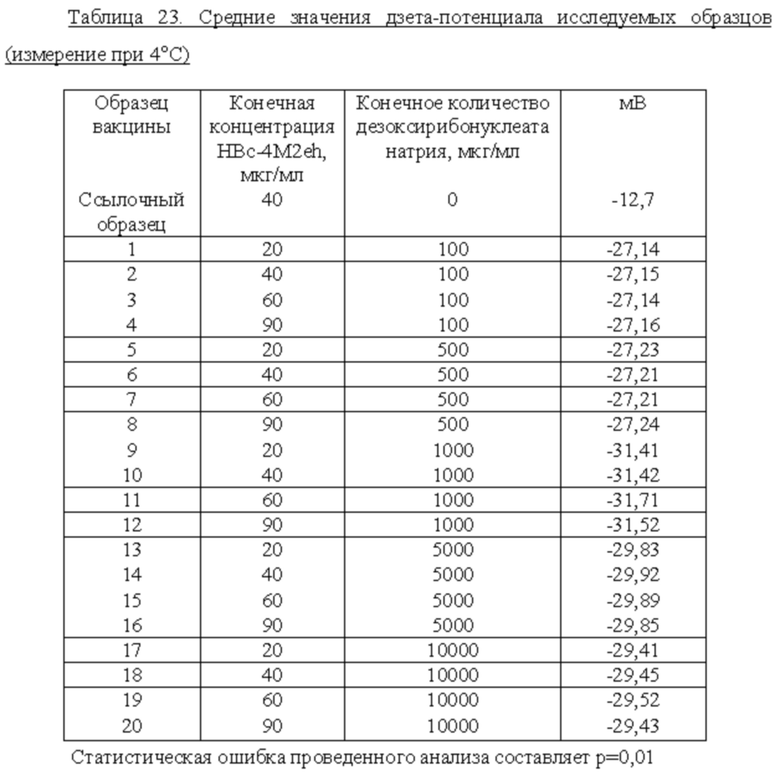
Для измерения дзета-потенциала известных вакцин на основе рекомбинантного белка HBc4M2eh (40 мкг) с содержанием 100 мкг и 500 мкг дезоксирибонуклеата натрия, а также белка HBc4M2eh без дезоксирибонуклеата натрия, поверхностный заряд частиц (дзета-потенциал) оценивали по стандартной методике, описанной в примере 2 ниже.
Полученные данные показали, что вакцина на основе рекомбинантного белка HBc4M2eh с содержанием 100 мкг дезоксирибонуклеата натрия имеет значение дзета-потенциала -27,15 мВ, и вакцина на основе рекомбинантного белка HBc4M2eh с содержанием 500 мкг дезоксирибонуклеата натрия имеет значение дзета-потенциала -27,21 мВ.
Таким образом, данные вакцины, содержащие дезоксирибонуклеат натрия в количестве 100 мкг и 500 мкг, не обеспечили достижение значения дзета потенциала, максимально близкого к 30 мВ, что свидетельствует о необходимости обеспечения вакцин, обладающих большей стабильностью.
Авторами настоящего изобретения неожиданно было обнаружено, что вакцины, содержащие рекомбинантный белок HBc-4M2eh в количестве от 20 до 90 мкг и адьювант дезоксирибонуклеат натрия в количестве 1000 до 10000 мкг, обладают улучшенной стабильностью, более высокой чем у описанной в настоящее время вакцины «Унифлю».
Получение образцов вакцин согласно настоящему изобретению и определение дзета потенциала описано в Примере 2 ниже.
Результаты описанных далее экспериментов неожиданно показали, что повышение дзета-потенциала происходит в строго определенном диапазоне содержания адъюванта дезоксирибонуклеата натрия: от 1000 до 10000 мкг.
Таким образом, авторы настоящего изобретения обнаружили, что содержание адъюванта дезоксирибонуклеата натрия от 1000 до 10000 мкг приводит к неожиданному увеличению дзета-потенциала, причем стабильные вакцины могут быть получены строго в указанном диапазоне содержания адъюванта дезоксирибонуклеата натрия, так как дальнейшее увеличение содержания адъюванта дезоксирибонуклеата натрия приводит к снижению дзета-потенциала.
В приведенных далее примерах раскрыты результаты реактогенности, безопасности и иммуногенности рекомбинантной универсальной гриппозной вакцины Унифлю у здоровых добровольцев в возрасте от 18 до 60 лет, а также подтверждена улучшенная стабильность и иммуногенность вакцины согласно настоящему изобретению.
Примеры
Список сокращений
Пример 1. Реактогенности, безопасности и иммуногенности рекомбинантной универсальной гриппозной вакцины Унифлю у здоровых добровольцев в возрасте от 18 до 60 лет
На основании описанных выше и опубликованных доклинических исследований кандидатной вакцины Унифлю были сделаны следующие выводы:
При оценке полипептидного состава вакцины Унифлю методом электрофореза в градиентном разделяющем полиакриламидном геле (8-16%) с додецилсульфатом натрия показано, что вакцина Унифлю содержит в своем составе только рекомбинантный белок с антигеном вируса гриппа (М2е); количество посторонних белковых примесей в вакцине составляет не более 7,5%; подлинность рекомбинантного белка (наличие целевого антигена эктодомена белка М2) подтверждена в иммуноблоте с использованием коммерческих моноклональных антител 14С2, специфичных для М2е. Структурно-морфологическая характеристика препарата была подтверждена с помощью электронной микроскопии, показано наличие вирусоподобных частиц, образуемых рекомбинантным белком диаметром 32-36 нм. По показателям острой токсичности вакцина Унифлю, введенная мышам и крысам, внутримышечно и внутривенно, в дозах, превышающих терапевтическую дозу для человека более чем в 3500 раз, не вызвала существенного повреждающего действия на организм опытных животных, его органы и системы. При изучении субхронической токсичности вакцины Унифлю у крыс обоего пола выявлено отсутствие токсического действия препарата на интегральные показатели жизнедеятельности и систему терморегуляции в течение всего периода наблюдения, значимых различий в уровне показателей функций сердечно-сосудистой и дыхательной систем; изучение гематологических, биохимических показателей крови крыс после введения вакцины и животных контрольной группы не выявило существенных различий. При изучении субхронической токсичности вакцины Унифлю у кроликов обоего пола выявлено отсутствие токсического воздействия на интегральные показатели жизнедеятельности организма и систему терморегуляции; показатели функции сердечно-сосудистой и дыхательной системы у опытных животных не отличались от таковых у контрольных; количественный и качественный морфологический состав периферической крови кроликов соответствовал физиологической норме; биохимические показатели периферической крови кроликов соответствовали видовой физиологической норме. Вакцина Унифлю не обладала местно-раздражающим действием. Результаты патоморфологических и гистопатологических исследований не выявили изменений в органах животных, связанных с иммунизацией. Вакцина Унифлю не оказала влияния на репродуктивную функцию крыс обоего пола и не обладала эмбриотоксическим действием. Вакцина Унифлю не обладала аллергизирующим и иммунотоксическим действием. При изучении на мышах и хорьках иммуногенных свойств вакцины Унифлю показано формирование антител к линейным и конформационным эпитопам эктодомена белка М2 вирусов гриппа А человека разных субтипов, М2е-специфического Т-клеточного ответа (Тх 1-го типа и цитотоксических Т-лимфоцитов). Показано 90-100% протективное действие вакцины у иммунизированных мышей гомологичными вирусами гриппа А человека различных субтипов: A/PR/8/34 (H1N1), A/Japan/305/57 (H2N2), A/Aichi/2/68 (H3N2) и 80% защита от гетерологичного по М2е вируса гриппа A/California/7/2009 (H1N1) pdm09; вакцина Унифлю снижала на 2,0-5,0 lg репродукцию вирусов гриппа А разных субтипов в легких мышей и носовых смывах хорьков.
Таким образом, было показано, что Унифлю - вакцина гриппозная рекомбинантная универсальная - является безопасным и эффективным препаратом в отношении различных субтипов вирусов гриппа А человека. Исследование было зарегистрировано на сайте ClinicalTrials.gov: UIV-I-01/2017 NCT03789539 Reactogenicity, Safety and Immunogenicity of an Universal Influenza Vaccine Uniflu. Public 12/25/2018 06:35.
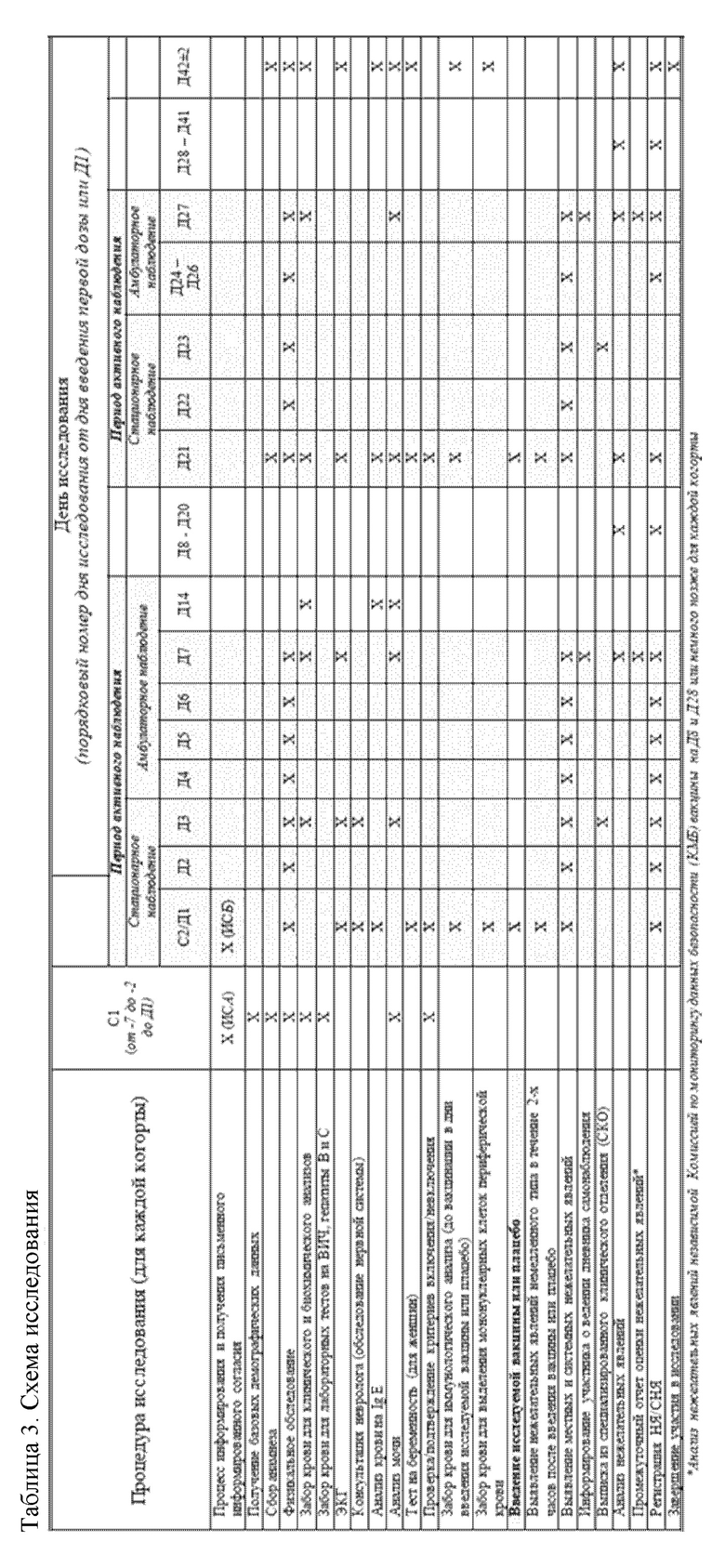
Процедуры исследования и визиты добровольцев в обобщенном виде были представлены в Схеме исследования (Таблица 3).
Для участия в исследовании были приглашены здоровые добровольцы (мужчины и женщины) в возрасте от 18 до 60 лет. Методом случайной выборки (рандомизация) 54 участника исследования были распределены в соотношении 2:1 для введения препарата вакцины двух уровней доз или плацебо. Были сформированы 2 когорты по 27 человек, в каждой из которых предусмотрено введение вакцины высокого или низкого уровня дозы и соответствующего плацебо (18 человек в группе вакцины и 9 человек в группе плацебо).
С целью оценки целесообразности проведения данного исследования Комиссией по мониторингу данных безопасности (КМБ) был проведен первоначальный анализ данных по безопасности каждой дозы вакцины у небольшого числа добровольцев каждой когорты. Для этой цели включение в исследование 54 человек происходило в четыре этапа.
На первом этапе в исследование было включено 12 добровольцев когорты 1. После завершения первого срока активного наблюдения с 1-го по 7-й день исследования КМБ провела первый промежуточный анализ показателей безопасности у 12 добровольцев когорты 1. КМБ провел анализ всех случаев нежелательных явлений, а также результатов лабораторных исследований (до и после вакцинации). На основании проведенного анализа КМБ представила свои рекомендации (i) о целесообразности проведения второй вакцинации у 12 добровольцев когорты 1 и (ii) включения в исследование следующих 15 человек когорты 1.
Затем КМБ провела второй промежуточный анализ всех случаев нежелательных явлений, а также результатов лабораторных исследований (до и после вакцинации) у всех добровольцев когорты 1. На основании проведенного анализа КМБ представила свои рекомендации о целесообразности введения второй дозы исследуемого препарата 15 добровольцам когорты 1 и начала исследования для небольшого числа добровольцев когорты 2 (12 добровольцев).
После окончания периода оценки безопасности препарата у 12 добровольцев когорты 2 КМБ представила третий промежуточный отчет о безопасности препарата высокого уровня дозы и дала рекомендации о (i) включении оставшихся 15 добровольцев когорты 2 и (ii) целесообразности проведения второй вакцинации 12 добровольцам данной когорты.
Ниже представлено краткое описание процедур исследования и указаны сроки их проведения в обеих когортах добровольцев.
В день первого скринингового обследования (С1), приблизительно за 5 дней (в период от - 2-го до - 7-го дня до начала исследования) до введения первой дозы исследуемой вакцины или плацебо было проведено обследование добровольцев по установлению их пригодности для участия в исследовании на основании данных медицинского анамнеза, физикального обследования добровольцев, серологического тестирования на наличие хронических вирусных инфекций [вирус иммунодефицита человека (ВИЧ), вирусы гепатита В (ВГВ) или С (ВГС)], стандартного биохимического и гематологического исследования крови и анализа мочи.
Обследование добровольцев, отвечающих критериям включения/невключения в исследование, было продолжено в день скринингового обследования 2 (С2). У женщин был проведен анализ образцов мочи на беременность. Всем добровольцам была проведена ЭКГ. У лиц, полностью отвечающих критериям включения/невключения и допущенных к вакцинации, были собраны образцы крови для проведения исследования на IgE, а также иммунологических исследований.
После введения исследуемой вакцины или плацебо всех участников исследования активно наблюдали в течение 7 дней. В этот период добровольцы находились в условиях стационара в течение первых 3-х дней после вакцинации (День 1 - День 3), затем совершали ежедневные визиты в исследовательский центр до 7 дня исследования, в ходе которых у добровольцев проводили тщательный мониторинг развития побочных реакций (нежелательных явлений). На 3-й и 7-й дни исследования у участников исследования были собраны образцы крови и мочи, а также проведено ЭКГ (только 7-й день). После периода активного наблюдения (с вечера 7-го дня исследования) каждый участник исследования вел записи в Дневнике наблюдения №1 для самостоятельного учета всех случаев развития у него НЯ, а также приема сопутствующих лекарственных препаратов.
Участники исследования возвращались в клинический центр через три недели после введения первой дозы исследуемой вакцины или плацебо (21-й день исследования). В этот день, до введения второй дозы исследуемой вакцины или плацебо, у добровольцев проводили сбор образцов крови для проведения стандартного биохимического и гематологического исследований крови и сбор образцов мочи, у женщин проводили тест на беременность. Помимо этого, был проведен анализ данных медицинского анамнеза добровольцев (в т.ч., записей в Дневниках наблюдения №1) за промежуточный период, начиная с 7-го дня исследования, когда было завершено активное наблюдение поле первой вакцинации.
Добровольцам, у которых по данным физикального обследования и клинической лабораторной диагностики не были выявлены неблагоприятные явления 2-й степени тяжести или выше, согласно шкале оценки уровня тяжести НЯ, а также имевшим отрицательный тест на беременность (у женщин), была проведена повторная вакцинация исследуемой вакциной или плацебо с последующим наблюдением и медицинским контролем в условиях стационара (с 21-го по 23-й дни) и амбулаторно (с 24-го по 27-й дни). В ходе визитов у добровольцев проводили тщательный мониторинг развития побочных реакций (нежелательных явлений). После завершения второго периода активного амбулаторного наблюдения за добровольцами (с 21-го по 27-й дни исследования) участники исследования вели записи в Дневниках самонаблюдения №2 по учету развития у них НЯ, а также приема сопутствующих лекарственных препаратов. Добровольцы возвращались в клинический центр через три недели после введения второй дозы исследуемой вакцины или плацебо для заключительного контрольного медицинского обследования (42-й день исследования). Процедуры заключительного дня исследования включали анализ данных медицинского анамнеза добровольцев (в т.ч., записей в Дневниках самонаблюдения №2) за второй промежуточный период, ЭКГ, а также заключительные процедуры по сбору у участников исследования образцов крови и мочи. У женщин был проведен анализ мочи на беременность. В этот день участие добровольцев в данном исследовании было завершено.
С целью оценки безопасности исследуемой вакцины медицинское наблюдение за состоянием здоровья добровольцев осуществляли в течение двух часов после каждого введения исследуемой вакцины или плацебо. В день вакцинации и последующие 6 дней после каждого введения исследуемой вакцины или плацебо контроль над развитием НЯ проводили во время пребывания в стационаре и ежедневных визитов добровольцев в клинический центр. Добровольцы вели записи в Дневниках самонаблюдения №1 и №2 по самостоятельному учету развития у них непредвиденных НЯ после каждого срока активного наблюдения до заключительного визита для контрольного медицинского обследования (42-й день исследования). С целью оценки параметров безопасности на 3-й, 7-й, 21-й (до введения второй дозы исследуемой вакцины/плацебо), 27-й и 42-й дни исследования у участников исследования осуществляли сбор образцов крови и мочи для стандартного биохимического и гематологического исследования крови и анализа мочи.
Для оценки содержания сывороточных антител IgG (иммуноглобулины класса G) к М2е антигену, сывороточных антител IgG к коровому антигену вируса гепатита В (HBc) и IgE в ИФА сбор образцов крови осуществляли в 1-й день (до введения первой дозы исследуемой вакцины/плацебо), 14-й день, 21-й день (до введения второй дозы исследуемой вакцины/плацебо) и 42-й день исследования. Для оценки клеточного иммунного ответа (продукция внутриклеточного ИФН-γ в активированных антигеном М2е CD3+CD4+ и CD3+CD8+ Т-лимфоцитах (по результатам анализа мононуклеарных клеток периферической крови (МПК) с использованием методов проточной цитометрии) осуществляли сбор периферической цельной крови в 1-й день (до введения первой дозы исследуемой вакцины/плацебо) и на 42-й день исследования.
В качестве плацебо использовали Деринат® (500 мкг), растворенный в вакцинном буфере (фосфатный буфер рН 7.2-7.4), содержащем 0,5% полиглюкин.
Препарат Деринат® представляет собой натриевую соль ДНК (олигодезоксинуклеотид - ОДН эукариотического происхождения), деполимеризованную ультразвуком до частиц с молекулярной массой 270-500 кДа (Р N002916/01 дата регистрации 27.02.2008).
В качестве показателя профиля безопасности вакцины была определена доля лиц, у которых было зарегистрировано развитие нежелательных явлений (НЯ) четырех нижеследующих категорий:
Нежелательные явления немедленного типа, возникающие в течение двух часов после введения какой-либо дозы вакцины и выявленные как медицинским персоналом, так и по информации, сообщенной добровольцем персоналу исследования.
Поствакцинальные реакции (предвиденные клинические проявления местного и системного характера), как правило, возникающие вследствие внутримышечной вакцинации в период через два часа и последующих 7 дней после введения какой-либо дозы исследуемой вакцины или плацебо и выявленные как медицинским персоналом, так и по информации, сообщенной добровольцем персоналу исследования.
Другие нежелательные явления (включая непредвиденные клинические проявления), возникающие в течение 7 дней после введения какой-либо дозы исследуемой вакцины или плацебо и выявленные как медицинским персоналом, так и по информации, сообщенной добровольцем персоналу исследования. При этом также учитывали патологические изменения данных лабораторного анализа образцов крови и мочи, собранных на 3-й, 7-й, и 27-й дни исследования, а также данные ЭКГ-обследования, проведенные на 3-й, 7-й, 21-й и 42-й дни исследования.
Все серьезные нежелательные явления (СНЯ), возникающие в период до 3-х недель после введения какой-либо дозы препарата и выявленные как медицинским персоналом, так и по информации, сообщенной добровольцем персоналу исследования или зарегистрированной вакцинируемым в Дневниках самонаблюдения. При этом также учитывали патологические изменения данных лабораторного анализа образцов крови и мочи, данные ЭКГ-обследования, проведенные на 3-й, 7-й, 21-й (перед второй вакцинацией) и 42-й дни исследования.
Был произведен подсчет общего количества всех нежелательных явлений (НЯ), которые затем были суммированы по степени тяжести, как "любое местное НЯ", или "любое системное НЯ", а также по взаимосвязи с введенной исследуемой вакциной или плацебо, исходя из заключения врача-исследователя при слепом методе исследования. Процент добровольцев с проявлениями НЯ или, по крайней мере, одним случаем НЯ, был рассчитан с 95% точным доверительным интервалом (ДИ).
По результатам проведенного клинического исследования по протоколу УГВ-I-01/2017 вакцина гриппозная универсальная рекомбинантная лиофилизированная Унифлю для внутримышечного введения характеризовалась хорошей переносимостью и была безопасна при двукратном внутримышечном введении. В ходе исследования серьезных нежелательных явлений, связанных с вакцинацией, отмечено не было. Возникновения местных нежелательных явлений у добровольцев, получивших исследуемый препарат и препарат сравнения, зарегистрировано не было. Системных нежелательных явлений, а также изменений инструментальных и лабораторных показателей (по результатам общего и биохимического анализов крови, анализа мочи, ЭКГ), связанных с вакцинацией выявлено не было.
Оценка иммуногенной активности
Исследование гуморального иммунного ответа на вакцину «Унифлю»
Гуморальный иммунный ответ, индуцируемый вакциной гриппозной универсальной рекомбинантной лиофилизированной Унифлю, оценивали по проценту привитых с достоверными 4-кратными и более приростами титров антител в сыворотках крови после второй вакцинации (сероконверсии), а также средним геометрическим титрам антител.
Поствакцинальный М2е-специфический гуморальный иммунный ответ изучали при помощи непрямого иммуноферментного анализа (ИФА). В сыворотках крови, полученных от добровольцев до вакцинации, на 21-е и 42-е сутки исследования (после однократного и двукратного введения 0,5 дозы, 1 дозы вакцины Унифлю или плацебо) определяли титры М2е-специфических антител изотипа IgG, подклассов IgG1 и IgG2. На 96-луночные планшеты с высокой сорбционной способностью (Greiner, Германия) сорбировали синтетический пептид M2e (SLLTEVETPIRNEWGCRCNDSSD), соответствующий последовательности М2е в составе вакцины. В качестве конъюгатов использовали вторичные козьи антитела меченые пероксидазой хрена (Abcam, UK): к IgG человека, в разведении 1:5000, к IgG1 человека, в разведении 1:500, к IgG2 в разведении 1:1000. Учет реакции проводили при длине волны 450нм. За титр принимали наибольшее разведение сыворотки, которое дает оптическую плотность, по крайней мере, в 2 раза больше, чем бланк.
Изучение специфического гуморального иммунного ответа проводили при помощи сравнения титров М2е-специфических антител класса IgG, подклассов IgG1, IgG2, а также их соотношения IgG1/IgG2 , формирующихся в ответ на иммунизацию вакциной Унифлю в разных дозах (0,5 и 1 доза) в сравнении с группой Плацебо. Кроме того, сопоставляли данные о титрах анти-М2е антител до иммунизации, после первой и второй вакцинации в пределах каждой группы (День 1, День 21 и День 42).
В таблице 4 представлены данные по титрам анти-М2е IgG в группах добровольцев на различных сроках исследования. Статистическую значимость различий в титрах антител на разных сроках исследования определяли с помощью дисперсионного анализа (ANOVA, Greenhouse-Geisser correction).
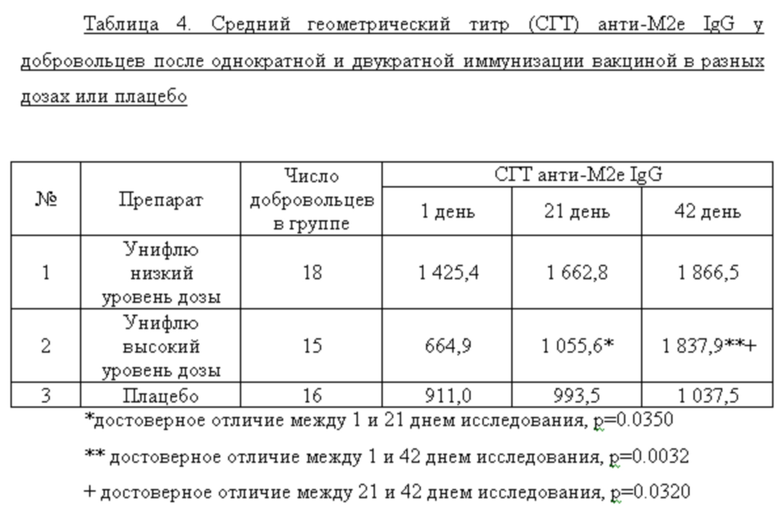
Согласно данным, представленным в таблице 4, у добровольцев, получавших вакцину высокой дозы (1 доза) отмечено достоверное нарастание титров анти-М2е IgG после первой (День 21), так и после второй (День 42) вакцинации. В группе добровольцев, получавших половинную дозу вакцины, а также в группе плацебо достоверных изменений в титрах специфических антител выявлено не было. Анализ соотношения подклассов IgG (IgG1/IgG2) на различные дни исследования показал возрастание доли анти-М2е IgG1 только у добровольцев, получавших высокую дозу вакцины (таблица 5). Это имеет принципиальное значение, так как у человека подкласс IgG1 связывается с высокой аффинностью с активирующими Fc рецепторами и является основным эффектором АЗКЦ. В этой связи возрастание доли анти-М2е IgG1 у добровольцев после иммунизации вакциной высокой дозы является положительным моментом формирования полноценного поствакцинального гуморального ответа.
В целом, анти-М2е IgG обеспечивают антивирусную защиту путем элиминации зараженных клеток с помощью механизма антитело-зависимой клеточной цитотоксичности (АЗКЦ) или комплемент обусловленного фагоцитоза. У мышей подкласс IgG2a - аналог человеческого IgG1 - обладает сильными эффекторными функциями и преимущественно участвует в АЗКЦ благодаря способности эффективно связываться со всеми тремя типами Fc рецепторов (FcγRI, FcγRIII, FcγRIV). Показано также, что высокий уровень анти-М2е IgG2a у мышей коррелирует с защитой против вирусов гриппа А.
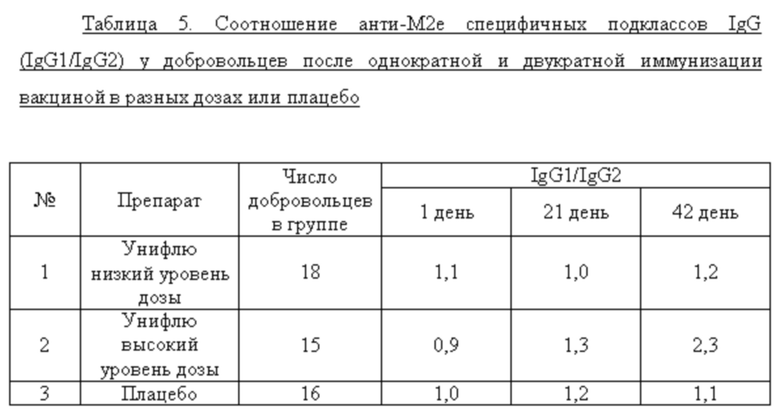
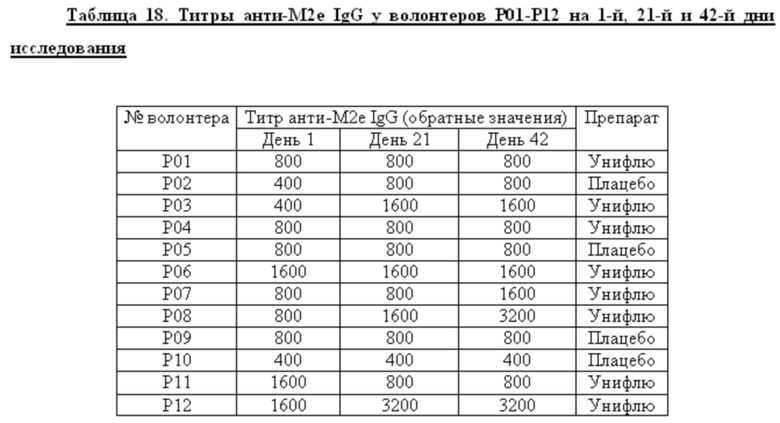
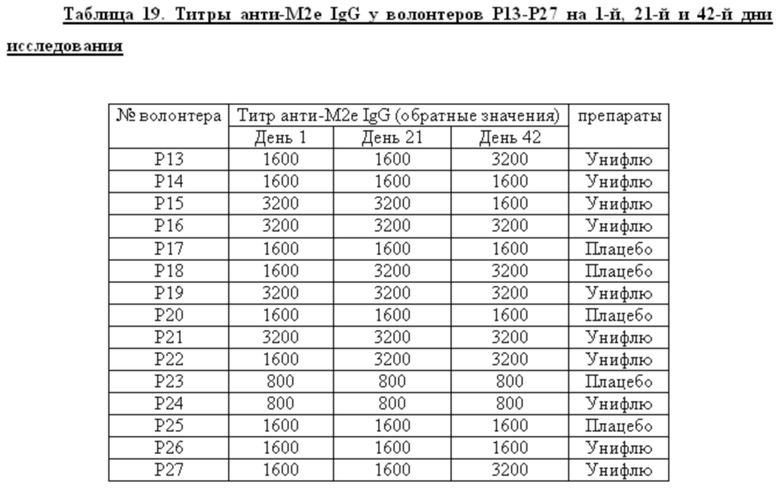
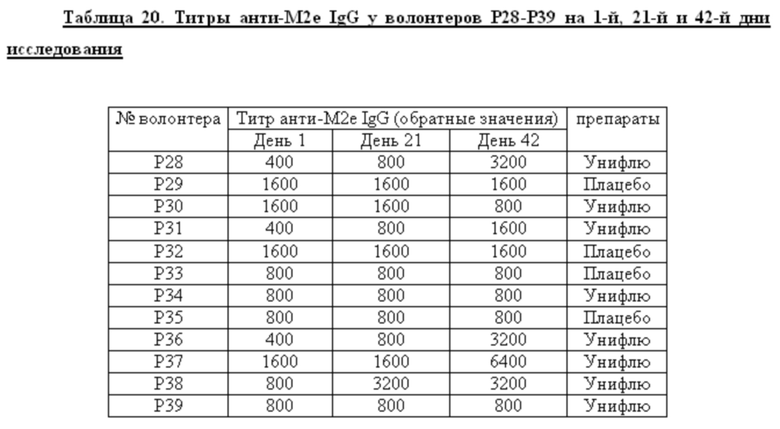
Среди добровольцев, получавших вакцину в разных дозах или плацебо, определяли долю лиц, имевших 4-х кратный прирост титров анти-М2е IgG к 42 дню исследования. У этих добровольцев определяли также СГТ анти-М2е IgG, IgG1/IgG2. Данные представлены в таблице 6. Анализ поствакцинального гуморального ответа у добровольцев, получавших вакцину в высокой и низкой дозах, показал наличие дозо-зависимого эффекта. Процент сероконверсий среди добровольцев, получавших 0,5 дозы вакцины, составил 11,1%, а среди добровольцев, получавших 1 дозу вакцины, возрос до 53,3%. Кроме того, среди добровольцев с сероковерсией отмечается возрастание доли анти-М2е антител подкласса IgG1.
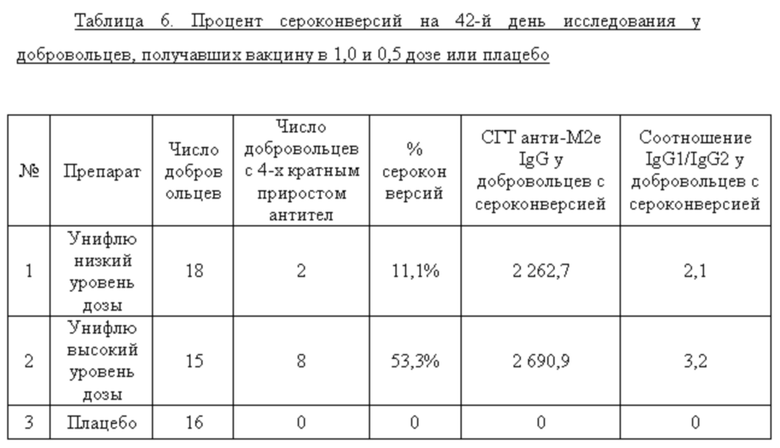
Показано, что формирование поствакцинального гуморального ответа у волонтеров, получавших вакцину Унифлю, носило дозо-зависимый характер. Вакцинация высокой дозой (1 прививочная доза) вызывала достоверный прирост титров специфических антител к 42-му дню исследования (процент сероконверсий составил 53,3%) и возрастанию доли антител подкласса IgG1, являющихся основным медиатором АЗКЦ у человека. «Ответчики» по гуморальному иммунитету (на 1 прививочную дозу): Р28, Р31, Р36, Р37, Р38, Р49, Р51, Р54 - 8 чел.
Определение антител (IgG+IgM) к коровому антигену (HBc) вируса гепатита В сыворотке крови волонтеров
Для выявления у добровольцев суммарных антител (IgM+IgG) к коровому антигену вируса гепатита В (HBc) использовали тест-систему Monolisa Anti-HBc PLUS (Bio-Rad, Франция; Lot 8A0616, годен до 13.05.2019). Тест-система Monolisa Anti-HBc PLUS позволяет выявить общие антитела (IgM+IgG) к HBc в сыворотке или плазме крови человека. На поверхность лунок 96-луночных планшетов наносится рекомбинантный HBc. Во время инкубации исследуемых проб в лунках планшета сывороточные антитела образуют комплекс с рекомбинантным HBc, адсорбированным на поверхность лунок. После промывки лунок от несвязанного материала в лунки вносится меченые пероксидазой хрена козьи антитела к человеческим IgG и IgM, которые связываются с комплексом HBc-антитела. На заключительном этапе несвязанный конъюгат вымывается и в лунки вносится субстрат. Последующее окрашивание прямо пропорционально количеству анти-HBc антител в исследуемых пробах. Постановка ИФА проводилась в соответствии с инструкцией к тест-системе. Отрицательный контроль (ОК) и сыворотки крови волонтеров исследовались в дубликатах. Положительный контроль (ПК) исследовался в трипликатах. Измерение оптических плотностей (ОП) исследуемых проб проводилось на планшетном ридере i-Mark при длине волны 450/655 нм.
В соответствие с инструкцией к тест-системе исследования считались правильными, если:
- для отрицательного контроля (ОК) каждое измеренное значение оптической плотности (ОП) меньше, чем 0,100;
- для положительного контроля (ПК) каждое измеренное значение ОП больше или равно 1,000 и меньше или равно 2,900.
Ни одно из трех значений ПК не выходило за пределы указанных значений или отличалось от среднего более чем на 30%.
Вычисляли среднее значение ОП (ОПср) лунок с ПК. Рассчитывали значение cut off (Vs) по формуле:
Vs = ОПсрПК:5.
Сыворотки волонтеров с ОП (оба значения) меньше, чем значение Vs считались отрицательными. Сыворотки волонтеров с ОП больше или равной Vs считались положительными.
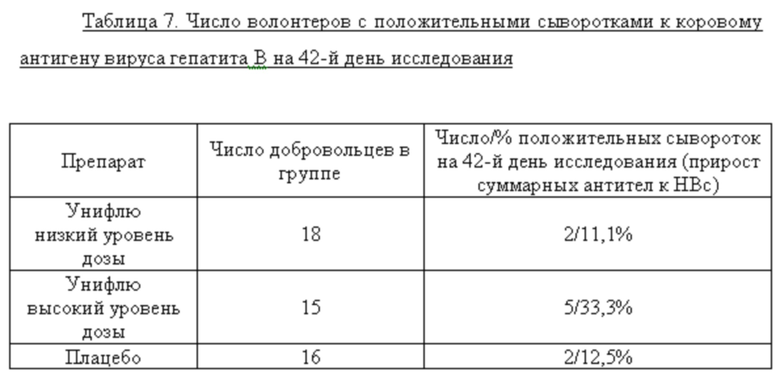
Данные по результатам определения анти-HBc IgM+IgG в сыворотке волонтеров на 42-й день исследования представлены в таблице 7. Показано, что двукратная иммунизация добровольцев вакциной Унифлю существенно не стимулирует образование антител к коровому антигену вируса гепатита В (белку-носителю антигена вируса гриппа в исследуемой вакцине).
Оценка антитело-зависимой комплемент обусловленной цитотоксичности (АЗКЦ) в сыворотках крови волонтеров на 42-й день исследования
Антитело-зависимую комплемент обусловленную цитотоксичность (АЗКЦ) использовали для определения анти-вирусного эффекта анти-М2е антител после двукратной иммунизации добровольцев вакциной Унифлю. Использовали метод, предложенный J. Atsmon et. al в модификации. Для учета результата (% лизированных клеток) использовали МТТ-тест, показывающий изменение митохондриальной активности и широко использующийся для измерения выживаемости клеток.
Суточный монослой клеточной линии MDCK заражали вирусом гриппа A/Aichi/2/68 (H3N2) с инфекционной активностью ТЦД50=3 lg и инкубировали в течение 18 часов при +370С и 5% CO2. В лунки вносили сыворотки добровольцев в разведениях 1:10, 1:20, 1:40, 1:80 и комплемент морской свинки (Комплемент сухой, МикроГен, Россия) в разведении 1:10. Инкубировали в течение 4 часов при +370С и 5% CO2. В качестве контролей использовали незараженную вирусом клеточную культуру MDCK (КК), клетки, инкубированные с вирусом гриппа (КВ), клетки, инкубированные с вирусом гриппа и комплементом (КВК), и клетки, инкубированные с вирусом гриппа и опытными сыворотками (КС). Все образцы брались в трех повторах, для каждого считалось среднее значение оптической плотности (ОП).
Процент специфического иммунного лизиса (СИЛ) рассчитывали по формуле:
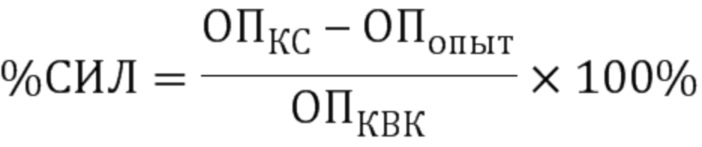 ,
,
где ОПКС - среднее значение ОП контроля КС (MDCK+вирус+сыворотка),
ОПопыт - среднее значение ОП опытной пробы (MDCK+вирус+сыворотка+комплемент),
ОПКВК - среднее значение ОП контроля комплемента (MDCK+вирус+комплемент).
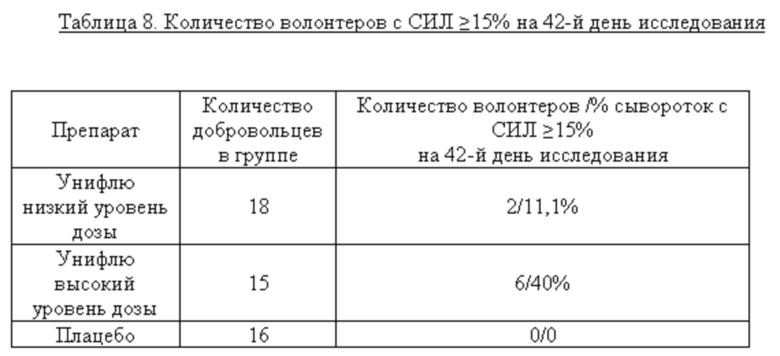
В таблице 8 представлены данные, показывающие % добровольцев, у которых выявлен СИЛ≥15%.
Исследование Т-клеточного иммунного ответа на вакцину «Унифлю»
Поствакцинальный Т-клеточный иммунный ответ исследовали при помощи метода многоцветной проточной цитометрии. Мононуклеарные клетки периферической крови (МПК), полученные от добровольцев до вакцинации и через 42 дня после введения двух доз вакцины или плацебо, окрашивали при помощи панели флуоресцентно-меченных моноклональных антител к поверхностным антигенам Т-лимфоцитов (CD4, CD8, CD45RO, CD69) и внутриклеточным цитокинам (IFN-γ, TNF-α).
Фракцию одиночных лимфоидных клеток выявляли на основании характеристик светорассеяния (FSC/SSC). Живые и мертвые клетки разделяли при помощи маркера жизнеспособности Zombie Aqua. По наличию маркеров CD4 или CD8 выявляли популяции Т-хелперов и цитотоксических Т-лимфоцитов. При помощи маркеров CD45RO и CCR7 указанные популяции подразделяли на наивные Т-клетки (Naïve), эффекторные клетки памяти (EM), центральные клетки памяти (CM) и терминально-дифференцированные эффекторы (TEMRA). Маркер CD69 использовали для выявления популяций, фенотипически соответствующих резидентным эффекторным и центральным Т-лимфоцитам памяти, а также в качестве маркера активации при анализе антиген-специфического иммунного ответа.
Использованная панель антител позволила выявить 12 популяций Т-лимфоцитов, относительное содержание которых отображено в таблице 9.
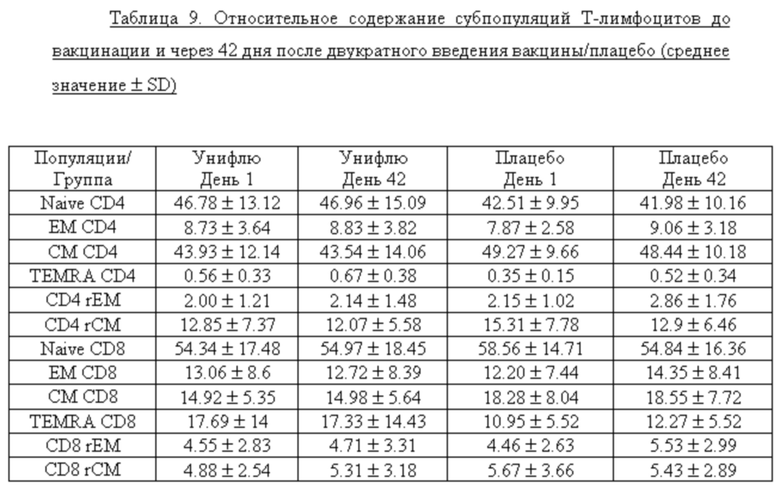
Согласно данным, представленным в таблице 7 наивные Т-клетки составляли в наибольшую субпопуляцию от общего количества CD4 или CD8 Т-лимфоцитов (41.98-58,56%). Кроме того, значительная часть CD4+ Т-лимфоцитов (до 49,27%) была представлена центральными клетками памяти. Наименьшая субпопуляция Т-клеток (0,35-0,67%) была представлена CD4 Т-лимфоцитами с фенотипом терминально-дифференцированных клеток. Изменений в относительном содержании субпопуляций Т-лимфоцитов в ответ на вакцинацию по изученным показателям отмечено не было.
Изучение Т-клеточного иммунного ответа проводили при помощи сравнения относительного содержания цитокин-продуцирующих Т-лимфоцитов, формирующихся в ответ на антиген-специфическую стимуляцию клеток пептидом М2е в группах добровольцев, получавших вакцину или плацебо. Кроме того, сопоставляли данные об уровне цитокин-продуцирующих клеток до и после вакцинации в пределах каждой группы (День 1 и День 42). Помимо продукции цитокинов IFN-γ и TNF-α также учитывали уровень экспрессии активационного маркера CD69 на клетках, секретирующих указанные цитокины.
При анализе антиген-специфического Т-клеточного ответа учитывали относительное содержание IFNg-TNFa+CD69+, IFNg-TNFa+CD69-, IFNg+TNFa-CD69+, IFNg+TNFa-CD69-, IFNg+TNFa+CD69- и IFNg+TNFa+CD69+ Т-лимфоцитов, а также общее число цитокин-продуцирующих Т-клеток, относящихся к субпопуляциям эффекторных клеток памяти (EM), центральных клеток памяти (CM) и терминально-дифференцированных эффекторов (TEMRA).
При проведении статистического анализа из значений, полученных после стимуляции клеток пептидом, вычитали соответствующие фоновые значения для нестимулированных клеток.
Формирование Т-клеточного иммунного ответа на вакцинацию
На первом этапе определяли долю лиц, имевших значимый прирост уровня цитокин-продуцирующих клеток к 42 дню исследования. В качестве потенциальных «ответчиков» были определены лица, у которых уровень цитокинового ответа какой-либо популяции Т-клеток статистически значимо превышал среднее значение по данной популяции (р<0.05). Статистическая достоверность отличий от среднего определяли при помощи одновыборочного t-теста Стьюдента.
Параметр Delta показывает разницу значений доли цитокинпродуцирующих Т-лимфоцитов на 42 день после введения вакцины и соответствующих значений, полученных до иммунизации. Красным цветом выделены номера волонтеров, у которых разница в уровне цитокинового ответа анализируемой популяции Т-клеток (Delta) статистически-значимо превышала среднее значение по данной популяции (р<0.05). Анализ прироста уровня цитокинового ответа проводился для популяций эффекторных клеток памяти (EM), центральных клеток памяти (CM) и терминально-дифференцированных эффекторов (TEMRA). Статистическая достоверность отличий от среднего определялась при помощи одновыборочного t-теста Стьюдента.
Далее потенциальных «ответчиков» тестировали на соответствие двум критериям:
Уровень клеточного ответа на 42 день после иммунизации должен достоверно превышать среднее значение соответствующего показателя в группе плацебо (использовали объединенные показатели группы плацебо, полученные на 1-й и 42-й дни).
Уровень клеточного ответа на 42 день после иммунизации должен достоверно превышать среднее значение соответствующего показателя до вакцинации (день 1).
В результате показано формирование Т-клеточного ответа у 6 из 15 добровольцев, получивших двукратно вакцину «Унифлю» в 1 дозе, т.е. у 40% привитых лиц. Для большинства добровольцев, получивших вакцину, было показано формирование антиген-специфичных клеток, относящихся к различным субпопуляциям (таблица 10).
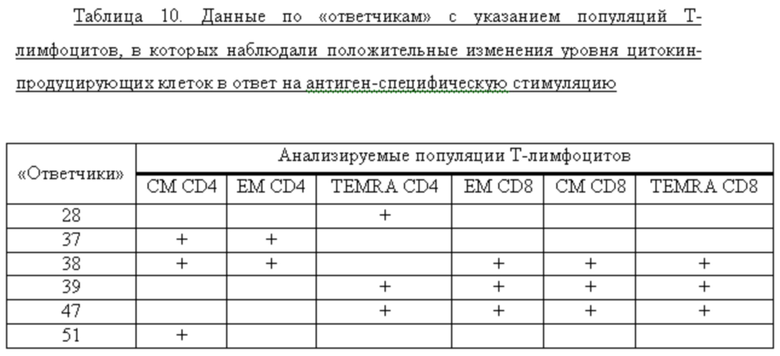
Показано, что вакцина «Унифлю» вызывала иммунный ответ со стороны CD8+ и CD4+ Т-лимфоцитов.
Т-клеточный иммунный ответ через 42 дня после вакцинации
Представлены значения доли клеток, продуцирующих цитокины в ответ на стимуляцию от общего числа CD8+ и CD4+ Т-лимфоцитов соответствующего фенотипа (CM/EM/TEMRA). Сравнение выполнено при помощи парного теста Вилкоксона. Достоверными считались различия при р<0.05 (*)
Установлено, что вакцинация приводит к достоверному увеличению доли цитокин-продуцирующих центральных и эффекторных CD4+ и CD8+ Т-лимфоцитов памяти. Дальнейший анализ популяционного состава цитокин-продуцирующих Т-лимфоцитов показал, что различия между показателями «ответчиков» до и после вакцинации обусловлены, преимущественно, увеличением числа IFNg+TNFa-CD69-, IFNg-TNFa+CD69- и IFNg+TNFa+CD69- CD4+ эффекторных Т-клеток памяти.
Показано, что вакцинация приводит к увеличению относительного содержания Т-лимфоцитов, продуцирующих одновременно два цитокина во всех изученных популяциях, однако достоверные различия отмечены только для CM и EM CD4+ Т-клеток. Полифункциональные IFNg+TNFa+ Т-лимфоциты являются важными медиаторами протекции при различных вирусных инфекциях. Также отмечено увеличение экспрессии активационного маркера CD69 на цитокин-продуцирующих Т-клетках, что может свидетельствовать о дифференцировке данных лимфоцитов в резидентные эффекторные и центральные клетки памяти.
Данные клинического и биохимического анализа крови
Таблица 11. Динамика показателей клинического анализа крови с физиологическими нормами, общими для мужчин и для женщин (M (SD))
Таблица 12. Динамика показателей клинического анализа крови с физиологическими нормами, общими для мужчин и для женщин (Me (Q1 - Q3))
Таблица 13. Динамика показателей биохимического анализа крови с физиологическими нормами, общими для мужчин и для женщин (M(SD))
Таблица 14. Динамика показателей биохимического анализа крови с физиологическими нормами, общими для мужчин и для женщин (Me (Q1 - Q3))
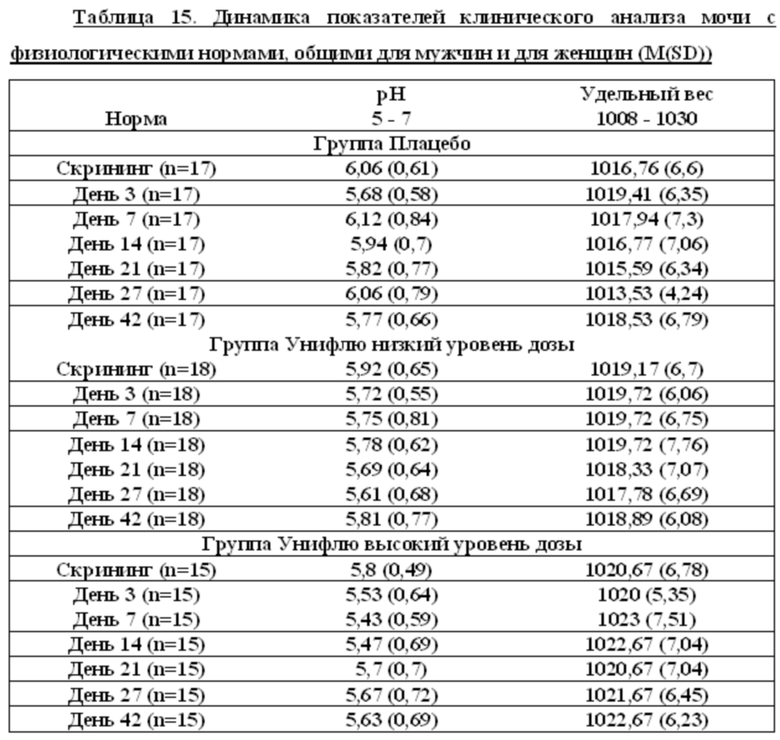
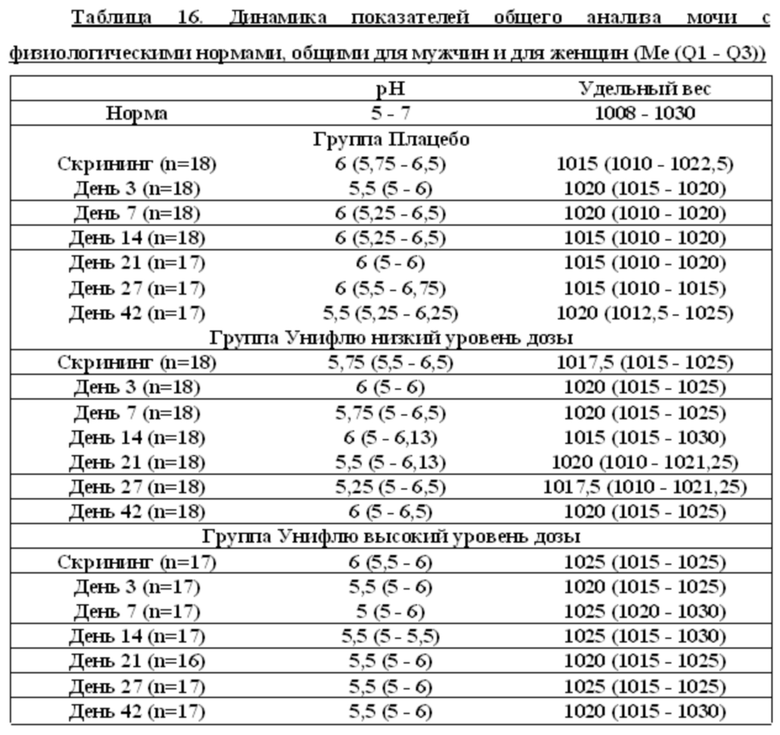
Данные клинического анализа мочи
Показатели белок и глюкоза равны нулю для всех испытуемых на всех визитах, исключение составляет доброволец Р25, для которого белок на 7 сутки равен 1.
Описательные характеристики IgE
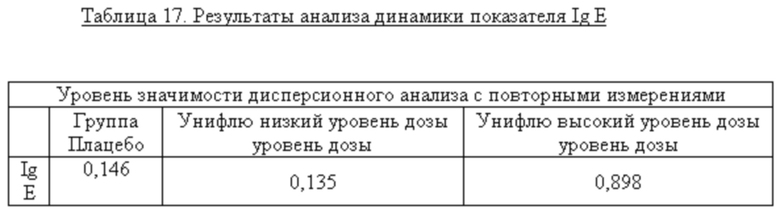
Данные анализа показали отсутствие значимых отклонений результатов клинико-лабораторных показателей у добровольцев от первоначальных показателей на дни Д7, Д14 и Д27, включая уровни IgE, зарегистрировано не было. Средние значения показателей периферической крови (клинический анализ крови с лейкоцитарной формулой) в группах привитых соответствовали нормальным значениям, и, несмотря на динамические изменения, не выходили за их пределы. Нежелательных явлений немедленного типа после введения добровольцам препаратов, а также системных НЯ ни в одной из групп (вакцина, плацебо) отмечено не было. Серьезных нежелательных явлений в ходе исследования отмечено не было.
Таким образом, по результатам проведенного клинического исследования по протоколу вакцина гриппозная универсальная рекомбинантная лиофилизированная Унифлю для внутримышечного введения характеризовалась хорошей переносимостью и была безопасна при двукратном внутримышечном введении. В ходе исследования серьезных нежелательных явлений, связанных с вакцинацией, отмечено не было. Возникновения местных нежелательных явлений у добровольцев, получивших исследуемый препарат и препарат сравнения, зарегистрировано не было. Системных нежелательных явлений, а также изменений инструментальных и лабораторных показателей (по результатам общего и биохимического анализов крови, анализа мочи, ЭКГ), связанных с вакцинацией выявлено не было.
Гуморальный ответ
Результаты
1. Вакцина гриппозная универсальная рекомбинантная лиофилизированная Унифлю для внутримышечного введения характеризовалась хорошей переносимостью и была безопасна при двукратном внутримышечном введении. В ходе исследования серьезных нежелательных явлений, связанных с вакцинацией, отмечено не было. Возникновения местных нежелательных и системных нежелательных явлений, а также изменений инструментальных и лабораторных показателей (по результатам общего и биохимического анализов крови, анализа мочи, ЭКГ), связанных с вакцинацией выявлено не было.
2. Анализ поствакцинального гуморального ответа у добровольцев, получавших вакцину в высокой и низкой дозах, показал наличие дозо-зависимого эффекта. Процент сероконверсий среди добровольцев к М2е антигену, получавших 0,5 дозы вакцины, составил 11,1%, а среди добровольцев, получавших 1 дозу вакцины, возрос до 53,3%. Кроме того, среди добровольцев с сероконверсией отмечается возрастание доли анти-М2е антител подкласса IgG1.
3. Показано формирование Т-клеточного ответа у 6 из 15 добровольцев, получивших двукратно вакцину «Унифлю» в высокой дозе, т.е. у 40% привитых лиц. Для большинства добровольцев, получивших вакцину, показано формирование антиген-специфичных клеток, относящихся к различным клеточным субпопуляциям (центральные и эффекторные CD4+ и CD8+ Т-клетки памяти).
Вакцина «Унифлю» является хорошим кандидатом для применения в качестве универсальной противогриппозной вакцины на основе рекомбинантного белка с точки зрения ее иммуногенности.
Пример 2. Измерение дзета-потенциала образцов вакцин на основе рекомбинантного белка и адьюванта дезоксирибонуклеата натрия
Стандартным образом, описанным в уровне техники и известным специалисту в данной области техники получили образцы вакцин, содержащие высокоочищенный рекомбинантный белок НВс4М2е с консенсной последовательностью М2е вирусов гриппа А человека (A/H1N1, A/H3N2, A/H2N2), адъювант - дезоксирибонуклеат натрия и фосфатно-буферный раствор (ФБР), рН 7,2-7,4.
Белки получены путем культивирования штаммов E. coli DLT1270 pQE60HBc/4Me2h и E.coli DLT1270 pQE60HBc соответственно и очищены с помощью гель-фильтрации и ионообменной хроматографии. В частности, применялась партия концентрированного очищенного раствора белка HBc-4M2eh от 10.01.2019 г., охарактеризованного по следующим параметрам:
концентрация белка - 350 мкг/мл
посторонние белковые примеси - 2%
содержание бактериальных эндотоксинов - 20 ЭЕ на дозу
содержание бактериальных нуклеиновых кислот - 30 пг на дозу
содержание бактериальных белков - 21 нг на дозу
Дезоксирибонуклеат натрия (адъювант) представляет собой натриевую соль ДНК (олигодезоксинуклеотид) эукариотического происхождения, деполимеризованную ультразвуком до фрагментов с молекулярной массой 270-500 кДа, не несущих генетической информации. В ходе исследований применялся препарат Деринат®. Регистрационный номер препарата Р N002916/01 (дата регистрации 27.02.2008). Применяли Деринат® серии 30130218 от 02.18 срок годности до 02.23.
Из концентрата рекомбинантного белка (таблица 22) были подготовлены образцы вакцины, содержащее 20, 40, 60 и 90 мкг рекомбинантного белка и 100, 500, 1000, 5000 и 10000 мкг адъюванта дезоксирибонуклеата натрия.
Таблица 22. Приготовление образцов вакцины с дезоксирибонуклеатом натрия
Поверхностный заряд частиц (дзета-потенциал) оценивали по стандартной методике измерения на приборе прибора Malvern Panalytical Ltd Zetasizer Nano ZSP. Определение дзета-потенциала образцов проводилось по методу электрофоретического рассеивания света. Программное обеспечение оборудования автоматически подбирает оптимальную взаимосвязь опорного сигнала и рассеиваемого частицами излучения. После помещения специальной кюветы в прибор, происходило измерение двух параметров рассеянного излучения: в присутствие электромагнитного поля и в его отсутствие, разность пиковых частот которых дает возможность вычислить величину доплеровского дрейфа. Полученное значение позволяет рассчитать среднюю подвижность частиц и их дзета-потенциал с помощью формулы Эйнштейна-Смолуховского.
Для образцов вакцин измерения дзета-потенциала проводили при температуре 4°C. Результаты представлены в Таблице 23.
Более того, неожиданно обнаруженный диапазон повышения дзета-потенцила был исследован более подробно, при этом образцы были получены описанным выше образом.
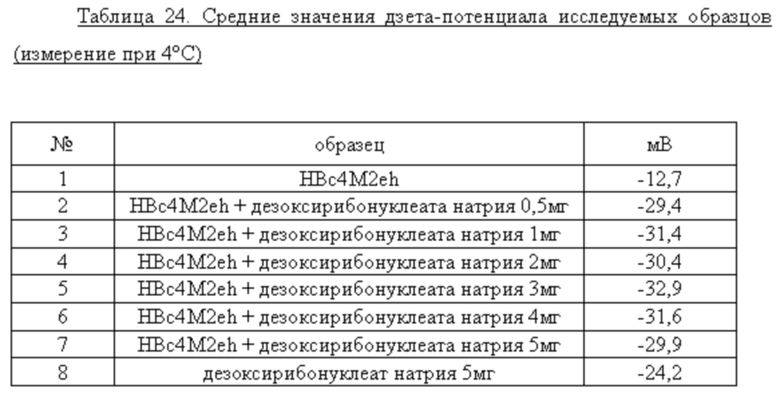
Измерение дзета-потенциала рекомбинантного белка HBc-4M2eh показало, что на поверхностный заряд вирусоподобных частиц оказывает влияние содержание адъюванта дезоксирибонуклеата натрия. Полученные значения дзета-потенциала показывают, что на увеличение дзета потенциала влияет определенный диапазон содержания дезоксирибонуклеата натрия от 1000 до 10000 мкг, для которого наблюдается получение наиболее стабильных вакцин.
Таким образом, авторам настоящего изобретения удалось создать наиболее стабильную универсальную противогрипозную вакцину на основе рекомбинантного белка, содержащую рекомбинантный белок HBc-4M2eh в количестве от 20 до 90 мкг и адьювант дезоксирибонуклеат натрия в количестве 1000 до 10000 мкг, которая может эффективно применяться для вакцинации населения против гриппа.
Пример 3. Исследование иммуногенности и протективного действия вакцин с содержанием HBc/4M2eh в количестве от 20 до 90 мкг и адьюванта дезоксирибонуклеата натрия в количестве от 1000 до 10000 мкг.
Для характеристики иммуногенности и протективного действия был проведен эксперимент по иммунизации мышей очищенными препаратами вирусоподобных частиц HBc/4M2ek частиц. Самки мышей BALB/c весом 16-18г были получены из Питомника лабораторных животных (Пущино, Россия). Иммунизацию проводили внутримышечно на 0, 14 и 28 дни вакцинами с концентрацией рекомбинантного белка 20, 40, 60 и 90 мкг/0,5 мл, содержащими 1000 мкг, 5000 мкг и 10000 мкг адьюванта дезоксирибонуклеата натрия (при проведении исследования применяли препарат Деринат®). Контрольной группе мышей вводили 0.5 мл фосфатно-солевого буфера. Образцы крови забирали из вентральной вены у 5 мышей через 2 недели после второй иммунизации. Через две недели после третьей иммунизации собирали образцы крови и бронхо-альвеолярных лаважей (БАЛ).
Титры М2е-специфичных антител в сыворотках и BAL мышей определяли с помощью иммуно-ферментного анализа (ИФА). Для определения титра анти-М2е антител в ИФА использовали синтетический пептид G-37, соответствующий M2ek. У мышей, иммунизированных препаратом ВПЧ HBc/4M2ek, уже после второй иммунизации развивается сильный иммунный ответ (Таблица 25).
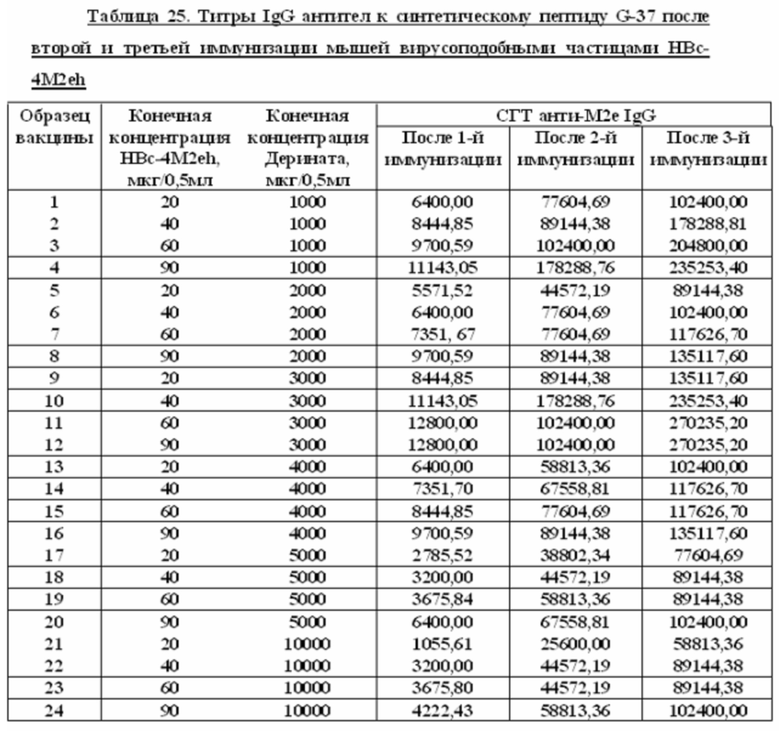
Таким образом, данные результаты подтверждают хорошую иммуногенность вакцины согласно настоящему изобретению.
Протективное действие вакцины согласно настоящему изобретению совпадает с протективным действием вакцины «Унифлю», описанным выше, то есть обеспечивается защита 75-100% иммунизированных животных после заражения вирусами гриппа А разных субтипов (H1N1. H2N2, H3N2, H5N1) высокими летальными дозами (5 LD50), и достигается достоверное снижение репродукции вирусов гриппа в легких.
Таким образом, авторам настоящего изобретения удалось разработать эффективную кросс-реактивную рекомбинантную вакцину против вируса гриппа А человека, обладающую превосходной стабильностью и улучшенной иммуногенностью.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА "СВИНОГО" ГРИППА H1N1 И СПСОБ ЕЕ ПОЛУЧЕНИЯ | 2010 |
|
RU2451027C2 |
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |
| ПРОТИВОГРИППОЗНАЯ ВАКЦИНА ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ПРОТИВ ПТИЧЬЕГО ГРИППА А НА ОСНОВЕ ЭКТОДОМЕНА БЕЛКА М2 | 2014 |
|
RU2571944C1 |
| Антиген для универсальной рекомбинантной вакцины против вируса гриппа птиц и человека | 2021 |
|
RU2839428C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЧУЖЕРОДНЫЕ АНТИГЕНЫ НА ПОВЕРХНОСТИ СФЕРИЧЕСКИХ НОСИТЕЛЕЙ, ПОЛУЧЕННЫХ ПРИ ТЕРМИЧЕСКОЙ ДЕНАТУРАЦИИ СПИРАЛЬНЫХ ВИРУСОВ | 2010 |
|
RU2440140C1 |
| Вакцина против ротавирусной инфекции человека на основе вирусоподобных частиц ротавируса | 2024 |
|
RU2829862C1 |
| ВИРИОНЫ И ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ ВИРУСА МОЗАИКИ АЛЬТЕРНАНТЕРЫ КАК УСИЛИТЕЛИ ИММУННОГО ОТВЕТА | 2015 |
|
RU2639491C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ И СПОСОБЫ | 2008 |
|
RU2468034C2 |
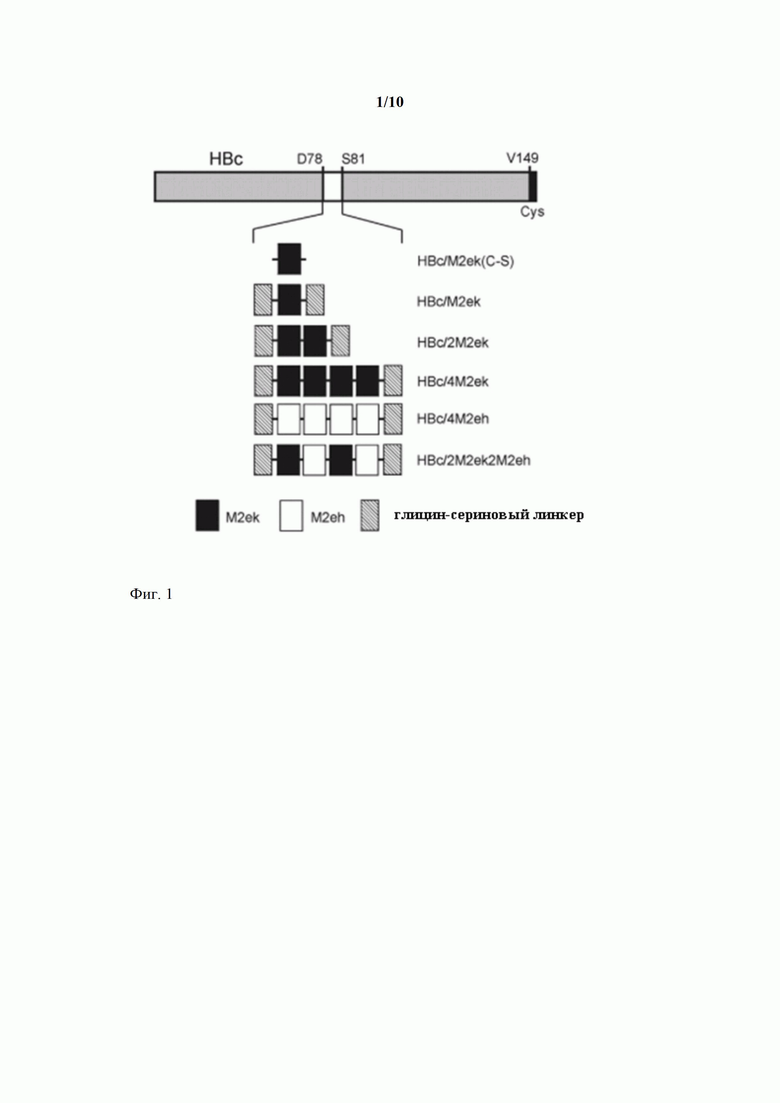
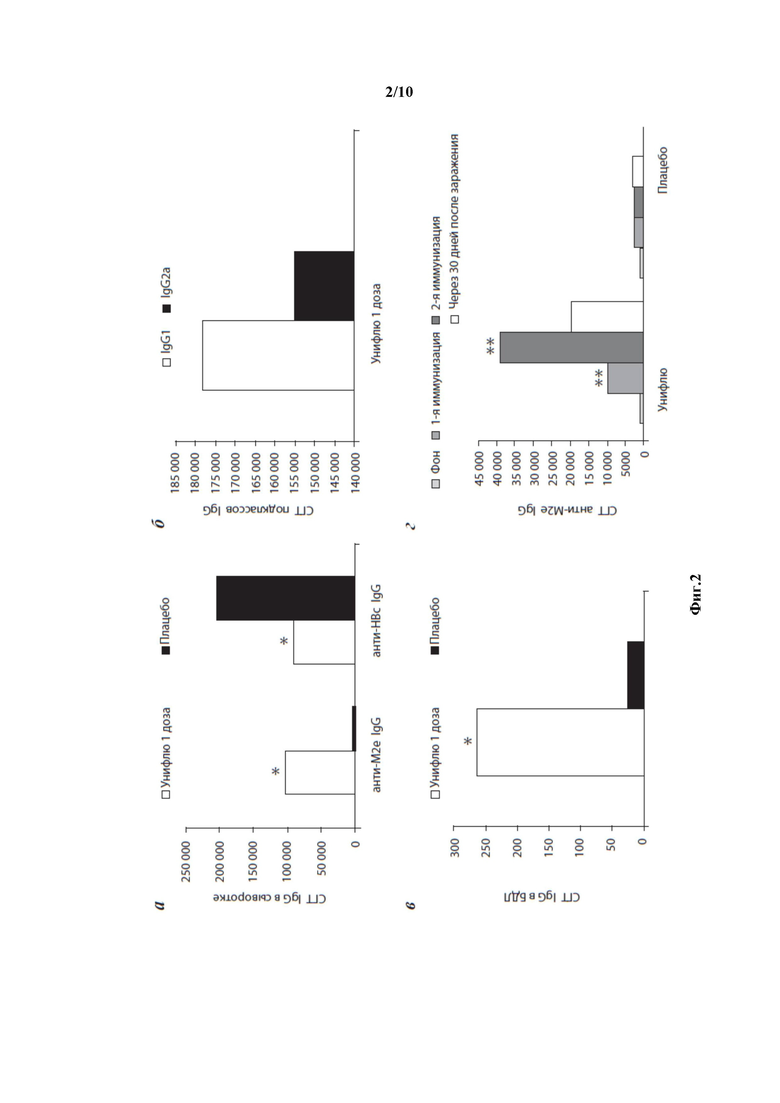








Изобретение относится к иммунологии и белковой инженерии. Описана кросс-реактивная рекомбинантная вакцина против вируса гриппа А человека, содержащая рекомбинантный белок HBc-4M2eh, который имеет аминокислотную последовательность SEQ ID NO 6, в количестве от 20 до 90 мкг и адъювант дезоксирибонуклеат натрия в количестве от 1000 до 10000 мкг, а также фосфатно-буферный раствор (ФБР), рН 7,2-7,4. Изобретение обеспечивает противогриппозную вакцину широкого спектра защиты («универсальную»), эффективную для борьбы с пандемией и эпидемией гриппа. 2 н. и 10 з.п. ф-лы, 25 табл., 3 пр., 5 ил.
1. Кросс-реактивная рекомбинантная вакцина против вируса гриппа А человека, содержащая рекомбинантный белок HBc-4M2eh, который имеет аминокислотную последовательность SEQ ID NO 6, в количестве от 20 до 90 мкг и адъювант дезоксирибонуклеат натрия в количестве от 1000 до 10000 мкг, а также фосфатно-буферный раствор (ФБР), рН 7,2-7,4.
2. Вакцина по п. 1, содержащая 20 мкг рекомбинантного белка HBc-4M2eh.
3. Вакцина по п. 1, содержащая 40 мкг рекомбинантного белка HBc-4M2eh.
4. Вакцина по п. 1, содержащая 60 мкг рекомбинантного белка HBc-4M2eh.
5. Вакцина по п. 1, содержащая 90 мкг рекомбинантного белка HBc-4M2eh.
6. Вакцина по п. 1, содержащая от 1000 до 3000 мкг адъюванта дезоксирибонуклеата натрия.
7. Вакцина по п. 1, содержащая от 1000 до 5000 мкг адъюванта дезоксирибонуклеата натрия.
8. Вакцина по п. 1, содержащая 1000 мкг адъюванта дезоксирибонуклеата натрия.
9. Вакцина по п. 1, содержащая 3000 мкг адъюванта дезоксирибонуклеата натрия.
10. Вакцина по п. 1, содержащая 5000 мкг адъюванта дезоксирибонуклеата натрия.
11. Вакцина по п. 1, содержащая 10000 мкг адъюванта дезоксирибонуклеата натрия.
12. Применение кросс-реактивной рекомбинантной вакцины по п. 1 для профилактики гриппа А человека.
| RU 2018118337 A, 25.11.2019 | |||
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |
| US 20120052082 A1, 01.03.2012 | |||
| US 20120052082 А1, 01.03.2012. | |||

