ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к биспецифическим антителам, содержащим по меньшей мере два Fab фрагмента, где первый Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого антигена, и второй Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении второго антигена, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab и где биспецифическое антитело лишено Fc домена; способам их получения, фармацевтическим композициям, содержащим указанные антитела, и их применению.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Моноклональные антитела (mAbs) представляют собой все более важный класс терапевтических агентов. Кроме продуктов mAb, состоящих из полноразмерных форм IgG, были разработаны разнообразные форматы мультиспецифических рекомбинантных антител, например, тетравалентные биспецифические антитела, созданные путем слияния, например, антител в формате IgG и одноцепочечных доменов (см., например, Coloma, M.J., et al., Nature Biotech 15 (1997) 159-163; WO 2001/077342; и Morrison, S.L, Nature Biotech 25(2007) 1233-1234).
Разработали несколько других новых форматов антител, способных связывать два или более антигенов, у которых структура остова антитела (IgA, IgD, IgE, IgG или IgM) не сохраняется, такие как диа-, триа- или тетратела, миниантитела, и несколько одноцепочечных форматов (scFv, Bis-scFv), (Holliger, Р., et al., Nature Biotech 23 (2005) 1126-1136; Fischer, N., Leger, 0., Pathobiology 74 (2007) 3-14; Shen, J., et al., Journal of Immunological Methods 318 (2007) 65-74; Wu, С., et al., Nature Biotech. 25 (2007) 1290-1297).
Во всех этих форматах используются линкеры либо для объединения остова антитела (IgA, IgD, IgE, IgG или IgM) с другим связывающим белком (например, scFv), либо для объединения, например, двух Fab фрагментов или scFv (Fischer, N., Leger, 0-, Pathobiology 74 (2007) 3-14). Тандемные scFV представляют собой два фрагмента scFv, связанные при помощи дополнительного пептидного линкера, они также обозначаются (scFv)2. За счет сокращения длины пептидного линкера между вариабельными доменами были получены диатела. Добавление дополнительного пептидного линкера между двумя полипептидами позволяет получить т.н. одноцепочечные диатела. US 2007/0274985 относится к антителам, содержащим одноцепочечные Fab (scFab) фрагменты. Использование фрагментов антител в качестве терапевтических средств имеет как преимущества, так и недостатки по сравнению с полноразмерными моноклональными антителами: одно из преимуществ состоит в том, что они обладают меньшим размером и быстрее проникают в ткани и опухоли. Кроме того, предполагается, что малый размер фрагментов позволяет связываться с эпитопами, недоступными для полноразмерных моноклональных антител. С другой стороны, фрагменты демонстрируют короткое время полужизни в кровотоке человека, возможно вследствие почечного клиренса. Более короткое время полужизни может препятствовать достаточному накоплению терапевтического средства в целевом участке. Получение фрагментов антител является непростой задачей, поскольку фрагменты склонны к формированию агрегатов и могут быть менее стабильными по сравнению с полноразмерными моноклональными антителами. Кроме того, нежелательное образование пар между чужеродными тяжелыми и легкими цепями приводит к формированию неактивных антигенсвязывающих сайтов и/или других нефункциональных нежелательных побочных продуктов, что представляет собой наибольшую трудность при масштабном производстве и терапевтическом применении фрагментов антител. В частности, невозможно получить конструкции тандемных Fab (Tandem-Fab), в которых два или более Fab объединены друг с другом посредством одного линкера, вследствие произвольной ассоциации двух легких цепей, приводящей к образованию нежелательных неактивных побочных продуктов. Новый формат антител по изобретению позволяет преодолеть эти трудности. В данной заявке описывается новый формат биспецифического антитела, которое можно легко получить с высоким выходом благодаря снижению количества ошибочно спаренных побочных продуктов, и которое демонстрирует пониженную агрегацию по сравнению с фрагментами биспецифических антител, известными в области техники. Применение подхода, основанного на взаимном обмене участками между тяжелой и легкой цепями (кроссовера), способствует правильной ассоциации легких цепей без необходимости создания общей легкой цепи. Подход с использованием общей легкой цепи невозможно применить к существующим антителам. Кроме того, биспецифические антитела нового формата имеют более высокий молекулярную массу по сравнению со многими стандартными фрагментами биспецифических антител, что препятствует избыточному почечному клиренсу и приводит к увеличению времени полужизни in vivo. Биспецифические антитела нового формата являются полностью функциональными и обладают сравнимым или улучшенным связыванием и активностью по сравнению с соответствующими стандартными биспецифическими антителами.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к биспецифическим антителам, содержащим по меньшей мере два Fab фрагмента, где первый Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого антигена, и второй Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении второго антигена, где произведен обмен вариабельными участками или константными участками между легкой и тяжелой цепями второго Fab и где биспецифическое антитело лишено Fc домена. В одном воплощении первый и второй Fab фрагменты соединены при помощи пептидного линкера. Предпочтительно, указанный пептидный линкер представляет собой линкер (G4S)2.
В одном воплощении указанное антитело дополнительно содержит третий Fab фрагмент. В другом воплощении указанный третий Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого или второго антигена, предпочтительно в отношении первого антигена.
В одном воплощении третий Fab фрагмент связан с легкой цепью или тяжелой цепью первого Fab фрагмента. В другом воплощении третий Fab фрагмент связан с N или С-концом легкой цепи или тяжелой цепи второго Fab фрагмента. В одном воплощении третий Fab фрагмент связан с первым или вторым Fab фрагментом посредством пептидного линкера. Предпочтительно, указанный пептидный линкер представляет собой линкер (G4S)2.
Биспецифические антитела по изобретению являются по меньшей мере бивалентными и могут быть тривалентными или мультивалентными, например, тетравалентными. В одном воплощении указанные биспецифические антитела являются бивалентными (формат 1+1) и имеют по одному сайту связывания, направленному против первого и второго антигена, соответственно. В другом воплощении указанные биспецифические антитела являются тривалентными (формат 2+1), с двумя сайтами связывания, каждый из которых направлен против первого антигена, и одним сайтом связывания, направленным против второго антигена, что будет подробно описано в следующем разделе.
Второй объект настоящего изобретения относится к фармацевтической композиции, содержащей биспецифическое антитело по настоящему изобретению.
Третий объект настоящего изобретения относится к биспецифическому антителу по настоящему изобретению для лечения рака. В другом воплощении предложено применение биспецифического антитела в качестве лекарственного препарата. Предпочтительно, указанное применение направлено на лечение рака.
Следующие объекты изобретения относятся к последовательности нуклеиновой кислоты, содержащей последовательность, кодирующую тяжелую цепь- биспецифического антитела по настоящему изобретению, последовательности нуклеиновой кислоты содержащей последовательность, кодирующую легкую цепь биспецифического антитела по настоящему изобретению, экспрессирующему вектору, содержащему последовательность нуклеиновой кислоты по настоящему изобретению, и к прокариотической или эукариотической клетке-хозяину, содержащей вектор по настоящему изобретению. Кроме того, предложен способ получения антитела, включающий культивирование клетки-хозяина с получением антитела.
В следующем воплощении предложен иммуноконъюгат, содержащий биспецифическое антитело по изобретению и цитотоксический агент.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1: Схематическая иллюстрация приведенных в качестве примера форматов биспецифических антител по изобретению, а) молекула Fab-кpoccFab, расположенный на С-конце, b) молекула Fab-кpoccFab, расположенный на N-конце с) молекула (Fab)2-кpoccFab, расположенный на С-конце d) молекула (Fab)2-кpoccFab, расположенный на N-конце е) молекула Fab-кpoccFab-Fab.
Фиг.2: Анализ получения и очистки hu Fab(MCSP)-кpoccFab(CD3): ДСН-ПАГ: 4-12% Бис/Трис (NuPage [invitrogen]; окрашивание Кумасси): а) 1 - маркер Mark 12 (invitrogen), 2 - hu Fab(MCSP)-кpoccFab(CD3) в невосстанавливающих условиях; b) 1 - маркер Mark 12 (invitrogen), 2 - hu Fab(MCSP)-кpoccFab(CD3) в восстанавливающих условиях.
Фиг.3: Анализ получения и очистки Fab(MCSP)-кpoccFab(CD3). Аналитическая эксклюзионная хроматография, хроматограмма А280 (Superdex 200 10/300 GL [GE Healthcare]; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс./об) NaCl; инжектировали 50 мкг образца).
Фиг.4: Анализ получения и очистки hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3): ДСН-ПАГ: 4-12% Бис/Трис (NuPage [invitrogen]; окрашивание Кумасси): а) 1 - маркер Mark 12 (invitrogen), 2 - hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3) в невосстанавливающих условиях; b) 1 - маркер Mark 12 (invitrogen), 2 - hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3) в восстанавливающих условиях.
Фиг.5: Анализ получения и очистки hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3). Аналитическая эксклюзионная хроматография, хроматограмма А280 (Superdex 200 10/300 GL [GE Healthcare]; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс./об) NaCl; инжектировали 50 мкг образца).
Фиг.6: Анализ получения и очистки hu Fab(MCSP)- кpoccFab(CD3) -Fab(MCSP). ДСН-ПАГ: 4-12% Бис/Трис (NuPage [invitrogen]; окрашивание Кумасси): а) 1 - маркер Mark 12 (invitrogen), 2 - hu Fab(MCSP)- кpoccFab(CD3)-Fab(MCSP) в невосстанавливающих условиях; b) 1 - маркер Mark 12 (invitrogen), 2 - hu Fab(MCSP)-кpoccFab(CD3) -Fab(MCSP) в восстанавливающих условиях.
Фиг.7: Анализ получения и очистки hu Fab(MCSP)-кpoccFab(CD3)-Fab(MCSP). Аналитическая эксклюзионная хроматография, хроматограмма А280 (Superdex 200 10/300 GL [GE Healthcare]; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс./об) NaCl; инжектировали 50 мкг образца).
Фиг.8: Анализ получения и очистки кpoccFab мыши (CD3) -Fab(MCSP)-Fab(MCSP). ДСН-ПАГ: 4-12% Бис/Трис (NuPage [invitrogen]; окрашивание Кумасси): а) 1 - маркер Mark 12 (invitrogen), 2 - кpoccFab мыши (CD3)-Fab(MCSP)-Fab(MCSP) в невосстанавливающих условиях; b) 1 - маркер Mark 12 (invitrogen), 2 - кpoccFab мыши (CD3)-Fab(MCSP)-Fab(MCSP) в восстанавливающих условиях.
Фиг.9: Анализ получения и очистки кpoccFab мыши (CD3) -Fab(MCSP)-Fab(MCSP). Аналитическая эксклюзионная хроматография, хроматограмма А280 (Superdex 200 10/300 GL [GE Healthcare]; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс./об) NaCl; инжектировали 50 мкг образца).
Фиг.10: Цитолитическая активность в отношении опухолевых клеток MDA-МВ-435 (оцениваемая по высвобождению ЛДГ) при совместном культивировании с общей популяцией Т-клеток человека (соотношение эффекторные клетки:клетки-мишеней=5:1) и активация в течение 20 часов при помощи различных концентраций hu Fab(MCSP)-кpoccFab(CD3) (="Fab-кpoccFab"), hu Fab(MCSP)-кpoccFab(CD3)-Fab(MCSP) (="Fab-кpoccFab-Fab"), hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3) (="(Fab)2-кpoccFab"), а также биспецифических молекул (scFv)2 (антиМСSР/антиhuCD3e) (="(scFv)2"). Конструкции, имеющие две валентности, направленные против MCSP, демонстрируют цитотоксическую активность сравнимую с конструкцией "(scFv)2", тогда как конструкция "Fab-кpoccFab", имеющая одну валентность связывания для MCSP, явно обладает меньшей эффективностью.
Фиг.11: Сравнение конструкций hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3) (="(Fab)2-кpoccFab") и (scFv)2 (антиМСSР/антиhuCD3e) (="(scFv)2"). Показано высвобождение ЛДГ из опухолевых клеток MDA-MB-435 при совместном культивировании с общей популяцией Т-клеток человека (соотношение эффекторные клетки:клетки-мишени=5:1) и активации в течение 21 часа с использованием биспецифических конструкций и соответствующих IgG в различных концентрациях. Присутствие "(Fab)2-кpoccFab" индуцировало апоптоз клеток-мишеней по меньшей мере так же эффективно, как и присутствие молекул (scFv)2.
Фиг.12: Сравнение конструкций hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3) (="(Fab)2-кpoccFab") и (scFv)2 (антиМСSР/антиhuCD3e) (="(scFv)2"). Показано высвобождение ЛДГ из опухолевых клеток меланомы человека MV-3 при совместном культивировании с человеческими МКПК (соотношение эффекторные клетки:клетки-мишени=10:1) и активации в течение 26 часов с использованием биспецифических конструкций и соответствующих IgG в различных концентрациях. Присутствие "(Fab)2-кpoccFab" индуцировало апоптоз клеток-мишеней по меньшей мере так же эффективно, как и присутствие молекул (scFv)2.
Фиг.13: Высвобождение ЛДГ из опухолевых клеток B16/F10-huMCSP Fluc2, клон 48, индуцированное активацией первичных Т-клеток мыши с использованием конструкции кpoccFab мыши (CD3)-Fab(MCSP)-Fab(MCSP) (=(Fab)2-кpoccFab), направленной против MCSP человека и CD3 мыши. Соотношение эффекторных клеток: клеток-мишеней составляло 5:1. Результаты теста анализировали после инкубации в течение 23,5 часов при 37°С, 5% CO2. Конструкция индуцирует зависимый от концентрации, опосредованный Т-клетками апоптоз клеток-мишеней, экспрессирующих MCSP человека.
Фиг.14: Высвобождение ЛДГ из опухолевых клеток B16/F10-huMCSP Fluc2, клон 48, индуцированное активацией первичных Т-клеток мыши с использованием конструкции кpoccFab мыши (CD3)-Fab(MCSP)-Fab(MCSP) (=(Fab)2-кpoccFab), направленной против MCSP человека и CD3 мыши, в концентрации 50 нМ. Соотношение эффекторных клеток: клеток-мишеней составляло 5:1. Результаты теста анализировали после инкубации в течение 23,5 часов при 37°С, 5% CO2. Конструкция индуцирует опосредованный Т-клетками апоптоз клеток-мишеней, экспрессирующих MCSP человека. При использовании конструкции в такой концентрации происходит только незначительная гиперактивация Т-клеток.
Фиг.15: Концентрации различных цитокинов, измеренные в супернатанте, после инкубации цельной крови с различными биспецифическими конструкциями CD3-MCSP (hu Fab(MCSP)-Fab(MCSP)-кpoccFab(CD3) (="(Fab)2-кpoccFab") и (scFv)2 (aHTHMCSP/антиhuCD3e) (="(scFv)2")) в концентрации 1 нМ в течение 24 часов в присутствии (А, В) или в отсутствие (С, D) опухолевых клеток Colo-38. В каждую лунку 96-луночного планшета помещали по 280 мкл цельной крови и добавляли по 30 000 клеток Colo-38, в соответствии с описанием. Основным цитокином, секретируемым при активации Т-клеток в присутствии опухолевых клеток Colo-38, был IL-6, затем следовал ИФН-гамма. Кроме того, при активации Т-клеток в присутствии клеток-мишеней также значительно повышался уровень гранзима В. В целом, конструкция "(scFv)2" повышала уровни ФНО и ИФН-гамма, а также гранзима В в присутствии клеток-мишеней (А и В) несколько более эффективно по сравнению с другими биспецифическими конструкциями.
При активации Т-клеток биспецифическими конструкциями в присутствии (или в отсутствие) клеток-мишеней не отмечалось выраженной секреции ТП2 цитокинов (IL-10 и IL-4). В этом тесте также наблюдалась незначительная секреция ИФН-гамма, индуцированная конструкцией "(Fab)2-кpoccFab" в отсутствие клеток-мишеней.
Фиг.16: Уровень поверхностной экспрессии позднего маркера активации CD25 в общей популяции Т-клеток мыши, выделенных из спленоцитов. Общую популяцию Т-клеток мыши инкубировали с конструкцией кpoccFab мыши (CD3)-Fab(MCSP)-Fab(MCSP) (=(Fab)2-кpoccFab) (биспецифической конструкцией, направленной против CD3 мыши и MCSP человека) в концентрации 50 нМ в присутствии или в отсутствие опухолевых клеток B16/F10-huMCSP Fluc2 клон 48, в соответствии с описанием (соотношение эффекторных клеток: клеток-мишеней 10:1). Показан уровень экспрессии позднего маркера активации CD25 на поверхности CD8+ Т-клеток через 70 часов. Повышение CD25 на поверхности CD8+ Т-клеток под воздействием конструкции (Fab)2-кpoccFab происходит только в присутствии клеток-мишеней. Референтные IgG, которые использовали в той же молярности, были не способны привести к повышению CD25.
Фиг.17: Анализ получения и очистки Fab(CD33)-кpoccFab (CD3). ДСН-ПАГ: а) 3-8% Трис/Ацетат (NuPage [invitrogen]; окрашивание Кумасси): а) 1 - маркер HiMark (invitrogen), 2 - Fab(CD33)-кpoccFab (CD3) в невосстанавливающих условиях; b) 4-12% Бис/Трис (NuPage [invitrogen]: 1 - маркер Mark 12 (invitrogen), 2 - Fab(CD33)-кpoccFab (CD3) в восстанавливающих условиях.
Фиг.18: Анализ получения и очистки Fab(CD33)-кpoccFab (CD3). Аналитическая эксклюзионная хроматография, хроматограмма А280 (Superdex 200 10/300 GL [GE Healthcare]; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс./об) NaCl; инжектировали 50 мкг образца).
Фиг.19: Цитолитическая активность в отношении опухолевых клеток MV-3 (оцениваемая по высвобождению ЛДГ) при совместном культивировании МКПК человека (соотношение эффекторные клетки: клетки-мишени=10:1) и активации в течение 24 часов с использованием биспецифических конструкций CD3-MCSP (hu Fab(MCSP)-кpoccFab(CD3); обозначенных "1+1 без Fc" и референтной молекулы (scFv)2 (антиМСSР/антиhuCD3e) (="(scFv)2")) в различных концентрациях. Конструкция "1+1 без Fc" индуцирует апоптоз клеток-мишеней MV-3 с рассчитанным значением ЕС50 25,4 пМоль, тогда как рассчитанное значение ЕС50 для референтной молекулы "(scFv)2" составляет 57 пМоль, что показывает чуть большую эффективность молекулы "1+1 без Fc" по показателю ЕС50.
Фиг.20: Активация CD4+ или CD8+Т-клеток, оцениваемая по повышению CD69 (А), соответствующему увеличению количества клеток, положительных по CD69 (В), в присутствии huMCSP-положительных опухолевых клеток MV-3 при совместном культивировании с МКПК человека (соотношение эффекторных клеток: клеток-мишеней=10:1), под воздействием биспецифических конструкций CD3-MCSP (hu Fab(MCSP)-кpoccFab(CD3); обозначенных "1+1 без Fc", и референтной молекулы (scFv)2 (антиМСSР/антиhuCD3e) (="(scFv)2"), соответственно) в течение ~24 часов. В целом, медианные значения CD69 выше у CD8+ Т-клеток по сравнению с CD4+ Т-клетками. Наблюдается четкое зависимое от концентрации повышение медианных значений CD69, а также процентного содержания положительных по CD69 клеток в случае обеих конструкций.
Фиг.21: Иллюстрация референтной молекулы (scFv)2.
Фиг.22: Анализ получения и очистки (scFv)2 (антиМСSР/антиhuCD3e). ДСН-ПАГ: 4-12% Бис/Трис (NuPage [invitrogen]; окрашивание Кумасси): 1 - маркер Mark 12 (invitrogen), 2 - (scFv)2 (антиМСSР/антиhuCD3e) в восстанавливающих условиях; 3 - (scFv)2 (антиМСSР/антиhuCD3e) в невосстанавливающих условиях.
Фиг.23: Анализ получения и очистки (scFv)2 (антиМСSР/антиhuCD3e). Аналитическая эксклюзионная хроматография, хроматограмма А280 (Superdex 75 10/300 GL [GE Healthcare]; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс./об) NaCl; инжектировали 50 мкг образца ((scFv)2 (антиМСSР/антиhuCD3e))).
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
I. Определения
"Каркасные участки" или "FR" (от англ. "framework") обозначают остатки вариабельного домена, отличные от остатков гипервариабельного участка (HVR). FR вариабельного домена, как правило, состоит из 4 FR доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в составе VH (или VL), как правило, располагаются в следующей последовательности: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
"Акцепторные каркасные участки человеческого происхождения" в целях данного описания представляют собой каркасные участки, содержащие аминокислотную последовательность каркасного участка вариабельного домена легкой цепи (VL) или каркасные участки вариабельного домена тяжелой цепи (VH), полученные на основе каркасных участков иммуноглобулина человека или консенсусных каркасных участков человеческого происхождения, определение которым приведено ниже. Акцепторные каркасные последовательности, "полученные на основе" каркасной последовательности иммуноглобулина человека или консенсусных каркасных участков человека, могут содержать ту же аминокислотную последовательность или могут иметь измененную аминокислотную последовательность. В некоторых воплощениях число аминокислотных замен равно 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее, или 2 или менее. В некоторых воплощениях акцепторная последовательность каркасных участков VL идентична последовательности каркасных участков VL иммуноглобулина человека или консенсусной последовательности каркасных участков человеческого происхождения.
"Консенсусные каркасные участки человеческого происхождения" представляют собой каркасные участки, представляющие наиболее часто встречающиеся аминокислотные остатки в наборе последовательностей каркасных участков VL или VH иммуноглобулинов человека. Как правило, последовательности VL или VH иммуноглобулинов человека выбирают из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу согласно описанию Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols.1-3. В одном воплощении, подгруппой для VL является подгруппа каппа I, согласно Kabat et а1.,см выше. В одном воплощении, подгруппой для VH является подгруппа III, согласно Kabat et al., см выше.
Термин "гипервариабельный участок" или "HVR", используемый здесь, обозначает каждый из участков вариабельного домена антитела, имеющих гипервариабельную последовательность и/или формирующих петли определенной структуры ("гипервариабельные петли"). Как правило, нативные антитела, состоящие из четырех цепей, имеют 6 HVR; три в VH (Н1, Н2, Н3), и три в VL (L1, L2, L3). Как правило, HVR содержат аминокислотные остатки гипервариабельных петель и/или "участков определяющих комплементарность" (CDR), последние характеризуются наибольшей вариабельностью последовательности и/или задействованы в распознавании антигена. Приведенные в качестве примера гипервариабельные петли находятся между аминокислотными остатками 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987). Приведенные в качестве примера CDR (CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-Н2 и CDR-H3) находятся между аминокислотными остатками 24-34 в L1, 50-56 в L2, 89-97 в L3, 31-35 В в Н1, 50-65 в Н2 и 95-102 в Н3. (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991).) Термины гипервариабельные участки (HVR) и участки, определяющие комплементарность (CDR), здесь используются взаимозаменяемо для обозначения частей вариабельного участка, формирующих антигенсвязывающие участки. Эти конкретные участки были описаны в публикациях Kabat et al., U.S. Dept. of Health and Human Services, "Sequences of Proteins of Immunological Interest" (1983) и Chothia et al., J. Mol. Biol. 196:901-917 (1987), и при сравнении друг с другом данные определения включают перекрывающиеся группы аминокислотных остатков. Тем не менее, применение любого из определений для обозначения CDR антитела или его вариантов находится в рамках термина, используемого в настоящем документе. Соответствующие аминокислотные участки, содержащие CDR, согласно любой из вышеупомянутых ссылок, для сравнения представлены ниже в Таблице 1. Точные номера аминокислотных остатков, содержащих конкретные CDR, могут варьировать в зависимости от последовательности и размера CDR. Специалисты в данной области могут легко определить по указанной аминокислотной последовательности вариабельного участка антитела, какие аминокислотные остатки содержат конкретный CDR.
Kabat et al. также разработали систему нумерации для последовательностей вариабельных участков, которую можно применить для любого антитела. Любой специалист в данной области может однозначно "пронумеровать по системе Кабат" любую последовательность вариабельного участка, не основываясь на каких-либо экспериментальных данных помимо самой последовательности. В данном документе "нумерация согласно Кабат" относится к системе нумерации, разработанной Кабат и др., U.S. Dept. of Health and Human Services, "Sequence of Proteins of Immunological Interest" (1983). Если не указано иначе, ссылки на номера позиций конкретных аминокислотных остатков в вариабельном участке антитела указаны в соответствии с системой нумерации согласно Кабат.
Как правило, CDR содержат аминокислотные остатки, образующие гипервариабельные петли, за исключением CDR1 в составе VH. CDR также включают "остатки, определяющие специфичность" или "SDR," которые представляют собой остатки, контактирующие с антигеном. SDR находятся в составе участков CDR, обозначаемых "укороченными CDR", или a-CDR (от англ. abbreviated-CDRs). Приведенные в качестве примера a-CDR (a-CDR-L1, a-CDR-L2, a-CDR-L3, a-CDR-H1, a-CDR-H2 и a-CDR-Н3) образованы аминокислотными остатками в положениях 31-34 в L1, 50-55 в L2, 89-96 в L3, 31-35 В в Н1, 50-58 в Н2 и 95-102 в НЗ (см. Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008).) Если не указано иначе, остатки HVR и иные остатки вариабельного домена (например, остатки каркасного участка) здесь пронумерованы в соответствии с Kabat et аl., см. выше.
Термин "антитело" здесь используется в широком смысле и включает антитела с различной структурой, включая моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они проявляют желаемую антигенсвязывающую активность, но не ограничивается ими. В частности, термин "антитело" также включает биспецифические антитела по изобретению, содержащие по меньшей мере два Fab фрагмента, но не имеющие Fc домена.
"Человеческое антитело" представляет собой антитело, имеющее аминокислотную последовательность, которая соответствует последовательности антитела, продуцируемого в организме или в клетках человека или полученного из источника, не относящегося к человеку, но задействующего репертуар антител человека или иные последовательности, кодирующие антитела человека. Данное определение человеческого антитела в частности исключает гуманизированные антитела, содержащие антигенсвязывающие остатки, не являющиеся человеческими.
Термин "рекомбинантное человеческое антитело" в данном документе включает все человеческие антитела, приготовленные, экспрессированные, созданные или выделенные с использованием рекомбинантных методов, такие как антитела, выделенные из клеток-хозяев, таких как клетки NSO или СНО, или из организма животного (например, мыши), трансгенного по генам человеческих иммуноглобулинов, или антитела, экспрессированные с использованием рекомбинантных векторов экспрессии, трансфецированных в клетку-хозяина. Такие рекомбинантные человеческие антитела имеют вариабельный и константный участки в перегруппированном виде. Рекомбинантные человеческие антитела согласно изобретению были подвергнуты соматической гипермутации in vivo. Таким образом, аминокислотные последовательности VH и VL доменов рекомбинантных антител представляют собой последовательности, которые получены на основе и близки последовательностям VH и VL зародышевой линии человека, но не могут существовать естественным образом в зародышевом наборе человеческих антител in vivo.
"Гуманизированное" антитело обозначает химерное антитело, содержащее аминокислотные остатки гипервариабельных участков, не являющихся человеческими, и аминокислотные остатки каркасных участков человеческого происхождения. В определенных воплощениях гуманизированное антитело содержит по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все гипервариабельные участки (например, CDR) соответствуют участкам антитела, не являющегося человеческим, и все или по существу все каркасные участки соответствуют участкам человеческого антитела. Гуманизированное антитело может содержать по меньшей мере часть константного участка антитела, полученного из антитела человека. "Гуманизированная форма" антитела, например, антитела не являющегося человеческим, обозначает антитело, прошедшее гуманизацию. Другие формы "гуманизированных антител", охватываемые настоящим изобретением, представляют собой такие, у которых константный участок был дополнительно модифицирован или изменен по сравнению с константным участком исходного антитела для получения свойств по изобретению, в частности, относящихся к связыванию C1q и/или связыванию Fc рецептора (FcR).
Термин "химерное" антитело обозначает антитело, в котором часть тяжелой и/или легкой цепи получена из конкретных источников или биологических видов, тогда как оставшаяся часть тяжелой и/или легкой цепи получена из другого источника или биологического вида, как правило полученное с использованием технологий рекомбинантной ДНК. Предпочтительными являются химерные антитела, содержащие мышиный вариабельный участок и человеческий константный участок. Другими предпочтительными формами "химерных антител", охватываемыми настоящим изобретением, являются те, у которых константный участок был модифицирован или изменен по сравнению с исходным антителом для получения свойств по изобретению, в частности, относящихся к связыванию C1q и/или связыванию Fc рецептора (FcR). Такие химерные антитела также обозначаются "антитела с переключенным классом". Химерные антитела являются продуктом экспрессии генов иммуноглобулинов, содержащих сегменты ДНК, кодирующие вариабельные участки иммуноглобулинов, и сегменты ДНК, кодирующие константные участки иммуноглобулинов. Способы получения химерных антител включают стандартные технологии рекомбинантной ДНК и трансфекции генов, хорошо известные в области техники. См., например, Morrison, S.L, et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855; патенты США US 5,202,238 и 5,204,244.
Термин "моноклональное антитело" в данном документе используется для обозначения антитела, полученного из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, идентичны и/или связывают один эпитоп, за исключением возможных вариантов антител, например, содержащих мутации, которые возникают естественным путем или в ходе получения препарата моноклонального антитела, при этом подобные варианты, как правило, присутствуют в минорных количествах. В противоположность препаратам поликлональных антител, обычно содержащим различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклональных антител направлено против единственной детерминанты или антигена. Таким образом, определение "моноклональное" указывает на характер антитела, полученного из по существу гомогенной популяции антител, и не должно рассматриваться как необходимость получения антитела определенным способом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, могут быть произведены с помощью различных технологий, включая гибридомную технологию, технологию рекомбинантной ДНК, технологию фагового дисплея и технологии с использованием трансгенных животных, имеющих все локусы человеческих иммуноглобулинов или их часть, но не ограничиваясь ими; данные способы и другие приведенные в качестве примера способы производства моноклональных антител описаны в данном документе.
"Фрагмент антитела" обозначает молекулу, отличную от интактного антитела, содержащую часть интактного антитела, связывающую антиген, с которым связывается интактное антитело. Примеры фрагментов антитела включают Fv, Fab, Fab', Fab'-SH, F(ab')2, диатела, линейные антитела, молекулы одноцепочечных антител (например, scFv) и мультиспецифические антитела, образованные из фрагментов антител, но не ограничиваются ими. Антитела scFv описаны, например, Houston, J.S., Methods in Enzymol. 203 (1991) 46-96). Кроме того, фрагменты антител содержат одноцепочечные полипептиды, обладающие характеристиками VH домена, а именно способные объединяться вместе с VL доменом, или характеристиками VL домена, а именно способные объединяться вместе с VH доменом с образованием функционального антигенсвязывающего сайта и таким образом обеспечивающие антигенсвязывающие свойства полноразмерных антител.
В данном документе "Fab фрагмент" обозначает фрагмент антитела, содержащий фрагмент легкой цепи, содержащий VL домен и константный домен легкой цепи (CL), а также VH домен и первый константный домен (СН1) тяжелой цепи. Биспецифические антитела по изобретению содержат по меньшей мере два Fab фрагмента, причем произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента. Вследствие обмена вариабельными участками или константными участками указанный второй Fab фрагмент также обозначается "кpoccFab (crossFab) фрагмент" или "xFab фрагмент" или "кроссовер-Fab (crossover Fab) фрагмент". Возможны два различных варианта компоновки молекулы кроссовер-Fab в составе биспецифических антител по изобретению: с одной стороны, может происходить обмен вариабельными участками между тяжелой и легкой цепями Fab, т.е. молекула кроссовер-Fab содержит пептидную цепь, состоящую из вариабельного участка легкой цепи (VL) и константного участка тяжелой цепи (СН1), и пептидную цепь, состоящую из вариабельного участка тяжелой цепи (VH) и константного участка легкой цепи (CL). Такая молекула кроссовер-Fab также обозначается кpoccFab (vlvh). С другой стороны, когда происходит обмен константными участками между тяжелой и легкой цепями Fab, молекула кроссовер-Fab содержит пептидную цепь, состоящую из вариабельного участка тяжелой цепи (VH) и константного участка легкой цепи (CL), и пептидную цепь, состоящую из вариабельного участка легкой цепи (VL) и константного участка тяжелой цепи (СН1). Такая молекула кроссовер-Fab также обозначается кpoccFab(CLCH1).
В одном воплощении указанные Fab фрагменты соединены при помощи пептидного линкера. "Соединены" означает, что Fab фрагменты связаны посредством пептидных связей, либо напрямую, либо при помощи одного или более пептидных линкеров.
Термин "пептидный линкер", используемый в изобретении, обозначает пептид с аминокислотной последовательностью, которая предпочтительно имеет искусственное происхождение. Эти пептидные линкеры по изобретению используются для соединения одного из Fab фрагментов с С- или N-концом другого Fab фрагмента с образованием мультиспецифического антитела по изобретению. Предпочтительно, указанные пептидные линкеры являются пептидами с аминокислотной последовательностью длиной по меньшей мере 5 аминокислот, предпочтительно длиной от 5 до 100, более предпочтительно от 10 до 50 аминокислот. В одном воплощении указанный пептидный линкер представляет собой (GxS)n или (GxS)nGm, где G=глицин, S=серин, а (х=3, n=3, 4, 5 или 6, и m=0, 1, 2 или 3) или (х=4, n=2, 3, 4 или 5, и m=0, 1, 2 или 3), предпочтительно х=4 и n=2 или 3, более предпочтительно х=4, n=2. Кроме того, линкеры могут содержать (часть) шарнирный(ого) участок(ка) иммуноглобулина. В одном воплощении указанный пептидный линкер представляет собой (G4S)2 (SEQ ID: NO 28). Другими пептидными линкерами, подходящими для соединения Fab фрагментов, являются, например, (G4S)6-GG (SEQ ID NO:147) или (SG3)2-(SEG3)4-(SG3)-SC (SEQ ID NO:148) или EPKSC(D)-(G4S)2 (SEQ ID NO 145 и 146).
Термин "антигенсвязывающий домен" обозначает часть антигенсвязывающей молекулы, содержащую область, которая специфически связывается и является комплементарной целому антигену или его части. В случаях, когда антиген имеет большие размеры, антигенсвязывающая молекула может связывать только отдельную часть антигена, которая называется эпитопом. Антигенсвязывающий домен может быть образован, например, одним или несколькими вариабельными доменами антитела (также называемыми вариабельными участками антитела). Предпочтительно, антигенсвязывающий домен содержит вариабельный участок легкой цепи антитела (VL) и вариабельный участок тяжелой цепи антитела (VH).
Термин "вариабельный участок" или "вариабельный домен" обозначает домен легкой или тяжелой цепи антитела, задействованный в связывании антитела с антигеном. Вариабельные домены тяжелой и легкой цепей нативного антитела (VH и VL, соответственно), как правило, имеют схожую структуру, каждый домен содержит четыре консервативных каркасных участка (FR) и три гипервариабельных участка (HVR) (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007).) Для обеспечения специфичности связывания антигена может быть достаточно одного VH или VL домена. Кроме того, антитела, связывающиеся с конкретным антигеном, можно выделить с использованием VH или VL домена антитела, связывающегося с антигеном, при скрининге библиотеки комплементарных доменов VL или VH, соответственно. См., например,, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
Термин "антигенсвязывающий сайт антитела" в данном документе обозначает аминокислотные остатки антитела, отвечающие за связывание антигена. Антигенсвязывающая часть антитела содержит аминокислотные остатки "участков, определяющих комплементарность" или "CDR". "Каркасные" или "FR" участки представляют собой участки вариабельных доменов, отличные от остатков "участков, определяющих комплементарность", согласно данному описанию. Таким образом, вариабельные домены легкой и тяжелой цепи антитела содержат в направлении от N- к С-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. В частности, CDR3 тяжелой цепи представляет собой участок, который вносит наибольший вклад в связывание антигена и определяет свойства антитела. Участки CDR и FR и/или остатки "гипервариабельной петли" определяются согласно стандартному описанию Kabat et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991).
Термин "эпитоп" включает любые полипептидные детерминанты, способные специфически связываться с антителом. В некоторых воплощениях эпитопные детерминанты включают химически активные поверхностные группировки молекул, такие как аминокислоты, углеводные боковые цепи, фосфорил или сульфонил, и, в некоторых воплощениях, могут иметь определенные трехмерные структурные характеристики и/или определенные характеристики заряда. Эпитоп представляет собой участок антигена, который связывается с антителом.
Термин "Fc домен" здесь обозначает С-концевой участок тяжелой цепи иммуноглобулина, содержащий по меньшей мере часть константного участка. Например, у антител естественного происхождения Fc домен состоит из двух одинаковых белковых фрагментов, происходящих из второго и третьего константных доменов двух тяжелых цепей антител с изотипом IgG, IgA и IgD; Fc домены IgM и IgE содержат три константных домена тяжелых цепей (CH домены 2-4) в составе каждой полипептидной цепи. Биспецифические антитела по изобретению лишены Fc домена. "Лишен Fc домена" здесь означает, что биспецифические антитела по изобретению не содержат СН2, СНЗ или СН4 домена; т.е. константный участок тяжелой цепи состоит только из одного или более СН1 доменов.
"Аффинность" обозначает суммарную силу всех нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иначе, термин "аффинность связывания" в данном документе обозначает собственную аффинность связывания, которая отражает взаимодействие между членами связывающейся пары (например, антителом и антигеном) в соотношении 1:1. Аффинность молекулы Х к своему партнеру Y обычно выражают через константу диссоциации (KD). Аффинность может быть измерена общепринятыми способами, известными в данной области техники, включая описанные в данном документе. Определенные иллюстративные и приведенные в качестве примеров воплощения для измерения аффинности связывания описаны ниже.
В данном документе термин "связывание" или "специфическое связывание" означает, что связывание антигена имеет избирательный характер и отличается от нежелательного или неспецифического взаимодействия. Способность антигенсвязывающих компонентов связывать определенные антигенные детерминанты можно измерить либо при помощи иммуноферментного анализа (ИФА) или других методов, известных специалистам в данной области, например, метода поверхностного плазменного резонанса (SPR) (анализируется при помощи биосенсора BIAcore) (Liljeblad et al., Glyco J 17, 323-329 (2000)) и традиционных тестов связывания (Heeley, Endocr Res 28, 217-229 (2002)). В одном воплощении степень связывания антигенсвязывающего компонента с посторонним белком составляет менее чем приблизительно 10% от связывания антигенсвязывающего компонента с антигеном, что измеряется, например, при помощи SPR. В некоторых воплощениях антигенсвязывающий компонент, который связывается с антигеном, или антигенсвязывающая молекула, содержащая этот антигенсвязывающий компонент, имеют константу диссоциации (KD) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
Антитело "подвергнутое аффинному созреванию" обозначает антитело с одной или несколькими модификациями в одном или нескольких гипервариабельных участках (HVR), по сравнению с материнским антителом, не имеющим таких модификаций, такие модификации приводят к повышению аффинности антитела к антигену.
В одном воплощении степень связывания биспецифического антитела, которое специфически связывается с первым антигеном и вторым антигеном, с посторонним белком составляет менее 10% от связывания антитела с первым или вторым антигеном, что измеряется, например, с помощью радиоиммуннологического анализа (РИА) или проточной цитометрии (FACS). В некоторых воплощениях биспецифическое антитело, специфически связывающееся с первым антигеном и вторым антигеном, имеет константу диссоциации (KD) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ, или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В некоторых воплощениях биспецифическое антитело, специфически связывающееся с первым антигеном и вторым антигеном, связывается с эпитопом первого антигена или второго антигена, который является консервативным у первого или второго антигена, происходящего из различных биологических видов.
Специфичность антитела обозначает избирательное распознавание антителом конкретного эпитопа антигена. Антитела природного происхождения, например, являются моноспецифическими. "Биспецифические антитела" по изобретению представляют собой антитела, у которых два антигенсвязывающих сайта обладают различной специфичностью. Антитела по настоящему изобретению являются специфическими в отношении двух различных антигенов, т.е. в отношении первого антигена и второго антигена.
Термин "моноспецифическое" антитело в данном документе обозначает антитело, которое имеет один или более сайтов связывания, каждый из которых связывается с тем же эпитопом того же антигена.
Термин "биспецифическое" антитело здесь обозначает антитело, которое имеет по меньшей мере два сайта связывания, каждый из которых связывается с различными эпитопами того же антигена или различных антигенов.
Предложенное здесь антитело представляет собой мультиспецифическое антитело, например биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, способные специфически связываться по меньшей мере с двумя различными сайтами. Здесь предложено биспецифическое антитело, способное специфически связываться с первым антигеном и вторым антигеном. В некоторых воплощениях биспецифические антитела могут связываться с двумя различными эпитопами первого антигена или второго антигена.
Биспецифические антитела также можно применять для сосредоточения цитотоксических агентов в клетках, экспрессирующих первый антиген или второй антиген.
Термин "валентность" в данном описании обозначает наличие определенного числа сайтов связывания в молекуле антитела. Так, термины "бивалентное", "тетравалентное" и "гексавалентное" обозначают наличие в молекуле антитела двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания, соответственно. Биспецифические антитела по изобретению являются по меньшей мере "бивалентными" и могут быть "тривалентными" или "мультивалентными" (например, "тетравалентными" или "гексавалентными").
Антитела по данному изобретению имеют два или более сайтов связывания и являются биспецифическими. Это означает, что антитела могут быть биспецифическими даже в случаях, когда существует более двух сайтов связывания (т.е. когда антитело является тривалентным или мультивалентным).
"Антитело, связывающееся с тем же эпитопом", что и референтное антитело, обозначает антитело, блокирующее связывание референтного антитела с его антигеном в тестах конкурентного связывания на 50% или более, и наоборот, референтное антитело блокирует связывание антитела с его антигеном в тестах конкурентного связывания на 50% или более. Примеры тестов конкурентного связывания приводятся ниже.
"Отсутствие существенной перекрестной реактивности" означает, что молекула (например, антитело) не распознает или не связывает специфически антиген, отличный от истинного антигена-мишени молекулы (например, антиген близкородственный антигену-мишени), в частности, при сравнении с этим антигеном-мишенью. Например, антитело может связывать от менее чем приблизительно 10% до менее чем приблизительно 5% антигена, отличного от истинного антигена-мишени, или может связывать указанный антиген, отличный от истинного антигена-мишени в количестве, выбранном из группы, состоящей из менее чем приблизительно 10%, 9%, 8% 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,5%, 0,2% или 0,1%, предпочтительно менее чем приблизительно 2%, 1% или 0,5%, и наиболее предпочтительно менее чем приблизительно 0,2% или 0,1% антигена, отличного от истинного антигена-мишени.
"Процент (%) идентичности аминокислотной последовательности" относительно референтной полипептидной последовательности определяется как процент аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в референтной полипептидной последовательности после выравнивания последовательностей и расстановки пропусков (гэпов), если это необходимо для достижения наибольшего процента идентичности последовательностей, и без принятия каких-либо консервативных замен за часть идентичной последовательности. Выравнивание в целях определения процента идентичности аминокислотной последовательности может выполняться различными способами, известными в данной области техники, например, с помощью общедоступных компьютерных программ, таких как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить надлежащие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Для целей настоящего описания, значения % идентичности аминокислотной последовательности получены при помощи компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана в компании Genentech, Inc., исходный код был представлен вместе с пользовательской документацией в бюро по охране авторских прав США, Washington D.C., 20559, где было зарегистрировано авторское право под номером TXU510087. Доступ к программе ALIGN-2 можно получить в компании Genentech, Inc., South San Francisco, California, или код программы может быть скомпилирован на основе исходного кода. Программа ALIGN-2 должна быть скомпилирована для работы на платформе UNIX, включая платформу digital UNIX V4.0D. Все параметры для сравнения последовательностей заданы программой ALIGN-2 и остаются неизменными.
В случаях, когда для сравнения аминокислотных последовательностей используется ALIGN-2, процент идентичности заданной аминокислотной последовательности А к последовательности В, с последовательностью В, или по сравнению с заданной аминокислотной последовательностью В (что можно также сформулировать как заданная аминокислотная последовательность А, имеющая или содержащая определенный процент идентичной аминокислотной последовательности к последовательности В, с последовательностью В, или по сравнению с заданной аминокислотной последовательностью В) рассчитывается по формуле:
X/Y×100,
где Х - это число аминокислотных остатков, которые были расценены программой для выравнивания последовательностей ALIGN-2 как абсолютные совпадения при выравнивании А и В в рамках этой программы, и где Y - это общее число аминокислотных остатков в В. Следует принимать во внимание, что когда длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, процент идентичности аминокислотной последовательности А с последовательностью В не будет равен проценту идентичности аминокислотной последовательности В с последовательностью А. Если не указано иначе, все значения % идентичности аминокислотных последовательностей, приведенные здесь, получены согласно описанию в предыдущем параграфе о применении компьютерной программы ALIGN-2.
"Выделенное" антитело обозначает антитело, изолированное от компонентов его естественного окружения. В некоторых воплощениях антитело очищено до степени чистоты более чем 95% или 99% согласно определению, например, электрофоретическими (например, с помощью ДСН-ПАГ, изоэлектрического фокусирования (IEF), капиллярного электрофореза) или хроматографическими методами (например, с помощью ионообменной или обратнофазной ВЭЖХ). Для обзора способов оценки чистоты антител см., например, Flatman et al., J. Chromatogr. В 848:79-87 (2007).
"Выделенная" нуклеиновая кислота обозначает молекулу нуклеиновой кислоты, изолированную от компонентов ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, как правило содержащих молекулу нуклеиновой кислоты, при этом молекула нуклеиновой кислоты находится внехромосомно или в участке хромосомы, отличном от естественного положения на хромосоме.
"Выделенная нуклеиновая кислота, кодирующая биспецифическое антитело по изобретению" обозначает одну или несколько молекул нуклеиновой кислоты, кодирующих тяжелые и легкие цепи антитела (или их фрагменты), включая такую(ие) молекулу(ы) нуклеиновой кислоты в одном векторе или в разных векторах, при этом такая(ие) молекула(ы) нуклеиновой кислоты находи(я)тся в одном или нескольких местоположениях в клетке-хозяине.
Термин "аминокислота" в данном описании обозначает группу карбокси а-амино кислот естественного происхождения, включающую аланин (трехбуквенный код: ala, однобуквенный код: А), аргинин (arg, R), аспарагин (asn, N), аспарагиновую кислоту (asp, D), цистеин (cys, С), глутамин (gln, Q), глутаминовую кислоту (glu, E), глицин (gly, G), гистидин (his, H), изолейцин (ile, I), лейцин (leu, L), лизин (lys, K), метионин (met, М), фенилаланин (phe, F), пролин (pro, Р), серин (ser, S), треонин (thr, Т), триптофан (trp, W), тирозин (tyr, Y) и валин (val, V).
Термин "вектор", используемый в данном документе, относится к молекуле нуклеиновой кислоты, способной передавать другую нуклеиновую кислоту, с которой она связана. Термин включает вектор как самореплицирующуюся структуру нуклеиновой кислоты, а также вектор, инкорпорированный в состав генома клетки-хозяина, в которую он был введен. Некоторые векторы способны направлять экспрессию нуклеиновых кислот, с которыми они функционально связаны. Такие векторы здесь обозначаются "экспрессирующие векторы".
В данном документе выражения "клетка", "клеточная линия" и "клеточная культура" используются взаимозаменяемо и все эти обозначения включают потомство клеток. Так, термины "трансфектанты" и "трансфецированные клетки" включают первичные исследуемые клетки, а также происходящие от них культуры, вне зависимости от количества пассажей. Следует понимать, что все потомство может не быть абсолютно идентичным по содержанию ДНК, вследствие неслучайных или случайных мутаций. Термины также охватывают варианты потомства, обладающие такой же функцией или биологической активностью, которая была определена при скрининге у исходно трансформированной клетки.
Термины "клетка-хозяин", "линия клеток-хозяев" и "культура клеток-хозяев" используются взаимозаменяемо и обозначают клетки, в которые была внедрена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают "трансформанты" и "трансформированные клетки", включающие первично трансформированные клетки и их потомков, независимо от числа пассажей. Потомство может быть не полностью идентично родительской клетке по содержанию нуклеиновой кислоты, но может иметь мутации. Сюда также включаются мутантные потомки, обладающие такими же функциями или биологической активностью при скрининге или отборе, как и исходные трансформированные клетки.
"Голое антитело" обозначает антитело, не конъюгированное с гетерологичным компонентом (например, цитотоксическим компонентом) или радиоактивной меткой. Голое антитело может присутствовать в фармацевтическом препарате.
"Иммуноконъюгат" представляет собой антитело, конъюгированное с одной или более гетерологичной(ыми) молекулой(ами), включая цитотоксический агент, но не ограничиваясь им.
Термин "цитотоксический агент" здесь используется для обозначения вещества, ингибирующего или предотвращающего осуществление клеточной функции и/или вызывающего гибель или разрушение клетки. Цитотоксические агенты включают радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты, ингибирующие рост; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или энзиматически активные токсины бактериального, грибкового, растительного или животного происхождения, включая фрагменты и/или их варианты; и различные противоопухолевые или противораковые агенты, описанные ниже, но не ограничиваются ими.
Термин "N-конец" обозначает последнюю аминокислоту N-конца, термин "С-конец" обозначает последнюю аминокислоту С-конца.
Термин "фармацевтический препарат" относится к препарату, который находится в такой форме, которая обеспечивает эффективное проявление биологической активности содержащегося в нем активного ингредиента, и не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому будет вводиться препарат.
"Фармацевтически приемлемый носитель" относится к ингредиенту в фармацевтическом препарате, отличному от активного ингредиента, который является нетоксичным для субъекта. Фармацевтически приемлемый носитель включает, буфер, эксципиент, стабилизатор или консервант, но не ограничивается ими.
Используемый в данном документе термин "лечение" (и его грамматические производные, такие как "лечить" или "проводить лечение") обозначает клиническое вмешательство в попытке изменить естественный ход заболевания у индивида, получающего лечение, и могут осуществляться как для профилактики, так и при наличии патологического состояния. Желательные эффекты от лечения включают предупреждение возникновения или повторного проявления заболевания, смягчение симптомов, уменьшение любых прямых или непрямых патологических последствий заболевания, предупреждение метастазирования, снижение скорости прогрессирования заболевания, улучшение или временное облегчение состояния, а также ремиссию или улучшение прогноза, но не ограничиваются ими. В некоторых воплощениях антитела по изобретению применяют, чтобы отсрочить развитие заболевания или чтобы замедлить прогрессирование заболевания.
"Индивид" или "субъект" является млекопитающим. Млекопитающие включают одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и не относящихся к человеку приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс), но не ограничиваются ими. В определенных воплощениях индивидом или субъектом является человек.
"Эффективное количество" агента, например фармацевтического препарата, обозначает количество, эффективное для достижения желаемого терапевтического или профилактического результата в необходимых дозировках и в течение необходимого периода времени.
Термин "рак" в данном документе обозначает пролиферативное заболевание, такое как лимфомы, лимфоцитарные лейкозы, рак легких, немелкоклеточный рак легких (NSCL), бронхоальвеолярный рак легкого, рак костей, рак поджелудочной железы, рак кожи, раковые образования головы или шеи, меланома кожи или внутриглазная меланома, рак матки, рак яичников, рак прямой кишки, рак анальной зоны, рак желудка, рак ЖКТ, рак толстого кишечника, рак молочной железы, рак матки, карцинома фаллопиевых труб, карцинома эндометрия, карцинома шейки матки, карцинома влагалища, карцинома вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, опухоли эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркома мягких тканей, рак уретры, рак полового члена, рак предстательной железы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточная карцинома, карцинома почечной лоханки, мезотелиома, гепатоцеллюлярный рак, рак желчного пузыря, новообразования центральной нервной системы (ЦНС), опухоли позвоночника, глиома ствола мозга, мультиформная глиобластома, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, плоскокпеточные карциномы, аденома гипофиза и саркома Юинга, включая рефракторные формы любого из видов рака, указанных выше, или сочетание одного или более видов рака, указанных выше.
Термин "листовка-вкладыш" используется для обозначения инструкций, обычно вкладываемых в коммерческую упаковку терапевтических продуктов, содержащих информацию о показаниях к применению, способах применения, дозировках, введении, комбинированной терапии, противопоказаниях и/или предупреждениях, касающихся применения таких терапевтических продуктов.
II. Композиции и способы
А. Приведенные в качестве примера биспецифические антитела
Настоящее изобретение относится к биспецифическим антителам, содержащим по меньшей мере два Fab фрагмента, где первый Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого антигена; и второй Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении второго антигена, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента и где биспецифическое антитело лишено Fc домена. В одном воплощении первый и второй Fab фрагменты соединены посредством пептидного линкера. Предпочтительно указанный пептидный линкер представляет собой пептид с аминокислотной последовательностью длиной по меньшей мере 5 аминокислот, предпочтительно длиной от 5 до 100, более предпочтительно от 10 до 50 аминокислот. В одном воплощении указанный пептидный линкер представляет собой (GxS)n или (GxS)nGm, где G=глицин, S=серин, а (х=3, n=3, 4, 5 или 6, и m=0, 1, 2 или 3) или (х=4, n=2, 3, 4 или 5, и m=0, 1, 2 или 3). Предпочтительно х=4 и n=2 или 3, более предпочтительно х=4, n=2. В одном воплощении указанный пептидный линкер представляет собой (G4S)2. Пептидный линкер используется для соединения первого и второго Fab фрагментов. В одном воплощении первый Fab фрагмент соединен с С- или N-концом второго Fab фрагмента.
В одном воплощении первый Fab фрагмент соединен с N-концом второго Fab фрагмента. В зависимости от того, произведен ли обмен вариабельными или константными доменами между тяжелой и легкой цепями второго Fab фрагмента, возможны различные варианты молекул биспецифического антитела, где первый Fab фрагмент соединен с N-концом второго Fab фрагмента.
В одном воплощении произведен обмен вариабельными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(VHVL)), а С-конец тяжелой или легкой цепи первого Fab фрагмента соединен с N-концом цепи VLCH1 второго Fab фрагмента. Предпочтительно, С-конец тяжелой цепи первого Fab фрагмента соединен с N-концом цепи VLCH1 второго Fab фрагмента. Таким образом, в одном воплощении биспецифическое антитело содержит три цепи: легкую цепь (VLCL) первого Fab фрагмента, тяжелую цепь первого Fab фрагмента, соединенную с цепью VLCH1 цепью второго Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VLCH1) и цепь VHCL второго Fab фрагмента.
В другом воплощении произведен обмен константными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(CLCH1)), а С-конец тяжелой или легкой цепи первого Fab фрагмента соединен с N-концом цепи VHCL второго Fab фрагмента. Предпочтительно, С-конец тяжелой цепи первого Fab фрагмента соединен с N-концом цепи VHCL второго Fab фрагмента. Таким образом, в одном воплощении биспецифическое антитело содержит три цепи: легкую цепь (VLCL) первого Fab фрагмента, тяжелую цепь первого Fab фрагмента, соединенную с цепью VHCL второго Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VHCL) и цепь VLCH1 второго Fab фрагмента.
В одном воплощении первый Fab фрагмент соединен с С-концом второго Fab фрагмента. В зависимости от того, произведен ли обмен вариабельными или константными доменами между тяжелой и легкой цепями второго Fab фрагмента, возможны различные варианты молекул биспецифического антитела, где первый Fab фрагмент соединен с С-концом второго Fab фрагмента.
В одном воплощении произведен обмен вариабельными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(VHVL)), a CH1 домен второго Fab фрагмента соединен с N-концом тяжелой или легкой цепи первого Fab фрагмента. Предпочтительно, CH1 домен второго Fab фрагмента соединен с N-концом тяжелой цепи первого Fab фрагмента. Таким образом, в одном воплощении биспецифическое антитело содержит три цепи: легкую цепь (VLCL) первого Fab фрагмента, цепь VLCH1 второго Fab фрагмента, соединенную с тяжелой цепью первого Fab фрагмента посредством пептидного линкера (VLCH1-линкер-VHCH1) и цепь VHCL второго Fab фрагмента.
В другом воплощении произведен обмен константными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(CLCH1)), а CL домен второго Fab фрагмента соединен с N-концом тяжелой или легкой цепи первого Fab фрагмента. Предпочтительно, CL домен второго Fab фрагмента соединен с N-концом тяжелой цепи первого Fab фрагмента. Таким образом, в одном воплощении биспецифическое антитело содержит три цепи: легкую цепь (VLCL) первого Fab фрагмента, цепь VHCL второго Fab фрагмента, соединенную с тяжелой цепью первого Fab фрагмента посредством пептидного линкера (VLCH1-линкер-VHCH1) и цепь VLCH1 второго Fab фрагмента.
Биспецифические антитела по изобретению являются по меньшей мере бивалентнымии и могут быть тривалентными или мультивалентными, например, тетравалентными. В одном воплощении указанные биспецифические антитела являются бивалентными (формат 1+1), у которых каждый из связывающихся сайтов направлен против первого антигена и второго антигена, соответственно. В другом воплощении указанные биспецифические антитела являются тривалентными (формат 2+1) с двумя связывающимися сайтами, направленными против первого антигена и одним связывающимся сайтом, направленным против второго антигена, как подробно описано в следующем разделе.
В одном воплощении указанное антитело дополнительно содержит третий Fab фрагмент. В другом воплощении указанный третий Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого или второго антигена, предпочтительно в отношении первого антигена.
В одном воплощении третий Fab фрагмент соединен с N- или С-концом первого Fab фрагмента. В одном воплощении третий Fab фрагмент соединен с первым Fab фрагментом посредством пептидного линкера. Предпочтительно, указанный пептидный линкер представляет собой линкер (G4S)2.
В одном воплощении третий Fab фрагмент соединен с N- или С-концом легкой цепи или тяжелой цепи первого Fab фрагмента. В зависимости от того, какой конец первого Fab фрагмента соединен со вторым Fab фрагментом (как подробно описано выше), третий Fab фрагмент соединен в противоположным (свободным) концом первого фрагмента.
В одном воплощении, биспецифическое антитело по изобретению содержит три Fab фрагмента, где указанные Fab фрагменты и указанный линкер соединены в следующем порядке в направлении от N-конца к С-концу: Fab фрагмент 3 - линкер - Fab фрагмент 1 - линкер - Fab фрагмент 2, где произведен обмен вариабельными участками или константными участками между тяжелой или легкой цепями второго Fab фрагмента. В этом воплощении С-конец третьего Fab фрагмента соединен с N-концом первого Fab фрагмента. Как подробно описано выше, Fab фрагменты могут быть соединены друг с другом через тяжелую или легкую цепи. В одном воплощении С-конец тяжелой цепи третьего Fab фрагмента соединен с N-концом тяжелой цепи первого Fab фрагмента посредством пептидного линкера; а С-конец первого Fab фрагмента соединен с N-концом второго Fab фрагмента, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента. В зависимости от того, произведен ли обмен вариабельными или константными доменами между тяжелой и легкой цепями второго Fab фрагмента, возможны различные варианты молекул биспецифического антитела.
В одном воплощении произведен обмен вариабельными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(VHVL)), а цепи трех Fab фрагментов соединены в следующем порядке в направлении от N-конца к С-концу: VHCH1-пинкер-VHCH1-линкер-VLCH1. В одном воплощении биспецифическое антитело содержит четыре цепи: легкую цепь (VLCL) третьего Fab фрагмента, легкую цепь (VLCL) первого Fab фрагмента, тяжелую цепь третьего фрагмента, соединенную с тяжелой цепью первого Fab фрагмента, которая в свою очередь соединена с цепью VLCH1 второго Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VHCH1-линкер-VLCH1), и цепь VHCL второго Fab фрагмента.
В одном воплощении произведен обмен константными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(CLCH1)), а цепи трех Fab фрагментов соединены в следующем порядке в направлении от N-конца к С-концу: VHCH1-пинкер-VHCH1-линкер-VHCL. В одном воплощении биспецифическое антитело содержит четыре цепи: легкую цепь (VLCL) третьего Fab фрагмента, легкую цепь (VLCL) первого Fab фрагмента, тяжелую цепь третьего фрагмента, соединенную с тяжелой цепью первого Fab фрагмента, которая в свою очередь соединена с цепью VHCL второго Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VHCH1-линкер-VHCL) и VLCH1 цепь второго Fab фрагмента.
В одном воплощении биспецифическое антитело по изобретению содержит три Fab фрагмента, где указанные Fab фрагменты и указанный линкер соединены в следующем порядке в направлении от N-конца к С-концу: Fab фрагмент 2 - линкер - Fab фрагмент 1 - линкер - Fab фрагмент 3, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента. В этом воплощении N-конец третьего Fab фрагмента соединен с С-концом первого Fab фрагмента. Как подробно описано выше, Fab фрагменты могут быть соединены друг с другом через тяжелую или легкую цепи. В одном воплощении N-конец тяжелой цепи третьего Fab фрагмента соединен с С-концом тяжели цепи первого Fab фрагмента посредством пептидного линкера; а N-конец первого Fab фрагмента соединен с С-концом второго Fab фрагмента, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента. В зависимости от того, произведен ли обмен вариабельными или константными доменами между тяжелой и легкой цепями второго Fab фрагмента, возможны различные варианты молекул биспецифического антитела.
В одном воплощении произведен обмен вариабельными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(VHVL)), а цепи трех Fab фрагментов соединены в следующем порядке в направлении от N-конца к С-концу: VLCH1-линкер-VHCH1-линкер-VHCH1. В одном воплощении биспецифическое антитело содержит четыре цепи: легкую цепь (VLCL) третьего Fab фрагмента, легкую цепь (VLCL) первого Fab фрагмента, цепь VLCH1 второго Fab фрагмента, соединенную с тяжелой цепью первого фрагмента, которая в свою очередь соединена с тяжелой цепью первого Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VHCH1-линкер-VHCH1) и цепь VHCL второго Fab фрагмента.
В одном воплощении произведен обмен константными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(CLCH1)), а цепи трех Fab фрагментов соединены в следующем порядке в направлении от N-конца к С-концу: VHCL-линкер-VHCH1-линкер-VHCH1. В одном воплощении биспецифическое антитело содержит четыре цепи: легкую цепь (VLCL) третьего Fab фрагмента, легкую цепь (VLCL) первого Fab фрагмента, цепь VHCL второго Fab фрагмента, соединенную с тяжелой цепью первого фрагмента, которая в свою очередь соединена с тяжелой цепью первого Fab фрагмента посредством пептидного линкера (VHCL-линкер-VHCH1-линкер-VHCH1) и цепь VLCH1 второго Fab фрагмента.
В другом воплощении третий Fab фрагмент соединен с N- или С-концом легкой цепи или тяжелой цепи второго Fab фрагмента. В одном воплощении третий Fab фрагмент соединен со вторым Fab фрагментом посредством пептидного линкера. Предпочтительно, указанный пептидный линкер представляет собой линкер (G4S)2. Как подробно описано выше, Fab фрагменты могут быть соединены друг с другом через тяжелую или легкую цепи.
В одном воплощении биспецифическое антитело по изобретению содержит три Fab фрагмента, где указанные Fab фрагменты и указанный линкер соединены в следующем порядке в направлении от N-конца к С-концу: Fab фрагмент 1 - линкер - Fab фрагмент 2 - линкер - Fab фрагмент 3, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента. В одном воплощении N-конец третьего Fab фрагмента соединен с С-концом второго Fab фрагмента.
В другом воплощении С-конец тяжелой цепи третьего Fab фрагмента соединен с N-концом второго Fab фрагмента посредством пептидного линкера; а N-конец первого Fab фрагмента соединен с С-концом второго Fab фрагмента, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента.
В зависимости от того, произведен ли обмен вариабельными или константными доменами между тяжелой и легкой цепями второго Fab фрагмента, возможны различные варианты молекул биспецифического антитела.
В одном воплощении произведен обмен вариабельными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(VHVL)), а цепи трех Fab фрагментов соединены в следующем порядке в направлении от N-конца к С-концу: VHCH1-линкер-VLCH1-линкер-VHCH1. В одном воплощении биспецифическое антитело содержит четыре цепи: легкую цепь (VLCL) третьего Fab фрагмента, легкую цепь (VLCL) первого Fab фрагмента, тяжелую цепь третьего фрагмента, соединенную с N-концом цепи VLCH1 второго Fab фрагмента, причем С-конец указанной цепи VLCH1 соединен с N-концом тяжелой цепи первого Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VLCH1-линкер-VHCH1), и цепь VHCL второго Fab фрагмента.
В одном воплощении произведен обмен константными доменами второго Fab фрагмента (т.е. второй Fab фрагмент представляет собой кpoccFab(CLCH1)), а цепи трех Fab фрагментов соединены в следующем порядке в направлении от N-конца к С-концу: VHCH1-линкер-VHCL-линкер-VHCH1. В одном воплощении биспецифическое антитело содержит четыре цепи: легкую цепь (VLCL) третьего Fab фрагмента, легкую цепь (VLCL) первого Fab фрагмента, тяжелую цепь третьего фрагмента, соединенную с N-концом цепи VHCL второго Fab фрагмента, причем С-конец указанной цепи VHCL соединен с N-концом тяжелой цепи первого Fab фрагмента посредством пептидного линкера (VHCH1-линкер-VHCL-линкер-VHCH1), и цепь VLCH1 второго Fab фрагмента.
В одном воплощении изобретения биспецифическое антитело представляет собой гуманизированное антитело, как подробно описано ниже.
В другом воплощении изобретения биспецифическое антитело представляет собой человеческое антитело, как подробно описано ниже.
Второй объект настоящего изобретения относится к фармацевтической композиции, содержащей биспецифическое антитело по настоящему изобретению.
Третий объект настоящего изобретения относится к биспецифическому антителу по настоящему изобретению для лечения рака. В другом воплощении предложено применение биспецифического антитела в качестве лекарственного препарата. Предпочтительно, указанное применение направлено на лечение рака.
Следующие объекты настоящего изобретения относятся к последовательности нуклеиновой кислоты, содержащей последовательность, кодирующую тяжелую цепь биспецифического антитела по настоящему изобретению, последовательности нуклеиновой кислоты, содержащей последовательность, кодирующую легкую цепь биспецифического антитела по настоящему изобретению, экспрессирующему вектору, содержащему последовательность нуклеиновой кислоты по настоящему изобретению, и к прокариотической или эукариотической клетке-хозяину, содержащей вектор по настоящему изобретению. Кроме того, предложен способ получения антитела, включающий культивирование клетки-хозяина с получением антитела.
В следующем воплощении предложен иммуноконъюгат, содержащий биспецифическое антитело по изобретению и цитотоксический агент.
В следующем аспекте, биспецифическое антитело по любому из указанных выше воплощений может обладать любыми признаками, описанными ниже в разделах 1-5, как в отдельности, так и в комбинации:
1. Аффинность антитела
Аффинность предложенного в данном документе биспецифического антитела к целевому антигену можно определить согласно способам, описанным в Примерах, с помощью поверхностного плазменного резонанса (SPR), с применением стандартного оборудования, такого как прибор BIAcore (GE Healthcare), а также рецепторов или белков-мишеней, которые могут быть получены путем рекомбинантной экспрессии. В альтернативном случае, связывание биспецифических антител с различными рецепторами или антигенами-мишенями можно оценивать с применением клеточных линий, экспрессирующих конкретный рецептор или антиген-мишень, например при помощи проточной цитометрии (FACS).
В некоторых воплощениях предложенное здесь биспецифическое имеет константу диссоциации (KD) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 M, например, от 10-9 М до 10-13 М).
Согласно одному воплощению, KD измеряют при помощи поверхностного плазмонного резонанса с применением BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С с иммобилизацией антигена на поверхности чипа СМ5 до уровня связывания ~10 единиц RU. Вкратце, карбоксиметилированные декстрановые чипы биосенсора (СМ5, BIACORE, Inc.) активировали с помощью N-этил-N'-(3-диметиламинопропил)-карбодиимида гидрохлорида (EDC) и N-гидроксисукцинимида (NHS) согласно инструкциям поставщика. Антиген разводили раствором 10 мМ ацетата натрия, рН 4,8, до концентрации 5 мкг/мл (~0,2 мкМ) перед инжектированием со скоростью 5 мкл/мин до достижения уровня связывания белка приблизительно 10 единиц RU. После инжектирования антигена инжектировали 1 М этаноламин для блокирования непрореагировавших групп. Для определения кинетики инжектировали Fab, последовательно разведенные в два раза (от 0,78 нМ до 500 нМ) в PBS, содержащем 0,05% сурфактант полисорбат 20 (TWEEN-20™) (PBST) при 25°С со скоростью приблизительно 25 мкл/мин. Скорости ассоциации (kd или kon) и скорости диссоциации (kd или koff) рассчитывали с использованием простой модели связывания один к одному, предложенной Ленгмюром (при помощи программного обеспечения (BIACORE ® Evaluation Software версии 3.2) при одновременной аппроксимации сенсограмм ассоциации и диссоциации. Константу равновесной диссоциации (KD) рассчитывали как отношение koff/kon См., например,, Chen et al., J. Mol. Biol. 293:865-881 (1999). Если скорость ассоциации превышала 106 М-1 с-1 при измерении методом поверхностного плазменного резонанса, описанным выше, скорость ассоциации определяли с использованием метода гашения флуоресценции, позволяющего определить увеличение или уменьшение интенсивности испускаемой флуоресценции (возбуждение=295 нм; эмиссия=340 нм, полоса пропускания 16 нм) 20 нМ антитела, направленного против антигена, (в форме Fab) в PBS, рН 7,2, при 25°С в присутствии возрастающих концентраций антигена, которую измеряли с помощью спектрофотометра, такого как спектрофотометра с функцией остановки потока (Aviv Instruments) или спектрофотометра SLM-AMINCO™ серии 8000 (ThermoSpectronic) с функцией перемешивания в кювете.
2. Химерные и гуманизированные антитела
В некоторых воплощениях предложенное здесь биспецифическое антитело представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в патенте США US 4,816,567 и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). В одном примере химерное антитело содержит вариабельный участок, не являющийся человеческим (например, вариабельный участок, происходящий из организма мыши, крысы, хомяка, кролика или не являющегося человеком примата, например, обезьяны), и константный участок человеческого происхождения. В следующем примере химерное антитело представляет собой антитело "с переключенным классом", у которого класс или подкласс были изменены по сравнению с родительским антителом. Химерные антитела включают также их антигенсвязывающие фрагменты.
В некоторых воплощениях химерное антитело представляет собой гуманизированное антитело. Как правило, антитело, не являющееся человеческим, подвергают гуманизации, чтобы снизить иммуногенность для человека, при этом сохраняя специфичность и аффинность родительского антитела, не являющегося человеческим. Как правило, гуманизированное антитело содержит один или более вариабельных доменов, в которых HVR, например, CDR, (или их части) получены из антитела, не являющегося человеческим, а каркасные участки FR (или их части) получены на основе последовательностей антитела человека. Возможно гуманизированное антитело также содержит по меньшей мере часть константного участка человеческого антитела. В некоторых воплощениях некоторые остатки FR в гуманизированном антителе заменены соответствующими остатками антитела, не являющегося человеческим (например, антитела из которого получены аминокислотные остатки HVR), например, для восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения представлены, например, в обзоре Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008) и более подробно описаны, например, Riechmann et al.. Nature 332:323-329 (1988); Queen et al., Proc. Nat'l Acad. Sci. USA 86:10029-10033 (1989); патентах США US 5, 821,337, 7,527,791, 6,982,321, и 7,087,409; Kashmiri et al., Methods 36:25-34 (2005) (описание пересадки SDR (a-CDR)); Padlan, Mol. Immunol. 28:489-498 (1991) (описание "изменения поверхности"); Dall'Acqua et al., Methods 36:43-60 (2005) (описание "перегруппировки FR") и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260 (2000) (описание "направленного отбора" для перегруппировки FR).
Каркасные участки человека, которые могут быть использованы для гуманизации, включают: каркасные участки, выбранные при помощи метода "максимального соответствия" (см., например, Sims et al. J. Immunol. 151:2296 (1993)); каркасные участки, полученные из консенсусных последовательностей человеческих антител с вариабельными участками легкой или тяжелой цепей определенной подгруппы (см., например, Carter et al. Proc. Natl. Acad. Sci. USA, 89:4285 (1992) и Presta et al. J. Immunol., 151:2623 (1993)); зрелые каркасные участки человека (с соматическими мутациями) или каркасные участки зародышевой линии (неперестроенные) (см., например, Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008)) и каркасные участки, полученные при скрининге библиотек FR (см., например, Васа et al., J. Biol. Chem. 272:10678-10684 (1997) and Rosok et al., J. Biol. Chem. 271:22611-22618 (1996)).
3. Человеческие антитела
В некоторых воплощениях предложенное в данном документе биспецифическое антитело представляет собой человеческое антитело. Человеческие антитела могут быть получены при помощи различных способов, известных в данной области техники. Человеческие антитела описаны van Dijk and van de Winkel, Curr. Opin. Pharmacol. 5: 368-74 (2001) и Lonberg, Curr. Opin. Immunol. 20:450-459 (2008).
Человеческие антитела могут быть получены путем введения иммуногена трансгенному животному, модифицированному для получения интактных человеческих антител или интактных антител с вариабельными участками человека в ответ на введение антигена. Такие животные обычно имеют все или часть локусов иммуноглобулинов человека, заменяющие локусы эндогенных иммуноглобулинов, находящиеся вне хромосом или случайным образом интегрированные в хромосомы животных. У таких трансгенных мышей локусы эндогенных иммуноглобулинов обычно бывают инактивированы. Обзор способов получения человеческих антител в трансгенных животных представлен Lonberg, Nat. Biotech. 23:1117-1125 (2005). См. также, например, патенты США US 6,075,181 и 6,150,584 описывающие технологию XENOMOUSE™; патент США US 5,770,429 описывающий технологию HuМАВ®; патент США US 7,041,870 описывающий технологию K-М MOUSE®, и заявку на патент США US 2007/0061900, описывающую технологию velocimouse®). Вариабельные участки человеческого происхождения в интактных антителах, полученных в таких животных, могут быть далее модифицированы, например, путем комбинации с различными константными участками человеческого происхождения.
Антитела человека могут также быть получены при помощи гибридомной технологии. Описаны клеточные линии миеломы человека и гетеромиеломы мышь-человек для продукции моноклональных человеческих антител (см.,, например,, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal AHmubody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987) и Boerner et al., J. Immunol., 147: 86 (1991).) Также описаны человеческие антитела, полученные с применением гибридомной технологии с использованием В-клеток человека Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006). Дополнительные способы включают те, что описаны, например, в патенте США US 7,189,826 (описание получения моноклональных антител человека класса IgM из гибридомных клеточных линий) и Ni, Xiandai Mianyixue, 26(4):265-268 (2006) (описание гибридом человек-человек). Технология человеческих гибридом (технология триом) также описана Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005) and Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3): 185-91 (2005).
Человеческие антитела могут быть получены путем выделения последовательностей вариабельных доменов Fv клонов, отобранных из библиотек фагового дисплея, созданных на основе генов человека. Такие последовательности вариабельных доменов затем могут быть объединены с необходимым константным доменом человека. Технологии для отбора человеческих антител из библиотек антител описаны ниже.
4. Антитела, полученные из библиотек
Биспецифические антитела по изобретению могут быть выделены путем скрининга комбинаторных библиотек на антитела с желаемой активностью или активностями. Например, в данной области техники известно большое количество способов создания библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие желаемыми характеристиками связывания. Такие способы рассматриваются, например, в обзоре Hoogenboom et al. in Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001) и также описываются, например, McCafferty et al., Nature 348:552-554; Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Marks and Bradbury, in Methods in Molecular Biology 248:161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004) и Lee et al., J. Immunol. Methods 284(1-2): 119-132(2004).
В некоторых способах фагового дисплея спектры генов VH и VL клонируют отдельно при помощи полимеразной цепной реакции (PCR) и рекомбинируют случайным образом в библиотеках фагового дисплея, и затем проводят их скрининг на связывание фаговой частицы с антигеном, согласно описанию Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). Как правило, фаги экспонируют фрагменты антител либо в виде одноцепочечных Fv (scFv) фрагментов, либо в виде Fab фрагментов. Библиотеки на основе генов, полученных из иммунизированных доноров, позволяют получить высокоаффинные антитела к иммуногену без необходимости создания гибридом. В альтернативном случае, для получения единого источника антител к широкому спектру "чужих" и "своих" антигенов можно клонировать репертуар генов первичных антител (например, человеческих), согласно описанию Griffiths et al., EMBO J, 12: 725-734 (1993). Наконец, "наивные" или первичные библиотеки также могут быть синтезированы путем клонирования неперестроенных сегментов, включающих V-гены стволовых клеток, и использования праймеров PCR, содержащих случайные последовательности, которые кодируют высоковариабельные участки CDR3 и обеспечивают перестройку in vitro, согласно описанию Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992). Публикации, описывающие фаговые библиотеки человеческих антител, включают, например, патент США US 5,750,373 и публикации US 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, здесь считаются человеческими антителами или фрагментами человеческих антител.
5. Варианты антител
В некоторых воплощениях рассматриваются варианты аминокислотных последовательностей предложенных в данном документе биспецифических антител. Например, может требоваться улучшение аффинности связывания и/или других биологических свойств биспецифического антитела. Варианты аминокислотных последовательностей биспецифического антитела могут быть получены путем введения соответствующих модификаций в нуклеотидную последовательность, кодирующую биспецифическое антитело, или путем пептидного синтеза. Такие модификации включают, например, делеции и/или инсерции и/или замены остатков в составе аминокислотной последовательности антитела. Для получения конечной конструкции можно осуществлять любые комбинации делеций, инсерций и замен, при условии, что конечная конструкция будет обладать желаемыми характеристиками, например, связыванием антигена.
А) Варианты с заменами, инсерциями и делециями
В некоторых воплощениях предложены варианты антитела, имеющие одну или более аминокислотных замен. Сайты, представляющие интерес для проведения замещающего мутагенеза включают HVR и FR. Консервативные замены представлены в Таблице 1 под заголовком "консервативные замены". Более значительные замены представлены в Таблице 2 под заголовком "примеры замен", и подробнее описаны ниже, со ссылкой на классы боковых цепей аминокислот. Аминокислотные замены можно вводить в антитело, представляющее интерес, и проводить скрининг полученных продуктов на предмет желаемой активности, например, сохранения/улучшения связывания антигена или понижения иммуногенности.
Аминокислотные остатки можно сгруппировать в соответствии общими свойствами боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены влекут за собой замещение представителя одного из этих классов представителем другого класса.
Один вид замен включает замену одного или нескольких остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный(е) вариант(ы), отбираемые для дальнейшего изучения, будут иметь модификации (например, улучшение) определенных биологических свойств (например, повышенную аффинность, пониженную иммуногенность) по отношению к родительскому антителу и/или в значительной степени сохраняют определенные биологические свойства родительского антитела. Примером варианта с заменами является антитело, подвергнутое аффинному созреванию, которое может быть легко получено, например, при помощи технологии увеличения аффинности с использованием фагового дисплея, подробно описанной здесь. Вкратце, один или более остатков HVR подвергают мутации и проводят скрининг вариантов антитела, экспрессируемых фагом, по определенной биологической активности (например, аффинности связывания).
Изменения (например, замены) могут осуществляться внутри HVR, например, для увеличения аффинности антитела. Такие изменения можно проводить в "горячих точках" HVR, т.е. в остатках, кодируемых кодонами, которые в процессе соматического созревания с высокой частотой подвергаются мутациям (см., например, Chowdhury, Methods Mol. Biol. 207:179-196 (2008)), и/или SDR (a-CDR), а у полученных вариантов VH или VL исследовать аффинность связывания. Повышение аффинности путем конструирования и повторного отбора из вторичных библиотек было описано, например, Hoogenboom et al. in Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, (2001).) В некоторых воплощениях для повышения аффинности повышают изменчивость вариабельных генов, выбранных для повышения аффинности при помощи любого из существующих разнообразных методов (например, ПЦР пониженной точности, перестройки цепей или олигонуклеотид-направленного мутагенеза). После этого создается вторичная библиотека. Затем проводят скрининг библиотеки для выявления любых вариантов антител с необходимой аффинностью. Другой способ повышения изменчивости включает методы, нацеленные на изменение HVR, при которых рандомизируют несколько остатков HVR (например, 4-6 остатков одновременно). HVR остатки, задействованные в связывании антигена, могут быть специально идентифицированы, например, при помощи аланин-сканирующего мутагенеза или моделирования. В частности, изменениям часто подвергаются CDR-H3 и CDR-L3.
В некоторых воплощениях замены, инсерции или делеции могут возникать в составе одного или нескольких HVR, при условии, что такие изменения существенным образом не снижают способность антитела связывать антиген. Например, могут быть выполнены консервативные замены в HVR (например, консервативные замены, описанные здесь), которые существенным образом не снижают аффинность связывания. Такие замены могут располагаться за пределами "горячих точек" HVR или SDR. В некоторых воплощениях, каждый HVR либо остается без изменений, либо содержит не более одной, двух или трех аминокислотных замен в составе вариантов последовательностей VH и VL, приведенных выше.
Полезный способ определения остатков или участков антитела, которые могут быть мишенями направленного мутагенеза, называется "аланин-сканирующий мутагенез", описанный Cunningham and Wells (1989) Science, 244:1081-1085. В этом способе выявляют аминокислотный остаток или группу таргетных остатков (например, заряженные остатки, такие как Arg, Asp, His, Lys и Glu) и замещают нейтральным или отрицательно заряженными аминокислотами (например, аланином или полиаланином), чтобы выяснить, изменится ли взаимодействие антитела с антигеном. Если при первоначальной замене аминокислот происходят изменения функциональных свойств молекулы, в этих позициях можно проводить дальнейшие замены. Дополнительно или альтернативно, может оказаться полезным провести анализ кристаллической структуры комплекса антиген-антитело для выявления точек контакта между антителом и антигеном. Такие остатки, задействованные в контактах, а также соседние с ними остатки могут быть выбраны в качестве кандидатов для замен. Можно проводить скрининг вариантов с целью выявления у них наличия желаемых свойств.
Инсерции в аминокислотных последовательностях включают сшивки в амино- и/или карбокси-терминальных участках, составляющие по длине от одного остатка до полипептидов, содержащих сотни или более остатков, а также инсерции внутри последовательности одного или нескольких аминокислотных остатков. Примеры инсерции в терминальных участках включают антитело с N-концевым остатком метионила. Другие варианты молекулы антитела с инсерциями включают сшивание N- или С-конца антитела с ферментом (например, ADEPT) или полипептидом, приводящее к повышению времени полужизни антитела в сыворотке.
Б) Модифицированные цистеином варианты антител
В некоторых воплощениях, может быть целесообразным создание модифицированных цистеином биспецифических антител, например, "thioMAbs," в которых один или несколько остатков биспецифического антитела заменены остатками цистеина. В конкретных воплощениях замененные остатки располагаются в доступных сайтах биспецифического антитела. При замене этих остатков на цистеин реакционно-способные тиоловые группы размещаются в доступных сайтах антитела и могут использоваться для конъюгирования антитела с другими компонентами, например, компонентами лекарств или фрагментами линкер-лекарство, для создания конъюгата антитела, согласно описанию ниже. В некоторых воплощениях один или несколько из следующих остатков могут быть заменены на цистеин: V205 легкой цепи (нумерация по Кабат) и АН 8 тяжелой цепи (нумерация EU). Модифицированные цистеином антитела могут быть получены, как описано, например, в патенте США US 7,521,541.
В) Производные антител
В некоторых воплощениях предложенное в данном документе биспецифическое антитело может быть также модифицировано с целью введения дополнительных небелковых компонентов, известных в данной области техники и легко доступных. Компоненты, подходящие для получения производных биспецифического антитела, включают водорастворимые полимеры, но не ограничиваются ими. Не являющиеся исчерпывающими примеры водорастворимых полимеров включают полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля и пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена и малеинового ангидрида, полиаминокислоты (как гомополимеры, так и случайные сополимеры) и декстран или поли(n-винил пирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры пролипропиленоксида и этилен оксида, полиоксиэтилированные полиолы (например, глицерол), поливиниловый спирт и их смеси, но не ограничиваются ими. Пропиональдегид полиэтиленгликоля может иметь преимущества при производстве благодаря своей стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к антителу, может варьировать и в случае присоединения более одного полимера, их молекулы могут быть идентичными или различаться. Как правило, число и/или тип полимеров, применяемых для получения производных, может определяться исходя из того, какие конкретные свойства или функции антитела, нуждаются в улучшении, и того, будет ли производное антитела применяться в терапии при определенных условиях, и т.д., но не ограничиваться данными соображениями.
В другом воплощении предложены конъюгаты биспецифического антитела и небелковые компоненты, которые могут быть избирательно нагреты под воздействием излучения. В одном воплощении, небелковый компонент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Излучение может иметь любую длину волны, включая, но не ограничиваясь длинами волн, не повреждающими обычные клетки, но нагревающими небелковый компонент до температуры, которая вызывает гибель клеток, расположенных вблизи небелкового компонента антитела.
Б. Рекомбинантные способы и композиции
Биспецифические антитела по изобретению могут быть получены, например, при помощи твердофазного синтеза пептидов (например, твердофазного синтеза, разработанного Меррифилдом) или получены рекомбинантным способом. Для получения рекомбинантным способом один или более полинуклеотидов, кодирующих биспецифическое антитело (фрагмент), например, согласно описаннию выше, выделяют и встраивают в один или более векторов для дальнейшего клонирования и/или экспрессии в клетках-хозяевах. Такой полинуклеотид можно легко выделить и секвенировать при помощи стандартных процедур. В одном воплощении предложен вектор, предпочтительно экспрессирующий вектор, содержащий один или более полинуклеотидов по изобретению. Для конструирования векторов экспрессии, содержащих кодирующую последовательность (фрагмента) биспецифического антитела вместе с соответствующими сигнальными последовательностями, контролирующими транскрипцию/трансляцию, могут применяться способы, хорошо известные специалистам в данной области. Эти способы включают in vitro технологию рекомбинантных ДНК, методы синтеза и рекомбинацию in vivo/генетическую рекомбинацию. См., например, методы, описанные Maniatis et al., molecular cloning: A laboratory manual, Cold Spring Harbor Laboratory, N.Y. (1989) и Ausubel et al., current protocols in molecular biology, Greene Publishing Associates and Wiley Interscience, N.Y (1989). Экспрессирующий вектор может быть частью плазмиды, вируса или может быть фрагментом нуклеиновой кислоты. Экспрессирующий вектор включает экспрессионную кассету, в которую клонирован полинуклеотид, кодирующий (фрагмент) биспецифическое(го) антитело(а) (т.е. кодирующая область), функционально связанный с промотором и/или другим элементом, контролирующим транскрипцию или трансляцию. В данном документе "кодирующая область" представляет собой часть нуклеиновой кислоты, которая состоит из кодонов, транслируемых в аминокислоты. Хотя "стоп-кодон" (TAG, TGA или ТАА) не транслируется в аминокислоту, его можно считать частью кодирующей области, однако любые фланкирующие последовательности, например, промоторы, сайты рибосом, терминаторы транскрипции, интроны, 5' и 3' нетранслируемые области и т.п. не являются частью кодирующей области. Две или более кодирующей области могут находиться в составе одной полинуклеотидной конструкции, например, в одном векторе, или в отдельных полинуклеотидных конструкциях, например, в отдельных (различных) векторах. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующие области, например, вектор по настоящему изобретению может кодировать один или более полипептидов, которые разделяются пост- или котрансляционно посредством протеолитического расщепления с образованием конечных белков. Кроме того, вектор, полинуклеотид или нуклеиновая кислота по изобретению могут кодировать гетерологичные кодирующие области, объединенные или не объединенные с полинуклеотидом, кодирующим (фрагмент) биспецифическое(го) антитело(а) по изобретению, или его вариант или производное. Гетерологичные кодирующие области включают специализированные элементы или мотивы, такие как сигнальная последовательность секретируемого пептида или гетерологичный функциональный домен, но не исчерпываются ими. Функциональная связь имеет место, когда область, кодирующая продукт гена, например, полипептид, связана с одной или более регуляторными последовательностями таким образом, что экспрессия продукта гена оказывается под влиянием или контролем регуляторной(ых) последовательности(ей). Два фрагмента ДНК (такие как кодирующая область полипептида и связанный с ней промотор) "функционально связаны", если индукция промотора приводит к транскрипции мРНК, кодирующей искомый продукт гена и если природа связи между двумя фрагментами ДНК не влияет на способность последовательностей, регулирующих экспрессию, направлять экспрессию продукта гена или не влияет на способность ДНК-матрицы транскрибироваться. Таким образом, область промотора будет функционально связана с нуклеиновой кислотой, кодирующей полипептид, если промотор способен осуществлять транскрипцию этой нуклеиновой кислоты. Промотор может быть специфическим для клетки промотором, направляющим транскрипцию ДНК по существу только в определенных клетках. Другие элементы, контролирующие транскрипцию, помимо промотора, например энхансеры, операторы, репрессоры и сигналы терминации транскрипции, могут быть функционально связаны с полинуклеотидом и направлять специфическую для клеток транскрипцию. Здесь описаны подходящие промоторы и другие области, контролирующие транскрипцию. Специалистам в данной области техники известно множество областей, контролирующих транскрипцию. Они включают, но не исчерпываются областями, контролирующими транскрипцию, которые функционируют в клетках позвоночных, такие как сегменты промоторов и энхансеров цитомегаловирусов (например, предранний промотор вместе с интроном-А), вируса обезьян sv 40 (например, ранний промотор) и ретровирусов (например, таких как вирус саркомы Рауса), но не ограничиваются ими. Другие области, контролирующие транскрипцию, включают области генов позвоночных, например, генов актина, белка теплового шока, бычьего гормона роста и кроличьего â-глобина, а также другие последовательности, способные контролировать экспрессию генов в эукариотических клетках. Дополнительные подходящие области, контролирующие транскрипцию, включают тканеспецифические промоторы и энхансеры, а также индуцибельные промоторы (например, тетрациклин-индуцибельные промоторы). Аналогично, специалистам в данной области техники известно множество элементов, контролирующих трансляцию. Они включают сайты связывания рибосом, кодоны инициации и терминации трансляции, а также элементы, полученные из вирусных систем (в частности, внутренний сайт связывания рибосом, или IRES, также обозначаемый последовательностью CITE (САР-независимый энхансер транскрипции)), но не ограничиваются ими. Экспрессионная кассета может также иметь другие признаки, такие как ориджины репликации, и/или элементы, обеспечивающие интеграцию в хромосомы, такие как длинные концевые повторы ретровирусов (LTR) или инвертированные концевые повторы (ITR) адено-ассоциированных вирусов (AAV).
Кодирующие области полинуклеотида и нуклеиновой кислоты по настоящему изобретению могут быть связаны с дополнительными кодирующими областями, которые кодируют обеспечивающие секрецию или сигнальные пептиды, направляющие секрецию полипептида, кодируемого полинуклеотидом по настоящему изобретению. Например, если необходима секреция биспецифической антигенсвязывающей молекулы, ДНК, кодирующую последовательность сигнального пептида, можно поместить в противоположном экспрессии направлении относительно нуклеиновой кислоты, кодирующей биспецифическое антитело по изобретению или его фрагмент. Согласно гипотезе о сигнальных пептидах, белки, секретируемые клетками млекопитающих, имеют сигнальный пептид или лидерную последовательность, которые отщепляются от зрелого белка после начала секреции растущей пептидной цепочки через шероховатый эндоплазматический ретикулум. Специалистам в данной области техники известно, что полипептиды, секретируемые клетками позвоночных, обычно имеют сигнальный пептид, объединенный с N-концом полипептида, который отщепляется от транслированного полипептида с получением секретируемой или "зрелой" формы полипептида. В некоторых воплощениях используется нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина или функциональное производное этой последовательности, которое сохраняет способность направлять секрецию полипептида, функционально связанного с ней. В альтернативном случае, можно использовать гетерологичный сигнальный пептид млекопитающих или его функциональное производное. Например, лидерную последовательность дикого типа можно заменить лидерной последовательностью человеческого тканевого активатора плазминогена (тАП) или мышиной β-глюкуронидазы.
ДНК, кодирующая короткую белковую последовательность, которую можно использовать для облегчения дальнейшей очистки (например, гистидиновый эпитоп) или способствования введению метки в биспецифическое антитело, может находиться внутри или на концах полинуклеотида, кодирующего (фрагмент) биспецифическое(го) антитело(а).
В следующем воплощении предложена клетка-хозяин, содержащая один или более полинуклеотидов по изобретению. В некоторых воплощениях предложена клетка-хозяин, содержащая один или более векторов по изобретению. Полинуклеотиды и векторы могут иметь любые признаки, указанные здесь в отношении полинуклеотидов и векторов, соответственно, как отдельно, так и в комбинации. В одном из таких воплощений клетка-хозяин содержит (например, была трансформирована или трансфецирована) вектор, содержащий полинуклеотид, который кодирует (часть) биспецифическое(го) антитело(а) по изобретению. В данном документе термин "клетка-хозяин" обозначает любой тип клеточной системы, которая может быть модифицирована для получения биспецифического антитела по изобретению или его фрагментов. Клетки-хозяева, подходящие для репликации и для поддержания экспрессии биспецифических антител, хорошо известны в области техники. Такие клетки можно трансфецировать или трансдуцировать соответствующим образом конкретными экспрессирующими векторами, и содержащие вектор клетки можно выращивать в большом количестве для засевания ферментеров для крупномасштабного производства биспецифического антитела в количестве, достаточном для клинического применения. Подходящие клетки-хозяева включают прокариотические микроорганизмы, такие как Е. coli, или различные эукариотические клетки, такие как клетки яичника китайского хомячка (СНО), клетки насекомых и т.п. Например, полипептиды можно продуцировать в бактериях, особенно когда не требуется их гликозилирование. После экспрессии полипептид может быть выделен из бактериальной массы в виде растворимой фракции и затем очищен. Кроме прокариот в качестве хозяев для клонирования или экспрессии векторов, кодирующих полипептиды, подходят микроорганизмы-эукариоты, такие как мицелиальные грибы или дрожжи, включая штаммы грибов и дрожжей, у которых пути гликозилирования были "гуманизированы" для обеспечения продукции полипептида с частично или полностью человеческим профилем гликозилирования. См. Gerngross, Nat Biotech 22, 1409-1414 (2004), and Li et al., Nat Biotech 24, 210-215 (2006). Клетки-хозяева, подходящие для экспрессии (гликозилированных) полипептидов также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Также были найдены многочисленные штаммы бакуловирусов, которые можно использовать совместно с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda. В качестве хозяев также можно использовать культуры клеток растений. См., например, патенты США US 5,959,177, 6,040,498, 6,420,548, 7,125,978 и 6,417,429 (описывающие технологию PLAHTHBODIES™ для получения антител в трансгенных растениях). Клетки позвоночных также могут использоваться в качестве хозяев. Например, можно использовать клеточные линии млекопитающих, адаптированные для роста в суспензии. Другими примерами клеточных линий млекопитающих, используемых в качестве хозяев, являются линия клеток почки обезьяны CV1, трансформированная SV40 (COS-7); линия человеческих эмбриональных клеток почки (клетки 293 или 293Т, согласно описанию, например, Graham et al., J. Gen Virol. 36:59 (1977)); клетки почки новорожденного хомяка (ВНК); мышиные клетки сертоли (клетки ТМ4, согласно описанию, например Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени крысы буффало (BRL 3А); клетки легких человека (W138); клетки печени человека (Нер G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI (согласно описанию, например Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; и клетки FS4. Другие клеточные линии млекопитающих, используемые в качестве хозяев, включают клетки яичников китайского хомячка (СНО), включая клетки dhfr- CHO cells (Uriaub et al., Proc Natl Acad Sci USA 77, 4216 (1980)) и линии клеток миеломы, такие как YO, NSO, P3X63 и Sp2/0. Обзор некоторых клеточных линий млекопитающих, подходящих в качестве хозяев для получения антител, представлен, например, Yazaki and Wu, Methods in Molecular Biology, Vol.248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp.255-268 (2003). Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, клетки дрожжей, клетки насекомых, бактериальные клетки и клетки растений, чтобы перечислить только некоторые из них, а также клетки, содержащиеся у трансгенного животного, трансгенного растения или в культуре тканей растения или животного. В одном воплощении клетка-хозяин представляет собой эукариотическую клетку, предпочтительно клетку млекопитающего, такую как клетка яичника китайского хомячка (СНО), человеческая эмбриональная клетка почки (НЕК) или лимфоидная клетка (например, клетка Y0, NS0, Sp20).
Стандартные методики, известные в области техники, позволяют экспрессировать чужеродные гены в этих системах. Клетки, экспрессирующие полипептид, содержащий тяжелую или легкую цепь антигенсвязывающего домена, такой как антитело, могут быть модифицированы с использованием генной инженерии таким образом, чтобы экспрессировать также другие цепи антитела, чтобы экспрессируемый продукт представлял собой антитело, имеющее и тяжелые, и легкие цепи.
В одном воплощении предложен способ получения биспецифического антитела по изобретению, где способ включает культивирование клетки-хозяина, содержащей полинуклеотид, кодирующий биспецифическое антитело, описанное здесь, при условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы, и выделение биспецифического антитела из клетки-хозяина (или среды, в которой культивируют клетку-хозяина).
Компоненты биспецифического антитела объединены друг с другом при помощи генетических методов. Биспецифическое антитело можно сконструировать таким образом, чтобы его компоненты были объединены друг с другом напрямую или опосредованно при помощи линкерной последовательности. Состав и длину линкера можно определить согласно способам, хорошо известным в области техники, и можно исследовать их эффективность. Примеры линкерных последовательностей, соединяющих различные компоненты биспецифических антител, можно найти среди последовательностей, представленных в данном документе. Также можно включать дополнительные последовательности, для введения сайта расщепления при необходимости разделения отдельных компонентов гибридного белка, например последовательность, распознаваемую эндопептидазой.
В некоторых воплощениях один или более антигенсвязывающих компонентов биспецифического антитела содержит по меньшей мере вариабельный участок антитела, способный связывать антигенные детерминанты. Вариабельные участки могут входить в состав антител естественного или искусственного происхождения и их фрагментов и быть получены из них. Способы продукции поликлональных антител и моноклональных антител хорошо известны в области техники (см., например, Harlow and Lane, "Antibodies, a laboratory manual", Cold Spring Harbor Laboratory, 1988). Антитела искусственного происхождения могут быть созданы с применением твердофазного синтеза пептидов, могут быть получены с помощью рекомбинантных методов (например, как описано в патенте США US 4,186,567) или могут быть получены, например, при скрининге комбинаторных библиотек, содержащих вариабельные участки тяжелых цепей и вариабельные участки легких цепей (см., например, патент США US 5,969,108, выданный McCafferty).
В составе биспецифических антител по изобретению можно использовать антитела, фрагменты антител, антигенсвязывающие домены или вариабельные участки любого вида животного. Антитела, фрагменты антител, антигенсвязывающие домены или вариабельные участки, которые можно использовать в данном изобретении, могут происходить из организма мыши, примата или человека, но не ограничиваться ими. Если антитело предназначено для применения у человека, можно использовать химерную форму антитела, где константные участки антитела имеют человеческое происхождение. Гуманизированные или полностью человеческие формы антитела могут быть изготовлены согласно способам, известным в данной области техники (см., например, патент США US 5,565,332, выданный Winter). Гуманизацию можно осуществлять различными способами, включая (а) пересаживание CDR, не являющихся человеческими (например, антитела-донора) на каркасные участки и константные участки человеческого происхождения (например, антитела-реципиента) с сохранением или без сохранения ключевых остатков каркасных участков (например, тех, которые важны для сохранения надлежащей аффинности связывания антигена или функций антитела), (б) пересаживание только участков, определяющих специфичность (SDR или a-CDR; остатков, имеющих ключевое значение для взаимодействия антиген-антитело), не являющихся человеческими, на каркасные участки и константные участки человеческого происхождения, или (в) трансплантацию не являющихся человеческими вариабельных доменов целиком и "укрывание" их антропоморфными участками путем замены поверхностных аминокислотных остатков, но не ограничиваясь ими. Гуманизированные антитела и способы их получения представлены, например, в обзоре Almagro and Fransson, Front Biosci 13, 1619-1633 (2008), и более подробно описаны, например, in Riechmann et al., Nature 332, 323-329 (1988); Queen et al., Proc Natl Acad Sci USA 86, 10029-10033 (1989); патентах США US 5,821,337, 7,527,791, 6,982,321 и 7,087,409; Jones et al., Nature 321, 522-525 (1986); Morrison et al., Proc Natl Acad Sci 81, 6851-6855 (1984); Morrison and Oi, Adv Immunol 44, 65-92 (1988); Verhoeyen et al., Science 239, 1534-1536 (1988); Padlan, Molec Immun 31(3), 169-217 (1994); Kashmiri et al., Methods 36, 25-34 (2005) (описание пересадки SDR (a-CDR)); Padlan, Mol Immunol 28, 489-498 (1991) (описание "изменения поверхности"); Dall'Acqua et al., Methods 36, 43-60 (2005) (описание "перегруппировки FR"); and Osbourn et al., Methods 36, 61-68 (2005) и Klimka et al., Br J Cancer 83, 252-260 (2000) (описание "направленного отбора" для перегруппировки FR). Человеческие антитела и человеческие вариабельные участки могут быть получены при помощи различных способов, известных в данной области техники. Человеческие антитела описаны van Dijk and van de Winkel, Curr Opin Pharmacol 5, 368-74 (2001) и Lonberg, Curr Opin Immunol 20, 450-459 (2008). Вариабельные участки человеческого происхождения могут входить в состав и могут быть получены из человеческих моноклональных антител, полученных при помощи гибридомной технологии (см. например, Monoclonal антиbody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987)). Человеческие антитела и вариабельные участки человеческого происхождения также можно получить путем введения иммуногена трансгенному животному, модифицированному для получения интактных человеческих антител или интактных антител с вариабельными участками человеческого происхождения в ответ на введение антигена (см. например, Lonberg, Nat Biotech 23, 1117-1125 (2005). Человеческие антитела и вариабельные участки человеческого происхождения также могут быть получены путем выделения последовательностей вариабельных участков Fv клонов, отобранных из библиотек фагового дисплея, созданных на основе генов человека (см., например, Hoogenboom et al. in Methods in Molecular Biology 178, 1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001); and McCafferty et al., Nature 348, 552-554; Clackson et al., Nature 352, 624-628 (1991)). Фаги, как правило, экспонируют фрагменты антител, либо в виде одноцепочечных фрагментов Fv (scFv), либо в виде Fab фрагментов.
В некоторых воплощениях биспецифические антитела по настоящему изобретению модифицированы с использованием генной инженерии таким образом, чтобы обладать повышенной аффинностью связывания, например, согласно способам, описанным в заявке на патент США US 2004/0132066, включенной сюда во всей полноте путем ссылки. Способность биспецифического антитела по изобретению связывать определенные антигенные детерминанты можно измерить либо при помощи иммуноферментного анализа (ИФА), либо других методов, известных специалистам в данной области техники, например, метода поверхностного плазменного резонанса (анализируется с использованием системы BIACORE T100) (Liljeblad, et al., Glyco J 17, 323-329 (2000)) и традиционных тестов связывания (Heeley, Endocr Res 28, 217-229 (2002)). Для идентификации антитела, фрагмента антитела, антигенсвязывающего домена или вариабельного домена, конкурирующего с референтным антителом за связывание с конкретным антигеном, например, антитела, конкурирующего с антителом V9 за связывание с CD3, можно использовать конкурентный анализ. В некоторых воплощениях, такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), с которым связано референтное антитело. Подробнее примеры способов картирования эпитопов, с которым связывается антитело, описаны Morris (1996) "Epitope Mapping Protocols," in Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ). В приведенном в качестве примера конкурентном анализе иммобилизованный антиген (например, CD3) инкубируют с раствором, содержащим первое меченое антитело, связывающееся с антигеном (например, антитело V9), и второе немеченое антитело, у которого определяют способность конкурировать с первым антителом за связывание с антигеном. Второе антитело может находиться в супернатанте гибридом. В качестве контроля иммобилизованный антиген инкубируют с раствором, содержащим первое меченое антитело, но не содержащим второго немеченого антитела. После инкубации в условиях, позволяющих связаться первому антителу с антигеном, избыток несвязанного антитела удаляют, и определяют количество метки, связавшейся с иммобилизованным антигеном. Если количество метки, связавшейся с иммобилизованным антигеном существенно снижается в исследуемом образце по отношению к контрольному образцу, это указывает на то, что второе антитело конкурирует с первым антителом за связывание с антигеном. См. Harlow and Lane (1988) Anybodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
Биспецифические антитела, полученные согласно данному описанию, могут быть очищены при помощи известных в области техники методов, таких как высокоэффективная жидкостная хроматография, ионообменная хроматография, электрофорез в геле, аффинная хроматография, эксклюзионная хроматография и т.п. Условия, используемые для очистки конкретного белка, будут зависеть, в частности, от таких факторов, как суммарный заряд, гидрофобность, гидрофильность и т.д., и будут очевидны для специалистов в данной области техники. Для очистки методом аффинной хроматографии можно применять антитело, лиганд, рецептор или антиген, с которым связывается биспецифическое антитело. Например, для очистки биспецифического антитела по изобретению методом аффинной хроматографии можно применять матрикс с белком А или белком G. Для выделения биспецифического антитела можно последовательно применять аффинную хроматографию с белком А или G и эксклюзионную хроматографию, по существу, как описано в Примерах. Степень чистоты биспецифического антитела можно определять при помощи любого из многочисленных аналитических способов, хорошо известных в области техники, включая электрофорез в геле, жидкостную хроматографию высокого давления и т.п.
В. Количественные методы анализа
Можно проводить идентификацию, скрининг или исследование физических/химических свойств и/или биологической активности предложенных здесь биспецифических антител при помощи различных количественных методов анализа, известных в области техники.
1. Анализ связывания и другие количественные методы анализа
В одном аспекте определяют антигенсвязывающую активность биспецифического антитела по изобретению, например, при помощи таких известных способов, как иммуноферментный анализ, вестерн-блоттинг и т.д.
В другом аспекте для идентификации антитела, конкурирующего со специфическим антителом за связывание с первым или вторым антигеном, можно применять конкурентный анализ. В некоторых воплощениях такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), с которым связано специфическое антитело. Подробнее примеры способов картирования эпитопов, с которым связывается антитело, описаны Morris (1996) "Epitope Mapping Protocols," in Methods in Molecular Biology vol.66 (Humana Press, Totowa, NJ).
2. Исследование активности
В одном аспекте предложены количественные методы анализа для выявления биологической активности биспецифических антител. Биологическая активность может включать, например, лизис клеток-мишеней или индукцию апоптоза. Также предложены анитела, проявляющие такую биологическую активность in vivo и/или in vitro.
В некоторых воплощениях исследуют биологическую активность биспецифического антитела по изобретению. Количественные методы анализа для исследования цитолиза (например, по высвобождению ЛДГ) или апоптоза (например, с помощью теста TUNEL) хорошо известны в данной области техники.
Г. Иммуноконъюгаты
В изобретении также предложены иммуноконъюгаты, содержащие биспецифическое антитело по изобретению, конъюгированное с одним или несколькими цитотоксическими агентами, например, химиотерапевтическими агентами или лекарствами, подавляющими рост агентами, токсинами (например, белковыми токсинами, энзиматически активными токсинами бактериального, грибкового, растительного или животного происхождения и их фрагментами) или радиоактивными изотопами.
В одном воплощении иммуноконъюгат представляет собой конъюгат антитело-лекарство (ADC), в составе которого антитело конъюгировано с одним или несколькими лекарствами, включая майтансиноид (см. патенты США US 5,208,020, 5,416,064 и европейский патент ЕР 0425235 В1); ауристатин, например, лекарственные фрагменты DE и DF монометилауристатина (ММАЕ и MMAF) (см. патенты США US 5,635,483 и 5,780,588 и 7,498,298); доластатин, калихеамицин или их производные (см. патенты США US 5,712,374, 5,714,586, 5,739,116, 5,767,285, 5,770,701, 5,770,710, 5,773,001 и 5,877,296; Hinman et al., Cancer Res. 53:3336-3342 (1993) и Lode et al., Cancer Res. 58:2925-2928 (1998)); антрациклин, например, дауномицин или доксорубицин (см. Kratz et al., Current Med. Chem. 13:477-523 (2006); Jeffrey et al., Bioorganic & Med. Chem. Letters 16:358-362 (2006); Torgov et al., Bioconj. Chem. 16:717-721 (2005); Nagy et al., Proc. Natl. Acad. Sci. USA 97:829-834 (2000); Dubowchik et al., Bioorg. & Med. Chem. Letters 12:1529-1532 (2002); King et al., J. Med. Chem. 45:4336-4343 (2002) США US 6,630,579); метотрексат; виндезин; таксам, например, доцетаксел, паклитаксел, ларотаксел, тезетаксел, ортатаксел; трихотецен; и СС1065, но не ограничиваясь ими.
В другом воплощении иммуноконъюгат содержит описанное в данном документе биспецифическое антитело, конъюгированное с энзиматически активным токсином или его фрагментом, включая А-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки диантины, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курицин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, ретстриктоцин, феномицин, эномицин и трихотецены, но не ограничиваясь ими.
В другом воплощении иммуноконъюгат содержит описанное в данном документе биспецифическое антитело, конъюгированное с радиоактивным изотопом и формирующее "радиоконъюгат". Для получения радиоактивных конъюгатов существует большое разнообразие радиоактивных изотопов. Примеры их включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если для детекции применяется радиоконъюгат, он может содержать радиоактивный изотоп для проведения сцинтиграфического исследования, например, tc99m или I123, или спиновую метку для проведения исследования методом ядерного магнитного резонанса (ЯМР) (также известного под названием магнитного резонансного исследования), например, иод-123, иод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты биспецифического антитела с цитотоксическим агентом могут быть получены с применением различных бифункциональных сшивающих агентов для присоединения белков, таких как N-сукцинимидил-3-(2-пиридилдитио)-пропионат (SPDP), сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональных производных имидоэфиров (таких как диметил адипимидат HCI), активных эфиров (таких как дисукцинимидил суберат), альдегидов (таких как глутаральдегид), бис-азидо соединений (таких как бис (р-азидобензоил) гександиамин), производных бисдиазония (таких как бис-(р-диазоний-бензоил)-этилендиамин), диизоцианатов (таких как толуол 2,6-диизоцианат) и соединений с двумя активными атомами фтора (таких как 1,5-дифлуоро-2,4-динитробензен). Например, иммунотоксин рицина может быть получен согласно описанию Vitetta et al., Science 238:1098 (1987). Меченая углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен триаминопентауксусная кислота (MX-DTPA) является примером хелатирующего агента для конъюгирования меченого радиоактивным изотопом нуклеотида с антителом. См. WO 94/11026. Линкер может быть "расщепляемым линкером", способствующим высвобождению цитотоксического лекарства в клетках. Например, можно применять кислотолабильный линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Res. 52:127-131 (1992); патент США US 5,208,020).
Иммуноконъюгаты или конъюгаты антитело-лекарство в данном документе явным образом предусматривают но не ограничиваются конъюгатами, полученными с помощью образующих поперечные связи реагентов, включающих, но не ограничивающихся BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)-бензоат), которые доступны для приобретения (например, в компании Pierce Biotechnology, Inc., Rockford, IL, U.S.A).
Д. Способы и композиции для диагностики и детекции
В некоторых воплощениях любое из предложенных здесь биспецифических антител можно применять для детекции наличия первого и/или второго антигена в биологическом образце. Термин "детекция", используемый в данном документе, включает количественное или качественное определение. В некоторых воплощениях биологический образец включает клетку или ткань.
В одном воплощении предложено биспецифическое антитело, которое может найти применение в способах диагностики или детекции. В следующем аспекте предложен способ детекции наличия первого и/или второго антигена в биологическом образце. В некоторых воплощениях способ включает приведение в контакт биологического образца с биспецифическим антителом по данному описанию в условиях, обеспечивающих связывание биспецифического антитела с первым и/или вторым антигеном, и определение, сформировался ли комплекс между биспецифическим антителом и первым и/или вторым антигеном. Такой способ может представлять собой способ для применения in vitro или in vivo. В одном воплощении биспецифическое антитело применяют для отбора субъектов, подходящих для лечения с применением биспецифического антитела, например, когда первый и/или второй антиген представляет собой биомаркер для отбора пациентов.
Примеры нарушений, которые можно диагностировать с применением антитела по изобретению, включают рак.
В некоторых воплощениях предложены меченные биспецифические антитела. Метки включают детектируемые напрямую метки или компоненты (например, флуоресцентные, хромофорные, электронно-плотные, хемилюминесцентные и радиоактивные метки), а также компоненты, например, ферменты или лиганды, которые детектируют опосредовано, например, при помощи ферментативной реакции или молекулярного взаимодействия, но не ограничиваются ими. Примеры меток включают радиоизотопы 32P, 14С, 125I, 3H и 131I, флуорофоры, например, редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светлячка и бактериальную люциферазу (патент США US 4,737,456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, р-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат дегидрогеназу, оксидазы гетероциклических соединений, такие как уриказа и ксантин-оксидаза, связанные с ферментом, использующим пероксид водорода для окисления хромогена, таким как пероксидаза хрена, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы, и им подобные, но не ограничиваются ими.
Е. Фармацевтические препараты
Фармацевтические препараты биспецифического антитела по данному описанию изготавливают путем смешивания такого биспецифического антитела, имеющего необходимую степень чистоты, с одним или несколькими возможными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в форме лиофилизированного препарата или водного раствора. Фармацевтически приемлемые носители, как правило, являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают: буферы, например, фосфатный, цитратный и содержащие Другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензил аммоний хлорид; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкил парабены, такие как метил или пропил парабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды;белки, такие как сывороточный альбумин, желатин или иммуноглобулины, гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; углеводы, такие как сахароза, маннитол, трегалоза или сорбитол; солеобразующие противоионы, такие как натрий; комплексы с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ), но не ограничиваются ими. Примеры фармацевтически приемлемых носителей здесь также включают диспергирующие агенты, такие как активные в нейтральных условиях растворимые гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы РН-20 человека, такие как rHuPH20 (HYLENEX®, Baxter International, Inc.). Примеры некоторых sHASEGP и способов их применения, включая rHuPH20, описаны в публикациях US 2005/0260186 и 2006/0104968. В одном аспекте sHASEGP дополнительно комбинирован с одним или более гликозаминогликаназами, такими как хондроитиназы.
Примеры лиофилизированных препаратов антител описаны в патенте США US 6,267,958. Водные препараты антител включают описанные в патенте США US 6,171,586 и WO 2006/044908, последние включают гистидин-ацетатный буфер.
Препараты могут также содержать несколько активных ингредиентов, предпочтительно имеющих комплементарную активность и не оказывающих друг на друга отрицательного влияния. Такие активные ингредиенты соответственно находятся в комбинации в количествах, эффективных для намеченного использования.
Активные ингредиенты могут быть заключены в микрокапсулы, приготовленные, например, с помощью технологии коацервации или полимеризации на границе раздела фаз, например, в микрокапсулы гидроксиметилцеллюлозы или желатина и микрокапсулы поли-(метилметакрилата), соответственно, в коллоидные системы доставки лекарственных препаратов (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методики описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно изготавливать препараты с пролонгированным высвобождением. Примеры подходящих препаратов с пролонгированным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антитело, в которых матрицы находятся в форме профилированных изделий, например, в виде пленок или микрокапсул.
Препараты, применяющиеся для введения in vivo, как правило, стерильны. Стерильность может быть достигнута, например, путем фильтрации через стерильные фильтрующие мембраны.
Ж. Терапевтические способы и композиции
Любое из предложенных в данном документе биспецифических антител можно применять в терапевтических способах.
В одном аспекте предложено биспецифическое антитело для применения в качестве лекарственного препарата. В других аспектах предложено биспецифическое антитело для применения в лечении рака. В некоторых воплощениях предложено биспецифическое антитело для применения в способе лечения. В некоторых воплощения изобретения предложено биспецифическое антитело для применения в способе лечения индивида, страдающего раком, который включает введение индивиду эффективного количества биспецифического антитела. В одном из таких воплощений способ также включает введение индивиду эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, согласно описанию ниже. "Индивидом" согласно любому из вышеперечисленных воплощений предпочтительно является человек.
В следующем аспекте изобретения предложено применение биспецифического антитела в производстве или изготовлении лекарственных препаратов. В одном воплощении лекарственный препарат предназначен для лечения рака. В следующем воплощении лекарственный препарат предназначен для применения в способе лечения рака, включающем введение эффективного количества лекарственного препарата индивиду, имеющему рак. В одном таком воплощении способ также включает введение индивиду эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, согласно описанию ниже. "Индивидом" согласно любому из вышеперечисленных воплощений может быть человек.
В следующем аспекте изобретения предложен способ лечения рака. В одном воплощении способ включает введение индивиду, имеющему рак, эффективного количества биспецифического антитела. В одном таком воплощении способ также включает введение индивиду эффективного количества по меньшей мере одного дополнительного терапевтического агента, согласно описанию ниже. "Индивидом" согласно любому из вышеперечисленных воплощений может быть человек.
В следующем аспекте изобретения предложены фармацевтические препараты, содержащие любое из предложенных здесь биспецифических антител, например, для применения в любом из вышеперечисленных терапевтических способов. В одном воплощении фармацевтический препарат содержит любое из предложенных здесь биспецифических антител и фармацевтически приемлемый носитель. В другом воплощении фармацевтический препарат содержит любое из предложенных здесь биспецифических антител и по меньшей мере один дополнительный терапевтический агент, например, согласно описанию ниже.
Биспецифические антитела по изобретению могут применяться для терапии либо отдельно, либо в комбинации с другими агентами. Например, антитело по изобретению можно вводить одновременно по меньшей мере с одним дополнительным терапевтическим агентом.
Такие комбинированные терапии, упомянутые выше, включают комбинированное введение (когда два или более терапевтических агентов включены в состав одного или нескольких препаратов) и раздельное введение, когда введение антитела по изобретению может осуществляться до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта. Биспецифические антитела по изобретению также можно применять в комбинации с лучевой терапией.
Биспецифическое антитело по изобретению (и любой дополнительный терапевтический агент) можно вводить любым подходящим способом, включая парентеральное, внутрилегочное и интраназальное и, при необходимости местного лечения, внутриочаговое введение. Парентеральное введение включает внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Дозирование может выполняться любым подходящим способом, например, посредством инъекций, таких как внутривенные или подкожные инъекции, в некоторой степени в зависимости от того, является ли введение кратковременным или длительным. Здесь предусматриваются различные режимы дозирования, включая однократное или многократное введение через различные промежутки времени, болюсное введение и пульсовое введение, но не ограничиваясь ими.
Биспецифические антитела по изобретению должны изготавливаться, дозироваться и вводиться в соответствии с надлежащей медицинской практикой. Факторы, которые необходимо при этом учитывать, включают конкретное заболевание, подлежащее лечению, конкретное млекопитающее, получающее лечение, клиническое состояние индивидуального пациента, причину нарушения, место введения агента, способ введения, схему введения и другие факторы, известные медицинскому персоналу. Биспецифическое антитело не обязательно, но возможно находится в сочетании с одним или несколькими агентами, используемыми на данный момент для предупреждения или лечения обсуждаемого нарушения. Эффективное количество этих иных агентов зависит от количества антитела, присутствующего в препарате, типа нарушения или лечения и других факторов, обсуждавшихся выше. Они как правило применяются в таких же дозировках и вводятся такими же способами, как описанные в данном документе, или как приблизительно от 1 до 99% описанных здесь дозировок, или в любой дозировке и любым способом, который эмпирически/клинически считается подходящими.
Соответствующая дозировка биспецифического антитела по изобретению (при использовании отдельно или в комбинации с одним или несколькими дополнительными терапевтическими агентами) для предупреждения или лечения заболевания будет зависеть от типа заболевания, подлежащего лечению, типа антитела, тяжести и течения заболевания, от того, вводят ли антитело в превентивных или в терапевтических целях, от предшествующего лечения, истории болезни пациента и ответа на биспецифическое антитело, а также от выбора лечащего врача. Соответственно, антитело вводят пациенту единовременно или на протяжении серии процедур. В зависимости от типа и тяжести заболевания возможная исходная дозировка для введения пациенту может составлять приблизительно от 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг-10 мг/кг) биспецифического антитела, например, как в виде одного или нескольких отдельных введений, так и в виде длительной инфузии. Одна типичная дозировка может варьировать приблизительно от 1 мкг/кг до 100 мг/кг или более, в зависимости от упомянутых выше факторов. При повторном введении на протяжении нескольких дней или более, в зависимости от патологического состояния, лечение должно длиться до тех пор, когда наступит желаемое подавление симптомов заболевания. Одним из примеров дозировки биспецифического антитела может быть диапазон приблизительно от 0,05 мг/кг приблизительно до 10 мг/кг. Таким образом, пациенту могут быть введены одна или более доз, составляющих приблизительно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любые их комбинации). Эти дозы могут быть введены с перерывами, например, каждую неделю или каждые три недели (например, таким образом, чтобы пациент получил приблизительно от двух до двадцати, или, например, приблизительно шесть доз биспецифического антитела). Может производиться введение первоначальной ударной дозы, за которой следует введение одной или нескольких более низких доз. Однако, могут применяться и другие режимы дозирования. Успех данного лечения легко отслеживается при помощи известных методик и тестов.
Очевидно, что любой из вышеупомянутых препаратов или терапевтических способов может осуществляться с применением иммуноконъюгата по изобретению вместо или в дополнение к биспецифическому антителу по изобретению.
3. Изделия
В другом аспекте изобретения предложены изделия, содержащие материалы, которые могут найти применение в лечении, предупреждении и/или диагностике нарушений, описанных выше. Изделия включают контейнер и этикетку или листовку-вкладыш на контейнере или внутри него. Подходящие контейнеры включают, например, бутыли, флаконы, шприцы, пакеты с раствором для внутривенного введения и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит композицию, которая самостоятельно или в комбинации с другой композицией является эффективной при лечении, предупреждении и/или диагностике патологических состояний, и может иметь отверстие для стерильного доступа (например, контейнер может представлять собой пакет с раствором для внутривенного введения или флакон с пробкой, которую можно проткнуть иглой для подкожных инъекций). По меньшей мере один активный агент в композиции представляет собой биспецифическое антитело по изобретению. На этикетке или листовке-вкладыше указано, что композицию применяют для лечения предпочтительного патологического состояния. Кроме того, изделие может содержать (а) первый контейнер с содержащейся в нем композицией, где композиция содержит биспецифическое антитело по изобретению и (б) второй контейнер с содержащейся в нем композицией, где композиция содержит цитотоксический или иной терапевтический агент. Изделие в данном воплощении изобретения может также содержать листовку-вкладыш, с указанием, что композиции могут применяться в лечении конкретного патологического состояния. Альтернативно или дополнительно изделие может также содержать второй (или третий) контейнер, содержащий фармацевтически-приемлемый буфер, например бактериостатическую воду для инъекций (BWFI), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Оно может также содержать другие материалы, привлекательные с коммерческой и с потребительской точки зрения, включая другие буферы, растворители, фильтры, иглы и шприцы.
Очевидно, что любое из вышеупомянутых изделий может содержать иммуноконъюгат по изобретению вместо или в дополнение к биспецифическому антителу по изобретению.
III. ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже приведены примеры способов и композиций по изобретению. Очевидно, что с учетом общего описания, приведенного выше, могут осуществляться многие другие воплощения.
В описании настоящего изобретения приведены графические материалы и примеры, позволяющие лучше понять изобретение, но при этом не ограничивающие объем изобретения. Описания всех патентов и научных публикаций, приведенные здесь, включены во всей полноте путем ссылок.
Пример 1: Получение Fab (MCSP)-кpoccFab(CD3)
Полученные последовательности ДНК вариабельных участков тяжелой и легкой цепей субклонировали в рамке считывания либо вместе с константным участком тяжелой цепи, либо вместе с константным участком легкой цепи, предварительно встроенными в соответствующий вектор-реципиент для экспрессии в клетках млекопитающих. Экспрессия антитела находилась под контролем промотора MPSV, а на 3' конце кодирующей последовательности находилась синтетическая сигнальная последовательность полиА. Кроме того, каждый вектор содержал последовательность ориджина репликации OriP вируса Эпштейна-Барр.
Для продукции молекулы осуществляли совместную трансфекцию клеток HEK293-EBNA векторами для экспрессии в клетках млекопитающих при помощи кальций-фосфатной трансфекции. Клетки HEK293-EBNA в фазе экспоненциального роста трансфецировали при помощи кальций-фосфатного метода. В альтернативном случае, клетки HEK293-EBNA, растущие в суспензии, трансфецировали при помощи полиэтиленимина. Клетки трансфецировали с использованием соответствующих экспрессирующих векторов в соотношении 1:1:1 ("вектор CH1-VH-CK-VH":"вектор, несущий легкую цепь":"вектор, несущий легкую цепь CH1-VL").
Для трансфекции с применением фосфата кальция клетки выращивали в виде адгезивного монослоя в культуральных флаконах, используя среду для культивирования среду Игла, модифицированную Дульбекко (DMEM), с добавлением 10% (об/об) эмбриональной телячьей сыворотки, и трансфецировали при конфлюентности от 50 до 80%. Для трансфекции высаживали 15 млн клеток во флаконы Т150 в 25 мл культуральной среды DMEM с добавлением эмбриональной телячьей сыворотки (в конечной концентрации 10% об/об) за 24 часа до трансфекции и помещали клетки на ночь в инкубатор с температурой 37°С и 5% CO2 в атмосфере. На каждый флакон Т150 для трансфекции готовили раствор ДНК, CaCl2 и воды путем добавления к 94 мкг суммарной плазмидной ДНК векторов, смешанных в соответствующих пропорциях, воды до конечного объема 469 мкл и 469 мкл раствора 1 М СаCl2. К этому раствору добавляли 938 мкл раствора 50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2HPO4 с рН 7,05, немедленно смешивали в течение 10 с и оставляли при комнатной температуре в течение 20 с. Суспензию разводили в 10 мл DMEM с добавлением 2% (об/об) эмбриональной телячьей сыворотки и добавляли во флаконы Т150 вместо находящейся там среды. Затем добавляли дополнительно 13 мл среды для трансфекции. Клетки инкубировали при 37°С, 5% CO2 в течение приблизительно 17-20 часов, затем среду заменяли 25 мл DMEM с 10% эмбриональной телячьей сыворотки. Приблизительно через 7 дней после замены среды собирали кондиционированную культуральную среду путем центрифугирования в течение 15 мин при 210×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (вес/об) и хранили при 4°С.
Для трансфекции с применением полиэтиленимина клетки HEK293 EBNA культивировали в суспензии в среде для культивирования CD CHO без добавления сыворотки. Для продукции высевали 400 млн клеток НЕК293 EBNA в 500-мл встряхиваемые колбы за 24 часа до трансфекции. Для трансфекции клетки центрифугировали в течение 5 мин при 210хд, надосадочную жидкость заменяли 20 мл предварительно прогретой среды CD CHO. Экспрессирующие векторы смешивали в 20 мл среды CD CHO для получения общего количества ДНК 200 мкг. После добавления 540 мкл ПЭИ раствор перемешивали на вортексе в течение 15 с и затем инкубировали в течение 10 мин при комнатной температуре. После этого клетки смешивали с раствором ДНК/ПЭИ, переносили в 500-мл встряхиваемую колбу и инкубировали в течение 3 часов при 37°С в инкубаторе с содержанием 5% CO2 в атмосфере. По истечении времени инкубации добавляли 160 мл среды F17 и культивировали клетки в течение 24 часов. Через день после трансфекции вносили 1 мМ вальпроевой кислоты и 7% добавки к среде Feed 1 (Lonza). Через 7 дней культивирования собирали надосадочную жидкость для очистки путем центрифугирования в течение 15 мин при 210×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (вес/об) и хранили при 4°С.
Секретируемый белок выделяли из культуральной надосадочной жидкости при помощи аффинной хроматографии с использованием белка А и белка G, с эксклюзионной хроматографией на последующем этапе. Для аффинной хроматографии надосадочную жидкость загружали в колонку HiTrap Protein A HP (объем колонки=5 мл, GE Healthcare), присоединенную к колонке HiTrap Protein G HP (объем колонки=5 мл, GE Healthcare), каждую колонку уравновешивали 30 мл 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5. Несвязавшийся белок удаляли путем пропускания через обе колонки 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5 в объеме, равном 6 объемам колонки. После этого требовался дополнительный этап отмывки для промывания только колонки HiTrap Protein G HP с использованием 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5 в объеме, равном по меньшей мере 8 объемам колонки. Целевой белок элюировали с колонки HiTrap Protein G HP с помощью ступенчатого градиента 8,8 мМ муравьиной кислоты, рН 3,0 в объеме, равном 7 объемам колонки. Раствор белка нейтрализовали добавлением 1/10 0,5 М фосфата натрия, рН 8.0. Целевой белок концентрировали и фильтровали перед загрузкой в колонку HiLoad Superdex 200 (GE Healthcare), уравновешенную раствором 25 мМ фосфата калия, 125 мМ хлорида натрия, 100 мМ глицина с рН 6,7.
Концентрацию белка в очищенных белковых образцах определяли путем измерения оптической плотности (OD) при 280 нм, с использованием молярного коэффициента экстинкции, рассчитанного на основании аминокислотной последовательности. Степень чистоты и молекулярную массу антител анализировали при помощи диск-электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПАГ) в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотрейтола) и окрашивания Кумасси (SimpleBlue™ SafeStain, Invitrogen). Систему гелей NuPAGE® Pre-Cast (Invitrogen, USA) использовали в соответствии с инструкциями производителя (4-12% Трис-ацетатные гели или 4-12% бис-Трис гели). Содержание агрегатов в образцах антител анализировали при помощи аналитической эксклюзионной хроматографии с использованием колонок Superdex 200 10/300GL (GE Healthcare, Sweden) в проточном буфере 2 мМ MOPS, 150 мМ NaCl, 0,02% (масс/об) NaN3, рН 7,3 при 25°С.
Анализ продукции и очистки приведенной в качестве примера молекулы Fab-кpoccFab (состоящей из трех цепей: VHCH1(MCSP)-VLCH1(CD3V9)=SEQ ID NO:25, VLCL(MCSP)=SEQ ID NO:17 и VHCL(CD3V9)=SEQ ID NO:23; расположенных, как показано на Фиг.1а)) представлен на Фиг.2 и 3. Эта молекула также обозначается Fab (MCSP)-кpoccFab (CD3) или hu Fab (MCSP)-кpoccFab (CD3).
Пример 2: Получение Fab (MCSP)-Fab (MCSP)-кpoccFab(CD3) и Fab (MCSP)-кpoccFab(CD3)-Fab (MCSP)
Полученные последовательности ДНК вариабельных участков тяжелой и легкой цепей субклонировали в рамке считывания либо вместе с константным участком тяжелой цепи, либо вместе с константным участком легкой цепи, предварительно встроенными в соответствующий вектор-реципиент для экспрессии в клетках млекопитающих. Экспрессия антитела находилась под контролем промотора MPSV, а на 3' конце кодирующей последовательности находилась синтетическая сигнальная последовательность полиА. Кроме того, каждый вектор содержал последовательность ориджина репликации OriP вируса Эпштейна-Барр.
Для продукции молекулы осуществляли совместную трансфекцию клеток HEK293-EBNA векторами для экспрессии в клетках млекопитающих при помощи кальций-фосфатной трансфекции. Клетки HEK293-EBNA в фазе экспоненциального роста трансфецировали при помощи кальций-фосфатного метода. В альтернативном случае, клетки HEK293-EBNA, растущие в суспензии, трансфецировали при помощи полиэтиленимина. Клетки трансфецировали с использованием соответствующих экспрессирующих векторов в соотношении 1:2:1 ("вектор CH1-VH-CH1-VH-CK-VH":"вектор, несущий легкую цепь":"вектор, несущий легкую цепь CH1-VL").
Для трансфекции с применением фосфата кальция клетки выращивали в виде адгезивного монослоя в культуральных флаконах, используя среду для культивирования Игла, модифицированную по способу Дульбекко (DMEM) с добавлением 10% (об/об) эмбриональной телячьей сыворотки и трансфецировали при конфлюентности от 50 до 80%. Для трансфекции высаживали 15 млн клеток во флаконы Т150 в 25 мл культуральной среды DMEM с добавлением эмбриональной телячьей сыворотки (в конечной концентрации 10% об/об) за 24 часа до трансфекции и помещали клетки на ночь в инкубатор с температурой 37 С и 5% CO2 в атмосфере. На каждый флакон Т150 для трансфекции готовили раствор ДНК, CaCl2 и воды путем добавления к 94 мкг суммарной плазмидной ДНК векторов, смешанных в соответствующих пропорциях, воды до конечного объема 469 мкл и 469 мкл раствора 1 М CaCl2. К этому раствору добавляли 938 мкл раствора 50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2HPO4 с рН 7,05, немедленно смешивали в течение 10 С и оставляли при комнатной температуре в течение 20 С. Суспензию разводили в 10 мл DMEM с добавлением 2% (об/об) эмбриональной телячьей сыворотки и добавляли во флаконы Т150 вместо находящейся там среды. Затем добавляли дополнительно 13 мл среды для трансфекции. Клетки инкубировали при 37°С, 5% CO2 в течение приблизительно 17-20 часов, затем среду заменяли 25 мл DMEM с 10% эмбриональной телячьей сыворотки. Приблизительно через 7 дней после замены среды собирали кондиционированную культуральную среду путем центрифугирования в течение 15 мин при 210×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (масс/об) и хранили при 4°С. Для трансфекции с применением полиэтиленимина клетки НЕК293 EBNA культивировали в суспензии в среде для культивирования CD CHO без добавления сыворотки. Для продукции высевали 400 млн клеток НЕК293 EBNA в 500-мл встряхиваемые колбы за 24 часа до трансфекции. Для трансфекции клетки центрифугировали в течение 5 мин при 210×g, надосадочную жидкость заменяли 20 мл предварительно прогретой среды CD CHO. Экспрессирующие векторы смешивали в 20 мл среды CD CHO для получения общего количества ДНК 200 мкг. После добавления 540 мкл полиэтиленимина (ПЭИ) раствор перемешивали на вортексе в течение 15 с и затем инкубировали в течение 10 мин при комнатной температуре. После этого клетки смешивали с раствором ДНК/ПЭИ, переносили в 500-мл встряхиваемую колбу и инкубировали в течение 3 часов при 37°С в инкубаторе с содержанием 5% СO2 в атмосфере. По истечении времени инкубации добавляли 160 мл среды F17 и культивировали клетки в течение 24 часов. Через день после трансфекции вносили 1 мМ вальпроевой кислоты и 7% добавки к среде Feed 1 (Lonza). Через 7 дней культивирования собирали надосадочную жидкость для очистки путем центрифугирования в течение 15 мин при 210×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (масс./об) и хранили при 4°С.
Секретируемый белок выделяли из культуральной надосадочной жидкости при помощи аффинной хроматографии с использованием белка А и белка G, с эксклюзионной хроматографией на последующем этапе.
Для аффинной хроматографии надосадочную жидкость загружали в колонку HiTrap Protein A HP (объем колонки=5 мл, GE Healthcare), присоединенную к колонке HiTrap Protein G HP (объем колонки=5 мл, GE Healthcare), каждую колонку уравновешивали 30 мл 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5. Несвязавшийся белок удаляли путем пропускания через обе колонки 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5 в объеме, равном 6 объемам колонки. После этого требовался дополнительный этап отмывки для промывания только колонки HiTrap Protein G HP с использованием 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5 в объеме, равном по меньшей мере 8 объемам колонки. Целевой белок элюировали с колонки HiTrap Protein G HP с помощью ступенчатого градиента 8,8 мМ муравьиной кислоты, рН 3,0 в объеме, равном 7 объемам колонки. Раствор белка нейтрализовали добавлением 1/10 0,5 М фосфата натрия, рН 8,0. Целевой белок концентрировали и фильтровали перед загрузкой в колонку HiLoad Superdex 200 (GE Healthcare), уравновешенную раствором 25 мМ фосфата калия, 125 мМ хлорида натрия, 100 мМ глицина с рН 6,7.
Концентрацию белка в очищенных белковых образцах определяли путем измерения оптической плотности (OD) при 280 нм, с использованием молярного коэффициента экстинкции, рассчитанного на основании аминокислотной последовательности. Степень чистоты и молекулярную массу антител анализировали при помощи ДСН-ПАГ в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотрейтола) и окрашивания Кумасси (SimpleBlue™ SafeStain, Invitrogen). Систему гелей NuPAGE® Pre-Cast (Invitrogen, USA) использовали в соответствии с инструкциями производителя (4-12% Трис-ацетатные гели или 4-12% бис-Трис гели). Содержание агрегатов в образцах антител анализировали при помощи аналитической эксклюзионной хроматографии с использованием колонок Superdex 200 10/300GL (GE Healthcare, Sweden) в проточном буфере 2 мМ MOPS, 150 мМ NaCl, 0,02% (масс./об) NaN3, рН 7,3 при 25°С и сравнивали с фрагментом антитела (scFv)2, известным из уровня техники (результаты приведены в таблице ниже).
Анализ продукции и очистки приведенной в качестве примера молекулы Fab-Fab-кpoccFab (состоящей из четырех цепей: VHCH1(MCSP)-VHCH1(MCSP)-VLCH1(CD3vg)=SEQ ID NO:26, 2 цепей VLCL(MCSP)=SEQ ID NO:17 и одной цепи VHCL(CD3v9)=SEQ ID NO:23; расположенных как показано на Фиг.1с)) представлен на Фиг.4 и 5. Эта молекула также обозначается Fab (MCSP)- Fab (MCSP)-кpoccFab (CD3) или hu Fab (MCSP)-Fab (MCSP)-кpoccFab (CDS).
Анализ продукции и очистки приведенной в качестве примера молекулы Fab-кpoccFab-Fab (состоящей из четырех цепей: VHCH1(MCSP)-VLCH1(CD3V9)-VHCH1(MCSP)=SEQ ID NO:27, 2 цепей VLCL(MCSP)=SEQ ID NO:17 и одной цепи VHCL(CD3V9)=SEQ ID NO:23; расположенных как показано на Фиг.1е)) представлен на Фиг.6 и 7. Эта молекула также обозначается Fab (MCSP)- Fab (MCSP)-кpoccFab (CD3) или hu Fab (MCSP)- Fab (MCSP)-кpoccFab (CD3).
Анализ продукции и очистки приведенной в качестве примера молекулы кpoccFab-Fab-Fab (состоящей из четырех цепей: VLCH1(CD32C11)- VHCHI(MCSP)-VHCH1(MCSP)=SEQ ID NO:42, 2 цепей VLCL(MCSP)=SEQ ID NO:17 и одной цепи VHCL(CD32C11)=SEQ ID NO:43; расположенных как показано на Фиг.1 d)) представлен на Фиг.8 и 9. Эта молекула также обозначается кpoccFab мыши (CD3)-Fab (MCSP)- Fab (MCSP).
Пример 3: Получение Fab(CD33)-кpoccFab (CD3)
Полученные последовательности ДНК вариабельных участков тяжелой и легкой цепей субклонировали в рамке считывания либо вместе с константным участком тяжелой цепи, либо вместе с константным участком легкой цепи, предварительно встроенными в соответствующий вектор-реципиент для экспрессии в клетках млекопитающих. Экспрессия антитела находилась под контролем промотора MPSV, а на 3' конце кодирующей последовательности находилась синтетическая сигнальная последовательность полиА. Кроме того, каждый вектор содержал последовательность ориджина репликации OriP вируса Эпштейна-Барр.
Для продукции молекулы осуществляли совместную трансфекцию клеток HEK293-EBNA векторами для экспрессии в клетках млекопитающих при помощи кальций-фосфатной трансфекции. Клетки HEK293-EBNA в фазе экспоненциального роста трансфецировали при помощи кальций-фосфатного метода. В альтернативном случае, клетки HEK293-EBNA, растущие в суспензии, трансфецировали при помощи полиэтиленимина. Клетки трансфецировали с использованием соответствующих экспрессирующих векторов в соотношении 1:1:1 ("вектор CH1-VH-CK-VH":"вектор, несущий легкую цепь":"вектор, несущий легкую цепь CH1-VL").
Для трансфекции с применением фосфата кальция клетки выращивали в виде адгезивного монослоя в культуральных флаконах, используя среду для культивирования DMEM с добавлением 10% (об/об) эмбриональной телячьей сыворотки и трансфецировали при конфлюентности от 50 до 80%. Для трансфекции высаживали 15 млн клеток во флаконы Т150 в 25 мл культуральной среды DMEM с добавлением эмбриональной телячьей сыворотки (в конечной концентрации 10% об/об) за 24 часа до трансфекции и помещали клетки на ночь в инкубатор с температурой 37°С и 5% CO2 в атмосфере. На каждый флакон Т150 для трансфекции готовили раствор ДНК, CaCl2 и воды путем добавления к 94 мкг суммарной плазмидной ДНК векторов, смешанных в соответствующих пропорциях, воды до конечного объема 469 мкл и 469 мкл раствора 1 М CaCl2. К этому раствору добавляли 938 мкл раствора 50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2HPO4 с рН 7,05, немедленно смешивали в течение 10 с и оставляли при комнатной температуре в течение 20 с. Суспензию разводили в 10 мл DMEM с добавлением 2% (об/об) эмбриональной телячьей сыворотки и добавляли во флаконы Т150 вместо находящейся там среды. Затем добавляли дополнительно 13 мл среды для трансфекции. Клетки инкубировали при 37°С, 5% CO2 в течение приблизительно 17-20 часов, затем среду заменяли 25 мл DMEM с 10% эмбриональной телячьей сыворотки. Приблизительно через 7 дней после замены среды собирали кондиционированную культуральную среду путем центрифугирования в течение 15 мин при 210×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (масс./об) и хранили при 4°С.
Для трансфекции с применением полиэтиленимина клетки HEK293 EBNA культивировали в суспензии в среде для культивирования CD CHO без добавления сыворотки. Для продукции высевали 400 млн клеток НЕК293 EBNA в 500-мл встряхиваемые колбы за 24 часа до трансфекции. Для трансфекции клетки центрифугировали в течение 5 мин при 210×g, надосадочную жидкость заменяли 20 мл предварительно прогретой среды CD CHO. Экспрессирующие векторы смешивали в 20 мл среды CD CHO для получения общего количества ДНК 200 мкг. После добавления 540 мкл ПЭИ раствор перемешивали на вортексе в течение 15 с и затем инкубировали в течение 10 мин при комнатной температуре. После этого клетки смешивали с раствором ДНК/ПЭИ, переносили в 500-мл встряхиваемую колбу и инкубировали в течение 3 часов при 37°С в инкубаторе с содержанием 5% CO2 в атмосфере. По истечении времени инкубации добавляли 160 мл среды F17 и культивировали клетки в течение 24 часов. Через день после трансфекции вносили 1 мМ вальпроевой кислоты и 7% добавки к среде Feed 1 (Lonza). Через 7 дней культивирования собирали надосадочную жидкость для очистки путем центрифугирования в течение 15 мин при 21×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (масс./об) и хранили при 4°С.
Секретируемый белок выделяли из культуральной надосадочной жидкости при помощи аффинной хроматографии с использованием белка А и белка G, с эксклюзионной хроматографией на последующем этапе.
Для аффинной хроматографии надосадочную жидкость загружали в колонку HiTrap Protein A HP (объем колонки=5 мл, GE Healthcare), присоединенную к колонке HiTrap Protein G HP (объем колонки=5 мл, GE Healthcare), каждую колонку уравновешивали 30 мл 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5. Несвязавшийся белок удаляли путем пропускания через обе колонки 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5 в объеме, равном 6 объемам колонки. После этого требовался дополнительный этап отмывки для промывания только колонки HiTrap Protein G HP с использованием 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5 в объеме, равном по меньшей мере 8 объемам колонки. Целевой белок элюировали с колонки HiTrap Protein G HP с помощью ступенчатого градиента 8,8 мМ муравьиной кислоты, рН 3,0 в объеме, равном 7 объемам колонки. Раствор белка нейтрализовали добавлением 1/10 0,5 М фосфата натрия, рН 8,0. Целевой белок концентрировали и фильтровали перед загрузкой в колонку HiLoad Superdex 200 (GE Healthcare), уравновешенную раствором 25 мМ фосфата калия, 125 мМ хлорида натрия, 100 мМ глицина с рН 6,7.
Концентрацию белка в очищенных белковых образцах определяли путем измерения оптической плотности (OD) при 280 нм, с использованием молярного коэффициента экстинкции, рассчитанного на основании аминокислотной последовательности. Степень чистоты и молекулярную массу антител анализировали при помощи ДСН-ПАГ в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотрейтол) и окрашивания Кумасси (SimpleBlue™ SafeStain, Invitrogen). Систему гелей NuPAGE® Pre-Cast (Invitrogen, USA) использовали в соответствии с инструкциями производителя (4-12% Трис-ацетатные гели или 4-12% бис-Трис гели). Содержание агрегатов в образцах антител анализировали при помощи аналитической эксклюзионной хроматографии с использованием колонок Superdex 200 10/300GL (GE Healthcare, Sweden) в проточном буфере 2 мМ MOPS, 150 мМ NaCl, 0,02% (масс./об) NaN3, рН 7,3 при 25°С.
Анализ продукции и очистки приведенной в качестве примера молекулы Fab-кpoccFab (состоящей из трех цепей: VHCH1(CD33)- VLCH1(CD3V9)=SEQ ID NO:102, VLCL(CD33)=SEQ ID NO:100 и VHCL(CD3V9)=SEQ ID NO:23 или SEQ ID NO:101; расположенных как показано на Фиг.1 а)) представлен на Фиг.17 и 18. Эта молекула также обозначается Fab(CD33)-кpoccFab (CD3) или hu Fab(CD33)-кpoccFab (CD3).
Пример 4: Получение референтной молекулы (scFv)2 Клонирование и продукция
Полученные последовательности ДНК вариабельных участков тяжелой и легкой цепей субклонировали в рамке считывания в соответствующий вектор-реципиент для экспрессии в клетках млекопитающих. Экспрессия антитела находилась под контролем промотора MPSV, а на 3'-конце кодирующей последовательности находилась синтетическая сигнальная последовательность полиА. Кроме того, каждый вектор содержал последовательность ориджина репликации OriP вируса Эпштейна-Барр.
Для продукции молекулы осуществляли трансфекцию клеток НЕК293-EBNA вектором для экспрессии в клетках млекопитающих с использованием полиэтиленимина. Клетки НЕК293 EBNA культивировали в суспензии в среде для культивирования CD CHO без добавления сыворотки. Для продукции высевали 400 млн клеток НЕК293 EBNA в 500-мл встряхиваемые колбы за 24 часа до трансфекции. Для трансфекции клетки центрифугировали в течение 5 мин при 210×g, надосадочную жидкость заменяли 20 мл предварительно прогретой среды CD CHO. Экспрессирующие векторы смешивали в 20 мл среды CD CHO для получения общего количества ДНК 200 мкг. После добавления 540 мкл ПЭИ раствор перемешивали на вортексе в течение 15 с и затем инкубировали в течение 10 мин при комнатной температуре. После этого клетки смешивали с раствором ДНК/ПЭИ, переносили в 500-мл встряхиваемую колбу и инкубировали в течение 3 часов при 37°С в инкубаторе с содержанием 5% СO2 в атмосфере. По истечении времени инкубации добавляли 160 мл среды F17 и культивировали клетки в течение 24 часов. Через день после трансфекции вносили 1 мМ вальпроевой кислоты и 7% добавки к среде Feed 1 (Lonza). Через 7 дней культивирования собирали надосадочную жидкость для очистки путем центрифугирования в течение 15 мин при 210×g, раствор стерилизовали фильтрованием (через фильтр с размером пор 0,22 мкм), добавляли азид натрия в конечной концентрации 0,01% (масс./об) и хранили при 4°С.
Очистка (scFv)2 (анти MCSP/анти huCD3)
Секретируемый белок выделяли из культуральной надосадочной жидкости при помощи аффинной хроматографии с использованием иммобилизованных ионов металлов (IMAC) и эксклюзионной хроматографии на последующем этапе.
Перед первым этапом очистки из надосадочной жидкости удаляли способные помешать очистке компоненты путем диафильтрации с применением систем фильтрации в тангенциальном потоке Sarcojet (Sartorius), снабженных мембранами, отсекающими компоненты с молекулярной массой 5.000 (Sartocon Slice Cassette, Hydrosart; Sartorius). Надосадочную жидкость концентрировали до 210 мл и затем разводили в 1 л 20 мМ фосфата натрия, 500 мМ хлорида натрия, рН 6,5. Раствор белка вновь концентрировали до 210 мл. Этот процесс повторяли дважды для осуществления полной замены буфера.
Для аффинной хроматографии ретентат, полученный после диафильтрации, загружали в картридж NiNTA Superflow (объем картриджа=5 мл, Qiagen), уравновешенный 25 мл 20 мМ фосфата натрия, 500 мМ хлорида натрия, 15 мМ имидазола, рН 6,5. Несвязанный белок удаляли путем пропускания через колонку 20 мМ фосфата натрия, 500 мМ хлорида натрия, 15 мМ имидазола, рН 6,5 в объеме, равном по меньшей мере 2 объемам колонки, с последующим дополнительным этапом отмывки с применением 20 мМ фосфата натрия, 500 мМ хлорида натрия, 62,5 мМ имидазола, рН 6,5 в объеме, равном 3 объемам колонки. Целевой белок элюировали 2 объемами 20 мМ фосфата натрия, 500 мМ хлорида натрия, 125 мМ имидазола, рН 6,5. Затем колонку промывали 20 мМ фосфата натрия, 500 мМ хлорида натрия, 250 мМ имидазола, рН 6,5.
Целевой белок концентрировали перед загрузкой в колонку HiLoad Superdex 75 (GE Healthcare), уравновешенную 25 мМ КН2Р04, 125 мМ NaCl, 200 мМ аргинина, рН 6,7.
Выход, содержание агрегатов после первого этапа очистки и содержание мономеров в конечном продукте показано в таблице выше. Сравнение содержания агрегатов после первого этапа очистки указывает, что конструкция Fab-кpoccFab имела наилучшую стабильность в противоположность (scFv)2.
Характеристика (scFv)2
Концентрацию белка в очищенных белковых образцах определяли путем измерения оптической плотности (OD) при 280 нм, с использованием молярного коэффициента экстинкции, рассчитанного на основании аминокислотной последовательности. Степень чистоты и молекулярную массу антител анализировали при помощи ДСН-ПАГ в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотрейтол) и окрашивания Кумасси (SimpleBlue™ SafeStain, Invitrogen). Систему гелей NuPAGE® Pre-Cast (Invitrogen, USA) использовали в соответствии с инструкциями производителя (4-12% Трис-ацетатные гели или 4-12% бис-Трис гели). Содержание агрегатов в образцах антител анализировали при помощи аналитической эксклюзионной хроматографии с использованием колонок Superdex 75 10/300GL (GE Healthcare, Sweden) в проточном буфере 2 мМ MOPS, 150 мМ NaCl, 0,02% (масс./об) NaN3, pH 7,3 при 25°С.
Схематичное изображение молекулы (scFv)2 показано на Фиг.21.
Анализ продукции и очистки приведенной в качестве примера молекулы (scFv)2 (антиMCSP/антиhuCD3; состоящей из двух одноцепочечных Fv: VL-VH (MCSP) и VH-VL (CD3v9)=SEQ ID NO:149); представлен на Фиг.22 и 23. Эта молекула также обозначается (scFv)2 (антиMCSP/антиhuCD3e).
Пример 5: Выделение из МКПК общей популяции первичных человеческих Т клеток
Мононуклеарные клетки периферической крови (МКПК) выделяли из обогащенных лимфоцитами препаратов (лейкоконцентрата), полученных из местных банков крови или из свежей крови доноров (здоровых людей) путем центрифугирования в градиенте плотности Histopaque.
Обогащение МКПК Т-клетками осуществляли с применением набора для выделения общей популяции Т-клеток Pan T Cell Isolation Kit II (Miltenyi Biotec #130-091-156), согласно инструкциям производителя. Вкратце, клеточный осадок разводили в 40 мкл ледяного буфера на 10 млн клеток (PBS с 0,5% БСА, 2 мМ EDTA - стерилизованный фильтрованием) и инкубировали с 10 мкл коктейля биотин-антитело на 10 млн клеток в течение 10 мин при 4°С.
Добавляли 30 мкл ледяного буфера и 20 мкл магнитных бусинок анти-биотин на 10 млн клеток и смесь инкубировали в течение 15 мин при 4°С.
Клетки отмывали путем добавления 10-20-кратного объема по сравнению с объемом, взятым для мечения, и затем центрифугировали при 300 g в течение 10 мин. Клетки в количестве до 100 млн ресуспендировали в 500 мкл буфера.
Магнитное разделение общей популяции немеченых Т клеток человека осуществляли с применением колонок LS (Miltenyi Biotec #130-042-401) согласно инструкциям производителя. Производили автоматический подсчет количества клеток в полученной популяции Т-клеток (ViCell) и хранили в среде AIM-V в инкубаторе с температурой 37°С 5% и CO2 в атмосфере до начала исследования (не более 24 ч).
Пример 6: Выделение мышиных пан-Т-клеток из спленоцитов
У мышей C57BL/6 выделяли селезенку, переносили в пробирки GentleMACS типа С (Miltenyi Biotech #130-093-237), содержащие буфер MACS (PBS+0,5% БСА+2 мМ EDTA) и гомогенизировали с помощью гомогенизатора GentleMACS для получения суспензии, состоящей из единичных клеток, в соответствии с инструкциями производителя.
Клеточную суспензию пропускали через фильтр для предварительной сепарации, чтобы избавиться от оставшихся негомогенизированных частиц ткани. После центрифугирования при 400 g в течение 4 мин при 4°С, добавляли лизирующий буфер АСК для лизиса эритроцитов (инкубация в течение 5 мин при комнатной температуре). Оставшиеся клетки дважды отмывали буфером MACS, подсчитывали и использовали для выделения общей популяции мышиных Т-клеток. Негативную (магнитную) селекцию осуществляли с применением набора для выделения общей популяции Т-клеток фирмы Miltenyi Biotec (#130-090-861), следуя инструкциям производителя. В полученной популяции Т-клеток выполняли автоматический подсчет количества клеток (ViCell) и немедленно использовали в дальнейших исследованиях.
Пример 7: Переадресация Т-клеточной цитотоксичности, опосредованная образованием поперечных связей биспецифическими конструкциями, направленными против CD3 Т-клеток и MCSP опухолевых клеток (исследование высвобождения ЛДГ)
Способность биспецифических конструкций, направленных против CD3 Т-клеток человека или мыши и MCSP опухолевых клеток человека, индуцировать опосредованный Т-клетками апоптоз клеток-мишеней исследовали по высвобождению ЛДГ.
Вкратце, клетки-мишени (человеческие Colo-38, человеческие MDA-MB-435, человеческие клетки меланомы MV-3 или мышиные клетки B16/F10-huMCSP Flue 2 клон 48, все из которых экспрессировали MCSP человека) собирали с использованием диссоциирующего буфера Cell Dissociation Buffer (MCSP чувствителен к трипсину) или трипсина (и затем высаживали за день до эксперимента), отмывали и ресуспендировали в соответствующей среде для культивирования клеток (см. подробное описание графических материалов). В круглодонные 96-луночные планшеты высаживали по 20000-30000 клеток на лунку и добавляли соответствующее антитело в указанном разведении (в трипликатах). Эффекторные клетки добавляли для получения итогового соотношения Эффекторные клетки:клетки-мишени=5:1 (для общей популяции Т-клеток человека), 10:1 (для МКПК человека).
Дополнительно, в качестве митогенного стимула для индукции активации Т-клеток человека или яванского макака использовали фитогемагглютинин РНА-М (Sigma #L8902), смесь изолектинов, выделенных из Phaseolus vulgaris, в концентрации 1-10 мкг/мл. В случае Т-клеток мыши, в качестве положительного контроля активации Т-клеток использовали 5% раствор активатора Т-клеток КонА (реагента "rat T-Stim with ConA") (BD #354115), представлявшего собой кондиционированную среду культуры спленоцитов крыс, стимулированных конканавалином А.
Для нормализации осуществляли максимальный лизис клеток-мишеней (=100%) путем инкубации клеток-мишеней с раствором Triton-X-100 в конечной концентрации 1%. Минимальный лизис (=0%) соответствовал инкубации клеток-мишеней вместе с эффекторными клетками в отсутствие какой-либо конструкции или антитела.
После инкубации в течение ночи по меньшей мере в течение 18 ч при 37°С, 5% CO2, в надосадочной жидкости измеряли концентрацию ЛДГ, выделившейся из апоптотических/некротических клеток-мишеней, при помощи набора для определения ЛДГ (Roche Applied Science, #11644793001), согласно инструкциям производителя.
Высвобождение ЛДГ в присутствии биспецифических конструкций Fab (MCSP)-кpoccFab (CD3) и Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3)
У очищенных Fab (MCSP)-кpoccFab (CD3), Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) и референтной молекулы (scFv)2 (антиMCSP/антиhuCD3e) исследовали способность индуцировать опосредованный Т-клетками апоптоз опухолевых клеток-мишеней при образовании поперечных связей конструкциями за счет связывания обоих нацеленных на антигены компонентов с соответствующими антигенами на клетках. Вкратце, клетки-мишени меланомы человека MDA-MB-435, экспрессирующие huMCSP, собирали с использованием буфера для диссоциации клеток, отмывали и ресуспендировали в среде AIM-V (Invitrogen #12055-091). В круглодонные 96-луночные планшеты высаживали по 30000 клеток на лунку и добавляли соответствующее антитело, разведенное до указанных концентраций. Все конструкции и контроли доводили до одинаковой молярности.
В качестве эффекторных клеток использовали общую популяцию Т-клеток человека при соотношении эффекторных клеток и клеток-мишеней 5:1. В качестве положительного контроля активации общей популяции Т-клеток человека использовали фитогемагглютинин РНА-М (Sigma #1-8902) в концентрации 1 мкг/мл. Для нормализации определяли максимальную степень лизиса клеток-мишеней (=100%) при инкубации клеток-мишеней с раствором Triton-X-100 в конечной концентрации 1%. Минимальный лизис (=0%) соответствовал инкубации клеток-мишеней вместе с эффекторными клеткам в отсутствие каких-либо конструкций или антител.
После инкубации в течение ночи на протяжении 20 ч при 37°С, 5% CO2 в надосадочной жидкости измеряли концентрацию ЛДГ, выделившейся из апоптотических/некротических клеток-мишеней, с использованием набора для определения ЛДГ (Roche Applied Science, #11644793001), согласно инструкциям производителя.
Как показано на Фиг.10, конструкции, имеющие две валентности, направленные против MCSP, демонстрировали цитотоксическую активность, сравнимую с таковой конструкции (scFv)2 (антиМСSР/антиhuCD3e), тогда как конструкция Fab (MCSP)-кpoccFab (CD3), имеющая одну валентность для связывания MCSP, оказывалась явно менее активной.
Высвобождение ЛДГ клетками-мишенями меланомы человека MDA-MB-435 в присутствии биспецифической конструкции Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3)
У очищенного Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) и референтной молекулы (scFv)2 (антиMCSP/антиhuCD3e) исследовали способность индуцировать опосредованный Т-клетками апоптоз опухолевых клеток-мишеней при образовании поперечных связей конструкциями за счет связывания обоих нацеленных на антигены компонентов с соответствующими антигенами на клетках.
Вкратце, клетки-мишени меланомы человека MDA-MB-435, экспрессирующие huMCSP, собирали с использованием буфера для диссоциации клеток, отмывали и ресуспендировали в среде AIM-V (Invitrogen #12055-091). В круглодонные 96-луночные планшеты высаживали по 30000 клеток на лунку и добавляли соответствующее антитело, разведенное до указанных концентраций. Все конструкции и контроли доводили до одинаковой молярности.
В качестве эффекторных клеток использовали общую популяцию Т-клеток человека при соотношении эффекторных клеток и клеток-мишеней 5:1. В качестве положительного контроля активации общей популяции Т-клеток человека использовали фитогемагглютинин PHA-M (Sigma #1-8902) в концентрации 5 мкг/мл. Для нормализации определяли максимальную степень лизиса клеток-мишеней (=100%) при инкубации клеток-мишеней с раствором Triton-X-100 в конечной концентрации 1%. Минимальный лизис (=0%) соответствовал инкубации клеток-мишеней вместе с эффекторными клеткам в отсутствие каких-либо конструкций или антител.
После инкубации в течение ночи на протяжении 21 ч при 37°С, 5% CO2, в надосадочной жидкости измеряли концентрацию ЛДГ, выделившейся из апоптотических/некротических клеток-мишеней, с использованием набора для определения ЛДГ (Roche Applied Science, #11644793001), согласно инструкциям производителя.
Как показано на Фиг.11, присутствие Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) индуцировало апоптоз клеток-мишеней по меньшей мере также эффективно, как присутствие молекул (scFv)2 (антиМСSР/антиhuCD3e).
Высвобождение ЛДГ человеческими клетками меланомы MV-3 в присутствии биспецифической конструкции Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3)
У очищенного Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) и референтной молекулы (scFv)2 (антиМСSР/антиhuCD3e) исследовали способность индуцировать опосредованный Т-клетками апоптоз опухолевых клеток-мишеней при образовании поперечных связей конструкциями за счет связывания обоих нацеленных на антигены компонентов с соответствующими антигенами на клетках.
Вкратце, клетки-мишени меланомы человека MV-3, экспрессирующие huMCSP, собирали с использованием трипсина за день до начала исследования высвобождения ЛДГ. Клетки отмывали и ресуспендировали в соответствующей среде для культивирования. В круглодонные 96-луночные планшеты высаживали по 30000 клеток на лунку. На следующий день удаляли супернатант и добавляли по 100 мкл/лунку среды AIM-V (Invitrogen #12055-091) и соответствующее антитело, разведенное до указанных концентраций. Все конструкции и контроли доводили до одинаковой молярности.
В качестве эффекторных клеток добавляли МКПК человека при соотношении эффекторных клеток и клеток-мишеней 10:1. В качестве положительного контроля активации общей популяции Т-клеток человека использовали фитогемагглютинин РНА-М (Sigma #1-8902) в концентрации 5 мкг/мл. Для нормализации определяли максимальную степень лизиса клеток-мишеней (=100%) при инкубации клеток-мишеней с раствором Triton-X-100 в конечной концентрации 1%. Минимальный лизис (=0%) соответствовал инкубации клеток-мишеней вместе с эффекторными клеткам в отсутствие каких-либо конструкций или антител.
После инкубации в течение ночи на протяжении 26 ч при 37°С, 5% СO2, в надосадочной жидкости измеряли концентрацию ЛДГ, выделившейся из апоптотических/некротических клеток-мишеней, с использованием набора для определения ЛДГ (Roche Applied Science, #11644793001), согласно инструкциям производителя.
Как показано на Фиг.12, присутствие Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) индуцировало апоптоз клеток-мишеней по меньшей мере так же эффективно, как присутствие молекул (scFv)2 (антиМСSР/антиhuCD3e).
Высвобождение ЛДН человечексими клетками меланомы MV-3 в присутствии биспецифической конструкции Fab (MCSP)-кpoccFab (CD3)
Высвобождение ЛДГ исследовали, как описано выше. На Фиг.19 показана цитолитическая активность в отношении положительных по huMCSP опухолевых клеток MV-3 при совместном культивировании с МКПК человека (соотношение эффекторные клетки:клетки-мишени=10:1), в присутствии Fab (MCSP)-кpoccFab (CD3), по сравнению с референтной молекулой (scFv)2 (антиМСSР/антиhuCD3e) в течение ~24 часов.
Высвобождение ЛДГ в присутствии биспецифической конструкции кpoccFab (CD3)-Fab (MCSP)-Fab (MCSP)
У очищенного кpoccFab мыши (CD3)-Fab (MCSP)-Fab (MCSP), направленного против CDS мыши и MCSP человека, исследовали способность индуцировать опосредованный Т-клетками апоптоз опухолевых клеток-мишеней при образовании поперечных связей конструкциями за счет связывания обоих нацеленных на антигены компонентов с соответствующими антигенами на клетках.
Вкратце, опухолевые клетки-мишени B16/F10-huMCSP Fluc2 клон 48, экспрессирующие huMCSP, собирали с использованием буфера для диссоциации клеток, отмывали и ресуспендировали в среде RPMI1640, содержащей 1х заменимые аминокислоты, 10 мМ Hepes, 50 мкМ 2-b-ME и 1 мМ пируват натрия.
В круглодонные 96-луночные планшеты высаживали по 20000 клеток на лунку и добавляли соответствующее антитело, разведенное до указанных концентраций. Биспецифические конструкции и различные контрольные IgG доводили до одинаковой молярности. В качестве дополнительного контроля активации Т-клеток мыши использовали реагент "Т Cell Stim with ConA" (BD #354115), разведенный 1:160 средой для количественного определения.
В качестве эффекторных клеток использовали общую популяцию Т-клеток мыши, выделенных из спленоцитов (мышей C57BL/6) при итоговом соотношении эффекторные клеток и клеток-мишеней 10:1. Для нормализации определяли максимальную степень лизиса клеток-мишеней (=100%) при инкубации клеток-мишеней с раствором Triton-X-100 в конечной концентрации 1%. Минимальный лизис (=0%) соответствовал инкубации клеток-мишеней вместе с эффекторными клеткам в отсутствие каких-либо конструкций или антител.
После инкубации в течение 70 ч при 37°С, 5% CO2, в надосадочной жидкости измеряли концентрацию ЛДГ, выделившейся из апоптотических/некротических клеток-мишеней, с использованием набора для определения ЛДГ (Roche Applied Science, #11644793001), согласно инструкциям производителя.
Как показано на Фиг.13, присутствие биспецифической конструкция вызывало зависящее от концентрации высвобождение ЛДГ из клеток-мишеней, сравнимое с наблюдавшимся в присутствии положительного контроля "Т Cell Stim with ConA".
Исследование высвобождения ЛДГ в присутствии биспецифической конструкции кpoccFab мыши (CD3)-Fab (MCSP)-Fab (MCSP)
У очищенного кpoccFab мыши (CD3)-Fab (MCSP)-Fab (MCSP), направленного против CD3 мыши и MCSP человека, исследовали способность индуцировать опосредованный Т-клетками апоптоз опухолевых клеток-мишеней при образовании поперечных связей конструкциями за счет связывания обоих нацеленных на антигены компонентов с соответствующими антигенами на клетках.
Вкратце, опухолевые клетки-мишени B16/F10-huMCSP Fluc2 клон 48, экспрессирующие huMCSP, собирали с использованием буфера для диссоциации клеток, отмывали и ресуспендировали в среде RPMI1640, содержащей 1х заменимые аминокислоты, 10 мМ Hepes, 50 мкМ 2-b-ME и 1 мМ пируват натрия.
В круглодонные 96-луночные планшеты высаживали по 20000 клеток на лунку и добавляли соответствующее антитело для достижения конечной концентрации 50 нМ. Биспецифические конструкции и различные контрольные IgG доводили до одинаковой молярности.
В качестве эффекторных клеток использовали общую популяцию Т-клеток мыши, выделенных из спленоцитов (мышей C57BL/6) при итоговом соотношении эффекторных клеток и клеток-мишеней 10:1. Для оценки степени гиперактивации Т-клеток мыши в отсутствие клеток-мишеней, в контрольные лунки, содержавшие биспецифическую конструкцию в концентрации 50 нМ, аналогичным образом высевали Т-клетки.
Для нормализации определяли максимальную степень лизиса клеток-мишеней (=100%) при инкубации клеток-мишеней с раствором Triton-X-100 в конечной концентрации 1%. Минимальный лизис (=0%) соответствовал инкубации клеток-мишеней вместе с эффекторными клеткам в отсутствие каких-либо конструкций или антител.
После инкубации в течение ночи на протяжении 70 ч при 37°С, 5% CO2, в надосадочной жидкости измеряли концентрацию ЛДГ, выделившейся из апоптотических/некротических клеток-мишеней, с использованием набора для определения ЛДГ (Roche Applied Science, #11644793001), согласно инструкциям производителя.
Как показано на Фиг.14, присутствие биспецифической конструкции вызывало значительное высвобождение ЛДГ из клеток-мишеней. В отсутствие клеток-мишеней наблюдалось только слабое повышение ЛДГ (отражающее гиперактивацию Т-клеток) по сравнению с необработанными Т-клетками мыши, которые инкубировали вместе с клетками-мишенями. Ни один из контрольных IgG не вызывал высвобождение ЛДГ из клеток-мишеней.
Пример 8: Исследование высвобождения цитокинов (цитометрический анализ с использованием мультиплексной технологии)
Для оценки секреции различных цитокинов de novo при активации Т клеток с использованием биспецифических конструкций, направленных против CD3, в присутствии или в отсутствие клеток-мишеней, выделяли МКПК человека из лейкоконцентрата и в круглодонные 96-луночные планшеты высаживали по 0,3 млн клеток на лунку. В альтернативном случае в лунки глубокодонных 96-луночных планшетов помещали по 280 мкл цельной крови здоровых доноров.
Опухолевые клетки-мишени (например, клетки MDA-MB-435 в случае биспецифических конструкций, направленных против CD3-MCSP) добавляли до достижения итогового соотношения эффекторные клетки/клетки-мишени 10:1. Биспецифические конструкции и контроли добавляли в соответствии с описанием. После инкубации на протяжении не более 24 ч при 37°С, 5% CO2, исследуемые планшеты центрифугировали в течение 5 мин при 350 g и переносили надосадочную жидкость в новые глубокодонные 96-луночные планшеты для последующего анализа.
Цитометрический анализ с использованием мультиплексной технологии (СВА) проводили согласно инструкциям производителя для FACS Cantoll, с использованием комбинации следующих наборов СВА Flex Sets для определения: гранзима В человека (BD 560304), ИФН-γ человека (BD 558269), ФНО человека (BD 558273), IL-10 человека (BD 558274), IL-6 человека (BD 558276), IL-4 человека(BD 558272).
Исследование высвобождения цитокинов с применением биспецифических конструкций, направленных против MCSP-CD3
У следующих биспецифических конструкций, направленных против MCSP человека и CD3 человека исследовали способность индуцировать опосредованную Т-клетками секрецию цитокинов de novo в присутствии (А, В) или в отсутствие (С, D) опухолевых клеток-мишеней: "Fab (MCSP)- Fab (MCSP)-кpoccFab (CDS) и референтной молекулы (scFv)2 (антиМСSР/анти huCDSe).
Вкратце, в каждую лунку 96-луночного планшета помещали по 280 мкл цельной крови здоровых доноров. Добавляли по 30000 опухолевых клеток-мишеней Colo-38, экспрессирующих MCSP человека, а также различные биспецифические конструкции и контрольные IgG в конечной концентрации 1 нМ. Клетки инкубировали в течение 24 ч при 37°С, 5% CO2 и затем центрифугировали в течение 5 мин при 350×g. Надосадочную жидкость переносили в новые глубокодонные 96-луночные планшеты для последующего анализа.
Цитометрический анализ с использованием мультиплексной технологии (СВА) выполняли согласно инструкциям производителя для FACS Cantoll, используя комбинацию следующих наборов СВА Flex Sets для определения: гранзима В человека (BD 560304), ИФН-γ человека (BD 558269), ФНО человека (BD 558273), IL-10 человека (BD 558274), IL-6 человека (BD 558276), IL-4 человека (BD 558272
На Фиг.15 показаны уровни различных цитокинов, измеренные в супернатанте, после инкубации цельной крови с различными биспецифическими конструкциями, направленными против CD3-MCSP, в концентрации 1 нМ (Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) и (scFv)2 (антиМСSР/антиhuCD3e)) в присутствии (А, В) или отсутствии (C,D) опухолевых клеток Colo-38 в течение 24 часов. В каждую лунку 96-луночного планшета добавляли по 280 мкл цельной крови и 30000 клеток Colo-38, в соответствии с описанием.
Основным цитокином, секретируемым при активации Т-клеток в присутствии опухолевых клеток Colo-38, был IL-6, затем следовал ИФН-гамма. Кроме того, при активации Т-клеток в присутствии клеток-мишеней наблюдалось значительное повышение уровня гранзима В. В целом, конструкция (scFv)2 (антиМСSР/антиhuCD3e) вызывала несколько более выраженное повышение уровней ФНО и ИФН-гамма, а также гранзима В в присутствии клеток-мишеней (А и В) по сравнению с другими биспецифическими конструкциями.
При активации Т-клеток с использованием биспецифических конструкций в присутствии (или в отсутствие) клеток-мишеней не наблюдалось существенной секреции Th2 цитокинов (IL-10 и IL-4).
В данном исследовании также наблюдалась слабая секреция ИФН-гамма, индуцированная конструкцией Fab (MCSP)-Fab (MCSP)-кpoccFab (CD3) в отсутствие клеток-мишеней.
Исследование высвобождения цитокинов с использованием биспецифических конструкций, направленных против MCSP и мышиного CD3
У очищенных биспецифических молекул, направленных против huMCSP-muCD3, представляющих собой кpoccFab мыши (CD3)-Fab (MCSP)- Fab (MCSP), при помощи проточной цитометрии исследовали способность повышать уровень поздних маркеров активации CD25 на поверхности CD8+ Т-клеток в присутствии опухолевых клеток, экспрессирующих MCSP человека.
Вкратце, положительные по MCSP опухолевые клетки B16/F10-huMCSP Fluc2 клон 48 собирали с помощью буфера для диссоциации клеток, подсчитывали и проверяли их жизнеспособность. Концентрацию клеток доводили до 0,3×106 (жизнеспособных) клеток на мл среды RPMI1640 (содержавшей 1х заменимые аминокислоты, 10 мМ Hepes, 50 мкМ 2-b-ME, 1 мМ пируват натрия), 100 мкл этой клеточной суспензии раскапывали по лункам 96-луночного планшета (в соответствии с описанием). В лунки, содержавшие клетки, добавляли биспецифические конструкции (в разведении) по 50 мкл до достижения конечной концентрации 50 нМ. Эффекторные Т-клетки мыши выделяли из спленоцитов (мышей C57BL/6) и доводили концентрацию до 3×106 (жизнеспособных) клеток на мл среды AIM-V. В каждую лунку аналитического планшета добавляли по 50 мкл этой суспензии (см. выше) для получения итогового соотношения эффекторные клетки: клетки-мишени 10:1. Для анализа способности биспецифической конструкции активировать Т-клетки только в присутствии клеток-мишеней, экспрессирующих huMCSP, в исследование включали лунки, содержавшие соответствующие биспецифические молекулы в концентрации 50 нМ, а также эффекторные Т-клетки, но не содержавшие клеток-мишеней.
После инкубации в течение 70 часов при 37°С, 5% CO2, клетки центрифугировали (5 мин, 350×g) и дважды отмывали PBS, содержащим 0,1% БСА, в объеме 150 мкл/лунку.
Поверхностное окрашивание на CD8a (IgG2a крысы; клон 53-6.7; BioLegend #100712) и CD25 (IgG2b крысы; клон 3С7; BD #553075) выполняли согласно рекомендациям производителя. Клетки дважды отмывали PBS, содержащим 0,1% БСА, в объеме 150 мкл/лунку и фиксировали в течение 15 мин при 4°С, с использованием 100 мкл/лунку фиксирующего буфера (BD ##554655).
После центрифугирования образцы ресуспендировали в PBS с 0,1% БСА в объеме 200 мкл/лунку и анализировали на приборе FACS Cantoll (программное обеспечение FACS Diva).
На Фиг.16 показано, что конструкция кpoccFab мыши (CD3)-Fab (MCSP)-Fab (MCSP) индуцирует повышение уровня CD25 только в присутствии клеток-мишеней.
Пример 9: Экспрессия поверхностных маркеров активации на первичных человеческих Т-клетках при использовании биспецифических конструкций
Для проверки возможности специфической активации Т-клеток при связывании биспецифических конструкций, направленных против CD3, исключительно в присутствии опухолевых клеток-мишеней, первичные МКПК человека (выделенные, как описано выше) инкубировали с биспецифическими конструкциями в указанных концентрациях в течение по меньшей мере 24 ч в присутствии или в отсутствие клеток-мишеней, положительных по опухолевым антигенам.
Вкратце, в лунки плоскодонного 96-луночного планшета, содержащие положительные по huMCSP клетки-мишени (опухолевые клетки MV-3) или среду, высаживали по 0,3 миллиона первичных МКПК человека. Итоговое соотношение эффекторных клеток и клеток-мишеней (Е:Т) составляло 10:1. Клетки инкубировали с биспецифическими конструкциями (Fab (MCSP)-кpoccFab (CD3), обозначенных "1+1 без Fc", и референтной молекулы (scFv)2 (антиМСSР/антиhuCD3e) (обозначенной "(scFv)2" направленными против CD3-MCSP, в указанных концентрациях в течение указанного времени инкубации при 37°С, 5% CO2. Эффекторные клетки окрашивали на CD8 и ранний маркер активации CD69 или поздний маркер активации CD25 и анализировали на FACS Cantoll.
На Фиг.20 показаны результаты этого эксперимента.
Следует понимать, что предпочтительные воплощения изобретения, приведенные и описанные в данном документе, не ограничивают объема изобретения, которое может иметь многие другие воплощения, находящиеся в рамках прилагаемой формулы.
Последовательности
Следует понимать, что предпочтительные воплощения изобретения, приведенные и описанные в данном документе, не ограничивают объема изобретения, которое может иметь многие другие воплощения, находящиеся в рамках прилагаемой формулы. Условные обозначения: GA201=антитело к рецептору эпидермального фактора роста EGFR, 3F2=антитело к белку активации фибробластов FAP, CH1A1A=антитело к раково-эмбриональному антигену СЕА.
Последовательности белков
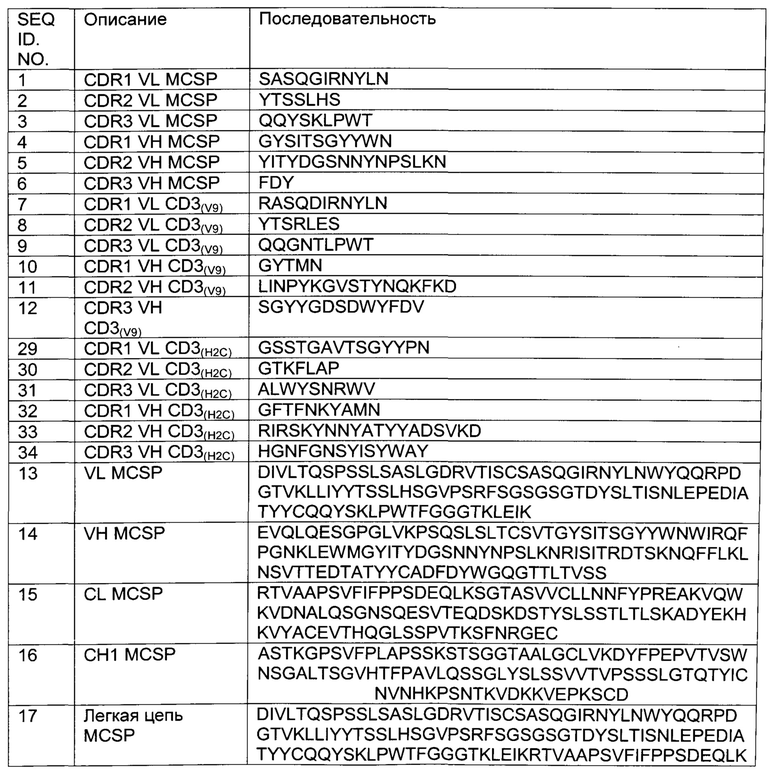
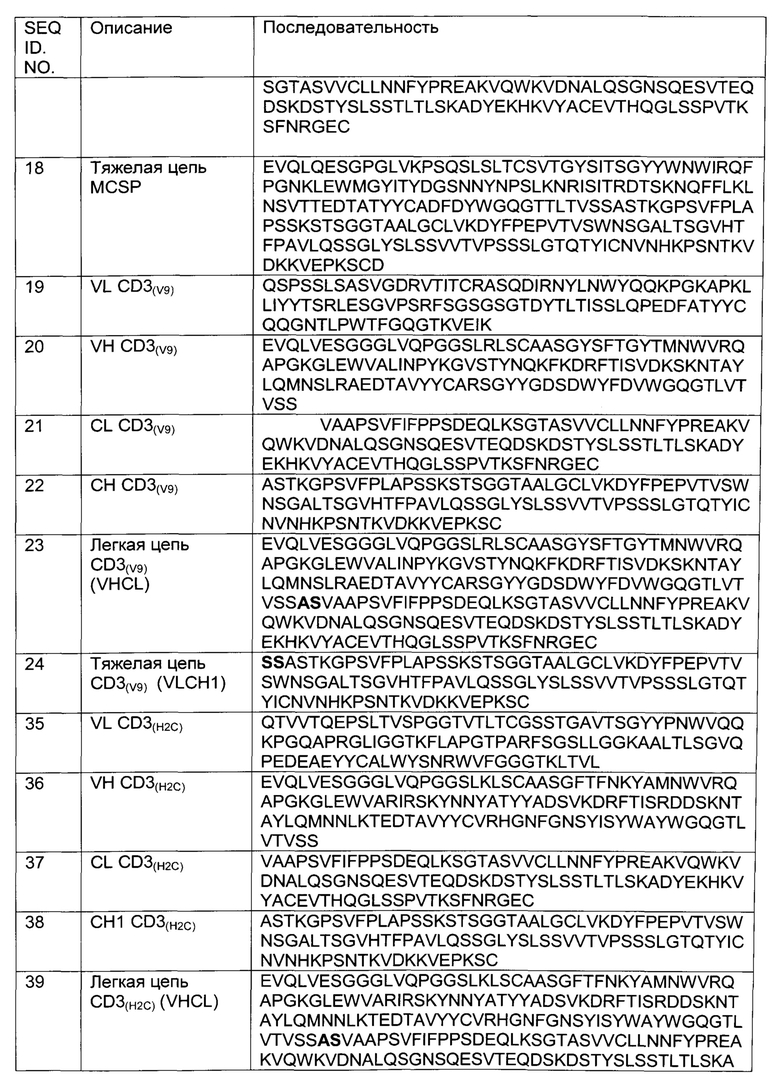

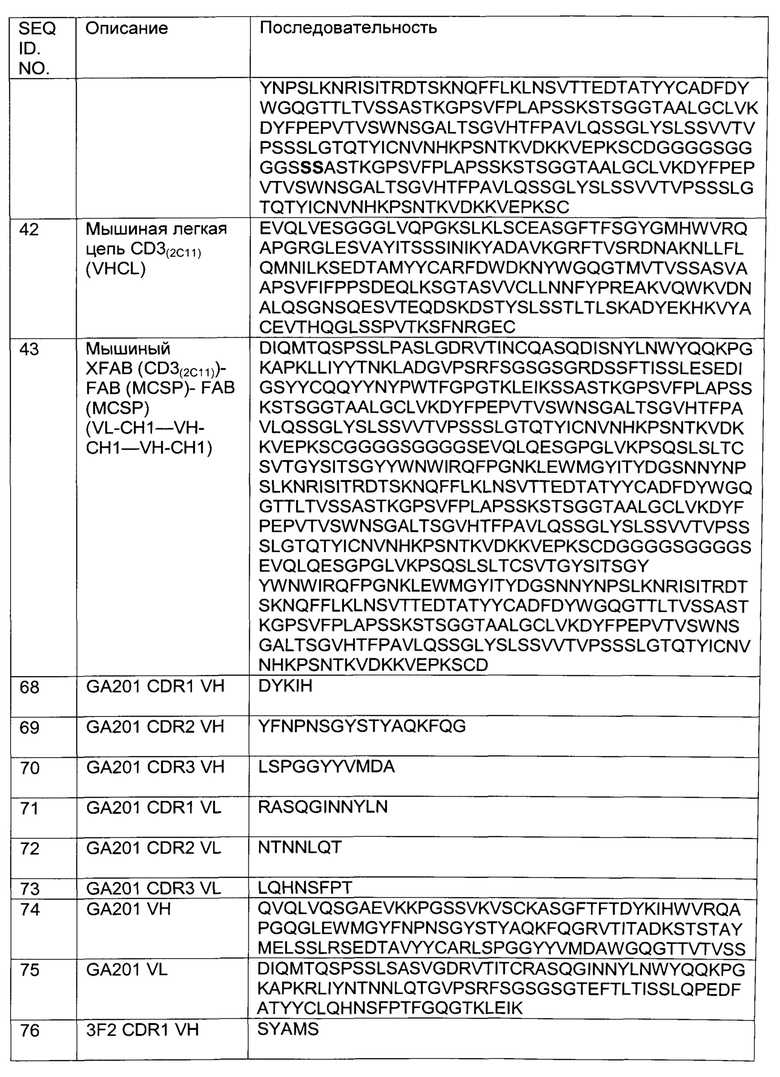


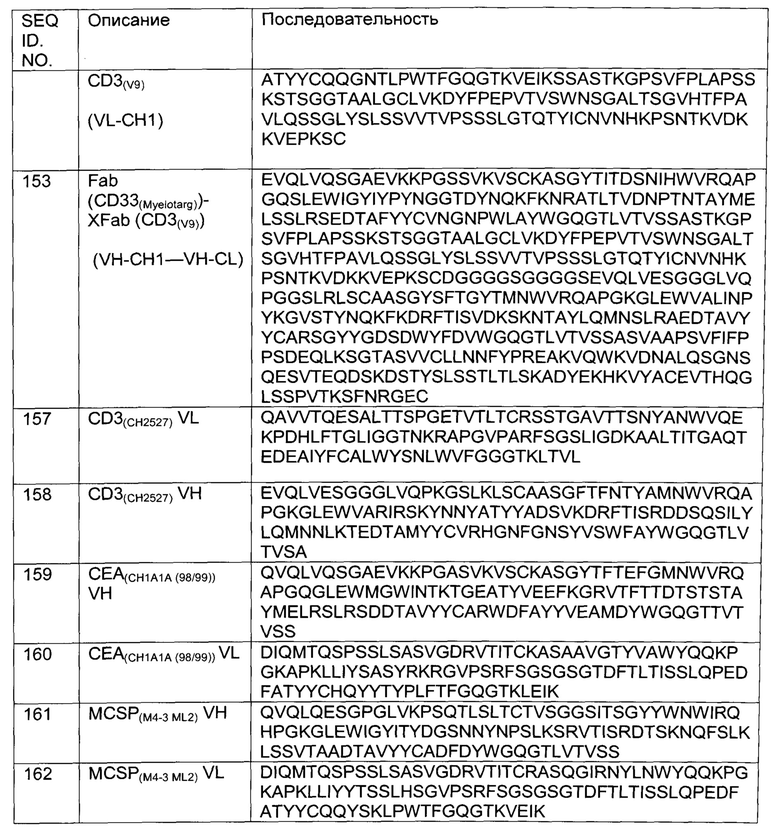
Последовательности ДНК

















Следует понимать, что предпочтительные воплощения изобретения, приведенные и описанные в данном документе, не ограничивают объема изобретения, которое может иметь многие другие воплощения, находящиеся в рамках прилагаемой формулы.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АНТИГЕНАМ, АКТИВИРУЮЩИМ Т-КЛЕТКИ, И ОПУХОЛЕВОМУ АНТИГЕНУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2605390C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2012 |
|
RU2650775C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| АНТИТЕЛО, НАЦЕЛИВАЮЩЕЕСЯ НА CD3, БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2808138C1 |
| СПОСОБ ОТБОРА И ПОЛУЧЕНИЯ СЕЛЕКТИВНЫХ И МУЛЬТИСПЕЦИФИЧЕСКИХ ТЕРАПЕВТИЧЕСКИХ МОЛЕКУЛ С ЗАДАННЫМИ СВОЙСТВАМИ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ НАЦЕЛИВАЮЩИЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2644263C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2011 |
|
RU2573588C2 |
| PSMA×CD3 БИСПЕЦИФИЧЕСКОЕ ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2009 |
|
RU2559531C2 |
| АНТИ-СD3-АНТИТЕЛО И МОЛЕКУЛА, СОДЕРЖАЩАЯ ЭТО АНТИТЕЛО | 2017 |
|
RU2790326C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИТИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2799436C1 |



































































































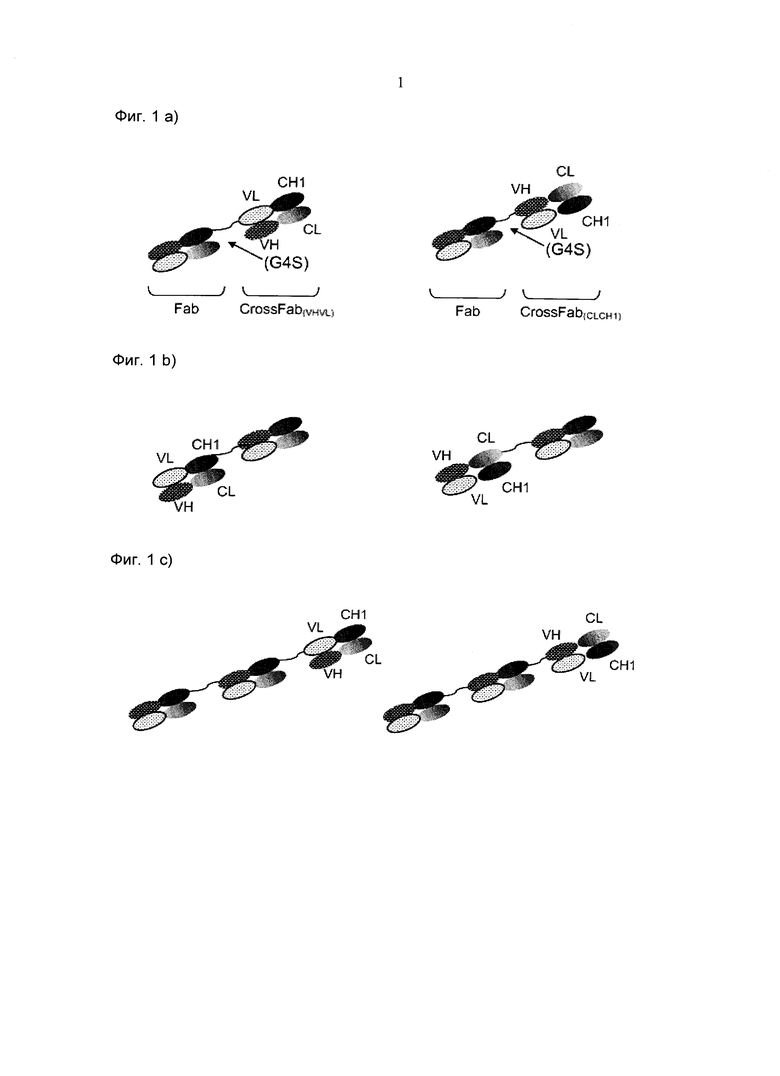
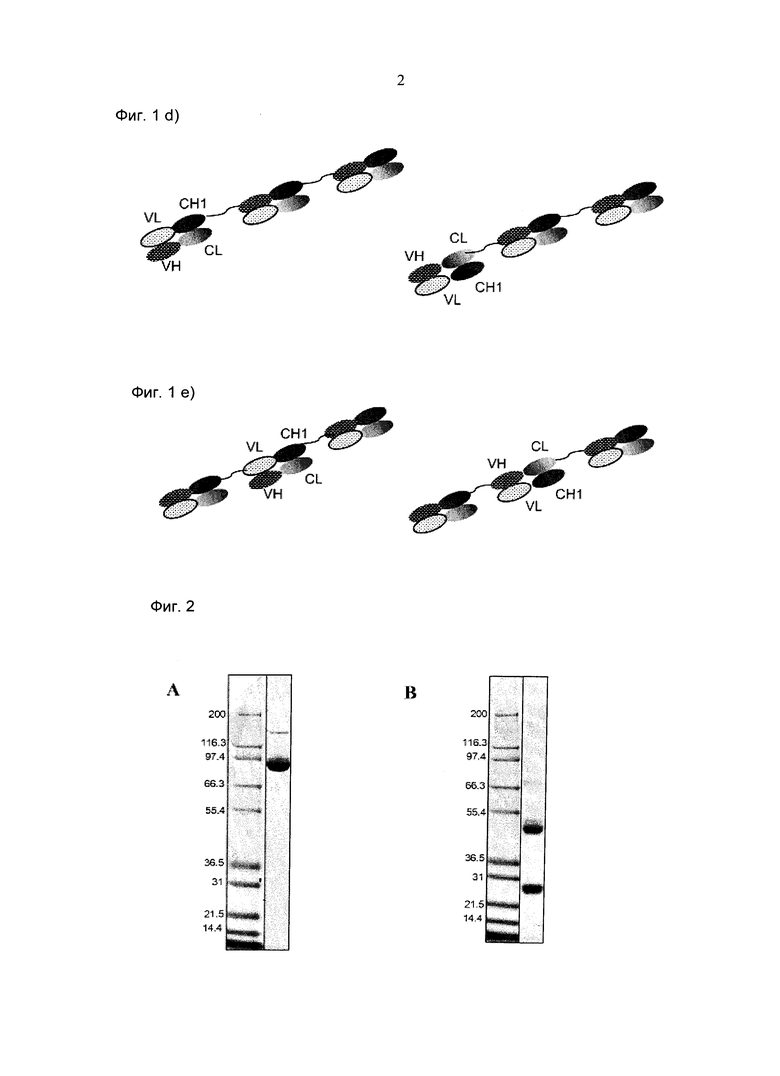
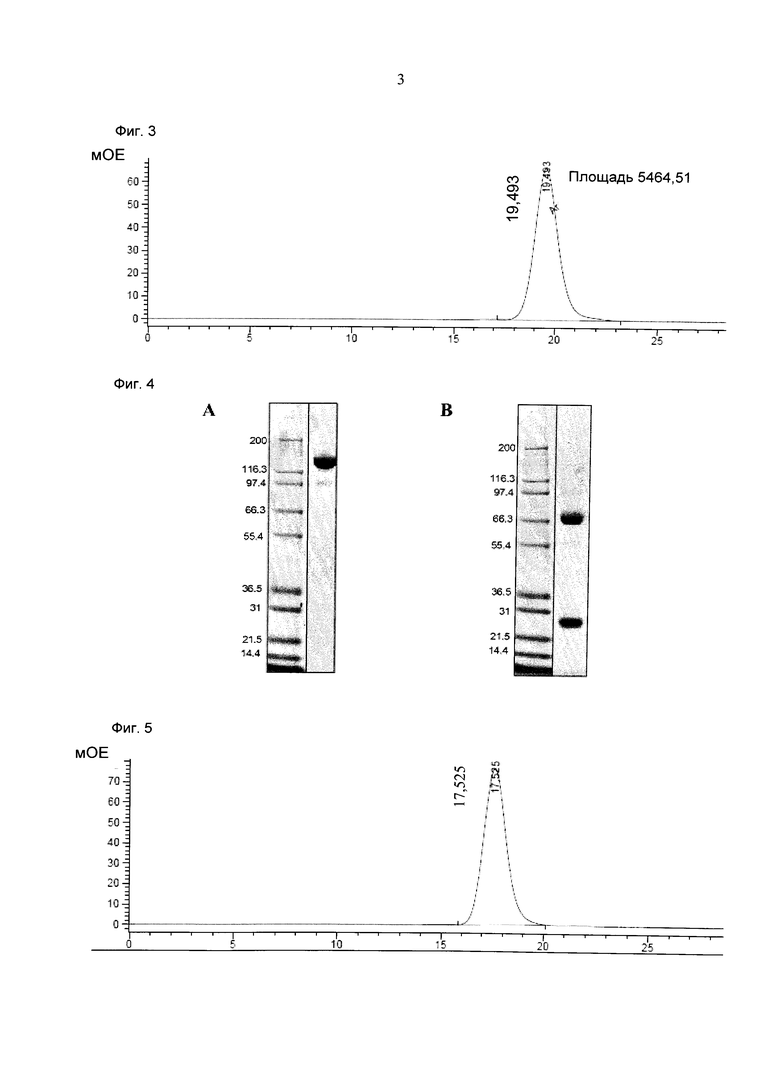
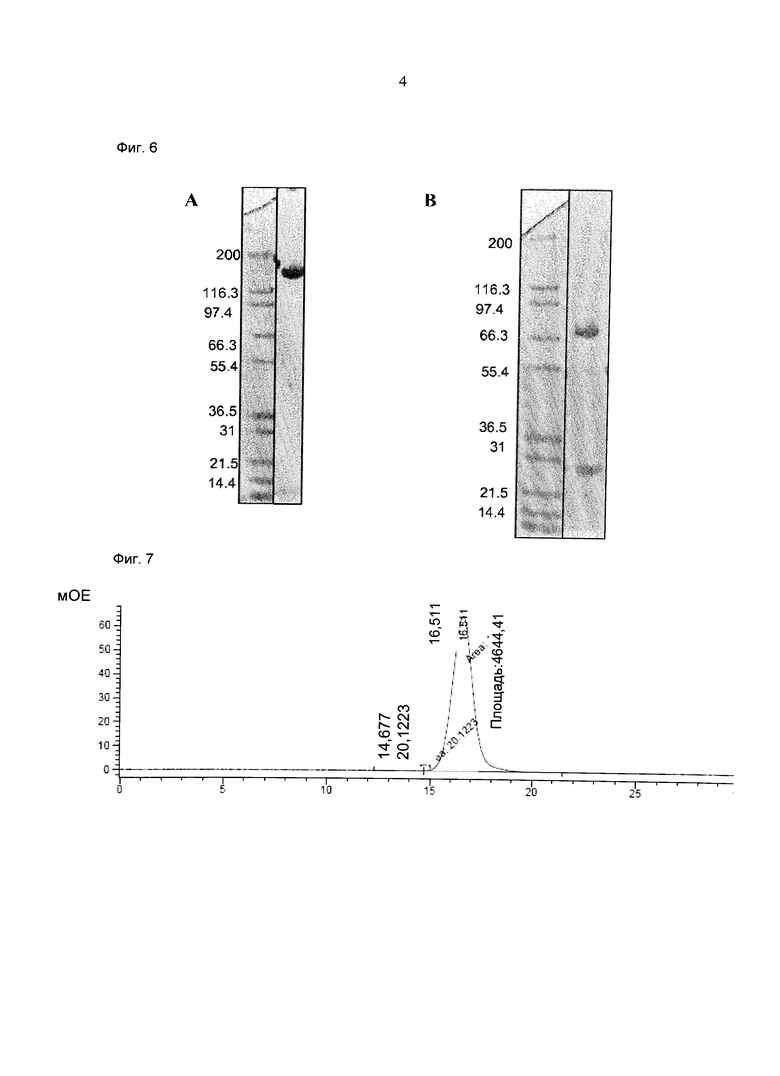
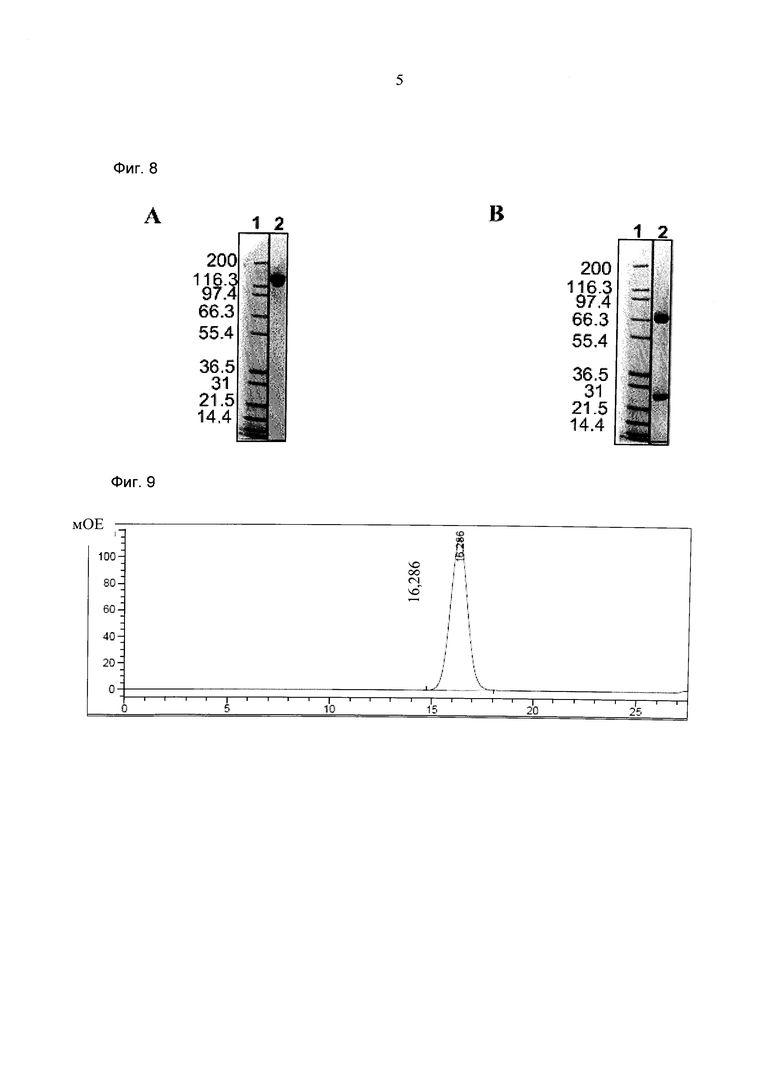
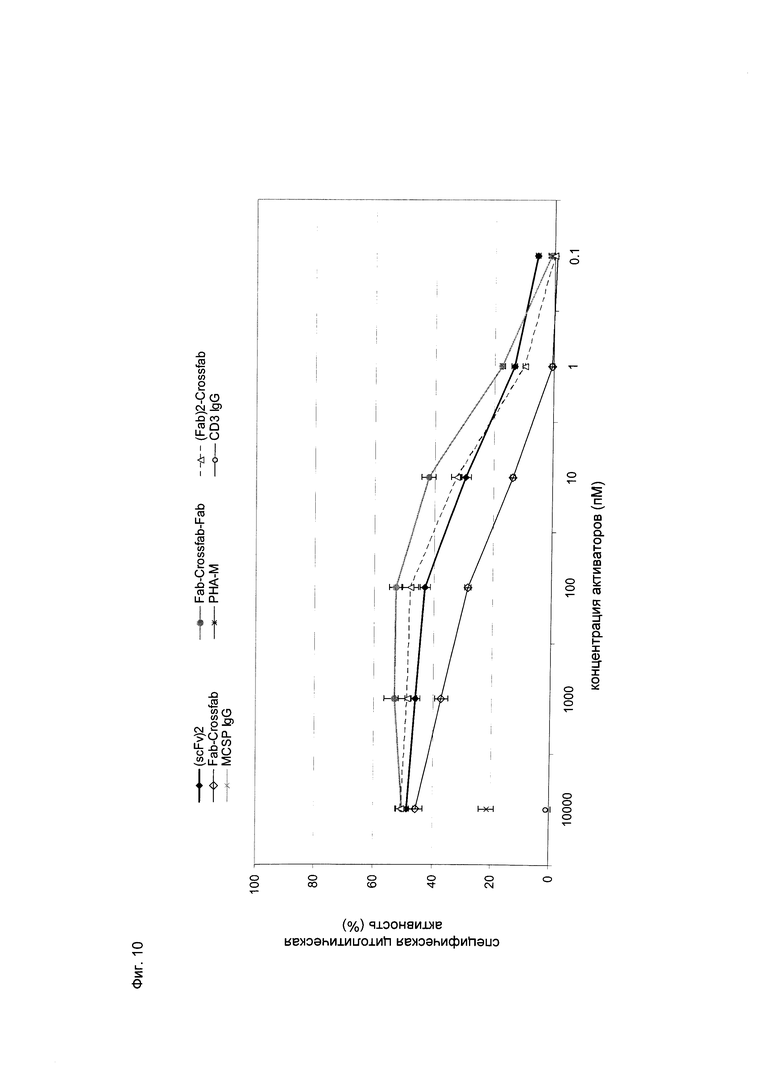
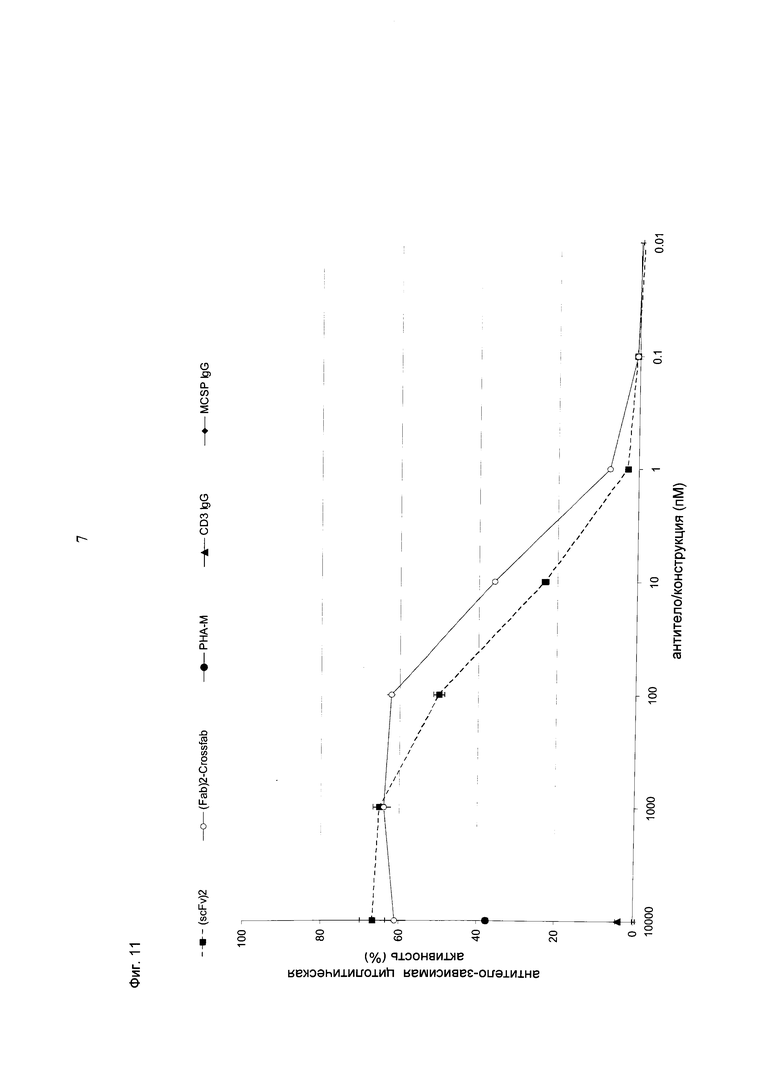
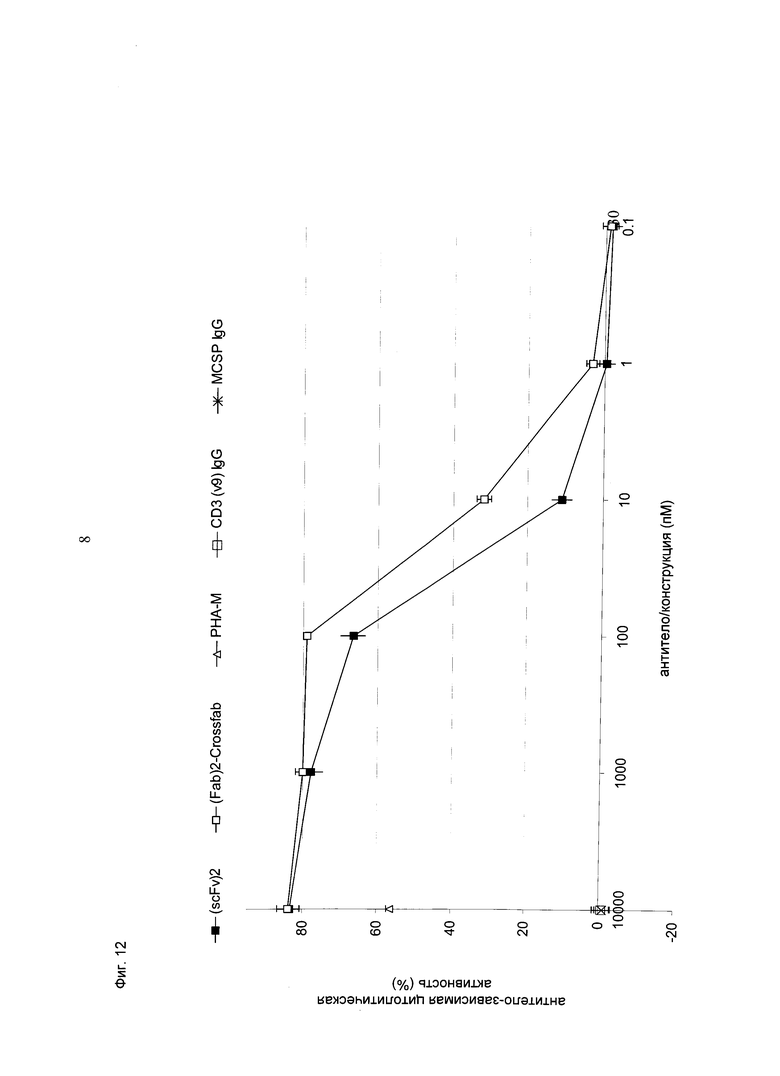
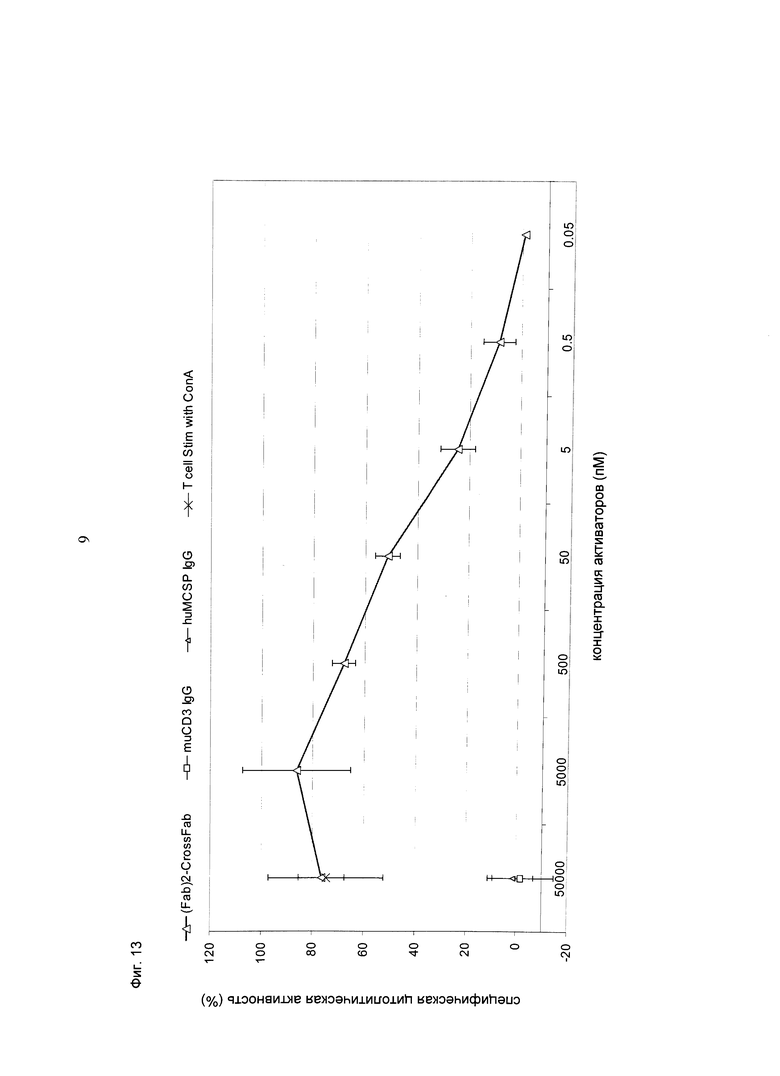
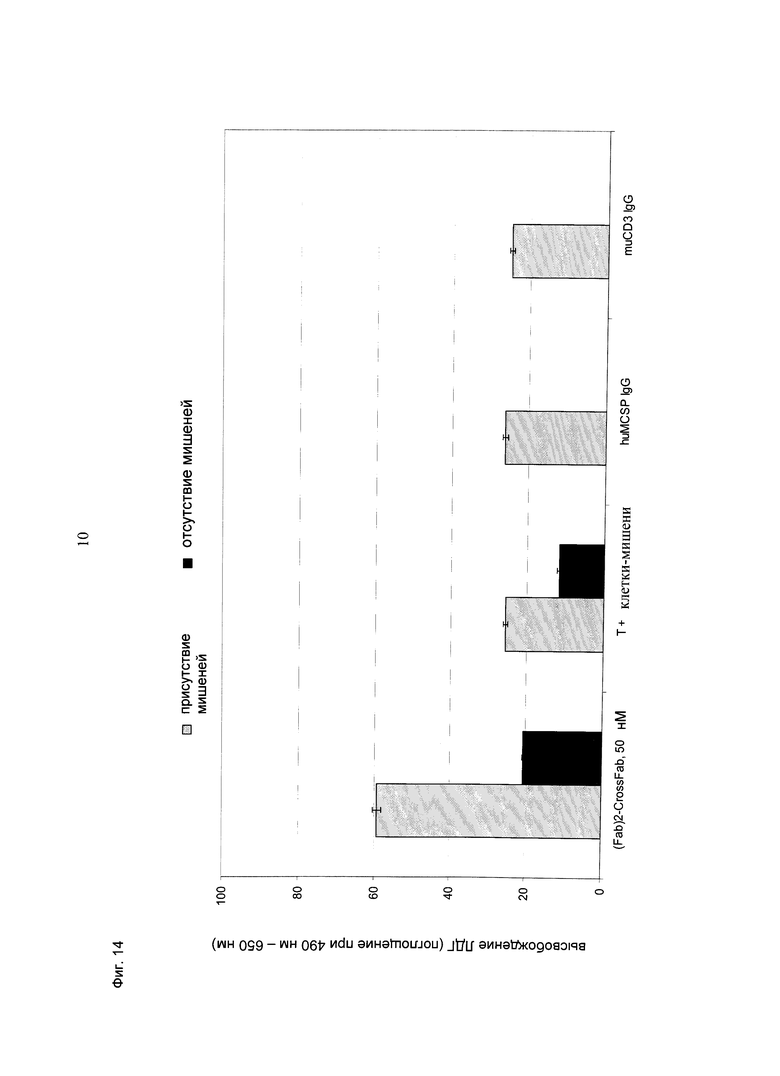
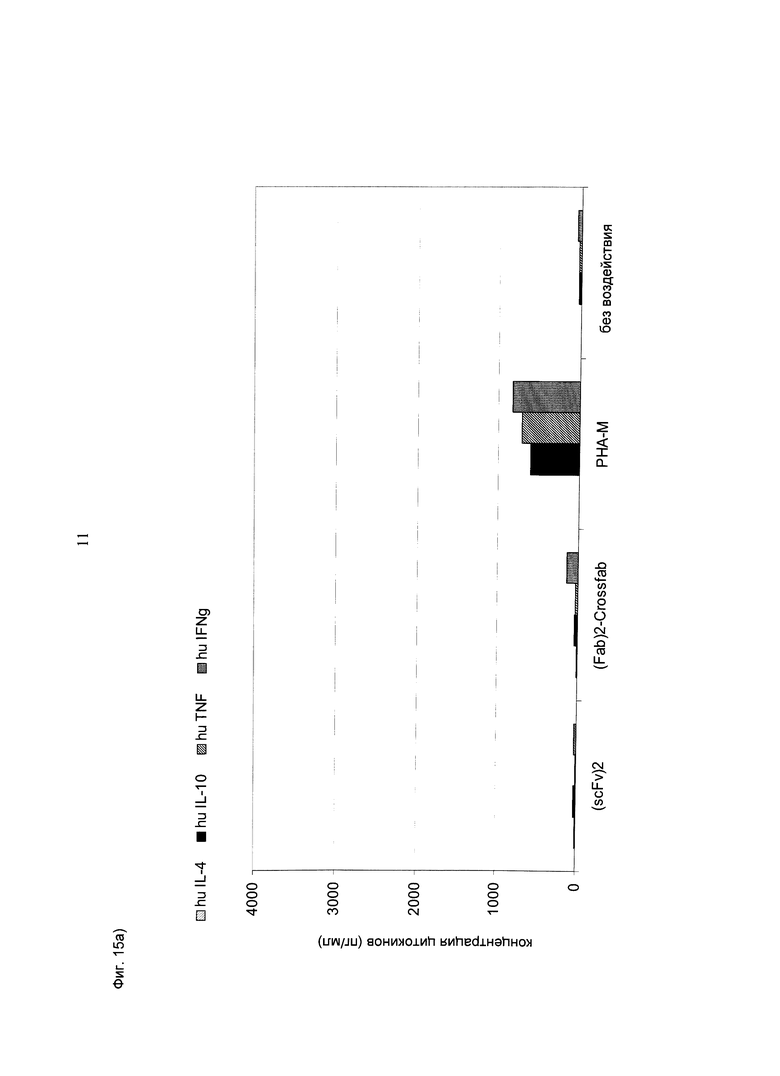
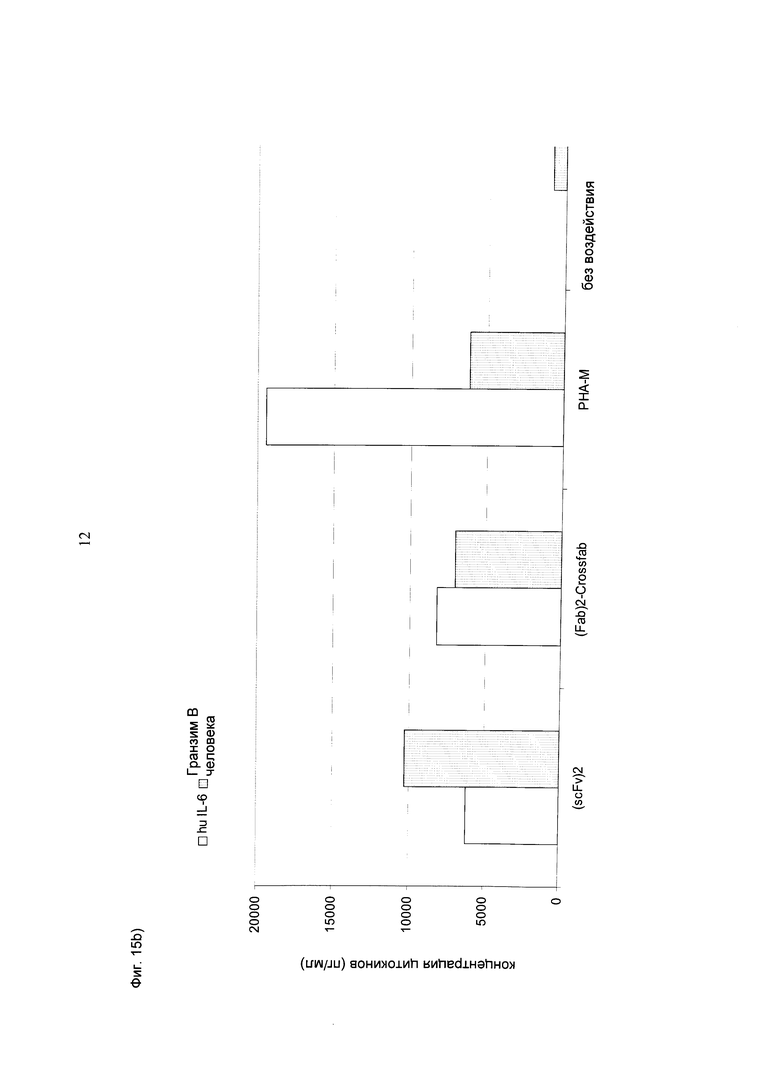


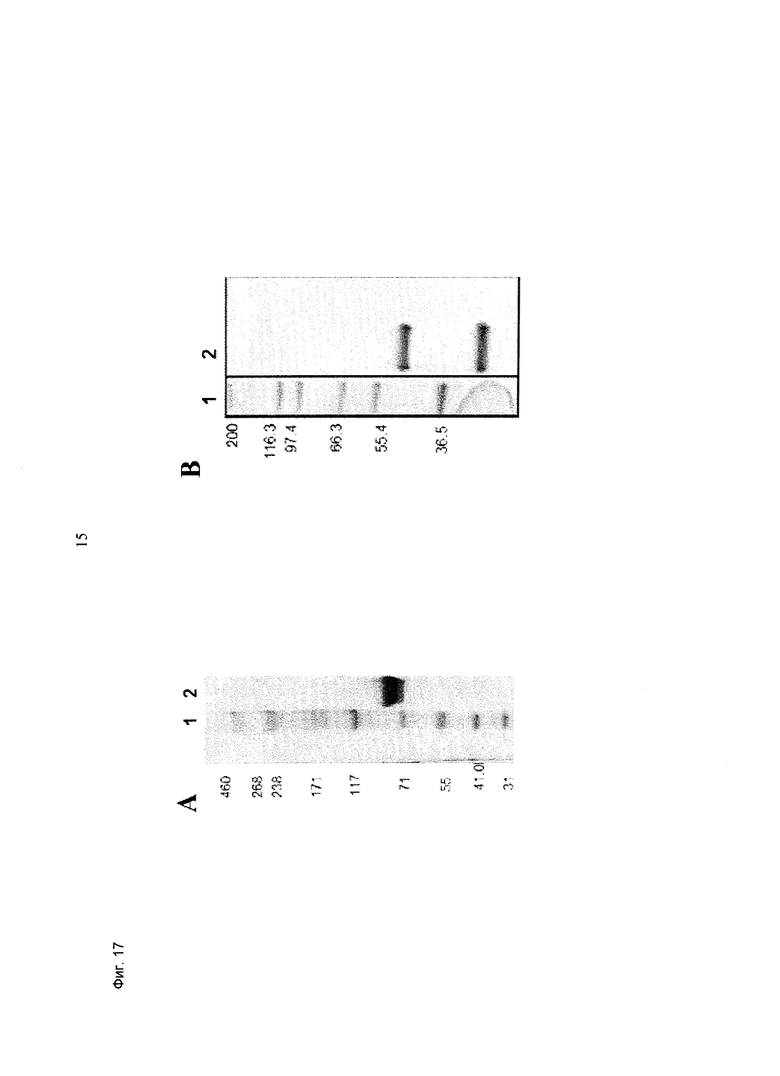
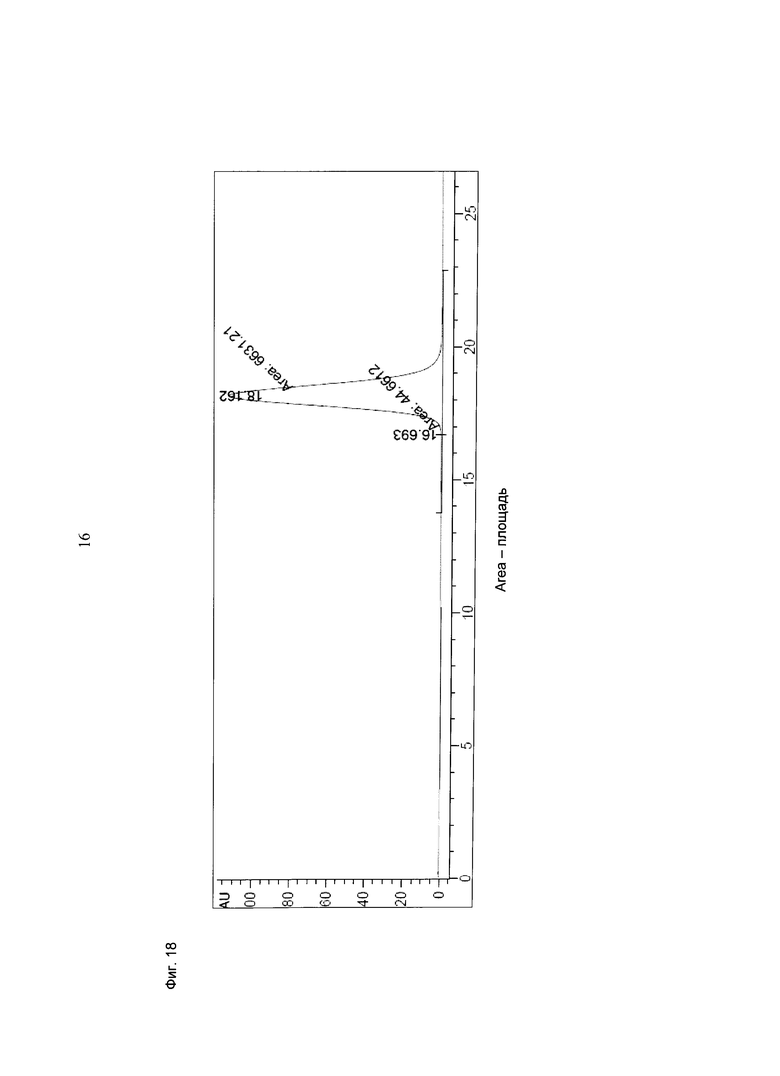
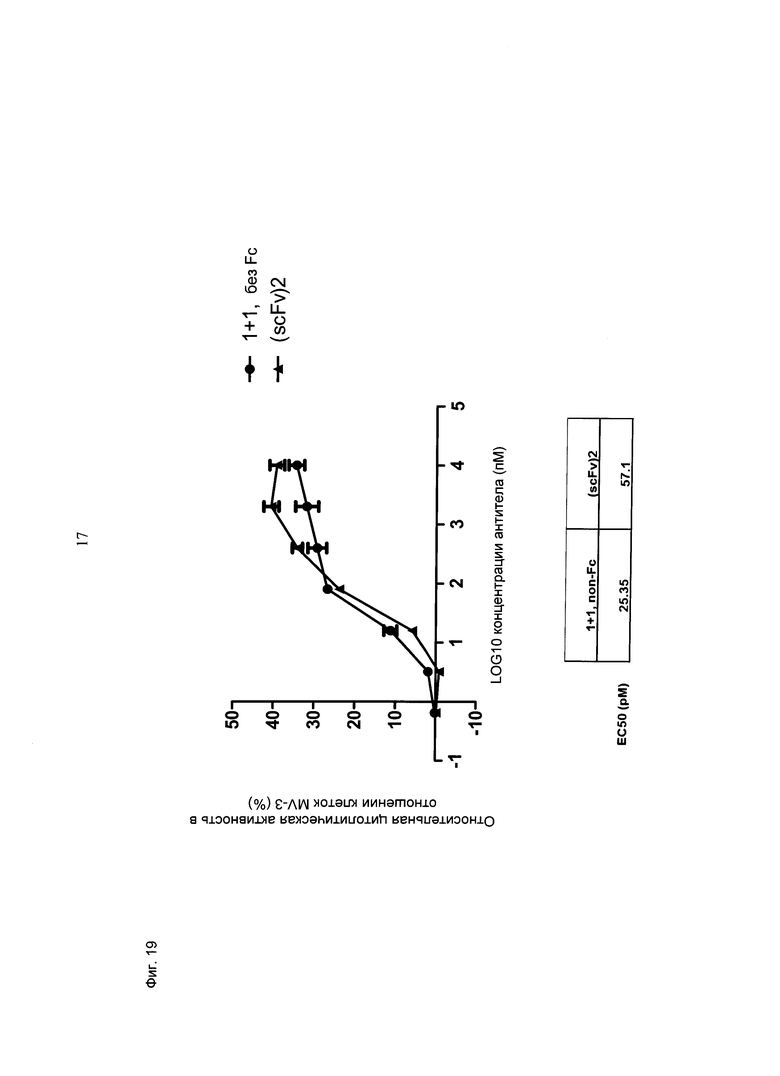
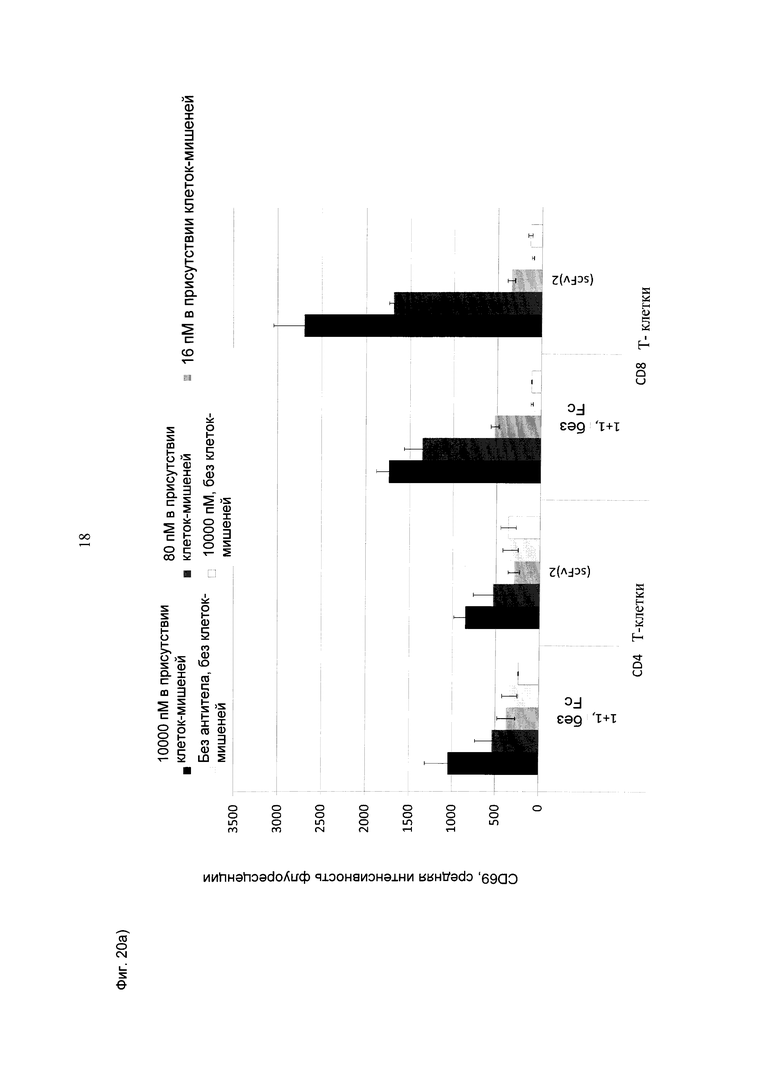
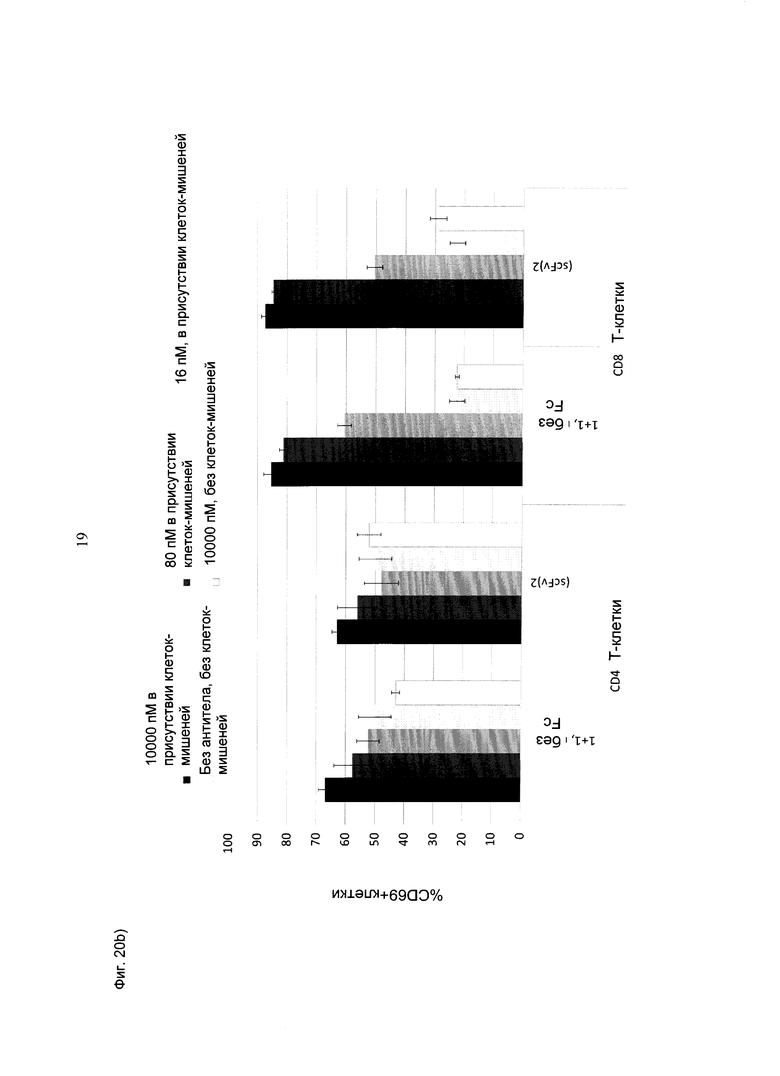

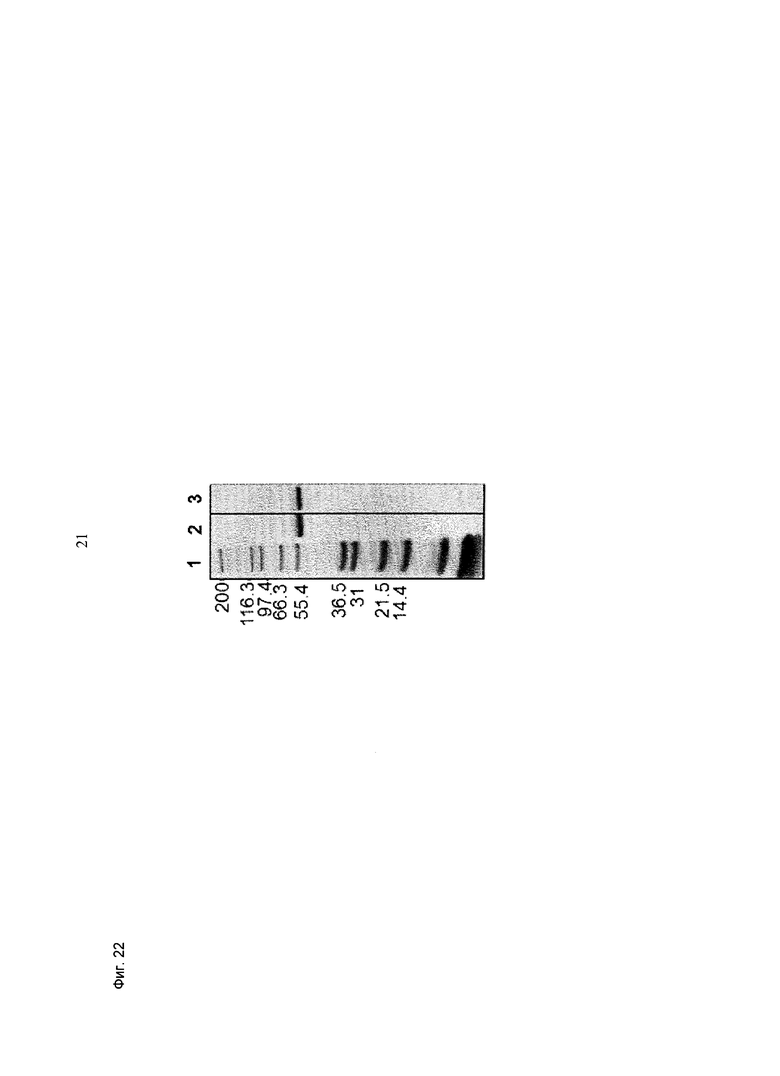
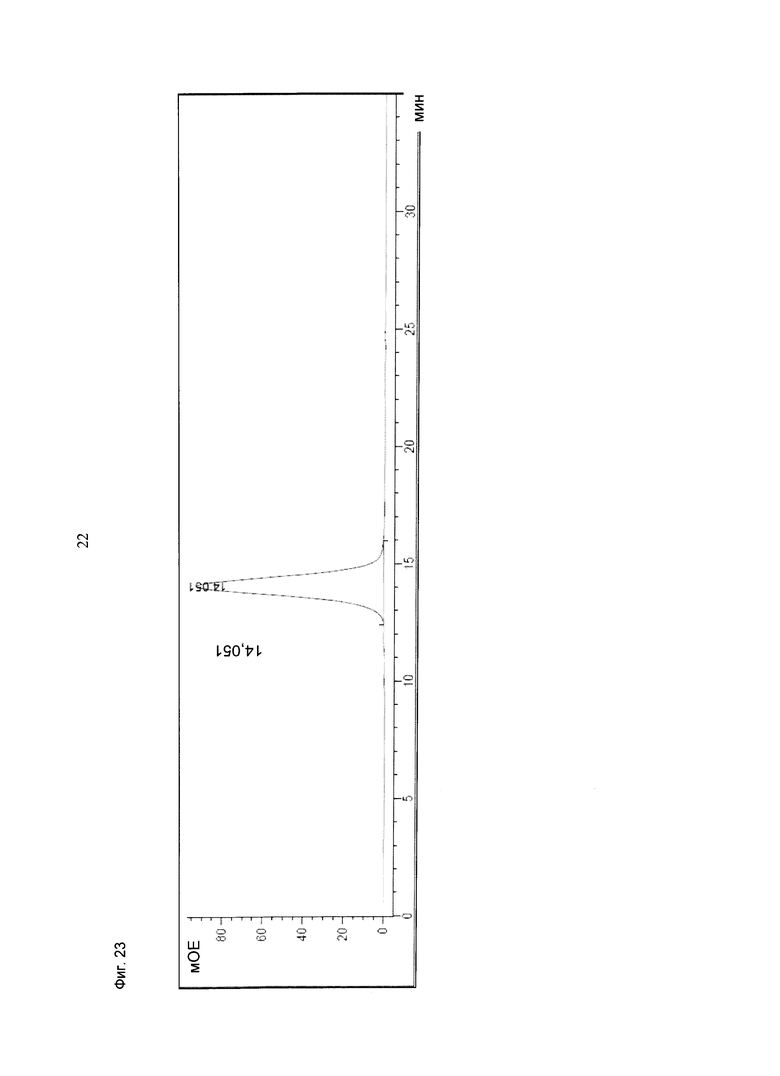
Группа изобретений относится к биотехнологии, в частности к биспецифическим антителам и их применению. Биспецифические антитела содержат два Fab фрагмента, причем первый Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого антигена, и второй Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении второго антигена, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента, причем биспецифическое антитело лишено Fc домена. Предложены также способ и клетка-хозяин для их получения, а также фармацевтическая композиция и иммуноконъюгат для лечения рака, содержащие указанные биспецифические антитела. Преимущество заявленных биспецифических антител состоит в пониженной агрегации фрагментов, повышенной стабильности и в повышении эффективности связывания с антигеном. Предлагаемые иммуноконъюгат и фармацевтическая композиция позволяют эффективно проводить терапию рака. 5 н. и 8 з.п. ф-лы, 23 ил., 2 табл., 9 пр.
1. Биспецифическое антитело, содержащее два Fab фрагмента, где первый Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого антигена, и второй Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении второго антигена, где произведен обмен вариабельными участками или константными участками между тяжелой и легкой цепями второго Fab фрагмента, где биспецифическое антитело лишено Fc домена таким образом, что константный участок тяжелой цепи состоит только из одного или более СН1 доменов, и где Fab фрагменты соединены посредством пептидного линкера.
2. Биспецифическое антитело по п. 1, дополнительно содержащее третий Fab фрагмент.
3. Биспецифическое антитело по п. 2, где третий Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого или второго антигена.
4. Биспецифическое антитело по п. 2, где третий Fab фрагмент содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении первого антигена.
5. Биспецифическое антитело по п. 2, где третий Fab фрагмент соединен с первым Fab фрагментом.
6. Биспецифическое антитело по п. 5, где С-конец третьего Fab фрагмента соединен с N-концом первого Fab фрагмента.
7. Биспецифическое антитело по п. 2, где третий Fab фрагмент соединен со вторым Fab фрагментом.
8. Биспецифическое антитело по п. 7, где N-конец третьего Fab фрагмента соединен с С-концом первого Fab фрагмента.
9. Биспецифическое антитело по п. 1, где пептидный линкер представляет собой линкер (G4S)2.
10. Фармацевтическая композиция для лечения рака, содержащая эффективное количество биспецифического антитела по пп. 1-9 и фармацевтически приемлемый носитель.
11. Прокариотическая или эукариотическая клетка-хозяин для получения биспецифического антитела по п. 1, содержащая векторы, включающие молекулы нуклеиновой кислоты, кодирующие легкие цепи и тяжелые цепи биспецифического антитела по любому из пп. 1-9.
12. Способ получения антитела, включающий культивирование клетки-хозяина по п. 11, приводящий к получению антитела по п. 1.
13. Иммуноконъюгат для лечения рака, содержащий антитело по любому из пп. 1-9 и цитотоксический агент.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| МОДИФИЦИРОВАННОЕ АГОНИСТИЧЕСКОЕ АНТИТЕЛО | 2001 |
|
RU2295537C2 |

