Область техники, к которой относится изобретение
Настоящее изобретение относится к области усовершенствованных анти-IgE антител и антигенсвязывающих фрагментов, а также к их композициям, которые нацелены на IgE, например, для применения в лечении заболеваний, вызываемых IgE (таких как аллергические реакции или некоторые аутоиммунные реакции), и, в частности, заболеваний, вызываемых взаимодействием IgE и рецептора FcεRI. В частности, настоящее изобретение относится к усовершенствованным анти-IgE антителам и антигенсвязывающим фрагментам, относящимся к новым мутантам омализумаба (ксолар®). Усовершенствованные анти-IgE антитела и антигенсвязывающие фрагменты по изобретению могут иметь усовершенствованную аффинность для IgE и/или усовершенствованное взаимодействие с Cε2-доменом IgE, и/или усовершенствованный модифицированный эпитоп на IgE (например, дополнительно включающий Cε2-домен IgE), и/или способность вызывать диссоциацию IgE от рецептора FcεRI при фармацевтически используемых концентрациях. В одном аспекте раскрыты усовершенствованные или новые способы лечения IgE-опосредованных заболеваний, в которых IgE является мишенью (например, свободный IgE и/или IgE в комплексе с рецептором FcεRI).
Предпосылки создания изобретения
IgE является представителем семейства иммуноглобулинов, который опосредует аллергические реакции, такие как астма, пищевая аллергия, гиперчувствительность 1 типа и семейное воспаление носовых пазух, которые распространены повсеместно. IgE секретируется B-клетками и экспрессируется на их поверхности. IgE, синтезируемый B-клетками, заякорен на B-клеточной мембране за счет трансмембранного домена, связанного с последовательностью зрелого IgE короткой мембраносвязанной областью. IgE также связан с B-клетками (а также моноцитами, эозинофилами и тромбоцитами) за счет связывания его Fc-области с низкоаффинным рецептором IgE (FcεRII). Когда млекопитающее подвергается воздействию аллергена, клонально размножаются B-клетки, синтезирующие IgE, который связывает аллерген. Этот IgE, в свою очередь, высвобождается в систему кровообращения B-клетками, где он связывается B-клетками (через FcεRII), а также тучными клетками и базофилами через так называемый высокоаффинный рецептор (FcεRI), присутствующий на поверхности тучных клеток и базофилов. В результате этого тучные клетки и базофилы сенсибилизируются аллергеном. При следующем воздействии аллергена происходит перекрестное связывание FcεRI на этих клетках и, вследствие этого, активация высвобождения ими гистамина и других факторов, являющихся причиной клинической гиперчувствительности и анафилаксии.
Омализумаб (ксолар®) представляет собой продуцируемое с рекомбинантной ДНК гуманизированное IgG1κ моноклональное антитело, которое селективно связывается с иммуноглобулином E человека (IgE) [Cε3-доменом]. Антитело имеет молекулярную массу примерно 149 кДа. Ксолар® продуцируется в суспензионной культуре клеток яичника китайского хомяка в питательной среде, содержащей антибиотик гентамицин. Ксолар® представляет собой стерильный белый, не содержащий консерванты лиофилизированный порошок, находящийся во флаконе одноразового использования, который восстанавливают стерильной водой для инъекций (СВДИ), USP, (или, альтернативно, в виде жидкого препарата в стерильном шприце) и вводят подкожной (п/к) инъекцией [смотри EP602126 (и SPC/GB06/005 на его основе); WO93/04173; US6267958 (и ксолар® PTE на основе данного патента); WO97/04807; WO97/04801; Presta et al. (1993) J. Immunol. 151:2623-2632].
Омализумаб в настоящее время назначают для лечения умеренной и тяжелой форм хронической астмы у пациентов в случае положительных результатов кожной пробы или in vitro реакции на круглогодичные аэроаллергены, а также в случае симптомов, которые не поддаются достаточному контролю ингаляционными кортикостероидами (из указаний по применению препарата ксолар®).
Проблемы, связанные с использованием омализумаба, заключаются в следующем: 1) он нацелен на свободный IgE, но не нацелен (или неэффективно нацелен) на патогенные варианты комплекса IgE/FcεRI при фармацевтически используемых дозах; 2) возможно, вследствие того, что патогенные варианты комплекса IgE/FcεRI не являются мишенью, «требуется по меньшей мере 12-16 недель для появления эффекта лечения ксоларом» (ксолар® 150 мг раствор - Краткая характеристика лекарственного средства, 2014) -или, фактически, для установления того, будет ли ксолар® эффективен для конкретного пациента, или будет необходимо другое лечение; 3) его не следует применять пациентам с высокими уровнями IgE (например, вследствие того, что патогенные варианты комплекса IgE/FcεRI не являются мишенью и не исчезают со временем из-за высоких уровней свободного IgE у пациента); 4) при приеме омализумаба могут возникать локальные типа I или системные реакции, включая анафилаксию и анафилактический шок» (ксолар® 150 мг раствор - Краткая характеристика лекарственного средства, 2014); 5) его аффинность для IgE не очень высока (примерно 2 нМ).
Целью настоящего изобретения является выявление новых антител, позволяющих решать одну или несколько из указанных проблем.
Следующей целью является выявление антител против новых эпитопов (имеющих более сильное взаимодействие с IgE Cε2, чем омализумаб) и/или антител на основе новых мутантов омализумаба с повышенной аффинностью и/или повышенной способностью вызывать диссоциацию комплекса IgE/FcεRI.
Следующей целью изобретения является выявление новых соединений, способов и композиций для лечения заболеваний, связанных с IgE, в частности, заболеваний, связанных с комплексом IgE/FcεRI, например, аллергических заболеваний.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном аспекте изобретение относится к анти-IgE антителу, или антигенсвязывающему фрагменту, которое контактирует с эпитопом, содержащим остатки T373, W374, S375, R376, A377, S378, G379, P381, Q417, C418, R419, T421, P426, R427, A428 Cε3-домена и остатки D278 и T281 Cε2-домена IgE человека. В следующих вариантах осуществления эпитоп может дополнительно содержать один или более из остатков K380 и/или M430 Cε3-домена IgE человека и/или один или более из остатков D276, V277, L279, S280, A282 и/или T298 Cε2-домена IgE человека.
Изобретение основано на результатах изучения кристаллической структуры в Примере 1, которая впервые продемонстрировала взаимодействие усовершенствованного антитела (на основе омализумаба) с IgE-Fc, где значительные взаимодействия наблюдались с Cε2-доменом IgE в области мутации. Это может приводить к усовершенствованным функциональным характеристикам анти-IgE антитела, или антигенсвязывающего фрагмента, в сравнении с омализумабом и/или Fab омализумаба. Например, анти-IgE антитело, или антигенсвязывающий фрагмент, может быть способно вызывать диссоциацию IgE человека от FcεRI при концентрациях (или пиковых сывороточных концентрациях) менее, чем 7, 3, 1, 0,66, 0,5 или 0,3 мкМ (например, при определении способом, описанным в Примере 2). Например, анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь усовершенствованную/более сильную аффинность (более низкую KD) для IgE человека (например, при использовании IgE-Fc) (например, при определении способом, описанным в Примере 6) в сравнении с омализумабом и/или Fab омализумаба; и/или усовершенствованную способность вызывать диссоциацию комплекса IgE/FcεRI (например, при определении способом, описанным в Примере 2) в сравнении с омализумабом и/или Fab омализумаба; и/или способность вызывать диссоциацию IgE человека от FcεRI при концентрациях (или пиковых сывороточных концентрациях), более низких, чем концентрации омализумаба и/или Fab омализумаба (например, при определении способом, описанным в Примере 2). «Усовершенствованная KD» означает по меньшей мере на 5, 10, 20, 30, 40 или 50% более низкое значение, чем значение для омализумаба и/или Fab омализумаба. KD анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению может составлять менее 2, 1,9, 1,8, 1,7, 1,6, 1,5, 1,4, 1,3, 1,2, 1,1, 1,0, 0,9, 0,8, 0,7, 0,6, 0,5, 0,4 или 0,3 нМ. «Усовершенствованная возможность или способность» вызывать диссоциацию IgE человека от FcεRI означает по меньшей мере на 5, 10, 20, 30, 40, 50, или 100% усовершенствованную способность в сравнении с омализумабом и/или Fab омализумаба (например, при измерении % диссоциации и/или кажущейся скорости диссоциации комплекса IgE/FcεRI, как описано в Примерах 2 и 7), и/или достижение диссоциации при концентрации, при которой омализумаб и/или Fab омализумаба не способен вызывать диссоциацию.
Во избежание сомнений, анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению не является омализумабом или Fab омализумаба.
В одном из вариантов осуществления эпитоп определяют методом кристаллографии (например, как описано в Примере 1) путем определения остатков IgE, находящихся в пределах 4 или 5 Å от анти-IgE антитела, или антигенсвязывающего фрагмента, в кристаллической структуре комплекса IgE-Fc/анти-IgE антитело, или антигенсвязывающий фрагмент. Используемая область IgE-Fc может иметь последовательность SEQ ID NO:108 (с дополнительными мутациями N265Q и N371Q).
В одном из вариантов осуществления анти-IgE антитело, или антигенсвязывающий фрагмент, в конкретном сайте связывания контактирует с эпитопом, при этом части эпитопа из Cε3-домена и Cε2-домена находятся на разных цепях IgE человека. IgE имеет две цепи в Fc-домене, каждая из которых имеет Cε3-домен и Cε2-домен.
В одном из вариантов осуществления анти-IgE антитело, или антигенсвязывающий фрагмент, в конкретном сайте связывания контактирует с эпитопом, при этом части эпитопа из Cε3-домена и Cε2-домена находятся на одной и той же цепи IgE человека.
Во избежание сомнений, два анти-IgE антитела, или антигенсвязывающих фрагмента, по изобретению могут связываться с IgE человека, но требуется взаимодействие лишь одного из них с эпитопом по изобретению, включающим Cε3 и Cε2-домены (другое может взаимодействовать лишь с другим Cε3-доменом, например).
В одном из вариантов осуществления (необязательно, также включающем признаки по первому аспекту изобретения) анти-IgE антитело, или антигенсвязывающий фрагмент, является специфическим для указанного эпитопа, содержащего остатки T373, W374, S375, R376, A377, S378, G379, P381, Q417, C418, R419, T421, P426, R427, A428 Cε3-домена и остатки D278 и T281 Cε2-домена IgE человека. Необязательно, указанный эпитоп может дополнительно содержать один или более из остатков K380 и/или M430 Cε3-домена IgE человека и/или один или более из остатков D276, V277, L279, S280, A282 и/или T298 Cε2-домена IgE человека. Во избежание сомнений, анти-IgE антитело, или антигенсвязывающий фрагмент, является специфичным для указанного эпитопа, если оно узнает и связывается со специфической структурой IgE человека, содержащей указанный эпитоп, а не в общем с IgE человека.
В следующем аспекте (необязательно, также включающем признаки по первому аспекту изобретения) предложено анти-IgE антитело, или антигенсвязывающий фрагмент, содержащее вариабельную область тяжелой цепи, содержащую определяющую комплементарность область, CDR-H3, с аминокислотной последовательностью SEQ ID NO:18, и вариабельную область легкой цепи, содержащую определяющую комплементарность область, CDR-L1, с аминокислотной последовательностью SEQ ID NO:29, при этом вариабельная область легкой цепи также содержит каркасную область, FR-L3, с аминокислотной последовательностью, выбранной из SEQ ID NO:32, которая имеет одну, две, три, четыре, пять, шесть, семь или более аминокислотных замен для усиления взаимодействия анти-IgE антитела, или антигенсвязывающего фрагмента, с Cε2-доменом IgE человека.
В следующем аспекте (необязательно, также включающем признаки по предшествующим аспектам изобретения) предложено анти-IgE антитело, или антигенсвязывающий фрагмент, содержащее вариабельную область тяжелой цепи, содержащую определяющую комплементарность область, CDR-H3, с аминокислотной последовательностью SEQ ID NO:18, и вариабельную область легкой цепи, содержащую определяющую комплементарность область, CDR-L1, с аминокислотной последовательностью SEQ ID NO:29, при этом вариабельная область легкой цепи также содержит каркасную область, FR-L1, с аминокислотной последовательностью SEQ ID NO:28, которая имеет одну, две, три, четыре, пять, шесть, семь или более аминокислотных замен для усиления взаимодействия анти-IgE антитела, или антигенсвязывающего фрагмента, с Cε2-доменом IgE человека.
Если области CDR-H3 и CDR-L1 заякоривают и ориентируют анти-IgE антитело, или антигенсвязывающий фрагмент, на Cε3-области IgE (как в случае омализумаба), изменение(я) в последовательностях FR-L3 и/или FR-L1 делает возможным более сильное взаимодействие с Cε2-доменом IgE человека. Более сильное взаимодействие мутанта в сравнении с омализумабом или Fab омализумаба можно оценивать путем измерения аффинности [более низкая KD] (например, при определении способом, описанным в Примере 6) и/или характеристик улучшенной диссоциации комплекса IgE/FcεRI (например, при определении способом, описанным в Примере 2).
Более сильное взаимодействие анти-IgE антитела, или антигенсвязывающего фрагмента, с Cε2-доменом IgE человека может быть охарактеризовано на основании усовершенствованных функциональных характеристик анти-IgE антитела, или антигенсвязывающего фрагмента, в сравнении с омализумабом и/или Fab омализумаба. Например, анти-IgE антитело, или антигенсвязывающий фрагмент, может обладать способностью вызывать диссоциацию IgE человека от FcεRI при концентрациях (или пиковых сывороточных концентрациях) менее, чем 7, 3, 1, 0,66, 0,5 или 0,3 мкМ (например, при определении способом, описанным в Примере 2). Например, анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь усовершенствованную/более сильную аффинность (более низкую KD) для IgE человека (например, при использовании IgE-Fc) (например, при определении способом, описанным в Примере 6) в сравнении с омализумабом и/или Fab омализумаба; и/или усовершенствованную способность вызывать диссоциацию комплекса IgE/FcεRI (например, при определении способом, описанным в Примере 2) в сравнении с омализумабом и/или Fab омализумаба; и/или способность вызывать диссоциацию IgE человека от FcεRI при концентрациях (или пиковых сывороточных концентрациях) ниже, чем в случае омализумаба и/или Fab омализумаба (например, при определении способом, описанным в Примере 2). «Усовершенствованная KD» означает по меньшей мере на 5, 10, 20, 30, 40 или 50% более низкое значение, чем значение для омализумаба и/или Fab омализумаба. KD анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению может составлять менее 2, 1,9, 1,8, 1,7, 1,6, 1,5, 1,4, 1,3, 1,2, 1,1, 1,0, 0,9, 0,8, 0,7, 0,6, 0,5, 0,4 или 0,3 нМ. «Усовершенствованная возможность или способность» вызывать диссоциацию IgE человека от FcεRI означает по меньшей мере на 5, 10, 20, 30, 40, 50, или 100% усовершенствованную способность в сравнении с омализумабом и/или Fab омализумаба (например, при измерении % диссоциации и/или кажущейся скорости диссоциации комплекса IgE/FcεRI, как описано в Примерах 2 и 7), и/или достижение диссоциации при концентрации, при которой омализумаб и/или Fab омализумаба не способен вызывать диссоциацию.
Во избежание сомнений, анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению не является омализумабом или Fab омализумаба.
В одном из вариантов осуществления область FR-L3 имеет мутацию относительно SEQ ID NO:129 в одном или более из положений S60, S63, S76, S77 и/или Q79 (Kabat) на одну из других природных аминокислот.
Например, область FR-L3 может иметь мутацию в положении S60 (Kabat) на одну из других природных аминокислот, например, на M, R, K, N, Q или T, в частности, M.
Например, область FR-L3 может иметь мутацию в положении S63 (Kabat) на одну из других природных аминокислот, например, W или Y, в частности, Y.
Например, область FR-L3 может иметь мутацию в положении S76 (Kabat) на одну из других природных аминокислот, в частности, N.
Например, область FR-L3 может иметь мутацию в положении S77 (Kabat) на одну из других природных аминокислот, например, R или K, в частности, R.
Например, область FR-L3 может иметь мутацию в положении Q79 (Kabat) на одну из других природных аминокислот, например, R или K, в частности, R.
Например, область FR-L1 может иметь мутацию относительно SEQ ID NO:20 в положении G16 и/или R18 (Kabat) на одну из других природных аминокислот.
В конкретных вариантах осуществления аминокислотную последовательность мутантной области FR-L3 анти-IgE антитела, или антигенсвязывающего фрагмента, выбирают из SEQ ID NO:43-49, 60-83, 131 или 138.
В следующем варианте осуществления область FR-L3 также имеет мутацию относительно SEQ ID NO:129 в положении S67 (Kabat) на одну из других природных аминокислот для повышения ее аффинности (более низкой KD) для IgE человека. В этом случае мутация может приводить к усилению взаимодействия анти-IgE антитела, или антигенсвязывающего фрагмента, с Cε3-доменом IgE. Например, область FR-L3 может иметь мутацию в положении S67 (Kabat) на M (в частности), E или D. В конкретных вариантах осуществления аминокислотную последовательность мутантной области FR-L3 анти-IgE антитела, или антигенсвязывающего фрагмента, выбирают из SEQ ID NO:53-59, 84-107, 131 или 138.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область легкой цепи, также содержащую определяющую комплементарность область, CDR-L2, с аминокислотной последовательностью SEQ ID NO:31.
В одном из вариантов осуществления область CDR-L2 имеет мутацию в положении S52 (Kabat) на одну из других природных аминокислот для повышения ее аффинности (более низкой KD) для IgE человека. В этом случае мутация может приводить к усилению взаимодействия анти-IgE антитела, или антигенсвязывающего фрагмента, с Cε3-доменом IgE. Например, область CDR-L2 может иметь мутацию относительно SEQ ID NO:129 в положении S52 (Kabat) на D (в частности), E, Q или R. В конкретных вариантах осуществления аминокислотную последовательность мутантной области CDR-L2 выбирают из SEQ ID NO:50 или SEQ ID NO:51.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, также содержащую определяющую комплементарность область, CDR-H1, с аминокислотной последовательностью SEQ ID NO:14.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, также содержащую определяющую комплементарность область, CDR-H2, с аминокислотной последовательностью SEQ ID NO:16.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область легкой цепи, также содержащую определяющую комплементарность область, CDR-L3, с аминокислотной последовательностью SEQ ID NO:33.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H1, с аминокислотной последовательностью SEQ ID NO:13.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H2, с аминокислотной последовательностью SEQ ID NO:15.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H3, с аминокислотной последовательностью SEQ ID NO:17.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H4, с аминокислотной последовательностью SEQ ID NO:19.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область легкой цепи, также содержащую каркасную область, FR-L2, с аминокислотной последовательностью SEQ ID NO:30.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область легкой цепи, также содержащую каркасную область, FR-L4, с аминокислотной последовательностью SEQ ID NO:34.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область легкой цепи, VL, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:35, SEQ ID NO:132 или SEQ ID NO:134, или SEQ ID NO:141, или SEQ ID NO:144, или SEQ ID NO:145, или SEQ ID NO:158, или SEQ ID NO:159.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, VH, имеющую аминокислотную последовательность SEQ ID NO:1.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может дополнительно содержать константную область легкой цепи.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь константную область легкой цепи, которая представляет собой константную область каппа.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь константную область легкой цепи, имеющую мутацию L154P (Kabat).
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область легкой цепи и константную область легкой цепи, VL-CL, имеющие аминокислотную последовательность, выбранную из SEQ ID NO:39 или SEQ ID NO:41, или SEQ ID NO:117, или SEQ ID NO:119, или SEQ ID NO:125, или SEQ ID NO:127, или SEQ ID NO:136, или SEQ ID NO:143, необязательно содержащую сигнальную последовательность, имеющую аминокислотную последовательность SEQ ID NO:160.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может дополнительно содержать константную область тяжелой цепи, CH1.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи и константную область тяжелой цепи, VH-CH1, имеющие аминокислотную последовательность SEQ ID NO:5.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может дополнительно содержать Fc-область тяжелой цепи, Fc.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь Fc из человеческого IgG1 или человеческого IgG4.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, константную область тяжелой цепи и Fc-область тяжелой цепи, VH-CH1-Fc, имеющие аминокислотную последовательность SEQ ID NO:9.
В следующем аспекте изобретения предложено анти-IgE антитело, или антигенсвязывающий фрагмент, содержащее вариабельную область тяжелой цепи, содержащую определяющую комплементарность область, CDR-H3, с аминокислотной последовательностью SEQ ID NO:18, и вариабельную область легкой цепи, содержащую определяющую комплементарность область, CDR-L1, с аминокислотной последовательностью SEQ ID NO:29, при этом:
a. вариабельная область легкой цепи также содержит каркасную область, FR-L3, с аминокислотной последовательностью SEQ ID NO:32, причем область FR-L3 имеет мутацию относительно SEQ ID NO:129 в положении S67 (Kabat) на одну из других природных аминокислот для повышения аффинности (более низкой KD) анти-IgE антитела, или антигенсвязывающего фрагмента, для IgE человека; и/или
b. вариабельная область легкой цепи также содержит определяющую комплементарность область, CDR-L2, с аминокислотной последовательностью SEQ ID NO:31, причем область CDR-L2 имеет мутацию относительно SEQ ID NO:129 в положении S52 (Kabat) на одну из других природных аминокислот для повышения аффинности (более низкой KD) анти-IgE антитела, или антигенсвязывающего фрагмента, для IgE человека.
Авторы настоящего изобретения обнаружили, что любая, или обе, из этих мутаций могут удивительно повышать аффинность (усовершенствованная или более низкая KD) анти-IgE антитела, или антигенсвязывающего фрагмента, на основе омализумаба или Fab омализумаба для IgE человека (например, при использовании IgE-Fc) (например, при определении способом, описанным в Примере 6). В частности, повышение аффинности имеет место в сравнении с омализумабом и/или Fab омализумаба. Мутации могут приводить к улучшению взаимодействия с Cε3-доменом IgE. «Усовершенствованная или более низкая KD» означает по меньшей мере на 5, 10, 20, 30, 40 или 50% более низкое значение, чем значение для омализумаба и/или Fab омализумаба. KD анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению может составлять менее 2, 1,9, 1,8, 1,7, 1,6, 1,5, 1,4, 1,3, 1,2, 1,1, 1,0, 0,9, 0,8, 0,7, 0,6, 0,5, 0,4 или 0,3 нМ.
Например, область FR-L3 может иметь мутацию относительно SEQ ID NO:129 в положении S67 (Kabat) на M (в частности), E или D.
В конкретных вариантах осуществления аминокислотную последовательность мутантной области FR-L3 анти-IgE антитела, или антигенсвязывающего фрагмента, выбирают из SEQ ID NO:52-59, 84-107, 131 или 138.
Например, область CDR-L2 может иметь мутацию относительно SEQ ID NO:129 в положении S52 (Kabat) на D (в частности), E, Q или R.
В конкретных вариантах осуществления аминокислотную последовательность мутантной области CDR-L2 анти-IgE антитела, или антигенсвязывающего фрагмента, выбирают из SEQ ID NO:50 (в частности) или SEQ ID NO:51.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, также содержащую определяющую комплементарность область, CDR-H1, с аминокислотной последовательностью SEQ ID NO:14.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, также содержащую определяющую комплементарность область, CDR-H2, с аминокислотной последовательностью SEQ ID NO:16.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи, также содержащую определяющую комплементарность область, CDR-L3, с аминокислотной последовательностью SEQ ID NO:33.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H1, с аминокислотной последовательностью SEQ ID NO:13.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H2, с аминокислотной последовательностью SEQ ID NO:15.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H3, с аминокислотной последовательностью SEQ ID NO:17.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, также содержащую каркасную область, FR-H4, с аминокислотной последовательностью SEQ ID NO:19.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи, также содержащую каркасную область, FR-L2, с аминокислотной последовательностью SEQ ID NO:30.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи, также содержащую каркасную область, FR-L4, с аминокислотной последовательностью SEQ ID NO:34.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи, VL, содержащую последовательные области FR-L1, CDR-L1, FR-L2, CDR-L2, FR-L3, CDR-L3 и FR-L4, и имеющую аминокислотную последовательность SEQ ID NO:20, за исключением того, что область CDR-L2 имеет аминокислотную последовательность, выбранную из SEQ ID NO:50 (в частности) или SEQ ID NO:51.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи, VL, содержащую последовательные области FR-L1, CDR-L1, FR-L2, CDR-L2, FR-L3, CDR-L3 и FR-L4, и имеющую аминокислотную последовательность SEQ ID NO:20, за исключением того, что область FR-L3 имеет аминокислотную последовательность SEQ ID NO:52.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи, VL, содержащую последовательные области FR-L1, CDR-L1, FR-L2, CDR-L2, FR-L3, CDR-L3 и FR-L4, и имеющую аминокислотную последовательность SEQ ID NO:20, за исключением того, что область CDR-L2 имеет аминокислотную последовательность, выбранную из SEQ ID NO:50 (в частности) или SEQ ID NO:51, и область FR-L3 имеет аминокислотную последовательность, которую выбирают из SEQ ID NO:52, 131 или 138.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь вариабельную область тяжелой цепи, VH, имеющую аминокислотную последовательность SEQ ID NO:1.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может дополнительно содержать константную область легкой цепи.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь константную область легкой цепи, которая представляет собой константную область каппа.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи и константную область легкой цепи, VL-CL, имеющие аминокислотную последовательность SEQ ID NO:24, за исключением того, что область CDR-L2 имеет аминокислотную последовательность, выбранную из SEQ ID NO:50 (в частности) или SEQ ID NO:51.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи и константную область легкой цепи, VL-CL, имеющие аминокислотную последовательность SEQ ID NO:24, за исключением того, что область FR-L3 имеет аминокислотную последовательность SEQ ID NO:52.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область легкой цепи и константную область легкой цепи, VL-CL, имеющие аминокислотную последовательность SEQ ID NO:24, за исключением того, что область CDR-L2 имеет аминокислотную последовательность, выбранную из SEQ ID NO:50 (в частности) или SEQ ID NO:51, и область FR-L3 имеет аминокислотную последовательность, которую выбирают из SEQ ID NO:52, 131 или 138.
В следующем аспекте изобретение относится к анти-IgE антителу, или антигенсвязывающему фрагменту, содержащему вариабельную область тяжелой цепи и вариабельную область легкой цепи, при этом:
a. вариабельная область тяжелой цепи содержит CDR-H1 с аминокислотной последовательностью SEQ ID NO:14, CDR-H2 с аминокислотной последовательностью SEQ ID NO:16 и CDR-H3 с аминокислотной последовательностью SEQ ID NO:18, и вариабельная область легкой цепи содержит CDR-L1 с аминокислотной последовательностью SEQ ID NO:29, CDR-L2 с аминокислотной последовательностью SEQ ID NO:50, CDR-L3 с аминокислотной последовательностью SEQ ID NO:33 и каркасную область FW-L3 с аминокислотной последовательностью SEQ ID NO:131 или 138; или
b. вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:1 и вариабельная область легкой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO:132 или 139.
В одном из вариантов осуществления анти-IgE антитело, или антигенсвязывающий фрагмент, может дополнительно содержать константную область легкой цепи, при этом вариабельная область легкой цепи и константная область легкой цепи VL-CL имеют аминокислотную последовательность, выбранную из SEQ ID NO:136 или 143, необязательно, содержащую сигнальную последовательность, имеющую аминокислотную последовательность SEQ ID NO:160.
Во всех вариантах осуществления, описанных в настоящем документе, анти-IgE антитело, или антигенсвязывающий фрагмент, может дополнительно содержать константную область тяжелой цепи, CH1.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи и константную область тяжелой цепи, VH-CH1, имеющие аминокислотную последовательность SEQ ID NO:5.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может дополнительно содержать Fc-область тяжелой цепи, Fc.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь Fc из человеческого IgG1 или человеческого IgG4.
Анти-IgE антитело, или антигенсвязывающий фрагмент, может иметь вариабельную область тяжелой цепи, константную область тяжелой цепи и Fc-область тяжелой цепи, VH-CH1-Fc, имеющие аминокислотную последовательность SEQ ID NO:9.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по всем аспектам изобретения можно выбирать из группы, состоящей из: целой молекулы антитела, имеющей полноразмерные тяжелую и легкую цепи, или ее фрагмента.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению можно выбирать из группы, состоящей из: Fab-фрагмента, модифицированного Fab'-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv, scFv, scab, диатела, биспецифического антитела, триатела, FabFv, Fab-Fv-Fv, триотела или (Fab-Fv)2-Fc. Без связи с конкретной теорией, анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может быть связано с меньшим риском вызывания анафилаксии, если оно имеет только один, а не несколько сайтов связывания антигена IgE.
В одном из вариантов осуществления анти-IgE антитело представляет собой Fab-фрагмент, связанный непосредственно или через линкер с фрагментом scFv, который связывается с сывороточным белком-носителем, таким как человеческий сывороточный альбумин.
В одном из вариантов осуществления scFv может содержать вариабельную область тяжелой цепи и вариабельную область легкой цепи, предпочтительно, связанные через линкер, имеющий SEQ ID NO:151, при этом вариабельная область тяжелой цепи содержит CDR-H1 с аминокислотной последовательностью SEQ ID NO:152, CDR-H2 с аминокислотной последовательностью SEQ ID NO:153 и CDR-H3 с аминокислотной последовательностью SEQ ID NO:154, и вариабельная область легкой цепи содержит CDR-L1 с аминокислотной последовательностью SEQ ID NO:155, CDR-L2 с аминокислотной последовательностью SEQ ID NO:156, CDR-L3 с аминокислотной последовательностью SEQ ID NO:157.
В одном из вариантов осуществления фрагмент scFv имеет аминокислотную последовательность SEQ ID NO:150. В одном предпочтительном варианте осуществления Fab-фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, при этом:
a. вариабельная область тяжелой цепи содержит CDR-H1 с аминокислотной последовательностью SEQ ID NO:14, CDR-H2 с аминокислотной последовательностью SEQ ID NO:16 и CDR-H3 с аминокислотной последовательностью SEQ ID NO:18, и вариабельная область легкой цепи содержит CDR-L1 с аминокислотной последовательностью SEQ ID NO:29, CDR-L2 с аминокислотной последовательностью SEQ ID NO:50, CDR-L3 с аминокислотной последовательностью SEQ ID NO:33 и каркасную область FW-L3 с аминокислотной последовательностью SEQ ID NO:131 или 138; или
b. вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:1 и вариабельная область легкой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO:132 или 139.
В другом варианте осуществления Fab-фрагмент также содержит константную область тяжелой цепи и константную область легкой цепи, при этом вариабельная область тяжелой цепи и константная область тяжелой цепи VL-CH1 имеют аминокислотную последовательность SEQ ID NO:5, и при этом вариабельная область легкой цепи и константная область легкой цепи VL-CL имеют аминокислотную последовательность, выбранную из SEQ ID NO:136 или 143, необязательно, содержащую сигнальную последовательность, которая имеет аминокислотную последовательность SEQ ID NO:160.
В другом варианте осуществления фрагмент scFv связан с CH1 Fab-фрагмента через линкер, имеющий аминокислотную последовательность SEQ ID NO:149.
В одном из вариантов осуществления вариабельная область тяжелой цепи и константная область тяжелой цепи, линкер и scFv имеют аминокислотную последовательность SEQ ID NO:147, необязательно, содержащую сигнальную последовательность, которая имеет аминокислотную последовательность SEQ ID NO:160.
В другом варианте осуществления тяжелая цепь Fab-фрагмента, связанного с scFv с SEQ ID NO:147, спарена с вариабельной и константной областью легкой цепи, которые имеют SEQ ID NO:136 или 143.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может иметь присоединенную к нему эффекторную или репортерную молекулу.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может быть гликозилированным (например, внутри Fc-домена) и/или может быть конъюгированным с полимером, выбранным из крахмала, альбумина и полиэтиленгликоля (ПЭГ). В одном из вариантов осуществления конъюгированный ПЭГ может иметь молекулярную массу в диапазоне 5-50 кДа.
Анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению может быть гуманизированным.
Следующий аспект изобретения относится к выделенной последовательности ДНК, кодирующей тяжелую и/или легкую цепь(и) анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению. Также предложен клонирующий или экспрессионный вектор, содержащий одну или более последовательностей ДНК по изобретению. Например, клонирующий или экспрессионный вектор может содержать одну или более последовательностей ДНК, выбранных из SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40 или SEQ ID NO:42, или SEQ ID NO:133, или SEQ ID NO:135, или SEQ ID NO:137, или SEQ ID NO:140, или SEQ ID NO:142, или SEQ ID NO:144, и, необязательно, может дополнительно содержать одну или более последовательностей ДНК, выбранных из SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10 или SEQ ID NO:12, или SEQ ID NO:148.
Следующий аспект изобретения относится к клетке-хозяину, содержащей один или более клонирующих, или экспрессионных, векторов по изобретению. Клетка-хозяин по изобретению может, необязательно, также содержать один или более клонирующих, или экспрессионных, векторов, содержащих одну или более последовательностей ДНК, выбранных из SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10 или SEQ ID NO:12, или SEQ ID NO:148.
Также предложен способ получения анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению, включающий культивирование клетки-хозяина по изобретению и выделение анти-IgE антитела, или антигенсвязывающего фрагмента.
Следующий аспект относится к фармацевтической композиции, содержащей анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению, в сочетании с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя. Предпочтительно, анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению присутствует в дозе 50-200, предпочтительно примерно, или точно, 150 мг на мл разбавителя. В конкретных вариантах осуществления эксципиент включает один или оба из L-аргинина, L-гистидина. Эксципиент может отдельно или в сочетании включать полисорбат 20. Разбавитель может представлять собой воду или водный изотонический раствор.
Фармацевтическая композиция по изобретению может находиться в стерильном флаконе в виде порошка, восстанавливаемого перед подкожным введением, или в стерильном шприце для немедленного подкожного введения.
Фармацевтическая композиция по изобретению может содержать общую дозу анти-IgE антитела, или антигенсвязывающего фрагмента, составляющую 75-600 мг - например, примерно, или точно, 100 или 150 мг.
Кроме того, фармацевтическая композиция по изобретению может содержать другие активные ингредиенты, находящиеся либо вместе с анти-IgE антителом, или антигенсвязывающим фрагментом, либо в отдельном препарате для совместного введения с анти-IgE антителом, или антигенсвязывающим фрагментом. Например, фармацевтическая композиция по изобретению может быть использована в контексте специальной иммунотерапии на основе аллергии, в которой анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению совместно вводят с отдельным (но, возможно, совместно упакованным) аллергеном. Таким образом, фармацевтическая композиция по изобретению может быть предназначена для специальной иммунотерапии на основе аллергии, в которой пациент получает фармацевтическую композицию по изобретению за 7, 6, 5, 4, 3, 2 или 1 дней до (или в тот же день) введения терапевтического аллергена.
Анти-IgE антитело, или антигенсвязывающий фрагмент, или композиция, по изобретению могут быть использованы в качестве лекарственного средства.
Анти-IgE антитело, или антигенсвязывающий фрагмент, или композиция, по изобретению могут быть использованы для лечения или профилактики заболевания.
Анти-IgE антитело, или антигенсвязывающий фрагмент, или композиция, по изобретению могут быть использованы для лечения или профилактики заболеваний, связанных с комплексом IgE человека и FcεRI.
Анти-IgE антитело, или антигенсвязывающий фрагмент, или композиция, по изобретению могут быть использованы для лечения или профилактики заболеваний за счет стимулирования диссоциации комплекса IgE человека и FcεRI и связывания IgE человека анти-IgE антителом, или антигенсвязывающим фрагментом.
Анти-IgE антитело, или антигенсвязывающий фрагмент, или композиция, по изобретению могут быть использованы для лечения или профилактики одного или более из: аллергии; аллергической астмы; тяжелой астмы; умеренной астмы; хронической спонтанной крапивницы; хронической идиопатической крапивницы; продолжительного аллергического ринита; сезонного аллергического ринита; обострений астмы; острого бронхоспазма; астматического статуса; гипер-IgE синдрома; аллергического бронхолегочного аспергиллеза; профилактики анафилактических реакций; пищевой аллергии; атопического дерматита; аллергического ринита; аллергии на пчелиный яд; идиопатической анафилаксии; аллергии на арахис; аллергии на латекс; воспалительных кожных заболеваний; крапивницы (вызываемой солнечным светом, холодом, локальным нагреванием и/или длительным сдавливанием); кожного мастоцитоза; системного мастоцитоза; эозинофил-связанного желудочно-кишечного заболевания; буллезного пемфигоида; интерстициального цистита; полипов носовой полости; идиопатического ангионевротического отека или неаллергической астмы.
Также предложен способ лечения или профилактики заболевания у индивида-человека, включающий введение индивиду эффективного количества анти-IgE антитела, или антигенсвязывающего фрагмента, или композиции по изобретению. Способ может быть предназначен для лечения или профилактики заболеваний, связанных с комплексом IgE человека и FcεRI. Способ по изобретению позволяет лечить или предотвращать заболевание за счет диссоциации комплекса IgE человека и FcεRI и связывания IgE человека анти-IgE антителом, или антигенсвязывающим фрагментом, по изобретению.
Способ по изобретению может быть предназначен для лечения или профилактики одного или более из: аллергии; аллергической астмы; тяжелой астмы; умеренной астмы; хронической спонтанной крапивницы; хронической идиопатической крапивницы; продолжительного аллергического ринита; сезонного аллергического ринита; обострений астмы; острого бронхоспазма; астматического статуса; гипер-IgE синдрома; аллергического бронхолегочного аспергиллеза; профилактики анафилактических реакций; пищевой аллергии; атопического дерматита; аллергического ринита; аллергии на пчелиный яд; идиопатической анафилаксии; аллергии на арахис; аллергии на латекс; воспалительных кожных заболеваний; крапивницы (вызываемой солнечным светом, холодом, локальным нагреванием и/или длительным сдавливанием); кожного мастоцитоза; системного мастоцитоза; эозинофил-связанного желудочно-кишечного заболевания; буллезного пемфигоида; интерстициального цистита; полипов носовой полости; идиопатического ангионевротического отека или неаллергической астмы.
В настоящем изобретении было установлено, что антитело, или антигенсвязывающий фрагмент, против первого полипептида, который осуществляет свой физиологический ответ за счет связывания со вторым полипептидом (таким как рецептор), может связываться как со свободным, так и со связанным, первым полипептидом, стабилизируя конформацию такого первого полипептида. Такая стабилизированная конформация имеет более слабую аффинность связывания для второго полипептида, чем в отсутствие антитела, или антигенсвязывающего фрагмента, таким образом, инициируется более быстрая диссоциация первого полипептида от второго полипептида.
В этом отношении, изобретение в следующем аспекте относится к антителу, или антигенсвязывающему фрагменту, способному связывать свободный и FcεRI-связанный человеческий IgE и стабилизировать конформацию IgE. Когда IgE находится в такой конформации, он имеет более слабую аффинность связывания для FcεRI, чем в отсутствие антитела, или антигенсвязывающего фрагмента, и при этом FcεRI-связанный человеческий IgE диссоциирует от FcεRI. Необязательно, когда IgE находится в такой конформации, IgE имеет более низкую аффинность связывания для омализумаба или его фрагмента, чем для антитела или антигенсвязывающего фрагмента по изобретению. Например, антитело, или антигенсвязывающий фрагмент, представляет собой антитело, описанное в настоящем документе.
В следующем аспекте изобретение относится к способу выбора таких антител, или антигенсвязывающих фрагментов, которые описаны в настоящем документе. Способ включает:
a. создание контакта тестируемого антитела, или антигенсвязывающего фрагмента, с образцом, содержащим человеческий IgE, связанный с FcεRI человека;
b. измерение константы диссоциации тестируемого антитела, или антигенсвязывающего фрагмента, для диссоциации IgE человека от человеческого FcεRI;
c. сравнение константы диссоциации, измеренной на этапе b), с константой диссоциации омализумаба, или его фрагмента, для диссоциации IgE человека от человеческого FcεRI;
d. выбор антитела, или антигенсвязывающего фрагмента, если указанное антитело, или антигенсвязывающий фрагмент, вызывает диссоциацию IgE от FcεRI быстрее, чем омализумаб или его фрагмент.
Альтернативно, способ выбора антител, или антигенсвязывающих фрагментов, по изобретению включает:
a. создание контакта тестируемого антитела, или антигенсвязывающего фрагмента, с образцом, содержащим человеческий IgE, связанный с FcεRI человека;
b. измерение аффинности связывания тестируемого антитела, или антигенсвязывающего фрагмента, для IgE человека, связанного с FcεRI человека;
c. сравнение аффинности связывания, измеренной на этапе b), с аффинностью связывания IgE человека для FcεRI;
d. выбор антитела, или антигенсвязывающего фрагмента, если указанное антитело, или антигенсвязывающий фрагмент, имеет более высокую аффинность связывания для IgE, чем аффинность IgE для FcεRI.
Необязательно, выбранные антитела, или антигенсвязывающие фрагменты, вызывает переход IgE, все еще связанного с FcεRI, в определенную конформацию, при этом IgE в указанной стабилизированной конформации может диссоциировать от FcεRI быстрее, чем IgE, связанный с FcεRI, в присутствии омализумаба или его фрагмента; и/или может иметь более высокую аффинность связывания для антитела, или антигенсвязывающего фрагмента, чем для FcεRI.
В завершающем аспекте настоящее изобретение относится к специфическим антителам, или антигенсвязывающим фрагментам, содержащим:
a. вариабельную область тяжелой цепи, содержащую SEQ ID NO:1, и вариабельную область легкой цепи, содержащую:
i. SEQ ID NO:109; или
ii. SEQ ID NO:113; или
iii. SEQ ID NO:121; или
iv. SEQ ID NO::132; или
v. SEQ ID NO::139; или
b. SEQ ID NO:5 и
i. SEQ ID NO:24, в которой S77 и S79 заменены на Q;
ii. SEQ ID NO:117 или
iii. SEQ ID NO:125; или
iv. SEQ ID NO::136; или
v. SEQ ID NO:143.
В одном из вариантов осуществления этого последнего аспекта изобретения анти-IgE антитело, или антигенсвязывающий фрагмент, контактирует, или контактирует и является специфичным для эпитопа, содержащего, со ссылкой на SEQ ID NO:108, остатки T373, W374, S375, R376, A377, S378, G379, P381, Q417, C418, R419, T421, P426, R427, A428 Cε3-домена и остатки D278 и T281 Cε2-домена IgE человека.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Ссылки и SEQ ID приведены в Примерах со ссылкой на Фигуры.
Фигура 1. Fab3 омализумаба содержит три точечные мутации. Fab3 омализумаба получен из омализумаба и содержит три точечные мутации дистально по отношению к антигенсвязывающим областям CDR, две в каркасной области домена VL (Ser81Arg, Gln83Arg) и одну в домене Cκ (Leu158Pro). Тяжелая и легкая цепи отмечены белым и синим, соответственно. Мутантные остатки отмечены красным, и CRDL1 отмечена зеленым, для указания на ориентацию Fab.
Фигура 2. Общая структура IgE-Fc в комплексе с Fab3 омализумаба. (A) Fab3 омализумаба связывается с IgE-Fc в стехиометрии 2:1. Fab1 (зеленый) связывается с цепью B IgE-Fc (розовый) исключительно через Cε3-домен. Fab2 (синий) взаимодействует с цепью A IgE-Fc (желтый) через Cε3-домен и незначительно взаимодействует с Cε2-доменом из цепи B IgE-Fc (розовый). (B) Два Fab образуют псевдосимметричный комплекс, примерно в два раза превышающий ось области Fcε3-4. Для ясности, Cε2-домены не показаны. (C) IgE-Fc ассиметрично изогнут в комплексе с Fab3 омализумаба. Cε2-домен из цепи B (розовый) контактирует с Fab2 (синий).
Фигура 3. Поверхность контакта между Fab3 омализумаба и IgE-Fc. Показана поверхность контакта между Fab2 Fab3 омализумаба (тяжелая и легкая цепи отмечены зеленым и желтым, соответственно) и Cε3-доменом из IgE-Fc (розовый). Маркировка остатков Fab3 омализумаба и Cε3-домена показана синим и черным цветом, соответственно. На поверхности контакта действуют водородные связи и ван-дер-ваальсовы силы. Примечательной особенностью поверхности контакта является катион-π взаимодействие между Arg419 (Cε3-домен) и Phe103 (CDRH3 Fab3 омализумаба). Боковая цепь Phe103 в основном погружена в «карман», создаваемый Thr373, Trp374, Ser375, Gln417 и Arg419 (Cε3-домен).
Фигура 4. Fab3 омализумаба и DARPin E2_79 связываются с перекрывающимися поверхностями контакта. Как Fab3 омализумаба, так и DARPin E2_7920, связываются с Cε3-доменом. Остатки IgE-Fc, которые образуют только часть поверхности контакта с Fab3 омализумаба, отмечены оранжевым, при этом те, которые образуют только часть поверхности контакта с DARPin E2_79, которая включает часть линкера Cε3-Cε4, отмечены синим. Остатки IgE-Fc отмеченные розовым, которые включают Arg419 и Met430, являются общими для поверхностей контакта с Fab3 омализумаба и DARPin E2_79.
Фигура 5. Конформационная гибкость в IgE-Fc. (A) Боковая проекция свободного IgE-Fc8, демонстрирующая его резкий, асимметричный изгиб. (B) Фронтальная проекция свободного IgE-Fc (изображение повернуто на 90° против часовой стрелки относительно изображения на (A). (C) Боковая проекция IgE-Fc из комплекса с Fab3 омализумаба, демонстрирующая его частично изогнутую конформацию. (D) Фронтальная проекция IgE-Fc в комплексе с Fab3 омализумаба (изображение повернуто на 90° против часовой стрелки относительно изображения на (C). (E) Боковая проекция полностью вытянутого IgE-Fc, захваченного анти-IgE-Fc Fab (Fab3 омализумаба)16. (F) Фронтальная проекция вытянутого IgE-Fc (изображение повернуто на 90° против часовой стрелки относительно изображения на (E).
Фигура 6. Конформационная гибкость в IgE-Fc. Гибкость IgE-Fc и разгибание из изогнутой в полностью вытянутую конформацию ранее было изучено методом молекулярной динамики16. Разгибание IgE-Fc представлено в виде свободной поверхностной энергии, как описано ранее16. (A) Вытянутая конформация IgE-Fc, захваченного в кристаллической структуре комплекса Fab3 омализумаба/IgE-Fc16. (B) Частично изогнутая конформация IgE-Fc, наблюдаемая в кристаллической структуре комплекса Fab3 омализумаба/IgE-Fc. (C) Изогнутая конформация свободного IgE-Fc7,8. Изогнутая конформация IgE-Fc занимает самый низкий энергетический уровень, в то время как частично изогнутая конформация, наблюдаемая в комплексе Fab3 омализумаба/IgE-Fc, занимает явно другой энергетический уровень (B).
Фигура 7. Нарушение взаимодействия между IgE-Fc и FcεRI. В комплексе с Fab3 омализумаба Cε3-домены принимают наиболее открытую конформацию, известную до настоящего времени для IgE-Fc, что предотвращает связывание с FcεRIα. Структура IgE-Fc в комплексе с FcεRIα8 отмечена желтым, и указаны два подсайта связывания с рецептором. Структура Fab3 омализумаба в комплексе с IgE-Fc была наложена на Cε4-домены, и Cε3-домены отмечены синим. Указаны положения His424 и Pro426 в двух структурах, чтобы подчеркнуть разные положения, принимаемые Cε3-доменами.
Фигура 8. Нарушение взаимодействия между IgE-Fc и CD23. (A) Остатки Cε3-домена, которые являются общими для поверхностей контакта как с Fab3 омализумаба, так и с CD23, обозначены розовым. (B) Совмещение изображений Cε3-доменов (темно-серые) из комплекса Fab3 омализумаба/IgE-Fc и комплекса CD23/Fcε3-411 выявило конкуренцию между CD23 (желтый) и Fab3 омализумаба (розовый).
Фигура 9. Исследование взаимодействия Fab3 омализумаба с IgE-Fc. (A) Связывание Fab3 омализумаба с IgE-Fc, захваченным через C-концевой His-маркер; Fab3 омализумаба был пропущен над IgE-Fc при следующих концентрациях: 100 нМ (черный), 50 нМ (красный), 25 нМ (зеленый), 12,5 нМ (синий), 6,2 нМ (голубой), 3,1 нМ (фиолетовый), 1,6 нМ (малиновый) и 0,8 нМ (темно-красный). Использовали стандартные методы двойного референса36; эксперименты для каждой концентрации проводили в двойном повторе. (B) Связывание второго сайта связывания Fab3 омализумаба характеризовали с использованием метода ППР в формате сэндвич-связывания. IgE-Fc был захвачен на поверхности Fab3 омализумаба, а затем добавляли вторую молекулу Fab3 омализумаба к комплексу IgE-Fc/Fab3 омализумаба при концентрациях 1000 нМ (черный), 500 нМ (красный), 250 нМ (зеленый), 125 нМ (синий), 62,5 нМ (голубой), 31,2 нМ (фиолетовый), 15,6 нМ (малиновый), 7,8 нМ (темно-красный) и 0 нМ (темно-синий). (C) Сравнение способности Fab3 омализумаба к связыванию IgE-Fc, захваченного через C-концевой His-маркер (красный), и IgE-Fc, захваченного за счет связывания с FcεRIα (синий); в каждом случае тестировали серию двукратных разведений, с наивысшей концентрацией 1000 нМ. На врезке видно, что Fab3 омализумаба все еще может связываться с комплексом IgE-Fc/FcεRIα, но с низким значением Bmax. (D) Ускоренная диссоциация комплекса IgE-Fc/FcεRIα, опосредованная Fab3 омализумаба в возрастающих концентрациях. Сначала был образован комплекс 1:1 IgE-Fc/FcεRIα путем захвата IgE-Fc на иммобилизованном FcεRIα, с последующим связыванием Fab3 омализумаба в следующих концентрациях: 5000 нМ (малиновый), 1000 нМ (фиолетовый), 200 нМ (голубой), 40 нМ (синий), 8 нМ (зеленый), 1,6 нМ (красный) и 0 нМ (черный). На врезке показан в увеличенном масштабе график ускоренного процесса диссоциации. Эксперименты для всех концентраций проводили в двойном повторе. Все эксперименты по связыванию проводили при 25°C, за исключением тех, в которых характеризовали второй сайт связывания Fab3 омализумаба (Фиг. 4B), их проводили при 5°C для сведения к минимуму аллостерического взаимодействия между двумя сайтами.
Фигура 10. Анализ прямого связывания, эксперименты по конкуренции и ускоренная диссоциация. Измеряли прямое связывание IgE-Fc с иммобилизованным Fab3 омализумаба (A), Fab омализумаба (B) и интактным омализумабом (C). Fab-фрагменты или интактное антитело ковалентно иммобилизовали при низкой плотности с использованием набора для связывания через аминогруппы (GE Healthcare); IgE-Fc пропускали над поверхностями в различных концентрациях, используя серийные двукратные разведения с наивысшей концентрацией 100 нМ. Эксперименты для всех концентраций проводили в двойном повторе. (D) Проводили эксперименты методом TR-FRET по конкурентному связыванию с IgE-Fc между Fab3 омализумаба и αγ-слитым белком. Связывание между меченым тербием αγ-слитым белком и меченым Alexa Fluor 647 IgE-Fc измеряли при возрастающих концентрациях немеченого Fab3 омализумаба в качестве ингибитора: 0 мкм (черный), 2,5 нМ (синий), 5 нМ (зеленый), 10 нМ (малиновый), 20 нМ (красный). В качестве ингибитора, Fab омализумаба влияет как на кажущуюся KD, так и на Bmax, взаимодействия между IgE-Fc и αγ-слитым белком, что указывает на некоторые свойства аллостерического ингибирования. (E) Сравнение ускоренной диссоциации комплекса IgE-Fc/sFcεRIα, опосредованной интактным омализумабом (черный), Fab омализумаба (красный) или Fab3 омализумаба (синий), каждый в концентрации 5 мкМ.
Фигура 11. Репрезентативная карта электронной плотности. Стереоизображение карты электронной плотности 2Fo-Fc, вычерченное с уровнем подрезки 1,1σ, показано для части Cε3-домена цепи A и ковалентно N-связанного олигосахаридного фрагмента на Asn394.
Фигура 12. Полученная с использованием Biacore сенсограмма диссоциации IgE-Fc от иммобилизованного sFcεRIεα. Диссоциацию контролировали в присутствии подвижного буфера (сплошная линия) или партнеров по связыванию IgE (все другие сенсограммы). Анализ проводили, как описано в методе анализа (1) Примера 2.
Фигура 13. Полученная с использованием Biacore сенсограмма диссоциации IgE-Fc от иммобилизованного sFcεRIα. Диссоциацию контролировали в присутствии контрольного супернатанта (сплошная линия) или партнеров по связыванию IgE (все другие сенсограммы). Анализ проводили, как описано в методе анализа (2) Примера 2.
Фигура 14. Анализ диссоциации меченого Alexa 488 IgE-Fc с поверхности клеток RBL-SX38. Измеренные данные по связыванию нормировали на 100% при t=0 и строили график для данных по диссоциации в виде изменения пропорции остающегося связанным IgE-Fc в зависимости от времени.
Фигура 15. Анализ эффекта терапевтического введения доз Fab омализумаба дикого типа и Fab3 омализумаба в 72-часовой модели ПКА.
Фигура 16. Остатки 224-547 человеческой последовательности IgE-Fc дикого типа (приведенной в SEQ ID NO:108), с остатками 224 и 547, выделенными жирным шрифтом. Нумерация приведена в соответствии со статьей Dorrington and Bennich (1978) Immunol. Rev. 41:3-25, вследствие чего L (Leu; лейцин) после L253 обозначен L235a (в рамке) и следующий остаток представляет собой C254. Остальные остатки пронумерованы последовательно без дополнительных добавлений. Остатки эпитопа отмечены звездочкой (*).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем документе будет использована нумерация аминокислот антитела либо из непрерывной аминокислотной последовательности антитела (например, омализумаба, содержащего последовательность VH SEQ ID NO:1 и последовательность VL SEQ ID NO:20 или SEQ ID NO:129) -так называемая нумерация «pdb» -или может быть использована общепринятая система нумерации Kabat. В случае описания обычных частей (CDR - определяющие комплементарность области или FR - каркасные области) последовательности VH или VL иммуноглобулина, они связаны в стандартном порядке (VH=FR-H1.CDR-H1.FR-H2.CDR-H2.FR-H3.CDR-H3.FR-H4; VL=FR-L1.CDR-L1.FR-L2.CDR-L2.FR-L3.CDR-L3.FR-L4). Для омализумаба нумерация «pdb» частей VH (SEQ ID NO:1) является следующей: FR-H1 (аминокислоты 1-25), CDR-H1 (26-36), FR-H2 (37-50), CDR-H2 (51-66), FR-H3 (67-98), CDR-H3 (99-110), FR-H4 (111-121); при этом нумерация Kabat является следующей: FR-H1 (аминокислоты 1-25), CDR-H1 (26-35), FR-H2 (36-49), CDR-H2 (50-65), FR-H3 (66-94), CDR-H3 (95-102), FR-H4 (103-113). Для омализумаба нумерация «pdb» частей VL (SEQ ID NO:20) является следующей: FR-L1 (аминокислоты 1-23), CDR-L1 (24-38), FR-L2 (39-53), CDR-L2 (54-60), FR-L3 (61-92), CDR-L3 (93-101), FR-L4 (102-111); при этом нумерация Kabat является следующей: FR-L1 (аминокислоты 1-23), CDR-L1 (24-34), FR-L2 (35-49), CDR-L2 (50-56), FR-L3 (57-88), CDR-L3 (89-97), FR-L4 (98-107).
Нумерация для IgE антитела является такой, как описано в Dorrington & Bennich (1978) Immunol. Rev. 41:3-25. Таким образом, полипептиды IgE-Fc, используемые по данному изобретению (смотри SEQ ID NO:108), состоят из V224-K547 (включая мутацию C225A). Как показано на Фигуре 16, используемая нумерация соответствует нумерации в Dorrington and Bennich (1978) Immunol. Rev. 41:3-25, гда L (Leu, лейцин) после положения 253 обозначен L253a и остальные остатки пронумерованы последовательно после L253a как C254 и так далее. В кристаллографических экспериментах следующие мутации также были введены в IgE-Fc для упрощения картины гликозилирования: N265Q и N371Q. Общепринято, что Cε2-область IgE-Fc занимает последовательность S226-D330. В настоящем документе ссылка на IgE может быть ссылкой на человеческий IgE (и наоборот), и также может быть ссылкой на IgE-Fc в контексте анализов и методов, описанных в настоящем документе. Последовательность Fab-плечей полноразмерного IgE человека антитела не включена в данное описание, поскольку они не присутствуют в кристаллических структурах.
В настоящем документе «омализумаб» означает имеющийся в продаже препарат ксолар® или полноразмерное IgG антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность VH SEQ ID NO:1, и легкую цепь, содержащую аминокислотную последовательность VL SEQ ID NO:20; или полноразмерное IgG антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность VH-CH1 SEQ ID NO:5, и легкую цепь, содержащую аминокислотную последовательность VL-CL SEQ ID NO:24; или полноразмерное IgG антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность VH-CH1-Fc SEQ ID NO:9, и легкую цепь, содержащую аминокислотную последовательность VL-CL SEQ ID NO:24. «Fab омализумаба» означает Fab-фрагмент, содержащий тяжелую цепь, содержащую аминокислотную последовательность VH SEQ ID NO:1, и легкую цепь, содержащую аминокислотную последовательность VL SEQ ID NO:20; или (в частности) Fab-фрагмент, содержащий тяжелую цепь, содержащую аминокислотную последовательность VH-CH1 SEQ ID NO:5, и легкую цепь, содержащую аминокислотную последовательность VL-CL SEQ ID NO:24.
Общие определения
Если нет иных указаний, все технические и научные термины, используемые в настоящем документе, имеют то значение, которое им обычно придают специалисты в области, к которой относится данное изобретение. Хотя методы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящем документе, могут быть использованы при применении на практике или при тестировании настоящего изобретения, подходящие методы и материалы описаны в настоящем документе. Все публикации, патентные заявки, патенты и другие литературные источники, упомянутые в настоящем документе, включены посредством ссылки в полном объеме. Кроме того, материалы, методы и примеры являются лишь иллюстративными и не должны быть ограничивающими.
Если иное не предусмотрено контекстом, термины в единственном числе должны включать также и термины во множественном числе, и термины во множественном числе должны включать также и термины в единственном числе. В данной заявке используемый союз «или» означает «и/или», если нет иных указаний. Кроме того, используемый термин «включая», а также его другие формы, такие как «включает» и «включено», не является ограничивающим. Также, такие термины, как «элемент» или «компонент», охватывают как элементы, так и компоненты, содержащие одну единицу, а также элементы и компоненты, содержащие более одной субъединицы, если специально не указано иначе.
В целом, используемые номенклатуры, а также методы культивирования клеток и тканей, молекулярной биологии, иммунологии, микробиологии, генетики, химии белков и нуклеиновых кислот, и гибридизации, описанные в настоящем документе, хорошо знакомы специалистам и обычно используются в данной области. Способы и методики по настоящему изобретению, как правило, используют общепринятым путем, хорошо известным в данной области, и так, как описано в различных литературных источниках общего или специального характера, которые цитируются и обсуждаются в тексте настоящей спецификации, если нет иных указаний. Ферментативные реакции и методы очистки можно выполнять в соответствии с инструкциями производителя, как обычно принято в данной области или как описано в настоящем документе. Используемые номенклатуры, а также лабораторные процедуры и методы аналитической химии, химии органического синтеза, а также медицинской и фармацевтической химии, описанные в настоящем документе, хорошо известны и широко используются в данной области. Для химического синтеза, химических анализов, создания, формулирования и доставки фармацевтических препаратов, а также для лечения пациентов используют стандартные методы.
Ниже приведено разъяснение некоторых терминов для лучшего понимания настоящего изобретения.
Используемый в настоящем документе термин «хозяин», как правило, означает индивида-человека, и особенно, когда в качестве акцепторной структуры используют человеческий или гуманизированный каркас антитела. Специалисты в данной области понимают, что в случае лечения другого хозяина могут потребоваться целенаправленные изменения антитела, или антигенсвязывающего фрагмента, для данного хозяина, чтобы избежать отторжения или для достижения большей совместимости. Известны способы использования областей CDR по настоящему изобретению и встраивания их генно-инженерными методами в надлежащий каркас или пептидную последовательность для осуществления планируемой доставки и функции в организме целого ряда хозяев. Другие хозяева могут включать млекопитающих или позвоночных других видов. Таким образом, термин «хозяин» может, альтернативно, относиться к таким животным, как мыши, обезьяны, собаки, свиньи, кролики, одомашненные свиньи (свиноматки и боровы), жвачные животные, лошади, домашняя птица, кошки, мыши, быки, собаки и тому подобное, при этом антитело, или антигенсвязывающий фрагмент, в случае необходимости, соответствующим образом конструируют для достижения совместимости с хозяином.
Используемый в настоящем документе термин «полипептид» означает любую полимерную цепь аминокислот. Термины «пептид» и «белок» используются взаимозаменяемо с термином «полипептид» и также означают полимерную цепь аминокислот. Термин «полипептид» охватывает природные и искусственные белки, белковые фрагменты и полипептидные аналоги белковой последовательности. Полипептид может быть мономерным или полимерным.
Используемый в настоящем документе термин «извлечение» означает процесс получения химического соединения, такого как полипептид, в форме, практически свободной от компонентов, связанных с ним естественным образом, путем выделения, например, с использованием методов очистки белков, хорошо известных в данной области.
Используемые в настоящем документе термины «специфическое связывание» или «специфически связывающие» со ссылкой на взаимодействию антитела, белка или пептида со вторым химическим соединением означают, что взаимодействие зависит от наличия конкретной структуры (например, «антигенной детерминанты» или «эпитопа», определение которым дано ниже) на химическом соединении; например, антитело узнает и связывается с конкретной белковой структурой, а не с любыми белками. Если антитело специфично для эпитопа «A», присутствие молекулы, содержащей эпитоп A (или свободного, немеченого A), в реакционной смеси, содержащей меченый «A» и антитело, будет приводить к уменьшению количества меченого A, связанного с антителом. Если в настоящем документе упомянут эпитоп, то анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению является специфичным для указанного эпитопа.
Используемый в настоящем документе термин «антитело» в широком смысле означает любую молекулу иммуноглобулина (Ig), состоящую из четырех полипептидных цепей, двух тяжелых (H) цепей и двух легких (L) цепей, или любой ее функциональный фрагмент, мутант, вариант или производное, которые сохраняют по меньшей мере некоторую часть особенностей связывания эпитопа, присущих молекуле Ig, что позволяет им специфически связываться с IgE. Такие форматы мутанта, варианта или производного антитела известны в данной области и описаны ниже. Неограничивающие варианты осуществления таких форматов известны в данной области и описаны ниже. Говорят, что антитело «способно связывать» молекулу (или эпитоп), если оно способно специфически реагировать с молекулой (или эпитопом), в результате чего происходит связывание молекулы (или эпитопа) с антителом.
Используемый в настоящем документе термин «моноклональное антитело» означает препарат молекул антитела, имеющих одни и те же аминокислотные последовательности тяжелой цепи и одни и те же аминокислотные последовательности легкой цепи, либо любого их функционального фрагмента, мутанта, варианта или производного, которые сохраняют по меньшей мере те же характерные для легкой цепи особенности связывания эпитопа, что и молекула Ig, в отличие от препаратов «поликлональных» антител, которые содержат смесь разных антител. Моноклональные антитела могут быть получены с помощью нескольких известных технологий, таких как фаговый, бактериальный, дрожжевой или рибосомный дисплей, а также классическими методами, например, антитела, полученные из гибридом (например, антитело, секретируемое гибридомой, полученной с использованием гибридомной технологии, такой как стандартная методология Kohler и Milstein ((1975) Nature 256:495-497).
В полноразмерном антителе каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (в настоящем документе используют аббревиатуру HCVR или VH) и константной области тяжелой цепи (CH). Константная область тяжелой цепи состоит из четырех доменов-или CH1, шарнир, CH2 и CH3 (тяжелые цепиγ, α иδ), или CH1, CH2, CH3 и CH4 (тяжелые цепи μ и ε). Каждая легкая цепь состоит из вариабельной области легкой цепи (в настоящем документе используют аббревиатуру LCVR или VL) и константной области легкой цепи (CL). Константная область легкой цепи состоит из одного домена, CL. Области VH и VL можно дополнительно подразделять на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые чередуются с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных в направлении от амино-конца к карбоксильному концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Молекулы иммуноглобулинов могут относиться к любому типу (например, IgG, IgE, IgM, IgD, IgA и IgY), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу.
Используемый в настоящем документе термин «антигенсвязывающий фрагмент» означает один или более фрагментов, или частей антитела, которые сохраняют способность специфически связываться с антигеном (например, IgE), либо синтетические модификации фрагментов антитела, которые сохраняют желаемую способность связываться с антигеном. Показано, что функция связывания антигена у антитела может осуществляться фрагментами или некоторыми частями полноразмерного антитела, либо их модификациями. Варианты осуществления включают биспецифические, с двойной специфичностью, а также мультиспецифические форматы, которые способны специфически связываться с двумя или более разными антигенами или с несколькими эпитопами или прерывистыми областями эпитопов антигена. Неограничивающие примеры антигенсвязывающих фрагментов включают (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab′)2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанные дисульфидной связью в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546, Winter et al., PCT публикация WO 90/05144 A1, содержание которых включено посредством ссылки), содержащий один вариабельный домен; (vi) выделенную определяющую комплементарность область (CDR), (vii) слитые продукты фрагментов антитела, такие как те, которые имеют иммуноглобулиновую природу, например, диатела, scAb, биспецифические антитела, триатела, Fab-Fv, Fab-Fv-Fv, триотела, (Fab-Fv)2-Fc, и (viii) части антитела, такие как CDR или петли антитела, привитые на не иммуноглобулиновые каркасы, например, фибронектин или лейциновые «застежки» (смотри Binz et al. (2005) Nat. Biotech. 23:1257-1268, содержание публикации включено посредством ссылки). Кроме того, хотя два домена Fv-фрагмента, VL и VH, закодированы разными генами, они могут быть соединены рекомбинантными или другими методами с помощью синтетического или природного линкера, который позволяет им существовать в виде одной белковой цепи, в которой области VL и VH попарно связаны, с образованием одновалентных молекул (известных как одноцепочечные Fv (scFv); смотри, например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такие одноцепочечные антитела также должны быть охвачены термином «антигенсвязывающий фрагмент». Другие формы одноцепочечных антител, такие как диатела, также охвачены этим термином. Диатела представляют собой двухвалентные, биспецифические антитела, в которых домены VH и VL экспрессируются в виде одной полипептидной цепи, но с использованием линкера, который является слишком коротким, чтобы допустить образование пар между двумя доменами на одной и той же цепи, в результате чего домены вынужденно образуют пары с комплементарными доменами другой цепи, с образованием двух антигенсвязывающих сайтов (смотри, например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak, R. J., et al. (1994) Structure 2:1121-1123). Такие связывающие фрагменты антитела известны в данной области (Kontermann and Dubel eds., Antibody Engineering (2001) Springer-Verlag. New York. 790 pp. (ISBN 3-540-41354-5).
Используемый в настоящем документе термин «конструкт антитела» означает полипептид, содержащий один или более антигенсвязывающих фрагментов по изобретению, связанные с линкерным полипептидом или константным доменом иммуноглобулина. Линкерные полипептиды содержат два или более аминокислотных остатков, связанные пептидными связями, и используются для связывания одного или более антигенсвязывающих фрагментов. Такие линкерные полипептиды хорошо известны в данной области (смотри, например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak, R. J., et al. (1994) Structure 2:1121-1123). Константный домен иммуноглобулина означает константный домен тяжелой или легкой цепи, например, константные домены IgA, IgD, IgE, IgG или IgM человека. Аминокислотные последовательности константных доменов тяжелой цепи и легкой цепи известны в данной области. Неограничивающие примеры константных областей тяжелой цепи γ1 Ig и легких цепей λ и κ Ig приведены в Таблицах 8 и 6, соответственно.
Кроме того, антитело, или его антигенсвязывающий фрагмент, может быть частью большей по размеру молекулы иммуноадгезина, образованной в результате ковалентной или нековалентной ассоциации антитела, или фрагмента антитела, с одним или более другими белками или пептидами. Примеры создания таких молекул иммуноадгезинов включают использование коровой области стрептавидина для создания тетрамерной молекулы scFv (Kipriyanov, S. M., et al. (1995) Human Antibodies and Hybridomas 6:93-101) и использование остатка цистеина, маркерного пептида и C-концевого полигистидинового маркера для создания двухвалентных и биотинилированных молекул scFv (Kipriyanov, S. M., et al. (1994) Mol. Immunol. 31:1047-1058). Фрагменты антитела, такие как фрагменты Fab и F(ab')2, могут быть получены из целых антител общепринятыми методами, такими как расщепление папаином или пепсином, соответственно, целых антител. Кроме того, антитела, фрагменты антител и молекулы иммуноадгезинов могут быть получены стандартными методами рекомбинантных ДНК, как описано в настоящем документе.
Используемый в настоящем документе термин «выделенное антитело» означает антитело, которое является практически свободным от других антител, имеющих другие антигенные специфичности (например, выделенное антитело, которое специфически связывает IgE, является практически свободным от других антител, которые специфически связывают антигены, отличные от IgE). Однако выделенное антитело, которое специфически связывает, например, человеческий IgE, может иметь перекрестную реактивность с другими антигенами, такими как молекулы IgE из других биологических видов. Кроме того, выделенное антитело может быть практически свободным от другого клеточного материала и/или химических реагентов.
Термин «CDR-привитое антитело» относится к антителам, которые содержат последовательности вариабельной области тяжелой и легкой цепи из одного биологического вида, но в которых последовательности одной или более из областей CDR VH и/или VL заменены последовательностями CDR из другого биологического вида, например, антитела, имеющие вариабельные области тяжелой и легкой цепи иммуноглобулина человека, в которых одна или более из человеческих CDR (например, CDR3) заменены на последовательности CDR мыши.
В настоящем документе термины «нумерация Kabat», «определения Kabat» и «маркировка Kabat» используют взаимозаменяемо. Эти термины, признанные в данной области, относятся к системе нумерации аминокислотных остатков, которые являются более вариабельными (то есть, гипервариабельными), чем другие аминокислотные остатки, в вариабельных областях тяжелой и легкой цепей антитела, или его антигенсвязывающего фрагмента (Kabat et al. (1971) Ann. NY Acad, Sci. 190:382-391 и Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, пятое издание, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). В вариабельной области тяжелой цепи гипервариабельная область включает аминокислотные положения 31-35 (CDR-H1), остатки 50-65 (CDR-H2) и остатки 95-102 (CDR-H3) в соответствии с системой нумерации Kabat. Однако в соответствии с системой Chothia (Chothia et al., (1987) J. Mol. Biol., 196, 901-917 (1987)), петля, эквивалентная CDR-H1, имеет протяженность от остатка 26 до остатка 32. Таким образом, если нет иных указаний, при использовании в настоящем документе «CDR-H1» соответствует остаткам 26-35, что является комбинацией системы нумерации Kabat и определения топологической петли Chothia. В вариабельной области легкой цепи гипервариабельная область включает аминокислотные положения 24-34 для CDRL1, аминокислотные положения 50-56 для CDRL2 и аминокислотные положения 89-97 для CDRL3.
Используемые в настоящем документе термины «акцептор» и «акцепторное антитело» означают антитело, или нуклеотидную последовательность, которое содержит, или кодирует, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% аминокислотных последовательностей одной или более из каркасных областей. В некоторых вариантах осуществления термин «акцептор» означает аминокислотную или нуклеотидную последовательность антитела, которая представляет собой, или кодирует, константную область(и). В другом варианте осуществления термин «акцептор» означает аминокислотную или нуклеотидную последовательность антитела, которая представляет собой, или кодирует, одну или более из каркасных областей и константных областей. В конкретном варианте осуществления термин «акцептор» означает аминокислотную или нуклеотидную последовательность антитела человека, которая представляет собой, или кодирует, по меньшей мере 80%, предпочтительно, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% аминокислотных последовательностей одной или более из каркасных областей. В данном варианте осуществления акцептор может содержать по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5 или по меньшей мере 10 аминокислотных остатков, которые не присутствуют в одном или более конкретных положениях в антителе человека. Акцепторная каркасная область и/или акцепторная константная область(и) могут, например, быть производными от, или быть получены из, гена антитела зародышевой линии, гена зрелого антитела, функционального антитела (например, антител, хорошо известных в данной области, антител, находящихся в процессе разработки, или антител, коммерчески доступных).
Используемый в настоящем документе термин «CDR» означает определяющую комплементарность область в составе вариабельных последовательностей антитела. Существуют три CDR в каждой из вариабельных областей тяжелой цепи и легкой цепи, которые обозначены CDRH1, CDRH2 и CDRH3 в случае CDR тяжелой цепи, и CDRL1, CDRL2 и CDRL3 в случае CDR легкой цепи. Используемый в настоящем документе термин «набор CDR» означает группу из трех CDR, которые присутствуют в одной вариабельной области, способной связывать антиген. Точные границы этих CDR определяют по-разному в соответствии с разными системами. Система, описанная Kabat (Kabat et al., Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987) и (1991)) не только предоставляет однозначную систему нумерации остатков, применимую к любой вариабельной области антитела, но также позволяет точно разграничивать остатки, составляющие три CDR. Эти CDR могут быть названы Kabat CDR. Chothia и сотрудники (Chothia & Lesk, J. Mol. Biol. 196:901-917 (1987) и Chothia et al., Nature 342:877-883 (1989)) обнаружили, что некоторые субфрагменты в составе Kabat CDR принимают почти идентичные конформации пептидного каркаса, несмотря на большие различия на уровне аминокислотной последовательности. Эти субфрагменты были обозначены L1, L2 и L3 или H1, H2 и H3, где «L» и «H» означают области легкой цепи и области тяжелой цепи, соответственно. Эти области могут быть названы Chothia CDR, они имеют границы, которые перекрываются с Kabat CDR. Другие границы, определяющие CDR, перекрывающиеся с Kabat CDR, были описаны Padlan (FASEB J. 9:133-139 (1995)) и MacCallum (J Mol Biol 262(5):732-45 (1996)). Другие определения границ CDR могут не следовать строго одной из вышеописанных систем, но, тем не менее, будет иметь место перекрывание с Kabat CDR, хотя они могут быть укороченными или удлиненными с учетом предсказания или экспериментальных результатов, указывающих на то, что конкретные остатки или группы остатков, или даже целые CDR не оказывают существенного влияния на связывание антигена. В способах, описанных в настоящем документе, могут быть использованы CDR, границы которых определены в соответствии одной из описанных систем, хотя в предпочтительных вариантах осуществления используют CDR, границы которых определены в соответствии с системой Kabat или Chothia, или их комбинацией.
Используемый в настоящем документе термин «канонический» остаток относится к остатку в CDR или каркасе, который определяет конкретную каноническую структуру CDR в соответствии с системой Chothia et al. (J. Mol. Biol. 196:901-907 (1987); Chothia et al., J. Mol. Biol. 227:799 (1992), обе публикации включены в настоящий документ посредством ссылки). В соответствии с Chothia et al., особо важные части CDR многих антител имеют почти идентичные конформации пептидного каркаса, несмотря на большие различия на уровне аминокислотной последовательности. Каждая каноническая структура определяет, в основном, набор углов поворота пептидного каркаса, чтобы непрерывный сегмент аминокислотных остатков образовывал петлю.
Используемые в настоящем документе термины «донор» и «донорское антитело» относятся к антителу, предоставляющему одну или более CDR. В предпочтительном варианте осуществления донорское антитело представляет собой антитело от биологического вида, иного, чем вид, от которого каркасные области антитела были получены или являются производными. В контексте гуманизированного антитела термин «донорское антитело» означает антитело, не являющееся человеческим, которое предоставляет одну или более CDR.
Используемый в настоящем документе термин «каркас» или «каркасная последовательность» относится к оставшимся последовательностям вариабельной области за вычетом областей CDR. Поскольку точные последовательности CDR могут быть определены в соответствии с разными системами, определение каркасной последовательности, соответственно, может иметь разные интерпретации. Шесть областей CDR (CDR-L1, -L2 и -L3 легкой цепи и CDR-H1, -H2 и -H3 тяжелой цепи) также разделяют каркасные области в легкой цепи и тяжелой цепи на четыре подобласти (FR1, FR2, FR3 и FR4) в каждой цепи, при этом CDR1 расположена между FR1 и FR2, CDR2 между FR2 и FR3, и CDR3 между FR3 и FR4. Без обозначения конкретных подобластей как FR1, FR2, FR3 или FR4, каркасная область, как отмечено другими авторами, представляет собой объединенные FR в вариабельной области одной природной цепи иммуноглобулина. При использовании в настоящем документе «область FR» означает одну из четырех подобластей, а «области FR» означает две или более из четырех подобластей, составляющих каркасную область.
Используемый в настоящем документе термин «ген антитела зародышевой линии» или «генный фрагмент» означает последовательность иммуноглобулина, закодированную в нелимфоидных клетках, которые не прошли процесс созревания, приводящий к генетической перегруппировке и мутации для экспрессии конкретного иммуноглобулина. Смотри, например, Shapiro et al., Crit. Rev. Immunol. 22(3):183-200 (2002); Marchalonis et al., Adv Exp Med Biol. 484:13-30 (2001). Одно из преимуществ, обеспечиваемых разными вариантами осуществления настоящего изобретения, основано на признании того, что в генах антител зародышевой линии с большей вероятностью, чем в генах зрелых антител, сохраняются необходимые структуры аминокислотной последовательности, характерные для индивидуумов в биологическом виде, следовательно, они будут с меньшей вероятностью распознаваться, как происходящие из постороннего источника, при терапевтическом использовании для данных видов.
Используемый в настоящем документе термин «ключевые» остатки относится к определенным остаткам в вариабельной области, которые оказывают большее воздействие на специфичность и/или аффинность связывания антитела, в частности, гуманизированного антитела. Ключевой остаток включает, но без ограничения, одно или более из следующего: остаток, примыкающий к CDR, потенциальный сайт гликозилирования (может быть сайтом либо N-, либо O-гликозилирования), редкий остаток, остаток, способный взаимодействовать с антигеном, остаток, способный взаимодействовать с CDR, канонический остаток, контактный остаток между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, остаток в зоне Верньера и остаток в области перекрывания между CDR1 вариабельной области тяжелой цепи по определению Chothia и первой каркасной областью тяжелой цепи по определению Kabat.
Термин «гуманизированное антитело», как правило, относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепей от биологического вида, отличного от человека (например, кролика, мыши и так далее), но в которых по меньшей мере часть последовательности VH и/или VL была изменена, чтобы стать более «похожей на человеческую», то есть, более похожей на вариабельные последовательности зародышевой линии человека. Одним видом гуманизированного антитела является CDR-привитое антитело, в котором человеческие последовательности CDR встроены в не принадлежащие человеку последовательности VH и VL для замены соответствующих последовательностей CDR, отличных от человеческих. Другим видом гуманизированного антитела является CDR-привитое антитело, в котором по меньшей мере одна не принадлежащая человеку CDR встроена в каркас человеческого иммуноглобулина. Настоящее изобретение, как правило, сфокусировано на последнем варианте.
В частности, используемый в настоящем документе термин «гуманизированное антитело» означает антитело, либо его вариант, производное, аналог или фрагмент, которое иммуноспецифически связывается с интересующим антигеном и которое содержит каркасную (FR) область, имеющую, по существу, аминокислотную последовательность человеческого антитела, и определяющую комплементарность область (CDR), имеющую, по существу, аминокислотную последовательность антитела, отличного от человеческого. Используемый в настоящем документе термин «по существу» в контексте CDR относится к CDR, имеющей аминокислотную последовательность, на по меньшей мере 50, 55, 60, 65, 70, 75 или 80%, предпочтительно по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичную аминокислотной последовательности CDR антитела, отличного от человеческого. В одном из вариантов осуществления гуманизированное антитело содержит область CDR, имеющую одну или более (например, 1, 2, 3 или 4) аминокислотных замен, добавлений и/или делеций по сравнению с CDR антитела, отличного от человеческого. Кроме того, не принадлежащая человеку CDR может быть генетически модифицирована, чтобы стать более «похожей на человеческую» или совместимой с организмом человека, с использованием известных методов. Гуманизированное антитело содержит практически все из по меньшей мере одного и, как правило, двух вариабельных доменов (Fab, Fab′, F(ab′)2, F(ab')c, Fv), в которых все или практически все из областей CDR соответствуют областям CDR иммуноглобулина, отличного от человеческого (то есть, донорского антитела), и все или практически все из каркасных областей представляют собой каркасные области консенсусной последовательности человеческого иммуноглобулина. Предпочтительно, гуманизированное антитело также содержит по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, человеческого иммуноглобулина. В некоторых вариантах осуществления гуманизированное антитело содержит как легкую цепь, так и по меньшей мере вариабельный домен тяжелой цепи. Антитело также может содержать CH1, шарнир, CH2 и CH3, или CH1, CH2, CH3 и CH4 тяжелой цепи. В некоторых вариантах осуществления гуманизированное антитело содержит только гуманизированную легкую цепь. В некоторых вариантах осуществления гуманизированное антитело содержит только гуманизированную тяжелую цепь. В конкретных вариантах осуществления гуманизированное антитело содержит только гуманизированный вариабельный домен легкой цепи и/или гуманизированную тяжелую цепь. Хотя некоторые мутации, описанные в настоящем документе, могут не быть обычными «очеловечивающими», это не является основанием для того, чтобы не считать анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению «гуманизированным».
Гуманизированное антитело можно выбирать из любого класса иммуноглобулинов, включая IgY, IgM, IgG, IgD, IgA и IgE, и любого изотипа, включая, без ограничения, IgA1, IgA2, IgG1, IgG2, IgG3 и IgG4. Гуманизированное антитело может содержать последовательности из иммуноглобулина более чем одного класса или изотипа, и конкретные константные домены можно выбирать для оптимизации желательных эффекторных функций с использованием методов, хорошо известных в данной области.
Области каркаса и CDR гуманизированного антитела не обязательно должны точно соответствовать исходным последовательностям, например, CDR донорского антитела или консенсусный каркас могут быть подвергнуты мутагенезу путем замены, вставки и/или делеции по меньшей мере одного аминокислотного остатка, так что остаток CDR или каркаса в данном сайте не соответствует точно ни донорскому антителу, ни консенсусному каркасу. В предпочтительном варианте осуществления такие мутации, однако, не будут обширными. Как правило, по меньшей мере 50, 55, 60, 65, 70, 75 или 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, и наиболее предпочтительно по меньшей мере 95%, 98% или 99% остатков гуманизированного антитела будут соответствовать остаткам в исходных последовательностях FR и CDR. В одном из вариантов осуществления одна или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) аминокислотных замен, добавлений и/или делеций могут иметь место в гуманизированном антителе по сравнению с исходными последовательностями FR и CDR (например, в сравнении с последовательностями омализумаба или Fab омализумаба). Используемый в настоящем документе термин «консенсуный каркас» означает каркасную область в консенсусной последовательности иммуноглобулина. Используемый в настоящем документе термин «консенсусная последовательность иммуноглобулина» означает последовательность, образованную из наиболее часто встречающихся аминокислот (или нуклеотидов) в семействе родственных последовательностей иммуноглобулинов (смотри, например, Winnaker, From Genes to Clones (Verlagsgesellschaft, Weinheim, Germany 1987). В семействе иммуноглобулинов каждое положение в консенсусной последовательности занято аминокислотой, встречающейся наиболее часто в этом положении в данном семействе. Если две аминокислоты встречаются с одинаковой частотой, любую из них можно включать в консенсусную последовательность.
Используемый в настоящем документе термин «зона Верньера» означает подгруппу каркасных остатков, которые могут регулировать структуру CDR и точно настраивать ее для соответствия антигену, как описано Foote и Winter (1992, J. Mol. Biol. 224:487-499, содержание публикации включено в настоящий документ посредством ссылки). Остатки зоны Верньера образуют слой, находящийся под CDR, и могут влиять на структуру CDR и аффинность антитела.
Используемый в настоящем документе термин «нейтрализация» означает нейтрализацию биологической активности IgE, когда анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению описанное в настоящем документе, специфически связывает белок IgE. Нейтрализация может быть результатом разных вариантов связывания указанного антитела с IgE. Предпочтительно, нейтрализующее антитело представляет собой антитело, связывание которого с IgE приводит к нейтрализации биологической активности IgE. Предпочтительно, нейтрализующий связывающий белок связывает IgE и уменьшает биологическую активность IgE на по меньшей мере примерно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 50%, 60%, 80%, 85% или более. Нейтрализацию биологической активности IgE нейтрализующим антителом можно оценивать путем измерения одного или более показателей биологической активности IgE, описанных в настоящем документе.
Используемый в настоящем документе термин «нейтрализующее моноклональное антитело» означает препарат молекул антитела, которые при связывании с IgE способны ингибировать или уменьшать биологическую активность IgE или частично, или полностью.
Используемые в настоящем документе термины «ослабление», «ослаблять», и тому подобные, означают снижение или уменьшение степени тяжести симптома или состояния, вызванного повышенными сывороточными уровнями IgE.
Термины «эпитоп» или «антигенная детерминанта» охватывают любую детерминанту полипептида, которая способна к специфическому связыванию с иммуноглобулином или T-клеточным рецептором. В конкретных вариантах осуществления антигенные детерминанты включают химически активные поверхностные группировки таких молекул, как аминокислоты, сахарные боковые цепи, фосфорил или сульфонил, и в конкретных вариантах осуществления они могут иметь специфические характеристики трехмерной структуры и/или специфические характеристики заряда. Эпитоп представляет собой область антигена, которая связывается антителом. В конкретных вариантах осуществления говорят, что антитело специфически связывает антиген, когда оно предпочтительно узнает свой антиген-мишень в сложной смеси белков и/или макромолекул.
Используемый в настоящем документе термин «kon» означает константу скорости ассоциации антитела с антигеном, с образованием комплекса антитело/антиген, как известно в данной области.
Используемый в настоящем документе термин «koff» означает константу скорости диссоциации антитела из комплекса антитело/антиген, как известно в данной области.
Используемый в настоящем документе термин «kd», или «kD», означает константу диссоциации для конкретного взаимодействия антитела с антигеном, как известно в данной области.
Сила, или аффинность, взаимодействия при иммунологическом связывании может быть выражена в виде константы диссоциации (kD или kd) взаимодействия, при этом меньшее значение kd соответствует большей или более высокой аффинности. Показатели иммунологического связывания выбранных полипептидов могут быть рассчитаны с использованием методов, хорошо известных в данной области. Один такой метод включает измерение скоростей образования и диссоциации комплекса антигенсвязывающий сайт/антиген, при этом данные скорости зависят от концентраций партнеров по комплексу, аффинности взаимодействия и геометрических параметров, которые в равной степени влияют на скорость обоих процессов. Таким образом, как «константу скорости ассоциации» («kon»), так и «константу скорости диссоциации» («koff»), можно определять путем расчета концентраций и фактических скоростей ассоциации и диссоциации. (Nature 361:186-87 (1993)). Отношение koff/kon позволяет отбрасывать все параметры, не связанные с аффинностью, и равно константе диссоциации kd. Davies et al. (1990) Annual Rev Biochem 59:439-473.
Термин «конъюгат антитела» означает связывающий белок, такой как антитело или фрагмент антитела, либо его связывающий фрагмент, химически связанный со вторым химическим соединением, таким как терапевтическое или цитотоксическое средство. Термин «средство» используют в настоящем документе для обозначения химического соединения, смеси химических соединений, биологической макромолекулы или экстракта, полученного из биологических материалов.
Используемые в настоящем документе термины «кристалл» и «кристаллизованные» относятся к антителу, или его антигенсвязывающему фрагменту, находящемуся в форме кристалла. Кристаллы представляют собой одну из форм твердого состояния вещества, отличную от других форм, таких как аморфное твердое состояние или жидкое кристаллическое состояние. Кристаллы состоят из регулярных, повторяющихся трехмерных решеток атомов, ионов, молекул (например, белков, таких как антитела) или молекулярных агрегатов (например, комплексов антиген/антитело). Эти трехмерные решетки расположены в соответствии с определенными математическими соотношениями, которые хорошо известны в данной области. Фундаментальная единица или строительный блок, который повторяется в кристалле, называется асимметричной единицей. Повторяющиеся асимметричные единицы в пространственной конфигурации, которая соответствует заданной четко определенной кристаллографической симметрии, создают «элементарную ячейку» кристалла. Повторяющиеся элементарные ячейки за счет регулярных переносов (трансляций) во всех трех измерениях создают кристалл. Смотри Giege, R. and Ducruix, A. Barrett, Crystallization of Nucleic Acids and Proteins, a Practical Approach, 2-е изд., pp. 201-16, Oxford University Press, New York, N.Y., (1999).
В настоящем документе термин «полинуклеотид» означает полимерную форму из двух или более нуклеотидов, либо рибонуклеотидов, либо дезоксинуклеотидов, либо модифицированных форм нуклеотидов каждого вида. Термин включает одно- и двухцепочечные формы ДНК, однако предпочтительно означает двухцепочечную ДНК.
Используемый в настоящем документе термин «выделенный полинуклеотид» означает полинуклеотид (например, геномного, кДНК или синтетического происхождения, либо их комбинацию), который, в силу своего названия, «выделенный полинуклеотид», не связан со всем или частью полинуклеотида, с которым «выделенный полинуклеотид» встречается в природе; функционально связан с полинуклеотидом, с которым он не связан в природе; или не существует в природе в виде части большей по размеру последовательности.
Используемый в настоящем документе термин «вектор» означает молекулу нуклеиновой кислоты, способную переносить другую нуклеиновую кислоту, с которой она была связана. Одним видом вектора является «плазмида», которая представляет собой кольцевую двухцепочечную петлю ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим видом вектора является вирусный вектор, в этом случае дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, не эписомные векторы млекопитающих) могут интегрироваться в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. В настоящем документе такие векторы называют «рекомбинантными экспрессионными векторами» (или просто, «экспрессионными векторами»). Как правило, экспрессионные векторы, используемые в методах рекомбинантных ДНК, часто имеют форму плазмиды. В настоящей спецификации термины «плазмида» и «вектор» могут быть использованы взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако изобретение должно охватывать и другие формы экспрессионных векторов, такие как вирусные векторы (например, репликативно-дефектные ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Термин «функционально связанные» относится к смежному расположению компонентов, при котором описанные компоненты находятся во взаимоотношениях, позволяющих им функционировать предназначенным для них образом. Последовательность контроля, «функционально связанная» с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, определяемых последовательностями контроля. «Функционально связанные» последовательности включают как последовательности контроля экспрессии, которые расположены в непосредственной близости от интересующего гена, так и последовательности контроля экспрессии, которые действуют в «транс» положении, или на расстоянии, контролируя интересующий ген. Используемый в настоящем документе термин «последовательность контроля экспрессии» означает полинуклеотидные последовательности, которые необходимы для осуществления экспрессии и процессинга кодирующих последовательностей, с которыми они лигированы. Последовательности контроля экспрессии включают соответствующие последовательности инициации, терминации транскрипции, последовательности промоторов и энхансеров; сигналы эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, повышающие эффективность трансляции (то есть, консенсусная последовательность Kozak); последовательности, повышающие стабильность белка, и, в случае необходимости, последовательности, повышающие секрецию белка. Природа таких последовательностей контроля бывает разной в зависимости от организма хозяина; у прокариот такие последовательности контроля, как правило, включают промотор, сайт связывания рибосомы и последовательность терминации транскрипции; у эукариот, как правило, такие последовательности контроля включают промоторы и последовательность терминации транскрипции. Термин «последовательности контроля» включает компоненты, присутствие которых необходимо для экспрессии и процессинга, и также может включать дополнительные компоненты, присутствие которых полезно, например, лидерные последовательности и последовательности партнера по слиянию.
Используемый в настоящем документе термин «трансформация» означает любой процесс, за счет которого экзогенная ДНК проникает в клетку-хозяина. Трансформация может происходить в естественных или искусственных условиях, при использовании различных методов, хорошо известных в данной области. Трансформация может быть основана на любом известном методе введения чужеродных нуклеотидных последовательностей в прокариотическую или эукариотическую клетку-хозяина. Метод выбирают с учетом подвергаемой трансформации клетки-хозяина, и он может включать, но не ограничивается ими, вирусную инфекцию, электропорацию, липофекцию и бомбардировку частицами. Такие «трансформированные» клетки включают стабильно трансформированные клетки, в которых введенная ДНК способна к репликации либо в виде автономно реплицирующейся плазмиды, либо в виде части хромосомы хозяина. Они также включают клетки, которые временно экспрессируют введенную ДНК или РНК в течение ограниченных периодов времени.
Используемый в настоящем документе термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») означает клетку, в которую была введена экзогенная ДНК. Следует понимать, что такие термины должны означать не только конкретную указанную клетку, но также и потомство такой клетки. Поскольку определенные модификации могут иметь место в последующих поколениях вследствие либо мутаций, либо влияния окружающей среды, такое потомство фактически может не быть идентичным исходной клетке, однако оно все-еще попадает под определение используемого в настоящем документе термина «клетка-хозяин». Предпочтительные клетки-хозяева включают прокариотические и эукариотические клетки, выбранные из любых царств живых организмов. Предпочтительные эукариотические клетки включают клетки простейших, грибков, растений и животных. Наиболее предпочтительно, клетки-хозяева включают, но не ограничиваются ими, прокариотическую линию клеток E. coli; линии клеток CHO, HEK 293 и COS млекопитающих; линию клеток Sf9 насекомых и клетки грибков Saccharomyces cerevisiae.
Можно использовать стандартные методы для получения рекомбинантных ДНК, олигонуклеотидного синтеза, а также культивирования тканей и трансформации (например, электропорацию, липофекцию). Ферментативные реакции и методы очистки можно использовать в соответствии с инструкциями производителя, как обычно принято в данной области или как описано в настоящем документе. Вышеупомянутые методы и процедуры, как правило, могут быть использованы общепринятым путем, хорошо известным в данной области, и так, как описано в различных литературных источниках общего или специального характера, которые цитируются и обсуждаются в тексте настоящей спецификации. Смотри, например, сборник Sambrook et al. Molecular Cloning: A Laboratory Manual (2-е издание, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)), содержание которого включено в настоящий документ посредством ссылки для любых целей.
Используемый в настоящем документе термин «эффективное количество» означает количество терапевтического средства, которое достаточно для ослабления или уменьшения степени тяжести и/или длительности заболевания, либо одного или более его симптомов, профилактики прогрессирования заболевания, вызывания регрессии заболевания, профилактики рецидива, развития, возникновения или прогрессирования одного или более симптомов, связанных с заболеванием, обнаружения заболевания, либо усиления или улучшения профилактического или терапевтического эффекта(ов) другого лекарственного средства (например, профилактического или терапевтического средства).
Карту конкретных областей или эпитопов человеческого белка IgE, описанного в настоящем документе, можно определять любым подходящим способом картирования эпитопов, известным в данной области, в сочетании с любым из антител, предложенных по настоящему изобретению. Примеры таких способов включают скрининг пептидов разной длины, полученных из IgE, на связывание с антителом по настоящему изобретению, при этом наименьший фрагмент, который специфически связывается с антителом, содержит последовательность эпитопа, узнаваемого антителом. Пептиды IgE могут быть получены путем синтеза или путем протеолитического расщепления белка IgE. Пептиды, которые связывают антитело, могут быть идентифицированы, например, при помощи масс-спектроскопического анализа. В другом примере можно использовать метод ЯМР-спектроскопии или рентгеновской кристаллографии для идентификации эпитопа, связанного антителом по настоящему изобретению. Методы кристаллизации и рентгеновской кристаллографии являются предпочтительными для определения структуры IgE и эпитопа на IgE, с которым связывается анти-IgE антитело, или антигенсвязывающий фрагмент, по изобретению.
Антитела, используемые по изобретению, можно получать методами получения антител из одиночных лимфоцитов путем клонирования и экспрессии кДНК вариабельных областей иммуноглобулинов, полученных из одиночных лимфоцитов, отобранных на основании продуцирования специфических антител, например, методами, описанными в Babcook, J. et al., 1996, Proc. Natl. Acad. Sci. USA 93(15):7843-7848l; WO92/02551; WO2004/051268 и международной патентной заявке номер WO2004/106377. Скрининг на антитела можно проводить, используя анализы для измерения связывания с IgE человека и/или анализы для измерения способности блокировать связывание IgE с его естественным рецептором. Примером анализа на связывание является ELISA.
Гуманизированные антитела (которые включают CDR-привитые антитела) представляют собой молекулы антител, имеющие одну или более определяющих комплементарность областей (CDR) иммуноглобулина от биологического вида, отличного от человека (например, кролика или мыши), и каркасную область из молекулы человеческого иммуноглобулина (смотри, например, US 5585089; WO91/09967). Следует понимать, что может быть необходимо переносить только определяющие специфичность остатки областей CDR, а не целые CDR (смотри, например, Kashmiri et al., 2005, Methods, 36, 25-34). Гуманизированные антитела могут, необязательно, также содержать один или более каркасных остатков из иммуноглобулина биологического вида, отличного от человека, из которого были получены CDR. Последние часто называют донорскими остатками. Молекулы антитела по настоящему изобретению предпочтительно имеют аффинность связывания (KD) менее 2 нМ. Аффинность можно измерять любым подходящим методом, известным в данной области, включая BIAcore, как описано в разделе «Примеры» настоящего документа (смотри Пример 6), с использованием выделенного природного или рекомбинантного IgE, либо подходящего слитого белка/полипептида.
Аффинность антитела, или антигенсвязывающего фрагмента, по настоящему изобретению, а также степень, до которой связывающее соединение (такое как антитело) ингибирует связывание, может определять специалист в данной области с использованием общепринятых методов, например, тех, которые описаны в Scatchard et al. (Ann. KY. Acad. Sci. 51:660-672 (1949)), или метода поверхностного плазмонного резонанса (ППР) с использованием таких систем, как BIAcore. В методе поверхностного плазмонного резонанса молекулы-мишени иммобилизуют на твердой фазе и экспонируют для лигандов в подвижной фазе, протекающей через проточную ячейку. Если происходит связывание лиганда с иммобилизованной мишенью, локальный показатель преломления изменяется, приводя к изменению угла ППР, которое можно контролировать в режиме реального времени путем детекции изменений интенсивности отраженного света. Скорости изменения сигнала ППР можно анализировать для определения кажущихся констант скорости фаз ассоциации и диссоциации реакции связывания. Отношение этих величин дает кажущуюся константу равновесия (аффинность) (смотри, например, Wolff et al., Cancer Res. 53:2560-65 (1993)).
Следует понимать, что аффинность антител, предложенных по настоящему изобретению, можно изменять любым подходящим способом, известным в данной области. Таким образом, настоящее изобретение также относится к вариантам молекул антител по настоящему изобретению, которые имеют усовершенствованную аффинность для IgE. Такие варианты могут быть получены с использованием целого ряда методов созревания аффинности, включая мутации CDR (Yang et al., J. Mol. Biol., 254, 392-403, 1995), перетасовку цепей (Marks et al., Bio/Technology, 10, 779-783, 1992), использование штаммов-мутаторов E. coli (Low et al., J. Mol. Biol., 250, 359-368, 1996), перетасовку ДНК (Patten et al., Curr. Opin. Biotechnol., 8, 724-733, 1997), фаговый дисплей (Thompson et al., J. Mol. Biol., 256, 77-88, 1996) и ПЦР с имитацией полового размножения (Crameri et al., Nature, 391, 288-291, 1998). В статье Vaughan et al. (выше) описаны эти методы созревания аффинности.
Гуманизированные антитела и антигенсвязывающие фрагменты
В одном аспекте настоящего изобретения предложены гуманизированные анти-IgE моноклональные антитела и антигенсвязывающие фрагменты. Гуманизированные антитела представляют собой антитела, в которых тяжелая и/или легкая цепь содержит одну или более CDR (включая, при необходимости, одну или более модифицированных CDR) из донорского антитела (например, антитела, не принадлежащего человеку, такого как мышиное или кроличье моноклональное антитело), привитые на каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, человеческого антитела). Для обзора смотри Vaughan et al, Nature Biotechnology, 16, 535-539, 1998.
В одном из вариантов осуществления вместо перенесения целой области CDR на каркас человеческого антитела переносят только один или более определяющих специфичность остатков из любой из CDR, описанных выше в настоящем документе (смотри, например, Kashmiri et al., 2005, Methods, 36, 25-34). В одном из вариантов осуществления на каркас человеческого антитела переносят только определяющие специфичность остатки из одной или более из CDR, описанных в настоящем документе. В другом варианте осуществления на каркас человеческого антитела переносят только определяющие специфичность остатки из каждой из CDR, описанных в настоящем документе.
Когда прививают CDR или определяющие специфичность остатки, можно использовать любую подходящую акцепторную каркасную последовательность вариабельной области, с учетом класса/типа донорского антитела, из которого получены CDR, включая каркасные области антител мыши, кролика, приматов и человека.
Предпочтительно, гуманизированное антитело по настоящему изобретению имеет вариабельный домен, содержащий человеческие акцепторные каркасные области, а также одну или более CDR, конкретно описанных в настоящем документе. Таким образом, в одном из вариантов осуществления предложено гуманизированное моноклональное антитело, которое связывает человеческий IgE, при этом вариабельный домен содержит человеческие акцепторные каркасные области (с необязательными мутациями, описанными в настоящем документе) и донорские CDR, не принадлежащие человеку.
Конструирование CDR-привитых антител, в целом, описано в Европейской патентной заявке EP-A-0239400, в которой раскрыт способ прививания CDR мышиного моноклонального антитела на каркасные области вариабельных доменов человеческого иммуноглобулина путем сайт-направленного мутагенеза с использованием длинных олигонуклеотидов, содержание заявки включено в настоящий документ. Области CDR определяют антигенсвязывающую специфичность антител и представляют собой относительно короткие пептидные последовательности, находящиеся на каркасных областях вариабельных доменов.
Самые ранние работы по гуманизации моноклональных антител путем прививания CDR были проведены на моноклональных антителах, узнающих синтетические антигены, например, NP. Однако примеры, в которых мышиное моноклональное антитело, узнающее лизоцим, и крысиное моноклональное антитело, узнающее антиген на человеческих T-клетках, были гуманизированы путем прививания CDR, были описаны Verhoeyen et al. (Science, 239, 1534-1536, 1988) и Riechmann et al. (Nature, 332, 323-324, 1988), соответственно. Гуманизацию антител осуществляют путем прививания CDR не принадлежащего человеку антитела, например, антитела мыши, крысы, козы или кролика, на «аналогичный» каркас (акцептор) человеческого антитела и выбора минимального числа ключевых каркасных остатков (обратные мутации), которые вручную отбирают из донорского моноклонального антитела и встраивают в акцепторный каркас человеческого антитела с целью сохранения оригинальной конформации CDR. Такие методы известны в данной области и включают те, которые описаны в публикациях Jones et al., Nature 321:522 (1986); Verhoeyen et al., Science 239:1534 (1988)), Sims et al., J. Immunol. 151: 2296 (1993); Chothia and Lesk, J. Mol. Biol. 196:901 (1987), Carter et al., Proc. Natl. Acad. Sci. U.S.A. 89:4285 (1992); Presta et al., J. Immunol. 151:2623 (1993), Padlan, Molecular Immunology 28(4/5):489-498 (1991); Studnicka et al., Protein Engineering 7(6):805-814 (1994); Roguska. et al., PNAS 91:969-973 (1994); PCT публикации WO 91/09967, PCT/: US98/16280, US96/18978, US91/09630, US91/05939, US94/01234, GB89/01334, GB91/01134, GB92/01755; WO90/14443, WO90/14424, WO90/14430, EP 229246, EP 592,106; EP 519,596, EP 239,400, патентах США №№ 5565332, 5723323, 5976862, 5824514, 5817483, 5814476, 5763192, 5723323, 5766886, 5714352, 6204023, 6180370, 5693762, 5530101, 5585089, 5225539; 4816567, содержание которых включено в настоящий документ.
Классификация подсемейств вариабельных последовательностей тяжелой и легкой цепей зародышевой линии человека может быть производной от обозначений подгрупп зародышевой линии в соответствии с системой Kabat: VH1, VH2, VH3, VH4, VH5, VH6 или VH7 для конкретной последовательности VH и JH1, JH2, JH3, JH4, JH5 и JH6 для конкретной соединительной группы вариабельной области тяжелой цепи для каркаса 4; VK1, VK2, VK3, VK4, VK5 или VK6 для конкретной последовательности VL каппа для каркаса 1, 2 и 3, и JK1, JK2, JK3, JK4 или JK5 для конкретной соединительной группы каппа для каркаса 4; или VL1, VL2, VL3, VL4, VL5, VL6, VL7, VL8, VL9 или VL10 для конкретной последовательности VL лямбда для каркаса 1, 2 и 3, и JL1, JL2, JL3 или JL7 для конкретной соединительной группы лямбда для каркаса 4.
Домены константной области молекулы антитела по настоящему изобретению, при наличии, могут быть выбраны с учетом предполагаемой функции молекулы антитела и, в частности, эффекторных функций, которые могут потребоваться. Например, домены константной области могут представлять собой домены IgA, IgD, IgE, IgG или IgM человека. В конкретных вариантах осуществления можно использовать домены константной области IgG человека, в частности, изотипов IgG1 и IgG3, если молекула антитела предназначена для терапевтического применения и необходимы эффекторные функции антитела. Альтернативно, можно использовать изотипы IgG2 и IgG4, если молекула антитела предназначена для терапевтических целей и эффекторные функции антитела не требуются. Следует понимать, что также можно использовать варианты последовательностей этих доменов константной области. Например, можно использовать молекулы IgG4, в которых серин в положении 241 был заменен на пролин, как описано в публикации Angal et al., Molecular Immunology, 1993, 30 (1), 105-108. Специалист в данной области также понимает, что антитела могут подвергаться различным посттрансляционным модификациям. Вид и степень таких модификаций часто зависят от линии клетки-хозяина, используемой для экспрессии антитела, а также от условий культивирования. Такие модификации могут включать вариации гликозилирования, окисления метионина, образования дикетопиперазина, изомеризации аспартата и дезамидирования аспарагина. Частой модификацией является утрата карбокси-концевого основного остатка (такого как лизин или аргинин) в результате действия карбоксипептидаз (как описано в Harris, R.J. Journal of Chromatography 705:129-134, 1995). Соответственно, C-концевой остаток лизина тяжелой цепи антитела может отсутствовать.
CDR и модификации человеческой каркасной области иммуноглобулина
Riechmann с соавторами установили, что перенос только CDR (как описано Kabat (Kabat et al. (выше) и Wu et al., J. Exp. Med., 132, 211-250, 1970)) не был достаточным для обеспечения удовлетворительной способности к связыванию антигена у CDR-привитого продукта. Было обнаружено, что необходимо изменение ряда каркасных остатков для того, чтобы они соответствовали остаткам донорской каркасной области. Предлагаемые критерии для выбора каркасных остатков, которые необходимо изменять, приведены в международной патентной заявке WO90/07861, содержание которой включено в настоящий документ.
Прививание не принадлежащих человеку CDR на каркас человеческого вариабельного домена с наибольшей вероятностью приведет к сохранению правильной пространственной ориентации CDR, если каркас человеческого вариабельного домена примет такую же или аналогичную конформацию, что и каркас не принадлежащего человеку вариабельного домена, из которого были взяты CDR. Это достигается путем получения человеческих вариабельных доменов из человеческих антител, каркасные последовательности которых имеют высокую степень идентичности последовательностей с не принадлежащими человеку каркасными последовательностями вариабельных доменов, из которых были взяты CDR. Как описано выше, каркасные области вариабельной области тяжелой и легкой цепей могут быть взяты из тех же самых или других последовательностей человеческого антитела. Последовательности человеческого антитела могут представлять собой последовательности природных человеческих антител или могут представлять собой консенсусные последовательности нескольких человеческих антител. Смотри Kettleborough et al, Protein Engineering 4:773 (1991); Kolbinger et al., Protein Engineering 6:971 (1993) и Carter et al., WO92/22653.
После идентификации определяющих комплементарность областей не принадлежащего человеку донорского иммуноглобулина и соответствующих акцепторных человеческих иммуноглобулинов, следующим этапом является определение того, какие, в случае необходимости в этом, остатки из этих компонентов должны быть заменены для оптимизации свойств полученного гуманизированного антитела. Как правило, замена характерных для человека аминокислотных остатков на аминокислотные остатки, не характерные для человека, должна быть сведена к минимуму, поскольку введение нехарактерных для человека остатков увеличивает риск того, что антитело вызовет у человека иммунный ответ против донорского антитела (HADA). Признанные в данной области методы определения иммунного ответа можно использовать для контролирования HADA-ответа у конкретного хозяина или при проведении клинических испытаний. Можно проводить оценку на иммуногенность у хозяев, которым вводят гуманизированные антитела, в начале исследования и в процессе введения указанного терапевтического средства. HADA-ответ определяют, например, путем обнаружения антител к гуманизированному терапевтическому средству в образцах сыворотки хозяина методом, известным в данной области, включая технологию поверхностного плазмонного резонанса (BIACORE) и/или твердофазный анализ ELISA.
Выбор аминокислотных остатков для замены (в настоящем документе также используют термин «мутации») частично определяется результатами компьютерного моделирования. В настоящем документе приведено описание компьютерных устройств и программного обеспечения для создания трехмерных изображений молекул иммуноглобулинов. Как правило, молекулярные модели создают, начиная с известных структур цепей иммуноглобулинов или их доменов. Моделируемые цепи сравнивают на предмет сходства аминокислотных последовательностей с цепями или доменами известных трехмерных структур, и цепи или домены, демонстрирующие наибольшее сходство последовательностей, выбирают в качестве отправных точек для построения молекулярной модели. Цепи или домены, имеющие по меньшей мере 50% идентичности последовательностей, выбирают для моделирования, и предпочтительно для моделирования выбирают те, которые имеют по меньшей мере 60%, 70%, 80%, 90% или более идентичности последовательностей. Известные исходные структуры модифицируют для учета различий между фактическими аминокислотами в цепях или доменах иммуноглобулинов, которые моделируют, и теми, которые присутствуют в исходной структуре. Затем модифицированные структуры собирают в комбинированный иммуноглобулин. И наконец, модель уточняют путем минимизации энергии и проверки того, что все атомы находятся на соответствующих расстояниях друг от друга, и что длины и углы связей находятся в приемлемых с точки зрения химии границах.
Выбор аминокислотных остатков для замены также может частично определяться результатами изучения характеристик аминокислот в конкретных местоположениях, либо эмпирического наблюдения эффектов замены или мутагенеза конкретных аминокислот. Например, если есть отличия между донорским остатком каркаса вариабельной области и выбранным остатком каркаса человеческой вариабельной области, аминокислоту человеческой каркасной области, как правило, следует заменять эквивалентной аминокислотой каркасной области из донорского антитела, если есть основания считать, что аминокислота:
(1) непосредственно нековалентно связывает антиген,
(2) находится рядом с областью CDR,
(3) иным образом взаимодействует с областью CDR (например, находится в пределах 3-6 ангстрем от области CDR, что определяют с помощью компьютерного моделирования), или
(4) находится в области контакта VL-VH.
Остатки, которые «непосредственно нековалентно связывают антиген» включают аминокислоты в положениях каркасных областей, которые с большой вероятностью непосредственно взаимодействуют с аминокислотами антигена в соответствии с известными химическими силами, такими как образование водородных связей, ван-дер-ваальсовы взаимодействия, гидрофобные взаимодействия и тому подобное. CDR и каркасные области соответствуют системе Kabat et al. или Chothia et al., выше. Если каркасные остатки, определенные по системе Kabat et al., выше, представляют собой остатки структурной петли в соответствии с системой Chothia et al., выше, то аминокислоты, присутствующие в донорском антителе, могут быть выбраны для заместительного введения в гуманизированное антитело. Остатки, которые «находится рядом с областью CDR» включают аминокислотные остатки в положениях, находящихся в непосредственной близости от одной или более CDR в основной последовательности цепи гуманизированного иммуноглобулина, например, в положениях, находящихся в непосредственной близости от CDR, определенной по системе Kabat, или CDR, определенной по системе Chothia (смотри, например, Chothia and Lesk 1MB 196: 901 (1987)). Существует большая вероятность того, что эти аминокислоты взаимодействуют с аминокислотами в CDR и, если будут выбраны из акцептора, будут вызывать искажение донорских CDR и приводить к уменьшению аффинности. Кроме того, расположенные в непосредственной близости аминокислоты могут напрямую взаимодействовать с антигеном (Amit et al., Science, 233:747 (1986), содержание публикации включено в настоящий документ посредством ссылки) и выбор таких аминокислот из донора может быть желательным для сохранения всех контактов с антигеном, которые обеспечивают аффинность исходного антитела. Как описано в настоящем документе, в последовательностях FR также можно производить замены/мутации для повышения аффинности анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению для IgE (и/или для расширения его области взаимодействия или эпитопа на Cε2 в IgE).
Остатки, которые «иным образом взаимодействуют с областью CDR», включают остатки, которые по результатам анализа вторичной структуры находятся в такой пространственной ориентации, что могут оказывать влияние на область CDR. В одном из вариантов осуществления остатки, которые «иным образом взаимодействуют с областью CDR», выявляют путем анализа трехмерной модели донорского иммуноглобулина (например, компьютерной модели). Трехмерная модель исходного донорского антитела, как правило, показывает, что определенные аминокислоты за пределами CDR расположены близко к CDR, и существует большая вероятность их взаимодействия с аминокислотами в CDR за счет образования водородных связей, ван-дер-ваальсовых взаимодействий, гидрофобных взаимодействий и так далее. Для таких аминокислотных положений предпочтительно выбирать аминокислоту донорского иммуноглобулина, а не аминокислоту акцепторного иммуноглобулина. Аминокислоты, соответствующие этому критерию, как правило, будут иметь атом боковой цепи в пределах примерно 3 ангстрем (A) от некоего атома в областях CDR и должны содержать атом, который мог бы взаимодействовать с атомами CDR в соответствии с известными химическими силами, такими как те, которые перечислены выше. В случае атомов, которые могут образовывать водородную связь, измеренное расстояние составляет 3 Å между их ядрами, но для атомов, которые не образуют связь, измеренное расстояние составляет 3 Å между их ван-дер-ваальсовыми поверхностями. Таким образом, в последнем случае ядра должны находиться на расстоянии в пределах 6 Å (3 Å плюс сумма ван-дер-ваальсовых радиусов), чтобы считать атомы способными на взаимодействие. Во многих случаях ядра будут находиться на расстоянии от 4 или 5 до 6 Å. При определении того, может ли аминокислота взаимодействовать с областями CDR, предпочтительно не рассматривать последние 8 аминокислот CDR2 тяжелой цепи как часть CDR, поскольку с точки зрения структуры эти 8 аминокислот являются скорее частью каркаса.
Аминокислоты, которые способны взаимодействовать с аминокислотами в областях CDR (или областях FR), можно идентифицировать и другим путем. Доступную для растворителя площадь поверхности каждой каркасной аминокислоты рассчитывают двумя способами: (1) в интактном антителе и (2) в гипотетической молекуле, представляющей собой антитело с удаленными областями CDR. Значительная разница между этими величинами, составляющая примерно 10 квадратных ангстрем или более, указывает на то, что доступ растворителя к этой каркасной аминокислоте, по меньшей мере частично, блокируется областями CDR и, следовательно, что эта аминокислота имеет контакт с областями CDR. Доступную для растворителя площадь поверхности аминокислоты можно рассчитывать на основании трехмерной модели антитела с использованием алгоритмов, известных в данной области (например, Connolly, J. Appl. Cryst. 16:548 (1983) и Lee and Richards, J. Mol. Biol. 55:379 (1971), содержание обеих публикаций включено в настоящий документ посредством ссылки). Каркасные аминокислоты также иногда могут взаимодействовать с областями CDR опосредованно, влияя на конформацию другой каркасной аминокислоты, которая, в свою очередь, контактирует с областями CDR.
Известно, что во многих антителах конкретные аминокислоты в некоторых положениях каркасной области способны взаимодействовать с областями CDR (Chothia and Lesk, выше, Chothia et al., выше и Tramontano et al., J. Mol. Biol. 215:175 (1990), содержание всех публикаций включено в настоящий документ посредством ссылки). В частности, известно, что во многих антителах аминокислоты в положениях 2, 48, 64 и 71 в легкой цепи, а также 71 и 94 в тяжелой цепи (нумерация по системе Kabat) способны взаимодействовать с областями CDR. Также возможно, что аминокислоты в положениях 35 в легкой цепи, а также 93 и 103 в тяжелой цепи взаимодействуют с областями CDR. Во всех этих перечисленных положениях для гуманизированного иммуноглобулина предпочтительно выбирать аминокислоту донора, а не аминокислоту акцептора (если они отличаются). С другой стороны, некоторые остатки, способные взаимодействовать с областью CDR, например, первые 5 аминокислот легкой цепи, иногда можно выбирать из акцепторного иммуноглобулина без потери аффинности гуманизированного иммуноглобулина.
Остатки, которые «находятся в области контакта VL-VH», или «остатки прокладки» включают остатки на границе контакта между VL и VH в соответствии с определением, например, Novotny and Haber, Proc. Natl. Acad. Sci. USA, 82:4592-66 (1985) или Chothia et al., выше. Как правило, необычные остатки прокладки следует сохранять в гуманизированном антителе, если они отличаются от остатков в человеческих каркасных областях.
Как правило, одну или более из аминокислот, удовлетворяющих вышеприведенным критериям, заменяют. В некоторых вариантах осуществления все или большинство из аминокислот, удовлетворяющих вышеприведенным критериям, заменяют. Иногда возникает некоторая неопределенность относительно того, удовлетворяет ли конкретная аминокислота вышеуказанным критериям, тогда получают альтернативные варианты иммуноглобулинов, один из которых имеет эту конкретную замену, а другой не имеет. Полученные таким образом альтернативные варианты иммуноглобулинов можно тестировать в любом из анализов, описанных в настоящем документе, на наличие желательной активности и выбирать предпочтительный иммуноглобулин.
Как правило, области CDR в гуманизированных антителах являются по существу идентичными и, чаще всего, идентичными соответствующим областям CDR донорского антитела. Хотя это обычно не желательно, иногда можно осуществлять одну или более консервативных аминокислотных замен остатков CDR без существенного влияния на аффинность связывания полученного гуманизированного иммуноглобулина. Консервативными или аналогичными заменами являются такие замены, как, например, замена лейцина на изолейцин или валин. Другие аминокислоты, которые часто могут заменять одна другую, включают, но не ограничиваются ими:
фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи);
лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи);
аспартат и глутамат (аминокислоты, имеющие кислые боковые цепи);
аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и,
цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи).
Дополнительными кандидатами на замену являются аминокислоты акцепторной человеческой каркасной области, которые являются необычными или «редкими» для человеческого иммуноглобулина в этом положении. Такие аминокислоты могут быть заменены аминокислотами из эквивалентного положения донорского антитела или из эквивалентных положений более типичных человеческих иммуноглобулинов. Например, замена может быть желательной, если аминокислота в человеческой каркасной области акцепторного иммуноглобулина редко встречается в этом положении, а соответствующая аминокислота в донорском иммуноглобулине является обычной для этого положения в последовательностях человеческих иммуноглобулинов; или если аминокислота в акцепторном иммуноглобулине редко встречается в этом положении и соответствующая аминокислота в донорском иммуноглобулине также встречается редко в сравнении с другими последовательностями человеческих иммуноглобулинов. Эти критерии помогают гарантировать то, что нетипичная аминокислота в человеческой каркасной области не нарушит структуру антитела. Более того, путем замены необычной аминокислоты человеческой акцепторной последовательности аминокислотой из донорского антитела, которая является типичной для человеческих антител, гуманизированное антитело можно сделать менее иммуногенным.
Используемый в настоящем документе термин «редкая» означает аминокислоту, которая занимает данное положение в менее чем примерно 20%, как правило, менее чем примерно 10% последовательностей в репрезентативном образце последовательностей, и используемый в настоящем документе термин «обычная» означает аминокислоту, которая занимает данное положение в более чем примерно 25%, как правило, более чем примерно 50% последовательностей в репрезентативном образце. Например, все человеческие последовательности вариабельных областей легкой и тяжелой цепей соответственно сгруппированы в «подгруппы» последовательностей, которые особенно гомологичны друг другу и имеют одни и те же аминокислоты в определенных важных положениях (Kabat et al., выше). При принятии решения о том, является ли аминокислота в человеческой акцепторной последовательности «редкой» или «обычной» для человеческих последовательностей, часто бывает предпочтительно рассматривать только человеческие последовательности, находящиеся в той же подгруппе, что и акцепторная последовательность.
Дополнительными кандидатами на замену являются остатки акцепторного каркаса, которые соответствуют редким или необычным остаткам донорского каркаса. Редкими или необычными остатками донорского каркаса являются остатки, являющиеся редкими или необычными (как определено в настоящем документе) для донорских антител в данном положении. Для донорских антител можно определять подгруппу в соответствии с системой Kabat и идентифицировать положения остатков, которые отличаются от консенсуса. Эти специфические отличия донорской последовательности могут указывать на соматические мутации в донорской последовательности, которые приводят к усилению активности. Необычные остатки, которые, согласно предсказанию, влияют на связывание, оставляют, в то время как остатки, которые, согласно предсказанию, не важны для связывания, могут быть заменены.
Дополнительными кандидатами на замену являются остатки не зародышевой линии, присутствующие в акцепторной каркасной области. Например, при выравнивании акцепторной цепи антитела (то есть, цепи человеческого антитела, имеющей значительную идентичность последовательности с донорской цепью антитела) с цепью антитела зародышевой линии (аналогичным образом имеющей значительную идентичность последовательности с донорской цепью), остатки, не совпадающие у каркаса акцепторной цепи и каркаса цепи зародышевой линии, могут быть заменены соответствующими остатками из последовательности зародышевой линии.
За исключением специфических аминокислотных замен, описанных выше, каркасные области гуманизированных иммуноглобулинов, как правило, являются по существу идентичными, и еще чаще, идентичными каркасным областям человеческих антител, из которых они происходят (за исключением описанных в настоящем документе для целей настоящего изобретения). Безусловно, многие из аминокислот в каркасной области вносят небольшой, или совсем не вносят, прямой вклад в специфичность или аффинность антитела. Следовательно, множество отдельных консервативных замен каркасных остатков можно осуществлять без заметных изменений специфичности или аффинности полученного гуманизированного иммуноглобулина. Таким образом, в одном из вариантов осуществления каркас вариабельной области гуманизированного иммуноглобулина имеет по меньшей мере 65, 75 или 85% сходства, или идентичности, последовательности с человеческой последовательностью каркаса вариабельной области или консенсусом таких последовательностей. В другом варианте осуществления каркас вариабельной области гуманизированного иммуноглобулина имеет по меньшей мере 90%, предпочтительно 95%, более предпочтительно 96%, 97%, 98% или 99% сходства, или идентичности, последовательности с человеческой последовательностью каркаса вариабельной области или консенсусом таких последовательностей. Однако, как правило, такие замены нежелательны (за исключением тех, которые описаны в настоящем документе).
Упоминаемую в настоящем документе степень идентичности и сходства можно с легкостью рассчитывать, например, как описано в Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, программе BLAST™, доступной от NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215:403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3:266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266:131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25:3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7:649-656, все публикации включены в настоящий документ посредством ссылки.
Опубликован ряд обзоров, в которых обсуждаются CDR-привитые антитела, включая публикацию Vaughan et al. (Nature Biotechnology, 16, 535-539, 1998), содержание которой включено в настоящий документ посредством ссылки.
Анти-IgE антитела по настоящему изобретению также могут содержать дополнительные связывающие домены, например, как молекула DVD-Ig, которая раскрыта в WO2007/024715, или так называемое антитело (FabFv)2Fc, описанное в WO2011/030107. Таким образом, антитело, используемое по настоящему изобретению, включает би-, три- или тетравалентные полноразмерные антитела.
Антигенсвязывающие фрагменты
Антигенсвязывающие фрагменты включают одноцепочечные антитела (то есть, полноразмерную тяжелую цепь и легкую цепь); Fab, модифицированные Fab, Fab', модифицированные Fab', F(ab')2, Fv, Fab-Fv, Fab-dsFv, однодоменные антитела (например, VH или VL или VHH), например, описанные в WO2001090190, scFv, би-, три- или тетравалентные антитела, бис-scFv, диатела, триотела, триатела, тетратела и связывающие эпитоп антигена фрагменты любых из вышеперечисленных (смотри, например, Holliger and Hudson, 2005, Nature Biotech. 23(9):1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217). Методы создания и производства таких фрагментов антител хорошо известны в данной области (смотри, например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Формат Fab-Fv был впервые описан в WO2009/040562, и его стабилизированные дисульфидными связями варианты, Fab-dsFv, были впервые описаны в WO2010/035012. Другие фрагменты антитела для использования по настоящему изобретению включают фрагменты Fab и Fab', описанные в международных патентных заявках WO2005/003169, WO2005/003170 и WO2005/003171. Мультивалентные антитела могут иметь несколько специфичностей, например, быть биспецифическими, или могут быть моноспецифическими (смотри, например, WO92/22583 и WO05/113605). Одним из примеров последних являются три-Fab (или TFM), описанные в WO92/22583.
Типичная молекула Fab' содержит пару тяжелой и легкой цепей, при этом тяжелая цепь содержит вариабельную область VH, константный домен CH1 и природную или модифицированную шарнирную область, а легкая цепь содержит вариабельную область VL и константный домен CL.
В одном из вариантов осуществления предложен димер Fab' по настоящему изобретению для создания F(ab')2, например, димеризация может быть произведена через природную шарнирную последовательность, описанную в настоящем документе, или ее производное, либо синтетическую шарнирную последовательность.
Связывающий домен антитела, как правило, будет содержать 6 областей CDR, три из тяжелой цепи и три из легкой цепи. В одном из вариантов осуществления области CDR вместе с каркасом образуют вариабельную область. Таким образом, в одном из вариантов осуществления антигенсвязывающий фрагмент содержит связывающий домен, специфичный для IgE, содержащий вариабельную область легкой цепи и вариабельную область тяжелой цепи.
Следует понимать, что одна или более (например, 1, 2, 3 или 4) аминокислотных замен, добавлений и/или делеций могут быть внесены в области CDR или другие последовательности (например, вариабельные домены), предложенные по настоящему изобретению, как описано выше или ниже, без существенного изменения способности антитела к связыванию с IgE. Влияние любых из аминокислотных замен, добавлений и/или делеций может быть с легкостью протестировано специалистом в данной области, например, с использованием методов, описанных в настоящем документе, в частности в разделе «Примеры».
В одном из вариантов осуществления одна или более (например, 1, 2, 3 или 4) аминокислотных замен, добавлений и/или делеций могут быть внесены в области CDR или каркасные области, используемые в антителе или фрагменте, предложенном по настоящему изобретению, таким образом, что аффинность связывания (KD) анти-IgE антитела, или антигенсвязывающего фрагмента, по изобретению с IgE составляет менее 2, 1,9, 1,8, 1,7, 1,6, 1,5, 1,4, 1,3, 1,2, 1,1, 1,0, 0,9, 0,8, 0,7, 0,6, 0,5, 0,4 или 0,3 нМ. В одном из вариантов осуществления предложено модифицированное гуманизированное антитело, в котором модификации были выполнены либо в областях CDR, каркасных областях, либо и тех, и других, с целью уменьшения KD, например, до менее 2, 1,9, 1,8, 1,7, 1,6, 1,5, 1,4, 1,3, 1,2, 1,1, 1,0, 0,9, 0,8, 0,7, 0,6, 0,5, 0,4 или 0,3 нМ.
Фрагмент антитела по настоящему изобретению включает Fab, Fab′, F(ab′)2, scFv, диатело, scFAb, dFv, однодоменную легкую цепь антитела, dsFv, пептид, содержащий CDR, и тому подобное.
Fab представляет собой фрагмент антитела, имеющий молекулярную массу примерно 50000 и антигенсвязывающую активность, в котором примерно половина N-концевой части H-цепи и полная L-цепь, в числе фрагментов, получаемых при обработке IgG протеазой папаином (расщепляющим по аминокислотному остатку в положении 224 H-цепи), связаны вместе дисульфидной связью.
Fab по настоящему изобретению можно получать путем обработки человеческого CDR-привитого антитела по настоящему изобретению, которое специфически реагирует с IgE, протеазой папаином. Кроме того, Fab можно получать путем вставки ДНК, кодирующей Fab антитела, в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и введения вектора в прокариотические или эукариотические клетки для экспрессии Fab.
F(ab′)2 представляет собой фрагмент антитела, имеющий молекулярную массу примерно 100000 и антигенсвязывающую активность, немного превышающий по размеру фрагменты Fab, связанные дисульфидной связью в шарнирной области, он находится в числе фрагментов, получаемых при обработке IgG протеазой пепсином.
F(ab′)2 по настоящему изобретению можно получать путем обработки человеческого CDR-привитого антитела, которое специфически реагирует с IgE, протеазой пепсином. Кроме того, F(ab′)2 можно получать путем связывания Fab′, описанных ниже, тиоэфирной связью или дисульфидной связью.
Fab′ представляет собой фрагмент антитела, имеющий молекулярную массу примерно 50000 и антигенсвязывающую активность, который получают путем разрыва дисульфидной связи в шарнирной области F(ab′)2.
Fab′ по настоящему изобретению можно получать путем обработки F(ab′)2, специфически реагирующего с IgE, восстанавливающим реагентом, дитиотреитолом. Кроме того, Fab′ по настоящему изобретению можно получать путем вставки ДНК, кодирующей Fab′ человеческого CDR-привитого антитела по настоящему изобретению, специфически реагирующего с IgE, в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и введения вектора в прокариотические или эукариотические клетки для экспрессии Fab′.
scFv представляет собой полипептид VH-P-VL или VL-P-VH, в котором одна цепь VH и одна цепь VL связаны с использованием соответствующего пептидного линкера (P) из 12 или более остатков, и который имеет антигенсвязывающую активность.
scFv по настоящему изобретению можно получать путем получения кДНК, кодирующих VH и VL человеческого CDR-привитого антитела, специфически реагирующего с IgE, по настоящему изобретению, конструирования ДНК, кодирующей scFv, встраивания ДНК в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и последующего введения экспрессионного вектора в прокариотические или эукариотические клетки для экспрессии scFv.
Fab-фрагмент по настоящему изобретению может быть связан, непосредственно или через линкер, с scFv. Используемый в настоящем документе термин «одноцепочечный вариабельный фрагмент», или «scFv», означает одноцепочечный вариабельный фрагмент, который стабилизирован пептидным линкером между вариабельными доменами VH и VL, например, пептидным линкером с аминокислотной последовательностью SEQ ID NO:151. Связь с Fab-фрагментом может быть образована за счет химической конъюгации, но наиболее предпочтительным является трансляционное слияние, то есть генетический слитый продукт с последовательностью каждого из компонентов, закодированной в экспрессионном векторе. Таким образом, линкер, как правило, представляет собой аминокислотный линкер, описанный в настоящем документе. scFv по настоящему изобретению, связанный с Fab-фрагментом, может связываться с сывороточным белком-носителем с целью увеличения времени полужизни слитого белка антитела in vivo. В этом случае увеличение времени полужизни не зависит от связывания IgE и может быть полезным.
Используемый в настоящем документе термин «сывороточный белок-носитель» означает любой подходящий белок-носитель в плазме, с которым может связываться scFv, в одном примере сывороточный белок-носитель выбирают из тироксинсвязывающего белка, транстиретина, α1-кислого гликопротеина, трансферрина, фибриногена и альбумина, либо фрагмента любого из них. Как правило, scFv связывается с альбумином, предпочтительно, сывороточным альбумином человека.
Любой подходящий альбумин-связывающий scFv можно включать в слитые белки антитела по изобретению. Подходящие альбумин-связывающие домены известны в данной области.
Диатело представляет собой фрагмент антитела, в котором фрагменты scFv, имеющие одну и ту же или разные специфичности связывания антигена, образуют димер, и который имеет двухвалентную антигенсвязывающую активность для одного и того же антигена, или две специфические антигенсвязывающие активности для разных антигенов.
Диатело по настоящему изобретению, например, двухвалентное диатело, специфически реагирующее с IgE, можно получать путем получения кДНК, кодирующих VH и VL антитела, специфически реагирующего с IgE, конструирования ДНК, кодирующей фрагмент scFv, имеющий полипептидный линкер из 3-10 остатков, встраивания ДНК в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и последующего введения вектора в прокариотические или эукариотические клетки для экспрессии диатела.
dsFv получают путем связывания полипептидов, в которых один аминокислотный остаток каждой из VH и VL заменен остатком цистеина, дисульфидной связью между остатками цистеина. Аминокислотный остаток, который заменяют остатком цистеина, может быть выбран на основании оценки трехмерной структуры антитела методом, описанным Reiter et al. (Protein Engineering, 7, 697 (1994)).
dsFv по настоящему изобретению можно получать путем получения кДНК, кодирующих VH и VL человеческого CDR-привитого антитела, специфически реагирующего с IgE, по настоящему изобретению, конструирования ДНК, кодирующей dsFv, встраивания ДНК в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и последующего введения вектора в прокариотические или эукариотические клетки для экспрессии dsFv.
Пептид, содержащий CDR, составляют путем включения по меньшей мере одной области из областей CDR H-цепи и L-цепи. Несколько областей CDR могут быть связаны напрямую или через соответствующий пептидный линкер.
Пептид, содержащий CDR, по настоящему изобретению можно получать путем получения кДНК, кодирующей CDR VH и VL человеческого CDR-привитого антитела, специфически реагирующего с IgE, конструирования ДНК, кодирующей CDR, встраивания ДНК в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и последующего введения вектора в прокариотические или эукариотические клетки для экспрессии пептида. Кроме того, пептид, содержащий CDR, также можно получать методом химического синтеза, таким как метод с использованием Fmoc (метод с использованием флуоренилметоксикарбонила), метод с использованием tBoc (метод с использованием t-бутилоксикарбонила) или тому подобное.
Антитело по настоящему изобретению включает производные антитела, в которых радиоизотоп, белок, средство или тому подобное, химически или генетически конъюгированы с антителом по настоящему изобретению.
Производные антитела по настоящему изобретению можно получать путем химической конъюгации радиоизотопа, белка или средства с N-концевой частью или C-концевой частью H-цепи или L-цепи антитела, или фрагмента антитела, специфически реагирующего с IgE, с соответствующей замещающей группой или боковой цепью антитела, или фрагмента антитела, либо с сахарной цепью в антителе, или фрагменте антитела (Antibody Engineering Handbook, под редакцией Osamu Kanemitsu, опубликовано Chijin Shokan (1994)).
Кроме того, их можно получать генно-инженерными методами путем связывания ДНК, кодирующей антитело, или фрагмент антитела, по настоящему изобретению, специфически реагирующее с IgE, с другой ДНК, кодирующей белок, который должен быть связан, встраивания ДНК в экспрессионный вектор и введения экспрессионного вектора в клетку-хозяина.
Радиоизотоп включает 131I, 125I и тому подобное, и он может быть конъюгирован с антителом, например, методом с использованием хлорамина T.
Средство предпочтительно представляет собой низкомолекулярное соединение. Примеры включают противораковые средства, такие как алкилирующие средства (например, азотистый иприт, циклофосфамид), метаболические антагонисты (например, 5-фторурацил, метотрексат), антибиотики (например, дауномицин, блеомицин, митомицин C, даунорубицин, доксорубицин), растительные алкалоиды (например, винкристин, винбластин, виндезин), гормональные лекарственные средства (например, тамоксифен, дексаметазон) и тому подобное (Clinical Oncology, под редакцией Japanese Society of Clinical Oncology, опубликовано Cancer and Chemotherapy (1996)); противовоспалительные средства, такие как стероидные средства (например, гидрокортизон, преднизон), нестероидные лекарственные средства (например, аспирин, индометацин), иммуномодуляторы (например, ауротиомалат, пеницилламин), иммунодепрессанты (например, циклофосфамид, азатиоприн) и антигистаминные средства (например, хлорфенирамин малеат, клемастин) (Inflammation and Anti-inflammatory Therapy, Ishiyaku Shuppan (1982)); и тому подобное. Методы конъюгации дауномицина с антителом включают метод, в котором дауномицин и аминогруппу антитела конъюгируют с помощью глутаральдегида, метод, в котором аминогруппу дауномицина и карбоксильную группу антитела конъюгируют с помощью водорастворимого карбодиимида, и тому подобные.
Кроме того, для непосредственного ингибирования раковых клеток можно использовать токсин, такой как рицин, дифтерийный токсин и тому подобное. Например, можно получать слитый продукт антитела с белком путем связывания кДНК, кодирующей антитело, или фрагмент антитела, с другой кДНК, кодирующей белок, конструирования ДНК, кодирующей слитое антитело, встраивания ДНК в экспрессионный вектор для прокариотических клеток или экспрессионный вектор для эукариотических клеток и последующего введения вектора в прокариотические или эукариотические клетки для экспрессии слитого антитела.
По настоящему изобретению также предусмотрены фрагменты антитела или антигенсвязывающие фрагменты, включая слитые продукты связывающих фрагментов, например, иммуноглобулин-подобные фрагменты и соединения, такие как диатела, scAb, биспецифические фрагменты, триатела, Fab-Fv-Fv, Fab-Fv, триотела, (Fab-Fv)2-Fc, а также фрагменты или части антител, такие как области CDR или петли антитела, содержащие области CDR, которые привиты на не-Ig каркасы, например, фибронектин или лейциновые «застежки», как описано в Binz et al., (2005) Nat. Biotech. 23:1257-1268, содержание публикации включено в настоящий документ в полном объеме.
Конъюгированные анти-IgE моноклональные антитела и антигенсвязывающие фрагменты
При необходимости, антитело, или антигенсвязывающий фрагмент, для использования по настоящему изобретению можно конъюгировать с одной или более эффекторными молекулами. Следует понимать, что эффекторная молекула может представлять собой одну эффекторную молекулу, либо две или более таких молекул, связанные в один фрагмент, который может быть присоединен к антителам по настоящему изобретению. Если нужно получить фрагмент антитела, связанный с эффекторной молекулой, его можно получать стандартными химическими методами или методами рекомбинантных ДНК, в которых фрагмент антитела связывают либо напрямую, либо через сшивающий агент, с эффекторной молекулой. Методы конъюгации таких эффекторных молекул с антителами хорошо известны в данной области (смотри, Hellstrom et al., Controlled Drug Delivery, 2-е издание, Robinson et al., eds., 1987, pp. 623-53; Thorpe et al., 1982, Immunol. Rev., 62:119-58 и Dubowchik et al., 1999, Pharmacology and Therapeutics, 83, 67-123). Конкретные химические методы включают, например, те, которые описаны в WO93/06231, WO92/22583, WO89/00195, WO89/01476 и WO03/031581. Альтернативно, если эффекторная молекула представляет собой белок или полипептид, связывание можно осуществлять методами рекомбинантных ДНК, например, как описано в WO86/01533 и EP0392745.
Используемый в настоящем документе термин «эффекторная молекула» включает, например, антинеопластические средства, лекарственные средства, токсины, биологически активные белки, например, ферменты, другие антитела или фрагменты антител, антигенсвязывающие фрагменты, синтетические (включая ПЭГ) или природные полимеры, нуклеиновые кислоты и их фрагменты, например, ДНК, РНК и их фрагменты, радионуклиды, в частности, радиоактивный йод, радиоактивные изотопы, хелатно связанные металлы, наночастицы и репортерные группы, такие как флуоресцентные соединения или соединения, которые могут быть обнаружены методами ЯМР или ЭСР-спектроскопии.
Примеры эффекторных молекул могут включать цитотоксины или цитотоксические средства, в том числе любое средство, которое наносит вред клеткам (например, убивает). Примеры включают комбрестатины, доластатины, эпотилоны, стауроспорин, майтанзиноиды, спонгистатины, ризоксин, халихондрины, роридины, гемиастерлины, таксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, а также их аналоги или гомологи.
Эффекторные молекулы также включают, но не ограничиваются ими, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), алкилирующие средства (например, мехлорэтамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиамин платину (II) (DDP, цисплатин), антрациклины (например, даунорубицин (прежнее название дауномицин) и доксорубицин), антибиотики (например, дактиномицин (прежнее название актиномицин), блеомицин, митрамицин, антрамицин (AMC), калихеамицины или дуокармицины), а также антимитотические средства (например, винкристин и винбластин).
Другие эффекторные молекулы могут включать хелатно связанные радионуклиды, такие как 111In и 90Y, Lu177, висмут213, калифорний252, иридий192 и вольфрам188/рений188; или лекарственные средства, такие как, но без ограничения, алкилфосфохолины, ингибиторы топоизомеразы I, таксоиды и сурамин.
Другие эффекторные молекулы включают белки, пептиды и ферменты. Представляющие интерес ферменты включают, но не ограничиваются ими, протеолитические ферменты, гидролазы, лиазы, изомеразы, трансферазы. Представляющие интерес белки, полипептиды и пептиды включают, но не ограничиваются ими, иммуноглобулины, токсины, такие как абрин, рицин A, экзотоксин псевдомонад или дифтерийный токсин, белки, такие как инсулин, фактор некроза опухолей, α-интерферон, β-интерферон, фактор роста нервов, тромбоцитарный фактор роста или тканевой активатор плазминогена, противотромбозные средства или антиангиогенные средства, например, ангиостатин или эндостатин, или модификаторы биологического ответа, такие как лимфокин, интерлейкин-1 (IL-1), интерлейкин-2 (IL-2), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), гранулоцитарный колониестимулирующий фактор (G-CSF), фактор роста нервов (NGF) или другие факторы роста и иммуноглобулины.
Другие эффекторные молекулы могут включать детектируемые вещества, полезные, например, в диагностике. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радионуклиды, позитронно-активные металлы (для использования в позитронно-эмиссионной томографии) и нерадиоактивные парамагнитные ионы металлов. Смотри, в основном, патент США № 4741900 для информации об ионах металлов, которые можно конъюгировать с антителами для использования в диагностике. Подходящие ферменты включают пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; подходящие простетические группы включают стрептавидин, авидин и биотин; подходящие флуоресцентные материалы включают умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансил хлорид и фикоэритрин; подходящие люминесцентные материалы включают люминол; подходящие биолюминесцентные материалы включают люциферазу, люциферин и экворин; и подходящие радионуклиды включают 125I, 131I, 111In и 99Tc.
В другом примере эффекторная молекула может увеличивать время полужизни антитела in vivo и/или уменьшать иммуногенность антитела, и/или облегчать доставку антитела через эпителиальный барьер к иммунной системе. Примеры подходящих эффекторных молекул данного типа включают полимеры, альбумин, альбумин-связывающие белки или альбумин-связывающие соединения, такие как те, которые описаны в WO05/117984.
В одном из вариантов осуществления время полужизни, которое обеспечивает эффекторная молекула, независимая от IgE или анти-IgE человеческого антитела, является преимуществом.
Если эффекторная молекула представляет собой полимер, она, как правило, может представлять собой синтетический или природный полимер, например, необязательно замещенный полиалкилен с прямой или разветвленной цепью, полиалкениленовый или полиоксиалкиленовый полимер, либо разветвленный или неразветвленный полисахарид, например, гомо- или гетерополисахарид.
Конкретные необязательные заместители, которые могут присутствовать на вышеуказанных синтетических полимерах, включают одну или более гидрокси, метильных или метокси групп.
Конкретные примеры синтетических полимеров включают необязательно замещенный поли(этиленгликоль), поли(пропиленгликоль), поли(виниловый спирт) с прямой или разветвленной цепью, или их производные, в частности, необязательно замещенный поли(этиленгликоль), такой как метоксиполи(этиленгликоль) или его производные.
Конкретные природные полимеры включают лактозу, амилозу, декстран, гликоген или их производные.
В одном из вариантов осуществления полимер представляет собой альбумин или его фрагмент, такой как человеческий сывороточный альбумин или его фрагмент. В одном из вариантов осуществления полимер представляет собой молекулу ПЭГ.
В настоящем документе термин «производные», используемый со ссылкой на конъюгатам, охватывает реакционноспособные производные, например, тиол-селективные реакционноспособные группы, такие как малеинимиды, и тому подобное. Реакционноспособная группа может быть связана с полимером напрямую или через сегмент линкера. Следует понимать, что остаток такой группы в некоторых случаях будет образовывать часть продукта в виде связывающей группы между фрагментом антитела и полимером.
Размер природного или синтетического полимера может варьироваться по мере необходимости, однако, как правило, будет находиться в диапазоне средних молекулярных масс от 500 Да до 50000 Да, например, 5000-40000 Да, например, 20000-40000 Да. Конкретный размер полимера можно выбирать, исходя из предполагаемого применения продукта, например, на основании способности к локализации в определенных тканях, таких как опухоли, или продолжительного периода полужизни в системе циркуляции (для обзора смотри Chapman, 2002, Advanced Drug Delivery Reviews, 54, 531-545). Таким образом, например, если продукт должен покидать систему циркуляции и проникать в ткань, например, для использования в лечении опухоли, может быть предпочтительным использование низкомолекулярного полимера, например, с молекулярной массой примерно 5000 Да. Для вариантов применения, в которых продукт остается в системе циркуляции, может быть предпочтительным использование более высокомолекулярного полимера, например, имеющего молекулярную массу в диапазоне от 20000 Да до 40000 Да.
Подходящие полимеры включают полиалкиленовый полимер, такой как поли(этиленгликоль) или, в частности, метоксиполи(этиленгликоль) или его производное, и особенно с молекулярной массой в диапазоне от примерно 15000 Да до примерно 40000 Да.
В одном примере антитела для использования по настоящему изобретению связаны с молекулами поли(этиленгликоля) (ПЭГ). В одном конкретном примере антитело представляет собой фрагмент антитела, и молекулы ПЭГ могут быть присоединены через любую доступную боковую цепь аминокислоты или функциональную группу концевой аминокислоты, находящейся в фрагменте антитела, например, любую свободную амино, имино, тиоловую, гидроксильную или карбоксильную группу. Такие аминокислоты могут существовать естественным образом во фрагменте антитела или могут быть искусственно введены во фрагмент с использованием методов рекомбинантных ДНК (смотри, например, US5219996; US5667425; WO98/25971, WO2008/038024). В одном примере молекула антитела по настоящему изобретению представляет собой модифицированный Fab-фрагмент, в котором модификация представляет собой добавление на C-конце его тяжелой цепи одной или более аминокислот, чтобы сделать возможным присоединение эффекторной молекулы. Предпочтительно, дополнительные аминокислоты образуют модифицированную шарнирную область, содержащую один или более остатков цистеина, с которыми может связываться эффекторная молекула. Можно использовать несколько сайтов для присоединения двух или более молекул ПЭГ.
Предпочтительно, молекулы ПЭГ ковалентно связаны через тиоловую группу по меньшей мере одного остатка цистеина, находящегося в фрагменте антитела. Каждая молекула полимера, присоединенная к модифицированному фрагменту антитела, может быть ковалентно связана с атомом серы остатка цистеина, находящегося в фрагменте. Ковалентная связь будет, как правило, представлять собой дисульфидную связь или, в частности, сероуглеродную связь. Если тиоловую группу используют в качестве точки присоединения соответствующим образом активированных эффекторных молекул, можно использовать, например, тиол-селективные производные, такие как малеинимиды, и производные цистеина. Активированный полимер можно использовать в качестве исходного материала для получения полимер-модифицированных фрагментов антител, как описано выше.
Активированный полимер может быть любым полимером, содержащим тиоловую реакционноспособную группу, такую как α-галокарбоновая кислота или сложный эфир, например, иодацетамид, имид, например, малеинимид, винил сульфон или дисульфид. Такие исходные материалы можно приобретать (например, у компании Nektar, бывшей Shearwater Polymers Inc., Huntsville, AL, USA) или можно получать из коммерчески доступных исходных материалов общепринятыми химическими методами. Конкретные молекулы ПЭГ включают 20K метокси-ПЭГ-амин (поставляемый компаниями Nektar, бывшей Shearwater; Rapp Polymere и SunBio) и M-ПЭГ-SPA (поставляемый компанией Nektar, бывшей Shearwater).
В одном из вариантов осуществления антитело представляет собой модифицированный Fab-фрагмент, Fab'-фрагмент или диFab, который пегилирован, то есть, содержит ПЭГ (поли(этиленгликоль)), ковалентно присоединенный к нему, например, методом, описанным в EP 0948544 или EP 1090037 [смотри также «Poly(ethyleneglycol) Chemistry, Biotechnical and Biomedical Applications», 1992, J. Milton Harris (ed), Plenum Press, New York, «Poly(ethyleneglycol) Chemistry and Biological Applications», 1997, J. Milton Harris and S. Zalipsky (eds), American Chemical Society, Washington DC и «Bioconjugation Protein Coupling Techniques for the Biomedical Sciences», 1998, M. Aslam and A. Dent, Grove Publishers, New York; Chapman, A. 2002, Advanced Drug Delivery Reviews 2002, 54:531-545]. В одном примере ПЭГ присоединен к остатку цистеина в шарнирной области. В одном примере модифицированный ПЭГ Fab-фрагмент имеет малеинимидную группу, ковалентно связанную с одной тиоловой группой в модифицированной шарнирной области. Остаток лизина может быть ковалентно связан с малеинимидной группой, и к каждой из аминогрупп на остатке лизина может быть присоединен полимер метоксиполи(этиленгликоль), имеющий молекулярную массу примерно 20000 Да. Общая молекулярная масса ПЭГ, присоединенного к Fab-фрагменту, таким образом, может составлять примерно 40000 Да.
Конкретные модифицирующие остаток лизина молекулы ПЭГ включают 2-[3-(N-малеимидо)пропионамидо]этиламид N,N'-бис(метоксиполи(этиленгликоля), ММ 20000), другое название PEG2MAL40K (поставляется компанией Nektar, бывшей Shearwater).
Альтернативные источники ПЭГ линкеров включают NOF, который предоставляет GL2-400MA3 (где m в структуре, приведенной ниже, равно 5) и GL2-400MA (где m равно 2), и n составляет примерно 450:
То есть, каждая молекула ПЭГ имеет молекулярную массу примерно 20000 Да.
Таким образом, в одном из вариантов осуществления ПЭГ представляет собой 2,3-бис(метилполиоксиэтиленокси)-1-{[3-(6-малеимидо-1-оксогексил)амино]пропилокси}гексан (разветвленный ПЭГ с 2 ветвями, -CH2)3NHCO(CH2)5-MAL, ММ 40000, известный как SUNBRIGHT GL2-400MA3.
Другие альтернативные эффекторные молекулы ПЭГ следующей формулы:
доступны от компаний Dr Reddy, NOF и Jenkem.
В одном из вариантов осуществления предложено антитело по изобретению, которое пегилировано (например, с использованием ПЭГ, описанных в настоящем документе) за счет присоединения ПЭГ к аминокислотному остатку цистеина в положении, или рядом с положением, 226 в цепи, например, в положении 226 тяжелой цепи (при последовательной нумерации).
В одном из вариантов осуществления настоящего изобретения предложена молекула Fab'-ПЭГ, содержащая один или более полимеров ПЭГ, например, 1 или 2 полимера, таких как 40 кДа полимер или полимеры.
Молекулы Fab'-ПЭГ по настоящему изобретению могут иметь особое преимущество, заключающееся в том, что они имеют время полужизни, независимое от Fc-фрагмента. В одном примере настоящего изобретения предложен способ лечения заболевания, которое облегчается за счет модулирования биологической активности IgE человека, включающий введение терапевтически эффективного количества анти-IgE антитела, или его антигенсвязывающего фрагмента, при этом антитело, или его антигенсвязывающий фрагмент, имеет время полужизни, которое не зависит от связывания Fc с IgE.
В одном из вариантов осуществления предложен фрагмент Fab', конъюгированный с полимером, таким как молекула ПЭГ, молекула крахмала или молекула альбумина.
В одном из вариантов осуществления предложен фрагмент scFv, конъюгированный с полимером, таким как молекула ПЭГ, молекула крахмала или молекула альбумина.
В одном из вариантов осуществления антитело, или фрагмент, конъюгировано с молекулой крахмала, например, для увеличения времени полужизни. Методы конъюгации крахмала с белком описаны в документе US8017739, содержание которого включено в настоящий документ посредством ссылки.
Полинуклеотиды
Настоящее изобретение также относится к выделенной последовательности ДНК, кодирующей тяжелую и/или легкую цепь(и) молекулы антитела по настоящему изобретению. Предпочтительно, последовательность ДНК кодирует тяжелую или легкую цепь молекулы антитела по настоящему изобретению. Последовательность ДНК по настоящему изобретению может включать синтетическую ДНК, например, полученную путем химической обработки, кДНК, геномную ДНК или любое их сочетание.
Последовательности ДНК, кодирующие молекулу антитела по настоящему изобретению, могут быть получены методами, хорошо известными специалистам в данной области. Например, последовательности ДНК, кодирующие часть или все из тяжелой и легкой цепей антитела, могут быть синтезированы, при необходимости, на основании определенных последовательностей ДНК или на основании соответствующих аминокислотных последовательностей.
ДНК, кодирующая акцепторные каркасные последовательности, широко доступна для специалистов в данной области и может быть с легкостью синтезирована на основании соответствующих известных аминокислотных последовательностей.
Можно использовать стандартные методы молекулярной биологии для получения последовательностей ДНК, кодирующих молекулу антитела по настоящему изобретению. Нужные последовательности ДНК могут быть синтезированы полностью или частично с использованием методов олигонуклеотидного синтеза. В соответствующих случаях можно использовать методы сайт-направленного мутагенеза и полимеразной цепной реакции (ПЦР).
Настоящее изобретение также относится к клонирующему или экспрессионному вектору, содержащему одну или более последовательностей ДНК по настоящему изобретению. Соответственно, предложен клонирующий или экспрессионный вектор, содержащий одну или более последовательностей ДНК, кодирующих антитело по настоящему изобретению. Предпочтительно, клонирующий или экспрессионный вектор содержит две последовательности ДНК, кодирующие легкую цепь и тяжелую цепь молекулы антитела по настоящему изобретению, соответственно, и соответствующие сигнальные последовательности. В одном примере вектор содержит межгенную последовательность между последовательностями тяжелой и легкой цепей (смотри WO03/048208).
Общие методы конструирования векторов, методы трансфекции и методы культивирования хорошо известны специалистам в данной области. В этом отношении, ссылка приводится на сборник «Current Protocols in Molecular Biology», 1999, F. M. Ausubel (ed), Wiley Interscience, New York и руководство Maniatis, изданное Cold Spring Harbour Publishing.
Клетки-хозяева, экспрессирующие анти-IgE антитела или их фрагменты
Также предложена клетка-хозяин, содержащая один или более клонирующих, или экспрессионных, векторов, содержащих одну или более последовательностей ДНК, кодирующих антитело по настоящему изобретению. Любую подходящую систему клетка-хозяин/вектор можно использовать для экспрессии последовательностей ДНК, кодирующих молекулу антитела по настоящему изобретению. Можно использовать бактериальные, например, E. coli, и другие микробные системы, либо можно использовать также и эукариотические системы экспрессии, например, с клетками-хозяевами млекопитающих. Подходящие клетки-хозяева млекопитающих включают клетки CHO, миеломы или гибридомы.
Подходящие виды клеток яичника китайского хомяка (клеток CHO) для использования по настоящему изобретению могут включать клетки CHO и CHO-K1, в том числе клетки dhfr- CHO, такие как клетки CHO-DG44 и клетки CHO-DXB11, которые могут быть использованы с селективным маркером DHFR, или клетки CHOK1-SV, которые могут быть использованы с селективным маркером глутамин-синтетазой. Другие виды клеток, используемых для экспрессии антител, включают линии лимфоцитарных клеток, например, клетки NSO миеломы и клетки SP2, клетки COS. Другие подходящие клетки могут включать фибробласты эмбриональной почки человека (hek), например, клетки hek293F и ExpiHek, которые известны в данной области.
Клетки CHO являются предпочтительными в случае полноразмерных Ат по изобретению, поскольку они являются стандартными клетками-хозяевами для продуцирования омализумаба (в одном из вариантов осуществления обеспечивая антителам по изобретению стандартную картину гликозилирования омализумаба) [смотри также WO2013/181577].
Получение анти-IgE антител или их фрагментов
Настоящее изобретение также относится к способу получения молекулы антитела по настоящему изобретению, включающему культивирование клетки-хозяина, содержащей вектор по настоящему изобретению, в условиях, подходящих для инициирования экспрессии белка с ДНК, кодирующей молекулу антитела по настоящему изобретению, и выделение молекулы антитела.
Молекула антитела может содержать только полипептид тяжелой или легкой цепи, в этом случае необходимо использовать только последовательность, кодирующую полипептид тяжелой цепи или легкой цепи, для трансфекции клеток-хозяев. Для получения продуктов, содержащих как тяжелую, так и легкую цепи, линию клеток можно трансфицировать двумя векторами, первым вектором, кодирующим полипептид легкой цепи, и вторым вектором, кодирующим полипептид тяжелой цепи. Альтернативно, можно использовать один вектор, содержащий последовательности, кодирующие полипептиды легкой цепи и тяжелой цепи.
Предложен способ культивирования клетки-хозяина и экспрессии антитела или его фрагмента, выделения последнего и, необязательно, его очистки для получения выделенного антитела или фрагмента. В одном из вариантов осуществления способ дополнительно включает этап конъюгации эффекторной молекулы с выделенным антителом или фрагментом, например, конъюгации с полимером ПЭГ, в частности, как описано в настоящем документе.
В одном из вариантов осуществления предложен способ очистки антитела (в частности, антитела или фрагмента по изобретению), включающий этапы: проведения анионообменной хроматографии в режиме отсутствия связывания таким образом, что примеси удерживаются на колонке, а антитело элюируется.
В одном из вариантов осуществления очистка включает аффинное удержание на колонке с белком A, с последующим титрованием. В одном из вариантов осуществления очистка включает аффинное удержание на колонке с белком G, с последующим титрованием методом ВЭЖХ. В одном из вариантов осуществления очистка включает аффинное удержание на колонке с IgE, с последующим титрованием.
В одном из вариантов осуществления очистка включает использование цибакрона синего или аналогичного реактива для очистки слитых или конъюгированных с альбумином молекул.
Подходящие ионообменные смолы для использования в способе включают смолу Q.FF (поставляемую GE-Healthcare). Этот этап можно, например, осуществлять при значении pH примерно 8.
Способ может дополнительно включать начальный этап захвата с использованием катионообменной хроматографии, выполняемый, например, при значении pH примерно 4-5, например, 4,5. Для катионообменной хроматографии можно, например, использовать такую смолу, как CaptoS или SP сефароза FF (поставляемые GE-Healthcare). Затем антитело или фрагмент можно элюировать со смолы с помощью ионного солевого раствора, такого как хлорид натрия, например, в концентрации 200 мМ.
Таким образом, этап или этапы хроматографии могут включать один или более этапов промывания, в зависимости от ситуации.
Способ очистки также может включать один или более этапов фильтрования, например, этап диафильтрования или этап ВЭЖХ-фильтрования.
Таким образом, в одном из вариантов осуществления предложено очищенное анти-IgE антитело или фрагмент, например, гуманизированное антитело или фрагмент, в частности, антитело или фрагмент по изобретению, в практически очищенной форме, в частности, свободной или практически свободной от эндотоксина и/или белка или ДНК клетки-хозяина.
Использованный выше термин «очищенная форма» должен означать степень чистоты по меньшей мере 90%, например, 91, 92, 93, 94, 95, 96, 97, 98, 99% содержания по массе, или еще большую степень чистоты.
«Практически свободные от эндотоксина», как правило, означает содержание эндотоксина, составляющее 1 МЕ на мг продукта антитела, или менее, например, 0,5 или 0,1 МЕ на мг продукта.
«Практически свободные от белка или ДНК клетки-хозяина» как правило, означает содержание белка и/или ДНК клетки-хозяина, составляющее 400 мкг на мг продукта антитела, или менее, например, 100 мкг на мг, или менее, в частности, 20 мкг на мг, в зависимости от ситуации.
Фармацевтические композиции
Поскольку антитела по настоящему изобретению полезны для лечения и/или профилактики патологического состояния, настоящее изобретение также относится к фармацевтической или диагностической композиции, содержащей антитело, или антигенсвязывающий фрагмент, по настоящему изобретению в сочетании с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя. Соответственно, предложено применение антитела, или антигенсвязывающего фрагмента, по изобретению для производства лекарственного средства. Композиция, как правило, будет предоставлена в виде части стерильной фармацевтической композиции, которая обычно содержит фармацевтически приемлемый носитель. Кроме того, фармацевтическая композиция по настоящему изобретению может содержать фармацевтически приемлемый эксципиент.
Настоящее изобретение также относится к способу получения фармацевтической или диагностической композиции, включающему добавление и смешивание антитела, или антигенсвязывающего фрагмента, по настоящему изобретению с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя.
Антитело, или антигенсвязывающий фрагмент, может быть единственным активным ингредиентом в фармацевтической или диагностической композиции, либо может сопровождаться другими активными ингредиентами, включая другие антитела или ингредиенты, не являющиеся антителами, такие как молекулы стероидов или других лекарственных средств, в частности, молекулы лекарственных средств, время полужизни которых не зависит от связывания IgE.
Предпочтительно, фармацевтические композиции содержат терапевтически эффективное количество антитела, или антигенсвязывающего фрагмента, по изобретению. Используемый в настоящем документе термин «терапевтически эффективное количество» означает количество терапевтического средства, необходимое для лечения, ослабления или профилактики целевого заболевания или состояния, либо для достижения заметного терапевтического или превентивного эффекта. Для любого описанного антитела, или антигенсвязывающего фрагмента, терапевтически эффективное количество можно оценивать первоначально либо в анализах на культурах клеток, либо в животных моделях, как правило, на грызунах, кроликах, собаках, свиньях или приматах. Животную модель также можно использовать для определения соответствующего диапазона концентраций и пути введения. Такую информацию можно использовать для определения используемых для человека доз и путей введения.
Точное терапевтически эффективное количество для индивида-человека будет зависеть от степени тяжести болезненного состояния, общего состояния здоровья индивида, возраста, массы тела и пола индивида, диеты, времени и частоты введения, сочетания(ий) лекарственных средств, чувствительности реакции и устойчивости/ответа на терапию. Это количество может быть определено путем обычного экспериментирования и находится в пределах компетенции лечащего врача. Как правило, терапевтически эффективное количество будет составлять от 0,01 мг/кг до 500 мг/кг, например, от 0,1 мг/кг до 200 мг/кг, например, 100 мг/кг. Фармацевтические композиции, предпочтительно, могут быть предоставлены в стандартных лекарственных формах, содержащих заранее определенное количество активного средства по изобретению в каждой дозе.
Терапевтические дозы антител, или антигенсвязывающих фрагментов, по настоящему изобретению не вызывают явные токсикологические эффекты in vivo.
Предпочтительно, активность IgE in vivo может поддерживаться на соответствующем пониженном уровне за счет последовательного введения доз антитела, или связывающего фрагмента, по изобретению.
Композиции можно вводить пациенту отдельно, либо можно вводить в сочетании (например, одновременно, последовательно или раздельно) с другими средствами, лекарственными средствами или гормонами.
Фармацевтическая композиция также может содержать фармацевтически приемлемый носитель для введения антитела или антигенсвязывающего фрагмента. Носитель сам по себе не должен вызывать выработку антител, вредных для индивидуума, получающего композицию, и не должен быть токсичным. Подходящие носители могут представлять собой крупные, медленно метаболизируемые макромолекулы, такие как белки, полипептиды, липосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы.
Можно использовать фармацевтически приемлемые соли, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, либо соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты.
Кроме того, фармацевтически приемлемые носители в терапевтических композициях могут содержать жидкости, такие как вода, солевой раствор, глицерин и этанол. Кроме того, в таких композициях могут присутствовать вспомогательные вещества, такие как увлажняющие средства или эмульгаторы, или регуляторы pH. Такие носители позволяют формулировать фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, густых суспензий и суспензий для приема внутрь пациентом.
Предпочтительные формы для введения включают формы, подходящие для парентерального введения, например, путем инъекции или инфузии, например, путем болюсной инъекции или непрерывной инфузии. Если препарат предназначен для инъекции или инфузии, он может быть в форме суспензии, раствора или эмульсии в масляной или водной среде, и он может содержать вспомогательные вещества, такие как суспендирующие средства, консерванты, стабилизаторы и/или диспергирующие средства. Альтернативно, молекула антитела может находиться в сухой форме, предназначенной для восстановления в соответствующей стерильной жидкости перед использованием.
После формулирования композиции по изобретению можно вводить непосредственно индивиду. Индивиды, получающие лечение, могут быть животными. Однако, предпочтительно, если композиции адаптированы для введения индивидам-людям.
Фармацевтические композиции по данному изобретению можно вводить самыми разными путями введения, включая, но без ограничения, пероральный, внутривенный, внутримышечный, внутриартериальный, интрамедуллярный, интратекальный, интравентрикулярный, трансдермальный, чрескожный (например, смотри WO98/20734), подкожный, внутрибрюшинный, интраназальный, энтеральный, топический, подъязычный, интравагинальный или ректальный пути. Для введения фармацевтических композиций по изобретению также можно использовать гипоспреи. Как правило, терапевтические композиции могут быть получены в виде инъекционных препаратов, либо в виде жидких растворов, либо в виде суспензий. Также можно готовить твердые формы, подходящие для растворения или суспендирования в жидких средах перед инъекцией.
Прямая доставка композиций, как правило, будет осуществляться путем подкожной (в частности), внутрибрюшинной, внутривенной или внутримышечной инъекции, или доставляться в интерстициальное пространство ткани. Композиции также можно вводить в очаг повреждения. Дозы можно вводить по системе однократных доз или системе многократных доз.
Понятно, что активным ингредиентом в композиции будет молекула антитела. В силу этого, она будет подвержена деградации в желудочно-кишечном тракте. Таким образом, если композицию предстоит вводить через желудочно-кишечный тракт, композиция должна будет содержать средства, защищающие антитело от деградации, но высвобождающие антитело после его абсорбции из желудочно-кишечного тракта.
Подробное описание фармацевтически приемлемых носителей можно найти в Remington's Pharmaceutical Sciences (Mack Publishing Company, N.J. 1991).
Структурно-функциональные свойства
В одном аспекте настоящего изобретения антитело, или антигенсвязывающий фрагмент, связывается со свободным и FcεRI-связанным IgE человека. Когда антитело (или антигенсвязывающий фрагмент) по изобретению связывается с FcεRI-связанным IgE человека, оно стабилизирует конформацию IgE. В такой стабилизированной конформации IgE имеет более слабую аффинность связывания для FcεRI или омализумаба (или его фрагментов), чем в отсутствие антитела, или антигенсвязывающего фрагмента, по настоящему изобретению и при этом FcεRI-связанный человеческий IgE диссоциирует от FcεRI. Предпочтительно, IgE после диссоциации от FcεRI остается связанным с антителом, или антигенсвязывающим фрагментом, описанным в настоящем документе. Как будет показано далее в настоящем документе (например, в Примере 1 и на Фигуре 2), антитело, или антигенсвязывающий фрагмент, по изобретению связывается с IgE в конформации, которая отличается от конформации, которую IgE имеет, будучи связанным с омализумабом.
Без связи с конкретной теорией, антитело по настоящему изобретению вынуждает IgE принимать частично изогнутую конформацию (Фиг. 2C и 5C-D), таким образом, вызывая разгибание структур, характерных для свободного или FcεRI-связанного IgE. Такое разгибание не дает возможность IgE связывать или сохранять связывание с FcεRI. Как будет показано в разделе «Примеры», поскольку антитело по настоящему изобретению конкурирует за сайт связывания на IgE с FcεRI, считается, что антитело может быть способно к образованию комплекса с IgE, связанным с FcεRI, за счет изменения структуры IgE, связанного с FcεRI, и стимуляции его диссоциации от FcεRI. Антитела по настоящему изобретению, обладающие этими свойствами, представляют собой антитела, описанные в настоящем документе, такие как антитела, или антигенсвязывающие фрагменты, содержащие:
a. вариабельную область тяжелой цепи, содержащую SEQ ID NO:1, и вариабельную область легкой цепи, содержащую:
i. SEQ ID NO:109; или
ii. SEQ ID NO:113; или
iii. SEQ ID NO:121; или
iv. SEQ ID NO:132; или
v. SEQ ID NO::139; или
b. SEQ ID NO:5 и
i. SEQ ID NO:24, в которой S77 и S79 заменены на Q;
ii. SEQ ID NO:117 или
iii. SEQ ID NO:125; или
iv. SEQ ID NO::136; или
v. SEQ ID NO:143.
Анти-IgE антитела, или антигенсвязывающие фрагменты, обладающие такими свойствами, контактируют, или контактируют и являются специфическими для эпитопа, содержащего, со ссылкой на SEQ ID NO:108, остатки T373, W374, S375, R376, A377, S378, G379, P381, Q417, C418, R419, P426, R427, A428 Cε3-домена и остатки D278 и T281 Cε2-домена IgE человека.
Антитела по настоящему изобретению имеют в некоторых положениях остатки метионина. Окисление остатков метионина является одним из наиболее распространенных путей деградации белков. Антитела по настоящему изобретению, в которых остатки метионина были введены в положениях S64 и S71, со ссылкой на SEQ ID NO:20, могут быть подвергнуты полному окислению без влияния на способность антител ускорять диссоциацию комплекса IgE-Fc:sFcRIα.
Таким образом, настоящее изобретение также относится к анти-IgE антителу, или антигенсвязывающему фрагменту, которое содержит вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO:1, и вариабельную область легкой цепи, содержащую CDR-L2 и FW-L3, при этом вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO:20, за исключением того, что CDR-L2 имеет аминокислотную последовательность SEQ ID NO:50, и того, что FW-L3 имеет аминокислотную последовательность SEQ ID NO:131 или 138, при этом остатки метионина в положениях 64 и/или 71, со ссылкой на SEQ ID NO:20, являются окисленными.
Настоящее изобретение также относится к анти-IgE антителу, или антигенсвязывающему фрагменту, содержащему вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO:1, и вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO:132 или 139, при этом остатки метионина в положениях 64 и/или 71, со ссылкой на SEQ ID NO:132 или 139, являются окисленными.
Другие остатки метионина в антителах по настоящему изобретению могут быть окислены без влияния на способность антител ускорять диссоциацию комплекса IgE-Fc:sFcRIα.
Далее изобретение будет описано с помощью примеров со ссылками на варианты осуществления, проиллюстрированные в сопроводительных чертежах.
Примеры
Пример 1: Структура мутанта терапевтического анти-IgE антитела омализумаба, связанного с IgE-Fc, позволяет понять механизм его действия
Аннотация
Иммуноглобулин E и его взаимодействие с рецепторами FcεRI и CD23 играет центральную роль в аллергических заболеваниях. Омализумаб, одобренное для клинического применения терапевтическое антитело, ингибирует взаимодействие между IgE и FcεRI, предотвращая активацию тучных клеток и базофилов, и блокирует связывание IgE с CD23. Авторы изобретения решили кристаллическую структуру комплекса 2:1 между полученным из омализумаба Fab и IgE-Fc, с одним Fab-фрагментом, связанным с каждым Cε3-доменом (но только одним из Fab-фрагментов, связанным с Cε2-доменом). Хотя свободный IgE-Fc в растворе преимущественно является резко изогнутым, в комплексе он лишь частично изогнут, что предотвращает взаимодействие с FcεRI; связывание CD23 ингибируется стерически вследствие перекрывания сайтов связывания на каждом Cε3-домене. Анализ взаимодействия в состоянии раствора демонстрирует ортостерическую и аллостерическую основу для ингибирования взаимодействий обоих рецепторов и, совместно со структурой, показывает, каким образом омализумаб (и, в частности, описанные мутанты омализумаба) может ускорять диссоциацию связанного с рецептором IgE от FcεRI, с использованием внутренней динамики и аллостерического потенциала IgE.
Введение
Антитела иммуноглобулины E (IgE) играют важную роль в аллергических заболеваниях, связываясь с аллергенами своими Fab-плечами и осуществляя свои эффекторные функции путем связывания с рецепторами для Fc-области1. Двумя основными рецепторами IgE являются FcεRI и CD23/FcεRII, которые обычно называют высоко- и низкоаффинными рецепторами, соответственно. На тучных клетках и базофилах IgE связывается с FcεRI настолько прочно (KD ≈ 10-10 M-1), что такие клетки сенсибилизируются заранее связанными IgE, и необходимо лишь присутствие аллергена для перекрестной сшивки комплексов IgE/FcεRI и вызывания немедленной реакции. CD23 представляет собой гомотример и, таким образом, исходно низкая аффинность каждого связывания IgE подобным лектину C-типа «головным» доменом (KD ≈ 10-7 M-1) может быть усилена за счет эффекта авидности при связывании с агрегированным IgE в иммунных комплексах, и почти соответствовать аффинности FcεRI для IgE2. CD23, экспрессируемый на B-клетках, участвует в регуляции IgE, и экспрессируемый на клетках дыхательных путей и кишечного эпителия, опосредует трансцитоз комплексов IgE/аллерген1,2. Как FcεRI, так и CD23, экспрессируются на разных антигенпредставляющих клетках. Таким образом, взаимодействия IgE-рецептор вовлечены в несколько аспектов аллергического ответа, и IgE является давней мишенью для терапевтического вмешательства3.
Fc-область IgE включает связанный дисульфидной связью димер трех доменов: Cε2, Cε3 и Cε4. Результаты ранних исследований методом FRET химерного IgE4,5 и изучения рентгеновского рассеяния растворов IgE-Fc6 указывали на компактную, изогнутую структуру, и более позднее изучение кристаллической структуры IgE-Fc позволило выявить резко и ассиметрично изогнутую конформацию, с парой доменов (Cε2)2, загнутых назад на домены Cε3 и Cε47. Изгиб, определяемый как угол между локальной сдвоенной осью пары доменов (Cε2)2 и таковой у Fcε3-4 (область, содержащая только домены Cε3 и Cε4), как обнаружено, становится еще более резким (от 62° до 54°) в кристаллической структуре IgE-Fc, связанного с sFcεRIα, растворимыми внеклеточными доменами IgE-связывающей α-цепи рецептора8. Недавние исследования методом FRET с меченым на N- и C-концах IgE-Fc подтвердили это увеличение изгиба при связывании sFcεRIα9.
Сайт связывания FcεRI охватывает оба Cε3-домена в области, проксимальной к Cε28,10, хотя Cε2-домен не вовлечен непосредственно; задействование обеих цепей определяет стехиометрию связывания 1:1. Напротив, две молекулы CD23 связываются с IgE-Fc, по одной для каждой цепи, и на другом, проксимальном к Cε4 конце Cε3-домена11,12,13. Связывание CD23 также вызывает конформационное изменение в IgE-Fc14, но не такое, которое сильно влияет на изгиб9. Однако относительно «закрытое» расположение Cε3-доменов в комплексе с растворимым головным доменом CD23 (sCD23), в сравнении со свободным IgE-Fc, несовместимо с более «открытым» расположением этих доменов, необходимым для связывания FcεRI. Это частично объясняет взаимное исключение связывания FcεRI и CD2311,12, хотя вероятно, что и другие факторы, такие как локальные конформационные изменения и модификации конформационной динамики15, также вносят вклад в аллостерическое взаимодействие между двумя сайтами связывания рецепторов2.
Более высокая степень гибкости в IgE-Fc недавно была обнаружена при изучении комплекса с анти-IgE-Fc Fab, называемым aεFab16. Две молекулы aεFab связываются с IgE-Fc симметричным образом, по одной с каждым Cε3-доменом, захватывая полностью вытянутую конформацию, в которой локальные сдвоенные оси доменов (Cε2)2 и область Fcε3-4 практически совпадают. Анализ образования комплекса в растворе, наряду с моделированием молекулярной динамики свободного IgE-Fc, свидетельствовали о том, что пара доменов (Cε2)2 может «переворачиваться» с одной стороны области Fcε3-4 на другую16. Конформация IgE-Fc, стабилизированная этим анти-IgE антителом, несовместима со связыванием FcεRI, что объясняет его ингибирующую активность.
Омализумаб представляет собой анти-IgE моноклональное IgG1 антитело, которое одобрено для терапевтического применения (ксолар®, Novartis)17. Он связывается со свободным IgE и ингибирует связывание как FcεRI, так и CD23; сайт связывания был картирован на Cε3-домен методами пептидного ингибирования и молекулярного моделирования18,19, однако механизм действия остается неизвестным. Тем не менее, связывание с FRET-меченым IgE-Fc указывает на небольшую степень разгибания9 и, следовательно, на возможность аллостерического, а не прямого ингибирования.
Недавно был обнаружен тип ингибитора, который активно разрушает предварительно образованные комплексы IgE/FcεRI: было обнаружено, что искусственный белок с анкириновыми повторами (Designed Ankyrin Repeat Protein (DARPin)) связывается с Cε3-доменом связанного с рецептором IgE и ускоряет его диссоциацию от FcεRI20. Кристаллическая структура комплекса 2:1 из DARPin E2_79 с молекулой Fcε3-4, ограниченного за счет искусственно созданной дисульфидной связи, позволила выяснить характер и расположение сайта связывания, однако механизм действия остался неизвестным. Впоследствии сообщалось, что омализумаб может аналогично способствовать диссоциации FcεRI-связанного IgE, хотя и при очень высоких концентрациях, которые значительно выше, чем те, которые могут быть достигнуты при терапевтическом применении21,22.
В настоящем документе авторы изобретения приводят кристаллическую структуру комплекса между IgE-Fc и новым фрагментом антитела, Fab, полученным из омализумаба (Fab3 омализумаба), который имеет три точечные мутации дистально по отношению к связывающим антиген (IgE-Fc) определяющим комплементарность областям (CDR). Мутациями являются S81R, Q83R и L158P со ссылкой на SEQ ID NO:125 (или S77R, Q79R и L154P со ссылкой на SEQ ID NO:129). Структура комплекса позволяет понять механизм действия омализумаба, и исследования растворов показывают, что данный механизм основан на внутренней динамике IgE.
Результаты
Несмотря на большие усилия, попытки кристаллизации IgE-Fc в комплексе с Fab омализумаба приводили к избирательной кристаллизации только Fab-фрагмента. Другие авторы также сообщали об аналогичных неудачных попытках кристаллизации данного комплекса23. Вследствие этого авторы изобретения сконструировали новое антитело, Fab, полученный из омализумаба, с тремя точечными мутациями, две в каркасной области домена VL (Ser81Arg, Gln83Arg) и одна в домене Cκ (Leu158Pro) (SEQ ID NO:125, нумерация PDB) (Фиг. 1), с целью нарушения благоприятных кристаллических контактов, наблюдаемых в кристаллической структуре Fab омализумаба (результаты описаны в другом документе). Авторы изобретения назвали этот полученный из омализумаба Fab-фрагмент «Fab3 омализумаба».
Общая структура комплекса IgE-Fc/Fab3 омализумаба
Авторы изобретения определили кристаллическую структуру комплекса между IgE-Fc и Fab3 омализумаба с разрешением 3,7 Å (Фиг. 2A). Две молекулы Fab3 омализумаба (Fab1 и Fab2) связываются с ассиметричной, частично изогнутой молекулой IgE-Fc, при этом каждый Fab связывает один Cε3-домен (Фиг. 2B-C). Fab1 связывает Cε3-домен цепи B IgE-Fc, в то время как Fab2 связывает Cε3-домен цепи A IgE-Fc. Вследствие частично изогнутой конформации IgE-Fc в комплексе легкая цепь Fab2 также вступает в незначительное взаимодействие с Cε2-доменом из цепи B IgE-Fc (далее в данном примере приведена более подробная информация о данном взаимодействии).
Граница контакта между IgE-Fc и Fab3 омализумаба
Каждая молекула Fab3 омализумаба связывает один край открытой поверхности Cε3-домена (цепи C, C', F и G, и основание связывающей рецептор FcεRI петли FG). Вовлечены как тяжелая, так и легкая цепь Fab3 омализумаба, при этом первая из них отвечает за ~60% площади контакта ~715 Å2 (Фиг. 2 и 3).
Контакты тяжелой цепи Fab3 омализумаба (SEQ ID NO:5), которые слегка различаются между двумя поверхностями контакта, можно кратко описать следующим образом: Gly32 и Tyr33 (CDRH1) вступают в ван-дер-ваальсовы взаимодействия с Ala377 и Ser378 (Cε3) (последовательность IgE-Fc приведена в SEQ ID NO:108 и на Фиг. 16), в то время как Tyr54 (CDRH2) контактирует с Gly379-Pro381 (Cε3). Остатки CDRH3 участвуют в наибольшей площади контакта и претерпевают значительное конформационное изменение при образовании комплекса, в сравнении с несвязанными структурами Fab (неопубликованные результаты19,23). Все из остатков Ser100, His101, Tyr102 и Trp 106 CDRH3 вступают в ван-дер-ваальсовы взаимодействия с остатками Cε3-домена, которые включают Ser375-Gly379, Gln417 и Arg419 (Cε3). Однако наиболее яркой особенностью этой части поверхности контакта является взаимодействие с Phe103 (CDRH3). Phe103 в основном погружен в «карман», создаваемый Thr373, основной цепью Trp374, Ser375, Gln417 и Arg419 (Cε3), и вступает в катион/π стэкинг-взаимодействие с Arg419 (Фиг. 3).
Arg419 (Cε3) также играет ключевую роль во взаимодействии с легкой цепью Fab3 омализумаба (SEQ ID NO:125) (Фиг. 3). Arg419 (Cε3) находится в пределах позволяющего образовывать водородные связи расстояния от карбонильных атомов кислорода основных цепей Tyr31 (CDRL1) и Asp32 (CDRL1), помимо контактирования с боковыми цепями Asp32, Asp34 и Tyr36 (с образованием водородной связи с гидроксильной группой Tyr36). Asp32 также вступает в ван-дер-ваальсовы взаимодействия с Thr373 и Thr421 (Cε3). Напротив, только два остатка CDRL2 участвуют в границе контакта: Tyr53 (CDRL2) контактирует с Gln417 (Cε3), и как Tyr53, так и Tyr57, вступают в ван-дер-ваальсовы взаимодействия с Met430 (Cε3); Tyr57 также образует водородную связь с остовом Met430. Что касается взаимодействий тяжелой цепи, имеют место небольшие различия в контактах с легкой цепью в случае Fab1 и Fab2.
Контактные остатки CDR на Fab3 омализумаба; нумерация в формате (pdb/Kabat/Chothia). Последовательность тяжелой цепи: SEQ ID NO:5; последовательность легкой цепи: SEQ ID NO:125
CDRH1: Ser (31/31/31), Gly (32/32/31a), Tyr (33/33/32)
CDRH2: Tyr (54/53/53)
CDRH3: Ser (100/96/96), His (101/97/97), Tyr (102/98/98), Phe (103/99/99), Trp (106/101B/101B)
CDRL1: Asp (30/27C/30), Tyr (31/27D/30A), Asp (32/28/30B), Gly (33/29/30C), Asp (34/30/30D), Tyr (36/32/32)
CDRL2: Tyr (53/49/49), Ser (56/52/52), Tyr (57/53/53), Ser (60/56/56)
CDRL1 и CDRH3 имеют самое большое число остатков, вовлеченных во взаимодействие, и, таким образом, определяют, каким образом омализумаб связывается и ориентируется относительно IgE-Fc. CDRL3 не вовлечена в связывание с IgE-Fc.
Сравнение границы контакта Fab3 омализумаба с другими анти-IgE комплексами
Сайты связывания на Cε3-домене для Fab3 омализумаба и недавно описанного DARPin E2_7920 перекрываются (Фиг. 4) и имеют сходный размер: ~715 Å2 и ~753 Å2, соответственно. Остатки Cε3-домена, являющиеся общими для двух поверхностей контакта, включают Ser375-Gly379, Gln417, Arg419, Arg427 и Met430, но в то время, как Fab3 омализумаба образует более тесные контакты с связывающей рецептор петлей FG Cε3, поверхность контакта DARPin E2_79 продолжается в противоположном направлении, включая линкер домена Cε3-4.
Перекрывающиеся сайты связывания Fab3 омализумаба и DARPin E2_79 заметно отличаются от поверхности контакта, недавно описанной для Fab3 омализумаба, который захватывает IgE-Fc в полностью вытянутой конформации16 (Фиг. 5). Не только площадь поверхности контакта Fab3 омализумаба примерно в два раза превышает таковую для Fab3 омализумаба и DARPin E2_79, ~1400 Å2, но Fab3 омализумаба связывает IgE-Fc в сайте с центром на Arg393 в Cε3 и также контактирует с остатками в Cε2-домене и линкером Cε2-Cε316. Кристаллическая структура другого анти-IgE Fab антитела, MEDI4212, в комплексе 2:1 с Fcε3-4 позволила обнаружить еще один сайт связывания антитела с Cε3-доменом, этот сайт включает N-связанный олигосахаридный фрагмент на Asn39424.
IgE-Fc принимает частично изогнутую конформацию при связывании с Fab3 омализумаба
IgE-Fc в растворе преимущественно находится в изогнутом состоянии5,6,9,25,26,27,28, и кристаллическая структура свободного IgE-Fc продемонстрировала резко изогнутую (62°), асимметричную конформацию, в которой пара доменов (Cε2)2 загнуты назад на домены Cε3 и Cε4 (Фиг. 5A-B), при этом Cε2-домен одной цепи (цепи B) контактирует с Cε4-доменом другой (цепи A)7,8. IgE-Fc становится еще более резко изогнутым (54°) при связывании FcεRIα8,9, и связанные конформационные изменения включают угловое смещение Cε3-домена цепи A вместе с парой доменов (Cε2)2, в виде жесткого блока, дальше от Cε3-домена цепи B8.
В отличие от комплекса с Fab3 омализумаба, в котором IgE-Fc принимает полностью вытянутую, линейную конформацию16, IgE-Fc принимает частично изогнутую конформацию в комплексе с Fab3 омализумаба (Фиг. 2C и 5C-D), что согласуется с полученными ранее результатами FRET-анализа, которые показали, что омализумаб вызывает разгибание IgE-Fc9. Сайт, с которым связывается Fab1, является экспонированным в свободном, резко изогнутом IgE-Fc, но дальнейшее разгибание IgE-Fc, до чуть более 90°, требуется, чтобы сайт, занимаемый Fab2, стал доступным. Это разгибание IgE-Fc в комплексе с Fab3 омализумаба связано с открытием обоих Cε3-доменов, что приводит к созданию почти симметричной области Fcε3-4 (Фиг. 2B). Пара доменов (Cε2)2 расположена между доменами Cε3 и Cε4 из каждой цепи, и больше не связана так близко с Cε3-доменом из цепи A.
Проведенное недавно моделирование молекулярной динамики, в котором исследовали разгибание IgE-Fc до вытянутой структуры, позволило установить, что хотя резко изогнутая конформация, наблюдаемая в кристаллической структуре свободного IgE-Fc, занимает самый низкий энергетический уровень, можно наблюдать и другой, отчетливый и определенный энергетический уровень, соответствующий частично изогнутым конформациям IgE-Fc16. Частично изогнутая конформация, принимаемая IgE-Fc в комплексе Fab3 омализумаба/IgE-Fc, занимает этот конкретный энергетический уровень (Фиг. 6).
Cε3-домены принимают отчетливо открытую конформацию в комплексе Fab3 омализумаба/IgE-Fc
В кристаллических структурах IgE-Fc и субфрагмента Fcε3-4 Cε3-домены принимают целый ряд разных ориентаций7,8,10,11,13,14,16,24,29, свойство, связанное с аллостерической регуляцией связывания IgE с его двумя основными рецепторами, FcεRI и CD238,11,12,14. Как расстояние между Cε3-доменами, так и их положение относительно Cε4-доменов, было использовано для описания многообразия конформаций, наблюдаемых для области Fcε3-429 (полное описание этих измерений приведено далее в данном примере). В комплексе Fab3 омализумаба/IgE-Fc Cε3-домены расположены дальше друг от друга и от Cε4-доменов, чем в любой другой кристаллической структуре, содержащей IgE-Fc или Fcε3-4, и, таким образом, принимают наиболее открытую конформацию, известную в настоящее время (Фиг. 5); эта конформация является значительно более открытой, чем конформация FcεRI-связанного IgE-Fc (Фиг. 7).
Эффект Fab3 омализумаба на связывание рецепторов FcεRI и CD23
Омализумаб ингибирует не только взаимодействие между IgE-Fc и FcεRI, но также и взаимодействие между IgE-Fc и CD2330. В соответствии с этим, сравнение комплексов Fab3 омализумаба/IgE-Fc и CD23/Fcε3-411 позволило выявить конкуренцию между Fab3 омализумаба и CD23 на обоих сайтах связывания CD23 на Fcε3-4. Более того, остатки Arg376, Ser378 и Lys380 Cε3-домена вовлечены в связывание как Fab3 омализумаба, так и CD2311,31.
В отличие от связывания CD23 с IgE, FcεRIα связывается через оба Cε3-домена. Однако в комплексе Fab3 омализумаба/IgE-Fc Cε3-домены принимают конформацию, которая является слишком открытой, чтобы допустить одновременное связывание обеих цепей (Фиг. 8). Более того, если произвести наложение комплексов Fab3 омализумаба/IgE-Fc и sFcεRIα/IgE-Fc на каждый из Cε3-доменов по очереди, в каждом случае происходят потенциальные стерические столкновения. При наложении на Cε3-домен цепи A Fab2 Fab3 омализумаба сталкивались бы с парой доменов (Cε2)2 в комплексе с FcεRIα, и линкер Cε2-Cε3 из комплекса с Fab3 омализумаба потенциально сталкивался бы с FcεRIα. При наложении на Cε3-домен цепи B, потенциально происходило бы столкновение как Fab1 Fab3 омализумаба, так и линкера Cε2-Cε3 (из комплекса с Fab3 омализумаба), с FcεRIα - хотя сайты связывания для Fab1 и FcεRIα фактически не перекрываются.
Однако остатки CDRL1 Fab3 омализумаба расположены в непосредственной близости от FcεRIα-связывающей петли FG Cε3-домена. Данная петля в цепи B участвует в гидрофобном взаимодействии типа «пролиновый сэндвич», в котором Pro426 в Cε3 располагается между двумя остатками триптофана в FcεRIα. Asp32 (CDRL1) контактирует с Thr421, Gly33 (CDRL1) контактирует с Pro426, Arg427 и Ala428, и Asp34 (CDRL1) контактирует с Arg427 и Ala428. Эти взаимодействия приводят к изменению положения петли FG Cε3-домена и дополнительно мешали бы связыванию IgE с FcεRI.
Недавно было сообщено о связывании омализумаба с FcεRIα-связанным IgE31,32, хотя сложно представить, как омализумаб может связывать FcεRI-связанный IgE, исходя из статических кристаллических структур IgE-Fc в комплексе с sFcεRIα8 и Fab3 омализумаба. Вследствие этого, авторы изобретения изучили связывание Fab3 омализумаба с IgE-Fc и охарактеризовали взаимодействие между Fab3 омализумаба и комплексом IgE-Fc/FcεRI. Результаты позволили составить представление о механизме действия омализумаба.
Взаимодействие Fab3 омализумаба с IgE-Fc в растворе
Авторы изобретения охарактеризовали взаимодействие IgE-Fc/Fab3 омализумаба двумя разными способами, либо путем непосредственной иммобилизации Fab3 омализумаба на поверхности и связывания IgE-Fc, либо путем связывания Fab3 омализумаба с His-маркированным IgE-Fc, захваченным на поверхности сенсора в анализе ППР. His-маркированный на C-конце конструкт IgE-Fc был захвачен с использованием антитела против His-маркера (GE Healthcare), и было проведено сравнение характеристик связывания Fab3 омализумаба, интактного омализумаба и Fab омализумаба. Не удивительно, что в экспериментах по конкурентному связыванию все три молекулы конкурировали за одни и те же сайты связывания и демонстрировали примерно сходные показатели аффинности связывания (данные не представлены). Конструкт Fab3 омализумаба имел слегка более высокую аффинность в сравнении с Fab3 омализумаба и интактным омализумабом (Фиг. 8A-C). В соответствии с кристаллической структурой, две молекулы Fab3 омализумаба связываются с IgE-Fc: связывание является явно двухфазным, с высокоаффинным взаимодействием (~1 нМ), наблюдаемым при низких концентрациях лиганда, и вторым (более слабым) сайтом связывания (~30 нМ), наблюдаемым при более высоких концентрациях (Фиг. 9A).
Эксперимент ППР в сэндвич-формате позволил отдельно охарактеризовать два сайта связывания IgE-Fc/Fab3 омализумаба. Используя данный подход, Fab3 омализумаба ковалентно иммобилизовали на поверхности сенсора и IgE-Fc пропускали над этой поверхностью. При низких концентрациях в данных условиях высокоаффинный сайт доминирует при взаимодействии и кривые связывания могут быть описаны в соответствии с кинетикой однофазного взаимодействия (KD ~ 1 нМ, kon ~ 1,2×106 M-1 с-1, koff ~ 8×10-4 с-1). Этот комплекс 1:1 IgE-Fc/Fab3 омализумаба, захваченный на поверхности биосенсора при ППР, затем может быть использован для измерения связывания второй молекулы Fab3 омализумаба, связывание которой является значительно более слабым (KD ~ 30 нМ, kon ~ 2×105 M-1 с-1, koff ~ 6×10-3 с-1), чем первой (Фиг. 9B).
Конкуренция между Fab3 омализумаба и FcεRIα за сайты связывания и образование комплекса Fab3 омализумаба/IgE-Fc/FcεRIα
Затем авторы изобретения исследовали способность Fab3 омализумаба влиять на взаимодействие между IgE-Fc и FcεRIα. В экспериментах по конкурентному связыванию в растворе Fab3 омализумаба в возрастающих концентрациях ингибировал связывание IgE-Fc с FcεRIα (Фиг. 8D). Механистически, Fab3 омализумаба влияет как на число доступных сайтов связывания (Bmax), так и на кажущуюся KD взаимодействия IgE-Fc/FcεRIα; это характерно для смешанного механизма ингибирования33. Уменьшение значений Bmax является показателем процесса аллостерического ингибирования, а уменьшение кажущейся аффинности взаимодействия чаще всего связано с прямой конкуренцией за общий сайт связывания (то есть, ортостерическое ингибирование), но также может наблюдаться для некоторых аллостерических ингибиторов. Принимая во внимание сайты связывания, наблюдаемые в кристаллической структуре, возможно, что Fab3 омализумаба ингибирует связывание IgE-Fc с FcεRI как по ортостерическому, так и по аллостерическому, механизмам.
Конкуренция между омализумабом и FcεRIα за сайты связывания была описана во многих публикациях, но всегда интерпретировалась как прямая конкуренция за один и тот же (или перекрывающийся) сайт связывания. Эту интерпретацию часто использовали для объяснения того, почему омализумаб не может связываться с комплексами IgE-FcεRI на клетках. Однако авторы изобретения установили, что Fab3 омализумаба может связываться с высокой аффинностью с IgE-Fc, который уже связан с FcεRIα (Фиг. 9C, врезка). В других исследованиях было высказано предположение о существовании комплекса омализумаб/IgE-Fc/FcεRI31,32, однако данный комплекс ранее не был экспериментально охарактеризован. Данные свидетельствуют о том, что хотя связывание IgE-Fc с FcεRIα не привело к сильному изменению аффинности Fab3 омализумаба для IgE-Fc, оно привело к заметному изменению числа доступных сайтов связывания для Fab3 омализумаба в популяции FcεRIα-связанных молекул IgE-Fc. Авторы изобретения сравнивали значения KD и Bmax связывания для молекулы IgE-Fc, захваченной через антитело против His-маркера, со значениями для молекулы, захваченной sFcεRIα, и установили, что FcεRIα-связанный IgE-Fc имеет менее 10% сайтов связывания Fab3 омализумаба в сравнении с IgE-Fc, захваченным через His-маркер, который, как и ожидалось, демонстрирует уровни связывания, согласующиеся со стехиометрией 2:1 (Фиг. 9C). Таким образом, в отличие от того, что предполагалось ранее, омализумаб не потому не связывается с IgE, связанным с тучными клетками, что сайты связывания FcεRIα и омализумаба перекрываются. Вместо этого, FcεRIα влияет на IgE-Fc аллостерически, изменяя динамическое равновесие разных конформаций IgE-Fc16, что приводит к значительному сокращению числа сайтов связывания омализумаба в популяции FcεRIα-связанных молекул IgE-Fc.
Механизм опосредованной Fab3 омализумаба ускоренной диссоциации комплекса IgE-Fc/FcεRIα
В публикации Kim et al.20 сообщалось, что DARPin E2_79 может ускорять распад ранее образовавшихся комплексов IgE-FcεRI. В продолжение этих результатов, в публикации Eggel et al.21 позже было продемонстрировано, что омализумаб также может стимулировать диссоциацию IgE от FcεRI. Аналогично, авторы изобретения обнаружили, что, когда Fab3 омализумаба связан с комплексом IgE-Fc/FcεRIα, он может ускорять диссоциацию IgE-Fc от FcεRIα (Фиг. 9D), и что Fab3 омализумаба делает это более эффективно, чем Fab3 омализумаба, и намного более эффективно, чем интактный омализумаб (Фиг. 10E). Один Fab связывается с комплексом IgE-Fc/FcεRIα, но не ускоряет диссоциацию IgE-Fc от FcεRIα. Поразительно, но похоже, что ускоренная диссоциация имеет место только после связывания со вторым сайтом связывания (то есть, низкоаффинным сайтом). Тетрамолекулярный комплекс (Fab3 омализумаба)2/IgE-Fc/FcεRIα должен изменять энергетическое окружение IgE-Fc так, чтобы был заметно снижен энергетический барьер для диссоциации IgE-Fc/FcεRIα, что приводит к быстрой диссоциации в других условиях очень стабильного комплекса.
Детали взаимодействия между легкой цепью Fab3 омализумаба и Cε2-доменом
В комплексе Fab3 омализумаба/IgE-Fc один Cε2-домен вступает в незначительно взаимодействие на площади примерно 260 Å2 (в сравнении со средней площадью взаимодействия ~715 Å2 между Fab3 омализумаба и Cε3-доменом) с двумя из мутантных остатков (Ser81Arg и Gln83Arg). Контакт между Pro158 и IgE-Fc отсутствует.
Боковая цепь Arg81 из легкой цепи Fab2 (один из мутантных остатков в Fab3 омализумаба; (SEQ ID NO:125, нумерация PDB)), укладывается напротив Val277 и Asp278 Cε2-домена из цепи B (SEQ ID NO:108). Ser80 (Fab3 омализумаба) укладывается напротив Asp278, Leu279 и Thr281 (Cε2-домен), в то время как Ser64 (Fab3 омализумаба) укладывается напротив Asp276 и Asp278. Ser64 и Ser80 являются идентичными в омализумабе и Fab3 омализумаба.
В комплексе Fab3 омализумаба/IgE-Fc Arg83 (один из мутантных остатков в Fab3 омализумаба), по-видимому, не контактирует с Cε2-доменом, вследствие нарушения в боковой цепи Asp278 (Cε2-домен). Однако, если боковая цепь Asp278 была бы в порядке, водородная связь или солевой мост потенциально могли бы образовываться между Arg83 и Asp278.
Определенные методом кристаллографии контакты в комплексе Fab3 омализумаба/IgE-Fc
Контакты между антителом и антигеном в пределах 4 Å в кристаллической структуре, как правило, являются характерными для поверхности контакта эпитоп/паратоп.
Остатки IgE-Fc в пределах 4 Å от областей CDR тяжелой и легкой цепей Fab3 омализумаба определяют следующий эпитоп:
T 373, W 374, S 375, R 376, A 377, 3 S 78, G 379, P 381, Q 417, C 418, R 419, P 426, R 427, A 428 (на цепи A).
Кроме того, остатки IgE-Fc в пределах 4 Å от остатков FR1 и FR3 легкой цепи Fab3 омализумаба продлевают эпитоп до:
D 278, T 281 (на цепи B - Cε2-домен) [с контактами с R18, S64, S80 и R81 антитела - и, кроме того, R83, как обнаружено при моделировании молекулярной динамики, описанном в Примере 5].
Контакты между антителом и антигеном в пределах 5 Å в кристаллической структуре также являются информативными для определения поверхности контакта антитело/антиген.
Дополнительные остатки IgE-Fc в пределах 5 Å от областей CDR тяжелой и легкой цепей Fab3 омализумаба представляют собой: K380, M430 (на цепи A).
Кроме того, дополнительными остатками IgE-Fc в пределах 5 Å от остатков FR1 и FR3 легкой цепи Fab3 омализумаба являются: D276, V277, L279, S280, A282 (на цепи B - Cε2-домен) [с дополнительными контактами с G16 в FR1 и R65 в FR3].
Анализ ориентации Cε3-домена в комплексе Fab3 омализумаба/IgE-Fc
В одном методе анализа положения Cε3-доменов относительно Cε4-доменов межатомное расстояние между Cα-атомом Asn394 Cε3-домена из одной цепи и Cα-атомом Lys497 Cε4-домена из другой цепи было использовано для описания «открытости» Cε3-доменов29. Межатомное расстояние между Cα Val336 было использовано для описания «размаха», или того, насколько близко Cε3-домены находятся друг к другу29.
Для FcεRI-связанного IgE-Fc и FcεRI-связанного Fcε3-4, в которых Cε3-домены принимают открытую конформацию, значения «открытости» находятся в диапазоне 23,5-28,4 Å, в то время как значения «размаха» составляют в среднем 23,3 Å8,10. Соответствующие значения для комплекса Fab3 омализумаба/IgE-Fc составляют в среднем 29,5 для «открытости» и 29,4 Å для «размаха». В комплексе с Fab3 омализумаба Cε3-домены принимают наиболее открытую конформацию (наиболее далеки друг от друга), описанную до настоящего времени.
Обсуждение
Авторы изобретения сообщают о полученной с разрешением 3,7 Å структуре комплекса между IgE-Fc и Fab-фрагментом, полученным из терапевтического анти-IgE антитела омализумаба; авторы изобретения называют данный Fab-фрагмент, содержащий три точечные мутации в каркасных областях, удаленных от антигенсвязывающего сайта, Fab3 омализумаба. Структура демонстрирует две молекулы Fab3 омализумаба в комплексе с IgE-Fc, по одной, связанной с каждым из двух Cε3-доменов (но только один из Fab связан с Cε2-доменом), и позволяет понять способность омализумаба к ингибированию связывания IgE как с FcεRI, так и с CD23. Также обнаружено, что IgE-Fc принимает частично изогнутую конформацию в комплексе с Fab3 омализумаба, это согласуется с результатами более ранних исследований авторов изобретения с использованием FRET-меченого IgE-Fc, которые указывают на легкое разгибание относительно свободного IgE-Fc9.
В растворе IgE-Fc находится преимущественно в изогнутом состоянии5,6,9,25,26,27,28, и на кристаллической структуре свободного IgE-Fc пара доменов (Cε2)2 загнуты назад напротив доменов Cε3 и Cε47,8. В последнее время было достигнуто более глубокое понимание конформационной гибкости IgE-Fc, когда авторы изобретения решили структуру полностью вытянутой конформации, которая имеет место в комплексе с анти-IgE-Fc Fab (aεFab)16. Моделирование молекулярной динамики, изучение разгибания IgE-Fc из полностью изогнутой в вытянутую конформацию, выявило энергетические уровни, соответствующие частично изогнутым конформациям (Фиг. 6). Комплексу Fab3 омализумаба/IgE-Fc, описанному в настоящем документе, в котором изгиб между парой доменов (Cε2)2 и доменами Fcε3-4 составляет ~90°, соответствует определенный энергетический уровень в данном моделировании16. Интересно, что расположение сайта связывания Fab3 омализумаба не препятствовало бы дальнейшему разгибанию до полностью вытянутой конформации, и, следовательно, возможно, что IgE-Fc может претерпевать значительные изменения в конформации, даже находясь в комплексе с омализумабом.
В дополнение к изгибу пары доменов (Cε2)2 относительно доменов Cε3 и Cε4, разные структуры комплекса IgE-Fc, Fcε3-4 и рецептора показали, что Cε3-домены могут принимать ряд относительных ориентаций, от закрытой до открытой7,8,10,11,13,14,16,24,29. Открытие и закрытие Cε3-доменов играет определенную роль в аллостерической регуляции связывания с рецептором IgE-Fc11,12: в комплексе с CD23 они относительно закрыты11,13,14, в то время как в комплексе с FcεRI они более открыты8,10. Сравнение структур комплексов CD23/Fcε3-4 и Fab3 омализумаба/IgE-Fc показывает, что сайты связывания CD23 и омализумаба перекрываются, и эксперименты по конкурентному связыванию показывают, что ингибирование связывания IgE с CD23 омализумабом является безусловно ортостерическим.
Однако ингибирование связывания FcεRI является более сложным. В комплексе с Fab3 омализумаба Cε3-домены принимают более открытую конформацию, чем можно было видеть на известной ранее структуре, настолько, что два подсайта взаимодействия между IgE-Fc и FcεRI, по одному с участием каждого Cε3-домена, не могут связываться одновременно. Другой вклад в данное ингибирование может являться результатом проксимального расположения молекулы Fab3 омализумаба (Fab1) относительно рецептор-связывающей петли FG в Cε3, что может непосредственно влиять на конформацию контактных остатков для FcεRIα. И наконец, даже если эпитопы связывания Fab3 омализумаба и FcεRIα на IgE-Fc строго не перекрываются, существует вероятность стерических столкновений при одновременном связывании обоих. Таким образом, кристаллическая структура свидетельствует о том, что механизм ингибирования омализумаба является, в основном, аллостерическим, но с возможным ортостерическим компонентом.
Исследования методом ППР позволили авторам изобретения оценивать кинетику и показатели аффинности для двух сайтов связывания Fab3 омализумаба. Два значения аффинности заметно отличались, с величинами KD ~1 нМ и ~30 нМ, первая из которых связана с более высокой константой скорости ассоциации (kon ~ 2×106 M-1 с-1 в сравнении с ~ 2×105 M-1 с-1) и немного более низкой константой скорости диссоциации (koff ~ 2×10-3 с-1 в сравнении с ~ 6×10-3 с-1). Можно рассуждать о том, что взаимодействие с более высокой аффинностью соответствует связыванию Fab1, который имел бы беспрепятственный доступ к изогнутой молекуле IgE-Fc, в то время как более низкая аффинность и более низкая скорость ассоциации характерны для Fab2, однако авторы изобретения не могут это утверждать с уверенностью.
Дополнительные ППР эксперименты для изучения механизма ингибирования связывания IgE-Fc с FcεRIα за счет Fab3 омализумаба продемонстрировали уменьшение числа доступных сайтов для Fab3 омализумаба на IgE-Fc (уменьшение Bmax), находящемся в комплексе с FcεRIα. Ингибирование связывания IgE с FcεRI омализумабом часто интерпретировалось как прямая конкуренция за перекрывающиеся сайты, однако были сообщения о том, что омализумаб может связываться со связанным с рецептором IgE21,32. Авторы изобретения напрямую продемонстрировали способность Fab3 омализумаба к связыванию IgE-Fc, который уже связан с FcεRIα, с образованием тримолекулярного комплекса. Эффект предварительного связывания IgE-Fc с FcεRIα заключается в уменьшении числа сайтов связывания Fab3 омализумаба на IgE-Fc до уровня менее 10% от числа сайтов, доступных в свободном IgE-Fc; такой эффект может являться следствием только аллостерической модуляции.
Характер взаимодействия Fab3 омализумаба с комплексом IgE-Fc/FcεRI дает представление о механизме ускоренной диссоциации. Впервые об этом явлении сообщалось для DARPin, а затем для омализумаба20,21, в случае последнего при значительно более высоких концентрациях, чем те, которые используют в терапии22, и теперь в настоящем документе это описано для Fab-фрагментов омализумаба. Авторы изобретения также продемонстрировали, что диссоциация происходит только после первого связывания второй (с меньшей аффинностью) молекулы Fab3 омализумаба. Иными словами, должен быть образован тетрамолекулярный комплекс - (Fab3 омализумаба)2/IgE-Fc/FcεRIα - чтобы происходило значительное ускорение диссоциации.
На основании наблюдений авторов изобретения для Fab3 омализумаба, IgE-Fc и sFcεRIα, они предполагают следующий механизм взаимодействия омализумаба, IgE и FcεRI: IgE связывается с FcεRI и, в этих условиях, небольшая популяция таких связанных молекул IgE принимают конформацию, с которой могут связываться молекулы омализумаба; когда связывается вторая молекула омализумаба, с образованием тетрамерного комплекса, энергетическое окружение IgE меняется так, что взаимодействие с FcεRI дестабилизируется и происходит быстрая диссоциация IgE от FcεRI. Ключ к пониманию данного механизма заключается в признании сложности энергетического окружения для IgE и того, что разные конформационные состояния существуют в динамическом равновесии.
Ингибирующая активность омализумаба, судя по всему, связана с характерной гибкостью IgE и, по меньшей мере в случае процесса ускоренной диссоциации, с динамикой комплекса IgE/FcεRI. IgE имеет ряд необычных структурных характеристик в сравнении с антителами других изотипов, включая наличие Cε2-доменов и уникально конформационно-динамический, подобный расплавленной глобуле, характер Cε3-доменов34. В совокупности, эти свойства создают путь аллостерического взаимодействия, который предотвращает одновременное связывание CD23 и FcεRI; это важно для избегания независимой от аллергена активации тучных клеток в результате перекрестного связывания FcεRI-связанного IgE тримерной молекулой CD2312. Другие функциональные преимущества, связанные с динамикой IgE, были предложены для связанного с мембраной B-клеточного рецептора IgE16. То наблюдение, что омализумаб не использует ожидаемый ортостерический механизм для ингибирования взаимодействия IgE/FcεRI, указывает на то, что он использует необычные динамические свойства IgE, как в качестве блокирующего антитела, так и с точки зрения его способности избегать перекрестного связывания с IgE, связанным с тучными клетками. И наконец, омализумаб может активно вызывать диссоциацию IgE от FcεRI, хотя и при концентрациях, более высоких, чем терапевтические21, за счет аллостерии и присущей IgE гибкости, которая сохраняется у него даже в комплексе с его рецепторами.
Методы
Клонирование, экспрессия и очистка белка. Fab человеческого IgG1 омализумаба и Fab3 омализумаба клонировали, экспрессировали и очищали, как описано в16. IgE-Fc получали, как описано ранее35. IgE-Fc имеет SEQ ID NO:108 (V224-K547 в соответствии с Dorrington & Bennich (1978) Immunol. Rev. 41:3-25, но со следующими мутациями, внесенными в IgE-Fc для упрощения картины гликозилирования: N265Q и N371Q). Омализумаб приобретали у компании Novartis Europharm Limited. Комплекс 2:1 Fab3 омализумаба/IgE-Fc очищали методом эксклюзионной хроматографии, элюировали в 25 мМ Tris-HCl, pH 7,5, 20 мМ NaCl и 0,05% (масс/об) NaN3, и концентрировали до 23 мг/мл.
Поверхностный плазмонный резонанс. Эксперименты методом ППР проводили на приборе Biacore T200 (GE Healthcare). Специфические поверхности готовили либо путем ковалентной сшивки белков с использованием протокола связывания через аминогруппы (GE Healthcare), с плотностью связывания <300 единиц ответа, либо путем захвата His-маркированных белков с использованием анти-His поверхности сенсора. Для захвата His-маркированных лигандов использовали моноклональное антитело против His-маркера, которое иммобилизовали в соответствии с инструкциями производителя (Biacore His Capture Kit, GE Healthcare). В экспериментах по связыванию, как правило, использовали время ассоциации 180-240 с, и диссоциацию компонентов контролировали в течение по меньшей мере 500 с. Инъекции выполняли при скорости потока 25 мкл мин-1 в подвижном буфере 20 мМ HEPES, pH 7,4, 150 мМ NaCl и 0,005% (по объему) сурфактанта P-20 (GE Healthcare). Большинство экспериментальных измерений проводили при 25°C; некоторые эксперименты по связыванию в формате сэндвич-анализа проводили при 5°C для сведения к минимуму явления ускоренной диссоциации. Использовали стандартные методы двойного вычитания референсных данных36, и кинетические кривые строили с использованием программы Origin (OriginLab).
TR-FRET. IgE-Fc метили донором флуорофора, проводя реакцию белка в концентрации 4 мг/мл в растворе 100 мМ натрия бикарбоната, 50 мМ NaCl, pH 9,3, с 5-кратным молярным избытком изотиоцианата хелата тербия (Invitrogen). Через 3 часа инкубации при комнатной температуре с перемешиванием избыток непрореагировавшего флуорофора удаляли диализом в PBS (20 мМ фосфатно-солевой буферный раствор, 150 мМ NaCl, pH 7,4). Слитый белок sFcεRIα-IgG4-Fc (α-γ)37 метили акцептором флуорофора, проводя реакцию белка в концентрации 3 мг/мл с 2,5-кратным молярным избытком сукцинимидил эфира Alexa Fluor 647 (Invitrogen) в течение 1 часа при комнатной температуре. Избыток флуорофора удаляли диализом в PBS.
TR-FRET анализы конкурентного ингибирования проводили, используя 1 нМ меченый тербием IgE-Fc и 0-20 нМ меченый Alexa Fluor 647 sFcεRIα-IgG4-Fc с Fab3 омализумаба в диапазоне концентраций. Анализы проводили в 384-луночных белых планшетах HiBase (Greiner BioOne) с использованием буфера Lanthascreen (Invitrogen) в качестве разбавителя. Планшет оставляли для инкубации на ночь при комнатной температуре и снимали показания на планшетном ридере Artemis (Berthold Technologies). Затем рассчитывали отношения TR-FRET для каждой лунки в виде частного от деления эмиссии акцептора при 665 нм на эмиссию донора при 620 нм, умноженного на 10000.
Кристаллизация. Кристаллы с прямоугольной морфологией, длиной до 400 мкм, выращивали при 18°C с использованием метода диффузии паров в сидячей капле. Резервуар содержал 50 мкл 4% (масс/об) ПЭГ 8000 и 0,03 M фторида натрия, и капля содержала 100 нл белка и 300 нл резервуара. Несмотря на значительные усилия по оптимизации, качество дифракции кристаллов не могло быть усовершенствовано более того, что было достигнуто в данном исследовании. Кристаллы, как правило, начинали расти через несколько дней, и часто растворялись в их каплях, однако могли быть стабилизированы в 4 M TMAO (триметиламин N-оксид), который был успешно использован в качестве криопротектора.
Сбор и обработка данных. Данные собирали с использованием пучков синхронизированного излучения I02 и I03 на источнике света Diamond (Harwell, UK). Интегрирование выполняли с использованием опции XDS38 в экспертной системе xia239. Для кристаллов была характерна анизотропная дифракция, и данные от нескольких кристаллов объединяли. Данные масштабировали до разрешения 3,7 Å с использованием AIMLESS из набора CCP440,41, а затем урезали до пределов разрешения 3,7 Å (a*), 3,9 Å (b*) и 4,2 Å (c*) с использованием сервера анизотропной дифракции UCLA42. Расчет коэффициента Мэтьюса показал, что содержание растворителя составляет ~62%, для одиночного комплекса 2:1 Fab3 омализумаба/IgE-Fc (молекулярная масса ~170 кДа) в асимметричной единице.
Определение структуры, построение и уточнение модели. Структура была решена путем молекулярной замены с помощью программ PHASER43 и MOLREP44 из пакета CCP440, с использованием атомов белка из PDB записи 2wqr8 и структуры Fab омализумаба с разрешением 1,9 Å (неопубликованные результаты) в качестве поисковых моделей. Уточнение первоначально выполняли с использованием REFMAC45, а затем PHENIX46, и чередовали с построением модели вручную в Coot47. Качество модели оценивали с помощью MolProbity48, POLYGON49 и других инструментов для валидации в графическом интерфейсе PHENIX50. Статистика обработки и уточнения данных представлена в Таблице 1. Область карты электронной плотности приведена на Фиг. 11. Поверхности контакта анализировали с использованием PISA51, и фигуры создавали при помощи PyMOL52.
Таблица 1. Статистика обработки и уточнения данных
a Значения в скобках относятся к слою наивысшего разрешения
bУточнение проводили с данными, урезанными до пределов разрешения 3,7 Å (a*), 3,9 Å (b*) и 4,2 Å (c*)
cRfree набор включает 5% отражений.
Список литературы
1. Gould, H.J., Sutton, B.J. IgE in allergy and asthma today. Nat. Rev. Immunol. 8,205-217 (2008).
2. Sutton, B.J., Davies, A.M. Structure and dynamics of IgE-receptor interactions: FcεRI and CD23/FcεRII. Immunol. Rev. 268,222-235 (2015).
3. Holgate, S.T. New strategies with anti-IgE in allergic diseases. World Allergy Organ. J. 7,17 (2014).
4. Holowka, D., Baird, B. Structural studies on the membrane-bound immunoglobulin E (IgE)-receptor complex. 2. Mapping of distances between sites on IgE and the membrane surface. Biochemistry 22,3475-3484 (1983).
5. Zheng, Y., Shopes, B., Holowka, D., Baird, B. Conformations of IgE bound to its receptor Fc epsilon RI and in solution. Biochemistry 30,9125-9132 (1991).
6. Beavil, A.J., Young, R.J., Sutton, B.J., Perkins, S.J. Bent domain structure of recombinant human IgE-Fc in solution by X-ray and neutron scattering in conjunction with an automated curve fitting procedure. Biochemistry 34,1449-1461 (1995).
7. Wan, T. et al. The crystal structure of IgE Fc reveals an asymmetrically bent conformation. Nat. Immunol. 3,681-686 (2002).
8. Holdom, M.D. et al., Conformational changes in IgE contribute to its uniquely slow dissociation rate from receptor FcεRI. Nat. Struct. Mol. Biol. 18, 571-576 (2011).
9. Hunt, J. et al. A fluorescent biosensor reveals conformational changes in human immunoglobulin E Fc: implications for mechanisms of receptor binding, inhibition, and allergen recognition. J. Biol. Chem. 287, 17459-17470 (2012).
10. Garman, S.C., Wurzburg, B.A., Tarchevskaya, S.S., Kinet, J.P., Jardetzky, T.S. Structure of the Fc fragment of human IgE bound to its high-affinity receptor FcεRIα. Nature 406,259-266 (2000).
11. Dhaliwal, B. et al. Crystal structure of IgE bound to its B-cell receptor CD23 reveals a mechanism of reciprocal allosteric inhibition with high affinity receptor FcεRI. Proc. Natl. Acad. Sci USA 109, 12686-12691 (2012).
12. Borthakur, S. et al. Mapping of the CD23 binding site on immunoglobulin E (IgE) and allosteric control of the IgE-FcRI interaction. J. Biol. Chem. 287,31457-31461 (2012).
13. Yuan, D. et al. Ca2+-dependent structural changes in the B-cell receptor CD23 increase its affinity for human immunoglobulin E. J. Biol. Chem. 288,21667-21677 (2013).
14. Dhaliwal, B., Pang, M.O., Yuan, D., Beavil, A.J., Sutton, B.J. A range of Cε3-Cε4 interdomain angles in IgE Fc accommodate binding to its receptor CD23. Acta Crystallogr. F70,305-309 (2014).
15. Cooper, A., Dryden, D.T. Allostery without conformational change. A plausible model. Eur. Biophys. J. 11,103-109 (1984).
16. Drinkwater, N. et al. Human immunoglobulin E flexes between acutely bent and extended conformations. Nat. Struct. Mol. Biol. 21, 397-404 (2014).
17. Holgate, S., Casale, T., Wenzel, S., Bousquet, J., Deniz, Y., Reisner, C. The anti-inflammatory effects of omalizumab confirm the central role of IgE in allergic inflammation. J. Allergy Clin. Immunol. 115,459-465 (2005).
18. Zheng, L. et al. Fine epitope mapping of humanized anti-IgE monoclonal antibody omalizumab. Biochem. Biophys. Res. Comm. 375,619-622 (2008).
19. Wright, J.D. et al. Structural and Physical Basis for Anti-IgE Therapy. Sci. Rep. 5,11581 (2015).
20. Kim, B. et al. Accelerated disassembly of IgE-receptor complexes by a disruptive macromolecular inhibitor. Nature 491,613-617 (2012).
21. Eggel, A. et al. Accelerated dissociation of IgE-FcεRI complexes by disruptive inhibitors actively desensitizes allergic effector cells. J. Allergy Clin. Immunol. 133,1709-1719 (2014).
22. Lowe, P.J., Tannenbaum, S., Gautier, A., Jimenez, P. Relationship between omalizumab pharmacokinetics, IgE pharmacodynamics and symptoms in patients with severe persistent allergic (IgE-mediated) asthma. Br. J. Clin. Pharmacol. 68,61-76 (2009).
23. Jensen, R.K. et al. Structure of the omalizumab Fab. Acta Crystallogr. F71,419-426 (2015).
24. Cohen, E.S. et al. A novel IgE-neutralizing antibody for the treatment of severe uncontrolled asthma. mAbs 6,755-763 (2014).
25. Zheng, Y., Shopes, B., Holowka, D., Baird, B. Dynamic conformations compared for IgE and IgG1 in solution and bound to receptors. Biochemistry 31,7446-7456 (1992).
26. Holowka, D., Baird, B. Structural studies on the membrane-bound immunoglobulin E-receptor complex. 2. Mapping of distances between sites on IgE and the membrane-surface. Biochemistry 22, 3475-3484 (1983).
27. Holowka, D., Conrad, D.H., Baird, B. Structural mapping of membrane-bound immunoglobulin-E receptor complexes: use of monoclonal anti-IgE antibodies to probe the conformation of receptor-bound IgE. Biochemistry 24, 6260-6267 (1985).
28. Davis, K.G., Glennie, M., Harding, S.E. & Burton, D.R. A model for the solution conformation of rat IgE. Biochem. Soc. Trans. 18, 935-936 (1990).
29. Wurzburg, B.A., Jardetsky, T.S. Conformational Flexibility in Immunoglobulin E-Fc3-4 Revealed in Multiple Crystal Forms. J. Mol. Biol. 393, 176-190 (2009).
30. Shiung, L.L. et al. An anti-IgE monoclonal antibody that binds to IgE on CD23 but not on high-affinity IgE.Fc receptors. Immunobiology 217, 676-683 (2012).
31. Borthakur, S., Andrejeva, G., McDonnell, J.M. Basis of the intrinsic flexibility of the Cε3 domain of IgE. Biochemistry 50,4608-4614 (2011).
32. Serrano-Candelas, E. et al. Comparable actions of omalizumab on mast cells and basophils. Clin. Exp. Allergy. 46,92-102(2016).
33. Fersht, A. In Structure and Mechanism in Protein Science. W. H. Freeman, New York, pp. 103-131 (1999).
34. Price, N.E., Price, N.C., Kelly, S.M., McDonnell, J.M. The key role of protein flexibility in modulating IgE interactions. J. Biol. Chem. 280,2324-2330 (2005).
35. Young, R.J. et al. Secretion of recombinant human IgE-Fc by mammalian cells and biological activity of glycosylation site mutants. Protein Eng. 8,193-199 (1995).
36. Myszyka, D.G. Improving biosensor analysis. Journal of Molecular Recognition 12, 279-284 (1999).
37. Shi, J. et al. Interaction of the Low-Affinity Receptor CD23/FcεRII Lectin Domain with the Fcε3-4 Fragment of Human Immunoglobulin E. Biochemistry 36,2112-2122 (1997).
38. Kabsch, W. XDS. Acta Crystallogr. D66,125-132 (2010).
39. Winter, G. xia2: an expert system for macromolecular crystallography data reduction. J. Appl. Cryst. 43, 186-190 (2010).
40. Winn, M.D. et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D67, 235-242 (2011).
41. Evans, P.R., Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. D69,1204-1214 (2013).
42. Strong, M., Sawaya, M.R., Wang, S., Phillips, M., Cascio, D., Eisenberg, D. Toward the structural genomics of complexes: Crystal structure of a PE/PPE protein complex from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA. 103,8060-8065 (2006).
43. McCoy, A.J. et al., Phaser crystallographic software. J. Appl. Cryst. 40, 658-674 (2007).
44. Vagin, A., Teplyakov, A. Molecular replacement with MOLREP. Acta. Cryst. D66, 22-25 (2010).
45. Murshudov, G.N. et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D67, 355-367 (2011).
46. P.V. Afonine et al., Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D68, 352-367 (2012).
47. Emsley, P., Lohkamp, B., Scott, W.G., Cowtan, K. Features and Development of Coot. Acta Crystallogr. D66, 486-501 (2010).
48. Chen, V.B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D66, 12-21 (2010).
49. Urhumsteva, L., Afonine, P.V., Adams, P.D., Urhumstev, A. Crystallographic model quality at a glance. Acta Crystallogr. D65, 297-300 (2009).
50. Echols, N. et al. Graphical tools for macromolecular crystallography in PHENIX. J. Appl. Cryst. 45, 581-586 (2012).
51. Krissinel, E., Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 372,774-797 (2007).
52. The PyMOL Molecular Graphics System, Version 1.1r1 Schrödinger, LLC.
Пример 2: Измерение ускоренной диссоциации IgE-Fc от иммобилизованного sFcεRIα с использованием Biacore
Технология Biacore позволяет измерять взаимодействие между биологическими молекулами без необходимости в мечении. Один из взаимодействующих компонентов, называемый лигандом, либо напрямую иммобилизован, либо захвачен на поверхности сенсора, в то время как другой, называемый аналитом, протекает в растворе над удерживающей поверхностью. Сенсор определяет изменение массы на поверхности сенсора, когда аналит связывается с лигандом и когда аналит диссоциирует от лиганда. Это соответствует процессам ассоциации и диссоциации. В анализе ускоренной диссоциации sFcεRIα является лигандом и его иммобилизуют на поверхности сенсора. IgE-Fc является аналитом и захватывается sFcεRIα. Диссоциацию IgE-Fc от sFcεRIα контролируют либо с протеканием буфера над поверхностью сенсора, либо с протеканием раствора связывающего партнера IgE над поверхностью сенсора. Подробная информация о методе:
Прибор: Biacore 3000, GE Healthcare AB, Uppsala, Sweden
Сенсорный чип: CM5. Каталожный номер BR100399
Раствор BIAnormalising: 70% (по массе) раствор глицерина. Часть набора BIAmaintenance. Каталожный номер BR100651. Набор BIAmaintenance хранили при 4°C.
Набор для связывания через аминогруппы: Каталожный номер BR100633. Этил-3-(3-диметиламинопропил)карбодиимид гидрохлорид (EDC) доводили до концентрации 75 мг/мл в дистиллированной воде и хранили в 200-мкл аликвотах при -70°C. N-гидроксисукцинимид (NHS) доводили до концентрации 11,5 мг/мл в дистиллированной воде и хранили в 200-мкл аликвотах при -70°C. Этаноламин гидрохлорид-NaOH, pH 8,5, хранили при 4°C.
Реагент для окисления sFcεRIα. Карбогидразид (SigmaAldrich, каталожный номер C11006) доводили до концентрации 5 мМ в дистиллированной воде. Цианоборогидрид натрия (SigmaAldrich, каталожный номер 156159) доводили до концентрации 100 мМ в ацетате натрия (BDH, кат. № S1104-500GM), 100 мМ, pH=4. m-периодат натрия (SigmaAldrich, каталожный номер S-1878) доводили до концентрации 50 мМ в ацетате натрия (BDH, кат. № S1104-500GM), 100 мМ, pH=5,5.
sFcεRIα разбавляли до концентрации 1 мг/мл в 0,1 M растворе ацетата натрия, pH 5,5. Затем 4 мкл периодата натрия (50 мМ, разведенного 1/50) добавляли к 200 мкл 1 мг/мл раствора sFcεRIα. Смесь оставляли на льду на 20 мин. Перед иммобилизацией раствор sFcεRIα разбавляли до концентрации 7 мкг/мл 10 мМ раствором ацетата натрия (GE Healthcare, каталожный номер BR100669), pH=4,0.
Буферы: Подвижный буфер представляет собой HBS-EP (а именно, 10 мМ HEPES, pH 7,4, 150 мМ NaCl, 3 мМ ЭДТА, 0,05% сурфактанта P20, восстановленный из 10x маточного раствора): каталожный номер BR100669. Буфер иммобилизации представляет собой ацетат 4,0 (а именно, 10 мМ раствор ацетата натрия, pH 4,0). Каталожный номер BR100349. Буфер хранили при 4°C.
Лиганд: sFcεRIα, представляющий собой внеклеточный фрагмент альфа-цепи человеческого высокоаффинного рецептора IgE. Экспрессирован в виде рекомбинантного белка в клетках CHO и очищен.
Аналит: IgE-Fc, представляющий собой Fc-фрагмент IgE человека, экспрессированный в виде рекомбинантного белка в клетках CHO и очищенный. Использовали человеческий IgE-Fc дикого типа (домены Cε2-Cε4 с нумерацией V224-K547 согласно Dorrington & Bennich (1978) Immunol. Rev. 41:3-25), несущий мутацию C225A (SEQ ID NO:108).
Номенклатура мутантов:
- Fab1 омализумаба: S81R, Q83R [S77R, Q79R в соответствии с нумерацией Kabat] (вариабельная область легкой цепи+константная область каппа, SEQ ID NO:24, где S77 и S79 заменены на Q; вариабельная область тяжелой цепи+CH1 константной области, SEQ ID NO:5)
- Fab2 омализумаба: L158P [L154P в соответствии с нумерацией Kabat] (вариабельная область легкой цепи+константная область каппа, SEQ ID NO:113; вариабельная область тяжелой цепи+CH1 константной области, SEQ ID NO:5)
- Fab3 омализумаба: S81R, Q83R, L158P [S77R, Q79R, L154P в соответствии с нумерацией Kabat] (вариабельная область легкой цепи, SEQ ID NO:121; вариабельная область тяжелой цепи+CH1 константной области, SEQ ID NO:5).
Связывающие партнеры IgE (1): полноразмерный омализумаб (Novartis); рекомбинантный Fab-фрагмент омализумаба, экспрессированный в клетках CHO и очищенный.
Связывающие партнеры IgE (2): рекомбинантный Fab-фрагмент омализумаба, с его мутациями, экспрессированный в клетках HEK-293 и анализируемый в виде культурального супернатанта. Культуральный супернатант концентрировали в десять раз перед анализом.
Метод анализа (1): sFcεRIα связывали с поверхностью сенсора методом связывания через альдегидные группы до уровня ~500 единиц ответа (ЕО). HBS-EP буфер использовали в качестве подвижного буфера при скорости потока 30 мкл/мин. IgE-Fc разбавляли до концентрации 10 нМ в HBS-EP и инжектировали над иммобилизованным sFcεRIα в течение 290 секунд, с последующими 3 инжекциями, каждая продолжительностью 690 секунд, подвижного буфера или связывающего партнера IgE, разведенного в подвижном буфере. Уровень захвата IgE-Fc составлял ~90 ЕО. Поверхность регенерировали двумя 60-секундными инжекциями 10 мМ раствора глицина-HCl, pH 2,5. Количество IgE-Fc, диссоциировавшего от иммобилизованного sFcεRIα, рассчитывали, как функцию от исходно связавшегося количества, и скорость диссоциации рассчитывали, как количество IgE-Fc, отделенного от иммобилизованного sFcεRIα, нормированное на исходно связавшееся количество, в зависимости от затраченного времени.
Метод анализа (2): sFcεRIα связывали с поверхностью сенсора методом связывания через альдегидные группы до уровня ~2000 единиц ответа (ЕО). HBS-EP буфер использовали в качестве подвижного буфера при скорости потока 30 мкл/мин. IgE-Fc разбавляли до концентрации 10 нМ в HBS-EP и инжектировали над иммобилизованным sFcεRIα в течение 180 секунд, с последующей 1 инжекцией в течение 180 секунд подвижного буфера или связывающего партнера IgE, разведенного в подвижном буфере. Уровень захвата IgE-Fc составлял ~275 ЕО. Поверхность регенерировали двумя 60-секундными инжекциями 10 мМ раствора глицина-HCl, pH 2,5. Количество IgE-Fc, диссоциировавшего от иммобилизованного sFcεRIα, рассчитывали, как функцию от исходно связавшегося количества, и скорость диссоциации рассчитывали, как количество IgE-Fc, отделенного от иммобилизованного sFcεRIα, нормированное на исходно связавшееся количество, в зависимости от затраченного времени.
буфер
Fab1 омализумаба 100 мкг/мл
Fab2 омализумаба 100 мкг/мл
Fab3 омализумаба 100 мкг/мл
100,0
100,0
100,0
100,0
87,6
61,3
79,4
45,3
81,9
51,0
71,5
34,6
77,9
47,3
69,5
30,8
1,02E-04
3,06E-04
1,48E-04
4,80E-04
Таблица 2: Расчет количества IgE-Fc, диссоциировавшего от иммобилизованного sFcεRIα. Начальное связывание IgE с sFcεRIα принимали за 100%, и диссоциацию рассчитывали относительно этого. Кажущаяся скорость диссоциации основана на предполагаемой односкоростной модели.
Проводили дальнейший мутагенез для определения того, могут ли быть идентифицированы дополнительные мутации, приводящие к ускорению диссоциации IgE-Fc от sFcεRIα, помимо тех, которые описаны для Fab3. В результате была идентифицирована мутация S64M (со ссылкой на SEQ ID NO:20), которая в контексте Fab3 была способна приводить к дальнейшему увеличению диссоциации IgE от sFcεRIα. Эти данные приведены на Фигуре 13 и в Таблице 3.
Таблица 3: Расчет количества IgE-Fc, диссоциировавшего от иммобилизованного sFcεRIα. Начальное связывание IgE с sFcεRIα принимали за 100%, и диссоциацию рассчитывали относительно этого. Кажущаяся скорость диссоциации основана на предполагаемой односкоростной модели. Mutant S64M: легкая цепь SEQ ID NO:39; Fab омализумаба ДТ: легкая цепь SEQ ID NO:20; Fab3 омализумаба: легкая цепь SEQ ID NO:125. Во всех трех тяжелая цепь SEQ ID NO:5.
Вывод:
В совокупности, полученные данные показывают, что мутантная форма Fab омализумаба может ускорять диссоциацию IgE от иммобилизованной формы высокоаффинного рецептора IgE, FcεRI. Мутации в легкой цепи, делающие это возможным, включают, но не обязательно ограничиваются ими, S64M, S81R, Q83R и L158P со ссылкой на SEQ ID NO:24, и результатом является SEQ ID NO:39.
Пример 3: Измерение ускоренной диссоциации IgE-Fc от FcεRI методом проточной цитометрии
Прибор: проточный цитометр FACSCanto II (Becton Dickinson)
Линия клеток: клетки RBL-SX38, экспрессирующие человеческий FcεRI, культивировали в минимальной поддерживающей среде с добавлением 10% эмбриональной бычьей сыворотки, 2 мМ GlutaMAX и 500 мкг/мл генетицина (Life Technologies). Во время анализа клетки промывали в PBS и инкубировали в Accutase до открепления, затем ресуспендировали при плотности 1×106 клеток/мл в культуральной среде. Все последующие этапы инкубации проводили в культуральной среде.
Метод анализа: клетки RBL-SX38 при плотности 1×106 клеток/мл инкубировали с 5 нМ раствором меченого Alexa 488 IgE-Fc в течение 1 часа при 37°C. Клетки дважды промывали в культуральной среде для удаления несвязанного IgE-488, затем суспендировали при плотности 1×106 клеток/мл в культуральной среде, или IgE-связывающие фрагменты разводили в культуральной среде в концентрации 100 мкг/мл. Затем клетки инкубировали при 37°C с постоянным перемешиванием. В каждой временной точке 0,5×105 клеток отбирали, промывали в ледяном PBS, а затем фиксировали, ресуспендируя в 1% растворе параформальдегида в PBS в течение 16 часов при 4°C. Количество флуоресценции связанного с клетками Alexa 488 определяли с использованием проточного цитометра FACSCanto II.
Проточная цитометрия: фиксированные клетки дважды промывали в FACS буфере (0,1% масс/об БСА, 0,01% масс/об NaN3 в PBS, pH 7,4) и ресуспендировали в 200 мкл FACS буфера. Проточную цитометрию проводили на цитометре FACSCanto II стандартными методами и рассчитывали среднее геометрическое интенсивности флуоресценции Alexa 488, связанного с интактными клетками, используя программу FlowJo. Скорость диссоциации меченого Alexa 488 IgE-Fc рассчитывали, как изменение среднего геометрического значения интенсивности флуоресценции с течением времени, в результате инкубации клеток либо в культуральной среде, избытке немеченого IgE-Fc, либо в присутствии IgE-связывающих фрагментов.
Номенклатура мутантов:
- Fab1 омализумаба: S81R, Q83R [S77R, Q79R в соответствии с нумерацией Kabat] (вариабельная область легкой цепи+константная область каппа, SEQ ID NO:24, в которой S77 и S79 заменены на Q; вариабельная область тяжелой цепи+CH1 константной области, SEQ ID NO:5)
- Fab2 омализумаба: L158P [L154P в соответствии с нумерацией Kabat] (вариабельная область легкой цепи+константная область каппа, SEQ ID NO:113; вариабельная область тяжелой цепи+CH1 константной области, SEQ ID NO:5)
- Fab3 омализумаба: S81R, Q83R, L158P [S77R, Q79R, L154P в соответствии с нумерацией Kabat] (вариабельная область легкой цепи+константная область каппа, SEQ ID NO:121; вариабельная область тяжелой цепи+CH1 константной области, SEQ ID NO:5).
Таблица 4: Рассчитанная скорость диссоциации меченого Alexa 488 IgE-Fc от поверхности клеток RBL-SX38. Кажущуюся скорость диссоциации определяли методом линейной регрессии на основании графика зависимости -ln(R1/R0) от времени для всех доступных временных точек.
Вывод:
В совокупности, полученные данные показывают, что мутантная форма Fab омализумаба может ускорять диссоциацию IgE от высокоаффинного рецептора IgE, FcεRI, экспрессированного на клеточной поверхности. Мутации в легкой цепи, делающие это возможным, включают, но не обязательно ограничиваются ими, S81R, Q83R и L158P.
Пример 4: Демонстрация того, что Fab3 омализумаба обладает более высокой эффективностью, чем Fab омализумаба дикого типа, при терапевтическом введении в 72-часовой модели ПКА.
Готовили 25 мкг/мл раствор человеческого анти-DNP IgE, добавляя 7,5 мкл маточного раствора в концентрации 6,68 мг/мл к 1992,5 мкл PBS. Инъекция 20 мкл этого раствора соответствовала бы дозе 500 нг IgE. Животным (Tg мыши hIgER) брили бока, а затем выполняли и/д инъекцию в каждый бок в 2 часа дня в день 0. 20 мкл PBS вводили инъекцией в левый бок каждого животного в качестве отрицательного контроля. Анти-DNP IgE (20 мкл) инъецировали в правый бок. В общей сложности инъекции выполняли 40 мышам. Введение либо Fab омализумаба дикого типа, либо Fab3 омализумаба (с мутациями S81R, Q83R, L158P [S77R, Q79R, L154P в соответствии с нумерацией Kabat]) начинали через 18 часов после введения IgE (8 часов утра). Мыши в двух группах (n=8/группу) получали дозу 100 мг/кг п/к либо Fab омализумаба дикого типа, либо Fab3 омализумаба. 8 мышей в другой группе получали п/к PBS. Мышам вновь вводили указанные дозы через 10 часов (6 часов вечера). В этой временной точке, через 28 часов после введения IgE, животным еще в двух группах (n=8/группу) п/к вводили либо Fab омализумаба дикого типа, либо Fab3 омализумаба, в дозе 100 мг/кг. Животным во всех группах повторно вводили дозы в 8 часов утра, 6 часов вечера и вновь в 8 часов утра в завершающий день эксперимента. Через 72 часа после и/д дозирования (2 часа дня) всем животным вводили в/в инъекцией 100 мкл раствора 1 мг/мл DNP-HSA, 2,5% масс/об красителя Evans blue, приготовленного в 100 МЕ/мл гепарина. Через 1 час животных умерщвляли в соответствии со схемой 1 метода. Кожу с боков вокруг зоны и/д инъекции удаляли и проводили пункционную биопсию. Образцы кожи помещали в 700 мкл формамида и проводили расщепление в течение ночи при 55°C. После расщепления 100 мкл x 2 жидкости из каждого образца отбирали и помещали в 96-луночный планшет для ELISA. Поглощение измеряли при 620 нм.
Вывод:
Полученные данные показывают, что мутантная форма Fab омализумаба, которая способна ускорять диссоциацию IgE от высокоаффинного рецептора IgE, FcεRI, также способна уменьшать пассивную кожную анафилаксию (о чем свидетельствует ингибирование утечки красителя Evans blue из зоны реакции) в статистически значимой степени при сравнении с Fab омализумаба дикого типа. Мутации в легкой цепи, делающие это возможным, включают, но не обязательно ограничиваются ими, S81R, Q83R и L158P со ссылкой на SEQ ID NO:24, и результатом является SEQ ID NO:39.
Пример 5: Моделирование молекулярной динамики Fab3 омализумаба в комплексе с IgE-Fc
Метод:
Кристаллическую структуру Fab3 омализумаба в комплексе с Fc-областью IgE (смотри Пример 1) получали с использованием программы Molecular Operating Environment (MOE) 2014.0901 (1), возмещая недостающие боковые цепи на некоторых остатках и недостающие петли между Cε2- и Cε3-доменами до проведения моделирования молекулярной динамики (МД) с использованием Amber 14 (2). Структуру комплекса дополняли водородом и проводили сольватацию с использованием явной водной модели TIP3P с 0,15 M растворенной солью NaCl в усеченном октаэдрическом боксе, в котором расстояние от каждой грани бокса до атомов белка составляло 10 Å. Систему настраивали с использованием силовых полей Amber ff12SB и олигосахаридов GLYCAM_06j-1 (3) и минимизировали путем сопряжения градиентного алгоритма для 50000 шагов с порогом отсечения 10,0 Å, установленным для кулоновских и ван-дер-ваальсовых взаимодействий, и основанным на решетке списком соседей. Затем систему постепенно нагревали от 0 до 300 K за 125 пс при постоянном объеме, с последующим 2,25-нс уравновешиванием в NPT ансамбле с ограничениями на все растворенные тяжелые атомы (ограничение гармонической силы составляет 5,0). Для электростатики, авторы изобретения использовали PME четвертого порядка с порогом отсечения 8,0 Å для кулоновских взаимодействий с настройками расстояния Фурье и допусков по умолчанию. Температуру контролировали с алгоритмом слабых взаимодействий, примененным к белку и растворителю, соответственно, с константой времени 1,0 пс, и давление контролировали с помощью изотропного баростата Берендсена, примененного ко всей системе, с константой времени 1,0 пс и сжимаемостью 4,46×10-5 бар-1. И наконец, проводили 1000-нс моделирование создания без каких-либо ограничений с использованием тех же параметров, что и для уравновешивания. Чтобы сделать возможным 4-фс временной шаг на инфраструктуре GPU, массу атомов водорода белка и сахаров перераспределяли до 3,024 дальтон с использованием ParmEd (4), в то время как массу атомов, с которыми они связаны, корректировали на количество, необходимое, чтобы общая масса осталась неизменной. Структуру Fab омализумаба дикого типа в комплексе с Fc-областью IgE моделировали на основании кристаллической структуры комплекса мутанта3 омализумаба с IgE-Fc путем виртуальной мутации R81 и R83 в легкой цепи антитела на остатки дикого типа серин и аспарагин, соответственно. Моделирование МД проводили с использованием того же протокола настроек, что и для мутанта3.
Использовали модуль AmberTools cpptraj (7) для кластерного анализа траекторий МД как для Fab3 омализумаба, так и для Fab омализумаба, в комплексе с IgE-Fc. Использовали иерархический алгоритм агломерации со средним расстоянием связи между всеми кластерами менее 2,0 Å. Расстояние между каркасами рассчитывали с помощью СКО наиболее точно подобранных координат между Cα-атомами в остатках V-области легкой цепи антитела. Кластеризацию проводили лишь для каждого 10 каркаса, и все остальные каркасы добавляли к кластерам на основании того, насколько они близки к центроидам кластеров после кластеризации.
Список литературы
1. Molecular Operating Environment (MOE), 2014.09; Chemical Computing Group Inc., 1010 Sherbooke St. West, Suite #910, Montreal, QC, Canada, H3A 2R7, 2015.
2. Case, D.A., et al. AMBER 2014, 2014, University of California, San Francisco.
3. Kirschner, K. et al. GLYCAM06: A generalizable biomolecular force field. Carbohydrates. J. Comput. Chem., 2008, 29, 622-655.
4. http://parmed.github.io/ParmEd/html/index.html.
Результаты:
Для изучения того, каким образом мутации S81R и Q83R в легкой цепи Fab3 омализумаба влияют на взаимодействие с IgE, проводили моделирование молекулярной динамики с интервалами в одну микросекунду для структур Fab омализумаба и Fab3 омализумаба в комплексе с IgE-Fc, соответственно. Моментальные снимки траектории кластеризовали, и 2 структуры с наиболее заполненными кластерными центрами анализировали, при этом было отчетливо видно, что в обоих кластерах два мутантных остатка аргинина в мутанте3 образуют интенсивные основанные на водородных связях сети с остатками D278 и S280 в соседнем Cε2-домене IgE, соответственно. Визуальная инспекция траектории подтвердила, что парные взаимодействия R81-S280 и R83-D278 являются очень консервативными и стабильными во время моделирования, вследствие этого, разогнутые конформации Cε2 относительно более близкого Cε3-домена, как видно в кристаллической структуре, зафиксированы за счет образования мостов с антителом. Интересно, что, хотя двойные мутантные остатки аргинина пространственно расположены вблизи D278 и S280 Cε2 в кристаллической структуре Fab3 омализумаба, они не связаны напрямую водородными связями, что предполагает моделирование МД. В случае Fab омализумаба, как и ожидалось, сети водородных связей, наблюдаемые в случае мутанта3, отсутствуют в 2 топовых структурах кластерного центра. Визуальная инспекция траектории показывает, что Cε2-домены становятся менее привязанными к каркасу легкой цепи антитела и, таким образом, их положения относительно более близкого Cε3-домена становятся более вариабельными, чем в случае Fab3 омализумаба. В дополнение к этому, поверхность контакта между Fab3 омализумаба и IgE-Fc в их комплексе подвергали анализу IOTA (IOTA является инструментом оценки статистического потенциала для определения вероятности контакта конкретного вида атома на поверхности контакта белка или сайте связывания), который предсказал, что положения S56, S64 и S71 легкой цепи в Fab3 омализумаба (со ссылкой на SEQ ID NO:113) могут быть мутированы для повышения аффинности омализумаба для IgE-Fc и, следовательно, будут увеличивать эффект, предсказанный для мутаций S81R и Q38R, подробно описанный выше.
Вывод:
Подводя итоги, на основании результатов моделирования МД можно высказать гипотезу, что мутации S81R и Q38R способствуют локализации Cε2-доменов IgE на антителе за счет прямых электростатических взаимодействий и водородных связей с соседними остатками D278 и S280 в Cε2, и влияют на пластичность IgE-Fc путем запирания разогнутой конформации Cε2. Наряду с этим, статистические методы позволяют предсказать, что мутация S64M, а также S56 (изменение на D, E, Q или R) и S71 (изменение на D, E или M) будет иметь аналогичный эффект.
Пример 6: Измерение аффинности анти-IgE Fab для IgE-Fc с использованием Biacore.
Технология Biacore позволяет измерять взаимодействие между биологическими молекулами без необходимости в мечении. Один из взаимодействующих компонентов, называемый лигандом, либо напрямую иммобилизован, либо захвачен на поверхности сенсора, в то время как другой, называемый аналитом, протекает в растворе над удерживающей поверхностью. Сенсор определяет изменение массы на поверхности сенсора, когда аналит связывается с лигандом и когда аналит диссоциирует от лиганда. Это соответствует процессам ассоциации и диссоциации. В кинетическом анализе анти-IgE Fab является лигандом, захваченным на поверхности сенсора. IgE-Fc является аналитом и захватывается анти-IgE Fab. Ассоциацию IgE-Fc с захваченным анти-IgE Fab и диссоциацию от него контролируют либо с протеканием IgE-Fc над поверхностью сенсора (фаза ассоциации), либо с протеканием буфера над поверхностью сенсора (фаза диссоциации). Подробная информация о методе:
Прибор: Biacore 3000, GE Healthcare AB, Uppsala, Sweden
Сенсорный чип: CM5. Каталожный номер BR100399
Раствор BIAnormalising: 70% (по массе) раствор глицерина. Часть набора BIAmaintenance. Каталожный номер BR100651. Набор BIAmaintenance хранили при 4°C.
Набор для связывания через аминогруппы: Каталожный номер BR100633. Этил-3-(3-диметиламинопропил)карбодиимид гидрохлорид (EDC) доводили до концентрации 75 мг/мл в дистиллированной воде и хранили в 200-мкл аликвотах при -70°C. N-гидроксисукцинимид (NHS) доводили до концентрации 11,5 мг/мл в дистиллированной воде и хранили в 200-мкл аликвотах при -70°C. Этаноламин гидрохлорид-NaOH, pH 8,5, хранили при 4°C.
Буферы: Подвижный буфер представляет собой HBS-EP (а именно, 10 мМ HEPES, pH 7,4, 150 мМ NaCl, 3 мМ ЭДТА, 0,05% сурфактанта P20, восстановленный из 10x маточного раствора): каталожный номер BR100669. Буфер иммобилизации представляет собой ацетат 4,0 (а именно, 10 мМ раствор ацетата натрия, pH 4,0). Каталожный номер BR100349. Буфер хранили при 4°C.
Лиганд: лигандом были молекулы анти-IgE Fab. Они были временно экспрессированы в виде рекомбинантного белка в клетках HEK-293 и использованы без дальнейшей очистки.
Аналит: IgE-Fc, представляющий собой Fc-фрагмент IgE человека, экспрессированный в виде рекомбинантного белка в клетках CHO и очищенный. Использовали человеческий IgE-Fc дикого типа (домены Cε2-Cε4 с нумерацией V224-K547 согласно Dorrington & Bennich (1978) Immunol. Rev. 41:3-25), несущий мутацию C225A (SEQ ID NO:108).
Fab2-фрагмент антитела козы против человеческого IgG1 (специфический для Fab2-фрагмента) (Jackson Immunolabs, каталожный номер 109-006-097) иммобилизовали на поверхности сенсора стандартными методами связывания через аминогруппы. Анти-IgE Fab разбавляли в подвижном буфере (HBS-EP), и примерно 200 единиц ответа было захвачено на поверхности. IgE-Fc разбавляли в подвижном буфере, и серийно разбавленные растворы с концентрациями от 2 нМ до 125 пM пропускали над захваченными анти-IgE Fab. Фаза ассоциации продолжалась 180 секунд, и фаза диссоциации продолжалась 300 секунд. Поверхность сенсора регенерировали 60-секундным воздействием 40 мМ HCl, с последующим 60-секундным воздействием 10 мМ NaOH, а затем еще одним 60-секундным воздействием 40 мМ HCl. Все данные по связыванию обрабатывали с использованием двойных референсов стандартными методами при помощи программы BiaEvaluation.
Таблица 5: Аффинность Fab омализумаба и мутантов для IgE-Fc при измерении в кинетическом анализе Biacore. Константа диссоциации (KD) рассчитана как kd/ka.
Таблица 6: Аффинность Fab омализумаба (ДТ) и мутантов для IgE-Fc при измерении в кинетическом анализе Biacore. Константа диссоциации (KD) рассчитана как kd/ka. Четыре мутации находятся в исходной последовательности дикого типа (легкая цепь, SEQ ID NO:20).
Таблица 7a: Аффинность Fab омализумаба (ДТ) и мутантов для IgE-Fc при измерении в кинетическом анализе Biacore. Константа диссоциации (KD) рассчитана как kd/ka. Мутации указаны со ссылкой на легкой цепи, SEQ ID NO:20.
Таблица 7b: Аффинность Fab омализумаба для IgE-Fc при измерении в кинетическом анализе Biacore. Константа диссоциации (KD) рассчитана как kd/ka. Мутации находятся в легкой цепи (со ссылкой на SEQ ID NO:20). Последовательности вариабельной области легкой цепи мутантов Fab указаны в левой колонке (VL SEQ ID NO:).
(SEQ ID NO:132)
Таблица 7c: Аффинность мутанта Fab омализумаба для IgE-Fc при измерении в кинетическом анализе Biacore. Константа диссоциации (KD) рассчитана как kd/ka. Мутации находятся в легкой цепи (со ссылкой на SEQ ID NO:20). Проводили исследования образцов в условиях усиленного окисления, и уровни окисления метионина в каждой временной точке определяли методом масс-спектрометрии.
Вывод:
Полученные данные показывают, что мутации Fab омализумаба могут приводить к повышению аффинности Fab омализумаба для IgE-Fc. Наилучшим сочетанием мутаций с точки зрения повышения аффинности является сочетание S71M с S56D. Это повышение аффинности в основном вызвано уменьшением скорости диссоциации Fab от IgE-Fc. Аффинность Fab1 омализумаба (с мутациями S81R, Q83R [S77R, Q79R в соответствии с нумерацией Kabat]) и Fab3 (с мутациями S81R, Q83R, L158P [S77R, Q79R, L154P в соответствии с нумерацией Kabat] со ссылкой на SEQ ID NO:125) также повышена, что обусловлено усовершенствованным взаимодействием антитела с Cε2 IgE-Fc в сравнении с не мутантным Fab омализумаба.
Пример 7: Измерение ускоренной диссоциация IgE-Fc от иммобилизованного sFcεRIα с использованием Biacore.
Эффект анти-IgE Fab на диссоциацию IgE-Fc от sFcεRIα определяли методами, описанными в Примере 2 (метод анализа 2). Все анти-IgE Fab экспрессировали в клетках HEK-293, очищали стандартными методами и количественно определяли по поглощению при 280 нм с использованием расчетных коэффициентов молярной экстинкции. В данном анализе концентрация IgE-Fc составляла 2 нМ и время диссоциации составляло 200 секунд. Количество IgE-Fc, диссоциировавшего от иммобилизованного sFcεRIα, рассчитывали, как функцию от исходно связавшегося количества, и скорость диссоциации рассчитывали, как количество IgE-Fc, отделенного от иммобилизованного sFcεRIα, нормированное на исходно связавшееся количество, в зависимости от затраченного времени.
Считается, что S80N (нумерация pdb) взаимодействует с D278 в Cε2 IgE и S67W/Y (нумерация pdb) взаимодействует с T298 в Cε2 IgE.
Таблица 8: Расчет количества IgE-Fc, диссоциировавшего от иммобилизованного sFcεRIα. Первоначальное связывание IgE с sFcεRIα принимали за 100%, и диссоциацию рассчитывали относительно этого. Кажущаяся скорость диссоциации основана на предполагаемой односкоростной модели.
Вывод:
Полученные данные продемонстрировали, что мутантная форма Fab омализумаба может ускорять диссоциацию IgE от иммобилизованной формы высокоаффинного рецептора IgE, FcεRI. Мутации в легкой цепи (SEQ ID NO:20), делающие это возможным, включают, но не обязательно ограничиваются ими, мутации в положениях S56, S64, S67, S71, S80, S81, Q83 и L158 (S52, S60, S63, S67, S76, S77, Q79 и L154, соответственно, в соответствии с нумерацией Kabat).
Пример 8: Усиленное окисление мутантного Fab омализумаба
Образцы анти-IgE Fab подвергали режиму усиленного окисления для выяснения эффекта окисления остатков метионина в вариабельной области легкой цепи на аффинность Fab для IgE-Fc и на способность ускорять диссоциацию комплекса IgE-Fc:sFcεRIα. Мутантный Fab омализумаба инкубировали с 0,1% и 1% (по объему) раствором пероксида водорода вплоть до 14 дней при комнатной температуре. После инкубации буфер образцов меняли обратно на PBS, pH 7,4, и определяли концентрацию по поглощению при 280 нм с использованием расчетного коэффициента экстинкции. Проводили масс-спектральный анализ для определения количества окисленных остатков метионина в вариабельных областях легкой цепи путем восстановления и алкилирования материала в денатурирующих условиях, с последующим расщеплением трипсином (50 мкг/мл трипсина в течение 180 минут при 37°C, с последующим гашением TFA), а затем ЖХ-МС анализ на масс-спектрометре Thermo Orbitrap Q Exactive Plus. Процент окисленных остатков метионина рассчитывали, исходя из предположения, что окисленные остатки метионина отсутствовали во время синтеза, и сравнивали с референсным материалом, который хранили при 4°C.
Таблица 9: Процентная доля окисленных остатков метионина в вариабельной области легкой цепи мутантного Fab омализумаба. Данный мутант Fab омализумаба имеет следующие мутации: S64M_S81R_Q83R_S56D_S71M (со ссылкой на SEQ ID NO:20).
Вывод:
Полученные данные показывают, что возможно получать мутантный Fab омализумаба, у которого имеет место практически полное окисление критических остатков метионина легкой цепи (M64 и M71). Исходя из этих данных, материал из образцов в день 1, день 3 и день 7 объединяли и использовали для определения влияния окисления метионина на аффинность мутантного Fab омализумаба для IgE-Fc и на способность ускорять диссоциацию комплекса IgE-Fc:sFcεRIα.
Пример 9: Измерение ускоренной диссоциации IgE-Fc от sFcεRIα с использованием FRET
Эффект анти-IgE Fab на диссоциацию IgE-Fc от sFcεRIα определяли в гомогенном анализе FRET. Все анти-IgE Fab экспрессировали в клетках HEK-293, очищали стандартными методами и количественно определяли по поглощению при 280 нм с использованием расчетных коэффициентов молярной экстинкции. В анализе FRET использовали Tb-меченый IgE-Fc в качестве донора и Alexa 488-меченый sFcεRIα в качестве акцептора. Оба реагента смешивали и уравновешивали при комнатной температуре в течение 60 минут с конечной концентрацией в анализе 1 нМ. Анти-IgE Fab добавляли к смеси до конечной концентрации в анализе 500 нМ и снимали показания флуоресценции (возбуждение при 330 нм, эмиссия при 495 и 520 нм) каждые 20 минут в течение 800 минут. Строили график зависимости эмиссии флуоресценции от времени, и скорость диссоциации IgE-Fc от sFcεRIα рассчитывали, как время полужизни комплекса. Полученные данные приведены в Таблицах 10 и 11.
Таблица 10: Эффект анти-IgE Fab (ДТ и мутантов) на диссоциацию IgE-Fc от sFcεRIα в гомогенном анализе FRET. Диссоциация комплекса IgE-Fc:sFcεRIα представляет собой двухфазный распад. Быстрая начальная фаза представляет собой фазу, в которой большая часть комплекса IgE-Fc:sFcεRIα диссоциирует, и на основании которой определяют время полужизни комплекса. Мутации находятся в легкой цепи (со ссылкой на SEQ ID NO:20). Последовательности вариабельной области легкой цепи мутантов Fab указаны в левой колонке (VL SEQ ID NO:).
Таблица 11: Эффект окисления метионина на способность анти-IgE Fab вызывать диссоциацию IgE-Fc от sFcεRIα в гомогенном анализе FRET. Диссоциация комплекса IgE-Fc:sFcεRIα представляет собой двухфазный распад. Быстрая начальная фаза представляет собой фазу, в которой большая часть комплекса IgE-Fc:sFcεRIα диссоциирует, и на основании которой определяют время полужизни комплекса.
Выводы:
Полученные данные показывают, что мутантные формы омализумаба способны вызывать диссоциацию IgE-Fc от sFcεRIα с большей скоростью, чем последовательность дикого типа. В частности, мутации в легкой цепи (со ссылкой на SEQ ID NO:20), делающие это возможным, включают, но не обязательно ограничиваются ими, мутации в положениях S64 и S67. Кроме того, окисление остатков метионина (M64 и M71, SEQ ID NO:20) не оказывает существенного эффекта на способность Fab ускорять диссоциацию комплекса IgE-Fc: sFcεRIα.
В данной спецификации описаны варианты осуществления изобретения. Однако специалист в данной области понимает, что различные модификации и изменения могут быть внесены без отклонения от сущности изобретения, изложенной в приведенной ниже формуле изобретения. Соответственно, спецификацию следует рассматривать в качестве иллюстрации, но не ограничения изобретения, и все такие модификации должны входить в объем данного изобретения.
Последовательности
тяжелая цепь
SEQ ID NO:1
v-область
evqlvesggg lvqpggslrl scavsgysit sgyswnwirq apgkglewva sitydgstny npsvkgriti srddskntfy lqmnslraed tavyycargs hyfghwhfav wgqgtlvtvs s
SEQ ID NO:2
v-область
gaagtgcagt tggtggagtc gggtggaggg ctggtgcagc ctggcggtag cctgaggctg tcctgtgccg tgtccggata ctccattacc tccggctact cgtggaactg gatcagacag gctcccggaa agggacttga gtgggtggcg tccatcacct acgacggctc aaccaactat aacccgtccg tgaagggccg catcaccatt tcgcgcgacg acagcaagaa tactttttac ctccaaatga acagcctgcg ggccgaagat actgccgtgt actactgcgc gcggggatca cattacttcg ggcactggca cttcgccgtc tggggacagg gcaccctcgt cactgtctcg agc
SEQ ID NO:3
v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysev qlvesggglv qpggslrlsc avsgysitsg yswnwirqap gkglewvasi tydgstnynp svkgritisr ddskntfylq mnslraEDTA vyycargshy fghwhfavwg qgtlvtvss
SEQ ID NO:4
v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagc
SEQ ID NO:5
v-область+CH1 константной области гамма-1
evqlvesggg lvqpggslrl scavsgysit sgyswnwirq apgkglewva sitydgstny npsvkgriti srddskntfy lqmnslraed tavyycargs hyfghwhfav wgqgtlvtvs sastkgpsvf plapssksts ggtaalgclv kdyfpepvtv swnsgaltsg vhtfpavlqs sglyslssvv tvpssslgtq tyicnvnhkp sntkvdkkve pksc
SEQ ID NO:6
v-область+CH1 константной области гамма-1
gaagtgcagt tggtggagtc gggtggaggg ctggtgcagc ctggcggtag cctgaggctg tcctgtgccg tgtccggata ctccattacc tccggctact cgtggaactg gatcagacag gctcccggaa agggacttga gtgggtggcg tccatcacct acgacggctc aaccaactat aacccgtccg tgaagggccg catcaccatt tcgcgcgacg acagcaagaa tactttttac ctccaaatga acagcctgcg ggccgaagat actgccgtgt actactgcgc gcggggatca cattacttcg ggcactggca cttcgccgtc tggggacagg gcaccctcgt cactgtctcg agcgcttcta caaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag cccaaatctt gt
SEQ ID NO:7
v-область+CH1 константной области гамма-1 с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysev qlvesggglv qpggslrlsc avsgysitsg yswnwirqap gkglewvasi tydgstnynp svkgritisr ddskntfylq mnslraEDTA vyycargshy fghwhfavwg qgtlvtvssa stkgpsvfpl apsskstsgg taalgclvkd yfpepvtvsw nsgaltsgvh tfpavlqssg lyslssvvtv pssslgtqty icnvnhkpsn tkvdkkvepk sc
SEQ ID NO:8
v-область+CH1 константной области гамма-1 с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagcgct tctacaaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa tcttgt
SEQ ID NO:9
v-область+полноразмерная константная область гамма-1
evqlvesggg lvqpggslrl scavsgysit sgyswnwirq apgkglewva sitydgstny npsvkgriti srddskntfy lqmnslraed tavyycargs hyfghwhfav wgqgtlvtvs sastkgpsvf plapssksts ggtaalgclv kdyfpepvtv swnsgaltsg vhtfpavlqs sglyslssvv tvpssslgtq tyicnvnhkp sntkvdkkve pkscdkthtc ppcpapellg gpsvflfppk pkdtlmisrt pevtcvvvdv shedpevkfn wyvdgvevhn aktkpreeqy nstyrvvsvl tvlhqdwlng keykckvsnk alpapiekti skakgqprep qvytlppsrE eMtknqvslt clvkgfypsd iavewesngq pennykttpp vldsdgsffl yskltvdksr wqqgnvfscs vmhealhnhy tqkslslspg k
SEQ ID NO:10
v-область+полноразмерная константная область гамма-1
gaagtgcagt tggtggagtc gggtggaggg ctggtgcagc ctggcggtag cctgaggctg tcctgtgccg tgtccggata ctccattacc tccggctact cgtggaactg gatcagacag gctcccggaa agggacttga gtgggtggcg tccatcacct acgacggctc aaccaactat aacccgtccg tgaagggccg catcaccatt tcgcgcgacg acagcaagaa tactttttac ctccaaatga acagcctgcg ggccgaagat actgccgtgt actactgcgc gcggggatca cattacttcg ggcactggca cttcgccgtc tggggacagg gcaccctcgt cactgtctcg agcgcttcta caaagggccc ctccgtgttc ccgctcgctc catcatcgaa gtctaccagc ggaggcactg cggctctcgg ttgcctcgtg aaggactact tcccggagcc ggtgaccgtg tcgtggaaca gcggagccct gaccagcggg gtgcacacct ttccggccgt cttgcagtca agcggccttt actccctgtc atcagtggtg actgtcccgt ccagctcatt gggaacccaa acctacatct gcaatgtgaa tcacaaacct agcaacacca aggttgacaa gaaagtcgag cccaaatcgt gtgacaagac tcacacttgt ccgccgtgcc cggcacccga actgctggga ggtcccagcg tctttctgtt ccctccaaag ccgaaagaca cgctgatgat ctcccgcacc ccggaggtca cttgcgtggt cgtggacgtg tcacatgagg acccagaggt gaagttcaat tggtacgtgg atggcgtcga agtccacaat gccaaaacta agcccagaga agaacagtac aattcgacct accgcgtcgt gtccgtgctc acggtgttgc atcaggattg gctgaacggg aaggaataca agtgcaaagt gtccaacaag gcgctgccgg caccgatcga gaaaactatc tccaaagcga agggacagcc tagggaacct caagtctaca cgctgccacc atcacgggaA gaaAtgacta agaatcaagt ctcactgact tgtctggtga aggggtttta ccctagcgac attgccgtgg agtgggaatc caacggccag ccagagaaca actacaagac tacccctcca gtgctcgact cggatggatc gttcttcctt tactcgaagc tcaccgtgga taagtcccgg tggcagcagg gaaacgtgtt ctcctgctcg gtgatgcatg aagccctcca taaccactat acccaaaagt cgctgtccct gtcgccggga aag
SEQ ID NO:11
v-область+полноразмерная константная область гамма-1 с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysev qlvesggglv qpggslrlsc avsgysitsg yswnwirqap gkglewvasi tydgstnynp svkgritisr ddskntfylq mnslraEDTA vyycargshy fghwhfavwg qgtlvtvssa stkgpsvfpl apsskstsgg taalgclvkd yfpepvtvsw nsgaltsgvh tfpavlqssg lyslssvvtv pssslgtqty icnvnhkpsn tkvdkkvepk scdkthtcpp cpapellggp svflfppkpk dtlmisrtpe vtcvvvdvsh edpevkfnwy vdgvevhnak tkpreeqyns tyrvvsvltv lhqdwlngke ykckvsnkal papiektisk akgqprepqv ytlppsrEeM tknqvsltcl vkgfypsdia vewesngqpe nnykttppvl dsdgsfflys kltvdksrwq qgnvfscsvm healhnhytq kslslspgk
SEQ ID NO:12
v-область+полноразмерная константная область гамма-1 с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagcgct tctacaaagg gcccctccgt gttcccgctc gctccatcat cgaagtctac cagcggaggc actgcggctc tcggttgcct cgtgaaggac tacttcccgg agccggtgac cgtgtcgtgg aacagcggag ccctgaccag cggggtgcac acctttccgg ccgtcttgca gtcaagcggc ctttactccc tgtcatcagt ggtgactgtc ccgtccagct cattgggaac ccaaacctac atctgcaatg tgaatcacaa acctagcaac accaaggttg acaagaaagt cgagcccaaa tcgtgtgaca agactcacac ttgtccgccg tgcccggcac ccgaactgct gggaggtccc agcgtctttc tgttccctcc aaagccgaaa gacacgctga tgatctcccg caccccggag gtcacttgcg tggtcgtgga cgtgtcacat gaggacccag aggtgaagtt caattggtac gtggatggcg tcgaagtcca caatgccaaa actaagccca gagaagaaca gtacaattcg acctaccgcg tcgtgtccgt gctcacggtg ttgcatcagg attggctgaa cgggaaggaa tacaagtgca aagtgtccaa caaggcgctg ccggcaccga tcgagaaaac tatctccaaa gcgaagggac agcctaggga acctcaagtc tacacgctgc caccatcacg ggaAgaaAtg actaagaatc aagtctcact gacttgtctg gtgaaggggt tttaccctag cgacattgcc gtggagtggg aatccaacgg ccagccagag aacaactaca agactacccc tccagtgctc gactcggatg gatcgttctt cctttactcg aagctcaccg tggataagtc ccggtggcag cagggaaacg tgttctcctg ctcggtgatg catgaagccc tccataacca ctatacccaa aagtcgctgt ccctgtcgcc gggaaag
SEQ ID NO:13
FR H1
evqlvesggg lvqpggslrl scavs
SEQ ID NO:14
CDRH1
gysitsgysw n
SEQ ID NO:15
FR H2
wirqapgkgl ewva
SEQ ID NO:16
CDRH2
sitydgstny npsvkg
SEQ ID NO:17
FR H3
ritisrddsk ntfylqmnsl raEDTAvyyc ar
SEQ ID NO:18
CDRH3
gshyfghwhf av
SEQ ID NO:19
FR H4
wgqgtlvtvs s
ЛЕГКАЯ ЦЕПЬ
SEQ ID NO:20
Омализумаб_v-область
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS SLQPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:21
Омализумаб_v-область
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat acgtttggcc agggaaccaa ggtcgaaatc aag
SEQ ID NO:22
Омализумаб_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaasylesgv pSrfsgsgsg tdftltisSl Qpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:23
Омализумаб_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt ggccagggaa ccaaggtcga aatcaag
SEQ ID NO:24
Омализумаб_v-область+константная область каппа
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS SLQPEDFATY YCQQSHEDPY TFGQGTKVEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
SEQ ID NO:25
Омализумаб_v-область+константная область каппа
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
SEQ ID NO:26
Омализумаб_v-область+константная область каппа с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysDI QLTQSPSSLS ASVGDRVTIT CRASQSVDYD GDSYMNWYQQ KPGKAPKLLI YAASYLESGV PSRFSGSGSG TDFTLTISSL QPEDFATYYC QQSHEDPYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG TASVVCLLNN FYPREAKVQW KVDNALQSGN SQESVTEQDS KDSTYSLSST LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:27
Омализумаб_v-область+константная область каппа с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg cctTCgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgTCGctg CAgccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt ggccagggaa ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg aaggtggata acgcccTCca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt
SEQ ID NO:28
FR L1
DIQLTQSPSS LSASVGDRVT ITC
SEQ ID NO:29
CDRL1
rasqsvdydg dsymn
SEQ ID NO:30
FR L2
WYQQKPGKAP KLLIY
SEQ ID NO:31
CDRL2
aasyles
SEQ ID NO:32
FR L3
GVPSRFSGSG SGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:33
CDRL3
qqshedpyt
SEQ ID NO:34
FR L4
FGQGTKVEIK
МУТАНТНАЯ ЛЕГКАЯ ЦЕПЬ
SEQ ID NO:35
S60M_S77R_Q79R_v-область (Kabat)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPMRFSGSG SGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:36
S60M_S77R_Q79R_v-область (Kabat)
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc ggcgtgccta tgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat acgtttggcc agggaaccaa ggtcgaaatc aag
SEQ ID NO:37
S60M_S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом (Kabat)
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaasylesgv pmrfsgsgsg tdftltisrl rpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:38
S60M_S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом (Kabat)
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg cctatgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgagactg aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt ggccagggaa ccaaggtcga aatcaag
SEQ ID NO:39
S60M_S77R_Q79R_v-область+константная область каппа, включающая L154P (Kabat)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPMRFSGSG SGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNAPQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
SEQ ID NO:40
S60M_S77R_Q79R_v-область+константная область каппа, включающая L154P (Kabat)
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc ggcgtgccta tgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cccgcaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
SEQ ID NO:41
S60M_S77R_Q79R_v-область+константная область каппа, включающая L154P, с сигнальной последовательностью, которая подчеркнута и выделена курсивом (Kabat)
mkwvtfisll flfssaysDI QLTQSPSSLS ASVGDRVTIT CRASQSVDYD GDSYMNWYQQ KPGKAPKLLI YAASYLESGV PMRFSGSGSG TDFTLTISRL RPEDFATYYC QQSHEDPYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG TASVVCLLNN FYPREAKVQW KVDNAPQSGN SQESVTEQDS KDSTYSLSST LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:42
S60M_S77R_Q79R_v-область+константная область каппа, включающая L154P, с сигнальной последовательностью, которая подчеркнута и выделена курсивом (Kabat)
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg cctatgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgagactg aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt ggccagggaa ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg aaggtggata acgccccgca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt
SEQ ID NO:43
FR L3 S64M (PDB) S60M (Kabat)
GVPMRFSGSG SGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:44
FR L3 S81R (PDB) S77r (Kabat)
GVPSRFSGSG SGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:45
FR L3 Q83R (PDB) q79r (Kabat)
GVPSRFSGSG SGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:46
FR L3 S64M S81R (PDB) S60M S77R (Kabat)
GVPMRFSGSG SGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:47
FR L3 S64M Q83R (PDB) S60M Q79R (Kabat)
GVPMRFSGSG SGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:48
FR L3 S81R Q83R (PDB) S77R Q79R (Kabat)
GVPSRFSGSG SGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:49
FR L3 S64M S81R Q83R (PDB) S60M S77R Q79R (Kabat)
GVPMRFSGSG SGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:50
CDRL2 S56D (PDB) S52d (Kabat)
aaDyles
SEQ ID NO:51
CDRL2 S56E (PDB) S52e (Kabat)
aaEyles
SEQ ID NO:52
FR L3 S71M (PDB) S67m (Kabat)
GVPSRFSGSG SGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:53
FR L3 S64M S71M (PDB) S60M S67m (Kabat)
GVPMRFSGSG MGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:54
FR L3 S81R S71M (PDB) S77r S67m (Kabat)
GVPSRFSGSG MGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:55
FR L3 Q83R S71M (PDB) q79r S67m (Kabat)
GVPSRFSGSG MGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:56
FR L3 S64M S81R S71M (PDB) S60M S77R S67m (Kabat)
GVPMRFSGSG MGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:57
FR L3 S64M Q83R S71M (PDB) S60M Q79R S67m (Kabat)
GVPMRFSGSG MGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:58
FR L3 S81R Q83R S71M (PDB) S77R Q79R S67m (Kabat)
GVPSRFSGSG MGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:59
FR L3 S64M S81R Q83R S71M (PDB) S60M S77R Q79R S67m (Kabat)
GVPMRFSGSG MGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:60
FR L3 S67Y (PDB) S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:61
FR L3 S64M S67Y (PDB) S60M S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:62
FR L3 S81R S67Y (PDB) S77r S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:63
FR L3 Q83R S67Y (PDB) q79r S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:64
FR L3 S64M S81R S67Y (PDB) S60M S77R S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:65
FR L3 S64M Q83R S67Y (PDB) S60M Q79R S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:66
FR L3 S81R Q83R S67Y (PDB) S77R Q79R S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:67
FR L3 S64M S81R Q83R S67Y (PDB) S60M S77R Q79R S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:68
FR L3 S80N (PDB) S76N (Kabat)
GVPSRFSGSG SGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:69
FR L3 S64M S80N (PDB) S60M S76N (Kabat)
GVPMRFSGSG SGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:70
FR L3 S81R S80N (PDB) S77r S76N (Kabat)
GVPSRFSGSG SGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:71
FR L3 Q83R S80N (PDB) q79r S76N (Kabat)
GVPSRFSGSG SGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:72
FR L3 S64M S81R S80N (PDB) S60M S77R S76N (Kabat)
GVPMRFSGSG SGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:73
FR L3 S64M Q83R S80N (PDB) S60M Q79R S76N (Kabat)
GVPMRFSGSG SGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:74
FR L3 S81R Q83R S80N (PDB) S77R Q79R S76N (Kabat)
GVPSRFSGSG SGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:75
FR L3 S64M S81R Q83R S80N (PDB) S60M S77R Q79R S76N (Kabat)
GVPMRFSGSG SGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:76
FR L3 S80N S67Y (PDB) S76N S63Y(Kabat)
GVPSRFYGSG SGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:77
FR L3 S64M S80N S67Y (PDB) S60M S76N S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:78
FR L3 S81R S80N S67Y (PDB) S77r S76N S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:79
FR L3 Q83R S80N S67Y (PDB) q79r S76N S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:80
FR L3 S64M S81R S80N S67Y (PDB) S60M S77R S76N S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:81
FR L3 S64M Q83R S80N S67Y (PDB) S60M Q79R S76N S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:82
FR L3 S81R Q83R S80N S67Y (PDB) S77R Q79R S76N S63Y (Kabat)
GVPSRFYGSG SGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:83
FR L3 S64M S81R Q83R S80N S67Y (PDB) S60M S77R Q79R S76N S63Y (Kabat)
GVPMRFYGSG SGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:84
FR L3 S67Y S71M (PDB) S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:85
FR L3 S64M S67Y S71M (PDB) S60M S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIS SLQPEDFATY YC
SEQ ID NO:86
FR L3 S81R S67Y S71M (PDB) S77r S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:87
FR L3 Q83R S67Y S71M (PDB) q79r S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:88
FR L3 S64M S81R S67Y S71M (PDB) S60M S77R S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIS RLQPEDFATY YC
SEQ ID NO:89
FR L3 S64M Q83R S67Y S71M (PDB) S60M Q79R S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIS SLRPEDFATY YC
SEQ ID NO:90
FR L3 S81R Q83R S67Y S71M (PDB) S77R Q79R S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:91
FR L3 S64M S81R Q83R S67Y S71M (PDB) S60M S77R Q79R S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIS RLRPEDFATY YC
SEQ ID NO:92
FR L3 S80N S71M (PDB) S76N S67M (Kabat)
GVPSRFSGSG MGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:93
FR L3 S64M S80N S71M (PDB) S60M S76N S67M (Kabat)
GVPMRFSGSG MGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:94
FR L3 S81R S80N S71M (PDB) S77r S76N S67M (Kabat)
GVPSRFSGSG MGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:95
FR L3 Q83R S80N S71M (PDB) q79r S76N S67M (Kabat)
GVPSRFSGSG MGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:96
FR L3 S64M S81R S80N S71M (PDB) S60M S77R S76N S67M (Kabat)
GVPMRFSGSG MGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:97
FR L3 S64M Q83R S80N S71M (PDB) S60M Q79R S76N S67M (Kabat)
GVPMRFSGSG MGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:98
FR L3 S81R Q83R S80N S71M (PDB) S77R Q79R S76N S67M (Kabat)
GVPSRFSGSG MGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:99
FR L3 S64M S81R Q83R S80N S71M (PDB) S60M S77R Q79R S76N S67M (Kabat)
GVPMRFSGSG MGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:100
FR L3 S80N S67Y S71M (PDB) S76N S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:101
FR L3 S64M S80N S67Y S71M (PDB) S60M S76N S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIN SLQPEDFATY YC
SEQ ID NO:102
FR L3 S81R S80N S67Y S71M (PDB) S77r S76N S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:103
FR L3 Q83R S80N S67Y S71M (PDB) q79r S76N S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:104
FR L3 S64M S81R S80N S67Y S71M (PDB) S60M S77R S76N S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIN RLQPEDFATY YC
SEQ ID NO:105
FR L3 S64M Q83R S80N S67Y S71M (PDB) S60M Q79R S76N S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIN SLRPEDFATY YC
SEQ ID NO:106
FR L3 S81R Q83R S80N S67Y S71M (PDB) S77R Q79R S76N S63Y S67M (Kabat)
GVPSRFYGSG MGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:107
FR L3 S64M S81R Q83R S80N S67Y S71M (PDB) S60M S77R Q79R S76N S63Y S67M (Kabat)
GVPMRFYGSG MGTDFTLTIN RLRPEDFATY YC
SEQ ID NO:108
Человеческий IgE-Fc дикого типа (домены Cε2-Cε4 с нумерацией V224-K547 согласно Dorrington & Bennich (1978) Immunol. Rev. 41:3-25), несущий мутацию C225A; Примечание, в кристаллографических экспериментах следующие мутации также были введены в IgE-Fc для упрощения картины гликозилирования: N265Q и N371Q (также в соответствии с нумерацией Dorrington & Bennich)
VASRDFTPPT VKILQSSCDG GGHFPPTIQL LCLVSGYTPG TINITWLEDG QVMDVDLSTA STTQEGELAS TQSELTLSQK HWLSDRTYTC QVTYQGHTFE DSTKKCADSN PRGVSAYLSR PSPFDLFIRK SPTITCLVVD LAPSKGTVNL TWSRASGKPV NHSTRKEEKQ RNGTLTVTST LPVGTRDWIE GETYQCRVTH PHLPRALMRS TTKTSGPRAA PEVYAFATPE WPGSRDKRTL ACLIQNFMPE DISVQWLHNE VQLPDARHST TQPRKTKGSG FFVFSRLEVT RAEWEQKDEF ICRAVHEAAS PSQTVQRAVS VNPGK
ЛЕГКАЯ ЦЕПЬ
Fab1
SEQ ID NO:109
S77R_Q79R_v-область (Fab1)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:110
S77R_Q79R_v-область (Fab1)
GATATTCAGC TGACTCAGAG CCCGAGCTCA CTCTCCGCTT CCGTGGGGGA TAGAGTGACC ATCACTTGCC GGGCATCCCA GTCGGTGGAC TACGACGGAG ACTCCTACAT GAACTGGTAC CAGCAGAAGC CCGGAAAAGC CCCAAAGTTG CTGATCTACG CCGCCTCCTA CCTTGAAAGC GGCGTGCCTT CACGCTTCTC GGGAAGCGGG TCTGGCACCG ATTTCACCCT GACCATTTCG AGACTGAGGC CGGAGGACTT CGCGACTTAC TACTGCCAAC AGTCCCACGA GGACCCCTAT ACGTTTGGCC AGGGTACCAA GGTCGAAATC AAG
SEQ ID NO:111
S77R_Q79R_v-область (Fab1) с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaasylesgv pSrfsgsgsg tdftltisrl rpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:112
S77R_Q79R_v-область (Fab1) с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccGATATT CAGCTGACTC AGAGCCCGAG CTCACTCTCC GCTTCCGTGG GGGATAGAGT GACCATCACT TGCCGGGCAT CCCAGTCGGT GGACTACGAC GGAGACTCCT ACATGAACTG GTACCAGCAG AAGCCCGGAA AAGCCCCAAA GTTGCTGATC TACGCCGCCT CCTACCTTGA AAGCGGCGTG CCTTCACGCT TCTCGGGAAG CGGGTCTGGC ACCGATTTCA CCCTGACCAT TTCGAGACTG AGGCCGGAGG ACTTCGCGAC TTACTACTGC CAACAGTCCC ACGAGGACCC CTATACGTTT GGCCAGGGTA CCAAGGTCGA AATCAAG
Fab2
Омализумаб_v-область+константная область каппа, включающая L154P
SEQ ID NO:113
Fab2_v-область
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS SLQPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:114
Fab2_v-область
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat acgtttggcc agggaaccaa ggtcgaaatc aag
SEQ ID NO:115
Fab2_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaasylesgv pSrfsgsgsg tdftltisSl Qpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:116
Fab2_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt ggccagggaa ccaaggtcga aatcaag
SEQ ID NO:117
Fab2_v-область+константная область каппа, включающая L154P
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS SLQPEDFATY YCQQSHEDPY TFGQGTKVEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNAPQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
SEQ ID NO:118
Fab2_v-область+константная область каппа, включающая L154P
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cccgcaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
SEQ ID NO:119
Fab2_v-область+константная область каппа, включающая L154P, с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysDI QLTQSPSSLS ASVGDRVTIT CRASQSVDYD GDSYMNWYQQ KPGKAPKLLI YAASYLESGV PSRFSGSGSG TDFTLTISSL QPEDFATYYC QQSHEDPYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG TASVVCLLNN FYPREAKVQW KVDNAPQSGN SQESVTEQDS KDSTYSLSST LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:120
Fab2_v-область+константная область каппа, включающая L154P, с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt ggccagggaa ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg aaggtggata acgccccgca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt
Fab3
S77R_Q79R_v-область (из Fab1)+константная область каппа, включающая L154P (из Fab2)
SEQ ID NO:121
S77R_Q79R_v-область
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:122
S77R_Q79R_v-область
GATATTCAGC TGACTCAGAG CCCGAGCTCA CTCTCCGCTT CCGTGGGGGA TAGAGTGACC ATCACTTGCC GGGCATCCCA GTCGGTGGAC TACGACGGAG ACTCCTACAT GAACTGGTAC CAGCAGAAGC CCGGAAAAGC CCCAAAGTTG CTGATCTACG CCGCCTCCTA CCTTGAAAGC GGCGTGCCTT CACGCTTCTC GGGAAGCGGG TCTGGCACCG ATTTCACCCT GACCATTTCG AGACTGAGGC CGGAGGACTT CGCGACTTAC TACTGCCAAC AGTCCCACGA GGACCCCTAT ACGTTTGGCC AGGGTACCAA GGTCGAAATC AAG
SEQ ID NO:123
S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaasylesgv pSrfsgsgsg tdftltisrl rpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:124
S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccGATATT CAGCTGACTC AGAGCCCGAG CTCACTCTCC GCTTCCGTGG GGGATAGAGT GACCATCACT TGCCGGGCAT CCCAGTCGGT GGACTACGAC GGAGACTCCT ACATGAACTG GTACCAGCAG AAGCCCGGAA AAGCCCCAAA GTTGCTGATC TACGCCGCCT CCTACCTTGA AAGCGGCGTG CCTTCACGCT TCTCGGGAAG CGGGTCTGGC ACCGATTTCA CCCTGACCAT TTCGAGACTG AGGCCGGAGG ACTTCGCGAC TTACTACTGC CAACAGTCCC ACGAGGACCC CTATACGTTT GGCCAGGGTA CCAAGGTCGA AATCAAG
SEQ ID NO:125
S77R_Q79R_v-область+константная область каппа, включающая L154P
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAASYLES GVPSRFSGSG SGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNAPQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
SEQ ID NO:126
S77R_Q79R_v-область+константная область каппа, включающая L154P
GATATTCAGC TGACTCAGAG CCCGAGCTCA CTCTCCGCTT CCGTGGGGGA TAGAGTGACC ATCACTTGCC GGGCATCCCA GTCGGTGGAC TACGACGGAG ACTCCTACAT GAACTGGTAC CAGCAGAAGC CCGGAAAAGC CCCAAAGTTG CTGATCTACG CCGCCTCCTA CCTTGAAAGC GGCGTGCCTT CACGCTTCTC GGGAAGCGGG TCTGGCACCG ATTTCACCCT GACCATTTCG AGACTGAGGC CGGAGGACTT CGCGACTTAC TACTGCCAAC AGTCCCACGA GGACCCCTAT ACGTTTGGCC AGGGTACCAA GGTCGAAATC AAGcgtacgg tagcggcccc atctgtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cccgcaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
SEQ ID NO:127
S77R_Q79R_v-область+константная область каппа, включающая L154P, с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysDI QLTQSPSSLS ASVGDRVTIT CRASQSVDYD GDSYMNWYQQ KPGKAPKLLI YAASYLESGV PSRFSGSGSG TDFTLTISRL RPEDFATYYC QQSHEDPYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG TASVVCLLNN FYPREAKVQW KVDNAPQSGN SQESVTEQDS KDSTYSLSST LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:128
S77R_Q79R_v-область+константная область каппа, включающая L154P, с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccGATATT CAGCTGACTC AGAGCCCGAG CTCACTCTCC GCTTCCGTGG GGGATAGAGT GACCATCACT TGCCGGGCAT CCCAGTCGGT GGACTACGAC GGAGACTCCT ACATGAACTG GTACCAGCAG AAGCCCGGAA AAGCCCCAAA GTTGCTGATC TACGCCGCCT CCTACCTTGA AAGCGGCGTG CCTTCACGCT TCTCGGGAAG CGGGTCTGGC ACCGATTTCA CCCTGACCAT TTCGAGACTG AGGCCGGAGG ACTTCGCGAC TTACTACTGC CAACAGTCCC ACGAGGACCC CTATACGTTT GGCCAGGGTA CCAAGGTCGA AATCAAGcgt acggtagcgg ccccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg aaggtggata acgccccgca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt
SEQ ID NO:129
Омализумаб_v-область
TQSPSSLSAS VGDRVTITCR ASQSVDYDGD SYMNWYQQKP GKAPKLLIYA ASYLESGVPS RFSGSGSGTD FTLTISSLQP EDFATYYCQQ SHEDPYTFGQ GTKVEIK
SEQ ID NO:130
Омализумаб_v-область
actcagagcc cgagctcact ctccgcttcc gtgggggata gagtgaccat cacttgccgg gcatcccagt cggtggacta cgacggagac tcctacatga actggtatca gcagaagccc ggaaaagccc caaagttgct gatctacgcc gcctcatacc ttgaaagcgg cgtgccttcg cgcttctcgg gaagcgggtc gggcaccgat ttcaccctga ccatttcgtc gctgcagccg gaggacttcg cgacttacta ctgccaacag tcccacgagg acccctatac gtttggccag ggaaccaagg tcgaaatcaa г
SEQ ID NO:131
FR3
GVPMRFSGSGMGTDFTLTISRLRPEDFATYYC
SEQ ID NO:132
S60M_S52D_S67M_S77R_Q79R_v-область
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAADYLES GVPMRFSGSG MGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:133
S60M_S52D_S67M_S77R_Q79R_v-область
GATATTCAGCTGACTCAGAGCCCGAGCTCACTCTCCGCTTCCGTGGGGGATAGAGTGACCATCACTTGCCGGGCATCCCAGTCGGTGGACTACGACGGAGACTCCTACATGAACTGGTATCAGCAGAAGCCCGGAAAAGCCCCAAAGTTGCTGATCTACGCCGCCGACTACCTTGAAAGCGGCGTGCCTATGCGCTTCTCGGGAAGCGGGATGGGCACCGATTTCACCCTGACCATTTCGAGACTGAGGCCGGAGGACTTCGCGACTTACTACTGCCAACAGTCCCACGAGGACCCCTATACGTTTGGCCAGGGAACCAAGGTCGAAATCAAG
SEQ ID NO:134
S60M_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaaDylesgv pMrfsgsgMg tdftltisrl rpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:135
S60M_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
ATGAAGTGGGTCACCTTCATCTCCCTGCTGTTTCTGTTCTCCTCCGCCTACTCCGATATTCAGCTGACTCAGAGCCCGAGCTCACTCTCCGCTTCCGTGGGGGATAGAGTGACCATCACTTGCCGGGCATCCCAGTCGGTGGACTACGACGGAGACTCCTACATGAACTGGTATCAGCAGAAGCCCGGAAAAGCCCCAAAGTTGCTGATCTACGCCGCCGACTACCTTGAAAGCGGCGTGCCTATGCGCTTCTCGGGAAGCGGGATGGGCACCGATTTCACCCTGACCATTTCGAGACTGAGGCCGGAGGACTTCGCGACTTACTACTGCCAACAGTCCCACGAGGACCCCTATACGTTTGGCCAGGGAACCAAGGTCGAAATCAAG
SEQ ID NO:136
S60M_S52D_S67M_S77R_Q79R_v-область+константная область каппа
DI QLTQSPSSLS ASVGDRVTIT CRASQSVDYD GDSYMNWYQQ KPGKAPKLLI YAADYLESGV PMRFSGSGMG TDFTLTISRL RPEDFATYYC QQSHEDPYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG TASVVCLLNN FYPREAKVQW KVDNALQSGN SQESVTEQDS KDSTYSLSST LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:137
S60M_S52D_S67M_S77R_Q79R_v-область+константная область каппа
GATATTCAGCTGACTCAGAGCCCGAGCTCACTCTCCGCTTCCGTGGGGGATAGAGTGACCATCACTTGCCGGGCATCCCAGTCGGTGGACTACGACGGAGACTCCTACATGAACTGGTATCAGCAGAAGCCCGGAAAAGCCCCAAAGTTGCTGATCTACGCCGCCGACTACCTTGAAAGCGGCGTGCCTATGCGCTTCTCGGGAAGCGGGATGGGCACCGATTTCACCCTGACCATTTCGAGACTGAGGCCGGAGGACTTCGCGACTTACTACTGCCAACAGTCCCACGAGGACCCCTATACGTTTGGCCAGGGAACCAAGGTCGAAATCAAGCGTACGGTAGCGGCCCCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGT
SEQ ID NO:138
FR3
GVPRRFSGSGMGTDFTLTISRLRPEDFATYYC
SEQ ID NO:139
S60R_S52D_S67M_S77R_Q79R_v-область
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAADYLES GVPRRFSGSG MGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:140
S60R_S52D_S67M_S77R_Q79R_v-область
gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagcggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaag
SEQ ID NO:141
S60R_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
mkwvtfisll flfssaysdi qltqspssls asvgdrvtit crasqsvdyd gdsymnwyqq kpgkapklli yaaDylesgv pRrfsgsgMg tdftltisrl rpedfatyyc qqshedpytf gqgtkveik
SEQ ID NO:142
S60R_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью, которая подчеркнута и выделена курсивом
ATGAAGTGGGTCACCTTCATCTCCCTGCTGTTTCTGTTCTCCTCCGCCTACTCCGATATTCAGCTGACTCAGAGCCCGAGCTCACTCTCCGCTTCCGTGGGGGATAGAGTGACCATCACTTGCCGGGCATCCCAGTCGGTGGACTACGACGGAGACTCCTACATGAACTGGTATCAGCAGAAGCCCGGAAAAGCCCCAAAGTTGCTGATCTACGCCGCCGATTACCTTGAAAGCGGCGTGCCTCGTCGCTTCTCGGGAAGCGGGATGGGCACCGATTTCACCCTGACCATTTCGAGACTGAGGCCGGAGGACTTCGCGACTTACTACTGCCAACAGTCCCACGAGGACCCCTATACGTTTGGCCAGGGAACCAAGGTCGAAATCAAG
SEQ ID NO:143
S60R_S52D_S67M_S77R_Q79R_v-область+константная область каппа
DI QLTQSPSSLS ASVGDRVTIT CRASQSVDYD GDSYMNWYQQ KPGKAPKLLI YAADYLESGV PRRFSGSGMG TDFTLTISRL RPEDFATYYC QQSHEDPYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG TASVVCLLNN FYPREAKVQW KVDNALQSGN SQESVTEQDS KDSTYSLSST LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:144
S60R_S52D_S67M_S77R_Q79R_v-область+константная область каппа
gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagcggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt
SEQ ID NO:145
S60K_S52D_S67M_S77R_Q79R_v-область (Kabat)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAADYLES GVPKRFSGSG MGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:146
S60Q_S52D_S67M_S77R_Q79R_v-область (Kabat)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAADYLES GVPQRFSGSG MGTDFTLTIS RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:147
v-область+CH1 константной области гамма-1 плюс линкер плюс CA645 gL4gH5 scFv с сигнальной последовательностью, которая подчеркнута и выделена курсивом
MKWVTFISLLFLFSSAYSEVQLVESGGGLVQPGGSLRLSCAVSGYSITSGYSWNWIRQAPGKGLEWVASITYDGSTNYNPSVKGRITISRDDSKNTFYLQMNSLRAEDTAVYYCARGSHYFGHWHFAVWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCSGGGGTGGGGSEVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKCLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSGGGGSGGGGSGGGGSGGGGSDIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGCGTKVEIKRT
SEQ ID NO:148
v-область+CH1 константной области гамма-1 плюс линкер плюс CA645 gL4gH5 scFv с сигнальной последовательностью, которая подчеркнута и выделена курсивом
atgaagtgggtcaccttcatctccctgctgtttctgttctccagcgcctactccgaagtgcagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctgtcctgtgccgtgtccggatactccattacctccggctactcgtggaactggatcagacaggctcccggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactataacccgtccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttacctccaaatgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatcacattacttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcgagcgcttctacaaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccagtgacggtgtcgtggaactcaggtgccctgaccagcggcgttcacaccttcccggctgtcctacagtcttcaggactctactccctgagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtcgataagaaagttgagcccaaatcttgtagtggaggtgggggcaccggtggaggtggcagcgaggttcaactgcttgagtctggaggaggcctagtccagcctggagggagcctgcgtctctcttgtgcagtaagcggcatcgacctgagcaattacgccatcaactgggtgagacaagctccggggaagtgtttagaatggatcggtataatatgggccagtgggacgaccttttatgctacatgggcgaaaggaaggtttacaattagccgggacaatagcaaaaacaccgtgtatctccaaatgaactccttgcgagcagaggacacggcggtgtactattgtgctcgcactgtcccaggttatagcactgcaccctacttcgatctgtggggacaagggaccctggtgactgtttcaagtggcggagggggtagtggagggggtggctctgggggtggcggaagcggtggcgggggttctgacatacaaatgactcagtctccttcatcggtatccgcgtccgttggcgatagggtgactattacatgtcaaagctctcctagcgtctggagcaattttctatcctggtatcaacagaaaccggggaaggctccaaaacttctgatttatgaagcctcgaaactcaccagtggagttccgtcaagattcagtggctctggatcagggacagacttcacgttgacaatcagttcgctgcaaccagaggactttgcgacctactattgtggtggaggttacagtagcataagtgatacgacatttgggtgcggtactaaggtggaaatcaaacgtacc
SEQ ID NO:149
Линкер между CH1 и CA645 gL4gH5 scFv
SGGGGTGGGGS
SEQ ID NO:150
CA645 gL4gH5 scFv
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKCLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSGGGGSGGGGSGGGGSGGGGSDIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGCGTKVEIKRT
SEQ ID NO:151
Линкер между CA645 gH5 и CA645 gL4 в scFv
GGGGSGGGGSGGGGSGGGGS
SEQ ID NO:152
CA645 CDRH1
GIDLSNYAIN
SEQ ID NO:153
CA645 CDRH2
IIWASGTTFYATWAKG
SEQ ID NO:154
CA645 CDRH3
TVPGYSTAPYFDL
SEQ ID NO:155
CA645 CDRL1
QSSPSVWSNFLS
SEQ ID NO:156
CA645 CDRL2
EASKLTS
SEQ ID NO:157
CA645 CDRL3
GGGYSSISDTT
SEQ ID NO:158
S60M_S52D_S67M_S77R_Q79R_S63W_S76N_v-область (Kabat)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAADYLES GVPMRFWGSG MGTDFTLTIN RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:159
S60M_S52D_S67M_S77R_Q79R_S63Y_S76N_v-область (Kabat)
DIQLTQSPSS LSASVGDRVT ITCRASQSVD YDGDSYMNWY QQKPGKAPKL LIYAADYLES GVPMRFYGSG MGTDFTLTIN RLRPEDFATY YCQQSHEDPY TFGQGTKVEI K
SEQ ID NO:160
Сигнальная последовательность
mkwvtfisll flfssays
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЮСБ Биофарма СПРЛ
<120> АНТИ-IgE АНТИТЕЛА
<130> PF0087-WO
<150> GB1610198.2
<151> 2016-06-10
<160> 160
<170> PatentIn version 3.5
<210> 1
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> V-область
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 2
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> V-область
<400> 2
gaagtgcagt tggtggagtc gggtggaggg ctggtgcagc ctggcggtag cctgaggctg 60
tcctgtgccg tgtccggata ctccattacc tccggctact cgtggaactg gatcagacag 120
gctcccggaa agggacttga gtgggtggcg tccatcacct acgacggctc aaccaactat 180
aacccgtccg tgaagggccg catcaccatt tcgcgcgacg acagcaagaa tactttttac 240
ctccaaatga acagcctgcg ggccgaagat actgccgtgt actactgcgc gcggggatca 300
cattacttcg ggcactggca cttcgccgtc tggggacagg gcaccctcgt cactgtctcg 360
agc 363
<210> 3
<211> 139
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> V-область с сигнальной последовательностью
<400> 3
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
20 25 30
Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr
35 40 45
Ser Gly Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro
65 70 75 80
Ser Val Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
85 90 95
Phe Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
100 105 110
Tyr Cys Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
130 135
<210> 4
<211> 417
<212> ДНК
<213> Искусственная последовательность
<220>
<223> V-область с сигнальной последовательностью
<400> 4
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg 60
cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt 120
gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc 180
ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg 240
tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa 300
atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac 360
ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagc 417
<210> 5
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> v-область + CH1 константной области гамма-1
<400> 5
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 6
<211> 672
<212> ДНК
<213> Искусственная последовательность
<220>
<223> v-область + CH1 константной области гамма-1
<400> 6
gaagtgcagt tggtggagtc gggtggaggg ctggtgcagc ctggcggtag cctgaggctg 60
tcctgtgccg tgtccggata ctccattacc tccggctact cgtggaactg gatcagacag 120
gctcccggaa agggacttga gtgggtggcg tccatcacct acgacggctc aaccaactat 180
aacccgtccg tgaagggccg catcaccatt tcgcgcgacg acagcaagaa tactttttac 240
ctccaaatga acagcctgcg ggccgaagat actgccgtgt actactgcgc gcggggatca 300
cattacttcg ggcactggca cttcgccgtc tggggacagg gcaccctcgt cactgtctcg 360
agcgcttcta caaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct 420
gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg 480
tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc 540
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 600
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag 660
cccaaatctt gt 672
<210> 7
<211> 242
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> v-область + CH1 константной области гамма-1 с сигнальной
последовательностью
<400> 7
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
20 25 30
Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr
35 40 45
Ser Gly Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro
65 70 75 80
Ser Val Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
85 90 95
Phe Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
100 105 110
Tyr Cys Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys
<210> 8
<211> 726
<212> ДНК
<213> Искусственная последовательность
<220>
<223> v-область + CH1 константной области гамма-1 с сигнальной
последовательностью
<400> 8
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg 60
cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt 120
gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc 180
ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg 240
tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa 300
atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac 360
ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagcgct 420
tctacaaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 480
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 540
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 600
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 660
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 720
tcttgt 726
<210> 9
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> v-область + полноразмерная константная область гамма-1
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 10
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> v-область + полноразмерная константная область гамма-1
<400> 10
gaagtgcagt tggtggagtc gggtggaggg ctggtgcagc ctggcggtag cctgaggctg 60
tcctgtgccg tgtccggata ctccattacc tccggctact cgtggaactg gatcagacag 120
gctcccggaa agggacttga gtgggtggcg tccatcacct acgacggctc aaccaactat 180
aacccgtccg tgaagggccg catcaccatt tcgcgcgacg acagcaagaa tactttttac 240
ctccaaatga acagcctgcg ggccgaagat actgccgtgt actactgcgc gcggggatca 300
cattacttcg ggcactggca cttcgccgtc tggggacagg gcaccctcgt cactgtctcg 360
agcgcttcta caaagggccc ctccgtgttc ccgctcgctc catcatcgaa gtctaccagc 420
ggaggcactg cggctctcgg ttgcctcgtg aaggactact tcccggagcc ggtgaccgtg 480
tcgtggaaca gcggagccct gaccagcggg gtgcacacct ttccggccgt cttgcagtca 540
agcggccttt actccctgtc atcagtggtg actgtcccgt ccagctcatt gggaacccaa 600
acctacatct gcaatgtgaa tcacaaacct agcaacacca aggttgacaa gaaagtcgag 660
cccaaatcgt gtgacaagac tcacacttgt ccgccgtgcc cggcacccga actgctggga 720
ggtcccagcg tctttctgtt ccctccaaag ccgaaagaca cgctgatgat ctcccgcacc 780
ccggaggtca cttgcgtggt cgtggacgtg tcacatgagg acccagaggt gaagttcaat 840
tggtacgtgg atggcgtcga agtccacaat gccaaaacta agcccagaga agaacagtac 900
aattcgacct accgcgtcgt gtccgtgctc acggtgttgc atcaggattg gctgaacggg 960
aaggaataca agtgcaaagt gtccaacaag gcgctgccgg caccgatcga gaaaactatc 1020
tccaaagcga agggacagcc tagggaacct caagtctaca cgctgccacc atcacgggaa 1080
gaaatgacta agaatcaagt ctcactgact tgtctggtga aggggtttta ccctagcgac 1140
attgccgtgg agtgggaatc caacggccag ccagagaaca actacaagac tacccctcca 1200
gtgctcgact cggatggatc gttcttcctt tactcgaagc tcaccgtgga taagtcccgg 1260
tggcagcagg gaaacgtgtt ctcctgctcg gtgatgcatg aagccctcca taaccactat 1320
acccaaaagt cgctgtccct gtcgccggga aag 1353
<210> 11
<211> 469
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> v-область + полноразмерная константная область гамма-1 с сигнальной
последовательностью
<400> 11
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
20 25 30
Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr
35 40 45
Ser Gly Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro
65 70 75 80
Ser Val Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
85 90 95
Phe Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
100 105 110
Tyr Cys Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
245 250 255
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Lys
465
<210> 12
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> v-область + полноразмерная константная область гамма-1 с сигнальной
последовательностью
<400> 12
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg 60
cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt 120
gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc 180
ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg 240
tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa 300
atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac 360
ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagcgct 420
tctacaaagg gcccctccgt gttcccgctc gctccatcat cgaagtctac cagcggaggc 480
actgcggctc tcggttgcct cgtgaaggac tacttcccgg agccggtgac cgtgtcgtgg 540
aacagcggag ccctgaccag cggggtgcac acctttccgg ccgtcttgca gtcaagcggc 600
ctttactccc tgtcatcagt ggtgactgtc ccgtccagct cattgggaac ccaaacctac 660
atctgcaatg tgaatcacaa acctagcaac accaaggttg acaagaaagt cgagcccaaa 720
tcgtgtgaca agactcacac ttgtccgccg tgcccggcac ccgaactgct gggaggtccc 780
agcgtctttc tgttccctcc aaagccgaaa gacacgctga tgatctcccg caccccggag 840
gtcacttgcg tggtcgtgga cgtgtcacat gaggacccag aggtgaagtt caattggtac 900
gtggatggcg tcgaagtcca caatgccaaa actaagccca gagaagaaca gtacaattcg 960
acctaccgcg tcgtgtccgt gctcacggtg ttgcatcagg attggctgaa cgggaaggaa 1020
tacaagtgca aagtgtccaa caaggcgctg ccggcaccga tcgagaaaac tatctccaaa 1080
gcgaagggac agcctaggga acctcaagtc tacacgctgc caccatcacg ggaagaaatg 1140
actaagaatc aagtctcact gacttgtctg gtgaaggggt tttaccctag cgacattgcc 1200
gtggagtggg aatccaacgg ccagccagag aacaactaca agactacccc tccagtgctc 1260
gactcggatg gatcgttctt cctttactcg aagctcaccg tggataagtc ccggtggcag 1320
cagggaaacg tgttctcctg ctcggtgatg catgaagccc tccataacca ctatacccaa 1380
aagtcgctgt ccctgtcgcc gggaaag 1407
<210> 13
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR H1
<400> 13
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser
20 25
<210> 14
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 14
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp Asn
1 5 10
<210> 15
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR H2
<400> 15
Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
1 5 10
<210> 16
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 16
Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val Lys Gly
1 5 10 15
<210> 17
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR H3
<400> 17
Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 18
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 18
Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val
1 5 10
<210> 19
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR H4
<400> 19
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 20
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> V-область легкой цепи
<400> 20
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 21
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> V-область легкой цепи
<400> 21
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc 180
ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg 240
tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aag 333
<210> 22
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> V-область легкой цепи с сигнальной последовательностью
<400> 22
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 23
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> V-область легкой цепи с сигнальной последовательностью
<400> 23
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg 240
ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg 300
cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaag 387
<210> 24
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> V-область легкой цепи + константная область каппа
<400> 24
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 25
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> V-область + константная область каппа
<400> 25
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc 180
ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg 240
tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 654
<210> 26
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> V-область + константная область каппа с сигнальной последовательностью
<400> 26
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 27
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> V-область + константная область каппа с сигнальной последовательностью
<400> 27
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg 240
ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg 300
cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc 420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac 480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccctcca atcgggtaac 540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt 708
<210> 28
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L1
<400> 28
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys
20
<210> 29
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 29
Arg Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn
1 5 10 15
<210> 30
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L2
<400> 30
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 31
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 31
Ala Ala Ser Tyr Leu Glu Ser
1 5
<210> 32
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3
<400> 32
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 33
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 33
Gln Gln Ser His Glu Asp Pro Tyr Thr
1 5
<210> 34
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L4
<400> 34
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 35
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область
<400> 35
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Met
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 36
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область
<400> 36
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc 180
ggcgtgccta tgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aag 333
<210> 37
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область с сигнальной последовательностью
<400> 37
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 38
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область с сигнальной последовательностью
<400> 38
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg 240
cctatgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaag 387
<210> 39
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область + константная область каппа, включающая L154P
<400> 39
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Met
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 40
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область + константная область каппа, включающая L154P
<400> 40
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc 180
ggcgtgccta tgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cccgcaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 654
<210> 41
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область + константная область каппа, включающая L154P,
с сигнальной последовательностью
<400> 41
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 42
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S77R_Q79R_v-область + константная область каппа, включающая L154P,
с сигнальной последовательностью
<400> 42
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg 240
cctatgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc 420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac 480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccccgca atcgggtaac 540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt 708
<210> 43
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S60M
<400> 43
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 44
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S77R
<400> 44
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 45
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R Q79R
<400> 45
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 46
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S60M S77R
<400> 46
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 47
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S60M Q79R
<400> 47
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 48
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S77R Q79R
<400> 48
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 49
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S60M S77R Q79R
<400> 49
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 50
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2 S56D S52D
<400> 50
Ala Ala Asp Tyr Leu Glu Ser
1 5
<210> 51
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2 S56E S52E
<400> 51
Ala Ala Glu Tyr Leu Glu Ser
1 5
<210> 52
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S71M S67M
<400> 52
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 53
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S71M S60M S67M
<400> 53
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 54
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S71M S77R S67M
<400> 54
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 55
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S71M Q79R S67M
<400> 55
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 56
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S71M S60M S77R S67M
<400> 56
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 57
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S71M S60M Q79R S67M
<400> 57
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 58
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S71M S77R Q79R S67M
<400> 58
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 59
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S71M S60M S77R Q79R S67M
<400> 59
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 60
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S67Y S63Y
<400> 60
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 61
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S67Y S60M S63Y
<400> 61
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 62
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S67Y S77R S63Y
<400> 62
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 63
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S67Y Q79R S63Y
<400> 63
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 64
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S67Y S60M S77R S63Y
<400> 64
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 65
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S67Y S60M Q79R S63Y
<400> 65
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 66
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S67Y S77R Q79R S63Y
<400> 66
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 67
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S67Y S60M S77R Q79R S63Y
<400> 67
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 68
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S80N S76N
<400> 68
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 69
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S80N S60M S76N
<400> 69
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 70
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S80N S77R S76
<400> 70
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 71
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S80N Q79R S76N
<400> 71
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 72
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S80N S60M S77R S76N
<400> 72
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 73
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S80N S60M Q79R S76N
<400> 73
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 74
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S80N S77R Q79R S76N
<400> 74
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 75
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S80N S60M S77R Q79R S76N
<400> 75
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 76
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S80N S67Y S76N S63Y
<400> 76
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 77
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S80N S67Y S60M S76N S63Y
<400> 77
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 78
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S80N S67Y S77R S76N S63Y
<400> 78
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 79
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S80N S67Y Q79R S76N S63Y
<400> 79
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 80
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S80N S67Y S60M S77R S76N S63Y
<400> 80
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 81
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S80N S67Y S60M Q79R S76N S63Y
<400> 81
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 82
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S80N S67Y S77R Q79R S76N S63Y
<400> 82
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 83
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S80N S67Y S60M S77R Q79R S76N S63Y
<400> 83
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 84
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S67Y S71M S63Y S67M
<400> 84
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 85
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S67Y S71M S60M S63Y S67M
<400> 85
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 86
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S67Y S71M S77R S63Y S67M
<400> 86
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 87
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S67Y S71M Q79R S63Y S67M
<400> 87
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 88
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S67Y S71M S60M S77R S63Y S67M
<400> 88
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 89
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S67Y S71M S60M Q79R S63Y S67M
<400> 89
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 90
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S67Y S71M S77R Q79R S63Y S67M
<400> 90
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 91
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S67Y S71M S60M S77R Q79R S63Y S67M
<400> 91
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 92
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S80N S71M S76N S67M
<400> 92
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 93
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S80N S71M S60M S76N S67M (
<400> 93
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 94
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S80N S71M S77R S76N S67M
<400> 94
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 95
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S80N S71M Q79R S76N S67M
<400> 95
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 96
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S80N S71M S60M S77R S76N S67M
<400> 96
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 97
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S80N S71M S60M Q79R S76N S67M
<400> 97
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 98
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S80N S71M S77R Q79R S76N S67M
<400> 98
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 99
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S80N S71M S60M S77R Q79R S76N S67M
<400> 99
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 100
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S80N S67Y S71M S76N S63Y S67M
<400> 100
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 101
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S80N S67Y S71M S60M S76N S63Y S67M
<400> 101
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 102
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R S80N S67Y S71M S77R S76N S63Y S67M
<400> 102
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 103
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 Q83R S80N S67Y S71M Q79R S76N S63Y S67M
<400> 103
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 104
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R S80N S67Y S71M S60M S77R S76N S63Y S67M
<400> 104
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 105
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M Q83R S80N S67Y S71M S60M Q79R S76N S63Y S67M
<400> 105
Gly Val Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Ser Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 106
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S81R Q83R S80N S67Y S71M S77R Q79R S76N S63Y S67M
<400> 106
Gly Val Pro Ser Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 107
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR L3 S64M S81R Q83R S80N S67Y S71M S60M S77R Q79R S76N S63Y
S67M
<400> 107
Pro Met Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr
1 5 10 15
Ile Asn Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 108
<211> 325
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Человеческий IgE-Fc дикого типа (домены CE2-CE4 с нумерацией V224-K547
согласно Dorrington & Bennich (1978)
<400> 108
Val Ala Ser Arg Asp Phe Thr Pro Pro Thr Val Lys Ile Leu Gln Ser
1 5 10 15
Ser Cys Asp Gly Gly Gly His Phe Pro Pro Thr Ile Gln Leu Leu Cys
20 25 30
Leu Val Ser Gly Tyr Thr Pro Gly Thr Ile Asn Ile Thr Trp Leu Glu
35 40 45
Asp Gly Gln Val Met Asp Val Asp Leu Ser Thr Ala Ser Thr Thr Gln
50 55 60
Glu Gly Glu Leu Ala Ser Thr Gln Ser Glu Leu Thr Leu Ser Gln Lys
65 70 75 80
His Trp Leu Ser Asp Arg Thr Tyr Thr Cys Gln Val Thr Tyr Gln Gly
85 90 95
His Thr Phe Glu Asp Ser Thr Lys Lys Cys Ala Asp Ser Asn Pro Arg
100 105 110
Gly Val Ser Ala Tyr Leu Ser Arg Pro Ser Pro Phe Asp Leu Phe Ile
115 120 125
Arg Lys Ser Pro Thr Ile Thr Cys Leu Val Val Asp Leu Ala Pro Ser
130 135 140
Lys Gly Thr Val Asn Leu Thr Trp Ser Arg Ala Ser Gly Lys Pro Val
145 150 155 160
Asn His Ser Thr Arg Lys Glu Glu Lys Gln Arg Asn Gly Thr Leu Thr
165 170 175
Val Thr Ser Thr Leu Pro Val Gly Thr Arg Asp Trp Ile Glu Gly Glu
180 185 190
Thr Tyr Gln Cys Arg Val Thr His Pro His Leu Pro Arg Ala Leu Met
195 200 205
Arg Ser Thr Thr Lys Thr Ser Gly Pro Arg Ala Ala Pro Glu Val Tyr
210 215 220
Ala Phe Ala Thr Pro Glu Trp Pro Gly Ser Arg Asp Lys Arg Thr Leu
225 230 235 240
Ala Cys Leu Ile Gln Asn Phe Met Pro Glu Asp Ile Ser Val Gln Trp
245 250 255
Leu His Asn Glu Val Gln Leu Pro Asp Ala Arg His Ser Thr Thr Gln
260 265 270
Pro Arg Lys Thr Lys Gly Ser Gly Phe Phe Val Phe Ser Arg Leu Glu
275 280 285
Val Thr Arg Ala Glu Trp Glu Gln Lys Asp Glu Phe Ile Cys Arg Ala
290 295 300
Val His Glu Ala Ala Ser Pro Ser Gln Thr Val Gln Arg Ala Val Ser
305 310 315 320
Val Asn Pro Gly Lys
325
<210> 109
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S77R_Q79R_v-область (Fab1)
<400> 109
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 110
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 77R_Q79R_v-область (Fab1)
<400> 110
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtac 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctccta ccttgaaagc 180
ggcgtgcctt cacgcttctc gggaagcggg tctggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggtaccaa ggtcgaaatc aag 333
<210> 111
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S77R_Q79R_v-область (Fab1) с сигнальной последовательностью
<400> 111
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 112
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S77R_Q79R_v-область (Fab1) с сигнальной последовательностью
<400> 112
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtaccagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cctaccttga aagcggcgtg 240
ccttcacgct tctcgggaag cgggtctggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggta ccaaggtcga aatcaag 387
<210> 113
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область
<400> 113
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 114
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область
<400> 114
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc 180
ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg 240
tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aag 333
<210> 115
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область с сигнальной последовательностью
<400> 115
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 116
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область с сигнальной последовательностью
<400> 116
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg 240
ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg 300
cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaag 387
<210> 117
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область + константная область каппа, включающая L154P
<400> 117
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 118
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область + константная область каппа, включающая L154P
<400> 118
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctcata ccttgaaagc 180
ggcgtgcctt cgcgcttctc gggaagcggg tcgggcaccg atttcaccct gaccatttcg 240
tcgctgcagc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cccgcaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 654
<210> 119
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область + константная область каппа, включающая L154P,
с сигнальной последовательностью
<400> 119
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 120
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab2_v-область + константная область каппа, включающая L154P,
с сигнальной последовательностью
<400> 120
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cataccttga aagcggcgtg 240
ccttcgcgct tctcgggaag cgggtcgggc accgatttca ccctgaccat ttcgtcgctg 300
cagccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc 420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac 480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccccgca atcgggtaac 540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt 708
<210> 121
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область
<400> 121
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 122
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область
<400> 122
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtac 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctccta ccttgaaagc 180
ggcgtgcctt cacgcttctc gggaagcggg tctggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggtaccaa ggtcgaaatc aag 333
<210> 123
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область с сигнальной последовательностью
<400> 123
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 124
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область с сигнальной последовательностью
<400> 124
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtaccagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cctaccttga aagcggcgtg 240
ccttcacgct tctcgggaag cgggtctggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggta ccaaggtcga aatcaag 387
<210> 125
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область + константная область каппа, включающая L154P
<400> 125
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 126
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область + константная область каппа, включающая L154P
<400> 126
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtac 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgcctccta ccttgaaagc 180
ggcgtgcctt cacgcttctc gggaagcggg tctggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggtaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cccgcaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 654
<210> 127
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область + константная область каппа, включающая L154P,
с сигнальной последовательностью
<400> 127
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 128
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab3_S77R_Q79R_v-область + константная область каппа, включающая L154P,
с сигнальной последовательностью
<400> 128
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtaccagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgcct cctaccttga aagcggcgtg 240
ccttcacgct tctcgggaag cgggtctggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggta ccaaggtcga aatcaagcgt acggtagcgg ccccatctgt cttcatcttc 420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac 480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccccgca atcgggtaac 540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgt 708
<210> 129
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Омализумаб_v-область
<400> 129
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
1 5 10 15
Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Tyr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His Glu Asp Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 130
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Омализумаб_v-область
<400> 130
actcagagcc cgagctcact ctccgcttcc gtgggggata gagtgaccat cacttgccgg 60
gcatcccagt cggtggacta cgacggagac tcctacatga actggtatca gcagaagccc 120
ggaaaagccc caaagttgct gatctacgcc gcctcatacc ttgaaagcgg cgtgccttcg 180
cgcttctcgg gaagcgggtc gggcaccgat ttcaccctga ccatttcgtc gctgcagccg 240
gaggacttcg cgacttacta ctgccaacag tcccacgagg acccctatac gtttggccag 300
ggaaccaagg tcgaaatcaa g 321
<210> 131
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR3
<400> 131
Gly Val Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 132
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_v-область
<400> 132
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Met
50 55 60
Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 133
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_v-область
<400> 133
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgccgacta ccttgaaagc 180
ggcgtgccta tgcgcttctc gggaagcggg atgggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aag 333
<210> 134
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью
<400> 134
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Met Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 135
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью
<400> 135
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgccg actaccttga aagcggcgtg 240
cctatgcgct tctcgggaag cgggatgggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaag 387
<210> 136
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_v-область + константная область каппа
<400> 136
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Met
50 55 60
Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 137
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_v-область + константная область каппа
<400> 137
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgccgacta ccttgaaagc 180
ggcgtgccta tgcgcttctc gggaagcggg atgggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 654
<210> 138
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FR3
<400> 138
Gly Val Pro Arg Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 139
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60R_S52D_S67M_S77R_Q79R_v-область
<400> 139
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Arg
50 55 60
Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 140
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60R_S52D_S67M_S77R_Q79R_v-область
<400> 140
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgccgatta ccttgaaagc 180
ggcgtgcctc gtcgcttctc gggaagcggg atgggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aag 333
<210> 141
<211> 129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60R_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью
<400> 141
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
20 25 30
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp
35 40 45
Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val
65 70 75 80
Pro Arg Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ser His Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys
<210> 142
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60R_S52D_S67M_S77R_Q79R_v-область с сигнальной последовательностью
<400> 142
atgaagtggg tcaccttcat ctccctgctg tttctgttct cctccgccta ctccgatatt 60
cagctgactc agagcccgag ctcactctcc gcttccgtgg gggatagagt gaccatcact 120
tgccgggcat cccagtcggt ggactacgac ggagactcct acatgaactg gtatcagcag 180
aagcccggaa aagccccaaa gttgctgatc tacgccgccg attaccttga aagcggcgtg 240
cctcgtcgct tctcgggaag cgggatgggc accgatttca ccctgaccat ttcgagactg 300
aggccggagg acttcgcgac ttactactgc caacagtccc acgaggaccc ctatacgttt 360
ggccagggaa ccaaggtcga aatcaag 387
<210> 143
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60R_S52D_S67M_S77R_Q79R_v-область + константная область каппа
<400> 143
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Arg
50 55 60
Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 144
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S60R_S52D_S67M_S77R_Q79R_v-область + константная область каппа
<400> 144
gatattcagc tgactcagag cccgagctca ctctccgctt ccgtggggga tagagtgacc 60
atcacttgcc gggcatccca gtcggtggac tacgacggag actcctacat gaactggtat 120
cagcagaagc ccggaaaagc cccaaagttg ctgatctacg ccgccgatta ccttgaaagc 180
ggcgtgcctc gtcgcttctc gggaagcggg atgggcaccg atttcaccct gaccatttcg 240
agactgaggc cggaggactt cgcgacttac tactgccaac agtcccacga ggacccctat 300
acgtttggcc agggaaccaa ggtcgaaatc aagcgtacgg tagcggcccc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 654
<210> 145
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60K_S52D_S67M_S77R_Q79R_v-область
<400> 145
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Lys
50 55 60
Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 146
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60Q_S52D_S67M_S77R_Q79R_v-область
<400> 146
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Gln
50 55 60
Arg Phe Ser Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 147
<211> 506
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> v-область + CH1 константной области гамма-1 плюс линкер плюс CA645
gL4gH5 scFv с сигнальной последовательностью
<400> 147
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
20 25 30
Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr
35 40 45
Ser Gly Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro
65 70 75 80
Ser Val Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
85 90 95
Phe Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
100 105 110
Tyr Cys Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys Ser Gly Gly Gly Gly Thr Gly Gly Gly Gly Ser Glu Val Gln
245 250 255
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
260 265 270
Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
275 280 285
Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile Gly Ile Ile
290 295 300
Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly Arg Phe
305 310 315 320
Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln Met Asn
325 330 335
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val
340 345 350
Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln
385 390 395 400
Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
405 410 415
Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp Tyr Gln
420 425 430
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala Ser Lys
435 440 445
Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
450 455 460
Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr
465 470 475 480
Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr Phe Gly
485 490 495
Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
500 505
<210> 148
<211> 1518
<212> ДНК
<213> Искусственная последовательность
<220>
<223> v-область + CH1 константной области гамма-1 плюс линкер плюс CA645
gL4gH5 scFv с сигнальной последовательностью
<400> 148
atgaagtggg tcaccttcat ctccctgctg tttctgttct ccagcgccta ctccgaagtg 60
cagttggtgg agtcgggtgg agggctggtg cagcctggcg gtagcctgag gctgtcctgt 120
gccgtgtccg gatactccat tacctccggc tactcgtgga actggatcag acaggctccc 180
ggaaagggac ttgagtgggt ggcgtccatc acctacgacg gctcaaccaa ctataacccg 240
tccgtgaagg gccgcatcac catttcgcgc gacgacagca agaatacttt ttacctccaa 300
atgaacagcc tgcgggccga agatactgcc gtgtactact gcgcgcgggg atcacattac 360
ttcgggcact ggcacttcgc cgtctgggga cagggcaccc tcgtcactgt ctcgagcgct 420
tctacaaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 480
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccagtgac ggtgtcgtgg 540
aactcaggtg ccctgaccag cggcgttcac accttcccgg ctgtcctaca gtcttcagga 600
ctctactccc tgagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 660
atctgcaacg tgaatcacaa gcccagcaac accaaggtcg ataagaaagt tgagcccaaa 720
tcttgtagtg gaggtggggg caccggtgga ggtggcagcg aggttcaact gcttgagtct 780
ggaggaggcc tagtccagcc tggagggagc ctgcgtctct cttgtgcagt aagcggcatc 840
gacctgagca attacgccat caactgggtg agacaagctc cggggaagtg tttagaatgg 900
atcggtataa tatgggccag tgggacgacc ttttatgcta catgggcgaa aggaaggttt 960
acaattagcc gggacaatag caaaaacacc gtgtatctcc aaatgaactc cttgcgagca 1020
gaggacacgg cggtgtacta ttgtgctcgc actgtcccag gttatagcac tgcaccctac 1080
ttcgatctgt ggggacaagg gaccctggtg actgtttcaa gtggcggagg gggtagtgga 1140
gggggtggct ctgggggtgg cggaagcggt ggcgggggtt ctgacataca aatgactcag 1200
tctccttcat cggtatccgc gtccgttggc gatagggtga ctattacatg tcaaagctct 1260
cctagcgtct ggagcaattt tctatcctgg tatcaacaga aaccggggaa ggctccaaaa 1320
cttctgattt atgaagcctc gaaactcacc agtggagttc cgtcaagatt cagtggctct 1380
ggatcaggga cagacttcac gttgacaatc agttcgctgc aaccagagga ctttgcgacc 1440
tactattgtg gtggaggtta cagtagcata agtgatacga catttgggtg cggtactaag 1500
gtggaaatca aacgtacc 1518
<210> 149
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер между CH1 и CA645 gL4gH5 scFv
<400> 149
Ser Gly Gly Gly Gly Thr Gly Gly Gly Gly Ser
1 5 10
<210> 150
<211> 253
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 gL4gH5 scFv
<400> 150
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln
130 135 140
Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val
145 150 155 160
Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu
180 185 190
Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr
225 230 235 240
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
245 250
<210> 151
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер между CA645 gH5 с CA645 gL4 в scFv
<400> 151
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 152
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 CDRH1
<400> 152
Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
1 5 10
<210> 153
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 CDRH2
<400> 153
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
1 5 10 15
<210> 154
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 CDRH3
<400> 154
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu
1 5 10
<210> 155
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 CDRL1
<400> 155
Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
1 5 10
<210> 156
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 CDRL2
<400> 156
Glu Ala Ser Lys Leu Thr Ser
1 5
<210> 157
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 CDRL3
<400> 157
Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
1 5 10
<210> 158
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_S63W_S76N_v-область
<400> 158
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Met
50 55 60
Arg Phe Trp Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Asn
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 159
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> S60M_S52D_S67M_S77R_Q79R_S63Y_S76N_v-область
<400> 159
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Asp Tyr Leu Glu Ser Gly Val Pro Met
50 55 60
Arg Phe Tyr Gly Ser Gly Met Gly Thr Asp Phe Thr Leu Thr Ile Asn
65 70 75 80
Arg Leu Arg Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 160
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сигнальная последовательность
<400> 160
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВНЕКЛЕТОЧНЫЙ ДОМЕН АЛЬФА-СУБЪЕДИНИЦЫ FC-РЕЦЕПТОРА IGE, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЕГО, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2019 |
|
RU2796162C2 |
| Антитела против ингибитора пути тканевого фактора и их применения | 2016 |
|
RU2815681C1 |
| ДИМЕР ПОЛИПЕПТИДА С ВЫСОКИМ СОДЕРЖАНИЕМ СИАЛОВОЙ КИСЛОТЫ, ВКЛЮЧАЮЩЕГО ЭКСТРАКЛЕТОЧНЫЙ ДОМЕН АЛЬФА-СУБЪЕДИНИЦЫ РЕЦЕПТОРА ДЛЯ Fc-ОБЛАСТИ IgE, И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2833598C2 |
| CD47-АНТИГЕНСВЯЗЫВАЮЩАЯ ЕДИНИЦА И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2780859C2 |
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПРОБИОТИКИ И ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ АФФИННОСТЬЮ СВЯЗЫВАНИЯ В ОТНОШЕНИИ IGE, И ЕЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2786578C2 |
| Композиции и способы лечения и предупреждения инфекций, вызванных Staphylococcus aureus | 2015 |
|
RU2764981C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| СЛИТАЯ С Fc α-ЦЕПЬ ВЫСОКОАФФИННОГО РЕЦЕПТОРА IgE | 2016 |
|
RU2715606C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ ПРОТИВ TIGIT И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2776714C2 |
| CD131-СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773927C2 |
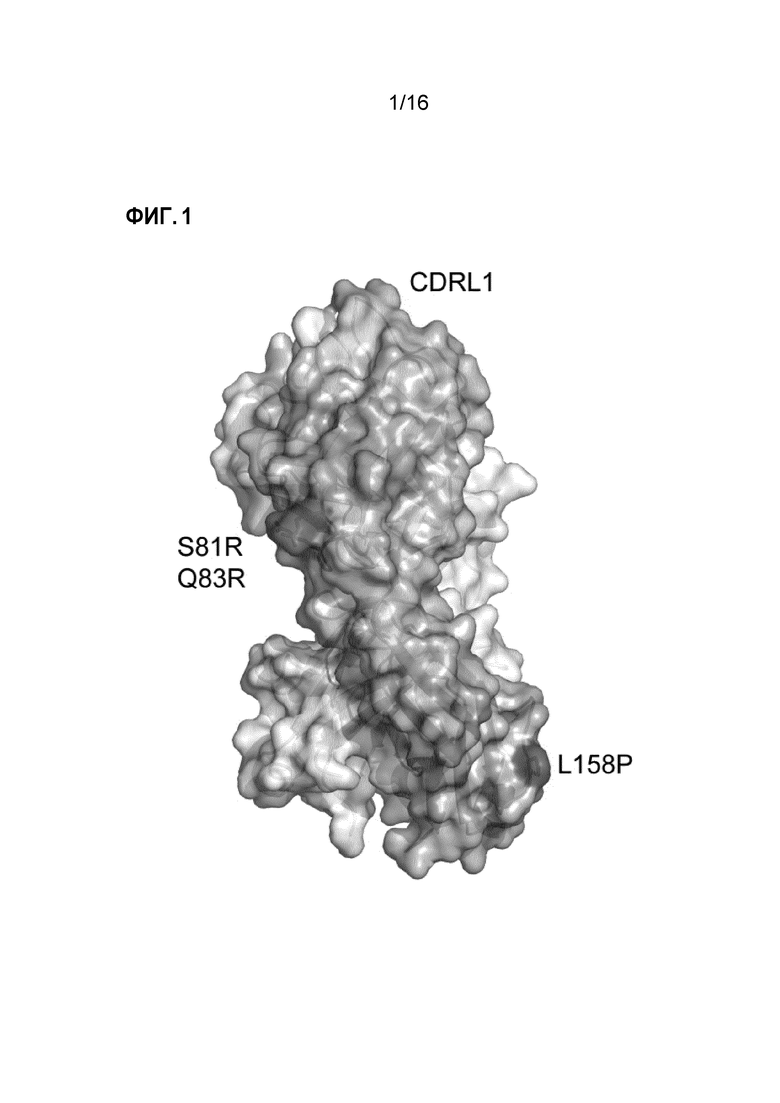
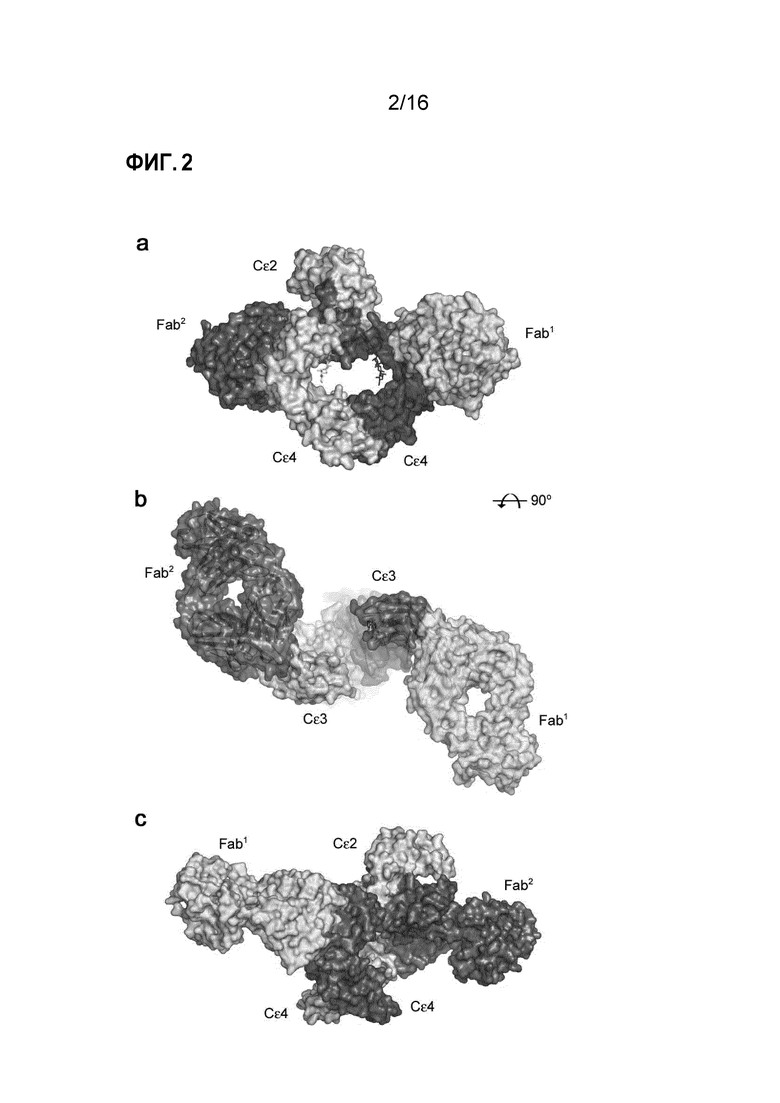
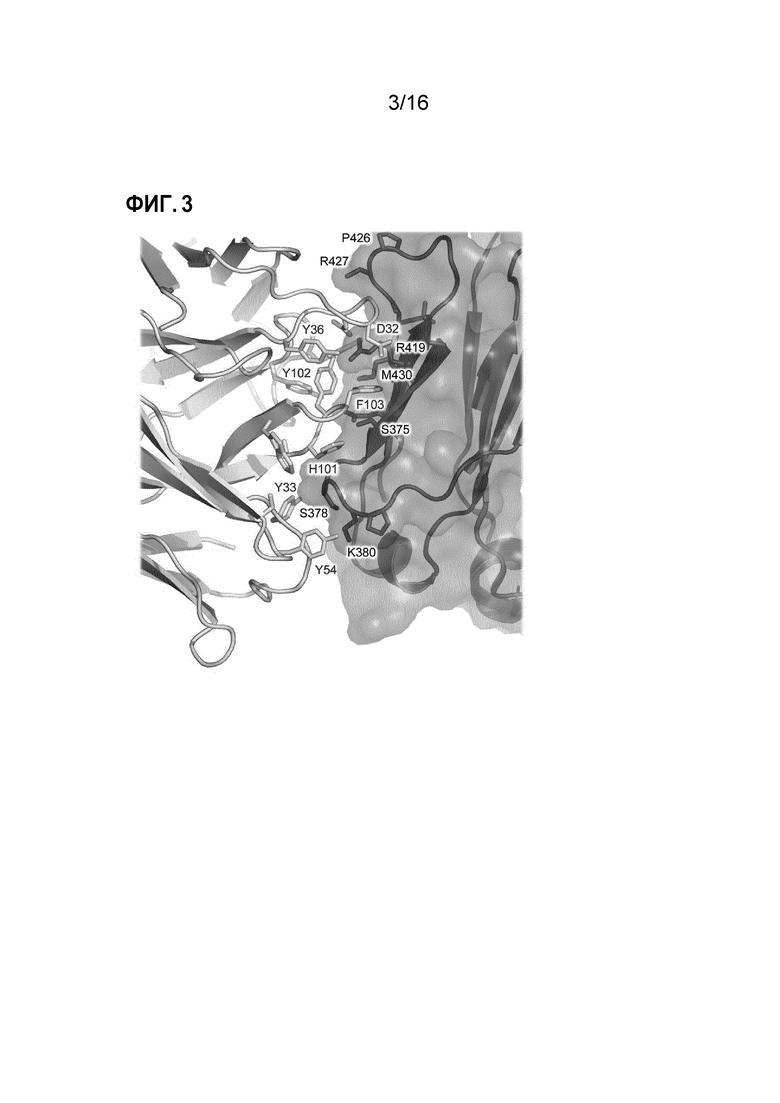
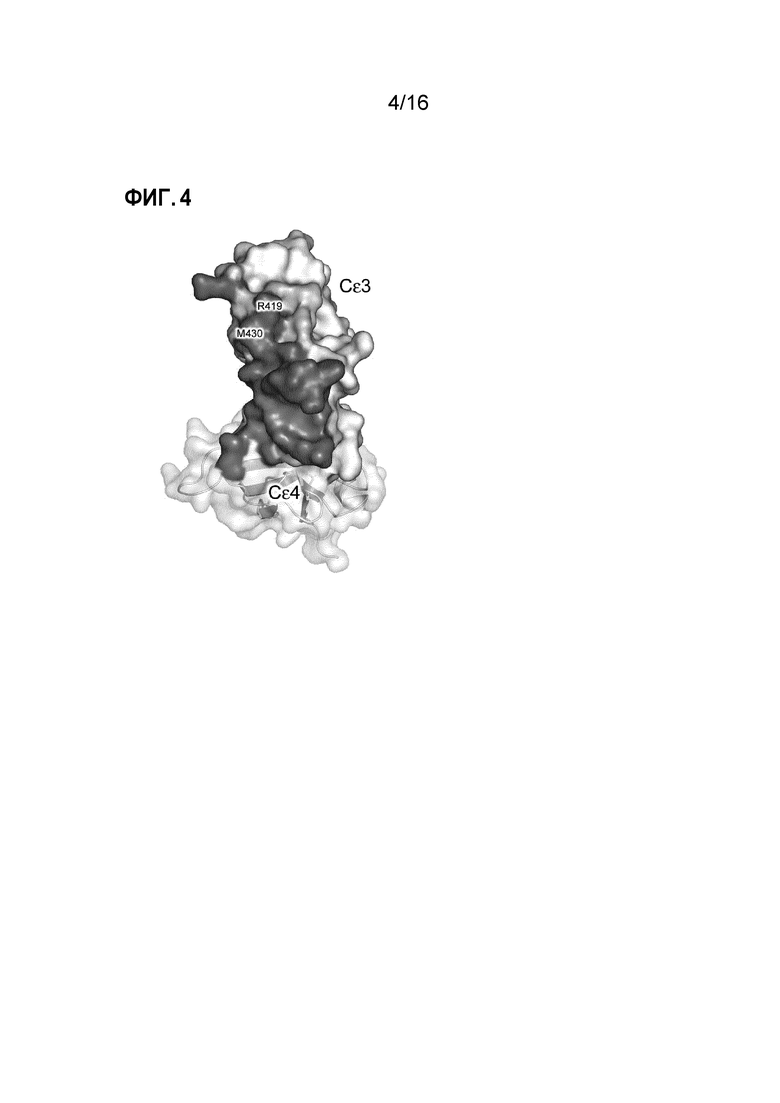
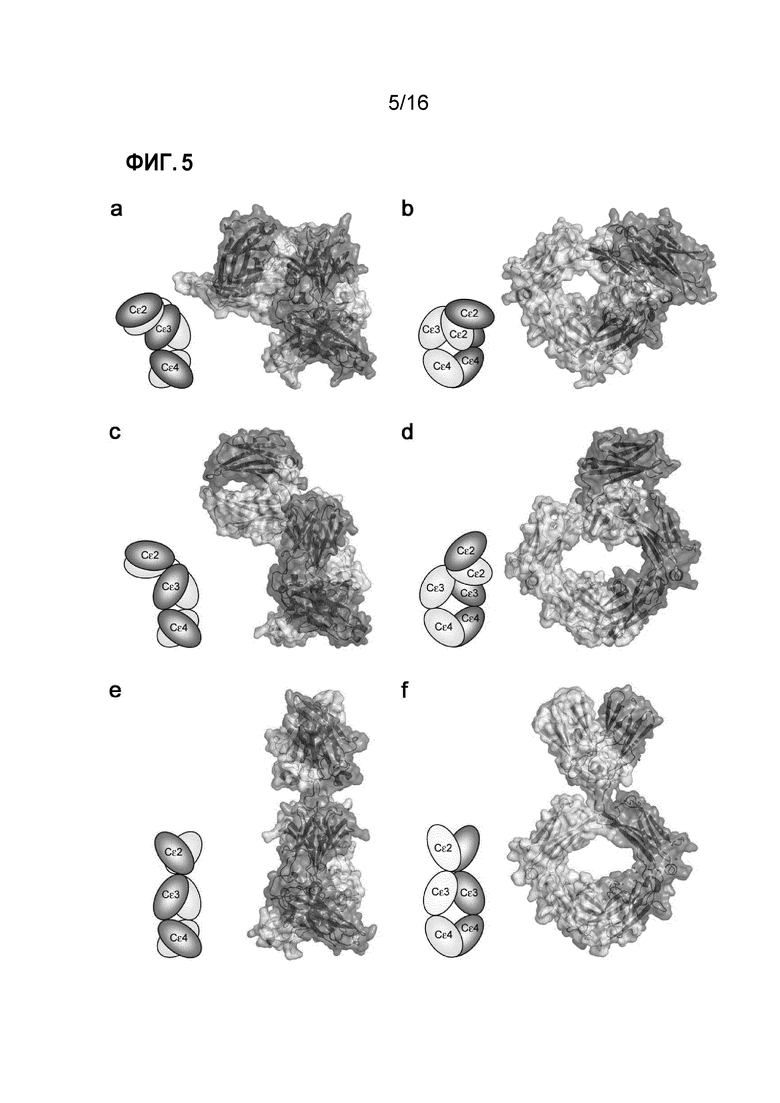
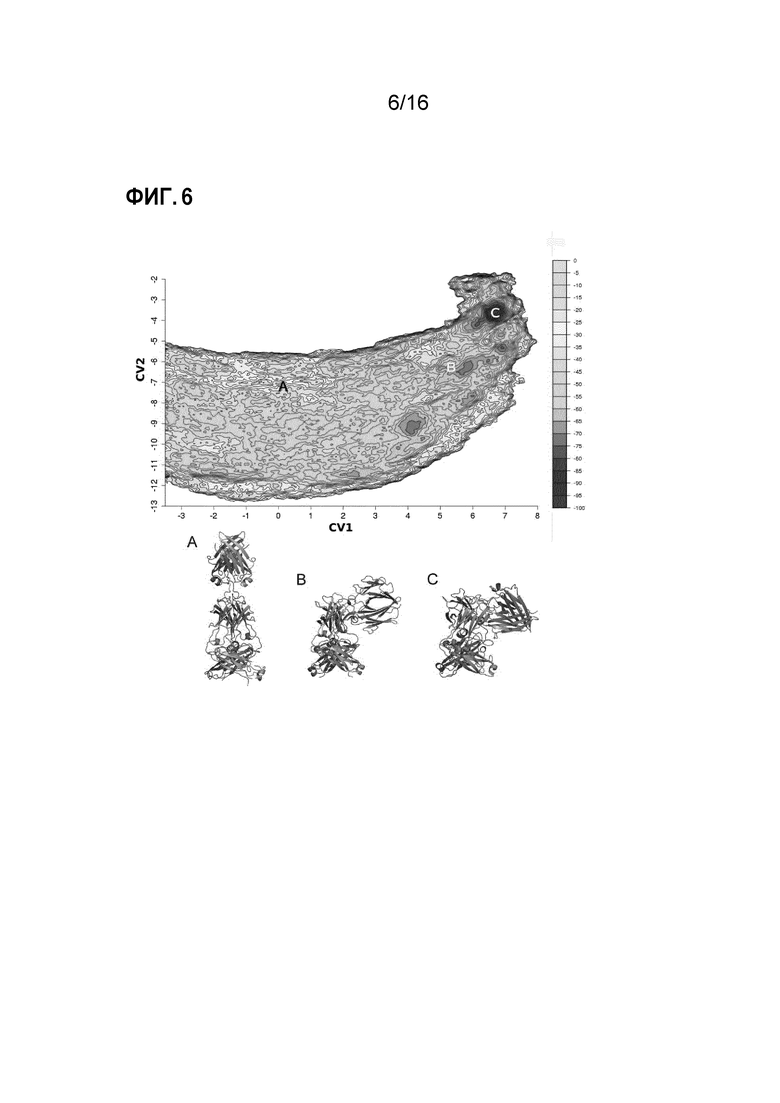
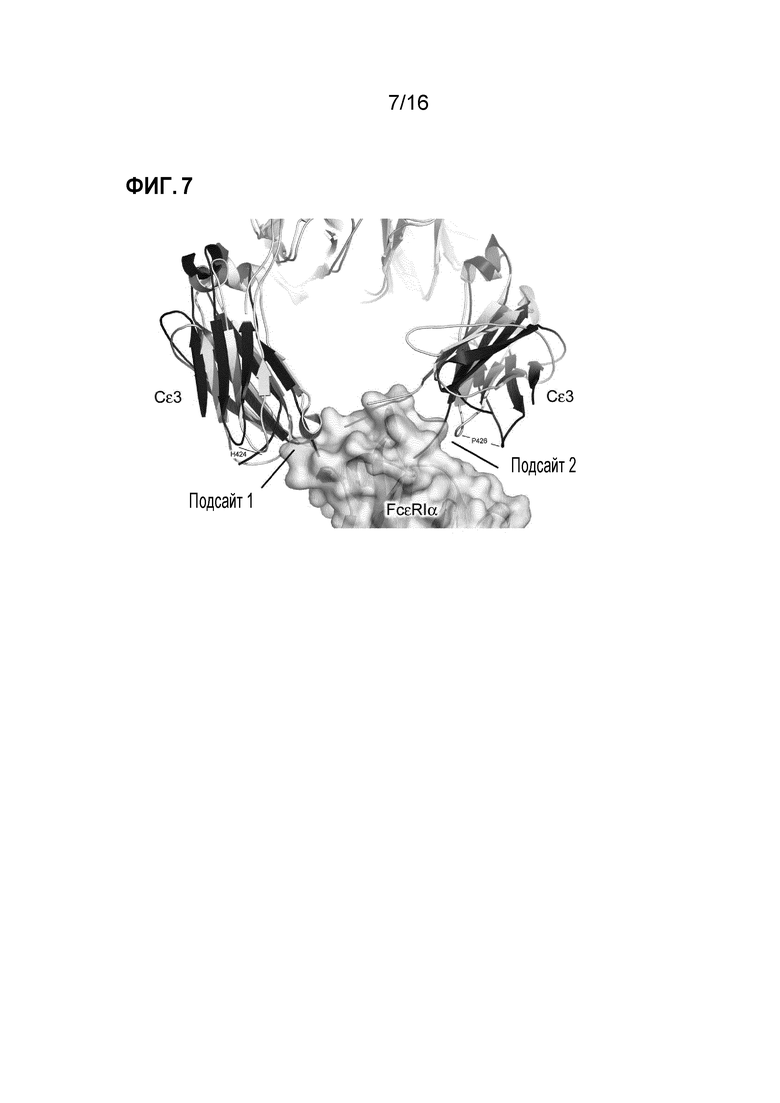
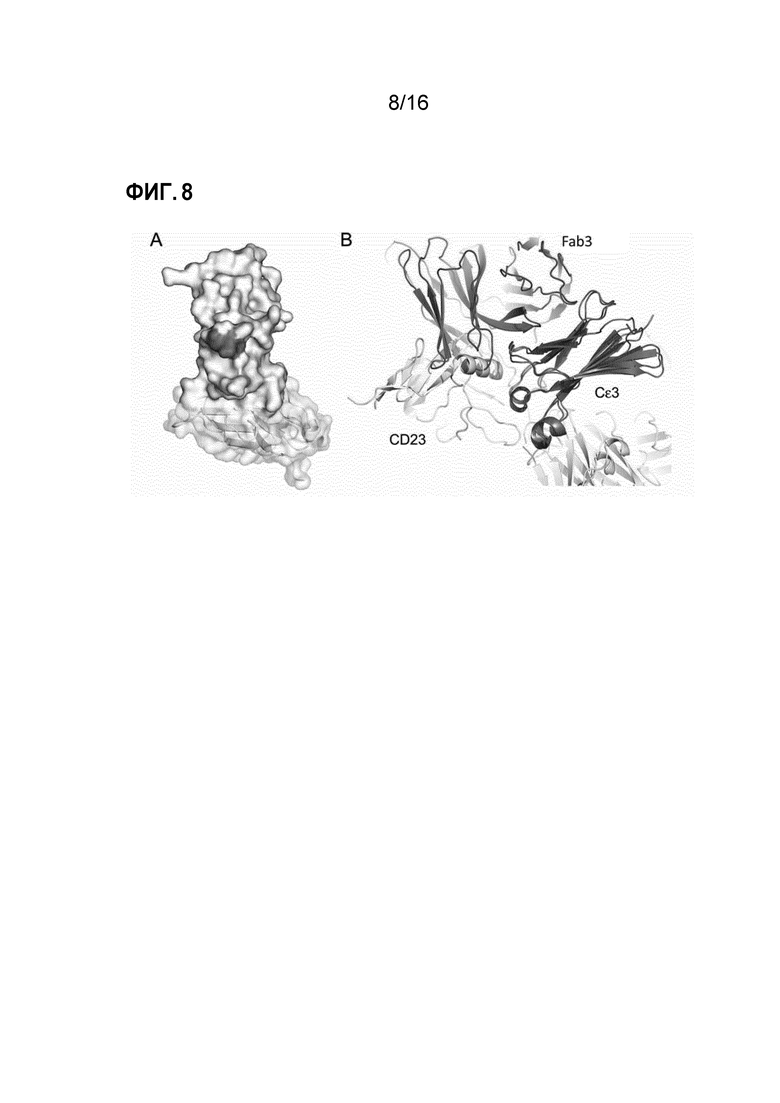
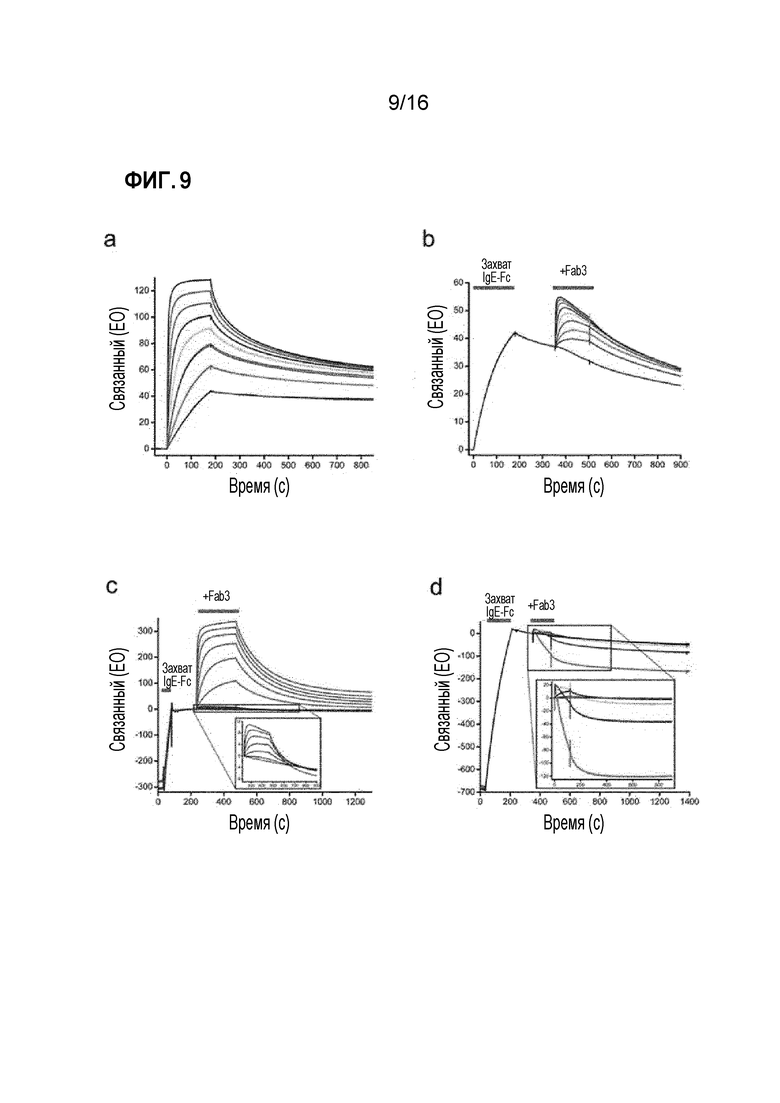
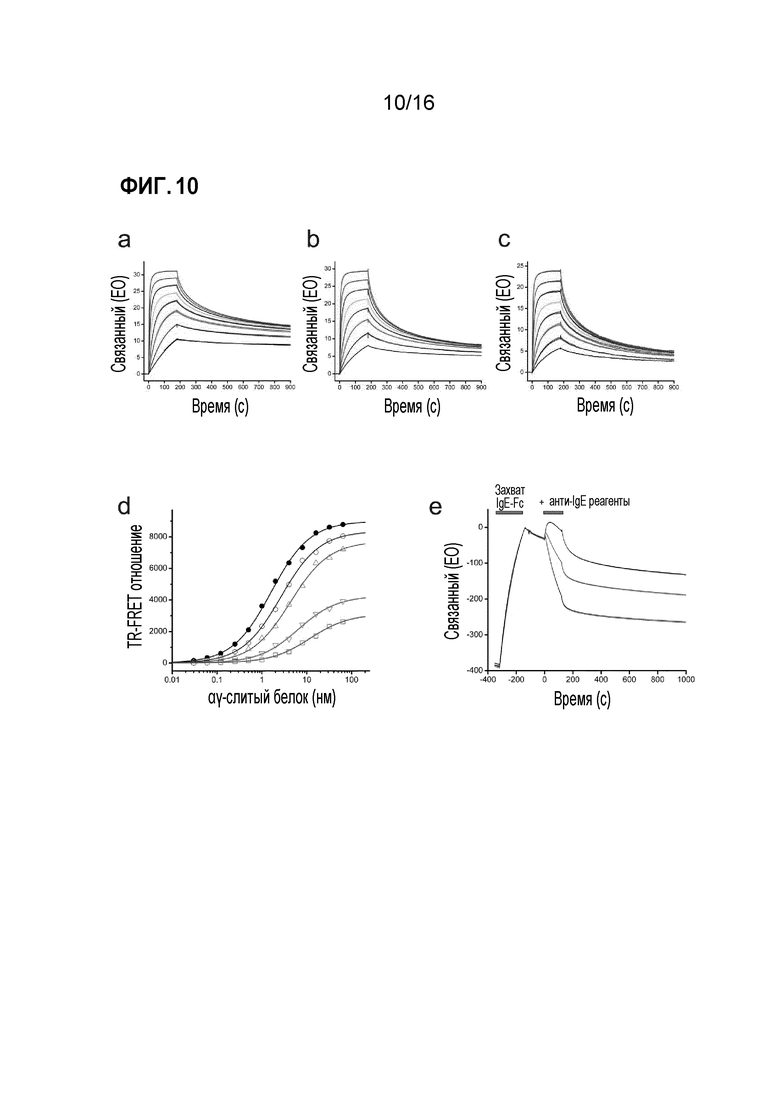

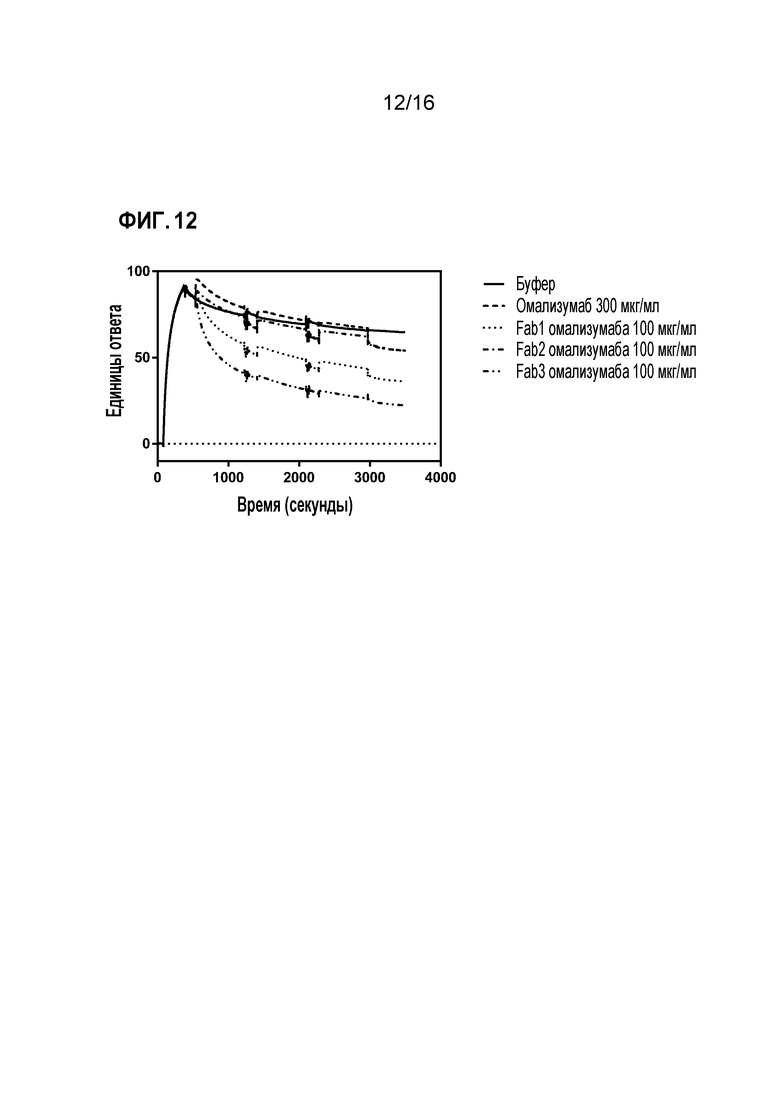
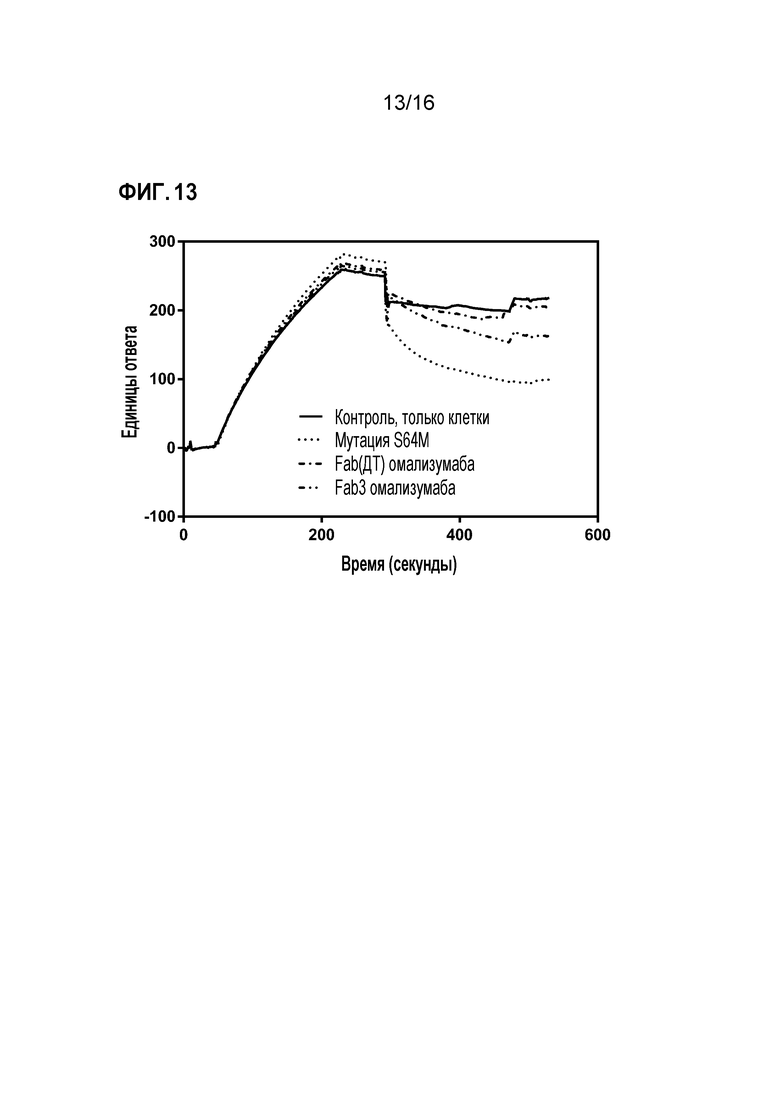
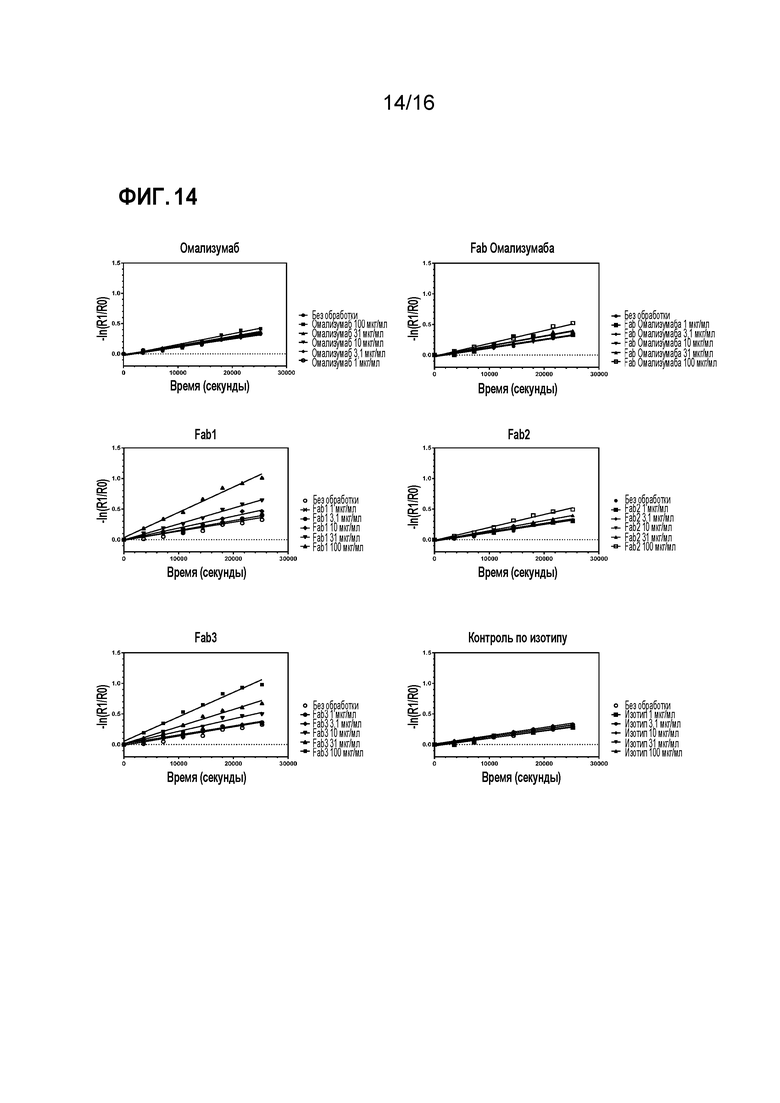
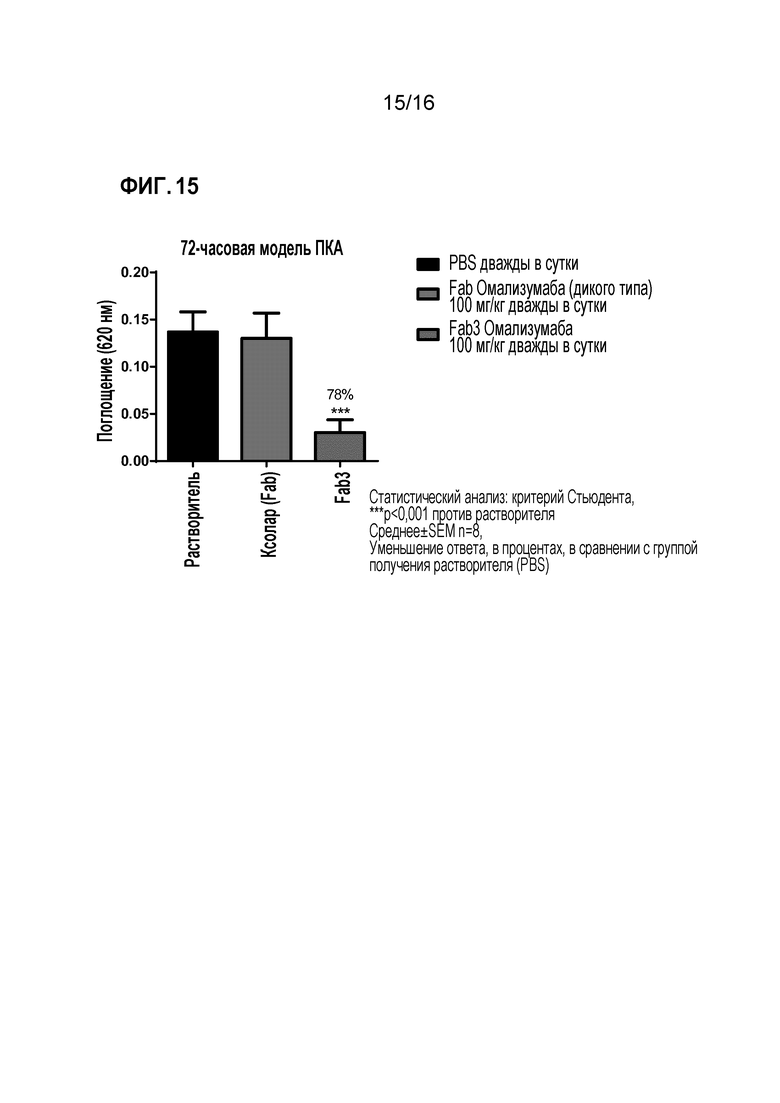

Изобретение относится к области биохимии, в частности к анти-IgE антителу или антигенсвязывающему агенту. Также раскрыты выделенная молекула ДНК, кодирующая указанное антитело; вектор для клонирования или экспрессии указанного антитела; фармацевтическая композиция, содержащая указанное антитело. Раскрыто применение указанных антитела и композиции для лечения. Изобретение позволяет эффективно лечить заболевания с помощью указанного анти-IgE антитела. 8 н. и 4 з.п. ф-лы, 16 ил., 13 табл., 8 пр.
1. Анти-IgE антитело, или антигенсвязывающий агент, содержащее
вариабельную область тяжелой цепи, содержащую
определяющую комплементарность область, CDR-H1, с аминокислотной последовательностью SEQ ID NO:14,
CDR-H2 с аминокислотной последовательностью SEQ ID NO:16,
CDR-H3, с аминокислотной последовательностью SEQ ID NO:18,
и
вариабельную область легкой цепи, содержащую
определяющую комплементарность область, CDR-L1, с аминокислотной последовательностью SEQ ID NO:29, CDR-L2, с аминокислотной последовательностью SEQ ID NO:50 или SEQ ID NO:51, CDR-L3, с аминокислотной последовательностью SEQ ID NO:33;
где вариабельная область легкой цепи также содержит каркасную область, FR-L3, с аминокислотной последовательностью, SEQ ID NO:32, которая имеет одну, две, три, четыре, пять, шесть, семь или более аминокислотных замен для усиления взаимодействия анти-IgE антитела, или антигенсвязывающего агента, с Сε2-доменом IgE человека; и где:
a. область FR-L3 имеет мутацию в положении S60 (Kabat) на М, R, К, N, Q или Т;
b. область FR-L3 имеет мутацию в положении S67 (Kabat) на М;
c. область FR-L3 имеет мутацию в положении S77 (Kabat) на R;
d. область FR-L3 имеет мутацию в положении Q79 (Kabat) на R.
2. Анти-IgE антитело по п.1, где scFv содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, предпочтительно, связанные через линкер с последовательностью SEQ ID NO:151,
где вариабельная область тяжелой цепи содержит
CDR-H1 с аминокислотной последовательностью SEQ ID NO:152,
CDR-H2 с аминокислотной последовательностью SEQ ID NO:153 и CDR-H3 с аминокислотной последовательностью SEQ ID NO:154,
и
вариабельная область легкой цепи содержит
CDR-L1 с аминокислотной последовательностью SEQ ID NO:155, CDR-L2 с аминокислотной последовательностью SEQ ID NO:156, CDR-L3 с аминокислотной последовательностью SEQ ID NO:157.
3. Анти-IgE антитело по п.1 или 2, где Fab-фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
a. вариабельная область тяжелой цепи содержит
CDR-H1 с аминокислотной последовательностью SEQ ID NO:14, CDR-H2 с аминокислотной последовательностью SEQ ID NO:16 и CDR-H3 с аминокислотной последовательностью SEQ ID NO:18, и вариабельная область легкой цепи содержит
CDR-L1 с аминокислотной последовательностью SEQ ID NO:29, CDR-L2 с аминокислотной последовательностью SEQ ID NO:50, CDR-L3 с аминокислотной последовательностью SEQ ID NO:33 и каркасную область FW-L3 с аминокислотной
последовательностью SEQ ID NO:131 или 138; или
b. вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:1 и
вариабельная область легкой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO:132 или 139.
4. Анти-IgE антитело, или антигенсвязывающий агент, которое выбрано из группы, состоящей из: Fab-фрагмента, модифицированного Fab'-фрагмента, Fab'-фрагмента, F(ab')2- фрагмента, Fv, scFv, scab, диатела, биспецифического антитела, триатела, FabFv, Fab-Fv-Fv, триотела или (Fab-Fv) 2-Fc.
5. Анти-IgE антитело по п.1, которое представляет собой Fab-фрагмент, связанный непосредственно или через линкер с scFv, который связывается с сывороточным белком-носителем, таким как сывороточный альбумин человека.
6. Выделенная молекула ДНК, кодирующая тяжелую и/или легкую цепь(и) анти-IgE антитела, или антигенсвязывающего агента, по пп.1-5.
7. Клонирующий вектор для клонирования антитела по п.1, содержащий одну или несколько последовательностей ДНК, выбранных из: SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40 или SEQ ID NO:42, или SEQ ID NO:133, или SEQ ID NO:135, или SEQ ID NO:137, или SEQ ID NO:140, или SEQ ID NO:142, или SEQ ID NO:144;
где, необязательно, клонирующий вектор также содержит одну или несколько последовательностей ДНК, выбранных из: SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10 или SEQ ID NO:12, или SEQ ID NO:148.
8. Вектор экспрессии для экспрессии антител по п.1, содержащий одну или несколько последовательностей ДНК, выбранных из: SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40 или SEQ ID NO:42, или SEQ ID NO:133, или SEQ ID NO:135, или SEQ ID NO:137, или SEQ ID NO:140, или SEQ ID NO:142, или SEQ ID NO:144;
где, необязательно, вектор экспрессии также содержит одну или несколько последовательностей ДНК, выбранных из: SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10 или SEQ ID NO:12, или SEQ ID NO:148.
9. Фармацевтическая композиция для лечения или профилактики одного или нескольких заболеваний: аллергии; аллергической астмы; тяжелой астмы; умеренной астмы; хронической спонтанной крапивницы; хронической идиопатической крапивницы; продолжительного аллергического ринита; сезонного аллергического ринита; обострений астмы; острого бронхоспазма; астматического статуса; гипер-IgE синдрома; аллергического бронхолегочного аспергиллеза; профилактики анафилактических реакций; пищевой аллергии; атопического дерматита; аллергического ринита; аллергии на пчелиный яд; идиопатической анафилаксии; аллергии на арахис; аллергии на латекс; воспалительных кожных заболеваний; крапивницы (вызываемой солнечным светом, холодом, локальным нагреванием и/или длительным сдавливанием); кожного мастоцитоза; системного мастоцитоза; эозинофил-связанного желудочно-кишечного заболевания; буллезного пемфигоида; интерстициального цистита; полипов носовой полости; идиопатического ангионевротического отека или неаллергической астмы, где антитело содержит анти-IgE антитело, или антигенсвязывающий агент, по пп.1-5, в комбинации с одним или несколькими фармацевтически приемлемыми эксципиентами, разбавителями или носителями.
10. Применение анти-IgE антитела, или антигенсвязывающего агента, по пп.1-5 в качестве лекарственного средства для лечения заболевания, которое облегчается за счет модулирования биологической активности IgE человека.
11. Применение анти-IgE антитела, или антигенсвязывающего агента, по пп.1-5 для лечения или профилактики одного или нескольких заболеваний: аллергии; аллергической астмы; тяжелой астмы; умеренной астмы; хронической спонтанной крапивницы; хронической идиопатической крапивницы; продолжительного аллергического ринита; сезонного аллергического ринита; обострений астмы; острого бронхоспазма; астматического статуса; гипер-IgE синдрома; аллергического бронхолегочного аспергиллеза; профилактики анафилактических реакций; пищевой аллергии; атопического дерматита; аллергического ринита; аллергии на пчелиный яд; идиопатической анафилаксии; аллергии на арахис; аллергии на латекс; воспалительных кожных заболеваний; крапивницы (вызываемой солнечным светом, холодом, локальным нагреванием и/или длительным сдавливанием); кожного мастоцитоза; системного мастоцитоза; эозинофил-связанного желудочно-кишечного заболевания; буллезного пемфигоида; интерстициального цистита; полипов носовой полости; идиопатического ангионевротического отека или неаллергической астмы.
12. Применение композиции по п. 9 для лечения или профилактики одного или нескольких заболеваний: аллергии; аллергической астмы; тяжелой астмы; умеренной астмы; хронической спонтанной крапивницы; хронической идиопатической крапивницы; продолжительного аллергического ринита; сезонного аллергического ринита; обострений астмы; острого бронхоспазма; астматического статуса; гипер-IgE синдрома; аллергического бронхолегочного аспергиллеза; профилактики анафилактических реакций; пищевой аллергии; атопического дерматита; аллергического ринита; аллергии на пчелиный яд; идиопатической анафилаксии; аллергии на арахис; аллергии на латекс; воспалительных кожных заболеваний; крапивницы (вызываемой солнечным светом, холодом, локальным нагреванием и/или длительным сдавливанием); кожного мастоцитоза; системного мастоцитоза; эозинофил-связанного желудочно-кишечного заболевания; буллезного пемфигоида; интерстициального цистита; полипов носовой полости; идиопатического ангионевротического отека или неаллергической астмы.
| WO 2011056606 A1, 12.05.2011 | |||
| RU 2013123927 A, 27.11.2014 | |||
| CN 104987412 A, 21.10.2015 | |||
| US 20130243750 A1, 19.09.2013. |

