Область техники
Изобретение относится к области биотехнологии, а именно к технологии получения биологически активных веществ (БАВ) методами генной инженерии, точнее к методам получения человеческого активатора плазминогена тканевого типа, соответствующему рекомбинантному человеческому делеционному варианту тканевого активатора плазминогена (ТАП) ретеплазе, в терапевтически достаточных количествах.
Предшествующий уровень техники.
Плазминоген - это гликопротеид, синтезируемый в печени и постоянно циркулирующий в плазме, способный связываться с фибрином тромба. После ферментативного превращения (активации) из неактивного плазминогена образуется плазмин - сериновая протеаза, которая расщепляет фибрин, фибриноген, а также V, VIII и XII факторы свертывания крови, уменьшает адгезию тромбоцитов и вызывает их дезагрегацию. Способностью активировать плазминоген обладают эндогенные (тканевый активатор плазминогена, урокиназа или тканевый активатор плазминогена урокиназного типа, фактор XII, калликреин, кинины) и экзогенные факторы (бактериальные стрептокиназа и стафилокиназа).
Современные фибринолитические (тромболитические) лекарственные средства основаны на стимуляции образования плазмина из эндогенного плазминогена. В этом случае системное действие лекарства будет сочетаться с местным, то есть препарат будет достигать наибольшей активности на поверхности тромба, где уже присутствует связанный с фибрином плазминоген. К числу тромболитиков - активаторов плазминогена относят препараты бактериальных ферментов (стрептокиназа, анистреплаза, стафилокиназа), препараты урокиназы и препараты тканевого активатора плазминогена, ТАП (рекомбинантный полноразмерный ТАП и делеционные варианты ТАП), а также другие, менее распространенные средства.
Активатор плазминогена тканевого типа или тканевый активатор плазминогена (tPA, ТАП) - это сериновая протеаза (шифр международного Классификатора Ферментов ЕС 3.4.21.68), которая катализирует превращение неактивного профермента плазминогена в активный фермент плазмин и является важным компонентом системы фибринолиза. ТАП синтезируется in vivo как одноцепочечный полипептид (72 кДа), который разрезается различными протеиназами, включая плазмин, тканевый калликреин и активированный фактор X, гидролизующими пептидную связь Arg275-Ile276 с образованием цепи А и цепи В, удерживающихся вместе за счет дисульфидной связи. N-концевая область (А- или тяжелая цепь) состоит из нескольких доменов: остатки 4-50 гомологичны пальцеподобному F-домену фибронектина, обеспечивающему связывание с фибрином; остатки 50-87 образуют Е-домен, гомологичный эпидермальному фактору роста; остатки 87-176 (крингл 1) и 176-262 (крингл 2) гомологичны крингл-доменам плазминогена. Р-домен, включающий остатки 276-527 (В- или тяжелая цепь), гомологичен сериновым протеиназам и содержит каталитический участок активного центра, включающий His322, Asp371 и Ser478 (J Thromb Haemost. 2004 Apr;2(4):541-6. Tissue-type plasminogen activator: a historical perspective and personal account. Collen D, Lijnen HR.). Отдельные домены ТАП обусловливают различные функции фермента, такие как связывание с фибрином, активация плазминогена, связывание с рецепторами эндотелиальных клеток и быстрое выведение из организма. Связывание ТАП с поверхностью фибрина опосредуется областью фибронектинового пальца и областью второго крингл-домена.
Эндогенный ТАП, определяемый в крови, представляет собой эндотелиальный активатор, высвобождаемый в кровоток под действием разных стимулов. Концентрация ТАП в крови равна 6,6±2,9 нг/мл. В результате частичной деградации фибрина плазмином его связывание с ТАП увеличивается. В присутствии фибрина в 1500 раз возрастает каталитическая эффективность (kкат/KM) ТАП в отношении плазминогена.
Существует несколько основных лекарственных препаратов, основанных на вариантах рекомбинантного человеческого ТАП (рчТАП): альтеплаза (патент США 5,869,314), ретеплаза (Европейская патентная заявка ЕР0242836А1), ТНКаза (патенты США 5,385,732 и 5,270,198). Концентрация природного ТАП в плазме очень низка и его выделение из донорской плазмы экономически невыгодно.
Ретеплаза - препарат на основе негликозилированного рекомбинантного делеционного варианта активатора плазминогена тканевого типа человека. Рекомбинантный полипептид, массой 39571 Да, включает 355 из 527 аминокислот природного аналога, продуцируется в системе Escherichia coli (вследствие чего негликозилирован), содержит каталитический и Крингл 2 домены природного активатора плазминогена тканевого типа человека и не содержит доменов крингл 1, эфр, фибронектиного пальца.
Структуры, которые усиливают аффинность природного ТАП к фибрину (фибронектиновый палец) и к рецепторам клеток печени, обеспечивающим ускоренный клиренс (крингл 1, домен Е, полисахаридные остатки), отсутствуют в рекомбинантном полипептиде, вследствие чего время полужизни ретеплазы увеличено (по сравнению с полноразмерным аналогом) до 13-16 минут. Увеличение времени полужизни позволяет использовать в клинике одно- или двукратное внутривенное введение препарата вместо постоянной инфузии (JAMA. 2001 Jul 25;286(4):442-9. Bolus fibrinolytic therapy in acute myocardial infarction. Llevadot J, Giugliano RP, Antman ЕМ). В свою очередь, снижение аффинности к фибрину позволяет свободному (не связавшемуся с фибрином) препарату проникать вглубь тромба, что обеспечивает ускорение тромболизиса по сравнению с полноразмерным ТАП. Ряд исследований свидетельствует о сравнительно большей эффективности ретеплазы по сравнению с полноразмерным вариантом ТАП (N Engl J Med. 1997 Oct 16;337(16):1118-23. A comparison of reteplase with alteplase for acute myocardial infarction. The Global Use of Strategies to Open Occluded Coronary Arteries (GUSTO III) Investigators)(Am J Health Syst Pharm. 1997 Nov 15;54 Suppl 1:S27-30. Clinical trials in thrombolytic therapy. Part 2: The open-artery hypothesis and RAPID-1 and RAPID-2).
Так, сравнительное исследование тромболитической активности ретеплазы и полноразмерного ТАП (альтеплазы) на модели in vitro показало, что полноразмерный ТАП по большей части после промывки остается на поверхности фибринового сгустка, тогда как делеционный ТАП проникает вглубь сгустка, активируя содержащийся внутри сгустка связанный с фибрином плазминоген. В ряде модельных исследований на собаках с тромбозом коронарной артерии было показано, что ретеплаза показывает максимальную эффективность по сравнению со стрептокиназой, урокиназой, анистреплазой, обеспечивая минимальное время реперфузии (восстановления кровотока) коронарной артерии.
Клинические испытания, в которых исследовалась сравнительная эффективность ретеплазы и препарата на основе полноразмерного ТАП при остром инфаркте миокарда показали, что при использовании ретеплазы скорость реперфузии коронарных артерий значительно выше и меньше доля пациентов, требующих последующей ангиопластики. При сравнительном клиническом исследовании RAPID 2, в ходе которого после введения ретеплазы или полноразмерного ТАП слепым методом проводили ангиографию сосудов сердца, было установлено, что ретеплаза вызывает более быстрое восстановление кровотока в пораженной коронарной артерии. Также было показано, что доля пациентов, потребовавших дополнительных вмешательств в течение первых 6 часов после начала терапии, в случае использования ретеплазы вдвое ниже, чем в случае использования полноразмерного ТАП (13.6% против 25.6%, р=0.004) (Lopez, LM, Am. J. Health Syst. Pharm. 1997 Nov 15;54 Suppl 1:S27-30. Clinical trials in thrombolytic therapy. Part 2: The open-artery hypothesis and RAPID-1 and RAPID-2).
Несмотря на отличные клинические показатели, ретеплаза в настоящий момент занимает ограниченную долю мирового рынка. Основной технологической проблемой в производстве ретеплазы является совокупность малоэффективного штамма-продуцента, основанного на использовании природной кДНК фрагмента ТАП и вспомогательной плазмиды с генами тРНК редких кодонов, а также весьма дорогостоящей процедуры рефолдинга, состоящей из конверсии неочищенной восстановленной формы ретеплазы в смешанно-дисульфидную с использованием больших количеств окисленного глютатиона и последующей ренатурации белка в очень разбавленном растворе в присутствии высоких концентраций аргинина (Европейская патентная заявка ЕР0242836А1). Следствием использования этих технологий получения является необходимость применения дорогостоящей аффинной очистки ренатурированного белка и весьма низкий выход готового продукта.
Краткое описание настоящего изобретения
Технической задачей, решаемой авторами, являлось создание технологии получения рчТАП с более высоким выходом.
Технический результат достигался путем создания технологии, включающей в себя новую экспрессионную плазмидную ДНК p10E-tPA, кодирующую фрагмент тканевого активатора плазминогена человека, создание штамма продуцента Е.coli на ее основе и технологию выделения рчТАП.
В основе данного решения лежат разработанные авторами плазмидная ДНК p10E-tPA длиной 6388 п.о., кодирующая отщепляемый N-концевой лидерный пептид длиной 23 аминокислоты, включающий декагистидиновый кластер и последовательность узнавания энтерокиназы, и слитую с ним в рамке последовательность, кодирующую фрагмент тканевого активатора плазминогена человека. Указанный фрагмент содержит оптимальные для Е.coli кодоны, позволяющие увеличить уровень экспрессии гетерологичного белка за счет эффективной трансляции всех аминокислот полипептида. Наличие лидерного пептида позволяет проводить очистку полипептида в денатурирующих условиях и проводить его рефолдинг не в растворе, а на поверхности металлохелатного сорбента, что приводит к значительному увеличению выхода очищенного активного целевого белка.
Целью настоящего изобретения является предоставление экспрессионной плазмиды, содержащей фрагмент ДНК, кодирующий фрагмент тканевого активатора плазминогена человека, включающий последовательность, кодирующую отщепляемый N-концевой лидерный пептид, включающий декагистидиновый кластер и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей фрагмент тканевого активатора плазминогена человека, под контролем промотора, функционирующего в бактериальной клетке.
Также целью настоящего изобретения является предоставление описанной выше экспрессионной плазмиды, где указанная плазмида выбрана из группы, состоящей из плазмид p10E-tPA и pM-tPA.
Также целью настоящего изобретения является предоставление бактерии, принадлежащей к роду Escherichia, трансформированной описанной выше плазмидой, - продуцента предшественника рекомбинантного фрагмента тканевого активатора плазминогена человека.
Также целью настоящего изобретения является предоставление описанной выше бактерии, где указанная бактерия представлена штаммами Е.coli BL21[DE3]/p10E-tPA и Е.coli BL21[DE3]/pM-tPA.
Также целью настоящего изобретения является предоставление предшественника рекомбинантного фрагмента тканевого активатора плазминогена человека, содержащего отщепляемый N-концевой лидерный пептид, включающий декагистидиновый кластер, и последовательность узнавания энтерокиназы.
Также целью настоящего изобретения является предоставление способа получения рекомбинантного фрагмента тканевого активатора плазминогена человека, включающий культивирование описанной выше бактерии в питательной среде, выделение телец включения, солюбилизацию белка-предшественника, металлохелатную хроматографию в денатурирующих условиях, рефолдинг белка-предшественника, получение зрелого белка обработкой энтерокиназой и выделение зрелого белка.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором культивируют штамм Е.coli BL21[DE3]/p10E-tPA или Е.coli BL21[DE3]/pM-tPA.
Подробное описание настоящего изобретения
Для реализации настоящего изобретения главной технической задачей явилось создание способа получения рекомбинантного фрагмента тканевого активатора плазминогена человека с использованием бактерии, трансформированной экспрессионной плазмидой, содержащей фрагмент ДНК, кодирующий предшественник рекомбинантного фрагмента тканевого активатора плазминогена человека, включающий последовательность, кодирующую отщепляемый N-концевой лидерный пептид, включающий декагистидиновый кластер и последовательность узнавания энтерокиназы, и слитую с ним в рамке последовательность, кодирующую рекомбинантный фрагмент тканевого активатора плазминогена человека, под контролем промотора, функционирующего в бактериальной клетке.
Термин «экспрессионная плазмида» означает плазмидную ДНК, содержащую все необходимые генетические элементы для экспрессии внедренного в него гена, например, такие как промотор, терминатор. Конкретным примером генетических элементов, необходимых для экспрессии предшественника рекомбинантного фрагмента тканевого активатора плазминогена человека в составе экспрессионной кассеты согласно настоящему изобретению, является промотор РНК-полимеразы бактериофага Т7, но не ограничивается им.
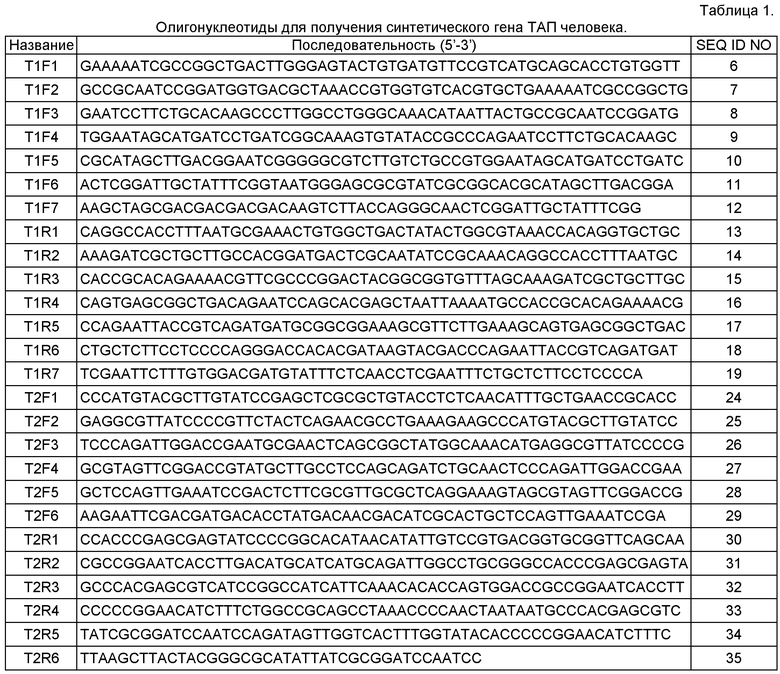
Фрагментом ДНК, кодирующим предшественник рекомбинантного фрагмента тканевого активатора плазминогена человека согласно настоящему изобретению, является, например, синтетический ген, кодирующий предшественник рекомбинантного фрагмента тканевого активатора плазминогена человека, включающий последовательность, кодирующую отщепляемый N-концевой лидерный пептид, включающий декагистидиновый кластер и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей рекомбинантный фрагмент тканевого активатора плазминогена человека. Указанный фрагмент ДНК может быть получен методом ПЦР (см. Пример 1, Рис.1). Также указанный фрагмент ДНК может быть получен с использованием технологии клонирования фирмы Sloning BioTechnology, описанной в заявке РСТ WO2005071077.
Чтобы обеспечить эффективную трансляцию клонированного гена в Е.coli, предпочтительно, чтобы в последовательности, кодирующей предшественник рекомбинантного фрагмента тканевого активатора плазминогена человека, все редкие кодоны были заменены синонимичными часто встречающимися кодонами.
Последовательность гена, кодирующего предшественник рекомбинантного фрагмента тканевого активатора плазминогена человека согласно настоящему изобретению, представлена в Перечне последовательностей под номером SEQ ID NO:1. Аминокислотная последовательность предшественника рекомбинантного ТАП человека согласно настоящему изобретению представлена в Перечне последовательностей под номером SEQ ID NO:2. Аминокислотная последовательность зрелого рекомбинантного фрагмента тканевого активатора плазминогена человека представляет собой последовательность под номером SEQ ID NO:2 без 23 первых аминокислот.
Фрагменты ДНК, которые кодируют по существу тот же белок, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК (SEQ ID NO:1), кодирующего предшественник рекомбинантного фрагмента тканевого активатора плазминогена человека, например, посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делегированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации. Фрагменты ДНК, которые кодируют по существу тот же белок, могут быть выявлены путем экспрессии фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке и установления активности экспрессируемого продукта.
Тканевый активатор плазминогена (синонимы: tPA, ТАП, активатор плазминогена тканевого типа) - это сериновая протеаза (шифр Международного Классификатора Ферментов ЕС 3.4.21.68), которая катализирует превращение неактивного профермента плазминоген в активный фермент плазмин.
ТАП синтезируется in vivo как одноцепочечный полипептид (72 кДа), который разрезается протеиназами, по связи Arg275-Ile276, с образованием цепи А и цепи В, удерживающихся вместе за счет дисульфидной связи. Отдельные домены ТАП обусловливают функции фермента, такие как связывание с фибрином, активация плазминогена, связывание с рецепторами эндотелиальных клеток и быстрое выведение из организма. Связывание ТАП с поверхностью фибрина опосредуется областью фибронектинового пальца и областью второго крингл-домена.
Экспрессируемый рекомбинантный полипептид, включает 355 из 527 аминокислот природного аналога, содержит каталитический и Крингл 2 домены природного активатора плазминогена тканевого типа человека (не содержит доменов крингл 1, эфр, фибронектинового пальца).
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами ТАП, определяются по его протеолитической активности (его способности гидролизовать пептидную связь плазминогена). Так, например, активность ТАП человека можно детектировать, например, по гидролизу синтетического субстрата Т2943 методом, описанным в Примере 7. Считается, что вариант белка обладает свойствами ТАП человека при условии, что активность указанного варианта составляет не ниже 1% активности нативного ТАП человека.
Экспрессионная плазмида согласно настоящему изобретению содержит фрагмент ДНК, кодирующий предшественник рекомбинантного фрагмента ТАП человека, включающий последовательность, кодирующую отщепляемый N-концевой лидерный пептид, включающий декагистидиновый кластер, и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей фрагмент ТАП человека, под контролем промотора, функционирующего в бактериальной клетке.
В качестве рекомбинантной плазмиды согласно настоящему изобретению могут использоваться различные плазмиды, обладающие способностью к экспрессии в клетке-реципиенте, такие как плазмиды pBR322, pMW119, pUC19, pET22b, pET28b и подобные им, но список плазмид не ограничивается ими.
Конкретным вариантом реализации настоящего изобретения является плазмида p10EK-tPA, которая состоит из:
1) фрагмента NheI-NcoI длиной 41 п.о., представляющего собой синтетический адаптер, кодирующий декагистидиновый кластер;
2) фрагмента NcoI-HindIII вектора рЕТ28а(+) длиной 5246 п.о., содержащего область начала репликации плазмиды pBR322, ген РНК-организующего белка Rop, участок инициации репликации бактериофага f1, последовательность, кодирующую аминогликозид-3'-фосфотрансферазу, промотор РНК-полимеразы бактериофага Т7; участок терминации транскрипции; последовательность, кодирующую репрессор лактозного оперона;
3) фрагмента NheI-HindIII длиной 1101 п.о., кодирующего фрагмент ТАП человека и слитую в рамке последовательность узнавания энтерокиназой.
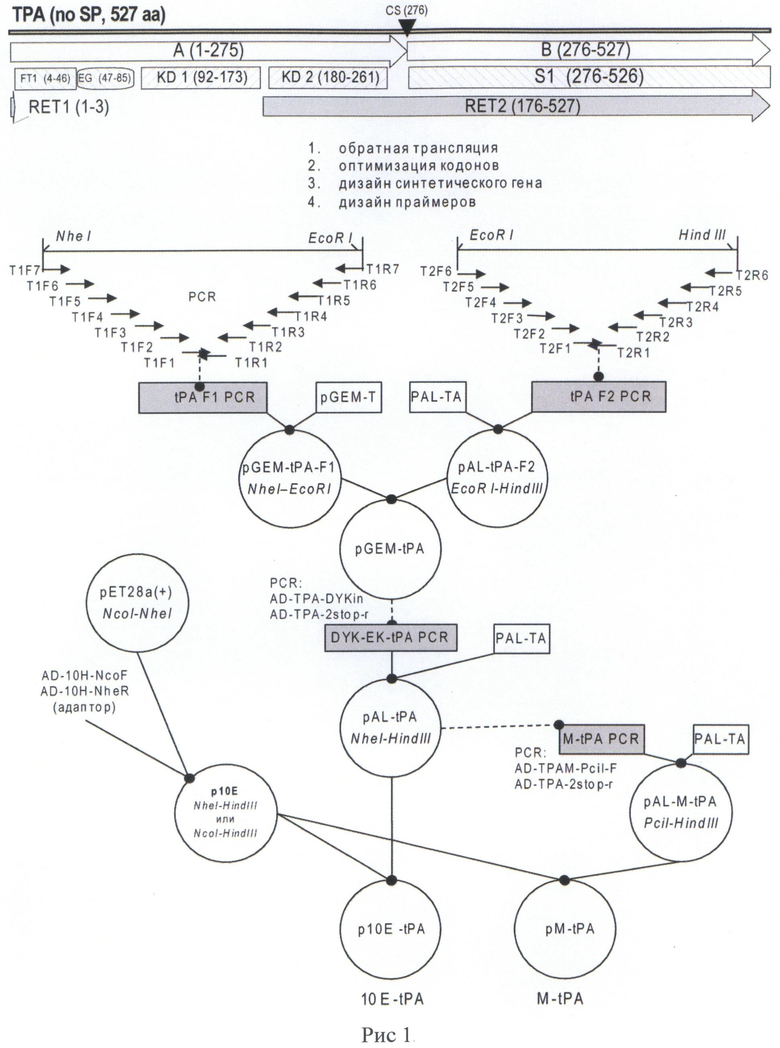
Указанная плазмида содержит уникальные сайты узнавания эндонуклеазами рестрикции: ClaI (1251), PciI (2142), ApaI (4040), NcoI (5070), NheI (5111), EcoRI (5708), HindIII (6212).
Структура плазмиды p10EK-tPA приведена на Рис.2.
Другим конкретным вариантом реализации настоящего изобретения является плазмида pM-tPA, которая состоит из:
1) фрагмента NcoI-HindIII вектора рЕТ28а(+) длиной 5246 п.о., содержащего область начала репликации плазмиды pBR322, ген РНК-организующего белка Rop, участок инициации репликации бактериофага f1, последовательность, кодирующую аминогликозид-3'-фосфотрансферазу, промотор РНК-полимеразы бактериофага Т7; участок терминации транскрипции; последовательность, кодирующую репрессор лактозного оперона;
2) фрагмента PciI-HindIII длиной 1076 п.о., кодирующего остаток метионина и фрагмент ТАП человека.
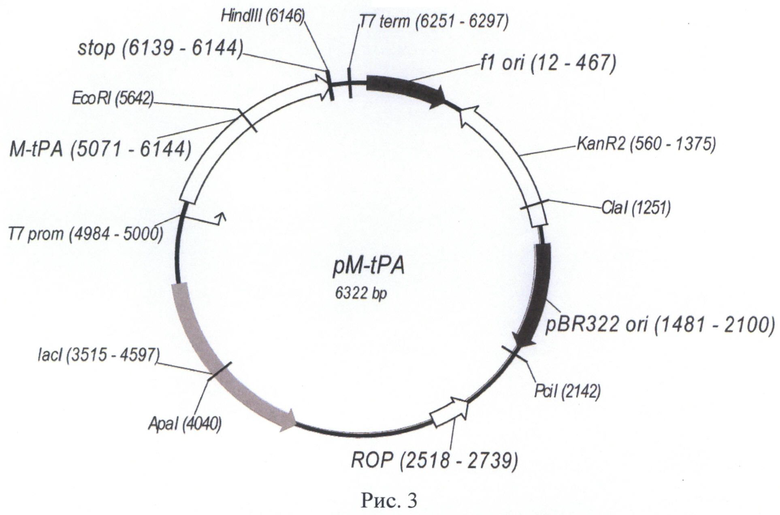
Указанная плазмида содержит уникальные сайты узнавания эндонуклеазами рестрикции: ClaI (1251), PciI (2142), ApaI (4040), NcoI (5004), NheI (5045), EcoRI (5642), HindIII (6146).
Структура плазмиды pM-tPA приведена на Рис.3.
При помощи созданных плазмид можно трансформировать бактериальную клетку, предпочтительно бактерию, принадлежащей к роду Escherichia, восприимчивую к подобной трансформации указанными плазмидами. Выбор конкретной клетки не является критическим, поскольку методология и приемы трансформации хорошо известны специалисту в данной области техники. И хотя в зависимости от вида клетки и условий культивирования полученного трансформанта уровень экспрессии предшественника фрагмента ТАП человека может варьироваться, факт экспрессии целевого белка будет иметь место при условии успешной трансформации клетки - реципиента.
«Трансформация клетки плазмидой» означает введение плазмиды в клетку с помощью методов, хорошо известных специалисту в данной области техники. Трансформация этой плазмидой приводит к экспрессии гена, кодирующего белок согласно настоящему изобретению, и к синтезу белка в бактериальной клетке. Методы трансформации включают любые стандартные методы, известные специалисту в данной области техники, например метод, описанный в Jac A. Nickoloff, Electroporation Protocols for Microorganisms (Methods in Molecular Biology) // Humana Press; 1st edition (August 15, 1995).
Согласно настоящему изобретению «бактериальная клетка - продуцент предшественника фрагмента ТАП человека» означает бактериальную клетку, обладающую способностью к продукции и накоплению предшественника фрагмента ТАП человека согласно настоящему изобретению, когда бактериальная клетка согласно настоящему изобретению выращивается в указанной питательной среде. Используемый здесь термин «бактериальная клетка - продуцент предшественника фрагмента ТАП человека» также означает клетку, которая способна накапливать продукт предшественника фрагмента ТАП человека в количестве не менее чем 1 мг/л (или 1 мкг/109 клеток), более предпочтительно, не менее чем 40 мг/л. Указанный предшественник фрагмента ТАП человека накапливается в указанной клетке предпочтительно в виде телец включения.
Предпочтительно использование бактерии, принадлежащей к роду Escherichia, для трансформации рекомбинантной плазмидой, содержащей фрагмент ДНК, кодирующий предшественник фрагмента ТАП человека.
Термин «бактерия, принадлежащая к роду Escherichia» может означать, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, может быть упомянута бактерия Escherichia coli (Е.coli).
Круг бактерий, принадлежащих к роду Escherichia, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть приведены в качестве примеров.
Конкретным примером штамма-реципиента для получения продуцента предшественника фрагмента ТАП человека согласно настоящему изобретению, является, но не ограничиваются им, штамм Escherichia coli BL21[DE3].
Штамм Escherichia coli BL21[DE3] характеризуется следующими культурально-морфологическими, физиолого-биохимическими признаками и генетическими признаками.
Культурально-морфологические особенности штамма: грамотрицательные палочки, образуют нити; на агаризованной среде - беловатые крупные колонии с неровным краем. Активность штамма определяется методом денситометрия электрофореграммы. Штамм хранится в следующих условиях: среда Лурье-Бертрана, 1% глюкозы, 10% глицерина. Штамм размножается в следующих условиях - среда Лурье-Бертрана, 1% глюкозы, канамицина сульфат 30 мкг/мл.
Генетические особенности штамма. Генотип штамма - F- ompT gal dcm lon hsdSB(rB - mB -) λ(DE3 [lacI lacUV5-T7 gene 1 ind1 sam7 nin5])
Трансформация штамма Escherichia coli BL21[DE3] плазмидой p10E-tPA или плазмидой pM-tPA приводит к получению штамма-продуцента BL21[DE3]/p10E-tPA или штамма-продуцента BL21[DE3]/pM-tPA, соответственно, каждый из которых обеспечивает синтез рекомбинантного белка-предшественника фрагмента ТАП человека в количестве 30-70% от суммарного содержания белка клеток.
Штамм Escherichia coli BL21[DE3]/p10E-tPA кодирует белок-предшественник 10Е-tPA, состоящий из аминокислотной последовательности фрагмента ТАП человека и слитого в рамке N-концевого лидерного пептида длиной 23 аминокислоты, содержащего декагистидиновый кластер и сайт расщепления энтерокиназой, находящийся непосредственно перед первой аминокислотой фрагмента ТАП человека.
Штамм Escherichia coli BL21[DE3]/pM-tPA кодирует белок- предшественник М-tPA, состоящий из аминокислотной последовательности [-1] метионил-фрагмента ТАП человека.
Способ получения фрагмента ТАП человека согласно настоящему изобретению включает культивирование описанной выше бактерии в питательной среде, подходящей для выращивания указанных прокариотических клеток, индукцию промотора гена-предшественника фрагмента ТАП, выделение телец включения, солюбилизацию белка-предшественника, металлохелатную хроматографию в денатурирующих условиях, рефолдинг белка-предшественника, получение зрелого белка обработкой энтерокиназой и выделение указанного зрелого белка рекомбинантного фрагмента ТАП человека.
Особенности плазмид и результаты их практического применения приведены на следующих чертежах.
На Рис. 1 показаны схема сборки синтетического гена фрагмента ТАП человека из олигонуклеотидных праймеров и схема получения экспрессионных плазмид p10E-tPA, pM-tPA. Используются следующие обозначения: "ТРА (no SP, 527 аа)" - тканевый активатор плазминогена, без сигнального пептида и отщепляемых пропептидов; "А", "В" - цепи А и В соответственно; "FT1" - пальцеподобный домен фибронектина ("фибронектиновый палец"); "EG" - домен, гомологичный эпидермальному фактору роста; "KD1" и "KD2" - крингл-домены 1 и 2 (гомологичны крингл-доменам плазминогена); "S1" - протеолитический домен, гомологичный сериновым протеиназам; "RET1" и "RET2" - фрагменты, входящие в состав ретеплазы. В скобках указаны первая и последняя аминокислоты фрагментов. Пунктирная линия обозначает полимеразную цепную реакцию, сплошная - рестрикцию и лигирование фрагментов ДНК.
На Рис. 2 показана карта экспрессионной плазмиды p10E-tPA. Используются следующие обозначения: «pBR322ori» - область начала репликации плазмиды pBR322; «ROP» - ген РНК-организующего белка Rop; «f1 ori» - участок инициации репликации бактериофага f1; «KanR2» - последовательность, кодирующая аминогликозид-3'-фосфотрансферазу, обеспечивающую устойчивость бактерий к канамицину; «Т7 prom» - промотор РНК-полимеразы бактериофага Т7; «T7term» - участок терминации транскрипции; «lacI» - последовательность, кодирующая репрессор лактозного оперона; «ЮЕ-tPA» - открытая рамка считывания (ОРС) полипептида белка-предшественника фрагмента ТАП человека, включающего отделяемый N-концевой дополнительный пептид «10Е leader» и фрагмент ТАП человека «tPA»; «EK-PRO» - участок, кодирующий сайт узнавания энтерокиназой, «stop» - блок стоп-кодонов. Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
На Рис. 3 показана карта экспрессионной плазмиды pM-tPA. Используются обозначения аналогично Рис. 2, а также «M-tPA» - открытая рамка считывания (ОРС) полипептида белка-предшественника [-1] метионил-фрагмента ТАП человека.
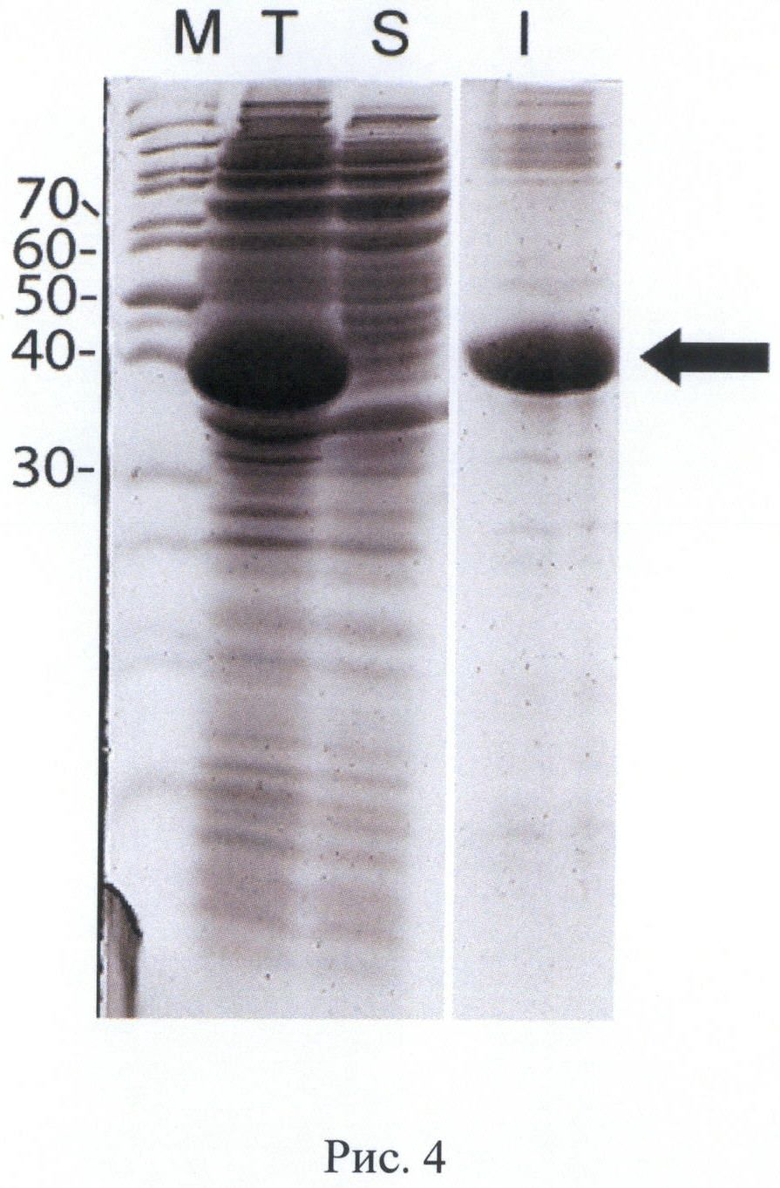
На Рис. 4 показана электрофореграмма тотального белка, фракций растворимого и нерастворимого белка клеток штамма-продуцента BL21[DE3]/p10E-tPA при индукции. Культивирование во встряхиваемой колбе, среда 2xYT, индукция 1 мМ ИПТГ, 37°С, 20 часов. Обозначения: Т - тотальный белок после индукции, S - фракция растворимых белков, I - фракция нерастворимых белков (тельца включения). М - маркер молекулярных масс. Молекулярные массы полос маркера указаны в кДа. Положение целевого белка указано стрелкой. Продуктивность: 300 мг/л, доля целевого белка: >60% (по денситометрии геля).
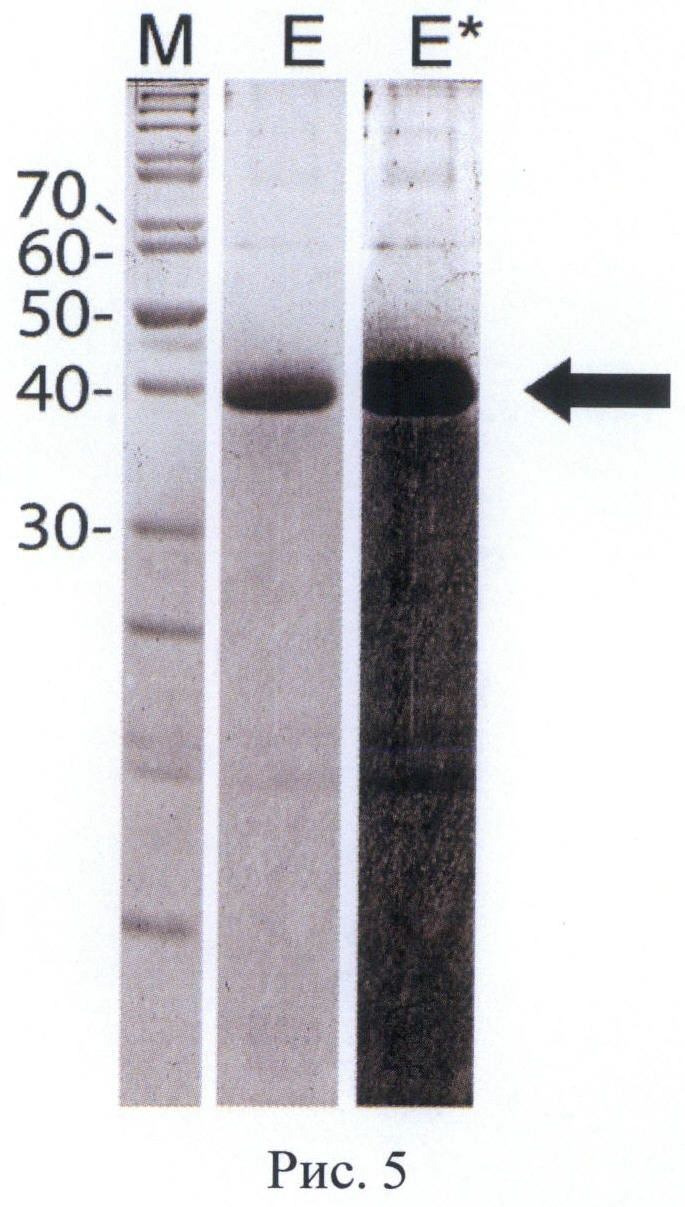
На Рис. 5 приведена электрофореграмма элюата, полученного при очистке белка-предшественника рекомбинантного фрагмента ТАП человека металлохелатной хроматографией в денатурирующих условиях. Обозначения: Е - фракция элюата раствором ЭДТА-Na; Е* - фракция элюата раствором ЭДТА-Na с усилением контраста изображения; М - маркер молекулярных масс. Молекулярные массы полос маркера указаны в кДа. Положение целевого белка указано стрелкой. Чистота: >98% (по денситометрии геля).
На Рис. 6 показана электрофореграмма рекомбинантного фрагмента ТАП человека после ренатурации на металлохелатной колонне. Обозначения: 1 - невосстанавливающие условия; 2 - восстанавливающие условия. 20% мономера по денситометрии геля.
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Получение плазмидной ДНК pAL-TPA
Для аминокислотной последовательности, включающей 355 из 527 аминокислот природного ТАП человека (аминокислоты 1-3 и 176-527) с добавленным 23-аминокислотным N-концевым лидерным пептидом (SEQ ID NO:3), содержащим последовательность узнавания энтерокиназой, была проведена обратная трансляция в последовательность нуклеотидов ДНК. При этом были использованы кодоны, оптимальные для экспрессии этого гена в Е.coli класса В, а также была проведена оптимизация структуры гена по вторичной структуре мРНК, GC составу, обеспечено отсутствие нежелательных регуляторных элементов (например, отсутствие внутренних сайтов связывания рибосом) а также отсутствие протяженных повторов, палиндромов.
Индекс CAI (Codon Adaptation Index), отражающий эффективность экспрессии гена в данном организме, для полученной последовательности составил 0,84, что является хорошим прогностическим показателем для промышленной пригодности полученного на его основе штамма-продуцента. Полученная нуклеотидная последовательность приведена в SEQ ID NO:4.
Синтетический ген ТАП собирали из двух фрагментов tPA-Fl (SEQ ID NO:5) и tPA-F2 (SEQ ID NO:6), а затем добавляли эпитоп для иммунохимической детекции методом ПЦР. Для этого получили конструкции pGEM-tPA-F1 и pAL-TPA-F2, содержащие фрагменты tPA-F1 и tPA-F2 соответственно, проводили их сборку в pGEM-tPA, проводили ПЦР и получали pAL-tPA с полным лидерным пептидом.
Фрагмент tPA-F1 (SEQ ID NO:5) собирали методом полимеразной цепной реакции (ПЦР) с праймерами T1F1-T1F7 и T1R1-T1R7 (SEQ ID NO:6-19), последовательности которых приведены в Таблице 1.
ПЦР проводили на приборе Терцик МС2 («ДНК-технология», Россия). Препаративные реакции проводили в объеме 50 мкл. Готовили инкубационную смесь следующего состава: 1х буфер для термостабильной ДНК-полимеразы; 10 пМ каждого праймерного олигонуклеотида; 2 мМ каждого дезоксирибонуклеотидтрифосфата; 2 ед. термостабильной ДНК-полимеразы. Поверх этой смеси наслаивали 50 мкл минерального масла и вели амплификацию по схеме: 1 цикл - денатурация - 94°С, 3 мин; 25 циклов - денатурация 94°С, 30 сек; отжиг 62°С, 30 сек; наращивание цепи 72°С, 120 сек; 1 цикл - наращивание цепи 72°С, 10 мин.
Продукты ПЦР выделяли из 1% агарозного геля, используя набор реактивов «Wizard SV Gel and PCR Clean-Up System» («Promega», США) по протоколу производителя, после чего лигировали в Т-вектор pGEM-T («Promega», США) с использованием ДНК-лигазы фага Т4 и стандартного буферного раствора («Fermentas», Литва). Лигирование вели в объеме 10 мкл при молярном соотношении вектора и вставки 1:10, в течение 2-20 часов при комнатной температуре. Полученными лигазными смесями трансформировали клетки Е.coli штамма DH5α, с генотипом F-φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17 (rk-, mk+) phoA supE44 λ- thi-1 gyrA96 relA1. Для этого к 200 мкл замороженной суспензии клеток Е.coli добавляли 5 мкл лигазной смеси, инкубировали на льду 20 мин для сорбции плазмидной ДНК, нагревали до 42°С 45 секунд и инкубировали на льду 5 мин. После чего добавляли 800 мкл питательного бульона LB и инкубировали при 37°С в течение 60 мин. После инкубации переносили суспензию на чашку Петри с твердой агаризованной средой, содержащей ампициллин в концентрации 100 мкг/1 мл агара и помещали в термостат на 18 часов при 37°С.
Колонии Е.coli, отобранные в результате бело-голубого скрининга, анализировали методом ПЦР с клонов, с использованием праймеров к последовательностям реципиентной плазмиды: M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21). Отобранные клоны наращивали в 5 мл питательного бульона LB и проводили выделение плазмидной ДНК набором реактивов «Wizard Plus SV Minipreps» («Promega», США) по протоколу производителя, первичную нуклеотидную последовательность конструкции pGEM-tPA-F1 подтверждали методом ПЦР-секвенирования с использованием праймеров T7prom (SEQ ID NO:22) и SP6 (SEQ ID NO:23).
Фрагмент tPA-F2 (SEQ ID NO:6) собирали методом ПЦР из праймеров T2F1-T2F6 и T2R1-T2R6 (SEQ ID NO:24-35), последовательности которых приведены в Таблице 1. ПЦР проводили так же, как при получении фрагмента tPA-F1. ПЦР продукт клонировали в Т-вектор pAL-TA (Евроген, Россия), проводили бело-голубой скрининг, анализировали методом ПЦР с колоний с использованием стандартных праймеров к последовательности вектора M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21), наращивали позитивные клоны в жидкой среде 2xYT-Amp, выделяли плазмидную ДНК при помощи набора «Gene JET Plasmid Miniprep Kit» («Fermentas», Литва), определяли нуклеотидную последовательность области вставки pAL-TA-TPAF2 методом ПЦР-секвенирования с использованием праймеров T7prom (SEQ ID NO:22) и SP6 (SEQ ID NO:23).
Получение конструкции pGEM-tPA (сборка фрагментов гена ТАП)
Плазмидную ДНК pGEM-tPA-F1, содержащую фрагмент tPA-F1 в прямой ориентации, расщепляли эндонуклеазой EcoRI («Permentas», Литва), дефосфорилировали с использованием щелочной фосфатазы («Fermentas», Литва); плазмидную ДНК pAL-TA-TPAF2 расщепляли избытком эндонуклеазы EcoRI. Продукты рестрикции плазмид pGEM-tPA-F1 и pAL-TA-TPAF2 подвергали разделению в 1% агарозном геле, выделяли набором WizardSV Gel and PCR Clean-Up System («Promega», США). Выделенные фрагменты лигировали с использованием Т-4 ДНК лигазы («Fermentas», Литва) по методике производителя. Анализировали методом ПЦР с колоний с использованием стандартных праймеров M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21), наращивали клоны, содержащие вставку, в жидкой среде 2xYT-Amp, выделяли плазмидную ДНК набором «GeneJET Plasmid Miniprep Kit» («Fermentas», Литва), подтверждали нуклеотидную последовательность области вставки pAL-TA-TPAP2 методом ПЦР-секвенирования с праймеров T7prom (SEQ ID NO:22) и SP6 (SEQ ID NO:23).
Введение в открытую рамку считывания рчТАП дополнительных аминокислот и второго стоп-кодона
Проводили 15 циклов ПЦР (по схеме: денатурация - 94°С, 3 мин; 15 циклов - денатурация 95°С, 15 сек, отжиг 55°С, 25 сек, наращивание цепи 72°С, 90 сек; финальное наращивание цепи 72°С, 10 мин) с использованием специфических праймеров AD-TPA-2stop-r (SEQ ID NO:36) и AD-TPA-DYKin-f (SEQ ID NO:37), взятых в количестве 10 пМ каждого, и 20 нг pGEM-tPA, использовавшейся в качестве матрицы. Продукты ПЦР разделяли в 1% агарозном геле, вырезали из геля нужный фрагмент и проводили выделение ДНК набором реактивов «Wizard SV Gel and PCR Clean-Up System» («Promega», CLUA) по протоколу производителя. Очищенные продукты лигировали в векторную плазмиду pAL-TA («Евроген», Россия) с использованием ДНК-лигазы фага Т4 и стандартного буферного раствора («Fermentas», Литва). Лигирование вели в объеме 20 мкл, при молярном соотношении вектора и вставки 1:10, в течение 10 часов при комнатной температуре. Полученными лигазными смесями трансформировали клетки Е.coli штамма DH5α. Колонии Е.coli, отобранные в результате бело-голубого скрининга, анализировали методом ПЦР с клонов, с использованием праймеров M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21). Отобранные клоны наращивали в 5 мл питательного бульона LB и проводили выделение плазмидной ДНК с использованием набора «Wizard Plus SV Minipreps» («Promega», США) по протоколу производителя. Полученные генетические конструкции pAL-tPA анализировали методом ПЦР-секвенирования с праймеров M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21).
После этого проводили препаративную рестрикцию плазмиды pAL-tPA эндонуклеазами рестрикции NheI и HindIII. Продукты реакции разделяли в 1% агарозном геле и выделяли с помощью набора реактивов Wizard SV Gel&PCR Clean-Up System no методике производителя донорный фрагмент для получения экспрессионной плазмиды p10E-tPA.
Пример 2. Получение векторной плазмиды р10Е
Последовательность, кодирующую фрагмент N-концевого лидерного пептида, включающего декагистидиновый кластер, получали методом отжига двух частично комплементарных олигонуклеотидов AD-10H-NcoF (SEQ ID NO:38) и AD-10H-NheR (SEQ ID NO:39) с образованием выступающих «липких» 5'-концов. Для этого вносили в пробирку по 100 пм каждого олигонуклеотида, нагревали до 95°С и медленно охлаждали до комнатной температуры.
Реципиентную плазмиду рЕТ28а (+) (Novagen) расщепляли последовательно каждой из эндонуклеаз NcoI и NheI. Сначала аликвоты плазмиды инкубировали 2 часа при 37°C с каждой из рестриктаз, затем контролировали электрофорезом в агарозном геле полную линеаризацию плазмид (в пределах чувствительности метода), после чего добавляли вторую рестриктазу и инкубировали еще 2 часа. Затем пробы объединяли, рестриктазы инактивировали прогреванием при 65°С в течение 20 минут. Рестрицированную плазмиду переосаждали 3 объемами этанола, центрифугировали 10 минут на скорости 13200 об/мин при комнатной температуре, промывали осадок 70% спиртом, растворяли в воде и использовали для постановки реакции лигирования в рабочей концентрации 10-2 мкг/мкл. Выделенные фрагменты лигировали с использованием Т-4 ДНК лигазы («Fermentas», Литва) по методике производителя. После этого проводили трансформацию клеток Е.coli штамма DH5α, как описано выше. Колонии Е.coli анализировали методом ПЦР с клонов, с использованием олигонуклеотидов AD-10H-NcoF (SEQ ID NO:38) и T7t (SEQ ID NO:40). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, выделяли плазмидную ДНК набором «GeneJET Plasmid Miniprep Kit» («Fermentas», Литва) по протоколу производителя. Для полученных генетических конструкций р10Е определяли нуклеотидную последовательность ДНК методом ПЦР-секвенирования с праймера T7t (SEQ ID NO:40).
Пример 3. Получение экспрессионной плазмидной ДНК p10E-tPA
Реципиентную плазмиду р10Е расщепляли последовательно каждой из эндонуклеаз NheI и HindIII. Сначала аликвоты плазмиды инкубировали при 37°С в течение 2 часов с каждой из рестриктаз, контролировали электрофорезом в агарозном геле полную линеаризацию плазмид, добавляли вторую рестриктазу и инкубировали еще 2 часа. Затем пробы объединяли, ферменты инактивировали прогреванием при 65°С в течение 20 минут и проводили дефосфорилирование щелочной фосфатазой («Fermentas», Литва) по протоколу производителя. Щелочную фосфатазу инактивировали прогреванием до 85°С в течение 20 минут. Расщепленную и дефосфорилированную плазмиду переосаждали 3 объемами этанола, центрифугировали 10 минут на скорости 13200 об/мин при комнатной температуре, промывали осадок 70% спиртом, растворяли в воде и использовали для постановки реакции лигирования в рабочей концентрации 10-2 мкг/мкл.
Реакцию лигирования очищенного фрагмента, соответствующего мини-гену tPA и реципиентной плазмиды проводили, как описано выше. После этого проводили трансформацию клеток Е.coli штамма DH5α, как описано выше.
Колонии Е.coli анализировали методом ПЦР с клонов с праймеров T7prom (SEQ ID NO:22) и T7t (SEQ ID NO:40). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, проводили выделение плазмидной ДНК с набором «Wizard Plus SV Minipreps» («Promega», США) по протоколу производителя. При помощи ПЦР-секвенирования для конструкции p10E-tPA определяли нуклеотидную последовательность обеих комплементарных цепей ДНК для области вставки с использованием праймеров T7prom (SEQ ID NO:22) и T7t (SEQ ID NO:40). В результате секвенирования установили, что в полученном препарате плазмиды не содержатся мутации в области вставки, то есть кодируется корректная последовательность гена ТАП. Структура плазмиды приведена на Фиг.2.
Пример 4. Получение плазмидной ДНК pM-tPA
Проводили 15 циклов ПЦР (по схеме: денатурация 94°С, 3 мин; 15 циклов денатурация 95°С, 15 сек; наращивание цепи 55°С, 25 сек; достройка 72°С, 90 сек; финальное наращивание цепи 72°С, 10 мин) с использованием специфических праймеров AD-TPAM-PciF (SEQ ID NO:41) и AD-TPA-2stop-r (SEQ ID NO:36), взятых в количестве 10 пМ каждого, и 20 нг pAL-tPA, использовавшейся в качестве матрицы. Продукты ПЦР разделяли в 1% агарозном геле, вырезали из геля нужный фрагмент и проводили его выделение набором «Wizard SV Gel and PCR Clean-Up System» («Promega», США). Очищенные продукты лигировали в векторную плазмиду pAL-TA («Евроген», Россия) с использованием ДНК-лигазы фага Т4 и стандартного буферного раствора («Fermentas», Литва). Лигирование вели в объеме 20 мкл при молярном соотношении вектора и вставки 1:10, в течение 18 часов при комнатной температуре, затем трансформировали лигазными смесями клетки Е.coli штамма DH5α. Колонии Е.coli, отобранные в результате бело-голубого скрининга, анализировали методом ПЦР с клонов с праймеров M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Amp и проводили выделение из них плазмидной ДНК набором «Wizard Plus SV Minipreps» («Promega», США). Полученные генетические конструкции pAL-M-tPA анализировали методом ПЦР-секвенирования с праймеров M13for (SEQ ID NO:20) и M13rev (SEQ ID NO:21).
После этого проводили препаративную рестрикцию плазмиды pAL-M-tPA эндонуклеазами рестрикции PciI и HindIII. Продукты реакции разделяли в 1% агарозном геле и выделяли с помощью набора реактивов «Wizard SV Gel&PCR Clean-Up System».
Реципиентную плазмиду р10Е расщепляли последовательно каждой из эндонуклеаз NcoI и HindIII, для этого аликвоты плазмиды инкубировали 2 часа при 37°C с каждой из рестриктаз, контролировали электрофорезом в агарозном геле линеаризацию плазмид, добавляли вторую рестриктазу и инкубировали еще 2 часа. Затем пробы объединяли, ферменты инактивировали прогреванием при 65°С в течение 20 минут и проводили дефосфорилирование щелочной фосфатазой («Fermentas», Литва) по протоколу производителя. Щелочную фосфатазу инактивировали прогреванием до 85°С в течение 20 минут. Расщепленную и дефосфорилированную плазмиду переосаждали 3 объемами этанола, центрифугировали 10 минут на скорости 13200 об/мин при комнатной температуре, промывали осадок 70% спиртом, растворяли в воде и использовали для постановки реакции лигирования в рабочей концентрации 10-2 мкг/мкл. Реакцию лигирования очищенного фрагмента, соответствующего мини-гену M-tPA и реципиентной плазмиды проводили, как описано выше. После этого проводили трансформацию клеток Е.coli штамма DH5α. Колонии Е.coli анализировали методом ПЦР с клонов с праймеров T7prom (SEQ ID NO:22) и T7t (SEQ ID NO:40). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, выделяли из них плазмидную ДНК набором «Wizard Plus SV Minipreps». При помощи ПЦР-секвенирования для конструкции pM-tPA определяли нуклеотидную последовательность обеих комплементарных цепей ДНК для области вставки с использованием праймеров T7prom (SEQ ID NO:22) и T7t (SEQ ID NO:40). В результате секвенирования установили, что в полученном препарате плазмиды pM-tPA не содержатся мутации в области вставки, то есть кодируется корректная последовательность гена [-1]М-ТАП. Структура плазмиды приведена на Рис.3.
Пример 5. Получение штаммов-продуцентов Е.coli BL21[DE3]/p10E-tPA и Е.coli BL21[DE3]/pM-tPA, оценка продуктивности штаммов-продуцентов и локализация целевого белка
Для получения штамма-продуцента рекомбинантного фрагмента ТАП конструкции, полученные в Примерах 3 и 4, использовали для трансформации компетентных клеток Escherichia coli BL21(DE3) (с генотипом F- ompT hsdSB (r-m-) gal dcm (DE3)) и проводили отбор клонов, сохраняющих уровень биосинтеза рекомбинантного полипептида после индукции не ниже 30% от суммарного клеточного белка в течение, по крайней мере, четырех последовательных пассажей.
Для получения штамма Е.coli BL21[DE3]/p10E-tPA - продуцента белка-предшественника фрагмента ТАП человека клетки штамма Е.coli BL21[DE3] трансформировали экспрессионной плазмидой p10E-tPA.
Для получения штамма Е.coli BL21[DE3]/pM-tPA - продуцента [-1]метионил-фрагмента ТАП человека клетки штамма Е.coli BL21[DE3] трансформировали экспрессионной плазмидой pM-tPA.
Трансформанты Е.coli BL21[DE3] высевали на агаризованную среду 2xYT с добавлением канамицина до 30 мкг/мл и глюкозы до 2%, проводили аналитическую экспрессию целевых белков для пяти случайно выбранных клонов типичного фенотипа. Клоны подращивали в питательном бульоне с добавлением канамицина до 30 мкг/мл и раствора глюкозы до 2% в течение 6-7 часов, инокулировали новую порцию питательной среды в соотношении 1:100, растили культуру до достижения оптической плотности 2 О.Е., индуцировали изопропилтио-β-D-галактозидом (ИПТГ) и культивировали в течение еще 2 часов. После окончания культивирования осадок клеток отделяли центрифугированием, ресуспендировали клетки в растворе 10 мМ Трис-HCl, 2 мМ ЭДТА-Na, 0,1% Тритона-Х100, 10 мкг/мл лизоцима в соотношении 10 мл раствора на 1 г клеточной пасты, выдерживали суспензию 30 мин на льду и проводили разрушение клеток ультразвуковым диспергатором до исчезновения видимой вязкости суспензии. Отбирали образцы для электрофоретического анализа, разделяли в них растворимую и нерастворимую фракцию белков центрифугированием в микроцентрифуге, дополнительно ресуспендировали осадок в том же растворе и осаждали центрифугированием. Результаты электрофоретического анализа тотального белка и фракций растворимых и нерастворимых белков для штамма BL21[DE3]/p10E-tPA приведены на Рис.4.
Электрофоретическая подвижность обоих целевых белков соответствует расчетным значениям, интенсивность полосы целевого белка для обоих целевых белков аналогична. По данным гель-электрофореза белковых фракций оба целевых белка практически полностью были локализованы в нерастворимой фракции белков, т.е. находились в форме «телец включения».
Пример 6. Выделение, очистка и ренатурация рекомбинантного ТАП
Штамм-продуцент BL21[DE3]/p10E-tPA высевали из музея петлей истощающим штрихом на чашку Петри с агаризованной средой LB, содержащей 30 мкг/мл канамицина и 1% глюкозы, растили в течение 14 часов при 37°С. Одну отдельную колонию штамма переносили в 5 мл жидкой среды LB, содержащей 30 мкг/мл канамицина и 1% глюкозы, и растили на качалке в течение 14 часов при 37°С. Содержимым инокулировали 250 мл среды 2xYT, содержащей 30 мкг/мл канамицина и 0,1% глюкозы, растили на качалке в течение 3,5 часов при 37°С, отбирали образцы бактериальной суспензии для анализа, добавляли ИПТГ до конечной концентрации 1 мМ и растили в течение еще 3-15 ч.
Осадок клеток отделяли центрифугированием, ресуспендировали в 20 мл раствора А (50 мМ Трис рН=7,4, 2 мМ ЭДТА), добавили лизоцим до 0,1 мг/мл и Тритон X100 до 0,1%, инкубировали 30 мин на льду. Проводили разрушение клеток и геномной ДНК при помощи ультразвукового диспергатора пульсами по 10 с до исчезновения повышенной вязкости суспензии. Отделяли осадок центрифугированием 10 мин при 20000 об/мин. Осадок ресуспендировали в растворе А, добавляли детергент NP-40 до 1%, суспензию обрабатывали ультразвуковым диспергатором до исчезновения частиц осадка крупнее 1 мм, отделяли осадок центрифугированием 10 мин при 20000 об/мин. Полученный осадок ресуспендировали в растворе А, добавляли NaCl до 500 мМ, суспензию обрабатывали ультразвуковым диспергатором до исчезновения частиц осадка крупнее 1 мм, отделяли осадок центрифугированием 10 мин при 20000 об/мин. Полученный осадок ресуспендировали в растворе 50 мМ Трис рН 7,4, суспензию обрабатывали ультразвуковым диспергатором до исчезновения частиц осадка крупнее 1 мм, отделяли осадок центрифугированием 10 мин при 20000 об/мин. Полученный осадок обогащенных телец включения хранили при -70°С.
Для проведения солюбилизации целевого белка к осадку телец включения добавляли раствор Б (6 М гуанидинхлорида, 50 мМ фосфата натрия, 50 мМ бета-меркаптоэтанола, рН 8,0) в соотношении 10 мл раствора на 1 г осадка. Суспензию инкубировали при перемешивании в течение 2 часов при 37°С, отделяли нерастворившийся клеточный дебрис центрифугированием 10 мин при 20000 об/мин. Супернатант разбавляли в 5 раз раствором В (6 М мочевины, 50 мМ фосфата натрия, 300 мМ хлорида натрия, рН 8,0) для уменьшения конечной концентрации бета-меркаптоэтанола, отделяли выпавший осадок центрифугированием 10 мин при 20000 об/мин и наносили супернатант на колонку с сорбентом Chelating Sepharose Fast Flow («GE Healthcare», CHIA), содержащим хелатированные ионы никеля, и уравновешенным раствором В. Последовательно промывали колонку раствором Г (6 М мочевины, 50 мМ фосфата натрия, 300 мМ хлорида натрия, рН 7.0) и раствором Д (6 М мочевины, 50 мМ фосфата натрия, 300 мМ хлорида натрия, 50 мМ имидазола рН 7,0) до стабилизации базовой линии. Элюировали очищенный белок раствором Е (6 М мочевины, 50 мМ фосфата натрия, 300 мМ хлорида натрия, 50 мМ ЭДТА-Na, рН 7,0). Элюат концентрировали ультрафильтрацией до конечной концентрации белка 20 мг/мл и обессоливали диафильтрацией, полученный раствор хранили в замороженном виде. Полипептид был очищен в денатурирующих условиях до видимой гомогенности (более 95% чистоты по денситометрии электрофореграммы, Рис.5).
Для рефолдинга в растворе к размороженному белку добавляли ДТТ до концентрации 10 мМ и инкубировали при 37°С в течение 2 часов. Затем белок разбавляли буферным раствором для рефолдинга (150 мМ ТРИС, рН 8,5, 0,5 М аргинина, 2 мМ ЭДТА, 0,2 мМ восстановленного глютатиона, 0,05% Твин-80 и 2,25 М мочевины) до концентрации 7 мкг/мл и инкубировали при комнатной температуре в течение трех суток. После этого белок вновь концентрировали до 2 мг в мл. Полученный препарат обладал 50-90% удельной протеолитической активности в сравнении со стандартом ретеплазы.
Рефолдинг белка-предшественника M-tPA проводили аналогично описанной выше процедуре, но не проводили металлохелатную хроматографию в денатурирующих условиях, а вводили в рефолдинг восстановленный денатурированный белок непосредственно из состава телец включения. Удельная протеолитическая активность 10Е-tPA и M-tPA не имела достоверных отличий.
Подлинность белка-предшественника 10Е-tPA исследовали при помощи масс-спектрометрии методом SELDI. Для этого аликвоту ренатурированного белка подвергали восстановлению дисульфидных связей дитиотрейтолом, блокировке свободных остатков цистеина иодоацетамидом и наносили на поверхность чипа Н-50 (Ciphergen, США), адсорбировали белки и промывали гнезда чипа по инструкции производителя и наносили на гнезда чипа насыщенный раствор матрицы. Масс-спектр образца снимали в приборе ProteinChip Reader II (Ciphergen, США). Была зафиксирована доминирующая молекулярная масса 43497 Да при значительной ширине и ассиметрии пика. Теоретически рассчитанная молекулярная масса белка 10Е-tPA с учетом проведенных химических модификаций составляет 43344 Да и находится в удовлетворительном соответствии с экспериментально найденной - ошибка определения 3,5%.
Пример 7. Очистка и ренатурация иммобилизованного 10Е-tPA
Рефолдинг на колонке проводили по общей схеме из Fan et. al. (J. Biophis. Biochem. meth., 2008) с изменениями.
1. Тельца включения растворяли в 8М гуанидин-гидрохлориде, 50 мМ Трис рН 7,5 и 50 мМ бета-меркаптоэтанола в соотношении 10 мл раствора на 1 г телец. Полученный раствор разводили в 5 раз раствором А (8 М мочевины, 0,5 М NaCl, 50 мМ Трис рН 7,5) и наносили на колонку с сорбентом Ni-NTA (Qiagen), уравновешенную тем же раствором, в соотношении 20 мг телец на 1 мл сорбента.
2. Промывали колонку 5 объемами раствора А и 5 объемами раствора А, дополнительно содержащими 0,5 М аргинина гидрохлорида.
3. В течение 10 объемов колонки проводили линейный градиент от раствора А (8 М мочевины, 0,5 М NaCl, 50 мМ ТРИС рН 7,5) к раствору В (100 мМ ТРИС рН 8,5, 0,5 М аргинина, 0,05% Твин 80).
4. Проводили элюцию целевого белка раствором 0,5 М аргинина, 2 М мочевины, 0,05% Твин 80, 50 мМ ЭДТА, 50 мМ ТРИС рН 8,5, 0,5 М NaCl.
Анализ элюированного ренатурированного 10Е-tPA проводили при помощи ДСН-ПААГ в невосстанавливающих условиях (Рис. 6). Чистота целевого белка после очистки составила >90% по денситометрии электрофореграмм. Типичный выход очищенного и ренатурированного 10Е-tPA составлял 30-50%. Протеолитическую активность 10Е-tPA измеряли методом гидролиза синтетического субстрата Т2943 (CH3SO2-D-HHT-Gly-Arg-pNA·AcOH, где ННТ - гексагидротирозин, pNA - п-нитроанилид, производство Sigma-Aldrich, США) по методике производителя субстрата. Удельная протеолитическая активность мономера ренатурированного 10Е-tPA составляла не менее 10% от стандарта ретеплазы.
Пример 8. Отделение лидерного пептида и очистка зрелого белка tPA
Отделение N-концевого пептида проводили при помощи рекомбинантной энтерокиназы ("Sigma", США), используя соотношение фермент:субстрат 1:10 (по массе). Реакцию вели в течение 2 ч при 37°С в присутствии 5 мМ хлорида кальция. Энтерокиназу, непрореагировавший белок-предшественник 10Е-tPA, свободный пептид 10Е и остатки примесных белков отделяли, пропуская реакционную смесь через колонку, заполненную сорбентом Chelating Sepharose FF ("GE Healthcare", США) с иммобилизованными ионами кобальта. Полученный полностью очищенный раствор зрелого белка tPA, находящегося во фракции проскока с колонки, концентрировали и обессоливали ультрафильтрацией и хранили при температуре -20°С. Удельная протеолитическая активность зрелого белка tPA и белка-предшественника 10Е-tPA не имела достоверных отличий.
Приведенные результаты показали, что включение в последовательность синтезируемого белка короткого дополнительного N-концевого пептида, кодируемого оптимальными для Е.coli кодонами, позволяет проводить хроматографическую очистку экспрессируемого белка в денатурирующих условиях методом металлохелатной хроматографии и совмещать с этой стадией очистки процедуру рефолдинга иммобилизованного белка, что, в свою очередь, позволяет получать полностью очищенный и ренатурированный продукт при помощи одной хроматографической стадии. При необходимости данный пептид может быть удален обработкой энтерокиназой и последующей повторной металлохелатной хроматографией.
Преимущества предлагаемого штамма Е.coli BL21[DE3] заключаются в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого de novo рекомбинантного ТАП и контаминации выделяемого белка наиболее активными протеазами Е.coli. Встроенный в геном штамма-реципиента ген РНК полимеразы бактериофага Т7 под контролем lacUV5 промотора при использовании Т7-1ас промотора и Т7 терминатора в плазмидах приводит к быстрой и эффективной продукции белка.
Еще одним общим преимуществом использованного штамма, экспрессионного вектора и стратегии биосинтеза является возможность проводить индукцию без изменения температуры культивирования.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.













Изобретение относится к биотехнологии и представляет собой способ получения предшественника рекомбинантного фрагмента тканевого активатора плазминогена (ТАП) человека (ретеплазы) с использованием бактерии, принадлежащей к роду Escherichia, трансформированной экспрессионной плазмидой, содержащей фрагмент ДНК, кодирующий предшественник рекомбинантного фрагмента тканевого активатора плазминогена (ТАП) человека (ретеплазы), включающий последовательность, кодирующую отщепляемый N-концевой пептид, включающий декагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназы, или фрагмент ДНК, кодирующий [-1]метионил-фрагмента ТАП человека (ретеплазы), под контролем промотора, функционирующего в указанной бактериальной клетке. Предложенное изобретение позволяет получать рекомбинантный фрагмент тканевого активатора плазминогена (ТАП) человека (ретеплазы) с высоким выходом. 4 н. и 6 з.п.ф-лы, 6 ил., 1 табл., 8 пр.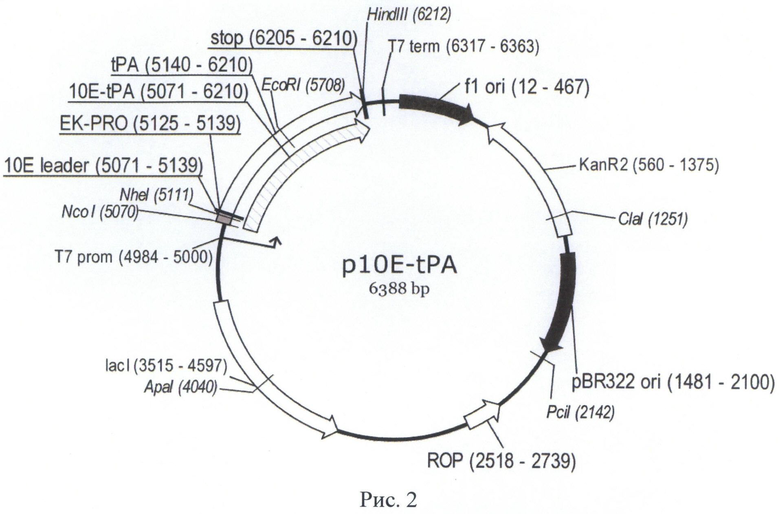
1. Плазмида для экспрессии в клетках бактерии рода Escherichia предшественника рекомбинантного фрагмента тканевого активатора плазминогена (ТАП) человека (ретеплазы), содержащего отщепляемый N-концевой пептид, включающий декагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, или [-1]метионил-фрагмента ТАП человека (ретеплазы), под контролем промотора, функционирующего в бактериальной клетке, в следующей последовательности по существу содержащая:
- фрагмент вектора рЕТ28а(+), содержащий участок инициации репликации бактериофага f1, последовательность, кодирующую аминогликозид-3'-фосфотрансферазу, область начала репликации плазмиды pBR322, ген РНК-организующего белка Rop;
- последовательность, кодирующую репрессор лактозного оперона;
- фрагмент ДНК, содержащий промотор, функционирующий в бактериальной клетке, и последовательность, кодирующую остаток [-1] метионина, или синтетический адаптор, кодирующий отщепляемый N-концевой лидер, включающий декагистидиновый кластер и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей предшественник рекомбинантного фрагмента ТАП человека (ретеплазу);
- фрагмент ДНК, кодирующий предшественник рекомбинантного фрагмента ТАП человека (ретеплазу) или [-1]метионил-фрагмент ТАП человека (ретеплазу); и
- участок терминации транскрипции.
2. Плазмида по п.1, отличающаяся тем, что указанным фрагментом ДНК, кодирующим предшественник рекомбинантного фрагмента ТАП человека с отщепляемым N-концевым пептидом, включающим декагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, является фрагмент ДНК, представленный в Перечне последовательностей под номером SEQ ID NO:1 или его вариант.
3. Плазмида по п.1, отличающаяся тем, что указанным промотором является промотор РНК-полимеразы бактериофага Т7
4. Плазмида по п.1, отличающаяся тем, что указанная плазмида выбрана из группы, состоящей из плазмид p10E-tPA, pM-tPA.
5. Бактерия, принадлежащая к роду Escherichia, трансформированная плазмидой по п.1, - продуцент предшественника рекомбинантного фрагмента ТАП человека (ретеплазы) или [-1]метионил-фрагмент ТАП человека (ретеплазы).
6. Бактерия по п.5, отличающаяся тем, что указанная бактерия выбрана из группы, включающей бактерии Е.coli BL21[DE3]/p10E-tPA, E.coli BL21[DE3]/pM-tPA.
7. Предшественник рекомбинантного фрагмента ТАП человека (ретеплазы) с отщепляемым N-концевым пептидом, включающим декагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, полученный путем культивирования бактерии по п.5 в питательной среде.
8. Предшественник рекомбинантного фрагмента ТАП человека (ретеплазы) по п.7, содержащий последовательность аминокислот, представленную в Перечне последовательностей под номером SEQ ID NO:2 или ее функциональный вариант.
9. Способ получения предшественника рекомбинантного фрагмента ТАП человека (ретеплазы) по п.7, включающий культивирование бактерии по п.5 в питательной среде, выделение телец включения, солюбилизацию белка-предшественника, металлохелатную хроматографию в денатурирующих условиях, рефолдинг белка-предшественника, получение зрелого белка обработкой энтерокиназой и выделение указанного зрелого белка фрагмента ТАП человека.
10. Способ по п.9, отличающийся тем, что культивируют штамм Е.coli BL21[DE3]/p10E-tPA.
| RU 2006110371 А, 10.10.2007 | |||
| ПРЕПАРАТ ЧЕЛОВЕЧЕСКОГО ЭНДОСТАТИНА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2004 |
|
RU2278688C1 |
| МОДИФИЦИРОВАННЫЕ ИНГИБИТОРЫ ПРОТЕИНАЗЫ | 1995 |
|
RU2182598C2 |
| База данных PatSearch: Tissue-type plasminogen activator chain B, номер E7ESF4, код E7ESF4-HUMAN, 08.03.2011 | |||
| Способ определения частотной характеристики рекордера | 1939 |
|
SU58123A1 |
| RU | |||

