УРОВЕНЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к иммунологически активным составам, которые могут стимулировать защитный иммунитет или толерантность. Состав, который стимулирует защитный иммунитет как у инфицированного, так и у неинфицированного хозяина, состоит из эпитопов антигенов, но не включает или не содержит эпитопы, которые участвуют в механизме ускользания от иммунологического контроля или стимулируют толерантность. Защитный иммунитет также можно стимулировать с помощью состава, который содержит ассоциированную с патогеном характерную молекулярную структуру (или структуры) и/или носитель с эпитопами антигенов или без них. Другой иммунологически активный состав, который стимулирует толерантность, включает эпитоп(ы) или молекулярную структуру (структуры), важные для ускользания патогена от иммунологического контроля, с носителем или без него. Кроме того, настоящее изобретение обеспечивает способы идентификации таких иммунологически активных молекул.
[0002] Прогресс в иммунобиологии обеспечил идентификацию иммунологических факторов, необходимых для разработки иммуномодуляторов, включая необходимость стимуляции врожденного и приобретенного иммунных ответов, которые подавляют патогены.
[0003] С появлением устойчивости к широкому спектру антибиотиков, применение иммуномодуляторов может быть наиболее эффективным подходом к обеспечению продолжительной защиты от микроорганизмов, включая внутриклеточные патогены. Современный интерес к иммуномодуляторам требует разработки новых векторов, эффективных носителей и систем адъювантов.
[0004] Понимание того, что патогены могут также вызывать ответ Тh2 и толерантность, можно использовать для разработки иммуномодуляторов для аутоиммунных заболеваний, трансплантации и других медицинских применений. Большинство патогенов попадает в организм через кожу и слизистые оболочки. Следовательно, эти пути введения наилучшим образом подходят для иммуномодуляции, направленной против инфекций, попадающих в организм через кожу, воздушные пути, желудочно-кишечный тракт или половые органы. Традиционные вакцины вводят парентерально, вдали от места локализации инфекции, и вызываемый ими ответ в слизистых менее выражен.
[0005] В настоящее время ясно, что помимо TLR-рецеторов (Toll-like receptors, TLR), существуют другие рецепторы и пути, которые играют важную роль в механизмах врожденного иммунного ответа. Примером являются белки, которые содержат домены олигомеризации нуклеотидов (NOD), которые распознают мотивы внутриклеточных микроорганизмов. Миндин, белок внеклеточного матрикса, также является медиатором воспалительного ответа на некоторые компоненты поверхности бактерий. Эти и другие исследования предполагают, что во врожденном иммунитете задействованы дополнительные факторы, не участвующие в передаче сигналов через рецепторы TLR, и что образования ядерного фактора каппа В (NFκB) или интерлейкина IL-1 может быть недостаточно для подавления инфекций.
[0006] Типичными элементами врожденного иммунитета, участвующими в подавлении инфекций, являются: (1) провоспалительный ответ: опосредуемый NFκB, активирует многихвозбудителей воспаления, повышенная стимуляция может привести к шоку; (2) катионные защитные белки хозяина: повышенная продукция пептидов, вызванная молекулярными структурами, ассоциированными с бактериальным патогеном (pathogen associated molecular fiatterns, PAMPs), и сигнальными молекулами; (3) активация фагоцитирующих клеток: повышенный внутриклеточный цитолиз в нейтрофилах и макрофагах (усиливаются как окислительный, так и не окислительный механизмы), повышенная продукция цитокинов; (4) хемтотаксис: повышенная адгезия фагоцитирующих клеток к эндотелию, миграция клеток к участку локализации инфекции, диапедез; (5) механизм внеклеточного цитолиза: активация комплемента, усиленное хелатирование железа, секреция противомикробных пептидов, продукция расщепляющих ферментов; (6) ограничение распространения инфекции: образование тромба через активацию фибриногена; (7) заживление раны: рост и адгезия фибробластов, ангиогенез; и (8) приобретенные иммунные ответы: активация В- и Т-клеток, часто опосредуемая дендритными клетками.
[0007] Стимуляции врожденного иммунитета можно достичь путем применения интерферонов, монофосфорил-липида А, имиквимода, нуклеотидов CpG или катионных пептидов. Однако врожденный иммунитет имеет лишь ограниченную возможность в предохранении от инфекций, и при таком сценарии основную роль играет приобретенный иммунный ответ.
[0008] Недавно обнаружили, что дендритные клетки необходимы для взаимодействия систем врожденного и приобретенного иммунитета, и это знание позволило иммунологам разработать стратегии иммуномодуляции против антигенов со слабой иммуногенностью. Дендритные клетки (ДК) образуются из предшественников миелоидной и лимфоидной линий, но являются основными антиген-презентирующими клетками (АПК). ДК присутствуют во всех тканях, и во время инфекции они являются ключевыми клетками иммунной системы, которые вступают в контакт с внедряющимся патогеном. Они представляют собой мостик между врожденными и приобретенными иммунными ответами.
[0009] Клетки эндотелия и эпителия, моноциты, макрофаги и другие клетки, включая незрелые ДК, экспрессируют рецепторы, распознающие характерные структуры патогенов (TLR-рецепторы, рецепторы с доменом лектина и другие рецепторы), которые связывают ассоциированные с патогенами консервативные молекулярные структуры (сокращенно РАМР), общие для патогенов, такие как липополисахарид из грамотрицательных бактерий, липотейхоевая кислота из грамположительных бактерий, пептидогликан, липопротеины ассоциированные с пептидогликаном, бактериальная ДНК и флагеллин из грамотрицательных и грамположительных бактерий, а также вирусная РНК. Различные клетки экспрессируют различные рецепторы, обеспечивая настройку ответа на патоген. После активации, незрелые антиген-захватывающие ДК дифференцируются в зрелые антиген-презентирующие ДК, способные презентировать антиген в составе основного комплекса гистосовместимости МНС II и I класса, а также повышать экспрессию костимулирующих молекул клеточной поверхности, таких как CD80 и CD86.
[0010] Зрелые и активированные ДК мигрируют во вторичные лимфоидные органы (лимфатические узлы, селезенку, Пейеровы бляшки), где они перемещаются в области скопления Т-клеток. Взаимодействие ДК с Т-клетками и стимуляция ими Т-клеток зависит от цитокинов, хемокинов и молекул адгезии, таких как внутриклеточные молекулы адгезии клеток (intracellular cell adhesion molecule, 1-CAM), функциональный лимфоцитарный антиген 1 типа (leukocyte function associated molecule, LFA-1) и специфичный для дендритных клеток I-CAM-связывающий неинтегринный рецептор DC-SIGN.
[0011] В зависимости от локального цитокинового окружения и самого антигена, запуск клеточного ответа Т-хелперов (Th1) и гуморального опосредуемого антителами, Th2- или Treg-ответа происходит в различной степени. Было показано, что доза антигена направляет дифференцировку Th1/Th2 клеток, при этом высокие дозы стимулируют в основном Th-1 ответ, а низкие - Th-2 ответ. Было показано, что искусственные носители, «представляющие» антигенные белки, и ДНК-вакцины, поглощаются незрелыми дендритными клетками и вызывают иммунный ответ. Следовательно, ДК представляют собой основную, но не единственную мишень для разработок средств модуляции иммунной системы.
[0012] В частности, ДК слизистых обеспечивают важную первичную защиту, поглощая внедряющиеся чужеродные агенты путем пиноцитоза и рецептор-опосредованного эндоцитоза. ДК играют решающую роль в иммунных свойствах слизистых оболочек, а слизистые оболочки тела действуют как барьер между внутренней средой организма и его окружением. ДК могут быть обнаружены в выстилке дыхательных путей и кишечника. Клетки Лангерганса представляют собой популяцию ДК, обнаруживаемых в коже и слизистой оболочке. ДК и М-клетки транспортируют антигены в лимфоидный фолликул, находящийся в толще слизистой оболочки, который представляет собой иммуноиндуцирующую область кишечника. В дыхательных путях были описаны аналогичные лимфоидные ткани, ассоциированные с носовой полостью и бронхами. В желудочно-кишечном тракте эта система играет важную роль, но в слизистой оболочке дыхательных путей сеть ДК может быть даже более важной.
В случае перорального введения иммуномодулятор должен пройти через желудок и верхние участки кишечного тракта неразрушенным. Такое разрушение маловероятно при назальном введении, введении через глаз или гениталии. Затем иммуномодулятор должен всосаться через эпителий кишечника, чтобы антиген-презентирующие клетки могли адсорбировать его и впоследствии представить иммунокомпетентным клеткам. Иммунокомпетентные клетки расположены в эпителии, в собственной пластинке слизистой оболочки (lamina propria) или под базальной мембраной. Следовательно, компоненты иммуномодулятора необходимо назначать совеместно с носителем, которой обеспечит их проникновение через указанный барьер. Считается, что когда молекулы связаны с носителями, представляющими собой частицы, они могут быть перенесены через указанный барьер М-клетками пейеровых бляшек.
[0013] Преждевременная деградация или высвобождение биоактивных молекул препятствовали разработке вакцин и способов доставки лекарственных средств на основе частиц. Это является возможным объяснением того, что согласно опубликованной литературе, для достижения эффекта, сравнимого с эффектом аналога, вводимого путем инъекции, необходима высокая доза антигена/лекарственного средства. Помимо плохой утилизации антигенов и лекарственных средств, в качестве основных недостатков указывают низкую способность М-клеток в пейровых бляшках (ПБ) транспортировать частицы и недостаточную стимуляцию иммунного и других ответов у человека. Известно, что М-клетки пейровых бляшек эпителия обеспечивают транспорт определенных бактерий, вирусов и простейших из кишечника. Некоторые исследования показали, что поглощение объекта, осуществляемое М-клетками, ДК и клетками Сасо-2, зависит от его размера, и максимальный диаметр поглощаемого объекта равен 10 мкм.
[0014] Новая информация по поглощению частиц М-клетками и различными типами дендритных клеток (ДК), присутствующих в ПБ и их окрестности, может обеспечить понимание задействованных в этом процессе механизмов. Веyеr с соавторами (Веyеr Т., et al; Bacterial carriers and virus-like-particles as antigen delivery devices: Role of dendritic cells in antigen presentation (Бактериальные носители и вирусоподобные частицы как средства доставки антигенов: Роль дендритных клеток в презентации антигенов). Curr. Drug Targets-Infect. Disord, 2001 1, 287-302) исследовали поглощение и кинетику клеток пекарских дрожжей (Saccharomyces cerevisiae) в ПБ, предполагая, что они представляют собой инертную модель транспорта через слизистую оболочку. Для распределения клеток дрожжей в М-клетках, межклеточном углублении под М-клетками и пространстве под базальной мембраной была обнаружена типичная временная зависимость, сходная с транспортом в различные типы фагоцитирующих, антигенпроцессирующих макрофагов или дендритных клеток. Обнаружили, что в зависимости от своего расположения, ДК имеют разные функции в микроокружении ПБ, продуцируя после активации Тh1- или Тh2-направляющие цитокины. Затем хемокиновое и цитокиновое микроокружение определит дифференцировку клеток-хелперов в субпопуляции Тh1 и Тh2 соответственно, а также влияет на выживание или апоптоз Т-клеток.
[0015] Кроме того, разные цитокины (IL-6 в дополнение к TGF-p, IL-4, IL-5 и IL-10) и специфический рецептор хоуминга a4b7, регулируют дифференцировку В-клеток и хоуминг плазматических клеток слизистой оболочки. Соответственно, можно заключить, что очевидно в слизистой оболочке существуют механизмы, через которые модуляция иммунной системы слизистой оболочкой может вызвать более дифференцированный иммунный ответ, лучше имитирующий ответ на природную инфекцию, чем полученный при применении других путей введения состава.
[0016] Несмотря на то, что многое известно о ДК, их предполагаемых предшественниках и субпопуляциях ДК, высокая степень функциональной сложности и пластичности ДК затрудняет предсказание действия конкретной вакцины на ДК и последующие Тh1, Тh2 и Treg ответы. Однако некоторые результаты, полученные для дифференцированных in vitro ДК, можно экстраполировать на зрелые ДК, выделенные из лимфоидных органов, поскольку они проявляют схожие свойства (Shortman K., et al; Mouse and human dentritic cell subtypes (Подтипы дендритных клеток мыши и человека). Nat. Rev. Immunol. 2002 Mar 2(3): 151-61). Например, in vitro была изучена способность липопептида микоплазмы MALP-2 модулировать ответ ДК (Weigt H., et al; Synthetic mycoplasma-derived lipopeptide MALP-2 induces maturation and function of dendritic cells (Синтетический липопептид MALP-2 микоплазменного происхождения стимулирует созревание и функцию дендритных клеток) Immunobiology 2003, 207(3): 223-33). Обработка дендритных клеток MALP-2 стимулировала экспрессию CD80, CD86 и высвобождение биоактивных TNF-α и IL-10, а также пролиферацию аутологичных лимфоцитов и продукцию последними IL-4, IL-5 и γ-интерферона. Эти признаки коррелируют со способностью стимулировать Т-клетки, и таким образом указывают на возможное действие MALP-2 на ДК in vivo.
[0017] Синтетические носители могут обеспечивать иммуностимулирующее действие антигенов аналогичное действию при презентации на молекулах комплекса гистосовместимости (МНС) класса I и II. На основе синтетических носителей можно разработать гибкую систему, которую можно адаптировать к разнообразным возможным приложениям. Характер этих носителей может существенно влиять на результат и эффективность иммунного ответа. Синтетические носители, такие как частицы, могут облегчить соответствие стандартам качества и сертификацию при разработке и производстве вакцин, и, соответственно, сократить время, необходимое для получения одобрения и выхода на рынок.
[0018] Для вакцинации можно применять различные стимулирующие иммунный ответ компоненты (пептиды, белки, липиды или полисахариды) различных инфекционных микроорганизмов. Эти компоненты могут быть синтезированы, выделены из микроорганизмов или получены по технологии рекомбинантных ДНК. Однако при их введении в свободной, растворимой форме перорально или парентеральнотребуются подходящие адъюванты.
[0019] В качестве носителей для различных антигенов и лекарственных средств исследовали несколько систем на основе частиц. Ранее в качестве систем-носителей лекарственных средств были описаны микрочастицы из хитозана, поли-DL-молочной кислоты или полиакрилового крахмала. Примеры таких систем описаны в Патентах США 5,603,960 и 6,521,431. В одном из источников сообщали, что микрочастицы из крахмала с ковалентно присоединенным сывороточным альбумином человека (HSA), который представлял собой модельный антиген, действовали как сильный адъювант при парентеральном введении мышам, а взятые отдельно микрочаскицы были неиммуногенны.
[0020] Необходимо отметить, что поглощение вероятнее всего зависит от структуры и также от возможных адгезивных свойств носителя. Агарозе и другим полисахаридам присущи мукоадгезивные свойства, которые могут улучшить их взаимодействие с различными слизистыми оболочками и облегчить поглощение.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0021] Согласно настоящему изобретению, описаны новые композиции для иммунологически активного состава или составов, которые могут стимулировать защитный иммунитет или толерантность. Состав для стимуляции защитного иммунитета у инфицированного или неинфицированного хозяина состоит из эпитопов антигенов, но в него не включены или из него удалены эпитопы, которые участвуют в «ускользании» от иммунной системы или стимулируют толерантность. Защитный иммунитет также можно стимулировать составом, который всодержит ассоциированную с патогеном молекулярную структуру/структуры и/или носитель с антигенными эпитопами или без них. Другой иммунологически активный состав, который стимулирует толерантность, содержит эпитоп(ы) ускользания или молекулярную структуру/структуры, важные для ускользания антигена, с носителем или без него. Кроме того, настоящее изобретение обеспечивает способы идентификации таких иммунологически активных молекул. Подходящим примером являются молекулы, распознающие ассоциированную с патогеном молекулярную структуру/структуры (PAMPs) с присоединенными модифицированными антигенами микоплазмы или без них. Как оказалось, такие молекулярные составы модулируют иммунные ответы против микоплазмы как у инфицированных, так и у неинфицированных хозяев. Некоторые из антигенов стимулировали толерантность или ускользание от иммунологического контроля.
[0022] Соответственно, в одном своем аспекте настоящее изобретение относится к иммунологически активному составу для стимуляции защитного иммунитета, который содержит:
(1) по меньшей мере одну ассоциированную с патогеном молекулярную структуру;
(2) возможно содержит по меньшей мере один иммунологически активный антиген или эпитоп антигена; и
(3) по меньшей мере один носитель, эффективно доставляющий состав в организм, что приводит к стимуляции защитного иммунитета.
[0023] Во многих случаях желательно включить по меньшей мере один активный антиген или эпитоп антигена. Подходящие примеры описаны ниже. Обычно указанный по меньшей мере один иммуннологически активный эпитоп антигена очищают от эпитопов ускользания. Это важно для предотвращения селективного отбора, который в противном случае привел бы к появлению патогенов, репликацию которых не блокирует не иммунный ответ. Примером патогенов, для которых важен селективный отбор, является вирус гриппа, который так быстро мутирует, что для каждого инфекционного сезона необходимо изготавливать новую вакцину, чтобы обеспечить иммунитет к конкретному штамму или штаммам гриппа, которые могут вызвать заболевание человека. Другой пример патогена, для которого, видимо, важенселективный отбора, - вирус иммунодефицита человека (ВИЧ), который также быстро мутирует.
[0024] В другом своем аспекте настоящее изобретение относится к иммунологически активному составу для стимуляции толерантности, который содержит:
(a) по меньшей мере одну ассоциированную с патогеном молекулярную структуру;
(b) по меньшей мере один иммунологически активный антиген или эпитоп антигена; и
(c) по меньшей мере один носитель, эффективно доставляющий состав в организм, что приводит к стимуляции толерантности.
[0025] Обычно иммунологически активный эпитоп антигена представляет собой пептид, белок, рекомбинантный пептид или мультипептид, рекомбинантный белок, липид, углевод, нуклеиновую кислоту или другую биологически активную молекулу, либо комбинацию любых из указанных молекул. Обычно, если иммунологически активный эпитоп антигена представляет собой пептид или белок, указанный пептид или белок имеет иммуномодулирующие посттранскрипционные модификации. Обычно посттранскрипционные модификации содержат фрагменты углеводов и/или липидов. Обычно посттранскрипционные модификации включают терминальное маннозилирование; в этом случае терминально маннозилированные иммуномодулирующие вещества обычно удаляют из иммунозащитного состава. Их можно удалить путем окисления, обработки ферментами или путем сахар-специфичного аффинного связывания. Альтернативно, если посттранскрипционные модификации включают фрагменты липидов, эти фрагменты липидов удаляют путем делипидизации.
[0026] Обычно иммунологически активные эпитопы антигенов представляют собой пептид или белок, причем указанный иммунологически активный пептид или белок не содержит иммуномодулирующих посттранскрипционных модификаций. Также иммунологически активный пептид или белок обычно не содержит последовательностей аминокислот, по которым может происходить N-гликозилирование и/или присоединение липида. В другом предпочтительном варианте иммунологически активный эпитоп антигена представляет собой пептид или белок, и указанный иммунологически активный пептид или белок содержит последовательность аминокислот, способную связываться с гликозаминогликанами (GAG) поверхности клетки. Обычно такие последовательности аминокислот имеют многоосновную природу и имеют общую формулу ХВВХВХ, ХВВВХХВХ, ВВХХВВВХХВВ, ВВВХХВ, ВХВХВ, ВВВ, ВХВХХХВХВ, или ВХВХХХХХВХВ, где В обозначает основную аминокислоту, а Х - любую другую аминокислоту. Обычно GAG-связывающие последовательности аминокислот применяют для получения антител к пептиду или белку, которые способны препятствовать связыванию патогена с поверхностью клетки. GAG может быть выбран из группы, состоящей из гепарина и его аналогов. Иммунологически активный пептид или белок может обладать комплементактивирующей активностью самостоятельно или в комбинации с антителами. Иммунологически активный эпитоп антигена может представлять собой множество пептидов, которые соединены в единый мультипептид. Иммунологически активные эпитопы антигенов могут включать как эпитопы Т-клеток, так и эпитопы В-клеток.
[0027] Обычно ассоциированную с патогеном молекулярную структуру выбирают из группы, включающей:
(1) агониста рецептора TLR 1;
(2) агониста рецептора TLR 2;
(3) агониста рецептора TLR 3;
(4) агониста рецептора TLR 4;
(5) агониста рецептора TLR 5;
(6) агониста рецептора TLR 6;
(7) агониста рецептора TLR 7;
(8) агониста рецептора TLR 8;
(9) агониста рецептора, TLR 9;
(10) агониста NOD-1;
(11) агониста NOD-2;
(12) DC-SIGN;
(13) L-SIGN; и
(14) рецептора маннозы.
[0028] Если ассоциированная с патогеном молекулярная структура представляет собой агонист NOD-1 или агонист NOD-2, такой агонист NOD-1 или агонист NOD-2 может быть выбран из группы, включающей бактериальный пептидогликан и производное бактериального пептидогликана.
[0029] В другом своем аспекте настоящее изобретение относится к способу идентификации иммунологически активных пептидов, способных препятствовать связыванию патогенов с гликозаминогликанами, включающему следующие этапы:
(1) проведение адсорбции гепарина;
(2) проведение иммуноаффинной селекции; и
(3) возможно, выполнение протеолитического расщепления белка или белков, выделенных путем иммуноаффинной селекции, для получения иммунологически активных пептидов. Иммуноаффинную селекцию можно осуществить способами, хорошо известными в данной области техники и описанными, например, у G.T. Hermanson et al., "Immobilized Affinity Ligand Techniques" («Способы с применением иммобилизованных аффинных лигандов» Academic Press, Inc., San Diego, 1992); в данной области известны также другие способы.
[0030] Еще один аспект настоящего изобретения относится к способу идентификации иммунологически активных пептидов, способных препятствовать связыванию патогенов с гликозаминогликанами, который включает стадию анализа информации о последовательности способом биоинформатики с использованием многоосновных линейных мотивов.
[0031] Еще один аспект настоящего изобретения относится к способу идентификации комплементактивирующих иммунологически активных пептидов, который включает следующие этапы:
(1) осуществление связывания комплементфиксирующих антител с белком комплемента;
(2) применение указанных антител для иммуноаффинной селекции белковых антигенов; и
(3) возможно, выполнение протеолитического расщепления выделенных белковых антигенов.
[0032] Полученные этими способами иммунологически активные пептиды также представляют собой аспекты настоящего изобретения. Кроме того, на основе идентифицированного иммунологически активного пептида можно создать защитное антитело для борьбы с заболеванием в организме хозяина.
[0033] В описанных выше составах молекулы могут присутствовать в смеси. В качестве альтернативы, молекулы могут быть химически связаны друг с другом. Носитель обычно представляет собой микрочастицу. Предпочтительно, микрочастицы имеют узкий диапазон распределения по размерам и являются пористыми. Обычно диаметр микрочастиц меньше 10 мкм, более предпочтительно диаметр микрочастиц меньше 5 мкм. Обычно частицы изготавливают из биополимера. В одном варианте иммунологически активные эпитопы антигенов нековалентно присоединены к микрочастицам. В другом варианте иммунологически активные эпитопы антигенов ковалентно присоединены к указанным микрочастицам. К микрочастицам можно присоединить более одного иммунологически активного эпитопа антигена и более одного агониста патогенраспознающего рецептора. К микрочастицам можно присоединить более одного патогенраспознающего рецептора.
[0034] В другом своем аспекте настоящее изобретение относится к способу индукции иммунного ответа у пациента, который включает этап введения иммунологически эффективной дозы состава, содержащего по меньшей мере один иммунологически активный эпитоп антигена и по меньшей мере один агонист патогенраспознающего (PR) рецептора, связанные с микрочастицами, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше. Состав может содержать более одного агониста патогенраспознающего рецептора.
[0035] Еще в одном своем аспекте настоящее изобретение относится к способу in vivo доставки иммунологически активного состава для индукции иммунного ответа у пациента, включающему этап введения иммунологически эффективного количества состава, содержащего по меньшей мере один агонист патогенраспознающего (PR) рецептора, связанный с микрочастицами, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
[0036] Еще в одном своем аспекте настоящее изобретение относится к способу in vivo доставки иммунологически активного состава для индукции иммунного ответа у пациента, включающему этап введения иммунологически эффективного количества состава, содержащего по меньшей мере один иммунологически активный эпитоп антигена и по меньшей мере один агонист патогенраспознающего (PR) рецептора, связанные с микрочастицами, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
[0037] В другом своем аспекте настоящее изобретение относится к способу индукции защитного иммунного ответа на по меньшей мере один патоген, включающему этап введения, в виде одной или нескольких доз, иммунологически эффективного количества состава, содержащего один или более иммунологически активный эпитоп антигена и комбинацию агонистов патогенраспознающих рецепторов, связанные с микрочастицами, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше, а иммунный ответ включает Тh1 или Th2 ответы, либо их комбинацию.
[0038] Еще в одном своем аспекте настоящее изобретение относится к способу индукции защитного иммунного ответа на по меньшей мере один патоген, включающему этап введения, в виде одной или нескольких доз, иммунологически эффективного количества состава, содержащего один или более иммунологически активный эпитоп антигена, но не содержащего антигены, которые участвуют в механизмах ускользания от иммунологического контроля, а также комбинацию агонистов патогенраспознающих рецепторов, связанные с микрочастицами, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
[0039] В способах введения или доставки согласно настоящему изобретению, введение состава можно осуществлять через слизистую оболочку, парентеральным путем, через кожу. В качестве альтернативы можно применять другие пути введения.
[0040] В способах индукции иммунитета согласно настоящему изобретению иммунный ответ обычно направлен на элементы патогенного микроорганизма, которые связывают гликозаминогликаны.
[0041] В качестве альтернативы, составы согласно настоящему изобретению можно применять в способах индукции иммунологической толерантности. Способ индукции толерантности к иммунологически активному агенту включает этап введения иммунологически эффективного количества состава, содержащего один или более иммунологически активный эпитоп антигена и комбинацию агонистов PRR, связанные с микрочастицами, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше, а иммунный ответ включает стимуляцию регуляторного ответа, или снижение иммунных функций, или ускользание от иммуннологического контроля. Состав может содержать иммунологически активный эпитоп антигена, имеющий липидсодержащий фрагмент. Иммунологически активный липидсодержащий фрагмент может быть присоединен к микрочастицам независимо от иммунологически активного эпитопа антигена. Иммунологически активный липидсодержащий фрагмент может иметь углеводные компоненты, углеводные компоненты могут являться результатом N-гликозилирования. Иммунологически активный эпитоп антигена может содержать мотивы, включающие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из аспарагина, треонина и серина, при этом указанные мотивы представляют собой Asp-X-Ser или Asp-X-Thr мотивы, где Х может быть любой аминокислотой, кроме пролина.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[0042] Настоящее изобретение будет более понятным, если обратиться к описанию, формуле и прилагающимся рисункам, где:
Фигура 1 представляет собой график, демонстрирующий распределение агарозных микрочастиц, подходящих для применения в составах, которые имеют микрочастицы в качестве носителя, согласно настоящему изобретению (Пример 1);
Фигура 2 представляет собой график, демонстрирующий секрецию TNF-α (фактор некроза опухолей альфа), мононуклеарами периферической крови (peripheral blood mononuclear cells, PBMC) - (А) или секрецию тканевого фактора - (В) после обработки пептидогликаном (PepG) (белый столбик) или бактериальной CpG-ДНК (bactDNA) (черный столбик) или обоими веществами одновременно в тех же концентрациях, в которых их вводили отдельно (заштрихованный столбик); числа после Р и D, соответственно, указывают концентрации пептидогликана и бактериальной ДНК в мкг/мл, соответственно; слева направо это P3+D15, P3+D5, P3+D1.5, P1+D15, P1+D5, P1+D1.5, P0.3+D15, P0.3+D5 и P0.3+D1.5;
Фигура 3 представляет собой график, демонстрирующий стимуляцию различных Th1 и Тh2 ответов частицами, приготовленными различными способами с очищенными Con А или обработанными перйодатом натрия антигенами, а также с различными комбинациям агонистов TLR2, TLR3, TLR4, и TLR9 (серый столбик, TNF-α×100, черный столбик, IL-10);
Фигура 4 представляет собой график, демонстрирующий соотношение TNF-α/IL-10 для моноцитов, обработанных частицами, приготовленными различными способами, как для Фигуры 3; и
Фигура 5 представляет собой график, демонстрирующий стимуляцию MHC-II и CD86 в дендритных клетках после воздействия на них препаратов микрочастиц.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНОГО СПОСОБА РЕАЛИЗАЦИИ
[0043] Настоящее изобретение описывает иммунологически активные составы, способы нацеливания молекул на определенные популяции клеток и стимуляции ответов в организме модельных животных, а также способы получения предложенных составов.
[0044] Существует потребность в создании более эффективных иммуномодулирующих агентов и систем доставки препаратов для ряда заболеваний или патологических состояний, а также для защиты от патогенов, вакцин против которых в настоящее время не существует или они не эффективны.
[0045] Традиционные вакцины, содержащие ослабленные, убитые или генетически модифицированные патогены, являются ограниченными для иммунного ответа, который определяют молекулярная структура патогена и его взаимодействие с патогенраспознающими рецепторами (PRR) иммунной системы. Патогены, которые способны вызывать хронические инфекции, имеют молекулярные структуры, которые позволяют им избегать иммунного ответа организма, в то время как другие патогены, такие как вирус гриппа, используют для самоподдержания мутации (антигенную изменчивость), приводящие к ускользанию от иммунологического контроля благодаря продукции антител, способствующих усилению поглощения вируса.
[0046] В целом, патоген-специфические антитела играют важную роль в борьбе с инфекциями путем ряда способов. Однако в некоторых случаях присутствие специфических антител может быть выгодно для патогена. Такая активность известна как антителозависимое усиление (АЗУ) инфекции. АЗУ инфекции - это явление, при котором патоген-специфические антитела усиливают вторжение патогена, а в некоторых случаях и репликацию патогена, например, вируса, в моноциты/макрофаги и в гранулоциты посредством взаимодействия с Fc рецепторами. Этот феномен описан in vitro и in vivo для патогенов, представляющих разнообразные семейства и роды, имеющие значение для здравоохранения и ветеринарии. Эти патогены, такие как М. gallisepticum имеют некоторые общие свойства, такие как предпочтительная репликация в макрофагах, способность к перисистированию и антигенному разнообразию.
[0047] Для некоторых патогенов АЗУ инфекции стало большой проблемой, мешающей борьбе с заболеванием путем вакцинации. Соответственно, было предпринято множество попыток разработки вакцин с минимальным риском АЗУ или вообще без него. Для этого важно идентифицировать или нейтрализовать эпитопы патогенов, связанные с АЗУ. Кроме того, ясное понимание событий, происходящих в клетке после попадания в нее патогена по механизму АЗУ стало критическим для разработки эффективного вмешательства. Однако понимание механизмов АЗУ все еще далеко от полного. Это обуславливает сложность разработки эффективных вакцин против таких патогенов. Поэтому мы идентифицировали мотивы, играющие важную роль в защитном иммунитете путем нарушения механизмов связывания с клеткой, а также мотивы, которые необходимо исключить из состава, чтобы вызвать защитный иммунитет без эффекта ускользания. Мы также идентифицировали мотивы, которые вызывали ускользание или толерантность.
[0048] Мы исследовали возможность нацеливания иммунологически активных молекул на определенные клетки при помощи носителя, состоящего из полисахарида, такого как нативная агароза, которое позволяет достичь модуляции как врожденного, так и приобретенного иммунного ответа и защиты от инфекции. Настоящее изобретение очевидно можно применять также в других приложениях, в которых нацеливание происходит на другие рецепторы при помощи других частиц. Преимуществом агарозы является то, что она представляет собой природный полисахарид, полимер D-галактозы, который подвержен биодеградации и, как было доказано, совместим с клетками млекопитающих. Было обнаружено, что микрочастицы агарозы при парентеральном введении демонстрируют слабую способность активировать макрофаги и сравнимые с гидроксидом алюминия свойства адъюванта (Gronlund H. et al., Carbohydrate-based particles: a new adjuvant for allergen-specific immunotherapy (Частицы на основе углеводов: новый адъювант для аллерген-специфичной иммунотерапии). Immunology, 2002, 107, 523-529).
[0049] С точки зрения конечного пользователя, важно, чтобы вакцина не требовала хранения в холодильнике и в то же время обладала длительным сроком хранения. Агарозные частицы способны удовлетворить этим требованиям. Также важно, чтобы введение вакцины или системы доставки лекарственного средства было максимально простым. Поэтому не требующие инъекций, вводимые через слизистую оболочку, в частности, перорально, составы имеют преимущество перед составами, водимыми парентерально. Пероральное применение, однако, столкнулось с проблемами стабильности состава, являющимися следствием работы пищеварительной системы.
[0050] Мы обнаружили, что присоединение антигенов к пористой агарозной матрице может защитить их от разрушения внутри желудочно-кишечного тракта. Связь между лигандами и частицами гарантирует, что адъювант и антиген будут поглощены одними и теми же антиген-процессирующими клетками. Размер агарозных микрочастиц (<5 мкм) также может обеспечить попадание частиц в Пейеровы бляшки (ПБ).
[0051] Эффективность неметилированной CpG-ДНК, поли(I:С) или MALP-2 в качестве адъювантов позволяет предположить, что мотивы ассоциированных с патогеном молекулярных структур (РАМР), при совместной иммобилизации с антигенами, орбеспечили бы специфичное поглощение иммунологически активных составов на поверхности слизистых различными АПК, экспрессирующими соответствующие рецепторы РАМР.
[0052] Различные субпопуляции ДК и других иммунокомпетентных клеток имеют различные «структурораспознающие рецепторы». Применение комбинаций лигандов позволяет достичь нацеливания на соответствующие субпопуляции клеток.
[0053] Соответственно, мы предположили, что молекулы-агонисты рецепторов (рецептор РАМР), такие как агонисты Toll-подобных рецепторов (TLR), рецепторов пектинов или рецепторов NOD, должны быть иммобилизованы на носителе совместно с биоактивными молекулами. Мы обнаружили, что характеристики присоединения, нацеливание и поглощение иммунологически активных составов можно улучшить, если подвергнуть иммунную систему действию искусственного «синтетического микроорганизма», что может значительно усилить направленное поглощение и эффективность таких иммуномодуляторов и обеспечить эффективный иммунный ответ. Специфичное нацеливание может также улучшить фармакокинетику и снизить возможные побочные эффекты разработанных таким образом искусственных иммуномодуляторов.
[0054] Toll-подобные рецепторы (TLR) и рецепторы NOD, -пектинов и т.д. являются рецепторами, распознающими структуры патогенов, у происходящих из микроорганизмов молекул. Они являются первичными чувствительными элементами систем врожденного и приобретенного иммунитета. В настоящее время идентифицировано 10 рецепторов TLR (TLR 1-10). Каждый из них распознает один или более специфических лигандов и осуществляет передачу сигналов. Периодически обнаруживают, что вновь открытые рецепторы и взаимодействия с участием рецепторов участвуют в активации клеток бактериальными продуктами. Накапливаются свидетельства того, что совместное действие этих рецепторов полезно для совершенствования распознавания лигандов и специфичности ответа. Также обнаружили, что после связывания лиганда происходит кластеризация рецепторов в липидных рафтах. Эти исследования также выявили пути, зависимые и независимые от гена первичного ответа миелоидной дифференцировки 88 (MyD88).
[0055] Каждый TLR - это трансмембранный белок I типа, обладающий внеклеточным доменом, богатым лейцином, и внутриклеточной частью, которая содержит консервативные участки, называемые доменом Toll/IL-IR (область гомологии с цитоплазматическим доменом Toll-белка дрозофилы и рецептором интерлейкина-1 млекопитающих, TIR), активация которого приводит к епривлечению белка MyD88 при МуD88-зависимых формах передачи сигнала. Некоторые микробные патогены могут также поступать в клетку путем эндоцитоза и начинать действовать непосредственно в цитоплазме, например, используя богатые лейцином домены TLR, NOD-1 и NOD-2, как это происходит в случае внутриклеточного распознавания пептидогликана грамположительных бактерий. Однако похоже, что все эти разнообразные пути сводятся транслокации в ядро ядерного фактора NF-κВ и активации генов воспаления и продукции различных цитокинов. Связывание рецепторов TLR 7, 8 и 9 вызывает продукцию IFN-α и IL-12p70, вызывая сильный Th1 ответ с кросс-презентацией/ цитотоксическими лимфоцитами (ЦТЛ). Активация TLR 3 вызывает продукцию IFN-α и Th1 ответ, а TLR5 индуцирует Th1 ответ через IL-12p70, а TLR4 индуцирует Th1 ответ через продукцию IL-12p70 и IFN-α, причем все эти процессы приводят к кросс-презентации ЦТЛ. Однако связывание рецепторов TLR 1, 2 и 6 стимулирует слабый ответ IL-12p70 с высокими уровнями IL-10 и вместе с некоторыми другими PRR (патогенраспознающий рецептор), такими как DC-SIGN, вызывает ответы Th0/Th2/TReg.
[0056] Наиболее тщательно из этого семейства рецепторов изучен TLR-4. Известно, что он распознает липополисахарид из грамотрицательных бактерий и липотейхоевую кислоту из грамположительных бактерий, в то время как TLR-2 связывает бактериальные липопротеины/липопептиды, компоненты микобактерий и микоплазм.
[0057] Мутация Lps2 выявила роль адаптерного белка TIR устойчивости (TIRAP) в TLR-3 и TLR-4 МуD88-независимом пути. TIRAP оказался также внутриклеточным участником цепи передачи сигнала от TLR-2 и TLR-4. Также показали, что Му088-независимый путь вовлечен в регуляцию LPS-опосредованного созревания дендритных клеток. Известно, что TLR-1 и TLR-6 функционируют в составе гетеродимера с рецептором TLR-2.
[0058] TLR-3 распознает двунитевую вирусную РНК. TLR-5 идентифицировали как рецептор флагеллина из Грам-отрицательных и положительных бактерий, участник цепи передачи сигнала посредством MyD88. TLR-7 чевствителен к однонитевой РНК и небольшим синтетическим иммунномодуляторам, таким как имиквимод, R-848, бропиримин (bropirimine) и локсорибин (loxoribine). Известно, что TLR-9 распознает неметилированную бактериальную ДНК. В настоящее время исследуют способность CpG-олигодезоксинуклеотидов выступать в качестве адъювантов и стимулировать дендритные клетки человека с целью разработки вакцин.
[0059] TLR-4, TLR-7 и TLR-9 особенно важны для разработки вакцин. TLR-8 человека недавно идентифицировали как рецептор однонитевой РНК и резиквимода (R-848). Недавно предложили рассматривать TLR-7, TLR 8 и TLR-9 как подгруппу семейства TLR рецепторов, поскольку распознавание их лигандов происходит в компартментах эндосом/лизосом.
[0060] Также имеет место экспрессия рецепторов пектинов С-типа (кальций-зависимые) CLR, которые связывают консервативные олигосахариды, которые обычно присутствуют на поверхностных гликопротеинах вирусов, бактерий и других патогенов. Рецепторы CLR, которые экпрессируют дендритные клетки, включают рецепторы маннозы (CD206), DEC-205 (CD205), лангерина (CD207) и специфичный для дендритных клеток связывающий IСАМ-3 неинтегриновый рецептор (DC-SIGN; CD209). Эти рецепторы различаются не только тем, что их экспрессируют различные субпопуляции дендритных клеток и других тканей, они также распознают различные олигосахариды, различая таким образом лиганды.
[0061] DC-SIGN представляет собой трансмембранный белок II типа, имеющий молекулярную массу 44 кДа, который связывает и и нтернализирует некоторые вирусы, такие как ВИЧ, вирус Эбола, цитомегаловирус (CMV), вирус Денге, вирус гепатита С и бактерии, такие как Микобактерии, хотя в этом также участвуют другие рецепторы. Другие патогены также могут взаимодействовать с DC-SIGN. Он очень важен для функционирования дендритных клеток, как для опосредования взаимодействия naive Т-клеток через ICAM-3, и в качестве «подвижного» (rolling) рецептора, который опосредует ДК-специфические ICAM-2-зависимые процессы миграции.
[0062] Было описано взаимодействие между TLR и другими иммуно-распознающими рецепторами. Примером этого является совместное формирование воспалительных ответов дектином-1 и TLR2. Сложные частицы, такие как клеточные стенки дрожжей, распознаются множеством рецепторов врожденного иммуннитета, включая TLR2-TLR6, дектин-1 и CD14. Гетеродимеры TLR2-TLR6 активируют NF-кВ и продукцию хемокинов и цитокинов, таких как TNF-α. Дектин-1 распознает α-глюканы в клеточной стенке и запускает фагоцитоз, а также активацию реакции образования реактивного кислорода NADPH-оксидазой. Кроме того, передача сигнала дектином-1 совместно с передачей сигналов TLR2-TLR6 усиливает образование специфических цитокинов, таких как IL-12.
[0063] Благодаря совместному действию рецепторов TLR друг с другом и с другими рецепторами, а также взаимодействию между запускаемым ими внутриклеточными молекулярными механизмами, не представляется удивительным проявление синергетического действия между соединениями микробных патогенов, таких как липотейхоевая кислота, CpG-ДНК и пептидогликан. Это позволяет предположить, что действие активаторов TLR в качестве адъювантов можно усилить, если применять их в комбинациях при разработке иммуномодуляторов.
[0064] Совместная иммобилизация подходящего лиганда PRR (патогенраспознающего рецептора) и биоактивной молекулы может обеспечить направленную модуляцию иммунного ответа, приводящую к мощному клеточному ответу (под сравнительно более сильным влиянием Тh1) и/или гуморальному ответу (под влиянием Th2). Альтернативно, при иммунизации другим составом можно получить иммунную толерантность.
[0065] Возможно, мощный клеточный ответ можно индуцировать даже при установившемся гуморальном иммунитете или толерантности. Таким образом, мы можем предположить возможность разработки иммуномодулятора, эффективного в организме как инфицированного, так и не инфицированного хозяина. Такой результат значительно увеличил бы пользу вакцинации.
[0066] В модельной системе (цыплята) мы установили, что при вакцинации птиц предшествующей инфицированию микоплазмой, можно достичь значительной степени защиты против инфекционного штамма Mycoplasma gallisepticum. Кроме того, мы наблюдали обращение характеристик патологических симптомов у ранее инфицированных животных, что указывает на эффективность такой вакцины при лечении инфицированной группы. Это имеет значение в связи с широко распространенной устойчивостью различных штаммов микроорганизмов к антибиотикам и возрастающего беспокойства общества и регулирующих органов относительно разрешения применения антибиотиков в профилактических целях в отношении сельскохозяйственных животных. Кроме того, мы обнаружили, что для индукции защитного ответа таким способом достаточно лишь небольшого количества антигена (10 мкг) на животное, тогда как в литературе для случая применения микрочастиц, несущих антиген, указано более 100 мкг(Brayden, D. 2001 European Journal of Pharmaceutical Sciences 14:183-189). Это позволяет предположить, что молекула(ы) иммуномодулятора не разрушается, проходя через кишечник животного, и что она эффективно доставляется к иммунной клетке-мишени на слизистой оболочке. Это является существенным улучшением по сравнению с существующими иммунологически активными составами.
[0067] Однако при увеличении доз антигена наблюдали снижение защитного действия, что позволяет предположить наличие иммунносупрессивного компонента среди иммунноафинно очищенных антигенов. DC-SIGN задействован в механизме ускользания патогенов от механизмов иммунологического надзора. DC-SIGN представляет собой лектин С-типа специфичный к молекулам липидов с высоким содержанием маннозы. Мембраны микоплазмы содержат большую долю липидов, при этом было показано, что различные микоплазмы связывают аффинную смолу с Конканавалином A (Concanavalin А), что указывает на присутствие маннозы на поверхности микоплазмы. Мы предположили, что этот эффект могут опосредовать молекулы, участвующие в ускользании от иммунологического контроля, включая молекулы, подвергающиеся посттранскрипционному N-гликозилированию, которые включают посттранскрипционную липидоманнановую модификацию очищенного белка (белках), содержащую терминальные фрагменты маннозы. Затем попробовали удалять терминальные остатки маннозы путем ферментативного отщепления или химического разрушения. Дополнительно терминально маннозилированные липопротеины адсорбировали на специфичной к маннозе колонке с иммобилизованным комплексом лектин-конканавалин А (ConA).
[0068] Колонка с СоnА удерживала приблизительно одну треть компонентов антигена, и вакцина, приготовленная из очищенного антигена, демонстрировала самый высокий защитный эффект и линейную зависимость ответа иммуногй системы от дозы при возрастании концентрации антигена. С другой стороны, антиген, выделенный с Con А колонки, вызывал подавление воспалительных реакций у животных, в то время как в их внутренних органах обнаруживали присутствие очень высокого уровня патогена. Видимо, эти маннозилированные компоненты М. gallisepticum участвуют в механизме ускользания этого патогена от иммунологического контроля, приводя к иммуносупрессии и развитию толерантности к указанному патогену. Мы также продемонстрировали, что подходящие свойства антигена могут помочь сдвинуть иммунный ответ в сторону защиты либо толерантности. Это подтвердило нашу гипотезу, а также подкреплило наблюдения, сделанные с применением иммобилизованных мембран микоплазмы о том, что посттранскрипционные липидоманнозные модификации могут вызвать у организма-хозяина толерантность к патогену. Понимание этого может быть использовано для приготовления иммуномодуляторных микрочастиц, обладающих потенциальным свойством стимулировать толерантность или подавлять имеющиеся иммунные ответы. Посттранскрипционные модификации антигенов с присоединением липидов также принимают участие в развитии толерантности к патогенам. Поэтому мы также предприняли попытку деацилирования очищенных антигенов, которая оказалась эффективной, подтверждая роль липидов в патологических механизмах действия микоплазм.
[0069] Основное вывод из этим экспериментов состоит в том, что вакцина для индукции защитного иммунитета, содержащая эпитопы и разработанная на основе анализа нативных белков, не должна содержать эпитопов, богатых аминокислотами, которые могут подвергаться гликозилированию и/или присоединению липидов (Asn, Thr, Ser), в то время как лиганды, содержащие эти аминокислотные мотивы могут быть использованы для индукции толерантности. Это обеспечивает множество способов применения композиций и способов согласно настоящему изобретению.
[0070] В дальнейших исследованиях мы применяли плазму и сыворотку крови защищенных вакциной животных для идентификациии эпитопов антигенов, ответственных за защиту. Таким образом, мы сосредоточили усилия на создании научных основ для разработки вакцины на основе эпитопов, так как получение микоплазмы в промышленных масштабах и очистка антигенных белков могут быть слишком затратными для многих применений. В более раннем исследовании мы показали, что различные микоплазмы способны связывать гепарин и аналоги гепарина (Szathmary, S. et al.: Binding of mycoplasmas to solid phase adsorbents (Связывание микоплазм с твердофазными адсорбентами). Ada Vet Hung 2005, 53(3):299-307). Гликозаминогликаны (GAG) экспрессируются на поверхности клеток млекопитающих, а гликозаминогликан-связывающие белки на поверхности патогенов опосредуют адгезию с клетками-мишенями (Wadstrom Т, Ljungh A: Glycosaminoglycan-binding microbial proteins in tissue adhesion and invasion: key events in microbial pathogenicity (Гликозамингликан-связывающие микробные белки в тканевой адгезии и инвазии: ключевые события патогенности микробов). J Med Microbiol 1999, 48(3):223-233). Были описаны консенсусные последовательности нескольких типов, характеризующиеся преимущественно основными аминокислотами: ХВВХВХ, ХВВВХХВХ, ВВХХВВВХХВВ, ВВВХХВ, ВХВХВ, ВВВ, ВХВХХХВХВ, или ВХВХХХХХВХВ, где В обозначает основную аминокислоту, а Х - любую другую аминокислоту. Другая возможность заключается в том, что линейные мотивы основных аминокислот вовлечены в механизм патогенеза. Эти последовательности необходимы для присоединения, которое, возможно, служит начальным этапом в механизме проникновения патогена через слизистую оболочку. Следовательно, нейтрализация этой способности может предотвратить инфекцию и, возможно, обеспечит разработку широкого спектра способов лечения от разнообразных патогенных организмов. Эту возможность ранее не рассматривали как основу для разработки вакцин. Мы обосновали тот факт, что антигенные эпитопы, расположенные по соседству или стерически близкие к GAG-связывающим доменам, могут быть нейтрализующими, и допустили возможность, что такие сайты связывания, могут быть введены в состав нейтрализующих эпитопов.
[0071] Соответственно, мы далее выделили гепарин-связывающие белки из М. gallisepticum путем аффинной хроматографии на колонке со смолой Heparin Actigel (Sterogene Bioseparations, Inc., Карлсбад, Калифорния, США). Затем мы получили очищенный IgG из нейтрализующей сыворотки, полученной из цыплят, вакцинированных против М gallisepticum. Очищенные IgG иммобилизовали на активированной смоле Actigel ALD (Sterogene Bioseparations, Inc., Карлсбад, Калифорния, США) и адсорбировали на этой колонке выделенные гепаринсвязывающие белки. Связанные белки расщепляли путем добавления трипсина в колонку, элюировали фрагменты пептидов, содержащие иммуногенные эпитопы, включающие гепаринсвязывающие последовательности, и анализировали путем лазерной десорбционно-ионизационной масс-спектрометрии с участием матрицы (MALDI-MS), чтобы получить информацию о последовательности. Иммуноглобулины G, очищенные при помощи таких антигенных эпитопов или моноклональные тела против таких эпитопов при введении in vivo также могут сообщать защитный иммунитет инфицированному хозяину. В качестве альтернативы вместо трипсина можно применять другие известные в данной области протеолитические ферменты, такие как химотрипсин, эластаза, бромелаин, протеаза V-8, пепсин и термолизин.
[0072] В параллельных экспериментах мы повторно адсорбировали выделенные гепаринсвязывающие белки на смоле Heparin Actigel и провели аналогичное расщепление связанных белков трипсином. Фрагменты, восстановленные со смолы, также анализировали путем MALDI-MS для получения информации о последовательности.
[0073] Важно также идентифицировать эптитопы, участвующие в опсофагоцитозе, которые задействует система комплемента. В частности, комплементфиксирующие антитела выделяют из сыворотки крови вакцинированных цыплят на колонке с иммобилизованным C1q, и применяют такие антитела для идентификации эпитопов, способных стимулировать такой ответ. Затем очищенный IgG иммобилизуют и применяют для выделения белковых антигенов из неочищенного лизата MG. Связанные белки расщепляют протеазой прямо на колонке, а затем поледовательности эпитопов элюируют и анализируют путем MALDI-MS для получения информации о последовательности. Иммуноглобулины G, очищенные при помощи таких антигенных эпитопов, или моноклональные антитела против таких эпитопов при введении in vivo также могут сообщать защитный иммунитет инфицированному хозяину.
[0074] Основная проблема некоторых вакцин на основе микроорганизмов заключается в одновременном введении иммуностимулирующих и иммуноподавляющих эпитопов вместе со смесью агонистов PRR, присутствующих на самом патогене, некоторые из которых стимулируют ответ Th1, а некоторые - ответы Th2 или Treg. Эти вакцины не могут преодолеть персистенцию некоторых инфекционных агентов в организме хозяина и могут превратить его в «фабрику патогенов» при отсутствии клинических проявлений заболевания. Это подход может даже вызвать селективный отбора патогенов, что может привести к образованию более вирулентных штаммов. Наша стратегия, напротив, основана на создании "имитирующей патоген" микрочастицы, которая содержит только иммуностимулирующие эпитопы, и не содержит эпитопы и/или агонисты PRR, которые участвуют в ускользании патогенов от иммунной системы, совместно с подходящими молекулами-агонистами PRR, направляющими иммунный ответ. Идентифицированные эпитопы можно также соединять в один мультипептид, содержащий объединяющие последовательности или линкеры между конкретными пептидами, которые представляют собой эпитопы антигена. Этот подход позволяет восстановить нарушенный иммунный ответ, вызванный персистирующими патогенами, и развить сбалансированную комбинацию клеточного и гуморального иммунных ответов, которые приводят к уничтожению инфекционных агентов. Эпитопы ускользания или агонисты PRR можно применять для развития толерантности в случаях, когда необходимо преодолеть нежелательную аутоиммунную реакцию.
[0075] Обычно биологически активная молекула - это молекула, которая воздействует на иммунную систему, такая как иммуноген или другая молекула, модулирующая иммунную функцию, такая как иммуностимулятор, иммуноингибитор или агент, который стимулирует иммунологическую толерантность.
[0076] Биологически активные молекулы можно ковалентно или нековалентно присоединять к микрочастицам. Способы ковалентного присоединения хорошо известны в данной области и описаны, например у Р. Tijssen, "Practice and Theory of Enzyme Immunoassays" («Теория и практика ферментного иммуноанализа» Elsevier, Amsterdam, 1985, pp.283-289), в S. S. Wong, "Chemistry of Protein Conjugation and Crosslinking" («Химия конъюгирования и сшивки белков», CRC Press, Boca Raton, Florida, 1993), в Т. Е. Creighton, ed., "Protein Function: A Practical Approach" («Функия белка: практический подход»,IRL Press, Oxford, 1989) и в G.T. Hermanson, "Bioconjugate Techniques" («Биоконъюгационные методики», Academic Press, San Diego, 1996). Все эти источники включены в настоящую заявку посредством ссылок. Обычно, если микрочастиы изготовлены из агарозы, биоактивную молекулу присоединяют к гидроксильной группе агарозы. В целом, гидроксильные группы полисахаридов можно активировать определенными соединениями, которые образуют промежуточные химически активные производные, содержащие подходящие уходящие группы для последующего нуклеофильного замещения. Реакция этих активированных гидроксилов с нуклеофилами, такими как амины (например, группы лизина в белках или пептидах) приводит к образованию стабильных ковалентных связей, которые сшивают биоактивную молекулу с агарозой. Подходящие реагенты включают карбонилдиимидазол, производные хлорформиата, трезил хлорид, тозил хлорид, бромциан, дивинилсульфон, цианурхлорид, и бис-эпоксиды. В качестве альтернативы, гидроксильные группы углеводов, таких как агароза, могут быть модифицированы хлоруксусной кислотой с образованием карбоксилатной функциональной группы. В качестве другой альтернативы на полисахаридах можно создать аминные функциональные группы; можно провести реакцию между восстановленными концами молекул углевода или полученных альдегидов и соединениями ряда диаминов с короткой цепью (т.е. обычно содержащими менее приблизительно 6 атомов углерода в цепи), чтобы получить короткие алкиламиновые спейсеры, которые затем можно применять в реакциях конъюгации. Гидразидные группы могут быть созданы аналогичным образом с применением бис-гидразидных соединений. Полученные функциональные группы можно затем соединять с биоактивной молекулой путем различных реакций. Например, если получены карбоксильные группы, их можно затем конъюгировать с белками или пептидами комбинированным ангидридным способом, карбодиимидным способом, с применением дициклогексилкарбодиимида и способом с применением N-гидроксисукцинимидного эфира. Алифатические амины можно конъюгировать с белками или пептидами различными способами с применением карбодиимида, толилен-2,4-диизоцианата или малеимидных соединений, в частности, N-гидроксисукцинимидовых эфиров производных малеимида. Примером такого соединения является 4-(N-малеимидометил)-циклогексан-1-карбоксильная кислота. Другим примером является метамалеимидобензоил-N-гидроксисукцинимидовый эфир. Еще один реагент, который можно применять - N-сукцинимидил-3-(2-пиридилдитио)-пропионат. Для связывания элементов, содержащих аминогруппы, с белками можно также применять бифункциональные эфиры, такие как диметилпимелимидат, диметиладипимидат или диметилсуберимидат. В данной области известны также другие способы ковалентного связывания соединений, включая пептиды, белки и углеводороды, а также другие соединения. Способы нековалентного присоединения основаны на множественных нековалентных взаимодействиях, таких как водородные связи, гидрофобные связи и ионные связи, которые могут стабилизировать взаимодействие.
[0077] Обычно биологически активная молекула представляет собой одну или более из следующих: пептида, белка, рекомбинантного пептида, рекомбинантного белка, липида, углевода, нуклеиновой кислоты, гликопротеина или гликолипида. Комбинации этих молекул можно применять таким образом, что к одним микрочастицам будет присоединено множество биологически активных молекул.
[0078] Если иммунологически активная молекула представляет собой пептид или белок, она, возможно, подверглась посттранскрипционным модификациям, влияющим на иммунные свойства. Обычно эти модификации включают элементы сахаров и/или липидов. Примером является терминальное маннозилирование. Тем не менее другие углеводные заместители могут также быть добавлены к белкам или пептидам. В качестве альтернативы, иммунологически активную молекулу можно выделить таким способом, что препарат этой иммунологически активной молекулы будет обеднен терминально маннозилированными молекулами. Удаление молекул можно осуществить путем окисления, например окисления периодатом, путем обработки ферментами, обычно гидролитическими ферментами, или путем сахар-специфического аффинного связывания и последующей очистки фракции, лишенной модифицированных молекул. В данной области также известны другие способы удаления терминальных остатков маннозы.
[0079] Аналогично, иммунологически активную молекулу можно выделить таким образом, что препарат иммунологически активной молекулы будет практически лишен иммуномодулирующих липидсодержащих элементов, являющихся результатом посттранскрипционной модификации. Этого можно достичь путем химического гидролиза или путем копреципитации с пентадекановой кислотой. В качестве альтернативы, иммуномодулирующие липидсодержащие посттранскрипционные модификации можно блокировать заряженным детергентом. Наиболее подходящим детергентом является хлорид цетил триметил аммония. В качестве альтернативы можно применять аналогичные четвертичные аммониевые детергенты.
[0080] Возможно присутствующая нацеливающая молекула-ь обычно представляет собой патогенраспознающую молекулу. В одном варианте патогенраспознающая молекула представляет собой агонист рецептора TLR, такой как агонист рецептора TLR1, агонист рецептора TLR2, агонист рецептора TLR3, агонист рецептора TLR4, агонист рецептора TLR5, агонист рецептора TLR6, агонист рецептора TLR7, агонист рецептора TLR8 или агонист рецептора TLR9. В другом варианте патогенраспознающая молекула представляет собой агонист белка NOD, такой как агонист NOD-1 или агонист NOD-2. Обычно агонист белка NOD представляет собой бактериальный пептидогликан или производное бактериального пептидогликана.
[0081] При необходимости можно вводить в состав и стабильно связывать с микрочастицами более одной биологически активной молекулы, такой как иммунологически активная молекула, а также более одной патогенраспознающей молекулы.
[0082] В другом своем аспекте настоящее изобретение относится к способу индукции у пациента иммунного ответа путем введения указанному пациенту иммунологически эффективного количества описанного выше состава. Обычно состав также включает нацеливающую молекулу, такую как патогенраспознающая молекула. В состав можно включать больше одной иммунологически активной молекулы и больше одной патогенраспознающей молекулы.
[0083] Еще в одном своем аспекте настоящее изобретение относится к способу доставки иммунологически активного состава in vivo, включающему этап введения эффективного количества описанного выше состава в организм, обладающий активной иммунной системой. Здесь также можно вводить в состав более одной иммунологически активной молекулы и более одной патогенраспознающей молекулы. Доставку иммунологически активного состава in vivo можно осуществлять через слизистую оболочку, парентеральным путем, через кожу или подкожно. Можно применять другие пути введения.
[0084] Еще в одном своем аспекте настоящее изобретение относится к способу индукции защитного иммунного ответа на по меньшей мере один патоген, включающему введение пациенту иммунологически эффективного количества состава, содержащего одну или более иммунологически активных молекул (т.е. иммуногенов) и комбинацию агонистов рецепторов TLR, стабильно соединенные с микрочастицами, как описано выше, с целью стимулировать у указанного пациента защитный иммунный ответ на указанный патоген. Защитный иммунный ответ включает Th1 или Тh2 ответы или комбинацию Th1 и Тh2 ответов. Вводить состав можно одной или несколькими дозами.
[0085] Еще в одном своем аспекте настоящее изобретение относится к способу индукции толерантности к иммунологически активному агенту, включающему введение пациенту иммунологически эффективного количества состава, содержащего одну или более иммунологически активных молекул (т.е. иммуногенов) и комбинацию агонистов рецепторов TLR и агонистов белка NOD, стабильно связанные с микрочастицами, с целью стимулировать у пациента толерантность к иммунологически активному агенту. Иммунологически активные молекулы могут иметь липидосодержащий фрагмент, который можно присоединить к микрочастицам независимо от оставшейся части иммунологически активных молекул. В качестве альтернативы иммунологически активный липидосодержащий фрагмент может дополнительно содержать углеводные группы.
[0086] Токсичность и терапевтическую эффективность составов согласно настоящему изобретению можно определить стандартными фармацевтическими способами на клеточных культурах или экспериментальных животных, для определения, например, LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Отношение доз, оказывающих терапевтическое и токсическое действие, называют терапевтическим индексом, его можно выразить как отношение LD50/ED50. Предпочтительными являются соединения с высокими терапевтическими индексами. Данные, полученные на клеточных культурах или экспериментальных животных, могут быть использованы для определения диапазона дозировок, приемлемых для людей или других животных. Дозировка таких составов предпочтительно определяется диапазоном циркулирующей концентрации, который включает ED50 при низкой или отсутствующей токсичности. В переделах этого диапазона дозировка может меняться в зависимости от применяемой дозы и применяемого пути введения состава.
[0087] Для любого состава согласно настоящему изобретению терапевтически эффективную дозу можно первоначально определить в результате эксперимента культурах клеток. Например, на модельных животных можно определить дозу, позволяющую достичь диапазона циркулирующей в плазме крови концентрации, который включает IC50, определенную для клеточной культуры (т.е. концентрацию исследуемого состава, которая позволяет достичь половины максимального ответа относительно воздействия состава на измеряемый параметр иммунной системы). Такую информацию можно применять для более точного определения применимых для людей доз. Уровни в плазме крови можно измерить, например, путем ВЭЖХ.
[0088] Точная лекарственная форма, путь введения и дозировка может быть выбрана каждым лечащим врачом на основании состояния пациента. (См., например, Fingl et al., in The Pharmacological Basis of Therapeutics (Фармакологические основы терапии), 1975, Ch. 1 p.1). Необходимо отметить, что лечащий врач будет знать, каким образом и когда следует прекратить, прервать или изменить применение состава из-за токсичности или нарушений функций органов. С другой стороны, лечащий врач также будет знать, когда следует начать вводить более высокие дозы при недостаточном клиническом ответе (избегая токсичности). Величина вводимой дозы при управлении течением представляющего интерес заболевания будет меняться в зависимости от тяжести заболевания и пути введения состава. Тяжесть заболевания можно частично оценить, например, стандартными способами прогностической оценки. Кроме того, дозу и, возможно, частоту введения дозы также можно варьировать в зависимости от возраста, массы тела и ответа на состав для конкретного пациента. Программу, аналогичную обсужденной выше, можно применять в ветеринарной медицине.
[0089] Применение фармацевтически приемлемых носителей для приготовления составов, описанных в настоящей заявке, с целью реализации настоящего изобретения, в лекарственных формах, подходящих для системного введения, входит в объем настоящего изобретения. При правильном выборе носителя и подходящего способа производства, составы согласно настоящему изобретению, в частности, изготовленные в форме растворов, можно вводить парентерально, например, путем внутривенных инъекций. Составы можно легко приготовить с применением фармацевтически приемлемых носителей, хорошо известных в данной области, подходящих для введения в слизистую оболочку или подкожно.
[0090] Настоящее изобретение иллюстрируют приведенные ниже Примеры. Эти Примеры приведены исключительно в пояснительных целях и не предназначены для ограничения настоящего изобретения.
ПРИМЕР 1
Выбор микрочастиц
[0091] Микрочастицы из агарозы с размерами в диапазоне 1-10 мкм производства Sterogene Bioseparations, Inc. (Carlsbad, Калифорния, США) исследовали на анализаторе размера частиц Saturn DigiSizer 5200 (Micromeritis Instrument Corp). Данные, показанные на Фигуре 1, демонстрируют следующее распределение частиц по размеру: 75% - меньше 5 мкм, 24% - от 5 до 10 мкм и 1% - больше 10 мкм.
ПРИМЕР 2
Культивирование Mycoplasma gallisepticum (MG)
[0092] К 0.1 мл среды для выращивания микоплазм добавляли лиофилизованную форму М. gallisepticum (K781R-16P) и помещали в 1.5 мл культуральной среды в инкубатор при 37°С. За ростом микоплазмы следили по изменению цвета среды (розовой на оранжевую или желтую) или помещая на чашки с агаром и проверяя колонии. Большие объемы (10-15 л) культур Mycoplasma выращивали путем перенесения зараженной культуральной среды в свежую среду. Затем культуры центрифугировали на 5500 об/мин в течение 30 мин. Осадок промывали PBS (20 мин на 5500 об/мин) до тех пор, пока оптическая плотность надосадочной жидкости на длине волны 280 нм (OD2so) не опускалась ниже 0.2. Осадок ресуспендировали в 20 мл PBS.
ПРИМЕР 3
Подготовка иммуноаффинной колонки для очистки MG антигена
Этап 1:
[0093] К 64 мл куриной сыворотки против M.gallisepticum добавляли 128 мл деионизованной (ДИ) воды в стеклянном мерном стакане. рН раствора доводили до 4.5 ледяной уксусной кислотой. Раствор перемешивали быстро, но осторожно, чтобы избежать разбрызгивания или образования пены. Преципитирующий раствор CAP-8 Precipitating Solution (Sterogene Bioseparations, Inc. Carlsbad, Калифорния, США) интенсивно перемешивали в течение 10 минут. Затем отмеряли 64 мл преципитирующего раствора САР-8 и медленно вливали по стенке воронки, образовавшейся в процессе перемешивания, в течение 1-2 минут. Затем перемешивание замедляли, чтобы раствор перемещался без образования воронки. Раствор перемешивали в течение 30 минут при комнатной температуре и затем переносили в центрифужную пробирку подходящего размера и осаждали преципитат центрифугированием на 5,500 об/мин в течение 15 минут. Надосадочную жидкость переносили в контейнер, а осадок промывали один раз 20 мл буфера, содержащего 20 мМ ацетата Na, рН 4.8. Надосадочную жидкость фильтровали через фильтр для шприца с размером пор 0.22 мкм.
Этап 2:
[0094] Катионообменную смолу SP Thruput Plus (Sterogene Bioseparations, Inc. Карлсбад, Калифорния, США) суспендировали в 3 объемах 1М NaOH в течение 10 минут, а затем промывали деионизованной водой до нейтральности. Затем ее промывали 10 объемами 0.5 М ацетата натрия (рН 4.8) а затем 20 объемами деионизованной воды. Смолу уравновешивали 15 объемами 20 мМ ацетата натрия, рН 4.8, и загружали на колонку, применяя загрузочный буфер, содержащий 20 мМ ацетата, рН 4.8. Полученную на Этапе 1 надосадочную жидкость загружали на колонку при скорости потока 3 мл/мин и промывали колонку буфером, содержащим 20 мМ ацетата натрия, рН 4.8. Комбинировали прокачивание жидкости с промывкой. Измеряли OD280 против 20 мМ ацетата натрия, рН 4.8. Колонку элюировали буфером, содержащим 50 мМ фосфата натрия, 300 мМ NaCl с рН 8.0 и измеряли OD280 элюанта.
Этап 3:
[0095] К 20 мл активированной смолы Actigel ALD (Sterogene Bioseparations, Inc. Карлсбад, Калифорния, США) добавляли раствор анти-М. gallisepticum очищенных куриных IgG, а затем 10.5 мл 1М цианоборогидрида натрия (Связывающий раствор ALD Coupling Solution, Sterogene Bioseparations, Inc. Карлсбад, Калифорния, США). Суспензию осторожно перемешивали в течение 20 часов при 2-8°С, а затем обильно промывали деионизованной водой. Смолу хранили в PBS с рН 7.0 при 2-8°С.
ПРИМЕР 4
Очистка MG антигенов
Этап 1:
[0096] К 20 мл промытого концентрата MG добавляли 0.2 г детергента Меда-10 и перемешивали в течение 20 часов при комнатной температуре. После 20 часов инкубации к суспензии также добавляли 1 мл Triton-X 100 и продолжали перемешивание при комнатной температуре в течение еще одного часа. Затем ее центрифугировали при 5,000 об/мин в течение 10 минут. Надосадочную жидкость отделяли от осадка и добавляли к ней 200 мл PBS с рН 7.2. Раствор белков из MG выдерживали при 2-8°С в течение 5 дней.
Step 2:
[0097] Колонку с объемом 20 мл, заполненную смолой Actigel с IgG кур против MG уравновешивали 5 объемами PBS с рН 7.2 при скорости потока 3 мл/мин. Раствор белков из MG загружали на колонку при скорости потока 8-15 мл/мин. Пропущенную через колонку жидкость собирали в отдельную бутылку. Колонку промывали 10 объемами PBS, рН 7.2, при скорости потока 8-15 мл/мин, а затем элюировали 20-40 мл 0.1М лимонной кислоты, рН 2.5, при той же скорости потока. рН элюата немедленно доводили до 7.2 2 М Tris-буфером. Такую очистку повторяли по меньшей мере 5 раз, используя пропущенную через колонку жидкость, чтобы адсорбировать все антигены. Все элюаты смешивали и концентрировали в диализном мешке, помещенном на ночь в сахарный порошок при температуре 2-8°С. Концентрированный раствор диализовали против 5 л PBS, рН 7.2, при 2-8°С в течение ночи. Проводили анализ концентрации белка по Брэдфорд (Bradford), чтобы определить концентрацию очищенного антигена. В качестве стандарта применяли сывороточный альбумин.
ПРИМЕР 5
Активированные микрочастицы из агарозы
[0098] Частицы активировали двумя разными способами. Активацию первым способом проводили с применением коммерчески доступного патентованного реагента для образования альдегидной связи фирмы Sterogene Bioseparations, Inc. (Карлсбад, Калифорния, США), который обеспечивает очень стабильное присоединение лигандов. Другое преимущество этого реагента - высокая воспроизводимость иммобилизации. Этот патентованный альдегидный реагент позволяет параллельно присоединять различные лиганды.
[0099] Другую активацию проводили с применением способа активации бромциана (CNBr). Вкратце, к 30 мл агарозных микрочастиц добавляли 30 мл 2 М раствора карбоната натрия и выдерживали в течение 3-5 минут на ледяной бане без перемешивания. Затем взвешивали 1.5 г CNBr и растворяли в 9 мл ацетонитрила. Раствор CNBr немедленно добавляли к смеси, содержащей смолу, и интенсивно перемешивали на ледяной бане в течение 2 минут. Затем смесь промывали 20 объемами ледяной воды путем центрифугирования при 4,500 об/мин, 2°С в течение 5 минут.
ПРИМЕР 6
Присоединение MG антигенов к микрочастицам
[0100] Реакция присоединения происходит между аминогруппами очищенных антигенов и альдегидными группами или CNBr-активированными группами микрочастиц. В случае применения активированных альдегидом микрочастиц присоединение осуществляли при помощи связывающего реагента, цианоборангидрида натрия, как описано в Примере 3. Антигены иммобилизовали при концентрациях 10 мкг/0.2 мл и 50 мкг/0.2 мл микрочастиц.
[0101] В другой реакции присоединения применяли активированные CNBr микрочастицы с теми же концентрациями антигенов. К 15 мл активированных CNBr микрочастиц добавляли раствор очищенного MG антигена при рН 8.0. Раствор аккуратно перемешивали при 2-8°С в течение 20 ч. Надосадочную жидкость отделяли путем центрифугирования и промывали смолу 10 объемами деионизованной воды.
Связанную смолу хранили в LAL-воде (вода, прошедшая специальный тест на бактериальные эндотоксины) при 2-8°С. Для определения количества несвязанного белка в надосадочной жидкости применяли анализ на белок по Брэдфорд.
ПРИМЕР 7
Приготовление и присоединение мембраны микоплазмы.
[0102] К 3.0 мл концентрированного антигена микоплазмы добавляли 80 мл автоклавированной деионизованной воды и перемешивали на магнитной мешалке при 37°С в течение 1 ч. Раствор центрифугировали при 5000 об./мин в течение 30 мин и дважды промывали осадок автоклавированной водой. Осадок восстанавливали в 10 мл PBS. К 1.5 мл активированных CNBr микрочастиц добавляли 0.5 мл очищенного осадка, разведенного 1:3 в 0.1 М NaHCO3 с рН 8. Раствор осторожно перемешивали при 2-8°С в течение 20 ч. Отделяли надосадочную жидкость центрифугированием и промывали смолу 10 объемами деионизованной воды. Связанную смолу хранили в LAL-воде при 2-8°С. Для определения количества несвязанного белка в надосадочной жидкости применяли анализ на белок по Брэдфорд
ПРИМЕР 8
Присоединение активатора рецептора NOD1 к микрочастицам
[0103] Пептидогликан (PG) в концентрации 2 мкг/0.2 мл смолы растворяли в 0.1М NaHCO3 и добавляли к активированным CNBr микрочастицам, приготовленным согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 9
Присоединение активатора TLR3 к микрочастицам
[0104] Поли(I:С) в концентрации 10 мкг/0.2 мл гранул иммобилизовали на активированных CNBr микрочастицах в 0.1 М NaHCO3 при рН 8, согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 10
Присоединение активатора TLR4 к микрочастицам
[0105] Бактериальный липополисахарид в концентрации 2 мкг/0.2 мл смолы растворяли в 0.1 М NaНСО3 при рН 8 и иммобилизовали на активированных CNBr микрочастицах, согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 11
Присоединение активатора TLR5 к микрочастицам
[0106] Флагеллин в концентрации 2 мкг/0.2 мл смолы растворяли в 0.1 М NаНСО3 при рН 8 и иммобилизовали на активированных CNBr микрочастицах согласно Примеру 6. Реакцию велив течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 12
Присоединение активатора TLR1 и TLR6 к микрочастицам
[0107] Антиген поверхности микоплазмы, содержащий MALP-2, в концентрации 2 мкг/0.2 мл смолы растворяли в 0.1М NаНСО3 при рН 8 и иммобилизовали на активированных CNBr микрочастицах согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 13
Присоединение активатора TLR7/TLR8 к микрочастицам
[0108] Однонитевую РНК или противовирусный имидазоквинолин Imiquimod (Aldara) в концентрации 2 мкг/0.2 мл смолы растворяли в 0.1М NаНСО3 при рН 8 и иммобилизовали на активированных CNBr микрочастицах согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 14
Присоединение активатора TLR9 к микрочастицам
[0109] CpG-ДНК в концентрации 10 мкг/0.2 мл смолы растворяли в 0.1М NаНСО3 при рН 8 и иммобилизовали на активированных CNBr микрочастицах согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 15
Иммобилизация комбинированных смесей молекул - агонистов рецептора распознавания РАМР на микрочастицах
[0110] Агонист NOD1 и агонисты 4 и 9 TLR смешивали в концентрацях 2 мкг/0.2 мл смолы, 10 мкг/0.2 мл смолы и 2 мкг/0.2 мл смолы, соответственно, в 0.1 М NаНСО3 при рН 8 и иммобилизовали на активированных CNBr микрочастицах согласно Примеру 6. Реакцию вели в течение ночи при 2-8°С. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 16
Иммобилизация MG антигена совместно с комбинированными смесями молекул - агонистов рецептора распознавания РАМР на микрочастицах
[0111] MG антигены присоединяли в условиях, описанных в Примере 6 в течение 1 часа. Затем добавляли смесь агонистов TLR, описанную в Примере 13, и реакцию вели в течение ночи at 2-8°C. Отделяли надосадочную жидкость путем центрифугирования и тщательно промывали смолу LAL-водой. Эту же воду применяли в качестве среды для хранения.
ПРИМЕР 17
Исследование на животных 1
[0112] Результаты усреднены по 3 экспериментам.
[0113] Вакцинировали трехдневных цыплят (не зараженных М. gallisepticum (MG) и М. synoviae (MS), у которых твердофазный иммуноферментный анализ ELISA не выявил материнских антител к MG и MS). Каждая группа состояла из 10 цыплят. На 14 день эксперимента цыплят инфицировали штаммом Rlow M. gallisepticum. Исследование завершали на 28 день.
[0114] Группы организовывали следующим образом:
G1-G5 введение составов вакцин на микрочастицах перед инфицированием
G1 = вводили перорально только микрочастицы (0.2 мл/цыпленка) и инфицировали
G2 = вводили перорально микрочастицы (0.2 мл/цыпленка) с аффинно очищенным антигеном M. gallisepticum (10 мкг/дозу) и инфицированли
- G3 = вводили перорально микрочастицы (0.2 мл/цыпленка) с агонистами рецептора (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали.
- G4 = вводили перорально микрочастицы (0.2 мл/цыпленка) с аффинно очищенным M. gallisepticum антигеном (10 мкг/дозу) и агонистами рейептора (10 мкг бактериальной ДНК, E.coli+2 мкг липополисазарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали.
- G5 = вводили перорально микрочастицы (0.2 мл/цыпленка) с мембраной M. gallisepticum (107/дозу)
G6-G7 введение составов вакцин на микрочастицах после инфицирования
- G6 = инфицировали и после этого перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным M. gallisepticum антигеном (10 мкг/дозу) и агонистами рецепторов (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу)
- G7 = инфицировали и после этого перорально вводили микрочастицы (0.2 мл/цыпленка) с агонистами рецепторов (10 мкг бактериальной ДНК, E.coli++2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу)
G8 и G9 = положительный и отрицательный контроли
G8 = инфицировали и ничего не вводили
G9 = не инфицировали и ничего не вводили
График
[0115] День-1: Формирование групп G1-G9. Умерщвление 10 цыплят для твердофазного иммуноферментного анализа, ПЦР и культивирования М. gallisepticum и М. synoviae для подтверждения того, что у подопытных цыплят нет материнских антител и отсутствуют М. gallisepticum и М. synoviae.
[0116] День 0: Вакцинация G1-G5 перед инфицированием.
[0117] День 14: Инфицирование групп G1-G8.
[0118] День 15: Вакцинирование групп G6-G7 после инфицирования.
[0119] День 28: G1-G9. Эвтаназия, аутопсия и культивирование определенных органов, трахеи, воздухоносного мешка и легкого для выделения М. gallisepticum (MG). Проводили гистологическое исследование трахеи и легкого.
[0120] Дни D14 и D28: У цыплят брали кровь, чтобы получить сыворотку для исследования на MG-специфические антитела путем проведения реакции агглютинации сыворотки на пластинке (SPA) и блокирующего твердофазного ИФА
[0121] В D-1 однодневных бройлерных цыплят общим количеством 90 делили на восемь групп (10 птиц/группу). Записывали массу тела каждого цыпленка. Цыплят делили на группы таким образом, чтобы средние массы в группах незначительно различались между собой. Каждой птице на крыло вешали цветной ярлычок с номером, согласно проводимому лечению и массе тела, записанных на соответствующем бланке.
Введение составов
[0122] В День 0, цыплятам групп G1-G5 вводили вакцин различного состава, 0.2 мл в 1 мл PBS на животное. Дозы описаны выше. Группам G6-G7 составы вводили в день 15 после инфицирования.
Инфицирование
[0123] В День D14 восемь групп животных G1-G8 инфицировали свежим культуральным бульоном вирулентного R-штамма М. gallisepticum, с титром, приблизительно равным 8.0 log10KОЕ/мл. Каждой из групп вводили десять мл этого свежего культурального бульона способом распыления. Вкратце, птиц помещали в изолированный блок объемом 0.22 кубических метров. Затем распыляли десять мл свежей культуры R-штамма М. gallisepticum под давлением 1 атмосфера в течение приблизительно 100 секунд и оставляли цыплят в блоке на 20 минут (В лаборатории при применении этой методики в нескольких экспериментах получили положительные результаты).
Эвтаназия и патология
[0124] В Д28, в конце экспериментального исследования все группы подвергали эвтаназии. Проводили аутопсию каждой птицы и определяли показатели макроскопических поражений, связанных с MG. Регистрировали наличие экссудата в трахее, левом и правом грудных воздухоносных мешках и брюшной полости. Показатель поражений определяли по следующей системе: В трахее: 0 = нет экссудата, 1 = легкое покраснение и небольшое количество экссудата, 2 = покраснение слизистой оболочки, экссудат.
Левый и правый воздухоносные мешки: 0 = нет поражений, 1 = сывороточный экссудат, 2 = сывороточный экссудат и небольшие скопления фибрина, 3 = сывороточный, фиброзный экссудаты, легкое утолщение стенки воздухоносного мешка, 4 = большое количество фиброзного экссудата, сильное утолщение стенки воздухоносного мешка.
Брюшная полость: 0 = нет экссудата, 1 = сывороточный экссудат, 2 = сывороточный экссудат с небольшими скоплениями фибрина, 3 = сывороточно-фиброзный экссудат.
Выделение MG
[0125] Во время исследования аутопсийного материала асептически, с помощью ватного тампона, изолировали образцы трахеи, грудных воздухоносных мешков, печени, легкого, селезенки, почки и сердца. Материал с ватных тампонов высевали на агар для микоплазм (МА) и инкубировали при 37°С в инкубаторе с 5% СO2. Чашки исследовали на микоплазму в дни 2, 4 и 7, а затем с недельными интервалами в течение максимум трех недель. Положительные колонии исследовали путем ПЦР, чтобы идентифицировать М. gallisepticum и М. synoviae.
Аутопсия
[0126] После инфицирования MG выявляли значительные патологические поражения в воздухоносном мешке и брюшной полости. Однако в группах, которым вводили частицы с агонистами рецепторов (G3, G6 p<0.001), частицами с очищенным антигеном (G2, p<0.001) и очищенным антигеном плюс агонисты рецепторов (G4, G6 p<0.001), регистрировали значительное снижение показателей поражений по сравнению с контрольной группой (G8), которой не вводили никаких веществ и инфицировали, а также с инфицированной группой, которой вводили только частицы (G1). Однако лучшие результаты получали с частицами плюс очищенный антиген или с частицами плюс очищенный антиген плюс агонисты рецепторов, если их вводили перед инфицированием, по сравнению с введением после инфицирования. При введении группе G5 мембраны М. gallisepticum, иммобилизованной на частицах, патологические поражения в некоторых случаях были более выраженными, чем в самой инфицированной группе. Это приводит к выводу о том, что вакцинация мембраной М. gallisepticum препятствовала адекватному иммунному ответу на инфицирование патогеном, вызывая подавление иммунного ответа и приводя к более выраженной инфекции. Повторное выделение микоплазмы
[0127] Микоплазму можно многократно повторно выделять из внутренних органов невакцинированных инфицированных цыплят контрольной группы. Наблюдали значительное снижение степени повторного выделения (из органов дыхания + внутренних органов) в группах, которым вводили частицы плюс очищенный антиген с агонистами рецепторов или без них (G2, G4, G6), по сравнению с контрольной группой, которую не вакцинировали (G8, p<0.01), и с группой, которой вводили только частицы (G1, p<0.001-0.01) или частицы плюс агонисты рецепторов (G3, G7 p<0.05). Однако степень повторного выделения существенно снижалась (p<0.05) в группе, которой вводили частицы плюс агонисты рецепторов (G3, G6) по сравнению с контролем (G8). Аналогичные результаты получали, когда сравнивали степень повторного выделения микоплазмы из дыхательных путей (трахея, легкие, воздухоносный мешок) или из других внутренних органов (печень, селезенка, почка и сердце) экспериментальных групп. Когда G5 вводили мембраны М. gallisepticum, иммобилизованные на частицах, степень повторного выделения М. gallisepticum из органов в некоторых случаях была выше, чем в самой инфицированной группе. Это приводит к выводу о том, что вакцинация мембраной М. gallisepticum препятствовала адекватному иммунному ответу на инфицирование патогеном, вызывая подавление иммунного ответа и приводя к более выраженной инфекции.
[0128] Результаты показаны в Таблице 1.
Результаты серологических исследований
[0129] В конце эксперимента серологическая реакция групп различалась. Реакция группы, которой не вводили никаких веществ, но инфицировали (G8), не отличалась от группы, которой вводили только частицы (G1). В то же время в группе, которой вводили частицы плюс очищенный антиген и агонисты PRR, наблюдали значительно более сильную реакцию (G4) (p<0.05), чем в группе, которой вводили только частицы (G1). По сравнению с группой, которой вводили частицы плюс очищенный антиген (G2), при добавлении PRR (G4, G6), наблюдали значительно более высокий серологический ответ (p<0.05). Значительной разницы между результатами серологических исследований в случаях, если частицами с антигеном и PRR применяли до (G4) или после (G6) инфицирования, не было.
Обсуждение
[0130] М. gallisepticum может вызывать сильное воспаление в воздухоносном мешке и брюшной полости, которое сопровождается колнизацией трахеи, воздухоносного мешка и легких. Микоплазму также можно часто обнаружить во внутренних органах. Мы разработали новый тип «имитирующих патоген» иммунологически активных составов, состоящих из микрочастиц, имеющих диапазон размеров, соответствующих размерам микроорганизмов (<5 мкм), несущих очищенные иммуноаффинным способом антигены, ковалентно иммобилизованные вместе с различными молекулами - агонистами PRR.
[0131] Полученные нами результаты показали, что частицы без какой-либо модификации не вызывают никакого иммунного ответа. Частицы сами по себе не защищали цыплят от перитонита и воспаления воздухоносных мешков, вызываемых М. gallisepticum, а также не предотвращали колонизации органов микоплазмой.
[0132] Когда частицы покрывали агонистом PRR без антигена, они не оказывали влияния на серологический ответ на инфицирование микоплазмой. Однако колонизация органов микоплазмой уменьшалось, а показатели патологических поражений снижались. Это подтверждает наши наблюдения in vitro о том, что агонисты PRR действительно стимулируют или усиливают врожденный иммунный ответ, усиливая защиту. Однако применяемое количество иммуномодулятора не могло полностью защитить животное от крупных инфицирующих доз высоко патогенного штамма. Такой эффект также известен для врожденного иммунитета, поскольку его обычно недостаточно для борьбы с большими дозами патогенов. Однако эти данные указывают на то, что этот новый иммуномодулятор, возможно, способен защитить от инфицирования более низкими дозами.
[0133] Когда к частицам, покрытым агонистами PRR, добавляли очищенный антиген, специфический серологический ответ на микоплазму усиливался. Колонизация органов значительно снижалась, а показатели патологических поражений были низкими. Этот эффект был более выраженным, когда вакцину вводили в слизистую и перед инфицированием, но аналогичный положительный ответ наблюдали, и когда вакцину вводили в слизистую оболочку после инфицирования.
[0134] Интересное наблюдение - увеличение концентрации антигена (до 50 мкг/дозу) фактически снижало защитное действие вакцины. Это позволяет сделать предположение о присутствии иммуносупрессивных компонентов в аффинно очищенном антигене. Чтобы выявить эти компоненты и избавиться от их действия, запланировали новые испытания на животных (см. ниже).
[0135] После вакцинации и до инфицирования, вакцинируемых цыплят ежедневно осматривали, чтобы оценить безопасность вакцины. Цыплят признали клинически здоровыми, побочных эффектов вакцины не обнаружили. Потребление пищи и воды птицами не изменилось по сравнению с группами, которые не вакцинировали. У животных, аутопсию которых проводили в ходе исследования, не обнаружили признаков воспаления или изменения размера/массы органов. Видимо, вакцина безопасна.
ПРИМЕР 18
[0136] Аффинно очищенный MG антиген готовили, как описано в предыдущих Примерах. Перед иммобилизацией очищенный антиген разделяли на три группы для обработки следующими способами.
1. Расщепление эндогликозидазой Н
[0137] К 53.6 мл антигенов (приблизительно 1.5 мг) добавляли 2.5 Ед. фермента и инкубировали при 37°С в течение ночи. На следующий день смесь прогоняли через охлажденную колонку с иммобилизованным маннаном (5 мл) и собирали пропущенную через нее жидкость. Этот образец обозначали Endo Н Antigens (антигены, расщепленные эндогликозидазой Н).
2. Обработка перйодатом
[0138] К 53.6 мл антигенов (приблизительно 1.5 мг) добавляли твердый перйодат натрия до конечной концентрации 15 мМ и после перемешивания выдерживали охлажденным в течение 1 часа. Добавляли двухкратный молярный избыток глицерина и инкубировали образец в течение еще одного часа. Проводили диализ против PBS в течение ночи. Диализованный образец обозначали Periodate (PJ) Antigens (антигены, обработанные перйодатом).
3. Удаление СоnА связывающих антигенов
[0139] Очищенные антигены (приблизительно 1.5 мг в 53.6 мл) прогоняли через колонку с иммобилизованным Con А объемом 2 мл. Собирали проточную жидкость. Этот образец обозначали Con A flowthrough (Проточная жидкость СоnА). Связанные антигены элюировали 1М альфа-метил манногликозидом в 50 мМ TRIS, рН 9.5. Этот образец обозначали Con A elution (СоnА Элюат).
Обработка антигена
[0140] Свойства антигенов показаны в Таблице 2.
ПРИМЕР 19
Исследование на животных II
[0141] Результаты представляют собой среднее по 3 экспериментам. Вакцинировали трехдневных цыплят (не зараженных М. gallisepticum (MG) и М. synoviae (MS), у которых твердофазный иммуноферментный анализ не выявляет материнских антител MG и MS). Каждая группа состояла из 10 цыплят.На 14 день эксперимента цыплят инфицировали штаммом М. gallisepticum Riow. Исследование завершали на 28 день.
Группы организовывали следующим образом:
G1 = не инфицировали и ничего не вводили
G2 = инфицировали и ничего не вводили Введение за 2 недели до инфицирования:
G3 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным антигеном М. gallisepticum (10 мкг/дозу) и агонистами TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали
G4 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным антигеном М. gallisepticum (50 мкг/дозу) и агонистами TLR (10 мкг бактериально ДНК, E.coli+2 мкг липополисахарида Е. coli и 2 мкг пептидогликана/дозу) и инфицировали
G5 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным адсорбированным СоnА антигеном М. gallisepticum (10 мкг) и инфицировали
G6 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным расщепленным эндогликозидазой Н антигеном М. gallisepticum (10 мкг) и инфицировали
G7 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным окисленным перйодатом антигеном М. gallisepticum (10 мкг) и инфицировали
G8 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным адсорбированным СоnА антигеном М. gallisepticum (10 мкг)+агонист TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали
G9 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным адсорбированным СоnА антигеном М. gallisepticum (50 мкг)+ агонист TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидосликана/дозу) и инфицировали
G10 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным расщепленным эндогликозидазой Н антигеном М. gallisepticum (10 мкг)+агонисты TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида Е.coli и 2 мкг пептидогликана/дозу) и инфицировали
G11 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным расщепленный эндогликозидазой Н М. gatlisepticum (50 мкг)+агонисты TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали
G12 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным окисленным периодатом антигеном М. gallisepticum (10 мкг)+агонисты TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали
G13 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным окисленным периодатом антигеном М. gallisepticum (50 мкг)+агонисты TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали
G14 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным расщепленным эндонуклеазой Н М. gallisepticum (10 мкг)+ агонисты TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) плюс аминогуанидин (внутрибрюшинно) в течение 7 дней и инфицировали
G15 = перорально вводили микрочастицы (0.2 мл/цыпленка) с Con А элюатом антигена М. gallisepticum на гранулах и инфицировали
График
[0142] День-1: Формирование групп G1-G15. Умерщвление 10 цыплят для твердофазного иммуноферментного анализа ELISA, ПЦР и культивирования М. gallisepticum и М. synoviae для подтверждения того, что у подопытных цыплят нет материнских антител и отсутствуют М. gallisepticum и М. synoviae.
[0143] День 0: Вакцинация G3-G14 перед инфицированием. День 14: инфицирование групп G2-G15.
[0144] День 28: G1-G15. Эвтаназия, аутопсия и посев для выделения М. gallisepticum (MG) из отдельных органов, трахеи, воздухоносного мешка и легкого. Проводили гистологические исследования трахеи и легкого.
[0145] Дни D14 и D28: У цыплят брали кровь, чтобы получить сыворотку для исследования на MG-специфические антитела путем проведения реакции агглютинации сыворотки на пластинке (SPA) и блокирующего твердофазного ИФА.
[0146] В Д-1 однодневных бройлерных цыплят общим количеством 150 делили на 15 групп (10 птиц/группу). Записывали массу тела каждого цыпленка. Цеплять делили на группы таким образом, чтобы средние массы групп не значительно различались между собой. Каждой птице на крыло вешали цветной ярлычок с номером согласно проводимому лечению и массе тела, записанных на соответствующем бланке.
Инфицирование
[0147] В День 14 четырнадцать групп животных G2-G15 инфицировали свежим культуральным бульоном вирулентного R-штамма М. gallisepticum, с титром, приблизительно равным 8.0 log10KОЕ/мл. Каждой из этих групп вводили десять мл этого свежего культурального бульона способом распыления. Вкратце, птиц помещали в изолирующий блок объемом 0.22 кубических метра. Затем распыляли десять мл свежей культуры R-штамма М. gallisepticum под давлением, равным 1 атмосфере, в течение приблизительно 100 секунд и оставляли цыплят в блоке на 20 минут (В лаборатории при применении этой методики в нескольких экспериментах получили положительные результаты).
Эвтаназия и патология
[0148] В Д28, в конце экспериментального исследования все группы подвергали эвтаназии. Проводили аутопсию каждой птицы и определяли показатели макроскопических поражений, связанных с MG. Регистрировали наличие экссудата в трахее, левом и правом грудных воздухоносных мешках и брюшной полости. Показатели поражений определяли по следующей системе:
В трахее: 0 = нет экссудата, 1 = легкое покраснение и небольшое количество экссудата, 2 = покраснение слизистой оболочки, экссудат.
Левый и правый воздухоносные мешки: 0 = нет поражений, 1 = сывороточный экссудат, 2 = сывороточный экссудат и небольшие скопления фибрина, 3 = сывороточный, фиброзный экссудаты, легкое утолщение стенки воздухоносного мешка, 4 = большое количество сывороточный экссудата, сильное утолщение стенки воздухоносного мешка.
Брюшная полость: 0 = нет экссудата, 1 = сывороточный экссудат, 2 = сывороточный экссудат с небольшими скоплениями фибрина, 3 = сывороточно-фиброзный экссудат.
Результаты
[0149] Результаты показаны в Таблице 3.
Выделение MG
[0150] Во время исследования аутопсийного материала асептически, с помощью ватного тампона, изолировали образцы трахеи, грудных воздухоносных мешков, печени, легкого, селезенки, почки и сердца. Материал с ватных тампонов высевали на агар для микоплазм (МА) и инкубировали при 37°С в инкубаторе с 5% CO2. Чашки исследовали на микоплазму в дни 2, 4 и 7, а затем с недельными интервалами в течение максимум трех недель. Положительные колонии определяли способом ПЦР, чтобы идентифицировать М. gallisepticum и М. synoviae.
Аутопсия и повторное выделение
[0151] После инфицирования MG выявляли значительные патологические поражения в воздухоносном мешке и брюшной полости. Однако в группах, которые вакцинировали обработанным очищенным антигеном плюс группы TLR, регистрировали значительное снижение показателей поражений и степеней повторного выделения, причем лучшие результаты по сравнению с контрольной группой, которой ничего не вводили, но инфицировали, были у антигена, очищенного на колонке с Con A (G9). Однако с Con А-элюированной фракцией G15 показатель вызванного патогеном поражения был высок, а степень повторного выделения была такой же, как в положительным контроле. Это приводит к заключению о том, что вакцинация этой фракцией антигена М. gallisepticum препятствовала адекватному иммунному ответу против инфицирования патогеном, вызывая иммуносупрессию и приведя к более выраженной инфекции.
Обсуждение
[0152] Результаты этих экспериментов показали, что удаление иммуносупрессивных антигенов из пула аффинно очищенных антигенов значительно улучшило защитное действие состава вакцины. Кроме того, химическая модификация или ферментативное расщепление сахаросодержащих посттранскрипционных модификаций на антигенах также привело к улучшению защитного иммунитета.
[0153] С другой стороны, мы обнаружили также, что введение выделенных иммуносупрессивных антигенов, присоединенных к микрочастицам, приводит к иммуносупрессии и развитию толерантности к патогену. Таким образом, мы продемонстрировали, что соответствующие свойства антигена могут сдвинуть иммунный ответ в сторону защиты или толерантности. Оба эти вида иммунитета обладают огромным значением в разработке рациональных подходов к модуляции иммунных ответов. Затем мы решили нацелить дополнительные поверхностные антигенные детерминанты, участвующие в развитии иммунных ответов.
ПРИМЕР 20
[0154] Аффинно очищенный антиген MG готовили, как описано в предыдущих Примерах. Перед иммобилизацией очищенный антиген разделяли на три группы для обработки следующими способами:
1. Деацилирование антигена MG
[0155] К 27 мл антигенов (приблизительно 0.65 мг) добавляли 8 мл 1 М NaOH и 10 мг пентадекановой кислоты, и осторожно взбалтывали раствор при 70°С в течение 45 мин. Затем рН доводили до 8.0 и удаляли преципитат центрифугированием при 3,000 об/мин в течение 5 минут. Этот образец обозначали как Deacylated Antigens (Деацилированные антигены).
2. Обработка перйодатом
[0156] По замыслу эта реакция должна была быть более жесткой, чем в Примере 18. К 25 мл антигенов (приблизительно 0.6 мг) добавляли твердый периодат натрия в конечной концентрации 80 мМ и после перемешивания выдерживали в течение 2.5 часов при комнатной температуре. Затем добавляли двукратный избыток глицерина и инкубировали образец еще один час. Проводили диализ против PBS в течение ночи. Диализованный образец обозначали Periodate Antigens (Антигены, обработанные перйодатом).
ПРИМЕР 21
Исследования на животных III
[0157] Результаты представляют собой среднее по 3 экспериментам. Вакцинировали трехдневных цыплят (не зараженные М. gallisepticum (MG) и М. synoviae (MS), у которых твердофазный иммуноферментный анализ не выявлял материнских антител MG и MS). Каждая группа состояла из 10 цыплят. На 14 день эксперимента цыплят инфицировали штаммом М. gallisepticum Rlow. Исследование завершали на 28 день.
[0158] Группы организовывали следующим образом:
G1 = не инфицировали и ничего не вводили
G2 = инфицировали и ничего не вводили
Вводили за 2 недели перед инфицированием:
G3 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным, адсорбированным СоnА антигеном М. gallisepticum (10 мкг/дозу)+агонисты TLR (10 мкг бактериальной ДНК, E.coli+2 мкг липополисахарида E.coli и 2 мкг пептидогликана/дозу) и инфицировали
G4 = перорально вводили микрочастицы (0.2 мл/цыпленка) аффинно очищенный деацилированный антиген М. gallisepticum (10 мкг/дозу)+агонисты TLR (10 мкг бактериальной ДНК, Е.coli+2 мкг липополисахарида Е.coli и 2 мкг пептидогликана/дозу) и инфицировали
G5 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным, адсорбированным СоnА антигеном М. gallisepticum (10 мкг)+TLR agonists (25 мкг поли(I:С)+10 мкг bacterial DNA and 2 мкг пептидогликана/дозу) и инфицировали
G6 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным, адсорбированным СоnА антигеном М. gallisepticum (10 мкг)+агонисты TLR (10 мкг поли(I:С)+10 мкг бактериальной ДНК и 2 мкг пептидогликана/дозу) и инфицировали.
Водили через 1 день после инфицирования:
G7 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенным, адсорбированным СоnА антигеном М. gallisepticum (10 мкг)+TLR agonists (25 мкг поли(I:С)+10 мкг бактериальной ДНК и 2 мкг пептидогликана/дозу)
G8 = перорально вводили микрочастицы (0.2 мл/цыпленка) с аффинно очищенный обработанным перйодатом и деацилированным антигеном М. gallisepticum (10 мкг)+агонисты TLR (25 мкг поли(I:С)+10 мкг бактериальной ДНК и 2 мкг пептидогликана/дозу) и инфицировали.
График
[0159] День-1: Формирование групп G1-G9. Умерщвление 10 цыплят для твердофазного иммуноферментного анализа ELISA, ПЦР и культивирования М. gallisepticum и М. synoviae для подтверждения того, что у подопытных цыплят нет материнских антител и отсутствует микоплазма.
[0160] День 0: Вакцинация G3-G6 перед инфицированием.
[0161] День 14: Инфицирование групп G2-G9.
[0162] День 15: Вакцинация групп G7-G9 после инфицированиия.
[0163] День 28: G1-G9. Эвтаназия, аутопсия и посев для выделения М. gallisepticum (MG) из различных органов, трахеи, воздухоносного мешка и легкого. Проводили гистологическое исследование трахеи и легкого.
[0164] Дни Д14 и Д28: У цыплят брали кровь, чтобы получить сыворотку для исследования на MG-специфические антитела путем проведения реакции агглютинации сыворотки на пластинке (SPA) и блокирующего твердофазного ИФА.
[0165] В Д-1 однодневных бройлерных цыплят общим количеством 90 делили на восемь групп (10 птиц/группу). Записывали массу тела каждого цыпленка. Цыплят делили на группы таким образом, чтобы средние массы групп незначительно различались между собой. Каждой птице на крыло вешали цветной ярлычок с номером, согласно проводимому лечению и массе и массе тела, записанных на соответствующем бланке.
Инфицирование
[0166] В день Д14 девять групп животных G2-G9 инфицировали свежим культуральным бульоном вирулентного R-штамма М. gallisepticum с титром, приблизительно равным 8.0 log10КОЕ/мл. Каждой из этих групп вводили десять мл этого свежего культурального бульона способом распыления. Вкратце, птиц помещали в изолирующий блок объемом 0.22 кубических метра. Затем распыляли десять мл свежей культуры R-штамма М. gallisepticum под давлением, равным 1 атмосфере, в течение приблизительно 100 секунд и оставляли цыплят в блоке на 20 минут.
Эвтаназия и патология
[0167] В Д28, в конце экспериментального исследования, все группы подвергали эвтаназии. Проводили аутопсию каждой птицы и определяли показатели макроскопических поражений, связанных с MG. Регистрировали наличие экссудата в трахее, левом и правом грудных воздухоносных мешках и брюшной полости. Показатели поражений определяли по следующей системе:
В трахее: 0 = нет экссудата, 1 = легкое покраснение и небольшое количество экссудата, 2 = покраснение слизистой оболочки, экссудат.
Левый и правый воздухоносные мешки: 0 = нет поражений, 1 = сывороточный экссудат, 2 = сывороточный экссудат и небольшие скопления фибрина, 3 = сывороточный, фиброзный экссудаты, легкое утолщение стенки воздухоносного мешка, 4 = большое количество сывороточного экссудата, сильное утолщение стенки воздухоносного мешка.
Брюшная полость: 0 = нет экссудата, 1 = сывороточный экссудат, 2 = сывороточный экссудат с небольшими скоплениями фибрина, 3 = сывороточно-фиброзный экссудат.
Результаты
[0168] Результаты показаны в Таблице 4.
Выделение MG
[0169] Во время исследования аутопсийного материала асептически, с помощью ватного тампона, изолировали образцы трахеи, грудных воздухоносных мешков, печени, легкого, селезенки и сердца. Материал с ватных тампонов высевали на агар для микоплазм (МА) и инкубировали при 37°С в инкубаторе с 5% CO2. Чашки исследовали на микоплазму в дни 2, 4 и 7, а затем с недельными интервалами в течение максимум трех недель. Положительные колонии исследовали ПЦР, чтобы идентифицировать М. gallisepticum и М. synoviae.
Аутопсия и повторное выделение
[0170] После инфицирования MG выявляли значительные патологические поражения в воздухоносном мешке и брюшной полости. Однако в группах, вакцинированных обработанным очищенным антигеном плюс группы TLR, регистрировали значительное снижение показателей поражений и степеней повторного выделения, при этом все группы давали сходные результаты, группа с деацилированным антигеном (G4) показала самый лучший результат по сравнению с контрольными группами, которые инфицировали, но которым ничего не вводили. В группах, которые вакцинировали после инфицирования, лучшие результаты давал антиген, обработанный Con А плюс TLR 3, 4, 9. По-видимому, эта композиция позволяет добиться защитной иммунной реакции даже после инфицирования.
Обсуждение
[0171] Результаты этих экспериментов показали, что удаление или блокирование иммуносупрессивных антигенов ускользания или антигенных детерминант от механизмов иммунной защиты из пула аффинно очищенных антигенов значительно улучшило защитное действие состава вакцины. Дополнительно, удаление и химические модификации сахаросодержащих посттранскрипционных модификаций (N-гликозилирование) на антигенах привело к улучшению защитного иммунитета. Кроме того, удаляли или блокировали химическими способами фрагменты липидов на антигенах, что также приводило к выраженному защитному иммунному эффекту.
[0172] Мы вновь продемонстрировали, что соответствующие модификации антигена позволяют сдвинуть иммунный ответ в сторону защиты. Это имеет огромное значение для разработки рациональных подходов к модулированию иммунных ответов. Затем мы разработали in vitro систему для воспроизведения соответствующих иммунных ответов в культуре ткани. Такая система может обладать прогностической ценностью в отношении иммунного ответа, вызываемого иммунологически активным составом, и, соответственно, может сократить цикл разработки.
ПРИМЕР 22
Исследования In vitro
[0173] Определяли секрецию TNF-α в мононуклеарных клеток периферической крови, выделенных из периферической крови и обработанных пептидогликаном (PepG) (Sigma, Сент-Луис, Мичиган, США), бактериальной CpG-ДНК (KM Biomedicals, Аврора, Огайо, США; липополисахариды<0.2ЭЕ/мл, согласно анализу с лизатом амебоцитов краба Limulus), или и тем и другим. TNF-α определяли путем твердофазного иммуноферментного анализа ELISA надосадочной жидкости культуры после 5 часов инкубации.
[0174] Тканевой фактор (ТФ) в моноцитах, представляющих систему врожденного иммунитета, оценивали в соответствующих культурах МНПК, которые использовали для анализа TNF-α. ТФ экстрагировали из адгезивных клеток детергентом Triton-Х100, следуя рекомендациям производителя набора ИФА для ТФ (Imubind kit, American Diagnostica, Гринвич, Коннектикут, США).
[0175] Результаты показаны на Фигуре 2 (числа после Р и D указывают концентрации пептидогликана и бактериальной ДНК в мкг/мл, соответственно, слева направо это P3+D15, P3+D5, P3+D1.5, P1+D15, P1+D5, P1+D1.5, P0.3+D15, P0.3+D5, и P0.3+D1.5). Секретируемый мононуклеарными клетками периферической крови TNF-α (А) и Тканевой Фактор (В) после обработки пептидогликаном (белый столбик), или бактериальной CpG-ДНК (черный столбик), или обоими одновременно в той же концентрации, в которой их вводили по отдельности (заштрихованный столбик). Числа после Р и D, соответственно, показывают концентрации пептидогликана и бактериальной ДНК в мкг/мл соотвтетственно. Р<0.05 для TNF-α и ТФ при сравнении суммы эффектов пептидогликана (3 или 1.5 мг/мл) и CpG-ДНК отдельно во всех исследованных концентрациях, и при совместном добавлении пептидогликана и бактериальной ДНК; t-критерий Стьюдента (n=4). Представлен один типичный эксперимент из трех.
[0176] Эти результаты демонстрируют сопутствующую стимуляцию TNF-α и ТФ пептидогликаном и бактериальной ДНК в МНПК и синергистическое действие этих двух молекул. В это могут быть вовлечены прямые эффекты на общих участников передачи сигналлов, или непрямые эффекты, опосредуемые соединениями, секретируемыми под действием пептидогликана или бактериальной ДНК. TNF-α - важный ранний медиатор ответа хозяина на пирогены и единственный эндогенный медиатор, способный запустить весь спектр метаблических, гемодинамических, тканевых и цитокиновых каскадных реакций септического шока. Синергистическое действие бактериальной ДНК и пептидогликана имеет значение для патогенеза сепсиса, где каждая молекула, хотя и присутствует in vivo в малых концентрациях, по-видимому усиливает эффект другой, как предполагали ранее.
Демонстрация стимуляции in vitro различных ответов Тh1 и Тh2 различными составами частиц
[0177] Мононуклеарные клетки периферической крови готовили путем выделения фиколлом из периферической крови здоровых добровольцев. Мононуклеарные клетки высевали на чашки при плотности 106 клеток/мл в 1.5 мл среды RPMI-1640 (Irvine Scientific, Ирвин, Калифорния, США), дополненной 100 Ед/мл смеси пенициллин/стрептомицин, 2 мМ глютамина и 3% фетальной сыворотки крупного рогатого скота, и инкубировали с 0.15 мл микрочастиц (в стерильном PBS) в течение 18 ч. Микрочастицы содержали очищенные СоnА или обработанные перйодатом натрия антигены, а также агонисты TLR 2, 3, 4 и 9, как показано на приведенным ниже графике. Приготовление этих гранул описано в Примере 19.
[0178] Результаты показаны на Фигуре 3 (серый столбик, TNF-α×100, черный столбик, IL-10) и на Фигуре 4, где приведены соотношения TNF-α/IL-10.
[0179] Анализ отношения TNF-α и IL-10 продемонстрировал, что очищенный Con А антиген иммобилизованный вместе с TLR 2, 4, 9 давал наиболее выраженный ответ Тh1, что, как оказалось, коррелирует с полученными in vivo результатами (см. Пример 19). Эти составы стабильно давали самый высокий эффект защиты животных от инфицирования микоплазмой.
ПРИМЕР 23
[0180] Чтобы далее описать клеточный ответ на микрочастицы с иммуномодулятором, мы подвергли дендритные клетки действию этих микрочастиц. Мы наблюдали активацию молекул МНС I и МНС II и костимуляторных молекул CD80 и CD86. Видимо, присутствие любой комбинации с агонистом PRR необходимо для увеличения количества МНС II на дендритных клетках после 18 часов обработки. Поскольку присутствие МНС на поверхности клеток представляет собой динамический процесс, для того чтобы наблюдать увеличение числа молекул ГКГС на поверхности клетки, может быть необходимо различное время инкубации с гранулами СоnА. МНС обеспечивает презентацию антигенов Т-клеткам, и CD86 взаимодействует с CD28 на Т-клетках. Взаимодействие сопутствующей презентации антигена дендритных клеток и взаимодействие В7-костимуляторного лиганда (такого как CD80 и CD86) приводит к активации Т-клеток. Эти данные in vitro указывают на то, что микрочастицы стимулируют изменения, которые представляют собой ключевые моменты созревания дендритных клеток и которые необходимы для стимуляции врожденного и приобретенного иммунитета.
[0181] Определяли экспрессию МНС I и МНС II, а также CD80 и CD86, на поверхности дендритных клеток после 18 часов воздействия различных препаратов микрочастиц: контрольные микрочастицы (контроль), на которых не иммобилизовали ни антиген, ни агонист PRR, и микрочастицы, на которых иммобилизовали антигены М. gallisepticum, очищенные от СоnА-связывающих антигенов (СоnА АГ) со следующей комбинацией агонистов PRR или без нее: 1) пептидогликан (PepG), липополисахарид (LPS), бактериальная ДНК (bactDNA) (СоnА АГ+PRR 2, 4, 9) или 2) поли I:С, липополисахарид, бактериальная ДНК (СоnА АГ+PRR 3, 4, 9). Стимуляция MHC-II и CD86 в дендритных клетках показана на Фигуре 5.
[0182] Результаты показывают действие микрочастиц на секрецию TNF-α дендритными клетками. Контрольные частицы не вызывали детектируемой секреции TNF-α в кондиционированной среде. Три другие типа микрочастиц (антигены М. gallisepticum после удаления СоnА-связывабщих антигенов и аналогичные антигены соиммобилизованные с пептидогликаном, липополисахаридом, бактериальной ДНК и поли(I:С)) стимулировали секрецию значительного количества TNF-α дендритными клетками по сравнению с контрольными микрочастицами. Интересно, что в присутствии соединений бактериального происхождения в качестве иммуномодулятора образовывалось меньше TNF-α, чем только с антигенами. Эти результаты указывают на то, что микрочастицы действительно взаимодействуют с дендритными клетками и способны активировать рецепторы TLR и NOD, вызывая физиологическую реакцию, соответствующую известному действию этих молекул на клетки иммунной системы. Мы продемонстрировали, что мишенью микрочастиц могут быть дендритные клетки.
ПРИМЕР 24
Выделение гепарин-связывающих белков из MG
[0183] М gallisepticum выращивали, как описано в Примере 2, концентрированную и промытую микоплазму инактивировали, как описано в этапе 1 Примера 4. Вкратце, к 75 мл неочищенного белка мембран MG добавляли 0.75 Mega-10 и перемешивали раствор в течение 20 часов при комнатной температуре. Затем к раствору добавляли 3.75 мл Triton X-100 и перемешивали еще один час при комнатной температуре. Раствор центрифугировали при 5,000 об/мин в течение 10 минут и отделяли надосадочную жидкость от осадка. Затем к надосадочной жидкости добавляли 75 мл буфера Tris (20 мМ Tris, 0.1M NaCl, pH 7.2). Колонку со смолой Heparin-Actigel уравновешивали 5 объемам буфера Tris при скорости потока 3 мл/мин. В колонку загружали раствор белка из MG при скорости потока 8-15 мл/мин и собирали протекающую жидкость в отдельную бутылку. Колонку промывали 10 объемами буфера Tris при скорости потока 8-15 мл/мин и элюировали буфером Tris, содержащим 2 М NaCl, при скорости потока 8-15 мл/мин. После промывки колонки 10 объемами буфера Tris на колонку снова наносили раствор белка из MG и повторяли элюирование, как описано выше. Собирали элюаты и диализировали их против буфера Tris при помощи диализной трубки, пропускающей молекулы с молекулярной массой 3,500 Да. Для того чтобы определить концентрацию элюированных белков, проводили количественный белковый анализ.
ПРИМЕР 25
Выделение IgG из нейтрализующей сыворотки и подготовка иммуноаффинной колонки
[0184] Тотальное выделение IgG из нейтрализующей сыворотки вакцинированных цыплят и иммобилизацию на Actigel ALD проводили согласно Примеру 3.
ПРИМЕР 26
Расщепление гепарин-связывающих белков MG, связанных в иммуноаффинной колонке, трипсином
[0185] Два мг очищенного белка MG добавляли на 2 мл иммуноаффинную колонку и оставляли связываться на 15 мин. Колонку промывали 10 мл Tris буфера В (50 мМ Tris, pH 7.5, 0.1М NaCl). Добавляли раствор трипсина (20 мкг/мл в Tris буфере В) и осторожно встряхивали смесь в течение 1 ч при 37°С. Фрагменты и свободный фермент отмывали Tris буфером В и элюировали связанные белки 0.1 М цитратом, pH 2.5. Затем элюат нейтрализовали NH4OHконц., концентрировали и очищали на фильтрационной обращенно-фазовой колонке с сорбентом С-18 (Pierce) и проводили анализ белкового состава путем масс-спектрометрии MALDI-TOF.
ПРИМЕР 27
Расщепление трипсином гепарин-связывающих белков из MG, связавшихся с гепариновой колонкой
[0186] Два мг очищенного белка из MG добавляли на 2 мл колонку Heparin Actigel и оставляли связываться на 1 час. Колонку промывали 10 мл Tris буфера В. Добавляли раствор трипсина (20 мкг/мл в Tris буфере В) и аккуратно встряхивали суспензию в течение 1 ч при 37°С. Фрагменты смывали Tris буфером В и элюировали связавшиеся пептиды Tris буфером В, содержащим 1 М NaCl. Элюат концентрировали и очищали на фильтрационной обращенно-фазовой колонке с сорбентом С-18 (Pierce) и проводили анализ белкового состава путем масс-спектрометрии MALDI-TOF.
ПРИМЕР 28
Очистка и иммобилизация белка C1q
[0187] Очистку C1q проводили, придерживаясь способа McKay, EJ:
простая двухэтапная процедура выделения C1q из плазмы различных видов животных. Immunol Letters 1981, 3:303-308. Очищенный C1q иммобилизовали на Aminogel (Sterogene Bioseparations, Inc., Карлсбад, Калифорния, США) в концентрации 3 мг/мл, с модификацией нашей процедуры, описанной в Патенте США 5,801,063.
ПРИМЕР 29
Очистка и иммобилизация комплемент-связыающих IgG
[0188] Поликлональные нейтрализующие антитела, очищенные согласно Примеру 24, наносили на 10 мл колонку с иммобилизованым C1q, уравновешенную Tris буфером В и оставляли связываться в течение 30 мин. Несвязанные антитела удаляли путем промывки 10 объемами Tris буфера В и элюировали связанные антитела тем же буфером, содержащим 1 М NaCl. Затем очищенные IgG иммобилизовали на Actigel ALD в концентрации 3 мг/мл.
ПРИМЕР 30
Расщепление трипсином активирующих комплемент белков из MG, связанных на иммуноаффинной колонке.
[0189] М gallisepticum выращивали, как описано в Примере 2, концентрированную и промытую микоплазму инактивировали, как описано в этапе 1 Примера 4. На 10 мл колонку с иммобилизованным антителом, активирующим комплемент, наносили 30 мл лизата MG при скорости потока 3 мл/мин. Колонку промывали 20 объемами слоя буфера Tris В при скорости потока 8 мл/мин, чтобы удалить неспецифически адсорбированные белки. Добавляли раствор трипсина (20 мкг/мл в Tris буфере В) и аккуратно встряхивали суспензию в течение 1 ч при 37°С. Фрагменты удаляли путем промывки Tris буфером В и элюировали связанные белки 0.1 М цитратом, рН 2.5. Затем элюат нейтрализовали NH4OH конц., концентрировали и очищали на фильтрационной обращенно-фазовой колонке с сорбентом С-18 (Pierce) и проводили анализ белкового состава путем масс-спектрометрии MALDI-TOF.
[0190] Показано, что пример иммунологически активного пептида имеет следующий состав: Lys-Leu-Ala-Leu-Thr-Ser-Glu-lle-Thr-Glu-Glu-Ile-Tyr-Pro-Ser-Ala-Pro-Lys-Val-Ser-Arg-Lys-Gln-Arg-Gly-Val-His-Gly-Phe-Ser-Glu-Pro-Thr-Ser (SEQ ID NO:1). Важно, что этот пептид содержит GAG/гепаринсвязывающий домен Arg-Lys-Gln-Arg. Другая полезная последовательность - Leu-Leu-Ala-Lys-Lys-Thr-Asp-Lys-Ser-Val-Ser-Pro-Gln-Ala-Ser-Leu-Thr (SEQ ID NO:2). Эти последовательности можно соединить пептидным линкером, который содержит сайты расщепления для эндопротеаз. Пример такой линкерной последовательности - LIKFRSN (Leu-Ile-Lys-Phe-Arg-Ser-Asn) (SEQ ID NO:3). В данной области известны и другие линкеры.
ПРИМЕР 31
Синтез эпитопов антигенных пептидов и пептидов с несколькими эпитопами
[0191] Антигенные пептиды синтезировали способом FMOC.
ПРИМЕР 32
Апробация защитных антигенных пептидов для вакцины
[0192] Антигенные пептиды смешивали с нейтрализующей сывороткой вакцинированных цыплят и способом конкурентного твердофазного иммуноферментного анализа определяли ингибирование связывания с иммобилизованным гепарином. Пробу пептида адсорбируют в лунках планшета для ИФА. Затем лунки блокируют блокирующим раствором, а затем добавляют в лунки иммунную и неиммунную контрольную антисыворотку и инкубируют в течение ночи при 4°С. Неспецифические белки удаляют промывочным раствором, после чего добавляют меченый биотином гепарин в концентрации 100 мкг/мл, разведенный на блокирующем буфере, и оставляют на 1 ч. После 3-кратной отмывки (каждая продолжительностью 10 минут) от несвязанного гепарина добавляют стрептавидин-пероксидазу, разведенную в отношении 1:3,000 на блокирующем буфере, и оставляют на 1 ч. Избыток конъюгата удаляют, промывая 3 раза по 10 минут промывочным буфером. Добиваются появления окраски добавлением раствора 3,3'-диаминобензидина. Сигнал обратно пропорционален содержанию нейтрализующих антител.
ПРИМЕР 33
[0193] Анализ способами биоинформатики антигенных белков патогенов также может обеспечить выявление эпитопов с высоким антигенным потенциалом. Такой анализ провели для белка MGA M. gallisepticum. Затем антигенные участки анализировали в контексте линейных основных мотивов. Примером такого предсказанного высокоантигенного линейного мотива является последовательность: эпитоп МНС I представляет собой 10-мер LLAKKTDKSV (SEQ ID NO:4), a MHC II представляет собой 15-мер LLAKKTDKSVSPAQAS (SEQ ID NO:5). Эти домены идентифицировали способом SYFPEITHI (www.syfpeithi.de). Это указывает на то, что линейные основные мотивы на самом деле включены в участки с высокой антигенной способностью. Анализ способами биоинформатики можно проводить по методикам, известным в данной области, например, описанным в J. Pevsner, "Bioinformatics and Functional Genomics" («Биоинформатика и функциональная геномика» Wiley-Liss, Hoboken, N.J., 2003), используя базы данных и методики компьютерного анализа для извлечения данных и поиска совпадений и сравнения последовательностей, известные в данной области.
ПРИМЕР 34
Очистка антител против эпитопов
[0194] Идентифицированные антигенные пептиды синтезировали с биотиновой меткой на конце. Пептиды с биотиновой меткой иммобилизовали в концентрации 1 мг/мл на Avidin Actigel (Sterogene Bioseparations, Inc. Карлсбад, Калифорния, США). Добавляли нейтрализующую антисыворотку из вакцинированных цыплят до насыщения и удаляли неспецифически связанные белки путем промывки фосфатным буферным раствором (PBS), рН 7.2. Специфически связанное антитело элюировали средой Actisep Elution Medium (Sterogene Bioseparations, Inc. Карлсбад, Калифорния, США).
ПРЕИМУЩЕСТВА ИЗОБРЕТЕНИЯ
[0195] Настоящее изобретение обеспечивает улучшенные иммуномодулирующие составы и способы, которые можно разработать, например, для стимуляции иммунного ответа, для того, чтобы вызвать защитный иммунный ответ, или чтобы вызвать толерантность. Эти составы стабильны и их можно приготовить с широким диапазоном иммуногенов и молекул-мишеней, чтобы вызвать желаемое воздействие на иммунный ответ. Они обеспечивают дополнительные пути введения (кроме парентерального) для таких агентов. Они предотвращают преждевременное расщепление или высвобождение иммуногенов и нацеливающих молекул. Частицы обладают мукоадгезивными свойствами, что улучшает их взаимодействие со слизистыми оболочками и облегчает поглащение. Они не требуют дополнительных адъювантов.
[0196] Способы и составы согласно настоящему изобретению основаны на роли N-гликозилирования (маннозилирования) в предотвращении ускользания от механизмов иммуннологического контроля путем устранения соответствующих эпитопов ускользания из состава, предназначенного для применения в вакцинах. В них также задействована идентификация линейных мотивов, связывающих глюкозаминогликан/гепарин, и способы, препятствующие такому связыванию. Способы и композиции согласно настоящему изобретению также задействуют способ, который был разработан для идентификации эпитопов, активирующих комплемент. Кроме того, способы и составы согласно настоящему изобретению применяют знание того факта, что применение нескольких агонистов TLR значительно усиливает иммунный ответ. Все антигены и агонисты TLR могут быть нацелены на одну клетку лимфоидной системы слизистой оболочки. Другой улучшенный аспект настоящего изобретения представляет собой применение синтетических эпитопов Т-клеток и В-клеток на одной частице.
[0197] Указанные изобретения, иллюстративно описанные в данной заявке, можно осуществить при отсутствии любого элемента или элементов, ограничения или ограничений, не раскрытых здесь конкретно. Так, например, термины "содержащий", "включающий", "состоящий из" и т.д. следует понимать широко и без ограничений. Кроме того, употребляемые здесь термины и выражения представляют собой термины описания, но не ограничения. Такие термины и выражения не предполагают исключения какого-либо возможного эквивалента, показанного и описанного, или любой его части. Подразумевается, что в пределах объема данного изобретения возможны различные модификации. Следовательно, необходимо понимать, что хотя настоящее изобретение было подробно раскрыто через предпочтительные способы реализации и факультативные свойства, специалист в данной области может обратиться к модификациям и вариациям изобретения, и такие модификации и вариации входят в объем раскрытого в настоящей заявке изобретения. Изобретения описаны в настоящей заявке ориентировочно и в общих чертах. Каждый из более узких видов и субродовых комбинаций, соответствующих общему описанию каждого изобретения, также является частью этих изобретений. Это включает общее описание каждого изобретения с оговоркой или отрицательным ограничением, исключающим из рода любой объект, вне зависимости от того, содержит ли он исключенные материалы.
[0198] Кроме того, если свойства или аспекты изобретения описаны посредством формулы Маркуша(Markush), специалист в данной области поймет, что настоящее изобретение можно, таким образом, описать в терминах любого отдельного члена или плодгруппы членов формулы Маркуша. Также следует понимать, что приведенное выше описание является лишь иллюстративным, но не лимитирующим. После ознакомления с описанием для специалиста в данной области станут ясны многие способы реализации настоящего изобретения. Следовательно, объем настоящего изобретения, таким образом, определен не приведенным выше описанием, а прилагающейся формулой изобретения, вместе с полным объемом эквивалентов, которые относятся к указанной формуле. Раскрытия всех статей и источников, включая патентные публикации, включены в данную заявку посредством ссылок.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОСТИМУЛИРУЮЩАЯ КОМБИНАЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ГЕПАТИТА С | 2006 |
|
RU2431499C2 |
| УЛУЧШЕННЫЙ ИММУНОЛОГИЧЕСКИЙ СОСТАВ | 2012 |
|
RU2664730C2 |
| ОПРЕДЕЛЕНИЕ ДОЗЫ ДЛЯ ИММУНОТЕРАПЕВТИЧЕСКИХ СРЕДСТВ | 2017 |
|
RU2771717C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ВАКЦИНЫ | 2010 |
|
RU2444570C1 |
| ПРИМЕНЕНИЕ МОНОМИКОЛИЛГЛИЦЕРИНА (MMG) В КАЧЕСТВЕ АДЪЮВАНТА | 2008 |
|
RU2479317C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ АДЪЮВАНТНЫХ ВИРОСОМ И АДЪЮВАНТНЫЕ ВИРОСОМЫ, ПОЛУЧАЕМЫЕ УКАЗАННЫМИ СПОСОБАМИ | 2014 |
|
RU2694367C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ КОМПОЗИЦИИ | 2016 |
|
RU2732118C2 |
| ФУНКЦИОНАЛЬНО РЕКОНСТРУИРОВАННЫЕ ВИРУСНЫЕ МЕМБРАНЫ, СОДЕРЖАЩИЕ АДЪЮВАНТ | 2004 |
|
RU2348428C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРЕЗЕНТАЦИИ МНОЖЕСТВЕННЫХ АНТИГЕНОВ, ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ И ПРИМЕНЕНИЯ | 2012 |
|
RU2619176C2 |
| РАСШИРЕНИЕ СПЕКТРА Т КЛЕТОК ДЛЯ ВКЛЮЧЕНИЯ СУБДОМИНАНТНЫХ ЭПИТОПОВ ПУТЕМ ВАКЦИНАЦИИ С АНТИГЕНАМИ, ДОСТАВЛЕННЫМИ КАК БЕЛКОВЫЕ ФРАГМЕНТЫ ИЛИ ПЕПТИДНЫЕ КОКТЕЙЛИ | 2007 |
|
RU2490024C2 |


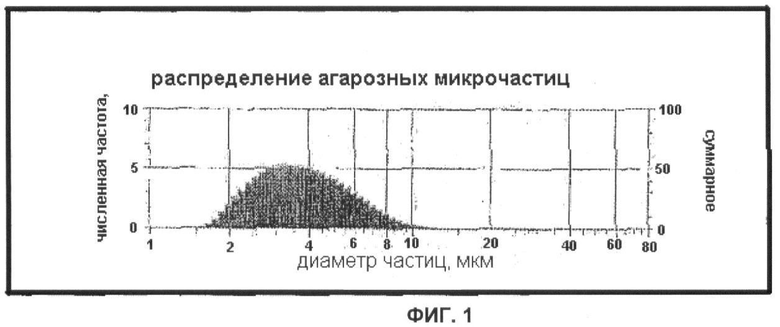
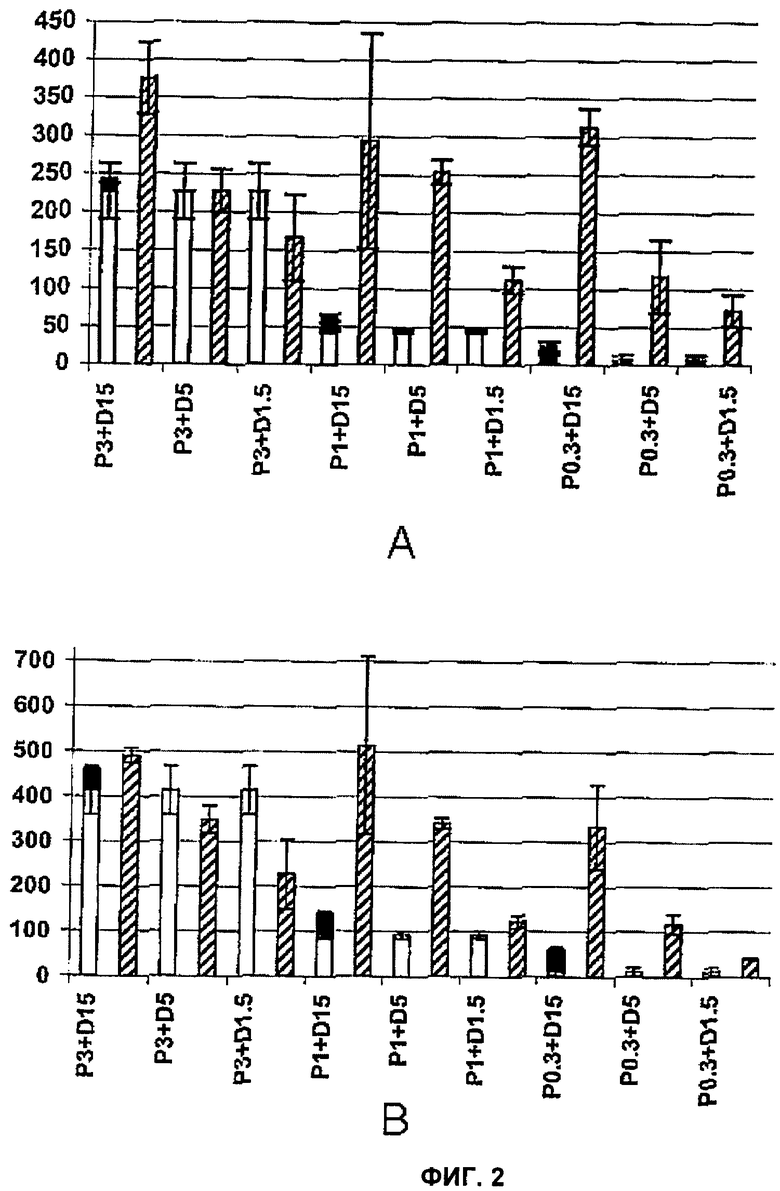
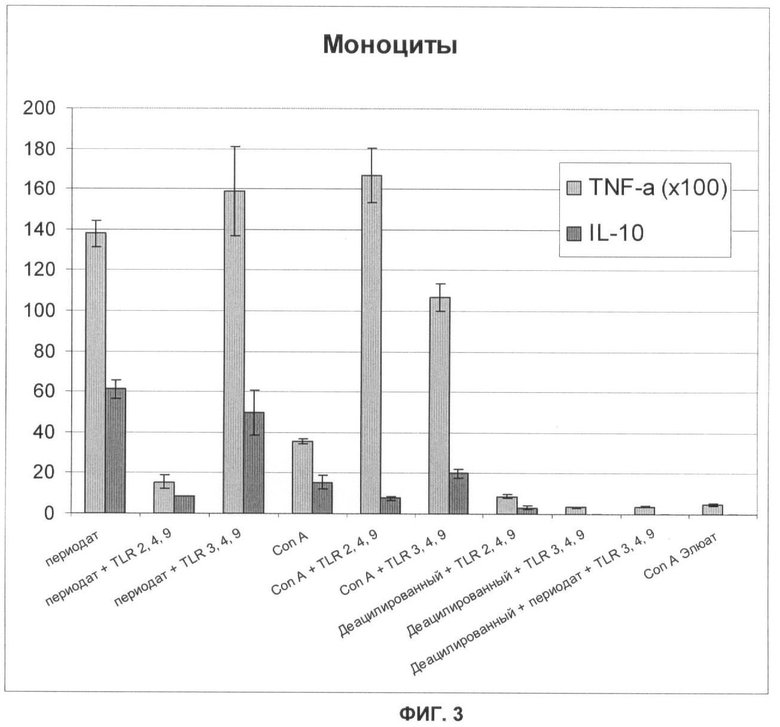
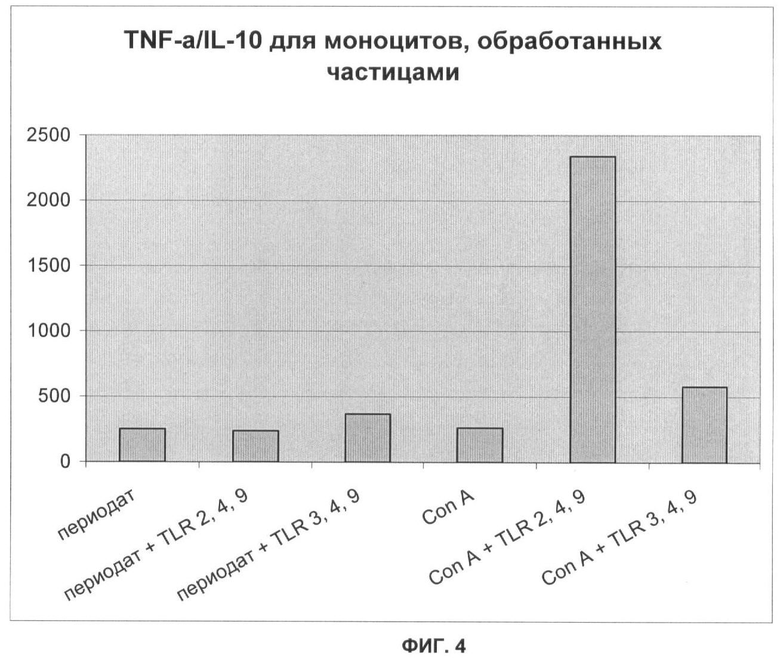
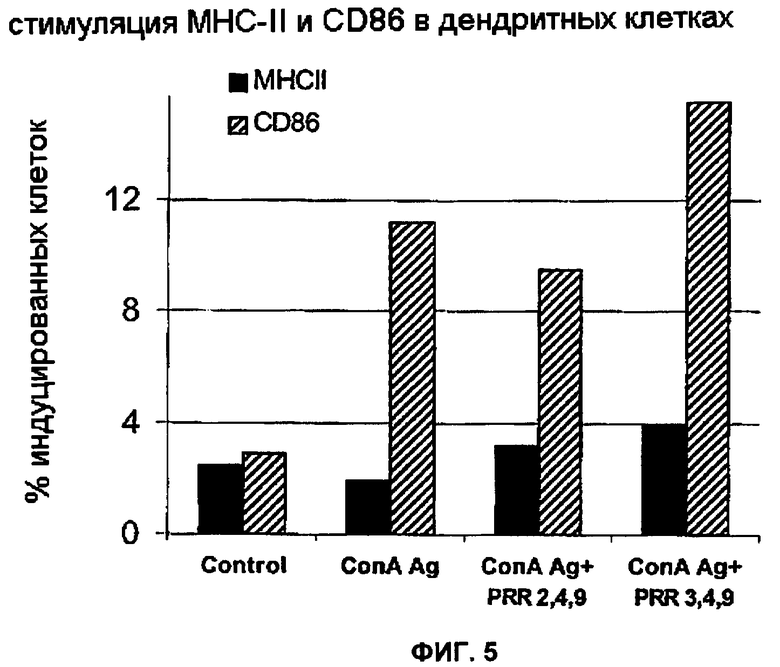
Изобретение описывает систему микрочастиц-носителей, содержащих один или более белков, пептидов, нуклеиновых кислот, углеводов, липидов или других биологически активных веществ с присоединенными нацеливающими молекулами или без них. В изобретении раскрыты иммуномодулирующие составы для индукции иммуномодулирующего эффекта - защитного иммунитета и толерантности. Иммунологически активный состав содержит ассоциированную с патогеном молекулярную структуру и носитель в виде микрочастиц агарозы. Молекулярная структура представляет по меньшей мере иммунологически активный антиген или эпитоп антигена, причем эпитоп антигена - это пептид, белок, рекомбинантный пептид или мультипептид, липид, углевод, нуклеиновая кислота или их комбинации. Описаны способы идентификации иммунологически активных пептидов для приготовления иммунологически активных составов и способы индукции толерантного иммунного ответа и обеспечения иммунитета с использованием описанных в изобретении составов. Изобретение обеспечивает стабильные составы с широким диапазоном иммуногенов и молекул-мишеней, позволяет стимулировать защитный иммунитет и толерантность как у инфицированных, так и у неинфицированных хозяев. 9 н. и 43 з.п. ф-лы, 5 ил., 4 табл., 34 пр.
1. Иммунологически активный состав для индукции иммуномодулирующего эффекта, выбранного из группы, состоящей из защитного иммунного ответа и толерантности, содержащий:
компонент А, содержащий
(a) по меньшей мере одну ассоциированную с патогеном молекулярную структуру и
(b) по меньшей мере один иммунологически активный антиген или эпитоп антигена;
компонент В, содержащий
по меньшей мере один носитель в виде микрочастиц агарозы, способный к эффективной доставке состава в организм с обеспечением ковалентного связывания по меньшей мере одной ассоциированной с патогеном молекулярной структуры и по меньшей мере одного иммунологически активного антигена или эпитопа антигена с микрочастицами агарозы с осуществлением индукции иммуномодулирующего состояния, выбранного из группы, состоящей из защитного иммунного ответа и толерантности, за счет взаимодействия ассоциированной с патогеном молекулярной структуры и по меньшей мере одного иммунологически активного антигена или эпитопа антигена в составе с клетками иммунной системы,
при этом индуцируемые защитный иммунный ответ или толерантность специфичны в отношении используемой в указанном составе ассоциированной с патогеном молекулярной структуры, а также в отношении иммунологически активного антигена или эпитопа антигена.
2. Состав по п.1, отличающийся тем, что дополнительно содержит по меньшей мере один иммунологически активный антиген или эпитоп антигена.
3. Состав по п.2, отличающийся тем, что указанный по меньшей мере один иммунологически активный антиген или эпитоп антигена представляет собой эпитоп антигена, причем указанный эпитоп антигена не является эпитопом ускользания.
4. Состав по любому из пп.2 или 3, отличающийся тем, что указанный по меньшей мере один иммунологически активный антиген или эпитоп антигена представляет собой эпитоп антигена, и при этом указанный эпитоп антигена представляет собой пептид, белок, рекомбинантный пептид или мультипептид, рекомбинантный белок, липид, углевод, нуклеиновую кислоту или другую биологически активную молекулу, либо комбинацию любых из указанных молекул.
5. Состав по п.2, отличающийся тем, что указанный по меньшей мере один иммунологически активный антиген или эпитоп антигена представляет собой эпитоп антигена, и при этом указанный иммунологически активный эпитоп антигена представляет собой пептид или белок, причем указанный пептид или белок содержит иммуномодулирующие мотивы, которые имеют гидрофобную природу, и к которым присоединены одна или более дополнительных молекул, причем указанные дополнительные молекулы, присоединенные к иммуномодулирующим мотивам, обладают способностью взаимодействовать с рецептором, представляющим собой рецептор, опосредующий попадание в клетку.
6. Состав по п.2, отличающийся тем, что указанный по меньшей мере один иммунологически активный антиген или эпитоп антигена представляет собой эпитоп антигена, причем указанный иммунологически активный эпитоп антигена представляет собой пептид или белок, причем указанный пептид или белок содержит иммуномодулирующие посттранскрипционные модификации.
7. Состав по п.6, отличающийся тем, что иммуномодулирующие посттранскрипционные модификации выбраны из группы, состоящей из:
(a) посттранскрипционных модификаций, затрагивающих фрагменты углеводов, и
(b) посттранскрипционных модификаций, затрагивающих фрагменты липидов, поддающихся удалению путем делипидации.
8. Состав по п.7, отличающийся тем, что посттранскрипционные модификации, затрагивающие фрагменты углеводов, включают посттранскрипционное маннозилирование, причем терминально маннозилированные иммуномодулирующие вещества могут быть удалены из состава способом, выбранным из группы, включающей: окисление, обработку ферментами и сахарспецифичное связывание.
9. Состав по п.2, отличающийся тем, что иммунологически активный антиген или эпитоп антигена представляет собой иммунологически активный эпитоп антигена, и при этом указанный иммунологически активный эпитоп антигена представляет собой пептид или белок, не содержащий иммуномодулирующих посттранскрипционных модификаций.
10. Состав по п.2, отличающийся тем, что иммунологически активный антиген или эпитоп антигена представляет собой иммунологически активный эпитоп антигена, при этом указанный иммунологически активный эпитоп антигена представляет собой пептид или белок, причем указанный иммунологически активный пептид или белок не содержит последовательности аминокислот, по которым может происходить N-гликозилирование и/или присоединение липида.
11. Состав по любому из пп.2 и 3, отличающийся тем, что иммунологически активный антиген или эпитоп антигена представляет собой иммунологически активный эпитоп антигена, при этом указанный иммунологически активный эпитоп антигена представляет собой пептид или белок, причем указанный иммунологически активный пептид или белок содержит последовательности аминокислот, способные связываться с фрагментом, выбранным из группы, состоящей из: рецептора клеточной поверхности, опосредующим попадание в клетку, и гликозаминогликанов (GAGs) клеточной поверхности, причем указанная последовательность аминокислот имеет многоосновную природу и соответствует общей формуле ХВВХВХ, ХВВВХХВХ, ВВХХВВВХХВВ, ВВВХХВ, ВХВХВ, ВВВ, ВХВХХХВХВ или ВХВХХХХХВХВ, где В обозначает основную аминокислоту, а Х любую другую аминокислоту.
12. Состав по п.11, отличающийся тем, что иммунологически активный пептид или белок содержит последовательности аминокислот, способные связываться с гликозаминогликанами (GAG) клеточной поверхности, и при этом GAG-связывающие последовательности аминокислот применяют для получения антител к пептиду или белку, которые способны препятствовать связыванию патогена с поверхностью клетки.
13. Состав по п.2, отличающийся тем, что иммунологически активный антиген или эпитоп антигена представляет собой иммунологически активный эпитоп антигена, и при этом указанный иммунологически активный эпитоп антигена применяют для получения антител к указанному эпитопу, которые способны препятствовать связыванию патогена со структурой, выбранной из группы, состоящей из поверхности клетки и рецепторов, опосредующих попадание в клетку.
14. Состав по п.2, отличающийся тем, что иммунологически активный антиген или эпитоп антигена представляет собой эпитоп антигена, и при этом указанный иммунологически активный эпитоп антигена применяют для получения антител к указанному эпитопу, которые способны препятствовать GAG-опосредованному связыванию патогена со структурами, выбранными из группы, состоящей из поверхности клетки и рецепторов, опосредующих попадание в клетку.
15. Состав по любому из пп.13 и 14, отличающийся тем, что рецептор, опосредующий попадание в клетку, представляет собой рецептор, отвечающий за ускользание от иммунной системы.
16. Состав по любому из пп.13 и 14, отличающийся тем, что рецептор, опосредующий попадание в клетку, представляет собой DC-SIGN.
17. Состав по п.11, отличающийся тем, что иммунологически активный пептид или белок содержит последовательность аминокислот, способную связывать гликозаминогликан, выбранный из группы, состоящей из гепарина и его аналогов.
18. Состав по п.11, отличающийся тем, что иммунологически активный пептид или белок обладает комплемент-активирующей активностью самостоятельно или в комбинации с антителами.
19. Состав по п.2, отличающийся тем, что иммунологически активный антиген или эпитоп антигена представляет собой эпитоп антигена, и при этом указанный иммунологически активный эпитоп антигена представляет собой множество пептидов, которые соединены в единый мультипептид.
20. Состав по п.2, обладающий по меньшей мере одним из следующих свойств или видов активности:
(a) иммунологически активный эпитоп антигена применяют для индукции антител, способных препятствовать попаданию патогена в клетку;
(b) иммунологически активный эпитоп антигена применяют для индукции антител, способных связывать и активировать комплемент;
(c) иммунологически активный эпитоп антигена содержит по отдельности или вместе эпитопы Т- и В-клеток;
(d) иммунологически активный эпитоп антигена представляет собой пептид или белок, способный вызывать образование антител, которые способны препятствовать связыванию последовательностей N-гликозилирования и/или присоединения липидов с их рецепторами в организме хозяина, причем указанные рецепторы могут представлять собой DC-SIGN или L-SIGN, и при этом нарушение связывания последовательностей N-гликозилирования и/или присоединения липидов с соответствующими рецепторами предотвращает или уменьшает образование IL-10.
21. Состав по любому из пп.1 и 3, отличающийся тем, что ассоциированную с патогеном молекулярную структуру выбирают из группы, включающей:
(a) агонист рецептора TLR 1;
(b) агонист рецептора TLR 2;
(c) агонист рецептора TLR 3;
(d) агонист рецептора TLR 4;
(e) агонист рецептора TLR 5;
(f) агонист рецептора TLR 6;
(g) агонист рецептора TLR 7;
(h) агонист рецептора TLR 8;
(i) агонист рецептора TLR 9;
(j) агонист NOD-1;
(k) агонист NOD-2;
(l) DC-SIGN;
(m) L-SIGN; и
(n) рецептор маннозы,
причем в случае, если ассоциированная с патогеном молекулярная структура представляет собой агонист NOD-1 или агонист NOD-2, указанный агонист NOD-1 или агонист NOD-2 выбран из группы, включающей бактериальный пептидогликан и производное бактериального пептидогликана.
22. Состав по любому из пп.1-3, отличающийся тем, что микрочастицы агарозы имеют одно из следующих свойств:
(a) микрочастицы имеют узкий диапазон распределения по размерам;
(b) микрочастицы являются пористыми;
(c) микрочастицы имеют диаметр меньше приблизительно 10 мкм;
(d) микрочастицы связаны с более чем одним иммунологически активным эпитопом, причем указанный более чем один иммунологически активный эпитоп может содержать по отдельности или вместе эпитопы Т- и В-клеток, и
(e) микрочастицы связаны с более чем одной ассоциированной с патогеном молекулярной структурой, и при этом указанная ассоциированная с патогеном молекулярная структура представляет собой агонист патогенраспознающего (PR) рецептора.
23. Состав по п.22, отличающийся тем, что микрочастицы имеют диаметр меньше приблизительно 5 мкм.
24. Способ идентификации иммунологически активных пептидов для использования в иммунологически активном составе по п.13 или 14, включающий следующие этапы:
(a) проведение адсорбции GAG;
(b) проведение иммуноаффинной селекции;
(c) проведение идентификации иммунологически активных пептидов из пептидов, отобранных при иммуноаффинной селекции.
25. Способ по п.24, дополнительно включающий стадию (d), представляющую собой выполнение протеолитического расщепления белка или белков, выделенных путем иммуноаффинной селекции с получением иммунологически активных пептидов.
26. Способ по любому из пп.24 или 25, в котором адсорбция GAG представляет собой адсорбцию гепарина и который дополнительно включает этап анализа информации о последовательности биоинформатическими методами с использованием многоосновных линейных мотивов для идентификации мотивов, опосредующих проникновение.
27. Способ идентификации иммунологически активных пептидов для использования в иммунологически активном составе по п.13 или 14, включающий следующие этапы:
(a) осуществление связывания комплементсвязывающих антител с белком комплемента;
(b) применение указанных антител для иммуноаффинной селекции; и
(c) проведение идентификации иммунологически активных пептидов из пептидов, отобранных при иммуноаффинной селекции.
28. Способ по п.27, дополнительно включающий стадию (d), представляющую собой выполнение протеолитического расщепления выделенных иммунологически активных пептидов.
29. Способ индукции иммунного ответа у субъекта, включающий этап введения иммунологически эффективного количества состава по п.1, в котором по меньшей мере одна ассоциированная с патогеном молекулярная структура представляет собой агонист патогенраспознающего (PR) рецептора, при этом носитель представляет собой микрочастицы, к которым присоединен указанный агонист патогенраспознающего (PR) рецептора, и при этом указанные микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
30. Способ по п.29, отличающийся тем, что состав содержит более одного агониста патогенраспознающего рецептора.
31. Способ по п.29, включающий in vivo доставку.
32. Способ по п.31, отличающийся тем, что он характеризуется одним или более из следующих эффектов или результатов:
(a) число патогенов в организме инфицированного животного снижают или патоген полностью элиминируют;
(b) блокируют проникновение патогена в клетки;
(c) введенные эпитопы обеспечивают образование блокирующего (нейтрализующего) антитела, способного активировать комплемент;
(d) блокирование эпитопов, которые способны препятствовать связыванию патогена или отдельных участков патогена с белком или рецептором, способным индуцировать IL-10, приводит к появлению защитного иммунитета;
(e) блокирование эпитопов приводит к уменьшению образования IL-10;
(f) введение указанного состава приводит к элиминации устойчивой инфекции;
(g) иммунный ответ направлен на элементы связывания с рецептором на патогенном микроорганизме;
(h) иммунный ответ направлен на элементы патогенного микроорганизма, которые связывают гликозамингликаны;
(i) иммунный ответ элиминирует клетки, зараженные патогеном.
33. Способ по п.31, отличающийся тем, что in vivo доставку осуществляют путем, выбранным из группы, состоящей из введения через слизистую оболочку, парентерального пути введения, введения через кожу.
34. Способ по п.31, отличающийся тем, что он индуцирует защитный иммунный ответ по меньшей мере на один патоген, и состав вводят в виде одной или нескольких доз, а иммунный ответ включает ответы Тh1 или Th2, либо их комбинацию.
35. Способ по п.34, который имеет одну из следующих характеристик:
(a) патоген представляет собой внутриклеточный патоген;
(b) патоген способен вызвать скрытую инфекцию.
36. Способ по п.35, отличающийся тем, что патоген представляет собой микоплазму.
37. Способ по п.36, отличающийся тем, что микоплазма представляет собой Mycoplasma gallisepticum.
38. Способ обеспечения иммунитета к патогену у животного, включающий этап введения животному иммунологически эффективного количества состава по п.1, в котором: по меньшей мере одна ассоциированная с патогеном молекулярная структура представляет собой по меньшей мере один агонист патогенраспознающего рецептора (PR), и при этом носитель представляет собой микрочастицы, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
39. Способ по п.38, отличающийся тем, что иммунный ответ, вызванный указанным составом, препятствует проникновению патогенного микроорганизма в клетку-мишень или направлен на гликозамингликансвязывающие элементы патогенного микроорганизма.
40. Способ по п.39, отличающийся тем, что животное выбрано из группы, состоящей из млекопитающего и домашней птицы.
41. Способ по п.40, отличающийся тем, что животное представляет собой домашнюю птицу, и при этом указанная птица является цыпленком.
42. Дендритная клетка, выбранная из изолированной клетки или клетки в культуре, полученная путем введения в указанную клетку или в организм, содержащий указанную клетку, иммунологически активного состава по пп.1, 2 или 3, причем указанная дендритная клетка обладает иммунологической активностью.
43. Иммунологически активный состав для индукции защитного иммунитета, содержащий:
компонент А, содержащий по меньшей мере одну ассоциированную с патогеном молекулярную структуру, и
компонент В, содержащий по меньшей мере один носитель в виде микрочастиц агарозы, способный к эффективной доставке состава в организм с обеспечением ковалентного связывания по меньшей мере одной ассоциированной с патогеном молекулярной структуры с микрочастицами агарозы с осуществлением индукции защитного иммунитета за счет взаимодействия ассоциированной с патогеном молекулярной структуры в составе с клетками иммунной системы,
при этом индуцируемый защитный иммунитет специфичен в отношении используемой в указанном составе ассоциированной с патогеном молекулярной структуры.
44. Состав по п.43, отличающийся тем, что ассоциированную с патогеном молекулярную структуру выбирают из группы, включающей:
(а) агонист рецептора TLR 1;
(b) агонист рецептора TLR 2;
(с) агонист рецептора TLR 3;
(d) агонист рецептора TLR 4;
(e) агонист рецептора TLR 5;
(f) агонист рецептора TLR 6;
(g) агонист рецептора TLR 7;
(h) агонист рецептора TLR 8;
(i) агонист рецептора TLR 9;
(j) агонист NOD-1;
(k) агонист NOD-2;
(l) DC-SIGN;
(m) L-SIGN; и
(n) рецептор маннозы,
причем в случае, если ассоциированная с патогеном молекулярная структура представляет собой агонист NOD-1 или агонист NOD-2, указанный агонист NOD-1 или агонист NOD-2 выбран из группы, включающей бактериальный пептидогликан и производное бактериального пептидогликана.
45. Состав по п.43, отличающийся тем, что микрочастицы агарозы имеют одно из следующих свойств:
(a) микрочастицы имеют узкий диапазон распределения по размерам;
(b) микрочастицы являются пористыми;
(c) микрочастицы имеют диаметр меньше приблизительно 10 мкм; и
(d) микрочастицы связаны с более чем одной ассоциированной с патогеном молекулярной структурой, при этом указанная ассоциированная с патогеном молекулярная структура представляет собой агонист патогенраспознающего (PR) рецептора.
46. Состав по п.45, отличающийся тем, что микрочастицы имеют диаметр меньше приблизительно 5 мкм.
47. Способ индукции толерантного иммунного ответа на молекулу, белок или патоген, причем иммунный ответ включает ответ Th reg, при этом указанный способ включает этап введения в виде одной или нескольких доз иммунологически эффективного количества состава по п.1, при этом:
- указанный состав содержит один или более иммунологически активных иммуносупрессивных антигенов или эпитопов антигенов,
- указанная по меньшей мере одна ассоциированная с патогеном молекулярная структура представляет собой агонист патогенраспознающего рецептора, и
- носитель представляет собой микрочастицы, с которыми связан указанный по меньшей мере один агонист патогенраспознающего рецептора, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
48. Способ индукции толерантного иммунного ответа на иммунологически активный агент, причем указанный иммунный ответ включает ответ, выбранный из группы, состоящей из: (i) индукции регуляторного ответа, или снижения иммунных функций, или ускользания от иммунологического контроля, и (ii) иммунного ответа, приводящего к повышению образования IL-10, при этом указанный способ включает этап введения иммунологически эффективного количества состава по п.1, при этом:
- указанный состав содержит один или более иммунологически активных иммуносупрессивных антигенов или эпитопов антигенов; и
- по меньшей мере одна ассоциированная с патогеном молекулярная структура представляет собой агонист патогенраспознающего рецептора, и
- носитель представляет собой микрочастицы, с которыми связан указанный по меньшей мере один агонист патогенраспознающего рецептора, причем микрочастицы имеют размер в том же диапазоне, что и патоген, или меньше.
49. Способ по п.48, отличающийся тем, что состав включает иммунологически активный эпитоп антигена, содержащий гидрофобный фрагмент.
50. Способ по п.49, отличающийся тем, что гидрофобный фрагмент представляет собой липидсодержащий фрагмент.
51. Способ по п.50, отличающийся тем, что липидсодержащий фрагмент обладает по меньшей мере одной из указанных характеристик:
(a) содержит углеводные компоненты, наличие которых является результатом N-гликозилирования;
(b) присоединен к микрочастицам независимо от иммунологически активного эпитопа антигена,
(c) к нему присоединены другие молекулы.
52. Способ по п.48, отличающийся тем, что иммунологически активный эпитоп антигена содержит мотивы, включающие по меньшей мере одну аминокислоту, выбранную из группы, состоящей из аспарагина, треонина и серина, при этом указанные мотивы представляют собой мотивы Asp-X-Ser или Asp-X-Thr, где Х может быть любой аминокислотой, кроме пролина, и указанное антитело способно блокировать указанные мотивы.
| O'HAGAN et al | |||
| Microparticles for the delivery of DNA vaccines | |||
| Immunological Rev., 2004, Vol.199, p.191-200 | |||
| GEIJENBEEK et al | |||
| Mycobacteria Target DC-SIGN to Suppres Dentritic Cell Function | |||
| J | |||
| Exp.Med., 2003,Vol.197, No.1, p.7-17 | |||
| ФОНТАЛИН Л.Н., ПЕВНИЦКИЙ Л.А | |||
| Иммунологическая толерантность, Медицина, 1978, с.121-249 | |||
| ИММУНОМОДУЛЯТОР | 2003 |
|
RU2245142C1 |
| ЛЕКАРСТВЕННЫЙ ПРЕПАРАТ, ОБЛАДАЮЩИЙ ИММУНОМОДЕЛИРУЮЩИМ, ИММУНОКОРРЕГИРУЮЩИМ, ПРОТИВОПАРАЗИТАРНЫМ, ПРОТИВОСКЛЕРОТИЧЕСКИМ, ПРОТИВОВИРУСНЫМ, ПРОТИВОБАКТЕРИАЛЬНЫМ, ПРОТИВОГРИБКОВЫМ, ПРОТИВОВОСПАЛИТЕЛЬНЫМ И ПРОТИВООПУХОЛЕВЫМ ДЕЙСТВИЕМ, И СПОСОБ ЕГО ПРИГОТОВЛЕНИЯ | 2001 |
|
RU2197248C2 |

