Изобретение относится к рецепторам, которые включают в себя фрагменты CD4, которые связывают ВИЧ, но не делают клетки хозяина восприимчивыми к ВИЧ-инфекции. Таким образом, настоящим изобретением создано новое и эффективное лечебное средство против ВИЧ.
Распознавание антигена T-клетками с помощью T-клеточного рецептора лежит в основе многочисленных иммунологических явлений. T-клетки управляют тем, что называют клеточно-опосредованным иммунитетом. Он включает в себя механизм уничтожения чужеродных тканей или инфицированных клеток с помощью клеток иммунной системы. Существуют разновидности T-клеток, включающие "хелперные" и "супрессорные" клетки, которые модулируют иммунный ответ, и цитотоксические (или "киллерные") клетки, которые непосредственно могут уничтожать аномальные клетки.
T-клетка, которая узнает и связывает соответствующий антиген, обнаруживаемый на поверхности другой клетки, становится активированной; она может затем размножиться и если она представляет собой цитотоксическую клетку, то она уничтожит присоединенную клетку.
ВИЧ и иммунопатогенез
В 1984 г. было установлено, что ВИЧ является этиологическим фактором СПИД. С тех пор определение СПИД неоднократно меняли много раз в соответствии с применяемым диагностическим критерием. Однако, несмотря на колебания диагностических параметров, очевидным общим действующим началом является ВИЧ-инфекция с последующим развитием персистентных конституциональных симптомов и заболеваний, определяющих СПИД, таких как вторичные инфекции, новообразования и неврологические заболевания.
Основы терапии Харрисона, 12-е изд. McGraw Hill (1991).
ВИЧ представляет собой человеческий ретровирус из группы лентивирусов. Четыре распознаваемые человеческие ретровирусы относятся к двум различным группам: T-лимфотропные (или лейкозные) ретровирусы человека. HTLV-1 и HTLV-2, и вирусы иммунодефицита человека, HIV-1 и HIV-2. Первая группа представлена трансформирующими вирусами, тогда как вторая группа представлена цитопатическими вирусами.
ВИЧ-4 идентифицировали в качестве наиболее распространенной причины во всем мире, вызывающей заболевание СПИД. Гомология последовательности между ВИЧ-2 и ВИЧ-1 составляет около 40% ВИЧ-2, который более близок к группе вирусов иммунодефицита человекообразных обезьян (SIV). Curran et al., Nature 324:572-575 (1986).
ВИЧ-2 представлен обычными ретровирусными генами (env, gag и pol), а также шестью другими генами, участвующими в репликации и других биологических функций вируса. Как указано выше, общим знаменателем СПИД является глубокая иммуносупрессия, приводящая к появлению разнообразных заболеваний, вызванных условно-патогенными микроорганизмами, в частности инфекционных и опухолевых.
Основную причину нарушения иммунитета при СПИД рассматривают как количественную и качественную недостаточность популяции T4-клеток, лимфоцитов производимых тимусом (Т). Эту группу клеток фенотипически выявляют по наличию поверхностных молекул CD4, которые являются клеточными рецепторами для ВИЧ. Dalgleich et al., Nature 312:763 (1984). Хотя T4-клетка представляет собой главный тип клетки, инфицируемой ВИЧ, фактически любая человеческая клетка, которая экспрессирует молекулу CD4 на своей поверхности, способна связываться и подвергаться инфицированию ВИЧ.
Традиционно молекулам CD4 T-клеток приписывают роль хелперов/индукторов, указывающей их функцию в обеспечении активированного сигнала для T-клеток или индуцирующей образование T-лимфоцитов, несущих аналогичный CD8-маркер для становления цитотоксических/супрессорных клеток. Reinherz and Schlossman, Cell 19:821-827 (1980); Goldstein et al., Immunol, rev. 68:5-42 (1982).
ВИЧ специфически связывается и с высоким сродством, в результате аминокислотного удлинения вирусной оболочки (gp120), с участком области VI молекулы CD4, локализованной вблизи ее N-конца. После связывания вирус сливается с мембраной клетки-мишени и интернализуется. Сразу после интернализации он использует фермент обратную транскриптазу для транскрибирования своей геномной РНК с ДНК, которая интегрирована в клеточную ДНК и где она существует в течение жизни клетки в качестве "провируса".
Провирус может оставаться латентным или активироваться, транскрибируя мРНК или геномную РНК, приводя к синтезу белка, сборке, образованию новых вирусных частиц и отделению вируса от клеточной поверхности. Хотя точный механизм, по которому вирус вызывает гибель клетки, не установлен, очевидно, что главный механизм представляет собой массовое отделение вирусных частиц от клеточной поверхности, приводящее к разрушению цитоплазматической мембраны и нарушению осморегуляции.
Во время инфекции организм хозяина вырабатывает антитела против вирусных белков, в том числе основных оболочечных гликопротеинов gp120 и gp41. Несмотря на этот гуморальный иммунитет, заболевание прогрессирует, приводя к летальной иммуносупрессии, характеризующейся множественными условно-патогенными инфекциями, паразитемии, деменции и смерти. Недостаток антивирусных антител хозяина, препятствующих развитию заболевания, является одним из наиболее мучительных и тревожных аспектов инфекции, и предвещает малую эффективность усилий вакцинирования традиционными способами.
Два фактора могут влиять на эффективность гуморального ответа при вирусном иммунодефиците. Во-первых, подобно другим РНК-вирусам (и в частности, подобно ретровирусам) иммунодефицитные вирусы проявляют высокую мутационную скорость в ответ на иммунный надзор хозяина. Во-вторых, оболочечные гликопротеины сами по себе являются сильно гликозилированными молекулами, представляющие несколько эпитопов, обладающие высоким сродством для связывания антител. Недостаточная антигенная мишень, которую представляет вирусная оболочка, оставляет хозяину мало возможности для ограничения вирусной инфекции путем образования специфических антител.
Клетки, инфицированные ВИЧ, экспрессируют на своей поверхности гликопротеин gp120, gp120 опосредует события слияния между CD4-клетками путем реакции, подобной той, с помощью которой вирус внедряется в непораженную клетку, приводя к образованию короткоживущих многоядерных гигантских клеток. Образование синцития зависит от непосредственного взаимодействия оболочечного гликопротеина gp120 с белком CD4. Dalgleish et al., см. выше; Klatzman et al., Nature 312:763 (1984); McDougal et al., Science 231:382 (1986); Sodroski et al., Nature 322:470 (1986); Lifson et al., Nature 323:725 (1986); Sodroski et al., Nature 321:412 (1986).
Доказательством того, что связывание CD4-gp120 несет ответственность за вирусную инфекцию клеток, несущих CD4-антиген, включает в себя обнаружение специфического комплекса, который образуется между gp120 и CD4 (McDougal et al. , см. выше). Другие исследователи показали, что клеточные линии, не инфицированные ВИЧ, превращались в инфицированные клеточные линии после трансфекции и экспрессии у человека гена CD4 кДНК. Maddon et al., Cell 46:333-348 (1986).
Терапевтические программы, основанные на растворимом CD4, в качестве пассивного агента для подавления вирусной адсорбции и межклеточной передачи, опосредованной синцитием, были предложены и успешно продемонстрированы ин витро целым рядом групп (Deen et al., Nature 331:82-84 (1988); Fisher et al. , Nature 331:76-78 (1988); Hussey et al., Nature 331:78-81 (1988); Smith et al. , Science 238: 1704-1707 (1987); Traunecker et al., Nature 331:84-86 (1988); a впоследствии создали слитые CD4-иммуноглобулиновые белки с удлиненным временем полужизни и умеренной биологической активностью (Capon et al., Nature 337:525-531 (1989); Traunecker et al., Nature 339:68-70 (1989); Byrn et al. , Nature 344:667-670 (1990); Zettlmeissl et al., DNA Cell Biol. 9: 347-353 (1990). Хотя конъюгаты CD4-иммунотоксин или слитые белки проявляют сильную цитотоксичность к инфицированным клеткам ин витро (Chaudhary et al., Nature 335: 369-372 (1988);Till et al., Science 242: 1166-1168 (1988), латентное состояние синдрома иммунодефицита делает маловероятным, чтобы какая-либо однократная терапия будет эффективной в ликвидации вирусного поражения, а антигенность чужеродных слитых белков, по-видимому, ограничит их переносимость при лечениях повторяющимися дозами. Испытания, проведенные на обезьянах, пораженных SIV, показали, что растворимый CD4 при введении животным без отчетливой CD4-цитопении может снизить титр SIV и улучшить ин витро уровень миелоидного потенциала (Watanabe et al., Nature 337:267-270 (1989). Однако индуцированное повторное вирусное проявление наблюдали после прерванного лечения, свидетельствующее, что пожизненный прием CD4, по-видимому, необходим для предупреждения прогрессивного истощения иммунной системы.
T-клетки и Fc-рецепторы
Клеточная поверхность экспрессирует самую обильную форму антигенного рецептора T-клеток (TCR), требующего коэкспрессии не менее 6 различных полипептидных цепей (Weiss et al. , J. Exp. Med. 160:1284-1299 (1984); Orloffhashi et al., Nature 516:606-609 (1985); Berkhout et al., J. Biol. Chem. 263: 8528-8556 (1988); Sussman et al., Cell 52:85-95 (1988), α/β-антиген, связывающий цепи, три полипептида CD3-комплекса, и s. Если какая-либо цепь отсутствует, устойчивая экспрессия остающихся членов комплекса не происходит. s представляет собой ограничивающий полипептид для поверхностной экспрессии полного комплекса (Sussman et al., Cell 52:85-95 (1988)) и предположительно опосредует фракцию, запускающую программы клеточной активации, с помощью рецептора узнающего лиганд (Weissman et al., EMBO J. 8:3651-5656 (1989); Frank et al., Science 249:174-177 (1990)). Гомодимер типа 1 с мол. массой 32 кДа, s (зета), закрепленный в мембране, имеет 9 внеклеточных остатков домена, не имеющего участков для N-присоединенного добавочного гликана, и 112 остатков (мышь) или 113 остатков (человек) внутриклеточного домена (Weissmaim et al., Science 238:1018-1020 (1988); Weissman et al., Proc. Natl. , Acad. Sci. UCA 85:9709-9713 (1988)). Изоформа s, названная η (эта) (Baniyash et al., J.Biol. Chem. 263:9874-9878 (1988); Orloff et al., J.Biol. Chem. 264:14812-14817 (1989), который возникла из альтернативного пути сплайсинга мРНК (Jin et al., Proc. Natl. Acad. Sci. UCA 87:3319-3255 (1990)), представлена в клетках, экспрессирующих антигенный рецептор, в сниженном количестве. Считают, что гетеродимеры s-η опосредуют образование инозитфосфатов, также как и инициированную рецептором запрограммированную гибель клетки, названную апоптозом (Mercep et al., Science 242:571-574 (1988); Mercep et al., Science 246:1162-1165 (1989).
Подобно s и η,γ (гамма)-цепь, ассоциированная с Fc-рецептором, экспрессирует на клеточной поверхности комплексы с дополнительными полипептидами, некоторые из них опосредует узнавание лиганда, а другие обладают неопределенной функцией, γ-цепь поддерживает гомодимерную структуру и общую организацию, очень сходную с той, которая образована s-цепью и является компонентом высокоаффинного рецептора IgE тучных/базофильных клеток, Fc RI, который состоит, как миниум, из трех разных полипептидных цепей (Blank et al. , Nature 337: 187-189 (1989); Ra et al., Nature 241:752-754 (1989), и одного низкоаффинного рецептора IgE, представленного у мышей FcyRII α (Ra et al. , J. Biol. Chem. 264:15323-15527 (1989), а у человека - CD17-субтипом, экспрессируемого макрофагами и природными клетками-киллерами, CD16ТМ (трансмембранные молекулы CD16) (Lanier et al., Nature 342:803-805 (1989); Anderson et al., Proc. Natl. Acad. Sci. UCA 87:2274-2278 (1990) и полипептидом с неизвестной функцией (Anderson et al., Proc. Natl. Acad. Sci. UCA 87: 2274-2278 (1990)). В недавней публикации сообщали, что γ-цепь экспрессируется мышиной линией T-клеток, CTL, в которой она образует гомодимеры, также как и гетеродимеры γ-s и γ-η (Orloff et al., Nature 347:189-191 (1990).
Fc-рецепторы опосредуют фагоцитоз иммунными комплексами, трансцитозом и антителозависимой клеточной цитотоксичностью (ADCC) (Ravetch and Kinet et al. , Annu. Rev. lmmunol. 9:547-492 (1991); Unkeless et al., Annu. Rev. Immunol. 6:251-281 (1988); и Mellman, Curr. Opin. Immunol. 1:16-25 (1988)). В последнее время показали, что одна из мышиных низкоаффинных изоформ Fc-рецептора, FcRylllBI, опосредует интернализацию мишеней, покрытых Ig, в ямках покрытых клатрином, а другой низкоаффинный рецептор, FcRylllA, опосредует ADCC через ассоциацию с одним или большим числом членов небольшого семейства "пусковых молекул" (Miettinen et al., Cell. 58:317-327 (1989); и Hunziber и Melllman, J. Cell Biol. 109:3291-3302 (1989)). Эти пусковые молекулы, s-цепь T-клеточного рецептора (TCR), η-цепь TCR, и γ- цепь Fc-рецептора взаимодействуют с лигандом, узнающим домены рецепторов отличающихся иммунных систем и может автономно инициировать программы клеточного эффектора, в том числе цитолиз с последующей агрегацией (Samelson et al., Cell 43:223-231 (1985); Weissman et al., Science 239:1018- 1020 (1988); Jin et al., Proc. Natl. Acad. UCA 87: 3319-3323 (1990); Blank et al., Nature 337:187-189 (1989); Lanier et al., Nature 342:803-805 (1989); Kurosaki и Pavetch, Nature 343: 805-807 (1989); Hibbs et al., Science 246:1608-1611 (1989); Anderson et al. , Proc. Natl. Acad. Sci. UCA 87:2274-2278 (1990); и Irving и Weiss, Cell 64:891-901 (1991).
Однако при проведении параллелей между низкоаффинными семействами Fc-рецептора мыши и человека становится очевидным, что изоформы FcRyllA и C человека не дублируют мышиные. Это отчасти потому, что их функцию еще не определили.
Поскольку гуморальные факторы, основанные только на CD4, могут иметь ограниченное применение in vivo, в предварительном исследовании зондировали возможность усиления клеточного иммунитета к ВИЧ. Идентифицировали препараты химерных белков (в которых внеклеточный доммен CD4 слили с трансмембранным и/или внутриклеточным доменами T-клеточного рецептора, Fc-рецептор lgG, или элементами, преобразующими сигнал рецептора B-клеток (U.S.S.N. 07/847566 и 07/665961, включенные в настоящее описание путем ссылки). Цитолитические T-клетки, экспрессирующие химеры, которые включают в себя внеклеточный домен CD4, проявляют сильную, независимую от МНС, способность уничтожать клеточные мишени, экспрессирующие оболочечные белки ВИЧ. Очень важным и новым компонентом этого способа явилась идентификация одиночного рецептора T-клетки, Fc-рецептора и цепей рецептора B-клетки, которые достаточно агрегированы, чтобы инициировать клеточный ответ. Одно из самых ценных достижений этого способа состоит в том, что создали химеры между CD4 и s, η, или γ, которые непосредственно разрушают T-лимфоциты при их узнавании и уничтожают клетки, экспрессирующие гликопротеин gp120 ВИЧ (U. S.S.N. 07/847566 и 07/665961, включенные в настоящее описание путем ссылки).
Краткое изложение существа изобретения
В соответствии с основной целью в настоящем изобретении отведено важнейшее место способу управления клеточным иммунным ответом, направленным против ВИЧ-инфицированной клетки млекопитающего. Данный способ включает в себя введение млекопитающему эффективного количества терапевтических клеток, а именно терапевтических клеток, экспрессирующих связанный с мембраной белковый химерный рецептор, включающий в себя (а) внеклеточную часть, которая включает в себя фрагмент CD4, который обладает способностью специфически узнавать и связывать ВИЧ-инфицированную клетку, но который не опосредует ВИЧ-инфицирование и (б) внутриклеточную часть, которая обладает способностью сигнализировать терапевтической клетке о необходимости уничтожить связанную с рецептором ВИЧ-инфицированную клетку.
В соответствии с целью, родственной этой, настоящее изобретение относится к клетке, которая экспрессирует связанный с мембраной белковый химерный рецептор, который включает в себя (а) внеклеточную часть, которая включает в себя фрагмент CD4, который обладает способностью специфически узнавать и связывать ВИЧ-инфицированную клетку, но который не опосредует ВИЧ-инфицирование и (б) внутриклеточную часть, которая обладает способностью сигнализировать терапевтической клетке о необходимости уничтожить связанную с рецептором ВИЧ-инфицированную клетку.
В соответствии со второй целью настоящее изобретение относится к способу воздействия на ВИЧ у млекопитающего, включающего в себя введение млекопитающему эффективного количества терапевтических клеток, а именно терапевтических клеток, экспрессирующих связанный с мембраной белковый химерный рецептор, включающий в себя внеклеточную часть, которая включает в себя фрагмент CD4, который обладает способностью специфически узнавать и связывать ВИЧ-инфицированную клетку, но который не опосредует ВИЧ-инфицирование.
В соответствии с целью, родственной этой, настоящее изобретение относится к клетке, которая экспрессирует связанный с мембраной белковый химерный рецептор, включающий в себя внеклеточную часть, которая включает в себя фрагмент CD4, который обладает способностью специфически узнавать и связывать ВИЧ-инфицированную клетку, но который не опосредует ВИЧ-инфицирование.
В предпочтительных вариантах осуществления основной и второй целей CD4-фрагмент представляет собой фрагмент последовательности аминокислот 1-394 молекулы CD4 или фрагмент последовательности аминокислот 1-200 молекулы CD4; данный CD4-фрагмент отделили от внутриклеточной части с помощью CD7-трансмембранного домена, представленного на фиг. 26 или с помощью шарнира, CH2- и CH3-доменов молекулы lgGI человека, представленной на фиг.25: а CD4-фрагмент отделили от терапевтической клетки расстоянием не менее 48 ангстрем (и предпочтительно - расстоянием не менее 72 ангстрем). В предпочтительных вариантах осуществления первой цели внутриклеточная часть представляет собой часть, преобразующую сигнал рецепторного белка T-клетки (к примеру, s), рецепторного белка B-клетки, или Fc-рецепторного белка; а терапевтические клетки выбраны из группы, состоящей из: (а) T-лимфоциты; (b) цитотоксические T-лимфоциты; (с) природные клетки-киллеры; (d) нейтрофилы; (е) гранулоциты; (f) макрофаги; (g) тучные клетки; и (i) эмбриональные стволовые клетки (ES).
В соответствии с другими целями, родственными этой, настоящее изобретение относится к ДНК, кодирующей химерный рецептор настоящего изобретения; и вектору, включающему в себя ДНК этого химерного рецептора.
Несмотря на то, что специфичным вариантом осуществления настоящего изобретения является химера между молекулами CD4 и zeta, любые рецепторные цепи, имеющие сходную с этими молекулами функцию, например, в гранулоцитах или B-лимфоцитах, могли бы использоваться для целей, раскрытых здесь. Отличительные признаки необходимой пусковой молекулы иммунной клетки включают в себя способность экспрессироваться автономно (т.е. в виде одиночной цепи), способность к слиянию с внеклеточной частью домена CD так, чтобы полученная химера присутствовала на поверхности терапевтической клетки, и способность к инициированию программ клеточного эффектора для вторичной агрегации при встрече с лигандом мишени.
В настоящее время большинство традиционных способов доставки химер к клеткам иммунной системы осуществляют посредством некоторых форм генетической терапии. Однако воспроизведение клеток иммунной системы с химерными рецепторами путем смешивания клеток с соответствующим растворенным очищенным химерным белком также будет приводить к образованию популяции биоинженерных клеток, способной отвечать на ВИЧ-инфицированные мишени. В терапевтических целях применили подобный способ, например, для интродуцирования CD4-молекулы в эритроциты. В этом случае популяция биоинженерных клеток не обладала способностью к самовоспроизведению.
Настоящее изобретение относится к функциональным и упрощенным химерам между CD-4 и фрагментами и T-клеточным рецептором, B-клеточным рецептором и субъединицами Fc-рецептора, которые обладают способностью управлять иммунными клетками для узнавания и лизиса ВИЧ-инфицированных клеток. Данный способ по управлению клеточным ответом у млекопитающих включает в себя введение эффективного количества терапевтических клеток (например, цитотоксических T-лимфоцитов) в организм млекопитающего, поскольку клетки способны разпознать и уничтожить ВИЧ-инфицированную клетку.
Настоящее изобретение включает в себя химерные рецепторные белки, которые направляют цитотоксические T-лимфоциты к ВИЧ-инфицированным клеткам для их узнавания и лизиса, клетки хозяина, трансформированные вектором, включающие в себя химерные рецепторы, и антитела к химерным рецепторам.
Эти и другие не ограничиваемые настоящим изобретением варианты осуществления настоящего изобретения будут понятны специалистам из нижеследующего подробного описания настоящего изобретения.
В нижеследующем детальном описании приведены ссылки на различные методики, известные специалистам в области молекулярной биологии и иммунологии. Публикации и другие изложенные материалы относительно таких известных методик, на которые ссылаются, включены в настоящее описание путем ссылки, приводимой в полном объеме.
Стандартные рабочие ссылки, излагающие технологию рекомбинантной ДНК, изложены в настоящем описании по Watson et al., Молекулярная биология гена, том I и II, the Benjamin/Cummings Publishing Company, Inc., publisher, Menlo Park, CA (1987); DarnelI et al., Молекулярная биология клетки. Scientific American Books, Inc., publisher, New York, N.Y. (1986); Lewin, Гены II, John Wiley & Sons, publishers. Неw York, N.Y. (1985); Old et al., Принципы работы с генами: Введение в генетическую инженерию, 2nd ed., University of California Press, publisher, Berkely, CA (1981); Maniatis et al., Молекулярное клонирование: Руководство для лабораторных работ, 2nd ed. Cold Spring Harbol Laboratory, publisher, Cold Spring Harbor, NY (1989); Ausubel et al., Современные методы в молекулярной биологии, Wiley Press, New York:, NY (1989).
ТЕРМИНОЛОГИЯ
Под "клонированием" подразумевают использование ин витро методов рекомбинации для внедрения отдельного гена или иной последовательности ДНК в молекулу вектора.
Под "кДНК" подразумевают комплементарную или копийную ДНК, полученную с матрицы РНК, в результате деятельности РНК-зависимой ДНК-полимеразы (обратная транскриптаза). Следовательно, "клон кДНК" означает последовательность двуцепочечной ДНК, комплементарную соответствующей молекуле РНК, представленную в клонирующем векторе.
Под "библиотекой кДНК" подразумевают коллекцию молекул рекомбинантной ДНК, содержащих вставки кДНК, которые включают в себя ДНК-копии мРНК, экспрессированную клеткой в период создания библиотеки кДНК. Такую кДНК можно приготовить с помощью методов, известным специалистам, описанным например у Ausubel et al. (см. выше) и у Maniatis et al. (см. выше). Вообще сначала выделяют РНК из клеток организма, из генома которого намерены проклонировать отдельный ген. Для цели настоящего изобретения предпочтительными являются линии лимфоцитарных клеток млекопитающего, в частности человека. В настоящее время предпочтительным вектором для этой цели является вирус коровьей оспы, штамм WR.
Под "вектором" подразумевают молекулу ДНК, выделенную, например, из плазмиды, бактериофага, или из вируса млекопитающего или насекомого, в которую можно вставить или проклонировать фрагменты ДНК. Вектор будет содержать один или больше уникальных сайтов рестрикции и может обладать способностью автономно реплицироваться в организме определенного хозяина или таком организме-посреднике, в котором клонируемая последовательность может репродуцироваться. Следовательно, под "экспрессионным вектором ДНК" подразумевают любой автономный элемент, способный управлять синтезом рекомбинантного пептида. Такие экспрессионные векторы ДНК включают в себя бактериальные плазмиды и фаги, а также плазмиды и вирусы млекопитающих и насекомых.
Под "практически чистым" подразумевают соединение, например, белок, полипептид или антитело, которое практически свободно от компонентов, естественно его сопровождающих. Вообще соединение является практически чистым, когда не менее 60%, более предпочтительно не менее 75%, и наиболее предпочтительно не менее 90% от общего объема в образце является соответствующим соединением. Степень очистки можно измерить любым подходящим способом, например, хроматографией на колонке, электрофорезом в полиакриламидном геле или HPLC-анализом. В контексте нуклеиновой кислоты термин "практически чистый" означает последовательность нуклеиновой кислоты, сегмент или фрагмент, которые непосредственно не контактируют с (т.е. не связаны ковалентно с) обоими кодирующими последовательностями, с которыми она непосредственно соприкасается (т.е. одна по 5'-концу, а другая по 3'-концу) в естественно встречаемом геноме организма, из которого выделили ДНК настоящего изобретения.
"Фрагмент" молекулы, как, например, любая из последовательностей кДНК настоящего изобретения, означает, что он относится к любому подклассу молекул нуклеотидного контига. "Аналог" молекулы означает, что он относится к неприродной молекуле, практически сходной с любой исходной молекулой или ее фрагментом. Указанная молекула является "практически сходной" с другой молекулой, если последовательность аминокислот обоих молекул является практической одной и той же. В частности "практически сходная" аминокислотная последовательность является той, которая проявляет не менее 50%, предпочтительно 85%, и наиболее предпочтительно 95% идентичности аминокислотной последовательности с естественной или референсной аминокислотной последовательностью и/или которая отличается от естественной или референсной последовательности лишь по консервативным аминокислотным заменам. Практически сходные аминокислотные молекулы обладают сходной биологической активностью. В том смысле как здесь используется, указанная молекула является "химически производной" другой молекулы, когда она содержит химически отличающиеся составные части отдельной молекулы. Такие составные части могут улучшить растворимость, абсорбцию, биологическое время полужизни молекулы и пр. Составные части могут альтернативно уменьшать токсичность молекулы, элиминировать или ослаблять любой нежелательный побочный эффект молекулы и т.п. Составные части, способные опосредовать такие эффекты, раскрыты, например, в фармацевтических науках Ремингтона, 16th ed., Mack Publishihg Co., Easton, P.A. (1980).
"Функциональное производное" гена химерного рецептора настоящего изобретения означает включение "фрагментов" или "аналогов" гена, которые являются "практически сходными" по нуклеотидной последовательности. "Практически сходная" нуклеиновая кислота кодирует практически сходные аминокислотные последовательности (как указано выше) и может также включать в себя любую последовательность нуклеиновой кислоты, способную гибридизировать с естественной или тест-последовательностью при соответствующих условиях гибридизации (см. , например, Ausubel et al., Современные методы в молекулярной биологии, Wiley Press, New York, NY (1989) для соответствующих жестких условий гибридизации).
"Практически сходный" химерный рецептор обладает сходной активностью с T-клеточным, B-клеточным или Fc-химерным рецептором "дикого типа". Наиболее предпочтительно, когда производное обладает 90%, более предпочтительно 70% и предпочтительно 40% активности химерного рецептора дикого типа. Активность производного функционального химерного рецептора включает в себя специфическое связывание (что касается его внеклеточной CD4-части) с ВИЧ-инфицированной клеткой и, в конечном итоге, уничтожении этой клетки; кроме того, химерный рецептор не делает клетку, несущую рецептор, восприимчивой к ВИЧ-инфицированию. Активность химерного рецептора можно проанализировать, используя, например, любой из методов, представленных здесь.
Последовательность ДНК, кодирующая химерный CD4-рецептор настоящего изобретения, или его функциональные производные, можно рекомбинировать с векторной ДНК в соответствии с традиционными методами, включающие тупление или наращивание концов для лигирования, расщепление ферментом рестрикции для получения соответствующих концов, достраивание липких концов надлежащим образом, обработку щелочной фосфатазой, чтобы избежать нежелательного соединения, и лигирование с помощью подходящих лигаз. Методики для таких манипуляций раскрыты у Maniatis et al. (см. выше) и хорошо известны специалистам в этой области.
Молекула нуклеиновой кислоты, такая как ДНК, как указано, "способна экспрессировать" полипептид, если она содержит нуклеотидные последовательности, содержащие информацию о регуляции транскрипции и трансляции, то такие последовательности "операбельно сцеплены" с нуклеотидными последовательностями, которые кодируют полипептид. Операбельное сцепление представляет собой сцепление, при котором регуляторные последовательности ДНК и экспрессируемая последовательность ДНК соединены таким образом, чтобы экспрессировать ген. Точная природа регуляторных областей, необходимых для генной экспрессии, может варьировать от организма к организму, но она всегда включает в себя область промотора, которая у прокариот содержит и промоторную последовательность (которая управляет инициацией транскрипции РНК), и последовательности ДНК, транскрибирование которых с РНК является сигналом инициации белкового синтеза. Такие области обычно включают 5'-некодирующие последовательности, вовлеченные в инициацию транскрипции и трансляции, и представлены TATA-боксом, кэппинг-последовательностью, CAAT-последовательностью и им подобными.
При необходимости с помощью вышеописанных методов можно получить соединение 3'-некодирующей области с последовательностью гена, кодирующего белок. Эту область можно использовать для регуляторных последовательностей, терминирующих ее транскрипцию с последовательностями, регулирующими терминирование транскрипции для ее терминирования и полиаденилирования. Таким образом, использованием 3'-области, естественно соприкасающейся с последовательностью ДНК, кодирующей белок, можно создать сигналы, терминирующие транскрипцию. Когда сигналы, терминирующие транскрипцию, функционируют неудовлетворительно в экспрессирующей клетке-хозяине, тогда функциональную 3'-область клетки-хозяина можно заменить.
Две ДНК последовательности (такие как последовательность области промотора и последовательность, кодирующую химерный CD4-рецептор), как указано, являются операбельно сцепленными, если характер сцепления между двумя последовательностями 1) не приводит к интродукции мутации в рамке считывания, 2) не подавляет способность последовательности области промотора управлять транскрипцией последовательности гена химерного рецептора или 3) не подавляет способность последовательности гена химерного рецептора быть транскрибированным с помощью последовательности области промотора. Область промотора будет операбельно сцеплена с последовательностью ДНК, если промотор способен усилить транскрипцию этой последовательности ДНК. Таким образом, при экспрессии белка необходимо, чтобы соответствующий хозяин узнавал сигналы транскрипции и трансляции.
Настоящее изобретение охватывает экспрессию белка химерного CD4-рецептора (или его функциональное производное) и в прокариотических, и в эукариотических клетках, но предпочтительно относится к экспрессии эукариотических клеток (и прежде всего к человеческим лимфтоцитам).
В соответствии с настоящим изобретением антитела можно приготовить любым имеющимся способом. Например, клетки, экспрессирующие белок химерного CD4-рецептора, или его функциональное производное, можно ввести животному с целью вызвать образование сыворотки, содержащей поликлональные антитела, которые способны связывать химеру.
В предпочтительном варианте осуществления способа антитела, в соответствии с настоящим изобретением, являются моноклональными антителами. Такие моноклональные антитела можно приготовить, применяя гибридомную технологию (Kohler et al., Nature 256:495 (1975); Kohler et al., Eur. J. Immunol. 6: 511 (1976); Kohler et al., Eur. J. Immunol. 6:292 (1976); Hammerling et al. , B: Моноклональные антитела и T-клеточные гибридомы, Elsevier, N.Y. pp. 565-684 (1981)). В основном такие методики предусматривают иимунизацию животного антигеном химерного CD4-рецептора. Полученные от таких животных спленоциты экстрагируют и подвергают слиянию с подходящей миеломной клеточной линией. В соответствии с настоящим изобретением можно использовать любую миеломную клеточную линию. После слияния полученные гибридомные клетки селективно поддерживаются на HAT-среде и затем клонируют методом лимитирующего разведения, как описано у Wands et al., (Castroenterology 80:225-232 (1981)). Полученные в результате такой селекции гибридомные клетки анализируют затем для идентификации клонов, которые секретируют антитела, способные связывать химеру.
В соответствии с настоящим изобретением антитела могут также быть поликлональными или, предпочтительно, регионспецифичными поликлональными антителами.
В соответствии с настоящим изобретением антитела к химерному CD4-рецептору можно использовать для контроля за количеством химерного рецептора (или клеток, несущих химерный рецептор) у пациента. Такие антитела хорошо подходят для использования в стандартных иммунодиагностических методах, известных в данной области, включая такие иммунометрические или "сэндвич"-методы в виде прямого сэндвич-анализа, обратного сэндвич-анализа и их сочетания. Эти антитела можно использовать в ряду любых комбинаций, выбранных специалистами, во избежание чрезмерного экспериментирования для осуществления иммуноанализа приемлемой специфичности, чувствительности и точности.
Ссылки на основные работы, формулирующие главные принципы иммунологии, включают Roitt, Основы иммунологии, 6th ed., Blackwell Scientific. Publications, publisher, Oxford (1988); Kimbal, Введение в иммунологию, 2nd ed., Macmillan Publishing Co. , publisher, New York (1986); Roitt et al., Иммунология, Gower Medical Publishing Ltd., publisher, London, (1985); Campbell, Техника работы с моноклональными антителами, у Burdon et al., eds., Техника лабораторных работ в биохимии и молекулярной биологии, том 13, Elsevier, publisher, Amsterdam (1984); Klein, Иммунология: The science of Self-Nonself Discimination, John Wiley & Sons, publisher. New York (1982) и Kennet et al. , Моноклональные антитела. Гибридома: новые подходы в биологических исследованиях, Plenum Press, publisher, New York (1980).
Под "детектированием" подразумевают наличие или отсутствие вещества или определяемое количество вещества. Поэтому данный термин относится к материалам, препаратам и способам настоящего изобретения, касающихся качественного и количественного определений.
Антитела и практически чистый антиген настоящего изобретения идеально подходит для создания набора. Такой набор может включать в себя переносной модуль для упаковки тесно расположенных одного или больше контейнеров, например в виде флаконов, тюбиков и т.п., каждый из указанных контейнеров включает в себя отдельные элементы, используемые в анализе.
Типы анализов, которые можно оформить в виде набора, многочисленны и представлены, например, конкурентным и неконкурентным анализом. Типичные примеры анализов, в которых можно использовать антитела настоящего изобретения: радиоиммунный анализ (РИА), иммуноферментный анализ (ИФА), твердофазный иммуносорбентный анализ (ELISA) и иммунометрические или "сэндвич"-иммуноанализы.
Под термином "иммунометрический анализ" или "сэндвич"-иммуноанализ подразумевают сочетание сэндвич-методов, прямой сэндвич-метод и обратный сэндвич-метод. Эти термины хорошо понятны специалистам данной области. Специалистам также понятно, что в соответствии с настоящим изобретением антитела пригодны для других вариантов и форм анализа, известных в настоящее время, или которые могут быть усовершенствованы в будущем. Подразумевают, что эти термины используются в рамках настоящего изобретения.
Под термином "специфически узнает и связывает" подразумевают, что антитело узнает и связывает полипептиды химерного рецептора, но практически не распознает и не связывает другие неродственные молекулы в образце, например, биологическом образце.
Под термином "терапевтическая клетка" подразумевают клетку, которая трансформирована химерным CD4-рецептором данного изобретения так, что она способна узнавать и уничтожать ВИЧ-инфицированную клетку; предпочтительно, чтобы такими терапевтическими клетками являлись клетки гемоноэтической системы.
Под термином "внеклеточный" подразумевают хотя бы часть молекулы, представленной на клеточной поверхности. Под термином "внутриклеточный" подразумевают по меньшей мере часть молекулы, представленной в цитоплазме терапевтической клетки. Под терминам "трансмембранный" подразумевают, как минимум, часть молекулы, пронизывающей плазматическую мембрану. Термины "внеклеточная часть", "внутриклеточная часть" и "трансмембранная часть", в том смысле как они используются здесь, могут включать в себя фланкирующие аминокислотные последовательности, которые простираются в соседние компартменты.
Под термином "олигомеризация" подразумевают образование комплекса с другими белками в виде димеров, тримеров, тетрамеров или олигомеров более высокого порядка. Такие олигомеры могут быть гомоолигомерами или гетероолигомерами. Термин "олигомеризационная часть" означает участок молекулы, которая управляет образованием комплекса (т.е., олигомера).
Под термином "цитолитический" подразумевают способность к уничтожению клетки (например, ВИЧ-инфицированной клетки или уничтожение инфекционного фактора (например, вируса ВИЧ)).
Под термином "вирус иммунодефицита" подразумевают ретровирус, который в форме дикого типа способен инфицировать T-клетки хозяина-примата, обладает морфогенезом и характерной морфологией подсемейства лентивирусов. Термин включает в себя, без ограничения, все варианты ВИЧ и SIV, в том числе ВИЧ-1, ВИЧ-2, SIVmac, SIVagm, SIVmnd, SIVsmm, SIVman, SIVmand и SIVcpz.
Под термином "МНС-независимый" подразумевают, что клеточный цитолитический ответ не требует присутствия МНС-антигена класса II на поверхности клетки-мишени.
Под термином "функциональное цитолитическое, трансдуцирующее сигнал производное" подразумевают функциональное производное (как определено выше), которое способно управлять не менее 40%, более предпочтительно 70% или наиболее предпочтительно не менее 90%, биологической активностью молекулы дикого типа. Используемое здесь "функциональное цитолитическое трансдуцирующее сигнал производное" может действовать в результате прямого сигнала терапевтической клетке уничтожить агент, связанный с рецептором, или клетку (например, в случае внутриклеточной части химерного рецептора) или действовать косвенно, способствуя олигомеризации с цитолитическими белками, трансдуцирующими сигнал (например, в случае трансмембранного домена). Такие производные можно протестировать на эффективность, например, используя описанные выше ин витро-анализы.
Под термином "функциональное производное, связывающее оболочку ВИЧ" подразумевают функциональное производное (как определено выше), которое способно связывать любой оболочечный белок ВИЧ. Функциональные производные можно идентифицировать используя, например, описанные выше ин витро-анализы
ТЕРАПЕВТИЧЕСКОЕ ВВЕДЕНИЕ
Трансформированные клетки настоящего изобретения использовали для терапии вирусного иммунодефицита. Современные способы введения таких трансформированных клеток включают адаптивную иммунотерапию или терапию переноса клеток. Эти способы позволяют возвратить трансформированные клетки иммунной системы в кровоток. Rosenberg, Sci. Am. 62 (May 1990); Rosenberg et al., N. Engl. J., Med. 325:570 (1990).
Фармацевтические препараты настоящего изобретения можно ввести любому животному, которое испытывает благоприятное воздействие соединений настоящего изобретения. На первом месте среди таких животных находится человек, хотя настоящее изобретение не намерено ограничиваться им.
Подробное описание
В первую очередь будут описаны чертежи
Краткое описание чертежей
Фиг. 1а представляет аминокислотную последовательность сайта слияния между CD4 (остатки 1-369) и разных рецепторных цепей (SEQ ID NOS: 58-41). Неподчеркнутая последовательность показывает положение аминокислот, кодируемых в рамках сайта BamHI, использованного для создания слитой конструкции. Начало трансмембранного домена отмечено вертикальной полосой. Последовательность η идентична последовательности s по аминоконцу, но отличается по карбоксиконцу (Jin et al., Proc. Natl. Acad.Sci USA 87:3319-3323 (1990)). Фиг. 1b представляет результаты проточного цитометрического анализа поверхностной экспрессии CD4, CD4:s, CD4:γ и
CD4: η в CVI-клетках. Клетки инфицировали вирусом, экспрессирующим CD4-химеры или CD16PI, инкубировали в течение 9 часов при температуре 37oC и окрашивали фикоэритрин-конъюгированными антиCD4 Mad Leu3A.
Фиг. 2 показывает поверхностную экспрессию CD16TM после коинфекции одним CD16ТМ (скученные точки), или коинфекции вирусов, экспрессирующем CD4: γ (штрихи) или CD4:s (сплошная линия). Редкие точки относятся к клеткам, инфицированными одним CD4:s и окрашенными 3G8 (Fleit et al., Proc. Natl. Acad. Sci USA 79:3275-3279 (1982)) (анти-CD16 Mad).
Фиг. 3 показывает поверхностную экспрессию CD16TM после коинфекции вирусами, экспрессирующими CD16ТМ и затем s-химерами: CD4:s (жирная линия), CD4: s C11G (сплошная линия), CD4: s (штриховая линия); CD4:s C11G/C11G (скрученные точки); без коинфекции (CD16ТМ только, редкие точки). Клетки инкубировали с анти-CD16 Mad 3G8 и Fab'-конъюгированными с фикоэритрином козьими антителами к мышиному lgG. Уровень экспрессии s-химер был практически идентичен для разных проанализированных мутантов, а коинфицированные клетки вирусами, экспрессирующие CD16ТМ и s-химеры, ощутимо не изменяют поверхностную экспрессию химер.
Фиг. 4a-d показывает увеличение ионов свободного внутриклеточного кальция после сшивки мутантных s-химер в T-клеточной линии. EE6-клетки Jurkat (Weiss et al., J. Immunol. 133:125-128 (1984)) инфицировали рекомбинантными вирусами коровьей оспы и анализировали с помощью проточной цитометрии. Представленные результаты относятся к пропущенной через затвор CD4-популяции, так что проанализированы только клетки, экспрессирующие нужный химерный белок. Среднее отношение фиолетовой и голубой Inbo-1 флуоресценции отражает внутриклеточную концентрацию свободного кальция в популяции в виде целых чисел и процентной доли отвечающих клеток, соответствующих доле клеток, которые превышают заданное пороговое отношение (брали так, чтобы 10% необработанных клеток были позитивными). Фиг. 4a и 4b показывают Jurkat-клетки, экспрессирующие CD4:S (сплошная линия) или CD16:s (штриховая линия), которые подвергли воздействию анти-CD4 Mag Leu3a (фикоэритриновый конъюгат), затем перекрестно сшили козье антитело с мышиным lgG. Точечная линия показывает ответ неинфицированных клеток к анти-CD3 MAt OKT3. Фиг. 4c и 4d показывает Jurkat-клетки, экспрессирующие CD4:SD15G (сплошная линия); CD4:SC11G/-D15G (штрихи); или CD4:SC11G (точки), которые обработали и проанализировали, как на фиг. 4a и 4b.
Фиг. 5a-c показываeт, что рецепторы CD4:s, CD4:η и CD4:γ позволяют цитолитическим т_лимфоцитам (ЦТЛ) уничтожать мишени, экспрессирующие молекулы gp120/41 ВИЧ-1. Фиг. 5a: черные кружки - это ЦТЛ, экспрессирующие CD4:s, проинкубированные с HeLa-клетками, экспрессирующими gp120/41; светлые кружки - это ЦТЛ, экспрессирующие CD4:s, проинкубированные с неинфицированными HeLa-клетками; черные квадраты - это неинфицированные ЦТЛ, проинкубированные с HeLa-клетками, экспрессирующими gp120/41; светлые квадраты - это неинфицированные ЦТЛ, проинкубированные с неинфицированными HeLa-клетками. Фиг. 5b: черные кружки - это ЦТЛ, экспрессирующие CD4:η, проинкубированные с HeLa-клетками, экспрессирующими gp120/41; светлые кружки - это ЦТЛ, экспрессирующие CD4:γ, проинкубированные с HeLa-клетками, экспрессирующими gp120/41; светлые квадраты - это ЦТЛ, экспрессирующие CD4; s-химеру двойного мутанта C11G/D15G, проинкубированные с HeLa-клетками, экспрессирующими gp120/41. Фиг. 5c: проточный цитометрический анализ CD4, экспрессируемых ЦТЛ, использованных на фиг. 5b. Для того чтобы уточнить соотношения мишени и эффектора, процентную долю клеток, экспрессирующих CD4-химеру, определяли вычитанием взвешенной величины негативной (неинфицированной) популяции путем наложения гистограмм; для сравнения на этой фигуре неинфицированные клетки обозначили произвольной пороговой величиной, которая дала приблизительно ту же долю позитивных клеток среди других клеточных популяций, как бы вычитанием гистограмм.
Фиг. 6a-b показывает специфичность CD4-направленного цитолиза. Фиг. 6a: черные кружки - это ЦТЛ, экспрессирующие CD4:s, проинкубированные с HeLa-клетками, экспрессирующими CD16PI; светлые кружки - это ЦТЛ, экспрессирующие CD4, проинкубированные с HeLa-клетками, экспрессирующими gp120/41; светлые квадраты - это ЦТЛ, экспрессирующие CD16PI, проинкубированные с HeLa-клетками, экспрессирующими gp120/41. Фиг. 6b: черные кружки - это ЦТЛ, экспрессирующие CD4:s, проинкубированные с клетками Raji (МНС класса II); светлые кружки - это неинфицированные ЦТЛ-клетки, проинкубированные с клетками RJ2.2.5 (МНС класса II Raji-мутанта); черные квадраты - это неинфицированные ЦТЛ, проинкубированные с клетками Raji (МНС класса II); светлые квадраты - это ЦТЛ, экспрессирующие CD4: s, проинкубированные с клетками RJ2.2.5 МНС класса II). Масштаб ординаты расширенный.
Фиг. 7a-b характеризует CD16:s-химерный рецептор. Фиг. 7a представляет собой схематическую диаграмму CD16:s-слитого белка. Внеклеточную часть, в форме сшитого с фосфатидилинозитолом мономерного CD16, присоединили к димеру s наружной части трансмембранного домена. Последовательность белка (SEQ ID NOSS: 42, 43) в точке присоединения показана ниже. Фиг. 7b показывает результаты проточного цитометрического анализа динамики кальция после сшивки CD16:s-химеры либо с TCR-позитивной, либо с TCR-негативной клеточной линией. Показано среднее соотношение фиолетовой и голубой флуоресценции (измеряет относительную ионную концентрацию кальция) в клеточных популяциях, обработанных антителами, в нулевой точке отсчета времени. Черные квадраты - это ответ Jurkat-клеток на анти-CD3 Mab OKT3; черные треугольники - это ответ CD16:s на анти-CD16 Mab 3G8, сшитый с REX33A TCR-мутанта; светлые квадраты - это ответ на CD16:s, сшитого с Jurkat TCR мутантной линией JRT3.T3.5; светлые тpeугольники - это ответ на CD16:s-сшивку в Jurkat-клeткax; и точки - это ответ на нехимерный CD16 в REX33A TCR-клеточной линии.
Фиг. 8а-b показывает анализ делеций цитолитического потенциала. Фиг. 8a показывает местоположения концевых точек делеций в s. Здесь также как и в другом месте, мутации в s представлены условным остатком, локализованного в мутанте оригинального остатка, так что D66* указывает, к примеру, замену Asn-66 терминирующим кодоном. Фиг. 8b показывает результаты цитолитического анализа неделетированного CD16: s и характерных s-делеций. Гибридомные клетки, экспрессирующие поверхностное антитело к CD16, нагружали 51Cr и инкубировали с увеличивающимся количеством человеческих цитолитических лимфоцитов (ЦТЛ), зараженных рекомбинантами вируса коровьей оспы, экспрессирующими химеры CD16: s. Процентная доля высвобожденного 51Cr графически представлена как функциональное отношение эффекторных клеток (ЦТЛ) к клеткам-мишеням (гибридома) (э/м). Черные кружки - это цитолиз, опосредованный клетками, экспрессирующими CD16:s Aсп66* (mfi 940,2); светлые квадраты - это цитолиз, опосредованный клетками, экспрессирующими CD16:s Глу60* (mfi 16,0); светлые кружки - это цитолиз, опосредованный клетками, экспрессирующими CD16: sТир51* (mfi 17,4); черные треугольники - это цитолиз, опосредованный клетками, экспрессирующими CD16:s Фен34*, (mfi 17,8); и светлые треугольники - это цитолиз, опосредованный клетками, экспрессирующими нехимерный CD16 (mfi 591). Хотя в этом эксперименте экспрессию CD16:s Асп66* не наблюдали, в отличие от других слитых белков цитолиз с помощью клеток, зкспрессирующих CD16:s на эквивалентных уровнях в одном и том же эксперименте давал практически идентичные результаты, проявленные клетками, экспрессирующими CD16:s Асп66.
Фиг. 9а-d показывает, что элиминацию потенциальных для трансмембраны взаимодействий обнаруживает короткий s-сегиент, способный опосредовать цитолиз. Фиг. 9a представляет собой схематическую диаграмму мономерных бираздельной и трираздельной химер. Верхняя часть представляет собой CD16:s-конструкцию, усеченную по остатку 65 и не имеющей трансмембранных остатков Цис и Асп. Ниже находятся CD16: CD5: s- и CD16:CD7:s-конструкции и родственные контрольные конструкции. Пептидные последовательности внутиклеточных доменов показаны ниже (SEQ ID NOS: 45-47). Фиг. 9b показывает цитолитическую активность мономерной химеры делетированных мутантов. Цитолитическая активность клеток, зкспрессирующих CD16:s (черные кружки; mfi 495) сравнили с активностью клеток, экспрессирующих CD16:5 Асп66* (черные квадраты; mfi 527) или с мутантами CD16:s Цис11Гли/Асп15Гли/Асп66* (светлые квадраты; mfi 338) и CD16: s Цис11Гли/Асп15Гли/Глу60* (черные треугольники; mfi 259). Фиг. 9с показывает цитолитическую активность, опосредованную трираздельными слитыми белками. Черные треугольники - это CD16:s Асп66*; светлые квадраты - это CD16: s(48-65); черные квадраты - это CD16:7:s (48-65); светлые треугольники - это CD16: 7: s (48-59); светлые кружки - это CD16:5; черные кружки - это CD16:7. Фиг. 9d показывает мобилизацию кальция мутантом и трираздельными химерами в TCR-негативной JRT3. T3.5-мутантной линии клеток Jurkat. Светлые кружки - это ответ клеток, экспрессирующих димерный CD16:sАсп66*; черные квадраты - это ответ клеток, зкспрессирующих CD16:sЦис11Гли/Асп15Гли/Асп66*; светлые квадраты - это ответ клеток, экспрессирующих CD16: sЦиc11Гли/Acп15Гли/Глe60*; черные треугольники - это ответ клеток, экспрессирующих CD16: 7: s (48-65); и светлые треугольники - это ответ клеток, зкспрессирующих CD16:s (48-59).
Фиг. 10a-f показываeт вид индивидуальных аминокислот в активность мотива из 18 аминокислот, трансдуцирующего цитолитический сигнал. Фиг. 10a и 10b показывают цитолитическую активность, а фиг. 10c показывает мобилизацию кальциевого иона, опосредованную химерами, несущими точечные мутации вблизи карбоксиконца тирозина (Y62). Фиг. 10a и 10b представляют данные, собранные для клеток, экспрессирующих, соответственно, низкие и высокие количества CD16: s-слитых белков. Идентичные обозначения использовали для анализа мобилизации кальция и цитолиза и показаны отдельным буквенным кодом справа. Черные кружки - это клетки, экспрессирующие CD16:s (mfi в A, 21; B, 376); черные квадраты - это клетки, экспрессирующие CD16:7:s (48-65) (mfi A, 31; B, 82); светлые квадраты - это CD16:7:s (48-65) Глу60Гли (mfi A, 33; B, 92), крестики - это CD16:7:s (48-64) Асп63Асн (mfi A, 30; B, 74); черные треугольники - это CD16:7:s (48-65) Тир62фен (mfi A, 24; B, 88); светлые кружки - это CD16:7:s (48-65) Глу61Глн (mfi A, 20; D, 62); светлые треугольники - это CD16: 7:s (48-65) Тир62Сер (mfi B, 64). Фиг. 10d и 10e показывают цитолитическую активность, а фиг. 10f показывает мобилизацию иона кальция с помощью химера, несущих точечные мутации вблизи аминоконца тирозина (Y51). Идентичные обозначения использовали для анализа мобилизации кальция и цитолиза, которые показаны справа. Черные кружки - это клетки, зкспрессирующие CD16: s (mfi в D, 21,2; в E, 672); черные квадраты - это клетки, экспрессирующие CD16: 7: s (48-65) (mfi D, 31,3; E, 179); черные треугольники - это CD16: 7: s (48-65) Асн48Сер (mfi D, 22,4; E, 209); светлые квадраты - это CD16: 7: s (48-65) Лей50Сер (mfi D, 25,0; E, 142); и светлые треугольники - это CD16:7:s (48-65) Тир51фен (mfi D, 32,3; E, 294).
Фиг. 11a-b показывает выстраивание внутренних повторов s и сравнивает их способность поддерживать цитолиз. Фиг. 11a представляет собой схематическую диаграмму химер, образованных в результате разделения внутриклеточного домена s на три части и присоединения их к трансмембранному домену CD16:7-химеры. Последовательности внутриклеточных доменов показаны ниже (SEQ ID NOS: 48-50), с разделенными, взятыми в рамку, остатками и связанными остатками, отмеченными звездочками. Фиг. 11b показывает цитолитическую эффективность трех s-субдоменов. Черные кружки - это клетки, экспрессирующие CD16:s (mfi 476); черные квадраты - это CD16:7:s (33-65) (mfi 68); светлые квадраты - это CD16: 7: s (71-104) (mfi 114); и черные треугольники - это CD16:7:s (104-138) (mfi 104).
Фиг. 12 представляет собой схематически диаграмму CD16:FcRyII-химер.
Фиг. 13a-b показываeт мобилизацию кальция после сшивки химер CD4:FcRyII и CD16:FcRyII. Фиг. 13a показывает соотношение фиолетовой и голубой флуоресценции, испускаемой клетками, нагруженных кальцием, чувствительного к флюорофору Indo-1, показанного в виде временной функции после сшивки внутриклеточной части CD16-домена с антителами, Фиг. 13b показывает сходные результаты анализа по увеличению в соотношении фиолетовой и голубой флуоресценции клеток, несущих CD4:FcRyII-химеры, после сшивки с антителами.
Фиг. 14a-b показываeт результаты анализов цитолиза химер CD4:GcRyII и CD16: FcRyII. Фиг. 14a показывает процентную долю 51Cr, высвобождаемого из анти-CD16 гибридомных (мишень) клеток, когда клетки подвергаются действию увеличивающегося количества цитотоксических T-лимфоцитов, экспрессирующих CD16:FcRyII-химеры (эффекторные клетки). Фиг. 14b показывает результаты аналогичного анализа, цитотоксически опосредованного с помощью CD41FcRyII-химер против клеток-мишеней, экспрессирующих оболочечные гликопротеины ВИЧ.
Фиг. 15a-e показываeт идентификацию остатков в A-хвосте FcRyII, который важен для цитолиза. Фиг. 15a представляет собой схематическую диаграмму делеционных конструкций. Фиг. 15b и 15c показывают мобилизацию кальция и цитолиз с помощью вариантов CD16:FcRyII A с карбоксиконцевой делецией. Фиг. 15d и 15e показывают мобилизацию кальция и цитолиз трираздельными химерами, несущими постепенно уменьшающееся количество аминоконцов во внутриклеточном A-хвосте CD16:FcRyII.
Фиг. 16 (SEQ ID NO: 24) показывает аминокислотную последовательность дельта-белка CD3-рецептора; ограниченная рамкой последовательность представляет предпочтительный цитолитический сигнал трансдуцирующей части.
Фиг. 17 (SEQ ID NO: 25) показывает аминокислотную последовательность гамма-белка T3-рецептора; ограниченная рамкой последовательность представляет предпочтительный цитолитический сигнал трансдуцирующей части.
Фиг. 18 (SEQ ID NO: 26) показывает аминокислотную последовательность белка mbi-рецептора, ограниченная рамкой последовательность представляет предпочтительный цитолитический сигнал трансдуцирующей части.
Фиг. 19 (SEQ ID NO: 27) показывает аминокислотную последовательность белка B29-рецептора; ограниченная рамкой последовательность представляет предпочтительный цитолитический сигнал трансдуцирующей части.
Фиг. 20 показывает схематическую диаграмму CD4-химер. Молекула "A" представляет собой CD4(D1-D4): Ig: CD7; молекула "B" представляет собой CD4(D1, D2): Ig:CD7; молекула "C" представляет собой CD4(D1, D2):Ig:CD7:s и молекула "E" представляет собой CD4:s. Внутриклеточный домен CD4-молекулы человека, соответствующий аминокислотам 1-394 предшественника, соединили с помощью BamHI-сайта с шарниром CH1- и CH2-доменами человеческого IgG1, как описано раньше (Zettlmeissl et al., DNA Cell Biol. 9:347 (1990)), за тем исключением, что вариант кДНК последовательности Ig человека использовали, чтобы сделать возможной экспрессию рекомбинантных вирусов коровьей оспы. Двухдоменный вариант CD4-химер создали с помощью вставки BamHI-адаптера по уникальному NheI-сайту (соответствующий 200-й аминокислоте) в кДНК CD4-предшественника. Последовательности, взаимодействующие с мембраной, состоят из 22 остатков из первого экзона человеческого мембранносвязанного IgG1 после остатков 146-205 маркера CD7. Аминокислоты 55-163 s-цепи служат в качестве пускового мотива терапевтических конструкций (C и D). В тетрараздельных конструкциях, содержащих s-цепь, внутриклеточную экспрессию s-цепи документально установили с помощью коммерческого антитела к внутриклеточному домеру (Coulter).
Фиг. 21 показывает цитолиз клетки-мишени, экспрессирующий оболочечный гликопротеин ВИЧ, опосредованный клоном цитотоксических T-клеток, WH3, экспрессирующих, в качестве эффекторных молекул, разнообразные химеры, производные CD4. Для анализа цитотоксичности маркеры CD8+, CD4- HLA B44, ограниченные T-клеточной линией WH3, поддерживали в среде IMD с добавлением 10% человеческой сыворотки, как описано здесь выше. Клетки стимулировали гамма-облученными (3000 рад) мононуклеарными клетками, несущими B44, и фитогемагглютинином (ФГА) при концентрации 1 мкг/мл. После одного дня стимуляции ФГА разбавляли до 0,5 мкг/мл путем добавления свежей среды; через 3 дня среду полностью меняли. Клетки росли не менее 10 дней до использования в цитотоксическом анализе. Клетки инфицировали соответствующими рекомбинантными вирусами коровьей оспы, как описано здесь для vPE16. Время инфицирования клеток в полной среде продлевали на 3-4 часа, после чего клетки собирали центрифугированием и ресуспендировали с плотностью 1·107/мл. 100 мкл этих клеток вносили в каждую лунку U-образной плашки для микротитрования, где в каждой лунке уже содержалось по 100 мкл полной среды, и разбавляли в 2 раза серийными операциями. Две лунки для образцов не содержали лимфоцитов, чтобы сделать возможным спонтанное высвобождение хрома и полностью измерить поглощение хрома. Клети-мишени, HeLa-клетки сублинии S3 (HeLa-S3, ATCC), инфицировали, как указано выше, vPE16, в 10 см чашках. 106 инфицированных клеток отделили с помощью PBS и 1 мМ ЭДТА, центрифугировали и ресуспендировали в 100 5-1 мкл 51Cr-хромата натрия (1 мКи/мл в PBS) в течение 1 часа при 37oC и затем отмывали три раза PBS. 100 мкл меченых клеток-мишеней вносили в каждую лунку. Плашку для микротитрования подвергали центрифугированию при 750·g в течение 1 минуты и инкубировали в течение 4 часов при 37oC. По окончании инкубационного периода клетки в каждой лунке ресуспендировали осторожным пипеттированием, образец удаляли для суммарного определения включенной метки и плашку для микротитрования центрифугировали при 750·g в течение 1 минуты. Извлекали аликвоты (100 мкл) супернатанта и метку считали на сцинтилляционном счетчике гамма-излучения. Уточняли соотношение эффектор:мишень для определения процентной доли инфицированных клеток с помощью проточной цитометрии.
Фиг. 22 показывает репликацию ВИЧ-1 в трансфицированной клеточной линии. В сублинин эмброиональных клеток почки человека линии 293 выявляли клеточные линии, стабильно экспрессирующую CD4 дикого типа и различные рекомбинентные химеры. Получили изолят IIIB вирусного штамма ВИЧ-1 с титром ≈106 инфекционных частиц/мл, измеренного с помощью метода разведения до конечной точки с использованием, в качестве индикатора, человеческой T-клеточной линии C8166. Инфицирование осуществляли примерно при 1 множественном заражении (MOI) в течение 8-12 часов при 37oC. На следующий день клетки отмывали три раза PBS, трипсинизировали, переносили в новые чашки и культуральный супернатант отбирали для p24-титрования (день обозначили 0). Каждые 3-4 дня собирали супернатанты культивируемых клеток и сохраняли до проведения p24-анализов. Клетки пополняли свежей средой, содержащей гигромицин B в концентрации 100 мкг/мл. Анализ культуральных супернатантов проводили, используя коммерческий набор для анализа, проверенного ELISA антигена p24 ВИЧ-1, в соответствии с инструкцией производителя (Coulter). Результаты представлены двумя независимыми экспериментами примерно одинаковой продолжительности.
Фиг. 23 показывает последовательность нуклеиновой кислоты (SEQ ID NO:28) и аминокислотную последовательность (SEQ ID NO:29) доменов D1-D4 молекулы CD4 (CD4 Bam).
Фиг. 24 показывает последовательность нуклеиновой кислоты (SEQ ID NO:30) и аминокислотную последовательность (SEQ ID NO:31) D1-D2-доменов CD4 (CD4 Nhe).
Фиг. 25 показывает последовательность нуклеиновой кислоты (SEQ ID NO:32) и аминокислотную последовательность (SEQ ID NO:33) шарнира, доменов CH2 и CH3 человеческого IgG1 (Igh23 Bam).
Фиг. 26 показывает последовательность нуклеиновой кислоты (SEQ ID NO:34) и аминокислотную последовательность (SEQ ID NO:35) трансмембранного домена CD7 (TM7 Bam Mlu).
Фиг. 27 показывает последовательность нуклеиновой кислоты (SEQ ID NO:36) и аминокислотную последовательность (SEQ ID NO:37) внутриклеточного домена zeta (Zeta Mlu Not).
Фиг. 28 показывает последовательность ДНК (SEQ ID NO:51) и первичную аминокислотную последовательность (SEQ ID NO:52) синтетической альфа-спирали.
ПРИМЕР I. Конструкция человеческих IgG1:рецепторных химер
Последовательности тяжелой цепи человеческого IgG1 готовили путей присоединения последовательностей Cн3-домена с фрагментом кДНК, полученной из 3'-конца мРНК трансмембранной формы антитела. Этот 3'-конец получили с помощью полимеразной цепной реакции, используя библиотеку кДНК небной миндалины в качестве субстрата и олигонуклеотиды, имеющие последовательности:
CGC GGG GTG ACC GTG CCC TCC AGC AGC TTG GGC (SEQ ID NO:7) и
CGC GGG GAT CCG TCG TCC AGA GCC CGT CCA GCT CCC CGT CCT GGG CCT CA (SEQ ID NO:8),
соответствующие 5'- и 3'-концам, соответствующих фрагментов ДНК. 5'-олигонуклеотид комплементарен сайту в Сн1-домене человеческого IgG1, а 3'-олигонуклеотид комплементарен именно сайту 5'-последовательности, кодирующей домен, пронизывающий мембрану. ПЦР-продукт обрабатывали BStXI и BamHI и лигировали между BStXI- и BamHI-сайтами полусинтетического гена антитела IgG1, несущего вариабельную и константную области. После встраивания BstXI в BamHI-фрагмент амплифицированные части конструкции заменили на SmaI-сайт в Сн3 путем рестрикции замененного фрагмента так, чтобы получить в ЦПР-реакции только часть амплификанта между SmaI-сайтом и 3'-олигонуклеотидом.
Для создания IgG1: ζ-химерного рецептора человека ген тяжелой цепи, оканчивающийся в BamHI-сайте, соединили с BamHI-сайтом ζ-химеры, описанной ниже, так, чтобы последовательности антитела образовали внутриклеточную часть. Проточная цитометрия COS-клеток, трансфицированных плазмидой, кодирующей химеру, показала высокий уровень экспрессии детерминант антитела, когда контрансфекцировали экспрессионную плазмиду, кодирующую легкую цепь кДНК, и умеренную экспрессию детерминант антитела, когда экспрессионная плазмида легкой цепи отсутствовала.
Подобные химеры, включающие в себя человеческий IgGl, слитый с η- или λ-цепью (см. ниже), или любую трансдуцирующую сигнал часть T-клеточного рецептора или Fc-рецепторный белок в принципе можно сконструировать, как описано выше, используя стандартные методики молекулярной биологии.
Для создания единственной транскрипционной единицы, которая позволяла бы и тяжелой, и легкой цепям экспрессироваться от одного промотора, из последовательностей, кодирующих тяжелую и легкую цепь и 5'-нетранслируемую часть мРНК, кодирующую 78 кДа регуляторный белок для глюкозы, известный также как grp78, или BiP, создали плазмиду, кодирующую двухцистронную мРНК, grp78-последовательности получили ПЦР из геномной ДНК человека, используя праймеры, имеющие следующие последовательности:
CGC GGG CGG CCG CGA CGC CGG CCA AGA CAG CAC (SEQ ID NO:9) и CGC GTT GAC GAG CAG CCA GTT GGG CAG CAG CAG (SEQ ID NO: 10) соответственно для 5'- и 3'-концов. Полимеразные цепные реакции с этими олигами осуществляли в присутствии 10% диметилсульфоксида. Полученный ПЦР-фрагмент обрабатывали NotI и HincII и встраивали между NotI- и HpaI-cайтами, расположенными ниже последовательностей, кодирующих человеческий IgG1. Последовательности кДНК, кодирующие легкие каппа-цепи человеческого IgG1, встраивали затем ниже лидерной последовательности grp78, используя HincII-сайт и другой сайт в векторе. Полученная в результате этих операций экспрессионная плазмида состояла из полусинтетического гена тяжелой цепи, следующего за лидерными последовательностями grp78, за последовательностями кДНК легких каппа-цепей, за сигналами полиаденилирования, полученными и ДНК-фрагмента SV40. COS-клетки, трансфецированные экспрессионной плазмидой, показавшие отчетливое увеличение экспрессии детерминант тяжелой цепи, сравнивали с трансфецирующей плазмидой, кодирующей только одни детерминанты тяжелой цепи.
Для создания двухцистронного гена, включающего в себя химеру тяжелая цепь/рецептор и легкую цепь, расположенную выше последовательности тяжелой цепи, можно заменить на любой вышеописанный химерный ген тяжелая цепь/химера.
ПРИМЕР II. Конструирование химерных CD4-рецепторов
С помощью полимеразной цепной реакции из библиотеки, полученной из опухолевой клеточной линии HPB-ALL (Aruffo et al., Proc. Natl. Acad. Sci. USA 84: 8573-8577 (1987b) и из человеческих естественных киллерных клеток, выделяли ζ-кДНК человека (Weissman et al., Proc. N t1. Acad. Sci. USA 85:9709-9713 (1988b)) и γ-кДНК (Kustler et al., J. Biol. Chem. 265:6448-6452 (1990), а η-кДНК выделяли из библиотеки мышиных тиномитов (Jin et al., Proc. Nаtl. Acad. Sci. USA 87:3319-3323 (1990)), ζ-,η- и γ-кДНК присоединяли к внеклеточному домену созданной формы CD4, обладающему BamHI-сайтом, расположенному непосредственно выше домена, пронизывающего мембрану (Aruffo et al., Proc, Natl. Acad. Sci. USA 84:8573-8577 (1987b)); Zettlmeissl et al., DNA Cell Biol. 9:347-353 (1990)), который присоединяли к BamHI-сайту, естественно присутствующему в ζ- и η-кДНК при сходном расположении нескольких остатков выше домена, пронизывающего мембрану (SEQ ID NO: 1, 3, 4 и 6). Для образования слитого белка с γ, BamHI-сайт встроили в последовательность с той же, приблизительно, локализацией (фиг. 1; SEQ ID NO: 2 и 5). Слитые гены интродуцировали в плазмиду, экспрессирующую вирус коровьей оспы, несущую gpt-ген E.coli в качестве селектируемого маркера, и встроенную в геном штамма WR коровьей оспы в результате гомологичной рекомбинации и селекции по росту в микрофенольной кислоте (Falkner et al., J. Virol. 62:1849-1854 (1988); Boyle et al., Gene 65:123-128 (1988)). Проточноцитометрический анализ показал, что рекомбинанты коровьей оспы управляют обильным продуцированием CD4: ζ и CD4: γ слитых белков на клеточной поверхности, тогда как экспрессия CD4: η практически редуцирована (фиг. 1b). Последнее наблюдение согласуется с недавним сообщением о том, что трансфекция η- кДНК экспрессионной плазмиды в мышиную гибридомную клеточную линию дала практически исчезающую экспрессию, чем трансфекция сравниваемой ζ- экспрессионной плазмиды (Clayton et al., J. Exp. Med. 172:1243-1253 (1990)). Иммунопреципитация клеток, инфицированных рекомбинантами коровьей оспы, обнаружила, что слитые белки образуются из ковалентных димеров, отличных от естественно встречаемого CD4-антигена. Молекулярные массы мономерных CD: ζ- и CD: γ-слитых белков и естественного CD4 равны, соответственно, 63, 55 и 53 кДа. Большие массы слитых белков примерно совпадают с большей длиной внутриклеточной части, которая превышает внутриклеточную часть CD4 на 75 (CD4: ζ- ) или на 5 CD4: γ остатков.
ПРИМЕР III. CD4-химеры можно ассоциировать с другими рецепторными цепями
Экспрессия клеточной поверхностью макрофаг/естественной киллерной клеткой человеческого Fc γ RIII (CD16ТМ), образуемого у трансфектантов, облегчается контрансфекцией мышиной (Kurosaki et al., Nature 342:805-807 (1989)) или человеческой (Hibbs et al., Science 246:1608-1611 (1989)) γ, также как и человеческой ζ (Lanier et al., Nature 342:803-805 (1989)).
Подтверждает эти сообщения экспрессия химер, делающих возможной также поверхностным экспрессию CD16TM при доставке к клетке-мишене либо в результате контрасфекции, либо коинфекции рекомбинантными вирусами коровьей оспы (фиг. 2). В исследованной клеточной линии стимуляция поверхностной экспрессии с помощью ζ была более выраженной, чем стимуляция с помощью γ (фиг. 2), тогда как естественный CD4 не усиливает поверхностную экспрессию CD16ТМ.
ПРИМЕР IV. ζ-Мутанты по Асп не коассоциируют с Fc-рецептором
Для создания химер, которые бы не ассоциировали с существующим антигеном или Fc-рецепторами, приготовили слитые белки ζ-мутанта, в которых недостает либо внутримембранного Асп-остатка, либо внутримембранного Цис-остатка. Проточная цитометрия показала, что интенсивность клеточно-поверхностной экспрессии различными мутантными химерами заметно не отличалась от немутантного предшественника, а эксперименты по иммунопреципитации показали, что суммарная экспрессия химер была схожей. Как и ожидалось, мутантные химеры, не имеющие трансмембранного цистеинового остатка, не обнаруживают образования димеров, сшитых дисульфидной связью. Две мутантные химеры, не имеющие Асп, были неспособны поддерживать поверхностную экспрессию CD16ТМ, тогда как мономерные химеры, не имеющие остатка Цис, но несущие остаток Асп, имели возможность коэкспрессировать CD16ТМ, но с пониженной эффективностью, чем родительский димер (фиг.3).
ПРИМЕР V. Мутантные рецепторы сохраняют способность инициировать кальциевый ответ
Чтобы выяснить, действительно ли сшивка слитых белков делает возможной накопление свободного внутриклеточного кальция способом, сходным с известным способом для антигенного рецептора T-клетки, клетки лейкозной T-клеточной линии человека, Jurkat E6 (ATCC, каталожный номер TIB 152, Американская типовая коллекция культур, Rockville, MD) инфицировали рекомбинантами коровьей оспы и измеряли отношение концентрации цитоплазматического кальция после сшивки внеклеточного домена с антителами. Проточноцитометрические измерения осуществляли с клетками, нагруженными чувствительным к кальцию красителем Indo-1 (Grynkiewich et al., J.Biol. Chem. 260:3340-3450 (1985); Rabinovitch et al., J. Immunol, 137:952-961 (1986)).
Фиг. 4a-d показывают результаты опытов кальциевого потока для клеток, инфицированных CD4: ζ и Асп- и Цис-мутантами ζ. Сшивка химер воспроизводимо увеличивает внутриклеточный кальций. Подобно этому CD4: η и CD4: γ делают возможным накопление внутриклеточного кальция в инфицированных клетках. Jurkat-клетки экспрессируют низкие уровни CD4 на клеточной поверхности, однако, сшивка естественного CD4 в присутствии или отсутствии CD16: ζ не меняет уровней внутриклеточного кальция (фиг. 4a-b).
ПРИМЕР VI. CD4: ζ,η-, и γ-химеры опосредуют цитолиз мишеней, экспрессирующих gp120/41 ВИЧ
Чтобы выяснить, действительно ли химерные рецепторы могли бы запустить цитолитические эффекторные программы, создали модель в виде системы мишень: эффектор, основанную на узнавании CD4 оболочечного комплекса gp120/gp41. Клетки HeLa инфицировали рекомбинантными вирусами коровьей оспы, экспрессирующими gp120/gp41 (Chakrabarti et al., Nature 320:535-537 (1986); Earl et al. , J.Virol. 64:2448-2451 (1990)) и метили 51Cr. Меченые клетки инкубировали с клетками аллоспецифической CD8+, CD4-) цитотоксической линией T-лимфоцитов, которая была инфицирована рекомбинантами коровьей оспы, экспрессирующими CD4: ζ-, CD: η- или CD4: γ- химеры, или двойную мутантную химеру CD4: ζ Цис11Гли:Асп15Гли. Фиг. 5a-c показываeт, что клетки HeLa, экспрессирующие gp120/41, были специфически лизированы цитотоксическими T-лимфоцитами (ЦТЛ), экспрессирующими CD4-xимepы. Неинфицированные клетки HeLa не были атакованы ЦТЛ, вооруженными CD4: ζ-химерами, а клетки HeLa, экспрессирующие gp120/41, не узнавались неинфицированными ЦТЛ. Для сравнения эффективности разных химер измерением в проточной цитометрии уточняли соотношения эффектора к мишени для фракции ЦТЛ, экспрессирующей CD-химеры, и для фракции клеток HeLa, экспрессирующих gp120/41. Фиг. 5c показывает результаты цитометрических анализов CD4-зкспрессии ЦТЛ, использованных в экспериментах по цитолизу, показанных на фиг. 5a и 5b. Хотя средняя плотность поверхностного CD4: ζ намного превышала среднюю плотность CD4: η, цитолитические эффективности экспрессирующих клеток были одинаковы либо близки. Для фракции мишеней, экспрессирующих gp120, уточненная эффективность цитолиза, опосредованная белками CD4: ζ и CD4: η, оказывается сравнимой с лучшими показателями эффективности, сообщенными для специфичных T-клеточных рецепторных пар мишень:эффектор (среднее соотношение эффектора к мишени при 50% секреции T-клетками, экспрессирующими CD4: ζ, составляло 1,9±10,99, n=10). Слияние CD4: γ было менее активным, как и слияние CD4: ζ, из-за отсутствия трансмембранных остатков Асп и Цис. Однако в обоих случаях наблюдали цитолиз (фиг. 5b-c).
Чтобы проверить возможность того, что инфицирование коровьей оспой может способствовать артефактному узнаванию с помощью ЦТЛ, подобные эксперименты по цитолизу осуществляли с клетками-мишенями, инфицированными рекомбинантами коровьей оспы, экспрессирующими фосфатидилинозитол-связанную форму CD16 (CD16PI) и меченую 51Cr, и с ЦТЛ, инфицированными для контроля рекомбинантами, экспрессирующими либо CD16PI или CD: ζ. Фиг. 6a показывает, что T-клетки, экспрессирующие не CD4-химеры, не узнают естественные клетки HeLa или клетки HeLa, экспрессирующие gp120/41, и подобно этому T-клетки, экспрессирующие CD4-химеры не узнают клетки HeLa, экспрессирующие другие поверхностные белки, кодируемые коровьей оспой. Кроме того, цитотоксические T-лимфоциты, экспрессирующие нехимерный CD4, незначительно лизируют клетки HeLa, зкспрессирующие пз120.41 (фиг. 6a).
ПРИМЕР VII. Клетки, несущие МНС класса II, не атакуются химерами
Предполагается, что CD4 взаимодействует с неполиморфной последовательностью, экспрессирующей антиген МНС класса II (Gay et al., Nature 328:626-629 (1987)); Sleckman et al., Nature 328:351-353 (1987)). Хотя специфическое взаимодействие между CD4 и антигеном класса II никогда не было документировано на очищенных белках, при определенных условиях адгезия между клетками, экспрессирующими CD4, и клетками, экспрессирующими молекулы класса II, все же показали (Doyle et al., Nature 330:256-259 (1987); Clayton et al., J. Exp. Med. 172:1243-1253 (1990); Lammare et al., Science 245:743-746 (1989)). Затем выясняли, действительно ли можно было бы детектировать уничтожение клеток, несущих молекулы класса II. Фиг. 6b показывает, что специфический цитолиз, направленный с помощью CD4: 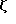 против клеточной линии Raji B, которая обильно экспрессирует антиген класса II, не наблюдается. Хотя умеренный (≈ 5%) цитолиз наблюдается, мутанты, негативные по классу II клеток Raji, RJ2.2.5 (Accolla, J. Exp. Med. 157:1053-1058 (1983)), показывают близкую восприимчивость с той, какую проявляют Raji-клетки, инкубированные с неинфицированными T-клетками.
против клеточной линии Raji B, которая обильно экспрессирует антиген класса II, не наблюдается. Хотя умеренный (≈ 5%) цитолиз наблюдается, мутанты, негативные по классу II клеток Raji, RJ2.2.5 (Accolla, J. Exp. Med. 157:1053-1058 (1983)), показывают близкую восприимчивость с той, какую проявляют Raji-клетки, инкубированные с неинфицированными T-клетками.
ПРИМЕР VIII. Последовательность, необходимая для индукции цитолиза zeta-цупью T-клеточного антиген/Fc-рецептора
Хотя химеры между CD4 и ζ могут вооружить цитотоксические T-лимфоциты (ЦТЛ) для уничтожения клеток-мишеней, экспрессирующих gp120 ВИЧ, для CD4 искали альтернативу для того, чтобы точно сравнить свойства zeta-химер, интродуцированных в человеческие T-клеточные линии. Такие линии клеток могут экспрессировать CD4, затрудняя возможность специфически определять взаимосвязь между типом или уровнем мобилизации кальция и цитотоксическим потенциалом разных химер. Чтобы обойти это затруднение, создавали химеры между ζ и CD16, в которых внеклеточный домен CD16 прикрепляли к трансмембране и внутриклеточной последовательности ζ (фиг. 7a). Слитой ген интродуцировали в вирус коровьей оспы, экспрессирующей плазмиду, несущей ген gpt E.coli в качестве селектируемого маркера, и вставленный в геном штамма WR коровьей оспы путем гомологичной рекомбинации и селекции по росту в микрофенольной кислоте (Falkner and Moss, J. Virol. 62:1849 (1988); Boyle and Coupar, Gene 65:125 (1988)).
T-клеточные линии инфицировали рекомбинантами коровьей оспы и измеряли отношение цитоплазматического свободного кальция после сшивки внутриклеточных доменов с антителами. Спектрофотометрические (основной части популяции) и проточноцитометрические (одиночной клетки) измерения осуществляли с клетками, нагруженными красителем Indo-1 (Grynkiewicz et al., J. Biol. Chem. 260: 3440 (1985); Rabinovitch et al., J. Immunol. 137:952 (1986)). Фиг. 7b показывает анализы данных, полученных на клетках Jurkat T-клеточной лейкозной линии, инфицированной рекомбинантами коровьей оспы, экспрессирующими слитой белок CD16: ζ. Сшивка химер воспроизводимо увеличивала внутриклеточный кальций, тогда как подобная обработка клеток, экспрессирующих нехимерный CD16, незначительно влияла на его концентрацию или была неэффективной. Когда химеру экспрессировали в мутантных клеточных линиях, не имеющих антигенного рецептора, либо в REX33A (Breitmeyer, et al., J. lmmunol. 138:726 (1987); Sancho et al. , L. Biol. Chem. 264:20760 (1989)), или в клетках Jurkat мутанта JRT3.T3.5 (Weiss et al., J. lmmunol. 135:123 (1984)), то наблюдали сильный ответ на сшивку CD16 с антителом. Аналогичные данные были получены для мутантной клеточной линии REX20A (Breitmeyer et al., см. выше, 1987; Blumberg et al., J. Biol. Chem. 265:14036 (1990)) и в этой лаборатории обнаружили негативный мутант CD3/Ti клеточной линии Jurkat. Инфицирование рекомбинантами, экспрессирующими CD16: ζ, не восстанавливает ответ к антителу анти-CD3, показывая, что слитой белок не действует при сохранении внутриклеточных цепей CD3-комплекса.
Чтобы оценить способность химер к переориентированию клеточноопосредованного иммунитета, цитотоксические T-лимфоциты инфицировали рекомбинантами коровьей оспы, экспрессирующими CD16-химеры, и использовали для специфического лизиса гибридочных клеток, экспрессирующих мембранносвязанные антитела анти-CD16. Этот анализ является продолжением анализа гибридомной цитотоксичности, первоначально разработанного для изучения эффекторных механизмов клеток, несущих Fc-рецепторы (Graziano and Fanger, J. Immunol. 138:945, (1987); Graziano and Fanger, J. lmmunol. 139:35-36 (1987); Shen et al., Mol. lmmunol. 26:959 (1989); Fanger et al., Immunol. Today 10:92 (1989)). Фиг. 8b показывает, что экспрессия CD16: ζ в цитотоксических T-лимфоцитах вооружает ЦТЛ для уничтожения 3G8-гибридомных клеток (анти-CD16; Fleit et al., Proc. Natl. Acad. Sci. USA 79:3275 (1982), тогда как ЦТЛ, экспрессирующие форму CD16, связанную с фосфатидилинозитолом, неактивны. ЦТЛ, вооруженные CD16: ζ, также не убивают гибридомные клетки, экспрессирующие несоответствующее антитело.
Для идентификации минимальных ζ- последовательностей, необходимых для цитолиза, готовили ряд делетированных мутантов, в которых удаления последовательно осуществляли с карбоксильного конца ζ- внутриклеточного домена (SEQ ID NO: 44). Самый большой внутриклеточный домен zeta можно было бы отщепить с маленькой последовательностью для реализации цитолитического потенциала; полной длины химера CD16: ζ была практически равна по эффективности химере, делетированной по остатку 65, CD16 ζ Асп66* (фиг. 8b). Существенное снижение цитотоксичности наблюдали при делеции в ζ остатка 59 (химера CD16: ζ Глу60*), а дополнительная делеция по остатку 50 приводила к незначительному уменьшению активности. Однако полной потери активности не наблюдали, даже когда внутриклеточный домен редуцировали на три остатка трансмембранного якоря (фиг. 8d).
Поскольку ζ представляет собой димер, связанный дисульфидной связью, единственным объяснением сохранения цитолитической активности было то, что эндогенный ζ образовал гетеродимеры с делетированной ζ- химерой, восстанавливая таким образом активность. Чтобы проверить эту идею, остатки 11 и 15 ζ-цепи, после Асп и Цис заменили, соответственно, на Гли (Цис11Гли/Асп15Гли) и затем провели иммунопреципитацию. Приблизительно 2·106 CVI-клеток вносили в течение одного часа в бессывороточную DME-среду, содержащую рекомбинант коровьей оспы, при множественности инфицирования (moi) не менее десяти. Через шесть-восемь часов после инфицирования клетки отделили от поверхности чашек с помощью PBS/1 мМ ЭДТА и поверхностно метили 0,2 мКи 125I на 2 · 106 клеток, используя лактопероксидазу и H2O2 по методу Clark и Einfield (Типирование лейкоцитов II, pp. 155-167, Springer-Verlag, NY, 1986). Меченые клетки собирали центрифугированием и лизировали в 1% NP-40, 0,1% ДСН, 0,15 М NaCl, 0,05 трис-буфере, pH 8,0, 5 мМ MgCl2 5 мМ KCl, 0,2 М иодацетамиде и 1 мМ PMSF. Ядра удаляли центрифугированием, а CD16-белки иммунопреципитировали антителом 3G8 (Flrit et al., см. выше, 1982; Medarex) и инти-мышиным IgG в агарозе (Cappel, Durham, NC). Образцы подвергали электрофорезу в 8% полиакриламидном ДСН-геле в невосстанавливающих условиях или в 10% геле в восстанавливающих условиях. Эти иммунопреципитаты подтвердили, что химера CD16: ζ Цис11Гли/Асп15Гли не ассоциирует в связанные дисульфидными связями димерные структуры.
Тестировали также цитолитическую активность мутантных рецепторов. Мутирововшая химера, делетированная по остатку 65 (CD16: ζ Цис11Гли/Асп15Гли/Асп66, ), была в зависимости от условий анализа в два-восемь раз менее активна в цитолитическом анализе, чем сравниваемая немутировавшая химера (CD16: ζ Асп66*), которая была обычно в пределах коэффициента два или не отличалась по активности от CD16: ζ (фиг. 9b). Это снижение активности мутантных химер сопоставимо с наблюдавшимся снижением для CD4-химер аналогичной структуры (см. выше) и скорее всего определяется низкой эффективностью ζ-мономеров, по сравнению с димерами. В противоположность этому Асп Цис-мутированная химера, делетированная по остатку 59, не имела цитолитической активности (фиг. 9b), подтверждая гипотезу о том, что ассоциация с другими цепями, опосредованными трансмембранными остатками Цис и/или Асп, ответственна за слабую персистенцию цитолитической активности при делециях более аминоконцевых остатков, чем остаток 65.
Проточноцитометрическое изучение показало, что делеционные мутанты, не имеющие трансмембранных остатков
Асп и Цис, могли бы все же способствовать увеличению свободных ионов внутриклеточного кальция как ответ на сшивку антитела в мутантной TCR-линии клеток Jurkat (фиг. 9d). Аналогичные результаты получили для химер, экспрессированных в родительскoй Jurkat-линии. Для случая CD16: ζ Цис11Гли/Асп15Гли/Глу60* эти данные свидетельствуют, что способность опосредовать кальциевую реакцию можно мутационно отделить от способности поддерживать цитолиз.
Чтобы окончательно исключить возможный вклад ζ- трансмембранных остатков, трансмембранные и первые 17 цитоплазматических остатков заменили последовательностями, кодирующими пронизывающую мембрану и, соответственно, первые 14 или первые 17 цитоплазматических остатков антргенов CD15 или CD7 (фиг. 9a). Полученные трираздельные слитые белки CD16:5: ζ (48-65) и CD16:7: ζ (48-65) не образовывали димеров, связанных дисульфидными связями, как это делают более простые химеры CD16: ζ, поскольку они не имели цистеинового остатка в ζ- трансмембранном домене. Обе трираздельные химеры были способны мобилизовать кальций в Jurkat- и TCR-негативной клеточных линиях (фиг. 9d) и для закрепления цитолитического ответа в ЦТЛ (фиг. 9c, но данные не показаны). Однако отсечение ζ- части по остатку 59 в химере CD16:7: ζ (48-59) аннулирует способность трираздельного слитого белка управлять кальциевым ответом в TCR-позитивных или негативных Jurkat-клетках или цитолизом в зрелых ЦТЛ (фиг. 9c и 9d без представления данных).
Для изучения вклада отдельных остатков в рамках мотива из 18 остатков мы получили ряд мутантных вариантов с помощью сайт-направленного мутагенена и оценили их способность опосредовать управляемое рецептором уничтожение в условиях низкой (фиг. 10a и 10d) или высокой (фиг. 10b и 10e) экспрессии химерного рецептора. Фиг. 10a-f показываeт, что в то время как ряд относительно консервативных замен (например, замещение кислотных остатков на их соответствующие амиды, или тирозина на фенилаланин), которые охватывают остатки от 59 до 63, приводили к умеренному компромиссу цитолитической эффективности, в целом эти варианты сохранили способность мобилизовать кальций. Однако совокупно эти остатки включают в себя важный субмотив, поскольку их делетирование ликвидирует цитолитическую активность. Конверсия Тир 62 либо в Фен, либо в Сер исключает и цитотоксический, и кальциевый ответ. На аминоконцевом участке из 18 остатков замена Тир 51 на Фен упраздняет и мобилизацию кальция, и цитолитическую активность, тогда как замена Лей на Сер в положении 50 исключает кальциевый ответ и лишь частично ослабляет цитолиз. Без существования связи с отдельными гипотезами сомнительно, чтобы неспособность Лей50Сер-мутанта мобилизовать кальций в короткие сроки проточноцитометрического анализа не полностью отражала его способность опосредовать существенное увеличение ионов свободного внутриклеточного кальция при удлинении времени цитолитического анализа. Однако для некоторых цитолитических T-клеточных линий сообщали о цитолитической активности, нечувствительной к кальцию, и возможно, что подобный феномен, подчеркивающий результаты, представленные выше, не был исключен. Замещение Асн48 на Сер частично ослабляет цитотоксичность в некоторых экспериментах, тогда как в других проявляет незначительный эффект.
Чтобы исследовать потенциальную роль повторенных элементов последовательности, внутриклеточный ζ-домен разделили на три сегмента, охватывающего остатки от 33 до 65, от 71 до 104 и от 104 до 138. Каждый из этих сегментов присоединили к CD16:Cd7-химере посредством Mlul-сайта, интродуцированного в дистальный участок заякоренных в основную мембрану последовательностей внутриклеточного домена CD7 (см. ниже; фиг. 11f). Сравнение цитолитической эффективности этих трех элементов показало, что они были фактически эквипотентными (фиг. 11b). Сравниваемая последовательность показывает, что второй мотив несет одиннадцать остатков между тирозинами, а первый и третий мотивы несут десять остатков.
Несмотря на то, что точное объяснение процесса T-клеточной активации не сделано, очевидно, что агрегация антигенного рецептора или рецепторных химер, которые несут ζ- внутриклеточные последовательности, запускает мобилизацию кальция, высвобождение цитокина и гранулы и появление маркеров активации клеточной поверхности. Активный сайт ζ-цепи в виде короткой линейной пептидной последовательности, по-видимому, очень маленький, чтобы иметь собственную энзиматическую активность, вероятно взаимодействует с одним или, самое большее, с несколькими белками, чтобы опосредовать клеточную активацию. Очевидно также, что мобилизация свободного кальция сама по себе недостаточна для клеточной активации, поскольку способность опосредовать цитолиз может быть мутационно отделена от способности опосредовать накопление кальция.
Как показано выше, присоединение 18 остатков, от внутриклеточного домена ζ- цепи до трансмембраны и внутриклеточного домена из двух несвязанных белков, делают возможным получение химер для перенацеливания цитолитической активности на клетки-мишени, которые связываются с внеклеточной частью слитых белков. Хотя химеры, несущие мотив из 18 остатков примерно в 8 раз менее активны, чем химеры, основанные на ζ полной длины, снижение активности можно отнести за счет потери трансмембранного взаимодействия, которое позволяет обычно ζ дикого типа образовывать димеры, связанные дисульфидной связью. То есть ζ-делеционные конструкты, которые имеют тот же карбокси-конец, что и мотив, и не имеют трансмембранных остатков Цис и Асп, обычно показывают несколько меньшую активность, чем химеры, несущие мотив из 18 остатков.
Цитолитический компетентный элемент, на котором мы сосредоточили внимание, имеет два тирозина и не имеет серинов или треонинов, ограничивая возможность вклада фосфорилирования в активность. Мутация по любому из тирозинов разрушает активность, однако, хотя предварительные эксперименты не указывают на важность тирозинового фосфорилирования после сшивки антигенов поверхности химеры, вероятность участия такого фосфорилирования на низком уровне исключить нельзя. В дополнение к эффектам, отмеченным по двум тирозиновым остаткам, ряд аминокислотных замен по амино- и карбокси-концу мотива ослаблял активность в условиях низкой рецепторной плотности.
Последовательности, подобные мотиву, активирующему ζ, можно найти в цитоплазматических доменах нескольких других трансмембранных белков, в том числе δ- и γ- молекулах CD3, ассоциированных с поверхностью IgM белках mb1 и B29, и β- γ- цепях рецептора Fc ε RI, обладающего высоким сродством IgE (Reth, Nature 338:383 (1989)). Хотя функция этих последовательностей не определена, при эффективной экспрессии каждая может автономно активировать T-клетки, и такую активность можно объяснить остаточным TCR-ответом, наблюдаемому в zeta-негативной мутантной клеточной линии (Sussman et al., Cell 52:85 (1988)).
Сама по себе ζ несет три такие последовательности, распределенные примерно одинаково, и приближенный трехсекционный внутриклеточный домен показывает, что каждая обладает способностью инициировать цитолитический ответ. Изоформа η, сращивающая ζ, не имеет половины карбокси-конца третьего мотива. Поскольку удаление половины карбокси-конца первого мотива ликвидирует активность, представляется вероятный, что большую часть биологической эффективности можно отнести на счет первых двух мотивов. Хотя при разных измерениях η по активности равно ζ в стимуляции высвобождения цитокина, опосредованного антигеном (Bauer et al. , Proc. Natl. Acad. Sci. USA 88:3842 (1991)), или в перенацеливании цитолизма (см. выше), η не фосфорилируется при ответе на рецепторную стимуляцию (Bauer et al., смотри выше, 1991). Следовательно, для фосфорилирования необходимо либо присутствие всех трех мотивов, или третьего мотива, представляющего благоприятный субстрат для неидентифицированной тирозинкиназы.
ПРИМЕР IX. Трансдукция цитолитического сигнала человеческим Fc-рецептором
Чтобы оценить действие разных субтипов человеческого Fc-рецептора, создали химерные молекулы, в которых внеклеточные домены человеческих CD4-, CD5- или CD16-антигенов соединили с трансмембранной и с субтипами внутриклеточных доменов FcRII γ A, A, B1, B2 и C (номенклатура Ravetch и Kinet, Ann. Rev. Immunol. 9:457 (1991)). Так, последовательности кДНК, соответствующие трансмембранному и цитоплазматическому доменам, ранее описанные как A, B1- и B2-изоформы FcRII, амплифицировали из предсуществующего клона PC23 и из библиотеки кДНК небной миндалины (сконструированной по стандартным методикам), используя следующие синтетические олигонуклеотидные праймеры:
CCC GGA TCC CAG CAT GGG CAG CTC TT (SEQ ID NO:18; FcRII A прямой);
CGC GGG GCG CCC GCT TTA GTT ATT ACT GTT GAC ATG GTC QTT (SEQ ID NO:19; FcRII A обратный);
GCG GGG GGA TCC CAC TGT CAA AGC TCC CAG CTC TTC ACC G (SEQ ID NO: 20 FcRII B1 и FcRII B2 обратный);
GCG GGG GCG GCC GCC TAA ATA CGG TTC TGG TC (SEQ ID NO:21: FcRII B1 и FcRII B2 обратный).
Эти праймеры содержат сайты расщепления, соответственно, для ферментов BamHI и NotI, с отступом на 6 остатков от 5'-конца. NotI-сайт расположен непосредственно за антисмысловым стоп-кодоном, либо CTA, либо TTA. Все праймеры содержат 18 или более остатков, комплементарных 5'- и 3'-концам соответствующих фрагментов. Фрагмент кДНК, соответствующий цитоплазматическому домену FcRIIγC, который отличается от IIA-изоформы только по одному аминокислотному остатку (L вместо P по остатку 268), получали с помощью сайт-направленного мутагенеза с помощью ПЦР, использующей праймеры с перекрывающимися последовательностями:
TCA GAA ACA GAC AAC CTG AAG AAA CCA ACA A (SEQ ID NO:22) и
TTG TTG GTT TCT TCA GGT TGT GTC TTT CTG A (SEQ ID NO:23).
Фрагменты ПЦР встраивали в экспрессионные векторы вируса коровьей оспы, которые содержали, соответственно, внеклеточные домены CD16 или CD4, и затем встраивали в коровью оспу дикого типа путем рекомбинации по локусу тимидинкиназы, применяя селекцию для коинтеграции gpt E.coli, чтобы облегчить идентификацию необходимых рекомбинантов. Идентичность всех изоформ (показано на фиг. 12) подтвердили дидезоксисеквенированием.
Образование химерных рецепторных белков подтвердили также с помощью исследований по иммунопреципитации. Примерно 107 клеток JRT3.T3.5 использовали для инфицирования в течение одного часа рекомбинанта коровьей оспы, содержащейся в бессывороточной IMDM-среде, при множественности инфицирования не менее десяти. Спустя двенадцать часов после инфицирования клетки собирали и поверхностно метили 0,5 мКи 125I на 107 клеток, применяя глюкозопероксидаза/глюкозооксидазный метод (Clark и Einfeld, см. выше). Меченые клетки собирали центрифугированием и лизировали 1% NP-40, 0,1 мМ MgCl2, 5 мМ KCl, 0,2 M иодацетамидом и 1 мМ PMSF. Ядра удаляли центрифугированием, a CD16-слитые белки иммунопреципитировали антителом 4G8 и азарозным анти-мышиным IgG. Образцы подвергали экстрофорезу в восстанавливающих условиях. Все иммунопреципитированные молекулы химерного рецептора имели ожидаемые молекулярные массы.
Чтобы проверить способность химерных рецепторов опосредовать увеличение в цитоплазме концентрации ионов свободного кальция, использовали рекомбинантные вирусы для инфицирования TCR-мутанта Jurkat-клеток линии JRT3.T3.5 (как описано выше), а цитоплазматический свободный кальций измеряли в клетках (как описано выше) после сшивки рецептора внеклеточного домена с моноклональным антителом 3G8 или Leu-3A (как описано выше). Эти эксперименты обнаружили, что внутриклеточные домены FcR γ II A и C способны опосредовать увеличение концентрации ионов свободного кальция в цитоплазме после сшивки внеклеточных доменов, тогда как внутриклеточные домены FcR γ II B1 и B2 при этих же условиях оказались неактивны (фиг. 13a и 13b). CD4-, CD5- и CD16-гибриды FcR γ II A практически поровну участвовали в усилении кальциевого ответа (фиг. 13a-b). Другие клеточные линии, происходящие от моноцитов и лимфоцитов, были способны отвечать на сигнал, инициированный сшивкой внеклеточных доменов.
Чтобы прозондировать вовлеченность разных внутриклеточных доменов FcR γ II в цитолиз, человеческие цитотоксические T-лимфоциты (ЦТЛ) инфицировали рекомбинантами коровьей оспы, экспрессирующими химеры CD16:FcR γ II A, B1, B2 и C. Затем инфицированные клетки сокультивировали с нагруженными 51Cr, гибридомными клетками (например, клетками 3G8 10-2), которые экспрессировали клеточной поверхностью антитело к CD16. В этом анализе цитотоксические T-лимфоциты, несущие CD16-химеру, уничтожали гибридомные клетки-мишени (позволяя высвобождать свободный 51Cr) при условии, что внеклеточный домен CD16-химеры присоединяли к внутриклеточному сегменту, способному активироваться по программе лимфоцитного эффектора; этот опыт по цитолизу подробно описан ниже. Фиг. 14a показывает, что ЦТЛ, вооруженные CD16:FcR γ II A и С, но не FcR γ II B1 или B2, обладают способностью лизировать клетки-мишени, экспрессирующие клеточной поверхностью антитело анти-CD16.
Чтобы исключить вероятность того, что специфичность цитолиза некоторым образом определялась взаимодействием с CD16-половиной, проводили эксперименты по цитолизу, в которых внутриклеточные домены FcRII прикрепляли к внеклеточному домену CD4. В этом случае клетками-мишенями были клетки HeLa, экспрессирующие ВИЧ-оболочечные белки gp120/41 (в частности, клетки HeLa инфицировали вектором vPE16 коровьей оспы (поставляемой Национальным Институтом аллергии и инфекционных болезней. Отдел СПИД, Bethesda, MD). Как и в CD16-системе, клетки-мишени, экспрессирующие ВИЧ-оболочку, были восприимчивы к лизису T-клетками, экспрессирующими химеру CD41:FcR γ II A, но не химеру FcR γ II B1 или B2 (фиг. 14b).
Внутриклеточные домены FcR γ II A не обладают последовательностью, достаточно гомологичной любому другому белку, в том числе членам распространенного семейства FcR γ /TCR ε. Чтобы определить последовательность элементов, ответственных за индукцию цитолиза, полученные делеции по 5'- и 3'-кодирующим последовательностям внутриклеточного домена (описано ниже и показано на фиг. 15a) оценивали в анализах по эффективности мобилизации кальция и цитолиза (как описано выше). Эксперименты, в которых удаляли амино-концевую часть внутриклеточного домена, трансмембранный домен FeR γ II заменяли на трансмембранный домен неродственного антигена CD7, чтобы исключить возможность содействия взаимодействию, опосредованного доменом, пронизывающим мембрану.
Фиг. 15b и 15c показывают, что удаление 14 карбокси-концевых остатков, включая тирозин 298, приводит к полной утрате цитолитической производительности и ощутимому снижению потенциала мобилизации кальция. Добавочная делеция сразу перед тирозином 282 давала идентичный фенотип (фиг. 15b и 15c). Делеция от N-конца внутриклеточного домена до остатка 268 не имела заметного эффекта ни на кальциевый профиль, ни на цитолитическую потенцию, тогда как делеция по остатку 275 отчетливо уменьшала высвобождение кальция, но мало влияла на цитолиз (фиг. 15d и 15e). Добавочная делеция по остатку 282 давала FcR γ-хвосты, которые теряли способность или мобилизовать кальций или запускать цитолиз (фиг. 15d и 15e). "Активный элемент", определенный с помощью этих приблизительных измерений, является относительно большим (36 аминокислот) и содержит два тирозина, разделенных 16 остатками.
ПРИМЕР Х. Цитолиз, обусловленный лимфоцитами, несущими химерные CD4-рецепторы, которые не содействуют инфицированию
Как обсуждалось выше, можно создать эффекторные молекулы, которые перенацеливают цитолитическую активность цитотоксических T-лимфоцитов способом, независимым от МНС. Например, химера, составленная из внеклеточного домена CD4, слитого ζ-цепью в клоне WH3 человеческих ЦТЛ, убивает именно клетки-мишени, обнаруживающие на своей поверхности оболочечный гликопротеин gp120 ВИЧ-1. Поскольку внеклеточный домен молекулы CD4 придает восприимчивость к ВИЧ-инфекции, то вооруженные цитотoксические T-лимфоциты могут становиться мишенями для вируса, приводя к снижению их потенции (Dalgleish et al., Nature 312:767 (1984); Klatzman et al., Nature 312:767 (1984)). Чтобы предотвратить такой исход, на основе CD4 создали химерные эффекторные молекулы, которые являются эффективными именно для ВИЧ-инфицированных клеток, убиваемых опосредованной клеткой, но которые не придают восприимчивость к инфицированию ВИЧ.
Трираздельный слитой белок создали путем генетического соединения внеклеточного домена CD4 (фиг. 23) с шарниром, вторым и третьим константными доменами тяжелой цепи IgG1 человека (Zettlmeissl et al., DNA Cell Biol. 9:347 (1990)) (фиг. 25), который соединили в данном случае с частью первого трансмембранного экзона человеческого IgG1, связанного с мембраной, находящегося за участком человеческого CD7-антигена, состоящего из последовательностей, между основанием Ig-подобного домена и стоп-трансферной последовательностью последующего трансмембранного домена (Aruffo and Seed, EMBO J. 6:3313 (1987)) (фиг. 26). Первичная аминокислотная последовательность внеклеточной составляющей CD7-сегмента состоит из пролин-богатой области, напоминающей стебельковоподобную структуру, которую Ig-подобный домен выставляет вне клеточной поверхности (Aruffo and Seed, EMBO J. 6:3313 (1987)) (фиг. 26). Рекомбинантные вирусы коровьей оспы готовили для экспрессии данной и родственной химер; как описано выше. В частности, рекомбинантные вирусы коровьей оспы получали путем гомологичной рекомбинации в клетках CV-1. Выполняли не менее двух циклов визуализации бляшек с OK14 или Leu3a с последующим выделением бляшек, которое проводили для каждого штамма перед получением штаммов клеток CV-1 с высоким титром.
Трираздельная химера (CD4(D1-D4):Ig:CD7) (фиг. 20, молекула "A") показала эффективную клеточно-поверхностную экспрессию и была испытана на способность действовать в качестве ВИЧ-рецептора в анализе образования синцития на основе осповакцины (Lifson et al., Nature 323:725 (1986)); Ashorn et al., J. Virol. 64:2149 (1990). Клетки HeLa, инфицированные рекомбинантным вирусом коровьей оспы (vPE16), кодирующим оболочечный гликопротеин ВИЧ-1 (Earl et al. , J. Virol. 64:2448 (1990)), сокультивировали с клетками HeLa, инфицированными CD4 либо CD4: ζ или CD4(D1-D4):Ig:CD7. Клетки HeLa (ATCC, Rockville, MD), из 6 см чашек при 50% слиянии, инфицировали в бессывороточной среде в течение 1 часа при приблизительной множественности 10 (MOI). Клетки дополнительно инкубировали 5-6 часов в полной среде и затем отслаивали фосфатно-солевым буферным раствором (PBS), содержащем 1 мМ ЭДТА. Клетки, экспрессирующие оболочку и CD4-химеру, смешивали в соотношении 1:1 и переносили в 6 см чашки с полной средой. Синцитий оценивали на 6-8 часов после сокультивирования и фотографировали.
Сокультивирование CD4 и vPE16 приводило к образованию легко обнаруживаемых многоядерных гигантских клеток. Кроме того, химера, содержащая внеклеточный домен CD4, слитый с ζ- цепью TCR (фиг. 27) (CD4 ζ), была способна опосредовать образование синцития, тогда как клетки, экспрессирующие CD4(D1-D4): Ig:CD7, не давали сигнала клеточного слияния. Мы также проверили конструкт, экспрессирующий только первый и второй домены CD4 (фиг. 24)), CD4(D10D4): Ig: CD7 (фиг. 20) молекула "B"), поскольку в другом контексте аминоконцы двух доменов CD4, как показано, были необходимы для создания инфекционности с помощью ВИЧ (Landau et al., Nature 334:159 (1988)). Эта молекула оказалась также нечувствительной к образованию ВИЧ-индуцированного синцития. Изучение связывания с растворимым gp120, меченым 125I, установило, что и CD4(D1-D4):Ig:CD7, и CD4(D1,D2):Ig:CD7 обладали исключительным сродством к gp120. Далее мы выясняли, действительно ли химерные молекулы, которые сопротивлялись образованию синцития, будут способны перенацелить уничтожение клетки, если их наделяли пусковой составляющей, как описано выше. Мы слили внутриклеточный домен ζ с 3'-концом CD4(D1-D4):Ig:CD7 и CD4(D1,D2):Ig:CD7 и приготовили соответствующие рекомбинантные вирусы коровьей оспы. Эти конструкты, CD4(D1-D4):Ig:CD7: и CD4(D1,D2):Ig:CD7:ζ (фиг.20, молекулы "C" и "D"), экспрессировали в человеческий клон WH3 цитотоксических лимфоцитов, и проверили их на способность мишенировать и уничтожать клетки HeLa, экспрессирующие поверхностный оболочечный гликопротеин ВИЧ (применяя способы, описанные выше). Фиг. 21 показывает, что внутриклеточный домен ζ, слитый с CD4(D1-D4): Ig: CD7 или с CD4(D1,D2):Ig:CD7, может придавать способность к уничтожению; конструкты, не имеющие ζ-цепи, были неспособны опосредовать эту активность. CD4: ζ, позитивный контроль, опосредовал несколько более эффективную цитотоксичность, а CD4(D1, D2):Ig:CD7: ζ- несколько меньшую эффективную цитотоксичность, чем CD4(D1-D4):Ig:CD7: ζ (фиг.21). Тем не менее очевидно, что и химера CD4(D1-D4):Ig:CD7: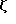 , и химера CD4(D1,D2):Ig:CD7:ζ обладают способностью опосредовать специфическое уничтожение клеток, экспрессирующих ВИЧ-оболочечные белки на своей поверхности. Четырехраздельные химеры были стабильно неспособны опосредовать образование синцития в опыте с коровьей оспой. Мы также продемонстрировали, что единственный вид мотива ζ, показанный на фиг. 11а, является достаточным для придания цитолитической активности химере CD4(D1-D4).
, и химера CD4(D1,D2):Ig:CD7:ζ обладают способностью опосредовать специфическое уничтожение клеток, экспрессирующих ВИЧ-оболочечные белки на своей поверхности. Четырехраздельные химеры были стабильно неспособны опосредовать образование синцития в опыте с коровьей оспой. Мы также продемонстрировали, что единственный вид мотива ζ, показанный на фиг. 11а, является достаточным для придания цитолитической активности химере CD4(D1-D4).
Эксперименты по радиоиммунопреципитации установили, что сливающиеся молекулы были преимущественно, если не всегда, димерами. В этих экспериментах агарозные гранулы белка A использовали для иммунопреципитации солюбилизированного экстракта метаболически меченых клеток HeLa, инфицированных рекомбинантной осповакциной экспрессирующей химеры CD4(D1-D4):Ig:CD7:ζ и CD4(D1, D2):Ig:CD7ζ. Иммунопреципитированный материал фракционировали электрофорезом в полиакриламидном геле в восстанавливающих и в невосстанавливающих условиях. В частности, приблизительно 5 · 106 клеток HeLa-S3 инфицировали vEP16, как описано выше, соответствующего штамма вируса коровьей оспы. Клетки метаболически метили 200 мКи/мл Tran35 S-меткой (ICN Радиохимикаты, lrvine, CA) в течение 6-8 часов в среде, дефицитной по цистеину и метионину, и отделяли с помощью PBS, содержащем 1 мМ ЭДТА. Клетки последовательно осадили и лизировали в 150 мМ NaCl, 50 мМ трис-буфере pH 7,5 5 мМ ЭДТА, 0,5% NP-40, 0,1% ДСН, 5 мМ ЭДТА, 1 мМ PMSF. После удаления ядер центрифугированием одну пятую часть каждого клеточного экстракта адсорбировали на отмытых гранулах агарозы, конъюгированных с белком A, в течение 2 часов при 4oC. Гранулы последовательно отмывали PBS, содержащем 1% NP-40, и элюировали в буферный образец, содержащий ДСН при наличии или отсутствии меркаптоэтанола. Результаты этих экспериментов свидетельствуют, что большинство иммунопреципитированных химер CD4(D1-D4):Ig:CD7:ζ и CD4(D1,D2):Ig:CD7:ζ, мигрировало в невосстанавливающих условиях как димеры с ожидаемой молекулярной массой.
Чтобы непосредственно оценить способность клеток, экспрессирующих слитые молекулы CD4, обеспечивать ВИЧ-инфекцию, мы провели долговременные исследования инфекционности на трансфектантах, экспрессирующих CD4(D1-D4):Ig:CD7 и CD4(D1, D2):Ig:CD7. Стабильные трансфектанты CD4(D1-D2):Ig:CD7 и CD4(D1,D2): Ig: CD7 и CD4 были получены в сублинии из 293 клеток легко трансфецируемой клеточной линии, выделенной из эмбриональной человеческой почки. Химерные молекулы субклонировали в двунаправленные векторы, в которых ген гигромицина В встраивали под промотор тимидинкиназы вируса простого герпеса. 60-70% сливных клеток из 10 см чашки трансфецировали 10 мкг ДНК этой плазмиды путем кальций-фосфатной концеприципитации. Перед трансфекцией плазмиды линеаризировали по уникальному сайту SfiI, а концы тупили с помощью Т4 ДНК-полимеразы. После 24 часов трансфекции клетки четырежды отслаивали и после 48 часов трансфекции подвергали селекции с гигромицином В (Sigma, St. Louis, Mo) при концентрации 400 мкг/мл. Каждые 3-4 дня клетки помещали в свежую среду, содержащую гигромицин.
Отбирали увеличенные в объеме резистентные колонии, и их экспрессию испытывали путем непрямой иммунофлуоресценции, после проточной цитометрии (Coulter, Hialeah, FL), используя анти-Fc IgG человека, конъюгированного с флуоресцеином (Organon Teknika, West Chester, PA), или Q4120, антитело, взаимодействующее с анти-CD (Sigma). Из каждого конструкта для анализа отобрали два независимых клона, сопоставимые по уровню CD4 на поверхности клетки с другими клеточными линиями. Фиг. 22 показывает, что при последующем выдерживании с ВИЧ p24 детектировали в культурах CD4-стабильных трансфектантов уже через 3 дня после инфицирования. Наличие полиядерных гигантских клеток и характерная шарообразность в этих культурах были явными уже через 5 дней после инфицирования. После 32 дней культивирования в родительской клеточной линии или в любом из двух независимо полученных изолятов, трансфецированных CD4(D1-D4): Ig: CD7 и CD4(D1,D2):Ig:CD7 (фиг. 22), не отмечено значительных уровней p24 или явного образования многоядерных гигантских клеток.
По окончании исследований по инфекционности клетки анализировали на экспрессию CD4 клеточной поверхностью. Плотность эпитопа поверхностного CD4 была существенно понижена в инфицированных культурах, экспрессирующих CD4, постоянная при вирусной низкой модуляции, и была не затронута в культурах, экспрессирующих CD4(D1-D4): Ig: CD7 и CD4(D1,D2):Ig:CD7. Эти эксперименты свидетельствуют о возможности создания химерных молекул, несущих два апикальных домена CD4, которые при слиянии с ζ-цепью рецептора T-клетки обладают способностью к мишенированим и уничтожению ВИЧ-инфицированных клеток, но которые не способствуют опосредованной CD4 ВИЧ-инфекции.
Дополнительные эксперименты подтверждают, что имеется физическое расстояние между внеклеточным доменом молекулы CD4 и липидным бислоем, что придает способность сопротивляться ВИЧ-инфекции. В первом эксперименте мы сконструировали химерную молекулу, несущую делецию стебля CD4 и трансмембранного домена; эта делеция удаляет пролин-богатую область трансмембранной части CD7. Когда этот домен слили с внеклеточным доменом CD4, он сохранил свою способность к эффективному заякориванию внеклеточного домена молекулы CD4, что установлено измерением экспрессии клеточной поверхностью молекулы CD4 (как описано выше). Однако потенциал к сопротивлению образования синцития, индуцированного гликопротеином ВИЧ-оболочки, был утрачен. Следовательно, делеция пропил-богатой области молекулы CD7, области, подходящей для образования α-спиральной структуры, эффективно уменьшала расстояние между внеклеточным доменом CD4 и липидным бислоем и аннулировала способность химеры сопротивляться образованию синцития.
Во втором эксперименте мы продемонстрировали, что способность сопротивляться ВИЧ-индуцированному образованию синцития может быть придана химере CD4/CD5, которую раньше рассматривали в качестве трансмембранного якоря для внеклеточного домена CD4, но которая была неспособна сопротивляться ВИЧ-индуцированному образованию синцития. В этом эксперименте шарнир, CH2- и CH3-домены тяжелой цепи человеческого IgG1 встраивали в молекулу CD4/CD5; полученная химера сопротивлялась образованию синцития, вновь подтверждая, что расстояние, предоставленное иммуноглобулиновыми доменами, является достаточным для придания резистентности к ВИЧ-индуцированному образованию синцития.
В третьем эксперименте CD4-домен удлинял изменяющиеся расстояния от клеточной мембраны, используя альфа-спирали разной длины. В частности, были сконструированы синтетические олигонуклеотиды, презентирующие повторенные альфа-спиральные мотивы остатков лизина и глутаминовой кислоты, фланкированные двумя аланиновыми остатками (см. фиг. 28 для первичных последовательностей нуклеиновой кислоты и аминокислот). В прежних исследованиях такие аминокислотные последовательности были обнаружены с высокой частотой в альфа-спиралях, подтверждая, что такие повторенные мотивы будут заимствовать конформацию альфа-спирали и что замена таких альфа-спиралей между трансмембранным доменом и внеклеточными доменами CD4 будут выталкивать CD4 из клеточной мембраны. Изменяя длину альфа-спирального сегмента, вычисление прогнозируемого расстояния, необходимого для сопротивления проникновения ВИЧ, осуществляли на основе известных значений удлинения и нарастания альфа-спирали. Эти результаты представлены в таблице 1.
В таблице 1 "CD4" соответствует CD4(D1-D4), за исключением особо отмеченных случаев; "H", "CH2" и "CH3" соответствуют шарниру, областям CH2 и CH3 тяжелой цепи человеческого IgG1, соответственно; "CD7tm и stk" соответствуют областям трансмембраны CD7 и стебля; "CD7tm (длинный вариант)" и "CD7tm (короткий вариант)" соответствуют трансмембранной области CB7 и трансмембранной области CD7, делетированной по пролин-богатому домену (как обсуждалось выше); "CD5tm" соответствует трансмембранной области CD5 и "CD34tm" соответствует трансмембранной области CD34. Для входов J-L длина альфа-спиральной области указана в ангстремах; эти значения основаны на том факте, что на виток альфа-спирали приходится 3,6 остатков, что соответствует 5,4 ангстрем (или 1,5 ангстрем на остаток). В соответствии с этим 16 остатков альфа-спирали будут выталкивать внеклеточный домен CD4 почти на 24 ангстрема. Альфа-спирали длиной 48 и 72 ангстрема были сконструированы в результате конкатемеризации BstY1-фрагмента во фрагмент уникального BamHI-сайта (см. фиг. 28) после селекции клонов с надлежащей ориентацией.
Образование синцития оценивали в опытах сокультивирования с клетками HeLa, экспрессирующими гликопротеин оболочки ВИЧ из конструкта vRE-16 вируса коровьей оспы (см. выше).
Экспрессию thy-1 измеряли следующим образом. На основе ВИЧ-клона hxb.2 сконструировали вектор живого ретровируса. В этом векторе несущественный nef-ген заменили на последовательность, кодирующую rat thy-1, молекулу, эффективно экспрессируемую клеточной поверхностью, которая заякорена в мембрану с помощью фосфатидинозитоловой связи. Вирус, полученный из этого молекулярного клона, обозначенный hxb/thy-1, оказался инфекционным, как следует из его цитопатологических эффектов и образования p24 в культуральных супернатантах инфицированных C8166-клеток (человеческая лейкозная T-клеточная линия CD4). Кроме того, при выдерживании с hxb/thy-1 клетки HeLa, транзиторно трансфецированные CD4, обнаружили сигналы экспрессии thy-1 уже через 18 часов после инфицирования, как-будто ожидали сигнализацию, регулируемую nef-образным способом. Сигналы, закодированные геном nef, обычно относятся к классу вирусных регуляторных белков, которые множественно сплайсированы и не имеют элемента ответа rev. Эти сигналы конститутивно накапливаются в цитоплазме в виде продуктов раннего вирусного гена. Предположили, что thy-1-сигналы регулируются аналогично, то есть встречаются рано в жизненном цикле вируса. Короче говоря, благодаря использованию thy-1-экспрессии в качестве суррогата вирусного проникновения эта система облегчает анализ проникновения ВИЧ. Основанные на CD4 разные химеры были транзиторно трансфецированы в клетки HeLa, используя стандартные ДЭАЭ-декстрановые способы. Трансфецированные клетки выдерживали с hxb/thy-1-вирусом в течение 48 часов после инфицирования и оценивали по thy-1-экспрессии в течение 24-48 часов после инфицирования. В этих результатах, показанных в таблице 1, thy-1-экспрессию измеряли в течение 24 часов после инфицирования, используя коммерческий препарат thy-1-моноклонального антитела (Точный).
Из данных, представленных в таблице 1, мы заключили, что внеклеточные домены CD4 оптимально должны выступать из клеточной мембраны не менее чем на 48 ангстрем и предпочтительно не менее чем на 72 ангстрем, для того чтобы сопротивляться инфицированию ВИЧ-1.
Применяя стратегию, аналогичную общей стратегии описанной выше, можно сконструировать химеры, основанные на оболочечных антителах анти-ВИЧ, которые мишенируют ВИЧ-инфицированные клетки. Примеры таких антител описаны у Gorny et al., Proc. Natl. Acad. Sci. USA 86:1624 (1989) и Marasco et al., J. Clin. Invest. 90:1467 (1992).
ПРИМЕР XI. Использование выступающих молекул CD4 в качестве ловушки для ВИЧ
Как показано выше, клетки, несущие CD4-домены, выступающие из клеточной поверхности, сопротивляются заражению ВИЧ. В соответствии с этим такие CD4-несущие клетки пригодны в качестве ловушки, чтобы связать циркулирующий ВИЧ и снижать титр вирусов у ВИЧ-инфицированных индивидуумов. Предпочтительно, чтобы CD4-домен был представлен в клетке, которая естественным образом проходит через лимфатические узлы; полезные клетки хозяина включают в себя любую клетку, которая играет роль в иммуноклеточном очищении, как и любую другую клетку, естественным образом присутствующую в фолликулах лимфатического узла. Отдельные примеры включают в себя, без ограничения, макрофаги, T-клетки (например, хелперные T-клетки), B-клетки, нейтрофилы, дендритные клетки и фолликулярные дендритные клетки.
Любой CD4-домен, способный связывать ВИЧ, в том числе домены D1-D4 и D1, D2, описанные выше, могут быть использованы в данном способе настоящего изобретения. Для, по крайней мере, некоторых вариантов осуществления (например, вариантов, описанных выше) части химеры внутриклеточного и/или трансмембранного домена могут быть выбраны из любого белка или любой аминокислотной последовательности.
ПРИМЕР XII. Дополнительные пусковые белки T-клеточного рецептора и B-клеточного рецептора
В соответствии с настоящим изобретением из рецепторных белков T-клетки-CD-дельта и T3-гамма, и из рецепторных белков B-клетки - mb1 и B29 можно получить другие внутриклеточные и трансмембранные домены, трансдуцирующие сигнал. Аминокислотные последовательности этих белков показаны на фиг. 16 (CD3-дельта; SEQ ID N: 24), фиг. 17 (T3-гамма; SEQ ID NО: 25), фиг. 18 (mb1; SEQ ID NО: 26) и фиг. 19 (B29; SEQ ID NО: 27). В скобках показаны участки последовательностей, достаточные для трансдукции цитолитического сигнала (и поэтому предпочтительно включенные в химерный рецептор настоящего изобретения). Химерные рецепторы, которые включают в себя эти белковые домены, сконструированы и использованы в терапевтических способах настоящего изобретения, в основном, описанных выше.
ПРИМЕР XIII. Методы экспериментов. Инфицирование осповакциной и радиоиммунопреципитация
Приблизительно 5 · 106 клеток CV1 инфицировали в течение одного часа в бессывороточной DME-среде рекомбинантной осповакциной при множественности инфицирования (moi) не менее десяти (титр измеряли в клетках CV1). После инфицирования клетки помещали в свежую среду и метаболически метили в течение шести часов 200 мКи/мл 35S-метионина плюс цистеин (Tran 35S-метка, ICN; Costo Mesa, CA) в бензметиониновой и безцистеиновой среде DMEM (Gibco; Grand Island, NY). Меченые клетки отделяли с помощью PBS, содержащего 1 мМ ЭДТА, собирали центрифугированием и лизировали в 1% NP-40, 0,1% ДСН, 0,15 NaCl, 0,05 трис-буфере pH 8,0 5 мМ ЭДТА и 1 мМ PMSF. Ядра удаляли центрифугированием и CD4-белки иммунопреципитировали OKT4-антителом и агарозным анти-мышиным IgG (Cappel, Durham, NC). Образцы подвергали ДСН-электрофорезу в 8% полиакриламидном геле в невосстанавливающих (NR) и в восстанавливающих (R) условиях. Перед радиоавтографией гели, содержащие 35sS-меченые образцы импрегнировали En3 Hance (New England Nuclear, Boston, MA). Содействие экспрессии трансмембранной формы CD16, CD16TM измеряли путем сравнения его экспрессии в клетках CV1, однократно инфицированных CD16TM для экспрессии в клетках, коинфицированных вирусами, кодирующими CD16TM и ζ- или γ-химеры. После заражения и инкубации в течение шести часов или больше клетки отслаивали от поверхности чашек с помощью PBS, 1 мМ ЭДТА и экспрессию CD16TM или химер измеряли непрямой иммунофлуоресценцией и проточной цитометрией.
Анализ поступления кальция
Jurkat-клетки сублинии E6 (Weiss et al., J. Immunol. 135:125-128 (1984)) инфицировали рекомбинантными вирусами коровьей оспы в течение одного часа в бессывороточной IMDM-среде при moi 10 и инкубировали от трех до девяти часов в IMDM-среде с 10% FBS. Клетки собирали центрифугированием и ресуспендировали при концентрации 3 · 106 клеток/мл в полной среде, содержащей 1 мМ Indo-1 ацетометоксиэфира (Grynkiewicz et al., J. Biol. Chem. 260:3340-3450 (1985)) (Молекулярные Пробы), и инкубировали при 37oC в течение 45 минут. Клетки, нагруженные Indo-1, осаждали и ресуспендировали при концентрации 1·106/мл в бессывороточной IMDM-среде при комнатной температуре в темноте. Клетки анализировали на наличие свободных ионов кальция путем одновременного измерения фиолетовой и голубой флуоресцентной эмиссии с помощью проточной цитометрии (Rabinovitch et al. , J. Immunol. 137:952-961 (1986)). Чтобы инициировать истечение кальция, к клеточной суспензии добавляют, при концентрации 1 мкг/мл, либо Leu-3A (анти-CD4), конъюгированный с фикоэритрином (PE), а затем 10 мкг/мл неконъюгированного анти-мышиного IgG, козы в нулевой временной точке, или к клеточной суспензии добавляют при концентрации 1 мкг/мл неконъюгированное моноклональное антитело 3G8 (анти-CD16), а затем 10 мкг/мл анти-мышиного IgG козы, Fab'-конъюгированного с фикоэритрином PE, в нулевой временной точке. Гистограммы соотношения фиолетовой/голубой эмиссии получали для популяции PE-позитивных (неинфицированных) клеток, которая обычно представлена 40-80% всех клеток. Рецепторный ответ T-клеточного антигена в неинфицированных клетках запускали с помощью антитела OKT3, без сшивки. Для экспериментов с участием CD16-химерных рецепторов образцы, проявляющие смешение базиса к снижению внутриклеточного кальция (без антитела), исключали из анализов. Гистограмму данных последовательно анализировали путем превращения бинарных данных в ASCII, применяя рукописное (Cooper City, FL) программное обеспечение, после анализа с помощью коллекционных программ FORTRAN. Соотношение эмиссии фиолетовый/голубой, перед добавлением реагентов второго антитела, использовали, чтобы стандартизованное начальное соотношение установить равным единице, а соотношение покоящегося порога установить так, чтобы 10% покоящейся популяции превышала бы порог.
Цитолитический анализ
Человеческую T-клеточную линию WH3, цитолитическая линия ограниченная CD8+CD4-HLA B44, поддерживали в IMDM, 10% человеческой сыворотке со 100 е/мл IL-2 и периодически стимулировали либо неспецифически, облученными (3000 рад) HLA-разрозненными лимфоцитами периферической крови и 1 мкг/мл фитогемагглютинином, или специфически, облученными мононуклеарными клетками, несущими B-44. После одного дня неспецифической стимуляции ФГА разбавляли до 0,5 мкг/мл добавлением свежей среды, в после трех дней среду меняли. После стимуляции клетки выращивали не менее 10 дней перед использованием в анализах на цитотоксичность. Клетки инфицировали рекомбинантной осповакциной при множественности инфицирования не менее 10 в течение одного часа в бессывороточной среде с последующей инкубацией в полной среде в течение трех часов. Клетки собирали центрифугированием и ресуспендировали при плотности 1·107 клеток/мл. 100 мкл этих клеток добавляли в каждую лунку U-образной планшетки для микротитрования, содержащих 100 мкл/лунку полной среды. Клетки разбавляли двумя последовательными операциями. Для каждого образца две лунки не содержали лимфоцитов, чтобы измерить спонтанное высвобождение хрома и суммарное поглощение хрома. Клетки-мишени из сублинии S3 клеток HeLa инфицировали в 6,0 или 10,0 см чашках при примерной множественности инфицирования (moi) 10 в течение одного часа в бессывороточной среде с последующей инкубацией в полной среде в течение трех часов. Затем их отслаивали от поверхности чашек с помощью PBS, содержащего 1 мМ ЭДТА, и подсчитывали. Аликвот 106 клеток-мишеней (HeLa, Raji, или RJ2.2.5-клетки для экспериментов с CD4-химерным рецептором, и клетки 3G8 10-2; Shen et al., Mol. Immunol. 26:959 (1989) для экспериментов с CD16-химерным рецептором) центрифугировали и ресуспендировали в 50 мкл стерильного 51Cr-хромата натрия (1 мКи/мл, Dupont Wilmington, DE) в течение одного часа при 37oC с периодическим перемешиванием и отмытых затем три раза с PBS. 100 мкл меченых клеток, ресуспендированных в среде при плотности 105 клеток/мл, добавляли в каждую лунку. Клетки-мишени Raji ь RJ2.2.5 метили таким же образом, что и клетки HeLa. Плашку для микротитрования центрифугировали при 750 · g в течение 1 минуты и инкубировали в течение 4 часов при 37oC. В конце инкубационного периода клетки в каждой лунке ресуспендировали осторожным пипеттированием, образец удаляли для подсчета включенной метки и плашку для микротитрования центрифугировали при 750 · g в течение 1 минуты. Отбирали 100 мкл аликвоты супернатантов и подсчитывали в сцинтилляционном счетчике гамма-излучения. Процент уничтожения корректировали по фракции инфицированных клеток-мишеней (обычно 50-90%) путем измерения проточной цитометрией. Для инфицированных эффекторных клеток соотношение эффектор:мишень корректировали по проценту инфицированных клеток (обычно 20-50% для экспериментов с CD4-химерным рецептором и >70% для экспериментов с CD16-химерным рецептором).
Мутагенез ζ-последовательности в условиях ин витро
Для создания точечных мутаций по аминокислотным остаткам 11 или 16 ζ- последовательности приготовили синтетические олигонуклеотидные праймеры, простирающиеся от BamHI-сайта выше ζ-трансмембранного домена и превращающие в естественной ζ-цепи в положении 11 остаток Цис на Гли (C11G) или остаток Асп на Гли в положении 15
(D15G) или оба (C11G/D15G), и использовали в реакциях ПЦР для получения мутантных фрагментов, которые вновь встраивали в CD4:ζ- конструкты дикого типа.
Чтобы создать ζ-делеции, последовательности кДНК ζ-цепи амплифицировали в ПЦР, используя синтетические олигонуклеотидные праймеры, предназначенные для создания стоп-кодона (UAG) после остатков 50, 59 или 65. Праймеры содержали сайт расщепления для фермента Notl, отрезающего пять или шесть остатков от 5'-конца, обычно в последовательности, построенной из CGC GGG CGG CCG CTA (SEQ ID NО: 11), в которой три последних остатка соответствуют стоп-антикодону. За последовательностями Notl и стоп-антикодона следуют 18 или больше остатков, комплементарных соответствующему 3'-концу этого фрагмента. Полученные химеры обозначили, соответственно, CD16:ζY51*, CD16:ζE60* и CD16: ζD66*. BamHI-сайт, расположенный выше трансмембранного домена, и Notl-сайт использовали для получения фрагментов, которые вновь встроили в CD16: ζ- конструкт дикого типа. Мономерные ζ- химеры создали в результате выделения ζ- трансмембранной и мембранной проксимальной внутриклеточной последовательностей путем расщепления с помощью BamHI и SacI, описанного выше Асп и Цис CD4: ζ-конструкта, и встраивания фрагмента, соответственно, в CD16:ζE60* и CD16:ζD66*-конструкт.
Конструирование CD16:7: ζ (48-65) и CD16:7: ζ (48-59) трираздельной химеры
Чтобы сконструировать CD16: ζ D66*, последовательность кДНК ζ- цепи, соответствующая трансмембранному домену, заменили соответствующим трансмембранным и цитоплазматическим доменом, полученным из кДНК CD5 и CD7.CD5- и CD7-фрагменты получили ПЦР-реакцией, используя прямые олигонуклеотиды, включающие в себя BamHI-сайт рестрикционного расщепления и соответствующий области, расположенной непосредственно выше трансмембранного домена, соответственно, CD5 и CD7, а затем обратные олигонуклеотиды, перекрывающие, соответственно, последовательности CD5 и CD7 и ζ-последовательность, которая содержит SacI-сайт рестрикционного расщепления.
CD5: ζ : CGC GGG GAG CTC GTT ATA GAG CTG GTT CTG GCG CTG CTT CTT CTG (SEQ ID NO: 12)
CD7: ζ : CGC GGG GAG CTC GTT ATA GAG CTG GTT TGC CGC CGA ATT CTT ATC CCG (SEQ ID NO: 13).
ПЦР-продукты CD5 и CD7 обрабатывали BamHI и SacI и лигировали с CD16: ζ E60*, обработанной BamHI и SacI, и замещали ζ- последовательность от BamHI до SacI на CD7-фрагмент. Для создания конструктор CD16:CD5 и CD16:CD7, фрагменты CD5 и CD7 получали ПЦР, используя олигонуклеотид, содержащий NotI-сайт рестрикционного расщепления и кодирующий стоп-кодон (UAA) после остатка Глн416 и Ала193, соответственно в CD5 и CD7. ПЦР-фрагмент CD5 и CD7 обрабатывали BamHI и SacI и встраивали в CD16:ζАсп66*-конструкт.
Мутагенез ин витро N-концевых остатков в пределах ζ-мотива, трансдуцирующего цитолитический сигнал
Синтетические олигонуклеотидные праймеры, простирающиеся от сайта SacI внутри ζ-мотива и превращающие естественный остаток 48 с Асп на Сер (N48S), остаток 50 - с Лей на Сер (L50S) и остаток 51 - с Тир на Фен (Y51F), синтезировали и использовали в ЦПР-реакции, чтобы получить фрагменты, которые реинтродуцировали в CD16:7: ζ (48-65)-конструкт дикого типа.
Мутагенез ин витро C-концевых остатков в пределах ζ-мотива, трансдуцирующего цитолитический сигнал
Синтетические олигонуклеотидные праймеры, простирающиеся от NotI-сайта до стоп-кодона и превращающие естественный остаток 60 с Глу на Глин (E60Q), остаток 61 - с Глу на Глн (E61Q), остаток 62 - с Тир на Фен или Сер (Y62F или Y62S) и остаток 63 - с Асп на Асн (D63N), синтезировали и использовали в ЦПР, чтобы получить фрагменты, которые субклонировали в CD16: ζ D66*-конструкт дикого типа от BamHI-сайта до NotI-сайта.
Химерные конструкции CD16: 7: ζ (33-65), CD16:7: ζ (71-104), CD16:7: ζ (104-137)
Трансмембранный фрагмент CD7, несущий MluI- и NotI-сайты на стыке между трансмембранным и внутриклеточным доменами, получили в ПЦР, используя олигонуклеотид следующей последовательности: CGC GGG GCG GCC ACG CGT CCT CGC CAG CAC ACA (SEQ ID NO: 14). Полученный ПЦР-фрагмент обрабатывали BamHI и NotI и вновь встраивали в CD16: 7: ζ (48-65)-конструкт. ζ- фрагменты, кодирующие остатки 33-65, 71-104 и 104-137, получали в ПЦР-реакции, используя пару праймеров, содержащих MluI-сайты на 5'-конце прямых праймеров и стоп-кодонов, следующими за NotI-сайтами на 5'-конце обратных праймеров. В каждом случае сайты рестрикции отрезали шесть остатков от 5'-концов праймеров, чтобы гарантировать расщепление ферментом рестрикции
ζ 33: CGC GGG ACG CGT TTC AGC CGT CCT CGC CAG CAC ACA (SEQ ID NO: 15)
ζ 71: CGC GGG ACG CGT GAC CCT GAG ATG GGG GGA AAG (SEQ ID NO: 16) и
ζ 104: CGC GGG ACG CGT ATT GGG ATG AAA GGC GAG CGC (SEQ ID NO: 17).
Конструирование FcR ζ IIA-делеционных мутантов
Карбокси-конец FeRIIA-делеционных мутантов конструировали с помощью ПЦР таким же образом, как и конструкты полной длины, превращая последовательности, кодирующие тирозин в положении 282 и 298, в стоп-кодоны (TAA). N-концевые делеции получали, амплифицируя с помощью ПЦР, фрагменты, кодирующие последовательно меньший внутриклеточный домен, используя олигонуклеотиды, которые позволяют полученным фрагментам встраиваться между сайтами рестрикции MluI и NotI в ранее сконструированную экспрессионую плазмиду, кодирующую внеклеточный домен CD16, слиты с трансмембранным доменом CD7, позднее терминированный по сайту MluI и присоединенный между трансмембранным и внутриклеточным доменом.
Другие варианты осуществления
Вышеописанные примеры свидетельствуют, что агрегация ζ-,η- или γ-химер достаточна для инициации цитолитического эффекторного клеточного ответа в T-клетках. Знание области экспрессии ζ,η или γ, которую включают T-лимфоциты, естественные клетки-киллеры, базофильные гранулоциты, макрофаги и тучные клетки, предполагает, что мотивы консервативной последовательности могут взаимодействовать с чувствительным аппаратом, обычным для клеток гемопоэтического происхождения и что важный компонент защитных сил организма в иммунной системе может быть опосредован исходом рецепторной агрегации.
Эффективность цитолитического ответа и отсутствие ответа на клетки-мишени, несущие рецепторы МНС класса II, свидетельствуют, что химеры, основанные на ζ,η и γ, создают основу для генетического воздействия на СПИД путем адаптивной иммунотерапии. Широкое распространение эндогенных ζ и η и очевидность, с которой Fc-рецепторы, ассоциированные с γ, опосредуют цитотоксичность в различных типах клеток (Fanger et al., Immunol. Today 10:92-99 (1989)), позволяют рассматривать разнообразие клеток применительно к этой цели. Например, нейтрофильные гранулоциты, которые обладают очень короткой продолжительностью жизни (≈4 ч) в циркулирующей крови и очень цитолитичны, представляют собой привлекательные клетки-мишени для экспрессии химер. Инфицирование нейтрофилов ВИЧ, вероятно, не приводит к высвобождению вируса, а изобилие этих клеток (преобладают над лейкоцитами) должно усиливать защитные силы организма. Другую привлекательную возможность для клеток организма открывают зрелые T-клетки, популяция, пригодная в настоящее время для ретровирусной инженерии (Rosenberg, Sci Am. 262:62-69 (1990)). С добавлением рекомбинантного IL-2 T-клеточная популяция при культивировании может относительно легко увеличиться в объеме, а при реинфузии такие популяции обычно имеют ограниченную продолжительность жизни (Rosenberg et al., N. Engl. J. Med. 323: 570-578 (1990)).
При соответствующих условиях узнавание ВИЧ-клетками, экспрессирующими CD4-химеры, должно также обеспечиваться митогенными стимулами, открывая возможность того, чтобы вооруженная клеточная популяция могла бы динамично реагировать на вирусное бремя. Несмотря на то, что мы сконцентрировали здесь внимание на поведении слитых белков в цитолитических T-лимфоцитах, экспрессия химер в хелперных лимфоцитах могла бы создать источник цитокинов, мобилизованный ВИЧ, который мог бы нейтрализовать разрушение подгруппы хелперных клеток при СПИД. Недавнее описание нескольких вариантов создания резистентности к инфекции на других стадиях, кроме проникновения вируса в клетку (Friedman et al., Nature 335:452-454 (1988); Green et al., Cell 58:215-223 (1989); Malim et al., Cell 58:205-214 (1989); Trono et al., Cell 59:113-120 (1989); Buonocore et al., Nature 345:625-628 (1990)), предполагает, что клетки, несущие CD4-химеры, могли бы быть предназначены для того, чтобы препятствовать вирусному размножению в результате экспрессии соответствующих агентов, имеющих внутриклеточный сайт воздействия.
Способность передавать сигналы для T-лимфоцитов посредством автономных химер также предусматривает способность регуляции ретровирусно созданных лимфоцитов ин виво. Стимулирующее действие сшивки, опосредованное, например, специфичными IgM-антителами, созданными для удаления доменов, связывающих комплемент, может позволить таким лимфоцитам количественно увеличиваться ин ситу, тогда как лечение подобными специфическими антителами IgG (например, для узнавания аминокислотной изменчивости, созданной в химерной цепи) могло бы истощить сконструированную популяцию. Кроме того, антитела IgM анти-CD4 не требуют дополнительной сшивки, чтобы мобилизовать кальций в Jurkat-клетках, экспрессирующих CD4: ζ-химеры. Способность регулировать клеточную популяцию, не прибегая к помощи повторной экстракорпоральной амплификации, может существенно расширить сферу и эффективность нынешнего использования, предложенного для геноинженерных T-клеток.
Хотя настоящее изобретение описано в связи со специфическими вариантами его осуществления, необходимо иметь в виду, что в него могут быть внесены дополнительные изменения, а настоящая заявка подразумевает внесение изменений, назначений или переделок настоящего изобретения, в том числе таких отклонений от настоящего существа изобретения, которые находятся в пределах техники, к которой относится настоящее изобретение и которые могут быть применены к существенным признакам, сформулированным выше, также как и в рамках нижеследующей формулы изобретения.



























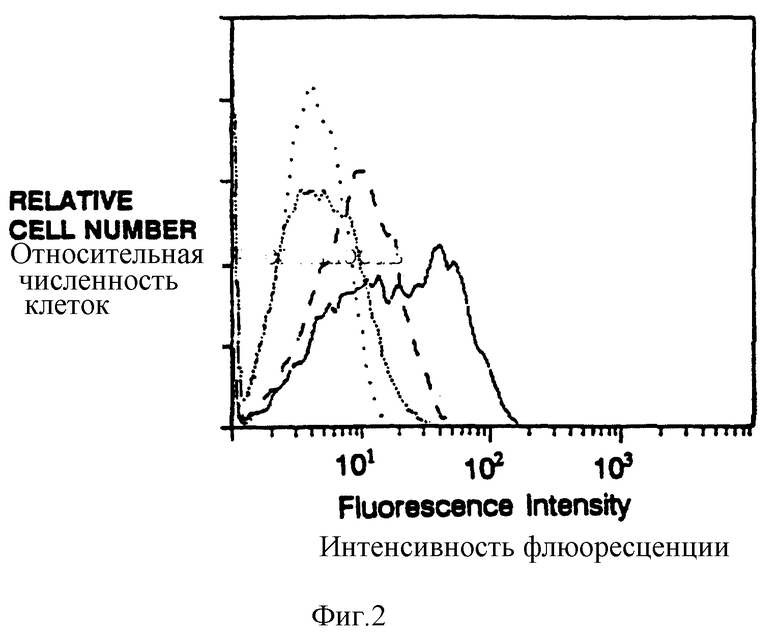
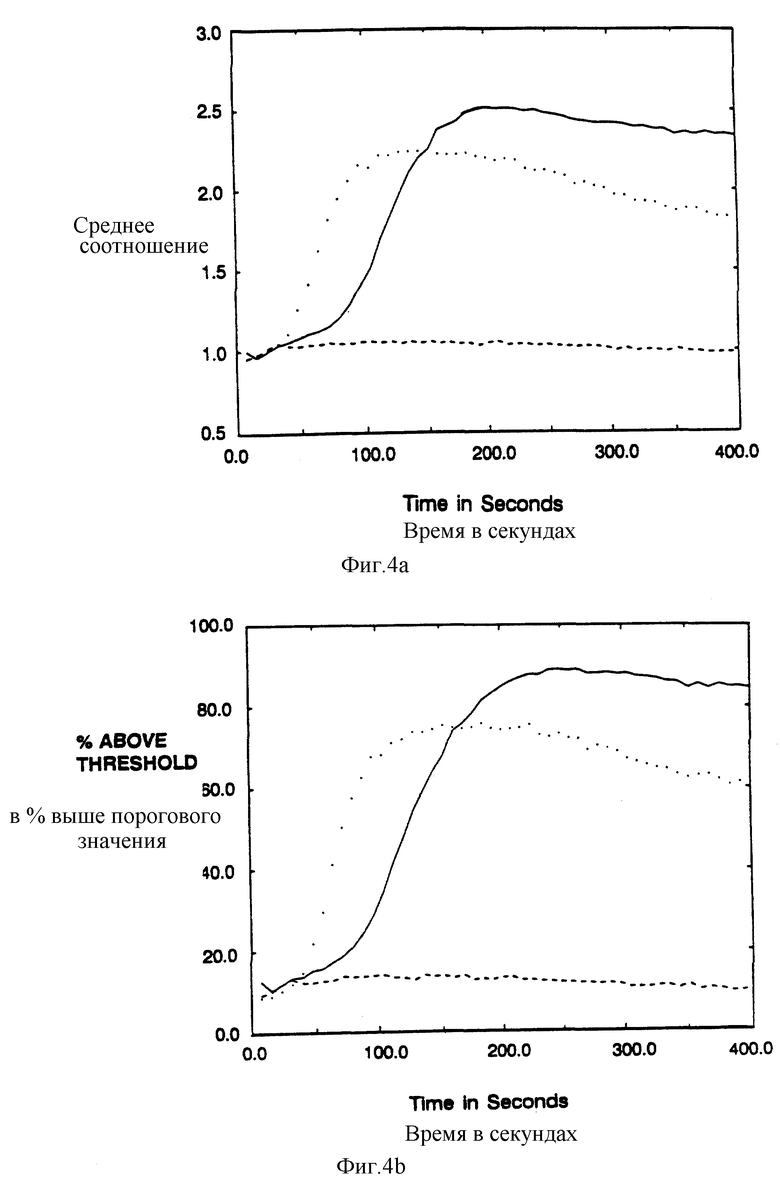
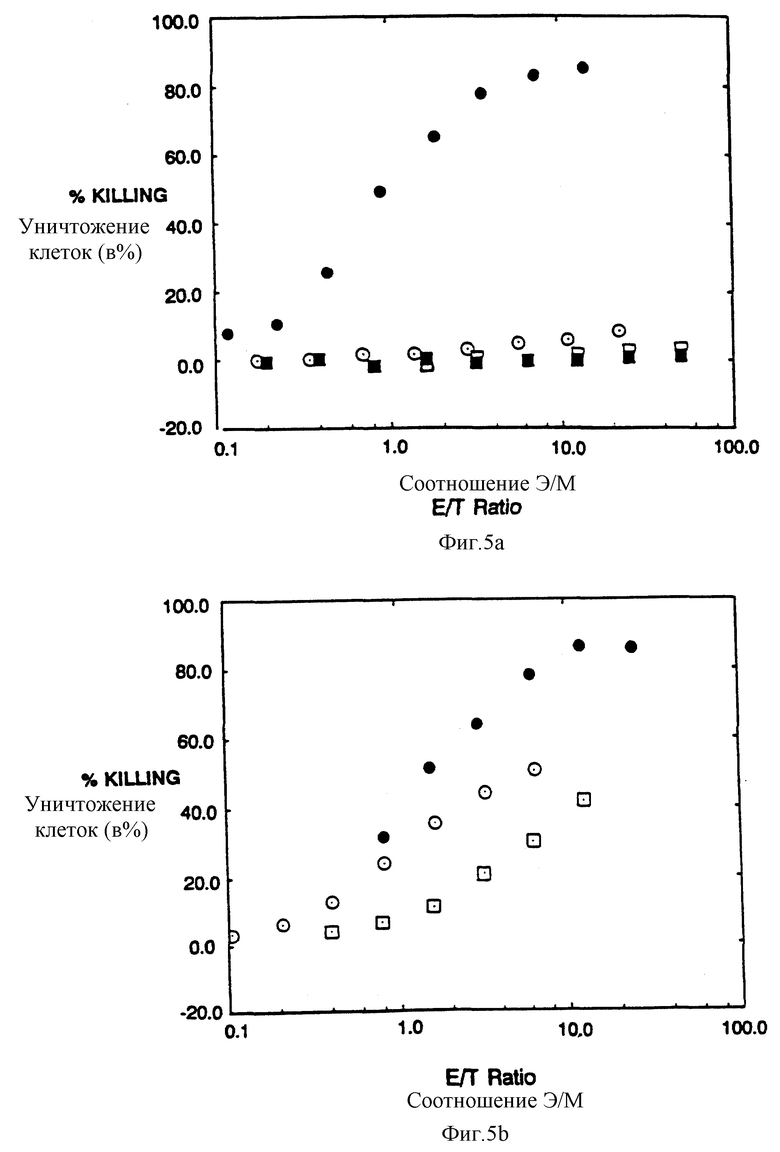
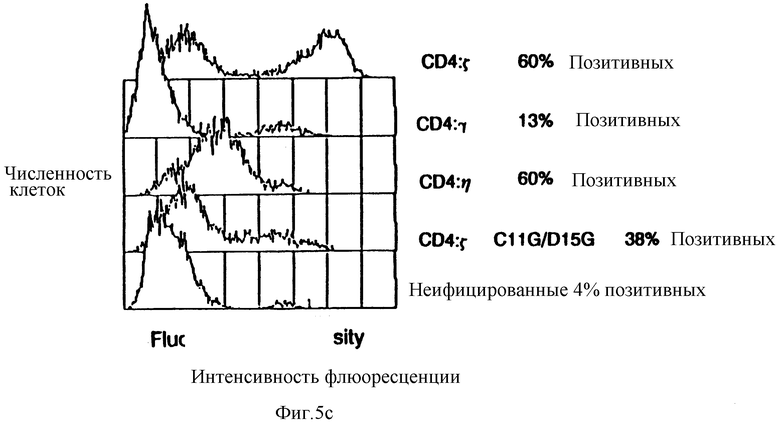

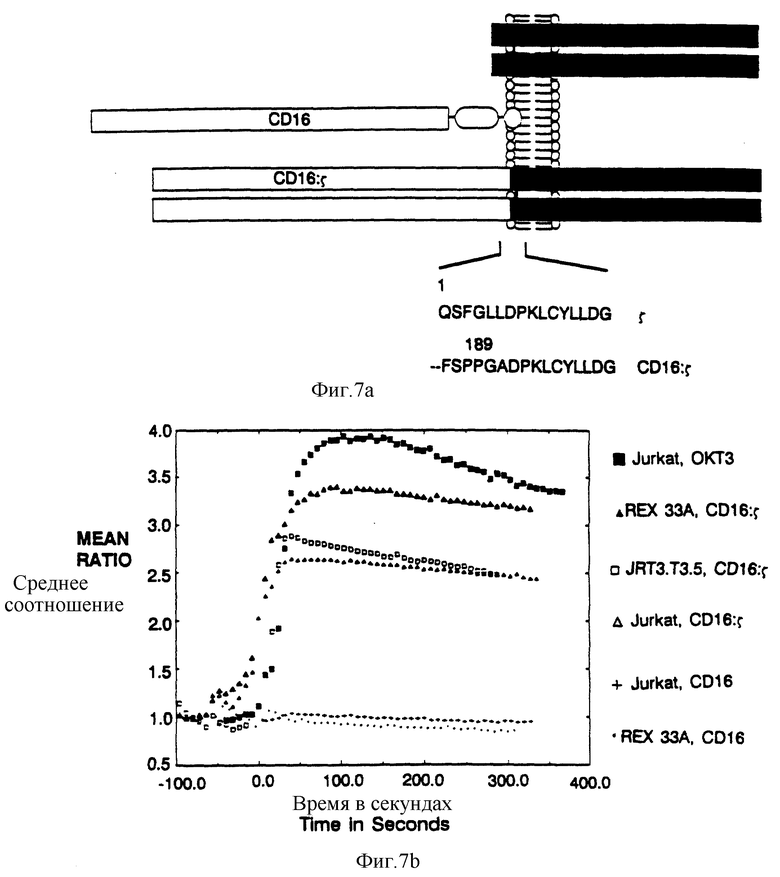
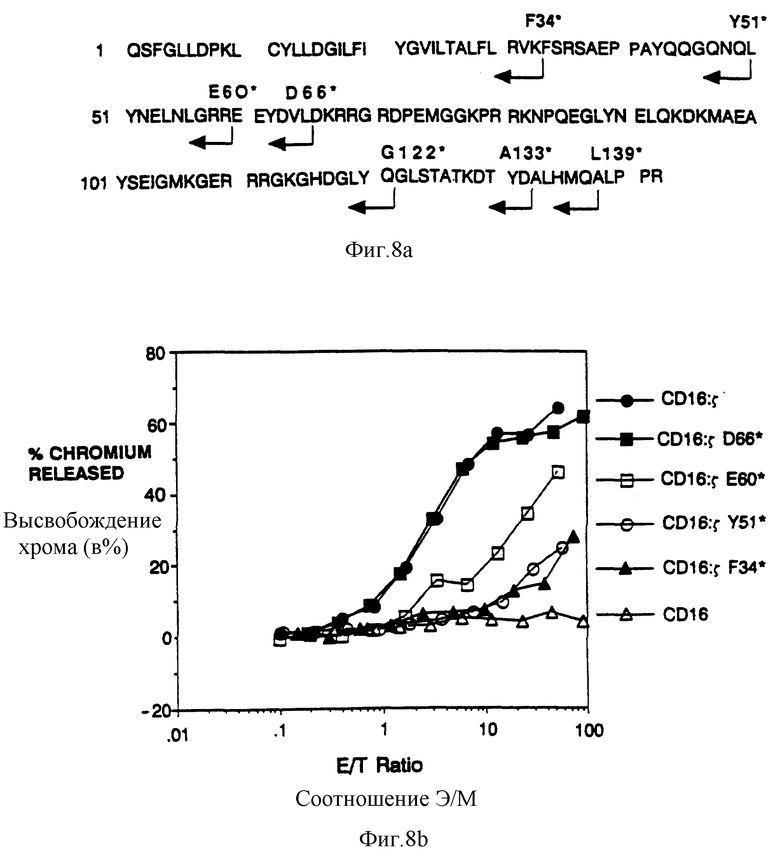
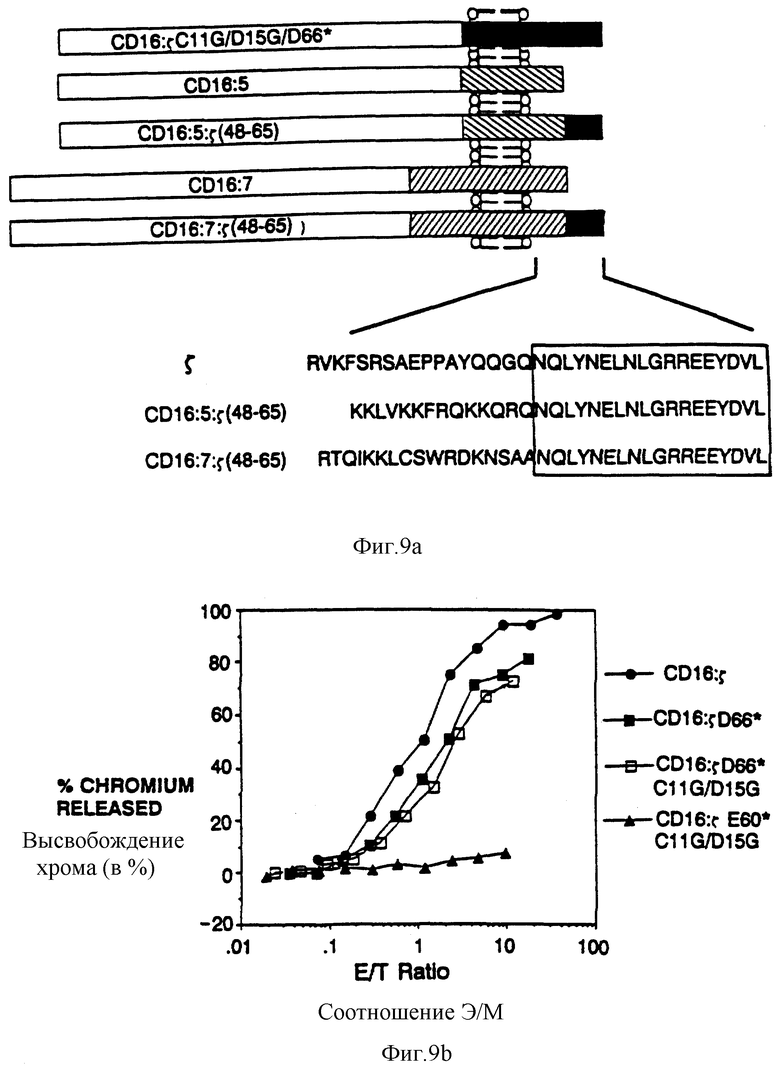
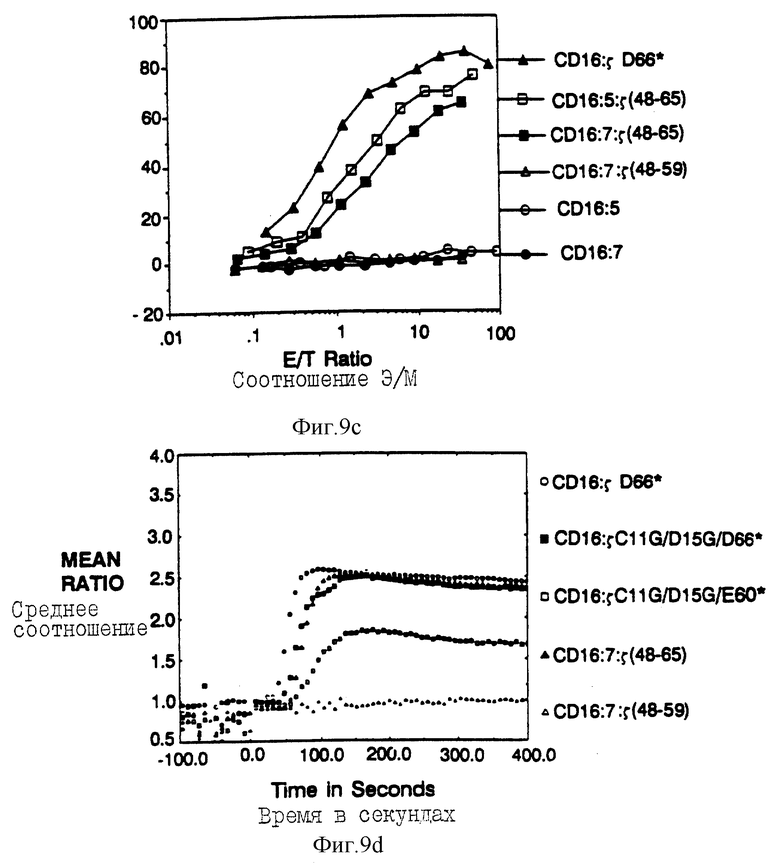
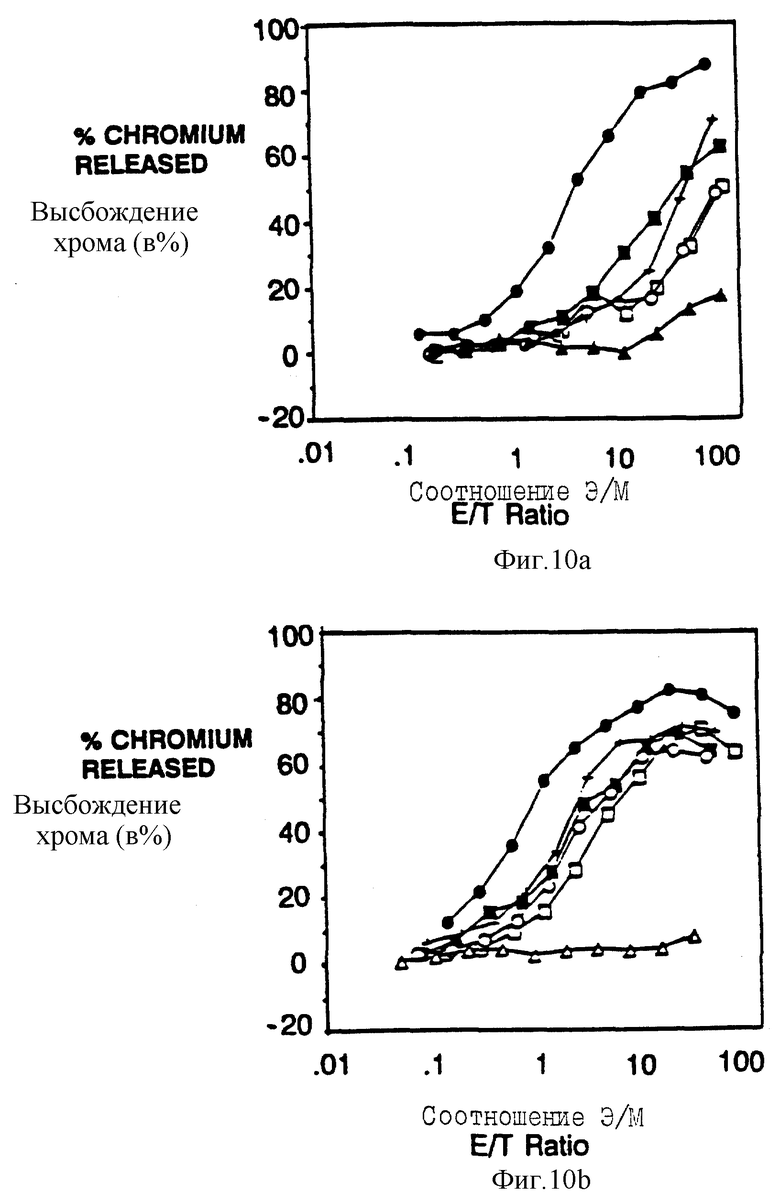
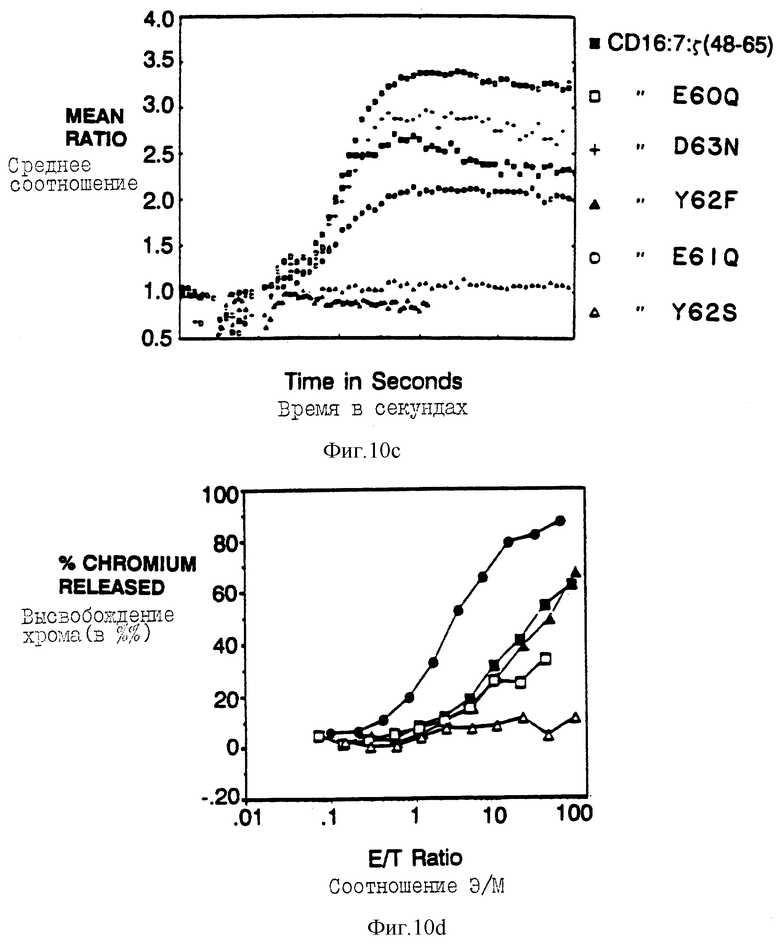
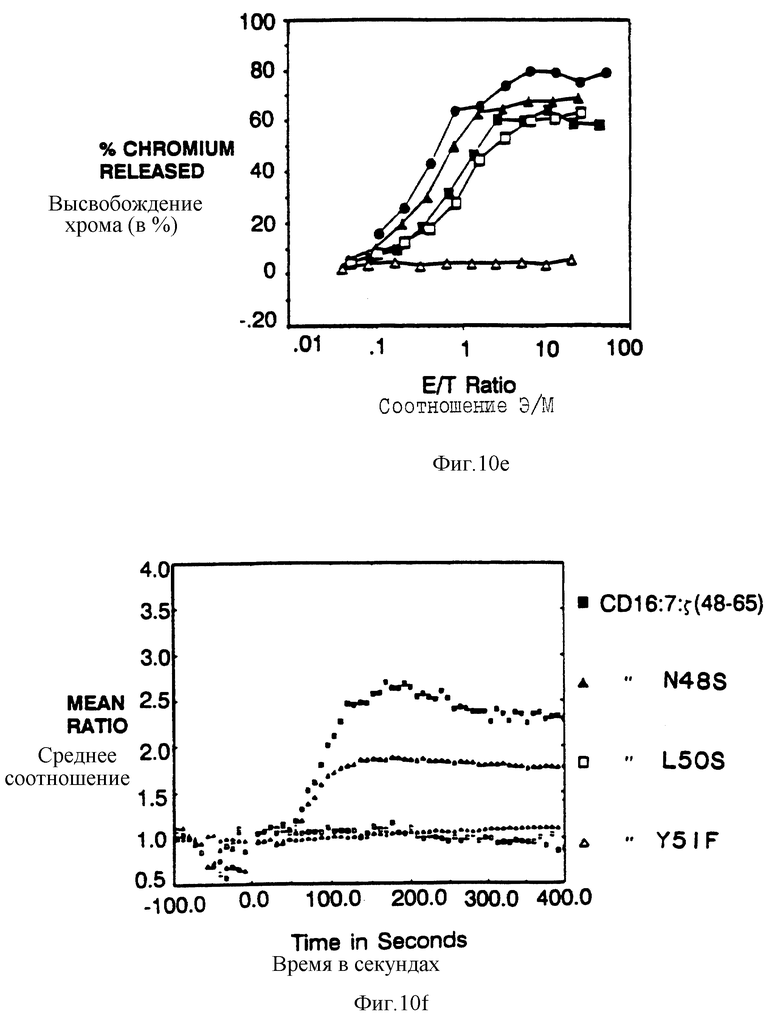
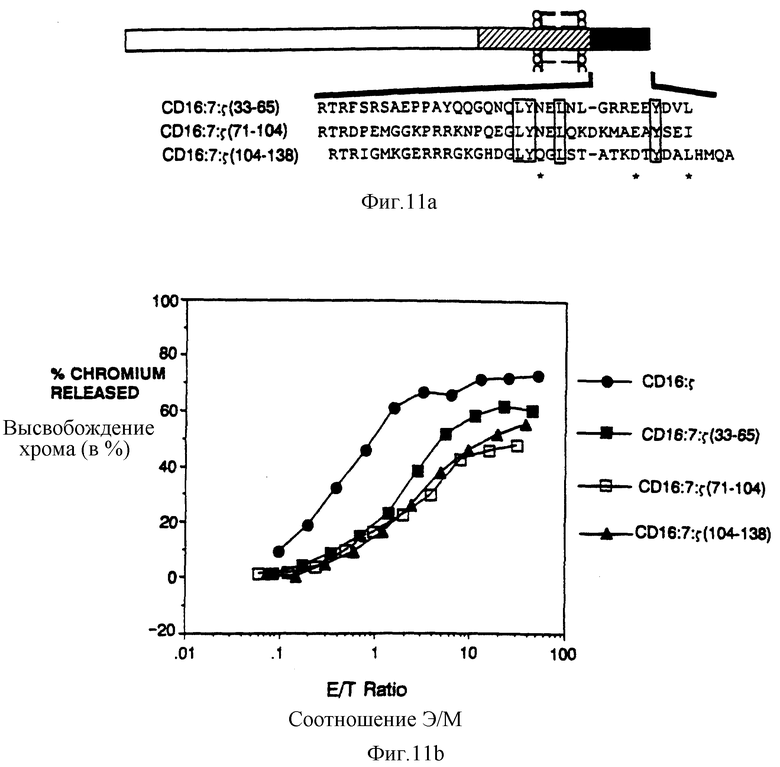
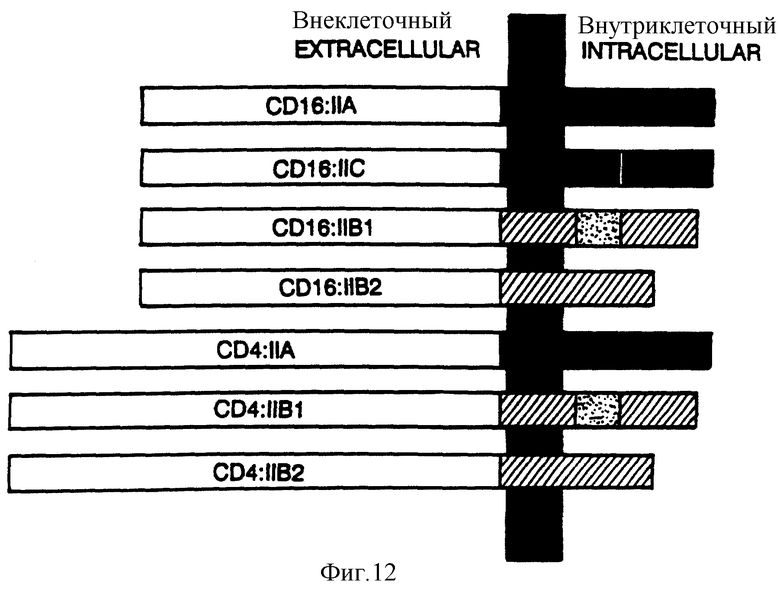
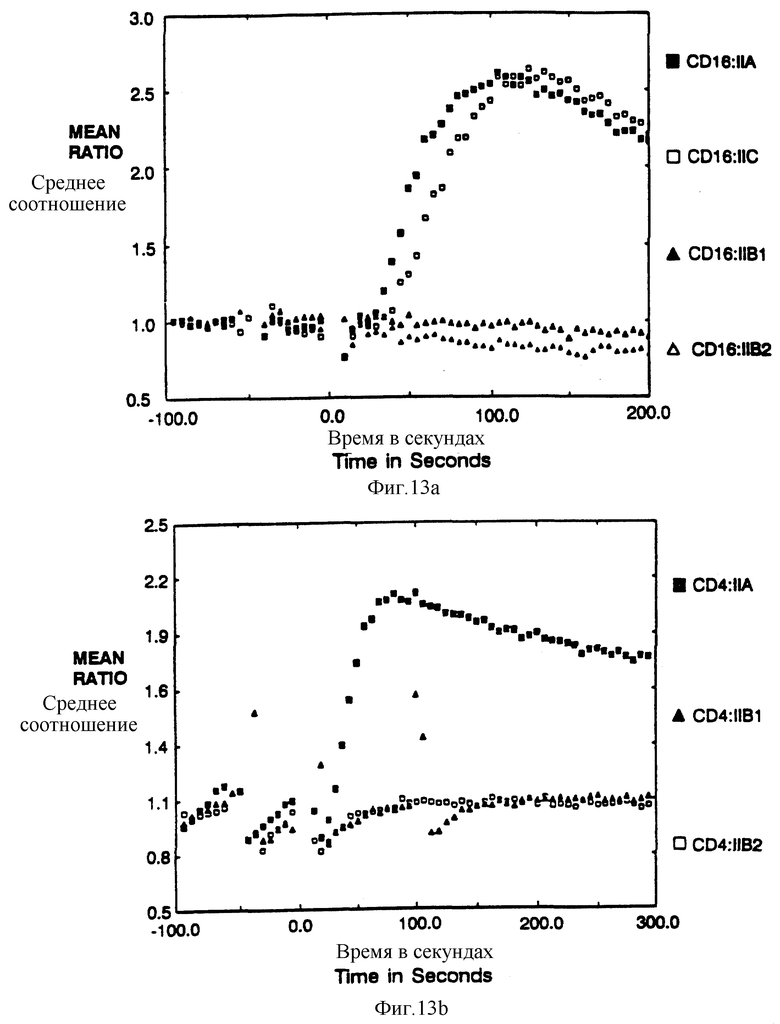
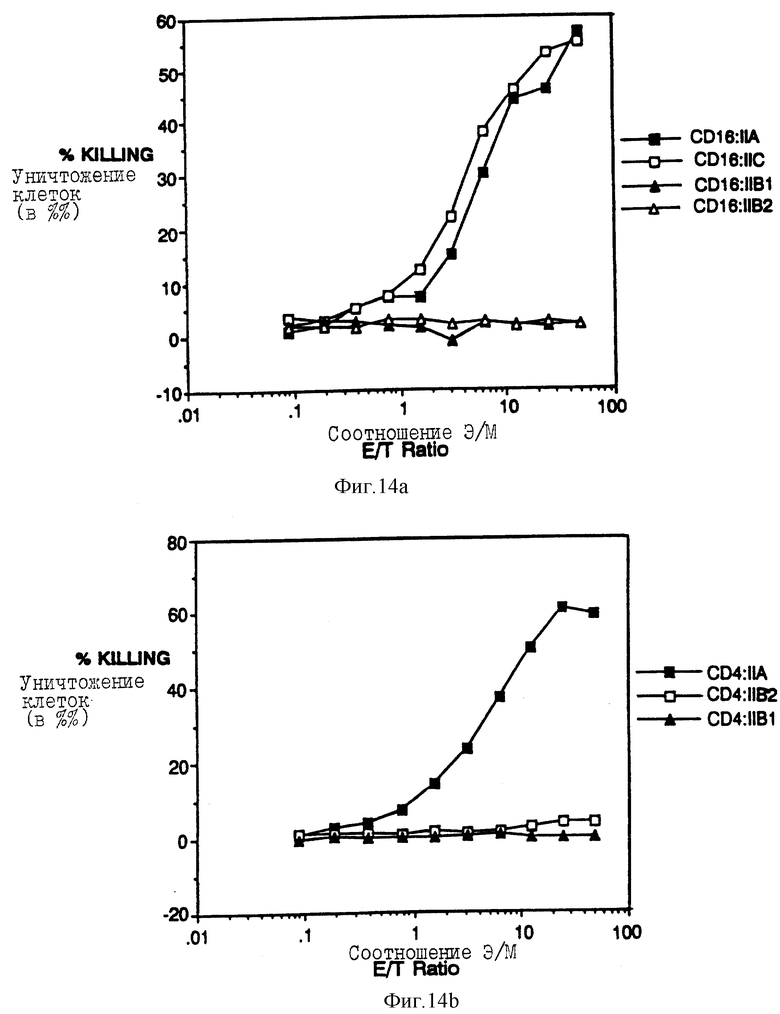
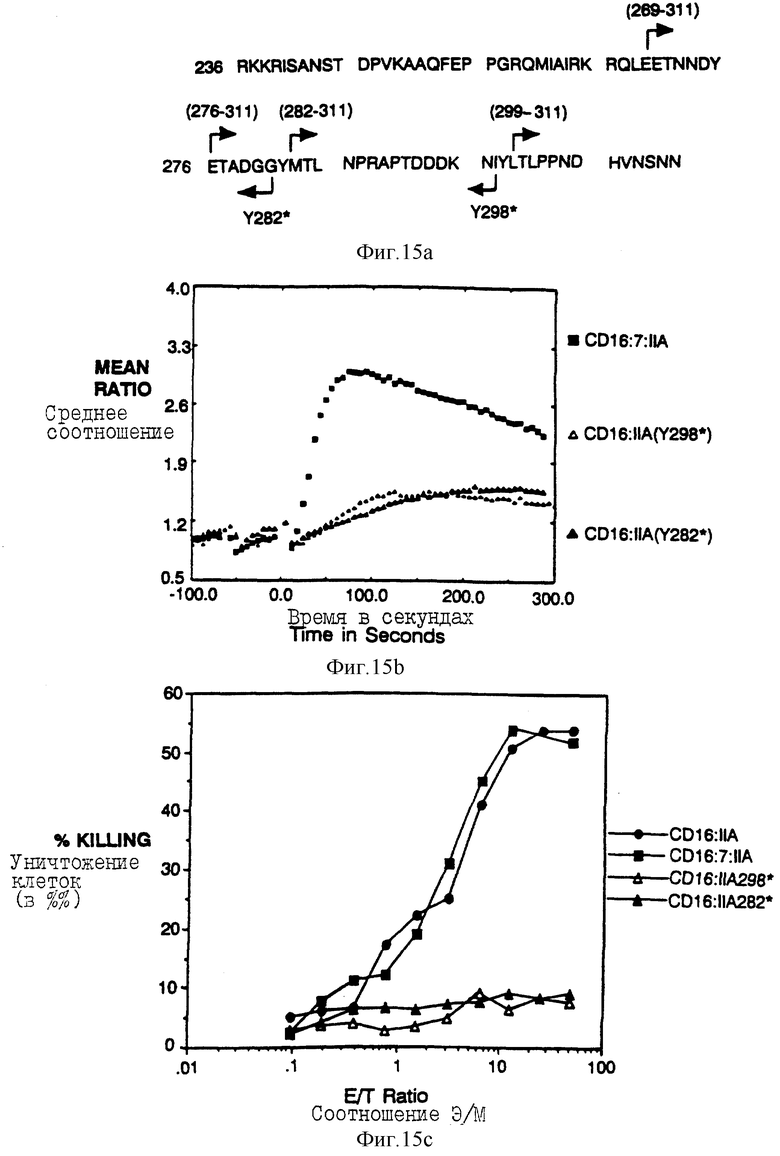
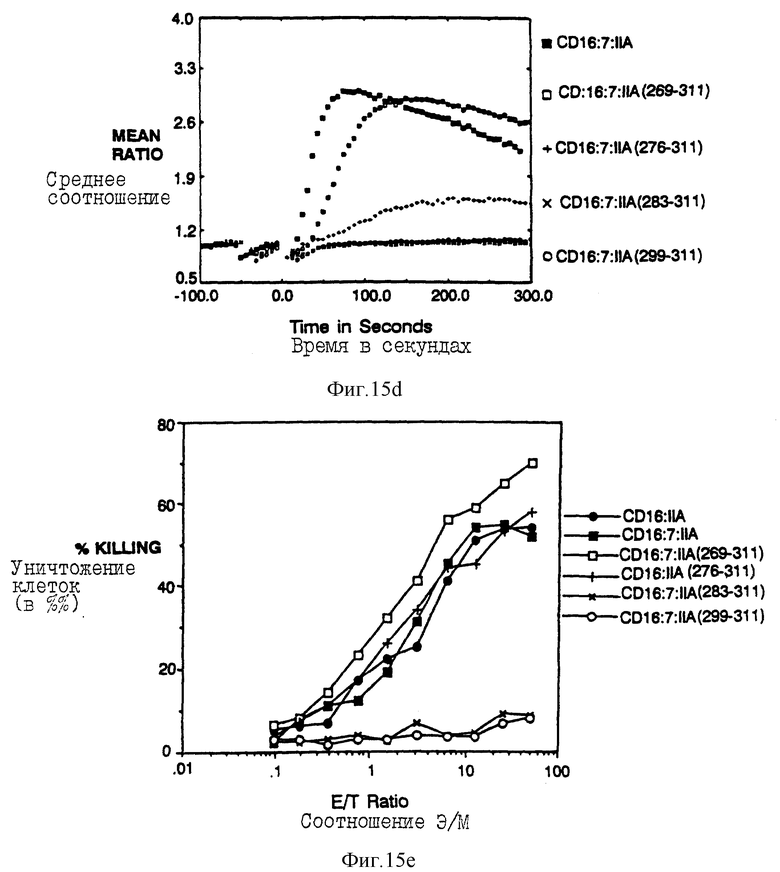
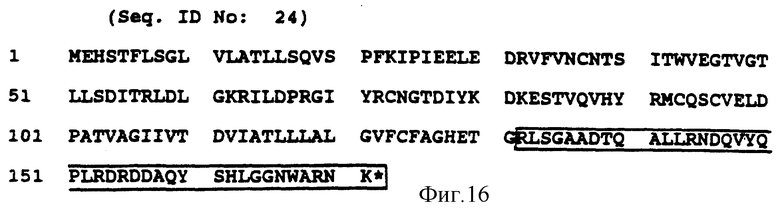

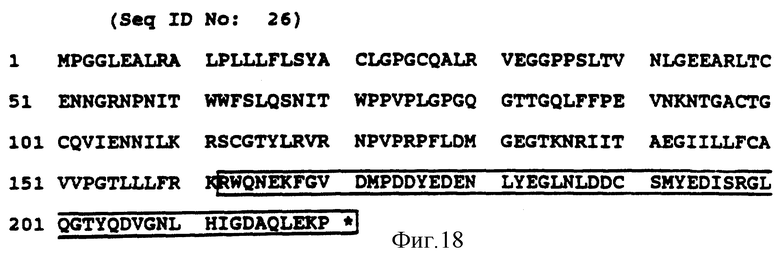

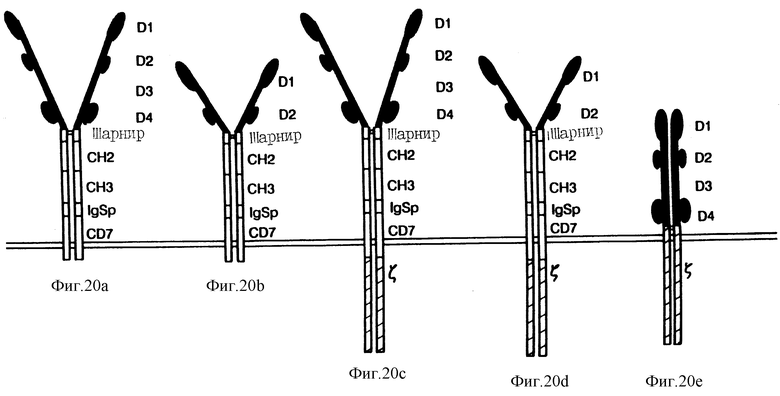
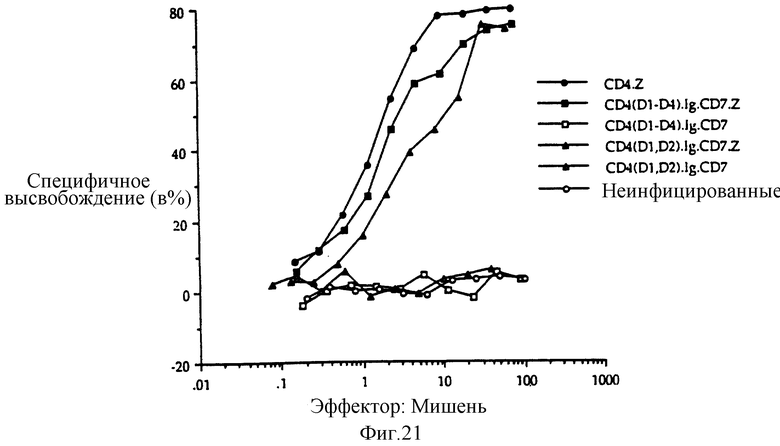
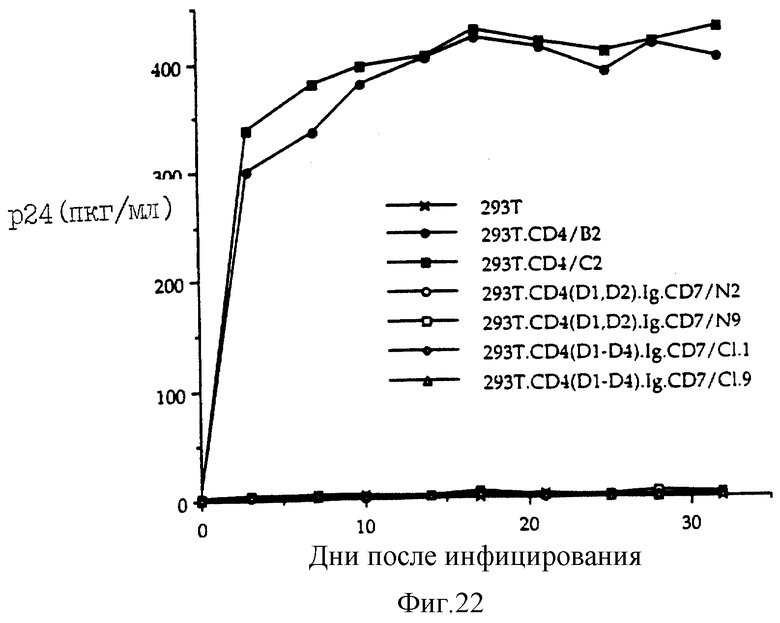






Изобретение относится к области генной инженерии. Предложенный способ включает введение млекопитающему эффективного количества терапевтических клеток, экспрессирующих связанный с мембраной белковый рекомбинантный рецептор, включающий в себя внеклеточную часть, которая включает в себя фрагмент CD4, который обладает способностью специфически узнавать и связывать ВИЧ-инфицированную клетку, но который не опосредует ВИЧ-инфицирование. Кроме того, описан белковый рекомбинантный рецептор, который также способен специфично распознавать и связывать ВИЧ-инфицированную клетку, но который не опосредует ВИЧ-инфекцию. Изобретение позволяет управлять клеточным иммунным ответом, направленным против ВИЧ-инфицированной клетки млекопитающего. 2 с. и 9 з.п. ф-лы, 28 ил., 1 табл.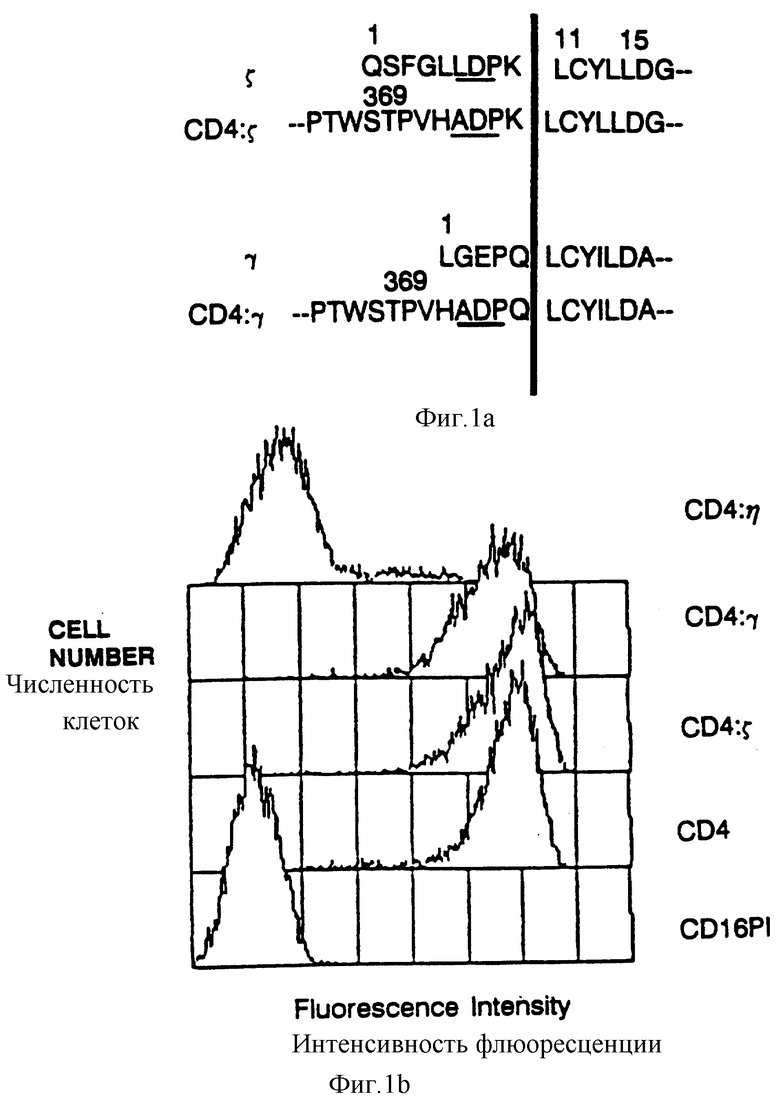
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| The EMBO Journal, 1992, vol 11, по | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| ПОДОГРЕВАТЕЛЬ ДЛЯ ИСПОЛЬЗОВАНИЯ ТЕПЛОТЫ ВЫХЛОПНЫХ ГАЗОВ ДЛЯ НАГРЕВА ВОДЫ | 1924 |
|
SU575A1 |
| Journal Biological Chemistry, 05 April 1989, vol.264, по | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
