Настоящее изобретение касается связывающих членов, особенно молекул антитела, которые ингибируют биологические эффекты IL-6. Связывающие члены полезны для лечения заболеваний, связанных с IL-6, включая воспалительные болезни и опухоли.
Интерлейкин 6 (IL-6) представляет собой 26 кДа плейотропный провоспалительный цитокин, продуцируемый рядом типов клеток, включая стимулируемые фибробласты, моноциты и эндотелиальные клетки, которые формируют главный источник IL-6 in vivo. Клетки, такие как Т клетки, В клетки, макрофаги, кератиноциты, остеобласты и несколько других, могут продуцировать IL-6 при стимуляции. IL-6 также экспрессируется из опухолевых клеточных линий и опухолевых клеток, например клеток из карциномы легкого, рака простаты, миеломы, гипернефромы и сердечной миксомы [1, 2]. При невоспалительных состояниях IL-6 секретируется из жировой ткани [3]. Регулирование экспрессии IL-6 зависит от типа клетки, которая его продуцирует. В клетках множественной миеломы IL-6, по-видимому, действует в положительной обратной петле стимулирование роста клеток, а также продуцирование больше IL-6 [4, 5]. В других типах клеток IL-6, по-видимому, ингибирует рост и активацию клеток и может действовать как отрицательный регулятор для некоторых провоспалительных цитокинов.
Чтобы инициировать передачу сигналов клетки, IL-6 связывается с низкой аффиностью с трансмембранным рецептором, рецептором альфа IL-6 (также обозначается как IL-6Rα, IL-6Ra, IL-6R, gp80 или CD126), чтобы сформировать комплекс "IL-6:IL-6Ra". Этот комплекс связывается с gp130 сигнальным рецептором; IL-6Rα и gp130 вместе формируют высокоаффинный IL-6 связывающий участок и индуцируют формирование гексамера, состоящего из двух копий каждого из IL-6, IL-6Ra и gp130 [6]. Трансмембранные и цитоплазматические домены IL-6Ra не требуются для трансдукции сигнала, поскольку IL-6Ra также существует в виде растворимой секретируемой форме (sIL-6R или sIL-6Ra). Растворимый рецептор продуцируется или путем дифференциального сплайсинга сообщения IL-6Ra или протеолитического слущивания. SIL-6R способен к формированию комплекса рецептор-лиганда с IL-6, "IL-6:sIL-6Ra". Этот комплекс может связывать gp130 на клетках и таким образом инициирует клеточную передачу сигналов в gp130 положительных клетках, даже если эти клетки не экспрессируют IL-6Ra. Таким образом, SIL-6R имеет потенциал для расширения диапазона клеток, чувствительных к IL-6, и, как считают, играет важную роль в IL-6-опосредованном воспалении [7].
Была объяснена кристаллическая структура человеческого лиганда IL-6 [6]. Кристаллическая структура внеклеточного домена человеческого IL-6Ra [8] и гексамерная структура IL-6/IL- 6R/gp130 комплекса [9] были также выявлены. Эти структуры, объединенные с исследованиями мутагенеза, идентифицировали три участка на поверхности IL-6, которые вовлечены в функциональную активность IL-6 в комплексе с различными компонентами рецептора. Остатки участка 1 вовлечены во взаимодействие между IL-6 и IL-6Ra. Остатки участка 2 вовлечены во взаимодействие между IL-6 и gp130 цитокин-связывающим доменом. Остатки в участке 3 из IL-6 вовлечены во взаимодействие с Ig-подобным доменом второго gp130 в гексамерном комплексе. Был также идентифицирован четвертый участок на IL-6, где IL-6 взаимодействует со второй молекулой IL-6 в гексамерном IL-6/IL-6R/gp130 комплексе [10].
Был выделен ряд лиганд анти-IL-6 моноклональных антител. Проводились исследования картирования, которые показывают, что они связываются с различными связывающими участками, описанными выше, на поверхности человеческого IL-6 [11, 12, 13, 14, 15].
Были также получены многие анти-IL-6Ra моноклональные антитела, и их связывающие участки на IL-6Ra картированы [16, 14, 15, 17].
IL-6 принадлежит к семейству цитокинов, которое включает Интерлейкин 11 (IL-11), цилиарный нейротрофный фактор (CNTF), Онкостатин М (OsM), фактор, ингибирующий лейкоз (LIF), кардиотрофин-подобный цитокин (CLC) и Кардиотрофин 1 (СТ-1). У каждого из членов этого семейства есть их собственные специфичные альфа-субъединицы рецептора, и они формируют комплексы с общей рецепторной субъединицей gp130. Нацеленное разрушение gp130 гена является эмбрионально летальным [18, 19]. Все члены семейства IL-6 могут индуцировать экспрессию белков острой фазы из гепатоцитов.
Передача сигналов IL-6 вовлекает фосфорилирование тирозина киназами семейства JAK, и последующую активацию двух главных внутриклеточных сигнальных каскадов, SHP2/ERK МАРК и STAT1/3 пути, приводя к генной экспрессии через NF-IL-6 и АР 1 [18, 20].
IL-6 показывает широкий спектр биологических функций, включая гематопоэз, индукцию ответов острой фазы, активацию Т клеток, стимуляцию секреции антител, защиту хозяина от инфекции, активацию клеток миеломы и остеокластов [21, 22]. Обзор эффектов IL-6 см [23]. IL-6 был первоначально идентифицирован в качестве фактора дифференцирования В-клеток, произведенных Т клетками [24], но был впоследствии идентифицирован как мощный активатор и рост-промотирующий фактор многих типов клеток. Он индуцирует заключительное созревание В клеток в антитело-продуцирующие клетки и является существенным дополнительным фактором для активации и пролиферации Т клеток. Исследования показали, что IL-6 вовлечен в активацию автореактивных Т лимфоцитов и пролиферацию и дифференцирование цитотоксических Т клеток. IL-6 был вовлечен в гематопоэз как кофактор, вызывающий активацию и дифференцирование гематопоэтических стволовых клеток. Эффект IL-6 на ответ острой фазы также хорошо задокументирован [25]. IL-6 индуцирует множество белков острой фазы, включая фибриноген, альфа-анти-химотрипсин, сывороточный амилоидный А и С-реактивный белок из человеческих гепатоцитов. Белки острой фазы контролируют иммунные ответы и воспаление и оказывают воздействие на ремоделлирование ткани. Сывороточный уровень IL-6 хорошо коррелирует с уровнем С-реактивного белка при разных патологиях, что свидетельствует о причинной роли IL-6 в ответе острой фазы. IL-6, как так же показано, продуцируется остеобластами и, по-видимому, вовлечен в активацию остеокластов и резорбцию костей [26, 27, 28]. Как это ни парадоксально, было предложено, что IL-6 действует не только как провоспалительный цитокин, но и может также, при определенных обстоятельствах и типах клеток, ослаблять эффекты других провоспалительных цитокинов, что приводит к снижению воспаления.
Поскольку у IL-6 есть множество биологических эффектов, повышение уровня IL-6 рассматривается как ключевой цитокин при множестве показаний болезней. Уровни циркулирующего IL-6, как показано, повышаются при болезнях, таких как ревматоидный артрит, болезнь Кастельмана, Ювенильный идиопатический артрит и Болезнь Крона [29]. Из-за этого IL-6 был вовлечен в ведение патологии при этих воспалительных показаниях. Кроме того, множество типов опухоли, как было показано, стимулировалось IL-6, включая меланому, почечно-клеточную карциному, саркому Капоши, карциному яичника, лимфому, лейкемию, множественную миелому и карциному простаты [30]. Кроме того, при нескольких типах рака сообщалось об увеличении циркулирующих уровней IL-6. При некоторых показаниях рака повышенные уровни IL-6 использовались в качестве прогностических индикаторов болезни.
Из-за роли IL-6 при болезни в качестве потенциальных методов лечения был разработан ряд мышиных и химерных античеловеческих IL-6 моноклональных антител.
US5856135 описывает измененное человеческое антитело к IL-6, полученное из мышиного моноклонального антитела "SK2".
JP-10-66582 сообщает о химерном антителе к IL-6, которое обозначается как распознавание спирального D участка IL-6 (участок 1).
WO2004/020633 (ЕР1536012) описывает молекулу человеческого scFv антитела к IL-6, выделенную с использованием технологии фагового дисплея. scFv, как сообщается, имеет аффинность, равную 13 нМ.
Мышиное антитело анти-IL-6, элсилимомаб (также известен как В-Е8) использовалось для лечения пациентов с Множественной миеломой [31, 32], почечно-клеточной карциномой [33] и ревматоидным артритом [34], и при определенных диагностических маркерах отмечались улучшения у леченых пациентов со всеми тремя болезнями. ВЕ-8 также использовался для лечения ВИЧ-положительных пациентов с иммунобластной или полиморфной крупноклеточной лимфомой [35] с облегчением системных симптомов (то есть лихорадки, потоотделения, кахексии) и супрессией спонтанного роста лимфомы приблизительно у 50% пациентов.
Однако быстрое выведение этого антитела из организма и возможные анафилактические реакции из-за продукции человеческих антимышиных антител (НАМА) к элсилимомабу ограничили его использование в клинической практике [36].
В целом, клиническое использование мышиных моноклональных антител ограничено, так как антитела часто вызывают НАМА. Часто продуцируются НАМА, направленные против части Fc иммуноглобулина мыши, что приводит к быстрому клиренсу анти-IL-6 mAb и возможному возникновению анафилактической реакции [36].
Также известно, что фармакокинетика антител мыши у людей отличается от человеческих антител, имеющих более короткие периоды полувыведения и увеличенные скорости клиренса.
Чтобы уменьшить иммуногенность мышиных антител у людей, были сконструированы химерные антитела с мышиными вариабельными участками и человеческими константными участками. Химерное человек-мышь антитело анти-IL-6 cCLB8 (известное как CNTO 328) использовалось для лечения пациентов с множественной миеломой [5, 37], при этом стабилизация болезни отмечалась у большинства пациентов.
Однако, хотя химерные антитела менее иммуногенны, чем мышиные MAbs, сообщалось об ответах человеческого антихимерного антитела (HACAs) [38].
Были выполнены исследования картирования cCLB8, которые показывают, что это ингибитор участка I активности IL-6. Brakenhoff и соавт. [39] продемонстрировали, что cCLB8 связывается с мутантами IL-6 аминотерминальной делеции Pro46, Ser49, Glu51, Ile53, Asp54 и также связывается с мутантами делеции Asp62
и Met77 (хотя при сниженной аффинности). Те же самые авторы показывают, что cCLB8 ингибирует IL-6 дикого типа, но не C-терминальную делецию 5 в исследовании В9-клеточной пролиферации и что cCLB8 не будет связывать IL-6 del С-4, у которого удалены последние 4 С-терминальные аминокислотные остатки. Эти данные свидетельствуют, что cCLB8 связывается с эпитопом, включающим С-терминальные остатки IL-6.
Kalai и соавт. [17] продемонстрировали, что cCLB8 был не в состоянии распознавать IL-6 мутанты F106E, F102E/F106E или R207E/R210E. Однако антитело действительно распознает IL-6 мутанты R207E и R207W. Связывание cCLB8 с мутантами R207W & R207E составляет приблизительно 50% от связывания по сравнению с диким типом, что свидетельствует о том, что остатки F106 и R210 вовлечены в CCLB8 связывающий эпитоп и остаток R207 вовлечен в связывание, но имеют меньший эффект, чем остатки F106 и R210. cCLB8 связывает мутанты IL-6 участка - I R196M, K199N/Q203L и Q203L со 100% активностью по сравнению с диким типом. Brakenhoff и соавт. [13] продемонстрировали, что cCLB8 связывает следующие варианты IL-6; Q182H, N183K, W185Q, W185G, W185R, Т190Р, Q182H/Q184P, W185R/S197N, Q187E/T190P, I164L/L186R/M189I, что не является удивительным, так как большинство из них являются дистально отделенными от остатка IL-6 участка 1.
Положительный эффект ингибирования IL-6 сигнального пути при раке и воспалительных заболеваниях был в дальнейшем подчеркнут при помощи гуманизированного анти-IL-6Ra антитела Тоцилизумаб (так же известном, как hPM-1, MRA и Actemra). Это гуманизированная версия мышиного анти-IL6Ra антитела РМ-1. Лечение пациентов с этим антителом оказалось эффективным при ряде болезней, включая ревматоидный артрит, Ювенильный идиопатический артрит, болезнь Крона, миелопролиферативное расстройство, болезнь Кастельмана и Системную красную волчанку [40].
Мы преуспели в выделении очень мощных, высокоаффинных связывающих членов для IL-6. Вследствие их высокой аффинности и мощности и их результата в функциональных исследованиях, описанного в настоящей заявке, связывающие члены согласно изобретению особенно подходят для использования в терапевтическом и/или диагностическом лечении организма животных или человека.
Связывающие члены полезны для лечения расстройств, связанных с IL-6, как подробно описано в настоящей заявке далее.
Человеческое анти-IL-6 антитело для лечения воспалительных заболеваний и рака обеспечивает существенные преимущества перед существующими подходами. Например, человеческие антитела не индуцируют ответы НАМА или НАСА и имеют более длительный период полувыведения in vivo по сравнению с нечеловеческими или химерными антителами.
Мы также установили, что связывающие члены для IL-6 предлагают существенные преимущества по сравнению со связывающими членами для IL-6Ra, особенно с точки зрения введения in vivo и лечения, как описано в настоящей заявке далее.
Как описано более подробно в Примерах, мы выделили родительскую молекулу антитела, обозначенную CAN022D10, с набором последовательностей CDR, как показано в Таблице 7. Через процесс оптимизации мы получили группу клонов антитела: Антитела 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23 с последовательностями CDR, полученными из родительских последовательностей CDR и имеющих замены в положениях, обозначенных в Таблице 7.
Таким образом, например, можно отметить из Таблицы 7, что у Антитела 2 есть родительская последовательность HCDR1, в которой остаток Kabat 35 заменен на Thr (SEQ ID NO:13). Антитела 14 и 22 содержат дополнительный остаток, то есть аминокислотную инсерцию, в HCDR3: Ilе на остатке Kabat 100D, которая не присутствует в родительской последовательности HCDR3 SEQ ID NO:5. Антитела 7, 8, 10, 16-19, 21 и 23 не содержат остаток Kabat 95 в LCDR3, тогда как родительский LCDR3 (SEQ ID NO:10) включает Pro в остатке Kabat 95. У родительских HCDR3 и HCDR3 последовательностей всех антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23 есть Trp в остатке Kabat 95 и Asp в остатке Kabat 101, что указывает на то, что Н95 Trp и Н101 Asp могут способствовать связыванию и/или силе действия для IL-6 в связывающих членах изобретения.
Домен VH, домен VL и последовательности CDR родительского антитела CAN022D10, и антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, описанных в настоящей заявке, показаны в приложенном списке последовательностей.
Как описано более подробно ниже, связывающие члены согласно изобретению, как показано, нейтрализовали IL-6 с высокой силой действия. Нейтрализация означает ингибирование биологической активности IL-6. Связывающие члены согласно изобретению могут нейтрализовать одну или несколько активностей IL-6. Ингибируемая биологическая активность типично представляет собой IL-6 связывание с одним или несколькими его связывающими партнерами. Например, ингибируемой биологической активностью может быть связывание IL-6 с трансмембранным и/или растворимым IL-6Ra. Это продемонстрировано в следующих испытаниях, которые кратко описаны в настоящей заявке и более подробно ниже: испытание TF-1 показывает, что связывающие члены согласно изобретению ингибируют IL-6 связывание с мембранным IL-6Ra, поскольку клетки TF-1, по-видимому, не продуцируют растворимый IL-6Ra. Также, поэтому связывающие члены согласно изобретению ингибируют IL-6 связывание с мембранным рецептором. В испытании синовиальных фибробластов связывающие члены согласно изобретению ингибируют IL-6 связывание с растворимым IL-6Ra, так как для того, чтобы оно работало, необходимо включать sIL-6Ra в это испытание. Добавленный IL-1бета вызывает выработку эндогенного IL-6, который при ингибировании связывающим членом согласно настоящему изобретению предотвращает выработку VEGF.
В соответствии с изобретением, связывание IL-6 человека или нечеловеческого примата, например обезьяны cynomolgus, с IL-6Rα, может быть ингибировано, например, связывающий член может ингибировать связывание зрелого человеческого IL-6 с IL-6Rα.
Ингибирование биологической активности может быть частичным или полным. Связывающие члены могут ингибировать биологическую активность IL-6 на 100%, или по крайней мере 95%, по крайней мере 90%, по крайней мере 85%, по крайней мере 80%, по крайней мере 75%, по крайней мере 70%, по крайней мере 60%, или по крайней мере 50% активности в отсутствии связывающего члена.
Может быть определена нейтрализирующая сила действия связывающего члена. Сила действия обычно выражается как значение IC50, в нМ, если не указано иначе. В функциональных испытаниях IC50 - концентрация связывающего члена, которая уменьшает биологический ответ на 50% от его максимума. В исследованиях связывания лиганда IC50 - концентрация, которая уменьшает формирование комплекса рецептор-лиганд на 50% от максимального уровня специфичного связывания. IC50 можно вычислить путем построения графика % максимального биологического ответа как функции логарифма концентрации связывающего члена и, используя программу, такую как Prism (GraphPad) или Origin (Origin Labs), чтобы приспособить сигмоидальную функцию к данным, для получения значений IC50. Сила действия может быть определена или измерена с использованием одного или нескольких испытаний, известных квалифицированному специалисту и/или описанных либо упомянутых в настоящей заявке.
Нейтрализация активности IL-6 связывающим членом в испытании, описанном в настоящей заявке, например испытании пролиферации TF-1 или других клеточных испытаниях, описанных ниже, указывает, что связывающий член связывает и нейтрализует IL-6. Другие методы, которые могут использоваться для определения связывания связывающего члена с IL-6, включают ELISA, Вестерн блот, иммунопреципитацию, аффинную хроматографию и биохимический анализ.
Связывающие члены, описанные в настоящей заявке, как было продемонстрировано, связываются и нейтрализуют биологические эффекты эндогенного человеческого IL-6, как показано в испытании ингибирования высвобождения VEGF из человеческих синовиальных фибробластов в ответ на эндогенный человеческий IL-6, что сообщается в Примерах 1.7 и 2.7 в настоящей заявке. В этом испытании синовиальные фибробласты от пациентов с ревматоидным артритом продуцируют IL-6 в ответ на стимуляцию с помощью IL-1β и растворимого IL-6Rα, что приводит к индуцированной IL-6 секреции VEGF. 1L-6, выработанный человеческими синовиальными фибробластами, таким образом, представляет собой эндогенный человеческий IL-6. Эндогенный IL-6 - молекулярная мишень для лечения у людей, таким образом, нейтрализация эндогенного IL-6 является важным индикатором терапевтического потенциала связывающих членов. Так как испытание проводилось с синовиальными фибробластами, полученными от пациентов с ревматоидным артритом, результаты особенно значимы для использования связывающих членов для лечения ревматоидного артрита. Нейтрализирующая сила действия оптимизированных молекул антитела, проверенных в испытании высвобождения VEGF, превзошла таковую для известного антитела анти Il-6 CNTO-328.
У связывающего члена согласно изобретению может быть IC50 меньше, чем 50 нМ, например меньше, чем 5 нМ, например меньше, чем 1 нМ в испытании ингибирования высвобождения VEGF из человеческих синовиальных фибробластов, стимулируемых с помощью 0,6 пкМ человеческого IL-1β и 2,4 нМ растворимого человеческого IL-6Rα.
Эндогенный IL-6, как известно, является смесью гликозилированных и негликозилированных форм. Связывание связывающего члена согласно изобретению с эндогенным IL-6 было продемонстрировано в испытании синовиальных фибробластов, так как это испытание использует IL-6 из человеческих синовиальных фибробластов, то есть эндогенный IL-6.
Связывающий член согласно изобретению может ингибировать индуцированную IL-6 пролиферацию клеток TF-1. TF-1 - человеческая предмиелоидная клеточная линия, полученная от пациента с эритролейкемией (Kitamura и соавт. 1989). Клеточная линия TF-1 требует присутствия фактора роста для выживания и пролиферации. Индивидуальные факторы роста, на которые могут ответить клетки TF-1, включают IL-6, GM-CSF и Онкостатин М. Связывающий член согласно изобретению может иметь IC50 меньше, чем 100 нМ, например меньше, чем 20 нМ, 10 нМ или 1 нМ, например меньше, чем 100 пМ, 70 пМ, 50 пМ, 40 пМ, 30 пМ, 20 пМ или 10 пМ, в испытании для ингибирования пролиферации клеток TF-1 в ответ на 20 пМ человеческого IL-6. Как описано в настоящей заявке (см. Пример 1.5), родительский IgG "CAN022D10", как показано, имел IC50 в испытании пролиферации TF-1, составляющую приблизительно 93 нМ, и мы впоследствии получили оптимизированные варианты CAN022D10, имеющие существенно увеличенную силу действия (IC50 вообще меньше, чем 100 пМ), как показано в Примерах 2.2, 2.5 и 2.6 (Таблицы 3, 4 и 5 соответственно). Особенно, измеренные для некоторых из оптимизированных клонов значения IC50 составили всего лишь 5 пМ или меньше, например, зародышевого IgG Антитела 7, Антитела 17 и Антитела 18, представляя чрезвычайно высокую нейтрализирующую силу действия этих антител.
Связывающий член согласно изобретению может ингибировать индуцированную IL-6 пролиферацию клеток В9. Клетки В9 - подклон мышиной В-клеточной линии гибридомы, В13.29, выбранные на основе их специфичного ответа на IL-6. Клетки В9 требуют IL-6 для выживания и пролиферации и отвечают на очень низкие концентрации IL-6. Также, может быть оценена пролиферация этих клеток в присутствии антитела IL-6 и определена аффинность антитела. Пример 2.10 авторов показывает, что Антитело 18 ингибировало пролиферацию клеток В9 в ответ на IL-6 и показало высокую аффинность в этом испытании.
При ревматоидном артрите происходит выработка аутоантитела, главным образом, класса IgM. SKW6.4 - кленовая IgM секретирующая человеческая лимфобластоидная В-клеточная линия.
При стимуляции с помощью IL-6 эти клетки секретируют IgM, таким образом, это испытание было воспринято как имеющее значение для ревматоидного артрита. Клетки SKW6.4 могут использоваться в испытании для определения силы действия связывающих членов для того, чтобы нейтрализовать IL-6 путем определения ингибирования секреции IgM в ответ на IL-6. Связывающий член согласно изобретению может иметь IC50, составляющую меньше, чем 10 пМ, например меньше, чем 5 пМ, в испытании с клетками SKW6.4 ингибирования секреции IgM в ответ на 100 пМ человеческого IL-6. Антитело 18, как показано, нейтрализует эффекты IL-6 в этом испытании - см. Пример 2.11 (Таблица 9).
Изобретение предлагает высокоаффинные связывающие члены для человеческого IL-6. Была также продемонстрирована высокая аффинность к IL-6 от обезьяны cynomolgus. Связывающий член согласно изобретению может связывать человеческий IL-6 и/или IL-6 cynomolgus с KD, составляющей не больше чем 1 нМ, например не больше чем 100 пМ, 50 пМ, 30 пМ или 10 пМ. KD может быть определена, например, с помощью поверхностного плазменного резонанса. BIAcore®. Измерения BIAcore® аффинности описаны в настоящей заявке в Примере 2.9. Характерно, аффинность Антител 7 и 18, как обнаружено, находилась вне предела, измеряемого с использованием инструмента BIAcore®, указывая на значение KD ниже 10 пМ.
Как описано в другом месте в настоящей заявке, поверхностный плазменный резонанс вовлекает прохождение аналита в жидкую фазу через лиганд, прикрепленный к основе, и определение связывания между аналитом и лигандом. Поверхностный плазменный резонанс может, например, быть выполнен посредством пропускания IL-6 в жидкой фазе через связывающий член, прикрепленный к основанию. Данные поверхностного плазменного резонанса могут быть подогнаны к модели данных одновалентного аналита. Константа аффинности Kd может быть вычислена из соотношения констант скорости kd/ka, определенных поверхностным плазменным резонансом с использованием модели данных одновалентного аналита.
Аффинность связывающего члена для IL-16 может альтернативно быть вычислена с помощью анализа Шильда, например, основанном на испытании ингибирования пролиферации клеток TF-1 в ответ на различные концентрации человеческого IL-6. Связывающий член согласно изобретению может иметь аффинность, составляющую меньше, чем 10 пМ, например меньше, чем 1 пМ, как вычислено анализом Шильда. Как сообщается в Примере 2.10 в настоящей заявке, аффинность Антитела 18 для человеческого IL-6 была вычислена как 0,4 пМ с использованием анализа Шильда.
Связывающий член согласно изобретению может (необязательно) не реагировать перекрестно с одним или несколькими или всеми из следующих: фактор, ингибирующий лейкоз (LIF), цилиарный нейротрофный фактор (CNTF), IL-11 или онкостатин М.
Связывающий член согласно изобретению может (необязательно) не реагировать перекрестно с крысиным IL-6, мышиным IL-6 и/или собачьим IL-6.
Перекрестная реактивность связывающих членов для связывания других белков или нечеловеческого IL-6 может быть проверена, например, в испытании флюоресценции с временным разрешением для ингибирования связывания человеческого IL-6 со связывающим членом, иммобилизированным на подложке, таком как испытание конкуренции эпитопа DELFIA®, как описано в Примере 1.6. Например, любой или все из LIF, CNTF, IL-11, онкостатина М, крысиного IL-6 и мышиного IL-6 могут показать отсутствие ингибирования меньше чем 50% ингибирование, или могут иметь IC50, составляющую больше чем 0,5 мМ, или больше чем 1 мМ в испытании флюоресценции с временным разрешением для ингибирования связывания меченого человеческого IL-6 со связывающим членом, иммобилизированным на подложке. Например, любой или все из LIF, CNTF, IL-11, онкостатина М, крысиного IL-6 и мышиного IL-6 могут показать отсутствие ингибирования или могут иметь IC50, по крайней мере 10- или 100-кратно больше, чем таковая для немеченого человеческого IL-6 в испытании флюоресценции с временным разрешением для проверки перекрестной реактивности. В этом испытании меченый зрелый человеческий IL-6 дикого типа используется в конечной концентрации Кd его взаимодействия со связывающим членом.
Связывающий член согласно изобретению может перекрестно реагировать с IL-6 cynomolgus. Перекрестная реактивность может быть определена как ингибирование связывания меченого человеческого IL-6 со связывающим членом, иммобилизированным на подложке, в испытании флюоресценции с временным разрешением, описанном выше. Например, IL-6 cynomolgus может иметь IC50, составляющую меньше, чем 5 нМ, например меньше, чем 2,5 нМ, например, приблизительно 1 нМ, в этом испытании флюоресценции с временным разрешением. У IL-6 Cynomolgus может быть IC50, менее чем отличающаяся в 10 раз, например менее чем 5-кратно отличная, от IC50 немеченого человеческого IL-6 в этом испытании.
Детальный протокол для испытания флюоресценции с временным разрешением для определения перекрестной реактивности предоставлен в разделе Материалы и Методы. Примеры данных перекрестной реактивности, полученных в этом испытании, показаны в Таблице 2 в Примере 1.6.
Как сообщается в Примере 2.8, связывающие члены, описанные в настоящей заявке, показали высокую перекрестную реактивность с IL-6 cynomolgus, и не показали никакой или ограниченную перекрестную реактивность с крысиным, мышиным или собачьим IL-6.
Данные перекрестной реактивности указывают, что связывающие члены, описанные в настоящей заявке, связывают эпитоп на IL-6, который сохранен между IL-6 последовательности человека и cynomolgus, и различен в мышиной, крысиной и собачьей последовательности IL-6 по сравнению с человеческой последовательностью.
Связывающие члены, описанные в настоящей заявке, как полагают, связывают участок "сайта 1" IL-6, который является участком, взаимодействующим с IL-6Rα. Связывающие члены согласно изобретению могут таким образом конкурентно ингибировать связывание IL-6 с IL-6Rα, нейтрализуя таким образом биологические эффекты IL-6, которые опосредуются через IL-6Rα.
Мы исследовали способность одного из антител, описанных в настоящей заявке. Антитела 18, связывать мутантный человеческий IL-6, в котором мутации были спроектированы в остатках сайта 1. Как описано в Примере 3, мы идентифицировали мутации в человеческом IL-6, что привело к снижению связывания Антителом 18, указывая на то, что мутированные остатки были вовлечены в распознавание Антителом 18 и могут являться частью эпитопа на IL-6, связанного этим антителом.
Например, в испытании флюоресценции с временным разрешением для ингибирования связывания меченого человеческого IL-6 дикого типа с Антителом 18, иммобилизированным на подложке, никакого ингибирования не наблюдалось для Arg207Glu мутантного человеческого IL-6 (SEQ ID NO:177), указывая на то, что Антитело 18 связывает человеческий IL-6 в остатке Arg207.
Так как Антитело 18 и Антитела 2, 3, 4, 5, 1, 8, 10, 14, 16, 17, 19, 21, 22 и 23 были все получены из родительского антитела CAN22C10, и все имеют структурно родственные CDR, все эти молекулы антитела, как ожидается, свяжут тот же самый или очень подобный перекрывающийся эпитоп. Соответственно, результаты картирования эпитопа, полученные с Антителом 18, как так же ожидается, будут характерными для CAN22D10 других оптимизированных антител, описанных в настоящей заявке.
Связывающий член согласно изобретению может связывать человеческий IL-6 в Phe102 и/или Ser204. Связывающий член согласно изобретению может также связать человеческий IL-6 в Arg207. Необязательно связывающий член может связать фланкирующие остатки или структурно соседние остатки в молекуле IL-6, в дополнение к связыванию Phe102 и/или Сера 204. По соглашению, нумерация остатков соответствует полноразмерному человеческому IL-6 (SEQ ID NO:161). Однако связывание может быть определено с использованием зрелого человеческого IL-6. Связывание с остатками IL-6 такое, как определено с помощью сайт-направленного мутагенеза, что объясняется ниже.
Мутагенез отдельных аминокислот и участков белков для того, чтобы скоррелировать структуру с активностью, хорошо известны специалисту, квалифицированному в данной области и использовались для определения участков белков, которые связываются с антителами [41]. Связывание и/или нейтрализация мутантного человеческого IL-6 может использоваться для оценки связывания связывающим членом Phe102, Ser204 и/или Arg207. Отсутствие связывания или нейтрализации, или значительно уменьшенное связывание или нейтрализация с мутантным IL-6 по сравнению с диким типом указывает на то, что связывающий член связывает мутированный остаток.
Связывание с остатком в IL-6 может быть определено с использованием IL-6, мутированного в выбранном остатке в испытании флюоресценции с временным разрешением ингибирования связывания меченого человеческого IL-6 дикого типа, со связывающим членом, иммобилизированным на подложке, причем меченый зрелый человеческий IL-6 дикого типа находится в конечной концентрации, равной Kd его взаимодействия со связывающим членом. Пример этого испытания и полученных данных по конкуренции показаны в Примере 3, с результатами, представленными в Таблице 10. В случае, когда мутантный IL-6 не ингибирует связывание меченого IL-6 дикого типа со связывающим членом, или когда мутантный IL-6 имеет IC50 больше чем IC50 немеченого IL-6 дикого типа (например, более чем 10-кратно или 100-кратно больше), это указывает на то, что мутированный остаток связан связывающим членом.
Phe102Glu мутантный человеческий IL-6 (SEQ ID NO:175), Ser204Glu мутантный человеческий IL-6 (SEQ ID NO:176) и/или Arg207Glu мутантный человеческий IL-6 (SEQ ID NO:177) могут показать отсутствие ингибирования или могут иметь IC50, которая больше чем 100-кратно больше, чем IC50 человеческого IL-6 дикого типа (SEQ ID NO:165), в испытании флюоресценции с временным разрешением для ингибирования связывания меченого человеческого IL-6 дикого типа, со связывающим членом изобретения, иммобилизированным на подложке, причем меченый человеческий IL-6 дикого типа находится в конечной концентрации, равной Kd его взаимодействия со связывающим членом.
Связывающий член согласно изобретению может (необязательно) не связывать и/или не нейтрализовать мутантный человеческий IL-6, имеющий мутацию в остатке Phe102, Ser204 и/или Arg207, например мутацию Phe102Glu, Ser204Glu, Ser204Tyr и/или Arg207Glu. Примеры последовательностей мутантного человеческого IL-6 включают SEQ ID NOS:175-177. Таким образом, связывающий член согласно изобретению может не ингибировать связывание одной или нескольких этих мутантных молекул IL-6 с IL-6Rα.
Связывающий член согласно изобретению может включать молекулу антитела, например молекулу человеческого антитела. Связывающий член обычно включает VH и/или VL домен антитела. VH и VL домены связывающих членов также предлагаются как часть изобретения. В пределах каждого VH и VL доменов находятся участки, определяющие комплементарность, ("CDR"), и каркасные участки, ("FR"). VH домен включает ряд HCDR, а VL домен включает ряд LCDR. Молекула антитела может включать VH домен антитела, включающий VH CDR1, CDR2 и CDR3 и каркас. Она может альтернативно или также включать VL домен антитела, включающий VL CDR1, CDR2 и CDR3 и каркас. Каркас VH или VL домена включает четыре каркасных участка, FR1, FR2, FR3 и FR4, рассеянных с CDR в следующей структуре:
FR1 - CDR1 - FR2 - CDR2 - FR3 - CDR3 - FR4.
Примеры VH и VL доменов антитела и CDR согласно настоящему изобретению перечислены в приложенном списке последовательностей, который является частью настоящего раскрытия. Дальнейшие CDR раскрыты ниже и в Таблице 7. Все VH и VL последовательности, CDR последовательности, наборы CDR и наборы HCDR и наборы LCDR, раскрытые в настоящей заявке, представляют аспекты и варианты осуществления изобретения. Как описано в настоящей заявке, "набор CDR" включает CDR1, CDR2 и CDR3. Таким образом, набор HCDR касается HCDR1, HCDR2 и HCDR3, и набор LCDR касается LCDR1, LCDR2 и LCDR3. Если не указано иначе, "набор CDR" включает HCDR и LCDR. Типично связывающие члены согласно изобретению представляют собой моноклональные антитела.
Связывающий член согласно изобретению может включать антиген-связывающий участок в пределах молекулы неантитела, обычно предоставляемой одним или несколькими CDRM, например набором CDR в остове белка неантитела, как обсуждается далее ниже.
Описанный в настоящей заявке связывающий член, включающий родительский набор CDR, как показано в Таблице 7, для родительского CAN022D10, в котором HCDR1 - SEQ ID NO:3 (остатки Kabat 31-35), HCDR2 - SEQ ID NO:4 (остатки Rabat 50-65), HCDR3 - SEQ ID NO:5 (остатки Rabat 95-102), LCDR1 - SEQ ID NO:8 (остатки Rabat 24-34), LCDR2 - SEQ ID NO:9 (остатки Kabat 50-56) и LCDR3 SEQ ID NO:10 (остатки Rabat 89-97).
Связывающий член согласно изобретению может включать один или несколько CDR, описанных в настоящей заявке, например CDR3, и необязательно также CDR1 и CDR2, чтобы сформировать набор CDR. CDR или набор CDR могут быть родительским CDR или родительским набором CDR, или могут быть CDR или набором CDR любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23, или могут быть их вариантом, как описано в настоящей заявке.
Например, связывающий член или VL домен согласно изобретению могут включать LCDR3, имеющий аминокислотную последовательность SEQ ID NO:120.
Связывающий член может включать набор Н и/или L CDR родительского антитела или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23 с одной или несколькими аминокислотными мутациями в пределах раскрытого набора Н и/или L CDR. Аминокислотные мутации представляют собой замены, делеции или инсерции одной аминокислоты. Например, может быть до 20, например до 12, 11, 10, 9, 8, 7, 6, 5, 4, 3 или 2 мутаций, например замен, в пределах набора Н и/или L CDR. Например, может быть до 6, 5, 4, 3 или 2 мутаций, например замен, в HCDR3 и/или может быть до 6, 5, 4, 3, или 2 мутаций, например, замен, в LCDR3. HCDR3 и/или LCDR3 могут необязательно содержать инсерцию или делецию одной аминокислоты по сравнению с раскрытым набором Н и/или LCDR. Замены могут, например, быть в положениях, замещенных в любом из Антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23, как показано в Таблице 7. Таким образом, замены могут необязательно быть в числах Kabat, выбранных из следующего:
Остаток Kabat 35 в HCDR1;
Остаток Kabat 64 в HCDR2;
Остаток Kabat 96, 97, 98, 99, 100, 100А, 100В, 100С и/или 102 в HCDR3;
Остаток Kabat 34 в LCDR1;
Остаток Kabat 89, 90, 91, 92, 93, 94, 96 или 97 в LCDR3.
Аминокислотные мутации могут включать мутации, как показано в Таблице 7, например, обозначенные аминокислотные замены.
Например, связывающий член или VH домен согласно изобретению могут включать родительский HCDR1 с остатком Kabat Ile 35, замещенным на Thr или Val.
Связывающий член или VH домен согласно изобретению могут включать родительский HCDR2 с остатком Kabat Lys 64, замененным на Arg.
Связывающий член или VH домен могут включать родительский HCDR3 с одной или несколькими из следующих мутаций:
остаток Kabat Ala 96 заменен на Glu;
остаток Kabat Asp 97 заменен на Glu или Asn;
остаток Kabat Asp 98 заменен на Gly, Glu или His;
остаток Kabat His 99 заменен на Gly или Thr;
остаток Kabat Tyr 100 заменен на Pro, Asn, Arg, Trp или Ala;
остаток Kabat Tyr 100А заменен на Ala, Arg, Thr, Gly, Asn, Pro или Ser;
остаток Kabat 100B заменен на His, Trp, Gln, Pro или Thr;
остаток Kabat Ile 100C заменен на Ala, Val, His, Tyr или Leu;
Ile вставленный в остаток Kabat 100D;
остаток Kabat Val 102 заменен на Leu, His, Met или Ile.
Таким образом, связывающий член или VH домен согласно изобретению могут включать HCDR3, причем остаток Kabat 100D представляет собой Ilе, или причем остаток Kabat 100D отсутствует.
Связывающий член или VL домен согласно изобретению могут включать родительский LCDR1, в котором остаток Kabat Ala 34 заменен на Thr.
Связывающий член VL домена изобретения может включать родительский LCDR3 с одной или несколькими из следующих мутаций:
остаток Kabat Gln 89 заменен на Met или Ala;
остаток Kabat Gln 90 заменен на Asn, Ser или Ala;
остаток Kabat Ser 91 заменен на Asn, Gly, Ala или His;
остаток Kabat Tyr 92 заменен на Trp, Ser, Lys или Phe;
остаток Kabat Ser 93 заменен на Leu, Lys, Arg или Ala;
остаток Kabat Thr 94 заменен на Ala, Gly или Pro;
остаток Kabat Pro 95 удален;
остаток Kabat Trp 96 заменен на Gly;
остаток Kabat Thr 97 заменен на Ser.
Таким образом, связывающий член или VL домен согласно изобретению могут включать LCDR3, в котором остаток Kabat 95 является Pro, или причем остаток Kabat 95 отсутствует.
Изобретение предоставляет выделенный связывающий член для человеческого IL-6, включающего набор CDR: HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, причем набор CDR имеет 22 или меньше аминокислотных изменений, например до 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2, 1 изменений или никаких изменений, от набора CDR, в котором:
HCDR1 имеет аминокислотную последовательность SEQ ID NO:3;
HCDR2 имеет аминокислотную последовательность SEQ ID NO:4;
HCDR3 имеет аминокислотную последовательность SEQ ID NO:115;
LCDR1 имеет аминокислотную последовательность SEQ ID NO:8;
LCDR2 имеет аминокислотную последовательность SEQ ID NO:9; и
LCDR3 имеет аминокислотную последовательность SEQ ID NO:120.
Аминокислотное изменение может быть заменой, инсерцией или делецией. Примеры положений Kabat, которые могут быть замещены, и примеры замен остатков обсуждаются ниже, и Таблица 7 иллюстрирует некоторые из замен.
Как показано в Таблице 7, длину HCDR3 и LCDR3 варьировали между различными оптимизированными антителами, описанными в настоящей заявке. Относительно родительских CDR CAN022D10 инсерция между остатками Kabat 100-102 (показаны в Таблице 7 в остатке Kabat 100D) наблюдалась в некоторых антителах, и делеция между остатками Kabat 92-97 наблюдалась в других антителах. Делеция в остатке Kabat 95 не наблюдалась в комбинации с инсерцией. Таким образом, может быть выгодно для более длинных 12-остатковых последовательностей HCDR3 быть комбинированными с более длинными 9- остатковыми последовательностями LCDR3, и может быть выгодно для более коротких 11-остатковых последовательностей HCDR3 быть комбинированными с более короткими 8-остатковыми последовательностями LCDR3.
Согласно системе нумерации Kabat, остатки LCDR3 пронумерованы от 89 до 97. Последовательности LCDR3 короче, чем 9 остатков, не предусмотрены системой нумерация Kabat. В настоящем изобретении у связывающих членов, например, может быть LCDR3 короче, чем 9 остатков. LCDR3 может быть 8 остатков в длину, как показано в Таблице 7. Мы нумеруем 8 остатков LCDR3 89, 90, 91, 92, 93, 94, 96 и 97 соответственно. В Таблице 7 делеция таким образом показана в остатке Kabat 95. Однако будет оценено, что влияние делеции должно уменьшить длину последовательности LCDR3, и что в принципе делеция, как могут полагать, может быть осуществлена в любом из остатков 89-97, например любом из остатков 92-97.
В HCDR3 системы нумерации Kabat приспосабливает изменчивость в длине CDR путем расширения системы нумерации между остатками Kabat 100 и 101, например, включая остаток 100А для HCDR3 9 остатков, плюс 100В для HCDR3 10 остатков, плюс 100С для HCDR3 11 остатков, плюс 100D для HCDR3 12 остатков соответствующим образом. В Таблице 7 инсерция дополнительной аминокислоты в HCDR3 некоторых из оптимизированных клонов относительно родительского HCDR3 показана в остатке Kabat 100D. Однако будет оценено, что в принципе эта инсерция, как могут полагать, будет осуществлена в любом из остатков Kabat 100-102.
Как демонстрируется в настоящей заявке, одна или несколько инсерций или делеций могут присутствовать в одном или нескольких CDR связывающего члена, например HCDR3 и/или LCDR3. Например, связывающий член согласно изобретению может включать набор CDR любого из Антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, или его вариант, описанный в настоящей заявке, причем каждый CDR необязательно имеет инсерцию для увеличения длины CDR на один остаток или имеет делецию одного остатка для уменьшения длины CDR на один остаток. Инсерции и/или делеции могут быть сделаны в HCDR и/или LCDR, например в HCDR3 и/или в LCDR3.
Например, связывающий член может, например, включать набор CDR, имеющих 20 или меньше аминокислотных замен в наборе CDR, в котором:
HCDR1 имеет аминокислотную последовательность SEQ ID NO:3;
HCDR2 имеет аминокислотную последовательность SEQ ID NO:4;
HCDR3 имеет аминокислотную последовательность SEQ ID NO:115;
LCDR1 имеет аминокислотную последовательность SEQ ID NO:8;
LCDR2 имеет аминокислотную последовательность SEQ ID NO:9; и
LCDR3 имеет аминокислотную последовательность SEQ ID NO:120,
причем у связывающего члена необязательно есть инсерция одного остатка для увеличения длины HCDR3 или делеция одного остатка для уменьшения длины HCDR3, и/или
имеет инсерцию одного остатка для увеличения длины LCDR3 или делецию одного остатка для уменьшения длины LCDR3.
Связывающий член согласно изобретению может иметь инсерцию одного остатка в HCDR3 SEQ ID NO:115 и/или инсерцию одного остатка в LCDR3 SEQ ID NO:120.
Инсерции или делеции могут быть сделаны в любой точке в CDR. Например, в HCDR3 инсерции или делеции может иметь любой из остатков Kabat 95-102, например любой из остатков Kabat 100-102. Например, в LCDR3 инсерции или делеции может иметь любой из остатков Kabat 89-97, например любой из остатков Kabat 92-97.
Связывающий член или VH домен согласно изобретению могут включать HCDR1, в котором остаток Kabat 35 является Ile, Thr или Val.
Связывающий член или VH домен согласно изобретению могут включать HCDR2, в котором остаток Kabat 64 является Lys или Arg.
Связывающий член или VH домен согласно изобретению могут включать HCDR3, в котором остаток Kabat 95 является Trp, и/или остаток Kabat 101 является Asp.
Связывающий член или VH домен согласно изобретению могут включать HCDR3, в котором:
Остатком Kabat 96 является Ala или Glu;
Остатком Kabat 97 является Asp, Glu или Asn;
Остатком Kabat 98 является Asp, Gly, Glu или His;
Остатком Kabat 99 является His, Gly или Thr;
Остатком Kabat 100 является Pro, Tyr, Asn, Arg, Trp или Ala;
Остатком Kabat 100A является Pro, Tyr, Ala, Arg, Thr, Gly, Asn, Pro или Ser;
Остатком Kabat 100B является Trp, Tyr, His, Gln, Pro или Thr;
Остатком Kabat 100C является Ile, Ala, Val, His, Tyr или Leu;
и Остатком Kabat 102 является Leu, Val, His, Met или Ile.
Связывающий член или VL домен согласно изобретению могут включать LCDR1, в котором остатком Kabat 34 является Ala или Thr.
Связывающий член или VL домен согласно изобретению могут включать LCDR3, в котором:
Остатком Kabat 89 является Gln, Met или Ala;
Остатком Kabat 90 является Gln, Asn, Ser или Ala;
Остатком Kabat 91 является Ser, Asn, Gly, Ala или His;
Остатком Kabat 92 является Trp, Tyr, Ser, Lys или Phe;
Остатком Kabat 93 является Leu, Ser, Lys, Arg или Ala;
Остатком Kabat 94 является Gly, Thr, Ala или Pro;
Остатком Kabat 96 является Gly или Trp; и
Остатком Kabat 97 является Ser или Thr.
Изобретение предлагает связывающие члены, включающие HCDR1, HCDR2 и/или HCDR3 родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, и/или LCDR1, LCDR2 и/или LCDR3 родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, например набор CDR родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, показанных в Таблице 7.
Например, связывающий член согласно изобретению может включать набор CDR: HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в котором:
HCDR1 - SEQ ID NO:113; HCDR2 - SEQ ID NO:114; HCDR3 - SEQ ID NO:115; LCDR1 - SEQ ID NO:118; LCDR2 - SEQ ID NO:119; и LCDR3 - SEQ ID NO:120, представляющие CDR Антитела 18.
Связывающий член может включать набор CDR VH одного из этих антител. Необязательно он может также включать набор CDR VL одного из этих антител, и CDR VL могут быть от того же самого или разных антител как CDR VH.
VH домен, включающий набор HCDR родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, и/или VL домен, включающий набор LCDR родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, также предлагаются в соответствии с изобретением.
Как правило, VH домен спарен с VL доменом для обеспечения антиген-связывающего участка антитела, хотя, как обсуждается далее ниже, VH или VL домен по отдельности может использоваться для связывания антигена. 2 VH домен антитела может быть спарен с 2 VL доменом антитела, таким образом, что образуется антиген-связывающий участок антитела, включающий 2 VH и VL домены антитела. Аналогичные варианты осуществления предоставлены для других VH и VL доменов, раскрытых в настоящей заявке. В других вариантах осуществления 2 VH антитела спарен с VL доменом, отличающимся от VL антитела. Разнородность легкой цепи хорошо установлена в данной области. Снова, аналогичные варианты осуществления предоставлены в соответствии с изобретением для других VH и VL доменов, раскрытых в настоящей заявке.
Таким образом, VH родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23 может быть спарен с VL родительского или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23.
Связывающий член может включать молекулу антитела, имеющую один или несколько CDR, например набор CDR, в пределах каркаса антитела. Например, один или несколько CDR или набор CDR антитела могут быть привиты в каркас (например, человеческий каркас), чтобы обеспечить молекулу антитела. Каркасные участки могут иметь человеческие зародышевые последовательности генного сегмента. Таким образом, каркас может быть зародышевый, причем один или несколько остатков в пределах каркаса изменены, чтобы соответствовать остаткам в эквивалентном положении в самом подобном человеческом зародышевом каркасе. Квалифицированный специалист может выбрать зародышевый сегмент, который является самым близким в последовательности к каркасной последовательности антитела перед «germlining» и проверить аффинность или активность антител для подтверждения того, что «germlining» значительно не уменьшает связывание антигена или силу действия в испытании, описанном в настоящей заявке. Человеческие зародышевые последовательности сегмента гена известны квалифицированным специалистам в данной области и могут быть оценены, например, из компиляции VBase.
Связывающий член согласно изобретению может быть выделенной молекулой антитела человека, имеющей VH домен, включающий набор HCDR в человеческой зародышевой структуре, например, Vh3_DP-86_(3-66). Таким образом, каркасные участки VH домена FR1, FR2 и/или FR3 могут включать каркасные участки человеческого зародышевого сегмента гена Vh3_DP-86_(3-66) и/или могут стать зародышевыми путем мутирования каркасных участков, чтобы соответствовать каркасным участкам этого человеческого зародышевого сегмента гена. FR4 может включать каркасный участок человеческого зародышевого j сегмента JH2. Аминокислотная последовательность VH FR1 может быть SEQ ID NO:167. Аминокислотная последовательность VH FR2 может быть SEQ ID NO:168. Аминокислотная последовательность VH FR3 может быть SEQ ID NO:169. Аминокислотная последовательность VH FR4 может быть SEQ ID NO:170.
Обычно, связывающий член также имеет VL домен, включающий набор LCDR, например, в человеческом зародышевом каркасе, например Vk1_L12. Таким образом, каркасные участки VL домена могут включать каркасные участки FR1, FR2 и/или FR3 человеческого зародышевого сегмента гена Vk1_L12 и/или могут стать зародышевыми путем мутирования каркасных участков, чтобы соответствовать каркасным участкам этого человеческого зародышевого сегмента гена. FR4 может включать каркасный участок человеческого зародышевого j сегмента JK2. Аминокислотная последовательность VL FR1 может быть SEQ ID NO:171. Аминокислотная последовательность VL FR2 может быть SEQ ID NO:172. Аминокислотная последовательность VL FR3 может быть SEQ ID NO:173. Аминокислотная последовательность VL FR4 может быть SEQ ID NO:174.
Зародышевый VL домен может быть зародышевым или нет в остатке или остатках Vernier, но обычно нет.
Молекула антитела или VH домен согласно изобретению могут включать следующий набор каркасных участков тяжелой цепи:
FR1 SEQ ID NO:167;
FR2 SEQ ID NO:168;
FR3 SEQ ID NO:169;
FR4 SEQ ID NO:170,
или может включать упомянутый набор каркасных участков тяжелой цепи с одной, двумя, тремя, четырьмя или пятью аминокислотными изменениями, например заменами.
Молекула антитела или VL домен согласно изобретению могут включать следующий набор каркасных участков легкой цепи:
FR1 SEQ ID NO:171;
FR2 SEQ ID NO:172;
FR3 SEQ ID NO:173;
FR4 SEQ ID NO:174,
или может включать упомянутый набор каркасных участков легкой цепи с одной, двумя, тремя, четырьмя или пятью аминокислотными изменениями, например заменами.
Аминокислотное изменение может представлять собой замену, инсерцию или делецию.
Например, молекула антитела изобретения может включать набор каркасных участков тяжелой и легкой цепи, причем
тяжелая цепь FR1 представляет собой SEQ ID NO:167;
тяжелая цепь FR2 представляет собой SEQ ID NO:168;
тяжелая цепь FR3 представляет собой SEQ ID NO:169;
тяжелая цепь FR4 представляет собой SEQ ID NO:170;
легкая цепь FR1 представляет собой SEQ ID NO:171;
легкая цепь FR2 представляет собой SEQ ID NO:172;
легкая цепь FR3 представляет собой SEQ ID NO:173;
легкая цепь FR4 представляет собой SEQ ID NO:174,
или может включать упомянутый набор каркасных участков тяжелой и легкой цепи с 10 или меньше, например пятью или меньше, аминокислотными изменениями, например заменами. Например, могут быть одна или две аминокислотные замены в упомянутом наборе каркасных участков тяжелой и легкой цепи.
Незародышевая молекула антитела имеет те же самые CDR, но различные каркасы по сравнению с зародышевой молекулой антитела. Из последовательностей антитела, показанных в настоящей заявке в приложенном списке последовательностей, последовательности антитела под номером 7, 10, 17 и 18 являются зародышевыми. Зародышевые антитела 2-5, 8, 14, 16, 19 и 21-23 могут быть получены путем «germlining» каркасных участков последовательностей VH и VL домена, показанных в настоящей заявке для этих антител.
3' cgt кодон, и соответствующий остаток Аргинина, показанные в нуклеотидной и аминокислотной последовательностях для каппа VL доменов Антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23 соответственно, были включены в экспрессированные scFv и IgG последовательности этих антител. C-терминальный остаток Аргинина последовательностей соответствует остатку Kabat 108. Происхождение этого остатка и его кодирующий триплет cgt объясняются ниже.
Чтобы экспрессировать легкую цепь IgG, была предоставлена нуклеотидная последовательность, кодирующая легкую цепь антитела, включающая первый экзон, кодирующий VL домен, второй экзон, кодирующий CL домен, и интрон, разделяющий первый экзон и второй экзон. При нормальных обстоятельствах интрон соединяется клеточной системой обработки мРНК, объединяя 3' конец первого экзона с 5' концом второго экзона. Таким образом, когда ДНК, имеющая упомянутую нуклеотидную последовательность, была экспрессирована как РНК, первый и второй экзоны были соединены сплайсингом вместе. При трансляции соединенной сплайсингом РНК получают полипептид, включающий VL домен и CL домен. Выбор константного домена является существенным, так как для легких цепей каппа, соединяющая аминокислота - аргинин, сформированная cgа кодоном, где первый цитозин закодирован в экзоне 1, а гуанин и аденин закодированы в экзоне 2.
После сплайсинга для Антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, Arg в остатке Kabat 108 закодирован последним основанием (с) каркасной 4 последовательности VL домена и первыми двумя основаниями (gt) CL домена.
Остаток Аргинина в остатке Kabat 108, как могут полагать, является С терминальным остатком VL домена молекулы антитела.
Связывающий член согласно изобретению может быть тем, который конкурирует за связывание с IL-6 с любым связывающим членом, который (i) связывает IL-6, и (ii) включает связывающий член, VH и/или VL домен, CDR, например HCDR3, и/или набор CDR, раскрытый в настоящей заявке.
Конкуренция между связывающими членами может быть легко оценена in vitro, например, используя ELISA и/или присоединения специфичной сигнальной молекулой к одному связывающему члену, которая может обнаруживаться в присутствии одного или нескольких других нетагговых связывающих членов, чтобы позволить идентификацию связывающих членов, которые связывают тот же эпитоп или перекрывающийся эпитоп. Такие методы хорошо известны среднему специалисту в данной области и описаны более подробно в настоящей заявке (см. Детальное Описание, и испытание конкуренции эпитопов в разделе Материалы и Методы Примеров). Таким образом, дальнейший аспект настоящего изобретения предлагает связывающий член, включающий человеческий антиген-связывающий участок антитела, который конкурирует с молекулой антитела, например молекулой антитела, включающей VH и/или VL домен, CDR, например HCDR3 или набор CDR родительского антитела или любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, для связывания с IL-6.
В дальнейших аспектах изобретение предлагает выделенную нуклеиновую кислоту, которая включает последовательность, кодирующую связывающий член, VH домен и/или VL домен согласно настоящему изобретению, и способы получения связывающего члена, VH домена и/или VL домена изобретения, которые включают экспрессию указанной нуклеиновой кислоты при условиях для инициации производства указанного связывающего члена, VH домена и/или VL домена, и его извлечения.
Другой аспект настоящего изобретения предлагает нуклеиновую кислоту, как правило, выделенную, кодирующую VH CDR или VL CDR последовательность, раскрытую в настоящей заявке.
Дальнейший аспект предлагает клетку-хозяин, содержащую нуклеиновую кислоту изобретения или трансформированную с ее помощью.
Дальнейшие аспекты настоящего изобретения предусматривают композиции, содержащие связывающие члены согласно изобретению, и их применение в способах связывания, ингибирования и/или нейтрализации IL-6, включая способы терапевтического лечения человека или животного.
Связывающие члены согласно изобретению могут использоваться в способе лечения или диагностики, таком как способ лечения (который может включать профилактическое лечение) заболевания или расстройства организма человека или животного (например, у пациента-человека), который включает введение указанному пациенту эффективного количества связывающего члена согласно изобретению. Состояния, поддающиеся лечению в соответствии с настоящим изобретением, включают любое состояние, в котором задействован IL-6, как обсуждается подробно в настоящей заявке в дальнейшем.
Эти и другие аспекты изобретения более подробно описаны ниже.
Терминология
Представляется возможным отметить, что "и/или" при использовании в настоящей заявке должно рассматриваться как определенное раскрытие каждой из двух указанных характеристик или компонентов с другим или без него. Например, "А и/или В" должно рассматриваться как определенное раскрытие каждого из (i) A, (ii) В и (iii) А и В, так же, если бы каждый был приведен в настоящей заявке индивидуально.
IL-6 и IL-6 рецептор
IL-6 - интерлейкин 6. IL-6 может также быть упомянут в настоящей заявке как "антиген".
Полноразмерная аминокислотная последовательность человеческого IL-6 представляет собой SEQ ID NO:161. Эта последовательность расщепляется in vivo для удаления N-терминального лидерного пептида, для получения зрелого IL-6. Зрелый человеческий IL-6 имеет аминокислотную последовательность SEQ ID NO:165. Зрелая последовательность представляет in vivo циркулирующий IL-6, который является антигеном-мишенью для терапевтического и in vivo диагностического применения, описанного в настоящей заявке. Соответственно, IL-6, обозначенный в настоящей заявке, представляет собой обычно зрелый человеческий IL-6, если в контексте не указано иначе.
IL-6 может быть конъюгирован с обнаружимой меткой, такой как HIS FLAG, например, для использования в испытании, описанном в настоящей заявке. Например, может использоваться слитый белок, включающий IL-6, конъюгированный с последовательностью HIS FLAG. Последовательность HIS FLAG, меченная человеческим IL-6, представляет собой SEQ ID NO:162.
Рецептор IL-6 a, IL-6Ra представляет собой рецептор для интерлейкина 6. IL-6Ra также известен как IL-6Ra, IL-6Ra, IL-6R и CD126. IL-6Ra существует in vivo в трансмембранной форме и в растворимой форме. Ссылками на IL-6Ra могут быть трансмембранный IL-6Ra и/или растворимый IL-6Ra, если в контексте не указано иначе.
Рецептор IL-6, упомянутый в настоящей заявке, обычно является рецептором человеческого IL-6, если не обозначено иначе. Аминокислотная последовательность человеческого растворимого IL-6Ra (sIL-6Ra, SIL-6R) представляет собой SEQ ID NO:163. Аминокислотная последовательность человеческого трансмембранного IL-6Ra представляет собой SEQ ID NO:164.
IL-6 связывает IL-6Ra с образованием комплекса IL-6:IL-6Ra. Комплекс может быть либо растворимым (с sIL-6Ra), либо связанным с мембраной (с трансмембранным IL-6Ra). Когда IL-6Ra находится в растворимой форме, комплекс обозначается как IL-6:sIL-6Ra. Ссылки на IL-6:IL-6Ra могут включать IL-6 в комплексе с трансмембранным IL-6Ra или с растворимым IL-6Ra, если в контексте не указано иначе.
gp130
gp130 представляет собой рецептор для комплекса IL-6:IL-6Ra. О клонировании и характеристике gp130 описано в Hibi и др., Cell 63:1149-1157 (1990). Последовательность человеческого gp130 изложена в SEQ ID NO:166.
Связывающий член
Описывает одного члена пары молекул, которые связывают друг друга. Члены связывающей пары могут быть получены природным путем или полностью, или частично синтетично. Один член пары молекул имеет область на своей поверхности, или полость, которая связывается со специфической пространственной и полярной организацией другого члена пары молекул и поэтому комплементарна ей. Примерами типов связывающих пар являются антиген-антитело, биотин-авидин, гормон-рецептор гормона, рецептор-лиганд, фермент-субстрат. Настоящее изобретение касается реакций типа антиген-антитело.
Связывающий член обычно включает молекулу, имеющую антиген-связывающий участок. Например, связывающий член может быть молекулой антитела или белком неантитела, который включает антиген-связывающий участок.
Антиген-связывающий участок может быть предоставлен посредством группировки CDR на остовах белка неантитела, таких как фибронектин или цитохром В и т.д. [42, 43, 44], или путем рандомизации или мутации аминокислотных остатков петли в пределах остова белка, чтобы обусловить связывающую специфичность для желаемой мишени. Остовы для создания новых связывающих участков в белках были рассмотрены подробно в Nygren и соавт. [44]. Остовы белка для имитаторов антитела раскрыты в WO/0034784, который включен в настоящую заявку путем ссылки в полном объеме, в котором изобретатели описывают белки (имитаторы антитела), которые включают домен III фибронектинового типа, имеющий по крайней мере одну рандомизированную петлю. Подходящий остов, в который прививается один или несколько CDR, например набор HCDR, может быть предоставлен любым членом домена суперсемейства генов иммуноглобулинов. Остов может быть человеческим или нечеловеческим белком. Преимущество остова белка неантитела состоит в том, что он может обеспечить антиген-связывающий участок в молекуле остова, которая меньше и/или легче для производства, чем по крайней мере некоторые молекулы антитела. Небольшой размер связывающего члена может обусловить полезные физиологические свойства, такие как способность входить в клетки, глубоко проникать в ткани или достигать мишени в пределах других структур, или связываться в пределах полостей белка антигена-мишени. Использование антиген-связывающих участков в остовах белка неантитела рассмотрено в Wess, 2004 [45]. Типичными являются белки, имеющие стабильную основную цепь и одну или несколько вариабельных петель, в которых аминокислотная последовательность петли или петель специфично или рандомизировано мутирована для создания антиген-связывающего участка, который связывает целевой антиген. Такие белки включают IgG-связывающие домены белка А от S. aureus, трансферрин, тетранектин, фибронектин (например, 10-й домен фибронектинового типа III), липокалины, а также гамма-кристаллин и другие остовы Affilin™ (Scil Proteins). Примеры других подходов включают синтетические "Микротела" на основе циклотидов - маленьких белков, имеющих внутримолекулярные дисульфидные связи. Микробелки (Versabodies™, Amunix) и белки анкириновых повторов (DARPins, Molecular Partners).
В дополнение к последовательностям антитела и/или антиген-связывающему участку, связывающий член согласно настоящему изобретению может включать другие аминокислоты, например, образуя пептид или полипептид, такие как свернутый домен, или для обеспечения молекулы другой функциональной характеристикой в дополнение к способности связывать антиген. Связывающие члены согласно изобретению могут нести обнаружимую метку, или могут быть конъюгированы с токсином или нацеливающим остатком или ферментом (например, через пептидильную связь или линкер). Например, связывающий член может включать каталитический участок (например, в домене фермента), а также антиген-связывающий участок, причем антиген-связывающий участок связывается с антигеном и таким образом нацеливает каталитический участок на антиген. Каталитический участок может ингибировать биологическую функцию антигена, например, путем расщепления.
Хотя, как отмечено, CDR могут нести остовы неантитела, структура для того, чтобы нести CDR или набор CDR изобретения вообще будет представлять собой тяжело- или легкоцепочечную последовательность антитела или ее существенную часть, в которой CDR или набор CDR расположены в положении, соответствующем CDR или набору CDR природных вариабельных VH и VL доменов антитела, закодированных перегруппированными генами иммуноглобулина. Структуры и положения вариабельных доменов иммуноглобулина могут быть определены, см. Kabat и соавт., 1987 [46], и обновления. Доступны многие академические и коммерческие ресурсы онлайн для запроса этой базы данных. Например, см. ссылку [47] и связанный ресурс онлайн в настоящее время по адресу в сети http://www.bioinf.org.uk/abs/simkab.html.
CDR участок или CDR обозначает гипервариабельные участки тяжелых и легких цепей иммуноглобулина, как определено Kabat и соавт. 1991 [48], и более поздними изданиями. Антитело типично содержит 3 CDR тяжелой цепи и 3 CDR легкой цепи. Термин CDR или CDR использован в настоящей заявке для обозначения, согласно случаю, одного из этих участков или нескольких, или даже всех этих участков, которые содержат большинство аминокислотных остатков, отвечающих за связывание аффинностью антитела для антигена или эпитопа, который оно распознает.
Среди шести коротких последовательностей CDR, третий CDR тяжелой цепи (HCDR3) имеет большую изменчивость размеров (большее разнообразие, главным образом из-за механизмов группировки генов, которые дают этому начало). Он может составлять всего 2 аминокислоты, хотя самый длинный известный размер равен 26. Длина CDR может также варьировать в зависимости от длины, которая может быть приспособлена определенным основным каркасом. Функционально, HCDR3 частично играет роль в определении специфичности антитела [ссылки. 49, 50, 51, 52, 53, 54, 55, 56].
HCDR1 может составлять 5 аминокислот в длину, состоя из остатков Kabat 31-35.
HCDR2 может составлять 17 аминокислот в длину, состоя из остатков Kabat 50-65.
HCDR3 может составлять 11 или 12 аминокислот в длину, состоя из остатков Kabat 95-102, необязательно включая остаток Kabat 100D.
LCDR1 может составлять 11 аминокислот в длину, состоя из остатков Kabat 24-34.
LCDR2 может составлять 7 аминокислот в длину, состоя из остатков Kabat 50-56.
LCDR3 может составлять 8 или 9 аминокислот в длину, состоя из остатков Kabat 89-97, необязательно включая остаток Kabat 95.
Молекула антитела
Описывает иммуноглобулин природный или полученный частично или полностью синтетическим путем. Термин также охватывает любой полипептид или белок, включающий антиген-связывающий участок антитела. Необходимо понимать здесь, что изобретение не касается антител в природной форме, то есть они не находятся в своей природной среде, а то, что их можно изолировать или получить очисткой из природных источников, или получить иным образом путем генетической рекомбинации, или химического синтеза, и что они могут тогда содержать неприродные аминокислоты, как будет описано далее. Фрагменты антитела, которые включают антиген-связывающий участок антитела, включают, кроме прочих, молекулы, такие как Fab, Fab', Fab'-SH, scFv, Fv, dAb и Fd. Были созданы генной инженерией различные другие молекулы антитела, включая один или несколько антиген-связывающих участков антитела, включая например, Fab2, Fab3, диатела, триатела, тетратела и минитела. Молекулы антитела и методы для их создания и использования описаны в [57].
Возможно применять моноклональные и другие антитела и методы использования технологии рекомбинантной ДНК с целью получения других антител или химерных молекул, которые связывают целевой антиген. Такие методы могут включать введение ДНК, кодирующую иммуноглобулиновый вариабельный участок, или CDR, антитела к константным участкам, или константным участкам плюс каркасным участкам, различного иммуноглобулина. См., например, ЕР-А-184187, GB 2188638A или ЕР-А-239400, и большой объем последующей литературы. Гибридома или другая клетка, вырабатывающая антитело, могут подвергаться генетической мутации или другим изменениям, которые могут (или нет) изменить связывающую специфичность выработанных антител.
Поскольку антитела могут быть модифицированы многими способами, термин "молекула антитела" должен быть рассмотрен как охватывающий любой связывающий член или вещество, имеющее антиген-связывающий участок антитела с необходимой специфичностью и/или связывания с антигеном. Таким образом, этот термин охватывает фрагменты и производные антитела, включая любой полипептид, содержащий антиген-связывающий участок антитела, природный или полученный частично или полностью синтетическим путем. Поэтому включаются химерные молекулы, содержащие антиген-связывающий участок антитела, или эквивалентный, слитый с другим полипептидом (например, полученным из другого вида или принадлежащим другому классу или подклассу антител). Клонирование и экспрессия химерных антител описаны в ЕР-А-0120694 и ЕР-А-0125023 и большом объеме последующей литературы.
Дальнейшие методы, доступные в области разработки антител, позволили изолировать человеческие и гуманизированные антитела. Например, человеческие гибридомы могут быть сделаны, как описано в Kontermann & Dubel [58]. Фаговый дисплей, другая установленная техника для создания связывающих членов была описана подробно во многих публикациях, таких как Kontermann & Dubel [58] и WO92/01047 (обсуждается далее ниже), и патенты США US5969108, US5565332, US5733743, US5858657, US5871907, US5872215, US5885793, US5962255, US6140471, US6172197, US6225447, US6291650, US6492160, US6521404.
Трансгенные мыши, в которых гены мышиного антитела инактивированы и функционально заменены генами антител человека, оставляя интактными другие компоненты иммунной системы мыши, могут использоваться для выделения человеческих антител [59]. Гуманизированные антитела могут быть получены с использованием методов, известных в данной области, таких как раскрытые, например, в WO 91/09967, US 5585089, EP592106, US 565 332 и WO93/17105. Далее, в WO 2004/006955 описаны методы для гуманизирования антител, основанные на выборе каркасных последовательностей вариабельного участка из человеческих генов антитела путем сравнивания канонических CDR структурных типов для последовательностей CDR вариабельного участка нечеловеческого антитела к каноническим CDR структурным типам для соответствующих CDR из библиотеки последовательностей человеческих антител, например зародышевых сегментов генов антител. Вариабельные участки человеческого антитела, имеющие подобные канонические CDR структурные типы к нечеловеческим CDR, формируют подмножество последовательностей человеческих антител членов для выбора человеческих каркасных последовательностей. Члены подмножества могут быть далее упорядочиваться по подобию аминокислот между человеческими и нечеловеческими последовательностями CDR. В способе согласно WO 2004/006955 главные упорядоченные человеческие последовательности выбираются для обеспечения каркасных последовательностей для конструирования химерного антитела, которое функционально заменяет человеческие последовательности CDR нечеловеческими копиями CDR с использованием отобранных человеческих каркасов членов подмножества, таким образом обеспечивая гуманизированное антитело высокой аффинности и низкой иммуногенности без необходимости сравнивать каркасные последовательности между нечеловеческими и человеческими антителами. Также описаны химерные антитела, сделанные согласно способу.
Синтетические молекулы антитела могут быть созданы путем экспрессии от генов, полученных посредством олигонуклеотидов, синтезированных и собранных в пределах подходящих векторов экспрессии, например, как описано Knappik и соавт. [60] или Krebs и соавт. [61].
Было показано, что фрагменты целого антитела могут выполнять функцию связывающих антигенов. Примеры связывающих фрагментов включают (i) фрагмент Fab, состоящий из VL, VHA, CL и СН1 доменов; (ii) фрагмент Fd, состоящий из VH и СН1 доменов; (iii) фрагмент Fv, состоящий из VL и VH доменов одного антитела; (iv) фрагмент dAb [62, 63, 64], который состоит из VH или VL домена; (v) изолированные участки CDR; (vi) фрагменты F(ab')2, бивалентный фрагмент, включающий два связанных фрагмента Fab (vii) одноцепочечные молекулы Fv (scFv), в которых VH домен и VL домен связаны пептидным линкером, который позволяет этим двум доменам ассоциировать, чтобы сформировать антиген-связывающий участок [65, 66]; (viii) биспецифичные одноцепочечные Fv димеры (PCT/US92/09965) и (ix) "диатела", многовалентные или мультиспецифичные фрагменты, сконструированные генным слиянием (WO94/13804; [67]). Fv, scFv или молекулы диатела могут быть стабилизированы путем введения дисульфидных мостиков, связывающих VH и VL домены [68]. Могут также быть сделаны минитела, включающие scFv, соединенный с СН3 доменом [69]. Другие примеры связывающих фрагментов включают Fab', который отличается от фрагментов Fab дополнением нескольких остатков на карбоксиконце СН1 домена тяжелой цепи, включая один или более цистеинов из шарнирного участка антитела, и Fab'-SH, который представляет собой фрагмент Fab', в котором остаток(ки) цистеина константных доменов содержат свободную тиольную группу.
Qui и соавт. [70] описали молекулы антитела, содержащие только два CDR, связанных каркасным участком. CDR3 из VH или VL домена был связан с петлей CDR1 или CDR2 другого домена. Связь осуществлялась через С конец выбранного CDR1 или CDR2 к N концу CDR3, через участок FR. Qui и соавт. выбрали участок FR, имеющий наименьшее количество гидрофобных участков. Лучшая комбинация для испытуемого антитела, как было обнаружено, была VL CDR1, связанный с помощью VH FR2 к VНУ CDR3 (VHCDR1-VHFR2-VLCDR3). При молекулярной массе, составляющей приблизительно 3 кДа, эти молекулы антитела предлагают преимущества с точки зрения улучшенного проникновения в ткани по сравнению с полноразмерными иммуноглобулинами (приблизительно 150 кДа) или scFv (приблизительно 28 кДа).
Могут быть получены фрагменты антитела изобретения, начиная с родительской молекулы антитела или любой из молекул антитела 2, 3, 4, 5, 1, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, с помощью методов, таких как расщепление ферментами, например пепсином или папаином, и/или расщеплением дисульфидных мостиков химическим восстановлением. Иным образом, фрагменты антител, содержащиеся в настоящем изобретении, могут быть получены методами генетической рекомбинации, аналогичными известным специалисту, квалифицированному в данной области или иначе пептидным синтезом посредством, например, автоматических синтезаторов пептидов, таких как реализуются компанией «Applied Biosystems», и т.д., или синтезом и экспрессией нуклеиновой кислоты.
Функциональные фрагменты антитела согласно настоящему изобретению включают любой функциональный фрагмент, период полувыведения которого увеличен химической модификацией, особенно ПЕГилированием, или введением в липосому.
dAb (антитело домена) является маленьким мономерным антиген-связывающим фрагментом антитела, а именно вариабельным участком тяжелой или легкой цепи антитела [64]. dAb VH встречаются в природе у верблюдовых (например, верблюде, ламе) и могут быть получены путем иммунизации верблюдовых целевым антигеном, изоляции антиген-специфичных В-клеток и непосредственного клонирования генов dAb из индивидуальных В-клеток. dAb также получают в клеточной культуре. Их небольшой размер, хорошая растворимость и температурная стабильность делают их особенно физиологически полезными и подходящими для выбора и созревания аффинности. dAb VH верблюдовых разрабатываются для терапевтического использования под названием "нанотела™". Связывающий член согласно настоящему изобретению может представлять собой dAb, включающее VH или VL домен, существенно такое, как изложено в настоящей заявке, или VH или VL домен, включающий набор CDR, существенно такое, как изложено в настоящей заявке.
Биспецифичные или бифункциональные антитела формируют второе поколение моноклональных антител, в которых два различных вариабельных участка объединены в той же молекуле [71]. Их использование было продемонстрировано и в диагностической области и в области терапии из-за их способности вербовать новые эффекторные функции или нацеливать несколько молекул на поверхности опухолевых клеток. При потребности использовать биспецифичные антитела, они могут быть обычными биспецифичными антителами, которые могут быть произведены множеством способов [72], например подготовлены химически или из гибридных гибридом, или могут быть любым из биспецифичных фрагментов антитела, упомянутых выше. Эти антитела могут быть получены химическими методами [73, 74] или соматическими ми методами [75, 76], но аналогично и предпочтительно методами генной инженерии, которые позволяют вызвать гетеродимеризацию и таким образом облегчить процесс очистки нужного антитела [77]. Примеры биспецифичных антител включают таковые технологии BiTE™, в которой могут использоваться связывающие домены двух антител с различной специфичностью и непосредственно связываться через короткие гибкие пептиды. Это комбинирует два антитела на короткой единственной цепи полипептида. Диатела и scFv могут быть сконструированы без участка Fc с использованием только вариабельных доменов, потенциально снижающих эффекты антиидиопатической реакции.
Биспецифичные антитела могут быть сконструированы как весь IgG, как биспецифичный Fab'2, как Fab'PEG, как диатела или иначе как биспецифичный scFv. Далее, два биспецифичных антитела могут быть связаны с использованием обычных методов, известных в данной области, чтобы сформировать тетравалентные антитела. Биспецифичные диатела, в противоположность бмспецифичным целым антителам, могут также быть особенно полезными, потому что они могут быть легко сконструированы и экспрессированы в E.coli. Диатела (и много других полипептидов, таких как фрагменты антитела) соответствующих связывающих специфичностей могут быть легко выбраны с использованием фагового дисплея (WO94/13804) из библиотек. Если одно плечо диатела необходимо сохранить постоянным, например, со специфичностью, направленной против IL-6, то может быть сделана библиотека, где другое плечо варьирует, и выбирается антитело соответствующей специфичности. Биспецифичные целые антитела могут быть созданы альтернативными методами инженерии, как описано в Ridgeway и соавт., 1996 [78].
В данной области техники доступны различные методы для того, чтобы получить антитела против IL-6. Антитела могут быть моноклональными антителами, особенно человеческого, крысиного, химерного или гуманизированного происхождения, которые могут быть получены согласно стандартным методам, известным специалисту, квалифицированному в данной области.
Вообще, для подготовки моноклональных антител или их функциональных фрагментов, особенно крысиного происхождения, можно обратиться к методам, которые описаны в особенности в руководстве "Antibodies" [79], или к методу получения из гибридом, описанному Köhler и Milstein [80].
Моноклональные антитела могут быть получены, например, из В клеток животного, иммунизированного против IL-6, или одного из его фрагментов, содержащих эпитоп, распознанный указанными моноклональными антителами. Подходящие фрагменты и пептиды или полипептиды, включающие их, описаны в настоящей заявке, и могут использоваться для иммунизации животных, чтобы получить антитела против IL-6. Указанный IL-6, или один из его фрагментов, может особенно быть получен согласно обычным рабочим методам, путем генетической рекомбинации, начинающейся с нуклеиновокислотной последовательности, содержащейся в последовательности кДНК, кодирующей IL-6 или его фрагмент, путем синтеза пептида, начинающегося с последовательности аминокислот, содержащейся в пептидной последовательности IL-6 и/или его фрагмента.
Моноклональные антитела могут, например, быть очищены на аффинной колонке, на которой был ранее иммобилизирован IL-6 или один из его фрагментов, содержащих эпитоп, распознанный указанными моноклональными антителами. Более подробно, моноклональные антитела могут быть очищены хроматографией на белке А и/или G, что сопровождается или не сопровождается ионнообменной хроматографией, нацеленной на устранение остаточных белковых загрязнителей, а также как ДНК и LPS, самих по себе, что сопровождается или не сопровождается эксклюзионной хроматографией на геле Сефароза, чтобы устранить потенциальные аггрегаты из-за присутствия димеров или других мультимеров. В одном варианте осуществления все эти методы могут использоваться одновременно или последовательно.
Антиген-связывающий участок
Описывает часть молекулы, которая связывается со всем или частью антигена-мишени и комплементарна ему. В молекуле антитела он упоминается как антиген-связывающий участок антитела и включает часть антитела, которая связывается со всем или частью антигена-мишени и комплементарна ему. Если антиген является большим, антитело может только связываться со специфической частью антигена, часть которого называют эпитопом. Антиген-связывающий участок антитела может быть предоставлен одним или несколькими вариабельными доменами антитела. Антиген-связывающий участок антитела может включать вариабельный участок легкой цепи антитела (VL) и вариабельный участок тяжелой цепи антитела (VH).
В WO2006/072620 описано создание с помощью инженерии антиген-связывающих участков в структурных (не CDR) петлях, простирающихся между бета-спиралями иммуноглобулиновых доменов. Антиген-связывающий участок может быть создан в участке молекулы антитела, отличного от природного положения CDR, например в каркасном участке VH или VL домена, или в константном домене антитела, например СН1 и/или СН3. Антиген-связывающий участок, спроектированный в структурном участке, может быть дополнительным к антиген-связывающему участку, сформированному наборами CDR VH и VL домена, или заменять его. При наличии множественных антиген-связывающих участков в молекуле антитела, они могут связывать тот же самый антиген (IL-6), таким образом увеличивая валентность связывающего члена. Альтернативно, множественные антиген-связывающие участки могут связывать различные антигены (IL-6 и один или несколько других антигенов), и это может использоваться для добавления эффекторных функций, продления периода полувыведения или улучшения in vivo поставки молекулы антитела.
Выделенный
Касается состояния, в котором связывающие члены согласно изобретению или нуклеиновая кислота, кодирующая такие связывающие члены, вообще будут находиться в соответствии с настоящим изобретением. Таким образом, связывающие члены, VH и/или VL домены и кодирующие молекулы нуклеиновой кислоты и векторы согласно настоящему изобретению можно представить выделенными и/или очищенными, например, из их природной среды, в практически чистой или гомогенной форме, или, в случае нуклеиновой кислоты, свободной или практически свободной от нуклеиновой кислоты или генов происхождения, отличных от последовательности, кодирующей полипептид с необходимой функцией. Выделенные члены и выделенная нуклеиновая кислота будут свободными или практически свободными от материала, с которым они природно ассоциированы, такие как другие полипептиды или нуклеиновые кислоты, с которыми они обнаружены в их природной среде, или среде, в которой они получены (например, клеточная культура), когда такое получение осуществляется технологией рекомбинантной ДНК in vitro или in vivo. Члены и нуклеиновая кислота могут быть приготовлены с разбавителями или адъювантами и все еще для практических целей быть выделенными - например, члены будут обычно смешиваться с желатином или другими носителями при использовании для покрытия микротитровальных планшетов для использования в иммуноисследованиях, или будут смешаны с фармацевтически приемлемыми носителями или разбавителями при применении в диагностике или терапии. Связывающие члены могут быть гликозилированными, природно или системами гетерологических эукариотических клеток (например, СНО или NS0 (ЕСАСС 85110503) клеток, или они могут быть (например, при получении экспрессией в прокариотической клетке) негликозилированными.
Гетерогенные препараты, включающие молекулы антитела анти-IL-6, также являются частью изобретения. Например, такие препараты могут быть смесями антител с полноразмерными тяжелыми цепями и тяжелыми цепями, не имеющими С-терминального лизина, с различными степенями гликозилирования и/или с дериватизованными аминокислотами, такими как циклизация N-терминальной глутаминовой кислоты, чтобы сформировать остаток пироглутаминовой кислоты.
Как используется в настоящей заявке, фраза "существенно как изложено" обозначает, что характеристика(и)соответствующих CDR VH или VL домена связывающих членов, описанных в настоящей заявке, будет идентичной или очень подобной указанным участкам, последовательность которых изложена в настоящей заявке. Как описано в настоящей заявке, фраза "очень подобный" относительно указанного участка (ов) одного или нескольких вариабельных доменов, подразумевает, что могут быть сделаны от 1 до приблизительно 5, например, от 1 до 4, включая 1-3, или 1 или 2, или 3 или 4, аминокислотные замены в CDR и/или VH или VL домена.
Краткое описание иллюстрации
Фигура 1. На этой фигуре показано, что эффект введения антитела анти-IL-6 (Антитело 18) на человеческом рекомбинантном IL-6 вызвало увеличение уровня гаптоглобина у мыши in vivo.
Детальное описание изобретения
Как отмечено выше, связывающий член в соответствии с настоящим изобретением модулирует и может нейтрализовать биологическую активность IL-6. Как описано в настоящей заявке, IL-6-связывающие члены согласно настоящему изобретению могут быть оптимизированы для нейтрализирующей силы действия. Вообще, оптимизация силы действия вовлекает мутацию последовательности выбранного связывающего члена (обычно последовательности вариабельного домена антитела), чтобы получить библиотеку связывающих членов, которые затем оцениваются на силу действия, и отбираются более мощные связывающие члены. Таким образом отобранные "оптимизированные по силе действия" связывающие члены имеют тенденцию иметь более высокую силу действия, чем связывающий член, от которого была получена библиотека. Однако связывающие члены с высокой силой действия могут также быть получены без оптимизации, например связывающий член с высокой силой действия может быть получен непосредственно из начального скрининга, например, испытания биохимической нейтрализации. "Оптимизированный по силе действия" связывающий член касается связывающего члена с оптимизированной силой действия нейтрализации специфической активности или нисходящей функции IL-6. Испытание и силы действия описаны более подробно в настоящей заявке в другом месте. Настоящее изобретение предлагает и оптимизированные по силе действия и неоптимизированные связывающие члены, а также способы оптимизации силы действия от отобранного связывающего члена. Настоящее изобретение таким образом позволяет квалифицированному специалисту получать связывающие члены, обладающие высокой силой действия.
В дальнейшем аспекте настоящее изобретение предлагает способ получения одного или нескольких связывающих членов, способных связывать антиген, способ, включающий контактирование библиотеки связывающих членов согласно изобретению, и указанного антигена, и отбор одного или нескольких связывающих членов библиотеки, способных связывать указанный антиген.
Библиотека может быть показана на частицах или молекулярных комплексах, например реплицируемых генетических пакетах, таких как дрожжевые, бактериальные или бактериофаговые (например, Т7) частицы, вирусы, клетки или ковалентные, рибосомные или другие системы дисплея in vitro, каждая частица или молекулярный комплекс, содержащий нуклеиновую кислоту, кодирующую VH вариабельный домен антитела, показанный на ней, и необязательно также показанный домен VL при наличии. Фаговый дисплей описан в WO92/01047, и, например, патентах США US5969108, US5565332, US5733743, US5858657, US5871907, US5872215, US5885793, US5962255, US6140471, US6172197, US6225447, US6291650, US6492160 и US6521404, каждый из которых включен в настоящую заявку путем ссылки в их полном объеме.
После выбора связывающих членов, способных связывать антиген и показанных на бактериофаге или других частицах библиотеки или молекулярных комплексах, нуклеиновая кислота может быть взята из бактериофага или другой частицы или молекулярного комплекса, показывающего указанный отобранный связывающий член. Такая нуклеиновая кислота может использоваться в последующем производстве связывающего члена или VH или VL вариабельного домена антитела путем экспрессии из нуклеиновой кислоты с последовательностью нуклеиновой кислоты, взятой из бактериофага или другой частицы или молекулярного комплекса, показывающего указанный отобранный связывающий член.
VH вариабельный домен антитела с аминокислотной последовательностью VH вариабельного домена антитела указанного отобранного связывающего члена может быть предоставлен в выделенной форме, как и связывающий член, включающий такой VH домен.
Может в дальнейшем испытываться способность связывать IL-6, а также способность конкурировать, например, с родительской молекулой антитела или молекулой антитела 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23 (например, в формате scFv и/или формате IgG, например, IgG1) для связывания с IL-6. Может испытываться способность нейтрализовать IL-6, как обсуждается далее в настоящей заявке в другом месте.
Связывающий член согласно настоящему изобретению может связывать IL-6 с аффинностью родительской или другой молекулы антитела, например scFv, или одного из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, например IgG1, или с аффинностью, которая лучше.
Связывающий член согласно настоящему изобретению может нейтрализовать биологическую активность IL-6 с силой действия родительской или другой молекулы антитела, одного из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, например scFv, или IgG1, или с силой действия, которая лучше.
Связывающая аффинность и сила действия нейтрализации различных связывающих членов могут сравниваться при соответствующих условиях.
Варианты VH и VL доменов и CDR настоящего изобретения, включая те, для которых аминокислотные последовательности изложены в настоящей заявке и которые могут использоваться в связывающих членах изобретения, могут быть получены посредством методов изменения последовательности или мутации и скрининга на антиген-связывающие члены с желательными характеристиками. Примеры желательных характеристик включают, кроме прочих:
- увеличенную связывающую аффинность к антигену относительно известных антител, которые являются специфичными для антигена;
- увеличенную нейтрализацию активности антигена относительно известных антител, которые являются специфичными для антигена, если активность известна;
- указанную конкурентную способность с известным антителом или лигандом к антигену при специфичном молярном отношении;
- способность к иммунопреципитации комплекса;
- способность связывать указанный эпитоп.
Линейный эпитоп, например последовательность пептида, идентифицированный с использованием сканирования связывания пептида, описанного в настоящей заявке, например, с использованием пептидов, скринингованных в линейной и/или ограниченной конформации;
конформационный эпитоп, сформированный ненепрерывными остатками;
способность модулировать новую биологическую активность IL-6 или нисходящей молекулы.
Такие способы также предлагаются в настоящей заявке.
Варианты молекул антитела, раскрытые в настоящей заявке, могут быть произведены и использоваться в настоящем изобретении. После лидерства вычислительной химии в применении многомерных методов анализа данных к отношениям структура/свойство-активность [81] количественные соотношения активность-свойство антител могут быть получены с использованием известных математических методов, таких как статистическая регрессия, распознавание образов и классификация [82, 83, 84, 65, 86, 87]. Свойства антител могут быть получены из эмпирических и теоретических моделей (например, анализ вероятных контактных остатков, или вычисленное физико-химическое свойство) последовательности антитела, функциональных и трехмерных структур, и эти свойства можно рассмотреть отдельно и в комбинации.
Антиген-связывающий участок антитела, состоящий из VH домена и VL домена, типично формируется шестью петлями полипептида: тремя из вариабельного домена легкой цепи (VL) и тремя из вариабельного домена тяжелой цепи (VH). Анализ антител известного атомного строения объяснил отношения между последовательностью и трехмерной структурой антитело-комбинирующих сайтов [88, 89]. Эти отношения подразумевают, что за исключением третьего участка (петли) в VH доменах, петли связывающего участка имеют одну из небольшого количества конформаций главной цепи: канонические структуры. Каноническая структура, сформированная в специфической петле, как показано, определяется ее размером и присутствием определенных остатков на ключевых участках и в петле, и в каркасных участках [88, 89].
Это исследование отношения последовательность-структура может использоваться для прогнозирования тех остатков в антителе известной последовательности, но неизвестной трехмерной структуры, которые важны в поддержании трехмерной структуры его петель CDR и, следовательно, поддерживают связывающую специфичность. Эти прогнозы могут быть поддержаны путем сравнения прогнозов с результатом из ведущих экспериментов по оптимизации. В структурном подходе модель может быть создана из молекулы антитела [90] с использованием любого свободно доступного или коммерческого пакета, такого как WAM [91]. Пакет программ анализа и визуализации белка, такой как Insight II (Accelrys, Inc.) или Deep View [92] могут тогда использоваться для оценки возможных замен в каждом положении в CDR. Эта информация может затем использоваться, чтобы сделать замены вероятно с минимальным или благоприятным воздействием на активность.
Методы, требуемые для осуществления замен в пределах аминокислотных последовательностей CDR, VH или VL доменов антитела и связывающих членов, вообще доступны в данной области. Могут быть сделаны вариантные последовательности с заменами, для которых можно или не возможно предсказать, что они имеют минимальное или благоприятное воздействие на активность, и их проверяют на способность связывать и/или нейтрализовать IL-6 и/или на любое другое желательное свойство.
Варианты аминокислотной последовательности вариабельного домена любого VH и VL домена, последовательности которых определенно раскрыты в настоящей заявке, могут использоваться в соответствии с настоящим изобретением, как обсуждается.
Варианты VL доменов изобретения и связывающие члены или молекулы антитела, включающие их, включают VL домены, в которых Аргинин не присутствует в остатке Kabat 108, например, где остаток Kabat 108 имеет другой остаток или он удален. Например, молекула антитела, такая как молекула антитела, не имеющая константного домена, например scFv, может включать VL домен, имеющий последовательность VL домена или его вариант, описанный в настоящей заявке, в котором Аргинин в остатке Kabat 108 аминокислотный остаток, отличный от Аргинина, или удален.
Дальнейший аспект изобретения представляет собой молекулу антитела, включающую VH домен, у которого есть по крайней мере 60, 70, 80, 85, 90, 95, 98 или 99%-ная идентичность аминокислотной последовательности с VH доменом любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, показанной в приложенном списке последовательности, и/или включающую VL домен, у которого есть по крайней мере 60, 70, 80, 85, 90, 95, 98 или 99%-ная идентичность аминокислотной последовательности с VL доменом любого из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, показанных в приложенном списке последовательности. Алгоритмы, которые могут использоваться для вычисления % идентичности двух аминокислотных последовательностей, включают, например, BLAST [93], FASTA [94], или алгоритм Смита-Ватермана [95], например, используя параметры по умолчанию.
Специфические варианты могут включать одно или несколько изменений аминокислотной последовательности (дополнение, делецию, замену и/или инсерцию аминокислотного остатка).
Изменения могут быть сделаны в одном или нескольких каркасных участках и/или одном или нескольких CDR. Изменения обычно не приводят к потере функции, таким образом, связывающий член, включающий измененную таким образом аминокислотную последовательность, может сохранить способность связывать и/или нейтрализовать IL-6. Он может сохранить то же самое количественное связывание и/или нейтрализацию способности как связывающий член, в котором не сделано изменение, например, как определено в испытании, описанном в настоящей заявке. У связывающего члена, включающего измененную таким образом аминокислотную последовательность, может быть улучшенная способность связывать и/или нейтрализовать IL-6.
Изменение может включить замену одного или нескольких аминокислотных остатков на неприродную или нестандартную аминокислоту, модификацию одного или нескольких аминокислотных остатков в неприродную или нестандартную форму, или инсерцию одной или нескольких неприродных или нестандартных аминокислот в последовательности. Примеры чисел и положений изменений в последовательностях изобретения описаны в другом месте в настоящей заявке. Природные аминокислоты включают 20 "стандартных" L-аминокислот, идентифицированных как G, А, V, L, I, М, Р, F, W, S, Т, N, Q, Y, С, K, R, H, D, Е по их стандартным однобуквенным кодам. Нестандартные аминокислоты включают любой другой остаток, который может быть включен в основную цепь полипептида или быть результатом модификации существующего остатка аминокислоты. Нестандартные аминокислоты могут встречаться в природе или не встречаться в природе. Несколько природных нестандартных аминокислот известны в данной области, такие как 4-гидроксипролин, 5-гидроксилизин, 3-метилгистидин, N-ацетилсерин и т.д. [96]. Те аминокислотные остатки, которые являются дериватизированными в их N-альфа-положении, будут расположены только в N-конце аминокислотной последовательности. Обычно в настоящем изобретении аминокислота - L-аминокислота, но это может быть D-аминокислота. Изменение поэтому может включить модификацию L-аминокислоты в D-аминокислоту, или ее замену на D-аминокислоту. Метилированные, ацетилированные и/или фосфорилированные формы аминокислот также известны, и аминокислоты в настоящем изобретении могут подвергаться такой модификации.
Аминокислотные последовательности в доменах антитела и связывающих членах изобретения могут включать неприродные или нестандартные аминокислоты, описанные выше. Нестандартные аминокислоты (например, D-аминокислоты) могут быть включены в аминокислотную последовательность во время синтеза, или путем модификации или замены "оригинальных" стандартных аминокислот после синтеза аминокислотной последовательности.
Использование нестандартных и/или неприродных аминокислот увеличивает структурное и функциональное разнообразие и может таким образом увеличить потенциал для достижения желаемых IL-6-связывающих и нейтрализующих свойств в связывающем члене изобретения. Дополнительно, у D-аминокислот и аналогов, как показано, различные фармакокинетические профили по сравнению со стандартными L-аминокислотами, вследствие in vivo деградации полипептидов, имеющих L-аминокислоты, после введения животному, например человеку, что означает, что D-аминокислоты выгодны для некоторых применений in vivo.
Новые VH или VL участки, несущие CDR-полученные последовательности изобретения, могут быть произведены с использованием случайного мутагенеза одного или нескольких отобранных VH и/или VL генов, чтобы получить мутации в пределах всего вариабельного домена. Такая техника описана Gram и др. [97], которые использовали подверженную ошибкам ПЦР. В некоторых вариантах осуществления одна или две аминокислотные замены делаются в пределах всего вариабельного домена или набора CDR.
Другой метод, который может использоваться, должен направить мутагенез на участки CDR VH или VL генов. Такие методы раскрыты Barbas и соавт. [98] и Schier и соавт. [99].
Все вышеописанные методы известны также в данной области, и квалифицированный специалист будет в состоянии использовать такие методы для обеспечения связывающих членов изобретения, используя обычную методологию в области.
Дальнейший аспект изобретения предлагает способ для получения антиген-связывающего участка антитела для IL-6, способ, включающий обеспечение посредством дополнения, делеции, замены или инсерции одной или нескольких аминокислот в аминокислотной последовательности VH домена, изложенной в настоящей заявке, VH домена, который является вариантом аминокислотной последовательности VH домена, необязательно комбинируя VH домен, таким образом предоставленный, с одним или несколькими VL доменами, и испытывая VH домен или комбинацию VH/VL или комбинации для идентификации связывающего члена или антиген-связывающего участка антитела для IL-6 и необязательно с одной или несколькими желательными свойствами, например способностью нейтрализовать активность IL-6. Указанный VL домен может иметь аминокислотную последовательность, которая существенно такая, как изложено в настоящей заявке. Может использоваться аналогичный метод, в котором один или несколько вариантов последовательности VL домена, раскрытого в настоящей заявке, комбинированы с одним или несколькими VH доменами.
Как отмечено выше, аминокислотная последовательность CDR, существенно такая, как изложена в настоящей заявке, может быть представлена в качестве CDR в вариабельном домене человеческого антитела или его существенной части. Последовательности HCDR3, существенно такие, как изложено в настоящей заявке, представляют варианты осуществления настоящего изобретения, и каждый из них может содержаться как HCDR3 в вариабельном домене тяжелой цепи человека или его существенной части.
Вариабельные домены, используемые в изобретении, могут быть получены или происходить от любого зародышевого или перегруппированного человеческого вариабельного домена, или могут быть синтетическим вариабельным доменом, основанным на консенсусных или фактических последовательностях известных человеческих вариабельных доменов. Вариабельный домен может быть получен из нечеловеческого антитела. Последовательность CDR изобретения (например, CDR3), может быть введена в репертуар вариабельных доменов, не имеющих CDR (например, CDR3), используя технологию рекомбинантной ДНК. Например, Marks и др. [100] описывают методы производства репертуаров вариабельных доменов антитела, в которых консенсусные праймеры, направленные на или смежную с 5' концом область вариабельного домена, используются в соединении с консенсусными праймерами к третьему каркасному участку человеческих VH генов, чтобы обеспечить репертуар VH вариабельных доменов, не имеющих CDR3. Marks и др. далее описывают, как этот репертуар может быть объединен с CDR3 специфического антитела. Используя аналогичные методы, CDR3-полученные последовательности настоящего изобретения могут быть перетасованы с репертуарами VH или VL доменов, не имеющих CDR3, и перетасованными полными VH или VL доменами, объединенными с родственным VL или VH доменом, чтобы обеспечить связывающие члены по изобретению. Репертуар может затем быть представлен в подходящей системе хозяина, такой как система фагового дисплея WO92/01047, который включен в настоящую заявку путем ссылки в полном объеме, или любого последующего большого объема литературы, включая Kay, Winter & McCafferty [101] так, чтобы могли быть отобраны подходящие связывающие члены. Репертуар может состоять из от 104 индивидуальных членов вверх, например, по крайней мере 105, по крайней мере 106, по крайней мере 107, по крайней мере 108, по крайней мере 109 или по крайней мере 1010 членов или больше. Другие подходящие системы хозяина включают, кроме прочих, дрожжевой дисплей, бактериальный дисплей, Т7 дисплей, вирусный дисплей, клеточный дисплей, рибосомный дисплей и ковалентный дисплей.
Предлагается способ получения связывающего члена для антигена IL-6, который включает:
(a) обеспечение стартового репертуара нуклеиновых кислот, кодирующих VH домен, который или включает CDR3, который будет заменен, или не содержит CDR3 кодирующего участка;
(b) комбинирование указанного репертуара с нуклеиновой кислотой донора, кодирующей аминокислотную последовательность, существенно такую, как изложено в настоящей заявке для VHA CDR3 таким образом, что указанная нуклеиновая кислота донора вставляется в участок CDR3 в репертуаре, чтобы обеспечить репертуар продукта нуклеиновых кислот, кодирующих VH домен;
(c) экспрессию нуклеиновых кислот указанного репертуара продукта;
(d) отбор связывающего члена для IL-6; и
(e) выделение указанного связывающего члена или нуклеиновой кислоты, кодирующей его.
Снова может использоваться аналогичный способ, в котором VL CDR3 по изобретению объединен с репертуаром нуклеиновых кислот, кодирующих VL домен, которые или включают CDR3, который будет заменен или не имеют CDR3 кодирующего участка.
Подобным образом, один или несколько, или все три CDR могут быть привиты в репертуар VH или VL доменов, которые затем проходят скрининг относительно связывающего члена или связывающих членов для IL-6.
Например, могут использоваться один или несколько HCDR1, HCDR2 и HCDR3 родительского или антитела 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23 или набора HCDR родительского или антитело 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23, и/или могут использоваться один или несколько LCDR1, LCDR2 и LCDR3 родительского или антитела 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23 или набор LCDR родительского или антитело 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 или 23.
Подобным образом, могут использоваться другие VH и VL домены, наборы CDR и наборы HCDR и/или наборы LCDR, раскрытые в настоящей заявке.
Существенная часть вариабельного домена иммуноглобулина может включать по крайней мере три участка CDR, вместе с их промежуточными каркасными участками. Часть может также включать по крайней мере приблизительно 50% одного или обоих первого и четвертого каркасного участков, 50% является C-терминальным 50% первого каркасного участка и N-терминальным 50% четвертого каркасного участка. Дополнительные остатки на N-терминальном или C-терминальном конце существенной части вариабельного домена могут быть теми, которые обычно не ассоциируются с природными участками вариабельного домена. Например, конструирование связывающих членов согласно настоящему изобретению с помощью методов рекомбинантной ДНК может привести к введению N- или C-терминальных остатков, закодированных линкерами, введенными для облегчения клонирования или других стадий манипуляции. Другие стадии манипуляции включают введение линкеров, чтобы присоединить вариабельные домены изобретения к дальнейшим последовательностям белка, включая константные участки антитела, другие вариабельные домены (например, при производстве диател) или обнаружимые/функциональные метки, как обсуждается в настоящей заявке более подробно в другом месте.
Хотя в небольшом количестве аспектов изобретения, связывающие члены включают пару VH и VL доменов, единственные связывающие домены, основанные на последовательностях или VH или VL домена, формируют дальнейшие аспекты изобретения. Известно, что единственные иммуноглобулиновые домены, особенно VH домены, способны к связыванию антигенов-мишеней определенным способом. Например, см. обсуждение dAb выше.
В случае любого из единственных связывающих доменов эти домены могут использоваться для скрининга на комплементарные домены, способные к формированию двухдоменного связывающего члена, способного связывать IL-6. Это может быть достигнуто путем методов скрининга фагового дисплея с использованием так называемого иерархического двойного комбинаторного подхода, как раскрыто в WO92/01047, включенном в настоящей заявке путем ссылки в полном объеме, в котором используется индивидуальная колония, содержащая клон или Н или цепи L, чтобы инфицировать всю библиотеку клонов, кодирующих другую цепь (L или Н), и получающийся двухдоменный связывающий член отбирается в соответствии с методами фагового дисплея, такими как описанные в той ссылке. Эта техника также раскрыта в Marks и др., там же [100].
Связывающие члены согласно настоящему изобретению могут далее включить константные участки антитела или их части, например константные участки человеческого антитела или их части. Например, VL домен может быть присоединен на своем C-терминальном конце к константным доменам легкой цепи антитела, включая человеческие Cκ или Cλ цепи. Подобным образом, связывающий член, основанный на VH домене, может быть присоединен на своем C-терминальном конце ко всей или части (например, СН1 домену) тяжелой цепи иммуноглобулина, полученной из любого изотипа антитела, например IgG, IgA, IgE и IgM и любого из подклассов изотипа, особенно IgG1 и IgG4. IgG1 выгоден из-за его эффекторной функции и легкости получения. Любой синтетический или другой вариант константного участка, который имеет эти свойства и стабилизирует вариабельные участки, может также быть полезным в настоящем изобретении.
Связывающие члены согласно изобретению могут быть мечены обнаруживаемыми или функциональными метками. Таким образом, связывающий член или молекула антитела могут присутствовать в форме иммуноконъюгата, чтобы получить обнаружимый и/или измеримый сигнал. Иммуноконъюгаты могут включать молекулу антитела изобретения, конъюгированную с обнаружимой или функциональной меткой. Метка может быть любой молекулой, которая продуцирует, или ее можно заставить продуцировать сигнал, включая, кроме прочих, люминофоры, радиоактивные метки, ферменты, хемилюминофоры или фотосенсибилизаторы. Таким образом, связывание может быть обнаружено и/или измерено путем обнаружения флюоресценции или люминесценции, радиоактивности, ферментной активности или световой абсорбции.
Подходящие метки включают, посредством иллюстрации и кроме прочих,
- ферменты, такие как щелочная фосфатаза, глюкоза-6-фосфатдегидрогеназа ("G6PDH"), альфа-D-галактозидаза, глюкозоксидаза, глюкозамилаза, карбоангидраза, ацетилхолинэстераза, лизозим, малатдегидрогеназа и пероксидаза, например пероксидаза хрена;
- красители;
- флуоресцентные метки или люминофоры, такие как флуоресцеин и его производные, флуорохром, соединения и производные родамина, GFP (GFP для "Зеленого Флуоресцентного Белка"), данзил, умбеллиферон, фикоэритрин, фикоцианин, аллофикоцианин, о-фтальальдегид и флуоресцамин; флуорофоры, такой как лантанид криптаты и хелаты, например Европий и т.д (Perkin Elmer and Cis Biointernational),
- хемилюминесцентные метки или хемилюминофоры, такие как изолюминол, люминол и диоксетаны;
- био-люминесцентные метки, такие как люцифераза и люциферин;
- сенсибилизаторы;
- коферменты;
- субстраты ферментов;
- радиоактивные метки, включая кроме прочих бром77, углерод14, кобальт57, флюорин8, галлий67, галлий 68, углерод3 (тритий), индий111, индий 113 м., йод123m, йод125, йод126, йод131, йод133, ртуть107, ртуть203, фосфор32, рений99m, рений101, рений105, рутений95, рутений97, рутений103, рутений105, скандий47, селен75, сера35, технеций99, технеций99m, теллур121m, теллур122m, теллур125m, тулий165, тулий167, тулий168, иттрий199 и другие радиоактивные метки, упомянутые в настоящей заявке;
- частицы, такие как латексные или углеродистые частицы; соли металлов; кристаллит; липосомы; клетки, и т.д., которые могут быть далее мечены красителем, катализатором или другой обнаружимой группой;
- молекулы, такие как биотин, дигоксигенин или 5-бромодезоксиуридин;
- остатки токсинов, такие как, например, остаток токсина, отобранный из группы экзотоксина Pseudomonas (РЕ или его цитотоксического фрагмента или мутанта), токсина Diptheria или его цитотоксического фрагмента или мутанта, ботулотоксина А, В, С, D, Е или F, рицина или его цитотоксического фрагмента, например рицина А, абрина или его цитостатического фрагмента, сапорина или его цитотоксического фрагмента, антивирусного токсина лаконоса или его цитотоксического фрагмента и бриодина 1 или его цитотоксического фрагмента.
Подходящие ферменты и коферменты раскрыты в Litman, и соавт., US 4275149, и Boguslaski, и соавт., US 4318980, каждый из которых включен в настоящую заявку путем ссылки в их полном объеме. Подходящие люминофоры и хемилюминофоры раскрыты в Litman, и соавт., US4275149, который включен в настоящую заявку путем ссылки в полном объеме. Метки далее включают химические остатки, такие как биотин, который может быть обнаружен через связывание со специфичным родственным обнаруживаемым остатком, например меченным авидином или стрептавидином. Обнаруживаемые метки могут быть присоединены к антителам изобретения с использованием обычной химии, известной в данной области.
Иммуноконъюгаты или их функциональные фрагменты могут быть подготовлены методами, известными специалисту, квалифицированному в данной области. Они могут быть спарены с ферментами или с флуоресцентными метками непосредственно или путем посредника группы-спейсера или связывающей группой, такой как полиальдегид, как глутаральдегид, этилендиаминтетрауксусная кислота (EDTA), диэтилен-триаминпентауксусная кислота (DPTA), или в присутствии агентов спаривания, таких как упомянутые выше для терапевтических конъюгатов. Конъюгаты, содержащие метки типа флуоресцеина, могут быть подготовлены реакцией с изотиоцианатом.
Методы, известные специалисту, квалифицированному в данной области, существующие для спаривания терапевтических радиоизотопов с антителами или непосредственно или через хелатирующего агента, такого как EDTA, DTPA, упомянутый выше, могут использоваться для радиоэлементов, которые могут использоваться в диагностике. Аналогично, возможно выполнить мечение с натрием125 методом хлорамина Т [102] или иначе технецием99 т техникой Crockford и соавт., (US 4424200, включен в настоящую заявку путем ссылки в полном объеме) или присоединением через DTPA, как описано Hnatowich (US 4479930, включен в настоящую заявку путем ссылки в полном объеме).
Существуют многочисленные методы, которыми метка может продуцировать сигнал, обнаруживаемый внешними средствами, например визуальной экспертизой, электромагнитным облучением, высокой температурой и химическими реактивами. Метка может также быть связана с другим связывающим членом, который связывает антитело изобретения, или с основой.
Метка может непосредственно продуцировать сигнал и поэтому дополнительные компоненты не обязаны давать сигнал. Многочисленные органические молекулы, например люминофоры, в состоянии поглощать ультрафиолетовый и видимый свет, при этом абсорбция света передает энергию этим молекулам и переводит их в возбужденное энергетическое состояние. Эта поглощенная энергия затем рассеивается эмиссией света на второй длине волны. Эта эмиссия второй длины волны может также передать энергию меченой акцепторной молекуле, и полученная энергия рассеивается от акцепторной молекулы эмиссией света, например передача энергии резонанса флюоресценции (FRET). Другие метки, которые непосредственно продуцируют сигнал, включают радиоактивные изотопы и красители.
Альтернативно, метка, возможно, нуждается в других компонентах, чтобы дать сигнал, и система продуцирования сигнала тогда включала бы все компоненты, требуемые для получения измеримого сигнала, которая может включать субстраты, коферменты, усиливающие агенты, дополнительные ферменты, вещества, которые реагируют с ферментными продуктами, катализаторы, активаторы, кофакторы, ингибиторы, уловители, ионы металлов, определенные связывающие вещества, требуемые для связывания генерирующих сигнал веществ. Детальное обсуждение подходящих систем производства сигнала может быть найдено в Ullman, и соавт. US 5185243, который включен в настоящую заявку путем ссылки в полном объеме.
Настоящее изобретение предлагает способ, включающий предоставление основания или возможности связывания связывающего члена, как предусмотрено в настоящей заявке с IL-6. Как отмечено, такое связывание может иметь место in vivo, например, после введения связывающего члена, или нуклеиновой кислоты, кодирующей связывающий член, или это может иметь место in vitro, например, в ELISA, Вестерн блоте, иммуноцитохимии, иммунопреципитации, аффинной хроматографии, и биохимическом или клеточном испытании, таком как испытание пролиферации TF-1 клеток.
Настоящее изобретение также предусматривает измерение уровней антигена непосредственно путем применения связывающего члена согласно изобретению, например, в системе биосенсора. Например, настоящее изобретение включает способ обнаружения и/или измерения связывания с IL-6, включающий (i) экспозицию указанного связывающего члена IL-6 и (ii) обнаружение связывания указанного связывающего члена с IL-6, причем связывание обнаруживается с использованием любого метода или обнаружимой метки, описанной в настоящей заявке. Этот и любой другой метод обнаружения связывания, описанный в настоящей заявке, могут интерпретироваться непосредственно человеком, выполняющим метод, например, путем визуального наблюдения обнаруживаемой метки. Альтернативно, этот метод, или любой другой метод обнаружения связывания, описанный в настоящей заявке, может предоставить отчет в виде авторадиограммы, фотографии, компьютерной распечатки, отчета проточной цитометрии, графика, диаграммы, пробирки или контейнера или лунки, содержащий результат, или любого другого визуального или физического представления результата метода.
Может быть определено количество связывания связывающего члена с IL-6. Количественное определение может касаться количества антигена в испытательном образце, который может представлять диагностический интерес. Скрининг на связывание IL-6 и/или его количественное определение может быть полезным, например в скрининге пациентов на болезни или расстройства, упомянутые в настоящей заявке и/или любой другой болезни или расстройства, включающее аберрантную экспрессию и/или активность IL-6.
Диагностический метод изобретения может включать (i) получение образца ткани или жидкости от субъекта, (ii) экспозицию указанного образца ткани или жидкости одному или нескольким связывающим членам настоящего изобретения; и (iii) обнаружение связанного IL-6 в сравнении с контрольным образцом, причем увеличение количества связывания IL-6 по сравнению с контролем может указывать на аберрантный уровень экспрессии или активности IL-6. Образцы ткани или жидкости, которые будут проходить испытание, включают кровь, сыворотку, мочу, материал биопсии, опухоли, или любую ткань, подозреваемую на содержание аберрантных уровней IL-6. Субъекты, дающие положительный результат на аберрантные уровни или активность IL-6, могут также извлечь пользу из способов лечения, раскрытых в настоящей заявке ниже. Квалифицированные специалисты в данной области смогут выбрать подходящий способ определения связывания связывающего члена с антигеном согласно их предпочтению и общему знанию в свете методов, раскрытых в настоящей заявке.
Реактивности связывающих членов в образце могут быть определены любыми соответствующими средствами. Радиоиммунологическое исследование (RIA) представляет собой одну из возможностей. Радиоактивный меченый антиген смешивается с немеченым антигеном (испытательный образец) и ему позволяют связаться со связывающим членом. Связанный антиген физически отделяется от несвязанного антигена, и определяется количество радиоактивного антигена, связанного со связывающим членом. Чем больше антигена находится в испытуемом образце, тем меньше радиоактивного антигена свяжется со связывающим членом. Испытание конкурентного связывания может также использоваться с нерадиоактивным антигеном с использованием антигена или аналога, связанного с сигнальной молекулой. Сигнальная молекула может представлять собой флуорохромом, фосфор или лазерный краситель со спектрально изолированными характеристиками абсорбции или эмиссии. Подходящие флуорохромы включают флуоресцеин, родамин, фикоэритрин и Техасский Красный, и хелаты или криптаты лантанида. Подходящие хромогенные красители включают диаминобензидин.
Другие сигнальные молекулы включают макромолекулярные коллоидные частицы или макрочастицы, такие как латексные гранулы, которые окрашены, магнитные или парамагнитные, и биологически или химически активные вещества, которые могут прямо или косвенно обусловить визуальное наблюдение, электронное обнаружение или иную регистрацию обнаружимых сигналов. Эти молекулы могут быть ферментами, которые катализируют реакции, которые развиваются, или изменяют окрашивание или, например, вызывают изменения в электрических свойствах. Они могут быть молекулярно легко возбудимыми таким образом, что электронные переходы между состояниями энергии приводят к характерным спектральным абсорбциям или эмиссиям. Они могут включать химические вещества, используемые в соединении с биосенсорами. Могут использоваться биотин/авидин или биотин/стрептавидин и системы обнаружения щелочной фосфатазы.
Сигналы, произведенные индивидуальными конъюгатами связывающий член-сигнальная молекула, могут использоваться для получения измеримых абсолютных или относительных данных связывания соответствующего связывающего члена в образцах (нормальном и испытуемом).
Комплект, включающий связывающий член согласно любому аспекту или варианту осуществления настоящего изобретения, также предлагается как аспект настоящего изобретения. В комплекте связывающий член может быть меченым, чтобы позволить определить его реактивности в образце, например, как описано далее ниже. Кроме того, связывающий член может быть присоединен к твердой основе или возможно нет. Компоненты комплекта, как правило, стерильны и находятся в запаянных флаконах или других контейнерах. Комплекты могут использоваться в диагностическом анализе или других методах, для которых полезны связывающие члены. Комплект может содержать инструкции для использования компонентов в методе, например методе в соответствии с настоящим изобретением. Вспомогательные материалы для того, чтобы помочь или позволить выполнить такой метод, могут быть включены в комплект изобретения. Вспомогательные материалы включают второй, отличный связывающий член, который связывается с первым связывающим членом и конъюгирован с обнаружимой меткой (например, Флуоресцентной меткой, радиоактивным изотопом или ферментом). Основанные на антителе комплекты могут также включать гранулы для проведения иммунопреципитации. Каждый компонент комплектов находится, как правило, в своем собственном подходящем контейнере. Таким образом, эти комплекты вообще включают разные контейнеры, подходящие для каждого связывающего члена. Кроме того, комплекты могут включать инструкции для выполнения испытания и методы для интерпретации и анализа данных, следующих из выполнения испытания.
Настоящее изобретение также предоставляет применение вышеуказанного связывающего члена для измерения уровней антигена в конкурентном количественном определении, то есть метод измерения уровня антигена в образце с применением связывающего члена, предоставленного настоящим изобретением, в конкурентном количественном определении. Это может применяться, если не требуется физическое отделение связанного антигена от несвязанного. Одной из возможностей является присоединение сигнальной молекулы к связывающему члену так, чтобы при связывании происходили физические или оптические изменения. Сигнальная молекула может прямо или косвенно генерировать воспринимаемые сигналы с возможностью количественной оценки. Присоединение сигнальной молекулы может быть прямым или косвенным, ковалентным, напр., пептидной связью, или нековалентным. Соединение посредством пептидной связи может быть результатом рекомбинантной экспрессии слияния генов, кодирующих антитело и сигнальную молекулу.
В различных аспектах и реализациях, настоящее изобретение распространяется на связывающий член, конкурирующий за связывание с IL-6 с любым из связывающих членов, определенных здесь, например с родительским антителом или любым из антител 2, 3, 4, 5, 7, 8, 10, 14, 16, 17, 18, 19, 21, 22 и 23, например, в виде IgG1. Конкуренция между связывающими членами может быть легко определена in vitro, например, посредством маркирования специфической сигнальной молекулой одного из связывающих членов, который может быть обнаружен в присутствии других немаркированных связывающих членов, с тем чтобы обеспечить идентификацию членов, связывающихся с тем же эпитопом или с перекрывающимися эпитопами. Конкуренция может быть определена, к примеру, с помощью ELISA, в котором IL-6 иммобилизуют на пластинке и добавляют первый маркированный или меченый связывающий член вместе с одним или более других немаркированных или немеченых связывающих членов. Присутствие немаркированного связывающего члена, конкурирующего с маркированным связывающим членом, обнаруживается по погашению сигнала, испускаемого маркированным связывающим членом.
Например, настоящее изобретение включает метод идентификации веществ, связывающихся с IL-6, включающий (i) иммобилизацию IL-6 на подложке, (ii) контакт иммобилизованного IL-6, одновременно или пошагово, как минимум с одним маркированным или меченым связывающим членом согласно изобретению и с одним или более немаркированными или немечеными испытуемыми связывающими членами, и (iii) идентификацию нового связывающегося с IL-6 вещества по уменьшению количества связанной метки из маркированного связывающего члена. Этот метод может осуществляться высокопроизводительным способом с использованием многоячеистых пластинок или комбинаторных матриц. Этот метод анализа также может осуществляться в растворе. См., к примеру, патент США 5814468, включенный в данную заявку полностью. Как описано выше, обнаружение связывания может интерпретироваться непосредственно персоналом, осуществляющим анализ, например, посредством визуального наблюдения обнаружимой метки или уменьшения ее присутствия. Или же, методы связывания согласно изобретению могут производить отчет в виде авторадиографии, фотографии, компьютерной распечатки, отчета потоковой цитометрии, графика, диаграммы, пробирки, контейнера или ячейки, содержащей результат, или любого другого визуального или физического представления результата метода.
Конкурентный количественный анализ также может использоваться для картирования эпитопов. В одном случае картирование эпитопов могло использоваться для идентификации эпитопа, связанного со связывающимся с IL-6 членом, который может опционально иметь оптимизированные нейтрализующие и/или модулирующие характеристики. Такой эпитоп может быть линейным или конформационным. Конформационный эпитоп может содержать как минимум два различных фрагмента IL-6, в котором указанные фрагменты располагаются поблизости один от другого, когда IL-6 складывается в третичную или четвертичную структуру, образуя конформационный эпитоп, который распознается ингибитором IL-6, таким как IL-6-связывающий член. В конкурентном анализе может применяться пептидный фрагмент антигена, особенно пептид, включающий или состоящий исключительно из интересующего эпитопа. Может использоваться пептид, имеющий последовательность эпитопа плюс одну или более аминокислот с любого конца. Связывающие члены, согласно настоящему изобретению, могут быть такими, что их связывание с антигеном ингибируется пептидом, имеющим или включающим данную последовательность.
Далее, настоящее изобретение предоставляет выделенную нуклеиновую кислоту, кодирующую связывающий член согласно настоящему изобретению. Нуклеиновая кислота может включать ДНК и/или РНК. Вкратце, настоящее изобретение предоставляет нуклеиновую кислоту, которая кодирует CDR, или ряд CDR, или VH домен, или VL домен, или антитело-антиген-связывающий центр, или молекулу антитела, например scFv или IgG1, как определено выше согласно изобретению.
Настоящее изобретение также предоставляет конструкции в виде плазмид, векторов, транскрипционных или экспрессионных кассет, которые содержат хотя бы один из вышеуказанных полинуклеотидов.
Настоящее изобретение также предоставляет рекомбинантную клетку-хозяин, содержащую одну или более вышеуказанных конструкций. Нуклеиновая кислота, кодирующая CDR, или ряд CDR, или VH домен, или VL домен, или антитело-антиген-связывающий центр, или молекулу антитела, например scFv или IgG1, как предусмотрено, сама по себе образует аспект настоящего изобретения, равно как и метод производства кодируемого продукта, который метод включает его экспрессию из кодирующей нуклеиновой кислоты. Экспрессию удобно производить, культивируя в подходящих условиях рекомбинантные клетки-хозяева, содержащие нуклеиновую кислоту. После производства посредством экспрессии VH или VL домен, или связывающий член могут быть выделены и/или очищены с помощью любой подходящей методики, а потом использованы по назначению.
Нуклеиновая кислота согласно настоящему изобретению может включать ДНК и/или РНК и быть полностью или частично синтетической. Ссылка на нуклеотидную последовательность, как изложено здесь, охватывает молекулу ДНК с указанной последовательностью, и молекулу РНК с указанной последовательностью, в которой Т заменено на U, если контекст не требует иного.
Еще один аспект предоставляет способ производства вариабельного домена VH антитела, метод, включающий экспрессию из кодирующей нуклеиновой кислоты. Такой метод может состоять в культивировании клеток-хозяев в условиях для продукции указанной вариабельного домена VH антитела.
Аналогичные методы производства вариабельных доменов VL и связывающих членов, включающих VH и/или VL домен, предоставляются как дальнейшие аспекты настоящего изобретения.
Метод производства может включать стадию выделения и/или очистки продукта. Метод производства может включать введение продукта в композицию, включающую как минимум один добавочный компонент, такой как фармацевтически приемлемый наполнитель.
Системы для клонирования и экспрессии полипептида во множестве клеток-хозяев хорошо известны. Подходящие клетки-хозяева включают бактерии, клетки млекопитающих, клетки растений, нитчатые грибы, дрожжи, бакуловирусные системы и трансгенные растения и животные. Экспрессия антител и фрагментов антител в прокариотических клетках хорошо известна из уровня техники. См., к примеру, обзор Plückthun [103]. Обычная бактерия-хозяин представляет собой Е.coli.
Экспрессия эукариотических клеток в культуре также доступна из уровня техники в качестве опции для производства связывающих членов [104, 105, 106]. Клеточные линии млекопитающих, доступные из уровня техники для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка (СНО), клетки HeLa, клетки почки молодого хомячка, клетки мышиной меланомы NS0, клетки крысиной миеломы YB2/0, клетки почки человеческого эмбриона, клетки сетчатки человеческого эмбриона и многие другие.
Могут быть выбраны или сконструированы векторы, содержащие подходящие регулирующие последовательности, включая последовательность промотора, последовательность терминатора, полиадениловую последовательность, разделительную последовательность, генетические маркеры и другие последовательности по обстоятельствам. Векторы могут быть плазмидами, например фагмид, или же вирусными, например 'фаг, по обстоятельствам [107]. Многие известные методики и процедуры для манипуляции нуклеиновыми кислотами, например приготовление конструкций нуклеиновых кислот, мутагенез, секвенирование, введение ДНК в клетки и экспрессия генов, подробно описаны в Ausubel и др. [108].
Дальнейший аспект настоящего изобретения предоставляет клетку-хозяин, содержащую нуклеиновую кислоту как изложено здесь. Такая клетка-хозяин может быть in vitro и может быть в культуре. Такая клетка-хозяин может быть in vivo. In vivo присутствие клеток-хозяев может позволять внутриклеточную экспрессию связывающих членов согласно настоящему изобретению в качестве "внутрител" или внутриклеточных антител. "Внутритела" могут использоваться для генотерапии.
Последующий аспект предоставляет метод, состоящий во введении нуклеиновой кислоты изобретения в клетку-хозяин. Для введения может применяться любая доступная методика. Для эукариотических клеток подходящие методики могут включать трансфекцию на фосфате кальция, DEAE-декстран, электропробой, трансфекцию через липосомы и трансдукцию с помощью ретровируса или другого вируса, например вакцинии, или для клеток насекомых, бакуловируса. Ввод нуклеиновой кислоты в клетку-хозяин, в частности в эукариотическую клетку, может производиться с помощью вирусной или плазмидной системы. Плазмидная система может поддерживаться эписомально, или может быть инкорпорирована в клетку-хозяин или в искусственную хромосому. Инкорпорация может быть осуществлена с помощью случайной или направленной интеграции одной или более копий в одном или множестве локусов. Для бактериальных клеток подходящие методики могут включать трансформацию хлоридом кальция, электропробой или трансфекцию с помощью бактериофага.
После ввода может следовать запуск экспрессии из нуклеиновой кислоты, например, культивированием клеток-хозяев в условиях для экспрессии гена. Очистка экспрессированного продукта может быть достигнута методами, известными из уровня техники.
Нуклеиновая кислота согласно изобретению может быть интегрирована в геном (например, в хромосому) клетки-хозяина. Интеграция может быть вызвана включением последовательности, вызывающей рекомбинацию генома, в соответствии со стандартными процедурами.
Настоящее изобретение также предоставляет метод, который включает применение вышеописанной конструкции в экспрессионной системе для того, чтобы экспрессировать связывающий член, как указано выше.
Существуют свидетельства участия IL-6 во множестве расстройств, что обсуждается здесь в других местах. Связывающие члены согласно настоящему изобретению могут, таким образом, использоваться в методе диагностики или лечения заболевания, связанного с IL-6. Таким заболеванием может быть, к примеру, воспалительное и/или автоиммунное заболевание, как, например, ревматоидный артрит, остеоартрит, кахексия, хроническое обструктивное заболевание легких, идиопатический ювенильный артрит, астма, системная красная волчанка, воспалительная болезнь кишечника, болезнь Крона или атеросклероз. Связывающие члены согласно настоящему изобретению могут также использоваться для лечения такого заболевания, как опухоль и/или рак.
Связывающие члены согласно настоящему изобретению могут также использоваться в методе диагностики или лечения как минимум одного из связанных с IL-6 заболеваний у пациента, животного, органа, ткани или клетки, включая, но не ограничиваясь:
(дыхательный тракт) обструктивные заболевания дыхательных путей, включая хроническое обструктивное заболевание легких (COPD); астма, например бронхиальная, аллергическая, внутренняя, внешняя и пылевая астма, в частности хроническая или застарелая астма (например, поздняя астма и гиперчувствительность дыхательных путей); бронхит; острый, аллергический, атрофический ринит и хронический ринит, включая творожистый ринит, гипертрофический ринит, гнойный ринит, сухой ринит и медикаментозный ринит; пленочный ринит, включая крупозный, фибринозный и лжепленочный ринит и золотушный ринит; сезонный ринит, включая нервный ринит (сенная лихорадка) и вазомоторный ринит, синусит, идиопатический фиброз легкого (IPF); саркоидоз, "легкое фермера" и родственные заболевания, синдром респираторного истощения взрослых, пневмонит с гиперчувствительностью, фиброидное легкое и идиопатическая промежуточная пневмония;
(кости и суставы) ревматоидный артрит, юношеский хронический артрит, системный перемежающийся юношеский артрит, сероотрицательные спондилоартропатии (включая анкилозирующий спондилит, псориатический артрит и болезнь Райтера), болезнь Бецета, синдром Сиогрена и системный склероз, подагра, остеопороз и остеоартрит;
(кожа) псориаз, атонический дерматит, контактный дерматит и другие экзематозные дерматозы, аллергический контактный дерматит, себорейный дерматит, плоский лишай, склеродермия, пузырчатка, пузырчатый пемфигоид, пузырчатый эпидермолиз, крапивница, ангиодермии, васкулиты, эритемы, кожные эозинофилии, увеит, пятнистая плешивость, аллергический конъюктивит и весенний конъюнктивит;
(желудочно-кишечный тракт) язва желудка, целиакия, проктит, эозинофильный гастроэнтерит, макроцитоз, воспалительная болезнь кишечника, болезнь Крона, язвенный колит, антифосфолипидный синдром, пищевые аллергии с удаленными от кишечника эффектами, например, с мигренью, ринитом и экземой;
(другие ткани и системные заболевания) кахексия, рассеянный склероз, атеросклероз, синдром приобретенного иммунодефицита (AIDS), мезангиальный пролиферирующий гломерулонефрит, нефротический синдром, нефрит, гломерулонефрит, острая почечная недостаточность, гемодиализ, уремия, локализованная или дискоидная красная волчанка, системная красная волчанка, болезнь Каслмана, тироидит Хашимото, тяжелая миастения, диабет I типа, инсулиноустойчивый диабет типа В, серповидноклеточная анемия, иридоциклит/увеит/зрительный неврит, нефритический синдром, эозинофильный фасцит, гипер-IgE синдром, системный васкулит/ гранулематоз Вегенера, орхит/ процедура обратной вазэктомии, лепроматозная проказа, алкогольный гепатит, синдром Сезари и идиопатическая пурпурная тромбоцитопения; послеоперационные спайки, нефроз, синдром системного воспалительного ответа, септический синдром, грамположительный сепсис, грамотрицательный сепсис, культуротрицательный сепсис, грибковый сепсис, нейтропеническая лихорадка, острый панкреатит, уросепсис, болезнь Грейвза, болезнь Рено, опосредованная антителом цитотоксичность, реакции гиперчувствительности III типа, синдром POEMS (синдром полинейропатии, органомегалии, эндокринопатии, моноклональной гаммопатии и кожных изменений), смешанные заболевания соединительной ткани, идиопатическая болезнь Аддисона, сахарный диабет, хронический активный гепатит, первичный желчный цирроз, витилиго, после-MI (кардиотомический) синдром, гиперчувствительность типа IV, гранулемы из-за внутриклеточных паразитов, болезнь Вильсона, гемахроматоз, недостаточная активность альфа-I-антитрипсина, диабетическая ретинопатия, тироидит Хашимото, оценка гипоталамически-питуитарно-надпочечниковой оси, тироидит, энцефаломиелит, хроническое легочное заболевание новорожденных, семейный гематофагоцитный лимфогистиоцитоз, алопеция, радиационная терапия (например, включая, но не ограничиваясь: астения, анемия, кахексия и т.п.), хроническое отравление салицилатами, ночная одышка, ожирение, сердечная недостаточность и менингококцемия;
(отторжение аллотрансплантата) острое или хроническое после, к примеру, трансплантации почки, сердца, печени, легкого, поджелудочной железы, костного мозга, кости, тонкой кишки, кожи, хряща и роговицы; и хроническая реакция "трансплантат против хозяина";
(злокачественные заболевания) лейкемия, острая лимфобластная лейкемия (ALL), острая лейкемия, Т-клеточная, В-клеточная, или FAB ALL, хроническая миелоцитная лейкемия (CML), острая миелоидная лейкемия (AML), хроническая лимфоцитная лейкемия (CLL), волосатоклеточная лейкемия, миелодиспластический синдром (MDS), любая лимфома, болезнь Ходжкина, неходжкиновская лимфома, любая злокачественная лимфома, лимфома Беркитта, множественная миелома, саркома Капоши, карцинома клеток почек, карцинома прямой кишки, карцинома простаты, карцинома поджелудочной железы, карцинома носоглотки, злокачественный гистиоцитоз, паранеопластический синдром/ гиперкальцемия злокачественности, солидные опухоли, аденокарциномы, саркомы, злокачественная меланома, гемангиома, метастатические заболевания, вызванное раком разрушение кости, вызванная раком костная боль; подавление ракового метастаза; облегчение раковой кахексии;
кистозный фиброз, инсульт, реперфузионное поражение сердца, мозга, конечностей и других органов;
ожоги, травмы/ кровотечения, воздействие ионизирующего излучения, хронические язвы кожи;
репродуктивные заболевания (например, расстройства овуляции, менструации и имплантации, преждевременные роды, преэклампсия, эндометриоз);
(инфекции) острые или хронические бактериальные инфекции, острые и хронические паразитарные или инфекционные процессы, включая бактериальные, вирусные и грибковые инфекции, ВИЧ-инфекцию/ВИЧ-нейропатию, менингит, гепатит (А, В или С, или другой подобный гепатит), септический артрит, перитонит, пневмония, эпиглоттит, эшерихиоз, вызванный е.coli 0157:h7, гемолитический уремический синдром/ тромботическая тромбоцитопеническая пурпура, малярия, геморрагическая лихорадка денге, лейшманиоз, проказа, синдром токсического шока, стрептококковый миозит, газовая гангрена, туберкулез, птичий внутриклеточный туберкулез, пневмония, вызванная pneumocystis carinii, воспаление тазовой полости, орхит/ эпидидимит, болезнь легионеров, болезнь Лайма, грипп а, вирус Эпштейна-Барра, витальный гемафагоцитический синдром, вирусный энцефалит/ асептический менингит и т.п.
Соответственно, изобретение предоставляет метод лечения связанного с IL-6 заболевания, заключающийся в назначении пациенту эффективного количества одного или более связывающих членов согласно настоящему изобретению или в комбинированном терапевтическом режиме вместе с другим подходящим лекарством, известным из уровня техники или описанным здесь.
Свидетельства участия IL-6 в некоторых заболеваниях подытожены в данной заявке в другом месте. Кроме того, данные, представленные в данной заявке далее, означают, что связывающие члены согласно изобретению могут применяться для лечения таких заболеваний, включая предупреждающее лечение и снижение тяжести заболеваний. Соответственно, изобретение предоставляет метод лечения или снижения тяжести хотя бы одного симптома любого из расстройств, упомянутых здесь, включая назначение нуждающемуся пациенту эффективного количества одного или более связывающих членов согласно настоящему изобретению, одного или в комбинированном терапевтическом курсе вместе с другим подходящим лекарством, известным из уровня техники или описанным здесь, так чтобы тяжесть хотя бы одного симптома любой из вышеуказанных болезней снизилась.
Таким образом, связывающие члены согласно настоящему изобретению полезны как терапевтические средства для лечения заболеваний или расстройств с участием экспрессии и/или активности IL-6 и/или IL-6Ra, особенно ненормальной экспрессии/активности. Метод лечения может включать назначение эффективного количества связывающего члена изобретения нуждающемуся пациенту, у которого снижается ненормальная экспрессия и/или активность IL-6 и/или IL-6Ra. Метод лечения может включать (i) идентификацию пациента, проявляющего ненормальные уровни или активность IL-6:IL-6Ra, например, с помощью вышеописанных диагностических методов, и (ii) назначение эффективного количества связывающего члена изобретения нуждающемуся пациенту, у которого снижается ненормальная экспрессия и/или активность IL-6 и/или IL-6Ra. Эффективное количество согласно изобретению представляет собой такое количество, которое снижает ненормальную экспрессию и/или активность IL-6 и/или IL-6Ra так, чтобы снизить или уменьшить тяжесть хотя бы одного симптома отдельного заболевания или расстройства, но не обязательно излечивает заболевание или расстройство.
Изобретение также предоставляет метод уменьшения хотя бы одного эффекта IL-6, включающий контакт или введение эффективного количества одного или более связывающих членов согласно настоящему изобретению, таких, что хотя бы один из эффектов IL-6 уменьшился. Эффекты IL-6, которые могут быть уменьшены методами изобретения, включают связывание IL-6 с gp130, и последующие эффекты, возникающие вследствие этого связывания.
Соответственно, дальнейшие аспекты изобретения предоставляют методы лечения, включающие введение связывающего элемента как предусмотрено, фармацевтические композиции, включающие такой связывающий член, и использование такого связывающего члена в производстве лекарства для применения, например, в методе приготовления лекарства или фармацевтической композиции, включающей сочетание связывающего члена и фармацевтически приемлемого наполнителя. Фармацевтически приемлемый наполнитель может быть веществом или смесью веществ, которые не вызывают вторичных реакций и позволяют, к примеру, облегчить введение активных веществ, увеличение времени жизни и/или эффективности внутри тела, повышение растворимости в растворе или улучшение сохранности. Эти фармацевтически приемлемые носители хорошо известны из уровня техники и будут адаптированы квалифицированным персоналом в соответствии с природой и способом введения выбранных активных веществ.
Связывающие члены согласно настоящему изобретению будут обычно применяться в виде фармацевтической композиции, которая может содержать как минимум один компонент в придачу к связывающему члену. Такие фармацевтические композиции согласно настоящему изобретению и для использования в соответствии с настоящим изобретением, могут содержать, вместе с активным ингредиентом, фармацевтически приемлемый эксципиент, носитель, буфер, стабилизатор или другой материал, известный из уровня техники. Такие материалы должны быть нетоксичны и не должны влиять на эффективность активного ингредиента. Конкретная природа носителя или другого материала будет зависеть от способа ввода, который может быть оральным, ингаляционным, внутритрахейным, местным, внутриполостным или инъекционным как обсуждается ниже.
Фармацевтические композиции для перорального применения, как, например, однодоменные молекулы антител (например, "нанотела™") и др. также предусмотрены в настоящем изобретении. Такие пероральные рецептуры могут быть в виде таблетки, капсулы, порошка, жидкости или мази. Таблетка может содержать твердый носитель, например желатин, или адъювант. Жидкие фармацевтические композиции обычно содержат носитель, такой как вода, нефть, животные или растительные масла, минеральное масло или синтетическое масло. Могут быть включены физиологический солевой раствор, раствор глюкозы или другого сахара или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной инъекции или инъекции в месте поражения активный ингредиент будет в виде парентерально приемлемого водного раствора, свободного от пирогенов и имеющего подходящий рН, изотоничность и стабильность. Квалифицированный персонал вполне способен приготовить необходимые растворы с помощью, например, изотонических носителей, таких как раствор хлорида натрия, раствор Рингера, раствор Рингера с лактатом. Консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки могут применяться при необходимости, включая такие буферы как фосфат, цитрат и другие органические кислоты; такие антиоксиданты как аскорбиновая кислота и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетония хлорид; фенол, бутиловый или бензиловый спирт; алкил парабензоаты, такие как метил- и пропилпарбен; катехин; резорцин; циклогексанол; 3'-пентанол; и m-крезол); низкомолекулярные полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагины, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные сурфактанты, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
Связывающие члены согласно настоящему изобретению могут быть формулированы в жидкой, мягкой или твердой форме в зависимости от физико-химических свойств молекулы и пути доставки.
Рецептуры могут включать эксципиенты, или комбинации эксципиентов, например сахара, аминокислоты и сурфактанты. Жидкие рецептуры могут включать широкий диапазон концентраций антитела и рН. Твердые формы могут производиться с помощью лиофилизации, распылительной сушки или сушки по сверхкритической технологии, например. Формы связывающих членов будут зависеть от намеченого способа доставки: например, формы для доставки в легкие могут состоять из частиц с физическими свойствами, обеспечивающими проникновение глубоко в легкие при ингаляции; местные рецептуры (например, для лечения рубцов, например, рубцов кожи) могут включать средства, модифицирующие вязкость, которые удлиняют время пребывания лекарства на месте действия. Связывающий член может быть приготовлен с носителем, который будет предохранять связывающий член от быстрого высвобождения, таким как форма с контролируемым высвобождением, включая имплантанты, трансдермальные пластыри и микрокапсулированные системы доставки. Могут использоваться биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие методы приготовления таких форм известны из уровня техники [109].
Лечение можно проводить перорально (как, к примеру, однодоменные молекулы антител (например, "нанотела™")), инъекционно (например, подкожно, внутрисуставно, внутривенно, внутрибрюшинно, внутриартериально или внутримышечно), ингаляционно, интратрахеально, внутрь мочевого пузыря, или местно (например, интраокулярно, интраназально, ректально, в раны, на кожу). Лечение может проводиться с помощью импульсной инфузии, особенно с уменьшающейся дозой связывающего члена. Путь введения может быть определен из физико-химических характеристик терапии, особых условий для заболевания или из необходимости оптимизировать эффективность или минимизировать побочные эффекты. Одним из отдельных путей введения является внутривенный. Еще одним путем введения фармацевтической композиции настоящего изобретения является подкожный. Подразумевается, что лечение не будет ограничиваться использованием в клинике. Таким образом, подкожные инъекции с помощью безыгольного прибора также полезны.
Композиция может применяться одна или вместе с другим лечением, одновременно или последовательно в зависимости от условий лечения.
Связывающий член согласно изобретению может использоваться как часть комбинированной терапии в сочетании с дополнительным лекарственным компонентом. Комбинированное лечение может применяться для обеспечения значительных синергических эффектов, особенно сочетание связывающего члена изобретения с одним или более других лекарств. Связывающий член согласно настоящему изобретению может применяться параллельно или последовательно, или в сочетании с другим терапевтическим средством или средствами, для лечения состояний, перечисленных здесь.
Связывающий член согласно изобретению может использоваться как хемосенсибилизатор и тем самым повышать терапевтическую эффективность цитотоксических средств, и таким образом может предоставляться для введения в сочетании с одним или более цитотоксических средств, одновременно или последовательно. Связывающий член также может использоваться как радиосенсибилизатор и тем самым повышать эффективность облучения, и таким образом может предоставляться для введения в сочетании с облучением, одновременно или последовательно.
Связывающий член согласно настоящему изобретению может предоставляться в сочетании или смеси с одним или более следующих средств:
- цитокин, или агонист либо антагонист цитокиновой функции (например, средство, действующее на цитокиновые сигнальные пути, типа модулятора SOCS-системы), такие как альфа-, бета- и/или гамма-интерферон; инсулиноподобный фактор роста I типа (IGF-1), его рецепторы и сопровождающие его связывающиеся белки; интерлейкины (IL), например один или более IL- от 1 до 33, и/или антагонист или ингибитор интерлейкина, такие как анакинра; ингибиторы рецепторов семейства интерлейкинов или ингибиторы специфических субъединиц таких рецепторов, ингибитор фактора некроза опухоли альфа (TNF-α), такой как моноклональные антитела против TNF (например, инфликсимаб, адалимумаб и/или CDP-870) и/или антагонист рецептора TNF, например молекула иммуноглобулина (например, этанерцепт) и/или низкомолекулярное вещество, такое как пентоксифиллин;
- модулятор В-клеток, например моноклональное антитело, нацеленное на В-лимфоциты (например, CD20 (ритуксимаб) или MRA-aILl6R) или на Т-лимфоциты (например, CTLA4-Ig, HuMax Il-15 или Абатацепт);
- модулятор, ингибирующий активность остеокластов, например антитело к RANKL;
- модулятор хемокина или функции хемокинового рецептора, такой как антагонист CCR1, CCR2, CCR2A, CCR2B, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10 или CCR11 (для семейства С-С); CXCR1, CXCR2, CXCR3, CXCR4, CXCR5 или CXCR6 (для семейства С-Х-С) и CX3CR1 для семейства С-Х3-С;
- ингибитор матричных металлопротеаз (MMPs), то есть один или более из стромелизинов, коллагеназ и желатиназ, а также аггреканаза, в особенности коллагеназа-1 (ММР-1), коллагеназа-2 (ММР-8), коллагеназа-3 (ММР-13), стромелизин-1 (ММР-3), стромелизин-2 (ММР-10) и/или стромелизин-3 (ММР-11) и/или ММР-9 и/или ММР-12, например средство типа доксициклина;
- ингибитор биосинтеза лейкотриенов, ингибитор 5-липоксигеназы (5-LO) или антагонист 5-липоксигеназактивирующего белка (FLAP), такой как зилеутон; АВТ-761;
фенлеутон; тепоксалин; Abbott-79175; Abbott-85761; N-(5-замещенные)-тиофен-2-алкилсульфонамиды; 2,6-ди-трет-бутилфенилгидразоны; метокситетрагидропираны, такие как Zeneca ZD-2138; вещество SB-210661; пиридилзамещенный 2-цианонафталин, такой как L-739,010; 2-цианохинолин, такой как L-746,530; индол и/или хинолин, такой как MK-591, MK-886 и/или BAY × 1005;
- антагонист рецептора для лейкотриенов (LT) B4, LTC4, LTD4 и LTE4, избранный из группы, состоящей из фенотиазинов-3-1, таких как L-651,392; амидинов, таких как CGS-25019с; бензоксаламинов, таких как ontazolast; бензолкарбоксимидоаминов, таких как BIIL 284/260; и вещества, такие как зафирлукаст, аблукаст, монтелукаст, пранлукаст, верлукаст (МК-679), RG-12525, Ro-245913, иралукаст (CGP 45715А) и BAY × 7195;
- ингибитор фосфодиэстеразы (PDE), такой как метилксантанин, например теофиллин и/или аминофиллин; и/или избирательный ингибитор изофермента PDE, например ингибитор PDE4 и/или ингибитор изоформы PDE4D и/или ингибитор PDE5;
- антагонист гистаминового рецептора типа 1, такой как цетиризин, лоратадин, дезлоратадин, фексофенадин, акривастин, терфенадин, астемизол, азеластин, левокабастин, хлорфенирамин, прометазин, циклизин и/или мизоластин (в основном применяемый перорально, местно или парэнтерально);
- ингибитор протонного насоса (такой как омепразол) или гастропротекторный антагонист гистаминового рецептора типа 2;
- антагонист гистаминового рецептора типа 4;
- агонист адренорецептора альфа-1/альфа-2 сосудосужающий симпатомиметик, такой как пропилгекседрин, фенилэфрин, фенилпропаноламин, эфедрин, псевдоэфедрин, нафазолина гидрохлорид, оксиметазолина гидрохлорид, тетрагидрозолина гидрохлорид, ксилометазолина гидрохлорид, трамазолина гидрохлорид и этилнорадреналина гидрохлорид;
- антихолинэргическое средство, например антагонист мускаринового рецептора (M1, M2 и М3), такой как атропин, гиосцин, гликопирролат, ипратропия бромид, тиотропия бромид, окситропия бромид, пирензепин и телензепин;
- агонист бета-адренорецептора (включая бета-рецептор подтипов 1-4), такой как изопреналин, сальбутамол, формотерол, сальметерол, тербуталин, орципреналин, битолтерол-мезилат и/или пирбутерол, например его хиральный энантиомер;
- хромон, например натрия хромогликат и/или недохромил-натрий;
- глюкокортикоид, такой как флунизолид, триамцинолона ацетонид, беклометазона дипропионат, будезонид, флутиказона пропионат, циклезонид и/или мометазона фуроат;
- средство, модулирующее рецепторы гормонов в ядре, такое как PPAR;
- иммуноглобулин (Ig), или препарат Ig, или антагонист либо антитело, модулирующее функцию Ig, такое как анти-IgE (например, омализумаб);
- другое системное или местно применяемое противовоспалительное средство, например талидомид или его производное, ретиноид, дитранол или кальципотриол;
- сочетание аминосалицилатов и сульфапиридинов, таких как сульфасалазин, месалазин, бальсалазид и ольсалазин; и иммуномодулирующих средств, таких как тиопурины; и кортикостероидов, таких как будезонид;
- противобактериальное средство, например производное пенициллина, тетрациклин, макролид, бета-лактам, фторхинолон, метронидазол и/или ингаляционный аминогликозид; и/или противовирусное средство, например ацикловир, фамцикловир, валацикловир, ганцикловир, цидофовир; амантадин, ремантадин; рибавирин; занамавир и/или оселтамавир; ингибитор протеазы, такой как индинавир, нелфинавир, ритонавир и/или саквинавир; нуклеозидный ингибитор обратной транскриптазы, такой как диданозин, ламивудин, ставудин, зальцитабин, зидовудин; ненуклеозидный ингибитор обратной транскриптазы, такой как невирапин, эфавиренз;
- сердечно-сосудистое средство, такое как блокатор кальциевых каналов, бета-адреноблокатор, ингибитор ангиотензин-превращающего фермента (АСЕ), антагонист ангиотензинового рецептора-2; липидопонижающеее средство, такое как статин и/или фибрат; модулятор морфологии клеток крови, такой как пентоксифиллин; тромболитик и/или антикоагулянт, например ингибитор аггрегации тромбоцитов;
- средство, воздействующее на ЦНС, такое как антидепрессант (такой как сертралин), антипаркинсоническое средство (такое как депренил, L-допа, ропинирол, прамипексол; ингибитор МАОВ, такой как селегилин и расагилин; ингибитор comP, такой как тасмар; ингибитор А-2, ингибитор обратного захвата дофамина, антагонист NMDA, агонист никотина, агонист дофамина и/или ингибитор нейронной NO-синтазы) и средство против болезни Альцгеймера, такое как донепезил, ривастигмин, такрин, ингибитор СОХ-2, пропентофиллин или метрифонат;
- средство для лечения острой и хронической боли, например центрально- или периферийно-действующий анальгетик, такой как аналог или производное опиоида, карбамазепин, фенитоин, натрия вальпроат, амитриптилин или другой антидепрессант, парацетамол, или нестероидное противовоспалительное средство;
- парентерально или местно применяемый (в том числе ингаляционный) местный анестетик, такой как лигнокаин или его аналог;
- средство против остеопороза, например гормональное средство, такое как ралоксифен, или бифосфонат, такой как алендронат;
- (i) ингибитор триптазы; (ii) антагонист тромбоцит-активирующего фактора (PAF); (iii) ингибитор интерлейкин-превращающего фермента (ICE); (iv) ингибитор IMPDH; (v) ингибиторы адгезии молекул, включая антагонист VLA-4; (vi) катепсин; (vii) ингибитор киназы, например ингибитор тирозинкиназ (таких как Btk, Itk, Jak3 MAP, примеры ингибиторов могут включать Гефитиниб, Иматиниб мезилат), серин/треонинкиназы (например, ингибитор МАР-киназы, такой как р38, JNK, протеинкиназы А, В и С и IKK), или киназы, участвующей в регуляции клеточного цикла (например, цилин-зависимая киназа); (viii) ингибитор глюкозо-6-фосфатдегидрогеназы; (ix) антагонист рецептора B.subl. - и/или В.sub2.-кинина; (х) антиподагрическое средство, например колхицин; (xi) ингибитор ксантиноксидазы, например аллопуринол; (xii) урикозурик, например пробенецид, сульфинпиразон и/или бензбромарон; (xiii) стимулятор секреции гормона роста; (xiv) трансформирующий фактор роста (TGFβ); (xv) фактор роста из тромбоцитов (PDGF); (xvi) фактор роста фибробластов, например основной фактор роста фибробластов (bFGF); (xvii) колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF); (xviii) капсаициновая мазь; (xix) антагонист рецептора тахикинина NK.sub1. и/или NK.sub3., такой как NKP-608C, SB-233412 (талнетант) и/или D-4418; (хх) ингибитор эластазы, например UT-77 и/или ZD-0892; (xxi) ингибитор TNF-альфа превращающего фермента (ТАСЕ); (xxii) ингибитор индуцированной NO-синтазы (iNOS) или (xxiii) хемоаттрактант - рецептор-гомологичная молекула, экспрессируемая клетками ТН2 (такой как антагонист CRTH2); (xxiv) ингибитор Р38; (xxv) средство, модулирующее функцию Toll-подобных рецепторов (TLR), и (xxvi) средство, модулирующее активность пуринэргических рецепторов, такое как Р2Х7; (xxvii) ингибитор активации фактора транскрипции, такой как NFkB, API и/или STATS.
Ингибитор может быть избирательным или смешанным ингибитором, например ингибитором, нацеленным на более чем одну молекулу (напр., рецепторы) или на вышеупомянутые классы молекул.
Связывающий член также может использоваться в сочетании с химиотерапевтическим средством или другим ингибитором тирозинкиназы в совместном введении или в виде иммуноконъюгата. Фрагменты указанного антитела также могут использоваться в биспецифичных антителах, полученных с помощью рекомбинантных механизмов или биохимического сочетания, с последующей ассоциацией специфичности вышеописанного антитела со специфичностью других антител, способных распознавать другие молекулы, участвующие в процессах, связанных с IL-6.
Для лечения воспалительных заболеваний связывающий член согласно изобретению может сочетаться с одним или более средств, таких как нестероидные противовоспалительные средства (далее везде NSAID), включая неселективные ингибиторы циклооксигеназы (СОХ)-1 / СОХ-2, применяемые местно или системно, такие как пироксикам, диклофенак, пропионовые кислоты, такие как напроксен, флурбипрофен, фенопрофен, кетопрофен и ибупрофен, фенаматы, такие как мефенамовая кислота, индометацин, сулиндак, азапропазон, пиразолоны, такие как фенилбутазон, салицилаты, такие как аспирин; селективные ингибиторы СОХ-2 (такие как мелоксикам, целекоксиб, рофекоксиб, вальдекоксиб, лумарококсиб, парекоксиб и эторикоксиб); ингибирующие циклооксигеназу доноры оксида азота (CINOD); глюкокортикостероиды (применяемые местно, орально, внутримышечно, внутривенно или внутрисуставно); метотрексат, лефлуномид; оксихлорохин, d-пеницилламин, ауранофин или другие парентеральные или оральные препараты золота; анальгетики; диацереин; внутрисуставные средства, такие как производные гиалуроновой кислоты; и пищевые добавки, такие как глюкозамин.
Связывающий член согласно изобретению может также использоваться в сочетании с существующим терапевтическим средством для лечения рака. Подходящие средства для комбинированного применения включают:
(i) антипролиферативные/ антинеопластические средства и их сочетания, используемые в медицинской онкологии, такие как Gleevec (иматиниб мезилат), алкилирующие средства (например, цисплатин, карбоплатин, циклофосфамид, азотистый иприт, мефалан, хлорамбуцил, бусульфан и нитрозомочевины); антиметаболиты (например, антифолаты, такие фторпиримидины как 5-фторурацил и тегафур, ралтитрексед, метотрексат, цитозинарабинозид, гидроксимочевина, гемцитабин и паклитаксел); противоопухолевые антибиотики (например, антрациклины, такие как адриамицин, блеомицин, доксорубицин, дауномицин, эпирубицин, идарубицин, митомицин-С, дактиномицин и митрамицин); антимитотические средства (например, алкалоиды барвинка, такие как винкристин, винбластин, виндезин и винорелбин, и таксоиды, такие как таксол и таксотер); и ингибиторы топоизомеразы (например, эпиподофиллотоксины, такие как этопозид, тенипозид, амсакрин, топотекан и камптотецины);
(ii) цитостатические средства, такие как антиэстрогены (например, тамоксифену торемифен, ралоксифен, дролоксифен и йодоксифен), отрицательные регуляторы эстрогенных рецепторов (например, фульвестрант), антиандрогены (например, бикалутамид, флутамид, нилутамид и ципротерона ацетат), антагонисты LHRH или агонисты LHRH (например, госерелин, лейпрорелин и бусерелин), прогестогены (например, мегестрола ацетат), ингибиторы ароматазы (например, анастрозол, летрозол, воразол и эксеместан) и ингибиторы 5α-редуктазы, такие как финастерид;
(iii) средства, ингибирующие инвазию раковых клеток (например, ингибиторы металлопротеазы, такие как маримастат, и ингибиторы функции рецептора активатора плазминогена урокиназы);
(iv) ингибиторы функции фактора роста, например ингибиторы, включающие антитела к фактору роста, антитела к рецептору фактора роста (например, анти-erbb2 антитело трастузумаб и анти-erbb1 антитело цетуксимаб [С225]), ингибиторы фарнезилтрансферазы, ингибиторы тирозинкиназы и ингибиторы серин/треонинкиназы, например ингибиторы семейства эпидермальных факторов роста (например, ингибиторы тирозинкиназ семейства EGFR, такие как N-(3-хлор-4-фторфенил)-7-метокси-6-(3-морфолинопропокси)хиназолин-4-амин (гефитиниб, AZD1839), N-(3-этинилфенил)-6,7-бис(2-метоксиэтокси)хиназолин-4-амин (эрлотиниб, OSI-774) и 6-акриламидо-N-(3-хлор-4-фторфенил)-7-(3-морфолинопропокси)хиназолин-4-амин (CI 1033)), например ингибиторы семейства тромбоцитных факторов роста и, например, ингибиторы семейства факторов роста гепатоцитов;
(v) антиангиогенные средства, такие как ингибиторы эффектов фактора роста сосудистого эндотелия (например, антитело против фактора роста клеток сосудистого эндотелия бевацизумаб, соединения, такие как описанные в патентах WO 97/22596, WO 97/30035, WO 97/32856 и WO 98/13354, каждый из которых полностью включен сюда) и вещества, действующие по другим механизмам (например, линомид, ингибиторы функции интегрина ανβ3 и ангиостатин);
(vi) средства, повреждающие сосуды, такие как комбретастатин А4 и соединения, описанные в патентах WO 99/02166, WO 00/40529, WO 00/41669, WO 01/92224, WO 02/04434 и WO 02/08213 (каждый из которых полностью включен сюда);
(vii) антисмысловые средства, например, направленные на вышеперечисленные мишени, такие как ISIS 2503, антисмысл анти-rаs;
(viii) генотерапевтические подходы, включая, например, методы замены неправильных генов, таких как неправильный р53 или неправильный BRCA1, или BRCA2, GDEPT (генонаправленное ферментное пролекарство), такие как подходы с использованием цитозиндезаминазы, тимидинкиназы или бактериального фермента нитроредуктазы, и способы повышения толерантности пациента к химиотерапии и облучению, такие как генотерапия мультилекарственной резистентности; и
(ix) иммунотерапевтические подходы, включая, к примеру, способы ex vivo и in vivo для повышения иммуногенности опухолевых клеток пациента, такие как трансфекция цитокинами, такими как интерлейкин 2, интерлейкин 4 или колониестимулирующий фактор гранулоцитов-макрофагов, способы снижении анергии Т-клеток, подходы с использованием трансфицированных иммунных клеток, таких как трансфицированные цитокином древовидные клетки, и подходы с использованием антиидиотипических антител.
Связывающий член согласно изобретению и один или более дополнительных лекарственных компонентов могут использоваться для приготовления лекарственного средства. Лекарство может быть для отдельного или комбинированного применения у пациента и соответственно может содержать связывающий член и дополнительный компонент в виде комбинированного препарата или как отдельные препараты. Отдельные препараты могут использоваться для облегчения отдельного, последовательного или одновременного введения и позволяет вводить компоненты разными путями, например орально и парентерально.
В соответствии с настоящим изобретением, предоставляемые композиции могут вводиться млекопитающим. Обычно назначается "терапевтически эффективное количество", что достаточно для принесения пользы пациенту. Эта польза может быть как минимум облегчением ходя бы одного симптома. Действительное вводимое количество, скорость и продолжительность введения будут зависеть от природы и тяжести заболевания, в особенности при лечении млекопитающих, от клинического состояния пациента, причины заболевания, места доставки композиции типа связывающего члена, метода ввода, расписания ввода и других факторов, известных практикующим врачам. Назначение лечения, например выбор дозировки и т.п., находится в пределах ответственности главных практикующих и других врачей и может зависеть от тяжести симптомов и/или прогрессированием заболевания. Подходящие дозы антитела хорошо известны из уровня техники [110, 111]. Могут использоваться специфические дозировки, указанные здесь или в Physician's Desk Reference (2003) в качестве подходящих для типа применяемого лекарства. Терапевтически эффективное количество или подходящая доза связывающего члена изобретения может быть определена сравнением его активности in vitro и активности in vivo на животной модели. Методы экстраполяции эффективных дозировок у мышей и других подопытных животных на людей известны. Точная доза будет зависеть от ряда факторов, включая применение антитела для диагностики, профилактики или лечения, размеры и расположение места для лечения, конкретная природа антитела (напр., целое антитело, фрагмент или двойное тело), и природы обнаружимой метки или другой молекулы, присоединенной к антителу. Типичная доза антитела будет в пределах от 100 мкг до 1 г для системного применения, и от 1 мкг до 1 мг для местного применения. Вначале может применяться более высокая доза, а потом одна или больше более низких доз. Типично, антитело будет целым антителом, напр., изотип IgG1. Это доза для разового лечения взрослого пациента, которая может быть установлена пропорционально для детей и подростков, а также пересчитана на другие форматы антител пропорционально молекулярному весу. Лечение может повторяться каждый день, два раза в неделю, еженедельно или ежемесячно, на усмотрение врача. Лечение может проводиться каждые две-четыре недели в случае подкожного введения, и каждые четыре-восемь недель в случае внутривенного введения. Лечение может быть периодическим, а период между введением около двух недель и более, например около трех недель и более, около четырех недель и более, или около раза в месяц. Лечение может проводиться перед и/или после операции, и/или может вводиться или применяться непосредственно на анатомическом месте хирургического вмешательства.
Связывающиеся с IL-6 элементы согласно изобретению могут предоставить преимущества в дозировке и требований к введению сравнительно с антителами к sIL-6Ra. Как здесь отмечается в другом месте, циркулирующие уровни IL-6 значительно ниже, чем циркулирующие уровни sIL-6Ra при заболевании. Соответственно, использование связывающегося с IL-6 элемента, в отличие от связывающего члена анти-IL-6R, имеет значительные преимущества в том, что количество лекарства, производимого на каждую дозу для пациентов, может быть меньше. Также если доза лекарства анти-IL6 ниже, могут быть значительные преимущества в том, что низкая доза облегчает как подкожные, так и внутривенные инъекции. Из уровня техники хорошо известно, что подкожная дозировка может быть ограничена количеством связывающего члена, например молекулы антитела, необходимого на одну дозу. Это потому, что подкожная инъекция ограничена объемом, который может быть введен в одном месте кожи. Обычно применяются подкожные инъекции объемом 1.2 мл или меньше. Так как может быть все труднее формулировать связывающий член для подкожной инъекции при концентрациях, больших чем 50 мг/мл, дозы выше 100 мг таким способом обычно требуют множественных инъекций и больше дискомфорта для пациента.
Для низкодозового лекарства анти-IL-6 может также потребоваться более низкая "загрузочная" доза антитела для ингибирования всего системного IL-6 сравнительно с системным sIL-6Ra, так как тот присутствует в более высоких концентрациях.
Дальнейшая выгода может быть связана с нацеленностью на IL-6, а не на рецептор IL-6, представляя дополнительные преимущества связывающих членов изобретения по сравнению со связывающими членами для IL-6Ra.
К примеру, есть литературные данные, показывающие, что циркулирующие уровни IL-6 значительно ниже, чем циркулирующие уровни sIL-6Ra при заболевании [112, 113]. Так как уровни SIL-6R существенно выше, чем уровни IL-6, больше связывающего члена анти-sIL-6R потребуется для нейтрализации sIL-6Ra по сравнению со связывающим членом анти-IL-6, необходимым для нейтрализации IL-6. Следовательно, будет нужна более низкая доза связывающего члена антилиганд по сравнению с использованием связывающего члена антирецептор.
Нацеливание на лиганд IL-6, а не на рецептор IL-6 может снизить уровни IL-6 при болезни, но все-таки позволяет уровням IL-6 повышаться при инфекции, при которой IL-6 регулируется в сторону повышения как часть иммунного ответа.
Kawano и др. [4] показали, что IL-6 является мощным фактором роста, а свежевыделенные из пациента клетки миеломы продуцируют IL-6 и экспрессируют его рецепторы. Более того, антитело анти-IL-6 ингибирует рост клеток миеломы in vitro. Это прямо свидетельствует об участии аутокринной петли в онкогенезе миелом человека. Вслед за этим исследованием Van Zaanen и др. [5] продемонстрировали, что производство IL-6 у пациентов с множественной миеломой снижается при лечении антителом антилиганд-IL-6.
Ряд дальнейших исследований показывает, что IL-6 участвует в автокринной петле обратной связи в других типах клеток, напр. в гладкомышечных клетках (SMC) [114], клетках астроглиомы U373-MG [115], адипоцитах 3Т3 [116], нейронах [117], клетках эндотелия [118] и в клетках саркомы Капоши [119]. Ингибирование IL-6 с помощью связывающего члена анти-IL6 при заболевании может таким образом вести к уменьшению базисного производства IL-6 при болезни.
Далее, связывающие члены анти-IL-6 связывают IL-6 в системном круговороте, в отличие от элементов, связывающихся с рецептором IL-6, которым необходимо проникнуть внутрь ткани, чтобы занять рецептор на поверхности клеток, вовлеченных в патологический процесс заболевания, подлежащего лечению.
Связывающиеся с IL-6 элементы могут установить равновесие с IL-6 в системном круговороте, обладая способностью вызывать градиенты поперек барьеров, напр., синовиальной мембраны, которая имеет сетевой эффект удаления активного IL-6 из соединения и образования неактивного комплекса со связывающим членом. Вследствие этого связывающийся с IL-6 элемент может действовать быстрее, а режим дозировки может быть другим и потенциально более легким для оптимизации сравнительно со связывающимся с IL-6R элементом.
Сигнал IL-6 опосредуется связыванием IL-6 с IL-6R и связыванием этого комплекса с gp130. Если связывание IL-6 и IL-6Ra имеет наномолярную аффинность (около 5 нМ), а связывание комплекса IL6:IL6R с gp130 имеет пикомолярную аффинность, связывающий член, нацеленный на IL-6, сталкивается с меньшим количеством конкуренции за связывание с IL-6 и таким образом может подавлять большую часть сигналов IL-6. Хотя это применимо также для связывающего члена, нацеленного на растворимый IL-6Ra и предотвращающего образование комплекса IL-6:IL-6Ra, если IL-6Ra связан с мембраной, то из-за стерических затруднений может быть более тяжело для анти-IL-6Ra связывать и ингибировать IL-6Ra, представленный на мембране.
Примеры
1. Пример 1. Выделение лидера
1.1 Селекция
Для селекции использовались выставочные библиотеки простого человеческого одноцеочечного Fv (scFv) фага, клонированные в фагмидном векторе, основанном на волокнистом фаге М13 [120, 121]). Специфические анти-IL-6 scFv антитела были выделены из фаговых выставочных библиотек с помощью ряда селекционных циклов на рекомбинантном человеческом IL-6 по существу как ранее описано Vaughan и др. [120] и Hawkins и др. [122]. Вкратце, для биоселекции, человеческий IL-6 в PBS (PBS-буфер Дульбекко, рН7.4) адсорбировался на ячейки микротитровальной пластинки в течение ночи при 4°C. Ячейки были промыты PBS, затем блокированы 1 ч с помощью PBS-Marvel (3% мас./об.). Очищенный фаг в PBS-Marvel (3% мас./об.) добавили в ячейки и оставили для связывания покрытого антигена на 1 ч. Несвязанный фаг был удален серией промывочных циклов с помощью PBS-Tween (0,1% об./об.) и PBS. Связанные фаговые частицы были смыты, инфицированы в бактерии и сохранены для следующего круга селекции [120].
1.2 Ингибирование связывания IL-6 с рецептором IL-6 неочищенным scFv
Репрезентативное количество индивидуальных клонов из второго круга селекции были выращены на 96-ячеистых пластинках. ScFvs экспрессировали в бактериальном периплазме и просеивали по ингибирующей активности методом количественного определения связывания IL-6/ человеческий рецептор IL-6 в HTRF® (гомогенная флуоресценция с разрешением по времени, CIS Bio international). В этом анализе образцы конкурировали за связывание с человеческим IL-6, меченым криптатом (R&D Systems), и с биотинилированным IL-6R (Peprotech). Стандартное антитело анти-IL-6 mAb (Biosource AHC0562) было включено во все определения активности в качестве положительного контроля. Подробный метод количественного определения предоставлен в разделе "Материалы и методы".
2.3 Переформатирование scFv к IgG1
Клоны были преобразованы из scFv в формат IgG с помощью субклонирования VH и VL доменов в векторах, экспрессирующих тяжелую и легкую цепи целого антитела соответственно. VH домен был клонирован в векторе (pEU15.1), содержащем постоянные домены тяжелой цепи человека и регуляторные элементы для экспрессии целой тяжелой цепи IgG в клетках млекопитающих. Аналогично, VL домен был клонирован в векторе pEU3.4 для экспрессии легкой каппа-цепи, или в pEU4.4 для экспрессии постоянных доменов легкой лямбда-цепи человека с регуляторными элементами для экспрессии целой легкой цепи IgG в клетках млекопитающих. Векторы для экспрессии тяжелых цепей и легких цепей были впервые описаны в [123]. Были разработаны векторы Cambridge Antibody Technology просто посредством введения элемента OriP. Для получения IgGs векторы, экспрессирующие тяжелую и легкую цепи IgG, были трансфицированы в клетки млекопитающих EBNA-HEK293. IgGs экспрессировались и секретировались в среду. Перед очисткой продукт был собран и профильтрован. IgG был очищен с помощью хроматографии на белке А. Супернатанты культур загружаются в колонку соответствующего размера из Ceramic Protein A (BioSepra) и промываются 50 ммоль трис-HCl рН 8,0, 250 ммоль NaCl. Связанный IgG был смыт с колонки с помощью 0,1 М цитрата натрия (рН 3,0) и нейтрализован прибавлением трис-HCl (рН 9,0). Смытый материал был перенесен в буфер PBS с помощью колонок Nap10 (Amersham, #17-0854-02), и концентрация IgG была определена спектрофотометрически с использованием коэффициента экстинкции, основанного на аминокислотной последовательности IgG [124]. Очищенные IgG анализировался на аггрегацию и деградацию с помощью SEC-HPLC и SDS-PAGE.
1.4 Ингибирование связывания IL-6 с рецептором IL-6 очищенным scFv и IgG
Была секвенирована ДНК тех ScFv, которые проявили значительный ингибирующий эффект на взаимодействие IL-6:IL-6R в виде неочищенных периплазмических экстрактов [120, 125]. Единичные scFvs были снова экспрессированы в бактериях и очищены аффинной хроматографией (как описано в Bannister и др. [126]. Очищенные образцы IgG этих клонов также были приготовлены как описано в разделе 1.3. Активности этих образцов были определены конкуренцией серии разбавлений очищенного препарата с биотинилированным SIL-6R за связывание с маркированным HIS FLAG человеческим IL-6 (происходящим из собственной Е.coli).
Результаты для клона CAN022D10 как scFv, так и IgG, содержащего тяжелую цепь человека и постоянный домен легкой каппа-цепи, приводятся в таблице 1. Подробные методики предоставлены в разделе "Материалы и методы".
1.5 Ингибирование индуцированной IL-6 пролиферации клеток TF-1 очищенным scFv и IgG
Нейтрализующая активность очищенных препаратов scFv против биоактивности IL-6 человека и макаки была оценена с помощью анализа пролиферации клеток TF-1. TF-1 представляет собой линию премиелоидных клеток человека, полученных от пациента с эритролейкемией [134]. Клеточная линия TF-1 зависима от фактора для выживания и пролиферации. Было показано, что клетки TF-1 отвечают на IL-6 человека и макаки (собственный, происходящий из E.coli); клетки содержались в среде, содержащей человеческий GM-CSF (4 нг/мл, R&D Systems). Ингибирование IL-6-зависимой пролиферации определялось посредством измерения уменьшения включения тритированного тимидина в новосинтезированную ДНК делящихся клеток. Подробное описание методики предоставлено в разделе "Материалы и методы".
Очищенные scFv препараты CAN022D10 были способны ингибировать IL-6-индуцированную пролиферацию клеток TF-1 при максимальной испытанной концентрации, хотя полное ингибирование не наблюдалось. Поэтому было невозможно вычислить точные данные по активности IC50 из полученных результатов. При испытании очищенных IgG, IC50 для CAN022D10 была рассчитана равной 93 нмоль.
1.6 Селективность и перекрестновидовая активность антител в тестах эпитопной конкуренции DELFIA®
Перекрестновидовая активность и селективность антител к членам семейства IL-6 определялась с помощью анализа эпитопной конкуренции DELFIA®, с помощью измерения ингибирования биотинилированного HIS FLAG IL-6 (собственный, из E.coli), связывающего каждое иммобилизованное антитело анти-IL-6.
Титры очищенного фактора ингибирования лейкемии (LIF) (Chemicon), цилиарного нейротрофного фактора (CNTF), IL-11 и онкостатина М (все R & D Systems) были проверены в каждом анализе для установления активности для каждого структурно родственного белка, как измерено в величинах IC50 в количественном определении.
Титры IL-6 видов, включая макак (собственный, из E.Coli), человека HIS FLAG IL-6 (собственный, из HEK-EBNA), крысы и мышиный IL-6 (оба R & D Systems) были тестированы в каждом анализе для установления перекрестновидовой активности антител. Примерные результаты этого опыта приведены в таблице 2. Подробности методики предоставлены в разделе "Материалы и методы".
1.7 Ингибирование индуцированного эндогенным IL-6 высвобождения VEGF из синовиального фибробласта человека очищенным IgG
Были определены активности антител при ингибировании индуцированного эндогенным IL-6 высвобождения VEGF из синовиальных фибробластов человека, взятых от доноров с ревматоидным артритом. Подробная методика этой процедуры предоставлена в разделе "Материалы и методы". Вкратце, титры испытуемого IgG были добавлены к культивированным фибробластам, которые затем были стимулированы добавкой человеческого IL-1β и растворимого человеческого IL-6Rα, чтобы вызвать экспрессию IL-6 и включить клеточную сигнализацию для индукции экспрессии VEGF. После 48 ч инкубации супернатанты были отделены и проверены с помощью ELISA на экспрессию VEGF с использованием продажного набора(R & D Systems). Эти данные использовались для определения IC50 для CAN022D10, которая была вычислена равной 45 нмоль.
Пример 2. Оптимизация антитела
2.1 Идентификация аминокислот, которые могут улучшать связывание лидирующего антитела с IL-6
Стратегия идентификации ключевых остатков в последовательности родительского антитела, которые могут улучшать связывание с IL-6, осуществлялась с помощью ввода случайных мутаций в последовательность CAN022D10 scFv. Это было достигнуто через два круга мутагенеза с помощью набора для случайного мутагенеза A DiversifyTM PCR (BD biosciences), следуя инструкциям производителя для ввода в среднем 8.1 мутаций на тысячу оснований за круг мутагенеза. Селекция производилась в основном как описано ранее (Hanes и др. 2000; Methods in Enzymology. 328. 404-430). Вкратце, библиотека случайного мутагенеза родительского клона была транскрибирована на мРНК, и с помощью процесса остановленной трансляции были образованы комплексы РНК-рибосома-scFv. Эти комплексы инкубировались с bio-huIL-6, и связавшиеся с антигеном были затем захвачены парамагнитными частицами, покрытыми стрептавидином. Неспецифические рибосомные комплексы были вымыты, из связанных рибосомных комплексов выделена мРНК, обратно транскрибирована в ДНК и затем размножена с помощью ПЦР. Эта ДНК использовались в следующем круге селекции и/или была клонирована для скрининга. Процесс селекции повторялся в присутствии снижающихся концентраций bio-huIL-6 (от 100 до 0,1 нмоль за 4 круга). ScFv, изолированный рибосомой, был клонирован в фагмидный вектор pCANTAB6 обработкой рестрикционной эндонуклеазой Nco1/Not1 (New England Biolabs) рибосомальной конструкции, с последующей сшивкой с pCANTAB6, обработанным Nco1/Not1, с помощью ДПК-лигазы Т4 (New England Biolabs) [127]. Сшитая ДНК затем была трансформирована в химически полноценные клетки TG-1, а неочищенный scFv из индивидуальных клонов конкурировал с CAN022D10 IgG за связывание с HIS/FLAG IL-6 в биохимическом анализе лиганд-антитело.
2.2 Идентификация улучшенных клонов с помощью биохимического анализа антитело-лиганд (с помощью CAN022D10 IgG)
Неочищенные препараты scFv из репрезентативного количества индивидуальных клонов из круга 3 и круга 4 были просеяны на ингибирующую активность в анализе связывания CAN022D10 IgG - IL-6 HTRF®. В этом анализе связывание биотинилированного антитела и FLAG-маркированного IL-6 определялось с помощью меченого криптатом моноклонального антитела анти-FLAG и стрептавидина XLent! (ТМ). Подробный метод количественного определения предоставлен в разделе "Материалы и методы".
ScFv, который проявлял значительный ингибирующий эффект, был секвенирован и произведен в виде очищенных препаратов, как описано в разделе 1.4. Величина IC50 для каждого scFv была вычислена из данных, полученных при тестировании серии разведении очищенного образца в биохимическом анализе HTRF антитело-лиганд и в анализе пролиферации TF-1. Самые активные в анализе пролиферации TF-1 клоны были конвертированы в IgG с постоянным доменом тяжелой цепи и постоянным доменом легкой каппа-цепи, как описано ранее, и вновь проверены в анализе пролиферации TF-1. Примерные данные по активности для очищенных scFv и IgG для каждого образца приведены в таблице 3.
2.3 Оптимизация родительского клона направленным мутагенезом
Лидирующие антитела были оптимизированы с помощью направленного мутагенеза с помощью аффинной фаговой селекции. Для подхода направленного мутагенеза были созданы большие библиотеки scFv-фагов, выведенных от лидирующих клонов, с помощью олигонуклеотид-направленного мутагенеза определяющих комплементарность областей 3 (CDR3) вариабельной тяжелой (VH) и легкой (VL) цепи с помощью стандартных методик молекулярной биологии [128]. Библиотеки подвергались аффинной фаговой селекции для отбора вариантов с высоким сродством к IL-6. Впоследствии они должны проявлять повышенную ингибирующую активность на связывание IL-6 с его рецептором. Селекция производилась в основном, как описано ранее [129]. Вкратце, частицы фага scFv инкубировались с рекомбинантным биотинилированным человеческим IL-6 в растворе (bio-huIL-6, собственный, из E.coli и модифицирован собственноручно). ScFv-фаг, связанный с антигеном, затем был захвачен парамагнитными частицами, покрытыми стрептавидином (Dynabeads® М 280) согласно рекомендациям производителя. Отобранные частицы scFv-фага были сохранены, как описано ранее [125], и процесс селекции повторялся в присутствии снижающихся концентраций bio-huIL-6 (от 500 до 0,1 нмоль за 3 круга).
По завершении 3 кругов селекции рандомизованные библиотеки VH и VL были объединены в одну библиотеку, в которой клоны содержали случайно спаренные индивидуально рандомизованные последовательности VH и VL. Затем селекция продолжалась, как описано ранее в присутствии снижающихся концентраций bio-huIL-6 (от 0,1 нмоль до 0,1 пмоль за 4 дальнейших круга).
2.4 Идентификация улучшенных клонов из направленного мутагенеза с помощью биохимического анализа антитело-лиганд (с помощью антитела 5 IgG)
Неочищенные scFv из клонов, выделенных из результатов селекции направленного мутагенеза, были тестированы с помощью биохимического анализа антитело-лиганд, по существу, как описано в разделе 2.2. Для этих результатов биохимический анализ был перенастроен на использование Антитела 5 IgG. Это антитело является улучшенным вариантом CAN02210 с более высокой активностью в анализе пролиферации TF-1. Вовлечение этого более активного IgG дает тест, способный различить клоны с более высокой активностью. Процедура этого модифицированного анализа была, как описано для исходного биохимического анализа антитело-лиганд с использованием CAN022D10, со следующими изменениями. Во-первых, используемая концентрация HIS FLAG IL-6 была уменьшена с 1 нмоль до 0,5 нмоль. Во-вторых, концентрации антитела анти-IL-6 и стрептавидина XLent! (ТМ) были увеличены с 1 нмоль и 20 нмоль до 16 нмоль и 40 нмоль соответственно. ScFv, который проявлял значительный ингибирующий эффект, был секвенирован и произведен в виде очищенных scFv и IgG, затем протестирован в анализе пролиферации TF-1.
2.5. Ингибирование индуцированной IL-6 пролиферации клеток TF-1 очищенным scFv и IgG оптимизированных клонов
Активности оптимизированных клонов определяли с помощью анализа индуцированной IL-6 пролиферации TF-1, как было описано ранее. Клоны тестировались в виде как очищенных препаратов scFv, так и переформатированных IgG. Примерные результаты для scFv и IgG даны в таблице 4.
Клоны проявляют значительный ингибирующий эффект, но точные значения IC50 не могут быть определены из серии разведений очищенного scFv.
2.6. Оптимизирование по зародышевой линии
Аминокислотные последовательности VH и VL доменов оптимизированных антител анти-IL-6 были соотнесены с известными последовательностями зародышевых линий человека в базе данных VBASE [130], и была идентифицирована ближайшая зародышевая линия по подобию последовательностей. Для Vн доменов антитела CANDY022D10 ближайший сегмент v зародышевой линии был Vh3_DP-86_(3-66), а ближайший сегмент j зародышевой линии был JH2. Для VL доменов ближайший сегмент v зародышевой линии был Vk1_L12, а ближайший сегмент j зародышевой линии был JK2.a
Без учета остатков Вернье [131], которые остались неизменными, было 3 изменения в каркасе VH доменов и 4 изменения в VL доменах, все из которых были превращены в ближайшие последовательности зародышевой линии для точного совпадения с антителами человека с помощью стандартных методик сайт-направленного мутагенеза с соответственными мутагенными праймерами.
Было всего обнаружено 5 остатков Вернье в последовательности scFv CAN022D10, мутировавших от зародышевой линии. Они были в тяжелой цепи у остатков Кабата 29 (I вместо V), 69 (М вместо I), 73 (I вместо N) and 78 (V вместо L). Одиночная мутация Вернье была обнаружена также в последовательности легкой цепи у остатка Кабата 46 (V вместо L).
Оптимизированный по зародышевой линии IgG затем был снова оценен в анализе индуцированной IL-6 пролиферации TF-1 для подтверждения отсутствия снижения активности. Примерные результаты для оптимизированных по зародышевой линии (GL) антител приведены в таблице 5.
2.7. Ингибирование индуцированного эндогенным IL-6 высвобождения VEGF из синовиального фибробласта человека оптимизированным IgG
Оптимизированные IgG были испытаны в анализе высвобождения VEGF синовиальными фибробластами для оценки активности против эндогенно экспрессируемого IL-6. Эта процедура упомянута в разделе и подробно описана в разделе "Материалы и методы". Примерные активности для испытанных IgG даны в таблице 6а. Средние данные по активности для испытанных IgG даны в таблице 6b.
2.8. Селективность и перекрестновидовая активность оптимизированных антител в тестах эпитопной конкуренции DELFIA®
Селективность и перекрестновидовая активность снова оценивалась для панели клонов в тестах эпитопной конкуренции DELFIA®, как было описано ранее (см. раздел 1.6 и "Материалы и методы"). IL-6 человека и макаки дают перекрывающиеся кривые ингибирования и таким образом неопределенные значения IC50 для всех испытанных IgG. He наблюдалось ингибирование для мышиного, крысиного, собачьего IL-6 и любого из родственных белков человека, испытанных на панели антител. Эти данные демонстрируют, что панель испытуемых клонов перекрестно реагирует с IL-6 макаки, но не связывается с IL-6 мыши, крысы и собаки или с человеческими белками, наиболее близкими к человеческому IL-6.
2.9 Вычисление аффинности оптимизированных клонов с помощью BIAcore
Связывающая аффинность очищенных образцов IgG репрезентативных антител 7 и 18 к IL-6 человека и макаки была определена методом поверхностного плазменного резонанса с помощью биодатчика BIAcore 2000 (BIAcore АВ), как описано в [132]. Вкратце, очищенные антитела присоединялись к поверхности датчика на чипе СМ5 с помощью аминосшивающего набора (BIAcore) для получения поверхностной плотности в пределах 220-225 Ru.
IL-6 человека и макаки в диапазоне концентраций от 200 нМ до 0,2 нМ в буфере HBS-EP пропускали над поверхностью датчика. Результирующие сенсорограммы оценивались с помощью программного обеспечения BIA evaluation 3.1 для получения относительных данных по связыванию.
Нижний предел измерения аффинности биодатчика BIAcore 2000™ составляет около 10 пМ (BIAcore 2000 Instrument handbook). Из полученных данных следует, что аффинность антител к IL-6 как человека, так и макаки, была ниже предела в 10 пМ, т.е. антитела были более активны, чем могло быть измерено. Поэтому точные значения аффинности не рассчитаны. Аффинность обоих антител к обоим видам IL-6 при таком подходе считается меньшей, чем 10 пМ.
2.10 Вычисление аффинности оптимизированных клонов с помощью анализа пролиферации клеток TF-1 In Vitro
Анализ TF-1 использовался для вычисления аффинности антитела 18 с помощью анализа Шильда. Стандартная кривая IL-6 (7,7×10-15 М до 3×10-9 М) была смешана с рядом концентраций IgG (2,67×10-13 М до 8,3×10-10 М) дважды. Из графика Log10 концентрации антитела против Log10 соотношения доз была определена аффинность IgG. С помощью такого подхода аффинность антитела 18 (GL) к человеческому IL-6 была вычислена равной 0,40 пМ (95% CI 0,12 пМ - 0,69 пМ, n=6).
2.11 Антагонистическая активность человеческого рекомбинантного IL-6 с помощью опосредованной IL-6 пролиферации клеток В9 In Vitro
Опосредованная IL-6 пролиферация клеток В9 осуществлялась в присутствии антитела 18 и изотипного контрольного антитела. Влияние ряда концентраций каждого антитела (1×10-13 М до 1×10-9 М) оценивалось на стандартной кривой IL-6 (диапазон концентраций от 1×10-14 М до 1×10-9 М). Точки данных были дублированы. Пролиферация В9 определялась после 4 дней инкубации по восстановлению аламара синего (флуоресцентный метод).
Показано, что антитело 18 ингибирует индуцированную IL-6 пролиферацию В9. Контрольный изотип не имеет ингибирующего эффекта. Средние данные приведены в таблице 8.
2.22 Антагонистическая активность человеческого рекомбинантного IL-6 с помощью опосредованного IL-6 высвобождения IgM из клеток SKW6.4 In Vitro
IL-6 индуцирует секрецию IgM из клеточной линии SKW 6.4 фибробластов человека В. Клетки SKW6.4, инкубированные в диапазоне концентраций IL-6 (от 1×10-13 М до 3×10-8,5 М), дают в среднем [А]50 77 пМ (n=3) для секреции IgM. Влияние антител 7, 17 и 18 против человеческого IL-6 и изотипного контрольного антитела на вызванную IL-6 секрецию IgM оценивалось по наблюдению ингибирования разных концентраций антитела (1×10-12,5 М до 1×10-8 М) в присутствии 100 пМ IL-6. Секреция IgM определялась через 4 дня с помощью античеловеческого IgM ELISA. Точки данных были дублированы.
Антитела 7, 17 и 18 ингибировали вызванную IL-6 секрецию IgM. Контрольный изотип не имеет ингибирующего эффекта в этих тестах. Средние данные приведены в таблице 9.
Пример 3. Картирование эпитопов
3.1 Сравнение эпитопа антитела анти-IL-6 с известными антителами анти-IL-6 человека
Эпитоп антитела 18 (GL) сравнивали с эпитопами двух антител против IL-6 человека - В-Е8 и cCLB8. Известно, что оба эти антитела ингибируют связывание IL-6 с IL-6Ra, и были исследованы как потенциальные терапевтические средства [5, 31, 34, 37, 133]. Чтобы обеспечить сравнение эпитопов этих трех антител, была сконструирована панель мутантов IL-6, каждый из которых содержал мутацию единичной аминокислоты по сравнению с последовательностью дикого типа (wt). Затем связывание этих мутантов с различными антителами оценивалось в биохимических конкурентных тестах. Эти эксперименты были основаны на биохимическом конкурентном методе, описанном в примере 1.6, с изменением, где это необходимо, концентраций антител и вариантов IL-6. Вкратце, антителами покрывали поверхность 96-ячеистой иммуноаналитической пластины Nunc Maxisorp при концентрации 2 нМ (антитело 18) или 4 нМ (В-Е8 и cCLB8) в PBS и инкубировали всю ночь при 4°C. После закрепления ячеек с помощью 3% (мас./об.) BSA в PBS к ячейкам, покрытым антителом, добавляли разведения ингибиторов в диапазоне концентраций от 200 нМ до 10 пМ в смеси с биотинилированным человеческим IL-6 при итоговой концентрации 0,15 нМ и оставили для связывания. Связывание биотинилированного IL-6 с антителами измерялось с помощью стрептавидина, меченного европием.
Сравнивая величины IC50, полученные для мутантов к немеченому человеческому IL-6 дикого типа, можно установить соотношение активностей для каждого мутанта. Далее, при сравнении этих соотношений касательно разных антител, можно оценить влияние индивидуальных мутаций на связывание антитела с молекулой IL-6. Типичные результаты этих опытов представлены в таблице 10 с экспериментами, повторенными в 2 дальнейших случаях.
ный
ный
S204E
Нерелевант
ный
wt IL-6
2.57Е-08
Нет
ингиб.
2.39E-10
09
1.90Е-
09
Нет
ингиб.
1.03Е-
09
08
4.83Е-
10
Нет
ингиб.
1.98Е-
10
107,754
-
1,000
1,848
-
1,000
2,434
-
1,000
Нумерация остатков в табл.10 приводится для аминокислотной последовательности полноразмерного IL-6 человека (SEQ ID NO:161).
Результаты демонстрируют, что три антитела имеют разные профили связывания с панелью мутантов IL-6 и, следовательно, связываются с разными эпигонами на поверхности цитокина. Kalai и др. (1997) ранее наблюдали, что cCLB8 не распознает мутантный IL-6 - F106E. Наши эксперименты подтвердили это; также он не ингибирует связывание биотинилированного IL-6 с антителом. С другой стороны, мутант F106E только в 5 раз менее активен, чем IL-6 дикого типа в конкурентном тесте, использующем антитело 18, что указывает на то, что он сильно связывается с этим антителом. Похожий результат наблюдался у мутанта Q211A, у которого соотношение активностей к антителу 18 было 1.5, сравнительно с 158 для cCLB8. Наоборот, мутанты F102E, R207E, R207L и S204E были мощными ингибиторами в анализе с cCLB8, но оказались значительно менее активными, чем дикий IL-6 в анализе с Антителом 18.
Различия в связывании Антитела 18 и В-Е8 наблюдались у мутантов R58E и S204E. Соотношение активностей для R58E было 2.009 для антитела 18 в сравнении с 79.083 для В-Е8, что указывает на то, что эта мутация уменьшает связывание В-Е8 с IL-6. Влияние мутации S204E оказывается специфическим для Антитела 18 из трех исследованных антител. Как и у cCLB8, эта мутация мало влияет на силу связывания IL-6 с В-Е8, однако мутант более чем в 100 раз менее активен, чем IL-6 дикого типа в биохимическом анализе для Антитела 18.
Пример 4. Введение антитела анти-IL-6 in vivo
4.1 Влияние введения антитела анти-IL-6 на рост нейтрофилов и гаптоглобина у мышей, вызванный человеческим рекомбинантным IL-6
Известно, что системное применение IL-6 вызывает системное повышение концентраций нейтрофилов и белков острой фазы. Была разработана модель in vivo, в которой человеческий IL-6 вводился внутрибрюшинно самцам мыши C57/B/6/J, и измерялись концентрации нейтрофилов и белка острой фазы гаптоглобина. Измерялась способность антитела 18 (GL), введенного подкожно, ингибировать эти ответы.
4.2 Гаптоглобиновый тест
Внутрибрюшинная инъекция человеческого IL-6 (5,2 нмоль/кг соответствует 12 мг/кг, b.i.d.) в течение 7 дней привела к значительному повышению уровней гаптоглобина плазмы от 0,02±0,01 мг/мл (контрольная группа) до 1,19±0,27 мг/мл в группе, получавшей IL-6 (Т-тест, Р<0,01). В то время как контрольный изотип IgG1 не проявлял эффекта, Антитело 18 дозозависимо ингибирует ответ со значительным ингибированием (ANOVA, Р<0,01 против IL-6 одного), замеченым в дозах 10,6 нмоль/кг (156 мг/кг) и выше (Фигура 1).
4.3 Нейтрофильный тест
Внутрибрюшинная инъекция человеческого IL-6 (5,2 нмоль/кг соответствует 12 мг/кг, b.i.d.) в течение 7 дней привела к значительному повышению количества нейтрофилов от 1,1±0,44×109 клеток/л (контрольная группа) до 2,47±0,12×109 клеток/л в группе, получавшей IL-6 (Т-тест, Р<0,01). В то время как контрольный изотип IgGI не проявлял эффекта, Антитело 18 дозозависимо ингибирует ответ со значительным ингибированием (ANOVA, Р<0,01 против IL-6 одного), замеченым в дозах 1,5 нмоль/кг (23 мг/кг) и выше.
Эти результаты подтверждают способность антител анти-Ib-6 ингибировать системные эффекты IL-6 in vivo.
Материалы и методы
Ингибирование связывания IL-6 с рецептором IL-6 неочищенным scFv
Продукты селекции подвергались скринингу в форме HTRF® (гомогенная флуоресценция с разрешением по времени) анализа на связывание рецептор-лиганд для ингибирования связывания человеческого IL-6, меченного криптатом (R&D Systems 206-IL), или человеческого IL-6, маркированного HIS FLAG (собственный из Е.coli) с биотинилированным IL-6R (Peprotech 200-06 R).
Продукты изоляции лидера подвергались скринингу в виде неразбавленных неочищенных scFv-содержащих периплазмических экстрактов, приготовленных в: 200 мМ hepes-буфере с рН 7,4, 0,5 мМ ЭДТА и 0,5 М сахарозы. 8 нМ биотинилированного человеческого IL-6R были проинкубированы в течение 30 минут при комнатной температуре в темноте с 8 нМ стрептавидина XLent! (ТМ) (CIS Bio International 611SAXLA). Все разбавления были сделаны на солевом растворе, забуференным фосфатом (PBS), содержащим 0,4 М фторида калия и 0,1% BSA (тестовый буфер).
После преинкубации реагентов 10 мкл неочищенного образца scFv поместили на 384-ячеистую малообъемную аналитическую пластинку (Costar 3676). После этого добавили 5 мкл проинкубированной смеси биотинилированного рецептора и стрептавидина XLent! (TM), а затем 5 мкл 11,2 нМ человеческого IL-6, меченного криптатом.
Аналитические пластинки центрифугировали при 1000 об/мин при комнатной температуре в течение 1 мин, затем инкубировали 2 ч при комнатной температуре перед считыванием данных флуоресценции с разрешением по времени при длинах волн испускания 620 и 665 нм с помощью считывателя пластинок Envision (Perkin Elmer).
Ингибирование связывания IL-6 с рецептором IL-6 очищенным scFv и IgG
Очищенные scFv и IgG из положительных клонов, идентифицированных при скрининге, были испытаны в анализе HTRF® на ингибирование связывания человеческого IL-6, маркированного HIS FLAG с биотинилированным IL-6R. 8 нМ биотинилированного человеческого IL-6R были проинкубированы в течение 30 минут при комнатной температуре в темноте с 8 нМ стрептавидина XLent! (TM). Все разбавления были сделаны на солевом растворе, забуференном фосфатом (PBS), содержащим 0,4 М фторида калия и 0,1% BSA (тестовый буфер).
Для установления активности клона использовалось титрование очищенного образца как измеренные величины IC50 в анализе. После преинкубации реагентов 10 мкл титра очищенного образца scFv поместили на 384-ячеистую малообъемную аналитическую пластинку (Costar 3676). После этого добавили 5 мкл проинкубированной смеси биотинилированного рецептора и стрептавидина XLent! (TM). 2 нМ человеческого IL-6, маркированного HIS FLAG, смешали с 1,732 нМ анти-flag IgG, меченого криптатом (CIS Bio International 61FG2KLB), и немедленно 5 мкл смеси добавили к аналитической пластинке.
Аналитические пластинки центрифугировали при 1000 об/мин при комнатной температуре в течение 1 мин, затем инкубировали 2 ч при комнатной температуре перед считыванием данных флуоресценции с разрешением по времени при длинах волн испускания 620 и 665 нм с помощью считывателя пластинок Envision (Perkin Elmer).
Анализ данных
Для анализа данных вышеописанных тестов HTRF® использовались следующие методы.
Данные анализировались посредством вычисления величин % Delta F для каждого образца. Delta F определяли согласно уравнению 1.
Уравнение 1:

Далее значения % Delta F использовались для вычисления % специфического связывания по уравнению 2.
Уравнение 2:
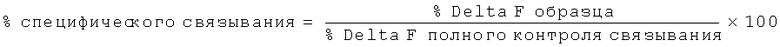
Значения IC50 были определены при помощи программного обеспечения GraphPad Prism методом подгонки кривой 4-параметрического логистического уравнения (Уравнение 3).
Уравнение 3:
Y=Низ+(Верх-Низ)/(1+10^((LogEC50-X)*Наклон))
X -- логарифм концентрации. Y -- специфическое связывание Y начинается снизу и идет вверх по сигмовидной кривой.
Стандартное антитело анти-IL-6 mAb (Biosource АНС0562) было включено во все тесты в качестве положительного контроля.
Ингибирование индуцированной IL-6 пролиферации клеток TF-1 очищенным scFv и IgG
Клетки TF-1 были предоставлены R&D Systems и содержались согласно прилагаемым процедурам. Среда для анализа включала RPMI-1640 с GLUTAMAX I (Invitrogen), содержащий 5% бычьей зародышевой сыворотки (JRH) и 1% пирувата натрия (Sigma). Перед каждым определением клетки TF-1 были гранулированы центрифугированием при 300×g в течение 5 мин, среды удалены отсасыванием, а клетки ресуспендированы в среде для анализа. Процесс был повторен дважды с ресуспендированием клеток с итоговой концентрацией 5×105 клеток/мл в аналитической среде. Клетки были посажены на 96-ячеистую аналитическую пластинку в количестве 100 мкл на ячейку. Пластинки инкубировали 24 часа при 37°C и 5% СО2 для истощения в них GM-CSF. Тестовые растворы очищенных scFv или IgG (дважды) были разбавлены аналитической средой до желаемой концентрации. Нерелевантное антитело, не направленное на IL-6, использовалось в качестве отрицательного контроля. Рекомбинантный IL-6 человека, произведенный бактериями (R&D) и IL-6 макаки (собственный), добавили в итоговой концентрации 20 пМ (человеческий IL-6) или 100 пМ (макака) при смешении с соответствующим тестовым антителом в общем объеме 100 мкл на ячейку. Концентрация IL-6, используемая в тесте, была выбрана как доза, которая при итоговой концентрации давала примерно 80% от максимального пролиферативного ответа. Все образцы инкубировали 30 мин при комнатной температуре. 100 мкл смеси IL-6 и антитела затем добавили к 100 мкл клеток, образуя итоговый объем анализа 200 мкл на ячейку. Пластинки инкубировали 24 часа при 37°C и 5% CO2. Затем к каждому образцу прибавили 20 мкл тритированного тимидина (5 мкКи/мл) и пластинки были возвращены в инкубатор на следующие 24 часа. Клетки были собраны на фильтрующих пластинках из стекловолокна (Perkin Elmer) с помощью собирателя клеток. Включение тимидина определяли с помощью микроплаты жидкостного сцинтилляционного счетчика Packard TopCount. Данные анализировались с помощью программного обеспечения Graphpad Prism.
Метод флуоресцентного анализа с разрешением по времени ингибирования связывания биотинилированного человеческого IL-6 с иммобилизованными антителами анти-IL-6
Специфический метод, используемый для этого анализа, результаты которого приводятся в примере 2.6, применяет реагенты DELFIA® и изложен выше. Метод также в более общем виде описывается ниже и пригоден для обнаружения и/или количественного определения связывания других форм IL-6 и родственных белков с анти IL-6 MAbs.
В этом способе моноклональное антитело анти-IL-6 связывается с твердой подложкой, например, соединяясь с подложкой через Fc. В качестве подложки могут использоваться полистирольные пластинки, связывающие белки, напр., пластинки Nunc Maxisorb.
- Покройте пластинки анти-IL-6 MAb в количестве 50 мкл на ячейку в PBS в течение ночи при 4°C.
- Все последующие операции производятся при комнатной температуре.
- Промойте пластинки три раза PBS, содержащим 0,05% Tween20 (PBST, продается Sigma P1379), потом закрепите с помощью 300 мкл на ячейку PBS содержащего 3% (мас./об.) BSA (продается Roche Diagnostics, 70129138) в течение 1 ч.
- Трижды промойте пластинки PBST.
- Приготовьте титры ингибитора в PBS, содержащем 3% (мас./об.) BSA и добавьте к 'разбавительной' пластинке (40 мкл на ячейку), потом 40 мкл на ячейку биотинилированного IL-6 так, чтобы итоговая концентрация биотинилированного IL-6 была эквивалентна KD белка для антитела. Перенесите 50 мкл образцов с разбавительной пластинки в соответствующие ячейки аналитической пластинки.
- Инкубируйте пластинки 1 ч.
- Трижды промойте пластинки PBST, затем в каждую ячейку добавьте 50 мкл на ячейку 0,1 мкг/мл стрептавидина, меченного европием, в 50 мМ трис-HCl, рН 7,5, содержащем 0,9% NaCl, 0,5% очищенного BSA, 0,1% Tween20 и 20 мкМ ЭДТА, инкубируйте 1 ч.
- Промойте пластинки семикратно промывочным буфером, состоящим из забуференного 0,05М трис солевого раствора (0,138М NaCl, 0,0027М KCl), 0,05% (об./об.) Tween20, рН8,0 (при 25°C).
- В каждую ячейку добавьте 50 мкл усиливающего раствора, подкисленного уксусной кислотой и содержащего Triton Х-100, вместе с хелаторами βNTA и ТОРО. Результирующий сдвиг рН от щелочного к кислому вызывает быструю диссоциацию ионов европия из стрептавидинового конъюгата. Затем свободные ионы европия образуют флуорогенные хелаты с присутствующими хелаторами. Вода удаляется в присутствии ТОРО, заставляя хелаты образовывать мицеллы, продлевая флуорогенность хелатов.
- Инкубируйте 5 мин, затем измерьте флуоресценцию с разрешением по времени при длине волны испускания 620 нм. Данные флуоресценции преобразуются в % специфического связывания согласно уравнению 1. Определите полное связывание по контрольной ячейке, содержащей биотинилированный huIL-6, но не конкурент. Определите неспецифическое связывание по ячейке, содержащей биотинилированный huIL-6 и 100-кратный избыток huIL-6. По полученным данным постройте сигмовидную кривую для вычисления величин IC50 согласно уравнению 2.
Определение покрытия антитела и концентраций биотинилированного huIL-6 для конкурентного биохимического анализа эпитопов
Концентрация антитела, используемого для покрытия, и концентрация биотинилированного huIL-6, используемого в конкурентном анализе эпитопов, будет зависеть от аффинности взаимодействия двух реагентов и эффективности иммобилизации антитела. Стандартная концентрация покрытия антитела и нужная концентрация биотинилированного huIL-6 должны поэтому быть определены для каждого испытуемого антитела.
Как общее правило, итоговая концентрация биотинилированного huIL-6, используемого в каждом тесте, эквивалентна KD лиганда для соответсвующего антитела, определяемая по анализу насыщения. Концентрация антитела, используемого для покрытия, должна быть такой, чтобы при добавлении биотинилированного huIL-6 при KD было получено соотношение сигнал/шум как минимум 10:1 при обнаружении в условиях конкурентного анализа.
Селективность и перекрестновидовая активность антител в тестах эпитопной конкуренции DELFIA®
Очищенный IgG адсорбировали на 96-ячеистые микротитровальные пластинки (Nunc) в PBS в концентрации, дающей значительный сигнал при добавлении биотинилированного человеческого IL-6 в количестве около его предполагаемого Кd для данного антитела IgG. Избыток IgG был вымыт PBS-Tween (0,1% об./об.), а ячейки закреплены с помощью PBS-Marvel (3% мас./об.) в течение 1 ч. Была приготовлена серия разведении каждого из следующих конкурентов в PBS, начиная с концентрации, примерно равной 200-кратному значению Kd взаимодействия между биотинилированным человеческим IL-6 и соответствующим IgG; человеческий IL-6, IL-6 макаки, IL-6 крысы (R & D Systems 506-RL/CF), мышиный IL-6 (R & D Systems 406-ML/CF), человеческий CNTF (R & D Systems 257-NT/CF), человеческий LIF (Chemicon, LIF1010), человеческий IL-11 (R & D Systems 518-IL/CF), человеческий онкостатин M (R & D Systems 295-OM/CF). Небиотинилированный человеческий IL-6 использовался в качестве положительного контроля. К этим сериям добавили равный объем биотинилированного рекомбинантного человеческого IL-6 в концентрации, приблизительно равной 2-кратному Kd (в результате серия начинается с соотношения конкурентный антиген:биотинилированный человеческий IL-6 около 100:1). Затем эти смеси переносили на закрепленный IgG и оставили на 1,5 ч для уравновешивания. Несвязанный антиген был удален промывкой PBS-Tween (0,1% об./об.), тогда как оставшийся биотинилированный человеческий IL-6 определялся по конъюгату стрептавидин-европий3+ (DELFIA®-детекция, PerkinElmer). Флуоресценция с разрешением по времени измерялась при 620 нм на считывающем устройстве Envision (PerkinElmer). Данные флуоресценции были преобразованы в % специфического связывания (100% определяли по контрольным ячейкам, содержащим биотинилированный человеческий IL-6, но не конкурент, 0% по ячейкам, содержащим биотинилированный человеческий IL-6 и 100-кратный избыток небиотинилированного человеческого IL-6). Результирующие данные были проанализированы с помощью программного обеспечения Prism (Graphpad) для определения значений IC50 согласно уравнению 3.
Метод флуоресцентного анализа с разрешением по времени ингибирования связывания биотинилированного человеческого IL-6 с иммобилизованными антителами анти-IL-6
Специфический метод, используемый для этого анализа, результаты которого приводятся в примере 2.8, применяет реагенты DELFIA® и изложен выше. Метод также в более общем виде описывается ниже и пригоден для обнаружения и/или количественного определения связывания других форм IL-6 и родственных белков с анти IL-6 MAbs.
В этом способе моноклональное антитело анти-IL-6 связывается с твердой подложкой, например, соединяясь с подложкой через Fc. В качестве подложки могут использоваться полистирольные пластинки, связывающие белки, напр., пластинки Nunc Maxisorb.
- Покройте пластинки анти-IL-6 МАb в количестве 50 мкл на ячейку в PBS, в течение ночи при 4°C.
- Все последующие операции производятся при комнатной температуре.
- Промойте пластинки три раза PBS, содержащим 0,05% Tween20 (PBST, продается Sigma P1379), потом закрепите с помощью 300 мкл на ячейку PBS, содержащего 3% (мас./об.) BSA (продается Roche Diagnostics, 70129138) в течение 1 ч.
- Трижды промойте пластинки PBST.
- Приготовьте титры ингибитора в PBS, содержащем 3% (мас./об.) BSA и добавьте к 'разбавительной' пластинке (40 мкл на ячейку), потом 40 мкл на ячейку биотинилированного IL-6 так, чтобы итоговая концентрация биотинилированного IL-6 была эквивалентна KD белка для антитела. Перенесите 50 мкл образцов с разбавительной пластинки в соответствующие ячейки аналитической пластинки.
- Инкубируйте пластинки 1 ч.
- Трижды промойте пластинки PBST, затем в каждую ячейку добавьте 50 мкл на ячейку of 0,1 мкг/мл стрептавидина, меченного европием, в 50 мМ трис-HCl, рН 7,5, содержащем 0,9% NaCl, 0,5% очищенного BSA, 0,1% Tween20 и 20 мкМ ЭДТА, инкубируйте 1 ч.
- Промойте пластинки семикратно промывочным буфером, состоящим из забуференного 0,05М трис солевого раствора (0,138М NaCl, 0,0027М KCl), 0,05% (об./об.) Tween20, рН 8,0 (при 25°C).
- В каждую ячейку добавьте 50 мкл усиливающего раствора, подкисленного уксусной кислотой и содержащего Triton X-100, вместе с хелаторами βNTA и ТОРО. Результирующий сдвиг рН от щелочного к кислому вызывает быструю диссоциацию ионов европия из стрептавидинового конъюгата. Затем свободные ионы европия образуют флуорогенные хелаты с присутствующими хелаторами. Вода удаляется в присутствии ТОРО, заставляя хелаты образовывать мицеллы, продлевая флуорогенность хелатов.
- Инкубируйте 5 мин, затем измерьте флуоресценцию с разрешением по времени при длине волны испускания 620 нм. Данные флуоресценции преобразуются в % специфического связывания согласно уравнению 1. Определите полное связывание по контрольной ячейке, содержащей биотинилированный huIL-6, но не конкурент. Определите неспецифическое связывание по ячейке, содержащей биотинилированный huIL-6 и 100-кратный избыток huIL-6. По полученным данным постройте сигмовидную кривую для вычисления величин IC50 согласно уравнению 2.
Определение покрытия антитела и концентраций биотинилированного huIL-6 для конкурентного биохимического анализа эпитопов
Концентрация антитела, используемого для покрытия, и концентрация биотинилированного huIL-6, используемого в конкурентном анализе эпитопов, будут зависеть от аффинности взаимодействия двух реагентов и эффективности иммобилизации антитела. Стандартная концентрация покрытия антитела и нужная концентрация биотинилированного huIL-6 должны поэтому быть определены для каждого испытуемого антитела.
Как общее правило, итоговая концентрация биотинилированного huIL-6, используемого в каждом тесте, эквивалентна KD лиганда для соответсвующего антитела, определяемая по анализу насыщения. Концентрация антитела, используемого для покрытия, должна быть такой, чтобы при добавлении биотинилированного huIL-6 при KD было получено соотношение сигнал/шум как минимум 10:1 при обнаружении в условиях конкурентного анализа.
Идентификация улучшенных клонов с помощью биохимического анализа антитело-лиганд
Продукты селекции при оптимизации лидера подвергались скринингу в форме анализа HTRF® на конкуренцию эпитопов для ингибирования связывания человеческого IL-6, маркированного HIS FLAG (собственный, из Е.coli) с биотинилированным антителом анти-IL-6 (собственный, из IgG через выделение лидера, CAN022D10).
Продукты изоляции лидера подвергались скринингу в виде неразбавленных неочищенных scFv-содержащих периплазмических экстрактов, приготовленных в: 50 нМ MOPS-буфере с рН 7,4, 0,5 мМ ЭДТА и 0,5 М сорбита. 1 нМ человеческого HIS FLAG IL-6 были проинкубированы в течение 30 минут при комнатной температуре в темноте с 1,732 нМ анти-flag IgG, меченного криптатом (CIS Bio International 61FG2KLB). Все разбавления производились на аналитическом буфере. Параллельно, 1 нМ биотинилированного анти-IL-6 IgG (против которого надо измерить конкуренцию тестового связывающего элемента) были проинкубированы в течение 30 минут при комнатной температуре в темноте с 20 нМ стрептавидина XLent! (TM) (CIS Bio International 611SAXLB).
После преинкубации реагентов 10 мкл неочищенного образца scFv поместили на черную 384-ячеистую оптическую пластинку (Perkin Elmer кат. №6007279). После этого добавили 10 мкл аналитического буфера на всю пластинку. Затем добавили 10 мкл проинкубированной смеси биотинилированного анти-IL-6 IgG и Стрептавидина XLent! (TM), и 10 мкл преинкубированной смеси маркированного HIS FLAG человеческого IL-6 и анти-flag криптата.
Аналитические пластинки центрифугировали при 1000 об/мин при комнатной температуре в течение 1 мин, затем инкубировали 2 ч при комнатной температуре перед считыванием данных флуоресценции с разрешением по времени при длинах волн испускания 620 и 665 нм с помощью считывателя пластинок EnVision (Perkin Elmer). Данные анализировались посредством вычисления % deltaF и % специфического связывания, как описано ранее.
После идентификации улучшенных лидеров из библиотеки случайного мутагенеза неочищенные неразбавленные продукты scFv от селекции на CDR3 направленным мутагенезом подвергались скринингу в модифицированной версии анализа HTRF® на конкуренцию эпитопов, включающей следующие изменения: 0,5 нМ человеческого HIS FLAG IL-6 были проинкубированы в течение 30 минут при комнатной температуре в темноте с 1,732 нМ анти-flag IgG, меченного криптатом (CIS Bio International 61FG2KLB). Параллельно, 16 нМ биотинилированного анти-flag IgG (Антитело 5, собственное, из IgG, идентифицированного из отбора CAN022D10 случайным мутагенезом) были проинкубированы в течение 30 минут при комнатной температуре в темноте с 40 нМ стрептавидина XLent! (TM) (CIS Bio International 611SAXLB). Все остальные условия, как описано для анализа конкуренции эпитопов CAN022D10. Данные анализировались посредством вычисления % deltaF и % специфического связывания, как описано ранее.
Ингибирование индуцированного эндогенным IL-6 высвобождения VEGF из синовиальных фибробластов человека очищенным IgG
Образцы колен с ревматоидным артритом после операции полной замены сустава были получены в DMEM, содержащем антибиотики. Синовиальная мембрана, погруженная в среду, была отсечена от сустава и мелко нарублена. Синовиальная ткань была промыта средой, снабженной 10% FCS. Суспензия клеток инкубировалась в растворе коллагеназы 2 часа в СО2 инкубаторе при 37°C. Переваренная суспензия синовиальных клеток была разрушена многократным всасыванием через 10 мл пипетку, клетки процежены и отцентрифугированы при 400 g при комнатной температуре в течение 5. Клетки были ресуспендированы в DMEM, содержащем 10% FCS, пропущены через клеточное сито, доведены до 1×106 клеток в мл и инкубированы в СО2 инкубаторе при 37°C в 225 см2 колбах для клеточных культур (3001, CoStar Corning Inc.). После прилипания большую часть среды отбрасывают, заменяют свежей и возвращают в инкубатор для длительной инкубации. Клетки были осмотрены через неделю и подвергнуты слиянию трипсинизацией при скорости прохождения 1 в 3.
Фибробласты (Р3-5) при слиянии были удалены из колб инкубированием с 10 мл 0,1% раствора трипсин-ЭДТА (25300-054, Gibco Life Sciences) на колбу в течение 5-10 минут при 37°C. Равный объем культуральной среды на основе DMEM, снабженной 10% FCS, прибавили к клеткам, которые затем были гранулированы центрифугированием при 330 g в течение 5 минут при комнатной температуре. После однократной промывки культуральной средой на основе DMEM, снабженного 10% FCS, суспензия клеток (1×105 клеток на 1 мл) была помещена (150 мкл на ячейку) в ячейки стерильных 96-ячеистых плоскодонных полистирольных пластинок (3598, Corning CoStar) в количестве 1.5×104 клеток на ячейку. Далее в каждую ячейку добавили культуральной среды на основе DMEM, снабженной 10% FCS (100 мкл на ячейку), полный объем становится 250 мкл на ячейку. Клетки инкубировали всю ночь при 37°C для обеспечения прилипания и неподвижности.
96-ячеистые пластинки были осмотрены, чтобы убедится, что все клетки сливаются и в хорошем состоянии (напр., без контаминации). Затем среду отсосали из ячеек и немедленно добавили 100 мкл культуральной среды на основе DMEM, снабженной 10% FCS. Далее, в ячейки добавили 50 мкл культуральной среды на основе DMEM с добавкой 10% FCS, содержащий образец IgG или саму среду (разбавлен 1 к 5 для анализа).
Далее добавили 50 мкл на ячейку культуральной среды на основе DMEM с добавкой 10% FCS, содержащую растворимый рекомбинантный человеческий (rhs)IL-6Rα (500 нг/мл; 12 нМ) и rhIL-1β (50 пг/мл; 2,95 пМ, разбавленный 1 к 5 для анализа).
В отдельные ячейки добавили 50 мкл культуральной среды на основе DMEM с добавкой 10% FCS, содержащую: rh-IL-6 (0,100 нг/мл; 21,5 нМ), sIL-6Rα (500 нг/мл; 12 нМ), rhIL-1β (50 пг/мл; 2,95 пМ), или саму среду (разбавлена 1 к 5 для анализа). Итоговый объем в каждой ячейке составил 250 мкл.
Пластинки были инкубированы 48 часов при 37°C. Инкубация производилась в двойных или тройных ячейках, как описано в формате пластинки. Пластинки были центрифугированы при 330 g в течение 5 минут при комнатной температуре, а супернатант был слит и сохранялся при -40°C в плоскодонных микротитровальных пластинках (611F96, Sterilin).
VEGF измерялся с помощью ELISA (DY293B, R&D Systems) согласно инструкциям производителя. Вкратце, пластинки ELISA были покрыты мышиным антителом против человеческого VEGF на ночь при 4°C и закреплены с помощью 1% BSA/PBS. Пластинки были промыты 0,05% Tween 20/PBS и инкубированы с супернатантами культуры синовиальных фибробластов человека и биотинилированным козлиным антителом к человеческому VEGF на ночь при комнатной температуре. После промывки VEGF был детектирован с помощью Стрептавидин пероксидазы хрена. Пластинки были проявлены с помощью 1:1 H2O2:тетраметилбензидин.
Реакция была остановлена 2 М H2SO4 и определили оптические плотности при 450 нм с коррекцией длины волны при 540 нм.
Измерения BIAcore
Исследования BIAcore были предприняты с помощью BIAcore 2000™. Антитела присоединялись к поверхности датчика на чипе СМ5 с помощью аминосшивающего набора для получения поверхностной плотности в пределах 220-225 Ru. IL-6 человека в диапазоне концентраций от 200 нМ до 0.2 нМ в буфере HBS-EP пропускали над поверхностью датчика. Результирующие сенсорограммы оценивались с помощью программного обеспечения BIA evaluation 3.1 для вычисления значений kon, koff и KD для испытуемых антител.
Анализ опосредованной IL-6 пролиферации клеток В9
Клетки В9 являются субклоном мышиной клеточной линии В-клеточной гибридомы, В13.29, отобранным на основе его специфического ответа на IL-6. Клетки В9 требуют IL-6 для выживания и пролиферации и откликаются на очень низкие концентрации IL-6.
Опосредованная IL-6 пролиферация клеток В9 осуществлялась в присутствии антитела 18 и изотипного контроля (CAT-002). Влияние ряда концентраций каждого антитела (1×10-13 М до 1×10-9 М) оценивалось на стандартной кривой IL-6 (диапазон концентраций от 1×10-14 М до 1×10-9 М). Точки данных были дублированы. Пролиферация В9 определялась после 4 дней инкубации по восстановлению аламара синего (флуоресцентный метод).
Клетки В9 были культивированы в RPMI-1640, содержащем 5% FCS, 2 мМ L-глутамина и 50 мкМ 2-меркаптоэтанола. Клетки делились каждые 2-4 дня до плотности между 0,05×106 мл-1 и 0,1×106 мл-1 и были снабжены 5×10-13 М человеческого IL-6. Клетки, используемые в эксперименте, не снабжались IL-6 как минимум 48 часов перед экспериментом, но должны снабжаться в течение 96 часов эксперимента. Клетки, используемые в анализе, были взяты из колбы с плотностью не более чем 0,8×106 мл-1.
Каждое антитело было разбавлено из исходного раствора до 10-кратной максимально необходимой концентрации соответственным разбавлением аналитической среды (RPMI+5%FCS, 2 мМ L-глутамина, 50 мкМ 2-меркаптоэтанол, пенициллин 100 Uмл-1 и стрептомицин 100 мгмл-1). Дальнейшие 10-кратные разведения в культуральной среде были проведены для получения необходимых концентраций каждого антитела.
IL-6 был воссоздан из лиофилизованного порошка в 1×10-5 M раствор добавлением соответствующего объема стерильного PBS+0,1% BSA. Дальнейшее разбавление до 1×10-8 M производится в культуральной среде. 1×10-8 М-аликвоты сохраняются замороженными, пока не понадобятся. В день анализа 1×10-8 М аликвоты разбавляются по необходимости до достижения диапазона растворов с необходимой итоговой концентрацией 10x.
Необходимые объемы клеток берут из культуральных колб и центрифугируют при 300 g в течение 8 минут. Супернатанты сливаются, а клетки ресуспендируются в соответствующем объеме культуральной среды до достижения плотности клеток 0.5×106 мл-1.
Анализ выполняется в плоскодонных полистирольных 96-ячеистых пластинках. Итоговый объем раствора равен 200 мкл. 20 мкл 10х раствора антитела (Антитело 18 или CAT-002) или культуральной среды поместили в соответствующие ячейки каждой пластинки, а затем добавили 140 мкл культуральной среды и 20 мкл соответствующей концентрации IL-6, или культуральной среды.
Пластинки поместили в увлажненный инкубатор с 5% СО2, 37°С на 2 часа. Затем в каждую ячейку добавили 20 мкл клеток. Итоговое количество клеток на ячейку равно 10000. Далее пластинки вернули в инкубатор на 4 дня. Пролиферация клеток определялась по восстановлению аламара синего. В каждую ячейку добавили 10% об./об. аламара синего и вернули пластинки в инкубатор на 6 часов. Флуоресценция пластинок измерялась на спектрофлуориметре при 590 нм с длиной волны возбуждения 544 нм.
Сырые данные были нормированы к контрольной кривой IL-6 на каждой пластинке так, что максимум флуоресценции был принят за 100%, а фоновая флуоресценция за 0%. Из нормированных данных была построена сигмовидная кривая доза-ответ с помощью нелинейной регрессии (переменный наклон) программы Graph Pad Prism 4.01. Контрольные значения pEC50 и значения pEC50 в присутствии каждой концентрации антитела были использованы для определения соотношения доз (DR). Значения Kb были определены для наименьшей концентрации антитела, которая проявляет 3-кратный или больший сдвиг на кривой эффекта концентрации IL-6, с помощью следующего уравнения химического антагонизма:
Kb=([Ab]/(DR-1))
(Kenakin ТР. In: Pharmacologic Analysis of Drug-Receptor Interactions. 1st ed. New York: Raven Press; 1987. p.205-24.).
Анализ опосредованного IL-6 высвобождения IgM из клеток SKW6.4
IL-6 участвует в окончательном созревании В-клеток в антителопродуцирующие клетки (дифференциация В-лимфоцитов). SKW-клетки ранее использовались в исследовании В-клеточного ответа (Nawata и др., Ann. N.Y. Acad. Sci. 557:230-238. 1989). Автоантитела, образующиеся при ревматоидном артрите, в основном принадлежат к классу IgM. SKW6.4 - это клональный секретирующий IgM человеческий лимфобластоид В-клеточной линии. Клетки получены в АТСС, ссылка #TIB 215. При стимуляции IL-6 эти клетки секретируют IgM, так что этот анализ считается важным при ревматоидном артрите.
Индуцированная IL-6 секреция IgM клетками SKW6.4 осуществлялась в присутствии САТ6001 и CAT-002 (изотипный контроль). Влияние ряда концентраций каждого антитела (1×10-12,5 М до 1×10-8 М) оценивалось в присутствии 100 пМ IL-6. Точки данных были дублированы. Секреция IgM в клеточном супернатанте определялась через 4 дня инкубации с помощью анализа античеловеческого IgM ELISA.
Клетки SKW 6.4 культивировались в RPMI1640, содержащем 2 мМ L-глутамина и 10% (об./об.) сыворотки зародыша теленка при 37°C и 95/5% (об./об.) воздух/СО2 и 95% относительной влажности. Клетки содержались при 0,4-2×106 клеток/мл. Для обычного пассажа клеток клетки были собраны 5-минутным центрифугированием при 300 х g при комнатной температуре, истощенная среда удалена, а клетки ресуспендированы в нужном объеме свежей среды.
Каждое антитело было разбавлено из исходного раствора до 50-кратной максимально необходимой концентрации соответственным разбавлением аналитической среды (RPMI+5%FCS, 2 мМ L-глутамина). Дальнейшие 10-кратные разведения в культуральной среде были проведены для получения необходимых концентраций каждого антитела.
Анализ выполняется в плоскодонных полистирольных 96-ячеистых пластинках. Клетки SKW 6.4 из хранилища были разбавлены до плотности клеток 0,3×106 мл-1 в свежей среде, и посажены в количестве 100 мкл на ячейку (30,000 клеток на ячейку). Затем в каждую ячейку добавили 2 мкл антитела указанной итоговой концентрации, а потом 2 µл IL-6 при итоговой концентрации 100 пМ.
Далее пластинки были возвращены в инкубатор при 37°C 5% CO2. Свободные от клеток супернатанты собирали центрифугированием после 4 дней инкубации и далее анализировали методом IgM ELISA в день сбора, или замораживали при -20°C до дальнейшего анализа.
ELISA генерировалась с помощью пары антител от Serotec. Покрывающее антитело было мышиным против человеческого IgM (МСА1662), а детектирующее антитело было козлиным против человеческого IgM: HRP связанный (STAR98P). Анализ был оптимизирован стандартными методами, обеспечив хорошее соотношение сигнал/ шум при использовании покрывающего антитела в разведении @ 1:2000 (5 мкг/мл) и детектирующего антитела в разведении @ 1:3500 (200 нг/мл).
Стандартный раствор IgM (Cat# PHP003 Human M Kappa purified protein) был приобретен у Serotec для получения стандартной кривой.
Данные анализировались с использованием полиномного приближения для данных стандартной кривой IgM при помощи стандартной вычислительной программы. Процент ингибирования каждого образца антител по сравнению с контрольным производством IgM в отсутствие антигена был вычислен и были генерированы значения IC50.
Генерирование IL-6 и мутантных белков IL-6 для картирования эпитопов
Клонирование ДНК человеческого и обезьяньего IL-6
Последовательности IL-6 человека и макаки были получены у Embl (No доступа: BC0155I1 и АВ000554 для человеческого и обезьяньего соответственно). С помощью этих последовательностей были разработаны олигонуклеотидные праймеры для амплификации кДНК, кодирующей IL-6 человека и макаки. N-терминальные праймеры были hIL6_5'NdeI и macIL6_5'NdeI для человека и обезьяны соответственно, а macIL6_3'NheI использовался как C-терминальный праймер для обоих (См. табл.11 для последовательностей олигонуклеотидов).
Для амплификации двух кДНК были проведены PCVR-реакции. Затравкой для каждой ПЦР было 10 нг кДНК, полученной из печени человека или печени макаки соответственно. Амплифицированная кДНК из каждой реакции была очищена и клонирована в pCR4blunt topo (Invitrogen) с помощью реакции сшивания топоизомеразы согласно производителю.
Положительные клоны были идентифицированы и секвенированы. Результирующие кДНК были субклонированы с помощью стандартных методик в различные векторы экспрессии Т7-промотора Е.coli таким способом, чтобы кДНК, кодирующие зрелые IL-6 человека или макаки, соединялись N-концом с N-концом метки HISG-FLAG непосредственно вслед за N-концевым валином зрелого IL-6.
Генерирование мутантов
Сайт-направленный мутагенез был осуществлен с помощью набора Quikchange XL от Stratagene согласно инструкции производителя.
Праймер для мутагенеза был разработан согласно инструкции производителя. Реакции мутагенеза были проведены согласно процедуре с использованием в качестве шаблона плазмиды pT7flagHISIL-6. Далее следовало переваривание с помощью DpnI и трансформация в химически компетентные клетки Тор10 с отбором на агаровых пластинках, содержащих соответствующие антибиотики при 37°C на ночь. Для каждой отдельной реакции мутагенеза были секвенированы несколько клонов, а плазмидная ДНК одного правильного клона из каждой реакции сохранялась для последующего использования.
Экспрессия IL-6 и мутантных белков IL-6
Плазмиды, экспрессирующие IL-6, были трансформированы в химически компетентные звездчатые клетки BL21 (DE3) (Invitrogen) при помощи метода производителя. Трансформированные клетки использовались для инокуляции культур 1L Terrific Broth, которые были инкубированы в орбитальном инкубаторе при 37°C, пока А600 не достиг 0,5. Затем добавили IPTG до 0,25 мМ, и инкубация продолжалась всю ночь при 22°C. Клетки были собраны центрифугированием, и осажденные клетки сохранялись при -80°C
Очистка IL-6 и мутантных белков IL-6
Гранулированные клетки оттаивали и ресуспендировали в 50 мл на гранулу 50 мМ фосфата калия, рН 7,4, 10 мМ имидазола, 0,3М NaCl, 5 мМ бета-меркаптоэтанола, 10% глицерина (буфер А)+полные ингибиторы протеаз без ЭДТА(Roche). Клетки были лизированы ультразвуком в течение 3×30 секунд во льду. Лизат был центрифугирован при 100,000 g и 4°C 30 минут, а супернатант подвергли аффинной хроматографии на Ni NTA. 5-мл колонка Ni-NTA Superflow (Qiagen) была уравновешена при 3 мл/мин (буфером А). Образец IL-6 был загружен в колонку, промыт 10 объемами колонки 15 мМ имидазола в буфере А. Затем следовала промывка 10 объемами колонки 30 мМ имидазола в буфере А. IL-6 был вымыт с колонки с помощью 5 объемов промывки в направлении снизу вверх 0,3М имидазола в буфере А. При промывках собирались фракции по 10 мл, а при элюировании по 5 мл. Колонка работала при 4°C с помощью AKTA Explorer 100 Air. Фракции, содержащие очищенный белок IL-6, были собраны и диализованы всю ночь при 4°C против 5 л PBS.
Диализованные белки IL-6 были далее очищены с помощью гель-фильтрационной хроматографии. При каждой очистке диализованный белок IL-6 был центрифугирован при 100,000 g и 4°C 20 минут. До 13 мл наносились на 318 мл колонку Superdex 200 26/60 (GE Healthcare), которая уравновешивалась PBS при 2,5 мл/мин. Колонка работала при 4°C с помощью АКТА Purifier. Фракции, содержащие мономерный пик белка IL-6, были собраны для дальнейшего анализа.
Каждый белок был проверен на чистоту с помощью стандартной SDS-хроматографии, измерялась концентрация белка, a Q-ToF масс-спектроскопия использовалась для измерения молекулярной массы белка. Очищенный IL-6 был заморожен в жидком азоте и хранился при -80°C.
Материалы и методы для исследований in vivo
Животные были случайно распределены между тестовыми группами. Мыши в каждой тестовой группе каждый день получали подкожные дозы (10 мл/кг) контрольного носителя (0,05% BSA в PBS), или 467 мкг/кг контрольного изотипа IgG1, или антитело 18 (в пределах от 467 мкг/кг до 8 мкг/кг). В то же время мыши получали внутрибрюшинную инъекцию (10 мг/кг) b.i.d. или другого контрольного носителя (0,05% BSA в PBS), или 12 мкг/кг человеческого рекомбинантного IL-6.
На 7 день, через два часа после последней дозы IL-6 в 09:00, мыши были забиты, и были взяты пробы крови из конечностей. Кровь была перенесена в 1 мл пробирки для крови Lab Tek с ЭДТА, которые поместили в роллер на 5 минут. Образцы хранились во льду до использования. Был произведен подсчет различных клеток с помощью счетчика клеток Sysmex. Остаток образца был перенесен в эппендорф и центрифугирован (300 g, 5 мин) для получения плазмы, из которой была отобрана аликвота, которая сохранялась при -20°C до анализа уровней гаптоглобина.
Гаптоглобиновый анализ может быть осуществлен по инструкциям, предоставленным в наборе PHASE™ RANGE TriDelta Format Biognosis (Hailsham, UK; cat. no. TP-801).
Все результаты были выражены в виде среднее±CKO. Анализ данных являлся неспаренным Т-тестом или односторонним ANOVA с последующим тестом Dunnett'a (GraphPad Instat).
Последовательности
VH домен, VL домен и CDR-последовательности связывающих членов приведены в прилагающемся списке последовательностей, в котором SEQ ID NOS соответствуют следующему:
1 CAN022D10 VH нуклеотидн.
2 CAN022D10 VH аминокислотн.
3 CAN022D10 VH CDR 1 аа
4 CAN022D10 VH CDR 2 аа
5 CAN022D10 VH CDR 3 аа
6 CAN022D10 VL нуклеотидн.
7 CAN022D10 VL аминокислотн.
8 CAN022D10 VL CDR 1 аа
9 CAN022D10 VL CDR 2 аа
10 CAN022D10 VL CDR 3 аа
11 Антитело 2 VH нуклеотидн.
12 Ab 2 VH аминокислотн.
13 Ab 2 VH CDR 1 аминокислотн.
14 Ab 2 VH CDR 2 аминокислотн.
15 Ab 2 VH CDR 3 аминокислотн.
16 Ab 2 VL нуклеотидн.
17 Ab 2 VL аминокислотн.
18 Ab 2 VL CDR 1 аминокислотн.
19 Ab 2 VL CDR 2 аминокислотн.
20 Ab 2 VL CDR 3 аминокислотн.
21 Антитело 3 VH нуклеотидн.
22 Ab 3 VH аминокислотн.
23 Ab 3 VH CDR 1 аминокислотн.
24 Ab 3 VH CDR 2 аминокислотн.
25 Ab 3 VH CDR 3 аминокислотн.
26 Ab 3 VL нуклеотидн.
27 Ab 3 VL аминокислотн.
28 Ab 3 VL CDR 1 аминокислотн.
29 Ab 3 VL CDR 2 аминокислотн.
30 Ab 3 VL CDR 3 аминокислотн.
31 Антитело 4 VH нуклеотидн.
32 Ab 4 VH аминокислотн.
33 Ab 4 VH CDR 1 аминокислотн.
34 Ab 4 VH CDR 2 аминокислотн.
35 Ab 4 VH CDR 3 аминокислотн.
36 Ab 4 VL нуклеотидн.
37 Ab 4 VL аминокислотн.
38 Ab 4 VL CDR 1 аминокислотн.
39 Ab 4 VL CDR 2 аминокислотн.
40 Ab 4 VL CDR 3 аминокислотн.
41 Антитело 5 VH нуклеотидн.
42 Ab 5 VH аминокислотн.
43 Ab 5 VH CDR 1 аминокислотн.
44 Ab 5 VH CDR 2 аминокислотн.
45 Ab 5 VH CDR 3 аминокислотн.
46 Ab 5 VL нуклеотидн.
47 Ab 5 VL аминокислоты.
48 Ab 5 VL CDR 1 аминокислотн.
49 Ab 5 VL CDR 2 аминокислотн.
50 Ab 5 VL CDR 3 аминокислотн.
51 Антитело 7 VH нуклеотидн.
52 Ab 7 VH аминокислотн.
53 Ab 7 VH CDR 1 аминокислотн.
54 Ab 7 VH CDR 2 аминокислотн.
55 Ab 7 VH CDR 3 аминокислотн.
56 Ab 7 VL нуклеотидн.
57 Ab 7 VL аминокислотн.
58 Ab 7 VL CDR 1 аминокислотн.
59 Ab 7 VL CDR 2 аминокислотн.
60 Ab 7 VL CDR 3 аминокислотн.
61 Антитело 8 VH нуклеотидн.
62 Ab 8 VH аминокислотн.
63 Ab 8 VH CDR 1 аминокислотн.
64 Ab 8 VH CDR 2 аминокислотн.
65 Ab 8 VH CDR 3 аминокислотн.
66 Ab 8 VL нуклеотидн.
67 Ab 8 VL аминокислотн.
68 Ab 8 VL CDR 1 аминокислотн.
69 Ab 8 VL CDR 2 аминокислотн.
70 Ab 8 VL CDR 3 аминокислотн.
71 Антитело 10 VH нуклеотидн.
72 Ab 10 VH аминокислотн.
73 Ab 10 VH CDR 1 аминокислотн.
74 Ab 10 VH CDR 2 аминокислотн.
75 Ab 10 VH CDR 3 аминокислотн.
76 Ab 10 VL нуклеотидн.
77 Ab 10 VL аминокислотн.
78 Ab 10 VL CDR 1 аминокислотн.
79 Ab 10 VL CDR 2 аминокислотн.
80 Ab 10 VL CDR 3 аминокислоты.
81 Антитело 14 VH нуклеотидн.
82 Ab 14 VH аминокислотн.
83 Ab 14 VH CDR 1 аминокислотн.
84 Ab 14 VH CDR 2 аминокислотн.
85 Ab 14 VH CDR 3 аминокислотн.
86 Ab 14 VL нуклеотидн.
87 Ab 14 VL аминокислотн.
88 Ab 14 VL CDR 1 аминокислотн.
89 Ab 14 VL CDR 2 аминокислотн.
90 Ab 14 VL CDR 3 аминокислотн.
91 Антитело 16 VH нуклеотидн.
92 Ab 16 VH аминокислотн.
93 Ab 16 VH CDR 1 аминокислотн.
94 Ab 16 VH CDR 2 аминокислотн.
95 Ab 16 VH CDR 3 аминокислотн.
96 Ab 16 VL нуклеотидн.
97 Ab 16 VL аминокислотн.
98 Ab 16 VL CDR 1 аминокислотн.
99 Ab 16 VL CDR 2 аминокислотн.
100 Ab 16 VL CDR 3 аминокислотн.
101 Антитело 17 VH нуклеотидн.
102 Ab 17 VH аминокислотн.
103 Ab 17 VH CDR 1 аминокислотн.
104 Ab 17 VH CDR 2 аминокислотн.
105 Ab 17 VH CDR 3 аминокислотн.
106 Ab 17 VL нуклеотидн.
107 Ab 17 VL аминокислотн.
108 Ab 17 VL CDR 1 аминокислотн.
109 Ab 17 VL CDR 2 аминокислотн.
110 Ab 17 VL CDR 3 аминокислотн.
111 Антитело 18 VH нуклеотидн.
112 Ab 18 VH аминокислотн.
113 Ab 18 VH CDR 1 аминокислотн.
114 Ab 18 VH CDR 2 аминокислоты.
115 Ab 18 VH CDR 3 аминокислотн.
116 Ab 18 VL нуклеотидн.
117 Ab 18 VL аминокислотн.
118 Ab 18 VL CDR 1 аминокислотн.
119 Ab 18 VL CDR 2 аминокислотн.
120 Ab 18 VL CDR 3 аминокислотн.
121 Антитело 19 VH нуклеотидн.
122 Ab 19 VH аминокислотн.
123 Ab 19 VH CDR 1 аминокислотн.
124 Ab 19 VH CDR 2 аминокислотн.
125 Ab 19 VH CDR 3 аминокислотн.
126 Ab 19 VL нуклеотидн.
127 Ab 19 VL аминокислотн.
128 Ab 19 VL CDR 1 аминокислотн.
129 Ab 19 VL CDR 2 аминокислотн.
130 Ab 19 VL CDR 3 аминокислотн.
131 Антитело 21 VH нуклеотидн.
132 Ab 21 VH аминокислотн.
133 Ab 21 VH CDR 1 аминокислотн.
134 Ab 21 VH CDR 2 аминокислотн.
135 Ab 21 VH CDR 3 аминокислотн.
136 Ab 21 VL нуклеотидн.
137 Ab 21 VL аминокислотн.
138 Ab 21 VL CDR 1 аминокислотн.
139 Ab 21 VL CDR 2 аминокислотн.
140 Ab 21 VL CDR 3 аминокислотн.
141 Антитело 22 VH нуклеотидн.
142 Ab 22 VH аминокислотн.
143 Ab 22 VH CDR 1 аминокислотн.
144 Ab 22 VH CDR 2 аминокислотн.
145 Ab 22 VH CDR 3 аминокислотн.
146 Ab 22 VL нуклеотидн.
147 Ab 22 VL аминокислотн.
148 Ab 22 VL CDR 1 аминокислоты.
149 Ab 22 VL CDR 2 аминокислоты.
150 Ab 22 VL CDR 3 аминокислотн.
151 Антитело 23 VH нуклеотидн.
152 Ab 23 VH аминокислотн.
153 Ab 23 VH CDR 1 аминокислотн.
154 Ab 23 VH CDR 2 аминокислотн.
155 Ab 23 VH CDR 3 аминокислотн.
156 Ab 23 VL нуклеотидн.
157 Ab 23 VL аминокислотн.
158 Ab 23 VL CDR 1 аминокислотн.
159 Ab 23 VL CDR 2 аминокислотн.
160 Ab 23 VL CDR 3 аминокислотн.
161 Полноразмерный IL-6 человека аминокислотн.
162 HIS FLAG меченый IL-6 человека
163 Растворимый IL-6Ra (человека)
164 Трансмембранный IL-6Ra (человека)
165 Зрелый IL-6 человека аминокислотн.
166 др 130 человека
167 Зародышевый VH FR1
168 Зародышевый VH FR2
169 Зародышевый VH FR3
170 Зародышевый VH FR4
171 Зародышевый VL FR1
172 Зародышевый VL FR2
173 Зародышевый VL FR3
174 Зародышевый VL FR4
175 F102E мутантный IL-6
176 S204E мутантный IL-6
177 R207E мутантный IL-6
178 F106E мутантный IL-6
179 Q211A мутантный IL-6
180 R58E мутантный IL-6
181 E200W мутантный IL-6
182 R207L мутантный IL-6
183 праймер macIL6_5'NdeI
184 праймер macIL6_3'NheI
185 праймер hIL6_5'NdeI
Последовательности антител 7, 10, 17 и 18 оптимизированы по зародышевой линии.
Все ссылки, упомянутые в любом месте в этом описании, включая любые места выше, включены в это описание в виде ссылок во всей их полноте и для любых целей.
Источники информации






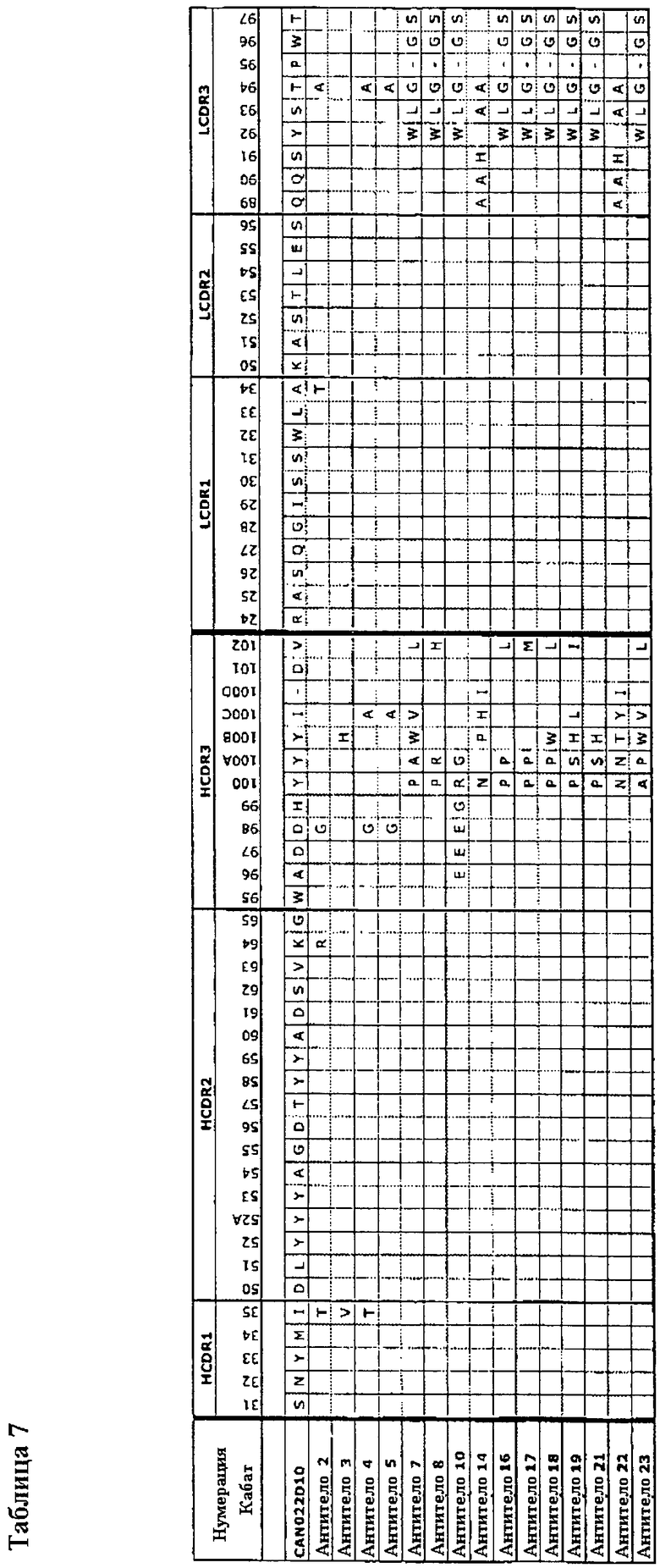
| название | год | авторы | номер документа |
|---|---|---|---|
| ЧЛЕНЫ СВЯЗЫВАНИЯ ПРОТИВ IL-1R1 | 2009 |
|
RU2555532C2 |
| МОЛЕКУЛЫ АНТИТЕЛ ЧЕЛОВЕКА К IL-13 | 2004 |
|
RU2387667C2 |
| НЕЙТРАЛИЗУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, НАПРАВЛЕННЫЕ ПРОТИВ ВИРУСА ГРИППА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2721706C2 |
| ЭЛЕМЕНТ, СВЯЗЫВАЮЩИЙСЯ С α-РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА-4 (IL-4Rα)-173 | 2008 |
|
RU2490278C2 |
| ИЗОЛИРОВАННОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, СПЕЦИФИЧЕСКОЕ(ИЙ) ДЛЯ ЧЕЛОВЕЧЕСКОГО ИММУНОГЛОБУЛИНА Е (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КЛЕТКА-ХОЗЯИН, СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ПРИМЕНЕНИЕ ЕЕ ДЛЯ ЛЕЧЕНИЯ РАССТРОЙСТВ, АССОЦИИРОВАННЫХ С IgE | 2008 |
|
RU2466140C2 |
| Антигены, ассоциированные с воспалительным заболеванием кишечника | 2012 |
|
RU2612878C2 |
| Антитело против IL-4R и его применение | 2018 |
|
RU2758091C1 |
| ПАРТНЕРЫ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С NGF | 2006 |
|
RU2406728C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ (ВАРИАНТЫ), СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ hTNFα С ИХ ПОСРЕДСТВОМ (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, РЕКОМБИНАНТНОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА (ВАРИАНТЫ) | 2005 |
|
RU2458704C9 |
| АНТИТЕЛА К БЕТА-АМИЛОИДУ | 2013 |
|
RU2651486C2 |
































































































































Настоящее изобретение относится к иммунологии и биотехнологии. Предложены варианты выделенной молекулы антитела для IL-6 человека, содержащие VH и VL; нуклеиновая кислота, включающая кодирующую антитело нуклеотидную последовательность; клетка для получения молекулы антитела, трансформированная указанной НК. Описан способ получения антител путем культивирования клетки; композиция и способ получения композиции молекулы антитела. Использование изобретения может найти применение в медицине в лечении заболеваний, связанных с IL-6. 11 н. и 23 з.п. ф-лы, 1 ил., 11 табл., 4 пр.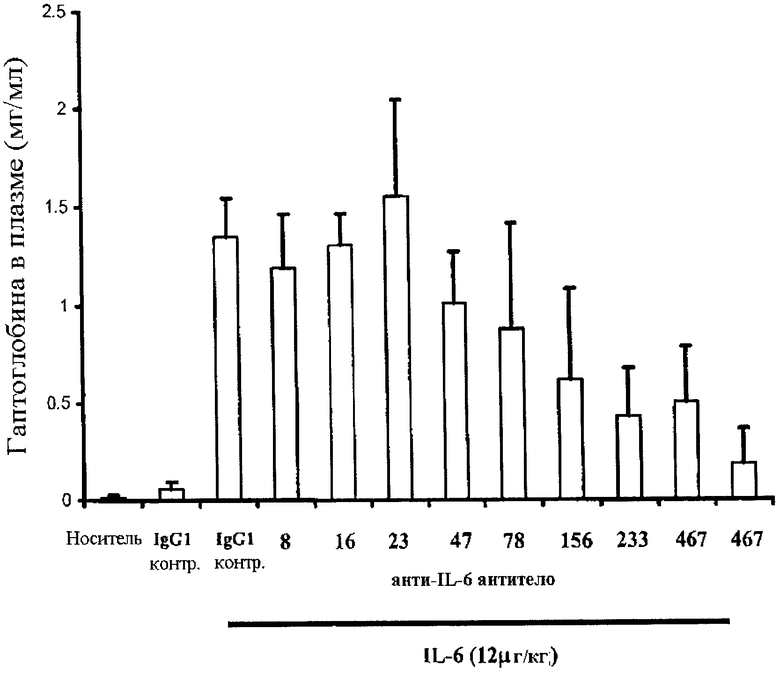
1. Выделенная молекула антитела для IL-6 человека, где указанное антитело включает вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), где VH и VL содержат 6 CDRs, выбранных из группы, включающей
(a) SEQ ID NO: 3; SEQ ID NO: 4; SEQ ID NO: 5; SEQ ID NO: 8; SEQ ID NO: 9 и SEQ ID NO: 10;
(b) SEQ ID NO: 13; SEQ ID NO: 14; SEQ ID NO: 15; SEQ ID NO: 18; SEQ ID NO: 19 и SEQ ID NO: 20;
(c) SEQ ID NO: 23; SEQ ID NO: 24; SEQ ID NO: 25; SEQ ID NO: 28; SEQ ID NO: 29 и SEQ ID NO: 30;
(d) SEQ ID NO: 33; SEQ ID NO: 34; SEQ ID NO: 35; SEQ ID NO: 38; SEQ ID NO; 39 и SEQ ID NO: 40;
(e) SEQ ID NO: 43; SEQ ID NO: 44; SEQ ID NO: 45; SEQ ID NO: 48; SEQ ID NO: 49 и SEQ ID NO: 50;
(f) SEQ ID NO: 53; SEQ ID NO: 54; SEQ ID NO: 55; SEQ ID NO: 58; SEQ ID NO: 59; SEQ ID NO: 60;
(g) SEQ ID NO: 63; SEQ ID NO: 64; SEQ ID NO: 65; SEQ ID NO: 68; SEQ ID.. NO: 69 и SEQ ID NO: 70;
(h) SEQ ID NO: 73; SEQ ID NO: 74; SEQ ID NO: 75; SEQ ID NO: 78; SEQ ID NO: 79 и SEQ ID NO: 80;
(i) SEQ ID NO: 83; SEQ ID NO: 84; SEQ ID NO: 85; SEQ ID NO: 88; SEQ ID NO: 89 и SEQ ID NO: 90;
(i) SEQ ID NO: 93; SEQ ID NO: 94; SEQ ID NO: 95; SEQ ID NO: 98; SEQ ID NO: 99 и SEQ ID NO: 100;
(k) SEQ ID NO: 103; SEQ ID NO: 104; SEQ ID NO: 105; SEQ ID NO: 108; SEQ ID NO: 109 и SEQ ID NO: 110;
(1) SEQ ID NO: 113; SEQ ID NO: 114; SEQ ID NO: 115; SEQ ID NO: 118; SEQ ID NO: 119 и SEQ ID NO: 120;
(m) SEQ ID NO: 123; SEQ ID NO: 124; SEQ ID NO: 125; SEQ ID NO: 128; SEQ ID NO: 129 и SEQ ID NO: 130;
(n) SEQ ID NO: 133; SEQ ID NO: 134; SEQ ID NO: 135; SEQ ID NO: 138; SEQ ID NO: 139 и SEQ ID NO: 140;
(о) SEQ ID NO: 143; SEQ ID NO: 144; SEQ ID NO: 145; SEQ ID NO: 148; SEQ ID NO: 149 и SEQ ID NO: 150;
(p) SEQ ID NO: 153; SEQ ID NO: 154; SEQ ID NO: 155; SEQ ID NO: 158; SEQ ID NO: 159 и SEQ ID NO: 160.
2. Выделенная молекула антитела для IL-6 человека, включающая набор CDRs;
HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в котором;
HCDR1 имеет аминокислотную последовательность SEQ ID NO: 3;
HCDR2 имеет аминокислотную последовательность SEQ ID NO: 4;
HCDR3 имеет аминокислотную последовательность SEQ ID NO: 115;
LCDR1 имеет аминокислотную последовательность SEQ ID NO: 8;
LCDR2 имеет аминокислотную последовательность SEQ ID NO: 9; и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 120.
3. Выделенная молекула антитела по п.1, где указанное антитело член имеет IC50 меньше, чем 1 нМ в анализе ингибирования выделения VEGF из синовиальных фибробластов, стимулированных при помощи 0,6 нМ IL-1β человека и 2,4 нМ растворимого IL-6Rα человека.
4. Выделенная молекула антитела по п.1, где указанное антитело связывает IL-6 макаки cynomolgus и где IL-6 макаки cynomolgus имеет IC50, отличающееся меньше, чем в 10 раз от IC50 немеченого IL-6 человека во флуоресцентном анализе с временным разрешением для ингибирования связывания меченого IL-6 человека с антителом, зафиксированным на подложке, причем указанный меченый IL-6 человека в конечной концентрации равен двойному значению Kd его взаимодействия со связывающим членом.
5. Выделенная молекула антитела по п.1, имеющий IC50 меньше, чем 100 нМ в анализе на ингибирование пролиферации клеток TF-1 в ответ на 20 нМ IL-6 человека.
6. Выделенная молекула антитела для IL-6 человека, включающая набор CDRs;
HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в которой:
HCDR1 имеет аминокислотную последовательность SEQ ID NO: 3;
HCDR2 имеет аминокислотную последовательность SEQ ID NO: 4;
HCDR3 имеет аминокислотную последовательность SEQ ID NO: 115;
LCDR1 имеет аминокислотную последовательность SEQ ID NO: 8;
LCDR2 имеет аминокислотную последовательность SEQ ID NO: 9; и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 120, и где указанное антитело имеет IC50 меньше, чем 100 нМ в анализе на ингибирование пролиферации клеток TF-1 в ответ на 20 нМ IL-6 человека.
7. Выделенная молекула антитела для IL-6 человека, включающая набор CDRs:
HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в котором:
HCDR1 представляет собой SEQ ID NO: 3;
HCDR2 представляет собой SEQ ID NO: 4;
HCDR3 представляет собой SEQ ID NO: 115;
LCDR1 представляет собой SEQ ID NO: 8;
LCDR2 представляет собой SEQ ID NO: 9; и
LCDR3 представляет собой SEQ ID NO: 80,
8. Выделенная молекула антитела по п.6, где указанная молекула антитела включает VH домен антитела и VL домен антитела, причем молекула антитела включает указанный ряд CDRs, где указанный VH домен включает HCDR1, HCDR2, HCDR3 и каркас, и указанный VL домен включает LCDR1, LCDR2, LCDR3 и каркас.
9. Выделенная молекула антитела по п.8, в которой указанная молекула антитела включает постоянную область антитела.
10. Выделенная молекула антитела по п.9, в которой указанная молекула антитела представляет собой IgG1.
11. Выделенная молекула антитела по п.8, в которой области каркаса VH и/или VL домена оптимизированы по зародышевой линии сегментных последовательностей гена зародышевой линии человека.
12. Выделенная молекула антитела по п.11, в которой VH домен антитела включает каркасные области, оптимизированные по зародышевой линии каркаса Vh3_DP-86_(3-66) зародышевой линии человека.
13. Выделенная молекула антитела по п.12, в которой VH домен включает каркасные области FR1, FR2, FR3 и FR4, имеющие аминокислотные последовательности, в которых FR1 представляет собой SEQ ID NO: 167, FR2 представляет собой SEQ ID NO: 168, FR3 представляет собой SEQ ID NO: 169 и FR4 представляет собой SEQ ID NO: 170.
14. Выделенная молекула антитела по п.13, в которой VH домен антитела имеет аминокислотную последовательность VH домена, показанную в SEQ ID NO: 112.
15. Выделенная молекула антитела по п.11, в которой VL домен антитела включает каркасные области, оптимизированные по зародышевой линии каркаса Vk1_L12 зародышевой линии человека.
16. Выделенная молекула антитела по п.15, в которой VL домен включает каркасные области FR1, FR2, FR3 и FR4, имеющие аминокислотные последовательности, в которых FR1 представляет собой SEQ ID NO: 171,FR2 представляет собой SEQ ID NO: 172, FR3 представляет собой SEQ ID NO: 173 и FR4 представляет собой SEQ ID NO: 174.
17. Выделенная молекула антитела по п.16, в котором VL домен антитела имеет аминокислотную последовательность VL домена, показанную в SEQ ID NO:117.
18. Выделенная молекула антитела, связывающая IL-6 человека или яванского макака, включающая тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 112 и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 117.
19. Молекула антитела по п.18, где указанная молекула антитела представляет собой IgG.
20. Молекула антитела по п.19, где IgG представляет собой IgGl.
21. Композиция, содержащая выделенную молекулу антитела по любому из пп.1-17, или молекулу антитела по любому из пп.18-20, и фармацевтически приемлемый наполнитель в количестве, эффективном для применения для лечения болезни или расстройств, относящихся к IL-6.
22. Композиция по п.21, где расстройство представляет собой воспалительное и/или аутоиммунное заболевание.
23. Композиция по п.22, где расстройство представляет собой ревматоидный артрит, остеоартрит, кахексию, хроническое обструктивное заболевание легких, идиопатический ювенильный артрит, астму, системную красную волчанку, воспалительную болезнь кишечника, болезнь Крона или атеросклероз.
24. Композиция по п.22, где расстройство представляет собой ревматоидный артрит.
25. Композиция по п.21, где расстройство представляет собой опухоль и/или рак.
26. Способ лечения расстройства, связанного с IL-6 у индивидуума, включающий введение индивидууму молекулы антитела по любому из пп.1-17 или по любому из пп.18-20.
27. Способ по п.26, в котором расстройство представляет собой воспалительное и/или аутоиммунное заболевание.
28. Способ по п.27, в котором расстройство представляет собой ревматоидный артрит, остеоартрит, кахексию, хроническое обструктивное заболевание легких, идиопатический ювенильный артрит, астму, системную красную волчанку, воспалительную болезнь кишечника, болезнь Крона или атеросклероз.
29. Способ по п.28, в котором расстройство представляет собой ревматоидный артрит.
30. Способ по п.26, в котором расстройство представляет собой опухоль и/или рак.
31. Выделенная молекула нуклеиновой кислоты, включающая нуклеотидную последовательность, кодирующую молекулу антитела по любому из пп.1-17 или по любому из пп.18-20.
32. Клетка-хозяин, трансформированная in vitro при помощи нуклеиновой кислоты по п.31, для получения молекулы антитела по любому из пп.1-17 или пп.18-20.
33. Способ продуцирования молекулы антитела по любому из пп.1-20 или VH или VL домена антитела по любому из пп.1-20, включающий культивирование клеток-хозяина по п.32.
34. Способ получения композиции молекулы антитела, включающий получение молекулы антитела, применяя способ по п.33, и приготовление молекулы антитела в виде композиции, содержащей по крайней мере один дополнительный компонент.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Приспособление для перемешивания горючей смеси в двигателях | 1926 |
|
SU4867A1 |
| СИНГЕР М | |||
| и др | |||
| Гены и Геномы, пер | |||
| с англ | |||
| - М.: Мир, 1998, с.63, т.1. | |||

