Настоящее изобретение относится к партнерам специфического связывания, в частности, с молекулами антител к NGF, в особенности с молекулами антител человека, и в особенности с теми, которые нейтрализуют активность NGF (фактор роста нервов). Оно далее относится к способам применения молекул антител к NGF в диагностике или лечении расстройств, связанных с NGF, включая боль, астму, хроническое обструктивное заболевание легких, фиброз легких, другие воспалительные заболевания дыхательных путей, диабетическую невропатию, сердечные аритмии, HIV, артрит, псориаз и рак.
Настоящее изобретение относится к молекулам антител, особенно ценных для связывания и нейтрализации NGF, и, таким образом, для применения в любом из множества терапевтических воздействий, как показано в содержащемся здесь экспериментальном материале и далее во вспомогательной технической литературе.
Фактор роста нервов (β-NGF, общеизвестный как NGF) играет хорошо известную ключевую роль в развитии нервной системы. У взрослых, однако, NGF играет более ограниченную роль, где он поддерживает здоровое состояние и выживаемость подмножества центральных и периферических нейронов (Huang & Reichardt, 2001). NGF также участвует в модулировании функциональных характеристик данных нейронов. Как часть этих последних процессов, NGF влияет на управление тонусом через чувствительность, или возбудимость, ноцицепторов (Priestley et al., 2002; Bennett, 2001). Данные периферические нейроны воспринимают и передают в центральную нервную систему разнообразные патологические стимулы, которые, в конечном итоге, вызывают ощущения боли (ноцицепция). Таким образом, средства, которые снижают уровни NGF, могут являться полезными в качестве обезболивающих лекарственных средств.
Социальные издержки неэффективного лечения боли дополнительно способствуют потенциальной полезности анальгетиков, основанных на действии против NGF. То есть, несмотря на существование и широко распространенное использование многочисленных обезболивающих лекарственных средств, существует отчетливая потребность в новых анальгетиках. Боль представляет собой один из наиболее общих симптомов, которые требуют медицинской помощи, и является основной жалобой половины из всех пациентов, посещающих врача. Высокие издержки боли для общества хорошо документированы. В США, например, хроническая боль причиняет страдание приблизительно 34 миллионам американцев. Боль приводит к потере 50 миллионов рабочих дней каждый год. Непосредственная медицинская стоимость, относящаяся только к боли в спине, боли в суставах и мигрени, насчитывает 40 миллиардов долларов ежегодно. Общая стоимость прописанных средств от боли на рынке лекарственных средств составляет приблизительно 15 миллиардов долларов в год (Pleuvry & Pleuvry).
Как следует из этих статистических данных, значительной процентной доле страдающих от боли не удается получить эффективное облегчение боли. Как следствие, остается значительная медицинская потребность в безопасных и эффективных анальгетиках с новыми механизмами действия (Pleuvry & Pleuvry).
Терапевтические средства, которые уменьшают тканевые уровни или ингибируют эффекты секретируемого NGF, обладают возможностью стать как раз такими новыми анальгетиками. Подкожные инъекции самого NGF вызывают боль у людей и животных. Таким образом, инъецированный NGF вызывает быструю тепловую гипералгезию, за которой следует отсроченная тепловая гипералгезия и механическая аллодиния (Petty et al., 1994; McArthur et al., 2000). Эндогенно секретируемый NGF сходным образом является проноцицептивным. Высвобождение NGF, вызываемое повреждением тканей, и его последующее действие на периферии играет основную роль в вызывании тепловой гипералгезии посредством процесса «периферической сенситизации» (Mendell & Arvanian, 2002). Повреждение тканей стимулирует высвобождение проноцицептивных и провоспалительных цитокинов, которые, в свою очередь, вызывают высвобождение NGF из кератиноцитов и фибробластов. Этот высвобожденный NGF действует непосредственно на ноцицепторы, вызывая болевые или ноцицептивные состояния в пределах минут патологического воздействия. Этот NGF также действует опосредованно, вызывая и поддерживая ноцицептивные/болевые состояния. Он запускает высвобождение гранул из тучной клетки, высвобождая проноцицептивные вещества, такие как гистамин и серотонин и, важно, дополнительное количество NGF, и может также стимулировать окончания симпатических нервов, высвобождая проноцицептивные нейротрансмиттеры, такие как норадреналин (Ma & Woolf, 1997).
Тканевые уровни NGF повышены у животных, инъецированных CFA и каррагинаном (Ma & Woolf, 1997; Amann & Schuligoi, 2000). Более того, повышение уровней NGF регистрировали у пациентов, страдающих ревматоидным артритом (Aloe & Tuveri, 1997) или циститом (Lowe et al., 1997). У грызунов повреждение периферического нерва повышает экспрессию мРНК NGF в макрофагах, фибробластах и шванновских клетках (Heumann et al., 1987). Сверхэкспрессия NGF у трансгенных мышей приводит к усилению невропатического болевого поведения вслед за повреждением нерва в большей степени, чем у мышей дикого типа (Ramer et al., 1998). В течение часов и суток повышенные уровни NGF играют роль в стимулировании «центральной сенситизации» - ускорении нейропередачи в синапсах в ноцицептивных путях спинного мозга. Центральная сенситизация приводит к персистирующей и хронической гипералгезии и аллодинии. В данный процесс, как предполагают, вовлечена интернализация комплексов NGF и его высокоаффинного рецептора, trkA (рецепторная тирозинкиназа A). Ретроградный транспорт данных комплексов к телам ноцицепторных клеток в ганглиях спинных корешков (DRG) усиливает секрецию ноцицептивных нейропептидов (например, вещества P, CGRP), активацию PKC и активацию рецептора NMDA в спинном роге спинного мозга (Sah et al., 2003) - все процессы, которые стимулируют сенситизацию ноцицептивных путей. NGF также играет роль в положительной регуляции и перераспределении потенциал-зависимых и управляемых лигандами ионных каналов, включая подтипы натриевых каналов, и рецептора капсаицина, VR1 (Mamet et al., 1999; Fjell et al., 1999; Priestley et al., 2002). Измененные активности и/или экспрессия трансмиттеров, рецепторов и ионных каналов лежат в основе повышенной чувствительности и возбудимости ноцицепторов, ассоциированных с состояниями невропатической боли.
NGF может также стимулировать спраутинг у симпатических нейронов и формирование аберрантной иннервации ноцицептивных нейронов. Эта иннервация, как предполагают, дает вклад в вызывание и поддержание хронических ноцицептивных/болевых состояний, таких как симпатически поддерживаемая боль или комплексный региональный болевой синдром (Ramer et al., 1999).
Ноцицепция/боль, индуцированная NGF, опосредована высокоаффинным рецептором NGF, trkA (рецепторной тирозинкиназой A) (Sah, et al., 2003). Приблизительно 40-45% тел клеток ноцицепторов в DRG экспрессируют trkA. Эти тела являются телами клеток волокон малого диаметра, или C-волокон, которые также экспрессируют секретируемые проноцицептивные пептиды, вещество P и CGRP. Данные волокна оканчиваются в слоях I и II спинного рога, где они передают в центральную нервную систему патологические стимулы, воспринимаемые периферическими ноцицепторами. Мутации или делеции в гене trkA приводят к фенотипу, характеризуемому утратой болевой чувствительности как у человека (Indo, 2002), так и у мышей с нокаутом trkA (de Castro et al., 1998). Существенно, что экспрессия trkA положительно регулируется у животных, являющихся моделями для боли при артрите (Pozza et al., 2000) или цистите (Qiao & Vizzard, 2002), или боли при воспалении, вызываемом инъекцией полного адъюванта Фрейнда (CFA) или каррагинана в конечность (Cho et al., 1996).
NGF также связывается с нейротрофиновым рецептором p75. Роль рецептора p75 зависит от клеточного окружения и наличия других рецепторов, с которыми, как предполагают, он выполняет вспомогательную или корецепторную функцию. Взаимодействие между рецепторами trkA и p75 приводит к формированию высокоаффинных участков связывания с NGF. Значение таких взаимодействий между рецепторами при болевой передаче сигнала, опосредованной NGF, не ясно, но согласно последним исследованиям рецептор p75 вовлечен в клеточные процессы, которые могут иметь значение (Zhang & Nicol, 2004). Однако, хотя мыши с нокаутом рецептора p75 демонстрируют повышение порогов к патологическим стимулам, они остаются восприимчивыми к гипералгезическим эффектам NGF, позволяя предположить, что одного рецептора trkA достаточно для опосредования данных эффектов (Bergmann et al., 1998).
Данные, цитированные выше, указывают, что процессы, опосредованные NGF, отвечают за вызывание острой боли, кратковременной боли, персистирующей ноцицептивной боли и персистирующей или хронической невропатической боли. Таким образом, показано, что средства против NGF полезны в качестве эффективных анальгетиков для лечения страдающих любым или всеми данными различными болевыми состояниями.
Одним из таких средств против NGF является trkA-Fc, который действует как ловушка или акцептор для связывания и, таким образом, инактивации эндогенного NGF. TrkA-Fc представляет собой слитый белок, состоящий из NGF-связывающей области trkA, связанной с константным доменным фрагментом (Fc) антител IgG. TrkA-Fc вызывает гипоалгезию у не подвергавшихся воздействию животных, снижает реакции ноцицептора и снижает спраутинг у немиелинизированных воспринимающих боль нейронов (Bennett et al., 1998).
Антисыворотки, полученные против NGF, могут также снижать уровни NGF при местной или системной инъекции. Как антисыворотки к NGF, так и trkA-Fc ослабляют воспалительную боль в конечности, вызываемую каррагинаном или CFA (Koltzenberg et al., 1999), и реакции при воспалении мочевого пузыря у крыс (Jaggar et al., 1999). Антисыворотка к NGF блокирует тепловую и холодовую гипералгезии, устраняет установившуюся тепловую гипералгезию и предотвращает коллатеральный спраутинг на модели невропатической боли при хроническом повреждении с помощью перетяжки (CCI) (Woolf, 1996; Ro et al., 1999). Также сообщалось о низкомолекулярных ингибиторах взаимодействия trkA-NGF. У крыс ингибитор NGF-trkA ALE-0540 снижает гипералгезию на модели термически индуцированной воспалительной боли и в формалиновом тесте острой и персистирующей боли (Owolabi et al., 1999). ALE-0540 также уменьшает механическую аллодинию на модели невропатической боли при повреждении седалищного нерва (Owolabi et al., 1999).
Терапевтические антитела, как правило, дают возможность степени избирательности к мишеням внутри семейства близкородственных рецепторов, лигандов рецепторов, каналов или ферментов, которая редко достижима с помощью низкомолекулярных лекарственных средств. Боль, опосредованная NGF, в частности, хорошо подходит для безопасного и эффективного лечения антителами, так как уровни NGF возрастают на периферии в ответ на патологические стимулы, и антитела обладают низкой проходимостью через гематоэнцефалический барьер. В то время как поликлональные антитела, как показано, являются эффективными на моделях боли у животных, моноклональные антитела к NGF наиболее перспективны для успешной разработки в качестве лекарственных средств у человека благодаря достижениям в получении и характеризации однородного, четко определенного химического вещества. Сообщали об антиноцицептивных эффектах мышиных моноклональных антител к NGF (Sammons et al., 2000), но не предоставили аминокислотные последовательности данных антител.
Последние данные позволяют предположить, что NGF стимулирует другие патологические состояния помимо боли. Таким образом, антитела к NGF могут также являться полезными для лечения других заболеваний, опосредованных NGF, включая в качестве неограничивающих примеров астму, хроническое обструктивное заболевание легких, фиброз легких, другие воспалительные заболевания дыхательных путей (Hoyle, 2003; Lommatzch et al., 2003), диабетическую невропатию (Yasuda et al., 2003), сердечные аритмии (WO 04/032852), HIV (Garaci et al., 2003), артрит, псориаз и рак (Nakagawara, 2001).
WO 02/096458 относится к антителам к NGF, в частности мышиным моноклональным антителам 911, и применению таких антител в лечении различных расстройств, связанных с NGF, включая боль, астму, артрит и псориаз. В нем утверждают, что антитела 911 не обладают побочным эффектом на иммунную систему на экспериментальной модели аллергии у мыши. Данные антитела также описаны Hongo et al., 2000.
В WO 04/032870 описано болеутоляющее действие мышиных моноклональных антител к NGF mab 911 и гуманизированных антител к NGF E3 на экспериментальных моделях послеоперационной боли. E3 отличается от константной области тяжелой цепи gamma2a человека на 2 аминокислоты.
В WO 04/032852 описаны способы профилактики внезапной сердечной смерти и лечения сердечных аритмий с использованием антагонистов к NGF.
В WO 01/78698 описано применение поликлональной антисыворотки к NGF для лечения хронической висцеральной боли.
Настоящее изобретение относится к партнерам, специфически связывающимся с NGF, предпочтительно с NGF человека. Таким образом, партнер специфического связывания по изобретению может связывать NGF человека или NGF, не относящийся к человеку (например, NGF не являющегося человеком примата и/или NGF крысы, и/или NGF мыши).
Партнерами специфического связывания по изобретению могут являться антитела к NGF человека, в особенности человеческие антитела, которые могут перекрестно связываться с NGF, не относящимся к человеку, включая NGF не являющегося человеком примата и/или NGF мыши, и/или NGF крысы.
Партнер специфического связывания согласно настоящему изобретению предпочтительно нейтрализует NGF. Нейтрализация означает снижение или ингибирование биологической активности NGF, например уменьшение или ингибирование связывания NGF с одним или более рецепторами (предпочтительно TrkA). Снижение биологической активности может являться частичным или полным. Степень, с которой антитела нейтрализуют NGF, обозначают как их нейтрализующую способность. Способность можно определить или измерить с использованием одного или более анализов, известных специалистам, и/или, как описано или приведено здесь, например:
- анализ мобилизации кальция «FLIPR» (см. здесь пример 2),
- анализ выживаемости PC12 (см. здесь пример 5),
- анализ пролиферации TF-1 (см. здесь пример 6),
- анализ ингибирования связывания с рецептором (см. здесь пример 9).
Анализы и характеристики описаны более подробно здесь в других местах.
Партнеров специфического связывания по настоящему изобретению можно оптимизировать по нейтрализующей способности. Как правило, оптимизация способности предусматривает мутацию последовательности выбранного партнера специфического связывания (обычно последовательности вариабельного домена антитела) для получения библиотеки партнеров специфического связывания, которую затем анализируют на способность и отбирают более способных партнеров специфического связывания. Таким образом, отобранные «оптимизированные по способности» партнеры специфического связывания обладают тенденцией к наличию лучшей способности, чем партнер специфического связывания, на основе которого получили библиотеку. Тем не менее, партнеров специфического связывания с хорошей способностью можно также получить без оптимизации, например, партнера специфического связывания с хорошей способностью можно получить непосредственно из начального скрининга, например, анализом на биохимическую нейтрализацию. Настоящее изобретение относится партнерам специфического связывания, как с оптимизированной способностью, так и неоптимизированным, а также к способам оптимизации способности для отобранного партнера специфического связывания. Настоящее изобретение, таким образом, позволяет специалисту получить партнеров специфического связывания, обладающих хорошей эффективностью.
Партнер специфического связывания согласно настоящему изобретению предпочтительно обладает антигипералгезивной и/или антиаллодинической активностью, например ингибирует тепловую гипералгезию, индуцированную каррагинаном.
В некоторых вариантах осуществления партнер специфического связывания по изобретению содержит молекулу антитела. В других вариантах осуществления партнер специфического связывания по изобретению содержит антигенсвязывающий участок в молекуле, не являющейся антителом, например набор CDR в белковом каркасе, не являющемся антителом, как дополнительно описано ниже.
В различных аспектах и вариантах осуществления изобретения представлено содержание формулы изобретения, включенной ниже.
Предпочтительными вариантами осуществления в настоящем изобретении являются молекулы антител или целые антитела (например, IgG, такой как IgG4), или фрагменты антител (например, scFv, Fab, dAb). Предпочтительно молекула антитела по изобретению является молекулой антитела человека. Предоставлены молекулы антител, содержащие антигенсвязывающие участки антител, такие как VH- и VL-домены антител. Внутри VH- и VL-доменов предоставлены области, определяющие комплементарность («CDR»), и каркасные области («FR»), для формирования VH- или VL-доменов в зависимости от обстоятельств. Антигенсвязывающий участок антител может состоять из VH-домена и/или VL-домена антител. Все VH- и VL-последовательности, CDR-последовательности, наборы CDR и наборы HCDR, и наборы LCDR, описанные здесь, представляют аспекты и варианты осуществления изобретения. «Набор CDR» включает CDR1, CDR2 и CDR3. Таким образом, набор HCDR означает HCDR1, HCDR2 и HCDR3, и набор LCDR означает LCDR1, LCDR2 и LCDR3. Если не утверждается обратное, «набор CDR» включает HCDR и LCDR.
Примеры VH- и VL-доменов антител и CDR согласно настоящему изобретению перечислены в приложенном списке последовательностей.
Здесь описан ряд линий антител, определенных со ссылкой на последовательности, например набор последовательностей CDR, не обязательно, с одной или более, например одной или двумя, или двумя заменами. Предпочтительной родительской линией является линия 1021E5. Линия 1021E5 включает предпочтительную молекулу антитела 1133C11 и другие молекулы антител «линии 1133C11», включая 1252A5. Также к родительской линии 1021E5 относятся молекулы антител 1165D4, 1230H7 и 1152H5. Авторы настоящего изобретения идентифицировали линии 1021E5, 1083H4 и, в особенности, 1133C11, как предоставляющие антигенсвязывающие участки человеческих антител к NGF, которые являются особенно ценными.
Линию 1133C11 определяют ссылкой на набор из шести последовательностей CDR из 1133C11, как указано далее: HCDR1 SEQ ID NO:193, HCDR2 SEQ ID NO:194, HCDR3 SEQ ID NO:195, LCDR1 SEQ ID NO:198, LCDR2 SEQ ID NO:199 и LCDR3 SEQ ID NO:200. Набор CDR, в котором HCDR1 обладает аминокислотной последовательностью SEQ ID NO:193, HCDR2 обладает аминокислотной последовательностью SEQ ID NO:194, HCDR3 обладает аминокислотной последовательностью SEQ ID NO:195, LCDR1 обладает аминокислотной последовательностью SEQ ID NO:198, LCDR2 обладает аминокислотной последовательностью SEQ ID NO:199 и LCDR3 обладает аминокислотной последовательность SEQ ID NO:200, здесь обозначен как «набор CDR 1133C11». HCDR1, HCDR2 и HCDR3 в наборе CDR 1133C11 обозначены как «набор HCDR 1133C11», и LCDR1, LCDR2 и LCDR3 в наборе CDR 1133C11 обозначены как «набор LCDR 1133C11». Набор CDR с набором CDR 1133C11, набором HCDR 1133C11 или LCDR 1133C11, или одной или двумя заменами в них, как говорят, является линией 1133C11.
Другие предпочтительные линии и наборы CDR определены ссылкой на аналогичные CDR, как представлено где-либо здесь, включая, как предпочтительные варианты осуществления, наборы CDR, описанные в таблице 2a (с SEQ ID NO, как представлено в таблице 2b). На таблице 2a и таблице 2b показаны наборы CDR (HCDR и LCDR) из оптимизированных клонов, полученных из клона 1021E5, поясняя, как последовательности CDR оптимизированных клонов отличаются от последовательностей 1021E5. Набор CDR из таблицы 2a/2b включает набор HCDR и/или набор LCDR из любого клона, представленного в таблице, не обязательно, включая сам 1021E5.
Данные наборы CDR предоставлены, как указано, как наборы CDR с описанными последовательностями, содержащими одну или две аминокислотные замены.
Настоящее изобретение также относится к партнерам специфического связывания и молекулам антител, содержащим определенные наборы CDR, набор HCDR или набор LCDR, как здесь описано, и наборы CDR с одной или двумя заменами в описанном наборе CDR. Соответствующий набор CDR предоставлен внутри каркаса антител или другого белкового каркаса, например фибронектина или цитохрома B (Koide et al., 1998; Nygren et al., 1997), как обсуждено ниже. Предпочтительно применяют каркасные области антител. Например, одну или более CDR или набор CDR антител можно встроить в каркас (например, каркас человека) для получения молекулы антитела или разных молекул антител. Например, молекула антитела может содержать CDR антитела линии 1021E5 и каркасные области последовательностей фрагмента гена зародышевой линии человека. Антитела линии можно предоставить с набором CDR внутри каркаса, который можно подвергнуть «превращению в зародышевую линию», где один или более остатков в каркасе изменяют, чтобы соответствовать остаткам в эквивалентном положении в наиболее сходном каркасе зародышевой линии человека (например, DP10 из семейства VH1) или каркасе семейства λ1, например DPL5. Таким образом, каркасные области антител происходят предпочтительно из зародышевой линии и/или человека.
Изобретение относится к выделенным антителам человека, специфичным к NGF, обладающим VH-доменом, содержащим набор HCDR в каркасе зародышевой линии человека, содержащей DP10. Обычно партнер специфического связывания также обладает VL-доменом, содержащим набор LCDR, предпочтительно в каркасе зародышевой линии человека, содержащей Vλ1, например DPL5. Предпочтительно CDR являются описанным здесь набором CDR.
«В основном, как представлено» означает, что соответствующая CDR или VH- или VL-домен по изобретению будут или идентичны или очень сходны с указанными областями, последовательность которых представлена здесь. «Очень сходны» предполагает, что можно осуществить аминокислотные замены от 1 до 5, предпочтительно от 1 до 4, такие как от 1 до 3 или 1 или 2, или 3, или 4 в CDR и/или VH- или VL-домене.
В одном из аспектов настоящее изобретение относится к партнеру специфического связывания с NGF, содержащему антигенсвязывающий участок антител, который сформирован из VH-домена антитела человека и VL-домена антитела человека и который содержит набор CDR, где VH-домен содержит HCDR1, HCDR2 и HCDR3, и VL-домен содержит LCDR1, LCDR2 и LCDR3, где HCDR1 обладает аминокислотной последовательностью SEQ ID NO:193, HCDR2 обладает аминокислотной последовательностью SEQ ID NO:194, HCDR3 обладает аминокислотной последовательностью SEQ ID NO:195, LCDR1 обладает аминокислотной последовательностью SEQ ID NO:198, LCDR2 обладает аминокислотной последовательностью SEQ ID NO:199, и LCDR3 обладает аминокислотной последовательностью SEQ ID NO:200; или где набор CDR содержит одну или две аминокислотные замены по сравнению с данным набором CDR.
Таким образом, изобретение относится к партнеру специфического связывания с NGF, содержащему антигенсвязывающий участок антител, который сформирован из VH-домена антитела человека и VL-домена антитела человека и который содержит набор CDR, где набор CDR является набором CDR 1133C11 или другим набором CDR, описанным здесь, или набором CDR, содержащим одну или две замены по сравнению с набором CDR 1133C11, или другим набором CDR, описанным здесь.
В предпочтительных вариантах осуществления одна или две замены являются одним или двумя из следующих остатков в CDR VH- и/или VL-доменов, с использованием общепринятой нумерации по Kabat (1991):
31, 34 в HCDR1;
51, 55, 56, 57, 58, 65 в HCDR2;
96 в HCDR3;
26, 27, 27A, 27B, 28, 29, 30 в LCDR1;
56 в LCDR2;
90, 94 в LCDR3.
В предпочтительных вариантах осуществления одну или две замены осуществляют посредством одного или двух из следующих остатков в наборе CDR 1133C11 согласно идентифицированным группам возможных замещающих остатков:
Остаток 29E в LCDR1 является особенно предпочтительным вариантом осуществления.
Предпочтительные варианты осуществления включают набор CDR 1133C11 или 1252A5, 1152H5, 1165D4, 1230H7 или 1021E5.
В одном из вариантов осуществления выделенный партнер специфического связывания содержит набор CDR, который содержит набор CDR 1133C11 с аминокислотной последовательностью FNSALIS (SEQ ID NO:532) или аминокислотной последовательностью MISSLQP (SEQ ID NO:533), замененными на аминокислотную последовательность LNPSLTA (SEQ ID NO:531) в HCDR3.
Любой набор HCDR линий, описанных здесь, можно предоставить в VH-домене, который можно применять в качестве партнера специфического связывания отдельно или в сочетании с VL-доменом. VH-домен можно предоставить с набором HCDR 1133C11, 1021E5 или антител других линий, например набором HCDR, как представлено в таблице 2a/2b, и если такой VH-домен находится в сочетании с VL-доменом, тогда VL-домен можно предоставить с набором LCDR 1133C11, 1021E5 или антител других линий, например набором LCDR, как представлено в таблице 2a/2b. Сочетание набора HCDR и набора LCDR может являться, как показано в таблице 2a/2b, давая антигенсвязывающий участок антител, содержащий набор CDR, как показано в таблице 2a/2b.
Каркасы VH- и VL-доменов содержат каркасные области, одна или более из которых могут являться каркасной областью зародышевой линии, обычно зародышевой линии человека. Каркас VH-домена является предпочтительно каркасом тяжелой цепи зародышевой линии человека, и каркас VL-домена является предпочтительно каркасом легкой цепи зародышевой линии человека. Каркасные области домена тяжелой цепи можно выбрать из семейства VH-1, и предпочтительным каркасом VH-1 является каркас DP-10. Каркасные области легкой цепи можно выбрать из семейства λ1, и предпочтительным каркасом является DPL5.
Одну или более CDR можно взять из VH- или VL-домена 1252A5 и вставить в подходящий каркас. Это обсуждено здесь далее. 1252A5 HCDR 1, 2 и 3 показаны на SEQ ID NO:393, 394, 395, соответственно. 1252A5 LCDR 1, 2 и 3 показаны на SEQ ID NO:398, 399, 400, соответственно.
Все это можно применить таким же образом к другим CDR и наборам CDR, как здесь описано, особенно к 1152H5, 1165D4 и 1230H7.
В вариантах осуществления по настоящему изобретению применяют VH- и/или VL-домен антител молекулы антитела линии 1021E5, например молекулы антитела 1021E5. Настоящее изобретение также относится к партнеру специфического связывания, содержащему антигенсвязывающий участок антител, содержащий такой VH- и/или VL-домен.
Предпочтительными вариантами осуществления являются следующие:
VH-домен, VL-домен, набор HCDR, набор LCDR или набор CDR: 112F1 (VH SEQ ID NO:102; VL SEQ ID NO:107), 1126G5 (VH SEQ ID NO:112; VL SEQ ID NO:117), 1126H5 (VH SEQ ID NO:122; VL SEQ ID NO:127), 1127D9 (VH SEQ ID NO:132; VL SEQ ID NO:137), 1127F9 (VH SEQ ID NO:142; VL SEQ ID NO:147), 1131D7 (VH SEQ ID NO:152; VL SEQ ID NO:157); 1131H2 (VH SEQ ID NO:162; VL SEQ ID NO:167), 1132A9 (VH SEQ ID NO:172; VL SEQ ID NO:177), 1132H9 (VH SEQ ID NO:182; VL SEQ ID NO:187), 1133C11 (VH SEQ ID NO:192; VL SEQ ID NO:197), 1134D9 (VH SEQ ID NO:202; VL SEQ ID NO:207), 1145D1 (VH SEQ ID NO:212; VL SEQ ID NO:217), 1146D7 (VH SEQ ID NO:222; VL SEQ ID NO:227), 1147D2 (VH SEQ ID NO:232; VL SEQ ID NO:237), 1147G9 (VH SEQ ID NO:242; VL SEQ ID NO:247), 1150F1 (VH SEQ ID NO:252; VL SEQ ID NO:257), 1152H5 (VH SEQ ID NO:262; VL SEQ ID NO:267), 1155H1 (VH SEQ ID NO:272; VL SEQ ID NO:277), 1158A1 (VH SEQ ID NO:282; VL SEQ ID NO:287), 1160E3 (VH SEQ ID NO:292; VL SEQ ID NO:297), 1165D4 (VH SEQ ID NO:302; VL SEQ ID NO:307), 1175H8 (VH SEQ ID NO:312; VL SEQ ID NO:317), 1211G10 (VH SEQ ID NO:322; VL SEQ ID NO:327), 1214A1 (VH SEQ ID NO:332; VL SEQ ID NO:337), 1214D10 (VH SEQ ID NO:342; VL SEQ ID NO:347), 1218H5 (VH SEQ ID NO:352; VL SEQ ID NO:357) и 1230Н7 (VH SEQ ID NO:362; VL SEQ ID NO:367).
Более того, предпочтительными являются VH-домен, VL-домен, набор HCDR, набор LCDR или набор CDR 1083H4 (VH SEQ ID NO:22; VL SEQ ID NO:27), 1227H8 (VH SEQ ID NO:372; VL SEQ ID NO:377) и 1230D8 (VH SEQ ID NO:382; VL SEQ ID NO:387).
В очень предпочтительном варианте осуществления VH-домен снабжен аминокислотной последовательностью SEQ ID NO:192; такой домен обозначен как «VH-домен 1133C11». В другом очень предпочтительном варианте осуществления VL-домен снабжен аминокислотной последовательность SEQ ID NO:197; такой домен обозначен как «VL-домен 1133C11». Очень предпочтительный антигенсвязывающий участок антител, предоставленный согласно настоящему изобретению, сформирован VH-доменом 1133C11, SEQ ID NO:192, и VL-доменом 1133C11, SEQ ID NO:197. Этот антигенсвязывающий участок антител можно предоставить с любым необходимым типом молекулы антитела, например scFv, Fab, IgG, IgG4 и т.п., как обсуждено далее в других местах данного описания.
В другом очень предпочтительном варианте осуществления VH-домен представлен аминокислотной последовательностью SEQ ID NO:392; такой домен обозначен как «VH-домен 1252A5». В другом очень предпочтительном варианте осуществления VL-домен представлен аминокислотной последовательностью SEQ ID NO:397, такой домен обозначен как «VL-домен 1252A5». Очень предпочтительный антигенсвязывающий участок антител, предоставленный согласно настоящему изобретению, сформирован VH-доменом 1252A5, SEQ ID NO:392, и VL-домен 1252A5, SEQ ID NO:397. Этот антигенсвязывающий участок антител можно предоставить с любым необходимым типом молекулы антитела, например scFv, Fab, IgG, IgG4 и т.п., как обсуждено далее в других местах данного описания.
В другом очень предпочтительном варианте осуществления настоящее изобретение относится молекуле антитела IgG4, содержащей VH-домен 1252A5, SEQ ID NO:392, и VL-домен 1252A5, SEQ ID NO:397. Данная молекула обозначена здесь как «IgG4 1252A5».
Другие IgG или другие молекулы антител, содержащие VH-домен 1252A5, SEQ ID NO:392, и/или VL-домен 1252A5, SEQ ID NO:397, относятся к настоящему изобретению, а также другие молекулы антител, содержащие набор HCDR 1252A5 (SEQ ID NO:393, 394 и 395) в VH-домене антитела, и/или набор LCDR 1252A5 (SEQ ID NO:398, 399 и 400) в VL-домене антитела.
Как указано, настоящее изобретение относится к партнеру специфического связывания, который связывается с NGF человека и который содержит VH-домен 1252A5 (SEQ ID NO:392) и/или VL-домен 1252A5 (SEQ ID NO:397). Здесь описаны свойства такого партнера специфического связывания.
Как правило, VH-домен находится в сочетании с VL-доменом для получения антигенсвязывающего участка антител, хотя, как обсуждено далее ниже, один VH-домен можно применять для связывания антигена. В одном из предпочтительных вариантов осуществления VH-домен 1252A5 (SEQ ID NO:392) находится в сочетании с VL-доменом 1252A5 (SEQ ID NO:397) так, что антигенсвязывающий участок антител сформирован содержащим как VH-, так VL-домены 1252A5. Аналогичные варианты осуществления предоставлены для других VH- и VL-доменов, описанных здесь. В других вариантах осуществления 1252A5 VH находится в сочетании с VL-доменом, отличным от VL 1252A5. Разнородность легких цепей твердо установлена в данной области. Более того, относятся к изобретению аналогичные варианты осуществления для других VH- и VL-доменов, описанных здесь.
Варианты VH- и VL-доменов и CDR по настоящему изобретению, включая те, для которых аминокислотные последовательности представлены здесь, и которые можно применять для партнеров специфического связывания с NGF, можно получить посредством способов изменения или мутации последовательности и скрининга. Такие способы также относятся к настоящему изобретению.
Согласно дополнительным аспектам настоящего изобретения предоставлен партнер специфического связывания, который конкурирует за связывание с антигеном с любым партнером специфического связывания, который как связывается с антигеном, так и содержит партнера специфического связывания, VH- и/или VL-домен, описанный здесь, или HCDR3, описанный здесь, или вариант любого из них. Конкуренцию между партнерами связывания можно легко анализировать in vitro, например, с использованием ELISA и/или посредством присоединения специфической репортерной молекулы к одному партнеру связывания, которого можно детектировать в присутствии одного или более других немеченых партнеров связывания, давая возможность идентификации партнеров специфического связывания, которые связываются с одинаковым эпитопом или с перекрывающимся эпитопом.
Таким образом, дополнительный аспект настоящего изобретения относится к партнеру специфического связывания, содержащему антигенсвязывающий участок антитела человека, который конкурирует с молекулой антитела, например, в особенности, с 1252A5 или другим предпочтительным scFv и/или IgG4 за связывание с NGF. В дополнительных аспектах настоящее изобретение относится к партнеру специфического связывания, содержащему антигенсвязывающий участок антитела человека, который конкурирует с антигенсвязывающим участком антител за связывание с NGF, где антигенсвязывающий участок антител сформирован VH-доменом и VL-доменом, и где VH- и VL-домены содержат набор CDR 1133C11, 1021E5, 1252A5 или другой линии, описанной здесь.
Различные способы доступны в данной области для получения антител к NGF, и которые могут конкурировать с 1252A5 или другой молекулой антитела, молекулой антитела с 1252A5 или другим набором CDR, или молекулой антитела с набором CDR 1252A5 или другой линии за связывание с NGF.
В дополнительном аспекте настоящее изобретение относится к способу получения одного или более партнеров специфического связывания, способных связываться с антигеном, где способ включает приведение в контакт библиотеки партнеров специфического связывания согласно изобретению и указанного антигена и отбор одного или более партнеров специфического связывания из библиотеки, способных связываться с указанным антигеном.
Библиотеку можно экспонировать на частицах или молекулярных комплексах, например реплицирующихся генетических структурах, таких как частицы из дрожжей, бактерий или бактериофагов (например, T7), или ковалентных, рибосомальных или других системах дисплеев in vitro; каждая частица или молекулярный комплекс содержит нуклеиновую кислоту, кодирующую вариабельный VH-домен антител, экспонированный на них, и, не обязательно, также экспонированный VL-домен в случае наличия.
После отбора партнеров специфического связывания, способных связываться с антигеном и экспонированных на бактериофаге или других частицах библиотеки, или молекулярных комплексах, нуклеиновую кислоту можно выделить из бактериофага или другой частицы, или молекулярного комплекса, экспонирующего указанного отобранного партнера специфического связывания. Такую нуклеиновую кислоту можно использовать в последующем получении партнера специфического связывания или вариабельного VH- или VL-домена антитела посредством экспрессии нуклеиновой кислоты с последовательностью нуклеиновой кислоты, выделенной из бактериофага или другой частицы, или молекулярного комплекса, экспонирующего указанного отобранного партнера специфического связывания.
Вариабельный VH-домен антител с аминокислотной последовательностью вариабельного VH-домена антител указанного отобранного партнера специфического связывания можно предоставить в выделенной форме, как можно предоставить партнера специфического связывания, содержащего такой VH-домен.
Способность связываться с NGF можно далее тестировать, а также способность конкурировать, например, с 1252A5 (например, в форме scFv и/или форме IgG, например, IgG4) за связывание с NGF. Способность нейтрализовать NGF можно тестировать, как обсуждено далее ниже.
Партнер специфического связывания согласно настоящему изобретению может связываться с NGF с аффинностью 1252A5 или другой молекулы антитела, например scFv, или предпочтительно 1252A5 или другого IgG4, или с лучшей аффинностью.
Партнер специфического связывания согласно настоящему изобретению может нейтрализовать NGF со способностью 1252A5 или другой молекулы антитела, например scFv, или предпочтительно 1252A5 или другого IgG4, или с лучшей способностью.
Аффинность связывания и нейтрализующую способность разных партнеров специфического связывания можно сравнивать в соответствующих условиях.
Антитела по настоящему изобретению обладают рядом преимуществ по сравнению с существующими коммерчески доступными антителами к NGF. Например, настоящее изобретение относится к антителам человека или зародышевой линии, которые, как ожидается, демонстрируют более низкую степень иммуногенности при хроническом или повторном введении человеку с терапевтической или диагностической целью. Далее, настоящее изобретение относится к антителам, которые обладают лучшей способностью нейтрализовать NGF, и, таким образом, желаемый терапевтический или диагностический эффект можно достичь с использованием меньшего количества антител. Кроме того, в одном из вариантов осуществления изобретения способность ингибировать взаимодействие NGF/рецептор trkA выше, чем та, которую наблюдают для ингибирования взаимодействия NGF/рецептор p75. Это может давать преимущества по сравнению с обработками с данной целью другими явно неселективными антагонистами к NGF, или в силе или природе достигаемого терапевтического эффекта, или в уменьшении нежелательных побочных эффектов.
Изобретение также относится к гетерогенным препаратам, содержащим молекулы антител к NGF. Например, такие препараты могут представлять собой смеси антител с полноразмерными тяжелыми цепями и тяжелыми цепями, у которых отсутствует C-концевой лизин, с различными степенями гликозилирования и/или с модифицированными аминокислотами, такими как циклизация N-концевой глутаминовой кислоты с формированием остатка пироглутаминовой кислоты.
В дополнительных аспектах изобретение относится к выделенной нуклеиновой кислоте, которая содержит последовательность, кодирующую партнера специфического связывания, VH-домен и/или VL-домен согласно настоящему изобретению, и способам получения партнера специфического связывания, VH-домена и/или VL-домена по изобретению, которые включают экспрессию указанной нуклеиновой кислоты в условиях для осуществления продукции указанного партнера специфического связывания, VH-домена и/или VL-домена, и его выделение.
Дополнительный аспект настоящего изобретения относится к нуклеиновой кислоте, как правило, выделенной, кодирующей вариабельный VH-домен антитела и/или вариабельный VL-домен антитела, описанные здесь.
Другой аспект настоящего изобретения относится к нуклеиновой кислоте, как правило, выделенной, кодирующей последовательность VH CDR или VL CDR, описанную здесь, в особенности, VH CDR, выбранную из: 1133C11 (VH CDR1 SEQ ID NO:193, VH CDR2 SEQ ID NO:194 и VH CDR3 SEQ ID NO:195), 1152H5 (VH CDR1 SEQ ID NO:263, VH CDR2 SEQ ID NO:264 и VH CDR3 SEQ ID NO:265) и 1252A5 (VH CDR1 SEQ ID NO:393, VH CDR2 SEQ ID NO:394 и VH CDR3 SEQ ID NO:395), или VL CDR, выбранную из: 1133C11 (VL CDR1 SEQ ID NO:198, VL CDR2 SEQ ID NO:199 и VL CDR3 SEQ ID NO:200), 1152H5 (VL CDR1 SEQ ID NO:268, VL CDR2 SEQ ID NO:269 и VL CDR3 SEQ ID NO:270) и 1252A5 (VL CDR1 SEQ ID NO:398, VL CDR2 SEQ ID NO:399 и VL CDR3 SEQ ID NO:400), наиболее предпочтительно 1252A5 (VH CDR3 SEQ ID NO:395). Нуклеиновая кислота, кодирующая набор CDR 1252A5, нуклеиновая кислота, кодирующая набор HCDR 1252A5, и нуклеиновая кислота, кодирующая набор LCDR 1252A5, также относятся к настоящему изобретению, как и нуклеиновые кислоты, кодирующие индивидуальные CDR, HCDR, LCDR и наборы CDR, HCDR, LCDR линий 1252A5, 1133C11 или 1021E5.
Дополнительный аспект относится к клетке-хозяину, трансформированной нуклеиновой кислотой по изобретению.
Другой дополнительный аспект относится к способу получения вариабельного VH-домена антител; где способ включает вызывание экспрессии кодирующей нуклеиновой кислоты. Такой способ может включать культивирование клеток-хозяев в условиях для продукции указанного вариабельного VH-домена антител.
Аналогичные способы получения вариабельных VL-доменов и партнеров специфического связывания, содержащих VH- и/или VL-домен, представлены как дополнительные аспекты настоящего изобретения.
Способ получения может включать стадию выделения и/или очистки продукта. Способ получения может включать составление продукта в композицию, содержащую, по меньшей мере, один дополнительный компонент, такой как фармацевтически приемлемый эксципиент.
Дополнительные аспекты настоящего изобретения относятся к композициям, содержащим партнеров специфического связывания по изобретению, и их применению в способах ингибирования или нейтрализации NGF, включая способы терапевтического лечения человека или животного.
Партнеров специфического связывания согласно изобретению можно применять в способе лечения или диагностики человека или животного, такого как способ лечения (который может включать профилактическое лечение) заболевания или расстройства у больного человека, который включает введение указанному пациенту эффективного количества партнера специфического связывания по изобретению. Состояния, поддающиеся лечению согласно настоящему изобретению, включают любое, в котором принимает участие NGF, в особенности, боль, астму, хроническое обструктивное заболевание легких, фиброз легких, другие воспалительные заболевания дыхательных путей, диабетическую невропатию, HIV, сердечные аритмии, артрит, псориаз и рак.
Эти и другие аспекты изобретения описаны более подробно ниже.
ТЕРМИНОЛОГИЯ
Удобно здесь отметить, что «и/или» при использовании здесь следует понимать как конкретное описание каждого из двух указанных признаков или компонентов с другим или без другого. Например, «A и/или B» следует понимать как конкретное описание каждого из (i) A, (ii) B и (iii) A и B, подобно тому, как каждый представлен здесь индивидуально.
NGF
NGF (также известный как beta-NGF) является фактором роста нервов. В контексте настоящего изобретения, NGF, как правило, представляет собой NGF человека, хотя он может являться NGF, не относящимся к человеку (например, NGF не являющегося человеком примата и/или NGF крысы, и/или NGF мыши). NGF также обозначают местами как «антиген».
NGF, используемый в анализе, описанном здесь, является, как правило, NGF человека, крысы или мыши, но можно использовать NGF другого, не являющегося человеком животного, например NGF не являющегося человеком примата.
Боль
Она характеризуется, как хорошо известно в данной области, ощущением боли и может включать одно или более, или все из следующего:
- гипералгезия (чрезмерная болевая реакция на нормальный болевой стимул);
- аллодиния (ощущение боли, вызываемое стимулом, который в норме не вызывает боль);
- спонтанное ощущение боли, вызываемое любым способом в отсутствие какого-либо явного внешнего влияния;
- боль, вызываемая физическими стимулами, такими как жара, тепло, холод, давление, вибрация, статическая или динамическая пальпация, или положение тела и движение;
- соматическая и висцеральная боль, вызываемая любым способом, например травмой, инфекцией, воспалением, метаболическим заболеванием, инсультом или неврологическим заболеванием.
Боль может являться, например, острой болью, кратковременной болью, персистирующей ноцицептивной болью или персистирующей, или хронической невропатической болью.
Партнер специфического связывания
Этот термин относится к компоненту пары молекул, которые обладают специфичностью связывания друг с другом. Компоненты специфически связывающейся пары могут обладать естественным происхождением, или их получают синтетически, полностью или частично. Один из компонентов пары молекул обладает областью на своей поверхности или углублением, которое специфически связывается, и, таким образом, комплементарно специфической пространственной и полярной организации другого компонента пары молекул. Таким образом, компоненты пары обладают свойством специфически связываться друг с другом. Примерами типов специфически связывающихся пар являются антиген-антитело, биотин-авидин, гормон-рецептор гормона, рецептор-лиганд, фермент-субстрат. Настоящее изобретение относится к типу взаимодействий антиген-антитело.
Партнер специфического связывания обычно содержит молекулу, обладающую антигенсвязывающим участком. Например, партнер специфического связывания может являться молекулой антитела или белком, не являющимся антителом, который содержит антигенсвязывающий участок. Антигенсвязывающий участок можно создать посредством размещения CDR на белковых каркасах, не являющихся антителами, таких как фибронектин или цитохром B и т.п. (Haan & Maggos, 2004; Koide et al., 1998; Nygren et al., 1997), или посредством рандомизации или мутагенеза аминокислотных остатков петли в белковом каркасе для придания специфичности связывания с требуемой мишенью. Каркасы для создания новых участков связывания в белках подробно рассмотрены в Nygren et al. (1997). Белковые каркасы для имитации антител описаны в WO 0034784, в котором авторы изобретения описывают белки (имитаторы антител), которые включают домен фибронектина III типа, обладающий, по меньшей мере, одной рандомизированной петлей. Подходящий каркас, в который встраивают одну или более CDR, например набор HCDR, можно получить из любого доменного члена суперсемейства генов иммуноглобулинов. Каркас может представлять собой белок, относящийся или не относящийся к человеку.
Преимущество белкового каркаса, не являющегося антителом, состоит в том, что он может обладать антигенсвязывающим участком в каркасной молекуле, который меньше и/или легче для получения, чем, по меньшей мере, некоторые молекулы антител. Небольшой размер партнера специфического связывания может придать полезные физиологические свойства, такие как способность проникновения в клетки, проникновения глубоко в ткани или достижения мишеней внутри других структур, или связывания внутри белковых углублений антигена-мишени.
Использование антигенсвязывающих участков в белковых каркасах, не являющихся антителами, рассмотрено в Wess, 2004. Обычно белки обладают неизменяемым остовом и одной или более вариабельными петлями, в которых аминокислотную последовательность петли или петель специфически или случайным образом мутагенизируют для создания антигенсвязывающего участка, обладающего специфичностью для связывания с антигеном-мишенью. Такие белки включают IgG-связывающие домены белка A из S. aureus, трансферрина, тетранектина, фибронектина (например, 10-й домен фибронектина III типа) и липокалинов. Другие способы включают синтетические «Microbodies» (Selecore GmbH), которые основаны на циклотидах - небольших белках, обладающих внутримолекулярными дисульфидными связями.
В дополнение к последовательностям антител и/или антигенсвязывающему участку, партнер специфического связывания согласно настоящему изобретению может содержать другие аминокислоты, например формирующие пептид или полипептид, такой как свернутый домен, или придающие молекуле другое функциональное свойство в дополнение к способности связываться с антигеном. Партнеры специфического связывания по изобретению могут нести детектируемую метку или их можно конъюгировать с токсином или направляющей группой, или ферментом (например, посредством пептидной связи или линкера). Например, партнер специфического связывания может содержать каталитический участок (например, в домене фермента), а также антигенсвязывающий участок, в котором антигенсвязывающий участок связывается с антигеном и, таким образом, нацеливает каталитический участок на антиген. Каталитический участок может ингибировать биологическую функцию антигена, например, посредством расщепления.
Хотя, как указано, каркасы, такие как фибронектин или цитохром B, могут нести CDR (Haan & Maggos, 2004; Koide et al., 1998; Nygren et al., 1997), структурой, содержащей CDR или набор CDR по изобретению, как правило, будет являться последовательность тяжелой или легкой цепи антител или существенная ее часть, в которой CDR или набор CDR размещены в положении, соответствующем CDR или набору CDR встречающихся в природе вариабельных доменов антител VH и VL, кодируемых перестроенными генами иммуноглобулинов. Структуры и местоположения вариабельных доменов иммуноглобулинов можно определить исходя из Kabat, et al., 1987, и ее обновлений, доступных в настоящее время в Интернет (http://immuno.bme.nwu.edu или найти «Kabat» с использованием любого средства поиска).
Молекула антитела
Ее описывают как иммуноглобулин, или природный, или частично, или полностью полученный синтетически. Термин также относится к любому полипептиду или белку, содержащему антигенсвязывающий участок антител. Фрагменты антител, которые содержат антигенсвязывающий участок антител, являются молекулами, такими как Fab, scFv, Fv, dAb, Fd и димерные антитела.
Возможно получение моноклональных и других антител и использование способов технологии рекомбинантных ДНК для получения других антител или химерных молекул, которые сохраняют специфичность исходных антител. Такие способы могут включать введение ДНК, кодирующую вариабельную область иммуноглобулина или CDR антител, в константные области или константные области плюс каркасные области другого иммуноглобулина. См., например, EP-A-184187, GB 2188638A или EP-A-239400, и значительное количество последующей литературы. Гибридому или другую клетку, вырабатывающую антитела, можно подвергнуть генетической мутации или другим изменениям, которые могут или не могут изменить специфичность связывания продуцированных антител.
Так как антитела можно модифицировать рядом способов, термин «молекула антитела» следует толковать, как относящийся к любому партнеру специфического связывания или веществу, обладающему антигенсвязывающим участком антител с необходимой специфичностью. Таким образом, данный термин относится к фрагментам антител и производным, включая любой полипептид, содержащий антигенсвязывающий участок антител, или природный, или полностью или частично синтетический. Химерные молекулы, содержащие антигенсвязывающий участок антител, или эквивалент, слитый с другим полипептидом, являются, таким образом, включенными. Клонирование и экспрессия химерных антител описаны в EP-A-0120694 и EP-A-0125023, и значительном количестве последующей литературы.
Другие способы, доступные в области технологии антител, сделали возможным получение человеческих и гуманизированных антител. Например, гибридомы человека можно получить, как описано в Kontermann & Dubel (2001). Фаговый дисплей, другой признанный способ получения партнеров специфического связывания, подробно описан во многих публикациях, таких как Kontermann & Dubel (2001) и WO 92/01047 (подробно обсуждены ниже). Для получения антител человека можно использовать трансгенных мышей, у которых гены мышиных антител инактивированы и функционально замещены генами антител человека, при этом оставляя нетронутыми другие компоненты иммунной системы мыши (Mendez et al., 1997).
Синтетические молекулы антител можно создать путем экспрессии генов, полученных посредством синтеза олигонуклеотидов и их сборки в подходящих экспрессирующих векторах, например, как описано в Knappik et al. (2000) или Krebs et al. (2001).
Показано, что фрагменты целого антитела могут выполнять функцию связывания с антигенами. Примерами связывающихся фрагментов являются (i) Fab-фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) Fd-фрагмент, состоящий из доменов VH и CH1; (iii) Fv-фрагмент, состоящий из доменов VL и VH одиночного антитела; (iv) dAb-фрагмент (Ward, 1989; McCafferty et al., 1990; Holt et al., 2003), который состоит из VH- или VL-домена; (v) изолированные области CDR; (vi) F(ab')2-фрагменты, двухвалентный фрагмент, содержащий два связанных Fab-фрагмента; (vii) одиночная цепь молекул Fv (scFv), в которой VH-домен и VL-домен связаны пептидным линкером, который позволяет двум доменам соединяться с образованием антигенсвязывающего участка (Bird et al., 1988; Huston et al., 1988); (viii) биспецифическая одиночная цепь димеров Fv (PCT/US92/09965); и (ix) «димерные антитела», поливалентные или полиспецифические фрагменты, сконструированные слиянием генов ( WO 94/13804; Holliger et al., 1993). Молекулы Fv, scFv или димерных антител можно стабилизировать включением дисульфидных мостиков, связывающих VH- и VL-домены (Reiter et al., 1996). Также, можно получить мини-антитела, содержащие scFv, присоединенный к CH3-домену (Hu et al., 1996).
dAb (доменное антитело) представляет собой небольшой мономерный антигенсвязывающий фрагмент антитела, а именно вариабельную область тяжелой или легкой цепи антитела (Holt et al., 2003). VH dAb образуется естественно у верблюжьих (например, верблюда, ламы), и его можно получить иммунизацией представителя верблюжьих антигеном-мишенью, выделением антигенспецифичных B-клеток и непосредственным клонированием генов dAb из индивидуальных B-клеток. dAb также получают в культуре клеток. Их небольшой размер, хорошая растворимость и температурная стабильность делают их особенно физиологически полезными и пригодными для селекции и аффинного созревания. Партнер специфического связывания по настоящему изобретению может являться dAb, содержащим VH- или VL-домен, в основном, как представлено здесь, или VH- или VL-домен, содержащий набор CDR, в основном, как представлено здесь.
В тех случаях, когда следует использовать биспецифические антитела, они могут представлять собой общепринятые биспецифические антитела, которые можно получать множеством способов (Holliger & Winter, 1993), например получать химически или из гибридных гибридом, или они могут являться любыми биспецифическими фрагментами антител, упомянутых выше. Примеры биспецифических антител включают антитела на основе технологии BiTETM, в которой можно использовать связывающие домены двух антител с разной специфичностью и непосредственно связывать их короткими гибкими пептидами. При этом объединяются два антитела на короткой одиночной полипептидной цепи. Димерные антитела и scFv можно конструировать без Fc-области, с использованием только вариабельных доменов, потенциально уменьшая эффекты антиидиотипической реакции.
Биспецифические димерные антитела, в противоположность биспецифическим целым антителам, могут также оказаться особенно полезными, так как их можно легко конструировать и экспрессировать в E.coli. Димерные антитела (и многие другие полипептиды, такие как фрагменты антител) подходящих специфичностей связывания можно легко отбирать с использованием фагового дисплея (WO 94/13804) из библиотек. Если одно плечо димерного антитела сохранять константным, например, со специфичностью, направленной против NGF, тогда можно получить библиотеку, в которой другое плечо вариабельно, и отбирать антитела подходящей специфичности. Биспецифические целые антитела можно получить способом knobs-into-holes (гребень/паз) (Ridgeway et al., 1996).
Антигенсвязывающий участок
Его описывают как часть молекулы, которая связывается и комплементарна всему антигену-мишени или его части. В молекуле антитела его обозначают как антигенсвязывающий участок антител, и он содержит часть антитела, которая специфически связывается и комплементарна всему антигену-мишени или его части. Если антиген большой, то антитела могут связываться только со специфической частью антигена, эту часть называют эпитопом. В создании антигенсвязывающего участка антител могут участвовать один или более вариабельных доменов антител. Предпочтительно антигенсвязывающий участок антител содержит вариабельную область легкой цепи антител (VL) и вариабельную область тяжелой цепи антител (VH).
Специфичный
Этот термин можно использовать для обозначения ситуации, в которой для одного компонента специфически связывающейся пары не будет показано какое-либо значительное связывание с молекулами, другими, чем его специфически связывающийся партнер(ы). Термин также применим там, где, например, антигенсвязывающий участок специфичен для конкретного эпитопа, который присутствует на ряде антигенов, в этом случае партнер специфического связывания, несущий антигенсвязывающий участок, будет способен связываться с различными антигенами, несущими эпитоп.
Выделенный
Этот термин относится к состоянию, в котором партнеры специфического связывания по изобретению или нуклеиновая кислота, кодирующая таких партнеров связывания, будут, как правило, находиться согласно настоящему изобретению. Выделенные партнеры и выделенная нуклеиновая кислота будут свободны или по существу свободны от материала, с которым они в естественных условиях ассоциированы, такого как другие полипептиды или нуклеиновые кислоты, с которым их находят в естественном окружении или окружении, в котором их получают (например, культура клеток), когда такое получение осуществляют технологией рекомбинантных ДНК, применяемой in vitro или in vivo. Партнеров и нуклеиновую кислоту можно смешать с разбавителями или адъювантами, и, тем не менее, для практических целей они являются выделенными - например, партнеров обычно будут смешивать с желатином или другими носителями при применении для покрытия планшетов для микротитрования для использования в иммунологических анализах, или будут смешивать с фармацевтически приемлемыми носителями или разбавителями при применении в диагностике или лечении. Партнеров специфического связывания можно гликозилировать, или естественным образом, или в системах гетерологичных эукариотических клеток (например, клетки CHO или NS0 (ECACC 85110503)), или они могут являться (например, при получении экспрессией в прокариотической клетке) негликозилированными.
ПОДРОБНОЕ ОПИСАНИЕ
Как указано выше, партнер специфического связывания согласно настоящему изобретению предпочтительно нейтрализует NGF. Степень, с которой антитело нейтрализует NGF, обозначают как нейтрализующую способность.
Способность обычно выражают как величину IC50, в нМ, если не утверждается обратное. IC50 представляет собой среднюю ингибирующую концентрацию молекулы антитела. В функциональных анализах IC50 представляет собой концентрацию, которая снижает биологический ответ на 50% от максимума. В исследованиях связывания лигандов IC50 представляет собой концентрацию, которая уменьшает связывание рецептора на 50% от максимального уровня специфического связывания.
IC50 можно вычислить построением % биологического ответа (представленного, например, мобилизацией ионов кальция в анализе FLIPR, выживаемостью в анализе PC12 или пролиферацией в анализе пролиферации TF-1) или % специфического связывания рецептора как функции от log концентрации партнера специфического связывания, и с использованием программного обеспечения, такого как Prism (GraphPad), построить сигмоидальную функцию исходя из данных для получения величин IC50, например, как описано здесь в примере 2, 5, 6 или 9.
Партнер специфического связывания согласно настоящему изобретению предпочтительно ингибирует внутриклеточную мобилизацию кальция, вызванную NGF человека, в клетках, экспрессирующих рецептор TrkA, например клетках, рекомбинантно трансфицированных геном TrkA, например клетках HEK. В анализе мобилизации кальция «FLIPR», как описано здесь в примере 2, партнер специфического связывания согласно изобретению предпочтительно обладает способностью (IC50) нейтрализовать NGF человека, характеризующейся величиной, равной или менее чем 600, 100, 90, 80, 70, 60, 50, 40, 30, 20 или 10 нМ. Обычно партнер специфического связывания по изобретению обладает способностью 5 нМ или менее, предпочтительно 2,5 нМ или менее, более предпочтительно 1 нМ или менее. В особенно предпочтительных вариантах осуществления способность равна 0,5 нМ или менее, например 0,4 нМ или менее, 0,3 нМ или менее, 0,2 нМ или менее или 0,15 нМ или менее. В некоторых вариантах осуществления способность может приблизительно составлять 0,1 нМ.
Способность может находиться между 0,1-100 нМ, 0,1-50 нМ, 0,1-10 нМ или 0,1-1,0 нМ. Например, способность может составлять 0,1-5,0 нМ, 0,2-5,0 нМ, 0,3-5,0 нМ или 0,3-0,4 нМ.
В некоторых вариантах осуществления изобретения нейтрализующая способность неоптимизированного по способности партнера специфического связывания в анализе на клетках HEK, как описано здесь, приблизительно равна от 1,8 до 560 нМ для NGF человека и/или приблизительно от 2,9 до 620 нМ для NGF крысы. В некоторых вариантах осуществления нейтрализующая способность оптимизированных по способности партнеров связывания в анализах на клетках HEK, как описано здесь, равна приблизительно от 0,12 до 120 нМ для NGF человека, приблизительно от 0,11 до 37 нМ для NGF крысы и приблизительно от 0,11 до 71 нМ для NGF мыши. Однако эти данные являются только примерами и можно достичь более высоких способностей. Хотя оптимизацию способности можно использовать для получения партнеров специфического связывания с более высокой способностью из данного партнера специфического связывания, также отмечают, что партнеров специфического связывания с хорошей способностью можно получить даже без оптимизации способности.
Партнер специфического связывания согласно настоящему изобретению предпочтительно ингибирует выживаемость лишенных сыворотки клеток PC12, поддерживаемых NGF. Нейтрализующая способность партнера специфического связывания по настоящему изобретению в анализе выживаемости PC12 для NGF человека, как описано здесь в примере 5, как правило, 1500 нМ или менее и предпочтительно 50 нМ или менее, или 10 нМ или менее. Как объяснено выше и как продемонстрировано здесь, оптимизацию способности можно использовать для достижения более высоких способностей против NGF. Предпочтительно партнер специфического связывания обладает способностью, равной или менее чем 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1,5 нМ, 1 нМ или 0,5 нМ. В некоторых вариантах осуществления способность приблизительно равна 0,1 нМ или более, 0,2 нМ или более. Таким образом, способность может находиться между 0,1 или 0,2 нМ и 0,5, 1,5, 5 или 50 нМ.
В некоторых вариантах осуществления изобретения нейтрализующая способность оптимизированного по способности партнера специфического связывания в анализе выживаемости PC12, как описано здесь, равна приблизительно от 0,2 до 670 нМ для NGF человека и приблизительно от 0,2 до 54 нМ для NGF крысы.
Партнер специфического связывания согласно настоящему изобретению предпочтительно ингибирует пролиферацию клеток TF-1, стимулированную NGF. Нейтрализующая способность партнера специфического связывания (обычно оптимизированного по способности партнера специфического связывания) по настоящему изобретению в анализе пролиферации TF-1 для NGF человека, как описано здесь в примере 6, как правило, равна 5 нМ или менее, предпочтительно 1 нМ или менее. Предпочтительно партнер специфического связывания по изобретению обладает способностью, равной или менее чем 0,7, 0,6, 0,5, 0,45, 0,4, 0,3, 0,2 или 0,1 нМ для NGF человека. Например, способность может находиться между 0,05-0,1 нМ, 0,05-0,2 нМ, 0,05-0,3 нМ, 0,05-0,4 нМ или 0,05-0,5 нМ.
В некоторых вариантах осуществления изобретения нейтрализующая способность оптимизированного по способности партнера специфического связывания в анализе пролиферации TF-1, как описано здесь, равна приблизительно от 0,08 до 0,7 нМ для NGF человека, приблизительно от 0,07 до 1,9 нМ для NGF крысы и приблизительно от 0,07 до 1,4 нМ для NGF мыши.
Партнер специфического связывания согласно настоящему изобретению предпочтительно ингибирует связывание NGF с TrkA и/или рецептором p75, предпочтительно TrkA и/или рецептором p75 человека. Изобретение также относится, в более широком смысле, к партнеру специфического связывания, который предпочтительно блокирует связывание NGF с рецептором TrkA по сравнению со связыванием NGF с рецептором p75. Нейтрализующая способность партнера специфического связывания (обычно оптимизированного по способности партнера специфического связывания) по настоящему изобретению в анализе связывания с рецептором TrkA, как описано здесь в примере 9, равна, как правило, 2,5 нМ или менее, предпочтительно 1 нМ или менее для нейтрализации NGF человека. Предпочтительно партнер специфического связывания по изобретению обладает способностью, равной или менее чем 0,5, 0,4, 0,3, 0,2, 0,1 или 0,075 нМ для нейтрализации связывания NGF человека с TrkA. Например, способность может находиться между 0,05-0,1 нМ, 0,05-0,2 нМ, 0,05-0,3 нМ, 0,05-0,4 нМ или 0,05-0,5 нМ.
Нейтрализующая способность партнера специфического связывания (обычно оптимизированного по способности партнера специфического связывания) по настоящему изобретению в анализе связывания рецептора p75, как описано здесь в примере 9, как правило, равна 1,5 нМ или менее, предпочтительно 1 нМ или менее для нейтрализации NGF человека. Предпочтительно партнер специфического связывания по изобретению обладает способностью, равной или менее чем 0,9, 0,8, 0,7, 0,6, 0,5, 0,4, 0,3, 0,2 или 0,1 нМ для нейтрализации связывания NGF человека с p75. Например, способность может находиться между 0,1-0,2 нМ, 0,1-0,3 нМ, 0,1-0,4 нМ, 0,1-0,5 нМ или 0,1-0,6 нМ.
Некоторые предпочтительные партнеры специфического связывания согласно настоящему изобретению ингибируют связывание NGF (например, NGF человека и/или крысы) с рецептором TrkA предпочтительно по сравнению со связыванием NGF с рецептором p75. Соответственно, в некоторых вариантах осуществления партнер специфического связывания по изобретению обладает меньшей константой ингибирования связывания, Ki, для ингибирования связывания NGF (например, NGF человека и/или крысы) с TrkA по сравнению со связыванием NGF с p75. Ki можно вычислить с использованием формулы, представленной в примере 9. Альтернативно, константы ингибирования связывания можно выразить как pKi, которую можно вычислить как -log10Ki. Таким образом, партнер специфического связывания по изобретению предпочтительно обладает большей величиной pKi для ингибирования связывания NGF с TrkA, чем с p75.
Предпочтительно партнер специфического связывания согласно изобретению связывается с NGF человека и/или NGF крысы с аффинностью, равной или менее чем 1, 0,8, 0,7, 0,6, 0,5, 0,4, 0,3 или 0,2 нМ. Например, партнер специфического связывания может связываться с NGF человека с аффинностью приблизительно 0,25-0,44 нМ и NGF крысы с аффинностью приблизительно 0,25-0,70 нМ.
Как указано выше, варианты молекул антител, описанных здесь, можно получать и применять по настоящему изобретению. Следуя за лидерами в вычислительной химии в применении способов многомерного анализа данных к взаимоотношениям структура/свойство-активность (Wold, et al. 1984), количественные связи активность-свойство для антител можно вывести с использованием широко известных математических способов, таких как статистическая регрессия, распознавание образов и классификация (Norman et al. 1998; Kandel & Backer, 1995; Krzanowski, 2000; Witten & Frank, 1999; Denison (Ed), 2002; Ghose & Viswanadhan). Свойства антител можно вывести из эмпирических и теоретических моделей (например, анализа вероятно контактирующих остатков или вычисления физико-химического свойства) последовательности антител, функциональных и трехмерных структур и данные свойства можно рассматривать по отдельности и в сочетании.
Антигенсвязывающий участок антител, образованный VH-доменом и VL-доменом, сформирован шестью петлями полипептида: три из вариабельного домена легкой цепи (VL) и три из вариабельного домена тяжелой цепи (VH). Анализ антител известной атомной структуры прояснил взаимоотношения между последовательностью и трехмерной структурой комбинированных участков антител (Chothia et al. 1992; Al-Lazikani, et al. 1997). Данные взаимоотношения подразумевают, что, за исключением третьей области (петли) в VH-доменах, петли участка связывания обладают одной из небольшого числа основных конформаций цепи: каноническими структурами. Каноническая структура, сформированная в конкретной петле, как показано, определяется ее размером и присутствием определенных остатков в ключевых положениях, как в петле, так и каркасных областях (Chothia et al. и Al-Lazikani et al., выше).
Это изучение взаимоотношения последовательность-структура можно использовать для предсказания таких остатков в антителах с известной последовательностью, но с неизвестной трехмерной структурой, которые важны для поддержания трехмерной структуры петель CDR и, таким образом, поддержания специфичности связывания. Данные предсказания можно подкрепить сравнением предсказаний результирующего продукта из экспериментов по оптимизации лидов. В структурном подходе можно создать модель молекулы антитела (Chothia, et al. 1986) с использованием любого свободно доступного или коммерческого пакета, такого как WAM (Whitelegg & Rees, 2000). Визуализацию белка и программный пакет анализа, такой как Insight II (Accelerys, Inc.) или Deep View (Guex & Peitsch, 1997), можно затем использовать для оценки возможных замен в каждом положении в CDR. Данную информацию можно затем использовать для получения замен, которые, вероятно, обладают минимальным или благоприятным эффектом на активность.
Способы, необходимые для получения замен в аминокислотных последовательностях CDR, VH или VL-доменов антител и партнеров специфического связывания, как правило, доступны в данной области. Можно получить варианты последовательностей с заменами, относительно которых можно или нельзя предсказать наличие минимального или благоприятного эффекта на активность, и тестировать на способность связывать и/или нейтрализовать NGF, и/или на любое другое требуемое свойство.
Варианты аминокислотных последовательностей вариабельных доменов любого из VH- и VL-доменов, чьи последовательности, в частности, описаны здесь, можно применять согласно настоящему изобретению, как рассмотрено. Конкретные варианты могут содержать одно или более изменений в аминокислотной последовательности (добавление, делецию, замену и/или вставку аминокислотного остатка), могут содержать менее чем приблизительно 20 изменений, менее чем приблизительно 15 изменений, менее чем приблизительно 10 изменений или менее чем приблизительно 5 изменений, возможно 5, 4, 3, 2 или 1. Изменения можно осуществить в одной или более каркасных областях и/или одной или более CDR.
Предпочтительно изменения не приводят к утрате функции, поэтому партнер специфического связывания, содержащий таким образом измененную аминокислотную последовательность, предпочтительно сохраняет способность связывать и/или нейтрализовать NGF. Более предпочтительно он сохраняет такую же количественную связывающую и/или нейтрализующую способность, как партнер специфического связывания, в котором не производили изменение, например, при измерении в анализе, описанном здесь. Наиболее предпочтительно партнер специфического связывания, содержащий таким образом измененную аминокислотную последовательность, обладает улучшенной способностью связывать или нейтрализовать NGF.
Изменение может включать замену одного или более аминокислотных остатков на не встречающуюся в природе или нестандартную аминокислоту, модификацию одного или более аминокислотных остатков в не встречающуюся в природе или нестандартную форму, или вставку одной или более не встречающихся в природе или нестандартных аминокислот в последовательность. Предпочтительные количества и положения изменений в последовательностях по изобретению описаны в других местах данного описания. Встречающиеся в природе аминокислоты включают 20 «стандартных» L-аминокислот, обозначенных как G, A, V, L, I, M, P, F, W, S, T, N, Q, Y, C, K, R, H, D, E посредством стандартного однобуквенного кода. Нестандартные аминокислоты включают любой другой остаток, который можно включить в полипептидный остов или получить в результате модификации существующего аминокислотного остатка. Нестандартные аминокислоты могут встречаться в природе или не встречаться в природе. Несколько встречающихся в природе нестандартных аминокислот известно в данной области, таких как 4-гидроксипролин, 5-гидроксилизин, 3-метилгистидин, N-ацетилсерин и т.п. (Voet & Voet, 1995). Те аминокислотные остатки, которые модифицированы в положении N-α, будут расположены только на N-конце аминокислотной последовательности. Обычно, по настоящему изобретению аминокислота является L-аминокислотой, но в некоторых вариантах осуществления она может являться D-аминокислотой. Изменение может, таким образом, включать модификацию L-аминокислоты в или замену ее на D-аминокислоту. Также известны метилированные, ацетилированные и/или фосфорилированные формы аминокислот, и аминокислоты по настоящему изобретению можно подвергать такой модификации.
Аминокислотные последовательности в доменах антител и партнерах специфического связывания по изобретению могут содержать неприродные или нестандартные аминокислоты, описанные выше. В некоторых вариантах осуществления нестандартные аминокислоты (например, D-аминокислоты) можно включить в аминокислотную последовательность в ходе синтеза, в то время как в других вариантах осуществления нестандартные аминокислоты можно ввести модификацией или заменой «исходных» стандартных аминокислот после синтеза аминокислотной последовательности.
Использование нестандартных и/или не встречающихся в природе аминокислот повышает структурное и функциональное разнообразие и может, таким образом, повышать потенциал для достижения требуемых свойств связывания и нейтрализации NGF у партнера специфического связывания по изобретению. Кроме того, D-аминокислоты и аналоги, как показано, обладают лучшими фармакокинетическими профилями по сравнению со стандартными L-аминокислотами вследствие деградации in vivo полипептидов, содержащих L-аминокислоты, после введения животному.
Как указано выше, аминокислотная последовательность CDR в основном, как представлено здесь, предпочтительно находится как CDR в вариабельном домене антител человека или его существенной части. Последовательности HCDR3 в основном, как представлено здесь, представляют предпочтительные варианты осуществления настоящего изобретения, и предпочтительно, чтобы каждая из них находилась как HCDR3 в вариабельном домене тяжелой цепи человека или его существенной части.
Вариабельные домены, применяемые по изобретению, можно получить или выделить из любого вариабельного домена зародышевой линии или перестроенного вариабельного домена человека, или они могут являться синтетическим вариабельным доменом, основанным на консенсусных или реальных последовательностях известных вариабельных доменов человека. Последовательность CDR по изобретению (например, CDR3) можно ввести в репертуар вариабельных доменов, у которых отсутствует CDR (например, CDR3), с использованием технологии рекомбинантных ДНК.
Например, в Marks et al. (1992) описаны способы получения репертуаров вариабельных доменов антител, у которых консенсусные праймеры, направленные на 5'-конец области вариабельного домена или расположенные рядом с ним, используют в сочетании с консенсусными праймерами к третьей каркасной области генов VH человека для получения репертуара вариабельных VH-доменов, лишенных CDR3. В Marks et al. далее описано, как данный репертуар можно объединять с CDR3 конкретного антитела. С использованием аналогичных способов последовательности по настоящему изобретению, полученные из CDR3, можно переставлять с репертуарами VH- или VL-доменов, лишенных CDR3, и переставленные полные VH- или VL-домены объединять с родственным VL- или VH-доменом для получения партнеров специфического связывания по изобретению. Репертуар можно затем экспонировать в подходящей системе-хозяине, такой как система фагового дисплея из WO 92/01047 или любой работы из последующего большого количества литературы, включая Kay, Winter & McCafferty (1996), так, что можно отбирать подходящих партнеров специфического связывания. Репертуар может состоять из любых от 104 индивидуальных членов и выше, например от 106 до 108 или 1010 членов. Другие подходящие системы-хозяева включают дрожжевой дисплей, бактериальный дисплей, дисплей на основе T7, рибосомный дисплей и ковалентный дисплей.
Аналогичные способы перестановки или сочетания также описаны в Stemmer (1994), где описан способ, относящийся к гену β-лактамазы, но замечено, что способ можно использовать для получения антител.
Другой альтернативой является получение новых VH- или VL-областей, несущих последовательности по изобретению, полученные из CDR, с использованием случайного мутагенеза одного или более выбранных VH- и/или VL-генов для получения мутаций в целом вариабельном домене. Такой способ описан в Gram et al. (1992), где использована PCR с низкой точностью. В предпочтительных вариантах осуществления одна или две аминокислотные замены получены в наборе HCDR и/или LCDR.
Другой способ, который можно использовать, представляет собой направление мутагенеза в области CDR VH- или VL-генов. Такие способы описаны в Barbas et al. (1994) и Schier et al. (1996).
Все вышеописанные способы, по существу, известны в данной области, и специалист будет способен применять такие способы для получения партнеров специфического связывания по изобретению с использованием общепринятой методологии в данной области.
Дополнительный аспект изобретения относится к способу получения антигенсвязывающего участка антител, специфичного для антигена NGF, где способ включает получение посредством добавления, делеции, замены или вставки одной или более аминокислот в аминокислотную последовательность VH-домена, представленного здесь, VH-домена, который является вариантом аминокислотной последовательности VH-домена, не обязательно, объединяя таким образом полученный VH-домен, с одним или более VL-доменами, и тестируя VH-домен, или сочетание или сочетания VH/VL, чтобы идентифицировать партнера специфического связывания или антигенсвязывающий участок антител, специфичный к антигену NGF, и, не обязательно, с одним или более предпочтительных свойств, предпочтительно способности нейтрализовать активность NGF. Указанный VL-домен может обладать аминокислотной последовательностью, которая является такой, как в основном, как представлено здесь.
Можно использовать аналогичный способ, в котором один или более вариантов последовательностей VL-домена, описанного здесь, объединяют с одним или более VH-доменов.
В предпочтительном варианте осуществления VH-домен 1252A5 (SEQ ID NO:392) можно подвергнуть мутагенезу для получения одного или более вариантов аминокислотной последовательности VH-домена, не обязательно, объединенного с VL 1252A5 (SEQ ID NO:397).
Дополнительный аспект изобретения относится к способу получения партнера специфического связывания, специфичного для антигена NGF, где способ включает:
(a) получение начального репертуара нуклеиновых кислот, кодирующих VH-домен, который или содержит CDR3, которую замещают, или лишен области, кодирующей CDR3;
(b) объединение указанного репертуара с донорной нуклеиновой кислотой, кодирующей аминокислотную последовательность в основном, как представлено здесь, VH CDR3, так, что указанная донорная нуклеиновая кислота встраивается в область CDR3 репертуара, так, чтобы получить результирующий репертуар нуклеиновых кислот, кодирующих VH-домен;
(c) экспрессию нуклеиновых кислот указанного результирующего репертуара;
(d) отбор партнера специфического связывания, специфичного для NGF; и
(e) выделение указанного партнера специфического связывания или нуклеиновой кислоты, его кодирующей.
Кроме того, можно использовать аналогичный способ, в котором VL CDR3 по изобретению сочетают с репертуаром нуклеиновых кислот, кодирующих VL-домен, который или содержит CDR3, которую замещают, или лишен области, кодирующей CDR3.
Сходным образом, одну или более, или все три CDR можно встроить в репертуар VH- или VL-доменов, который затем скринируют на партнера специфического связывания или партнеров специфического связывания, специфичных для NGF.
В предпочтительном варианте осуществления можно применять одну или более HCDR1 (SEQ ID NO:393), HCDR2 (SEQ ID NO:394) и HCDR3 (SEQ ID NO:395) 1252A5, или набор HCDR 1252A5, и/или можно применять одну или более LCDR1 (SEQ ID NO:398), LCDR2 (SEQ ID NO:399) и LCDR3 (SEQ ID NO:400) 1252A5, или набор LCDR 1252A5.
В других аналогичных вариантах осуществления 1152H5, 1165D4 или 1230H7 заменены на 1252A5.
Сходным образом, можно применять другие VH- и VL-домены, наборы CDR и наборы HCDR и/или наборы LCDR, описанные здесь.
Существенная часть вариабельного домена иммуноглобулина будет содержать, по меньшей мере, три области CDR вместе с промежуточными каркасными областями. Предпочтительно часть будет также содержать, по меньшей мере, приблизительно 50% любой или обеих первой и четвертой каркасных областей, где 50% являются C-концевыми 50% первой каркасной области и N-концевыми 50% четвертой каркасной области. Дополнительными остатками на N-конце или C-конце существенной части вариабельного домена могут являться те, которые обычно ассоциированы с встречающимися в природе областями вариабельных доменов. Например, конструирование партнеров специфического связывания по настоящему изобретению, осуществляемое способами рекомбинантных ДНК, может приводить к включению N- или C-концевых остатков, кодируемых линкерами, введенных для облегчения клонирования или других стадий манипуляции. Другие стадии манипуляции включают введение линкеров для соединения вариабельных доменов по изобретению с другими белковыми последовательностями, включая константные области антител, другие вариабельные домены (например, при получении димерных антител) или детектируемые/функциональные метки, как более подробно обсуждено в других местах данного описания.
Хотя в предпочтительном аспекте изобретения предпочтительны партнеры специфического связывания, содержащие пару VH- и VL-доменов, одиночные связывающие домены, основанные на последовательностях или VH-, или VL-доменов, образуют дополнительные аспекты изобретения. Известно, что одиночные домены иммуноглобулинов, в особенности VH-домены, способны к связыванию антигенов-мишеней специфическим образом. Например, см. обсуждение dAb выше.
В случае любого из одиночных специфически связывающихся доменов, данные домены можно использовать для скрининга комплементарных доменов, способных образовывать двухдоменного партнера специфического связывания, способного связываться с NGF.
Этого можно достичь посредством способов скрининга фагового дисплея с использованием так называемого иерархического двойного комбинаторного способа, как описано в WO 92/01047, в котором индивидуальную колонию, содержащую клон с или H- или L-цепью, используют для инфицирования полной библиотеки клонов, кодирующих другую цепь (L или H), и полученного двухцепочечного партнера специфического связывания отбирают согласно способам фагового дисплея, как описано в данной ссылке. Данный способ также описан в Marks et al, ibid.
Партнеры специфического связывания по настоящему изобретению могут дополнительно содержать константные области антител или их части, предпочтительно константные области антител человека или их части. Например, VL-домен можно присоединить своим C-концом к константным доменам легкой цепи антител, включая цепи Cκ или Cλ человека, предпочтительно цепи Cλ. Сходным образом, партнера специфического связывания, основанного на VH-домене, можно присоединить своим C-концом ко всей или части (например, CH1 домену) тяжелой цепи иммуноглобулина, полученной из любого изотипа антитела, например IgG, IgA, IgE и IgM, и любого из подклассов изотипа, в частности IgG1 и IgG4. Предпочтителен IgG4. IgG4 предпочтителен, так как он не связывается с комплементом и не осуществляет эффекторных функций. Любой вариант синтетической или другой константной области, который обладает данными свойствами и стабилизирует вариабельные области, также предпочтителен для применения в вариантах осуществления настоящего изобретения.
Партнеров специфического связывания по изобретению можно метить детектируемой или функциональной меткой. Детектируемые метки включают радиоактивные метки, такие как 131I или 99Tc, которые можно присоединить к антителам по изобретению с использованием общепринятых химических способов, известных в данной области визуализации антител. Метки также включают ферментные метки, такие как пероксидаза хрена. Метки далее включают химические группы, такие как биотин, который можно детектировать посредством связывания со специфической родственной детектируемой группой, например меченым авидином.
Партнеры специфического связывания по настоящему изобретению предназначены для применения в способах диагностики или лечения субъектов, являющихся людьми или животными, предпочтительно людьми.
Соответственно, дополнительные аспекты изобретения относятся к способам лечения, включающим введение указанного партнера специфического связывания, фармацевтических композиций, содержащих такого партнера специфического связывания, и применение такого партнера специфического связывания для изготовления лекарственного средства для введения, например, в способе создания лекарственного средства или фармацевтической композиции, содержащей смесь партнера специфического связывания с фармацевтически приемлемым эксципиентом.
Клинические показания, по которым антитела к NGF можно применять для получения терапевтической пользы, включают боль, астму, хроническое обструктивное заболевание легких, фиброз легких, другие воспалительные заболевания дыхательных путей, диабетическую невропатию, артрит, псориаз, сердечные аритмии, HIV и рак. Как уже объяснено, лечение против NGF показано для всех этих заболеваний.
Лечение против NGF можно осуществлять перорально, инъекцией (например, подкожно, внутривенно, внутрибрюшинно или внутримышечно), ингаляцией, внутрипузырным способом (инстилляцией в мочевой пузырь) или местно (например, интраокулярно, интраназально, ректально, в раны, на кожу). Способ введения можно установить исходя из физико-химических характеристик лечения, особых соображений для заболевания или необходимости оптимизировать эффективность или минимизировать побочные эффекты.
Предусмотрено, что лечение против NGF не будет ограничено применением в клинике. Таким образом, также предпочтительна подкожная инъекция с использованием безыгольного устройства.
Сочетанные способы лечения можно использовать для получения значительных синергических эффектов, в частности сочетание партнера специфического связывания с NGF с одним или более другими лекарственными средствами. Партнера специфического связывания согласно настоящему изобретению можно предоставить в сочетании или в дополнение к кратковременно или долговременно действующим обезболивающим, противовоспалительным, противоаллергическим, противоастматическим, противофиброзным, противовирусным, химиотерапевтическим средствам и иммунотерапевтическим средствам.
Сочетанное лечение одним или более, кратковременно или долговременно действующими анальгетиками и/или противовоспалительными средствами, такими как опиаты и нестероидные противовоспалительные лекарственные средства (NSAID), можно применять для лечения состояний, характеризуемых болью и/или воспалением, например ревматоидного артрита или послеоперационной боли. Антитела по настоящему изобретению можно также применять в сочетании с противоастматическими, противоаллергическими или противофиброзными лекарственными средствами, такими как ингаляционные агонисты β-адренорецепторов, стероиды, антагонисты цитокинов, или другими новыми терапевтическими способами лечения астмы, аллергической астмы, других аллергических состояний или любого состояния, характеризуемого патологическим фиброзом. Антитела по настоящему изобретению можно также применять в сочетании с противоинфекционными средствами, например противовирусными средствами для лечения HIV-инфекции.
Согласно настоящему изобретению предоставленные композиции можно вводить индивидуумам. Введение производят предпочтительно в «терапевтически эффективном количестве», которого достаточно для проявления благоприятного воздействия на пациента. Такое благоприятное воздействие может, по меньшей мере, смягчать один из симптомов. Реальное количество введенного средства, и быстрота и продолжительность введения будут зависеть от природы и тяжести того, что подлежит лечению. Рекомендации по лечению, например решения относительно дозировки и т.п., находятся в пределах компетенции врачей общей практики и других врачей и могут зависеть от тяжести симптомов и/или прогрессирования заболевания, подлежащего лечению. Подходящие дозировки антител хорошо известны в данной области; см. Ledermann et al. (1991) и Bagshawe (1991). Можно применять конкретные дозировки, указанные здесь или в Physician's Desk Reference (2003), как соответствующие типу вводимого лекарственного средства. Терапевтически эффективное количество или подходящую дозировку партнера специфического связывания по изобретению можно определить сравнением его активности in vitro и активности in vivo на модели у животного. Известны способы экстраполяции эффективных дозировок у мышей и других подопытных животных на человека.
Конкретная дозировка будет зависеть от ряда факторов, включая то, предназначены ли антитела для диагностики или для лечения, размер и положение области, подлежащей лечению, конкретную природу антитела (например, целое антитело, фрагмент или димерное антитело) и природу любой детектируемой метки или другой молекулы, присоединенной к антителу. Типичная дозировка антител будет находиться в диапазоне от 100 мкг до 1 г для системных применений и от 1 мкг до 1 мг для местных применений. Как правило, антитела будут целыми антителами, предпочтительно изотипа IgG4. Это дозировка для однократного лечения взрослого пациента, которую можно пропорционально приспособить для детей, в том числе грудных, и также приспособить для других видов антител пропорционально молекулярной массе. Лечение можно повторять с периодичностью - ежедневно, два раза в неделю, еженедельно или ежемесячно, по решению врача. В предпочтительных вариантах осуществления по настоящему изобретению лечение является периодическим, и период между введениями составляет приблизительно две недели или более, предпочтительно приблизительно три недели или более, более предпочтительно приблизительно четыре недели или более, или приблизительно один раз в месяц. В других предпочтительных вариантах осуществления изобретения лечение можно осуществлять до и/или после операции и более предпочтительно можно вводить или наносить препарат непосредственно на анатомический участок хирургического вмешательства.
Партнеров специфического связывания по настоящему изобретению будут обычно вводить в форме фармацевтической композиции, которая может содержать, по меньшей мере, один компонент в дополнение к партнеру специфического связывания.
Таким образом, фармацевтические композиции согласно настоящему изобретению и для применения согласно настоящему изобретению могут содержать в дополнение к активному ингредиенту фармацевтически приемлемый эксципиент, носитель, буфер, стабилизатор или другие вещества, хорошо известные специалистам в данной области. Такие вещества должны являться нетоксичными и не должны препятствовать эффективному действию активного ингредиента. Конкретная природа носителя или другого вещества будет зависеть от способа введения, который может являться пероральным или инъекцией, например, внутривенной.
Фармацевтические композиции для перорального введения могут находиться в виде таблетки, капсулы, порошка, жидкости или полутвердой формы. Таблетка может содержать твердый носитель, такой как желатин, или адъювант. Жидкие фармацевтические композиции, как правило, содержат жидкий носитель, такой как вода, нефть, животные или растительные масла, минеральное масло или синтетическое масло. Можно включать физиологический солевой раствор, раствор глюкозы или других сахаров, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной инъекции или инъекции в место повреждения, активный ингредиент будет в форме парентерально приемлемого водного раствора, который является апирогенным и обладает подходящим pH, изотоничностью и стабильностью. Специалисты в данной области в состоянии получить подходящие растворы с использованием, например, изоосмотических наполнителей, таких как хлорид натрия для инъекций, раствор Рингера для инъекций, раствор лактата Рингера для инъекций. Если необходимо, можно включать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки.
Композицию можно вводить по отдельности или в сочетании с другими воздействиями, или одновременно или последовательно, в зависимости от состояния, подлежащего лечению.
Партнеров специфического связывания по настоящему изобретению можно предоставлять в жидких, полутвердых или твердых формах в зависимости от физико-химических свойств молекулы и способа доставки. Составы могут содержать эксципиенты или сочетания эксципиентов, например: сахара, аминокислоты и поверхностно-активные вещества. Жидкие составы могут содержать антитела в широком диапазоне концентраций и pH. Твердые составы можно получить, например, лиофилизацией, сушкой распылением или сушкой по технологии надкритической жидкости. Составы анти-NGF будут зависеть от предполагаемого способа доставки: например, составы для доставки в легкие могут состоять из частиц с физическими свойствами, которые обеспечивают проникновение глубоко в легкие при вдыхании; местные составы могут содержать модификаторы вязкости, которые увеличивают время, в течение которого лекарственное средство находится на месте действия. В некоторых вариантах осуществления партнера специфического связывания можно получить с носителем, который будет защищать партнера специфического связывания от быстрого высвобождения, таким как состав с контролируемым высвобождением, включая имплантаты, чрезкожные пластыри и микроинкапсулированные системы доставки. Можно использовать биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Множество способов получения таких составов известны специалистам в данной области. См., например, Robinson, 1978.
Настоящее изобретение относится к способу, включающему обеспечение связывания или предоставление возможности связывания партнера специфического связывания, как представлено здесь, с NGF. Как отмечено, такое связывание может иметь место in vivo, например, после введения партнера специфического связывания или нуклеиновой кислоты, кодирующей партнера специфического связывания, или оно может иметь место in vitro, например, в ELISA, вестерн-блоттинге, иммуноцитохимии, иммунопреципитации, аффинной хроматографии или анализах с использованием клеток, таких как анализ TF-1.
Можно определить степень связывания партнера специфического связывания с NGF. Количественное определение может соотноситься с количеством антигена в тестируемом образце, который может представлять собой диагностический интерес.
Набор, содержащий партнера специфического связывания или молекулу антитела согласно любому из аспектов или вариантов осуществления настоящего изобретения, также представлен как аспект настоящего изобретения. В наборе по изобретению партнера специфического связывания или молекулу антитела можно пометить, чтобы позволить определить его реактивность в образце, например, как описано далее ниже. Компоненты набора, как правило, стерильные и находятся в запаянных флаконах или других емкостях. Наборы можно применять в диагностическом анализе или других способах, для которых пригодны молекулы антител. Набор может содержать инструкции для использования компонентов в способе, например в способе согласно настоящему изобретению. Можно включать в набор по изобретению вспомогательные вещества для облегчения или возможности осуществления такого способа.
Реакционноспособность антител в образце можно определять любыми подходящими способами. Радиоиммунный анализ (RIA) представляет собой одну из возможностей. Радиоактивно меченный антиген смешивают с немеченым антигеном (тестируемым образцом) и позволяют связаться с антителами. Связанный антиген физически отделяют от несвязанного антигена и определяют количество радиоактивного антигена, связанного с антителами. Чем больше антигена присутствует в тестируемом образце, тем меньше радиоактивного антигена будет связываться с антителами. Анализ конкурентного связывания можно также использовать с нерадиоактивным антигеном, с использованием антигена или аналога, связанного с репортерной молекулой. Репортерная молекула может являться флуорохромом, люминофором или лазерным красителем, со спектрально обособленными характеристиками поглощения или эмиссии. Подходящие флуорохромы включают флуоресцеин, родамин, фикоэритрин и техасский красный. Подходящие хромогенные красители включают диаминобензидин.
Другие репортеры включают макромолекулярные коллоидные частицы или дисперсный материал, такой как гранулы латекса, которые являются окрашенными, магнитными или парамагнитными, и биологически или химически активные вещества, которые могут непосредственно или опосредованно вызывать детектируемые сигналы для визуального наблюдения, электронного детектирования или другого способа регистрации. Данные молекулы могут являться ферментами, которые, например, катализируют реакции, которые вызывают появление или изменение окраски или вызывают изменения электрических свойств. Они могут являться молекулярно возбудимыми, так что электронные переходы между энергетическими состояниями приводят к характеристическим спектрам поглощений или эмиссий. Они могут включать химические вещества, используемые в соединении с биосенсорами. Можно использовать системы детекции на основе биотин/авидина или биотин/стрептавидина и щелочной фосфатазы.
Сигналы, производимые индивидуальными конъюгатами антитело-репортер, можно использовать для получения измеримых абсолютных или относительных данных для соответствующего связывания антител в образцах (нормальном и тестируемом).
Настоящее изобретение также относится к применению партнера специфического связывания, как указано выше, для измерения уровней антигена в конкурентном анализе, то есть в способе измерения уровня антигена в образце с применением партнера специфического связывания, как представлено согласно настоящему изобретению, в конкурентном анализе. Это можно применять там, где не требуется физическое отделение связанного от несвязанного антигена. Одной из возможностей является присоединение репортерной молекулы к партнеру специфического связывания, так что при связывании происходит физическое или оптическое изменение. Репортерная молекула может непосредственно или опосредованно давать детектируемые и предпочтительно измеримые сигналы. Присоединение репортерной молекулы может являться непосредственным или опосредованным, ковалентным, например, посредством пептидной связи, или нековалентным. Присоединение посредством пептидной связи может происходить в результате рекомбинантной экспрессии слитого гена, кодирующего антитело и репортерную молекулу.
Настоящее изобретение также относится к непосредственному измерению уровней антигена посредством применения партнера специфического связывания согласно изобретению, например, биосенсорной системе.
Способ определения связывания не относится к настоящему изобретению, и специалисты в данной области в состоянии выбрать подходящий способ согласно своему предпочтению и общедоступному знанию.
Как указано, в различных аспектах и вариантах осуществления настоящее изобретение распространяется на партнера специфического связывания, который конкурирует за связывание с NGF с любым партнером специфического связывания, определенным здесь, например IgG4 1252A5. Конкуренцию между партнерами связывания можно легко анализировать in vitro, например, мечением посредством специфической репортерной молекулы одного из партнеров связывания, которого можно детектировать в присутствии другого немеченого партнера(ов) связывания, чтобы дать возможность идентифицировать партнеров специфического связывания, которые связываются с одинаковым эпитопом или перекрывающимся эпитопом.
Конкуренцию можно определять, например, с использованием ELISA, в котором NGF иммобилизован на планшете, и в планшет добавляют первого меченого партнера связывания вместе с одним или более другими немечеными партнерами связывания. Наличие немеченого партнера связывания, который конкурирует с меченым партнером связывания, наблюдают по уменьшению сигнала, порождаемого меченым партнером связывания.
При тестировании конкуренции можно использовать пептидный фрагмент антигена, в особенности, пептид, содержащий интересующий эпитоп или существенным образом состоящий из него. Можно использовать пептид, обладающий последовательностью эпитопа плюс одна аминокислота или более на каждом конце. Партнеры специфического связывания согласно настоящему изобретению могут являться такими, что их связывание с антигеном ингибируется пептидом с данной последовательностью или включающим ее. В таком тестировании можно использовать пептид с любой последовательностью плюс одна аминокислота или более.
Партнеров специфического связывания, которые связывают специфический пептид, можно выделять, например, из библиотеки фагового дисплея посредством пэннинга с пептидом(ами).
Настоящее изобретение далее относится к выделенной нуклеиновой кислоте, кодирующей партнера специфического связывания по настоящему изобретению. Нуклеиновая кислота может содержать ДНК и/или РНК. В предпочтительном аспекте настоящее изобретение относится нуклеиновой кислоте, которая кодирует CDR или набор CDR, или VH-домен, или VL-домен, или антигенсвязывающий участок антител или молекулу антитела, например scFv или IgG4, по изобретению, определено выше.
Настоящее изобретение также относится к конструкциям в форме плазмид, векторов, транскрипционных или экспрессирующих кассет, которые содержат, по меньшей мере, один полинуклеотид, как указано выше.
Настоящее изобретение также относится к рекомбинантной клетке-хозяину, которая содержит одну или более конструкций, как указано выше. Нуклеиновая кислота, кодирующая любую CDR, или набор CDR, или VH-домен, или VL-домен, или антигенсвязывающий участок антител или молекулу антитела, например scFv или IgG4, как представлено, сама образует аспект настоящего изобретения, как и способ получения кодируемого продукта, где способ включает экспрессию кодирующей его нуклеиновой кислоты. Экспрессию можно подходящим образом осуществить культивированием в соответствующих условиях рекомбинантных клеток-хозяев, содержащих нуклеиновую кислоту. После продукции путем экспрессии VH- или VL-домен или партнера специфического связывания можно выделить и/или очистить с использованием любого подходящего способа, затем использовать соответствующим образом.
Партнеров специфического связывания, VH- и/или VL-домены, и молекулы кодирующей нуклеиновой кислоты, и векторы согласно настоящему изобретению можно предоставить выделенными и/или очищенными, например, от их естественного окружения, в, по существу, чистой или гомогенной форме, или, в случае нуклеиновой кислоты, лишенной или, по существу, лишенной нуклеиновой кислоты или генов другого происхождения, чем последовательность, кодирующая полипептид с требуемой функцией. Нуклеиновая кислота согласно настоящему изобретению может включать ДНК или РНК и может являться полностью или частично синтетической. Ссылка на нуклеотидную последовательность, как представлено здесь, включает молекулу ДНК с указанной последовательностью и включает молекулу РНК с указанной последовательностью, в которой U замещает T, если в контексте не требуется по-другому.
Хорошо известны системы для клонирования и экспрессии полипептида во множестве разных клеток-хозяев. Подходящие клетки-хозяева включают бактерии, клетки млекопитающих, растительные клетки, дрожжевые и бакуловирусные системы и трансгенные растения и животных. Экспрессия антител и фрагментов антител в прокариотических клетках хорошо обоснована в данной области. Для обзора см., например, Plückthun (1991). Общепринятым предпочтительным бактериальным хозяином является E. coli.
Экспрессия в культуре эукариотических клеток также доступна для специалистов в данной области как средство выбора для получения партнера специфического связывания, например Chadd & Chamow (2001), Andersen & Krummen (2002), Larrick & Thomas (2001). Линии клеток млекопитающих, доступные в данной области для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомяка (CHO), клетки HeLa, клетки эмбриональной почки хомяка, клетки меланомы мыши NS0, клетки миеломы крысы YB2/0, клетки эмбриональной почки человека, клетки эмбриональной сетчатки человека и многие другие.
Подходящие векторы можно выбрать или сконструировать содержащими подходящие регуляторные последовательности, включая промоторные последовательности, терминаторные последовательности, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие последовательности в соответствующих случаях. Векторы могут являться плазмидами, например фагемидами, или вирусными, например, фагами, в соответствующих случаях. Дополнительные подробности см., например, в Sambrook & Russell (2001). Многие известные способы и протоколы для манипуляции с нуклеиновой кислотой, например получение конструкций нуклеиновой кислоты, мутагенез, секвенирование, введение ДНК в клетки и экспрессия генов, и анализ белков подробно описаны в Ausubel et al., 1988 и Ausubel et al., 1999.
Таким образом, дополнительный аспект настоящего изобретения относится к клетке-хозяину, содержащему нуклеиновую кислоту, как описано здесь. Такая клетка-хозяин может находиться in vitro и может находиться в культуре. Такая клетка-хозяин может находиться in vivo. Существование in vivo клетки-хозяина может позволить внутриклеточную экспрессию партнеров специфического связывания по настоящему изобретению в качестве «intrabodies» или внутриклеточных антител. Внутриклеточные антитела можно использовать для генной терапии.
Еще один аспект относится к способу, включающему введение такой нуклеиновой кислоты в клетку-хозяина. Введение можно осуществлять любым доступным способом. Для эукариотических клеток подходящие способы могут включать трансфекцию с фосфатом кальция, DEAE-декстраном, электропорацию, трансфекцию с помощью липосом и трансдукцию с использованием ретровируса или другого вируса, например вируса коровьей оспы или, для клеток насекомых, бакуловируса. Для введения нуклеиновой кислоты в клетку-хозяина, в частности эукариотическую клетку, можно использовать систему, основанную на вирусах или плазмидах. Плазмидную систему можно поддерживать эписомно или можно встраивать в клетку-хозяина или искусственную хромосому. Встраивание может являться или случайной, или направленной интеграцией одной или более копий в одиночные или множественные локусы. Для бактериальных клеток подходящие способы могут включать трансформацию с хлоридом кальция, электропорацию и трансфекцию с использованием бактериофага.
Вслед за введением можно вызвать или позволить экспрессию нуклеиновой кислоты, например, культивированием клеток-хозяев в условиях для экспрессии гена.
В одном из вариантов осуществления нуклеиновую кислоту по изобретению встраивают в геном (например, хромосому) клетки-хозяина. Интеграцию можно стимулировать включением последовательностей, которые способствуют рекомбинации с геномом согласно общепринятым способам.
Настоящее изобретение также относится к способу, который включает применение конструкции, как описано выше, для системы экспрессии, чтобы экспрессировать партнера специфического связывания или полипептид, как указано выше.
Аспекты и варианты осуществления настоящего изобретения будут сейчас проиллюстрированы посредством примеров со ссылкой на следующие экспериментальные и сопроводительные чертежи, в которых:
на фиг.1 показаны кривые концентрации-ингибирования для нейтрализации антителами NGF человека (фиг.1A) и крысы (фиг.1B) в анализе мобилизации кальция посредством рецептора TrkA человека. Антитела IgG4 человека к NGF сравнивали с коммерческими антителами к NGF G1131, Mab5260Z и MAB256 для ингибирования внутриклеточной мобилизации кальция, вызванной 1 нМ NGF. Значения данных представляют собой результаты одиночного эксперимента и являются средним значением ± стандартное отклонение для определений в трех параллелях для каждой концентрации антител.
На фиг.2 показано ингибирование внутриклеточной мобилизации кальция в клетках HEK-293, рекомбинантно экспрессирующих рецептор TrkA человека. Антитела IgG4 человека, оптимизированные по способности, оценивали по ингибированию реакций, вызванных 1 нМ NGF человека (фиг.2A), крысы (фиг.2B) и мыши (фиг.2C). Значения данных представляют собой результаты одиночного эксперимента и являются средним значением ± стандартное отклонение для определений в трех параллелях для каждой концентрации антител.
На фиг.3 показан ингибиторный эффект антител IgG4 человека к NGF в анализе на выживаемость клеток PC12. Жизнеспособность клеток поддерживали в присутствии 1 нМ NGF человека (фиг.3A) или NGF крысы (фиг.3B). Данные представляют собой среднее значение ± стандартное отклонение для определений в трех параллелях в одиночном эксперименте.
На фиг.4 показано ингибирование NGF-опосредованной пролиферации клеток TF-1 антителами IgG4 человека к NGF из зародышевой линии и не из зародышевой линии и с эталонными антителами к NGF MAB256. Клетки стимулировали 200 пМ NGF человека (фиг.4A), NGF крысы (фиг.4B) или NGF мыши (фиг.4C). Данные представляют собой среднее значение ± стандартное от среднего для определений в трех параллелях в одиночном эксперименте.
На фиг.5 показано отсутствие перекрестной рeактивности антител IgG4 1252A5 человека к NGF с нейротрофинами BDNF, NT-3 и NT-4. В лунку добавляли 100 нг нейтрофина. Каждое значение данных представляет собой среднее значение ± стандартное отклонение для определений в трех параллелях в одиночном эксперименте.
На фиг.6 показаны кривые насыщения связывания для связывания 125I-NGF человека с антителами IgG4 1252A5 (фиг.6A) человека к NGF и G1152H5 (фиг.6B). Вычисленные значения Kd соответственно представляют собой 0,35 нМ и 0,37 нМ и являются результатом одиночного эксперимента.
На фиг.7 показаны кривые насыщения связывания для связывания 125I-NGF крысы с антителами IgG4 1252A5 (фиг.7A) человека к NGF и G1152H5 (фиг.7B). Вычисленные значения Kd соответственно представляют собой 0,44 нМ и 0,50 нМ и являются результатом одиночного эксперимента.
На фиг.8 показано ингибирование в зависимости от концентрации связывания 125I-NGF человека (фиг.8A) или крысы (фиг.8B) со слитым белком рецептора TrkA посредством IgG4 человека или эталонных антител. Концентрация радиоактивно меченного NGF в каждой измеряемой лунке составляла приблизительно 150 пМ. Данные являются результатом одиночного эксперимента. См. также таблицы 6 и 7.
На фиг.9 показано ингибирование в зависимости от концентрации связывания 125I-NGF человека (фиг.9A) или крысы (фиг.9B) со слитым белком рецептора p75 посредством IgG4 человека или эталонных антител. Концентрация радиоактивно меченного NGF в каждой измеряемой лунке составляла приблизительно 150 пМ. Данные являются результатом одиночного эксперимента. См. также таблицы 8 и 9.
На фиг.10 показано дозозависимое ингибирование тепловой гипералгезии, индуцированной каррагинаном, у мыши; 48 ч после системного введения человеческого IgG4 1252A5 к NGF.
ПРИМЕР 1
Выделение scFv к NGF
Репертуар антител ScFv
Для селекций использовали три библиотеки большой одиночной цепи Fv (scFv) антител человека, клонированных в фагемидный вектор. Библиотеки происходили из (A) лимфоцитов селезенки (Hutchings, 2001), (B) смеси лимфоцитов периферической крови, B-клеток миндалин и B-клеток костного мозга (Vaughan et al., 1996) и (C) легких цепей и областей VH CDR3 из A, объединенных с каркасом тяжелой цепи зародышевой линии DP47.
Селекция scFv
Способ используемой фаговой селекции являлся, в существенной степени, как описано в Hutchings, выше. ScFv, которые распознавали NGF, выделяли из библиотек фагового дисплея в сериях циклов селекции на NGF человека и крысы. Вкратце, немодифицированным антигеном покрывали пробирки Nunc Maxisorb. Фаги инкубировали с антигеном в общем объеме 500 мкл 1 час перед промывкой для удаления несвязанного фага. Связанный фаг затем выделяли, как описано в Vaughan et al., выше, и повторяли процедуру селекции. Для облегчения выделения перекрестно-реактивного scFv человека/грызуна осуществляли поочередные раунды селекции соответствующими видовыми изоформами NGF. С любой из библиотек осуществляли максимально четыре раунда селекции с использованием данной стратегии селекции со сменяющими друг друга изоформами. β-NGF или человека, или крысы использовали в качестве начального антигена для селекций первого раунда.
Результирующие продукты из раундов 2-4 располагали в порядке очередности из биохимического скрининга, основываясь на процентной доле выделенных NGF-специфичных клонов и разнообразии последовательностей этих клонов. Процентную долю NGF-специфичных клонов определяли в каждом случае фаговым ELISA. Различия последовательностей определяли секвенированием ДНК.
Протокол фагового ELISA
Культуры бактерий, трансдуцированных фагами, получали в 1 мл среды 2xYT, содержащей 100 мкг/мл ампициллина и 50 мкг/мл канамицина, при встряхивании при 30°C в течение 16 ч. Супернатант фагов получали центрифугированием культуры (10 мин при 3000 об/мин) и блокировали 3% мас./об. сухого молока в PBS 1 ч. После блокирования фаги добавляли в планшеты, предварительно покрытые 1 мкг/мл антигена или постороннего антигена. Планшеты отмывали между каждой стадией тремя промывками PBS-Tween 20 (0,1% об./об.) с последующими тремя промывками PBS. Связанный фаг детектировали инкубацией с конъюгатом пероксидазы хрена (HRP) с анти-M13 (Amersham UK), разведенным 1:5000 в 3% мас./об. сухого молока в PBS 1 ч, и проявляли инкубацией с субстратом тетраметилбензидином (TMB) (Sigma). Колориметрическую реакцию останавливали после соответствующего периода добавлением 0,5 М серной кислоты. Показатели поглощения снимали при 450 нм. Клоны, которые специфично связывались с антигеном, идентифицировали по обладанию сигналом на антиген, большим чем или равным 5x сигналу постороннего антигена.
Секвенирование ДНК
Двухцепочечную ДНК-матрицу для секвенирования получали посредством PCR scFv с использованием праймеров FDTETSEQ24 (TTTGTCGTCTTTCCAGACGTTAGT - SEQ ID NO:534) и PUCreverse (AGCGGATAACAATTTCACACAGG - SEQ ID NO:535). Избыток праймеров и dNTP из первичного PCR удаляли с использованием 96-луночных планшетов для PCR Macherey-Nagel Nucleofast (Millipore) согласно инструкциям производителя. Гены VH секвенировали при помощи праймера Lseq (GATTACGCCAAGCTTTGGAGC SEQ ID NO:536). Гены VL секвенировали при помощи праймера MYC Seq 10 (CTCTTCTGAGATGAGTTTTTG SEQ ID NO:537). Каждая реакционная смесь содержала 20-40 нг ДНК, 3-20 пмол праймера и 4 мкл Big Dye Terminator V3,0 (Applied Biosystems, UK) в объеме 20 мкл. Реакции секвенирования состояли из 25 циклов 96°C 10 с; 50°C, 5 с; 60°C, 4 мин. Образцы разделяли и анализировали при помощи анализатора ДНК Applied Biosystems 3700. Неоднозначные области анализировали вручную с использованием программного обеспечения Continuity собственной разработки (Cambridge Antibody Technology, UK) и программного обеспечения для манипуляции с последовательностями ДНК SeqEd (Applied Biosystems, UK).
Биохимический скрининг на scFv, нейтрализующий NGF
Результирующий продукт из процедуры фаговой селекции далее скринировали для идентификации клонов, которые ингибируют связывание NGF со слитым белком внеклеточного домена рецептора TrkA.
Необработанные образцы scFv получали из периплазматических лизатов бактерий E.coli TG-1, трансфицированных селектированным фагом для оценки в анализе связывания. 96-луночные планшеты Nunc Maxisorb покрывали в течение ночи слитым белком внеклеточного домена рецептора TrkA человека (R&D Systems; концентрация для покрытия: анализ NGF человека - 0,25 нМ; анализ NGF крысы - 1 нМ). Планшеты для анализа промывали PBS/Tween 20, забивали 2 час с использованием 1% бычьего сывороточного альбумина (BSA) в PBS и промывали снова. Образцы ScFv преинкубировали 30 мин с 1 нМ человеческого или крысиного рекомбинантного β-NGF (R&D Systems) в 1% BSA. Образцы переносили в объеме 100 мкл в анализируемые планшеты и инкубировали 60 мин при комнатной температуре. Планшеты промывали и NGF, который остался связанным с планшетами, метили с использованием 0,3 мкг/мл или биотина к NGF человека (Peprotech), или биотина к NGF крысы (R&D Systems), разведенным в 1% BSA, с последующей инкубацией 60 мин при комнатной температуре. Меченные биотином антитела к NGF детектировали с использованием системы детектирования флуоресценции с временным разрешением DELFIA (Wallac). Вкратце, планшеты промывали и добавляли в каждую лунку 100 мкл стрептавидина-Eu3+, разведенного 1/1000 в буфере для анализа DELFIA. Планшеты инкубировали дополнительно 60 мин при комнатной температуре и промывали промывочным буфером DELFIA, с последующим добавлением 100 мкл усиливающего раствора DELFIA в каждую лунку. С планшетов снимали показания с использованием флуориметрического планшетного анализатора Wallac Victor (длина волны возбуждения 314 нм; длина волны эмиссии 615 нм).
Клоны, которые ингибировали связывание NGF как человека, так и крысы более чем на 70% в качестве препаратов периплазматических лизатов scFv, пересчитывали на очищенный scFv в анализе связывания. Очищенные препараты scFv получали, как описано в примере 3 WO 01/66754. Концентрации белка в очищенных препаратах scFv определяли с использованием способа BCA (Pierce). Повторным анализом выявили следующие антитела ScFv, которые являлись способными нейтрализовать связывание NGF человека и крысы со слитым белком внеклеточного домена рецептора TrkA человека:
1064F8 (VН SEQ ID NO:2; VL SEQ ID NO:7),
1022E3 (VН SEQ ID NO:12; VL SEQ ID NO:17),
1083H4 (VН SEQ ID NO:22; VL SEQ ID NO:27),
1021E5 (VН SEQ ID NO:32; VL SEQ ID NO:37),
1033G9 (VН SEQ ID NO:42; VL SEQ ID NO:47),
1016A8 (VН SEQ ID NO:52; VL SEQ ID NO:57),
1028F8 (VН SEQ ID NO:62; VL SEQ ID NO:67),
1033B2 (VН SEQ ID NO:72; VL SEQ ID NO:77),
1024C4 (VН SEQ ID NO:82; VL SEQ ID NO:87) и
1057F11 (VН SEQ ID NO:92; VL SEQ ID NO:97).
ПРИМЕР 2
Экспрессия антител IgG4 человека и функциональная оценка in vitro способности нейтрализовать NGF
Нейтрализующие NGF scFv 1064F8, 1022E3, 1083H4, 1021E5, 1033G9, 1016A8, 1028F8, 1033B2, 1024C4 и 1057F11 переделали как антитела IgG4 человека и проанализировали на способность нейтрализовать NGF в системе с интактными клетками.
Конверсия IgG
Векторы конструировали для клонов scFv с наибольшей способностью, чтобы дать возможность их реэкспрессировать как целые антитела IgG4 человека, в существенной степени, как описано в Persic et al. (1997). Клетки EBNA-293, поддерживаемые в кондиционированной среде, котрансфицировали конструкциями, экспрессирующими домены тяжелой и легкой цепей. Целые антитела очищали от среды с использованием аффинной хроматографии на основе белка A (Amersham Pharmacia). Очищенные препараты антител стерилизовали фильтрованием и хранили при 4°C в фосфатно-солевом буферном растворе (PBS) перед измерением способности in vitro. Концентрацию белка определяли спектрофотометрически согласно Mach et al. (1992).
Анализ внутриклеточной мобилизация кальция FLIPR
Способность и эффективность IgG человека для нейтрализации NGF определяли во флуоресцентном анализе мобилизации кальция с использованием клеток. Способность антител человека сравнивали с мышиными антителами к NGF человека (MAB256; R&D Systems), крысиными антителами к NGF мыши (G1131; Promega) и мышиными антителами к NGF мыши (MAB5260Z; Chemicon). Анти-NGF IgG совместно инкубировали с рекомбинантным β-NGF человека (Calbiochem, 480275) или рекомбинантным β-NGF крысы (R&D Systems, 556-NG-100). Комплекс затем добавляли к клеткам HEK293, экспрессирующим рекомбинантные рецепторы TrkA человека, нагруженным чувствительным к кальцию красителем Fluo-4, и затем анализировали Ca2+-зависимую флуоресценцию.
Клетки HEK (peak-S, Edge Biosystems), трансфицированные рекомбинантным TrkA человека (полученным от M. Chao, Skirball Institute, NY), выращивали в среде Игла, модифицированной по Дульбекко (MEM, Cellgro, MT10-017-CV), с добавлением 10% эмбриональной телячьей сыворотки (Hyclone, SH30071.03), 1,5 мкг/мл пуромицина (Edge Biosystems, 80018) и 1% пенициллина-стрептомицина. Конфлюэнтные клетки собирали отделением клеток от подложки в фосфатно-солевом буферном растворе Дульбекко (DPBS) и затем нагружали нагружающим буфером, содержащим 6 мкМ Fluo-4 (Molecular Probes) при 37°C 1,5 час в присутствии ингибитора анионного транспорта (2,5 мМ пробенецида в 1% FBS/MEM). После однократной промывки клеток буфером для анализа (2,5 мМ пробенецида в 0,1% BSA в растворе Хэнка/HEPES) клетки помещали на покрытые поли-D-лизином 96-луночные планшеты с прозрачным дном (Costar № 3904) из расчета приблизительно 60000 клеток/лунку в 120 мкл. Клетки инкубировали в темноте 30 мин при комнатной температуре и планшеты затем центрифугировали при 1200 об/мин (290×g) 5 мин. Перед тестированием планшеты предварительно прогревали при 35°C 20 мин. Тестируемые IgG анализировали при 7 концентрациях в трех параллельных лунках. Тридцать пять микролитров 10X анти-NGF IgG и равные количества 10 нМ 2,5S NGF человека, крысы или мыши преинкубировали при комнатной температуре приблизительно 1 час. Планшеты, содержащие предварительные комплексы с IgG, и планшеты, содержащие 100 мкл/лунка буфера для анализа, предварительно прогревали при 35°C 20 мин перед тестированием на флуориметрическом планшетном анализаторе (FLIPR; Molecular Devices). После добавления 80 мкл/лунка буфера для анализа к планшету с клетками и инкубации 5 мин при 35°C, 50 мкл/лунка разведенного комплекса IgG человека/NGF добавляли к планшету с клетками в FLIPR с непрерывным анализированием Ca2+-зависимой флуоресценции. Флуоресценцию, индуцированную NGF, подсчитывали как разницу между интенсивностью фоновой флуоресценции непосредственно перед добавлением NGF и наивысшей интенсивностью флуоресценции, достигаемой через 2 минуты после добавления NGF. Среднее значение в трех параллелях величин флуоресценции, индуцированной NGF, в отсутствие антител определяли как 100% мобилизацию кальция. В присутствии антител, среднее значение ± стандартное отклонение в трех параллелях величин флуоресценции, индуцированной NGF, подсчитывали как процентную долю от контрольной мобилизации кальция. Процентную долю величин контрольной мобилизация кальция строили как функцию от log концентрации IgG. Значения IC50 подсчитывали аппроксимацией сигмоидальной функции доза-эффект (вариабельный наклон) с использованием Prism (GraphPad).
Все тестируемые антитела демонстрировали ингибирование в зависимости от концентрации стимулированной NGF человека внутриклеточной мобилизация кальция. Порядок ранжирования по способности нейтрализовать NGF человека являлся 1064F8>1022E3>1083H4≥1033G9≥1016A8≥1028F8≥1021E5≥1033B2>>1024C4>>1057F11 (таблица 1). Пять антител далее оценивали по функциональной нейтрализации NGF крысы, и они оказались приблизительно равносильными против изоформ обоих видов (фиг.1A и 1B; таблица 1).
ПРИМЕР 3
Выделение оптимизированных антител IgG4 человека к NGF
Оптимизация способности scFv посредством рибосомного дисплея
Большие библиотеки рибосомного дисплея создали и селектировали на scFv, который специфически узнавал рекомбинантный NGF человека (R&D Systems), существенным образом, как описано в Hanes et al. (2000). Вначале клоны 1064F8, 1022E3, 1083H4, 1021E5, 1033G9, 1016A8, 1028F8, 1033B2 и 1024C4 преобразовали в формат рибосомного дисплея, где матрицы последовательно использовали для создания библиотеки. Клон 1057F11 не выбрали для оптимизации способности, и, таким образом, матрицу рибосомного дисплея не получили на это антитело. На уровне ДНК добавляли промотор T7 на 5'-конец для эффективной транскрипции мРНК. На уровне мРНК конструкция содержала прокариотический участок для связывания с рибосомой (последовательность Шайна-Дальгарно). На 3'-конце одиночной цепи удаляли стоп-кодон и добавляли часть gIII, чтобы она действовала как пространственный разделитель (Hanes et al., выше).
Библиотеки рибосомного дисплея, происходящие из 1064F8, 1022E3, 1083H4, 1021E5, 1033G9, 1016A8, 1028F8, 1033B2 и 1024C4, получали мутагенезом scFv HCDR3. Реакции PCR осуществляли с Taq полимеразой без корректирующей активности. Селекции, основанные на аффинности, осуществляли следующим образом, после инкубации с библиотекой биотинилированный NGF человека присоединяли к парамагнитным бусам, покрытым стрептавидином (Dynal M280). Связанные четверные комплексы (мРНК-рибосома-scFv) выделяли магнитным разделением, при котором отмывались несвязанные комплексы. мРНК, кодирующую связанный scFv, затем выделяли посредством RT-PCR, как описано в Hanes et al., (выше), и процедуру селекции повторяли с уменьшением концентраций (100 нМ-10 пМ в ходе пяти раундов) биотинилированного NGF человека, присутствующего в ходе селекции.
PCR с низкой точностью также использовали для дальнейшего увеличения размера библиотеки. В условиях селекции использовали уровень ошибок из расчета 7,2 мутации на 1000 п.н. (DiversifyTM, Clontech). Реакции PCR с низкой точностью осуществляли перед началом селекций на раундах три и четыре с использованием биотинилированного NGF человека в концентрации 1 нМ и 0,1 нМ, соответственно.
Представительную пропорцию scFv из результирующего продукта после третьего, четвертого и пятого раундов селекции лигировали в вектор pCantab6 (Vaughan et al., 1996) и клонировали в штамме TG1 E.coli. Образец ДНК данных scFv секвенировали, как описано в примере 1, для подтверждения отличия последовательности результирующего продукта перед скринингом in vitro на активность по нейтрализации NGF. Клоны скринировали в виде неочищенного scFv в анализе связывания NGF/слитый белок внеклеточного домена рецептора TrkA, как описано в примере 1. Концентрацию препаратов scFv из периплазматических лизатов уменьшали до 0,5%-5% окончательного объема в анализе, и клоны, которые ингибировали >95% связывание NGF как человека, так и крысы, выделяли для дальнейшего изучения. Данным способом выделяли панель оптимизированных по способности, перекрестно реактивных нейтрализаторов NGF. Неожиданно, наиболее способные нейтрализаторы NGF получили из родительских клонов 1021E5 и 1083H4, которые не являлись наиболее способными из родительских антител. Оптимизированные клоны секвенировали и повторно анализировали в виде очищенного scFv для подтверждения способности перед преобразованием в IgG4 человека, как описано в примере 2.
Антитела, полученные из родительского клона 1021Е5 (VH SEQ ID NO:32; VL SEQ ID NO:37) и преобразованные в формат IgG4 человека, представляли собой 1126F1 (VH SEQ ID NO:102; VL SEQ ID NO:107), 1126G5 (VH SEQ ID NO:112; VL ID NO:117), 1126H5 (VH SEQ ID NO:122; VL SEQ ID NO:127), 1127D9 (VH SEQ ID NO:132; VL SEQ ID NO:137), 1127F9 (VH SEQ ID NO:142; VL SEQ ID NO:147), 1131D7 (VH SEQ ID NO:152; VL SEQ ID NO:157), 1131H2 (VH SEQ ID NO:162; VL SEQ ID NO:167), 1132A9 (VH SEQ ID NO:172; VL SEQ ID NO:177), 1132H9 (VH SEQ ID NO:182; VL SEQ ID NO:187), 1133C11 (VH SEQ ID NO:192; VL SEQ ID NO:197), 1134D9 (VH SEQ ID NO:202; VL SEQ ID NO:207), 1145D1 (VH SEQ ID NO:212; VL SEQ ID NO:217), 1146D7 (VH SEQ ID NO:222; VL SEQ ID NO:227), 1147D2 (VH SEQ ID NO:232; VL SEQ ID NO:237), 1147G9 (VH SEQ ID NO:242; VL SEQ ID NO:247), 1150F1 (VH SEQ ID NO:252; VL SEQ ID NO:257), 1152H5 (VH SEQ ID NO:262; VL SEQ ID NO:267), 1155H1 (VH SEQ ID NO:272; VL SEQ ID NO:277), 1158A1 (VH SEQ ID NO:282; VL SEQ ID NO:287), 1160E3 (VH SEQ ID NO:292; VL SEQ ID NO:297), 1165D4 (VH SEQ ID NO:302; VL SEQ ID NO:307), 1175H8 (VH SEQ ID NO:312; VL SEQ ID NO:317), 1211G10 (VH SEQ ID NO:322; VL SEQ ID NO:327), 1214A1 (VH SEQ ID NO:332; VL SEQ ID NO:337), 1214D10 (VH SEQ ID NO:342; VL SEQ ID NO:347), 1218H5 (VH SEQ ID NO:352; VL SEQ ID NO:357) и 1230Н7 (VH SEQ ID NO:362; VL SEQ ID NO:367).
Антитела, полученные из родительского клона 1083H4 (VH SEQ ID NO:22; VL SEQ ID NO:27) и преобразованные в формат IgG4 человека, представляли собой 1227H8 (VH SEQ ID NO:372; VL SEQ ID NO:377) и 1230D8 (VH SEQ ID NO:382; VL SEQ ID NO:387).
Превращение в зародышевую линию каркасных областей 1133C11 для получения 1252A5 и других вариантов 1021E5
Изучение информации о последовательностях VH и VL CDR для оптимизированных клонов, полученных из 1021E5, выявило, что значительная пропорция этих антител содержала аминокислотную последовательность LNPSLTA (SEQ ID NO:531) в VH CDR3 (т.е. аминокислоты от 100A до 100G согласно нумерационной системе Kabat). Данные клоны приведены в таблице 2a, демонстрируя, как они различаются по аминокислотной последовательности областей VH и VL CDR, вместе с оценкой их NGF-нейтрализующей способностью, при анализе в виде очищенного scFv. Из этих клонов 1133C11 отличался по областям CDR от 1021E5 только 7 последовательными аминокислотами от 100A до 100G, как описано. Таким образом, 1133C11 выбрали для превращения в зародышевую линию, чтобы, во-первых, подтвердить, что способность сохраняется при модификации каркаса, и, во-вторых, позволить ввести последовательные мутации в CDR, если требуется, для получения других интересующих антител зародышевой линии из той же самой линии.
Полученные аминокислотные последовательности VH и VL 1133C11 выравнивали с известными последовательностями зародышевой линии человека в базе данных VBASE (Tomlinson [1997], MRC Centre for Protein Engineering, Cambridge, UK) и идентифицировали наиболее близкую зародышевую линию. Наиболее близкую зародышевую линию для VH 1133C11 идентифицировали как DP10, члена семейства VH1. 1133C11 VH обладает 5 аминокислотными изменениями в каркасных областях по сравнению с зародышевой линией DP10. Наиболее близкую зародышевую линию для VL 1133C11 идентифицировали как DPL5, члена семейства Vλ1. 1133C11 VL обладает только 4 изменениями в каркасных областях по сравнению с зародышевой линией. Каркасные области 1133C11 возвратили к зародышевой линии сайт-специфическим мутагенезом scFv для получения scFv 1252A5 (VH SEQ ID NO:392; VL SEQ ID NO:397). Его превратили в IgG4 человека, как описано в примере 2. Превращение в зародышевую линию для других вариантов клонов, полученных из 1021E5, достигли введением мутаций CDR на превращенный в зародышевую линию остов IgG4 1252A5. Данный способ привел к получению превращенных в зародышевую линию антител G1152H5 (VH SEQ ID NO:402; VL SEQ ID NO:407), G1165D4 (VH SEQ ID NO:412; VL SEQ ID NO:417) и G1230H7 (VH SEQ ID NO:422; VL SEQ ID NO:427).
ПРИМЕР 4
Оценка оптимизированных антител IgG4 человека в анализе FLIPR внутриклеточной мобилизации кальция
Оптимизированные антитела IgG4 человека к NGF оценивали в анализе внутриклеточной мобилизации кальция, вызванной NGF, в клетках, рекомбинантно экспрессирующих рецептор TrkA человека, как описано в примере 2. Антитела анализировали на нейтрализующую активность против NGF человека, крысы и мыши (фиг.2A, 2B и 2C; таблица 3).
Все оптимизированные тестированные антитела человека ингибировали внутриклеточную мобилизацию кальция, вызванную 1 нМ NGF. Оптимизированные антитела демонстрировали субнаномолярные величины IC50, в большинстве случаев представляя более чем стократное усиление способности нейтрализовать NGF по сравнению с родительскими IgG (таблица 3). Нейтрализующие способности (IC50) превращенных в зародышевую линию антител IgG4 человека к изоформам NGF человека, крысы и мыши являлись, соответственно:
1252А5 - 0,33 нМ, 0,29 нМ и 0,26 нМ;
G1152H5 - 0,22 нМ, 0,27 нМ и 0,18 нМ;
G1165D4 - 0,32 нМ, 0,33 нМ и 0,27 нМ;
G1230H7 - 0,31 нМ, 0,34 нМ и 0,25 нМ.
Данные результаты выявляют эффективность и ценность способа рибосомного дисплея для оптимизации способности антител. Более общепринятым способом оптимизации антител в прошлом являлось получение библиотек фагового дисплея вариантов антител ScFv. Данный способ требует интенсивного труда и медленнее, чем способ рибосомного дисплея, что часто означает, что только один родительский scFv используют в качестве стартовой точки для конструирования библиотеки. Относительная простота получения библиотек рибосомного дисплея позволяет оптимизировать одновременно много родительских scFv, и, как продемонстрировано в примере 3, это может приводить к выделению антител с высокой способностью, полученных из родительских клонов, которыми в ином случае могли пренебречь для оптимизации.
ПРИМЕР 5
Оценка оптимизированных антител IgG4 человека в анализе на выживаемость клеток PC12
В анализе PC12 NGF поддерживает жизнеспособность лишенных сыворотки клеток PC12 крысы, экспрессирующих нативные рецепторы TrkA и p75, в течение двух суток. Нейтрализующие NGF антитела уменьшают выживаемость клеток, измеряемую с помощью AlamarBlue.
Клетки феохромоцитомы крысы PC12 выращивали в RPMI 1640 (Cellgro, 18040181) с добавлением 5% эмбриональной телячьей сыворотки (JRH,12103-78P), 10% донорской лошадиной сыворотки, инактивированной нагреванием (JRH, 12446-77P), и 1% пенициллина-стрептомицина. Клетки собирали гомогенизацией и затем промывали дважды бессывороточной RPMI 1640, содержащей 0,01% BSA (Sigma, A7030). Клетки помещали в 96-луночные планшеты, покрытые коллагеном из хвоста крысы (Biological Technology Institute, BT-274), из расчета 50000 клетки/лунка в 120 мкл бессывороточной среды с 0,01% BSA. Осуществляли серийные разведения 5X IgG к NGF человека с использованием бессывороточной среды и 40 мкл/лунка добавляли к планшету с клетками. 40 мкл/лунка 0,5 нМ β-NGF человека (Calbiochem, 480275) или рекомбинантного NGF крысы (R&D Systems, 556-NG-100) добавляли к планшету и общий объем доводили до 200 мкл/лунка бессывороточной средой. Максимальную клеточную гибель определяли посредством 80 мкл/лунка бессывороточной среды в трех параллельных лунках. 100% жизнеспособность определяли посредством 40 мкл/лунка бессывороточной среды и 40 мкл/лунка 0,5 нМ NGF в трех параллельных лунках. Планшеты инкубировали при 37°C в 5% CO2 48 час. Для измерения жизнеспособности клеток добавляли 22 мкл/лунка AlamarBlue и сразу снимали показания с планшетов для определения фоновой флуоресценции в каждой лунке флуориметрическим планшетным анализатором (BMG) при длине волны возбуждения 530 нМ и длине волны испускания 590 нМ. После инкубации 6~7 час при 37°C повторно снимали показания с планшета для определения полной флуоресценции. Флуоресценцию Alamarblue вычисляли как разницу между фоновой и полной интенсивностями флуоресценции. В присутствии антител, среднее значение ± стандартное отклонение величин флуоресценции в трех параллелях вычисляли как процентную долю от среднего значения величин флуоресценции при 100% выживаемости в трех параллелях. Процентную долю от контрольных величин выживаемости строили как функцию от log концентрации IgG. Величины IC50 вычисляли аппроксимацией сигмоидальной функции доза-эффект (вариабельный наклон) с использованием Prism (GraphPad).
Результаты анализа PC12 дополнительно подтверждали повышенную способность оптимизированных антител IgG4 человека к NGF по сравнению с родительскими IgG. Антитела ингибировали выживаемость клеток PC12, поддерживаемую NGF человека и крысы, зависящим от концентрации образом (фиг.3A и 3B; таблица 3). Превращение в зародышевую линию, как оказалось, уменьшает способность нейтрализовать NGF у тестируемых антител, в особенности, против изоформы NGF крысы. Например, среднее значение IC50 антител зародышевой линии G1152H5 для ингибирования выживаемости клеток, опосредованной 1 нМ NGF человека или крысы, составляла 1,1 нМ и 7,3 нМ, соответственно. В то же время, среднее значение IC50 для нейтрализации 1 нМ NGF человека или крысы посредством 1152H5 (т.е. антителами не из зародышевой линии) составляла 0,40 нМ и 0,38 нМ, соответственно.
ПРИМЕР 6
Активность по нейтрализации NGF в анализе пролиферации клеток TF-1
Линия клеток TF-1 представляет собой премиелоидную линию клеток человека, которую можно стимулировать к пролиферации экзогенными факторами роста и цитокинами. Клетки TF-1 экспрессируют рецептор TrkA человека и пролиферируют в ответ на активацию NGF. Анализ пролиферации клеток TF-1 использовали для дальнейшего исследования функциональной способности к нейтрализации in vitro антителами IgG4 человека к NGF.
Клетки TF-1 получали от R&D Systems и культивировали согласно приложенным протоколам. Среды для анализа состояли из RPMI-1640 с GLUTAMAX I (Invitrogen), содержащей 5% эмбриональной телячьей сыворотки (Hyclone) и 1% пирувата натрия (Sigma). Перед каждым анализом клетки TF-1 осаждали центрифугированием при 300×g в течение 5 минут, среды удаляли отсасыванием и клетки ресуспендировали в средах для анализа. Данную процедуру повторяли три раза с клетками, ресуспендированными в конечной концентрации 105/мл в средах для анализа, и добавляли 100 мкл к каждой лунке 96-луночного планшета с плоским дном для анализа культур тканей, чтобы получить конечную плотность клеток 1×104/лунка. Тестируемые растворы антител (в трех параллелях) разводили, чтобы получить конечную концентрацию для анализа 1 мкг/мл в средах для анализа, и титровали 1:5 во всем анализируемом планшете. Посторонние антитела (CAT-001), не нацеленные на NGF, использовали как отрицательный контроль. Кроме того, эталонные моноклональные антитела MAB256 (R&D Systems) использовали как положительный контроль. 50 мкл тестируемых антител затем добавляли к каждой лунке перед добавлением 50 мкл нативного очищенного NGF из мышиных (7S форма; Invitrogen), крысы (Sigma) или человека (Sigma), разведенного, чтобы получить конечную концентрацию для анализа 200 пМ. Анализируемые планшеты инкубировали 68 час при 37oC в 5% CO2 в инкубаторе с увлажнением. Двадцать микролитров меченного тритием тимидина (5,0 мКи/мл, NEN) затем добавляли в каждую анализируемую лунку и анализируемые планшеты возвращали в инкубатор дополнительно на 5 час. Клетки собирали в 96-луночные планшеты со стекловолоконным фильтром (Perkin Elmer) с использованием коллектора клеток. Затем добавляли MicroScint 20TM (50 мкл) в каждую лунку планшета с фильтром и количественно определяли включение [3H]-тимидина с использованием жидкостного сцинтилляционного счетчика для микропланшетов Packard TopCount. В присутствии антител включение тимидина (количественно определяемое, как импульсы в минуту) вычисляли как разность между средним значением фона (т.е. у клеток без обработки NGF) и средним значением общего числа импульсов в минуту (т.е. клеток, стимулированных NGF) и выражали как процентную долю от максимальной пролиферации. Процентную долю от максимальной пролиферации строили как функцию от log концентрации IgG. Величины IC50 вычисляли аппроксимацией сигмоидальной функции доза-эффект (вариабельный наклон) с использованием Prism (GraphPad).
Антитела IgG4 человека к NGF являлись активными ингибиторами пролиферации клеток TF-1, опосредованной изоформами NGF человека, крысы и мыши (фиг.4A, 4B и 4C). Данные результаты демонстрируют, что антитела, полученные из линии 1021E5, могут нарушать передачу сигнала посредством NGF через активацию нативного рецепторов NGF человека in vitro. Согласно наблюдаемой активности антител к NGF человека в анализе выживаемости клеток PC12 (пример 5), антитела не из зародышевой линии являлись более активными, чем их аналоги из зародышевой линии в анализе пролиферации TF-1 (таблица 3). Основываясь на средних значениях данных по IC50, порядок ранжирования по способности тестируемых антител ингибировать пролиферацию, опосредованную 200 пМ NGF человека, представлял собой 1133C11>1152H5>1252A5=G1152H5>>MAB256.
ПРИМЕР 7
Перекрестная реактивность анти-NGF IgG с другими нейтрофинами
Для определения перекрестной реактивности анти-NGF IgG с другими нейтрофинами осуществляли ELISA. ELISA включали в себя покрывание планшетов из расчета 100 нг/лунка NGF человека (R&D systems, 256-GF) нейротрофическим фактором головного мозга (BDNF; R&D systems, 248-BD), нейротрофином-3 (NT-3; R&D systems, 257-N3) или нейротрофином-4 (NT4; R&D systems, 257-N4) при комнатной температуре 5-6 час, перед блокированием планшетов 0,25% HSA при 4°C в течение ночи. Возрастающие концентрации анти-NGF IgG, варьирующие в пределах 0,03-10 нМ, инкубировали при комнатной температуре 2 час, чтобы позволить связаться с каждым нейротрофином. Анти-NGF IgG детектировали с помощью биотинилированных поликлональных антител против человека (1:300) (Rockland 609-1602), щелочной фосфатазой, соединенной со стрептавидином (1:1000), и флуоресцентным субстратом A. В положительных контролях, демонстрирующих связывание нейротрофина с планшетом, использовали коммерческие биотинилированные поликлональные антитела против человека (R&D Systems анти-NGF BAF 256, анти-BDNF BAM 648, анти-NT-3 BAF 267, анти-NT-4 BAF 268), которые детектировали непосредственно с использованием щелочной фосфатазы, соединенной со стрептавидином, и последующим добавлением субстрата A. Неспецифическое связывание определяли с использованием лунок, покрытых BSA вместо нейротрофина. Образование продукта отслеживали во временном интервале 0-60 мин после добавления субстрата A. Анти-NGF IgG 1064F8 использовали для оптимизации анализа. Для 1064F8 наблюдали линейное образование продукта в течение 15 мин, которое затем выравнивалось с течением времени, вероятно, вследствие истощения субстрата. Перекрестные реактивности вычисляли как процентную долю от специфического связывания нейротрофина, родственного NGF, для общих концентраций IgG. Для высокоаффинных IgG, таких как 1064F8, процентную долю перекрестных реактивностей подсчитывали с использованием данных по образованию продукта в течение 15 мин. Для низкоаффинных IgG, таких как 1016A8, процентную долю перекрестных реактивностей подсчитывали с использованием данных по образованию продукта в течение 60 мин.
Определены перекрестные реактивности семи антител IgG4 анти-NGF человека к BDNF, NT-3 и NT-4 относительно NGF. При тестируемых концентрациях для всех семи антител показана незначительная перекрестная реактивность (таблица 4). Например, для 1252A5 наиболее высокие уровни отмеченной перекрестной реактивности с NT-3, NT-4 и BDNF являлись 1,1%, 0,9% и 1,4% соответственно (фиг.5).
ПРИМЕР 8
Определение аффинности связывания с NGF для антител человека к NGF
Аффинности связывания с NGF антител IgG4 человека к NGF определяли с использованием формата анализа связывания радиоактивно меченного лиганда, осуществляемого при комнатной температуре. Вкратце, разовые планшеты (Perkin Elmer SMP200) покрывали из расчета 100 мкл/лунка 2,2 мг/мл козьего IgG против человека (Sigma-Aldrich, UK) в фосфатно-буферном солевом растворе (PBS) 1 час. Лунки промывали PBS и затем блокировали 1 час 200 мкл/лунка PBS, содержащим 3% мас./об. бычьего сывороточного альбумина (BSA; Sigma-Aldrich, UK). Лунки промывали PBS и добавляли 10 нг антител человека к NGF к каждой лунке в объеме 0,1 мл PBS, содержащего 0,5% мас./об. BSA. После инкубации 1 час планшеты промывали PBS.
Меченный радиоактивным иодом NGF человека и крысы получали от Amersham UK (125I-NGF человека, Amersham cat. № IM286; 125I-NGF крысы, меченный по заказу рекомбинантный β-NGF крысы, приобретенный от R&D Systems, cat. № 556-GF-100). Каждую изоформу 125I-NGF серийно разводили в буфере для анализа (PBS, содержащем 0,5% мас./об. BSA и 0,05% об./об. Tween 20) и 100 мкл образцов в двух параллелях добавляли к анализируемому планшету для получения измерения «общего связывания» в диапазоне концентраций 2 пМ-15 нМ. Неспецифическое связывание (NSB) определяли для каждой концентрации 125I-NGF измерением связывания в присутствии большого избытка нерадиоактивного NGF. Лунки для NSB содержали 125I-NGF (2 пМ-15 нМ) вместе с конечной концентрацией 500 нМ немеченого β-NGF человека (R&D Systems Cat. № 256-GF-100) или β-NGF крысы (R&D Systems Cat. № 556-GF-100), в соответствующих случаях. Планшеты инкубировали в течение ночи и снимали показания с лунок в течение 1 мин на гамма-счетчике (TopCount NXT, Perkin Elmer). Специфическое связывание вычисляли согласно формуле «специфическое связывание = общее связывание - неспецифическое связывание». Строили кривые связывания и параметры связывания определяли согласно модели насыщаемого связывания с одним участком с использованием программного обеспечения Prism (GraphPad Software Inc., USA).
Для 125I-NGF человека и крысы показано насыщаемое высокоаффинное связывание с антителами IgG4 человека к NGF 1252A5 и G1152H5 (фиг.6 и 7). Вычисленные величины Kd для связывающего взаимодействия с 125I-NGF человека составляли 0,35 нМ для 1252A5 и 0,37 нМ для G1152H5. Величины Kd для связывания с 125I-NGF крысы составляли 0,44 нМ для 1252A5 и 0,50 нМ для G1152H5.
ПРИМЕР 9
Определение величин Ki для ингибирования связывания NGF с рецепторами TrkA и p75 человека
Проводили эксперименты для определения, обладают ли антитела 1252A5 и G1152H5 различным ингибированием связывания NGF с рецепторами TrkA и p75. Проводили эксперименты по конкурентному связыванию, чтобы вычислить величины констант ингибирования связывания (Ki). IC50 вычисляли для опосредованного антителами ингибирования связывания радиоактивно меченного NGF человека или крысы со слитыми белками TrkA- или p75-рецепторов человека. Величины Ki затем вычисляли с использованием уравнения Cheng-Prusoff.
Слитые белки рецепторов TrkA и p75 человека (R&D Systems) разводили в PBS по Дульбекко (Gibco) до конечных концентраций 10 нМ и 0,6 нМ, соответственно. Белые 96-луночные планшеты для микротитрования Maxisorp Nunc (Nalge Nunc) покрывали в течение ночи при 4°C из расчета 100 мкг/лунка разведенным раствором рецептора TrkA или p75. Планшеты промывали 3 раза PBS-Tween 20 и затем блокировали 200 мкл/лунка 3% мас./об. бычьего сывороточного альбумина (BSA) в PBS. Планшеты промывали после 1 час инкубации при комнатной температуре. Тестируемые антитела разводили до требуемой концентрации в буфере для анализа (0,5% мас./об. BSA и 0,05% об./об. Tween 20 в PBS). Посторонние антитела, не нацеленные на NGF, применяли как отрицательный контроль, в то время как не радиоактивный NGF использовали как эталонный ингибитор связывания радиоактивно меченного лиганда. Использовали лунки в двух параллелях для каждой концентрации тестируемого образца. 125I-NGF человека или крысы (Amersham Biosciences) разводили буфером для анализа так, что конечная концентрация в анализируемых лунках, при смешивании с тестируемым образцом, составляла 150 пМ в общем анализируемом объеме 100 мкл. Анализируемые планшеты инкубировали при комнатной температуре 2 часа перед промывкой PBS/Tween 20 для удаления несвязанного 125I-NGF. Связанную радиоактивную метку количественно определяли добавлением 100 мкл/лунка Microscint 20 (Perkin Elmer) перед снятием показаний с использованием жидкостного сцинтилляционного счетчика для микропланшетов Packard TopCount. Данные наносили и анализировали с использованием программного обеспечения Graphpad Prism для вычисления величин IC50 для каждого эксперимента и выводили соответствующую Ki согласно уравнению Cheng-Prusoff:
Ki=IC 50 /(1+D/Kd)
где D = концентрация NGF в анализе (номинально 150 пМ, но фактические концентрации в анализе определяли для каждого эксперимента)
Kd = аффинность NGF к рецептору TrkA или p75 при одинаковых условиях анализа. Ее определяли анализом насыщаемого связывания для связывания 125I-NGF с рецепторами TrkA и p75 в индивидуальных экспериментах.
Все оцениваемые антитела, за исключением контрольного IgG4 человека, ингибировали связывание 125I-NGF человека и крысы с рецепторами TrkA и p75 человека (фиг.8 и 9; таблица 5). В таблице 6 приведены величины IC50 и Ki, вычисленные из данных, приведенных на фиг.8A; в таблице 7 приведены величины IC50 и Ki, вычисленные из данных, приведенных на фиг.8B; в таблице 8 приведены величины IC50 и Ki, вычисленные из данных, приведенных на фиг.9A; в таблице 9 приведены величины IC50 и Ki, вычисленные из данных, приведенных на фиг.9B. Антитела 1252A5 последовательно демонстрировали наибольшую способность ингибирования связывания 125I-NGF. Интересно, что величины констант ингибирования связывания, определенные для ингибирования связывания NGF с рецепторами TrkA и p75, опосредованного 1252A5, значимо отличались. Таким образом, вычисленное среднее значение pKi для ингибирования связывания NGF человека с TrkA и p75 составило 10,26±0,08 и 9,85±0,04, соответственно (P<0,01, критерий Стьюдента; оба n=3). Вычисленное среднее значение pKi для ингибирования связывания NGF крысы с рецепторами TrkA и p75 составило 9,79±0,04 и 9,55±0,03, соответственно (P<0,05, критерий Стьюдента; оба n=3). Данный результат позволяет предположить, что 1252A5 является предпочтительным ингибитором взаимодействия между NGF и рецептором TrkA, и неожиданно контрастирует с результатами, полученными для G1152H5, для которого отсутствовала значимая разница между соответствующими величинами pKi (таблица 5).
ПРИМЕР 10
Антигипералгезивная активность антител IgG4 человека к NGF
Антигипералгезивную активность антител к NGF оценивали на мышиной модели для индуцированной каррагинаном тепловой гиперчувствительности. Самцов мышей (20-25 г массы тела) вначале акклиматизировали в тестируемом устройстве 2 ч. На следующие сутки производили исходные измерения ответной реакции на тепловую стимуляцию обеих задних лап. Фокусируемый источник тепла прикладывали к подошвенной поверхности задних лап и регистрировали время до отдергивания согласно способу Hargreaves et al. (1988). Исходные величины вычисляли как среднее значение определений в трех параллелях для каждой лапы, зарегистрированные спустя 10 мин. Мыши затем получали внутрибрюшинную инъекцию NGF-нейтрализующих антител IgG4 человека или контрольных, совпадающих по изотипу, неактивных антител в фосфатно-буферном солевом растворе (PBS) как носителе. Спустя двадцать четыре часа вызывали воспалительную гипералгезию подподошвенной инъекцией каррагинана (2% мас./об. в PBS; объем инъекции 30 мкл). После дополнительного периода в 24 ч снова определяли задержки отдергивания для воспаленной и невоспаленной задних лап.
Тепловую гипералгезию, наблюдаемую 24 час после инъекции каррагинана, в зависимости от дозы ингибировала предварительная обработка мышей антителами IgG4 1252A5 человека к NGF (фиг.10).
Все цитированные документы приведены здесь в виде ссылки.
ССЫЛКИ
Al-Lazikani, et al. Journal Molecular Biology (1997) 273(4), 927-948 Aloe, L. and Tuveri, M.A. (1997) Clin. Exp. Rheumatol., 15(4):433-8.
Amann, R. and Schuligoi R. (2000) Neurosci Lett, 278(3); 173-6.
Andersen DC and Krummen L (2002) Current Opinion in Biotechnology 13:117.
Ausubel et al. eds., Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, John Wiley & Sons, 4th edition 1999.
Bagshawe K.D. et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4:915-922.
Barbas et al., 1994, Proc. Natl. Acad. Sci., USA, 91:3809-3813.
Bennett, D.L. et al. (1998) Eur. J Neurosci, 10(4):1282-91.
Bennett, D.L. (2001) Neuroscientist, 7(1):13-7.
Bergmann I. et al., Neurosci Lett., 255(2) 87-90, 1998.
Bird et al., Science, 242, 423-426, 1988;
de Castro, F. et al. (1998) Eur. J Neurosci, 10(1):146-52.
Chadd HE and Chamow SM (2001) Current Opinion in Biotechnology 12:188-194.
Cho, H.J. et al. (1996) Brain Res, 716(1-2):197-201.
Chothia, et al. Science, 223, 755-758 (1986).
Chothia С. et al. Journal Molecular Biology (1992) 227, 799-817.
Current Protocols in Molecular Biology, Second Edition, Ausubel et al. eds., John Wiley & Sons, 1988.
Denison David G. T. (Editor), Christopher C, Holmes, Bani K.
Mallick, Adrian F. M. Smith. Bayesian Methods for Nonlinear Classification and Regression (Wiley Series in Probability and Statistics). John Wiley & Sons; (July 2002), ISBN: 0471490369.
Fjell, J. et al. (1999) J Meurosci Res, 57(1):39-47.
Garaci, E. et al. (2003) Proc. Natl. Acad. Sci. USA, 100(15):8927-8932.
Ghose, Arup K. & Viswanadhan, Vellarkad N. Combinatorial Library Design and Evaluation Principles, Software, Tools, and Applications in Drug Discovery. ISBN: 0-8247-0487-8.
Gram et al., 1992, Proc. Natl. Acad. Sci., USA, 89:3576-3580.
Guex, N. and Peitsch, M.C. Electrophoresis (1997) 18, 2714-2723.
Haan & Maggos (2004) BioCentury, 12(5); A1-A6.
Hanes et al., Methods in Enzymology, 328:24, (2000).
Hargreaves et al., Pain, 32:77 (1988).
Heumann, R. et al. (1987) J Cell Biol., 104(6):1623-31.
Holliger, P. and Winter G. Current Opinion Biotechnol 4, 446-449, 1993.
Holliger, P. et al, Proc. Natl. Acad. Sci. USA 90 6444-6448, 1993.
Holt et al. (2003) Trends in Biotechnology 21, 484-490.
Hongo, J.S. et al. (2000) Hybridoma 19(3):215-227.
Hoyle, G.W, (2003) Cytokine Growth Factor Rev, 14(6); 551-8.
Hu, S. et al., Cancer Res., 56, 3055-3061, 1996.
Huang, E.J. and Reichardt, L. F. (2001) Ann. Rev. Neurosci., 24:677-736.
Huston et al., PNAS USA, 85, 5879-5883, 1988.
Hutchings, in Antibody Engineering, Konterman and Dubel eds., Springer, Berlin, pp.93-108 [2001].
Indo, Y. (2002) Clin Auton Res, 12 Suppl 1: I20-32.
Jagger, S.I. et al. (1999) Br J Anaesth, 83(3):442-8.
Kabat, E.A. et al, Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health and Human Services. 1987.
Kabat, E.A. et al. (1991) Sequences of Proteins of Immunological Interest, 5th Edition. US Department of Health and Human Services, Public Service, NIH, Washington.
Kandel, Abraham & Backer, Eric. Computer-Assisted Reasoning in Cluster Analysis. Prentice Hall PTR, (May 11, 1995), ISBN; 0133418847.
Kay, В.K., Winter, J., and McCafferty, J. (1996) Phage Display of Peptides and Proteins; A Laboratory Manual, San Diego: Academic Press.
Knappik et al. J. Mol. Biol. (2000) 296, 57-86.
Koide et al. (1998) Journal of Molecular Biology, 284:1141-1151.
Koltzenburg, M. et al. (1999) Eur. J Neurosci, 11(5):1698-704.
Kontermann, R & Dubel, S, Antibody Engineering, Springer-Verlag New York, LLC; 2001, ISBN: 3540413545.
Krebs et al. Journal of Immunological Methods 254, 2001, 67-84.
Krzanowski, Wojtek. Principles of Multivariate Analysis; A User's Perspective (Oxford Statistical Science Series, № 22 (Paper)). Oxford University Press; (December 2000), ISBN; 0198507089.
Larrick JW and Thomas DW (2001) Current Opinion in Biotechnology 12:411-418.
Ledermann J.A. et al. (1991) Int. J. Cancer 47:659-664.
Lommatzsch, M. et al. (2003) Ann. NY Acad. Sci., 992:241-9.
Lowe, E.M. et al. (1997) Br. J Urol., 79(4):572-577.
Ma, Q.P. and Woolf, C.J. (1997) Neuroreport, 8(4):807-10.
Mach et al., Analytical Biochemistry, 200:74, (1992).
Mamet, J. et al. (2003) J Biol. Chem., 278(49):48907-13.
Marks et al Bio/Technology, 1992, 10:779-783.
McArthur, J.C. et al. (2000) Neurology, 54(5):1080-8.
McCafferty et al (1990) Nature, 348, 552-554.
Mendell, L.M. and Arvanian, V.L. (2002) Brain. Res. Rev., 40(1-3):230-9.
Mendez, M. et al. (1997) Nature Genet, 15(2):146-156.
Nakagawara, A. (2001) Cancer Lett., 169(2):107-14.
Norman et al. Applied Regression Analysis. Wiley-Interscience; 3rd edition (April 1998) ISBN: 0471170828.
Nygren et al. (1997) Current Opinion in Structural Biology, 7:463-469.
Owolabi, J.B. et al. (1999) J Pharmacol. Exp. Ther., 289(3):1271-6.
Persic et al., Gene, 187:9, (1997).
Petty, B.G. et al. (1994) Ann Neurol, 36(2):244-6.
Pleuvry В. & Pleuvry A., (2000) Analgesia: Markets and Therapies, ISBN 1860674143.
Pluckthun, A. Bio/Technology 9:545-551 (1991).
Pozza, M., et al. (2000) J Rheumatol, 27(5):1121-7.
Priestley, J.V. et al. (2002) Can. J Physiol. Pharmacol., 80(5):495-505.
Qiao, L.y. and Vizzard, M.A. (2002) J Comp. Neurol., 454(2):200-11.
Ramer, M.S. et al. (1998) Neurosci Lett, 251(1):53-6.
Ramer, M.S. et al. (1999) Pain, Suppl 6: S111-20.
Reiter, Y. et al., Nature Biotech, 14, 1239-1245, 1996.
Ridgeway, J. В. В. et al., Protein Eng., 9, 616-621, 1996.
Ro, L.S. et al. (1999) Pain, 79(2-3):264-74.
Robinson, J. R. ed., Sustained and Controlled Release Drug Delivery Systems, Marcel Dekker, Inc., New York, 1978.
Sah, D.W. et al. (2003) Nat. Rev. Drug. Discov, 2(6):460-72.
Sambrook and Russell, Molecular Cloning: a Laboratory Manual: 3rd edition, 2001, Cold Spring Harbor Laboratory Press.
Sammons, M.J. et al. (2000) Brain Res, 876(1-2):48-54.
Schier et al., 1996, J. Mol. Biol. 263:551-567.
Stemmer, Nature, 1994, 370:389-391.
Vaughan et al., Nature Biotechnology 14, 309-314, 1996.
Voet & Voet, Biochemistry, 2nd Edition, (Wiley) 1995.
Ward, E.S. et al., Nature 341, 544-546 (1989).
Wess, L. In: BioCentury, The Bernstein Report on BioBusiness, 12(42), A1-A7, 2004.
Whitelegg, N.R.u. and Rees, A.R (2000). Prot. Eng., 12, 815-824.
Witten, Ian H. & Frank, Elbe. Data Mining: Practical Machine Learning Tools and Techniques with Java Implementations. Morgan Kaufmann; (October 11, 1999), ISBN; 1558605525.
Wold, et al. Multivariate data analysis in chemistry. Chemometrics - Mathematics and Statistics in Chemistry (Ed.; B. Kowalski), D. Reidel Publishing Company, Dordrecht, Holland, 1984 (ISBN 90-277-1846-6).
Woolf, C.J. (1996) Philos Trans R Soc. Lond В Biol. Sci., 351(1338):441-8.
Yasuda, H. et al. (2003) Prog. Neurobiol, 69(4):229-85.
Zhang YH, Nicol GD. Neurosci. Lett. 2004 Aug. 12:366 (2):187-92.
Нейтрализующая способность антител IgG4 человека к NGF в анализе мобилизации кальция

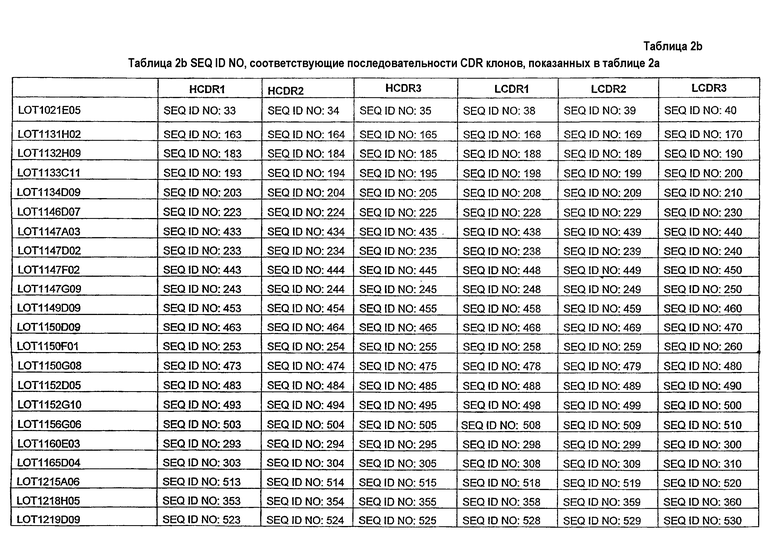
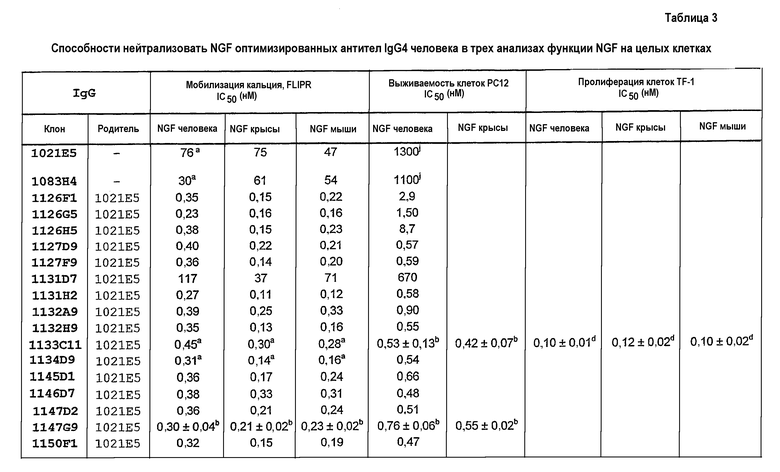
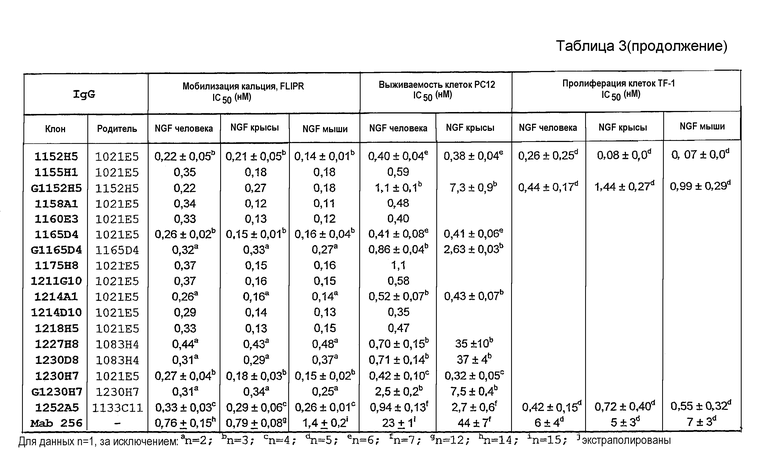
Перекрестная реактивность оптимизированных антител IgG4 человека к NGF с другими нейротрофинами
Данные в колонках представляют собой диапазон вычисленных перекрестных реактивностей антител. Величины вычислены как процентная доля от сигнала, наблюдаемого для каждого нейротрофина по сравнению с сигналом для связывания NGF, при одинаковой концентрации тестируемого антитела. Нейротрофинами покрывали анализируемые планшеты в концентрации 100 нг/лунка и связывание тестируемых антител измеряли в диапазоне концентраций 0,03-10 нМ. Данные представляют результат одиночного эксперимента.
Сводная таблица определений констант ингибирования связывания для 1252А5 и G1152H5. Данные представляют среднее значение ± стандт. ошибку средн. трех независимых экспериментов
** Р<0,05 для взаимодействия NGF/p75 крысы;
§ недостаточно для p75;
Непарный Т-критерий Стьюдента.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ АНТИТЕЛ ЧЕЛОВЕКА К IL-13 | 2004 |
|
RU2387667C2 |
| ХИМЕРНЫЙ БЕЛОК, СОСТАВЛЕННЫЙ ИЗ ДОМЕНА АНТАГОНИСТА NGF И ДОМЕНА АНТАГОНИСТА TNFα | 2015 |
|
RU2678810C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2644235C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2640252C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2788616C2 |
| АНТИТЕЛА ЧЕЛОВЕКА С ВЫСОКОЙ АФФИННОСТЬЮ К ФАКТОРУ РОСТА НЕРВОВ ЧЕЛОВЕКА | 2008 |
|
RU2473564C2 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ GDF-8 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2360925C2 |
| Антигены, ассоциированные с воспалительным заболеванием кишечника | 2012 |
|
RU2612878C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА РЕЦЕПТОРА С-МЕТ | 2011 |
|
RU2608644C2 |
| АНТИГЕН, АССОЦИИРОВАННЫЙ С РЕВМАТОИДНЫМ АРТРИТОМ | 2013 |
|
RU2649368C2 |








































































































































































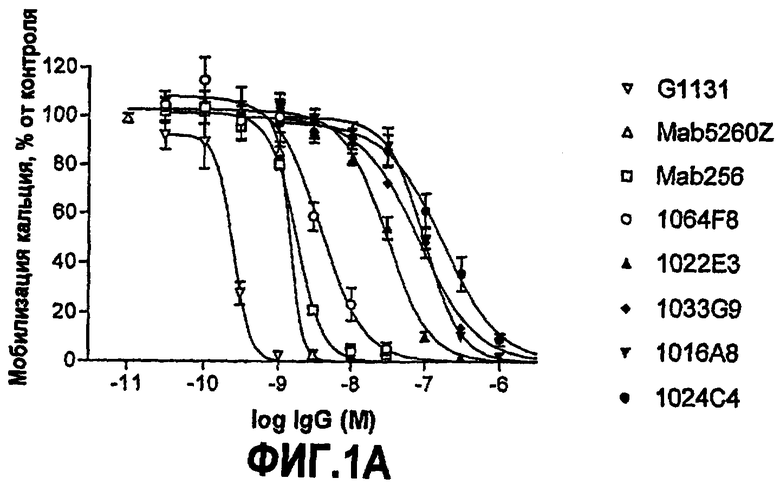
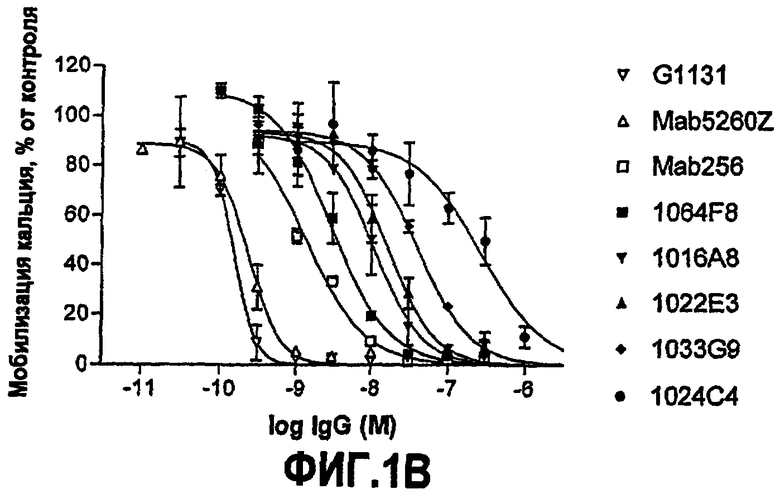
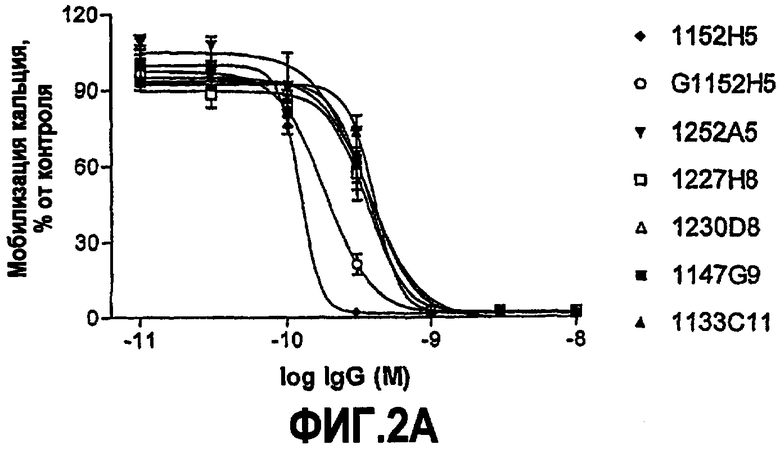
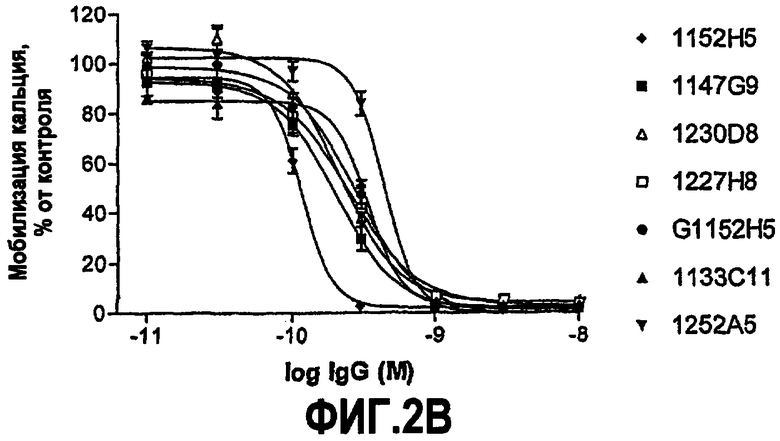
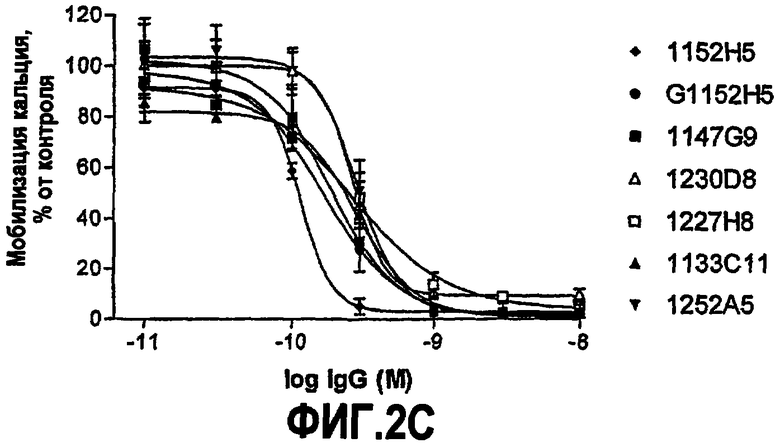
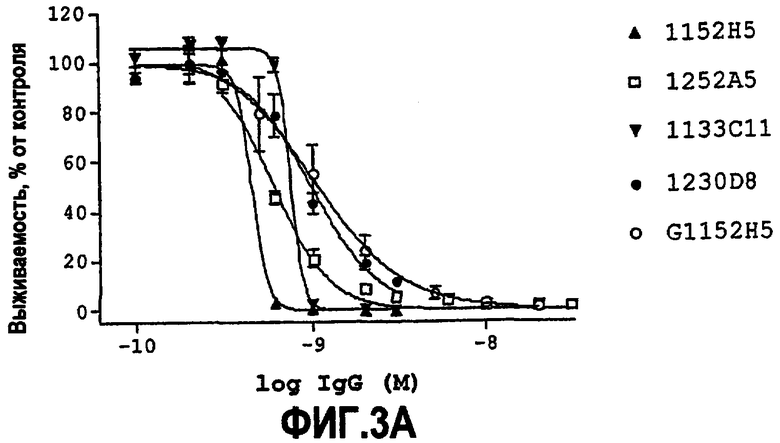
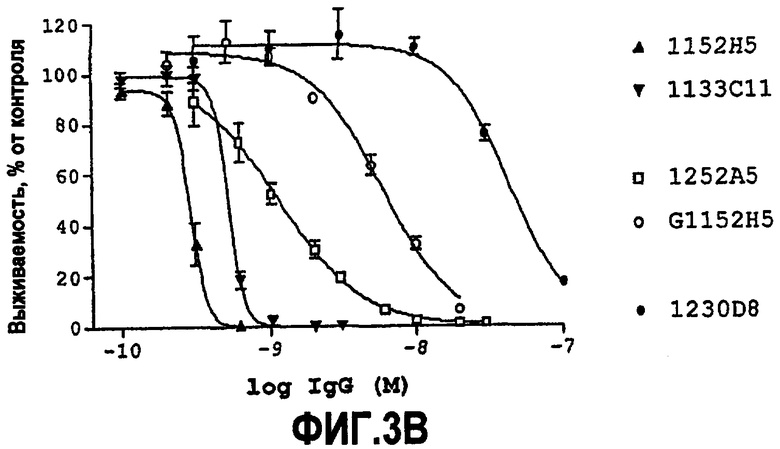
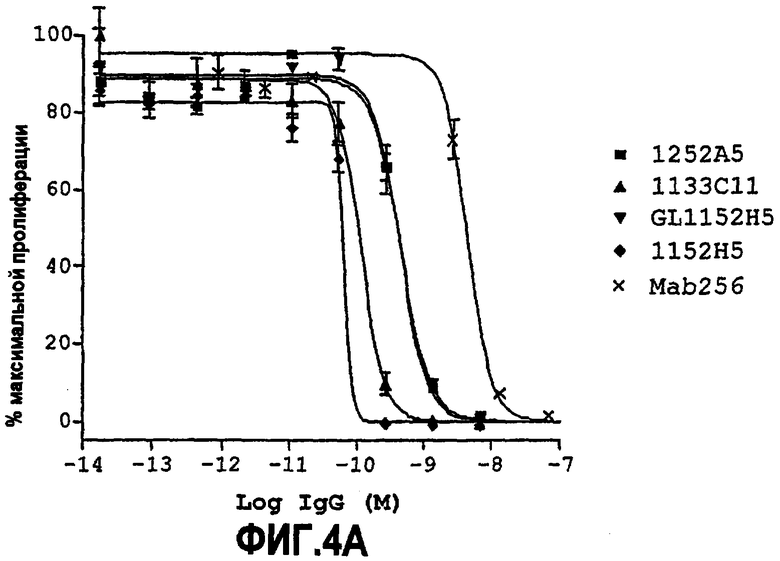
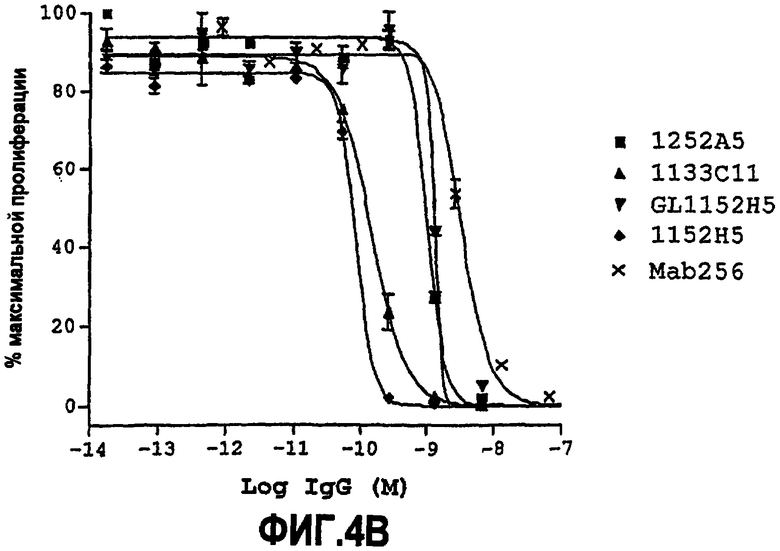
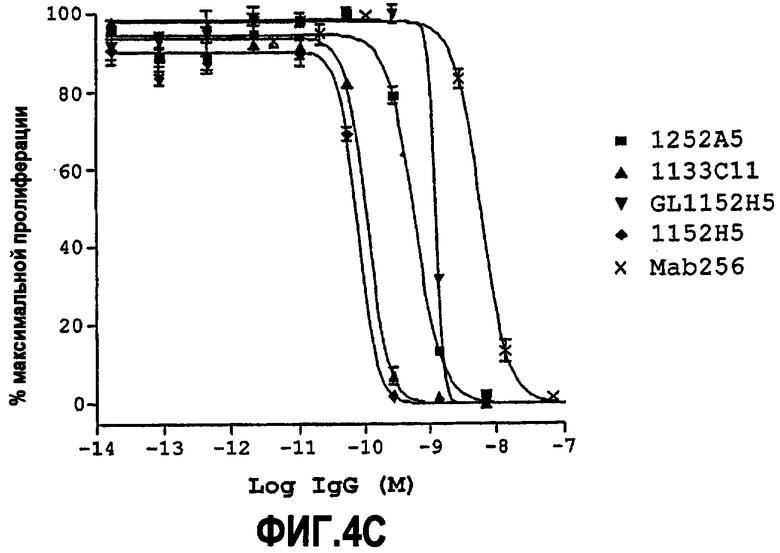
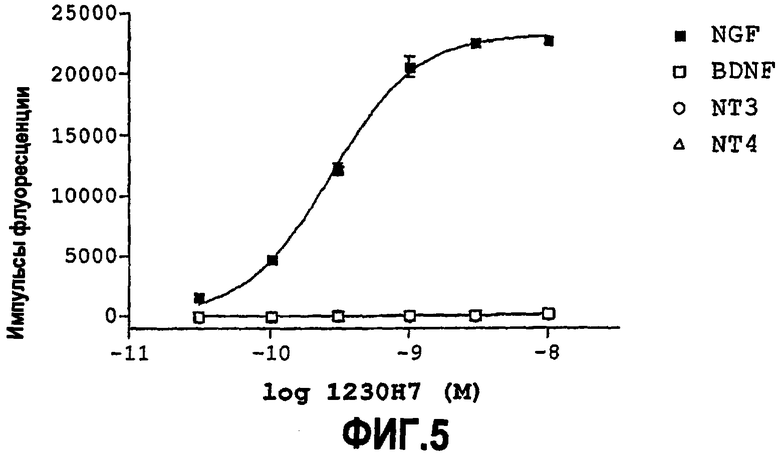
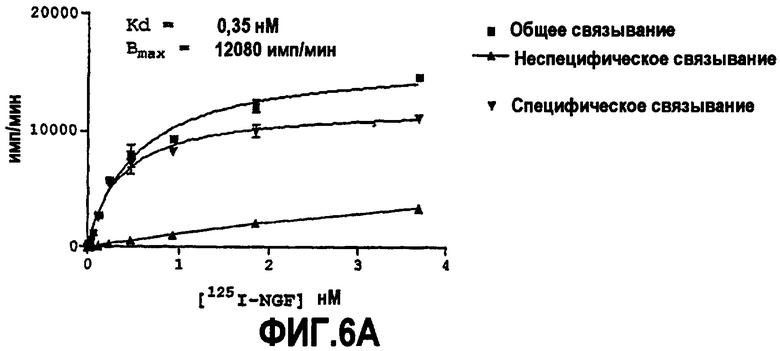
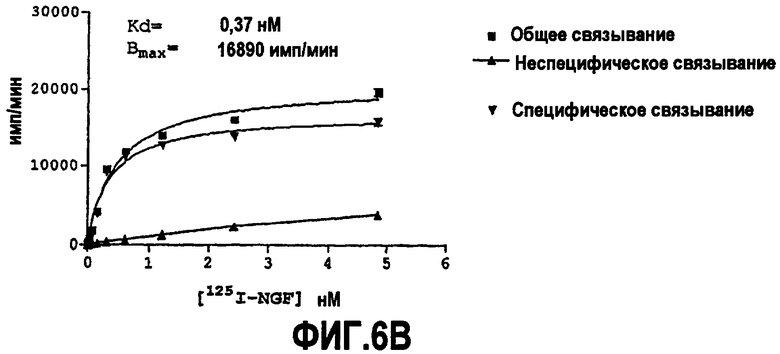
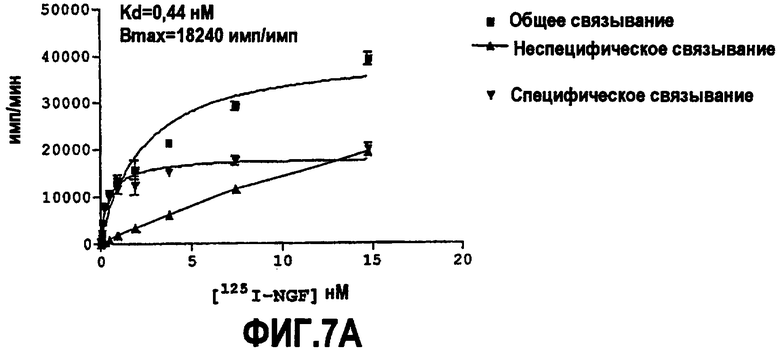
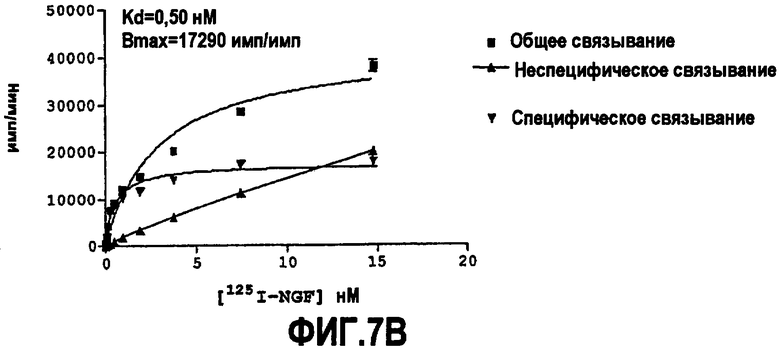
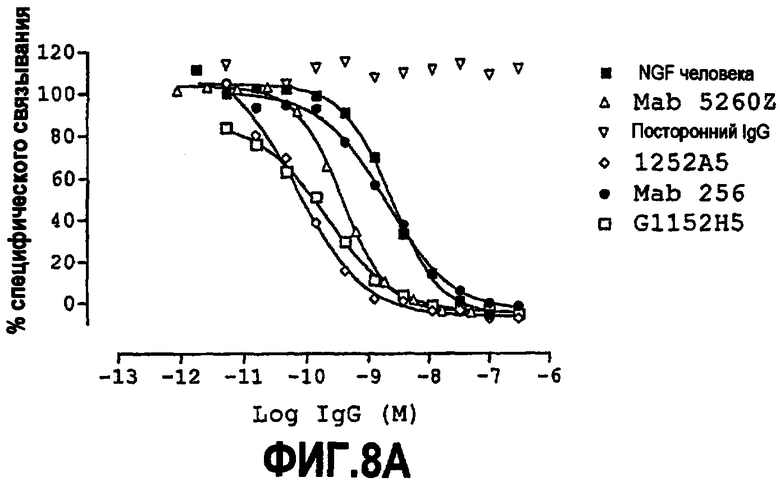
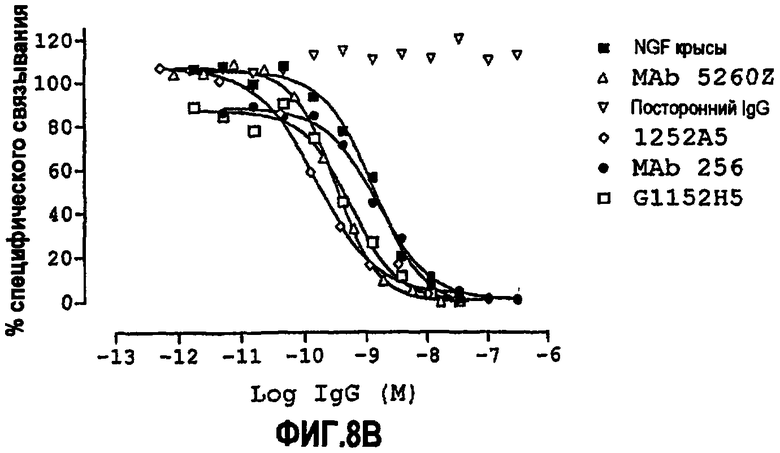
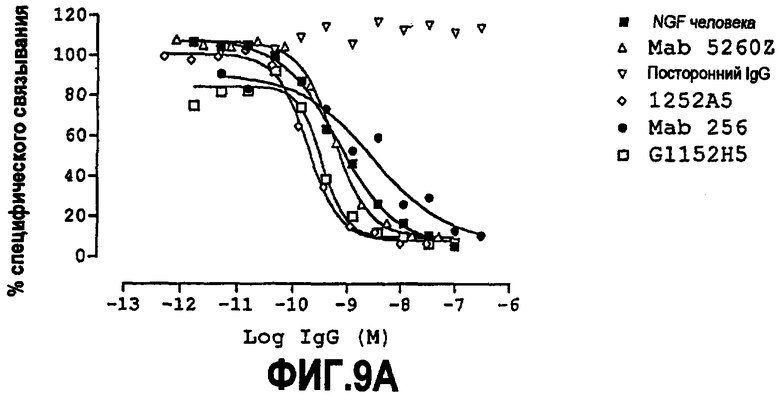
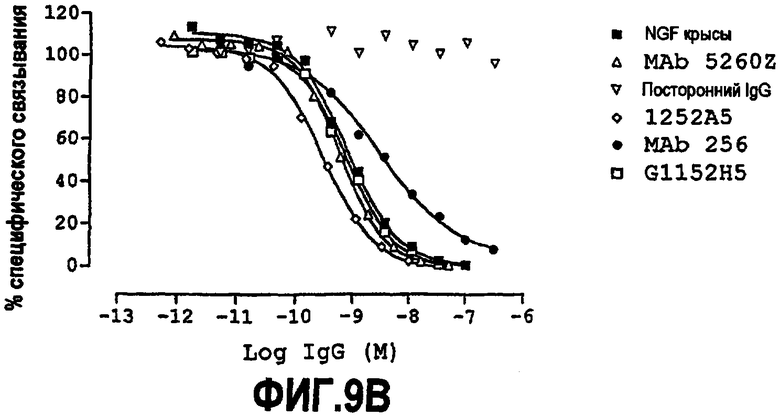
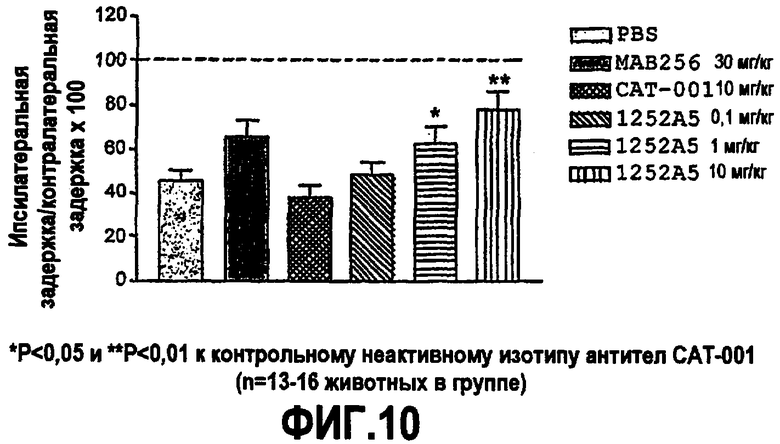
Изобретение относится к области биотехнологии и представляет собой партнеры специфического связывания с фактором роста нервов (NGF), в частности молекулы антител против NGF, в особенности, молекулы антитела человека и, в особенности, те, которые нейтрализуют активность NGF. Кроме того, изобретение относится к способам применения молекул антител против NGF. Изобретение можно эффективно использовать в диагностике или лечении расстройств, связанных с NGF, включая боль, астму, хроническое обструктивное заболевание легких, фиброз легких, другие воспалительные заболевания дыхательных путей, диабетическую невропатию, сердечные аритмии, HIV, артрит, псориаз и рак. 11 н. и 36 з.п. ф-лы, 10 ил., 9 табл.
1. Выделенный партнер специфического связывания фактора роста нервов (NGF) человека, содержащий антигенсвязывающий участок антитела, который состоит из VH-домена антител человека и VL-домена антитела человека и который содержит набор CDR HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, в котором VH-домен содержит HCDR1, HCDR2, HCDR3 и каркас и VL-домен содержит LCDR1, LCDR2, LCDR3 и каркас, где набор CDR состоит из набора CDR, выбранного из группы, состоящей из:
набора CDR 1133C11, который отличается тем, что HCDR1 имеет аминокислотную последовательность SEQ ID NO:193, HCDR2 имеет аминокислотную последовательность SEQ ID NO:194, HCDR3 имеет аминокислотную последовательность SEQ ID NO:195, LCDR1 имеет аминокислотную последовательность SEQ ID NO:198, LCDR2 имеет аминокислотную последовательность SEQ ID NO:199, и LCDR3 имеет аминокислотную последовательность SEQ ID NO:200;
набора CDR, который содержит одну или две аминокислотные замены, по сравнению с набором CDR 1133C11;
любого набора CDR, как показано для индивидуальных клонов в таблице 2; и
набора CDR, который содержит набор CDR 1133C11 с аминокислотной последовательностью SEQ ID NO:533 или аминокислотной последовательностью SEQ ID NO:532, замененными на аминокислотную последовательность SEQ ID NO:531 в HCDR3,
где одна или две замены остатков в CDR сделаны в следующих положениях
стандартной нумерации Kabat:
2. Выделенный партнер специфического связывания по п.1, в котором остаток 29 в LCDR1 является Е.
3. Выделенный партнер специфического связывания по п.1, содержащий набор CDR, который содержит набор CDR 1133C11 с аминокислотной последовательностью SEQ ID NO:533, замененной на аминокислотную последовательность SEQ ID NO:531 в HCDR3.
4. Выделенный партнер специфического связывания по п.1, содержащий набор CDR, который содержит набор CDR 1133C11 с аминокислотной последовательностью SEQ ID NO:532, замененной на аминокислотную последовательность SEQ ID NO:531 в HCDR3.
5. Выделенный партнер специфического связывания по п.1, содержащий набор CDR 1133C11.
6. Выделенный партнер специфического связывания по п.1, в котором каркас VH-домена представляет собой каркас зародышевой линии тяжелой цепи человека и/или каркас VL-домена представляет собой каркас зародышевой линии легкой цепи человека.
7. Выделенный партнер специфического связывания по п.6, в котором каркас зародышевой линии тяжелой цепи содержит VH1 DP10.
8. Выделенный партнер специфического связывания по п.6 или 7, в котором каркас зародышевой линии легкой цепи содержит VL Vλ1.
9. Выделенный партнер специфического связывания по п.1, который связывается с NGF крысы или мыши.
10. Выделенный партнер специфического связывания по п.1, который предпочтительно блокирует связывание NGF с рецептором TrkA по сравнению со связыванием NGF с рецептором р75.
11. Выделенный партнер специфического связывания по п.7, содержащий VH-домен 1252А5 с аминокислотной последовательностью SEQ ID NO:392.
12. Выделенный партнер специфического связывания по п.7, содержащий VL-домен 1252А5 с аминокислотной последовательностью SEQ ID NO:397.
13. Выделенный партнер специфического связывания по п.1, который связывается с NGF человека с аффинностью, равной или лучшей, чем аффинность антигенсвязывающего участка для NGF человека, сформированного VH-доменом 1252А5 с аминокислотной последовательностью SEQ ID NO:392 и VL-доменом 1252А5 с аминокислотной последовательностью SEQ ID NO:397, где аффинность партнера специфического связывания и аффинность антигенсвязывающего участка определены в одинаковых условиях.
14. Выделенный партнер специфического связывания по п.1, который связывает и/или нейтрализует NGF человека.
15. Выделенный партнер специфического связывания по п.14, который нейтрализует NGF человека, со способностью равной или лучшей, чем способность антигенсвязывающего участка для NGF, сформированного VH-доменом 1252А5 с аминокислотной последовательностью SEQ ID NO:392 и VL-доменом 1252А5 с аминокислотной последовательностью SEQ ID NO:397, где способность партнера специфического связывания и способность антигенсвязывающего участка определены в одинаковых условиях.
16. Выделенный партнер специфического связывания по п.7, содержащий VH-домен 1152Н5 с аминокислотной последовательностью SEQ ID NO:262.
17. Выделенный партнер специфического связывания по п.7 или 16, содержащий VL-домен 1152Н5 с аминокислотной последовательностью SEQ ID NO:267.
18. Выделенный партнер специфического связывания по п.7, содержащий VH-домен 1165D4 с аминокислотной последовательностью SEQ ID NO:302.
19. Выделенный партнер специфического связывания по п.7 или 18, содержащий VL-домен 1165D4 с аминокислотной последовательностью SEQ ID NO:307.
20. Выделенный партнер специфического связывания по п.7, содержащий VH-домен 1230Н7 с аминокислотной последовательностью SEQ ID NO:362.
21. Выделенный партнер специфического связывания по п.7 или 20, содержащий VL-домен 1230Н7 с аминокислотной последовательностью SEQ ID NO:367.
22. Выделенный партнер специфического связывания по п.1, который представляет собой молекулу антитела scFv.
23. Выделенный партнер специфического связывания по п.1, который содержит константную область антитела.
24. Выделенный партнер специфического связывания по п.23, который представляет собой целое антитело.
25. Выделенный партнер специфического связывания по п.24, в котором целое антитело представляет собой IgG4.
26. Партнер специфического связывания по любому из пп.1-25 для применения в лечении состояния, в котором принимает участие NGF, где состояние выбрано из группы, состоящей из боли, астмы, хронического обструктивного заболевания легких, фиброза легких, других воспалительных заболеваний дыхательных путей, диабетической невропатии, сердечных аритмий, HIV, артрита, псориаза и рака.
27. Партнер специфического связывания по п.26, где указанное лечение применяют в отношении боли.
28. Выделенная молекула антитела, которая связывает NGF человека, где молекула антитела содержит VH-домен и VL-домен, и где аминокислотная последовательность VH-домена представляет собой последовательность SEQ ID NO:392 и аминокислотная последовательность VL-домена представляет собой последовательность SEQ ID NO:397.
29. Выделенная молекула антитела по п.28, где молекула антитела представляет собой IgG4.
30. Молекула антитела по п.28 или 29 для применения в лечении состояния, в котором принимает участие NGF, где состояние выбрано из группы, состоящей из боли, астмы, хронического обструктивного заболевания легких, фиброза легких, других воспалительных заболеваний дыхательных путей, диабетической невропатии, сердечных аритмий, HIV, артрита, псориаза и рака.
31. Молекула антитела по п.30, где указанное лечение применяют в отношении боли.
32. Фармацевтическая композиция для связывания NGF человека, содержащая терапевтически эффективное количество партнера специфического связывания по любому из пп.1-25 или молекулу антитела по п.28 или 29, и, по меньшей мере, один дополнительный фармацевтически приемлемый эксципиент, основу или носитель, в котором партнер специфического связывания или молекула антитела способны связывать NGF.
33. Выделенная кодирующая нуклеиновая кислота, представляющая собой нуклеотидную последовательность, которая кодирует партнера специфического связывания по любому из пп.1-25 или молекулу антитела по п.28 или 29.
34. Клетка-хозяин, трансформированная in vitro нуклеиновой кислотой по п.33, экспрессирующая указанную нуклеиновую кислоту.
35. Клетка-хозяин, трансформированная in vitro нуклеиновой кислотой, включающей последовательность, кодирующую VH-домен партнера специфического связывания по любому из пп.1-25, или кодирующую VH-домен молекулы антитела по п.28 или 29, и нуклеиновой кислотой, включающей последовательность, кодирующую VL-домен партнера специфического связывания по любому из пп.1-25, или кодирующую VL-домен молекулы антитела по п.28 или 29, где клетка предназначена для экспрессии указанной нуклеиновой кислоты.
36. Способ получения партнера специфического связывания по любому из пп.1-25, или молекулы антитела по п.28 или 29, где способ включает культивирование клеток-хозяев по п.34 или 35 в условиях для продуцирования указанного партнера специфического связывания или молекулы антитела.
37. Способ по п.36, дополнительно включающий выделение и/или очистку указанного партнера специфического связывания или молекулы антитела.
38. Способ по п.36 или 37, дополнительно включающий составление партнера специфического связывания или молекулы антитела в композицию, содержащую, по меньшей мере, один дополнительный компонент.
39. Способ связывания NGF человека или его фрагмента, включающий контактирование и связывание партнера специфического связывания по любому из пп.1-25, или молекулы антитела по п.28 или 29 с NGF человека или фрагментом NGF человека.
40. Способ по п.39, в котором указанное связывание происходит in vitro.
41. Способ по п.39 или 40, включающий количественное определение связывания партнера специфического связывания или антитела с NGF или фрагментом NGF.
42. Применение партнера специфического связывания по любому из пп.1-25 для изготовления лекарственного средства для лечения состояния, в котором принимает участие NGF, где состояние выбрано из группы, состоящей из боли, астмы, хронического обструктивного заболевания легких, фиброза легких, других воспалительных заболеваний дыхательных путей, диабетической невропатии, сердечных аритмий, HIV, артрита, псориаза и рака.
43. Применение по п.42, где указанное лечение применяют в отношении боли.
44. Применение молекулы антитела по п.26 или 27 для изготовления лекарственного средства для лечения состояния, в котором принимает участие NGF, где состояние выбрано из группы, состоящей из боли, астмы, хронического обструктивного заболевания легких, фиброза легких, других воспалительных заболеваний дыхательных путей, диабетической невропатии, сердечных аритмий, HIV, артрита, псориаза и рака.
45. Применение по п.44, где указанное лечение применяют в отношении боли.
46. Способ лечения состояния, в котором принимает участие NGF, где способ включает введение партнера специфического связывания по любому из пп.1-25 или молекулы антитела по п.28 или 29 пациенту с состоянием или риском развития этого состояния, где состояние выбрано из группы, состоящей из боли, астмы, хронического обструктивного заболевания легких, фиброза легких, других воспалительных заболеваний дыхательных путей, диабетической невропатии, сердечных аритмий, HIV, артрита, псориаза и рака.
47. Способ по п.46, где указанное лечение применяют в отношении боли.
| WARREN S.L | |||
| ET AL | |||
| Inhibition of biological activity of mouse beta-nerve growth factor by monoclonal antibody | |||
| Science | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| OKISHIO M | |||
| ET AL | |||
| Establishment of monoclonal antibodies against human nerve growth factor | |||
| Biochem | |||
| Biophys | |||
| Res Commun | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| RO L.S | |||
| ET AL | |||
| Effect of NGF and anti-NGF on | |||

