Изобретение относится к области биомолекулярной фармакологии, биотехнологии и генетической инженерии и представляет собой рекомбинантный гибридный полипептидный препарат, включающий фрагменты плазминогена человека и эндостатина человека, обладающие специфической антипролиферационной активностью в отношении эндотелиальных клеток кровеносных сосудов, и способ получения этого препарата путем экспрессии его гена в Е. coli в составе плазмидной ДНК с последующей очисткой. Изобретение может быть использовано в медицине и в медицинской промышленности для создания новых лекарственных препаратов с антиангиогенным терапевтическим эффектом.
Одной из важнейших задач, стоящих перед медициной, является поиск эффективных методов для лечения больных со злокачественными заболеваниями. Одним из ключевых достижений науки XX века в этой области является доказательство необходимости процесса ангиогенеза для роста злокачественных солидных опухолей, а также создание концепции противоопухолевой терапии, основанной на подавлении ангиогенеза, разработанной Дж. Фолкманом [Folkman J. Nat. Med., 1:27-31, 1995].
Ангиогенез - процесс роста капилляров из кровеносных сосудов, в результате которого образуются новые сосудистые сети. В здоровом организме взрослого человека ангиогенез не встречается в большинстве органов и тканей, или его интенсивность пренебрежимо мала, кроме процессов тканевой регенерации, а также образования желтого тела, эндометрия и плаценты. Однако при экспрессии факторов роста (таких как фактор роста эндотелия сосудов (VEGF), факторы роста фибробластов (FGF) и др.), эндотелиальные клетки сосудов, находящиеся в покоящемся состоянии, могут вступать в клеточный цикл, пролиферировать, мигрировать и образовывать новые кровеносные сосуды. При этом патологический рост новых сосудов обусловливает прогрессию многих заболеваний, прежде всего рост и метастазирование солидных опухолей [Carmeliet P. and Jain R. K. Nature, 407:249-257, 2000].
При нормальных физиологических условиях интенсивность ангиогенеза зависит не только от уровня экспрессии указанных и некоторых других факторов роста, но и от уровня экспрессии ингибиторов ангиогенеза. Низкая интенсивность ангиогенеза обеспечивается паритетом в экспрессии как факторов роста, стимулирующих ангиогенез, так и ингибиторов последнего. Среди специфических ингибиторов ангиогенеза, воздействующих на пролиферирующие эндотелиальные клетки сосудов, одним из самых мощных является ангиостатин - белок с молекулярной массой 38-40 кДа, выделенный М. О'Рейлли и соавт. из крови и сыворотки мышей с солидными опухолями в организме [O'Reilly M.S., Holmgren L, et al. Cell, 79:315-328, 1994].
Путем специфического ингибирования ангиогенеза можно проводить терапию злокачественных новообразований и заболеваний, связанных с неоваскуляризацией сетчатки глаза, таких как диабетическая и серповидноклеточная ретинопатия и др. Ряд антиангиогенных препаратов, например, ангиостатин, эндостатин, скваламин, 2-метоксиэстрадиол и др., находятся в настоящее время на стадии клинических испытаний в странах Западной Европы и США [Ziche М. et al. Development of new drags in angiogenesis, Curr Drag Targ, 5:389-406, 2004]. В связи с этим поиск новых ингибиторов ангиогенеза и разработка методов их получения в количествах, достаточных для проведения доклинических исследований, становится важной научно-прикладной задачей современной медицины.
При изучении ангиостатина, одного из самых мощных ингибиторов пролиферации эндотелиальных клеток in vitro и ангиогенеза in vivo, было обнаружено, что этот белок является фрагментом одного из компонентов системы свертывания крови, а именно плазминогена. Ангиостатин образуется в организме, несущем раковую опухоль, путем протеолиза плазмина рядом ферментов, специфически экспрессирующихся в опухолевой ткани, например, матриксными металлопротеиназами (желатиназы А и В, стромелизин и др.), простатоспецифическим антигеном, а также в результате аутолиза плазмина. Поскольку при протеолизе плазмина различными ферментами образуется ряд различных полипептидов, обладающих антиангиогенной активностью, внимание исследователей привлек вопрос сравнительного изучения антипролиферативной активности по отношению к эндотелиальным клеткам различных фрагментов молекулы плазминогена.
Плазминоген человека - протеин с молекулярной массой 90 кДа, который содержит пять крингл-доменов (обозначаемых номерами от 1 до 5), представляющих собой особую жесткую структуру полипептидной цепи, свернутую в два кольца и поддерживаемую тремя дисульфидными связями. Длина цепи каждого крингл-домена составляет приблизительно 80 аминокислотных остатков. Ангиостатин представляет собой структуру, соответствующую крингл-доменам 1-4 плазминогена (К1-3). При исследованиях фрагментов плазминогена (ангиостатин, крингл-домен 1 (К1), крингл-домен 3 (К3), крингл-домены 2-3 (К2-3) и т.д.) было показано, что наибольшей ингибирующей активностью обладают крингл-домен 5 (К5) [Cao Y., Chen A. et al. J Biol Chem, 272:22924-22928, 1997] и крингл-домены 1-3.
Помимо высокой антипролиферативной активности, очевидны и другие преимущества крингл-домена 5 и К1-3 плазминогена человека как антиангиогенных препаратов. Во-первых, они проявляют ингибирующую активность специфично, действуя только на пролиферирующие эндотелиоциты кровеносных сосудов, и поэтому нетоксичны для других типов клеток [Zhang D., Kaufman P. et al. Diabetologia, 44:757-765, 2001]. Во-вторых, плазминоген является эндогенным белком тканей человека, и поэтому его пептидные фрагменты К5 и К1-3 не вызывают иммунного ответа. В-третьих, фрагмент К5, как полипептид с небольшой молекулярной массой (12-13 кДа), может быть легко получен в виде рекомбинантного белка в клетках Е. coli.
Еще одним мощным ингибитором ангиогенеза является фрагмент коллагена-XVIII-эндостатин, представляющий собой белок с молекулярной массой 20-22 кДа [M.S. O'Reilly et al. Cell, 88:277-285, 1997]. Фрагменты эндостатина обладают антиангиогенными и другими биологическими свойствами, характерными для эндостатина. Некоторые фрагменты обладают более высокой антиангиогенной активностью по сравнению с нативным эндостатином [Cattaneo M.G. et al. Exp Cell Res, 283:230-236 (2003)].
В настоящее время описан метод лечения некоторых заболеваний, сопровождающихся неоваскуляризацией роговицы, с помощью генотерапии лентивирусной конструкцией, экспрессирующей гибридный протеин, состоящий из эндостатина и К5 [Murthy R.C. et al. Investigative Ophthalmology & Visual Science, 44(5): 1837-1842, (2003)]. Однако генная терапия, лежащая в сути описанного R.C. Murthy и соавт. метода, известна своими недостатками, прежде всего неизученными последствиями внедрения ксеногенов и низкой эффективностью.
Задачей изобретения являлось создание новых нетоксичных высокоэффективных полипептидных препаратов, обладающих антиэндотелиальной активностью.
Для решения изобретательской задачи предложены препараты, представляющие собой гибридные белки, содержащие в различных вариантах фрагменты эндостатина (полипептид 1) и плазминогена (полипептид 2), ковалентно связанные между собой. В предлагаемых антиангиогенных препаратах сравнительно со структурой нативного эндостатина человека в полипептиде 1 осуществлена замена аминокислотного остатка цистеина в положении 173 на аланин (фиг.1А), что позволяет существенно облегчить получение препарата генно-инженерными методами, но не сказывается на проявлении им антиангиогенной активности. Полипептид 2 в предлагаемых гибридных препаратах представляет собой последовательность плазминогена человека с 82 по 341 аминокислоту (фиг.2А) или с 463 по 511 аминокислоту (фиг.2Б).
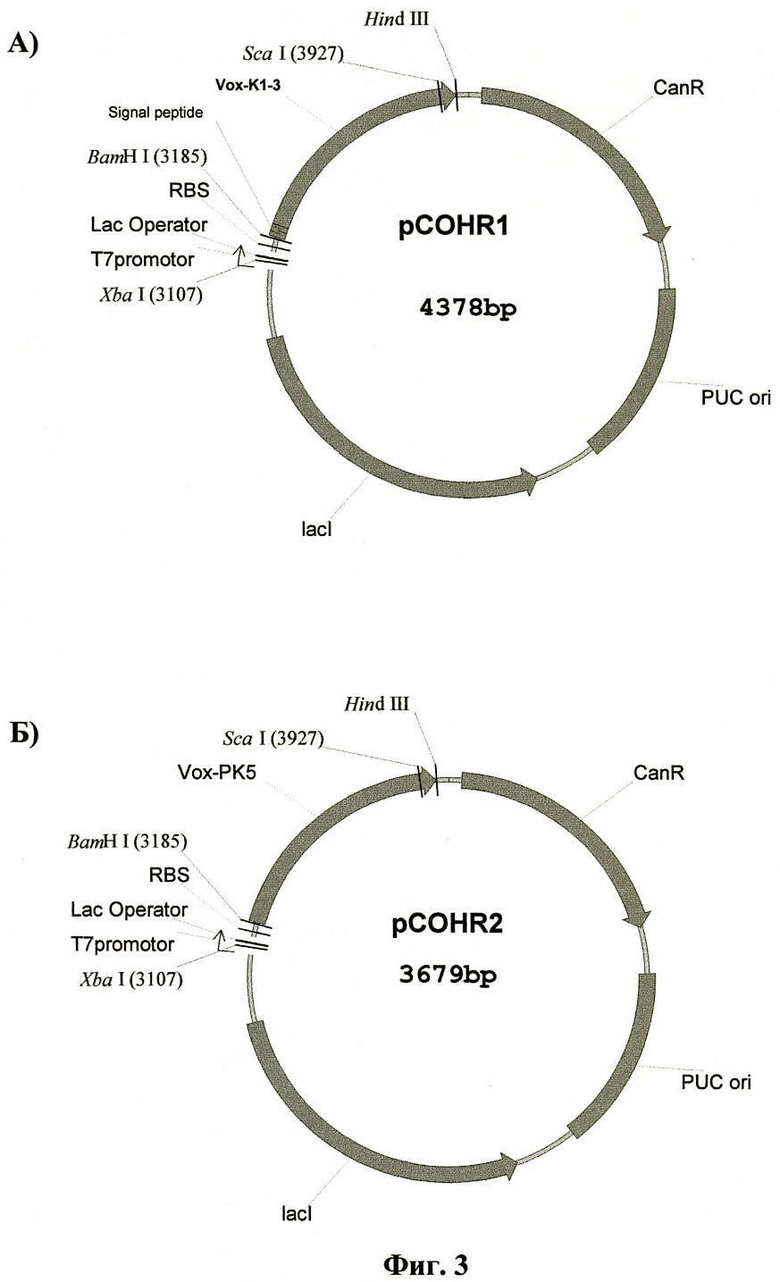
В рамках изобретения предлагаются также способы получения предлагаемых препаратов в качестве рекомбинантных продуктов с использованием продуцента Е. coli, включающие методы их выделения и очистки. Рекомбинантные гибридные полипептиды получают путем экспрессии их генов в составе рекомбинантных плазмидных ДНК pCOHRl (4378 пар оснований) и pCOHR2 (3679 пар оснований) в клетках Е. coli, причем экспрессию осуществляют в среде для роста Е. coli сначала в отсутствие индуктора 1ас-промотора изопропилтио-β-D-галактопиранозида (IPTG), а по достижении мутности среды 0,4-0,6 OD, добавляя индуктор IPTG до концентрации 0,1-0,3 мМ и продолжая экспрессию под контролем индуцированного lac-промотора рекомбинантной плазмиды. При этом препарат, получаемый из периплазмы клеток Е. coli, трансформированных рекомбинантной плазмидной ДНК pCOHRl, очищают с помощью методов аффинной и гельфильтрационной хроматографии, а препарат, выделяемый в виде телец включения из клеток Е. coli, трансформированных рекомбинантной плазмидной ДНК pCOHR2, ренатурируют и очищают с применением высокоэффективной жидкостной хроматографии.
С получением предлагаемых препаратов открываются возможности применения их в терапии онкологических и других заболеваний, сопровождающихся несбалансированной неоваскуляризацией, таких как диабетическая ретинопатия, болезнь Крона и др.
Изобретение иллюстрируется следующими примерами:
Пример 1. Синтез последовательности ДНК, содержащей сайт рестрикции фермента BamHI, старт-кодон, ген, кодирующий сигнальный пептид, последовательность, кодирующую слитый белок, состоящий из фрагмента эндостатина с произведенной заменой (фиг.1Б) и фрагмента плазминогена с 82 по 341 аминокислоту, а также стоп-кодон и сайт рестрикции фермента Hind III (последовательность назвали SPEK13).
На первой стадии проводили синтез последовательности, кодирующей 3 штрих часть гена фрагмента плазминогена (82-341) (последовательность назвали 3К13). Синтез 3К13 проводили с помощью праймеров 3К13 и 3К13 - R и библиотеки кДНК человека:

Последовательность ДНК, названную SPE, содержащую сайт рестрикции фермента BamHI (подчеркнутый), старт-кодон, ген, кодирующий сигнальный пептид, последовательность, приведенную на фиг.1Б и названную Vox, а также фрагмент, кодирующий 5 штрих часть гена фрагмента плазминогена (82-341) (выделенный) с сайтом рестрикции фермента SacII (пунктирное подчеркивание), получали с помощью синтезатора олигонуклеотидов:

Пример 2. Синтез последовательности ДНК, содержащей сайт рестрикции фермента BamHI, старт-кодон, участок, кодирующий слитный белок, состоящий из фрагмента плазминогена с 463 по 511 аминокислоту и фрагмента эндостатина с произведенной заменой (фиг.1Б), а также стоп-кодон и сайт рестрикции фермента Hind III (последовательность назвали PK5Vox).
С помощью синтезатора олигонуклеотидов получали фрагменты, названные 5РК5 и 3K5Vox:
5РК5 - последовательность ДНК, содержащей сайт рестрикции фермента BamHI, старт-кодон, участок, кодирующий 5' часть фрагмента плазминогена с 463 по 511 аминокислоту с сайтом рестрикции фермента AvaII.

Пример 3. Продукция гибридных полипетидов в штаммах Е. coli
Для создания экспрессионных векторов последовательность SPEK13 или PK5Vox обрабатывали эндонуклеазами рестрикции BamHI и Hindlll и лигировали с вектором pBSH2, который предварительно был обработан этими же рестриктазами. Полученные векторы назвали pCOHRl (фиг.3А) и pCOHR2(фиг.3Б).
Экспрессионный вектор pCOHRl либо pCOHR2 был инкорпорирован в штамм Е. coli BL21/DE3. Для осуществления экспрессии целевого белка 30 мл трансфецированного штамма выдерживали при покачивании в течение 2-3 ч в среде LB при 37°C с добавлением канамицина (50 мкг/мл) и глюкозы (0,2%). Затем разбавляли эту культуру в 20 раз свежей средой LB, содержащей канамицин (40 мкг/мл), и наращивали 2-3 ч при покачивании. По достижении культуральной средой плотности OD280нм=0,5 вносили в нее изопропилтио-β-D-галактопиранозид (IPTG) до концентрации 0,5 мМ для индукции биосинтеза. Затем инкубировали данную культуру около 5 ч при 37°С при покачивании, центрифугировали при 10000 g в течение 15 мин, супернатант отбрасывали.
Пример 4. Очистка продукта экспрессии вектора pCOHRl
Осадок клеток, полученный, как описано в примере 3, ресуспендировали в буфере А (20 мМ Трис, рН 8.0, 15% галактоза и 1 мМ ЭДТА) в течение 10 мин при 20°C. Суспензию клеток затем центрифугировали при 10000 g в течение 15 мин; супернатант удаляли и ресуспендировали клетки в 7 мМ растворе магния (II) сульфата при 0°C в течение 20 мин. Высвободившиеся в результате белки периплазматического пространства отделяли затем от клеток с помощью центрифугирования при 10000 g в течение 20 мин при 4°C; осадок отбрасывали. Полученный раствор белков периплазмы концентрировали с помощью ультрафильтрации в ячейке для концентрирования и диализовали против буфера Б (12 мМ Трис, рН 7.5) в течение 5 ч при 4°C. Затем очистку целевого продукта, полученного в штаммах, трансфецированных рекомбинантной плазмидой pCOHRl, осуществляли методом аффинной хроматографии. Раствор, содержащий целевой продукт, наносили на хроматографическую колонку, заполненную сорбентом лизин-сефарозой, предварительно уравновешенную буфером В (50 мМ натрий гидрофосфат, рН7,4). После нанесения раствора целевого белка колонку промывали двумя объемами буфера В, а затем буфером Г (50 мМ натрий гидрофосфат, 0,15 М натрий хлорид, рН 7,4) до регистрации базовой линии. Затем элюировали целевой продукт 0,2 М раствором ε-аминокапроновой кислоты в буфере Г. Фракции, содержащие целевой продукт, собирали и концентрировали на мембране для ультрафильтрации (MW=10 кДа). Дальнейшую очистку проводили способом гельфильтрационной хроматографии. При очистке продукта, полученного из штамма-продуцента, трансфецированного экспрессионным вектором pCOHRl, сконцентрированные фракции наносили на хроматографическую колонку, заполненную сорбентом Sephacryl S - 300 (Amersham Biosciences, USA), предварительно уравновешенную буфером Д (10 мМ натрий гидрофосфат, 2,7 мМ калий хлорид, 0,137 М натрий хлорид, рН 7,4) (процедуру проводили при 4°C). Собирали фракции, содержащие очищенный целевой продукт.
Пример 5. Очистка продукта экспрессии вектора pCOHR2
Осадок клеток, полученный так, как описано в примере 3, обрабатывали ультразвуком с помощью щелевого зонда в буфере А (50 мМ Трис, рН 8,0, 1 мМ ЭДТА). Полученную суспензию центрифугировали при 10000 g в течение 15 мин, супернатант отбрасывали. Осадок затем ресуспендировали в буфере Б (50 мМ Трис, рН 8,0, 1 мМ ЭДТА, 1 мМ дитиотреитол, 1% натрий N-лаурилсаркозин) и центрифугировали при 10000 g в течение 15 мин, супернатант отбрасывали. Осадок ресуспендировали в буфере В (20 мМ Трис, рН 8,0, 2% натрий деоксихолат) и центрифугировали при 10000 g в течение 15 мин, супернатант отбрасывали; данную процедуру повторяли трижды. Затем полученный осадок телец включения, содержащих целевой компонент, растворяли в буфере Г (50 мМ Трис, рН 8,0, 8 М гуанидин гидрохлорид, 70 мМ 2-меркаптоэтанол). После полного растворения осадка полученный раствор разбавляли буфером Д (50 мМ Трис, рН 8,0, 2 М мочевина,
2 мМ восстановленный глутатион, 300 мкМ окисленный глутатион, 5 мМ L-лизин гидрохлорид) в соотношении 1:30. Полученный раствор оставляли для прохождения ренатурации рекомбинантного белка при 4°C на 24 ч. Затем раствор продукта помещали в диализные мешки с проницаемостью пор молекулярным весом 8 кДа и диализовали 8 ч против буфера Е (10 мМ натрий гидрофосфат, 15 мМ натрий хлорид, 0,02% натрий азид, рН 7,0); процедуру повторяли, сменив буфер Е. Раствор затем изымали из диализной системы и дальнейшую очистку проводили методом высокоэффективной жидкостной хроматографии.
Пример 6. Определение биологической активности полученных гибридных антиэндотелиальных препаратов
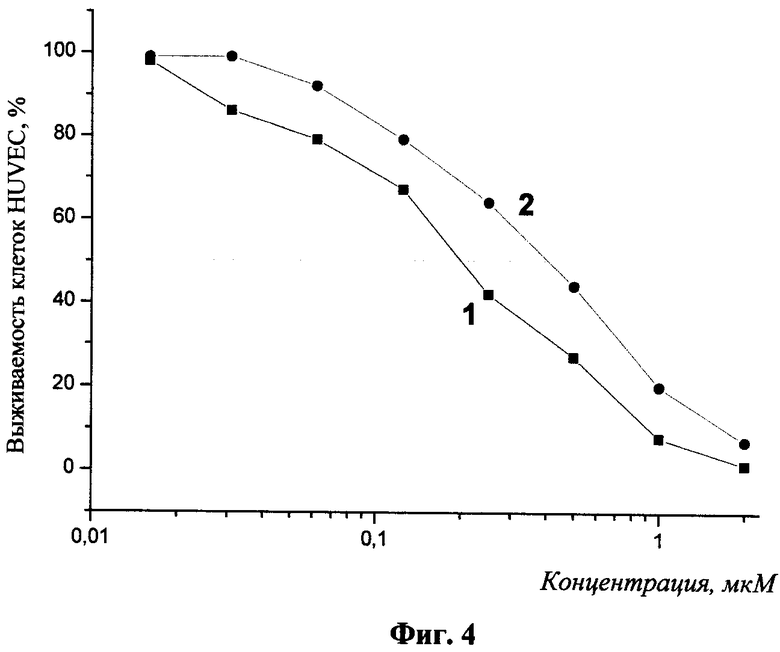
Определение биологической активности полученных препаратов проводили путем оценки их способности ингибировать пролиферацию эндотелиальных клеток человека in vitro. Исследование пролиферации эндотелиальных клеток проводили описанным ранее методом [O'Reilly М. et al. Cell, 88:277-285 (1997)]. Клетки HUVEC (эндотелиальные клетки пупочной вены человека) из монослоя трипсинизировали и ресуспендировали в среде M199, содержащей 5% фетальной бычьей сыворотки. Затем клетки рассеивали в 96-луночные планшеты, покрытые желатином, в плотности 5000 клеток в лунку. Через 24 ч добавляли раствор гибридных полипептидов в различных концентрациях. По прошествии 20 мин культуры клеток обрабатывались 5 нг/мл раствором основного фактора роста фибробластов (Life Technilogies Inc., USA) в присутствии гепарина (1 мкг/мл). Выживаемость клеток в контрольной и в обработанной препаратами группах определяли с помощью МТТ-теста через 72 ч инкубации. МТТ-тест эффективно определяет метаболитическую активность митохондрий, и его результат прямо коррелирует с числом живых клеток. По результатам анализа полученных препаратов, представленных на фиг.4, определена их IC50: препарат, полученный при трансфецировании штаммов Е. coli плазмидой pCOHR1, имел IC50=190 нМ; препарат, полученный при трансфецировании штаммов Е. coli плазмидой pCOHR2, имел IC50=400нМ. Таким образом, настоящее изобретение позволяет с высоким выходом получать новый гибридный полипептидный препарат, проявляющий антиэндотелиальную активность.


Изобретение относится к области биомолекулярной фармакологии, биотехнологии и генетической инженерии и касается рекомбинантного гибридного полипептида и способа его получения. Предложенный гибридный полипептид представляет собой полипептид 1, к которому ковалентно присоединен полипептид 2, где полипептид 1 представляет собой последовательность эндостатина человека с 135 по 184 аминокислотный остаток, причем в положении 173 произведена замена Цис на Ала сравнительно с нативной последовательностью эндостатина, а полипептид 2 представляет собой последовательность плазминогена человека с 82 по 341 или с 463 по 511 аминокислотный остаток. Так же предложен способ получения гибридного полипептида с использованием продуцента Е. coli, включающий методы его выделения и очистки. Представленный полипептид способен ингибировать пролиферацию эндотелиальных клеток человека in vitro и может быть использован при изготовлении нетоксичных препаратов для ингибирования ангиогенеза. 2 н. и 2 з.п. ф-лы, 4 ил., 6 пр.
1. Рекомбинантный гибридный полипептид, способный ингибировать пролиферацию эндотелиальных клеток человека in vitro, представляющий собой полипептид 1, к которому ковалентно присоединен полипептид 2, где полипептид 1 представляет собой последовательность эндостатина человека с 135 по 184 аминокислотный остаток, причем в положении 173 произведена замена Цис на Ала сравнительно с нативной последовательностью эндостатина, а полипептид 2 представляет собой последовательность плазминогена человека с 82 по 341 или с 463 по 511 аминокислотный остаток.
2. Способ получения полипептида по п.1, отличающийся тем, что полипептид получают путем экспрессии его гена в клетках Е. coli BL21 (DE3), трансформированных рекомбинантными плазмидными ДНК pCOHR1 (4378 п.н.) или pCOHR2 (3679 п.н.), для чего клетки инкубируют в ростовой среде до достижения мутности 0,4-0,6 OD, затем продолжают инкубацию в присутствии изопропилтио-β-D-галактопиранозида в концентрации 0,1-0,3 мМ, а целевой полипептид выделяют из периплазмы клеток или из телец включения.
3. Способ по п.2, отличающийся тем, что полипептид выделяют из периплазмы клеток Е. coli, трансформированных рекомбинантной плазмидной ДНК pCOHR1, и очищают с помощью аффинной хроматографии и гельфильтрационной хроматографии.
4. Способ по п.2, отличающийся тем, что полипептид выделяют из клеток Е. coli, трансформированных рекомбинантной плазмидной ДНК pCOHR2, в виде телец включения и осуществляют его ренатурацию с последующей очисткой высокоэффективной жидкостной хроматографией.
| ГОМОДИМЕРНЫЙ СЛИТЫЙ БЕЛОК, ИМЕЮЩИЙ АКТИВНОСТЬ ИНГИБИТОРА АНГИОГЕНЕЗА (ВАРИАНТЫ), МОЛЕКУЛА ДНК, КОДИРУЮЩАЯ ГОМОДИМЕРНЫЙ СЛИТЫЙ БЕЛОК, ВЕКТОР ДЛЯ ЭКСПРЕССИИ ГОМОДИМЕРНОГО СЛИТОГО БЕЛКА, СПОСОБ ТРАНСФЕКЦИИ КЛЕТКИ МЛЕКОПИТАЮЩЕГО И СПОСОБ ПОЛУЧЕНИЯ ГОМОДИМЕРНОГО СЛИТОГО БЕЛКА | 1999 |
|
RU2240328C2 |
| RAGHU С.MURTHY et al., Corneal Transduction to Inhibit Angiogenesis and Graft Failure, Investigative Ophthalmology & Visual Science, 2003 May, Vol.44, No.5, p.p.1837-1842. | |||

