Изобретение относится к биотехнологии и, в частности, к генетической инженерии и представляет собой штамм рекомбинантного вируса осповакцины, обуславливающий синтез структурного белка вируса гепатита В человека (HBsAg), в котором пре-S2-область заменена на искусственный полипептид, кодирующий девять иммуногенных эпитопов двух основных структурных белков вируса иммунодефицита человека типа 1 (ВИЧ-1) в инфицированных клетках, и Т- и В-клеточный иммунный ответ против HBsAg и ВИЧ-1 у вакцинированных им лабораторных животных.
Вирус гепатита В человека широко распространен по всему миру. Лечение и предупреждение гепатита В имеет большое значение для медицины, так как он, видимо, является самым распространенным виновником хронических заболеваний печени, в том числе гепатоцеллюлярной карциномы у человека [1].
Вирус иммунодефицита человека вызывает все более распространяющееся эпидемически опасное заболевание, методы лечения и предотвращения которого пока отсутствуют и оно часто завершается летальным исходом. До 1995 г. территория бывшего Советского Союза и стран Восточной Европы считалась зоной с низким уровнем ВИЧ-инфицированности населения. После 1995 г., когда вспышки ВИЧ-инфекции произошли сначала на Украине, а затем в Белоруссии и в Российской Федерации, эти страны стали характеризоваться как имеющие самый высокий относительный прирост числа случаев ВИЧ-инфекции [2].
Создание эффективных вакцин против этих вирусных инфекций является актуальной задачей. Одним из подходов к решению этой проблемы является использование геномов некоторых вирусов в качестве векторов для экспрессии протективных антигенов патогенных вирусов в клетках млекопитающих.
В современной литературе описано несколько векторов, позволяющих экспрессировать чужеродные гены в эукариотических клетках. Созданные к настоящему времени вектора на основе ретровирусов, аденовирусов, папилломавирусов и паповавирусов обладают целым рядом существенных недостатков: малой скоростью размножения на культуре клеток, небольшой емкостью относительно размера встраиваемых генетических последовательностей, неспособностью размножаться в организме тех или иных животных.
Определенные успехи по созданию рекомбинантных вакцин были достигнуты при использовании вируса осповакцины (BOB). BOB широко используется для экспрессии чужеродных белков, представления их иммунной системе и определения роли индивидуального антигена в иммунном ответе [3, 4]. Были сконструированы рекомбинантные ВОВ, экспрессирующие отдельно индивидуальные белки ВИЧ-1: gpl60, gpl20, tat и обратную транскриптазу [5-9]. Полученные рекомбинантные ВОВ были использованы для иммунизации лабораторных животных и выявления белков ВИЧ-1, способных вызывать индукцию вируснейтрализующих антител. Кроме того, это давало возможность оценить относительный вклад и взаимодействие гуморальной и клеточной составляющей иммунитета, поскольку ВОВ является эффективным индуктором как Т-, так и В-клеточного иммунного ответа, т. е. фактически выполняет роль адъюванта. Результаты проведенных исследований свидетельствуют, что рекомбинантные ВОВ, экспрессирующие иммунологически значимые антигены ВИЧ-1, способны вызывать индукцию гуморального и клеточного иммунного ответа, что является определяющим для выработки протективного иммунитета и, следовательно, для создания вакцины.
Однако ни в одной из опубликованных работ с помощью рекомбинантных ВОВ не удалось индуцировать полноценный протективный иммунный ответ против ВИЧ-инфекции. По-видимому это обусловлено низкой иммуногенностью индивидуальных вирусных белков. Для индукции полноценного ответа иммунной системе, по-видимому, необходимо представлять антигенные детерминанты одновременно нескольких вирусных белков и в более иммуногенном виде, чем тот, который имеют эти детерминанты в природных вирусных белках.
Продуктивным подходом для преодоления возникающих трудностей является создание надмолекулярных вирусоподобных белковых структур, на поверхности которых многократно представлены те или иные антигенные детерминанты. При этом необходимо наличие белка-носителя, который при определенных условиях способен осуществить самосборку вирусоподобных частиц. Подходящим кандидатом на роль такого белка-носителя является поверхностный вирионый белок вируса гепатита В человека HBsAg (иначе называемый S), который при синтезе в эукариотических клетках формирует высокоиммуногенные сферические частицы диаметром около 22 нм (S-частицы) [10]. Одна из форм поверхностного белка, называемая npeS2-S, при синтезе в эукариотических клетках образует надмолекулярные структуры, на поверхности которых многократно представлена последовательность пpeS2. Такие частицы индуцируют при вакцинации еще более выраженный иммунный ответ против гепатита В по сравнению с S-частицами [11].
Известна рекомбинантная плазмидная ДНК и рекомбинантный ВОВ, экспрессирующий пептид размером 15 аминокислотных остатков из состава gp120 ВИЧ-1 (пептид P18IIIB) в составе химерного белка-гемагглютинина вируса гриппа Н-1 (НА) [12, прототип] . Иммунофлюоресцентные исследования с антисыворотками против H1N1 вируса гриппа и пептида P18IIIB позволяют выявить химерный белок внутриклеточно, но пептид P18IIIB не обнаруживается на внешней поверхности инфицированных клеток. Эти рекомбинанты определяют в основном цитотоксическую специфичность Т-лимфоцитов CD8+ к P18IIIB и НА вируса гриппа; наблюдается стимуляция продукции специфичных антител против НА, но не против P18IIIB. Однако такие Т-лимфоциты способны лизировать клетки, инфицированные ВИЧ-1.
Недостатком рассмотренного выше реализованного подхода является то, синтезируемый рекомбинантным ВОВ химерный белок не секретируется из инфицированных клеток и не формирует надмолекулярных образований, которые потенциально могут быть высокоиммуногенными.
Одним из перспективных направлений в создании эффективных и безопасных вакцин нового поколения является использование синтетических полипептидов, несущих протективные Т- и В-эпитопы. Для повышения эффективности синтетических вакцин используют такие приемы, как многократное повторение протективного эпитопа, соединение в одну цепь нескольких В-эпитопов, пришивку к В-эпитопу Т-детерминанты, экспрессию эпитопа в составе некоторого белка-носителя. При этом считается, что вакцины, эффективные в гетерогенной по HLA антигенам человеческой популяции, должны содержать по крайней мере несколько Т-детерминант и несколько В-детерминант [13].
В России разработан универсальный подход к конструированию синтетических вакцин в виде белков с оптимальным составом эпитопов инфекционного агента и с заранее заданной третичной структурой. В рамках этого подхода осуществлен дизайн 4-спирального белка-кандидата в вакцины против ВИЧ-1 [14, 15]. Этот белок назван TBI.
Белок-иммуноген (TBI) включает следующие Т-клеточные эпитопы ВИЧ-1: 105-117 HEDIISLWDQSLK, 421-437 KQIINMQEVGKAMYA, 827-841 DRVIEVVQGAYRAIR из ENV, 291-305 EPFRDYVDRFYKTLR из GAG (штамм ВН10). В белке присутствуют 5 различных нейтрализующих В-эпитопов: 255-265, 309-317, 730-744 из ENV и 92-109, 351-361 из GAG. Белок TBI - кандидат в вакцины против ВИЧ - удовлетворяет основным требованиям к эффективным вакцинам (мультивалентность, конформационная адекватность В-эпитопов, отсутствие нежелательных эпитопов).
Технической задачей изобретения является создание штамма рекомбинантного вируса осповакцины, осуществляющего экспрессию in vivo искусственного гена, кодирующего основные экспериментально локализованные эпитопы вируса ВИЧ, способные вызывать индукцию гуморального и клеточного иммунного ответа против ВИЧ-1, в составе химерного белка, белком-носителем которого является HBsAg, обладающий способностью к самосборке в высокоиммуногенные вирусоподобные частицы.
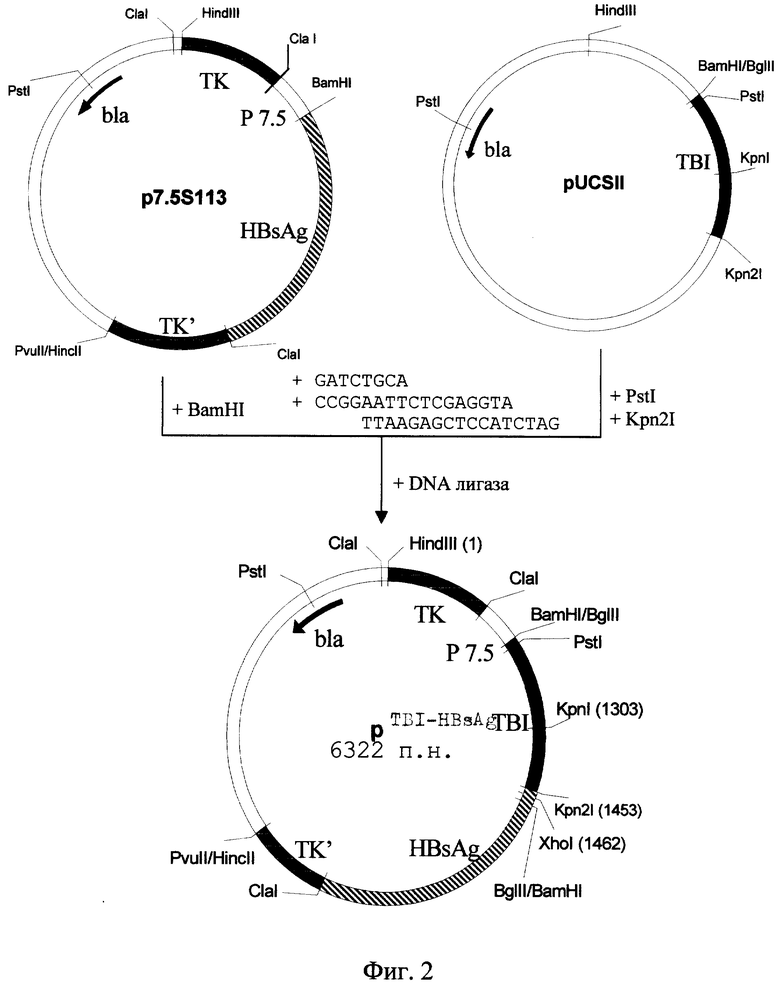
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК p TBI-HBsAg размером 6322 п.н., несущей химерный ген HBsAg, в котором вместо пре-S2-области находится последовательность, кодирующая ген исскуственного белка TBI, под контролем промотора белка 7.5К вируса осповакцины, и состоящей из (фиг.1-3):
- ClaI-BamHI векторного фрагмента ДНК плазмиды p7.5S113 размером 4647 п. н. , обеспечивающего селекцию и размножение гибридной плазмиды в клетках бактерии E. coli, а также рекомбинационное встраивание и экспрессию в клетках млекопитающих чужеродных генов в составе гена тимидинкиназы вируса осповакцины;
- Bg1II-PstI синтетического олигонуклеотида размером 8 п.н., обеспечивающего правильную стыковку гена TBI с сайтом инициации трансляции, находящимся в последовательности векторной плазмиды, имеющего структуру
5'-GATCTGCA-3';
- PstI-Kpn2I фрагмента ДНК размером 432 п.н., кодирующего искусственный полипептид TBI, иммуногенный против ВИЧ-1;
- Kpn2I-BgIII синтетического фрагмента ДНК размером 17 п.н., соединяющего гены белков TBI и S в правильной рамке трансляции, что обеспечивает синтез химерного белка TBI-HBsAg, имеющего структуру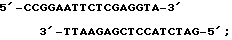
- BamHI-ClaI фрагмента генома вируса гепатита В, кодирующего поверхностный вирионный белок S (HBsAg), размером 1226 п.н.;
Для получения целевого рекомбинантного штамма вируса осповакцины, обеспечивающего экспрессию in vivo исскуственного гена TBI, кодирующего основные антигенные эпитопы ВИЧ-1 в составе химерного HBsAg, описанную плазмиду р TBI-HBsAg встраивают в ген тимидинкиназы коммерческого штамма ЛИВП вируса осповакцины под контролем промотора 7.5К. Полученный в результате рекомбинантный штамм ВОВ, обозначенный VR18, содержит в геноме, по сравнению с исходным штаммом ЛИВП, дополнительную последовательность, состоящую из:
- фрагмента длиной 284 н. п., включающего район промотора гена белка 7.5К;
- фрагмента длиной 1675 н.п., содержащего ген HBsAg, в котором вместо пре-S2-области находится последовательность гена исскуственного белка TBI.
Нуклеотидная последовательность встроенного фрагмента приведена на фиг. 1.
Полученный штамм рекомбинантного вируса осповакцины VR18 характеризуется следующими признаками.
Морфологические признаки.
Штамм VR18 обладает свойствами типичного представителя семейства ортопоксвирусов. Вирионы штамма имеют размер 200•300 нм, характерную брикетообразную форму и по данным электронной микроскопии не отличаются от исходного штамма ЛИВП.
Физиолого-биохимическая характеристика и культуральные свойства штамма.
ДНК штамма VR18 имеет длину около 190 т.п.н., при HindIII-рестрикционном анализе данной вирусной ДНК вместо HindIII-K-фрагмента ДНК штамма ЛИВП (5008 п.н.) выявляется фрагмент большего молекулярного веса (6967 п.н.), по данным ДНК-ДНК блот-гибридизации содержащий последовательности генов HBsAg и TBI.
При размножении VR18 на развивающихся куриных эмбрионах характер поражений на хориоаллантоисных оболочках такой же, как и при размножении исходного штамма ЛИВП. Кроме того, штамм VR18 не отличается от исходного по продуктивности в монослое перевиваемых линий клеток CV-1 и Н-143ТК-. В отличие от штамма ЛИВП рекомбинант VR18 обладает фенотипом ТК-, не способен к размножению на перевиваемой линии клеток Н-143ТК- в присутствии 25 мкг/мл бромдезоксиуридина.
Патогенность для животных.
Исследование свойств штамма VR18 на лабораторных животных показало, что в стандартных тестах на токсичность, безвредность, некротическую активность и специфичность рекомбинантный штамм VR18 не отличается от исходного штамма ЛИВП.
Иммунобиологические свойства штамма VR18.
Штамм VR18 высокоиммуногенен, он вызывает гуморальный (антительный) и Т-клеточный иммунные ответы против HBsAg и ВИЧ при иммунизации мышей линии BALB/c в дозе 1•106 БОЕ/животное. Антитела определяют методом иммуноферментного анализа, а клеточный иммунный ответ - в реакции бласттрансформации спленоцитов.
Основным отличием штамма VR18 от ЛИВП является способность к синтезу химерного белка TBI-HBsAg при инфицировании культур клеток млекопитающих, чувствительных к ВОВ (например, CV-1). Индикацию экспрессии осуществляют с помощью метода иммуноферментного анализа.
Штамм описанного рекомбинанта VR18 депонирован в Коллекции культур микроорганизмов Государственного научного центра вирусологии и биотехнологии "Вектор" за номером V-312 от 06.10.2000 г.
Исследование экспрессии химерного белка TBI-HBsAg проводилось в клетках животных CV-1 и Н-143ТК- при инфицировании их рекомбинантным ВОВ VR18, полученным из вышеуказанных компонентов. Нуклеотидная последовательность встроенного в геном ВОВ химерного гена TBI-HBsAg была подтверждена секвенированием. Кроме того, было проведено изучение Т- и В- клеточного иммунного ответа у мышей линии BALB/c, иммунизированных рекомбинантным BOB VR18.
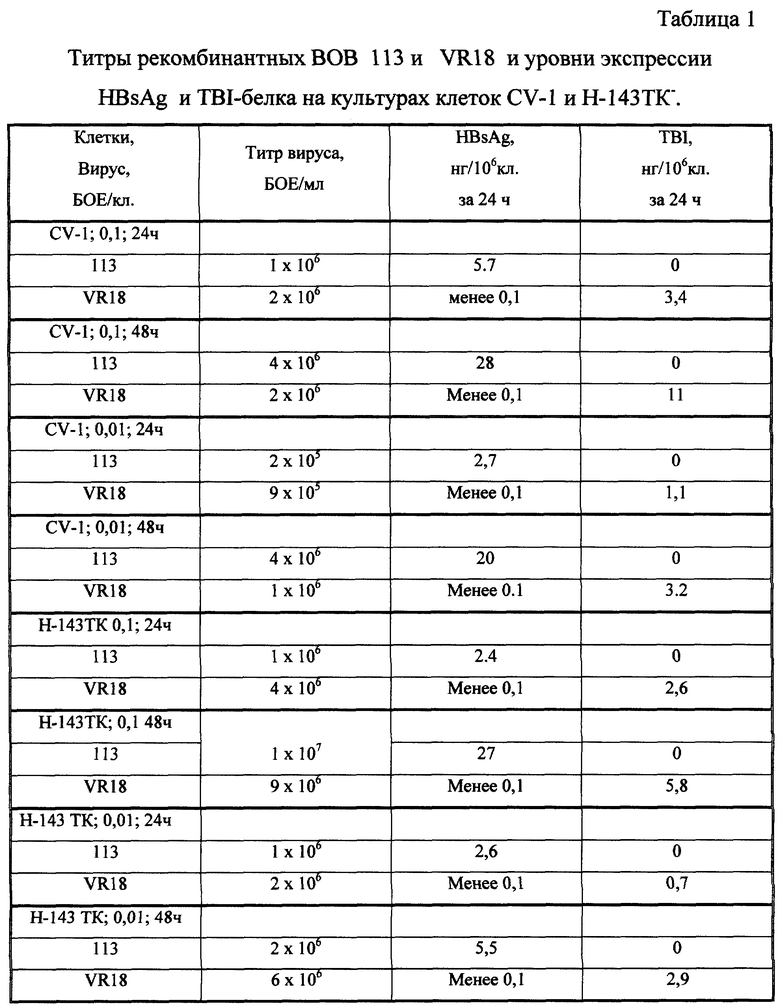
Определение количества HBsAg в образцах проводили методом иммуноферментного анализа. При этом продукция HBsAg достигала 28-27 нг за 24 часа в пересчете на миллион клеток CV-1 или Н-143ТК (множественность заражения 0,1 БОЕ/клетку; время культивирования - 48 часов; см. табл. 1).
Определение количества TBI-белка проводили методом конкурентного иммуноферментного анализа с использованием мышиной сыворотки против TBI-белка бактериального происхождения. Продукция TBI-белка достигала 11 нг и 5,8 нг за 24 часа в пересчете на миллион клеток CV-1 и Н-143 ТК соответственно (множественность заражения 0,1 БОЕ/клетку; время культивирования - 48 часов; см. табл. 1).
Попытки выявить HBsAg в инфицированных культурах клеток в составе химерного белка с использованием коммерческой тест-системы успехом не увенчались. Независимо от типа клеток, множественности заражения и времени инкубации HBsAg тест-системой не выявлялся. Это говорит о том, что антигенная детерминанта "а" HBsAg, по которой работает использованная тест-система, в составе химерного белка экранирована встроенным вместо пре-S2-области синтетическим TBI-белком. Можно предположить, что при вакцинации животных такой рекомбинантной вакциной будет несколько понижен гуморальный иммунный ответ на HBsAg и повышен на TBI-белок. Эксперименты по иммунизации мышей линии BALB/c подтвердили это предположение (см. табл. 2). У животных, иммунизированных препаратом вируса 113 (препарат рекомбинантного ВОВ, содержащий полный ген HBsAg с пре-S2-областью), формируются антитела только к HBsAg. Напротив, у животных, иммунизированных рекомбинантным BOB VR18, формируются антитела прежде всего к антигену ВИЧ (HIV) на всех сроках наблюдения и незначительно к HBsAg, что отмечается только на 35 сутки.
Таким образом, сущность изобретения заключается в том, что сконструирован рекомбинантный ВОВ, экспрессирующий химерный белок HBsAg, в котором пре-S2-область заменена на синтетический белок TBI, содержащий девять Т- и В-клеточных эпитопов главных структурных белков вируса HIV-1. Молекулы HBsAg способны собираться в вирусоподобные частицы, обладающие высокой иммуногенностью, в которых пре-S2-область экспонирована на поверхности. При замене указанной области на синтетический ген белка-кандидата в вакцины против СПИД/ВИЧ в химерном белке на поверхности вирусоподобных частиц экспонируются молекулы TBI, что позволяет получить высокий уровень иммунного ответа. Последнее было продемонстрировано в экспериментах на животных.
Изобретение иллюстрируется следующими чертежами:
Фиг. 1. Нуклеотидная последовательность искусственного гена химерного белка TBI-HBsAg. Инициирующий и терминирующий триплеты трансляции химерного белка обведены рамкой. Отмечены районы генов TBI, HBsAg и стыкующего их линкера (последовательность выделена жирным шрифтом). Обозначены сайты гидролиза для некоторых рестриктаз.
Фиг. 2. Рекомбинантная плазмидная ДНК рTBI-HDsAg и схема ее конструирования.
Фиг.3. Схема стыковки нуклеотидных кодирующих последовательностей белков TBI и HBsAg между собой и сайтом инициации трансляции векторной плазмиды p7.5S113 с помощью синтетических олигонуклеотидов (выделены жирным шрифтом). Липкие одноцепочечные концы, образованные после гидролиза соответствующими рестриктазами, названия которых приведены сверху или снизу последовательности, обозначены ломаными линиями.
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование рекомбинантной плазмидной ДНК р18.
Для конструирования целевой плазмиды p TBI-HBsAg используют плазмиду p7,5S113 [16], которая получена на основе вектора pVAR15 [17] путем встройки гена HBsAg под контроль промотора Р7.5К. В качестве источника гена TBI используют плазмиду pUCSII [15]. 30 мкг указанной плазмиды расщепляют совместным гидролизом рестриктазами PstI и Крn21, полученный фрагмент ДНК, кодирующий TBI, выделяют методом электрофореза на агарозном геле с последующей элюцией на диализную мембрану и лигируют в стандартных условиях с гидролизованной рестриктазой BamHI и обработанной фосфатазой плазмидой p7,5S113 и добавленными синтетическими нуклеотидными адаптерами (фиг.2, 3). Лигазной смесью трансформируют компетентные клетки штамма Е. coli JM103 и из клонов, выросших на среде с ампициллином, выделяют плазмидную ДНК и подвергают рестрикционному анализу с помощью ферментов PstI, EcoRI, Xhol, BamHI. Соответствие последовательности нуклеотидов встройки в отобранной рекомбинантной плазмиде теоретической проверяют методом Максама-Гилберта (фиг.1). Полученная плазмида обозначена р TBI-HBsAg.
Пример 2. Получение рекомбинантного вируса осповакцины VR18.
Культуральные сосуды с монослоем клеток почек зеленой мартышки CV-1 заражают вирусом осповакцины штамма ЛИВП с множественностью 0,05-0,1 БОЕ/клетку. Адсорбцию вируса на клетках проводят в течение 1 ч при 37oС, затем удаляют неадсорбированный вирус и клетки промывают дважды культуральной средой ДМЕМ(м), не содержащей сыворотки. К плазмидной ДНК р18 (0,05 мг/мл, 25 мкл) добавляют равный объем липофектина (0,2 мг/мл) и приготовленную смесь оставляют на 15 мин при комнатной температуре для формирования липосом. Сформировавшиеся липосомы разбавляют 1 мл среды ДМЕМ(м) без сыворотки и переносят на подготовленные клетки. Через 16 ч инкубации при 37oС к клеткам добавляют 4 мл среды ДМЕМ(м), содержащей 2% эмбриональной сыворотки и 80 ед/мл гентамицина и инкубацию продолжают в течение 24 ч. Клетки после трансфекции замораживают, оттаивают и клонируют вирусное потомство на клетках перевиваемой культуры Н-143ТК- под агаровым покрытием, содержащим 25 мкг/мл 5-бромдезоксиуридина. Через 48 ч инкубации при 37oС в атмосфере, содержащей 5% СО2, наносят второе покрытие, содержащее 0,01% красителя нейтрального красного. Окрашивание монослоя клеток для визуализации вирусных бляшек проводят в течение 3 ч при 37oС в атмосфере, содержащей 5% СО2. Вирусные клоны из отдельных бляшек подращивают в 24-луночных планшетах и анализируют методом ДНК-ДНК дот-гибридизации на наличие чужеродной последовательности. Положительные по гибридизации клоны повторно клонируют и препаративно нарабатывают на культуре клеток CV-1.
Пример 3. Определение уровня экспрессии химерного белка TBI-HBsAg.
Монослой клеток CV-1 или Н-143ТК- в лунках 6-луночных планшетов инфицируют рекомбинантными вирусами осповакцины 113 (содержит ген HBsAg с пре-S2-областью) и VR18 (содержит химерный ген TBI-HBsAg) с множественностью 0,1 или 0,01 БОЕ/клетку. Инфицированные клетки инкубируют в течение 24 или 48 часов при 37oС в атмосфере, содержащей 5% СО2. Во всех экспериментах на культурах клеток используют культуральную среду ДМЕМ(м), содержащую 2% эмбриональной сыворотки и по 100 мкг/мл пенициллина и стрептомицина. После окончания инкубации клетки разрушают двукратным замораживанием-оттаиванием. Клеточный дебрис осаждают центрифугированием при 600g в течение 10 мин. Супернатанты используют для определения количества HBsAg и TBI- белка методами иммуноферментного анализа.
Определение количества HBsAg в образцах проводят методом иммуноферментного анализа по калибровке, сделанной по разведениям препарата HBsAg с известной концентрацией, с использованием коммерческой тест-системы (ЗАО "Вектор-Бест", п. Кольцово). Определение количества TBI-белка в образцах проводят методом конкурентного иммуноферментного анализа с использованием мышиной сыворотки против TBI-белка бактериального происхождения.
Результаты определения титров рекомбинантных вирусов и уровни экспресии HBsAg и TBI-белка на культурах клеток CV-1 и Н-143ТК-представлены в таблице 1. Как видно, выходы обоих рекомбинантных вирусов сравнимы для клеток CV-1 и Н-143ТК- и различаются в 2-10 раз в зависимости от множественности заражения и времени инкубации клеток после их инфицирования. Уровни экспрессии HBsAg для рекомбинантного вируса 113 и TBI-белка для рекомбинантного вируса VR18 хорошо коррелируют с титрами соответствующих вирусов. Продукция HBsAg достигает 28-27 нг за 24 ч в пересчете на миллион клеток CV-1 или Н-143ТК- (множественность заражения 0,1 БОЕ/клетку; время культивирования - 48 ч). Продукция TBI-белка достигает 11 и 5,8 нг за 24 ч в пересчете на миллион клеток CV-1 и Н-143ТК- соответственно (множественность заражения 0,1 БОЕ/клетку; время культивирования - 48 ч).
Попытки выявить HBsAg в составе химерного белка с использованием коммерческой тест-системы успехом не увенчались. По-видимому, антигенная детерминанта "а" HBsAg, по которой работает тест-система, в составе химерного белка экранирована встроенным TBI-белком. Основываясь на этих данных, предполагаем, что при вакцинации животных рекомбинантным вирусом VR18 будет несколько понижен гуморальный иммунный ответ на HBsAg и повышен на TBI-белок. Последующие эксперименты по иммунизации мышей линии BALB/c подтвердили это предположение (см. табл. 2).
Пример 4. Изучение иммуногенных свойств рекомбинантного вируса VR18.
В эксперименте использовались мыши линии BALB/c (галотип Н-2b) массой 12 граммов в количестве 30 голов, которые содержались на стандартном рационе.
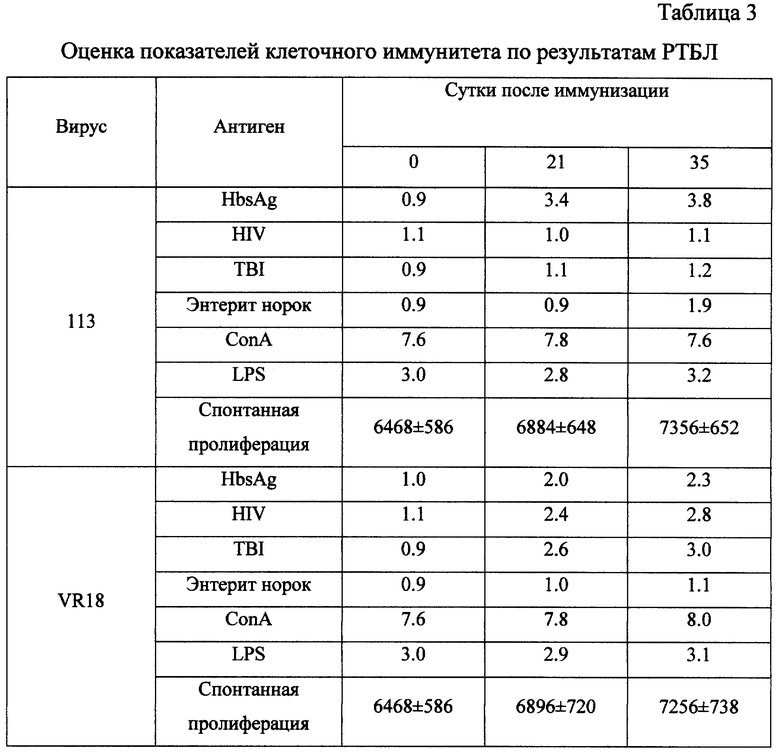
Изучение иммуногенных свойств препаратов рекомбинантных вирусов 113 и VR18 проводилось на 0, 21 и 35 сутки после иммунизации (животных разделили на 2 группы). Иммунизацию проводили подкожно в корень хвоста (0,1 мл) дозой 10 БОЕ. На указанные сроки изучались показатели гуморального иммунитета в реакции иммуноферментного анализа с использованием стрипов с сорбированными антигенами ВИЧ и HBsAg и видоспецифического конъюгата иммуноглобулинов кролика против IgG мыши фирмы "Sigma".
Как видно из данных, приведенных в таблице 2, у животных, иммунизированных препаратом вируса 113, формируются антитела только к HBsAg. Напротив, у животных, иммунизированных препаратом вируса VR18, формируются антитела прежде всего к антигену HIV (на всех сроках наблюдения) и незначительно к HBsAg, что отмечается только на 35 сутки.
Показатели клеточного иммунитета изучались в реакции бласттрансформации спленоцитов. Для проведения этой реакции из селезенок животных экспериментальных групп выделяют клетки, которые двукратно отмывают в среде культивирования (среда RPMI 1640 с 2 mM L-глугамина, 5•10-5 М 2-меркаптоэтанола, 80 мг/л гентамицина, 5% прогретой при 56oС эмбриональной сывороткой). После этого концентрацию спленоцитов доводят до 2 млн/мл и используют для реакции бласттрансформации в 96-луночных планшетах. В лунки добавляют по 2•105 клеток в объеме 100 мкл. В качестве антигенов используют:
- лизат вируса HIV в концентрации 2 мкг/мл;
- белок TBI в концентрации 2 мкг/мл;
- инактивированный антиген вируса энтерита норок в концентрации 2 мкг/мл.
В качестве митогенов используют конканавалин A ("Sigma") в концентрации 10 мкг/мл и липополисахарид E.coli (серотип 055:В5, "Sigma") в концентрации 25 мкг/мл. Антигены и митогены добавляют в объеме 100 мкл в лунки с клетками, используя на каждый антиген или митоген по 4 лунки.
Спленоциты, стимулированные антигенами, культивируют в течение 96 ч, а стимулированные митогенами - 78 ч. За 18 ч до окончания культивирования добавляют 3H-тимидин в концентрации 2 мкКи на лунку. По окончании срока культивирования клетки замораживают, оттаивают и проводят сбор на "Cell-Hawester" ("Flow" Великобритания). Счет радиоактивности проводят в толуольном сцинтилляторе на счетчике "Mark III". Пролиферативную активность спленоцитов оценивают по абсолютным числам стимулированных и нестимулированных спленоцитов в динамике наблюдения и индексу стимуляции, определяемому как отношение счета стимулированных спленоцитов к счету нестимулированных спленоцитов.
Как следует из результатов, представленных в табл. 3, у животных, иммунизированных вирусом 113, на 21 и 35 сутки после иммунизации отмечается выраженная пролиферация спленоцитов in vitro при стимуляции белком HBsAg, пролиферативный ответ на стимуляцию белками HIV и вируса энтерита норок отсутствует. Это является подтверждением специфической пролиферации на HBsAg и свидетельствует о наличии гена этого белка в препарате вируса 113. У животных, иммунизированных препаратом сконструированного вируса VR18, на протяжении всей динамики наблюдения - 21 и 35 сутки - отмечается пролиферативная активность спленоцитов в ответ на стимуляцию как белками HIV, так и HBsAg. Этот показатель достоверно превосходит аналогичный показатель у интактных животных (0 сутки). Специфичность реакции спленоцитов этой группы подтверждается отсутствием выраженной пролиферации при стимуляции антигеном энтерита норок.
Таким образом, получен новый рекомбинантный штамм вируса осповакцины, экспрессирующий химерный белок HBsAg, в котором пре-S2-область заменена на синтетический белок TBI, содержащий протективные Т- и В-клеточные эпитопы вируса иммунодефицита человека типа 1, и вызывающий иммунный ответ против вирусов ВИЧ и гепатита В в организме животных. Полученный рекомбинантный вирус может быть использован в качестве кандидата для создания на его основе поливалентной вакцины против ВИЧ-1, вируса гепатита В человека и ортопоксвирусов. Ожидается, что полученная на основе такого подхода вакцина будет характеризоваться высокой иммуногенностью одновременно против трех указанных выше вирусных инфекционных заболеваний, а по отношению к ВИЧ-1, близостью конформаций В-эпитопов в вакцине и вирусном антигене, эффективностью в гетерогенной по HLA-антигенам человеческой популяции, стимуляцией ответа к разным В-эпитопам, отсутствием нежелательных эпитопов белков ВИЧ-1, а также возможностью оптимизации протективных свойств за счет изменения состава и расположения Т- и В-детерминант и технологичностью получения.
Список литературы
1. Szmuness W. // Prog. Med. Virol., 1978, v.24, p.40-69.
2. Козлов А.П. // Русский журнал ВИЧ/СПИД и родственные проблемы, 2000, т.4, 1, стр.11-14.
3. Moss В., and Flexner С. // Annu. Rev. Immunol., 1987, v.5, p.305.
4. Moss B. // Seminars in Virology, 1992, v.3, p.277.
5. Falkner F.G., et al. // Virology, 1988, v.164, p.450.
6. Flexner C., et al. // Virology, 1988, v.166, p.339.
7. Takahashi H. , et al. // Proc. Natl. Acad. Sci. USA, 1988, v.85, p. 3105.
8. Walker B.D., et al. // Science, 1988, v.240, p.64.
9. Willey R.L., et al. // J. Virol., 1988, v.62, p. 139.
10. Heerman K.H., et al. // J. Virol., 1984, v.52, p.396.
11. Prange R., et al. // J. Gen. Virol., 1995, v.76, p.2131.
12. Chiba M. // Arch. Virol., 1999, v.144, p.1469.
13. Berzofsky J.A., et al. // Curr. Top. Microbiol. Immunol., 1986, v. 130, p.13.
14. Eroshkin A.M., et al. // Protein Eng., 1993, v.6, p.997.
15. Eroshkin A.M., et al. // Protein Eng., 1995, v.8, 2, p. 167.
16. Беляев А.С. и др. // Докл. АН СССР, 1990, т.314, стр.488.
17. Фодор И.И. и др. // Биотехнология, 1987, т.3, стр.302.


Изобретение относится к биотехнологии, в частности к генной инженерии. Изобретение представляет собой штамм рекомбинантного вируса осповакцины и рекомбинантную плазмидную ДНК pTBI-HBsAg. Рекомбинантная плазмидная ДНК кодирует девять иммуногенных эпитопов двух основных структурных белков вируса иммунодефицита человека типа-1. Рекомбинантный штамм вируса осповакцины, модифицированный плазмидой pTBI-HBsAg, вызывает Т- и В-клеточный иммунный ответ против HBsAg и ВИЧ-1 у вакцинированных им животных. Штамм может быть использован в качестве основы для поливалентной вакцины против ВИЧ-1, вируса гепатита В человека и ортопоксвирусов. 1 з.п.ф-лы, 3 ил., 3 табл.
BamHI-ClaI фрагмент генома вируса гепатита В, кодирующий поверхностный вирионный белок S (HBsAg), размером 1226 п. н. ; генетические маркеры: bla-ген ампициллинрезистентности - ген β-лактамазы, определяющий устойчивость к ампициллину при трансформации клеток E. coli; TК - прерванный ген тимидинкиназы вируса осповакцины, обуславливающий ТК фенотип при рекомбинационной встройке в геном ТК вируса осповакцины; уникальные сайты рестрикции: HindIII - положение 1; KpnI - 1303; Kpn21 - 1453; XhoI - 1462.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ HBS AG ВИРУСА ГЕПАТИТА B, И ШТАММ ВИРУСА ОСПОВАКЦИНЫ - ПРОДУЦЕНТ HBS AG ВИРУСА ГЕПАТИТА B | 1988 |
|
SU1515698A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PVAX2 ДЛЯ ПЕРЕНОСА И ЭКСПРЕССИИ В ГЕНОМЕ ВИРУСА ОСПОВАКЦИНЫ ГЕНА ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В И ШТАММ ВИРУСА ОСПОВАКЦИНЫ, ЭКСПРЕССИРУЮЩИЙ ПОВЕРХНОСТНЫЙ АНТИГЕН ВИРУСА ГЕПАТИТА В | 1985 |
|
SU1354709A1 |
| RU 209148927, 27.09.1997 | |||
| ЖИВАЯ РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ГЕПАТИТА В И НАТУРАЛЬНОЙ ОСПЫ ДЛЯ НАКОЖНОГО ПРИМЕНЕНИЯ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 1992 |
|
RU2073524C1 |

