Настоящее изобретение относится к специфическим связывающим партнерам, в частности к молекулам анти-IL-13-антител человека и особенно молекулам, которые нейтрализуют активность IL-13. Изобретение, кроме того, относится к способам применения молекул анти-IL-13-антител в диагностике или лечении связанных с IL-13 расстройств, включая астму, атопический дерматит, аллергический ринит, фиброз, воспалительное заболевание кишечника и лимфому Ходжкина.
В предпочтительных вариантах осуществления настоящего изобретения используют домен VH и/или VL антитела из молекулы антитела, называемого в данном описании BAK502G9, и других молекул антител линии BAK502G9 и линии BAK278D6, которые определены в данном описании. В следующих предпочтительных вариантах используют определяющие комплементарность области (CDR) линии BAK278D6 и предпочтительно BAK502G9, в частности CDR3 VH, в каркасных областях других антител. Следующие аспекты настоящего изобретения относятся к композициям, содержащим специфические связывающие партнеры согласно изобретению, и к их применениям в способах ингибирования или нейтрализации IL-13, включая способы терапевтического лечения организма человека или животного.
Настоящее изобретение относится к молекулам антител, имеющим особое значение в связывании и нейтрализации IL-13 и, следовательно, имеющим применение в любом из множества терапевтических способов лечения, как продемонстрировано в экспериментальной работе, приведенной в данной описании, и, кроме того, подтверждается технической литературой.
Интерлейкин (IL) 13 является цитокином из 114 аминокислот с немодифицированной молекулярной массой примерно 12 кД [1, 2]. IL-13 наиболее близок IL-4, с которым он обладает 30% сходством последовательностей на аминокислотном уровне. Ген IL-13 человека расположен в хромосоме 5q31 рядом с геном IL-4 [1], [2]. Указанная область хромосомы 5q содержит последовательности генов других, происходящих из лимфоцитов Th2 цитокинов, включая GM-CSF и IL-5, уровни которых вместе с IL-4, как показано, коррелируют с тяжестью заболевания у больных астмой и моделях аллергического воспаления у грызунов [3], [4], [5], [6], [7], [8].
Хотя IL-13 первоначально идентифицирован как производимый лимфоцитами Th2CD4+ цитокин, он также продуцируется T-клетками Th1CD4+, T-лимфоцитами CD8+ NK-клетками и популяциями клеток, не являющихся T-клетками, такими как тучные клетки, базофилы, эозинофилы, макрофаги, моноциты и клетки гладкой мускулатуры дыхательных путей.
Сообщается, что IL-13 опосредует свои эффекты через систему рецепторов, которая включает α-цепь рецептора IL-4 (IL-4Rα), которая сама по себе может связывать IL-4, но не IL-13, и, по меньшей мере, два других белка клеточной поверхности, IL-13Rα1 и IL-13Rα2 [9], [10]. IL-13Rα1 может связывать IL-13 с низкой аффинностью, затем мобилизируя IL-4Rα с образованием высокоаффинного функционального рецептора, который передает сигнал [11], [12]. В базе данных Genbank зарегистрирована аминокислотная последовательность и последовательность нуклеиновой кислоты IL-13Rα1 в виде NP_001551 и Y10659 соответственно. Исследования дефицитных по STAT6 (сигнальный трансдуктор и активатор транскрипции 6) мышей показали, что IL-13 сходным с IL-4 образом передает сигнал, используя путь JAK-STAT6 [13], [14]. IL-13Rα2 обладает 37% идентичностью последовательностей с IL-13Rα1 на аминокислотном уровне и связывает IL-13 с высокой аффинностью [15], [16]. Однако IL-13Rα2 имеет более короткий цитоплазматический хвост, в котором отсутствуют известные мотивы для передачи сигнала. Клетки, экспрессирующие IL-13Rα2, не отвечают на IL-13 даже в присутствии IL-4Rα [17]. Поэтому предполагается, что IL-13Rα2 действует в качестве рецептора-ловушки, регулирующего функционирование IL-13, но не IL-4. Это подтверждается исследованиями дефицитных по IL-13Rα2 мышей, фенотип которых соответствует повышенной чувствительности к IL-13 [18], [19]. В базе данных Genbank приведены аминокислотная последовательность и последовательность нуклеиновой кислоты IL-13Rα2 в виде NP_000631 и Y08768 соответственно.
Передающий сигналы комплекс рецепторов IL-13Rα1/IL-4Rα экспрессируется на B-клетках, тучных клетках, моноцитах/макрофагах, дендритных клетках, эозинофилах, базофилах, фибробластах, эндотелиальных клетках, эпителиальных клетках дыхательных путей и гладкомышечных клетках дыхательных путей человека.
Бронхиальная астма является распространенным персистирующим воспалительным заболеванием легких, характеризующимся гиперчувствительностью дыхательных путей, избыточным образованием слизи, фиброзом и повышенными уровнями IgE в сыворотке. Гиперчувствительность дыхательных путей (AHR) представляет собой чрезмерное сужение дыхательных путей в ответ на неспецифичные стимулы, такие как холодный воздух. Считают, что как AHR, так и избыточное образование слизи ответственны за непостоянную обструкцию дыхательных путей, которая приводит к затруднению дыхания, характерному для приступов астмы (обострений), и которая является причиной смертности, связанной с данным заболеванием (около 2000 смертей/год в Великобритании).
Заболеваемость астмой наряду с другими аллергическими заболеваниями значительно возросла в последние годы [20], [21]. Например, в настоящее время около 10% популяции в Великобритании диагностированы как больные астмой.
Современные рекомендации Британского торакального общества (BTS) и Глобальной инициативы против астмы (GINA) предлагают поэтапный подход к лечению астмы [22, 23]. Слабо и умеренно выраженную астму, как правило, можно контролировать, используя ингалируемые кортикостероиды в комбинации с бета-агонистами или ингибиторами лейкотриена. Однако из-за документально подтвержденных побочных эффектов кортикостероидов пациенты склонны не соблюдать режим лечения, что снижает эффективность лечения [24-26].
Существует очевидная потребность в новых способах лечения субъектов с более тяжелым заболеванием, которые часто получают очень ограниченную пользу либо от высоких доз ингалируемых, либо от пероральных кортикостероидов, предлагаемых в рекомендациях по астме. Долговременное лечение пероральными кортикостероидами связано с побочными эффектами, такими как остеопороз, замедленные скорости роста у детей, диабет и пероральный кандидоз [88]. Как целебные, так и неблагоприятные эффекты кортикостероидов опосредованы одним и тем же рецептором, и лечение представляет собой баланс между безопасностью и эффективностью. Госпитализация указанных пациентов, которые составляют около 6% популяции астматиков в Великобритании, в результате тяжелых обострений определяет основную часть значительного экономического бремени, связанного с астмой, для органов здравоохранения [89].
Предполагается, что патология астмы вызвана продолжающимся воспалением, опосредованным лимфоцитами Th2, которое возникает в результате неадекватных ответов иммунной системы на неопасные антигены. Накапливаются доказательства о вовлечении IL-13, а не классического образуемого клетками Th2 цитокина IL-4, в качестве ключевого медиатора патогенеза развившегося заболевания дыхательных путей.
Введение рекомбинантного IL-13 в дыхательные пути нативных несенсибилизированных грызунов вызывало множество признаков фенотипа астмы, включая воспаление дыхательных путей, образование слизи и AHR [27], [28], [29], [30]. Сходный фенотип наблюдали у трансгенной мыши, у которой IL-13 специфично сверхэкспрессировался в легких. В данной модели дополнительное воздействие IL-13 также приводило к фиброзу [31].
Кроме того, в моделях аллергического заболевания у грызунов многие признаки фенотипа астмы были связаны с IL-13. Показано, что мышиный IL-13Rα2, сильный нейтрализатор IL-13, ингибирует AHR, повышенное выделение слизи и приток воспалительных клеток, которые являются характерными признаками данной модели на грызунах [27], [28], [30]. В дополнительных исследованиях у мышей, у которых был делегирован ген IL-13, не развивалась индуцированная аллергеном AHR. AHR можно было восстановить у указанных дефицитных по IL-13 мышей введением рекомбинантного IL-13. Напротив, у мышей, дефицитных по IL-4, в данной модели развивалось заболевание дыхательных путей [32], [33].
Используя долговременную модель индуцированного аллергеном воспаления легких, Taube et al. продемонстрировали эффективность растворимого мышиного IL-13Rα2 против развившегося заболевания дыхательных путей [34]. Растворимый мышиный IL-13Rα2 ингибировал AHR, повышенное образование слизи и в меньшей степени воспаление дыхательных путей. Напротив, растворимый IL-4Rα, который связывает и антагонизирует IL-4, мало влиял на AHR или воспаление дыхательных путей в данной системе [35]. Указанные данные подтвердили Blease et al., которые разработали модель хронической астмы грибкового генеза, в которой поликлональные антитела против IL-13, но не против IL-4, были способны снижать повышенное образование слизи, AHR и субэпителиальный фиброз [36].
Ряд типов генетического полиморфизма в гене IL-13 также связывали с аллергическим заболеванием. В частности, вариант гена IL-13, в котором остаток аргинина в положении аминокислоты 130 заменен глутамином (R130Q), связывали с бронхиальной астмой, атопическим дерматитом и повышенными уровнями IgE в сыворотке [37], [38], [39], [40]. Указанный конкретный вариант IL-13 также назван вариантом Q110R (остаток аргинина в положении аминокислоты 110 заменен глутамином) некоторыми группами, которые исключили 20-аминокислотную сигнальную последовательность из нумерации аминокислот. Arima et al. [41] сообщают, что указанный вариант связан с повышенными уровнями IL-13 в сыворотке. Вариант IL-13 (R130Q) и антитела к данному варианту обсуждаются в WO 01/62933. Полиморфизм промотора IL-13, который изменяет продукцию IL-13, также связывали с аллергической астмой [42].
Повышенные уровни IL-13 также измеряли у людей с астмой, атопическим ринитом (сенной лихорадкой), аллергическим дерматитом (экземой) и хроническим синуситом. Например, обнаружено, что уровни IL-13 выше в биопсии бронхов, мокроте и клетках бронхоальвеолярного лаважа (BAL) у больных астмой по сравнению с контрольными субъектами [43], [44], [45], [46]. Кроме того, уровни IL-13 в образцах BAL повышались у больных астмой в случае провокации аллергеном [47], [48]. Кроме того, показано, что способность Т-клеток CD4(+) продуцировать IL-13 является полезным маркером риска последующего развития аллергического заболевания у новорожденных [49].
Li et al. [114] недавно сообщили о воздействиях нейтрализующего антитела против IL-13 мыши в модели хронической астмы у мышей. Хронический подобный астматическому ответ (такой как AHR, тяжелое воспаление дыхательных путей, повышенное образование слизи) индуцировали у сенсибилизированных OVA мышей. Li et al. сообщают, что введение IL-13-антитела во время каждой провокации OVA подавляет AHR, инфильтрацию эозинофилов, уровни IgE в сыворотке, уровни провоспалительных цитокинов/хемокинов и ремоделирование дыхательных путей [14].
В заключение, полученные данные представляют свидетельство того, что IL-13, а не IL-4 является более привлекательной мишенью для лечения аллергического заболевания человека.
IL-13 может играть роль в патогенезе воспалительного заболевания кишечника. Heller et al. [116] сообщают, что нейтрализация IL-13 введением растворимого IL-13Rα2 уменьшало воспаление ободочной кишки в мышиной модели язвенного колита человека [116]. Соответственно экспрессия IL-13 была выше в образцах биопсии прямой кишки от пациентов с язвенным колитом по сравнению с контролями [117].
Кроме астмы, IL-13 связывали с другими фиброзными состояниями. Повышенные уровни IL-13 до уровней в 1000 раз выше, чем IL-4, измерены в сыворотке пациентов с системным склерозом [50] и в образцах BAL от пациентов, пораженных другими формами легочного фиброза [51]. Соответственно сверхэкспрессия IL-13, но не IL-4, в легких мышей приводила к выраженному фиброзу [52], [53]. Вклад IL-13 в фиброз в других тканях, отличных от легких, широко исследован в мышиной модели индуцированного паразитами фиброза легких. Специфичное ингибирование IL-13 введением растворимого IL-13Rα2 или нарушением гена IL-13, но не исключение образования IL-4, предотвращало фиброгенез в печени [54], [55], [56].
Хроническое обструктивное легочное заболевание (COPD) охватывает популяции пациентов с различной степенью хронического бронхита, заболевания малых дыхательных путей и эмфиземы и характеризуется прогрессирующим необратимым снижением функции легких, которое плохо поддается современной терапии, основанной на лечении астмы [90]. Частота заболеваемости COPD сильно возросла в последние годы, и заболевание стало четвертым среди главных причин смертности во всем мире (Всемирная организация здравоохранения). Поэтому лечение COPD представляет собой большую неудовлетворенную потребность для медицины.
Причины, лежащие в основе COPD, остаются плохо понятными. «Датская гипотеза» предполагает, что существует общая чувствительность к COPD и астме и, следовательно, такие сходные механизмы могут вносить вклад в патогенез обоих расстройств [57].
Zheng et al. [58] показали, что сверхэкспрессия IL-13 в легких мыши вызывала эмфизему, повышенное образование слизи и воспаление, отражая признаки COPD человека. Кроме того, показано, что AHR, IL-13-зависимый ответ в мышиных моделях аллергического воспаления, является прогностическим признаком снижения функции легких у курильщиков [59]. Также была установлена связь между полиморфизмом промотора IL-13 и чувствительностью к развитию COPD [60].
Следовательно, имеются свидетельства того, что IL-13 играет важную роль в патогенезе COPD, особенно у пациентов с подобными астме признаками, включая AHR и эозинофилию. Показано, что уровни мРНК IL-13 выше в аутопсийных образцах тканей от субъектов с COPD в истории болезни по сравнению с образцами легких от субъектов, у которых не зарегистрировано легочное заболевание (J. Elias, Oral communication at American Thoracic Society Annual Meeting, 2002). В другом исследовании повышенные уровни IL-13 показаны посредством иммуногистохимии в периферических срезах легких от пациентов с COPD [91].
Болезнь Ходжкина является распространенным типом лимфомы, которая составляет примерно 7500 случаев в год в Соединенных Штатах. Болезнь Ходжкина является необычной среди злокачественных опухолей, так как неопластические клетки Рид-Штернберга, часто происходящие из B-клеток, составляют только небольшую часть клинически выявляемой массы. Полученные в случае болезни Ходжкина линии клеток и первичные клетки Рид-Штернберга часто экспрессируют IL-13 и его рецептор [61]. Так как IL-13 стимулирует жизнеспособность клеток и пролиферацию нормальных B-клеток, то сделано предположение, что IL-13 может действовать в качестве фактора роста для клеток Рид-Штернберга. Skinnider et al. показали, что нейтрализующие антитела против IL-13 могут ингибировать рост полученных в случае болезни Ходжкина линий клеток in vitro [62]. Указанные наблюдения свидетельствуют, что клетки Рид-Штернберга могут повышать свою собственную жизнеспособность посредством аутокринной и паракринной цитокиновой петли IL-13. В соответствии с данной гипотезой повышенные уровни IL-13 выявлены в сыворотке некоторых пациентов с болезнью Ходжкина по сравнению с нормальными контролями [63]. Поэтому ингибиторы IL-13 могут предотвращать прогрессирование заболевания в результате ингибирования пролиферации злокачественных клеток Рид-Штернберга.
Многие клетки злокачественной опухоли человека экспрессируют иммуногенные специфичные для опухоли антигены. Однако несмотря на то, что многие опухоли спонтанно регрессируют, некоторые избегают иммунной системы (иммунологического надзора), подавляя опосредованный T-клетками иммунитет. Terabe et al. [64] продемонстрировали роль IL-13 в иммуносупрессии в мышиной модели, в которой опухоли спонтанно регрессируют после начального роста и затем рецидивируют. Специфичное ингибирование IL-13 растворимым IL-13Rα2 защищало указанных мышей от рецидива возникновения опухоли. Terabe et al. [64] далее показали, что IL-13 подавляет дифференцировку специфичных для опухоли цитотоксических лимфоцитов CD8+, которые опосредуют противоопухолевые иммунные ответы.
Поэтому ингибиторы IL-13 могут быть использованы терапевтически для предотвращения рецидивов опухолей или метастазов. Показано, что ингибирование IL-13 усиливает противовирусные вакцины в животных моделях и может быть полезным при лечении ВИЧ и других инфекционных заболеваний [65].
Следует отметить, что, как правило, в данном описании указание интерлейкин-13 или IL-13 за исключением случаев, когда контекст диктует иное, относится к IL-13 человека. В некоторых случаях его также называют «антигеном». Настоящее изобретение относится к антителам к IL-13 человека, в частности антителам человека, которые перекрестно реагируют с IL-13 примата, отличного от человека, включая IL-13 макак-крабоедов и макак-резус. Антитела согласно некоторым вариантам осуществления настоящего изобретения распознают вариант IL-13, в котором остаток аргинина в положении аминокислоты 130 заменен глутамином. В других аспектах и вариантах осуществления настоящего изобретения предлагаются специфические связывающие партнеры против IL-13 мышей и крыс, в частности против мышиного IL-13.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана эффективность нейтрализации (% ингибирования) в случае BAK167A11 (заштрихованные квадраты) и его производного BAK615E3 (незаштрихованные квадраты) в виде scFv против 25 нг/мл IL-13 человека в анализе пролиферации клеток TF-1. Треугольники означают нерелевантный scFv. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.2 показана эффективность нейтрализации (% ингибирования) в случае BAK278D6 (заштрихованные квадраты) и его производного BAK502G9 (незаштрихованные квадраты) в виде scFv против 25 нг/мл IL-13 человека в анализе пролиферации клеток TF-1. Треугольники означают нерелевантный scFv. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.3 показана эффективность нейтрализации (% ингибирования) в случае BAK209B11 (заштрихованные квадраты) в виде scFv против 25 нг/мл мышиного IL-13 в анализе пролиферации клеток TF-1. Треугольники означают нерелевантный scFv. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.4 показана эффективность нейтрализации (% ингибирования) в случае BAK278D6 (заштрихованные квадраты) в виде scFv против IL-13 в анализе пролиферации клеток TF-1. Треугольники означают нерелевантный scFv. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.4A показана эффективность против 25 нг/мл IL-13 человека.
На фиг.4B показана эффективность против 25 нг/мл варианта IL-13 человека.
На фиг.4C показана эффективность против 50 нг/мл IL-13 примата, отличного от человека.
На фиг.5 показано сравнение эффективности анти-IL-13-антител человека в анализе пролиферации TF-1. Данные представляют собой средние значения эффективности нейтрализации с линиями стандартных ошибок на основании 5-7 экспериментов против 25 нг/мл IL-13 человека. Эффективность относительно коммерчески доступного антитела B-B13 оценивали статистически, осуществляя однофакторный ANOVA с использованием критерия Даннета. *P<0,05, **P<0,01 по сравнению с B-B13.
На фиг.6 показана эффективность нейтрализации (% ингибирования) в случае BAK502G9 (заштрихованные квадраты), BAK1167F2 (заштрихованные треугольники) и BAK1183H4 (заштрихованные перевернутые треугольники) в виде IgG4 человека против целевого IL-13 в анализе пролиферации клеток TF-1. Незаштрихованные треугольники означают нерелевантный IgG4. Данные представляют собой средние значения с линиями стандартных ошибок на основе трех отдельных экспериментов.
На фиг.6A показана эффективность против 25 нг/мл IL-13 человека.
На фиг.6B показана эффективность против 25 нг/мл варианта IL-13 человека.
На фиг.6C показана эффективность против 50 нг/мл IL-13 примата, отличного от человека.
На фиг.7 показана эффективность нейтрализации (% ингибирования) в случае BAK502G9 (заштрихованные квадраты), BAK1167F2 (заштрихованные треугольники), BAK1183H4 (заштрихованные перевернутые треугольники) в виде IgG4 человека и коммерческих анти-IL-13-антител человека (B-B13 - незаштрихованные квадраты; JES10-5A2 - незаштрихованные перевернутые треугольники) в анализе зависимой от нативного IL-13 пролиферации клеток HDLM-2. Незаштрихованные треугольники означают нерелевантный IgG4. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.8 показано сравнение эффективности анти-IL-13-антител человека в анализе NHLF. Данные представляют собой средние значения эффективности нейтрализации (IC50 пМ) с линиями стандартных ошибок на основании 4-5 экспериментов против 10 нг/мл IL-13 человека в анализе высвобождения эотаксина NHLF. Эффективность по сравнению с коммерчески доступным антителом B-B13 оценивали статистически, осуществляя однофакторный ANOVA с использованием критерия Даннета. *P<0,05, **P<0,01 по сравнению с B-B13.
На фиг.9 показана эффективность нейтрализации (% ингибирования) в случае BAK502G9 (заштрихованные квадраты), BAK1167F2 (заштрихованные треугольники), BAK1183H4 (заштрихованные перевернутые треугольники) в виде IgG4 человека против повышающей регуляции VCAM-1 на поверхности HUVEC в ответ на 10 нг/мл IL-13 человека. Незаштрихованные треугольники означают нерелевантный IgG4. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.10 показана эффективность нейтрализации (% ингибирования) в случае BAK502G9 (заштрихованные квадраты), BAK1167F2 (заштрихованные треугольники), BAK1183H4 (заштрихованные перевернутые треугольники) в виде IgG4 человека против высвобождения эотаксина из HUVEC с повышенной регуляцией VCAM-1 на поверхности в ответ либо на 1 нг/мл IL-4 человека (фиг.10A), либо на 0,5 нг/мл IL-1β человека (фиг.10B). Незаштрихованные треугольники означают нерелевантный IgG4. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.11 показана эффективность нейтрализации (% ингибирования) в случае BAK209B11 (квадраты) в виде IgG4 человека против 1 нг/мл мышиного IL-13 в анализе зависимой от факторов пролиферации клеток B9. Незаштрихованные треугольники означают нерелевантный IgG4. Данные представляют собой средние значения с линиями стандартных ошибок на основе определений в трех повторах в одном и том же эксперименте.
На фиг.12 показан относительный уровень IL-13 в гомогенатах легких от сенсибилизированных (s) (столбик справа) и несенсибилизированных (ns) (столбик слева) мышей после провокации в мышиной модели острого аллергического воспаления легких. Влияние сенсибилизации оценивали на основе t-критерия Стьюдента, используя количественные данные по IL-13. *<0,05, **<0,01 по сравнению с нечувствительными контрольными животными (n=5-6 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
Фиг.13 иллюстрирует влияние в/в введения BAK209B11 в виде IgG4 человека в разных количествах по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на индуцированную овальбумином мобилизацию лейкоцитов в легкие сенсибилизированных овальбумином мышей. Показано количество лейкоцитов (×104). Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные дифференцированного подсчета клеток. *<0,05, **<0,01 по сравнению с контрольными животными, которых подвергали провокации овальбумином и вводили PBS (=0% ингибирования; n=5-8 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
Фиг.14 иллюстрирует влияние в/в введения BAK209B11 в виде IgG4 человека в различных количествах по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на индуцированную овальбумином мобилизацию эозинофилов в легкие сенсибилизированных овальбумином мышей. Показано количество эозинофилов (×104). Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные дифференцированного подсчета клеток. *<0,05, **<0,01 по сравнению с контрольными животными, которых подвергали провокации овальбумином и вводили PBS (=0% ингибирования; n=5-8 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
Фиг.15 иллюстрирует влияние в/в введения BAK209B11 в виде IgG4 человека в различных количествах по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на индуцированную овальбумином мобилизацию нейтрофилов в легкие сенсибилизированных овальбумином мышей. Показано количество нейтрофилов (×104). Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные дифференцированного подсчета клеток. *<0,05, **<0,01 по сравнению с контрольными животными, которых подвергали провокации овальбумином и вводили PBS (=0% ингибирования; n=5-8 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
Фиг.16 иллюстрирует влияние в/в введения BAK209B11 в виде IgG4 человека в различных количествах по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на индуцированную овальбумином мобилизацию лимфоцитов в легкие сенсибилизированных овальбумином мышей. Индукция лимфоцитов зависимым от дозы образом ингибировалась BAK209B11 с максимальным ингибированием при 3 мкг/мл BAK209B11. Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные дифференцированного подсчета клеток. *<0,05, **<0,01 по сравнению с контрольными животными, которых подвергали провокации овальбумином и вводили PBS (=0% ингибирования; n=5-8 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
Фиг.17 иллюстрирует влияние в/в введения BAK209B11 в виде IgG4 человека в различных количествах по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на индуцированную овальбумином мобилизацию моноцитов/макрофагов в легкие сенсибилизированных овальбумином мышей. Не было значимого увеличения уровней моноцитов/макрофагов у сенсибилизированных животных по сравнению с контрольными животными. Однако такие фоновые уровни указанных клеток понижались при >36 мкг/мл BAK209B11 у сенсибилизированных животных. Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные дифференцированного подсчета клеток. *<0,05, **<0,01 по сравнению с контрольными животными, которых подвергали провокации овальбумином и вводили PBS (=0% ингибирования; n=5-8 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
На фиг.18 показано влияние коммерческого нейтрализующего анти-IL-13-антитела JES10-5A2 на приток клеток (показано количество лейкоцитов (×104)) к воздушному мешочку мыши, вызванный введением полученного в бактериях рекомбинантного IL-13 человека. Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные дифференцированного подсчета клеток. *<0,05, **<0,01 по сравнению с контрольными животными CMC (=0% ингибирования; n=11-13 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
На фиг.19 показано выравнивание последовательностей IL-13 макак-крабоедов по сравнению с аминокислотными последовательностями IL-13 человека. Заштрихованы семь аминокислотных остатков, которые различны в IL-13 человека и макак-крабоедов. IL-13 макак-резус и макак-крабоедов имели идентичную аминокислотную последовательность.
Фиг.20 иллюстрирует влияние однократной болюсной дозы 10 мг/кг в/в BAK502G9 в виде IgG4 человека на уровни IgE в сыворотке у 4 страдающих аллергией, но не подвергнутых провокации приматов макак-крабоедов (2 самца/2 самки) в течение 29 дней. Концентрация IgE в сыворотке значимо снижается со 100% (до введения дозы) до 66±10% от контрольных значений (p<0,05) на 4 и 5 день после введения дозы. Указанное снижение концентрации IgE в сыворотке восстанавливается до 88±8% от контрольных уровней на 22 день. *=p<0,05 по сравнению с уровнями IgE до введения дозы, многократные измерения ANOVA с последующим использованием критерия множественных сравнений Даннета (n=4 животных).
На фиг.20B показаны относительные уровни IgE в сыворотке самцов и самок приматов макак-крабоедов относительно времени после однократной внутривенной дозы 10 мг/кг BAK502G9. Относительные данные для IgE в сыворотке выражены в виде арифметической средней ± SEM в процентах от значения исходного уровня.
Фиг.21 иллюстрирует влияние внутрибрюшинного введения BAK209B11 в разных количествах (H=237 мкг/сутки, M=23,7 мкг/сутки и L=2,37 мкг/сутки) по сравнению с совпадающим по изотипу IgG1 нерелевантным контрольным антителом на функцию легких сенсибилизированных овальбумином и подвергнутых провокации мышей. На фиг.21A функция легких представлена log PC50 (log концентрации метахолина, требуемой для увеличения PenH исходного уровня на 50%) перед любой обработкой (0 день) и после сенсибилизации, провокации и обработки лекарственным средством (25 день). На фиг.21A показаны исходные данные, используемые для расчета конечного результата исследования, показанного на фиг.21B (разность log PC50). Данные представляют собой средние значения с линиями стандартных ошибок для n=8.
На фиг.21B изменение функции легких показано в виде изменения log PC50 отдельной мыши (разность log PC50). Разность log PC50 определяют как изменение log PC50 индивидуумов на 25 день по сравнению с 0 днем. Данные представляют собой средние для группы значения разности log PC50 (индивидуальные изменения, усредненные по группам обработки) с линиями стандартных ошибок. Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные о разности log PC50. **<0,01 по сравнению с сенсибилизированными овальбумином и подвергнутыми провокации контрольными животными (n=8 мышей).
Фиг.22 иллюстрирует влияние локального (i. po.) и системного (в/в) введения BAK502G9 в виде IgG4 человека в разных количествах по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на мобилизацию суммарных лейкоцитов (фиг.22A) и мобилизацию эозинофилов (фиг.22B) в воздушный мешочек мышей BALB/C. Данные представляют собой средние значения с линиями стандартных ошибок для n=10. Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя преобразованный в виде log данные. *<0,05, **<0,01 по сравнению с мышами, подвергнутыми провокации huIL-13 (n=10).
Фиг.23 иллюстрирует влияние в/б введения BAK502G9 в виде IgG4 человека по сравнению с совпадающим по изотипу IgG4 нерелевантным контрольным антителом на развитие AHR после внутритрахеального введения IL-13 человека в дыхательные пути мышей. Влияние обработки антителами оценивали статистически, осуществляя однофакторный ANOVA с применением критерия Даннета, используя данные для метахолина PC200. *<0,05, **<0,01 по сравнению с группой позитивного контроля с IL-13 человека (n=6-8 мышей). Данные представляют собой средние значения с линиями стандартных ошибок.
На фиг.24 показана эффективность нейтрализации (% максимального ответа) в случае BAK502G9 (заштрихованные квадраты) в виде IgG4 против 30 нг/мл IL-13 в анализе продуцирования IgE B-клетками человека. Незаштрихованные квадраты означают нерелевантный IgG4. Данные представляют собой средние значения с линиями стандартных ошибок для шести доноров из отдельных экспериментов.
На фиг.25 показано влияние BAK502G9 на индуцированное IL-13 усиление индуцированной агонистом передачи сигналов Ca2+ в гладкомышечных клетках бронхов. Площадь под кривой (AUC) ответа на гистамин в виде передачи сигнала Ca2+ определяли для каждого антитела +/- условия предварительной обработки IL-13. Объединенные данные из трех независимых экспериментов показаны для нерелевантного антитела CAT-001 (a) и BAK502G9 (b) в виде различий в процентах в AUC±SD с необработанными клетками (ns=не значимы (p>0,05), *p<0,05, **p<0,01). Результаты оценивали статистически, используя однофакторный дисперсионный анализ (ANOVA) и апостериорный критерий для множественных сравнений Бонферрони.
На фиг.26 показано влияние введенного в фазе II BAK502G9.
На фиг.26A показано влияние на AHR, измеренное по изменению площади под кривой дозовой зависимости ответа от гистамина (n=14).
На фиг.26B показано влияние на AHR, измеренное по изменению PC30 (n=18).
На фиг.26C показано влияние примирования антигеном (n=20).
На фиг.26D показано влияние воспаления BAL (n=21).
На фиг.27 показано влияние BAK502G9 на индуцированную IL-13 экспрессию CD23. Данные представляют ответ в процентах на IL-13 отдельно (100%) и выражены в виде среднего ± SEM % от контроля для 6 отдельных экспериментов от 6 отдельных доноров (выполнено в трех повторах).
На фиг.28 показано влияние BAK502G9 и нерелевантного IgG4 на индуцированную IL-13 и/или IL-4 экспрессию CD23 в PBMC. Данные представлены в виде ответа в процентах на IL-4 отдельно (100%) и выражены в виде среднего ± SEM % от контроля для 4 отдельных экспериментов от 4 отдельных доноров (выполнено в трех повторах).
На фиг.29A показано влияние BAK502G9 на продукцию эотаксина-1 NHLF, индуцированную 48-часовым культивированием в среде, содержащей IL-13/TNF-α/TGF-β1. Данные показаны в виде среднего арифметического ± SEM из определений в трех повторах для сред, используемых в данном исследовании для того, чтобы индуцировать изменение формы лейкоцитов.
На фиг.29B показано влияние BAK502G9 на изменение формы эозинофилов человека, индуцированное кондиционированными средами в разведении 1:16. Точки представленных данных означают средние ± SEM % по сравнению с изменением формы в контрольных средах для отдельных экспериментов от четырех отдельных доноров.
На фиг.30 показано выравнивание IL-13 человека и мышиного IL13 с отмеченными мутациями, которые вводили в IL-13 человека, чтобы получить первую панель химер IL-13. Четыре альфа-спирали выделены прямоугольниками и отмечены петля 1 и петля 3. Получали пять химерных белков, в которых спирали B, C и D и петлю 1 и петлю 3 заменяли мышиной последовательностью. Получены четыре дополнительных химерных белка и пронумерованы в соответствии с аминокислотами в пре-белке человека (но не с нумерацией множественного выравнивания, представленного выше), в которых аргинин в положении остатка 30 (положение 34 выше) мутирован, остатки 33 и 34 (положение 37 и 38 выше) мутированы, остатки 37 и 38 (VH) мутированы (положение 41 и 42 выше) и остатки 40 и 41 (TQ) мутированы (положение 44 и 45 выше).
На фиг.31 показано выравнивание IL-13 человека и мышиного IL-13 с отмеченными мутациями, которые вводили в IL-13 человека, чтобы получить вторую панель химер IL-13. Получали шесть химер, в которых остатком(ками) человека заменяли мышиный остаток(ки) (выделены прямоугольниками). Получены четыре следующих химерных белка (нумерация согласно положениям аминокислот в пре-белке человека), в которых лейцин в положении остатка 58 (62 на фигуре выше) мутирован, лейцин в положении остатка 119 (остаток 123 выше) мутирован, лизин в положении 123 (остаток 127 выше) мутирован и аргинин в положении остатка 127 (остаток 132 выше) мутирован.
На фиг.32 показаны мутации, сделанные в IL-13 человека. Мутации, обозначенные темно-серым, уменьшали связывание с BAK502G9, мутации, обозначенные светло-серым, не изменяли связывание. Показана линейная последовательность пре-IL-13 человека с мутантными остатками.
В различных аспектах и вариантах осуществления изобретения предлагаются предметы изобретения согласно формуле изобретения, приведенной далее.
Настоящее изобретение относится к специфическим партнерам, связывающим IL-13, в частности IL-13 человека, и/или примата, и/или вариант IL-13 (R130Q) и IL-13 мыши. Предпочтительными вариантами настоящего изобретения являются молекулы антител, либо полное антитело (например, IgG, такой как IgG4), либо фрагменты антител (например, scFv, Fab, dAb). Предлагаются антигенсвязывающие области антител, которые представляют собой домены VH и VL антител. В пределах доменов VH и VL предлагаются определяющие комплементарность области, CDR, которые могут быть представлены в разных каркасных областях, FR, чтобы образовать домены VH или VL в зависимости от обстоятельств. Антигенсвязывающий участок может состоять из домена VH и/или домена VL антитела.
Антигенсвязывающий участок может быть получен с помощью размещения CDR в каркасах белков, не являющихся антителами, таких как фибронектин или цитохром B и т.д. [115, 116]. Подробный обзор каркасов для конструирования новых связывающих участков в белках приведен в Nygren et al. [116]. Белковые каркасы для миметиков антител описаны в WO/0034784, где авторы изобретения описывают белки (миметики антител), которые содержат домен фибронектина типа III, имеющий, по меньшей мере, одну рандомизированную петлю. Подходящий каркас, в который прививают один или несколько CDR, например набор HCDR, может быть представлен любым представителем доменов надсемейства генов иммуноглобулинов.
Предпочтительными вариантами настоящего изобретения являются варианты, называемые в данном описании «линией BAK278D6». Линию определяют с указанием следующего набора последовательностей CDR BAK278D6: HCDR1 (SEQ ID NO: 1), HCDR2 (SEQ ID NO: 2), HCDR3 (SEQ ID NO: 3), LCDR1 (SEQ ID NO: 4), LCDR2 (SEQ ID NO: 5) и LCDR3 (SEQ ID NO: 6). В одном аспекте настоящее изобретение относится к специфическому партнеру, связывающему IL-13 человека, содержащему антигенсвязывающий участок антитела, который состоит из домена VH антитела человека и домена VL антитела человека и который содержит набор CDR, в котором домен VH содержит HCDR1, HCDR2 и HCDR3 и домен VL содержит LCDR1, LCDR2 и LCDR3, где HCDR1 имеет аминокислотную последовательность SEQ ID NO: 1, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 2, HCDR3 имеет аминокислотную последовательность SEQ ID NO: 3, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 4, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 5 и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 6; или в котором набор CDR содержит оду или две аминокислотных замены по сравнению с набором CDR, в котором HCDR1 имеет аминокислотную последовательность SEQ ID NO: 1, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 2, HCDR3 имеет аминокислотную последовательность SEQ ID NO: 3, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 4, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 5 и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 6.
Набор CDR, в котором HCDR1 имеет аминокислотную последовательность SEQ ID NO: 1, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 2, HCDR3 имеет аминокислотную последовательность SEQ ID NO: 3, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 4, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 5 и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 6, в данном описании называют «набором CDR BAK278D6». HCDR1, HCDR2 и HCDR3 в наборе CDR BAK278D6 называют «набором HCDR BAK278D6» и LCDR1, LCDR2 и LCDR3 в наборе CDR BAK278D6 называют «набором LCDR BAK278D6». Набор CDR с набором CDR BAK278D6, набором HCDR BAK278D6 или LCDR BAK278D6 или с одной или двумя заменами называют линией BAK278D6.
Как указано в одном аспекте, изобретение относится к партнеру, специфично связывающему IL-13 человека, содержащему антигенсвязывающий участок антитела, который состоит из домена VH антитела человека и домена VL антитела человека и который содержит набор CDR, при этом набор CDR представляет собой набор CDR BAK278D6 или набор CDR, содержащий одну или две замены по сравнению с набором CDR BAK278D6.
В предпочтительных вариантах одна или две замены находятся в одном или двух из следующих остатков в CDR доменов VH и/или VL, пронумерованных с использованием стандартной нумерации по Кабату [107].
31, 32, 34 в HCDR1
52, 52A, 53, 54, 56, 58, 60, 61, 62, 64, 65 в HCDR2
96, 97, 98, 99, 101 в HCDR3
26, 27, 28, 30, 31 в LCDR1
56 в LCDR2
95A, 97 в LCDR3
Предпочтительные варианты имеют две замены по сравнению с набором CDR BAK278D6, в HCDR3 остаток 99 и в LCDR1 остаток 27. Из указанных вариантов предпочтительные варианты имеют замену N на S в остатке 99 в HCDR3 и/или замену N на I в остатке 27 в LCDR1. Дополнительные варианты имеют замену в остатке 99 в HCDR3, выбранную из группы, состоящей из S, A, I, R, P и K, и/или замену в остатке 27 в LCDR1, выбранную из группы, состоящей из I, L, M, C, V, K, Y, F, R, T, S, A, H и G.
В предпочтительных вариантах одну или две замены осуществляют в одном или двух из следующих остатков в наборе CDR BAK278D6 в соответствии с известными группами остатков возможных заместителей:

Предпочтительные варианты имеют набор CDR BAK278D6 с заменой N на S в положении остатка 99 в HCDR3 и заменой N на I в положении остатка 27 в LCDR1. Таким образом, характеризуемый набор CDR представляет собой следующий набор: HCDR1 - SEQ ID NO: 7; HCDR2 - SEQ ID NO: 8, HCDR3 - SEQ ID NO: 9; LCDR1 - SEQ ID NO: 10, LCDR2 - SEQ ID NO: 11; LCDR3 - SEQ ID NO: 12. Указанный набор CDR в данном описании называют «набором CDR BAK502G9».
Следующие предпочтительные варианты имеют набор CDR BAK278D6 с одной или двумя заменами в CDR при условии, что исключена пара замен N на S в положении остатка 99 в HCDR3 и N на I в положении остатка 27 в LCDR1.
Другие предпочтительные варианты представляют собой следующее.
BAK1166G2: HCDR1 - SEQ ID NO: 67, HCDR2 - SEQ ID NO: 68, HCDR3 - SEQ ID NO: 69, LCDR1 - SEQ ID NO: 70, LCDR2 - SEQ ID NO: 71; LCDR3 - SEQ ID NO: 72.
BAK1167F2: HCDR1 - SEQ ID NO: 61, HCDR2 - SEQ ID NO: 62, HCDR3 - SEQ ID NO: 63, LCDR1 - SEQ ID NO: 64, LCDR2 - SEQ ID NO: 65; LCDR3 - SEQ ID NO: 66.
BAK1184C8: HCDR1 - SEQ ID NO: 73, HCDR2: SEQ ID NO: 74, HCDR3 - SEQ ID NO: 75. LCDR1 - SEQ ID NO: 76, LCDR2 - SEQ ID NO: 77; LCDR3 - SEQ ID NO: 78.
BAK1185E1: HCDR1 - SEQ ID NO: 79, HCDR2 - SEQ ID NO: 80, HCDR3 - SEQ ID NO: 81. LCDR1 - SEQ ID NO: 82, LCDR2- SEQ ID NO: 83; LCDR3 - SEQ ID NO: 84.
BAK1167F4: HCDR1 - SEQ ID NO: 85, HCDR2 - SEQ ID NO: 86, HCDR3 - SEQ ID NO: 87. LCDR1 - SEQ ID NO: 88, LCDR2 - SEQ ID NO: 89; LCDR3 - SEQ ID NO: 90.
BAK1111D10: HCDR1 - SEQ ID NO: 91, HCDR2-SEQ ID NO: 92, HCDR3-SEQ ID NO: 93. LCDR1 - SEQ ID NO: 94, LCDR2 - SEQ ID NO: 95; LCDR3 - SEQ ID NO: 96.
BAK1183H4: HCDR1 -SEQ ID NO: 97, HCDR2 - SEQ ID NO: 98, HCDR3 - SEQ ID NO: 99. LCDR1 - SEQ ID NO: 100, LCDR2 - SEQ ID NO: 101; LCDR3 - SEQ ID NO: 102.
BAK1185F8: HCDR1 - SEQ ID NO: 103, HCDR2 - SEQ ID NO: 104, HCDR3 - SEQ ID NO: 105. LCDR1- SEQ ID NO: 106, LCDR2 - SEQ ID NO: 107; LCDR3 - SEQ ID NO: 108.
Все указанные варианты получены из BAK502G9 рандомизацией CDR1 и CDR2 тяжелой цепи и, следовательно, являются линией BAK502G9.
Домен VH содержит набор CDR HCDR1, HCDR2 и HCDR3 любого клона, которые указаны в таблице 1. В данном изобретении в таблице 1 также представлен отдельно домен VL, содержащий набор CDR LCDR1, LCDR2 и LCDR3 клонов, показанных в таблице 1. Предпочтительно указанный домен VH образует пару с указанным доменом VL, и наиболее предпочтительно пары доменов VH и VL такие же, как в случае клонов, которые указаны в таблице 1.
Кроме того, в настоящем изобретении предлагается домен VH, содержащий набор CDR HCDR1, HCDR2 и HCDR3, при этом набор CDR соответствует набору любого клона, показанного в таблице 1, с одной или двумя аминокислотными заменами.
Кроме того, в настоящем изобретении предлагается домен VL, содержащий набор CDR LCDR1, LCDR2 и LCDR3, при этом набор CDR соответствует набору любого клона, показанного в таблице 1, с одной или двумя аминокислотными заменами.
Также в настоящем изобретении предлагается специфический связывающий партнер, содержащий антигенсвязывающий домен антитела, содержащий такой VH- и/или VL-домен.
Авторы настоящего изобретения идентифицировали линию BAK278D6 как обеспечивающую антигенсвязывающие домены антител человека против IL-13, которые являются особенно полезными. В линии идентифицировали BAK502G9 как особенно полезные. Наборы CDR BAK278D6 и BAK502G9 уже идентифицированы выше.
Следуя инструкции в области вычислительной химии по применению способов многофакторного анализа данных относительно взаимосвязи структура/свойство-активность [94], можно количественно оценить взаимосвязи активность-свойство для антител, используя хорошо известные математические способы, такие как статистическая регрессия, распознавание и идентификация образов [95-100]. Свойства антител могут быть получены на основе эмпирических и теоретических моделей (например, анализ вероятно контактирующих остатков или рассчитанное физико-химическое свойство) последовательности антител, при этом функциональные и трехмерные структуры и указанные свойства можно рассматривать отдельно и в комбинации.
Антигенсвязывающий участок антитела, состоящий из домена VH и домена VL, образуется шестью петлями полипептида: три из вариабельного домена легкой цепи (VL) и три из вариабельного домена тяжелой цепи (VH). С помощью анализа антител с известной атомарной структурой выяснены взаимосвязи между последовательностью и трехмерной структурой связывающих участков антитела [101, 102]. Указанные взаимосвязи свидетельствуют, что за исключением третьей области (петли) в доменах VH петли связывающих участков имеют одну из небольшого количества конформаций основной цепи: канонических структур. Показано, что каноническая структура, образованная в конкретной петле, определяется ее размером и наличием определенных остатков в ключевых участках как в петле, так и в областях каркаса [101, 102].
Указанное исследование взаимосвязи последовательность-структура может быть использовано для предсказания тех остатков в антителе с известной последовательностью, но неизвестной трехмерной структурой, которые важны для поддержания трехмерной структуры его петель CDR и, следовательно, сохраняют специфичность связывания. Указанные предсказания могут подтверждаться сравнением предсказаний с результатом приведенных в качестве примеров экспериментов по оптимизации.
В случае структурного подхода может быть создана модель молекулы антитела [103] с использование легкодоступного или коммерческого пакета программ, такого как WAM [104]. Затем можно использовать пакет компьютерных программ для визуализации и анализа белка, такой как Insight II [105] или Deep View [106], чтобы оценить возможные замены в каждом положении в CDR. Затем полученную информацию можно использовать для получения замен, которые, вероятно, оказывают минимальное или полезное влияние на активность.
Авторы настоящего изобретения анализировали данные о последовательности для панели клонов, наборы CDR для которых показаны таблице 1.
В анализе проверяли гипотезу о том, что любая из двойных комбинаций перечисленных аминокислотных изменений в CDR из представленного набора вариантов scFv приводит к варианту scFv, по меньшей мере, с начальной эффективностью исходного scFv BAK278D6.
Все варианты scFv на панели, показанной в таблице 1, были отобраны в отношении повышенной аффинности, и было подтверждено, что они проявляют более высокую эффективность.
Наблюдаемые аминокислотные изменения могут быть либо благоприятными, неблагоприятными, либо нейтральными по их влиянию на исходную эффективность scFv BAK278D6 в анализе TF-1 при 44 нМ.
Не наблюдали связи между любыми двумя аминокислотными изменениями, что подтверждало отсутствие синергии как «положительной», так и «отрицательной», между любыми двумя выбранными изменениями.
Существует четыре сценария, при которых такая двойная комбинация удовлетворяет гипотезе, и три сценария, при которых гипотеза будет неправомерна. Синергические аминокислотные варианты не рассматриваются, так как связи не наблюдалось.
Гипотеза правомерна в том случае, когда:
A1: мутация 1 является благоприятной и мутация 2 является благоприятной,
A2: мутация 1 является благоприятной, а мутация 2 является нейтральной,
A3: мутация 1 является нейтральной и мутация 2 является нейтральной,
A4: мутация 1 является благоприятной, а мутация 2 является неблагоприятной (при этом влияние 1 превосходит влияние 2).
Гипотеза неправомерна в том случае, когда:
B1: мутация 1 является неблагоприятной, а мутация 2 является нейтральной,
B2: мутация 1 является неблагоприятной и мутация 2 является неблагоприятной,
B3: мутация 1 является благоприятной, а мутация 2 является неблагоприятной (при этом влияние 2 превосходит влияние 1).
Для того чтобы была возможна ситуация A4, мутация 1 должна быть в высокой степени благоприятной, чтобы уравновесить отрицательное влияние мутации 2 на эффективность. Так как такая в высокой степени благоприятная мутация может присутствовать в библиотеке вариантов, используемых для отбора, она может быть отобрана и поэтому может часто встречаться на панели вариантов. Поскольку синергию можно исключить, такая мутация может быть полезной в любом контексте последовательностей и поэтому должна повторно встречаться в разных вариантах scFv. Примером такой частой замены аминокислоты является замена в CDR1 легкой цепи Asn27Ile. Однако данная мутация сама по себе (в клоне BAK531E2) оказывает только умеренное 2-кратное влияние на эффективность (конечная IC50 23,2 нМ). Сама по себе такая мутация не могла бы обеспечить возможность сценария, изображенного в A4, так как она на является в высокой степени благоприятной мутацией. Это означает, что каждый клон в представленном наборе связывающих IL-13 клонов (таблица 1), который имеет замену Asn27Ile в CDR1 легкой цепи вместе с одной или несколькими дополнительными мутациями, является, по меньшей мере, таким же эффективным, как вариант, имеющий одну мутацию Asn27Ile в CDR1 легкой цепи. Другие мутации являются либо нейтральными, либо положительными, но не оказывают отрицательного или вредного влияния.
Следующим примером является Asn99Ser в CDR3 тяжелой цепи (смотри таблицу 1). Поскольку клон, несущий указанное конкретное изменение одной аминокислоты, не был выявлен, то эффективность такого клона оценили как 12,0 нМ по следующей причине:
Эффективность BAK278D6 составляет 44 нМ. Изменения N27I CDR1 VL+N99S CDR3 VH приводит к BAK502G9 с эффективностью 8 нМ, т.е. 5,5-кратному улучшению.
Эффективность BAK278D6 составляет 44 нМ. Изменение N27I CDR1 VL приводит к BAK531E2 с эффективностью 23 нМ, т.е. к 1,9-кратному улучшению.
Эффективность BAK278D6 составляет 44 нМ. Изменение N99S CDR3 VH дает возможный клон с эффективностью 12,2 нМ, т.е. 2,9-кратное улучшение (5,5/1,9=2,9).
Двойная комбинация Asn99Ser CDR3 тяжелой цепи с Asn27Ile CDR1 легкой цепи дает scFv BAK0502G9 с эффективностью 8 нМ. Так как синергия исключается, то, следовательно, вклад замены Asn99Ser в CDR3 тяжелой цепи в BAK502G9 является аддитивным.
Следовательно, каждый клон в представленном наборе связывающих IL-13 клонов (таблица 1), который имеет замену AsnH99Ser в CDR3 тяжелой цепи вместе с одной или несколькими дополнительными мутациями, должен обладать эффективностью, составляющей, по меньшей мере, 12 нМ или больше, в 2,5-кратном разрешенном интервале анализа для n=1-2.
Таким образом, авторы изобретения отмечают, что в высокой степени благоприятное аминокислотное изменение, которое можно было бы предпочтительно выбрать, не наблюдается. Как обсуждалось выше, два изменения, которые выделяются в таблице 1 вариантов scFv, анализировали более тщательно. Любой вариант scFv в таблице 1 с любой из указанных мутаций вместе с одной или несколькими дополнительными мутациями проявлял эффективность, которая была настолько же улучшена, как и в случае клона, содержащего любую одну из указанных двух отдельных аминокислотных изменений в исходном BAK278D6. Поэтому нет свидетельства того, что в высокой степени благоприятное аминокислотное изменение, которое могло бы обеспечить возможность сценария A4, присутствует на панели.
Указанное наблюдение привело авторов изобретения к выводу, что в данном наборе вариантов scFv не присутствуют неблагоприятные мутации. Это означает, что сценарии A4 и B1-B3 не подходят и гипотеза правомерна.
Соответственно, как уже отмечалось, настоящее изобретение относится к специфически связывающим партнерам, содержащим определенные наборы CDR, в частности набор CDR BAK278D6, и наборы CDR линии BAK278D6 с одной или двумя заменами в наборе CDR, например с набором CDR BAK502G9.
Предлагается подходящий набор CDR в каркасных областях антитела или другом белковом каркасе, например в фибронектине или цитохроме B [115, 116]. Предпочтительно используют каркасные области антител, и в случае их использования они предпочтительно представляют собой каркасные области зародышевой линии, более предпочтительно каркасной областью антитела для тяжелой цепи может быть DP14 из семейства VH1. Предпочтительной каркасной областью для легкой цепи может быть λ3-3H. Для набора CDR BAK502G9 предпочтительно, чтобы каркасные области антитела представляли собой для FR1 VH SEQ ID NO: 27, для FR2 VH SEQ ID NO: 28, для FR3 VH SEQ ID NO 29, для FR1 легкой цепи SEQ ID NO: 30, для FR2 легкой цепи SEQ ID NO: 31, для FR3 легкой цепи SEQ ID NO: 32. В высокопредпочтительном варианте предлагается домен VH с аминокислотной последовательностью SEQ ID NO: 15, его называют «доменом VH BAK502G9». В следующем высокопредпочтительном варианте предлагается домен VL с аминокислотной последовательностью SEQ ID NO: 16, его называют «доменом VL BAK502G9». Высокопредпочтительный антигенсвязывающий участок антитела, предлагаемый согласно настоящему изобретению, состоит из домена VH BAK502G9, SEQ ID NO: 15, и домена VL BAK502G9, SEQ ID NO: 16. Указанный антигенсвязывающий участок антитела может быть представлен в любой требуемой форме молекулы, например scFv, Fab, IgG, IgG4, dAb и т.д., которые дополнительно обсуждаются в данном описании.
В следующем высокопредпочтительном варианте настоящее изобретение относится к молекуле антитела IgG4, содержащей домен VH BAK502G9, SEQ ID NO: 15, и домен VL BAK502G9, SEQ ID NO: 16. Молекулу называют в данном описании «IgG4 BAK502G9».
В настоящем изобретении предлагается другой IgG4 или другие молекулы антител, содержащие домен VH BAK502G9, SEQ ID NO: 15, и/или домен VL BAK502G9, SEQ ID NO: 16, которые представляют собой другие молекулы антител, содержащие набор HCDR BAK502G9 (SEQ ID NO: 7, 8 и 9) в VH-домене антитела и/или набор LCDR BAK502G9 (SEQ ID NO: 10, 11 и 12) в VL-домене антитела.
Следует указать, что «и/или» в случае использования в данном описании необходимо воспринимать как конкретное описание каждого из двух конкретных признаков или компонентов вместе друг с другом или отдельно. Например, «A и/или B» следует воспринимать как конкретное описание каждого варианта (i) A, (ii) B и (iii) A и B так же, как в случае, когда каждый отдельно указан в данном описании.
Как указано, настоящее изобретение относится к специфическому связывающему партнеру, который связывает IL-13 человека и который содержит VH-домен BAK502G9 (SEQ ID NO: 15) и/или VL-домен BAK502G9 (SEQ ID NO: 16).
Как правило, образуют пару домена VH с доменом VL, чтобы получить антигенсвязывающий участок антитела, хотя, как обсуждается далее, можно использовать отдельно домен VH, чтобы связать антиген. В одном предпочтительном варианте образуют пару VH-домена BAK502G9 (SEQ ID NO: 15) с VL-доменом BAK502G9 (SEQ ID NO: 16) для того, чтобы образовать антигенсвязывающий участок антитела, содержащий как VH-, так и VL-домены BAK502G9. В других вариантах образуют пару VH BAK502G9 c другим VL-доменом, отличным от VL BAK502G9. Получение разнородных легких цепей хорошо разработано в данной области.
Подобным образом любой набор HCDR линии BAK278D6 может быть представлен в домене VH, который используют в качестве специфического связывающего партнера отдельно или в комбинации с доменом VL. Может быть получен домен VH с набором HCDR антитела линии BAK278D6, например, показанным в таблице 1, и если такой VH-домен объединяют с доменом VL, то домен VL может быть получен с набором LCDR антитела линии BAK278D6, например, показанным в таблице 1. Объединение набора HCDR и набора LCDR может быть таким, как показано в таблице 1, с образованием антигенсвязывающего участка антитела, содержащего набор CDR, который показан в таблице 1. Каркасные области доменов VH и/или VL могут быть каркасами зародышевой линии. Каркасные области домена тяжелой цепи могут быть выбраны из семейства VH-1, и предпочтительным каркасом VH-1 является каркас DP-14. Каркасные области легкой цепи могут быть выбраны из семейства λ3, и таким предпочтительным каркасом является λ3-3H.
Один или несколько CDR могут быть взяты из домена VH или VL BAK502G9 и включены в подходящий каркас. Это дополнительно обсуждается в данном описании. HCDR 1, 2 и 3 BAK502G9 показаны в SEQ ID NO: 7, 8 и 9 соответственно. LCDR 1, 2 и 3 BAK502G9 показаны в SEQ ID NO: 10, 11 и 12 соответственно.
То же самое применимо для других CDR линии BAK278D6 и наборов CDR, которые показаны в таблице 1.
Следующие варианты осуществления изобретения относятся к специфическому связывающему партнеру, содержащему домен VH и/или VL, или антигенсвязывающий участок, содержащий CDR домена VH и/или VL молекулы антитела, называемого в данном описании 167A11 (VH: SEQ ID NO: 23 и VL: SEQ ID NO: 24), и его производных 615E3 (VH: SEQ ID NO: 33 и VL: SEQ ID NO: 34), BAK582F7 (CDR VH SEQ ID 141-143) и BAK612B5 (CDR VH SEQ ID 147-149). Указанные партнеры распознают IL-13 человека. Производные 167A11, полученные в результате рандомизации CDR3 VH, являются эффективными молекулами scFv (5-6 нМ). Линию 167A11 можно использовать в любом аспекте и варианте осуществления настоящего изобретения, которые приведены в данном описании для других молекул, например в способах мутирования и отбора антигенсвязывающих участков с повышенной эффективностью.
Варианты доменов VH и VL и CDR согласно настоящему изобретению, включая варианты, аминокислотные последовательности которых приведены в данном описании и которые могут быть использованы в специфических партнерах, связывающих IL-13, могут быть получены с помощью способов изменения или мутирования последовательностей и скрининга. Такие способы также предлагаются в настоящем изобретении.
Варианты аминокислотной последовательности вариабельного домена любого из доменов VH и VL, последовательности которых специально приведены в данном описании, могут быть использованы согласно настоящему изобретению, как обсуждается. Конкретные варианты могут включать одно или несколько изменений аминокислотной последовательности (присоединение, делецию, замену и/или инсерцию аминокислотного остатка), могут иметь примерно менее 20 изменений, примерно менее 15 изменений, примерно менее 10 изменений или примерно менее 5 изменений, 4, 3, 2 или 1. Изменения могут быть осуществлены в одной или нескольких каркасных областях и/или одной или нескольких CDR.
Согласно следующим аспектам настоящего изобретения предлагается специфический связывающий партнер, который конкурирует за связывание с антигеном с любым специфическим связывающим партнером, который связывает антиген и содержит специфический связывающий партнер, VH- и/или VL-домен, приведенный в данном описании, или HCDR3, приведенный в данном описании, или вариант любого из указанных. Конкурирование между связывающими партнерами можно легко анализировать in vitro, например используя ELISA и/или посредством одного связывающего партнера специфичной репортерной молекулой, которую можно выявлять в присутствии другого немеченого связывающего партнера(ров), чтобы обеспечить возможность идентификации специфических связывающих партнеров, которые связывают один и тот же эпитоп или перекрывающийся эпитоп.
Таким образом, следующий аспект настоящего изобретения относится к специфическому связывающему партнеру, содержащему антигенсвязывающий участок антитела человека, который конкурирует с молекулой антитела BAK502G9, в частности scFv и/или IgG4 BAK502G9, в отношении связывания с IL-13. В следующих аспектах настоящее изобретение относится к специфическому связывающему партнеру, содержащему антигенсвязывающий участок антитела человека, который конкурирует с антигенсвязывающим участком антитела за связывание с IL-13, при этом антигенсвязывающий участок антитела состоит из домена VH и домена VL, и при этом домены VH и VL содержат набор CDR линии BAK278D6.
В данной области имеются различные способы получения антител против IL-13, и указанные антитела могут конкурировать с молекулой антитела BAK502G9, молекулой антитела с набором CDR BAK502G9 или молекулой антитела с набором CDR линии BAK278D6 за связывание с IL-13.
В следующем аспекте настоящее изобретение относится к способу получения одного или нескольких специфических связывающих партнеров, способных связывать антиген, при этом способ включает в себя осуществление контакта библиотеки специфических связывающих партнеров согласно изобретению и указанного антигена и отбор одного или нескольких специфических связывающих партнеров из библиотеки, способных связывать указанный антиген.
Библиотека может быть представлена на поверхности частиц бактериофага, при этом каждая частица содержит нуклеиновую кислоту, кодирующую вариабельный домен VH антитела, представленный на ее поверхности, и необязательно также представленный домен VL, если он присутствует.
После отбора специфических связывающих партнеров, способных связывать антиген и представленных на частицах бактериофага, из частицы бактериофага, представляющего указанный выбранный специфический связывающий партнер, может быть взята нуклеиновая кислота. Такая нуклеиновая кислота может быть использована при последующем получении специфического связывающего партнера или вариабельного домена VH антитела (необязательно вариабельного домена VL антитела) посредством экспрессии нуклеиновой кислоты с последовательностью нуклеиновой кислоты, взятой из частицы бактериофага, представляющего указанный выбранный специфический связывающий партнер.
Вариабельный домен VH антитела с аминокислотной последовательностью вариабельного домена VH антитела указанного выбранного специфического связывающего партнера может быть получен в изолированной форме, также как в виде специфического связывающего партнера, содержащего такой домен VH. Затем может быть проверена способность связывать IL-13, а также способность конкурировать с BAK502G9 (например, в форме scFv и/или в форме IgG, например, IgG4) за связывание с IL-13. Может быть проверена способность нейтрализовать IL-13, как обсуждается далее.
Специфический связывающий партнер согласно настоящему изобретению может связывать IL-13 с аффинностью молекулы антитела BAK502G9, например scFv или предпочтительно IgG4 BAK502G9, или с более высокой аффинностью.
Специфический связывающий партнер согласно настоящему изобретению может нейтрализовать IL-13 с эффективностью молекулы антитела BAK502G9, например scFv или предпочтительно IgG4 BAK502G9, или с более высокой эффективностью.
Специфический связывающий партнер согласно настоящему изобретению может нейтрализовать встречающийся в природе IL-13 с эффективностью молекулы антитела BAK502G9, например scFv, или предпочтительно IgG4 BAK502G9, или с более высокой эффективностью.
Аффинность связывания и эффективность нейтрализации различных специфических связывающих партнеров можно сравнить в подходящих условиях.
Антитела согласно настоящему изобретению имеют ряд преимуществ по сравнению с существующими коммерческими анти-IL-13-антителами, в частности тремя коммерческими антителами грызунов против IL-13 человека, а именно JES10-5A2 (BioSource), B-B13 (Euroclone) и клоном 321166 (R and D Systems). Эффективность антител согласно настоящему изобретению сравнивали с коммерческими антителами JES10-A2 и B-B13. Клон 321166 не оценивали, так как предварительные эксперименты показали, что данный клон был значительно менее эффективным, чем известные коммерческие антитела.
Эффективность и применение коммерческих IL-13-антител грызунов на человеке, вероятно, ограничены вследствие повышенной возможности индуцировать иммуногенные ответы и, следовательно, более быстрого клиренса из организма. Кинетический анализ антител согласно настоящему изобретению у приматов, отличных от человека, свидетельствует, что указанные антитела обладают скоростью клиренса, которая сходна со скоростью клиренса других известных человеческих или гуманизированных антител.
Антитела, предлагаемые в различных вариантах осуществления настоящего изобретении, узнают IL-13 приматов, отличных от человека, включая IL-13 макак-резус и макак-крабоедов. Определение профилей эффективности и безопасности антитела у приматов, отличных от человека, чрезвычайно важно, так как обеспечивает возможность предсказания безопасности, фармакокинетического и фармакодинамического профиля антител у людей.
Кроме того, антитела согласно различным вариантам осуществления настоящего изобретения дополнительно узнают вариант IL-13 человека R130Q, который связан с астмой. Перекрестная реактивность с вариантом IL-13 позволяет использовать антитела согласно настоящему изобретению и композиции, содержащие антитела согласно настоящему изобретению, для лечения пациентов с IL-13 дикого типа и вариантом IL-13.
Предпочтительный вариант осуществления настоящего изобретения включает антитела, которые нейтрализуют встречающийся в природе IL-13 с эффективностью, которая равна или превышает эффективность связывающего антиген IL-13 участка, образованного доменом VH BAK502G9 (SEQ ID NO: 15) и доменом VL BAK502G9 (SEQ ID NO: 16). Например, авторы изобретения показали, что типичные клоны, такие как BAK502G9, 1167F2 и 1183Н4, являются в значительной степени более эффективными против встречающегося в природе IL-13, чем известные коммерческие антитела (фиг.7).
Кроме последовательностей антител, специфический связывающий партнер согласно настоящему изобретению может содержать другие аминокислоты, например формирующие пептид или полипептид, такой как подвергнутый фолдингу домен, или придающие молекуле другое функциональное свойство в дополнение к способности связывать антиген. Специфические связывающие партнеры согласно изобретению могут нести регистрируемую метку или могут быть конъюгированы с токсином или направляющим к мишени остатком или ферментом (например, посредством пептидной связи или линкера).
В следующих аспектах изобретение относится к изолированной нуклеиновой кислоте, которая содержит последовательность, кодирующую специфический связывающий партнер, домен VH и/или домены VL согласно настоящему изобретению, и к способам получения специфического связывающего партнера, VH-домена и/или VL-домена согласно изобретению, которые включают в себя экспрессию указанной нуклеиновой кислоты в условиях, который приводят к получению указанного специфического связывающего партнера, VH-домена и/или VL-домена, и его извлечение.
Специфические связывающие партнеры согласно изобретению могут быть использованы в способе лечения или диагностики организма человека или животного, таком как способ лечения (которое может включать профилактическое лечение) заболевания или расстройства у больного человека, который включает в себя введение указанному пациенту эффективного количества специфического связывающего партнера согласно изобретению. Состояния, поддающиеся лечению согласно настоящему изобретению, включают любое состояние, при котором IL-13 играет роль, в частности астму, атопический дерматит, аллергический ринит, фиброз, хроническое обструктивное заболевание легких, склеродермию, воспалительное заболевание кишечника и лимфому Ходжкина. Кроме того, антитела согласно настоящему изобретению также можно использовать при лечении опухолей и вирусных инфекций, поскольку указанные антитела будут ингибировать опосредованную IL-13 иммуносупрессию [64, 65].
Следующий аспект настоящего изобретения относится к нуклеиновой кислоте, как правило, изолированной, кодирующей вариабельный домен VH и/или вариабельный домен VL антитела, охарактеризованные в данном описании.
Другой аспект настоящего изобретения относится к нуклеиновой кислоте, как правило, изолированной, кодирующей приведенную в данном описании последовательность CDR VH или CDR VL, в частности CDR VH, выбранную из SEQ ID NO: 7, 8 и 9, или CDR VL, выбранную из SEQ ID NO: 10, 11 и 12, наиболее предпочтительно CDR3 VH BAK502G9 (SEQ ID NO: 9). Также в настоящем изобретении предлагается нуклеиновая кислота, кодирующая набор CDR BAK502G9, нуклеиновая кислота, кодирующая набор HCDR BAK502G9, и нуклеиновая кислота, кодирующая набор LCDR BAK502G9, которые представляют собой нуклеиновые кислоты, кодирующие отдельные CDR, HCDR, LCDR и наборы CDR, HCDR, LCDR линии BAK278D6.
Следующий аспект относится к клетке-хозяину, трансформированной нуклеиновой кислотой согласно изобретению.
Еще один аспект относится к способу получения вариабельного домена VH антитела, при этом способ заключается в том, чтобы вызвать экспрессию кодирующей нуклеиновой кислоты. Такой способ может заключаться в культивировании клеток-хозяев в определенных условиях для получения указанного вариабельного домена VH антитела.
Также предлагаются аналогичные способы получения вариабельных доменов VL и специфических связывающих партнеров, содержащих VH- и/или VL-домен, в качестве следующих аспектов настоящего изобретения.
Способ получения может включать в себя стадию выделения и/или очистки продукта.
Способ получения может заключаться в приготовлении продукта в виде композиции, содержащей, по меньшей мере, один дополнительный компонент, такой как фармацевтически приемлемый эксципиент.
Указанные и другие аспекты изобретения более подробно описаны ниже.
ТЕРМИНОЛОГИЯ
Специфический связывающий партнер
Данный термин описывает член пары молекул, который обладает специфичностью связывания с другим членом пары. Партнеры в специфично связывающейся паре могут быть природного происхождения или полностью или частично получены путем синтеза. Один партнер в паре молекул имеет площадь на своей поверхности или углубление, которое специфично связывается и, следовательно, является комплементарным по отношению к конкретной пространственной и полярной организации другого члена пары молекул. Таким образом, члены пары обладают свойством специфично связываться друг с другом. Примерами типов специфично связывающихся пар являются антиген-антитело, биотин-авидин, гормон-рецептор гормона, рецептор-лиганд, фермент-субстрат. Настоящее изобретение относится к реакциям типа антиген-антитело.
Молекула антитела
Данный термин описывает иммуноглобулин, либо природный, либо получаемый полностью или частично с помощью синтеза. Термин также охватывает любой полипептид или белок, содержащий связывающий домен антитела. Фрагменты антител, которые содержат антигенсвязывающий домен, представляют собой такие молекулы, как Fab, scFv, Fv, dAb, Fd, и диантитела.
Можно получить моноклональные и другие антитела и использовать способы технологии рекомбинантной ДНК, чтобы получить другие антитела или химерные молекулы, которые сохраняют специфичность исходного антитела. Такие способы могут заключаться во введении ДНК, кодирующей вариабельную область иммуноглобулина или определяющие комплементарность области (CDR) антитела в константные области или константные области плюс каркасные области другого иммуноглобулина. Смотри, например, EP-A-184187, GB2188638A или EP-A-239400, и большое количество более поздней литературы. Гибридома или другая клетка, продуцирующая антитело, может быть подвергнута генетической мутации или другим изменениям, которые могут изменять или не изменять специфичность связывания продуцируемого антитела.
Поскольку антитела могут быть модифицированы рядом способов, термин «молекула антитела» следует рассматривать как охватывающий любой специфический связывающий партнер или вещество, имеющее антигенсвязывающий домен антитела требуемой специфичности. Таким образом, данный термин охватывает фрагменты и производные антител, включая любой полипептид, содержащий связывающий домен иммуноглобулина, либо природный, либо полностью или частично полученный с помощью синтеза. Следовательно, включены химерные молекулы, содержащие связывающий домен иммуноглобулина или эквивалент, слитый с другим полипептидом. Клонирование и экспрессия химерных антител описаны в EP-A-0120694 и EP-A-0125023 и в большом количестве более поздней литературы.
Следующие способы, имеющиеся в области конструирования антител, сделали возможным выделение человеческих и гуманизированных антител. Например, гибридомы человека могут быть получены, как описано в Kontermann et al. [107]. Фаговый дисплей, другой разработанный способ создания специфических связывающих партнеров, подробно описан во многих публикациях, таких как Kontermann et al. [107] и WO 92/01047 (обсуждаемая ниже). Трансгенных мышей, у которых гены антител мыши инактивированы и функционально заменены генами антител человека и при этом оставлены интактными другие компоненты иммунной системы мышей, могут быть использованы для выделения антител человека к антигенам человека [108].
Синтетические молекулы антител могут быть созданы посредством экспрессии генов, образованных из олигонуклеотидов, синтезированных и собранных в подходящих экспрессирующих векторах, например, как описано Knappik et al. J. Mol. Biol.(2000) 296, 57-86 или Krebs et al. Journal of Immunological Methods 254, 2001, 67-84.
Показано, что фрагменты целого антитела могут выполнять функцию связывания антигенов. Примерами связывающих фрагментов являются (i) Fab-фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) Fd-фрагмент, состоящий из доменов VH и CH1; (iii) Fv-фрагмент, состоящий из доменов VL и VH одного антитела; (iv) dAb-фрагмент (Ward, E.S. et al., Nature 341, 544-546 (1989), McCafferty et al. (1990) Nature, 348, 552-554), который состоит из домена VH; (v) изолированные области CDR; (vi) F(ab')2- фрагменты, бивалентный фрагмент, содержащий два связанных Fab-фрагмента; (vii) одноцепочечные молекулы Fv (scFv), в которых домен VH и домен VL связаны пептидным линкером, который позволяет двум доменам объединяться с образованием антигенсвязывающего участка (Bird et al., Science, 242, 423-426, 1988; Huston et al., PNAS USA, 85, 5879-5883, 1988); (viii) биспецифичные одноцепочечные димеры Fv (PCT/US 92/09965) и (ix) «диантитела», поливалентные или полиспецифичные фрагменты, сконструированные посредством слияния генов (WO 94/13804; P.Holliger et al., Proc. Natl. Acad. Sci. USA 90, 6444-6448, 1993). Молекулы Fv, scFv или диантитела могут быть стабилизированы введением связывания доменов VH и VL дисульфидными мостиками (Y.Reiter et al., Nature Biotech, 14, 1239-1245, 1996). Также могут быть получены мини-антитела, содержащие scFv, связанный с доменом CH3 (S.Hu et al., Cancer Res., 56, 3055-3061, 1996).
В том случае, когда используют биспецифичные антитела, они могут представлять собой обычные биспецифичные антитела, которые могут быть созданы множеством способов (Holliger, P.and Winter G. Current Opinion Biotechnol. 4, 446-449 (1993)), например полученные химическим путем или из гибридных гибридом, или могут представлять собой любой из биспецифичных фрагментов антител, указанных выше. Примеры биспецифичных антител включают антитела, полученные с использованием методики BiTE™, при которой могут быть использованы связывающие домены двух антител с разной специфичностью, и непосредственно связаны короткими гибкими пептидами. При этом объединяют два антитела в одной короткой полипептидной цепи. Диантитела и scFv могут быть сконструированы без области Fc с использованием только вариабельных доменов, что потенциально снижает влияние анти-идиотипической реакции.
Биспецифичные диантитела, в отличие от биспецифичных целых антител, также могут быть особенно полезными, поскольку их можно легко конструировать и экспрессировать в E.coli. Диантитела (и многие другие полипептиды, такие как фрагменты антител) с подходящими специфичностями связывания легко могут быть выбраны с использованием фагового дисплея (WO 94/13804) из библиотек. Если одно плечо диантитела необходимо сохранить постоянным, например, со специфичностью, направленной против IL-13, то может быть создана библиотека, в которой другое плечо варьируется, и выбирают антитело подходящей специфичности. Биспецифичные целые антитела могут быть получены конструированием способом «knobs-into-holes» («головка-в углубление» (J.B.B.Ridgeway et al., Protein Eng., 9, 616-621, 1996).
Антигенсвязывающий домен
Термин описывает часть молекулы антитела, которая содержит область, которая специфично связывается и является комплементарной по отношению к части или всему антигену. В том случае, когда антиген крупный, антитело может связываться только с конкретной частью антигена, и указанную часть называют эпитопом. Антигенсвязывающий домен может быть обусловлен одним или несколькими вариабельными доменами антитела (например, так называемый Fd-фрагмент антитела, состоящий из домена VH). Предпочтительно антигенсвязывающий домен содержит вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH).
Специфический
Термин может быть использован, по отношению к ситуации, в которой один член специфично связывающейся пары не будет проявлять какого-либо существенного связывания с другими молекулами, отличными от его специфического партнера(ров) в связывании. Термин также применим, например, в том случае, когда антигенсвязывающий домен является специфичным по отношению к конкретному эпитопу, который несут ряд антигенов, и в этом случае специфический связывающий партнер, несущий антигенсвязывающий домен, будет способен связываться с различными антигенами, несущими эпитоп.
Содержат
Термин обычно используют в смысле включают в себя, то есть в смысле, допускающем наличие одного или нескольких признаков или компонентов.
Изолированный
Термин относится к состоянию, в котором согласно настоящему изобретению будут находиться специфические связывающие партнеры согласно изобретению или нуклеиновая кислота, кодирующая такие связывающие партнеры. Изолированные партнеры и изолированная нуклеиновая кислота не будут содержать или по существу не будут содержать вещества, с которым они ассоциированы в природных условиях, такого как другие полипептиды или нуклеиновые кислоты, с которыми их обнаруживают в их природной среде или в среде, в которой их получают (например, в культуре клеток), когда такой препарат получают на практике способом на основе рекомбинантной ДНК in vitro или in vivo. Партнеры и нуклеиновая кислота могут быть приготовлены с разбавителями или адъювантами и все еще быть изолированными для практических целей - например, партнеры, как правило, будут смешаны с желатином или другими носителями при использовании для того, чтобы покрыть планшеты для микротитрования для применения иммуноанализов, или будут смешаны с фармацевтически приемлемыми носителями или разбавителями при использовании в диагностике или терапии. Специфические связывающие партнеры могут быть гликозилированы, либо в естественных условиях, либо посредством систем гетерологичных эукариотических клеток (например, клеток CHO или NSO (ECACC 85110503), или они могут быть (например, в случае получения в результате экспрессии в прокариотической клетке) негликозилированными.
Встречающийся в природе IL-13
Термин, как правило, относится к состоянию, в котором могут встречаться белок IL-13 или его фрагменты. Встречающийся в природе IL-13 означает белок IL-13, который в природных условиях продуцируется клеткой без предварительного введения кодирующей нуклеиновой кислоты с использованием рекомбинантной технологии. Таким образом, встречающийся в природе IL-13 может быть продуцирован в естественных условиях, например T-клетками CD4+, и/или может быть выделен из млекопитающего, например человека, примата, отличного от человека, грызуна, такого как крыса или мышь.
Рекомбинантный IL-13
Термин относится к состоянию, в котором могут встречаться белок IL-13 или его фрагменты. Рекомбинантный IL-13 означает белок IL-13 или его фрагменты, полученные с помощью рекомбинантной ДНК в гетерологичном хозяине. Рекомбинантный IL-13 может отличаться от встречающегося в природе IL-13 гликозилированием.
Рекомбинантные белки, экспрессированные в прокариотических бактериальных системах экспрессии, не гликозилируются, тогда как белки, экспрессированные в эукариотических системах, таких как клетки млекопитающих или насекомых, являются гликозилированными. Однако белки, экспрессированные в клетках насекомых, отличаются по гликозилированию от белков, экспрессированных в клетках млекопитающих.
Под «по существу такой, который указан» подразумевается, что подходящий CDR или VH- или VL-домен согласно изобретению будут либо идентичны, либо в высокой степени сходны с конкретными областями, последовательность которых указана в данном описании. Термин «в высокой степени сходный» предполагает, что от 1 до 5, предпочтительно от 1 до 4, например от 1 до 3, или 1, или 2, или 3, или 4 аминокислотные замены могут быть осуществлены в CDR и/или VH- или VL-домене.
Структура, несущая CDR или набор CDR согласно изобретению, как правило, будет представлять собой последовательность тяжелой цепи или легкой цепи антитела или ее основную часть, в которой CDR или набор CDR локализованы в положении, соответствующем CDR или набору CDR встречающихся в природе вариабельных доменов антител VH и VL, кодируемых подвергнутыми реаранжировке генами иммуноглобулинов. Структуры и положения вариабельных доменов иммуноглобулинов можно получить, обратившись к публикации (Kabat, E.A. et al., Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health и Human Services. 1987, и ее обновленной редакции, доступной в настоящее время в Интернете (http://immuno.bme.nwu.edu или при поиске «Kabat» с использованием любого поискового сервера).
Носителем CDR также могут быть другие каркасы, такие как фибронектин или цитохром B [115, 116].
Предпочтительно аминокислотную последовательность CDR, по существу такую, как указана в данном описании, в виде CDR несет вариабельный домен человека или его основная часть. Последовательности HCDR3, по существу такие, которые указаны в данном описании, представляют собой предпочтительные варианты осуществления настоящего изобретения, и предпочтительно, чтобы каждая из таких последовательностей в качестве HCDR3 находилась в вариабельном домене тяжелой цепи человека или его основной части.
Вариабельные домены, используемые в изобретении, могут быть получены из любого вариабельного домена зародышевой линии человека или вариабельного домена человека, подвергнутого реаранжировке, или могут представлять собой синтетический вариабельный домен, основанный на консенсусных последовательностях известных вариабельных доменов человека. Последовательность CDR согласно изобретению (например, CDR3) может быть введена в репертуар вариабельных доменов, в которых отсутствует CDR (например, CDR3), с использованием методики рекомбинантной ДНК.
Например, Marks et al. (Bio/Technology, 1992, 10: 779-783) описывают способы получения репертуаров вариабельных доменов антител, в которых используют консенсусные праймеры, направленные или расположенные рядом с 5'-концом области вариабельного домена, вместе с консенсусными праймерами к третьей каркасной области генов VH человека, чтобы получить репертуар вариабельных доменов VH, в которых отсутствует CDR3. Marks et al., кроме того, описывают, каким образом можно комбинировать данный репертуар с CDR3 конкретного антитела. Используя аналогичный способ, можно осуществлять перетасовку полученных из CDR3 последовательностей согласно настоящему изобретению с репертуарами VH- или VL-доменов, в которых отсутствует CDR3, и подвергнутые перетасовке полные домены VH или VL могут быть объединены с родственным VL- или VH-доменом, чтобы получить специфические связывающие партнеры согласно изобретению. Затем репертуар может быть представлен в подходящей системе хозяина, такой как система фагового дисплея согласно WO 92/01047 или согласно любой из огромного множества более поздней литературы, включая Kay, B.K., Winter, J. and McCafferty, J. (1996) Phage Display of Peptides and Proteins: A Laboratory Manual, San Diego: Academic Press, для того, чтобы можно было отобрать подходящие специфические связывающие партнеры. Репертуар может состоять из любого количества от 104 отдельных партнеров и выше, например из 106-108 или 1010 партнеров. Другие подходящие системы хозяев включают дрожжевой дисплей, бактериальный дисплей, дисплей в T7, рибосомный дисплей и тому подобные. Обзор относительно рибосомного дисплея смотри в Lowe D. and Jermutus L., 2004, Curr. Pharm, Biotech, 517-27, а также WO 92/01047.
Аналогичные способы перетасовки или комбинаторные способы описаны Stemmer (Nature, 1994, 370: 389-391), который описывает способ в связи с геном β-лактамазы, но отмечает, что способ может быть использован для создания антител.
Следующей альтернативой является создание новых областей VH или VL, несущих полученные из CDR последовательности согласно изобретению, с использованием случайного мутагенеза одного или нескольких выбранных генов VH и/или VL, чтобы создать мутации в полном вариабельном домене. Такой способ описан Gram et al. (1992, Proc. Natl. Acad. Sci., USA, 89: 3576-3580), которые использовали склонную к ошибкам ПЦР. В предпочтительных вариантах получают одну или две аминокислотные замены в наборе HCDR и/или LCDR.
Другим способом, который можно использовать, является направленный мутагенез в CDR-областях генов VH или VL. Такие способы описаны Barbas et al.,(1994, Proc. Natl. Acad. Sci., USA, 91: 3809-3813) и Schier et al. (1996, J. Mol. Biol. 263: 551-567).
Все описанные выше способы как таковые известны в данной области и сами по себе не являются частью настоящего изобретения. Специалист в данной области сможет применить такие способы, чтобы получить специфические связывающие партнеры согласно изобретению, используя обычную в данной области методику.
Следующий аспект изобретения относится к способу получения антигенсвязывающего домена антитела, специфичного по отношению к антигену IL-13, при этом способ включает в себя получение с помощью присоединения, делеции, замены или инсерции одной или нескольких аминокислот в аминокислотной последовательности домена VH, указанного в данном описании, домена VH, который представляет собой вариант аминокислотной последовательности домена VH, необязательно объединение полученного таким образом домена VH с одним или несколькими доменами VL и тестирование домена VH или комбинации или комбинаций VH/VL, чтобы идентифицировать специфический связывающий партнер или антигенсвязывающий домен антитела, специфический по отношению к антигену IL-13 и необязательно обладающий одним или несколькими предпочтительными свойствами, предпочтительно способностью нейтрализовать активность IL-13. Указанный домен VL может иметь аминокислотную последовательность, которая по существу является такой, как указано в данном описании.
Можно применять аналогичный способ, в котором один или несколько вариантов последовательности домена VL, приведенных в данном описании, объединяют с одним или несколькими доменами VH.
В предпочтительном варианте домен VH BAK502G9 (SEQ ID NO: 15) может быть подвергнут мутации, чтобы получить один или несколько вариантов аминокислотной последовательности домена VH и/или VL BAK502G9 (SEQ ID NO: 16).
Следующий аспект изобретения относится к способу получения специфического связывающего партнера, специфичного по отношению к антигену IL-13, и указанный способ включает в себя:
(a) получение исходного репертуара нуклеиновых кислот, кодирующих домен VH, который либо содержит CDR3, которую нужно заменить, либо не содержит области, кодирующей CDR3;
(b) объединение указанного репертуара с донорной нуклеиновой кислотой, кодирующей аминокислотную последовательность, по существу такую, которая указана в данном описании, для CDR3 VH, так чтобы встроить указанную донорную нуклеиновую кислоту в область CDR3 в репертуаре, для того, чтобы получить продукт в виде репертуара нуклеиновых кислот, кодирующих домен VH;
(c) экспрессию нуклеиновых кислот указанного полученного в результате репертуара;
(d) отбор специфического связывающего партнера, специфичного по отношению к IL-13; и
(e) извлечение указанного специфического связывающего партнера или кодирующей его нуклеиновой кислоты.
Кроме того, можно использовать аналогичный способ, в котором CDR3 VL согласно изобретению объединяют с репертуаром нуклеиновых кислот, кодирующих домен VL, который либо содержит CDR3, которую необходимо заменить, либо не содержит области, кодирующей CDR3.
Подобным образом одна, или несколько, или все три CDR могут быть привиты в репертуар доменов VH или VL, которые затем подвергают скринингу в отношении специфического связывающего партнера или специфических связывающих партнеров, специфичных по отношению к IL-13.
В предпочтительном варианте можно использовать одну или несколько HCDR1 (SEQ ID NO: 7), HCDR2 (SEQ ID NO: 8) и HCDR3 (SEQ ID NO: 9) BAK502G9 или набор HCDR BAK502G9 и/или одну или несколько LCDR1 (SEQ ID NO: 10), LCDR2 (SEQ ID NO: 11) BAK502G9 или набор LCDR BAK502G9.
Основная часть вариабельного домена иммуноглобулина будет содержать, по меньшей мере, три области CDR вместе с расположенными между ними каркасными областями. Предпочтительно часть также будет содержать, по меньшей мере, примерно 50% одной из двух или обеих первой и четвертой каркасных областей, при этом 50% представляют собой C-концевые 50% первой каркасной области и N-концевые 50% четвертой каркасной области. Дополнительными остатками на N-конце или C-конце основной части вариабельного домена могут быть остатки, обычно не связанные с встречающимися в природе областями вариабельного домена. Например, конструирование специфических связывающих партнеров согласно настоящему изобретению, осуществляемое способами на основе рекомбинантной ДНК, может приводить к введению N- или C-концевых остатков, кодируемых линкерами, введенными для того, чтобы облегчить клонирование или другие стадии обработки. Другие стадии обработки включают введение линкеров, чтобы соединить вариабельные домены согласно изобретению с дополнительными последовательностями белка, включая тяжелые цепи иммуноглобулина, другие вариабельные домены (например, при получении диантител) или белковые метки, которые более подробно обсуждаются в другой части данного описания.
Хотя в предпочтительном аспекте изобретения предпочтительными являются специфические связывающие партнеры, содержащие пару доменов VH и VL, отдельные связывающие домены, основанные на последовательностях доменом либо VH, либо VL, образуют следующие аспекты изобретения. Известно, что отдельные домены иммуноглобулина, в частности домены VH, способны специфичным образом связывать антигены-мишени.
В случае любого из отдельных специфических связывающих доменов указанные домены могут быть использованы для скрининга в отношении комплементарных доменов, способных образовывать состоящий из двух доменов специфический связывающий партнер, способный связывать IL-13.
Это можно осуществить способами скрининга на основе фагового дисплея с использованием так называемого иерархического двойного комбинаторного способа, который описан в WO 92/01047, при котором отдельную колонию, содержащую клон либо H-, либо L-цепи, используют для инфекции полной библиотеки клонов, кодирующих другую цепь (L или H), и полученный в результате двухцепочечный специфический связывающий партнер отбирают согласно способу на основе фагового дисплея, такому как способы, описанные в указанной публикации. Такой способ также описан в Marks et al., там же.
Специфические связывающие партнеры согласно настоящему изобретению, кроме того, могут содержать константные области антитела или их части. Например, домен VL домен может быть связан на своем C-конце с константными областями легкой цепи антитела, включая Cκ- или Cλ-цепи человека, предпочтительно Cλ-цепи. Подобным образом, специфический связывающий партнер, основанный на домене VH, может быть связан на своем C-конце с полной или с частью (например, доменом CH1) тяжелой цепи иммуноглобулина, полученной из антитела любого изотипа, например IgG, IgA, IgE и IgM, и любого подкласса изотипов, в частности IgG1 и IgG4. Предпочтительным является IgG4. IgG4 является предпочтительным, так как он не связывает комплемент и не создает эффекторных функций. Любой синтетический или другой вариант константной области, который обладает указанными свойствами и стабилизирует вариабельные области, также является предпочтительным для применения в вариантах осуществления настоящего изобретения.
Специфические связывающие партнеры согласно изобретению можно метить регистрируемой или функциональной меткой. Регистрируемые метки включают радиоактивные метки, такие как 131I или 99Tc, которые могут быть связаны с антителом согласно изобретению с использованием обычных химических способов, известных в области визуализации антител. Метки также включают ферментные метки, такие как пероксидаза хрена. Метки, кроме того, включают химические остатки, такие как биотин, которые можно регистрировать посредством связывания со специфичным родственным регистрируемым остатком, например меченным авидином.
Специфические связывающие партнеры согласно настоящему изобретению конструируют для применения в способах диагностики или лечения человека или животного, предпочтительно человека.
Соответственно следующие аспекты изобретения относятся к способам лечения, включающим в себя введение предлагаемого специфического связывающего партнера, фармацевтических композиций, содержащих такой специфический связывающий партнер, и к применению такого специфического связывающего партнера для производства лекарственного средства для введения, например, в способе получения лекарственного средства или фармацевтической композиции, включающем в себя приготовление композиции специфического связывающего партнера с фармацевтически приемлемым эксципиентом.
Клинические показания, при которых можно применять анти-IL-13-антитело, чтобы получить терапевтический эффект, включают астму, атопический дерматит, аллергический ринит, фиброз, хроническое обструктивное заболевание легких, воспалительное заболевание кишечника, склеродермию и лимфому Ходжкина. Как уже отмечалось, лечение с помощью анти-IL-13-антител является эффективным в случае всех указанных заболеваний.
Анти-IL-13-лечение можно осуществлять перорально, путем инъекции (например, подкожно, внутривенно, внутрибрюшинно или внутримышечно), путем ингаляции или местно (например, внутриглазным введением, интраназальным, ректальным, в рану, на кожу). Путь введения можно определить на основе физико-химических параметров лечения, принимая во внимание заболевание или требования оптимизации эффективности или минимизации побочных эффектов.
Предполагается, что анти-IL-13-лечение не будет ограничено применением в клинике. Поэтому также предпочтительна подкожная инъекция с использованием безыгольного устройства.
Комбинированное лечение может быть использовано для получения значимых синергических эффектов, в частности комбинирование специфического связывающего партнера против IL-13 с одним или несколькими другими лекарственными средствами. Специфический связывающий партнер согласно настоящему изобретению может применяться при комбинировании или в дополнение к быстро действующим или длительно действующим бета-агонистам, кортикостероидам, кромогликату, антагонистам лейкотриена (рецептора), метилксантинам и их производным, ингибиторам IL-4, антагонистам мускариновых рецепторов, ингибиторам IgE, гистаминным ингибиторам, ингибиторам IL-5, ингибиторам эотаксина/CCR3, ингибиторам PDE4, антагонистам TGF-бета, интерферону-гамма, перфенидону, химиотерапевтическим средствам и иммунотерапевтическим средствам.
Комбинированную терапию с одним или несколькими быстро действующими или длительно действующими бета-агонистами, кортикостероидами, кромогликатом, антагонистами лейкотриена (рецептора), ксантинами, ингибиторами IgE, ингибиторами IL-4, ингибиторами IL-5, ингибиторами эотаксина/CCR3, ингибиторами PDE4 можно использовать для лечения астмы. Антитела согласно настоящему изобретению также можно использовать в комбинации с кортикостероидами, антиметаболитами, антагонистами TGF-бета и нижележащего пути передачи сигнала для лечения фиброза. Комбинированная терапия указанными антителами с ингибиторами PDE4, ксантинами и их производными, антагонистами мускариновых рецепторов, быстро и длительно действующими бета-антагонистами может быть применима для лечения хронического обструктивного заболевания легких. Сходные рассуждения в отношении комбинирования приемлемы в случае применения анти-IL-13-лечения атопического дерматита, аллергического ринита, хронического обструктивного заболевания легких, воспалительного заболевания кишечника, склеродермии и лимфомы Ходжкина.
Согласно настоящему изобретению, предлагаемые композиции могут быть введены людям. Введение предпочтительно осуществляют в «терапевтически эффективном количестве», которое является достаточным для проявления полезного для пациента действия. Таким полезным действием может быть, по меньшей мере, ослабление, по меньшей мере, одного симптома. Реальное вводимое количество и скорость и временной курс введения будут зависеть от природы и тяжести состояния, подвергаемого лечению. Назначение лечения, например решение относительно дозы и т.д., возлагается на терапевтов и других лечащих врачей. Подходящие дозы антитела хорошо известны в данной области; смотри Ledermann J.A. et al. (1991) Int. J. Cancer 47: 659-664; Bagshawe K.D. et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4: 915-922.
Точная доза будет зависеть от ряда факторов, включая тот факт, предназначено ли антитело для диагностики или лечения, размер и положение области, подвергаемой лечению, точной природы антитела (например, целое антитело, фрагмент или диантитело) и природы любой регистрируемой метки или другой молекулы, связанной с антителом. Типичная доза антитела будет в диапазоне от 100 мкг до 1 г для системных применений и от 1 мкг до 1 мг для местных применений. Обычно антитело будет представлять собой целое антитело, предпочтительно изотипа IgG4. Указанное представляет собой дозу для однократного лечения взрослого пациента, которая может быть пропорционально скорректирована для детей и новорожденных, а также скорректирована для других форм антител пропорционально молекулярной массе. Лечение может повторяться ежедневно, дважды в неделю, еженедельно или с интервалами в месяц по предписанию лечащего врача. В предпочтительных вариантах осуществления настоящего изобретения лечение является периодическим, и период между введениями составляет примерно две недели или больше, предпочтительно примерно три недели или больше, более предпочтительно примерно четыре недели или больше или примерно раз в месяц.
Специфические связывающие партнеры согласно настоящему изобретению, как правило, будут вводиться в форме фармацевтической композиции, которая может содержать, по меньшей мере, один компонент, кроме специфического связывающего партнера.
Такие фармацевтические композиции согласно настоящему изобретению и для применения согласно настоящему изобретению могут содержать, кроме активного ингредиента, фармацевтически приемлемый эксципиент, носитель, буфер, стабилизатор или другие вещества, хорошо известные специалистам в данной области. Такие вещества должны быть нетоксичными и не должны препятствовать проявлению эффективности активного ингредиента. Конкретная природа носителя или другого вещества будет зависеть от пути введения, которое может быть пероральным или посредством инъекции, например внутривенной.
Фармацевтические композиции для перорального введения могут быть в форме таблетки, капсулы, порошка или жидкости. Таблетка может содержать твердый носитель, такой как желатин или адъювант. Жидкие фармацевтические композиции, как правило, содержат жидкий носитель, такой как вода, нефть, животные и растительные масла, минеральное масло или синтетическое масло. Могут быть включены физиологический раствор соли, раствор декстрозы или другого сахарида, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной инъекции или инъекции в место поражения активный ингредиент будет в форме парентерально приемлемого водного раствора, который является апирогенным и имеет подходящий pH, изотоничность и стабильность. Специалисты, имеющие отношение к данной области, легко могут приготовить подходящие растворы с использованием, например, изотоничных наполнителей, таких как раствор хлорида натрия для инъекций, раствор Рингера для инъекций, раствор Рингера на основе лактата для инъекций. Могут быть включены консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки, которые требуются.
Композиция может быть введена отдельно или в комбинации с другими терапевтическими средствами либо одновременно, либо последовательно, в зависимости от состояния, подвергаемого лечению.
Специфические связывающие партнеры согласно настоящему изобретению могут быть приготовлены в жидкой или твердой формах в зависимости от физико-химических свойств молекулы и пути доставки. Препараты могут содержать эксципиенты или комбинации эксципиентов, например сахара, аминокислоты и поверхностно-активные вещества. Жидкие препараты могут содержать антитело в широком диапазоне концентраций и иметь широкий диапазон pH. Твердые препараты могут быть получены, например, лиофилизацией, распылительной сушкой или сушкой с использованием сверхкритической флюидной технологии. Препараты анти-IL-13 будут зависеть от предполагаемого пути доставки: например, препараты для легочной доставки могут состоять из частиц с физическими свойствами, которые обеспечивают проникновение глубоко в легкие при ингаляции; местные препараты могут содержать модифицирующие вязкость агенты, которые продлевают время, в течение которого лекарственное средство находится в месте действия.
Настоящее изобретение относится к способу, вызывающему или обеспечивающем возможность связывания специфического связывающего партнера, предлагаемого в данном изобретении, с IL-13. Как указано, такое связывание может иметь место in vivo, например, после введения специфического связывающего партнера или нуклеиновой кислоты, кодирующей специфический связывающий партнер, или может иметь место in vitro, например в ELISA, при Вестерн-блоттинге, иммуноцитохимии, иммунопреципитации, аффинной хроматографии или в основанных на клетках анализах, таких как анализ TF-1.
Можно количественно определить связывание специфического связывающего партнера с IL-13. Количественное определение может быть связано с количеством антигена в тестируемом образце, что может представлять интерес для диагностики.
Также предлагается набор, содержащий специфический связывающий партнер или молекулу антитела согласно любому аспекту или варианту осуществления настоящего изобретения, в качестве аспекта настоящего изобретения. В наборе согласно изобретению специфический связывающий партнер или молекула антитела могут быть мечеными, чтобы обеспечить выявление его реактивности в образце, например как описано далее. Компоненты набора, как правило, являются стерильными и находятся в герметично закрытых флаконах или других емкостях. Наборы могут быть использованы в диагностических анализах или других способах, в которых применимы молекулы антител. Набор может содержать инструкции по применению компонентов в способе, например в способе, согласно настоящему изобретению. Вспомогательные материалы, помогающие или обеспечивающие возможность осуществления такого способа, могут быть включены в набор согласно изобретению.
Реактивности антител в образце можно определить подходящими способами. Одной из возможностей является радиоиммуноанализ (РИА). Радиоактивно меченный антиген смешивают с немеченым антигеном (тестируемым образцом) и дают возможность связаться с антителом. Связанный антиген физически отделяют от несвязанного антигена и определяют количество радиоактивного антигена, связанного с антителом. Чем больше антигена имеется в тестируемом образце, тем меньше радиоактивного антигена будет связываться с антителом. Анализ конкурентного связывания также можно использовать с нерадиоактивным антигеном, используя антиген или аналог, связанный с репортерной молекулой. Репортерной молекулой может быть флуорохром, люминофор или лазерный краситель с спектрально изолированными характеристиками поглощения и эмиссии. Подходящие флуорохромы включают флуоресцеин, родамин, фикоэритрин и техасский красный. Подходящие хромогенные красители включают диаминобензидин.
Другие репортеры включают макромолекулярные коллоидные частицы или состоящий из частиц материал, такой как латексные шарики, которые являются окрашенными, магнитными или парамагнитными, и биологически или химически активные агенты, которые могут непосредственно или опосредованно вызывать регистрируемые сигналы, наблюдаемые визуально, регистрируемые электронами способами или регистрируемые иным образом. Указанными молекулами могут быть ферменты, которые катализируют реакции, которые, например, вызывают развитие или изменение окраски или вызывают изменения электрических свойств. Они могут быть легковозбудимыми на уровне молекул, так что электронные переходы между энергетическими состояниями приводят к характерным спектральным параметрам поглощения или эмиссии. Они могут включать химические элементы, используемые для конъюгации с биосенсорами. Могут быть использованы системы регистрации биотин/авидин или биотин/стрептавидин и система щелочной фосфатазы.
Сигналы, генерируемые отдельными конъюгатами антитело-репортер, можно использовать для получения количественно измеряемых абсолютных или относительных данных о связывании релевантных антител в образцах (нормальных и тестируемых).
Настоящее изобретение также относится к применению специфического связывающего партнера, указанного выше, для измерения уровней антигена в конкурентом анализе, то есть к способу измерения уровня антигена в образце с использованием специфического связывающего партнера, предлагаемого в настоящем изобретении, в конкурентном анализе. Это возможно в том случае, когда не требуется физическое разделение связанного и несвязанного антигена. Одной из возможностей является связывание репортерной молекулы со специфическим связывающим партнером, так что происходит физическое или оптическое изменение при связывании. Репортерная молекула может непосредственно или опосредованно генерировать регистрируемые и предпочтительно измеряемые сигналы. Связывание репортерных молекул может быть прямым или опосредованным, ковалентным, например посредством пептидной связи, или нековалентным. Связывание посредством пептидной связи может быть результатом рекомбинантной экспрессии слияния генов, кодирующего антитело и репортерную молекулу.
Настоящее изобретение также относится к измерению уровней антигена непосредственно с использованием специфического связывающего партнера согласно изобретению, например в биосенсорной системе.
Способ определения связывания не является признаком настоящего изобретения и специалисты в данной области способны выбрать подходящий способ согласно своим предпочтениям и общим знаниям.
Как указано в различных аспектах и вариантах, настоящее изобретение распространяется на специфический связывающий партнер, который конкурирует за связывание с IL-13 с любым специфическим связывающим партнером, определенным в данном описании, например IgG4 BAK502G9. Конкуренцию между связывающими партнерами легко можно проанализировать in vitro, например мечением специфичной репортерной молекулой одного связывающего партнера, который может быть выявлен в присутствии другого немеченого связывающего партнера(ров), делая возможным идентификацию специфических связывающих партнеров, которые связывают тот же самый эпитоп или перекрывающийся эпитоп.
Конкуренцию можно определить, например, с использованием ELISA, в котором IL-13 иммобилизуют на планшете, в планшет добавляют первый меченый связывающий партнер вместе с одним или несколькими другими немечеными связывающими партнерами. Присутствие немеченого связывающего партнера, который конкурирует с меченым связывающим партнером, наблюдают по уменьшению сигнала, испускаемого меченым связывающим партнером.
При тестировании в отношении конкуренции можно использовать пептидный фрагмент антигена, в частности, пептид, содержащий представляющий интерес эпитоп. Можно использовать пептид, имеющий последовательность эпитопа плюс одну или несколько аминокислот на любом конце. Можно сказать, что такой пептид «по существу состоит» из указанной последовательности. Специфические связывающие партнеры согласно настоящему изобретению могут быть такими, что их связывание с антигеном ингибируется пептидом с приведенной последовательностью или пептидом, содержащим приведенную последовательность. При указанном тестировании можно использовать пептид с любой последовательностью плюс одна или несколько аминокислот.
Специфические связывающие партнеры, которые связывают конкретный пептид, могут быть выделены, например, из библиотеки в фаговом дисплее посредством пэннинга с использованием пептида(дов).
Настоящее изобретение, кроме того, относится к изолированной нуклеиновой кислоте, кодирующий специфический связывающий партнер согласно настоящему изобретению. Нуклеиновая кислота может включать ДНК и/или РНК. В предпочтительном аспекте настоящее изобретение относится к нуклеиновой кислоте, которая кодирует CDR или набор CDR, или домен VH или домен VL, или антигенсвязывающий участок антитела, или молекулу антитела, например scFv или IgG4, согласно изобретению, которые определены выше.
Настоящее изобретение также относится к конструкциям в форме плазмид, векторов, кассет транскрипции или экспрессии, которые содержат, по меньшей мере, один полинуклеотид, указанный выше.
Настоящее изобретение также относится к рекомбинантной клетке-хозяину, которая содержит одну или несколько конструкций, указанных выше. Нуклеиновая кислота, кодирующая любой CDR, или набор CDR, или домен VH, или домен VL, или антигенсвязывающий участок антитела или молекулу антитела, например scFv или IgG4, которые предлагаются, как таковая образует аспект настоящего изобретения, также как и способ получения кодируемого продукта, и данный способ включает в себя экспрессию продукта с кодирующей нуклеиновой кислоты. Экспрессия может быть легко осуществлена культивированием в подходящих условиях рекомбинантных клеток-хозяев, содержащих нуклеиновую кислоту. После продуцирования посредством экспрессии домен VH или VL или специфический связывающий партнер могут быть выделены и/или очищены с использованием любого подходящего способа и затем применены соответствующим образом.
Специфические связывающие партнеры, домены VH и/или VL и молекулы кодирующих нуклеиновых кислот и векторы согласно настоящему изобретению могут быть выделенными и/или очищенными, например, из их природной среды, в значительной степени очищенной или гомогенной форме, или в случае нуклеиновой кислоты не содержащими или по существу не содержащими нуклеиновой кислоты или генов другого происхождения, отличного от последовательности, кодирующей полипептид с требуемой функцией. Нуклеиновая кислота согласно настоящему изобретению может содержать ДНК или РНК и может быть полностью или частично синтетической. Указание нуклеотидной последовательности, которая приведена в данном описании, охватывает молекулу ДНК с указанной последовательностью и охватывает молекулу РНК с указанной последовательностью, в которой T заменен на U, если контекст не требует иного.
Системы клонирования и экспрессии полипептида во многих различных клетках-хозяевах хорошо известны. Подходящие клетки-хозяева включают бактерии, клетки млекопитающих, клетки растений, дрожжевые и бакуловирусные системы и трансгенные растения и животные. Линии клеток млекопитающих, доступные в данной области для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка (CHO), клетки HeLa, клетки почки детенышей хомячка, клетки меланомы мышей NS0, клетки миеломы крыс YB2/0, клетки эмбриональной почки человека, клетки эмбриональной сетчатки человека и многие другие. Общеизвестным предпочтительным бактериальным хозяином является E.coli.
Экспрессия антител и фрагментов антител в прокариотических клетках, таких как E.coli, хорошо разработана в данной области. Обзор смотри, например, в Pluckthun, A. Bio/Technology 9: 545-551 (1991). Экспрессия в эукариотических клетках в культуре также доступна для специалистов в данной области в качестве выбора для получения специфического связывающего партнера, например Chadd HE and Chamow SM (2001) 110 Current Opinion in Biotechnology 12: 188-194, Andersen DC and Krummen L (2002) Current Opinion in Biotechnology 13: 117, Larrick JW and Thomas DW (2001) Current opinion in Biotechnology 12: 411-418.
Могут быть выбраны или сконструированы подходящие векторы, содержащие подходящие регуляторные последовательности, включая промоторные последовательности, последовательности терминаторов, последовательности полиаденилирования, энхансерные последовательности, маркерные гены или другие последовательности в зависимости от ситуации. Векторы могут представлять собой плазмиды, вирусные векторы, например фаг или фагмиду в зависимости от обстоятельств. Более подробное описание смотри, например, в Molecular Cloning: a Laboratory Manual: 3rd edition, Sambrook and Russell, 2001, Cold Spring Harbor Laboratory Press. Многие известные способы и протоколы обработки нуклеиновой кислоты, например получение конструкций нуклеиновых кислот, мутагенез, секвенирование, введение ДНК в клетки и экспрессия генов, и анализ белков подробно описаны в Current Protocols in Molecular Biology, Second Edition, Ausubel et al. eds., John Wiley & Sons, 1988, Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Ausubel et al. eds., John Wiley & Sons, 4th edition 1999. Описания Sambrook et al. и Ausubel et al. (оба) включены в данное описание в виде ссылки.
Таким образом, следующий аспект настоящего изобретения относится к клетке-хозяину, содержащей нуклеиновую кислоту, указанную в данном описании. Такая клетка-хозяин может находится в условиях in vitro и может быть в культуре. Такая клетка-хозяин может находится в условиях in vivo. Присутствие клетки-хозяина in vivo может обеспечивать возможность внутриклеточной экспрессии специфических связывающих партнеров согласно настоящему изобретению в виде «внутренних антител» или внутриклеточных антител. Внутренние антитела могут быть использованы для генной терапии [112].
Еще один аспект относится к способу, включающему в себя введение такой нуклеиновой кислоты в клетку-хозяина. Для введения можно использовать любой способ. В случае эукариотических клеток подходящие способы могут включать трансфекцию на основе фосфата кальция, способ с использованием DEAE-декстрана, электропорацию, опосредованную липосомами трансфекцию и трансдукцию с использованием ретровируса или другого вируса, например вируса вакцинии или, в случае клеток насекомых, бакуловируса. Для введения нуклеиновой кислоты в клетку-хозяина, в частности эукариотическую клетку, можно использовать вирусную или основанную на плазмиде систему. Плазмидная система может поддерживаться в виде эписом или может быть внедрена в хромосому клетки-хозяина или в искусственную хромосому [110, 111]. Внедрение может представлять собой либо случайную, либо целенаправленную интеграцию одной или нескольких копий в одном или во многих локусах. В случае бактериальных клеток подходящие способы могут включать трансформацию с использованием хлорида кальция, электропорацию и трансфекцию с использованием бактериофага.
После введения может быть вызвана экспрессия или обеспечена возможность экспрессии нуклеиновой кислоты, например, посредством культивирования клеток-хозяев в условиях экспрессии гена.
В одном варианте нуклеиновая кислота согласно изобретению интегрируется в геном (например, хромосому) клетки-хозяина. Интеграцию можно стимулировать введением последовательностей, которые стимулируют рекомбинацию с геномом, согласно стандартным способам.
Настоящее изобретение также относится к способу, который включает в себя использование конструкции, которая указана выше, в экспрессирующей системе, чтобы экспрессировать специфический связывающий партнер или полипептид, которые указаны выше.
Аспекты и варианты осуществления настоящего изобретения теперь будут проиллюстрированы в виде примера со ссылкой на следующие эксперименты.
ПРИМЕР 1
Выделение анти-IL-13-scFv
Репертуар scFv-антител
Для отбора использовали большую библиотеку антител человека в виде одноцепочечных Fv (scFv), полученную из лимфоцитов селезенки от 20 доноров и клонированную в фагмидном векторе [66].
Отбор scFv
ScFv, которые узнают IL-13, выделяли из библиотек на основе фагового дисплея в серии повторяющихся циклов селекции с использованием рекомбинантного полученного в бактериях IL-13 человека или мыши (Peprotech), по существу как описано в [67]. Коротко, после инкубации с библиотекой иммобилизованный антиген, который был предварительно связан с парамагнитными шариками, и связанный фаг извлекали посредством магнитного разделения, при этом несвязанный фаг удаляли промывкой. Затем связанный фаг спасали, как описано Vaughan et al. [67], и процесс отбора повторяли. В разных раундах селекции использовали различные твердые поверхности и способы захвата, чтобы уменьшить неспецифичное связывание. Антиген либо ковалентно связывали с шариками (Dynabeads M-270, карбоновая кислота), либо модифицировали биотинилированием с последующим вторичным улавливанием на покрытых стрептавидином шариках (Dynabeads M-280) согласно протоколам производителя (Dynal). Часть типичных клонов, полученных в результате раундов селекции, подвергали секвенированию ДНК, как описано в Vaughan et al. [67] и Osbourn et al. [70]. Уникальные клоны анализировали в отношении их способности нейтрализовать IL-13 в виде очищенных препаратов scFv в анализах зависимой от IL-13 пролиферации клеток.
Создавали библиотеки на основе рибосомного дисплея и подвергали скринингу в отношении scFv, которые специфично узнают рекомбинантный, полученный в бактериях IL-13 человека или мыши (Peprotech), по существу, как описано в Hanes et al. [113]. Сначала лучший клон BAK278D6, полученный в результате в исходных отборах, превращали в форму рибосомного дисплея и затем полученную матрицу использовали для создания библиотеки. На уровне ДНК добавляли промотор T7 к 5'-концу для эффективной транскрипции мРНК. На уровне мРНК конструкция содержала прокариотический участок связывания рибосомы (последовательность Шайна-Далгарно). На 3'-конце одной цепи удаляли стоп-кодон и добавляли часть gIII (ген III) в качестве спейсера [113].
Библиотеки в рибосомном дисплее, полученные из BAK278D6, создавали мутагенезом определяющих комплементарность областей антитела (CDR), при этом осуществляли реакции ПЦР с полимеразой Tag, не обладающей корректирующей активностью. Осуществляли селекцию на основе аффинности, после инкубации с библиотекой биотинилированный IL-13 человека улавливали парамагнитными шариками, покрытыми стрептавидином (Dynal M280) и связанные тройные комплексы (мРНК-рибосома-scFv-IL-13) извлекали посредством магнитного разделения, при этом несвязанные комплексы удаляли промывкой. Затем мРНК, кодирующую связанные scFv, извлекали посредством ОТ-ПЦР, как описано в Hanes et al. [113], и процесс отбора повторяли с уменьшающимися концентрациями (100 нМ - 100 пМ на протяжении 5 раундов) биотинилированного IL-13 человека, присутствующего во время селекции.
Склонную к ошибкам ПЦР также использовали для дальнейшего увеличения размера библиотеки. В схеме селекции использовали три интенсивности ошибок (2,0, 3,5 и 7,2 мутации на 1000 п.о. после стандартной реакции ПЦР, которая описана в протоколе производителя (Clontech)). Исходные реакции склонной к ошибкам ПЦР имели место перед циклом из одной селекции, начинаемой при 100 нМ. Следующий раунд склонной к ошибкам ПЦР осуществляли перед циклом из трех селекций при 10 нМ биотинилированного IL-13 человека. Как указано выше, часть типичных клонов, полученных в результате раундов селекции, подвергали секвенированию ДНК, как описано в Vaughan et al. [67] и Osbourn et al. [70]. Уникальные клоны анализировали в отношении их способности нейтрализовать IL-13 в виде очищенных препаратов scFv в анализах зависимой от IL-13 пролиферации клеток.
ПРИМЕР 2
Нейтрализующая эффективность анти-IL-13-scFv в анализе зависимой от IL-13 пролиферации клеток TF-1
Нейтрализующую эффективность очищенных препаратов scFv против биоактивности IL-13 человека или мыши оценивали, используя анализ пролиферации клеток TF-1. Очищенные препараты scFv получали, как описано в примере 3 в WO 01/66754. Концентрации белка очищенных препаратов scFv определяли, используя способ BCA (Pierce). TF-1 представляет собой линию премиелоидных клеток человека, полученных от пациента с эритролейкозом [68]. Линия клеток TF-1 является фактор-зависимой в отношении выживаемости и пролиферации. В этой связи клетки TF-1, отвечающие либо на человеческий, либо на мышиный IL-13 [69], поддерживали в среде, содержащей GM-CSF человека (4 нг/мл, R and D Systems). Ингибирование зависимой от IL-13 пролиферации определяли посредством измерения снижения включения меченного тритием тимидина в новосинтезированную ДНК делящихся клеток.
Протокол анализа клеток TF-1
Клетки TF-1 получали из R and D Systems и поддерживали согласно прилагаемым протоколам. Среды для анализа содержали RPMI-1640 с GLUTAMAX I (Invitrogen), содержащую 5% фетальную сыворотку теленка (JRH) и 1% пируват натрия (Sigma). Перед каждым анализом клетки TF-1 осаждали центрифугированием при 300×g в течение 5 мин, среду удаляли отсасыванием и клетки ресуспендировали в среде для анализа. Указанный процесс повторяли дважды и клетки ресуспендировали в конечной концентрации 105 клеток/мл в среде для анализа. Тестируемые растворы антитела (в трех повторах) разбавляли до требуемой концентрации в среде для анализа. В качестве негативного контроля использовали нерелевантное антитело, не направленное к IL-13. Рекомбинантный полученный в бактериях IL-13 человека или мыши (Peprotech) добавляли до конечной концентрации 50 нг/мл при смешивании с подходящим тестируемым антителом в общем объеме 100 мкл/лунку в 96-луночном планшете для анализа. Концентрацию IL-13, используемую в анализе, выбирали в такой дозе, которая при конечной концентрации в анализе давала примерно 80% от максимального пролиферативного ответа. Все образцы инкубировали в течение 30 минут при комнатной температуре. Затем в каждую анализируемую точку добавляли 100 мкл ресуспендированных клеток, получая общий объем анализа 200 мкл/лунку. Анализируемые планшеты инкубировали в течение 72 часов при 37°C в 5% CO2. Затем в каждую анализируемую точку добавляли 25 мкл меченного тритием тимидина (10 мкКюри/мл, NEN) и анализируемые планшеты возвращали в инкубатор еще на 4 часа. Клетки собирали на фильтровальные пластины из стекловолокна (Perkin Elmer), используя устройство для сбора клеток. Включение тимидина определяли, используя жидкостной сцинтилляционный счетчик для микропланшетов Packard TopCount. Данные анализировали, используя компьютерную программу Graphpad Prism.
Результаты
Несмотря на периодические циклы селекции с использованием антигенов человека и мыши, не получили перекрестно реагирующих нейтрализующих антител. В результате селекции получили два различных нейтрализующих scFv против IL-13 человека и один против IL-13 мыши. BAK278D6 (VH SEQ ID NO: 13; VL SEQ ID NO: 14) и BAK167A11 (VH SEQ ID NO: 23; VL SEQ ID NO: 24) узнавали IL-13 человека, тогда как BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26) узнавало IL-13 мыши. BAK278D6 (фиг.2) и BAK167A11 (фиг.1) в виде scFv нейтрализовали 25 нг/мл IL-13 человека с IC50, составляющей 44 нМ и 111 нМ соответственно. BAK209B11 (фиг.3) в виде scFv нейтрализовали 25 нг/мл IL-13 мыши с IC50, составляющей 185 нМ.
ПРИМЕР 3
Нейтрализующая эффективность лучших клонов, полученная в результате целенаправленной оптимизации CDR3 тяжелой цепи исходных клонов, в анализе зависимой от IL-13 пролиферации клеток TF-1
Osbourn et al. [70] показали, что целенаправленный мутагенез остатков в CDR3 тяжелой цепи может в значительной степени повысить аффинность антител. Селекцию осуществляли, как описано в примере 1, на репертуаре scFv, в котором остатки в CDR3 тяжелой цепи CDR3 BAK278D6 (SEQ ID NO: 6), BAK167A11 (SEQ ID NO: 57) были рандомизированы мутагенезом. Уникальные клоны, полученные в результате селекции, идентифицировали секвенированием ДНК и их нейтрализующую эффективность оценивали в виде scFv в анализе пролиферации клеток TF-1, как описано в примере 2.
Результаты
Значительного увеличения эффективности достигали в обеих линиях. Наиболее эффективными клонами из линии BAK167A11 были BAK615E3, BAK612B5 и BAK582F7, которые в виде scFv имели IC50, составляющую 3 нМ (фиг.1), 6,6 нМ, 6,65 нМ соответственно против 25 нг/мл IL-13 человека в анализе пролиферации клеток TF-1. Из линии BAK278D6 наиболее эффективным был клон BAK502G9, который в виде scFv имел IC50, равную 8 нМ, против 25 нг/мл IL-13 человека в анализе пролиферации клеток TF-1 (фиг.2).
ПРИМЕР 4
Нейтрализующая эффективность линий BAK167A11 и BAK278D6 против IL-13 приматов, отличных от человека, и варианта IL-13, связанного с астмой, в анализе зависимой от фактора пролиферации клеток TF-1
Ни линия BAK167A11, ни линия BAK278D6, нейтрализующие IL-13 человека, не были перекрестно реагирующими по отношению к мыши.
Поэтому авторы изобретения выбрали следующие критерии для линии, отбираемой для дальнейшей оптимизации и клинической разработки: она должна быть предпочтительно перекрестно реагирующей с IL-13 примата, отличного от человека, и должна узнавать вариант IL-13, в котором аргинин в положении аминокислоты 130 заменен глутамином (R130Q). Указанный вариант генетически связан с астмой и другими аллергическими заболеваниями [37, 39, 41, 71]. Перекрестную реактивность определяли по способности очищенных препаратов scFv связываться с IL-13 примата, отличного от человека, и вариантом IL-13 с использованием анализа резонанса поверхностного плазмона (BIAcore). Функциональную активность определяли с использованием анализа пролиферации клеток TF-1.
Получение IL-13 дикого типа, варианта и IL-13 примата, отличного от человека
кДНК IL-13 человека дикого типа получали из InvivoGen и модифицировали сайт-направленным мутагенезом (набор Stratagene Quikchange®), чтобы получить кДНК, кодирующую вариант IL-13. Кодирующую последовательность как для IL-13 макак-резус, так и для IL-13 макак-крабоедов получали с помощью ПЦР на матрице геномной ДНК, используя вырожденные праймеры, основанные на последовательности IL-13 человека. Обе последовательности приматов, отличных от человека (резус и крабоедов), были идентичны друг другу, но отличались от IL-13 человека семью аминокислотами (фиг.19). Затем экспрессировали рекомбинантный IL-13 дикого типа, вариант и IL-13 примата, отличного от человека, используя бакуловирусную систему экспрессии (Invitrogen). К экспрессирующей конструкции добавляли аффинную метку на карбоксильном конце, чтобы экспрессировать белок, который обеспечивает возможность очистки из кондиционированной среды клеток насекомых почти до гомогенности.
Количественный анализ связывания с использованием BIAcore
Аффинность связывания очищенных препаратов scFv по отношению к IL-13 примата, отличного от человека, варианту и IL-13 дикого типа определяли посредством измерений резонанса поверхностного плазмона с использованием биосенсора BIAcore 2000 (BIAcore AB), как описано в Karlsson et al. [72]. Коротко, IL-13 связывали с сенсорными чипами СМ5, используя набор для связывания амина (BIAcore), с поверхностной плотностью примерно 200 усл.ед., и три концентрации тестируемого scFv (примерно 350 нМ, 175 нМ и 88 нМ) в НВ3-ЕР-буфере пропускали через поверхность сенсорного чипа. Полученные в результате сенсограммы оценивали с использованием компьютерной программы для оценки BIA 3.1, получая относительные данные о связывании.
Протокол анализа TF-1
Анализ осуществляли, по существу, как описано в примере 2, со следующими модификациями: IL-13 примата, отличного от человека, вариант IL-13 человека (R130Q) и IL-13 человека дикого типа использовали в концентрациях 50 нг/мл, 25 нг/мл и 25 нг/мл соответственно.
Результаты
Результаты анализа связывания BIAcore свидетельствуют, что линия BAK278D6, но не линия ВАК167А11, имела требуемый профиль перекрестной реактивности для дальнейшей разработки в терапевтических целях (таблица 2). Полученные данные подтверждались результатами биоанализа, свидетельствующими, что BAK278D6 (фиг.4) и BAK502G9 (фиг.6) были способны нейтрализовать IL-13 человека, вариант IL-13 человека (R130Q) и IL-13 примата, отличного от человека, в анализе пролиферации клеток TF-1 почти с равной эффективностью. Напротив, хотя ВАК615ЕЗ (VH SEQ ID NO: 33; VL SEQ ID NO: 34) обладали в значительной степени повышенной эффективностью против IL-13 человека по сравнению с исходным ВАК167А11 (VH SEQ ID NO: 23; VL SEQ ID NO: 24) в анализе пролиферации клеток TF-1 (фиг.1), ни один из клонов не связывал IL-13 примата, отличного от человека, или вариант IL-13 в анализе связывания BIAcore.
Каркасные области BAK278D6 и BAK502G9 зародышевой линии
Полученную аминокислотную последовательность VH BAK278D6 (SEQ ID NO: 13) и VL (SEQ ID NO: 14) выравнивали с известными последовательностями зародышевой линии человека в базе данных VBASE [73] и идентифицировали наиболее близкую зародышевую линию по сходству последовательностей. Наиболее близкую последовательность зародышевой линии для домена VH BAK278D6 (SEQ ID NO: 14) и его производных идентифицировали как DP14, член семейства VH1. VH BAK278D6 имеет 9 отличий от DP14 зародышевой линии в каркасных областях. Наиболее близкую последовательность зародышевой линии для VL BAK278D6 идентифицировали как Vλ3 3h. VL-домен BAK278D6 (SEQ ID NO: 13) имеет только 5 изменений по сравнению с зародышевой линией в каркасных областях. Каркасные области BAK278D6 и его производных возвращали к областям зародышевой линии сайт-направленным мутагенезом (набор Stratagene Quikchange) для идентичного соответствия с нативным антителом человека.
ПРИМЕР 5
Нейтрализующая эффективность лучших клонов, полученных в результате целенаправленной оптимизации последовательностей CDR1 тяжелой цепи и CDR2 тяжелой цепи BAK502G9 в анализе зависимой от IL-13 человека пролиферации клеток TF-1
Вторую фазу оптимизации осуществляли с использованием в качестве матрицы последовательности BAK502G9 с каркасными областями, приближенными к зародышевой линии. Отбор осуществляли, по существу, как описано в примере 1, из репертуара scFv, в котором любые остатки в CDR1 тяжелой цепи или CDR2 тяжелой цепи BAK502G9 подвергали рандомизации мутагенезом. Уникальные клоны, полученные в результате селекции, идентифицировали секвенированием ДНК и оценивали их нейтрализующую эффективность в виде очищенных препаратов scFv в анализе пролиферации клеток TF-1, как описано в примере 2. Конструировали векторы для наиболее эффективных клонов scFv, чтобы обеспечить возможность повторной экспрессии в виде целого IgG4-антитела человека, как описано в Persic et al. (1997 Gene 187; 9-18), с небольшими модификациями. В векторы включали фрагмент oriP, чтобы облегчить применение с клетками HEK-EBNA 293 и обеспечить репликацию эписом. Вариабельный домен VH клонировали в полилинкере между лидерной последовательностью секреции и константной областью гамма 4 человека в экспрессирующем векторе pEU8.1(+). Вариабельный домен VL клонировали в полилинкере между лидерной последовательностью секреции и константной областью лямбда человека экспрессирующего вектора pEU4.1(-).
Целое антитело очищали из кондиционированной среды клеток EBNA-293, совместно трансфицированных конструкциями, экспрессирующими тяжелые и легкие цепи, посредством аффинной хроматографии с использованием белка A (Amersham Pharmacia). Очищенные препараты антител стерильно фильтровали и хранили при 4°C в фосфатно-солевом буфере (PBS) до проведения оценки. Концентрацию белка определяли измерением поглощения при 280 нм с использованием способа BCA (Pierce). Преобразованные целые антитела IgG4 человека сравнивали с коммерчески доступными анти-IL-13-антителами человека в анализе пролиферации TF-1, описанном в примере 2.
Результаты
Как проиллюстрировано на фиг.5, показали, что коммерческое антитело B-B13, (мышиный IgG1 - Euroclone 5) значительно более эффективно против IL-13 человека, чем коммерческое антитело JES10-5A2 (IgG1 крыс - Biosource) с IC50, составляющей 1021 пМ и 471 пМ соответственно. Восемь клонов, а именно BAK1111D10, BAK1166G02, BAK1167F02, BAK1167F04, BAK1183H4, BAK1184C8, BAK1185E1, BAK1185F8, полученных из BAK502G9 (и поэтому «линия BAK502G9»), в которых CDR1 или CDR2 тяжелой цепи были целенаправленно изменены, проявляли повышенную эффективность в виде scFv по сравнению с коммерческими антителами. Указанное повышение сохранялось при превращении в целое IgG4-антитело человека. Каждый из указанных доменов VH и VL отдельно и в соответствующих парах согласно указанным пунктам формулы изобретения составляет аспект или вариант осуществления настоящего изобретения, также как специфические связывающие партнеры для IL-13, которые содержат один или несколько из указанных доменов, а также специфические связывающие партнеры, содержащие одну или несколько CDR из клонов линии BAK502G9, предпочтительно домен VH, содержащий набор HCDR линии BAK502G9 и/или домен VL, содержащий набор LCDR линии BAK502G9. Указанные домены могут быть использованы в любом и во всех аспектах изобретения, которые указаны на протяжении данного описания. Производные BAK502G9 в виде целых антител (IgG4) имеют IC50 в диапазоне от 244 пМ до 283 пМ. BAK502G9 в виде целого антитела IgG4 имеет IC50 384 пМ. В заключение, основные улучшения эффективности могут быть получены целенаправленным изменением CDR1 (SEQ ID NO: 7) или CDR2 (SEQ ID NO: 8) тяжелой цепи BAK502G9. Статистические сравнения с B-B13 осуществляли с использованием ANOVA с последующим анализом апостериорного критерия Даннета (компьютерная программа InStat).
Дополнительная характеристика
Выбранные анти-человеческие антитела из линии BAK278D6 подвергали дополнительной характеристике, чтобы определить их специфичность. Указанные антитела включали BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16) и его производные BAK1167F2 (VH SEQ ID NO: 35; VL SEQ ID NO: 36) и BAK1183H4 (VH SEQ ID NO: 37; VL SEQ ID NO: 38), которые представляют собой типичные примеры клонов с модификациями в CDR1 тяжелой цепи и CDR2 тяжелой цепи BAK502G9 соответственно.
ПРИМЕР 6
Нейтрализующая эффективность лучших клонов, полученных в результате целенаправленной оптимизации последовательностей CDR1 тяжелой цепи и CDR2 тяжелой цепи BAK502G9, против IL-13 приматов, отличных от человека, и варианта IL-13, связанного с астмой, в анализе зависимой от факторов пролиферации клеток TF-1
Перекрестную реактивность антител против IL-13 человека определяли по их способности ингибировать опосредованную IL-13 примата, отличного от человека, и вариантом IL-13 пролиферацию клеток TF-1, как описано в примере 4.
Результаты
Оптимизированные антитела против IL-13 человека BAK1167F2 (VH SEQ ID NO: 35; VL SEQ ID NO: 36) и BAK1183H4 (VH SEQ ID NO: 37; VL SEQ ID NO: 38) сохраняли специфичность своего исходного BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16) (фиг.6). Увеличение эффективности против IL-13 дикого типа отражалось на их способности нейтрализовать IL-13 примата, отличного от человека, и вариант IL-13 по существу с равной эффективностью. IC50 в случае BAK502G9 против IL-13 человека, варианта IL-13 человека и IL-13 примата, отличного от человека, составляли 1,4 нМ, 1,9 нМ и 2,0 нМ соответственно. IC50 BAK1167F2 против IL-13 человека, варианта IL-13 человека и IL-13 примата, отличного от человека, составляли 1,0 нМ, 1,1 нМ и 1,3 нМ соответственно. IC50 BAK1183H4 против IL-13 человека, варианта IL-13 человека и IL-13 примата, отличного от человека, составляли 0,9 нМ, 1,0 нМ и 1,6 нМ соответственно. Указанные клоны подходят для терапевтического применения.
ПРИМЕР 7
Нейтрализующая эффективность лучших антител против IL-13 человека, направленных против нативного IL-13 человека в анализе пролиферации клеток HDLM-2
Последовательность IL-13 человека имеет четыре возможных сайта N-гликозилирования. Авторы изобретения показали способность BAK278D6 и его производных нейтрализовать рекомбинантный IL-13, экспрессируемый либо бактериальной, либо в бакуловирусной системах экспрессии. Хотя существует доказательство того, что многие события процессинга, известные в системах млекопитающих, также происходят у насекомых, имеются ключевые отличия в гликозилировании белков, в частности в N-гликозилировании [74].
Авторы изобретения исследовали способность производных BAK278D6 нейтрализовать нативный IL-13, высвобождаемый из клеток человека.
Клетки HDLM-2 выделяли по Drexler et al. [75] от пациента с болезнью Ходжкина. Skinnider et al. [76] показали, что пролиферация клеток HDLM-2 отчасти зависела от аутокринного и паракринного высвобождения IL-13. Лучшие антитела против IL-13 человека оценивали по их способности ингибировать пролиферацию клеток HDLM-2, опосредованную высвобождением нативного (или встречающегося в природе) IL-13.
Протокол анализа клеток HDLM-2
Клетки HDLM-2 получали из Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) и поддерживали согласно прилагаемым протоколам. Среда для анализа состояла из RPI-1640 с Glutamax I (Invitrogen), содержащей 20% фетальной сыворотки теленка. Перед каждым анализом клетки осаждали центрифугированием при 300×g в течение 5 мин, среду удаляли отсасыванием и клетки ресуспендировали в свежей среде. Указанный процесс повторяли три раза и, наконец, клетки ресуспендировали до конечной концентрации 2×105 клеток/мл в среде для анализа. 50 мкл ресуспендированных клеток добавляли в каждую точку для анализа в 96-луночном планшете для анализа. Тестируемые растворы антитела (в трех повторах) разбавляли до требуемой концентрации в среде для анализа. В качестве негативного контроля использовали антитело нерелевантного изотипа, не направленное к IL-13. К клеткам добавляли соответствующее тестируемое антитело в общем объеме 50 мкл/лунку, при этом в каждой точке для анализа получали общий объем для анализа 100 мкл/лунку. Анализируемые планшеты инкубировали в течение 72 часов при 37°C в 5% CO2. Затем в каждую анализируемую точку добавляли 25 мкл меченного тритием тимидина (10 мккюри/мл, NEN) и анализируемые планшеты возвращали в инкубатор еще на 4 часа. Клетки собирали на фильтровальные пластины из стекловолокна (Perkin Elmer), используя устройство для сбора клеток. Включение тимидина определяли, используя жидкостной сцинтилляционный счетчик для микропланшетов Packard TopCount. Данные анализировали, используя компьютерную программу Graphpad Prism.
Результаты
Как показано на фиг.7, BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16) и его производные BAK1183H4 (VH SEQ ID NO: 37; VL SEQ ID NO: 38) и BAK1167F2 (VH SEQ ID NO: 35; VL SEQ ID NO: 36) были способны вызывать зависимое от дозы ингибирование пролиферации клеток с относительными эффективностями, сходными с эффективностями, наблюдаемыми в биоанализах. IC50 для BAK502G9, BAK1183H4, BAK1167F2 в виде IgG4 человека составляли 4,6 нМ, 3,5 нМ и 1,1 нМ соответственно. IC50 для коммерческих антител JES10-5A2 и B-B13 составляли 10,7 нМ и 16,7 нМ соответственно.
ПРИМЕР 8
Нейтрализующая эффективность лучших антител против IL-13 человека по отношению к зависимым от IL-13 ответам в первичных клетках, имеющих отношение к заболеванию
Вторичные биоанализы осуществляли с использованием первичных клеток и индикаторов, имеющих большее отношение к заболеванию дыхательных путей. Такие индикаторы включали высвобождение эотаксина из нормальных фибробластов легких человека (NHLF) и повышенную регуляцию молекул адгезии сосудистых клеток 1 (VCAM-1) на поверхности эндотелиальных клеток пупочной вены человека (HUVEC). Оба зависимых от IL-13 ответа могли вносить вклад в мобилизацию эозинофилом, признак фенотипа астмы [92].
Протокол анализа NHLF
Показано, что IL-13 вызывает высвобождение эотаксина из фибробластов легких [77], [78], [79]. Зависимое от фактора высвобождение эотаксина из NHLF определяли в ELISA.
NHLF получали из Biowhittaker и поддерживали согласно прилагаемым протоколам. Средой для анализа была среда FGM-2 (Biowhittaker). Тестируемые растворы антитела (в трех повторах) разбавляли до требуемой концентрации в среде для анализа. В качестве негативного контроля использовали нерелевантное антитело, не направленное к IL-13. Затем добавляли рекомбинантный полученный в бактериях IL-13 человека (Peprotech) до конечной концентрации 10 нг/мл при смешивании с подходящим тестируемым антителом в общем объеме 200 мкл. Концентрацию IL-13, используемую в анализе, выбирали в такой дозе, которая давала примерно 80% от максимального ответа. Все образцы инкубировали в течение 30 минут при комнатной температуре. Затем анализируемые образца добавляли к NHLF, которые предварительно высевали при плотности 1×104 клеток на лунку в 96-луночные планшеты для анализа. Анализируемые планшеты инкубировали при 37°C в течение 16-24 часов в 5% CO2. Планшеты для анализа центрифугировали при 300×g в течение 5 минут, чтобы осадить открепившиеся клетки. Уровни эотаксина в надосадках определяли в ELISA, используя реагенты и способы, описанные производителем (R and D Systems). Данные анализировали, используя компьютерную программу Graphpad Prism.
Результаты
Клоны линии BAK278D6 обладали способностью ингибировать зависимое от IL-13 человека высвобождение эотаксина из NHLF. Относительная эффективность была сходной с эффективностью, наблюдаемой в анализе пролиферации клеток TF-1 (фиг.8). BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16), BAK1183H4 (VH SEQ ID NO: 37; VL SEQ ID NO: 38), BAK1167F2 (VH SEQ ID NO: 35; VL SEQ ID NO: 36) имели IC50 207 пМ, 118 пМ и 69 пМ соответственно против 10 нг/мл IL-13 человека. Коммерческие антитела JES10-5A2 и B-B13 имели IC50 623 пМ и 219 пМ соответственно.
Протокол анализа HUVEC
Показано, что IL-13 повышает экспрессию VCAM-1 на поверхности клеток HUVEC [80, 81]. Зависимую от фактора экспрессию VCAM-1 определяли, регистрируя повышающую регуляцию клеточной экспрессии рецептора VCAM-1, используя данные флуоресценции с разрешением во времени.
HUVEC получали из Biowhittaker и поддерживали согласно прилагаемым протоколам. Средой для анализа была среда EGM-2 (Biowhittaker). Тестируемые растворы антитела (в трех повторах) разбавляли до требуемой концентрации в среде для анализа. В качестве негативного контроля использовали нерелевантное антитело, не направленное к IL-13. Рекомбинантный полученный в бактериях IL-13 человека (Peprotech) добавляли до конечной концентрации 10 нг/мл при смешивании с подходящим тестируемым антителом в общем объеме 200 мкл. Концентрацию IL-13, используемую в анализе, выбирали в такой дозе, которая давала примерно 80% от максимального ответа. Все образцы инкубировали в течение 30 минут при комнатной температуре. Затем анализируемые образца добавляли к HUVEC, которые предварительно высевали при плотности 4×104 клеток на лунку в 96-луночные планшеты для анализа. Анализируемые планшеты инкубировали при 37°C в течение 16-20 часов в 5% CO2. Затем среду для анализа удаляли отсасыванием и заменяли блокирующим раствором (PBS, содержащий 4% сухого порошка молока Marvel®). Планшеты для анализа инкубировали при комнатной температуре в течение 1 часа. Лунки три раза промывали PBST с твином, затем в каждую лунку добавляли 100 мкл (разведение 1:500 в PBST/1% Marvel®) биотинилированного анти-VCAM-1-антитела (Serotec). Планшеты для анализа инкубировали при комнатной температуре в течение 1 часа. Лунки три раза промывали буфером для промывки Delfia (Perkin Elmer), затем в каждую лунку добавляли 100 мкл меченного европием стрептавидина или антитела против IgG1 мыши (разведение 1:1000 в буфере для анализа Delfia, Perkin Elmer). Затем планшеты для анализа инкубировали при комнатной температуре в течение 1 часа. Лунки 7 раз промывали буфером для промывки Delfia (Perkin Elmer). Наконец в каждую лунку добавляли 100 мкл усиливающего раствора (Perkin Elmer) и определяли интенсивность флуоресценции, используя устройство для считывания планшетов Wallac 1420 VICTOR2 (стандартный протокол для европия). Данные анализировали, используя компьютерную программу Graphpad Prism.
Результаты
Типичные данные для BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16), BAK1183H4 (VH SEQ ID NO: 37; VL SEQ ID NO: 38), BAK1167F2 (VH SEQ ID NO: 35; VL SEQ ID NO: 36) в виде целого антитела IgG4 человека показаны на фиг.9. Относительная эффективность была сходна с эффективностью, наблюдаемой в анализе пролиферации клеток TF-1. IC50 для BAK502G9, BAK1183H4 и BAK1167F2 составляли 235 пМ, 58 пМ и 55 пМ соответственно против 10 нг/мл IL-13 человека.
ПРИМЕР 9
Нейтрализующая эффективность анти-IL-13-антител по отношению к зависимой от IL-1β и IL-4 повышающей регуляции VCAM-1
Специфичность клонов линии BAK278D6 оценивали в модифицированном биоанализе HUVEC. Показано, что наряду с IL-13 и IL-4 и IL-1β увеличивают экспрессию VCAM-1 на поверхности клеток HUVEC [80, 81].
Протокол анализа HUVEC
Анализ осуществляли, по существу, как описано в примере 5, со следующими модификациями. Рекомбинантный IL-1β человека и IL-4 (R and D Systems) использовали вместо IL-13 человека в концентрации 0,5 нг/мл и 1 нг/мл соответственно и вводили в дозе, которая давала примерно 80% максимального ответа.
Результаты
Ни один из оцениваемых клонов линии BAK278D6 не нейтрализовал повышающую регуляцию VCAM-1 в ответ на IL-1β или IL-4 человека и таким образом проявлял специфичность в отношении IL-13 (фиг.10). IL-4 является наиболее близко родственным IL-13, проявляя 30% идентичность последовательностей на уровне аминокислот [82].
ПРИМЕР 10
Нейтрализующая эффективность BAK209B11 в виде IgG4 человека в анализе зависимой от IL-13 мыши пролиферации мышиных клеток B9
BAK209B11, идентифицированный как нейтрализующий клон против IL-13 мыши в виде scFv, как описано в примере 1, преобразовывали в целое антитело в виде IgG4 человека, как описано в примере 5, и оценивали его эффективность в анализе зависимой от IL-13 мыши пролиферации клеток B9. B9 представляет собой линии клеток B-клеточной гибридомы мышей [83]. B9 зависит от факторов в отношении выживаемости и пролиферации. В этой связи B-клетки, отвечающие на IL-13 мыши, поддерживали в среде, содержащей IL-6 человека (50 пг/мл, R and D Systems). Ингибирование зависимой от IL-13 мыши пролиферации определяли посредством измерения снижения включения меченного тритием тимидина в новосинтезированную ДНК делящихся клеток.
Протокол анализа клеток B9
Клетки B9 получали из Европейской коллекции культур клеток животных ECACC и поддерживали согласно прилагаемым протоколам. Анализ осуществляли, по существу, как описано для анализа TF-1 в примере 2, но со следующими модификациями. Среда для анализа состояла из RPMI-1640 с GLUTAMAX I (Invitrogen), содержащей 5% фетальной сыворотки теленка (Hyclone) и 50 мкМ 2-меркаптоэтанола (Invitrogen). IL-13 человека заменяли рекомбинантным полученным в бактериях IL-13 мыши (Peprotech) в конечной концентрации в анализе 1 нг/мл.
Результаты
BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26) в виде IgG4 человека нейтрализовали 1 нг/мл IL-13 мыши с IC50, равной 776 пМ, в анализе B9 (фиг.11). Таким образом, BAK209B11 представляет собой полезное средство для исследования роли IL-13 в мышиных моделях заболевания. Это ясно показано в примере 12, который демонстрирует эффективность BAK209B11 в мышиной модели острого воспаления легких.
ПРИМЕР 11
Определение аффинности анти-IL-13-антитела в анализе BIAcore
Аффинность BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16), BAK1167F2 (VH SEQ ID NO: 35; VL SEQ ID NO: 36) и BAK1183H4 (VH SEQ ID NO: 37; VL SEQ ID NO: 38) по отношению к IL-13 человека и BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26) по отношению к IL-13 мыши в виде IgG4 человека определяли посредством измерений резонанса поверхностного плазмона с использованием биосенсора BIAcore 2000 (BIAcore AB), по существу, как описано в [72]. Коротко, антитела связывали с сенсорными чипами CM5, используя набор для связывания амина (BIAcore), с поверхностной плотностью примерно 500 усл. ед., и серийные разведения IL-13 (от 50 нМ до 0,78 нМ) в HBS-EP-буфере пропускали через поверхность сенсорного чипа. Полученные в результате сенсограммы оценивали с использованием компьютерной программы для оценки BIA 3.1, получая кинетические данные.
Результаты
IgG4 BAK502G9, BAK1167F2 и BAK1183H4 связывали IL-13 человека с высокой аффинностью с Kd 178 пМ, 136 пМ и 81 пМ соответственно, что соответствует их относительной эффективности в основанных на клетках анализах. BAK209B11 связывали IL-13 мыши с аффинностью 5,1 нМ (таблица 3).
ПРИМЕР 12
Эффективность BAK209B11 в мышиной модели острого аллергического воспаления легких
Мышиная модель острого аллергического воспаления легких
Влияние BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26), нейтрализующего IgG4-антитела человека против IL-13 мыши, исследовали у мышей с острым аллергическим воспалением легких. Данную модель осуществляли, по существу, как описано Riffo-Vasquez et al. [84], и конечный результат характеризовали по увеличению содержания IL-13 в бронхоальвеолярном лаваже (BAL) (фиг.12), инфильтрации клеток в легкое и BAL (фиг.13), повышенным уровням IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Самок мышей Balb/C (Charles River UK) обрабатывали либо антителом BAK209B11 против IL-13 мыши (в дозах 12, 36, 119 или 357 мкг), либо совпадающим по изотипу контрольным антителом (доза 357 мкг). В 0 и 7 дни мышей в каждой группе сенсибилизировали внутрибрюшинной инъекцией 10 мкг овальбумина (Ova) в 0,2 мл наполнителя (физиологический раствор, содержащий 2% Al2O3 (Rehydragel) в качестве адъюванта). Отдельная контрольная группа несенсибилизированных мышей получала равный объем наполнителя. Мышей подвергали провокации овальбумином на 14, 15 и 16 день. Овальбумин разбавляли до 1% (мас./об.) в стерильном физиологическом растворе перед распылением. Все провокации посредством ингаляции проводили в камере для обработки из оргстекла. Ova распрыскивали в виде аэрозоля, используя распылитель deVilbiss Ultraneb 2000 (Sunrise Medical) в виде серии из трех воздействий по 20 минут, разделенных 1-часовыми интервалами.
BAK209B11 или нерелевантный IgG4 человека вводили внутривенно за 1 день до первой провокации и затем за 2 часа до каждой последующей провокации (всего 4 дозы). Модель заканчивали на 17 день через 24 часа после последней провокации. Собирали кровь (сыворотку) и BAL. Сыворотку анализировали в отношении суммарного IgE. BAL получали, инъецируя 3 аликвоты физиологического раствора (0,3 мл, 0,3 мл и 0,4 мл) и объединяя образцы. Подсчет суммарных лейкоцитов и дифференциальный подсчет клеток получали на основе клеток BAL.
Результаты
Провокация овальбумином сенсибилизированных мышей вызывала значимое (p<0,05) увеличение мобилизации суммарных клеток в BAL по сравнению с несенсибилизированными, но подвергнутыми провокации животными. Указанная мобилизация зависимым от дозы образом ингибировалась BAK209B11; значимое (p<0,05) ингибирование наблюдали при ≥36 мкг BAK209B11, но не контрольного антитела (фиг.13). Сходные эффекты также наблюдали на эозинофилах (фиг.14) и нейтрофилах (фиг.15) со значимым (p<0,05) ингибированием притока клеток при минимальной дозе BAK209B11 36 мкг. Указанное ингибирование не наблюдали в случае контрольного антитела. Лимфоциты также были индуцированы при провокации у сенсибилизированных мышей и не индуцировались у несенсибилизированных мышей. Указанная индукция зависимым от дозы образом ингибировалась BAK209B11 с максимальным ингибированием, наблюдаемым при 36 мкг BAK209B11. Контрольное антитело не оказывало влияния (фиг.16). Хотя моноциты/макрофаги не индуцировались у сенсибилизированных животных по сравнению с несенсибилизированными животными, исходные уровни были понижены при действии ≥36 мкг BAK209B11, но не понижены при действии контрольного антитела (фиг.17). Уровни IgE в сыворотке были значимо повышены у сенсибилизированных животных по сравнению с несенсибилизированными после провокации (p<0,05). Указанное увеличение снижалось после обработки 36 мкг BAK209B11, но не контрольным антителом.
В заключение, системное введение BAK209B11, антитела, нейтрализующего мышиный IL-13, но не контрольного антитела, ингибировало приток воспалительных клеток и повышающую регуляцию уровней IgE в сыворотке, вызванную сенсибилизацией и последующей провокацией овальбумином в мышиной модели аллергического воспаления.
Примеры 13-20 являются прогностическими
ПРИМЕР 13
Эффективность BAK209B11 в мышиной модели Ллойда острого воспаления легких
Мышиная модель острого аллергического воспаления легких
Влияние BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26), нейтрализующего антитела против IL-13 мыши, исследовали во второй мышиной модели острого аллергического воспаления легких. Данную модель осуществляли, по существу, как описано McMillan et al. [85], и характеризовали конечный результат по повышенному содержанию IL-13 в BAL и легочной ткани, клеточной инфильтрации в легкие и BAL, повышенным уровням содержания IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Самкам мышей Balb/C (Charles River UK) вводили разные дозы антитела BAK209B11 против IL-13 мыши или совпадающего по изотипу контрольного антитела следующим образом. В 0 и 12 день мышей в каждой группе сенсибилизировали (SN) внутрибрюшинной инъекцией 10 мкг овальбумина (Ova) в 0,2 мл наполнителя (физиологический раствор, содержащий 2 мг Al(OH)3 в качестве адъюванта [рассчитывали, как описано в примере 12]). Отдельная контрольная группа несенсибилизированных мышей (NS) получала равный объем наполнителя. Мышей подвергали провокации овальбумином в течение 20 минут в 19, 20, 21, 22, 23 и 24 день. Овальбумин разбавляли до 5% (мас./об.) в физиологическом растворе перед распылением. Все провокации посредством ингаляции проводили в камере для обработки из оргстекла. Ova распрыскивали в виде аэрозоля, используя распылитель deVilbiss Ultraneb 2000 (Sunrise Medical). На 18, 19, 20, 21, 22, 23 и 24 день мышам вводили разные внутрибрюшинные дозы (237 мкг, 23,7 мкг или 2,37 мкг; указанные на фиг.21 в виде H, M или L) антитела BAK209B11 в виде muIgG1 против IL-13 мыши или совпадающего по изотипу контрольного антитела (237 мкг). Функцию дыхательных путей оценивали в 0 и 25 день посредством провокации возрастающими количествами метахолина и регистрировали, используя плетизмографию животных, находящихся в сознании (Buxco). PC50 (концентрацию метахолина, требуемую для увеличения исходного уровня PenH на 50%) оценивали для отдельных мышей в 0 день и 25 день на основании подгонки кривых по 4 нефиксированным параметрам для кривых зависимости ответа от дозы метахолина.
Модель заканчивали на 25 день через 24 часа после последней провокации. Собирали образцы крови, сыворотки, BAL и легочной ткани.
Результаты
Функцию легких оценивали для отдельных животных в 0 день (перед обработкой) и на 25 день (после провокации) и количественно измеряли, рассчитывая значения PC50 (концентрации метахолина, требуемой для повышения исходного уровня PenH на 50%) (фиг.21A). Гиперчувствительность дыхательных путей отдельных животных (AHR) определяли по изменению log PC50 на 25 день по сравнению с 0 днем (log PC50 на 25 день - log PC50 в 0 день). Указанная разница log PC50 представляла собой основной конечный результат исследования; данные PC50 преобразовывали в виде log из-за требований ANOVA для конечного результата. Индивидуальные изменения усредняли по группам, чтобы получить среднюю разность log PC50 (которая показана на фиг.21B).
Провокация овальбумином сенсибилизированных мышей вызывала значимую AHR по сравнению с несенсибилизированными и подвергнутыми провокации мышами (p<0,01). BAK209B11 вызывали явное и зависимое от дозы снижение AHR, тогда как контрольное антитело не оказывало влияния.
ПРИМЕР 14
Эффективность BAK209B11 в мышиной модели Жерара острого воспаления легких
Мышиная модель острого аллергического воспаление легких
Влияние BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26), нейтрализующего антитела против IL-13 мыши в виде IgG4 человека, исследовали в третьей мышиной модели острого аллергического воспаления легких. Данную модель осуществляли, по существу, как описано Humbles et al. [86], и конечный результат характеризовали по повышенному содержанию IL-13 в BAL и легочной ткани, клеточной инфильтрации в легкие и BAL, повышенным уровням содержания IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Самкам мышей Balb/C (Charles River UK) вводили разные дозы антитела BAK209B11 против IL-13 мыши или совпадающего по изотипу контрольного антитела. В 0, 7 и 14 день мышей в каждой группе сенсибилизировали (SN) внутрибрюшинной инъекцией 10 мкг овальбумина (Ova) в 0,2 мл наполнителя (физиологический раствор, содержащий 1,125 мг Al(OH)3 в качестве адъюванта (рассчитывали, как описано в примере 12). Отдельная контрольная группа несенсибилизированных мышей (NS) получала равный объем наполнителя. Мышей подвергали провокации овальбумином в течение 20 минут в 21, 22, 23 и 24 день. Овальбумин разбавляли до 5% (мас./об.) в физиологическом растворе перед распылением. Все провокации посредством ингаляции проводили в камере для обработки из оргстекла. Ova распрыскивали в виде аэрозоля, используя распылитель deVilbiss Ultraneb 2000 (Sunrise Medical).
Модель заканчивали на 25 день через 24 часа после провокации. Собирали образцы крови, сыворотки, BAL и легочной ткани.
ПРИМЕР 15
Эффективность BAK209B11 в модели Ллойда хронического воспаления легких
Мышиная модель острого аллергического воспаление легких
Влияние BAK209B11 (VH SEQ ID NO: 25; VL SEQ ID NO: 26), нейтрализующего антитела против IL-13 мыши в виде IgG4 человека, исследовали в модели острого аллергического воспаления легких. Данную модель осуществляли, по существу, как описано Temelkovski et al. [87], и конечный результат характеризовали по клеточной инфильтрации в легкие и BAL, повышенным уровням содержания IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Самкам мышей Balb/C (Charles River UK) вводили разные дозы антитела BAK209B11 против IL-13 мыши или совпадающего по изотипу контрольного антитела. В 0 и 11 день мышей в каждой группе сенсибилизировали (SN) внутрибрюшинной инъекцией 10 мкг овальбумина (Ova) в 0,2 мл наполнителя (физиологический раствор, содержащий 2 мг Al(OH)3 в качестве адъюванта (рассчитывали, как описано в примере 12). Отдельная контрольная группа несенсибилизированных мышей (NS) получала равный объем наполнителя. Мышей подвергали провокации овальбумином в течение 20 минут на 18, 19, 20, 21, 22, 23, 28, 30, 32, 35, 37, 39, 42, 44, 46, 49 и 51 день. Овальбумин разбавляли до 5% (мас./об.) в физиологическом растворе перед распылением. Все провокации посредством ингаляции проводили в камере для обработки из оргстекла. Ova распрыскивали в виде аэрозоля, используя распылитель deVilbiss Ultraneb 2000 (Sunrise Medical).
Модель заканчивали на 52 день через 24 часа после провокации. Собирали образцы крови, сыворотки, BAL и легочной ткани.
ПРИМЕР 16
Эффективность антител против IL-13 человека против экзогенного IL-13 человека, введенного в модель воздушных мешочков мышей
Влияние антител против IL-13 человека на противовоспалительное действие IL-13 человека исследовали в основной мышиной модели. Указанную модель осуществляли, по существу, как описано Edwards et al. [93], и характеризовали конечный результат по инфильтрации клеток в воздушный мешочек.
Протокол модели
Воздушный мешочек создавали на спине самок мышей Balb/C посредством подкожной инъекции 2,5 мл стерильного воздуха в 0 день. Воздушный мешочек снова надували дополнительным объемом 2,5 мл стерильного воздуха на 3 день. 2 мкг huIL-13 в 0,75% CMC инъецировали непосредственно в мешочек на 6 день. Через 24 часа мышей умерщвляли, воздушный мешочек промывали 1 мл физиологического раствора с гепарином. Обработки антителами либо проводили вместе с huIL-13 (в мешочек), либо проводили системно.
Результаты
IL-13 человека, инъецированный в воздушный мешочек (i. po.), вызывал значимое увеличение инфильтрации суммарных лейкоцитов (p<0,01) и эозинофилов (p<0,01) через 24 часа после провокации по сравнению с мышами, обработанными наполнителем (0,75% карбоксиметилцеллюлоза (CMC) в физиологическом растворе i. po.).
Локально введенное BAK502G9 (200 мг, 20 мг или 2 мг внутрь мешочка) значимо и зависимым от дозы образом ингибировало инфильтрацию суммарных лейкоцитов (p<0,01) и эозинофилов (p<0,01) в воздушный мешочек, вызванную 2 мкг huIL-13 в 0,75% CMC.
Системно введенное BAK209B11 (30 мг/кг, 10 мг/кг и 1 мг/кг) также значимо и зависимым от дозы образом ингибировало инфильтрацию суммарных лейкоцитов (p<0,01) и эозинофилов (p<0,01) в воздушный мешочек, вызванную 2 мкг huIL-13 в 0,75% CMC.
ПРИМЕР 17
Создание трансгенных мышей с внедренным геном IL-13 человека/нокаутированных по IL-13 мыши в целях оценки эффективности антитела против IL-13 человека в моделях аллергического воспаления легких
Авторы настоящего изобретения создали мышей, которые экспрессируют человеческий, а не мышиный IL-13, посредством целенаправленного изменения генов. Ген IL-13 мыши заменяли от стартового кодона до стоп-кодона соответствующей частью гена IL-13 человека. Полученная линия мышей экспрессирует IL-13 человека, а не мышиный IL-13, в ответ на такие же стимулы, как и мышь дикого типа, так как эндогенный промотор IL-13 и pA-хвост IL-13 оставались неизменными. Показано, что IL-13 человека может связываться и передавать сигнал через рецепторы IL-13 мыши, приводя к таким же физиологическим последствиям, как и передача сигнала, вызванная IL-13 мыши, связанным с рецепторами IL-13 мыши. Например, экзогенный IL-13 человека вызывал мобилизацию воспалительных клеток в воздушный мешочек мышей (фиг.18). Полученные трансгенные животные позволили авторам изобретения оценивать немышиные перекрестно реагирующие антитела против IL-13 человека в разработанных мышиных моделях заболевания.
Полученную мышь использовали в моделях острого аллергического воспаления дыхательных путей (которые описаны в примерах 18 и 19) и моделях хронического аллергического воспаления дыхательных путей (которые описаны в примере 20), что обеспечило возможность оценки фармакологического лечения антителом против IL-13 человека при аллергическом заболевании дыхательных путей.
ПРИМЕР 18
Эффективность антител против IL-13 человека в мышиной модели Ллойда острого воспаления легких на основе трансгенных по huIL-13 мышей
Мышиная модель острого аллергического воспаления легких
Влияние нейтрализующих IL-13 человека антител в виде IgG4 человека исследовали в мышиной модели острого аллергического воспаления легких, используя трансгенных мышей, созданных в примере 17. Данную модель осуществляли, по существу, как описано McMillan et al. [85] и как описано в примере 13. Конечный результат модели характеризовали по повышенному содержанию IL-13 в BAL и легочной ткани, клеточной инфильтрации в легкие и BAL, повышенным уровням содержания IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Протокол данной модели был таким же, как описано в примере 13, за исключением того, что вводили дозы антитела против IL-13 человека вместо BAK209B11.
ПРИМЕР 19
Эффективность антител против IL-13 человека в мышиной модели Жерара острого воспаления легких на основе трансгенных по huIL-13 мышей
Мышиная модель острого аллергического воспаления легких
Влияние нейтрализующих IL-13 человека антител в виде IgG4 человека исследовали в другой мышиной модели острого аллергического воспаления легких, используя трансгенных мышей, созданных в примере 17. Данную модель осуществляли, по существу, как описано Humbles et al. [86] и как описано в примере 14. Конечный результат модели характеризовали по повышенному содержанию IL-13 в BAL и легочной ткани, клеточной инфильтрации в легкие и BAL, повышенным уровням содержания IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Протокол данной модели был таким же, как описано в примере 14, за исключением того, что вводили дозы антитела против IL-13 человека вместо BAK209B11.
ПРИМЕР 20
Эффективность антител против IL-13 человека в мышиной модели Ллойда хронического воспаления легких на основе трансгенных по huIL-13 мышей
Влияние нейтрализующих IL-13 человека антител в виде IgG4 человека исследовали в модели хронического аллергического воспаления легких, используя трансгенных мышей, созданных в примере 17. Данную модель осуществляли, по существу, как описано Temelkovski et al. [87] и как описано в примере 15, и конечный результат характеризовали по клеточной инфильтрации в легкие и BAL, повышенным уровням содержания IgE в сыворотке и гиперчувствительности дыхательных путей (AHR).
Протокол модели
Протокол данной модели был таким же, как описано в примере 15, за исключением того, что вводили дозы антитела против IL-13 человека вместо BAK209B11.
ПРИМЕР 21
Фармакокинетика и фармакодинамика антител против IL-13 человека у макак-крабоедов с аллергией на Ascaris suum
Фармакокинетику и фармакодинамику 502G9 оценивали у 4 аллергичных, но не подвергнутых провокации приматов макак-крабоедов (2 самца/2 самки) после однократной болюсной дозы 10 мг/кг в/в. Эксперимент продолжался в течение 29 дней. Фармакокинетические параметры антитела определены на основе кривой среднегеометрической концентрации лекарственного средства в сыворотке и подробно описаны ниже в таблице 4.
В этом же исследовании также следили за концентрациями IgE в сыворотке, используя набор для анализа IgE человека в ELISA (Bethyl laboratories, USA).
Результаты
Концентрации IgE в сыворотке значимо снижались после однократной болюсной дозы 10 мг/кг в/в BAK502G9, от уровней 100% в контроле (до введения дозы) до 66±10% от значений в контроле (p<0,05) на 4 и 5 день после введения дозы. Указанное снижение концентрации IgE в сыворотке восстанавливалось до 88±8% от уровней в контроле на 22 день (смотри фиг.20). Указанные данные также получали нормализацией концентрации IgE в сыворотке каждого животного по отношению к уровням до введения дозы, при этом концентрации до введения дозы составляли 100%, и затем усреднением кривых для 4 тестируемых животных.
Два самца обезьян имели относительно низкие суммарные уровни IgE в сыворотке до введения дозы (60 нг/мл и 67 нг/мл). Указанные уровни IgE не изменялись таким образом, который свидетельствует о какой-либо тенденции после обработки BAK502G9 (фиг.20B). Две самки обезьян имели относительно высокие уровни суммарного IgE в сыворотке (1209 нг/мл и 449 нг/мл). Указанные уровни IgE снижались после обработки BAK502G9 максимально на 60% на 7 день и возвращались примерно до уровней перед введением дозы на 28 день после введения (фиг.20B). Полученные данные показывают, что BAK502G9 снижает концентрации IgE в сыворотке у животных с относительно высокими уровнями циркулирующего IgE.
ПРИМЕР 22
Эффективность антител против IL-13 человека в моделях кожной аллергии у макак-крабоедов
Влияние нейтрализующих IL-13 человека антител в виде IgG4 человека исследовали в модели острого аллергического воспаления кожи приматов. Указанную модель осуществляли посредством инъекции IL-13 человека и антигена A. suum интрадермально макакам-крабоедам. Через 24-96 час брали биопсии кожи и образцы сыворотки. Конечный результат в модели характеризовали по инфильтрации клеток в кожу.
ПРИМЕР 23
Эффективность антител против IL-13 человека в модели легочной аллергии у макак-крабоедов
Эффективность нейтрализующих IL-13 человека антител в виде IgG4 человека исследовали в модели острого аллергического воспаления легких у приматов. Указанную модель осуществляли, воздействуя распыляемым антигеном A. suum на макак-крабоедов с аллергией на A. suum, таким образом вызывая аллергическую реакцию. Указанную аллергию характеризовали в конченом итоге по инфильтрации клеток в легкие и BAL, повышенным уровням IgE в сыворотке и гиперчувствительности дыхательных путей.
Дополнительно оценивали фармакодинамику ex vivo, используя способ проточной цитометрии. CD23 является высокоаффинным рецептором IgE и может быть экспрессирован на мононуклеарных клетках периферической крови человека. Экспрессия CD23 может быть индуцирована IL-13 и IL-4 в отношении количества клеток, экспрессирующих CD23, а также в отношении того, сколько CD23 экспрессирует каждая клетка. Ответ, опосредованный IL-13, но не IL-4, может быть ингибирован антителом против IL-13 человека.
Животных предварительно отбирали для введения во 2-фазу исследования на основе предварительно вызванной AHR после провокации распыляемым антигеном (экстракт Ascaris suum). В фазе I функцию дыхательных путей оценивали во время внутривенной провокации гистамином в 1 и 11 день. PC30, дозу гистамина, необходимую для того, чтобы вызвать 30% увеличение резистентности легких (RL) выше исходного уровня, определяли на основе каждой кривой зависимости ответа от дозы гистамина. На 9 и 10 день животных подвергали провокации индивидуально рассчитанными дозами распыляемого антигена, которые, как показывали предварительно, вызывают 40% увеличение RL, а также 35% снижение динамической пластичности (CDYN). Исторически в данной модели более высокую RL наблюдали после второй провокации заданной дозой аллергена, чем после первой; это означает примирование антигеном. Две провокации антигеном вызывали AHR, которую измеряли по увеличенной площади под кривой зависимости ответа от дозы гистамина и/или падению PC30 и BAL, а также эозинофилии на 11 день по сравнению с 1 днем. Животных, проявляющих AHR-фенотип, отбирали для введения в фазу II.
Фазу II поводили точно так же, как фазу I, за исключением того, что все животные получали инфузию 30 мг/кг BAK502G9 в 1, 5 и 9 день. Влияние BAK502G9 оценивали посредством сравнения изменений, наблюдаемых в фазе II, с изменениями, наблюдаемыми в фазе I, для отдельных животных.
Собирали образцы крови, сыворотки, BAL и ткани легкого. Уровни IgE в сыворотке проверяли с помощью ELISA. Показано, что сыворотка обработанных BAK502G9 макак-крабоедов ингибирует экспрессию CD23 на мононуклеарных клетках периферической крови человека, индуцированную IL-13, но не IL-4. Величина указанного ингибирования соответствовала уровням BAK502G9 в сыворотке, рассчитанным с помощью ELISA PK.
Результаты
BAK502G9 значимо ингибировало AHR, которую измеряли по AUC RL (p<0,05) (фиг.26A; таблица 7). Наблюдали ингибирующее влияние BAK502G9 на AHR, измеренное по PC30, но оно не достигало статистической значимости (фиг.26B; таблица 7). BAK502G9 также значимо ингибировало примирование обоими антигенами (p<0,01) (фиг.26C; таблица 7) и воспаление BAL. BAK502G9 значимо ингибировало приток суммарных клеток (p<0,05), и эозинофилов (p<0,05) но не макрофагов, лимфоцитов или тучных клеток в BAL (фиг.26D; таблица 7).
ПРИМЕР 24
Эффективность антител против IL-13 человека в отношении астматического фенотипа, который развивается при введении IL-13 человека в легкие мыши
Мышиная модель гиперчувствительности дыхательных путей
Исследовали эффективность нейтрализующего антитела BAK502G9 против IL-13 человека, направленную против развития гиперчувствительности дыхательных путей (AHR), после введения IL-13 человека в легкие мышей. Указанную модель осуществляли, по существу, как описано Yang et al. [119], за исключением того, что использовали IL-13 человека вместо мышиного IL-13.
Протокол модели
Для развития фенотипа на мышей BALB/c действовали двумя дозами IL-13 человека, разделенными 48-часовым интервалом. Коротко, мышей анестезировали внутривенной инъекцией 100 мкл раствора саффана (разведение 1:4 в воде). Мышам проводили интубацию катетерной иглой калибра 22, через которую вливали рекомбинантный IL-13 человека (25 мкг, растворенных в 20 мкл фосфатно-солевого буфера (PBS)) или наполнитель в качестве контроля (PBS). Функцию дыхательных путей оценивали через 24 часа после последнего введения IL-13 при возрастающих провокациях метахолином и регистрировали, используя плетизмографию животных, находящихся в сознании (Buxco). PC200 (концентрацию метахолина, требуемую для увеличении исходного уровня PenH на 200%) определяли на основании подгонки кривых по 4 нефиксированным параметрам для кривых зависимости ответа от дозы метахолина. Обработки антителом проводили посредством внутрибрюшинной инъекции за 24 часа до введения каждой дозы IL-13.
Результаты
Интратрахеальное вливание IL-13 человека нативным мышам дикого типа приводило к развитию значимой (p<0,05) гиперчувствительности дыхательных путей по сравнению с контрольными животными, что определяли на основе концентраций метахолина PC200. Системно введенное BAK502G9 (1 мг/кг) значимо ингибировало (p<0,01) развитие AHR, тогда как антитело нулевого контроля не оказывало влияния (фиг.23).
ПРИМЕР 25
Нейтрализующая эффективность BAK502G9 в виде IgG4 человека против зависимого IL-13 человека высвобождения IgE из B-клеток человека
Протокол анализа переключения B-клеток
Показано, что IL-13 индуцирует синтез IgE в B-клетках человека in vitro [120]. Зависимое от фактора высвобождение IgE из B-клеток человека определяли в ELISA. Эффективность нейтрализации BAK502G9 в виде IgG4 человека оценивали по отношению к зависимому от IL-13 человека высвобождению IgE из B-клеток человека.
Мононуклеарные клетки периферической крови (PBMC) очищали из лейкоцитной пленки человека (Blood Transfusion Service) центрифугированием в градиенте плотности 1,077 г/л. B-клетки очищали из PBMC с использованием набора для выделения B-клеток II (Miltenyi Biotec), используя реагенты и способы, описанные производителем. Среда для анализа содержала среду Дульбекко, модифицированную по способу Исков (Life Technologies), содержащую 10% фетальной сыворотки теленка и 20 мкг/мл трансферрина человека (Serologicals Proteins Inc). После очистки B-клетки ресуспендировали в конечной концентрации 106/мл в среде для анализа. 50 мкл ресуспендированных клеток добавляли в каждую анализируемую точку в 96-луночный планшет для анализа. В соответствующие анализируемые лунки добавляли 50 мкл раствора 4 мкг/мл анти-CD40-антитела EA5 (Biosource). Тестируемые растворы антител (шесть повторов) разбавляли до требуемой концентрации в среде для анализа. В качестве негативного контроля использовали нерелевантное антитело, не направленное к IL-13. К клеткам добавляли 50 мкл/лунку соответствующего тестируемого антитела. Затем добавляли рекомбинантный, полученный в бактериях IL-13 человека (Peprotech) до конечной концентрации 30 нг/мл, получая общий объем для анализа 200 мкл/лунку. Концентрацию IL-13, используемую для анализа, выбирали так, чтобы она давала максимальный ответ. Планшеты для анализа инкубировали в течение 14 дней при 37°C в 5% CO2. Уровни IgE в надосадках определяли в ELISA, используя реагенты и протоколы, прилагаемые производителем (BD Biosciences, Bethyl Laboratories). Данные анализировали с использованием компьютерной программы Graphpad prism.
Результаты
Как показано на фиг.24, BAK502G9 (VH SEQ ID NO: 15; VL SEQ ID NO: 16) способно ингибировать зависимую от IL-13 человека продукцию IgE B-клетками человека. BAK502G9 в виде IgG4 человека имело IC50 1,8 нМ против 30 нг/мл IL-13 человека.
ПРИМЕР 26
Эффективность BAK502G9 против опосредованного IL-13 усиления индуцированной гистамином передачи сигнала Ca2+ в первичных клетках гладкой мускулатуры бронхов человека
Показано, что IL-13 непосредственно модулирует сократимость гладкой мускулатуры дыхательных путей [121, 122]. Внутриклеточная мобилизация кальция является необходимым условием сокращения гладкой мускулатуры. Недавние исследования показали, что способность IL-13 изменять сократимость гладкой мускулатуры отчасти опосредована модулированием индуцированной сократительным агонистом передачи сигнала Ca2+ [123, 124].
Эффективность BAK502G9, антитела против IL-13 человека, в форме IgG4, направленная против опосредованных IL-13 изменений сигнальных ответов гладкомышечных клеток бронхов человека (BSMC) на сократительный агонист гистамин, исследовали в анализе передачи сигнала Ca2+.
Протокол анализа передачи сигнала Ca2+ в BSMC
Первичные BSMC человека, среду роста 2 для гладкомышечных клеток (SmGM-2) и основную среду для гладкомышечных клеток (SmBM) получали из Bio Whittaker. BSMC поддерживали в SmGM-2 согласно рекомендациям поставщика. BSMC высевали при плотности 2×104 клеток/лунку в 96-луночный планшет для микротитрования для культуры ткани и давали возможность прикрепиться в течение 24 часов, затем снова подкармливали и инкубировали еще в течение 24 часов. Перед экспериментом, связанным с передачей сигнала Ca2+, BSMC стимулировали IL-13 (Peprotech) в конечной концентрации 50 нг/мл в присутствии или в отсутствие антитела и инкубировали в течение 18-24 часов. BAK502G9 и совпадающее по изотипу нерелевантное контрольное моноклональное антитело, CAT-001, оценивали в конечной концентрации 10 мкг/мл. Изменения внутриклеточных концентраций Ca2+ в ответ на гистамин (Calbiochem), титруемый, начиная с 20 мкМ, измеряли, используя стандартный способ с чувствительным к Ca2+ красителем Fluo-4 (Molecular Probes) и используя считывающее устройство для визуализации флуоресценции на 96-луночном планшете (FLIPR) (Molecular Devices). Площадь под кривой (AUC) ответа в виде передачи сигнала Ca2+ на гистамин определяли для каждого условия предварительной обработки клеток. Анализ данных осуществляли с использованием GraphPad Prism, версия 4 для Windows (компьютерная программа GraphPad).
Результаты
Предварительная инкубация BSMC с IL-13 значимо усиливала передачу сигнала Ca2+ в ответ на гистамин. Предварительная инкубация BAK502G9 (фиг.25B) (но не нерелевантного контрольного антитела определенного изотипа (фиг.25A)) с IL-13 значимо ингибировала усиление передачи сигнала Ca2+ в ответ на гистамин (фиг.25).
ПРИМЕР 27
Нейтрализующая эффективность анти-IL-13-антител в анализе зависимой от IL-13 человека экспрессии CD23 PBMC
Эффективность типичного IL-13-антитела оценивали в анализе зависимой от IL-13 человека экспрессии CD23 в мононуклеарных клетках периферической крови (PBMC). PBMC отвечали как на IL-13, так и на IL-4 увеличением экспрессии CD23 на клеточной поверхности [120]. CD23 (FceRII) является низкоаффинным рецептором IgE и экспрессируется на различных воспалительных клетках, включая моноциты. Ингибирование зависимой от IL-13 человека повышающей регуляции экспрессии CD23 определяли посредством измерения уменьшения связывания флуоресцентно меченного моноклонального антитела к CD23 с PBMC с помощью проточной цитометрии.
Протокол анализа
Кровь человека получали из Blood Transfusion Service и избавлялись от эритроцитов с помощью 40-минутного осаждения в декстране-T500 (Pharmacia) (конечная концентрация 0,6%). Затем отделяли фракцию, обогащенную лейкоцитами и тромбоцитами, с помощью 20-минутного центрифугирования при 1137g в ступенчатом градиенте Percoll 3 мл 64% и 5 мл 80% (100% состоит из 9 частей Percoll и 1 части 10×PBS). PBMC собирали сверху 64% слоя, промывали и ресуспендировали в буфере для анализа (Invitrogen RPMI 1640, 10% FCS, 200 IU/мл пенициллина, 100 мкг/мл стрептомицина, 2 мМ L-глутамин). Анализ осуществляли в 24-луночных планшетах с использованием 2×106 клеток, ±80 пМ рекомбинантного IL-13 человека (Peprotech) или 21 пМ рекомбинантного IL-4 человека (R and D Systems), BAK502G9 или нерелевантного IgG4 в конечном объеме 500 мкл. Клетки культивировали в течение 48 час при 37°C, затем собирали и красили CD23-PE (BD Pharmingen) в течение 20 минут при 4°C. Наконец клетки регистрировали в проточном цитометре. Экспрессию CD23 определяли с помощью «оценки» CD23; по проценту CD23-позитивных клеток, умноженному на яркость окраски (средняя геометрическая флуоресценции). «Оценку» CD23 без стимулятора не вычитали, и данные представлены в виде процента от ответа на IL-13, добавленный отдельно (100%). Данные выражали в виде средней ± SEM, полученной на основе 4-6 отдельных экспериментов с использованием клеток от 4-6 отдельных доноров, проводимых в трех повторах для каждой точки.
Результаты
Инкубация PBMC с 80 пМ IL-13 или 21 пМ IL-4 в течение 48 часов приводила к выраженной экспрессии CD23 (фиг.27 и фиг.28). BAK502G9 зависимым от дозы образом ингибировала индуцированную IL-13 экспрессию CD23 с геометрической средней 120,2 пМ (фиг.27). Напротив, BAK502G9 не обладала способностью ингибировать экспрессию CD23, индуцированную 21,4 пМ IL-4 (n=4 отдельных донора, фиг.28). Нерелевантный IgG4 не ингибировал ни IL-13-, ни IL-4-зависимую экспрессию CD23 на PBMC (фиг.27 и фиг.28). Совместная стимуляция PBMC 80 пМ IL-13 и 21,4 пМ IL-4 давала аддитивный CD23-ответ. BAK502G9, но не CAT-001, снижало уровни экспрессии CD23 до уровней, наблюдаемых при стимуляции отдельно IL-4 (фиг.28).
ПРИМЕР 28
Нейтрализующая эффективность антитела против IL-13 человека в анализе зависимого от IL-13 человека изменения формы эозинофилов
Целью данного исследования была оценка влияния IL-13-антитела на изменение формы эозинофилов, индуцированной медиаторами, высвобождаемыми из NHLF после стимуляции факторами, обнаруженными в легких больных астмой, такими как IL-13 [125, 126], TNF-α [127], TGF-β1 [128]. IL-13 действует синергически с TNF-α [129] или TGF-β1 [130], индуцируя фибробласты к продуцированию эотаксина-1, который затем может действовать непосредственно как хемоаттрактант эозинофилов. Изменение формы лейкоцитов опосредуется перестройкой цитоскелета клеток и имеет важное значение для процесса миграции лейкоцитов из микроциркуляции в места воспаления. Ингибирование зависимого от IL-13 высвобождения фактора, индуцирующего изменение формы, из NHLF определяли посредством измерения снижения секреции эотаксина-1 в ELISA и снижения изменения формы эозинофилов с помощью проточной цитометрии.
Протокол анализа
Клетки NHLF культивировали в среде, не содержащей добавок, или в среде, содержащей стимуляторы (9,6 нМ IL-13, 285,7 пМ TNF-α (R and D Systems) и 160 пМ TGF-β1 (R and D Systems) в отсутствие или в присутствии BAK502G9 (диапазон концентраций 875 нМ - 6,84 нМ). Затем клетки культивировали еще в течение 48 час при 37°C, затем полученную в результате кондиционированную среду отсасывали и хранили при -80°C. Концентрацию эотаксина-1 в кондиционированной среде оценивали, используя системы ELISA R and D systems Duoset (R and D Systems).
Кровь человека получали из Blood Transfusion Service и избавлялись от эритроцитов с помощью 40-минутного осаждения в декстране-T500 (Pharmacia) (конечная концентрация 0,6%). Затем отделяли фракцию, обогащенную лейкоцитами и тромбоцитами, с помощью 20-минутного центрифугирования при 1137g в ступенчатом градиенте Percoll 3 мл 64% и 5 мл 80% (100% состоит из 9 частей Percoll и 1 части 10×PBS). Гранулоциты собирали на границе раздела 64%:80%, промывали и ресуспендировали в буфере для анализа (Sigma PBS, 1 мМ CaCl2, 1 мМ MgCl2, 10 мМ HEPES, 10 мМ D-глюкоза, 0,1% БСА Sigma, pH 7,3). Анализ осуществляли в пробирках для FACS, содержащих 5×105 клеток, ±3 нМ рекомбинантного эотаксина-1 человека (R and D Systems) или кондиционированную среду в конечном объеме 400 мкл. Клетки инкубировали в течение 8,5 минут при 37°C, затем переносили на 4°C и фиксировали фиксатором (CellFix, BD Biosciences) и наконец регистрировали в проточном цитометре. Эозинофилы идентифицировали по их FL-2-аутофлуоресценции и регистрировали параметр прямого рассеяния (FSC). Параметр FSC эозинофилов изменялся в ответ как на эотаксин-1, так и на кондиционированную среду, обеспечивая возможность измерения изменения формы. Содержимое пробирок анализировали при высокой скорости потока и сбор данных прекращали после 1000 событий встречаемости эозинофилов или через 1 минуту, независимо от того, что было быстрее. Изменение формы рассчитывали в виде процента по отношению к FSC, вызванному только одним буфером для изменения формы (100% изменение формы в контроле). Данные выражали в виде среднего % изменения формы в контроле ± SEM, полученного на основе 4 отдельных экспериментов. В каждом эксперименте использовали клетки из отдельных лейкоцитарных пленок (и следовательно, от отдельного донора) и эксперимент выполняли в двух повторах для каждой точки.
Результаты
Клетки NHLF, костимулированные 9,6 нМ IL-13, 285,7 пМ TNF-α и 160 пМ TGF-β1 и культивированные в течение 48 час, секретировали 9,6 нМ эотаксина-1 в культуральную среду. Напротив, клетки NHLF, культивированные только поддерживающей рост среде, секретировали 0,1 нМ эотаксина-1 в культуральную среду. Указанное продуцирование эотаксина-1 зависело от IL-13, так как BAK502G9 зависимым от дозы образом ингибировала продуцирование эотаксина-1 клетками NHLF, костимулированными IL-13/TNF-α/TGF-β1, с IC50 32,4 нМ (фиг.29A).
Основной целью данной части исследования было определение изменения формы эозинофилов. Величина изменения формы эозинофилов в ответ на 3 нМ эотаксина (позитивный контроль) составляла 122,2±2,1% (n=4). Индуцированное эотаксином-1 изменение формы полностью ингибировалось 100 нМ антителом против эотаксина CAT-213, среднее изменение формы 101,0±1,0% (n=4).
Среда от клеток NHLF, костимулированных 9,6 нМ IL-13, 285,7 пМ TNF-α и 160 пМ TGF-β1 и культивированных в течение 48 час (кондиционированная среда), индуцировала явное изменение формы эозинофилов (фиг.29B). Напротив, среда от клеток NHLF, культивированных в течение 48 час только в поддерживающей рост клеток NHLF среде, не индуцировала изменения формы эозинофилов (фиг.29B).
Добавление анти-IL-13-антитела BAK502G9 к среде для костимулирования перед культивированием NHLF приводило к зависимому от дозы ингибированию изменения формы эозинофилов с геометрической средней IC50 16,8 нМ при анализе в разведении 1:16 (фиг.29B).
Также исследовали способность стимуляторов (IL-13, TNF-α и TGF-β1), не вводимых в культуру клеток NHLF, индуцировать изменение формы эозинофилов и нейтрофилов. 9,6 нМ IL-13, 285,7 пМ TNF-α и 160 пМ TGF-β1 не индуцировали видимое изменение формы эозинофилов. Это свидетельствует о том, что способность кондиционированной среды изменять форму эозинофилов, которое развивается во время культивирования клеток NHLF со стимуляторами, возникает не из-за какого-либо стимулятора отдельно или в комбинации (фиг.29B).
ПРИМЕР 29
Картирование анти-IL-13-антител на IL-13 человека
Эпитопное картирование типичного IL-13-антитела BAK502G9 осуществляли, используя молекулярный способ и стандартное вырезание пептидов.
Молекулярный способ
Конструировали химеры IL-13, в которых части последовательности IL-13 человека заменяли мышиной последовательностью. Полученные химеры использовали в исследованиях связывания с типичными IL-13-антителами, чтобы идентифицировать специфический эпитоп.
Получали две панели химер IL-13. Первая панель содержала девять химер (фиг.30) и была использована для того, чтобы локализовать общее положение эпитопа. Вторая панель содержала десять химер (фиг.31) и была использована для тонкого картирования эпитопа.
Химерные последовательности IL-13 собирали, используя ПЦР, и клонировали в векторе для введения Gateways®, который затем рекомбинировали с целевым вектором pDEST8 (модифицированным так, чтобы он кодировал регистрируемую и аффинную метку на C-конце рекомбинантного белка). Указанные экспрессирующие векторы использовали для трансформации химически компетентных E.coli DH10Bac™, обеспечивающих возможность сайт-специфичной транспозиции меченого химерного IL-13 в бакуловирусный челночный вектор (бакмиду). Рекомбинантную бакмидную ДНК выделяли для каждой химеры IL-13 и трансфицировали в клетки насекомых Sf9 (Spodoptera frugiperda), используя реагент Cellfectin®. Рекомбинантные бакуловирусы собирали через 72 часа после трансфекции и пассировали на клетках насекомых Sf9 еще два раза.
Надосадок культуры объемом 2000-500 мл очищали на аффинной колонке и элюированный материал концентрировали от 16 до 1 мл и наносили на колонку для эксклюзионной хроматографии по размеру Superdex 200 HR10/300GL для окончательной обработки и замены буфера.
Разработали однородный конкурентный анализ, используя биотинилированный IL-13 человека, стрептавидин-антофиоцинат и меченное европием BAK502G9. Анализ осуществляли следующим образом: Eu-BAK502G9 связывает биотинилированный IL-13 человека, затем комплекс распознается конъюгатом стрептавидин-APC, и в случае применения вспышки света происходит перенос энергии от APC-метки к европию благодаря близости, и может быть измерена флуоресценция с временным разрешением. Конкуренцию указанному связыванию создают посредством немеченого IL-13 человека (в качестве контроля) и химерных конструкций. Указанную конкуренцию оценивают количественно, чтобы рассчитать относительные аффинности мутантов IL-13 по отношению к IL-13-антителам, что делает возможным идентификацию мутаций, изменяющих связывание.
Результаты
Обнаружено, что химерная конструкция IL13-спираль D (таблица 5) является наиболее слабым конкурентом для биотинилированного IL-13 человека за связывание BAK502G9, свидетельствуя, что спираль D в молекуле IL-13 вовлечена в эпитопное связывание BAK502G9 (таблица 5). Пониженная активность также наблюдалась в случае мутантов 4041 и 3334, в которых остатки 40, 41 и 33, 34 исходной последовательности соответственно были изменены, что свидетельствует о возможном участии спирали A в узнавании BAK502G9. Пониженные активности петли 3 не принимали в расчет, так как указанная петля имеет меньшее количество аминокислот в мутанте по сравнению с молекулой человека, и, вероятно, может быть изменена общая структура белка. Другие снижения способности химерных молекул IL-13 конкурировать за связывание BAK502G9 не считали существенными в случае таких аминокислотных изменений.
Затем тестировали набор более целенаправленных мутаций в спирали D (фиг.26). Полученные результаты показаны в таблице 6 и представляют собой следующее.
Результаты показывают, что химерные конструкции 116117TK (в которой лизин в положении 116 заменен треонином и аспартат в положении 117 заменен лизином), 123KA (в которой заменен лизин в положении 123) и 127RA (в которой заменен аргинин в положении 127) обладают наименьшей способностью конкурировать за связывание с BAK502G9 (123KA и 127RA не конкурируют при 1 мкМ). Другие остатки, вовлеченные в связывание с BAK502G9 на основе пониженных эффективностей в конкурентном анализе, включают остатки спирали D 124Q (где лизин заменен глутамином) и 120121SY (пара лейцин-гистидин заменена на пару серин-тирозин). Мутация лейцина в положении 58L также уменьшает связывание, и анализ 3D-структур показал, что указанный остаток упакован вплотную к спирали D и может либо непосредственно контактировать с BAK502G9, или может влиять на выравнивание спирали D.
Проведенные эксперименты показывают, что остатки в спирали D важны для связывания BAK502G9 с IL-13. В частности, лизин в положении 123 и аргинин в положении 127 важны для указанного связывания, так как мутация в любом из них прекращает связывание BAK502G9.
Вырезание эпитопа
Эпитопное картирование BAK502G9 также осуществляли, используя стандартный способ вырезания пептида. В данном случае IgG иммобилизовали на твердой фазе и давали возможность улавливать лиганд IL-13. Затем образованный комплекс подвергали специфичному протеолитическому расщеплению, во время которого расщеплялись доступные пептидные связи, однако связи, защищенные областью контакта IgG:лиганд, оставались интактными. Таким образом, пептид, содержащий эпитоп, оставался связанным с IgG. Затем его можно было десорбировать, собрать и идентифицировать масс-спектрометрией (МС).
Использовали два дополнительных способа, в первом использовали масс-спектрометр MALDI-TOF для считывания белковых чипов Ciphergen, при этом можно было ковалентно связывать IgG с чипом для масс-спектрометра и затем осуществлять расщепление и экстракцию in situ. Во втором способе использовали биотинилированный BAK502G9, связанный с покрытыми стрептавидином шариками, и обеспечивали возможность сбора достаточного количества пептида для подтверждения последовательности тандемной масс-спектрометрией (МС/МС).
Два способа хотя и отличались по реальным деталям и масштабу, по существу включали одни и те же стадии, связывание IgG, блокирование непрореагировавших участков связывания, промывку, улавливание лиганда, удаление несвязанного лиганда, расщепление и конечную стадию промывки.
В способе MALDI-TOF-МС использовали запатентованные МС-чипы, активированные карбонилдиимидазолом, который ковалентно связывается со свободными первичными аминогруппами, с которыми связывали IgG в концентрации 1-2 мг/мл в PBS в течение ночи при 4°C. Затем чип блокировали раствором этаноламина при комнатной температуре в течение 1 часа и затем тщательно промывали PBS или HBS плюс подходящий детергент. Затем на чип наносили аликвоту, содержащую один пикомоль IL-13, либо в PBS, либо в HBS, и давали возможность связаться с химически иммобилизованным IgG в течение 2 часов при комнатной температуре. После этого проводили дополнительные промывки в PBS или HBS с детергентом или без него, чтобы удалить любой неспецифично связанный IL-13. Затем добавляли раствор трипсина в диапазоне от 200 до 3,1 мкг/мл в PBS или HBS к комплексу IgG:лиганд и обеспечивали возможность расщепления в течение 30 минут при комнатной температуре, после чего чип промывали в PBS или HBS плюс детергент, PBS или HBS и наконец водой. Затем после применения подходящей матрицы для MALDI-TOF МС чип помещали непосредственно в масс-спектрометр и анализировали.
Основанный на шариках способ начинали с биотинилирования IgG, используя соединение NHS-биотин в молярном соотношении 1 IgG на 4 молекулы биотина. После этого следовало удаление несвязанного биотина и побочных продуктов реакции с использованием гель-фильтрации. Затем давали возможность биотинилированному IgG связаться с покрытыми нейтравидином агарозными шариками, при этом пытались максимизировать захват IgG. Затем аликвоты покрытых IgG шариков распределяли на концентрирующие центрифужные колонки и промывали PBS Дульбекко + 0,05% твин 20, затем ресуспендировали в PBS Дульбекко + 0,05% твин 20. Затем к ресуспендированным IgG-шарикам импульсно добавляли IL-13 и обеспечивали возможность для протекания связывания в течение 10 минут, после этого жидкую фазу удаляли центрифугированием и шарики промывали PBS Дульбекко + 0,05% твин 20 с последующим ресуспендированием в PBS Дульбекко + 0,05% твин 20.
Затем комплекс шарик:IgG:лиганд подвергали протеолизу либо трипсином, либо химотрипсином, инкубируя при комнатной температуре или при 37°C. После этого шарики снова промывали в PBS Дульбекко + 0,05% твин 20 с последующими дополнительными промывками в PBS Дульбекко с детергентом. Затем шарики ресуспендировали в смеси воды, ацетонитрила, трифторуксусной кислоты и извлекали надосадок. Затем надосадок анализировали различными способами, либо посредством MALDI-TOF МС, либо ВЭЖХ с обращенной фазой и масс-спектрометрией, включая тандемную (МС/МС) фрагментацию с использованием масс-спектрометра с ионной ловушкой ThermoQuest LCQ ESI. Затем предпринимали попытку найти соответствие полученной в результате картины фрагментации с последовательностью IL-13 человека и отдельной последовательностью тяжелой и легкой цепи IgG BAK502G9.
Во время серии экспериментов использовали ряд контролей первоначально пустых поверхностей, IgG отдельно и контролей изотипа, чтобы показать, что идентифицированные пептиды получали специфично из IgG-захваченного IL-13, и они не являются продуктом расщепления BAK502G9 или неспецифично связанного IL-13.
Результаты
В серии экспериментов одинаковым образом получали одиночные специфичные пептиды IL-13 для каждого расщепления. Данные, полученные с помощью прибора с ионной ловушкой LCQ, показали, что полученный при обработке трипсином фрагмент имел моноизотопную массу 3258 Д (MH+), а фрагмент, полученный обработкой химотрипсином - моноизотопную массу 3937 Д (MH+).
Поиск указанных масс по сравнению с подходящим расщеплением in silico IL-13 человека давал близкие совпадения с родственными пептидами в C-концевой части молекулы.
Совпадение массы пептида, полученного обработкой трипсином:3258 Д
При допуске 1000 м.д. 3258 Д совпадает с последовательностью от аспарагиновой кислоты в положении 106 до C-концевого аспарагина в положении 132. При таком допуске других совпадений нет. Указанная область выделена жирным шрифтом в последовательности формы предшественника IL-13 человека ниже.

Совпадение массы пептида, полученного обработкой химотрипсином:3937 Д
При допуске 1000 м.д. 3937 Д совпадает с последовательностью от серина в положении 99 до C-концевого аспарагина в положении 132. Указанная область выделена жирным шрифтом в последовательности формы предшественника IL-13 человека ниже.

Оба приведенных совпадения показывают, что IgG BAK502G9 сохраняет C-концевую часть молекулы IL-13 во время протеолиза комплекса антитело:лиганд.
Идентичность обоих пептидов успешно подтвердили в МС/МС, ни для одного из них не наблюдали какой-либо существенной параллели последовательностей с BAK502G9. На карте фрагмента МС/МС, разработанной для того, чтобы идентифицировать либо Y-, либо B-ионы, совпадало 26 из 104 возможных ионов в одном заряженном участке в случае пептида, полученного обработкой трипсином, и 19 из 128 возможных ионов в случае пептида, полученного обработкой химотрипсином. Обзор всех заряженных участков демонстрирует идентификацию 23 из 27 аминокислотных остатков в случае трипсинового фрагмента и 29 из 33 остатков в случае химотрипсинового фрагмента. Этого достаточно для подтверждения идентичности.
В серии экспериментов в целом идентифицировали такую часть эпитопа BAK502G9 в IL-13 человека, которая расположена в пределах двадцати семи C-концевых аминокислотных остатков. Полученные результаты подтверждают результаты молекулярного способа, подробно описанного выше.








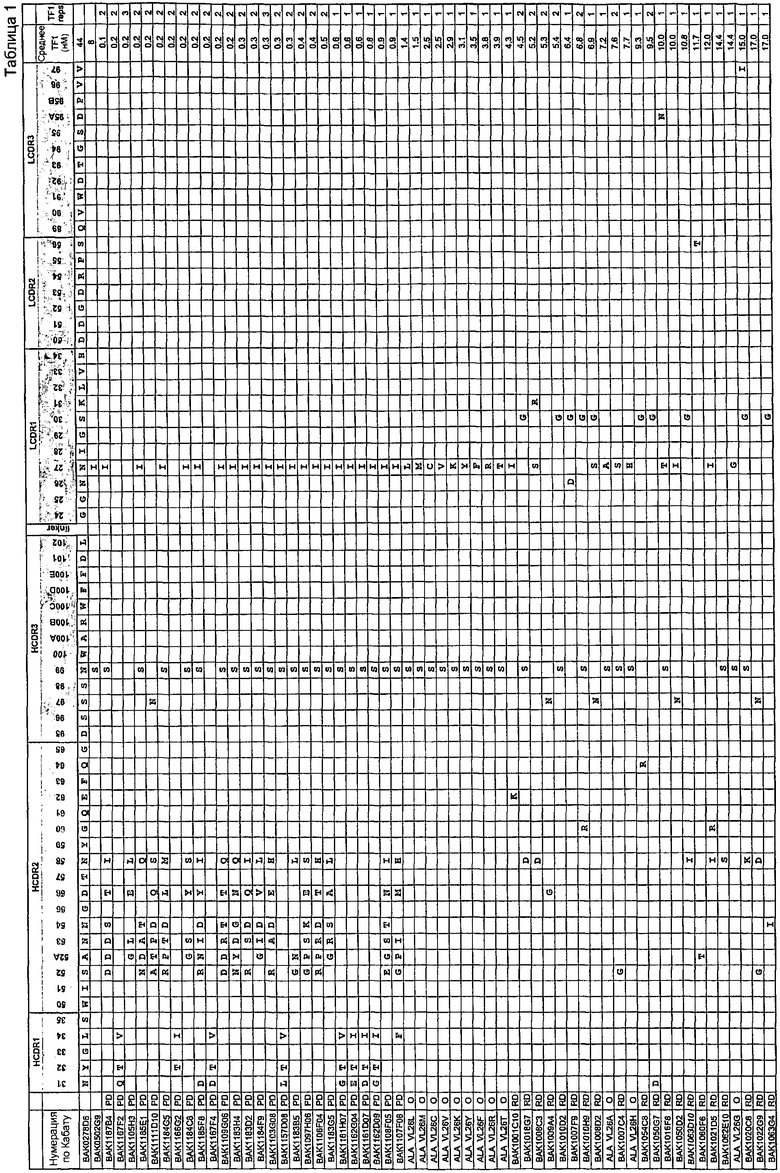
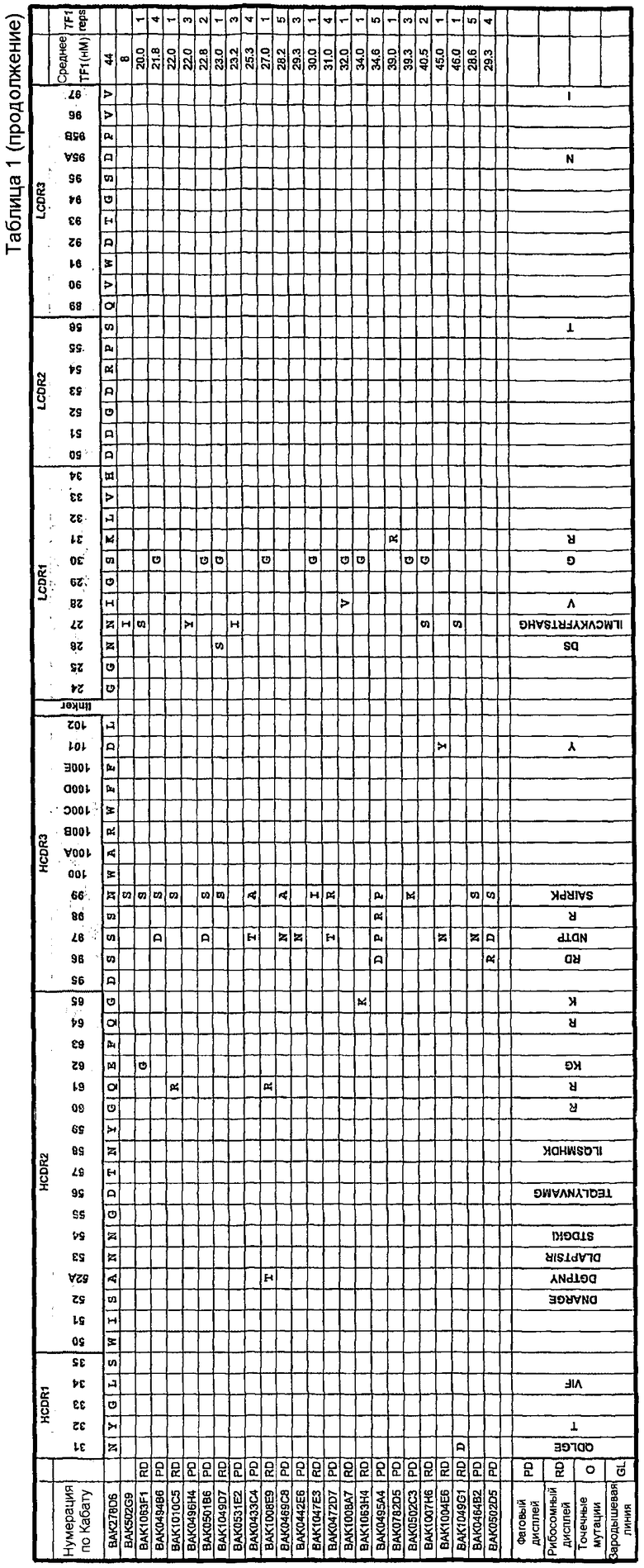
Cl
inf = клиренс в течение времени 0-бесконечность, рассчитанный на основе экстраполированной AUC.
AUC
inf = площадь под кривой (измерение суммарной экспозиции с лекарственным средством) в течение времени 0-бесконечность, включая экстраполированный период, основанный на константе скорости элиминации (k) и последней определяемой концентрации лекарственного средства в сыворотке.
AUC
ext = процент AUC от суммарной AUC, которая экстраполирована.
T
0,5 = время полужизни лекарственного средства в конечной фазе элиминации.
a5 животных исключали из анализа AUC, так как они не проявляли AHR (увеличенная AUC) в фазе I.
3 следующих животных исключали вследствие технической неисправности при сборе данных о функционировании дыхательных путей в фазе II.
c2 животных исключали из анализа примирования антигеном из-за технической неисправности при сборе данных о функционировании дыхательных путей в фазе I.
d1 животное исключали из анализа BAL вследствие заметного воспаления BAL в начале исследования.


















| название | год | авторы | номер документа |
|---|---|---|---|
| ПАРТНЕРЫ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С NGF | 2006 |
|
RU2406728C2 |
| ЭЛЕМЕНТ, СВЯЗЫВАЮЩИЙСЯ С α-РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА-4 (IL-4Rα)-173 | 2008 |
|
RU2490278C2 |
| АНТИГЕН, АССОЦИИРОВАННЫЙ С РЕВМАТОИДНЫМ АРТРИТОМ | 2013 |
|
RU2649368C2 |
| СОЕДИНЕНИЯ | 2007 |
|
RU2466139C2 |
| Антитело против IL-4R и его применение | 2018 |
|
RU2758091C1 |
| НЕЙТРАЛИЗУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, НАПРАВЛЕННЫЕ ПРОТИВ ВИРУСА ГРИППА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2721706C2 |
| АНТИТЕЛА, ОБЛАДАЮЩИЕ СПЕЦИФИЧНОСТЬЮ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2794996C2 |
| Антигены, ассоциированные с воспалительным заболеванием кишечника | 2012 |
|
RU2612878C2 |
| ЧЛЕНЫ СВЯЗЫВАНИЯ ПРОТИВ IL-1R1 | 2009 |
|
RU2555532C2 |
| ЛЕЧЕНИЕ ИНФАРКТА МИОКАРДА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ TGF-БЕТА | 2011 |
|
RU2637088C2 |




























































































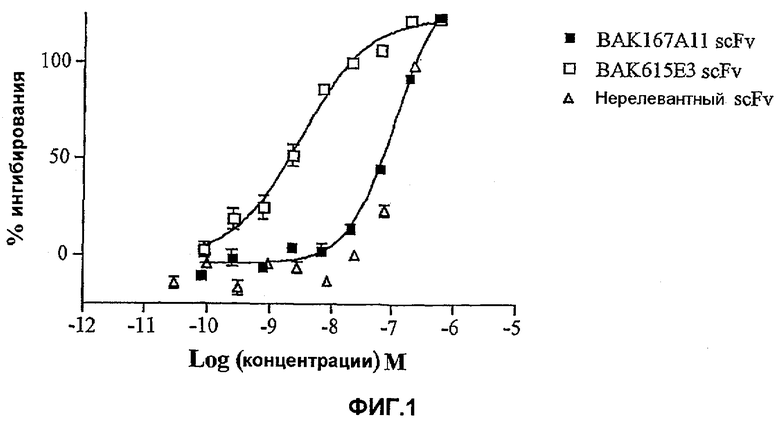

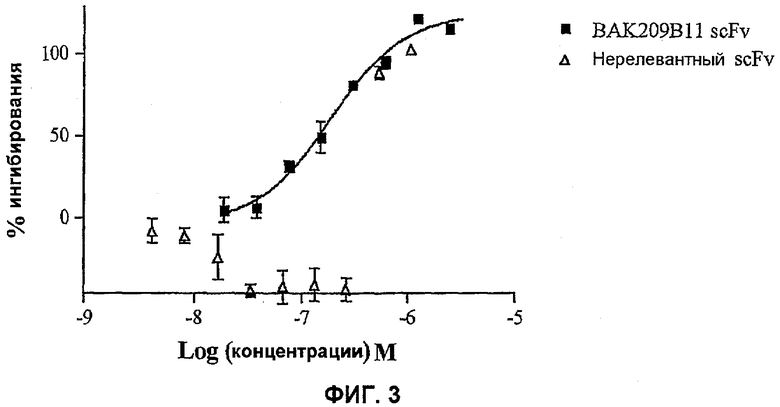
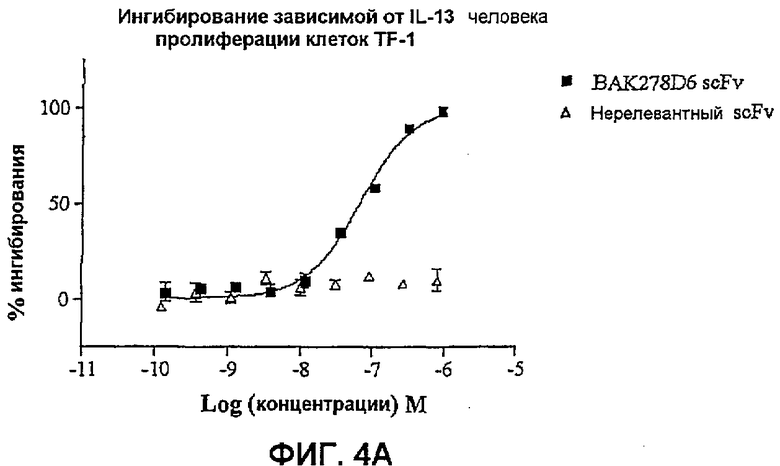
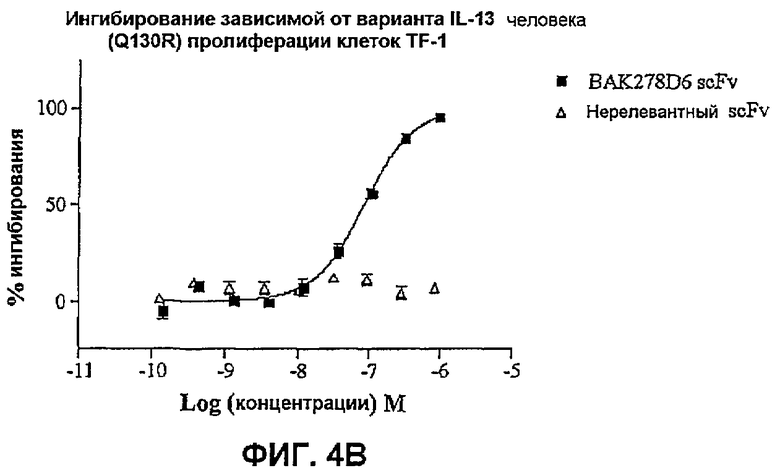
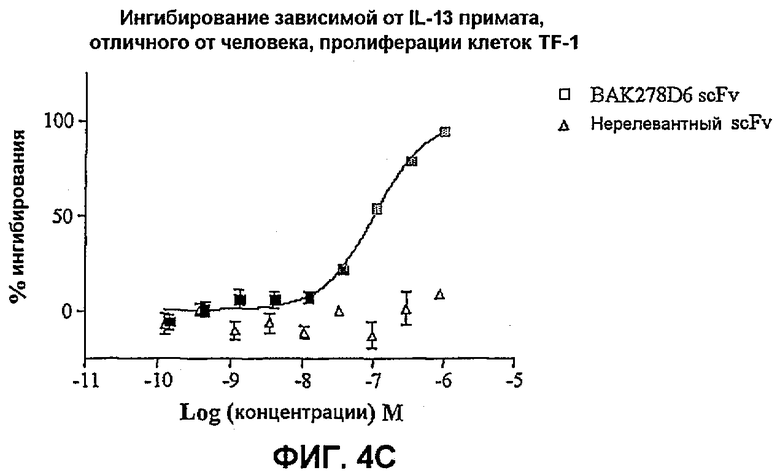
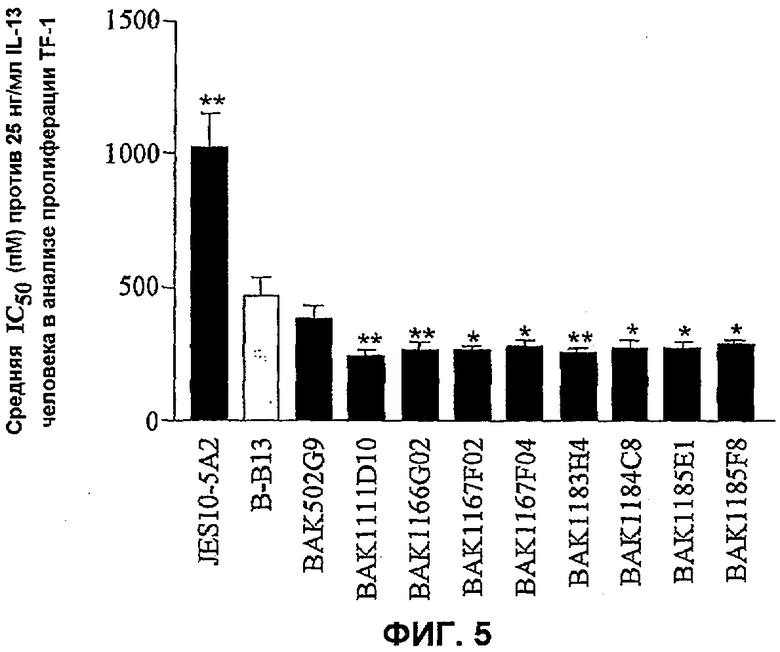
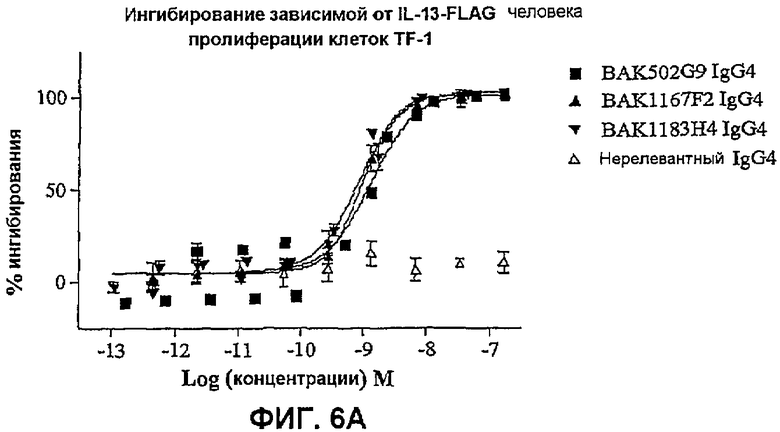
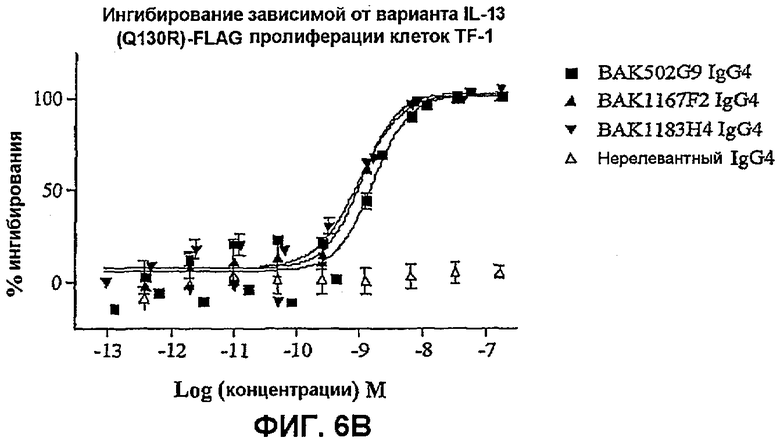
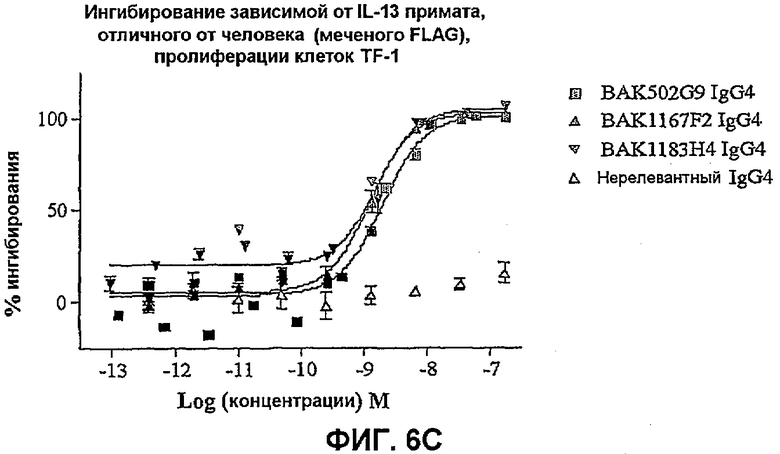
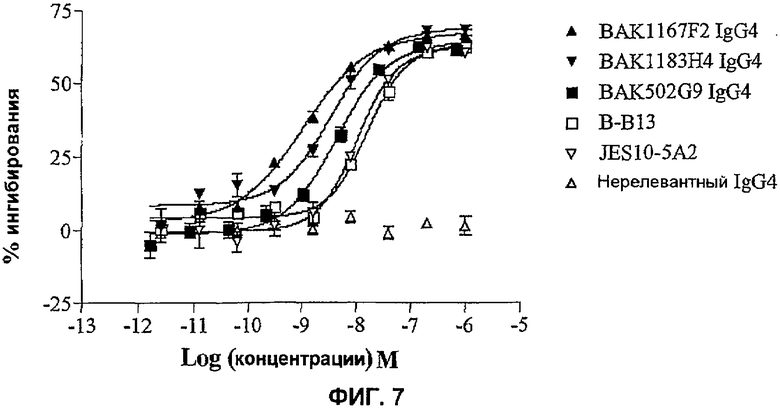
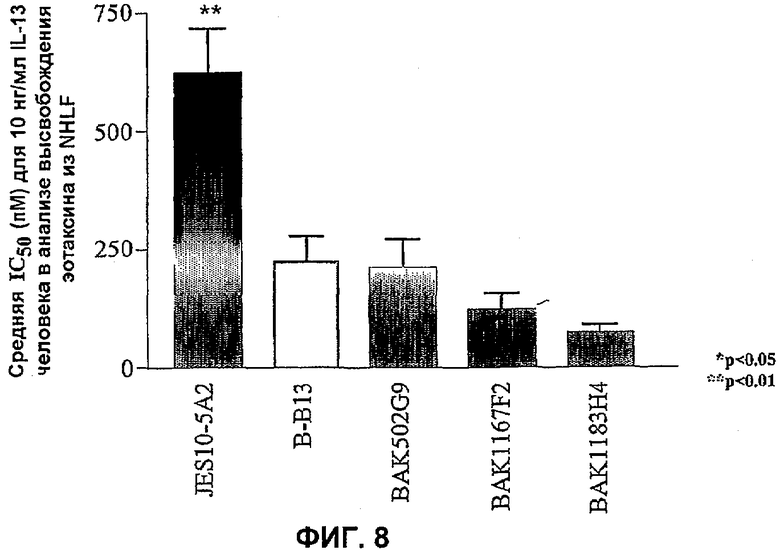
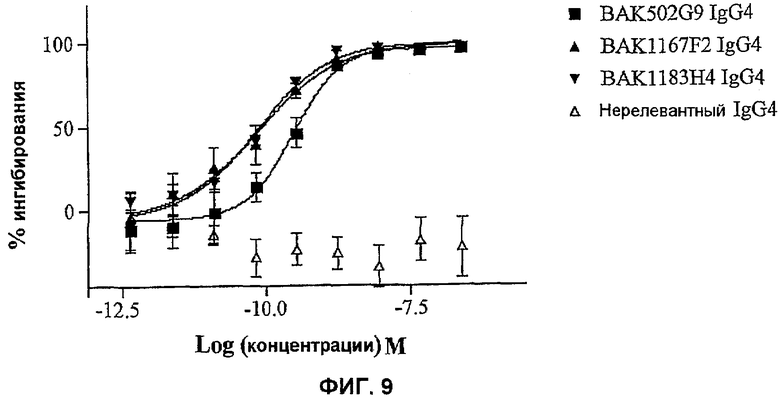
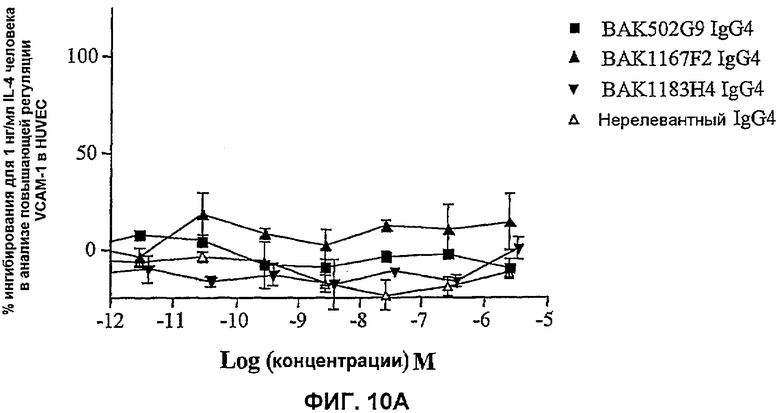
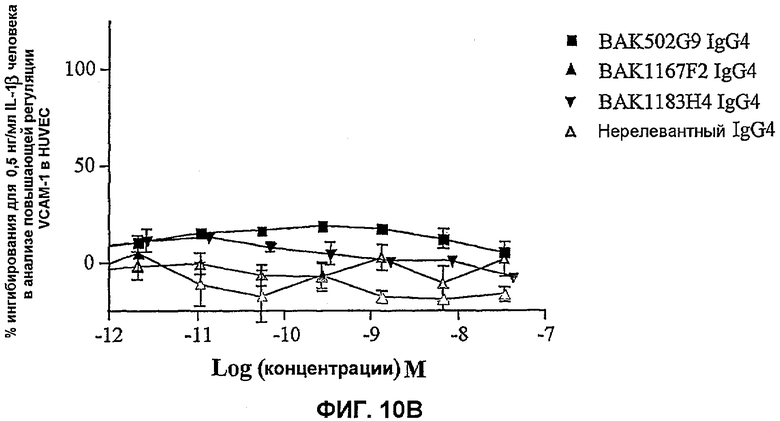
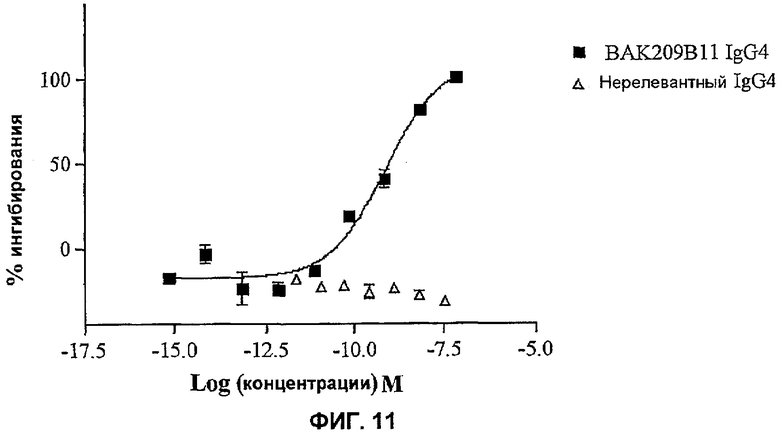
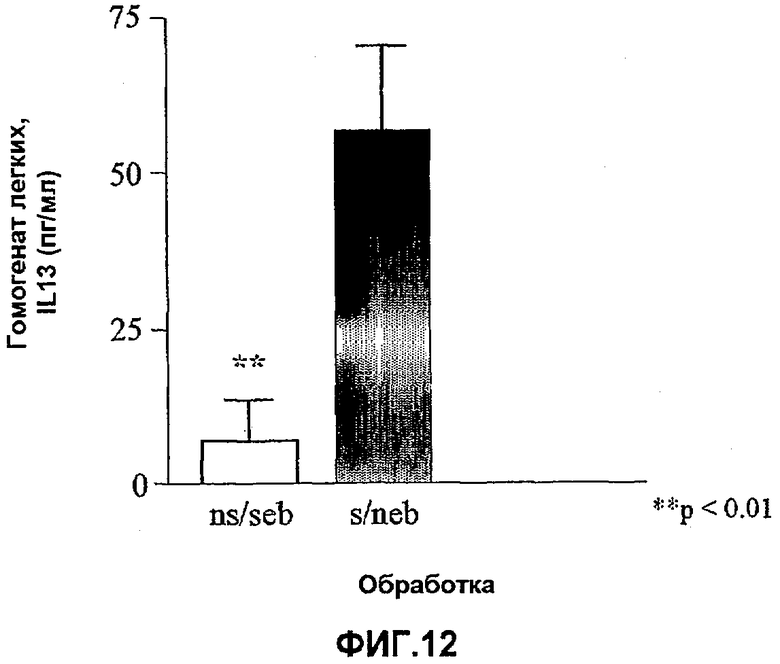
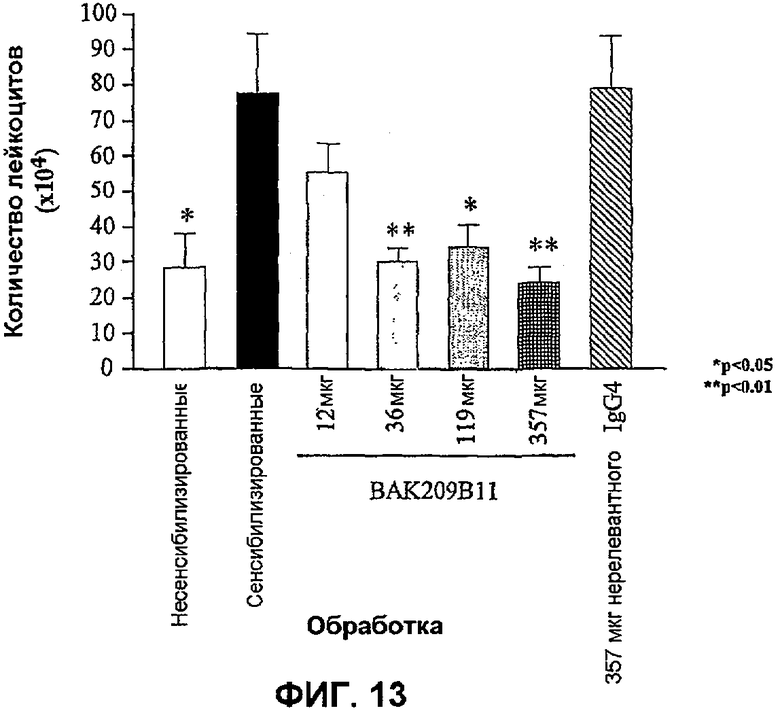
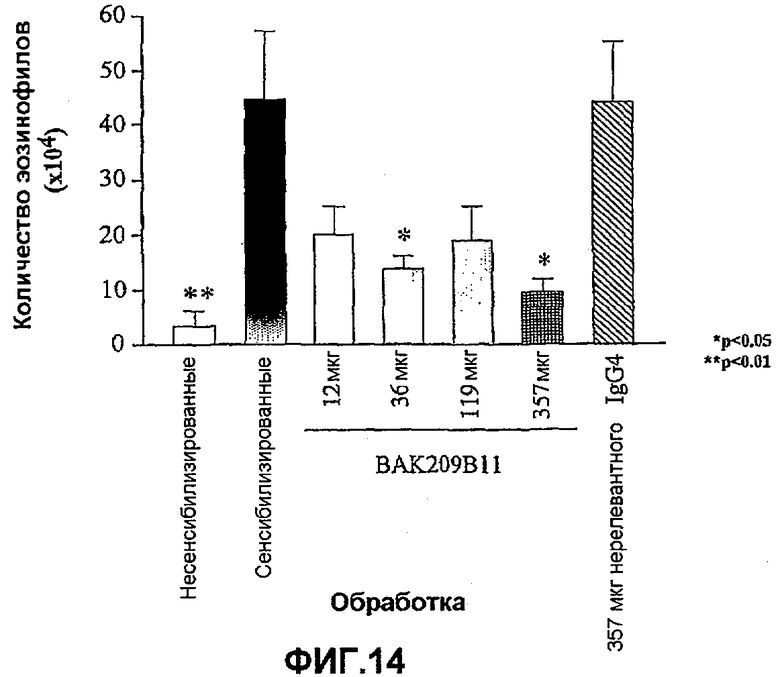
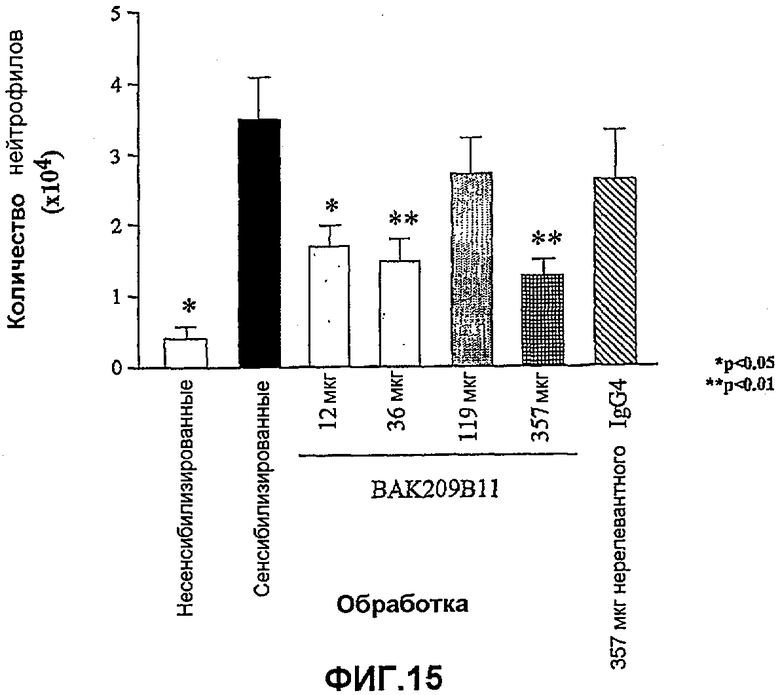
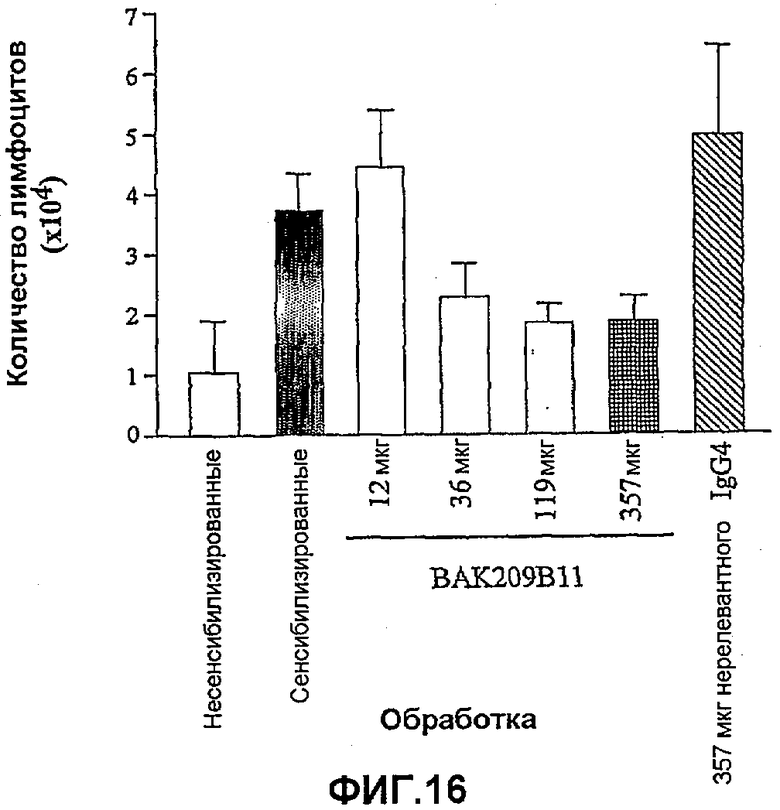
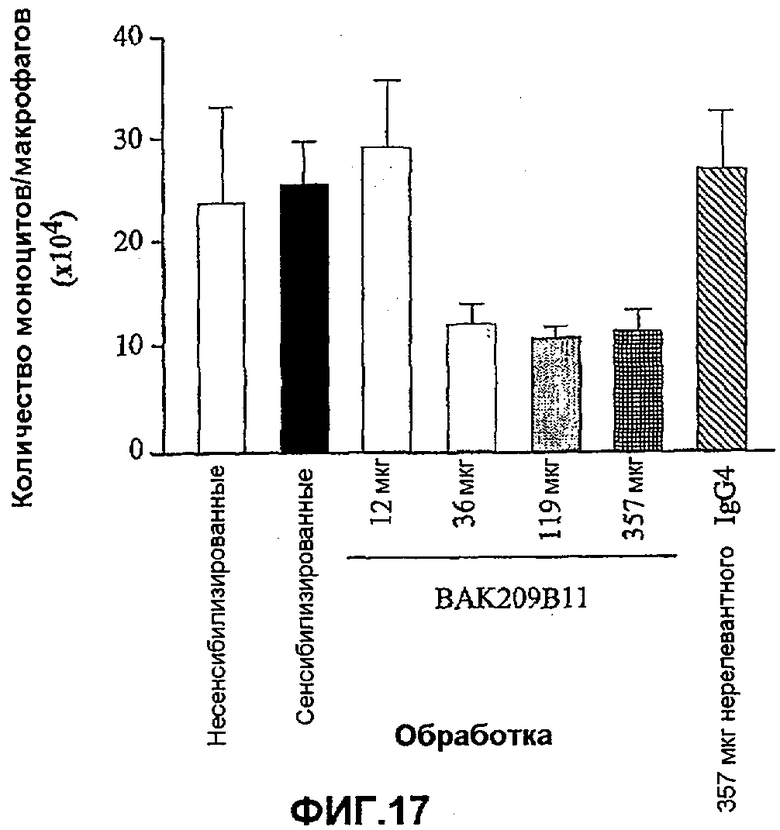
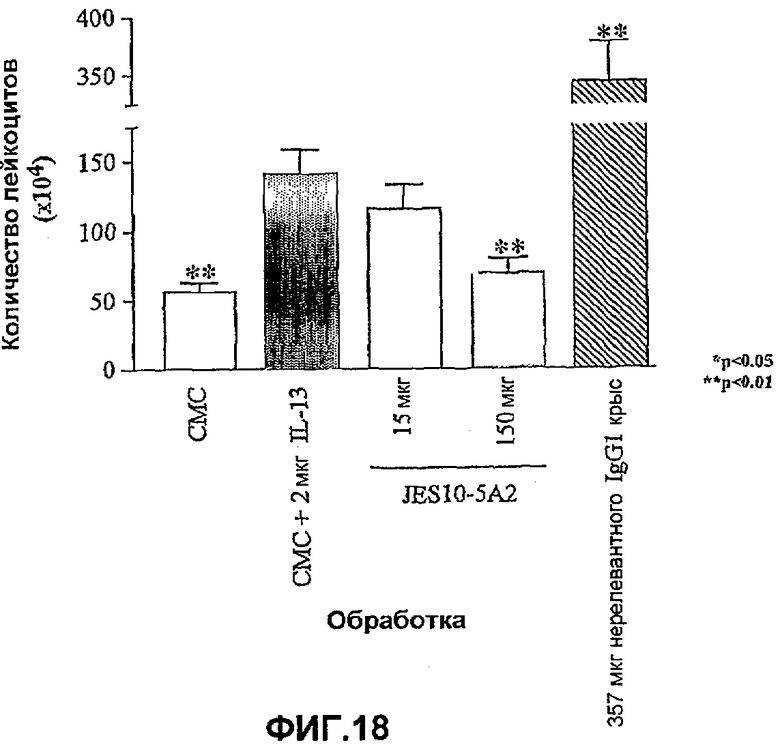
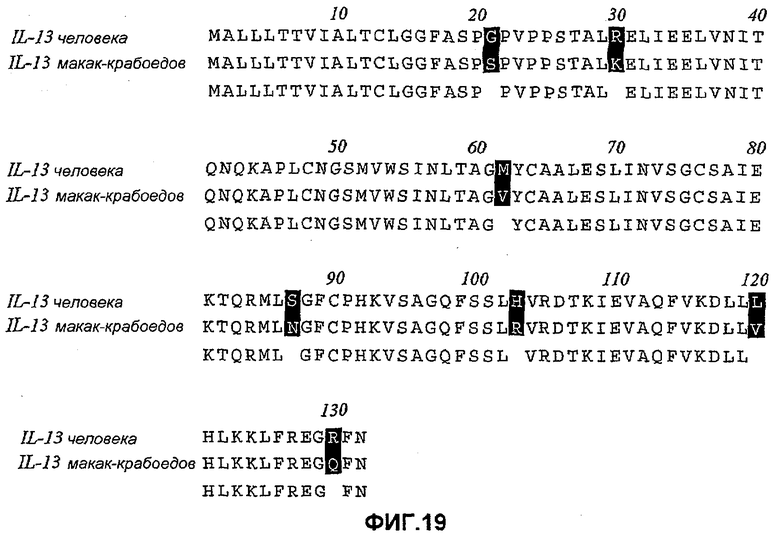
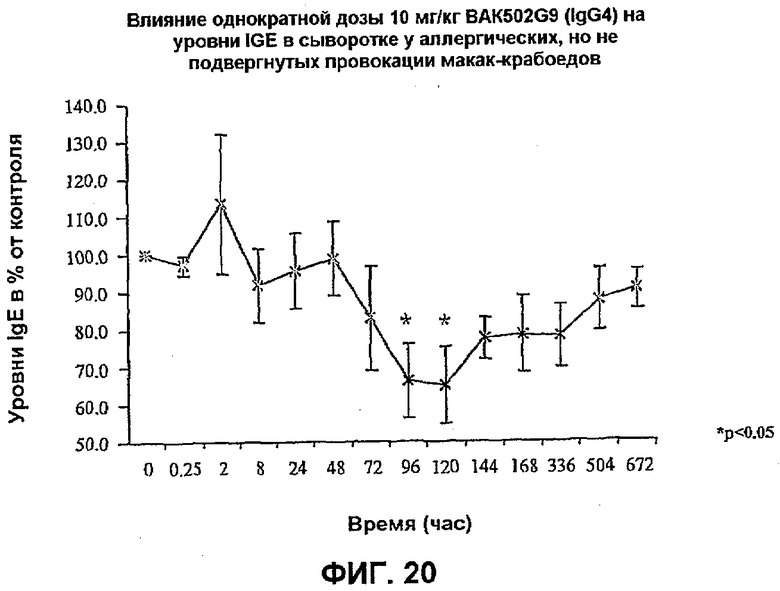
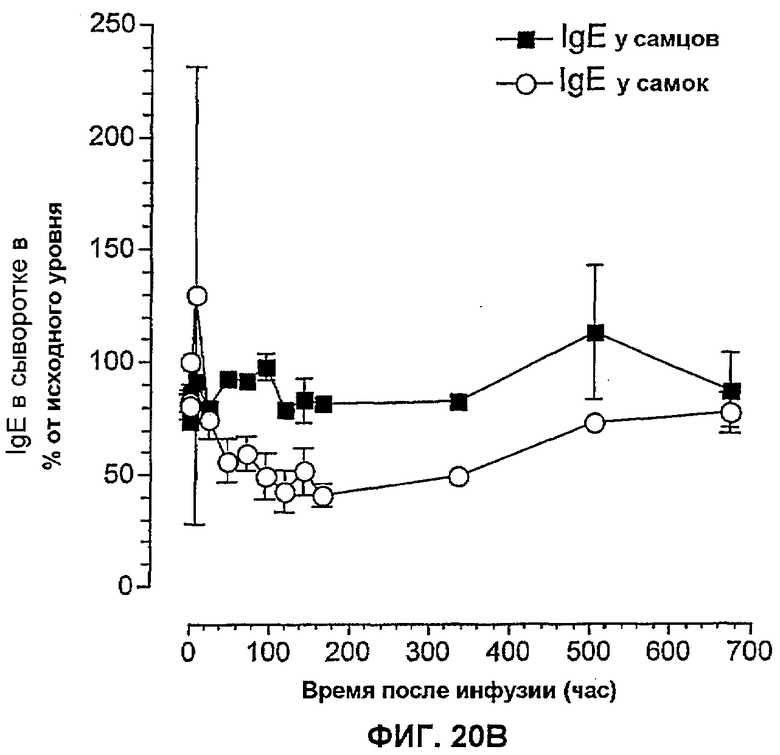
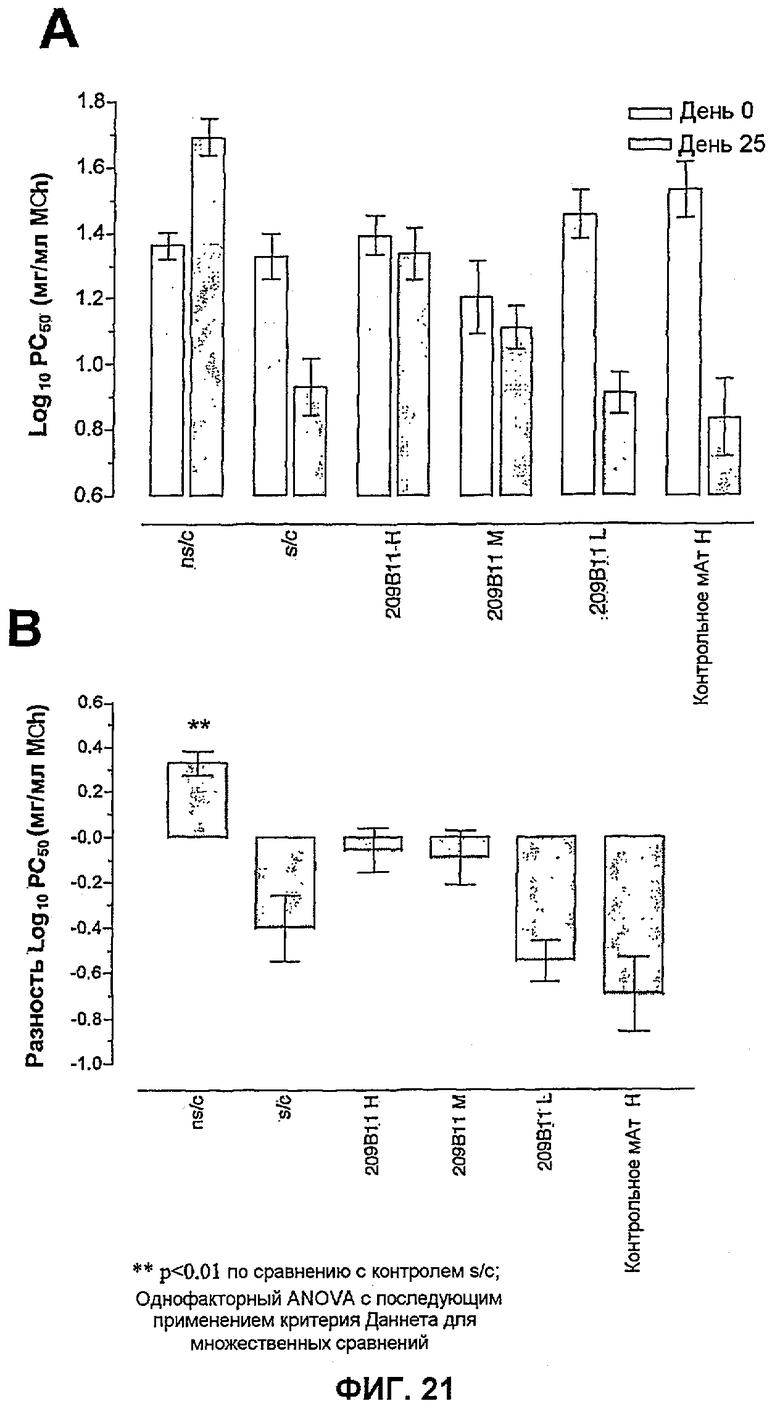
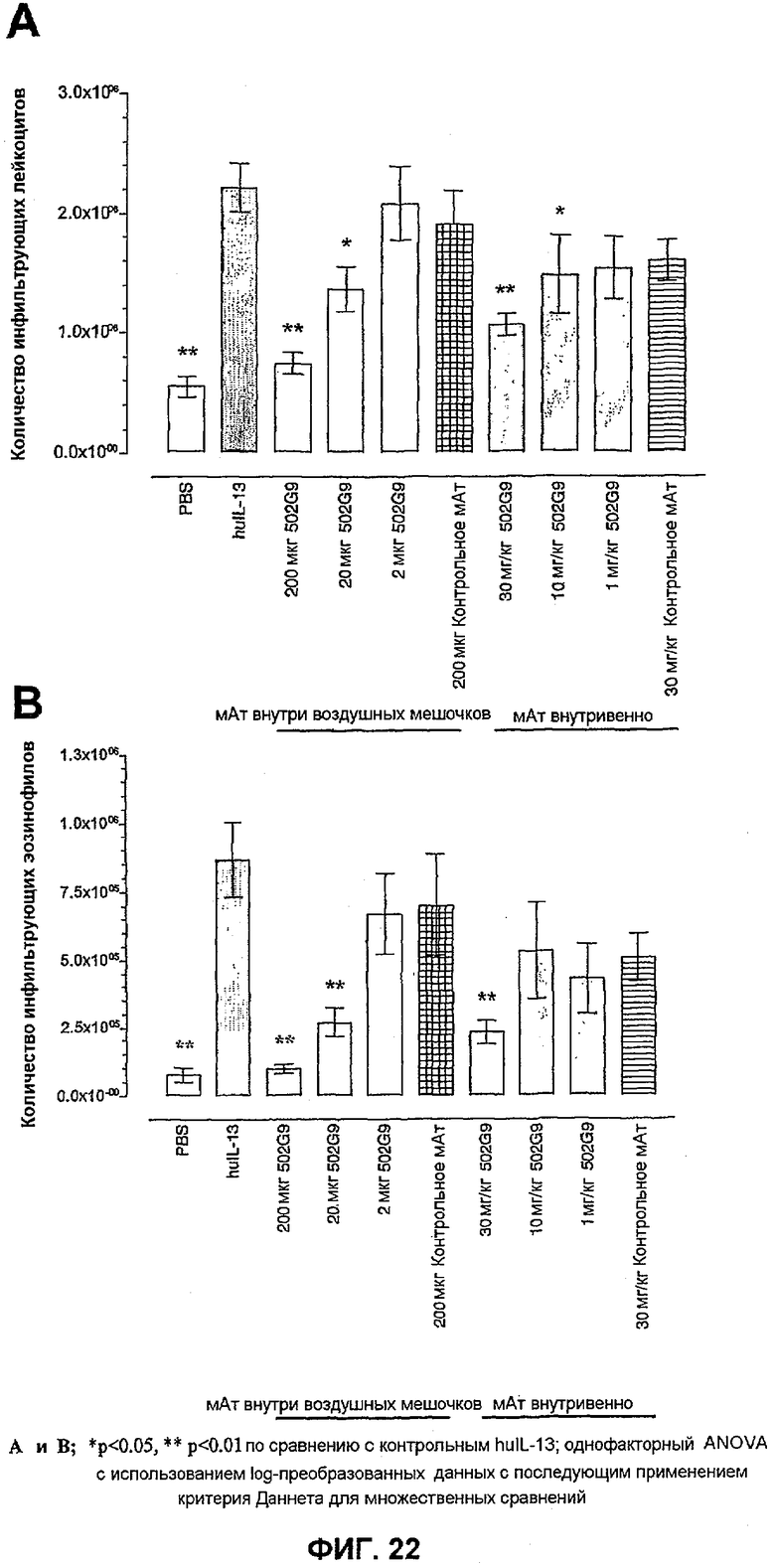
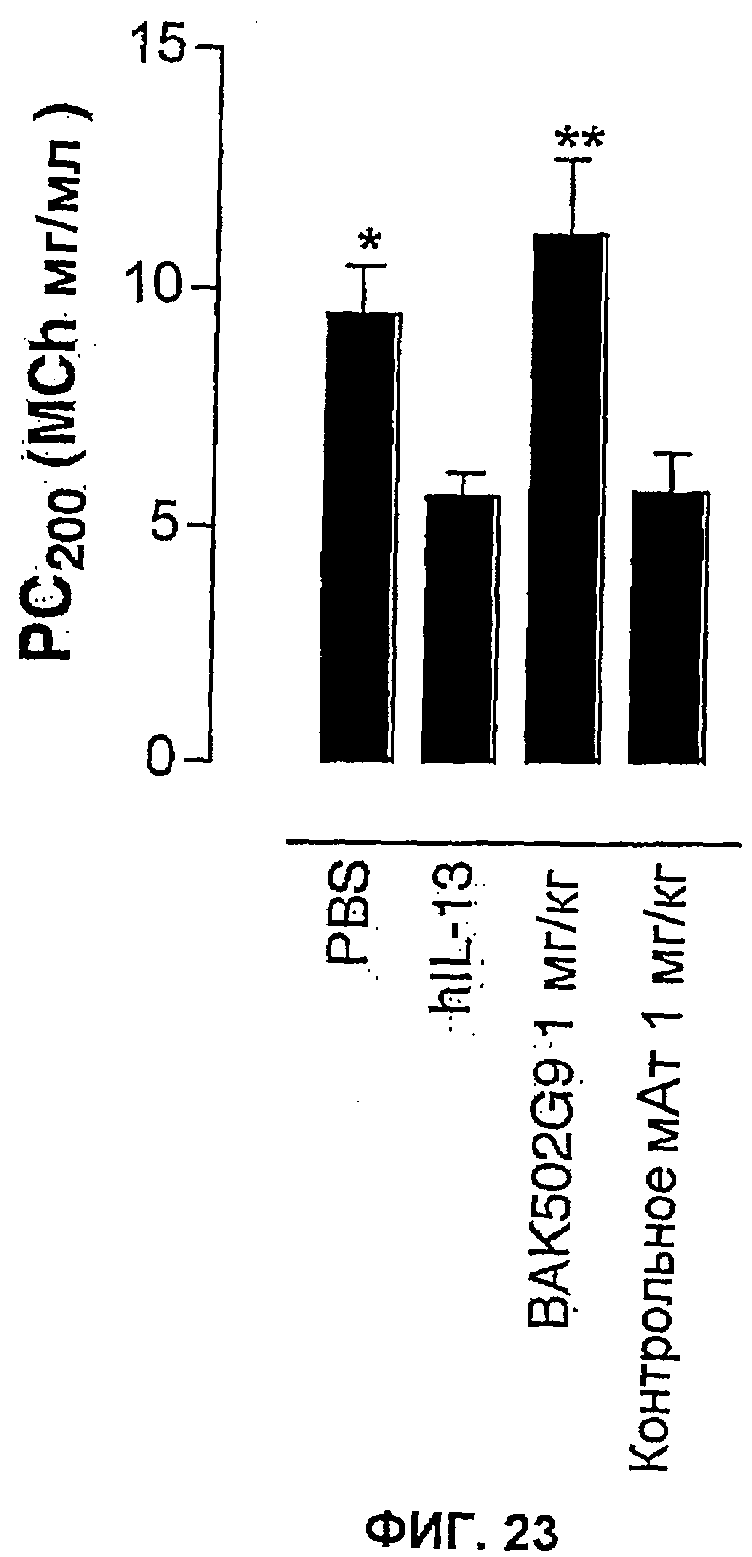
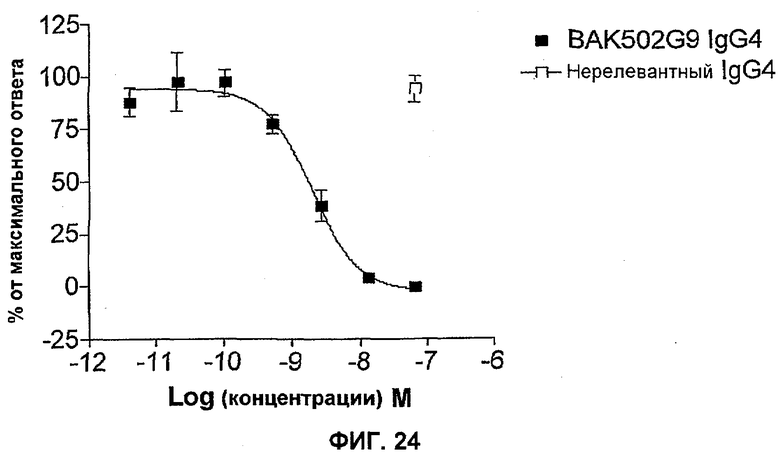
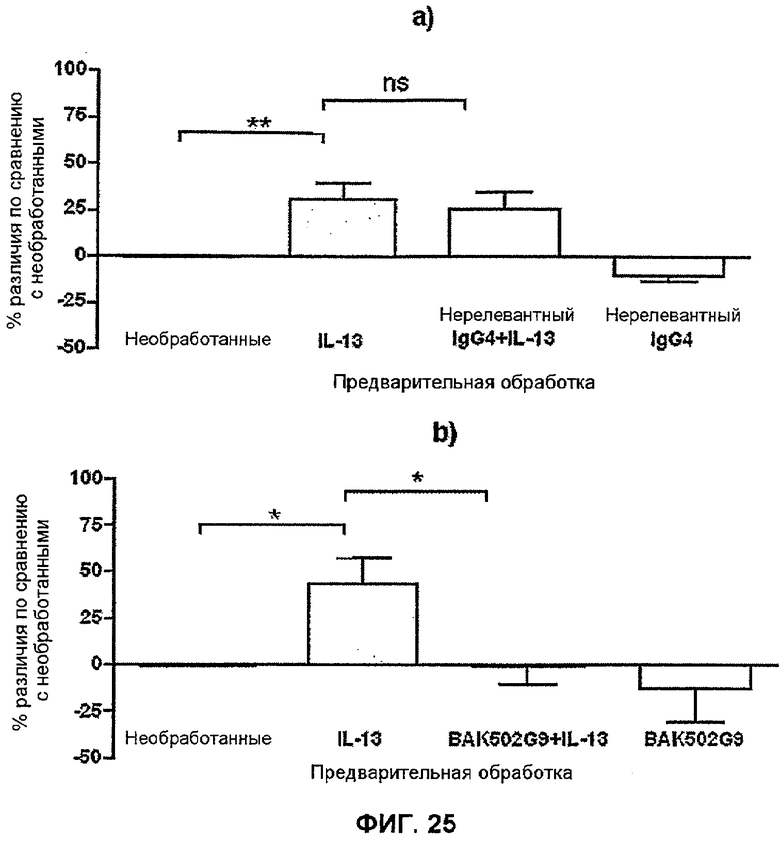
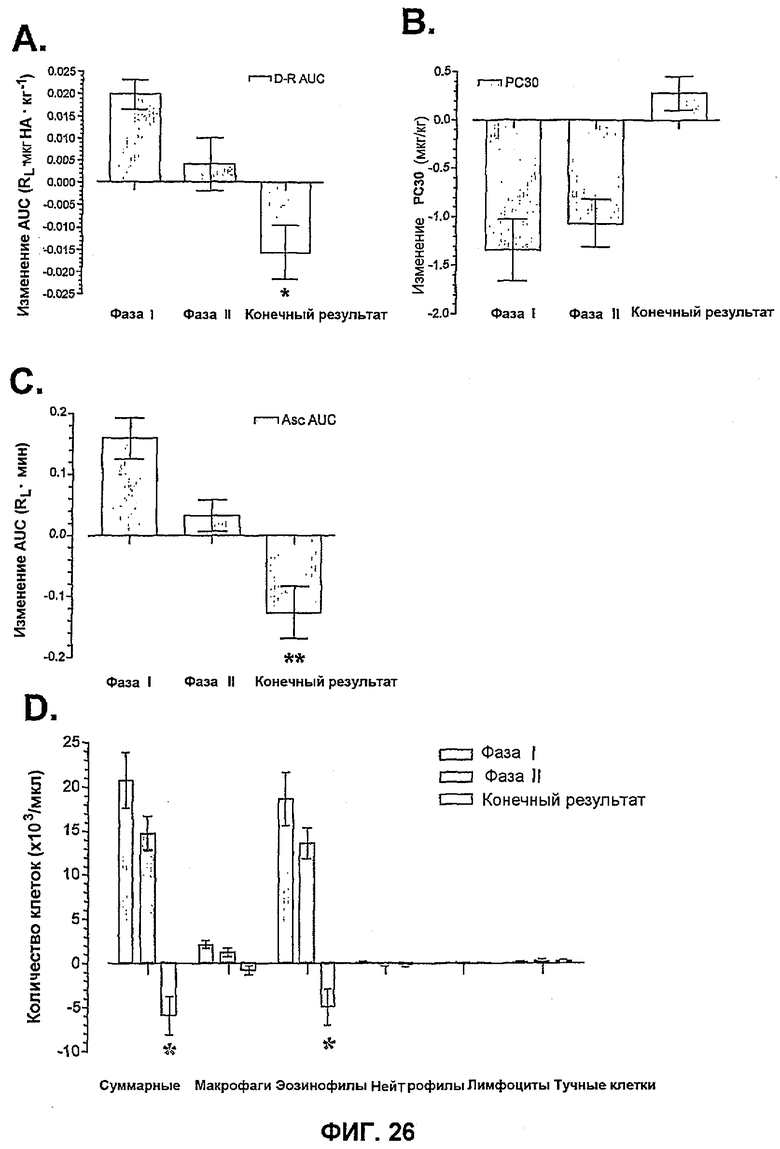
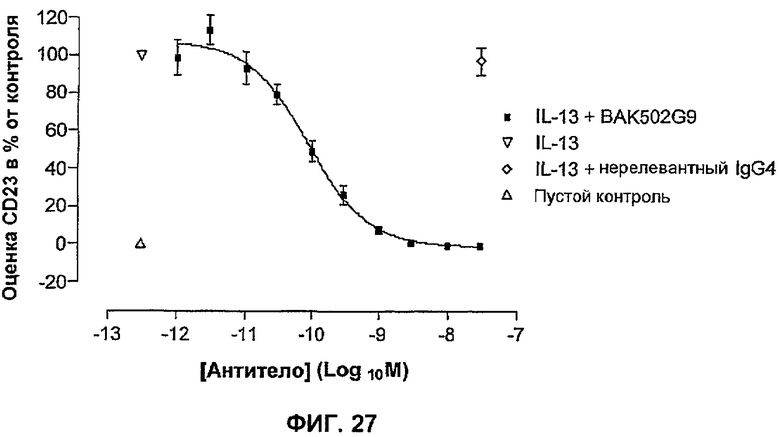
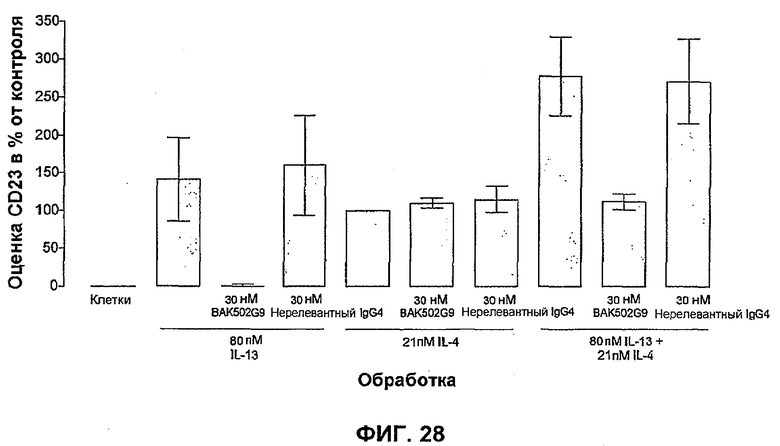
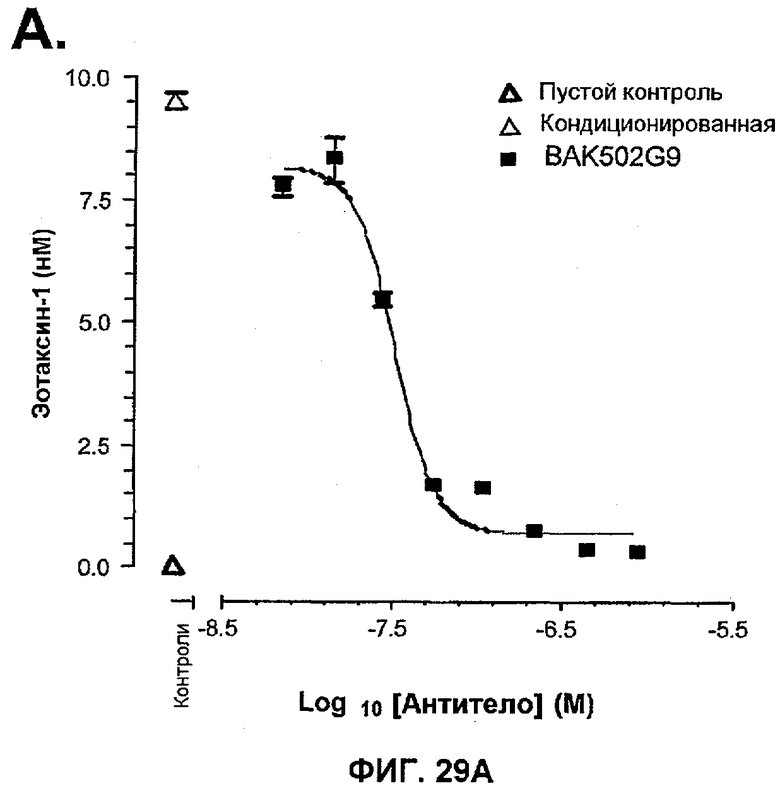
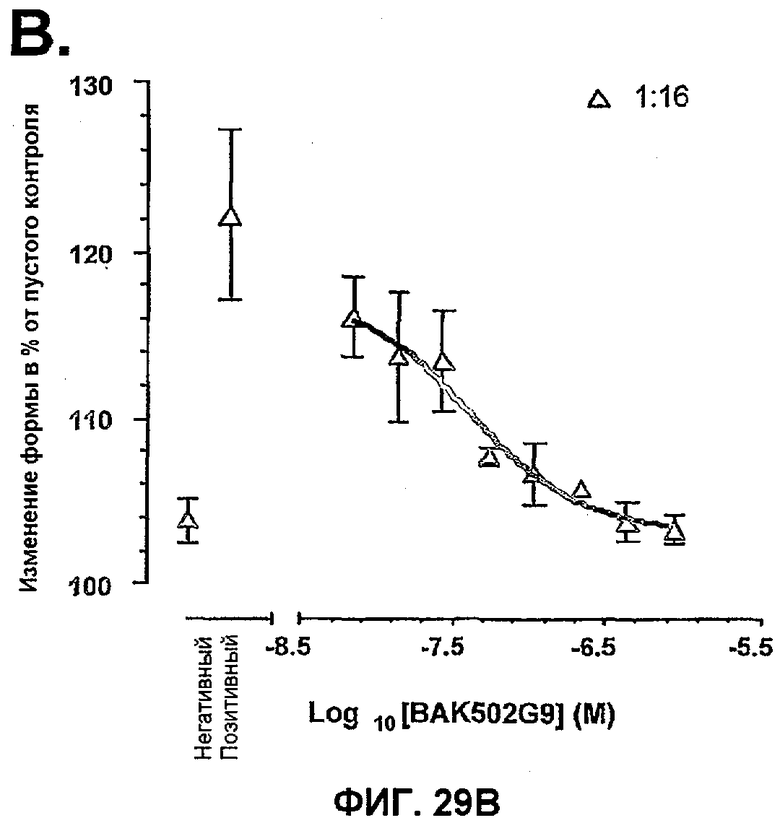
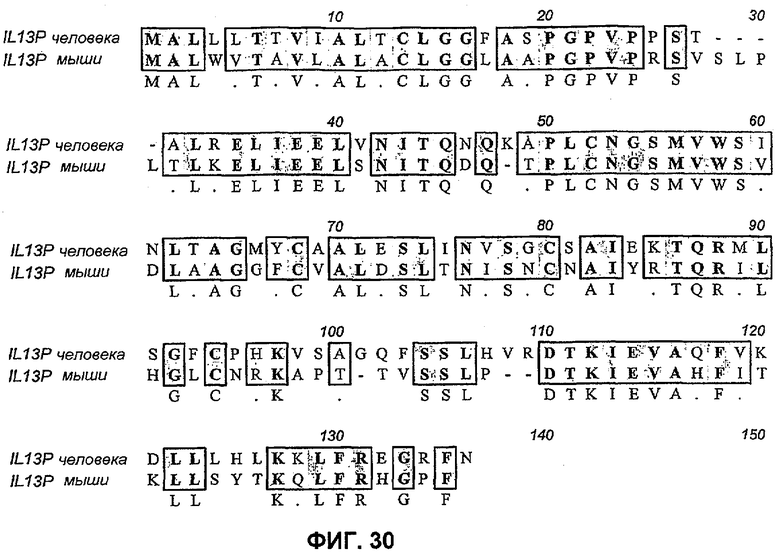
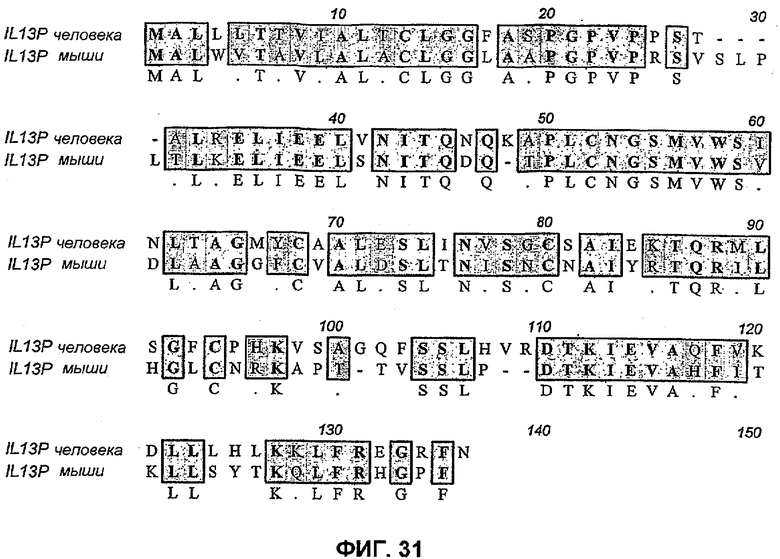

Изобретение относится к иммунологии и биотехнологии. Предложены варианты антител к IL-13 человека, в том числе на основе CDR антитела BAK278D6. Описана композиция на их основе, а также изолированная нуклеиновая кислота, клетка-хозяин для получения антител и варианты способа получения антител. Раскрыто использование антител для производства лекарственного средства и композиции для лечения различных заболеваний, опосредованных активностью IL-13. Использование изобретения обеспечивает антитела, нейтрализующие IL-13, что может найти применение в медицине для приготовления вакцины. 13 н. и 39 з.п. ф-лы, 32 ил., 7 табл.
1. Изолированный специфический связывающий партнер для IL-13 человека, содержащий антигенсвязывающий участок антитела, который состоит из домена VH антитела человека и домена VL антитела человека и который содержит набор CDR HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, при этом домен VH содержит HCDR1, HCDR2 и HCDR3, а домен VL содержит LCDR1, LCDR2 и LCDR3, причем набор CDR состоит из набора CDR, выбранного из группы, состоящей из
набора CDR BAK278D6, в котором, согласно определению,
HCDR1 имеет аминокислотную последовательность SEQ ID NO: 1,
HCDR2 имеет аминокислотную последовательность SEQ ID NO: 2,
HCDR3 имеет аминокислотную последовательность SEQ ID NO: 3,
LCDR1 имеет аминокислотную последовательность SEQ ID NO: 4,
LCDR2 имеет аминокислотную последовательность SEQ ID NO: 5 и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 6,
набора CDR из таблицы 1 с дополнительной одной или двумя аминокислотными заменами по сравнению с набором CDR BAK278D6, и
каждого набора CDR, который показан для отдельных клонов в таблице 1.
2. Изолированный специфический связывающий партнер по п.1, где одна или две замены затрагивают один или два из следующих остатков в CDR, при использовании стандартной нумерации согласно Кабату:
31, 32, 34 в HCDR1
52, 52А, 53, 54, 56, 58, 60, 61, 62, 64, 65 в HCDR2
96, 97, 98, 99, 101 в HCDR3
26, 27, 28, 30, 31 в LCDR1
56 в LCDR2
95A, 97 в LCDR3.
3. Изолированный специфический связывающий партнер по п.2, где одна или две замены осуществлены в следующих положениях из числа идентифицированных групп возможных остатков заместителей для каждого положения:
4. Изолированный специфический связывающий партнер по п.3, где имеется две замены по сравнению с набором CDR BAK278D6, в HCDR3 замена остатка в положении 99 и в LCDR1 замена остатка в положении 27.
5. Изолированный специфический связывающий партнер по п.4, содержащий набор CDR BAK278D6 с заменой остатка в положении 99 в HCDR3, выбранной из группы, состоящей из S, A, I, R, Р и К, и/или заменой в положении остатка 27 в LCDR1, выбранной из группы, состоящей из I, L, M, С, V, K, Y, F, R, T, S, A, H и G.
6. Изолированный специфический связывающий партнер по п.4, содержащий набор CDR BAK278D6 с заменой остатка N на S в положении 99 в HCDR3 и/или с заменой остатка N на I в положении 27 в LCDR1.
7. Изолированный специфический связывающий партнер по п.2, в котором HCDR1, HCDR2 и HCDR3 домена VH находятся в каркасе зародышевой линии и/или LCDR1, LCDR2 и LCDR3 домена VL находятся в каркасе зародышевой линии.
8. Изолированный специфический связывающий партнер по п.7, в котором HCDR1, HCDR2 и HCDR3 домена VH находятся в каркасе зародышевой линии VH1 DP14.
9. Изолированный специфический связывающий партнер по п.7, в котором HCDR1, HCDR2 и HCDR3 домена VH находятся в каркасе зародышевой линии VL Vλ3 3h.
10. Изолированный специфический связывающий партнер по п.2, который дополнительно связывает вариант IL-13 человека, в котором аргинин в положении 130 заменен глутамином.
11. Изолированный специфический связывающий партнер по любому из пп.1-10, который дополнительно связывает IL-13 отличного от человека примата.
12. Изолированный специфический связывающий партнер по п.11, в котором IL-13 отличного от человека примата является IL-13 макаки-резус или макаки-крабоеда.
13. Изолированный специфический связывающий партнер по п.8, содержащий VH-домен BAK502G9 (SEQ ID NO: 15).
14. Изолированный специфический связывающий партнер по п.8, содержащий VL-домен BAK502G9 (SEQ ID NO: 16).
15. Изолированный специфический связывающий партнер по п.13, содержащий VL-домен BAK502G9 (SEQ ID NO: 16).
16. Изолированный специфический связывающий партнер по любому из пп.1-10, 13 или 15, который связывает IL-13 с аффинностью, равной или большей, чем аффинность антигенсвязывающего участка для IL-13, образованного VH-доменом BAK502G9 (SEQ ID NO: 15) и VL-доменом BAK502G9 (SEQ ID NO: 16), причем аффинность специфического связывающего партнера и аффинность антигенсвязывающего участка определяют в одних и тех же условиях.
17. Изолированный специфический связывающий партнер по любому из пп.1-10, 13 или 15, связывание которого с IL-13 человека приводит к нейтрализации IL-13 человека.
18. Изолированный специфический связывающий партнер по п.17, который нейтрализует IL-13 человека с эффективностью, равной или большей, чем эффективность антигенсвязывающего участка для IL-13, образованного VH-доменом BAK502G9 (SEQ ID NO: 15) и VL-доменом BAK502G9 (SEQ ID NO: 16), причем эффективность специфического связывающего партнера и эффективность антигенсвязывающего участка определяют в одних и тех же условиях.
19. Изолированный специфический связывающий партнер по любому из пп.1-10, 13 или 15, который содержит молекулу антитела scFv или константную область антитела, или целое антитело, где целое антитело необязательно является IgG4.
20. Изолированный VH-домен антитела в составе специфического связывающего партнера по любому из пп.1-19.
21. Изолированный VL-домен антитела в составе специфического связывающего партнера по любому из пп.1-19.
22. Композиция для лечения связанного с IL-13 расстройства, содержащая специфический связывающий партнер, домен VH или VL антитела по любому из пп.1-21 и по меньшей мере один дополнительный компонент.
23. Композиция по п.22, содержащая фармацевтически приемлемый эксципиент, наполнитель или носитель.
24. Изолированная нуклеиновая кислота, кодирующая специфический связывающий партнер или домен VH или VL антитела в составе специфического связывающего партнера по любому из пп.1-21.
25. Клетка-хозяин, трансформированная in vitro нуклеиновой кислотой по п.24, для получения специфического связывающего партнера, или домена VH, или VL антитела по любому из пп.1-21.
26. Способ получения специфического связывающего партнера, или домена VH, или VL антитела по любому из пп.1-21 путем культивирования клеток-хозяев по п.25.
27. Способ по п.26, дополнительно включающий в себя выделение и/или очистку указанного специфического связывающего партнера или вариабельного домена VH или VL антитела.
28. Способ получения антигенсвязывающего домена антитела, специфичного в отношении IL-13 человека, при этом способ включает в себя
получение посредством дополнительной замены согласно таблице 1 одной или двух аминокислот в аминокислотных последовательностях CDR исходного домена VH, содержащего HCDR1, HCDR2 и HCDR3, где HCDR1, HCDR2 и HCDR3 исходного домена VH представляют собой набор HCDR BAK278D6, в котором, согласно определению, HCDR1 имеет аминокислотную последовательность SEQ ID NO: 1, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 2, HCDR3 имеет аминокислотную последовательность SEQ ID NO: 3, или набор HCDR BAK502G9, в котором, согласно определению, HCDR1 имеет аминокислотную последовательность SEQ ID NO: 7, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 8, HCDR3 имеет аминокислотную последовательность SEQ ID NO: 9, домена VH, который представляет собой вариант аминокислотной последовательности исходного домена VH, и, необязательно, объединение полученного таким образом домена VH с одним или несколькими доменами VL, с получением одной или нескольких комбинаций VH/VL;
и тестирование указанного домена VH, который является вариантом аминокислотной последовательности исходного домена VH, или комбинации или комбинаций VH/VL, с идентификацией антигенсвязывающего домена антитела, специфичного в отношении IL-13 человека.
29. Способ по п.28, в котором аминокислотная последовательность исходного домена VH выбрана из группы, состоящей из SEQ ID NO: 13 и SEQ ID NO: 15.
30. Способ по п.28, в котором указанный один или несколько доменов VL получают посредством дополнительной замены согласно таблице 1 одной или двух аминокислот в аминокислотных последовательностях CDR исходного домена VL, содержащего LCDR1, LCDR2 и LCDR3, где LCDR1, LCDR2 и LCDR3 исходного домена VL представляют собой набор LCDR BAK278D6, в котором, согласно определению, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 4, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 5, LCDR3 имеет аминокислотную последовательность SEQ ID NO: 6, или набор LCDR BAK502G9, в котором, согласно определению, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 10, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 11, LCDR3 имеет аминокислотную последовательность SEQ ID NO: 12, с получением одного или нескольких доменов VL, каждый из которых является вариантом аминокислотной последовательности исходного домена VL.
31. Способ по п.30, в котором аминокислотная последовательность исходного домена VL выбрана из группы, состоящей из SEQ ID NO: 14 и SEQ ID NO: 16.
32. Способ по любому из пп.28-31, дополнительно включающий в себя получение антигенсвязывающего участка антитела в молекуле антитела IgG, scFv или Fab.
33. Способ получения специфического связывающего партнера, который связывает IL-13 человека, при этом способ включает в себя
получение исходной нуклеиновой кислоты, кодирующей домен VH, или исходного репертуара нуклеиновых кислот, каждая из которых кодирует домен VH, при этом домен VH или домены VH либо содержат HCDR1, HCDR2 и/или HCDR3, которые подлежат замене, либо лишены области HCDR1, HCDR2 и/или HCDR3;
объединение указанной исходной нуклеиновой кислоты или исходного репертуара с донорной нуклеиновой кислотой или донорными нуклеиновыми кислотами, кодирующими аминокислотную последовательность HCDR1 (SEQ ID NO: 1) или HCDR1 (SEQ ID NO: 7), HCDR2 (SEQ ID NO: 2) или HCDR2 (SEQ ID NO: 8) и/или HCDR3 (SEQ ID NO: 3) или HCDR3 (SEQ ID NO: 9) или полученными посредством дополнительной замены согласно таблице 1 одной или двух аминокислот в указанных аминокислотных последовательностях,
так, чтобы указанная донорная нуклеиновая кислота или указанные донорные нуклеиновые кислоты были встроены в область CDR1, CDR2 и/или CDR3 в исходной нуклеиновой кислоте или исходном репертуаре, с тем, чтобы в результате получить продукт в виде репертуара нуклеиновых кислот, кодирующих домены VH;
экспрессию нуклеиновых кислот полученного продукта в виде репертуара, с получением продуктов в виде доменов VH;
необязательно объединение указанного продукта в виде доменов VH с одним или несколькими доменами VL;
отбор специфического связывающего партнера, специфичного в отношении IL-13 человека, содержащего полученный продукт в виде домена VH и, необязательно, домен VL; и
извлечение указанного специфического связывающего партнера или кодирующей его нуклеиновой кислоты.
34. Способ по п.33, согласно которому донорные нуклеиновые кислоты получают в результате мутации указанной HCDR1 и/или HCDR2.
35. Способ по п.33, согласно которому донорную нуклеиновую кислоту получают в результате мутации HCDR3.
36. Способ по п.35, включающий в себя получение донорной нуклеиновой кислоты посредством мутации нуклеиновой кислоты, кодирующей аминокислотную последовательность HCDR3 (SEQ ID NO: 3) или HCDR3 (SEQ ID NO: 9).
37. Способ по любому из пп.33-36, дополнительно включающий в себя связывание полученного домена VH, который находится в извлеченном специфическом связывающем партнере, с константной областью антитела.
38. Способ по любому из пп.33-36, включающий в себя получение молекулы антитела IgG, scFv или Fab, содержащей полученный домен VH и домен VL.
39. Способ по любому из пп.28 или 33, дополнительно включающий в себя тестирование антигенсвязывающего домена антитела или специфического связывающего партнера, который связывает IL-13 человека, в отношении способности нейтрализовать IL-13 человека.
40. Способ по п.32, дополнительно включающий в себя тестирование антигенсвязывающего домена антитела или специфического связывающего партнера, который связывает IL-13 человека, в отношении способности нейтрализовать IL-13 человека.
41. Способ по п.37, дополнительно включающий в себя тестирование антигенсвязывающего домена антитела или специфического связывающего партнера, который связывает IL-13 человека, в отношении способности нейтрализовать IL-13 человека.
42. Способ по п.38, дополнительно включающий в себя тестирование антигенсвязывающего домена антитела или специфического связывающего партнера, который связывает IL-13 человека, в отношении способности нейтрализовать IL-13 человека.
43. Способ по п.39, согласно которому получают специфический связывающий партнер, который содержит фрагмент антитела, который связывает и нейтрализует IL-13 человека.
44. Способ по п.43, согласно которому фрагментом антитела является молекула антитела scFv или молекула антитела Fab, где необязательно указанный способ дополнительно включает в себя получение домена VH и/или домена VL фрагмента антитела в целом антителе.
45. Применение специфического связывающего партнера или вариабельного домена VH или VL антитела, полученных способом по п.27, или специфического связывающего партнера, который связывает IL-13, антигенсвязывающего участка антитела или вариабельного домена VH или VL антитела, которое содержит специфически связывающий партнер, или антигенсвязывающего участка антитела, который связывает IL-13, полученных способом по любому из пп.28, 32, 33, 37 и 38, для получения композиции для нейтрализации IL-13, содержащей по меньшей мере один дополнительный компонент.
46. Способ по любому из пп.28 или 33, дополнительно включающий в себя связывание специфического связывающего партнера, который связывает IL-13 человека, с IL-13 или фрагментом IL-13, где указанное связывание происходит in vitro.
47. Способ по п.32, дополнительно включающий в себя связывание специфического связывающего партнера, который связывает IL-13 человека, с IL-13 или фрагментом IL-13, где указанное связывание происходит in vitro.
48. Способ по п.37, дополнительно включающий в себя связывание специфического связывающего партнера, который связывает IL-13 человека, с IL-13 или фрагментом IL-13, где указанное связывание происходит in vitro.
49. Способ по п.38, дополнительно включающий в себя связывание специфического связывающего партнера, который связывает IL-13 человека, с IL-13 или фрагментом IL-13, где указанное связывание происходит in vitro.
50. Применение специфического связывающего партнера, который связывает IL-13 по любому из пп.1-21, в способе определения количества IL-13 в тестируемом образце in vitro, согласно которому создают условия для связывания указанного специфического связывающего партнера с IL-13 человека или фрагментом IL-13 человека и определяют количество связавшегося специфического связывающего партнера с IL-13 или фрагментом IL-13.
51. Применение специфического связывающего партнера, полученного способом по любому из пп.28, 32, 33, 37 и 38, для производства лекарственного средства для лечения заболевания или расстройства, выбранного из группы, состоящей из астмы, атопического дерматита, аллергического ринита, фиброза, воспалительного заболевания кишечника и лимфомы Ходжкина.
52. Применение специфического связывающего партнера по любому из пп.1-19 в производстве лекарственного средства для лечения заболевания или расстройства, выбранного из группы, состоящей из астмы, атопического дерматита, аллергического ринита, фиброза, воспалительного заболевания кишечника и лимфомы Ходжкина.
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| КОМПОЗИЦИЯ ДЛЯ ИНГИБИРОВАНИЯ АНГИОГЕНЕЗА, МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, ПОЛИПЕПТИД, СПОСОБ ИНГИБИРОВАНИЯ РОСТА ОПУХОЛИ (ВАРИАНТЫ) | 1992 |
|
RU2170589C2 |

