Изобретение относится к области биомолекулярной фармакологии и биотехнологии, конкретно к препарату эндостатина человека и способу его получения.
Эндостатин обладает особенно большим потенциалом в антиангиогенной терапии. Ангиогенез - процесс образования новых кровеносных сосудов из предсуществующих, который играет ключевую роль в прогрессии и метастазировании злокачественных новообразований. В организме здорового взрослого человека наблюдается низкая интенсивность ангиогенеза в большинстве органов и тканей, за исключением физиологически значимых процессов, связанных с репарацией поврежденных тканей и репродуктивным циклом у женщин.
Во взрослом здоровом организме ангиогенез находится под жестким контролем регуляторных систем организма, обеспечивающих баланс между позитивными и негативными регуляторами ангиогенеза, и в основном поддерживается на низком уровне или носит периодический и кратковременный характер. Неадекватная или несбалансированная васкуляризация является причиной многочисленных патологических состояний, приводящих к таким заболеваниям как ишемическая болезнь сердца, ретинопатия, атеросклероз и др. Особенно важную роль ангиогенез играет в процессе развития и метастазирования злокачественных новообразований, в связи с чем разъяснение механизмов опухолевого ангиогенеза и разработка терапевтических агентов для воздействия на них приобретают особую актуальность.
Условием прогрессии и метастатической активности опухоли является привлечение кровеносных капилляров из предсуществующих сосудов. В противном случае опухоль не достигает крупных размеров и остается в "дремлющем" состоянии, при котором индекс пролиферации опухолевых клеток уравновешивается индексом их апоптоза. В таком состоянии опухоль достигает размеров 1-3 мм3, и для ее прогрессии необходимо "переключение" опухолевых клеток на ангиогенный фенотип. В качестве "антиангиогенного переключателя" выступает соотношение между проангиогенными и антиангиогенными молекулами. Таким образом, "антиангиогенный переключатель" находится в выключенном состоянии, когда превалируют антиангиогенные факторы, и во включенном, когда преобладают ангиогенные факторы. В настоящее время идентифицировано более 20 эндогенных активаторов и ингибиторов ангиогенеза [1, 2, 3, 4].
Даже при длительном терапевтическом применении эндостатин не вызывает возникновения резистентности у опухолевых клеток и при этом обладает ярко выраженным терапевтическим эффектом. Это особенно важно, поскольку такие свойства открывают возможности для долговременного воздействия на опухоли, характеризующиеся высокой биологической агрессивностью и метастатическим потенциалом. Очевидно, что этот антиангиогенный агент может быть использован не только в противоопухолевой терапии, но и при лечении других ангиогенных болезней. Важной особенностью антиангиогенной терапии является возможность ее комбинированного применения с другими видами терапии, такими как радио-, иммуно- или химиотерапия. Поскольку значение ангиогенеза во взрослом здоровом организме не столь велико, как в развивающемся, применение антиангиогенной терапии без существенных побочных эффектов имеет самые широкие перспективы.
Эндостатин является мощным ингибитором ангиогенеза и представляет собой фрагмент коллагена XVIII типа α-1, являющегося компонентом базальных мембран и внеклеточного матрикса [5]. Эндостатин специфически ингибирует пролиферацию и миграцию эндотелиальных клеток капилляров и может вызывать апоптоз пролиферирующих эндотелиальных клеток. Непосредственного эффекта на опухолевые клетки различных линий in vitro эндостатин не оказывает [6]. In vivo эндостатин оказывает сильный ингибирующий эффект в отношении роста различных первичных опухолей и их метастазов. Пролонгированная терапия высокими дозами эндостатина вызывает практически полную блокаду опухолевого ангиогенеза и приводит к регрессии опухоли до микроскопических размеров. В дальнейшем микроопухоли впадают в "дремлющее" состояние, в котором индекс пролиферации опухолевых клеток уравновешивается индексом их апоптоза. Даже после пролонгированной терапии не наблюдается возникновения резистентности к препарату или каких-либо токсических реакций [7]. Механизм действия эндостатина до сих пор не ясен. Эндостатин может ингибировать индуцируемый ангиогенными факторами роста каскад внутриклеточной передачи сигналов, а также блокировать активацию и каталитическую активность матриксных металлопротеиназ [8]. В настоящее время рекомбинантный эндостатин проходит начальную стадию клинических испытаний в качестве антиангиогенного противоопухолевого препарата. Эндостатин может также найти применение при лечении болезней, связанных с нарушением ангиогенеза (атеросклероз, ревматоидный артрит, псориаз, диабетическая ретинопатия и др.) [9].
Широкие терапевтические перспективы диктуют необходимость разработки эффективных систем получения отечественного рекомбинантного эндостатина. К настоящему времени был разработан ряд систем экспрессии эндостатина, и целевой белок был экспрессирован в Е.coli, клетках Spodoptera frugiperda, трансфецированных бакуловирусом, Pichia pastoris, стабильно трансформированных клетках Drosophila melanogaster, трансфецированных эмбриональных человеческих клетках почек 293-EBNA. Из всех перечисленных систем экспрессия эндостатина в клетках Е.coli является наименее затратной и наиболее технологически простой системой, способной обеспечить высокие выходы биологически активного белка.
Аналогом технической сущности изобретения является рекомбинантная ДНК и ее штамм-продуцент на базе Е.coli, которые пригодны для получения препаративных количеств биологически активного мышиного эндостатина [10]. Рекомбинантная ДНК содержит нуклеотидную последовательность гена эндостатина, слитую с нуклеотидной последовательностью сигнального пептида щелочной фосфатазы, под контролем фосфатазного промотора (phoA) E.coli. Вектор, содержащий рекомбинантную ДНК, устойчиво экспрессируется в Е.coli, обеспечивает стабильную секрецию белка в культуральную среду и после выделения дает биологически активный правильно процессированный мышиный эндостатин. Однако используемый штамм-продуцент обеспечивает получение продукта, требующего дополнительной стадии концентрирования культуральной среды и двустадийной хроматографической очистки, включающей последовательные хроматографии на дорогостоящем носителе гепарин-сефарозе. Кроме того, в результате использования сильнозагрязненной различными примесями культуральной среды для нанесения на колонку с гепарин-сефарозой последняя быстро приходит в негодность, что значительно увеличивает затраты на производство эндостатина. Для получения высокоочищенного белка с использованием описанной авторами технологии необходима дополнительная хроматографическая стадия с привлечением высокоэффективной жидкостной хроматографии (ВЭЖХ).
Наиболее близким аналогом нашей разработки является способ производства эндостатина человека и мыши, описанный в патенте РФ №2225728, опубл. в марте 2004 г., включающий культивирование клеток-хозяев, которые экспрессируют ген эндостатина, выделение продукта экспрессии, рефолдинг генного продукта при рН 6,0-8,5, очистку конечного продукта методом ионообменной хроматографии или ВЭЖХ. Однако этот способ предусматривает применение довольно сложных и дорогостоящих векторов, а по оптимальным режимам отдельных этапов суммарно вся процедура получения является более длительной.
Технической задачей настоящего изобретения является создание апирогенного препарата человеческого эндостатина отечественного производства, полученного с применением более дешевых исходных материалов и сокращенным по времени процессом производства.
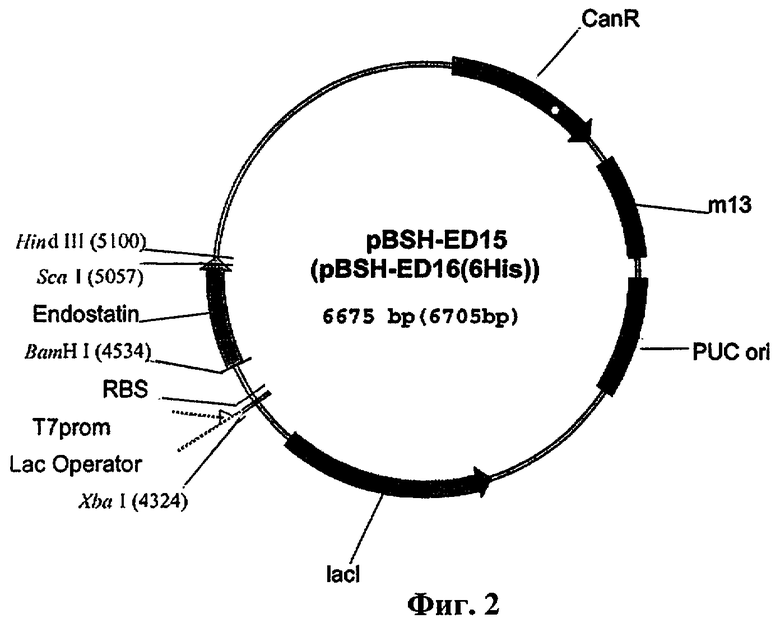
Технический результат достигается тем, что препарат человеческого эндостатина, представляющий собой рекомбинантный белок с молекулярной массой 20 кДа, полученный из трансформированных плазмидным вектором клеток Е.coli и обладающий ингибирующим действием в отношении пролиферации эндотелиальных клеток, отличается тем, что препарат выделен после инкубации клеток Е.coli BL21 (ВЕЗ) в ростовой среде до уровня плотности 0,4-0,6 OD600, при этом клетка содержит плазмидный вектор pBSH-ED15 или pBSH-ED16 размером 6675 пар нуклеотидов (п.н.) и ген устойчивости к канамицину, вектор содержит репликативные ориджины pUC ori и М13 и ген, кодирующий lac-репрессор под контролем Т7-промотора, а конечный продукт очищен в зависимости от использованной встроенной плазмиды, имеет 99% чистоты и обладает апирогенными свойствами. В плазмидный вектор оперативно встроена кДНК гена человеческого эндостатина размером 552 нуклеотида между сайтами рестриктаз BamH I и Hind III, плазмидный вектор pBSH-ED16 содержит лидерную последовательность, кодирующую шесть гистидиновых остатков.
Другим объектом настоящего изобретения является способ получения препарата человеческого эндостатина, включающий экспрессию его гена в клетках Е.coli BL21 (DE3), трансформированных рекомбинантной плазмидной ДНК pBSH-ED15 или pBSH-ED16 с лидерной последовательностью, кодирующей шесть гистидиновых остатков, клетки культивируют в жидкой среде до мутности 0.4-0.6 OD600, после чего в среду добавляют 0,1-0,3 мМ изопропил-бета-D-тиогалактозида (ИПТГ) и продолжают культивирование до накопления целевого продукта в течение 2-3 часов, далее клетки подвергают воздействию ультразвука в течение 50-70 секунд при 0°С в присутствии 0,1%-ного деоксихолата натрия, растворение телец проводят в присутствии 1%-ного натрия N-лаурилсаркозина, суспензию многократно промывют и центрифугируют при 5000 g, при рефолдинге окисление кислородом воздуха осуществляют не более 35 часов, а очистку целевого продукта проводят в зависимости от используемой встроенной в ДНК клетки плазмиды. Очистку конечного продукта, полученного при культивировании клеток, содержащих встроенную плазмиду pBSH-ED15, осуществляют методом гель-фильтрационной хроматографии с участием сорбента Superose 12. Очистку конечного препарата, полученного при культивировании клеток, содержащих встроенную плазмиду рВ SHED 16, осуществляют в одну стадию методом катионообменной хроматографии на агарозном носителе.
Нуклеотидная последовательность, кодирующая полученный препарат, представлена на фиг.1. Общая схема генетической конструкции представлена на фиг.2.
Предлагается также способ получения человеческого эндостатина путем экспрессии его гена в составе рекомбинантной ДНК в Е.coil, отличием которого является то, что экспрессию осуществляют в рекомбинантной плазмиде pBSH-ED15 или рВ SH-ED 16 в ростовой среде до ее плотности 0,4-0,6 OD600, после чего в нее добавляют 0,1-0,3 мМ ИПТТ - индуктора lac-промотора - и экспрессию продолжают под контролем индуцированного lac-промотора рекомбинантной плазмиды.
Непосредственно связанным с этим изобретением является описание методов и способов, которыми эндостатин, содержащийся как главный компонент во внутриклеточных тельцах экспрессионных систем - клеток бактерий, может быть переведен в растворимое состояние, после чего очищен и с помощью рефолдинга возвращен к нативной структуре, благодаря которой реализуется его биологическая активность.
Стадии растворения внутриклеточных телец, содержащих эндостатин человека, предпочтительно выполняются при значениях рН, близких к нормальному, в пределах примерно от 7.0 до 8.0. Наиболее предпочтительно значение рН, равное 7.8.
Стадии растворения и восстановления нативной пространственной структуры эндостатина человека выполняются в присутствии восстанавливающих агентов, способных восстанавливать дисульфидные связи, образованные SH-группами полипептидных цепей. Например, восстанавливающий агент выбирается из группы агентов, состоящей из 2-меркаптоэтанола, DL-дитиотреитола и восстановленного глутатиона. Наиболее предпочтительным восстанавливающим агентом является DL-дитиотреитол.
Предпочтительно во время стадии растворения DL-дитиотреитол присутствует в концентрации от 0.5 мМ до 2 мМ. Наиболее предпочтительной концентрацией DL-дитиотреитола является концентрация около 1 мМ.
Предпочтительно во время стадии рефолдинга DL-дитиотреитол присутствует в концентрации приблизительно от 0.05 мМ до 0.2 мМ. Наиболее предпочтительной концентрацией DL-дитиотреитола является концентрация 0.1 мМ. На стадии рефолдинга эндостатина человека предпочтительно присутствует агент, способный усиливать обмен дисульфидными связями. Предпочтительно в качестве такого агента выбирается окисленный глутатион. Предпочтительно на стадии рефолдинга окисленный глутатион присутствует в концентрации от 2 мкМ до 20 мкМ. Наиболее предпочтительной концентрацией окисленного глутатиона на стадии рефолдинга является 10 мкМ. На стадии рефолдинга дисульфидные связи между SH-группами полипептидных цепей эндостатина человека образуются при окислении кислородом воздуха.
Наиболее предпочтительной продолжительностью по времени стадии окисления кислородом воздуха является около 35 часов.
Эндостатин человека после рефолдинга далее подвергается очистке с помощью любого из нижеперечисленных процессов: гель-фильтрационная хроматография, катионнообменная хроматография, высокоэффективная жидкостная хроматография с обращенной фазой либо аффинная хроматография. В нашем случае предпочтительно после стадии ренатурации очистку целевого продукта, полученного в плазмидной ДНК pBSH-ED15, осуществляют методом гель-фильтрационной хроматографии на сорбенте Superose 12, а очистку целевого продукта, полученного в плазмидной ДНК pBSH-ED16, осуществляют в одну стадию методом катионообменной хроматографии на агарозном носителе.
Использование в конструкции pBSH-ED16 лидерной последовательности, кодирующей шесть гистидиновых остатков, позволяет обеспечить надежную экспрессию слитого с ней эндостатина, отсутствие необходимых в таких случаях точечных замен для адаптации к аппарату биосинтеза белка клетки-хозяина, упрощение конечного выделения и очистки слитого белка. Присутствие гистидиновой последовательности не создает препятствий для корректной ренатурации зрелого белка.
Изобретение иллюстрируется следующими примерами:
Пример 1. Клонирование гена эндостатина человека
В качестве источника гена эндостатина была использована библиотека генов человека. Фрагмент гена коллагена, соответствующий последовательности, кодирующей эндостатин, был выделен методом амплификации с помощью следующих праймеров:
EndoXho5'-ACATGGATCCATGCATCATCATCACCATCATGATGATGA
TAAACCACCCGCCCACAGCCACCGCGA
Endo-Bam 5'-ACATGGATCCCCACCCGCCCACAGCCACCGCGA
Endo-Hind 5'-TCCGAAGCTTCAGGCAGTCATGAAGCTGTTCTCA
Праймер Endo-Xho 5' предназначен для синтезирования гена эндостатина человека с лидерной последовательностью 6-His и эпитопом узнавания энтерокиназой.
Подчеркнутые последовательности вносят сайты рестрикции Bam HI и Hind III для клонирования в вектор для секвенирования последовательности гена эндостатина pBS+. Фрагмент был обработан рестриктазами Bam HI-+-Hind III и клонирован между Bam HI и Hind III сайтами вектора pBS+.
Фрагмент, синтезированный с помощью праймеров Endo-Xho I и Endo-Hind, содержит в себе лидерную последовательность, кодирующую 6-His, и эпитоп узнавания энтерокиназой. Его обработали рестриктазами Xho I и Hind III, и заклонировали в вектор pBS+, между сайтами Xho I и Hind III.
В каждом случае получено и проанализировано по 20 независимых клонов.
Была получена следующая нуклеотидная последовательность, соответствующая гену эндостатина (фиг.1). Последовательность вставили по сайтам рестрикции BamH I и Hind III в вектор pBSH. Полученный вектор назвали pBSH-ED15 (pBSH-ED16, в случае с 6-His) (фиг.2).
Индукция синтеза эндостатина оценивалась в 10 мл ИПТГ-индуцированных культур (выращивание до OD600 0-5, внесение ИПТГ до 0.2 мМ, затем культивировали 2-3 часа при 37°С на качалке). Продукция белка ожидаемого размера проверена по методу SDS-гель-электрофореза в полиакриламидном геле (SDS-PAGE).
Для экспрессии рекомбинантных белков был использованы клетки Е.coli BL21(DE3).
Пример 2. Синтез эндостатина человека в Е.coli BL21(DE3)
Уровень синтеза эндостатина человека определяли в штамме Е.coli BL21(DE3), содержащем плазмиды pBSH-ED15 и pBSH-ED16. В качестве контроля использовали штамм Е.coli с рекомбинантной плазмидой pBSH, не несущей гена эндостатина. Клетки Е.coli штамма BL21(DE3) трансформировали плазмидой pBSH-ED15 либо плазмидой pBSH-ED16 и выращивали в течение 12 часов при 37°С в среде LB с добавлением канамицина (100 мкг/мл) и глюкозы (0.2%). Затем разбавляли ночную культуру в 25 раз свежей средой LB с канамицином и подращивали 2-3 часа до плотности OD600, затем вносили ИПТГ до конечной концентрации 0.2 мМ для индукции синтеза эндостатина. Растили еще 3 часа, центрифугировали культуру при 5000 g в течение 10 минут, супернатант отбрасывали.
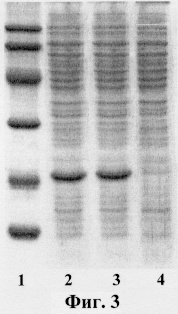
Как следует из данных, представленных на фиг.3, клетки Е.coli, содержащие плазмиды pBSH-ED15 и pBSH-ED16, синтезируют рекомбинантный эндостатин. На фиг.3 представлен электрофорез белков штаммов Е.coli в SDS-PAGE, где
1 - маркеры молекулярной массы;
2 - штамм Е.coli, содержащий рекомбинантную плазмиду pBSH-ED15;
3 - штамм Е.coli, содержащий рекомбинантную плазмиду pBSH-ED16;
4 - штамм Е.coli BL21(DE3), не несущий плазмиды pBSH-ED15 или pBSH-ED16.
Пример 3. Выделение препарата эндостатина человека из штамма-продуцента Е.coli, содержащего рекомбинантную плазмиду pBSH-ED15
Осадок клеток, полученный после культивирования Е.coli, ресуспендировали в буфере для обработки ультразвуком (100 мМ трис-HCl, рН 8.0, 5 мМ ЭДТА), после чего добавляли лизоцим до конечной концентрации 50 мкг/мл и инкубировали в этом буфере при комнатной температуре в течение 15 минут. Затем обработанные указанным способом клетки подвергали воздействию ультразвука с использованием зонда прибора для генерации ультразвука в течение около 50-70 секунд при 0°С в присутствии натрия деоксихолата в концентрации 0,1%. Полученную суспензию затем центрифугировали при 5000 g в течение 20 минут. Супернатант отбрасывали и осадок ресуспендировали в 15 мл буфера для обработки ультразвуком, с добавлением 0.1% натрия деоксихолата, после чего снова центрифугировали при указанных условиях; супернатант отбрасывали.
Дальнейшие этапы выполняли при +4°С. Внутриклеточные тельца, полученные из культуры клеток Е.coli штамма BL21(DE3), трансформированного плазмидой pBSH-ED15 или pBSH-ED16, растворяли в буфере А (50 мМ трис, рН 8.0, 1% натрий N-лаурилсаркозин, 1 мМ дитиотреитол) при концентрации эндостатина 1 мг/мл. Этот раствор перемешивали около 10 минут, после чего центрифугировали при 5000 g в течении 10 минут. Чистый супернатант переносили в диализные мешки с проницаемостью пор молекулярным весом менее 3 кДа и диализировали против большого объема буфера Б (50 мМ трис-HCl, рН 8.0, 0.1 мМ дитиотреитол) в течение 5 часов, после чего меняли буфер диализа на свежий и диализировали в течение 6 часов. Затем белковые продукты диализировали 4 часа против большого объема буфера В (50 мМ трис-HCl, рН 8.0), после чего диализировали против большого объема буфера Г (50 мМ трис-HCl, рН 8.0, 0.01 мМ окисленного глутатиона и 1 мМ восстановленного глутатиона) (дважды по 4 часа). На последнем этапе дважды диализировали против большого объема буфера В в течение 12 часов, чтобы полностью удалить редокс-пару глутатиона. Конечный продукт отбирали из диализных мешков, центрифугировали при 5000 g в течение 10 минут и отделяли супернатант, представляющий собой раствор препарата эндостатина человека.
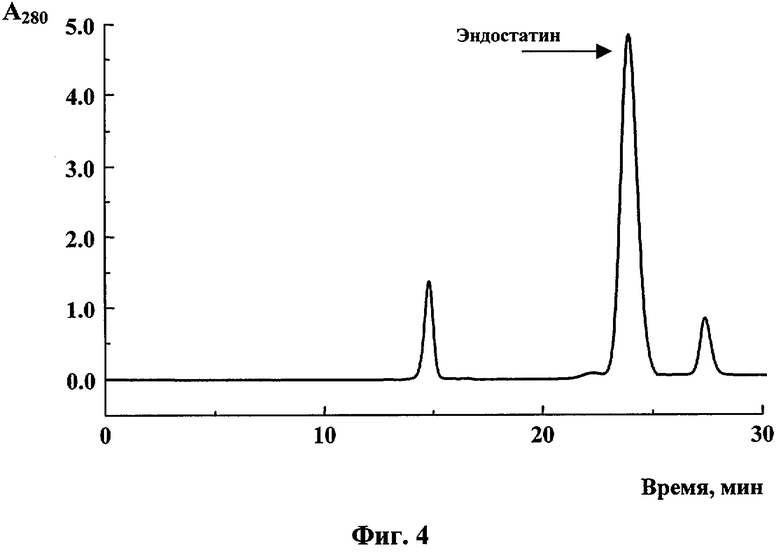
Очистку целевого продукта, полученного в плазмидной ДНК pBSH-ED15, осуществляли методом гель-фильтрационной хроматографии с помощью системы FPLC на хроматографическом сорбенте Superose 12. Профиль элюции эндостатина представлен на фиг.4. Уровень продукции эндостатина человека в штамме Е.coli, трансформированном плазмидой pBSH-ED15, составлял около 70 мг/л культуры.
Пример 4. Выделение препарата эндостатина человека из штамма-продуцента Е.coli, содержащего рекомбинантную плазмиду pBSH-ED16
Стадии дезинтеграции клеток и рефолдинга эндостатина проводили, как описано в примере 3. Очистку целевого продукта, полученного в плазмидной ДНК pBSH-ED16, осуществляли методом катионообменной хроматографии на агарозном носителе. Колонку, заполненную сорбентом IDA-агарозой (объем около 1 мл), уравновешивали раствором никеля (II) сульфата, затем буфером Д (10 мМ трис, рН 8.0, мочевина 6 М, натрия хлорид 400 мМ, 2-меркаптоэтанол 10 мМ, имидазол 5 мМ). Супернатант, полученный после рефолдинга, как описано в этом разделе ранее, наносили на колонку, промывали последнюю 5 объемами буфера Д, затем 10 объемами буфера Е (буфер Д, где концентрация имидазола 20 мМ). Элюцию проводили буфером Ж (10 мМ трис, рН 8.0, мочевина 2 М, натрия хлорид 400 мМ, 2-меркаптоэтанол 10 мМ, имидазол 300 мМ). Фракции, содержащие эндостатин человека, объединяли и диализировали в течение 14 часов против 100-кратного объема буфера 3 (10 мМ трис, рН 8.0, 150 мМ NaCl).
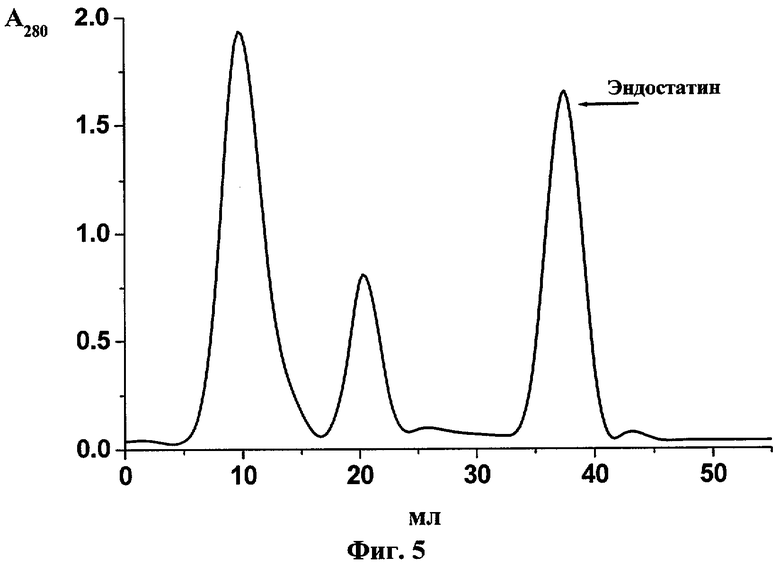
Профиль элюции эндостатина с катионообменной колонки с агарозным носителем представлен на фиг.5. Уровень продукции эндостатина человека в штамме Е.coli, трансформированном плазмидой pBSH-ED16, составлял около 80 мг/л культуры.
Пример 5. Определение концентрации остаточного детергента в препаратах эндостатина
Определение концентрации остаточного детергента - натрия N-лаурилсаркозина - в препаратах эндостатина человека после серии диализов на стадии рефолдинга проводили турбидиметрическим методом. Для этого раствор белка в различных соотношениях разбавляли деионизованной водой, после чего добавляли к испытуемым образцам 10 об.% 1 М хлороводородной кислоты и инкубировали 5 минут при комнатной температуре. Затем измеряли поглощение световых лучей образцов при λ450 нм. Расчет количества натрия N-лаурилсаркозина в растворе белка осуществляли с использованием калибровочной кривой, полученной путем измерения А450нм серии растворов детергента с известными концентрациями в диапазоне от 0,005% до 0,025%. В указанном диапазоне поглощение при λ450 нм линейно зависит от концентрации натрия N-лаурилсаркозина. Измерение остаточной концентрации натрия N-лаурилсаркозина показывало, что в конечном продукте его концентрация была менее 0,0035% (0,035 мг/мл), что говорило о почти полном (более 99,7%) удалении детергента. При этом эндостатин человека сохранял растворимость в водных растворах.
Пример 6. Определение степени ренатурации целевого препарата эндостатина
Количество свободных (неокислившихся) SH-групп в препаратах очищенного эндостатина определяли, используя реактив Эллмана (5,5'-дитиобис(2-нитробензойная кислота)), как было описано ранее [11]. С использованием указанной методики было установлено, что степень ренатурации полученного препарата эндостатина составляла более 95%.
Пример 7. Электрофоретический анализ конечного препарата эндостатина
На фиг.6 представлены данные электрофоретического анализа конечного препарата эндостатина в 15%-ном полиакриламидном геле в присутствии 0,1% натрия додецилсульфата, где:
1 - маркеры молекулярных масс;
2 - препарат эндостатина из штамма с плазмидой pBSH-ED15 после заключительной стадии очистки;
3 - препарат эндостатина из штамма с плазмидой pBSH-ED16 после заключительной стадии очистки.
Из фиг.6 видно, что оба препарата представляют собой практически гомогенные белки (чистота более 99%) с соответствующей молекулярной массой (20 кДа).
Пример 8. Определение апирогенности препарата
Учитывая использование эндостатина человека, полученного с использованием описываемой методики, преимущественно в качестве терапевтического агента, вводимого парентерально в организм животных, эндостатин человека иссследовали на качественное содержание бактериальных эндотоксинов (липополисахаридов) (ЛАЛ-тест). Анализ на ЛАЛ-тест проводили в апирогенных пробирках из натриевого стекла размером 10×75 мм. Готовили реакционную смесь, представляющую собой 0.1 мл раствора ЛАЛ-реактива (Limulus Amebocyte Lysate, Pyrotell, США) и 0.1 мл раствора эндостатина человека. После внесения реакционной смеси в пробирки и перемешивания пробирки инкубировали при 37°С в течение 60 минут. Затем вынимали пробирки из инкубатора и переворачивали на 180°. Разрушение геля, образовавшегося на дне пробирки, при ее переворачивании, или отсутствие такового позволяло сделать заключение об апирогенности анализируемого раствора эндостатина.
Пример 9. Оценка биологической активности полученного препарата эндостатина.
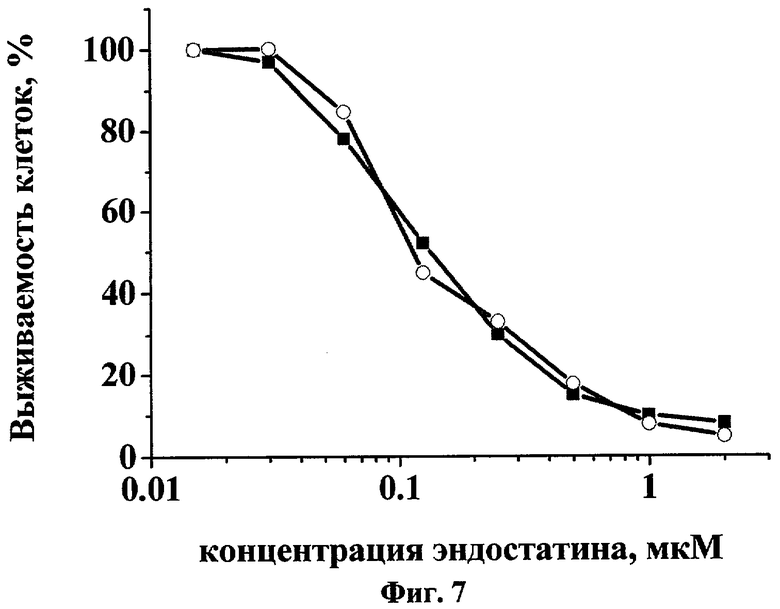
Биологическую активность полученного эндостатина оценивали по его способности ингибировать пролиферацию эндотелиальных клеток in vitro. Эндотелиальные клетки пупочной вены зародыша человека (HUVEC) из монослоя трипсинизировали и ресуспендировали в среде M199 с добавкой 5%-ного раствора фетальной бычьей сыворотки. Затем клетки рассеивали в 96-луночные планшеты, покрытые желатином, в плотности 5000 клеток в лунку. Через 24 часа добавляли полученный препарат эндостатина в различных концентрациях. По прошествии 20 минут культуры клеток обрабатывали раствором основного фактора роста фибробластов в концентрации 5 нг/мл в присутствии гепарина (1 мкг/мл). Выживаемость клеток в контрольной и в обработанной полученным препаратом группах определяли с помощью МТТ-теста через 72 часа инкубации [12].
Данные о влиянии препаратов эндостатина на пролиферацию клеток HUVEC приведены на фиг.7. Полученные препараты эндостатина из клеток с плазмидами pBSH-ED15 и pBSH-ED16 оказывали выраженное дозозависимое ингибирующее действие в отношении пролиферации клеток исследованной линии.
Таким образом, нами получен очищенный рекомбинантный препарат эндостатина человека по оригинальной методике, являющийся апирогенным и имеющем чистоту более 99%, что позволяет использовать его для предклинических испытаний в качестве антиангиогенного вещества при различных патологиях.
ЛИТЕРАТУРА
1. Bouck N. et al., Adv. Cancer Res. 69: 135-174, 1996.
2. Folkman J., Nature Med. 1: 27-31,1995.
3. Hanahan D. and Weinberg R.A., Cell 100: 57-70, 2000.
4. Kerbel R. and Folkman J., Nat. Rev. Cancer 2: 727, 2002.
5. Muragaki Y. et al., Proc. Natl. Acad. Sci. USA 92: 8763-8767, 1995.
6. O'Reilly M.S. et al., Cell 88: 277-285, 1997.
7. Sim B.K. et al., Cancer Metastasis Rev. 19: 181-190, 2000.
8. Jiang L. et al., Am. J. Physiol. Cell Physiol. 280: 1140-1150, 2001.
9. Kirn Y.M. et al., Cancer Res. 60: 5410-5413, 2000.
10. Xu R. et al., Protein Expression and Purification 24: 453-459, 2000.
11. Riddles P.W. et al., Meth. Enzymol. 91: 49-60, 1983.
12. Mosmann T.J. Immunol. Meth., 65: 55-63, 1983.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА БЕЛКА ИЗ СЕМЕЙСТВА СТРЕССОВЫХ И ПРЕПАРАТ БЕЛКА HSP70, ПОЛУЧЕННЫЙ ЭТИМ СПОСОБОМ | 2005 |
|
RU2283128C1 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ГИБРИДНОГО БЕЛКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2006 |
|
RU2316342C1 |
| РЕКОМБИНАНТНЫЙ ГИБРИДНЫЙ ПОЛИПЕПТИД, СПОСОБНЫЙ ИНГИБИРОВАТЬ ПРОЛИФЕРАЦИЮ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК ЧЕЛОВЕКА IN VITRO, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2011 |
|
RU2465283C1 |
| ШТАММ E.coli BL21(DE3)/pET-hHSP70 - ПРОДУЦЕНТ БЕЛКА ТЕПЛОВОГО ШОКА ЧЕЛОВЕКА HSP70 | 2006 |
|
RU2333956C1 |
| ПЛАЗМИДНАЯ ДНК pBSH2EGF, КОДИРУЮЩАЯ СИНТЕЗ ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА ЧЕЛОВЕКА, И СПОСОБ ЕГО ПОЛУЧЕНИЯ С ПОМОЩЬЮ ДАННОЙ ДНК | 2004 |
|
RU2289623C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИАНГИОГЕННОГО ПОЛИПЕПТИДА | 2012 |
|
RU2488635C1 |
| ПРОТИВООПУХОЛЕВЫЙ ПЕПТИДНЫЙ ПРЕПАРАТ НА ОСНОВЕ ФРАГМЕНТА АЛЬФА-ФЕТОПРОТЕИНА, ЕГО КОНЪЮГАТ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБ ЛЕЧЕНИЯ ГОРМОНЗАВИСИМЫХ ОПУХОЛЕЙ | 2005 |
|
RU2285537C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ БИОСИНТЕЗ ЧЕЛОВЕЧЕСКОГО АЛЬФА 2-ИНТЕРФЕРОНА, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2002 |
|
RU2230117C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ФРАГМЕНТА ПЛАЗМИНОГЕНА ЧЕЛОВЕКА, ОБЛАДАЮЩЕГО АНТИАНГИОГЕННЫМ ДЕЙСТВИЕМ | 2011 |
|
RU2458987C1 |
| ШТАММ Escherichia coli BL21(DE3)/pAFP11D3 - ПРОДУЦЕНТ ФРАГМЕНТА С 404 ПО 609 АМИНОКИСЛОТУ АЛЬФА-ФЕТОПРОТЕИНА ЧЕЛОВЕКА | 2010 |
|
RU2422512C1 |


Изобретение относится к области молекулярной фармакологии, в частности к препарату человеческого эндостатина и способу его получения. Сущность изобретения состоит в том, что препарат выделен после инкубации клеток E.coli BL21 (DE3) в ростовой среде до уровня плотности 0,4-0,6 OD600, клетка содержит плазмидный вектор pBSH-ED15 или pBSH-ED16 размером 6675 п.н. и ген устойчивости к канамицину, вектор содержит репликативные участки pUC ori и M13 и ген, кодирующий lac-репрессор под контролем Т7-промотора, а конечный продукт - 99% чистоты и обладает апирогенными свойствами. Еще одним объектом изобретения является способ получения препарата. Сущность способа состоит в следующем: проводят экспрессию гена эндостатина с помощью плазмидной ДНК в составе клеток E.coli, рефолдингом и очисткой продукта, при этом экспрессию гена осуществляют в клетках E.coli BL21 (DE3), трансформированных рекомбинантной плазмидной ДНК pBSH-ED15 или pBSH-ED16, клетки культивируют в ростовой среде до уровня плотности 0-4-0.6 OD600, после чего в среду добавляют 0,1-0,3 мМ ИПТГ и культивируют до накопления продукта в течение 2-3 часов, далее клетки подвергают воздействию ультразвука в течение 50-70 секунд при 0°С в присутствии 0,1%-ного деоксихолата натрия, растворение телец проводят в присутствии 1%-ного натрия N-лаурилсаркозина, суспензию многократно промывют и центрифугируют при 5000 g, при рефолдинге окисление кислородом воздуха осуществляют в течение 35 часов, а очистку целевого продукта проводят хроматографически в зависимости от используемой встроенной плазмиды. Технический результат - получение апирогенного продукта высокой чистоты. 2 н. и 4 з.п. ф-лы, 7 ил.
| СПОСОБ ПРОИЗВОДСТВА ЭНДОСТАТИНА МЫШИ И ЧЕЛОВЕКА | 1999 |
|
RU2225728C2 |
| US 5854205 А, 29.12.1998 | |||
| US 6653098 A, 25.11.2003. | |||

