Родственные заявки
В настоящей заявке испрашивается преимущество предварительной заявки на патент США No. 61/858402, поданной 25 июля 2013, содержание которой во всей своей полноте вводится в настоящее описание посредством ссылки.
Область, к которой относится изобретение
В общих чертах, настоящее изобретение относится к мультиспецифическим антителам и к мультиспецифическим активируемым антителам, которые специфически связываются с двумя или более различными мишенями или эпитопами, а также к способам получения и применения этих мультиспецифических антител и/или мультиспецифических активируемых антител в различных терапевтических, диагностических и профилактических целях.
Предшествующий уровень техники
Моноклональные антитела были получены как молекулы для терапевтического лечения различных расстройств. Однако, нацеливание на отдельный белок или его нейтрализация не всегда являются достаточными для эффективного лечения некоторых расстройств, и это ограничивает терапевтическое применение моноспецифических моноклональных антител. Кроме того, было установлено, что терапия на основе антител является эффективной для лечения некоторых заболеваний, но, в некоторых случаях, токсичность, вызываемая экспрессией мишеней широкого ряда, ограничивает терапевтическую эффективность этих антител. Более того, терапия на основе антител имеют и другие ограничения, такие как быстрый клиренс этих антител из кровотока после их введения.
В соответствии с этим возникает необходимость в получении антител и терапевтических средств, которые были бы способны к нацеливанию на множество мишеней и/или множество эпитопов в одной молекуле, а также обладали бы повышенной селективностью по отношению к рассматриваемым мишеням, но, при этом, не вызывали бы серьезных побочных эффектов после их введения.
Описание сущности изобретения
Настоящее изобретение относится к мультиспецифическим антителам и к мультиспецифическим активируемым антителам. Описанные здесь мультиспецифические антитела представляют собой антитела, которые распознают два или более различных антигенов или эпитопов. Описанные здесь мультиспецифические активируемые антитела представляют собой мультиспецифические антитела, которые включают по меньшей мере одну маскирующую группу (ММ), связанную по меньшей мере с одним антиген- или эпитоп-связывающим доменом мультиспецифического антитела, а также связывание с ММ приводит к снижению способности антиген- или эпитоп-связывающего домена связываться с мишенью. В некоторых вариантах осуществления изобретения, ММ связывается с антиген- или эпитоп-связывающим доменом мультиспецифического антитела посредством отщепляемой группы (CM), которая служит в качестве субстрата для протеазы. Описанные здесь активируемые мультиспецифические антитела являются стабильными в кровотоке, и активируются в нужных участках, подвергаемых лечению и/или диагностике, но не в нормальной, то есть, здоровой ткани, и в случае такой активации, эти антитела обладают способностью связываться с мишенью на уровне, по меньшей мере сравнимом с уровнем связывания соответствующего немодифицированного мультиспецифического антитела.
В некоторых вариантах осуществления изобретения, мультиспецифические антитела и/или мультиспецифические активируемые антитела конструируют так, чтобы они связывались с иммунными эффекторными клетками, и такие антитела называются здесь мультиспецифическими антителами, связывающимися с иммунным эффектором, и/или мультиспецифическими активируемыми антителами, связывающимися с иммунным эффектором. В некоторых вариантах осуществления изобретения, мультиспецифические антитела и/или мультиспецифические активируемые антитела конструируют так, чтобы они связывались с лейкоцитами, и такие антитела называются здесь мультиспецифическими антителами, связывающимися с лейкоцитами, и/или мультиспецифическими активируемыми антителами, связывающимися с лейкоцитами. В некоторых вариантах осуществления изобретения, мультиспецифические антитела и/или мультиспецифические активируемые антитела конструируют так, чтобы они связывались с Т-клетками, и такие антитела называются здесь мультиспецифическими антителами, связывающимися с Т-клетками, и/или мультиспецифическими активируемыми антителами, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, мультиспецифические антитела и/или мультиспецифические активируемые антитела связываются с поверхностным антигеном на лейкоцитах, например, на Т-клетках, на природных клетках-киллерах (NK), на миелоидных мононуклеарных клетках, на макрофагах и/или на других иммунных эффекторных клетках. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является лейкоцит. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является T-клетка. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является NK-клетка. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является мононуклеарная клетка, такая как миелоидная мононуклеарная клетка. В некоторых вариантах осуществления изобретения, мультиспецифические антитела и/или мультиспецифические активируемые антитела конструируют так, чтобы они связывались или как-либо иначе взаимодействовали с более, чем одной мишенью и/или с более, чем одним эпитопом, и такие антитела также называются здесь антителами, нацеленными на множество антигенов, и/или активируемыми антителами, нацеленными на множество антигенов. Используемые здесь термины «мишень» и «антиген» являются синонимами.
В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с иммунной эффекторной клеткой, являются нацеливающее антитело или его антигенсвязывающий фрагмент, и антитело, связывающееся с иммунной эффекторной клеткой, или его антигенсвязывающая часть. В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с иммунной эффекторной клеткой, являются антитело, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, и антитело, связывающееся с иммунной эффекторной клеткой, или его антигенсвязывающая часть. В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с иммунной эффекторной клеткой, являются антитело IgG, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, и scFv, связывающийся с иммунной эффекторной клеткой. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является лейкоцит. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является T-клетка. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является NK-клетка. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является миелоидная мононуклеарная клетка.
В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с Т-клеткой, являются нацеливающее антитело или его антигенсвязывающий фрагмент, и антитело, связывающееся с Т-клеткой, или его антигенсвязывающая часть. В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с Т-клеткой, являются антитело, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, и антитело, связывающееся с Т-клеткой, или его антигенсвязывающая часть. В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с Т-клеткой, являются антитело IgG, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, и scFv, связывающийся с Т-клеткой. В некоторых вариантах осуществления изобретения, мультиспецифическим антителом, связывающимся с Т-клеткой, является scFv-фрагмент антитела против CD3 эпсилон (CD3ε, также обозначаемого здесь CD3e и CD3) и нацеливающее антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления изобретения, мультиспецифическим антителом, связывающимся с Т-клеткой, является scFv-фрагмент антитела против CD3ε, и антитело, нацеленное на раковую опухоль или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления изобретения, мультиспецифическим антителом, связывающимся с Т-клеткой, является анти-CD3ε scFv и антитело IgG, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления изобретения, мультиспецифическим антителом, связывающимся с Т-клеткой, является scFv-фрагмент антитела против CD3 эпсилон (CD3ε), который происходит от OKT3. В некоторых вариантах осуществления изобретения, мультиспецифическим антителом, связывающимся с Т-клеткой, является анти-CTLA-4 scFv.
В некоторых вариантах осуществления изобретения, мультиспецифическими антителами, связывающимися с иммунной эффекторной клеткой согласно изобретению, является нацеливающее антитело или его антигенсвязывающий фрагмент, и антитело, связывающееся с иммунной эффекторной клеткой, или его антигенсвязывающая часть, где по меньшей мере одно из нацеливающих антител или его антигенсвязывающих фрагментов и/или по меньшей мере одно из антител, связывающихся с иммунной эффекторной клеткой, или его антигенсвязывающих частей является маскированным. В некоторых вариантах осуществления изобретения, антителом, связывающимся с иммунной эффекторной клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с иммунной эффекторной клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью. В некоторых вариантах осуществления изобретения, нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, антителом, связывающимся с иммунной эффекторной клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с иммунной эффекторной клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью, а нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, антителом, связывающимся с неиммунной эффекторной клеткой, является антитело, нацеленное на раковую опухоль. В некоторых вариантах осуществления изобретения, антителом против неиммунных эффекторных клеток является IgG. В некоторых вариантах осуществления изобретения, антителом, связывающимся с иммунной эффекторной клеткой, является scFv. В некоторых вариантах осуществления изобретения, нацеливающим антителом (например, антителоом против неиммунных эффекторных клеток) является IgG, а антителом, связывающимся с иммунной эффекторной клеткой, является scFv. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является лейкоцит. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является Т-клетка. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является NK-клетка. В некоторых вариантах осуществления изобретения, иммунной эффекторной клеткой является миелоидная мононуклеарная клетка.
В некоторых вариантах осуществления изобретения, мультиспецифическими активируемыми антителами, связывающимися с Т-клеткой согласно изобретению, является нацеливающее антитело или его антигенсвязывающий фрагмент, и антитело, связывающееся с Т-клеткой, или его антигенсвязывающая часть, где по меньшей мере одно из нацеливающих антител или его антигенсвязывающих фрагментов и/или по меньшей мере одно из антител, связывающихся с Т-клеткой, или его антигенсвязывающих частей является маскированным. В некоторых вариантах осуществления изобретения, антителом, связывающимся с Т-клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью. В некоторых вариантах осуществления изобретения, нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, антителом, связывающимся с Т-клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью, а нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью.
В некоторых вариантах осуществления изобретения, мультиспецифическими активируемыми антителами, связывающимися с Т-клеткой согласно изобретению, является антитело, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, и антитело, связывающееся с Т-клеткой, или его антигенсвязывающая часть, где по меньшей мере одно из антител, нацеленных на раковую опухоль, или его антигенсвязывающих фрагментов и/или по меньшей мере одно из антител, связывающихся с Т-клеткой, или его антигенсвязывающих частей является маскированным. В некоторых вариантах осуществления изобретения, антителом, связывающимся с Т-клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью. В некоторых вариантах осуществления изобретения, антителом, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциирующейся с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциирующейся с раковой опухолью. В некоторых вариантах осуществления изобретения, антителом, связывающимся с Т-клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью, а антителом, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциирующейся с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциирующейся с раковой опухолью.
В некоторых вариантах осуществления изобретения, мультиспецифическими активируемыми антителами, связывающимися с Т-клеткой согласно изобретению, является антитело IgG, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, и scFv, связывающийся с Т-клеткой, где по меньшей мере одно из антител IgG, нацеленных на раковую опухоль, или его антигенсвязывающих фрагментов и/или по меньшей мере одно из антител, связывающихся с Т-клеткой, или его антигенсвязывающих частей является маскированным. В некоторых вариантах осуществления изобретения, антителом, связывающимся с Т-клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью. В некоторых вариантах осуществления изобретения, антителом IgG, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью. В некоторых вариантах осуществления изобретения, антителом, связывающимся с Т-клеткой, или его антигенсвязывающим фрагментом является первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клеткой, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с первой мишенью, а антителом IgG, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью.
В некоторых вариантах осуществления изобретения, относящихся к мультиспецифическому активируемому антителу, связывающемуся с иммунной эффекторной клеткой, антигеном обычно является антиген, присутствующий на поверхности опухолевой клетки или клетки другого типа, ассоциированной с заболеванием, и такими антигенами являются, но не ограничиваются ими, любые мишени, перечисленные в таблице 1, такие как, но не ограничивающиеся ими, EGFR, erbB2, EpCAM, Jagged, PD-L1, B7H3 или CD71 (рецептор трансферрина), а другим антигеном обычно являются стимулирующий или ингибирующий рецептор, присутствующий на поверхности Т-клетки, природной клетки-киллера (NK), миелоидной мононуклеарной клетки, макрофага и/или других иммунных эффекторных клеток, где таким антигеном являются, но не ограничиваются ими, B7-H4, BTLA, CD3, CD4, CD8, CD16a, CD25, CD27, CD28, CD32, CD56, CD137, CTLA-4, GITR, HVEM, ICOS, LAG3, NKG2D, OX40, PD-1, TIGIT, TIM3 или VISTA. В некоторых вариантах осуществления изобретения, антигеном является стимулирующий рецептор, присутствующий на поверхности Т-клетки или NK-клетки, и примерами таких стимулирующих рецепторов являются, но не ограничиваются ими, CD3, CD27, CD28, CD137 (также называемый 4-1BB), GITR, HVEM, ICOS, NKG2D и OX40. В некоторых вариантах осуществления изобретения, антигеном является ингибирующий рецептор, присутствующий на поверхности Т-клетки, и примерами таких ингибирующих рецепторов являются, но не ограничиваются ими, BTLA, CTLA-4, LAG3, PD-1, TIGIT, TIM3 и KIR, экспрессируемые NK. Домен антитела, сообщающий специфичность к антигену Т-клеточной поверхности, может быть также заменен лигандом или доменом лиганда, который связывается с рецептором Т-клеток, рецептором NK-клеток, рецептором макрофагов и/или рецептором других иммунных эффекторных клеток, и таким лигандом являются, но не ограничиваются ими, B7-1, B7-2, B7H3, PD-L1, PD-L2 или TNFSF9.
Одним из вариантов осуществления изобретения является мультиспецифическое активируемое антитело, которое активируется в микроокружении раковой опухоли, и которое представляет собой антитело, например, IgG или scFv, направленное на опухолевую мишень, и антитело-агонист, например IgG или scFv, направленное на костимулирующий рецептор, экспрессируемый на поверхности активированной Т-клетки или NK-клетки, где по меньшей мере одно из антител, нацеленных на раковую опухоль и/или антител-агонистов, является маскированным. Примерами костимулирующих рецепторов являются, но не ограничиваются ими, CD27, CD137, GITR, HVEM, NKG2D и OX40. В этом варианте осуществления изобретения, мультиспецифическое активируемое антитело, после его активации опухоле-ассоциированными протеазами, эффективно и перекрестно связывается с костимулирующими рецепторами, экспрессируемыми Т-клеткой или NK-клеткой, и активирует эти рецепторы по опухоле-зависимому механизму, что приводит к повышениею активности Т-клеток, отвечающих на любой опухолевый антиген посредством эндогенных Т-клеточных антигенов или NK-активирующих рецепторов. Зависимость этих костимулирующих рецепторов Т-клеток или NK-клеток от активации позволяет направлять активность активированного мультиспецифического активируемого антитела на опухолеспецифические Т-клетки, но не на все Т-клетки, независимо от антигенной специфичности этих антител. В одном из вариантов осуществления изобретения, по меньшей мере одно из мультиспецифических активируемых антител против костимулирующего рецептора маскируют для предупреждения активации аутореактивных Т-клеток, которые могут присутствовать в тканях, также экспрессирующих антиген, распознаваемый антителом против опухолевой мишени, которое представляет собой мультиспецифическое активируемое антитело, но активность которого ограничена отсутствием связывания с ко-рецептором.
Одним из вариантов осуществления изобретения является мультиспецифическое активируемое антитело, которое активируется при заболевании, характеризующимся сверхстимуляцией Т-клеток, и таким заболеванием являются, но не ограничиваются ими, аутоиммунное заболевание или микроокружение, вызываающее воспалительное заболевание. Таким мультиспецифическим активируемым антителом является антитело, например, IgG или scFv, направленное на мишень, содержащую поверхностный антиген, экспрессируемый в ткани, являющейся мишенью для Т-клеток, при аутоиммунном или воспалительном заболевании, и антитело, например, IgG или scFv, направленное против ингибирующего рецептора, экспрессируемого на поверхности Т-клетки или NK-клетки, где по меньшей мере одно из антител, нацеленных на пораженную ткань и/или антител против ингибирующего рецептора Т-клеток, является маскированным. Примерами таких ингибирующих рецепторов являются, но не ограничиваются ими, BTLA, CTLA-4, LAG3, PD-1, TIGIT, TIM3 и KIR, экспрессируемые NK. Примерами тканевого антигена-мишени для Т-клеток при аутоиммунном заболевании являются, но не ограничиваются ими, поверхностный антиген, экспрессирующийся на миелиновых или нервных клетках при рассеянном склерозе, или поверхностный антиген, экспрессирующийся на клетках панкреатических островков при диабете типа 1. В этом варианте осуществления изобретения, мультиспецифическое активируемое антитело, если оно локализуется в ткани при аутоимунной атаке или при воспалении, активируется и связывается с ингибирующим ко-рецептором Т-клеток или NK-клеток, что приводит к подавлению активности аутореактивных Т-клеток, отвечающих на любые антигены-мишени в пораженной ткани посредством эндогенных TCR или активирующих рецепторов. В одном из вариантов осуществления изобретения, по меньшей мере одно или множество антител маскируют для предупреждения ингибирования желаемых Т-клеточных ответов в непораженных тканях, в которых может также экспрессироваться антиген-мишень.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является scFv-фрагмент антитела против CD3 эпсилон (CD3ε, также обозначаемого здесь CD3e и CD3) и нацеливающее антитело или его антигенсвязывающий фрагмент, где по меньшей мере один из анти-CD3ε scFv и/или одно из нацеливающих антител или его антигенсвязывающих частей являются маскированными. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε. В некоторых вариантах осуществления изобретения, нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε, а нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является анти-CD3ε scFv и антитело, нацеленное на раковую опухоль или его антигенсвязывающий фрагмент, где по меньшей мере один из анти-CD3ε scFv и/или одно из антител, нацеленных на раковую опухоль, или его антигенсвязывающих частей являются маскированными. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε. В некоторых вариантах осуществления изобретения, антителом, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε, а антителом, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является анти-CD3ε scFv и антитело IgG, нацеленное на раковую опухоль или его антигенсвязывающий фрагмент, где по меньшей мере один из анти-CD3ε scFv и/или одно из антител IgG, нацеленных на раковую опухоль, или его антигенсвязывающих частей являются маскированными. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε. В некоторых вариантах осуществления изобретения, антителом IgG, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε, а антителом IgG, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является scFv-фрагмент антитела против CD3 эпсилон (CD3ε), который происходит от OKT3, где по меньшей мере одно из нацеливающих антител или его антигенсвязывающих фрагментов и/или один из анти-OKT3 scFv и/или scFv, происходящих от OKT3, являются маскированными. В некоторых вариантах осуществления изобретения, анти-OKT3 scFv или scFv, происходящий от OKT3, представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε. В некоторых вариантах осуществления изобретения, нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, анти-OKT3 scFv или scFv, происходящий от OKT3, представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε, а нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является анти-OKT3 scFv и/или scFv, происходящий от OKT3, и антитело, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, где по меньшей мере один из анти-OKT3 scFv и/или scFv, происходящих от OKT3, и/или одно из антител, нацеленных на раковую опухоль, или его антигенсвязывающих частей являются маскированными. В некоторых вариантах осуществления изобретения, анти-OKT3 scFv или scFv, происходящий от OKT3, представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε. В некоторых вариантах осуществления изобретения, антителом, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью. В некоторых вариантах осуществления изобретения, анти-OKT3 scFv или scFv, происходящий от OKT3, представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε, а антителом, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является анти-OKT3 scFv и/или scFv, происходящий от OKT3, и антитело IgG, нацеленное на раковую опухоль, или его антигенсвязывающий фрагмент, где по меньшей мере один из анти-OKT3 scFv и/или scFv, происходящих от OKT3, и/или одно из антител IgG, нацеленных на раковую опухоль, или его антигенсвязывающих частей являются маскированными. В некоторых вариантах осуществления изобретения, анти-OKT3 scFv или scFv, происходящий от OKT3, представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε. В некоторых вариантах осуществления изобретения, антителом IgG, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью. В некоторых вариантах осуществления изобретения, анти-OKT3 scFv или scFv, происходящий от OKT3, представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CD3ε, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CD3ε, а антителом IgG, нацеленным на раковую опухоль, или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, ассоциированной с раковой опухолью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью, ассоциированной с раковой опухолью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является анти-CTLA-4 scFv, где по меньшей мере одно из нацеливающих антител или его антигенсвязывающих фрагментов и/или один из анти-CTLA-4 scFv являются маскированными. В некоторых вариантах осуществления изобретения, анти-CTLA-4 scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CTLA-4, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CTLA-4. В некоторых вариантах осуществления изобретения, нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, анти-CTLA-4 scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CTLA-4, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CTLA-4, а нацеливающим антителом или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом, связывающимся с Т-клеткой, является анти-CTLA-4 scFv и нацеливающее антитело IgG или его антигенсвязывающий фрагмент, где по меньшей мере один из анти-CTLA-4 scFv и/или по меньшей мере одно из нацеливающих антител IgG или его антигенсвязывающих частей являются маскированными. В некоторых вариантах осуществления изобретения, анти-CTLA-4 scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CTLA-4, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CTLA-4. В некоторых вариантах осуществления изобретения, нацеливающим антителом IgG или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, анти-CTLA-4 scFv представляет собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с CTLA-4, где AB1 связывается с маскирующей группой (MM1), и такое связывание с MM1 приводит к снижению способности AB1 связываться с CTLA-4, а нацеливающим антителом IgG или его антигенсвязывающим фрагментом является второе антитело или его фрагмент, которые представляют собой второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где AB2 связывается с маскирующей группой (MM2), и такое связывание с MM2 приводит к снижению способности AB2 связываться со второй мишенью.
В некоторых вариантах осуществления изобретения, антителами, нацеленными на множество антигенов, и/или активируемыми антителами, нацеленными на множество антигенов, являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент, которые связываются с первой мишенью и/или с первым эпитопом, и второе антитело или его антигенсвязывающий фрагмент, которые связываются со второй мишенью и/или со вторым эпитопом. В некоторых вариантах осуществления изобретения, антитела, нацеленные на множество антигенов, и/или активируемые антитела, нацеленные на множество антигенов, связываются с двумя или более различными мишенями. В некоторых вариантах осуществления изобретения, антитела, нацеленные на множество антигенов, и/или активируемые антитела, нацеленные на множество антигенов, связываются с двумя или более различными эпитопами на одной и той же мишени. В некоторых вариантах осуществления изобретения, антитела, нацеленные на множество антигенов, и/или активируемые антитела, нацеленные на множество антигенов, связываются с комбинацией двух или более различных мишеней и двух или более различных эпитопов на одной и той же мишени.
Различные варианты мультиспецифических активируемых антител согласно изобретению представлены на фигурах 3А и 5-9. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело, включающее IgG, имеет маскированные вариабельные домены IgG. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело, включающее scFv, имеет маскированные домены scFv. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где по меньшей мере один из вариабельных доменов IgG связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где по меньшей мере один из доменов scFv связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где по меньшей мере один из вариабельных доменов IgG связан с маскирующей группой, и по меньшей мере один из доменов scFv связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где вариабельные домены IgG и домены scFv связаны со своей собственной маскирующей группой. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к антигену мишени, а другой домен этого антитела обладает специфичностью к антигену Т-клеточной поверхности. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к антигену мишени, а другой домен этого антитела обладает специфичностью к другому антигену мишени. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к эпитопу антигена мишени, а другой домен этого антитела обладает специфичностью к другому эпитопу антигена мишени.
В мультиспецифическом активируемом антителе, scFv может быть присоединен к карбоксильному концу тяжелой цепи активируемого антитела IgG, к карбоксильному концу легкой цепи активируемого антитела IgG или к карбоксильным концам тяжелой и легкой цепей активируемого антитела IgG. В мультиспецифическом активируемом антителе, scFv может быть присоединен к амино-концу тяжелой цепи активируемого антитела IgG, к амино-концу легкой цепи активируемого антитела IgG или к амино-концам тяжелой и легкой цепей активируемого антитела IgG. В мультиспецифическом активируемом антителе, scFv может быть присоединен к любой комбинации одного или более карбоксильных концов и одного или более амино-концов активируемого антитела IgG. В некоторых вариантах осуществления изобретения, маскирующая группа (MM), связанная с отщепляемой группой (CM), присоединена к антигенсвязывающему домену IgG и маскирует этот домен. В некоторых вариантах осуществления изобретения, маскирующая группа (MM), связанная с отщепляемой группой (CM), присоединена к антигенсвязывающему домену по меньшей мере одного scFv и маскирует этот домен. В некоторых вариантах осуществления изобретения, маскирующая группа (MM), связанная с отщепляемой группой (CM), присоединена к антигенсвязывающему домену IgG и маскирует этот домен, и маскирующая группа (MM), связанная с отщепляемой группой (CM), присоединена к антигенсвязывающему домену по меньшей мере одного scFv и маскирует этот домен.
В описании настоящего изобретения приводятся примеры структур мультиспецифических активируемых антител, которыми являются, но не ограничиваются ими, следующие структуры: (VL-CL)2:(VH-CH1-CH2-CH3-L4-VH*-L3-VL*-L2-CM-L1-MM)2; (VL-CL)2:(VH-CH1-CH2-CH3-L4-VL*-L3-VH*-L2-CM-L1-MM)2; (MM-L1-CM-L2-VL-CL)2:(VH-CH1-CH2-CH3-L4-VH*-L3-VL*)2; (MM-L1-CM-L2-VL-CL)2:(VH-CH1-CH2-CH3-L4-VL*-L3-VH*)2; (VL-CL)2:(MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL)2:(MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL-CL)2:(VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL-CL)2:(VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL*-L3-VH*-L4-VL-CL)2:(VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VH*-L3-VL*-L4-VL-CL)2:(VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; или (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2, где: VL и VH представляют собой вариабельные домены легкой и тяжелой цепей первой специфичности, содержащиеся в IgG; VL* и VH* представляют собой вариабельные домены второй специфичности, содержащиеся в scFv; L1 представляет собой линкерный пептид, соединяющий маскирующую группу (MM) и отщепляемую группу (CM); L2 представляет собой линкерный пептид, соединяющий отщепляемую группу (CM) и антитело; L3 представляет собой линкерный пептид, соединяющий вариабельные домены scFv; L4 собой линкерный пептид, соединяющий антитело первой специфичности и антитело второй специфичности; CL представляет собой константный домен легкой цепи, а CH1, CH2, CH3 представляют собой константные домены тяжелой цепи. Первая и вторая специфичности могут относиться к любому антигену или эпитопу.
В некоторых вариантах изобретения, относящихся к мультиспецифическому активируемому антителу, связывающемуся с T-клеткой, одним антигеном обычно является антиген, присутствующий на поверхности опухолевой клетки или клетки другого типа, ассоциированной с заболеванием, и такими антигенами являются, но не ограничиваются ими, любые мишени, перечисленные в таблице 1, такие как, но не ограничивающиеся ими, EGFR, erbB2, EpCAM, Jagged, PD-L1, B7H3 или CD71 (рецептор трансферрина), а другим антигеном обычно являются стимулирующий рецептор (также называемый здесь активирующим рецептором) или ингибирующий рецептор, присутствующий на поверхности Т-клетки, природной клетки-киллера (NK), миелоидной мононуклеарной клетки, макрофага и/или других иммунных эффекторных клеток, где таким антигеном являются, но не ограничиваются ими, B7-H4, BTLA, CD3, CD4, CD8, CD16a, CD25, CD27, CD28, CD32, CD56, CD137 (также называемый TNFRSF9), CTLA-4, GITR, HVEM, ICOS, LAG3, NKG2D, OX40, PD-1, TIGIT, TIM3 или VISTA. Домен антитела, сообщающий специфичность к антигену Т-клеточной поверхности, может быть также заменен лигандом или доменом лиганда, который связывается с рецептором Т-клеток, рецептором NK-клеток, рецептором макрофагов и/или рецептором других иммунных эффекторных клеток, и такими лигандами являются, но не ограничиваются ими, B7-1, B7-2, B7H3, PD-L1, PD-L2 или TNFSF9. В некоторых вариантах осуществления изобретения, относящихся к активируемому антителу, нацеленному на множество антигенов, один антиген выбран из группы мишеней, перечисленных в таблице 1, и второй антиген выбран из группы мишеней, перечисленных в таблице 1.
В некоторых вариантах осуществления изобретения, нацеливающим антителом является анти-EGFR антитело. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225v5, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225v4, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225v6, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является анти-Jagged антитело. В некоторых вариантах осуществления изобретения, нацеливающим антителом является 4D11, которое специфически связывается с человеческим и мышиным Jagged 1 и Jagged 2. В некоторых вариантах осуществления изобретения, нацеливающим антителом является 4D11v2, которое специфически связывается с человеческим и мышиным Jagged 1 и Jagged 2.
В некоторых вариантах осуществления изобретения, нацеливающее антитело может присутствовать в форме активируемого антитела. В некоторых вариантах осуществления изобретения, scFv может (могут) присутствовать в форме Pro-scFv (см., например, WO 2009/025846, WO 2010/081173).
В некоторых вариантах осуществления изобретения, scFv специфически связывается с CD3ε, и/или происходит от антитела или его фрагмента, связывающихся с CD3ε, например, с CH2527, FN18, H2C, OKT3, 2C11, UCHT1 или V9. В некоторых вариантах осуществления изобретения, scFv специфически связывается с CTLA-4 (также называемым здесь CTLA и CTLA4).
В некоторых вариантах осуществления изобретения, анти-CD3ε scFv имеет последовательность, выбранную из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5. В некоторых вариантах осуществления изобретения, анти-CD3ε scFv имеет аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5.
В некоторых вариантах осуществления изобретения, scFv специфически связывается с одной или более Т-клетками, одной или более NK-клетками и/или с одним или более макрофагами. В некоторых вариантах осуществления изобретения, scFv специфически связывается с мишенью, выбранной из группы, состоящей из B7-H4, BTLA, CD3, CD4, CD8, CD16a, CD25, CD27, CD28, CD32, CD56, CD137, CTLA-4, GITR, HVEM, ICOS, LAG3, NKG2D, OX40, PD-1, TIGIT, TIM3 или VISTA.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью Jagged, например, Jagged 1 и/или Jagged 2, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH и последовательности CDR3 VH, где по меньшей мере одна из последовательностей CDR1 VH, последовательностей CDR2 VH и последовательностей CDR3 VH выбрана из последовательности CDR1 VH, которая включает по меньшей мере аминокислотную последовательность SYAMS (SEQ ID NO: 6); последовательности CDR2 VH, которая включает по меньшей мере аминокислотную последовательность SIDPEGRQTYYADSVKG (SEQ ID NO: 7); последовательности CDR3 VH, которая включает по меньшей мере аминокислотную последовательность DIGGRSAFDY (SEQ ID NO: 8), и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью Jagged, например, Jagged 1 и/или Jagged 2, и которые содержат комбинацию последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где по меньшей мере одна из последовательностей CDR1 VL, последовательностей CDR2 VL и последовательностей CDR3 VL, выбрана из последовательности CDR1 VL, которая включает по меньшей мере аминокислотную последовательность RASQSISSY (SEQ ID NO: 9); последовательности CDR2 VL, которая включает по меньшей мере аминокислотную последовательность AASSLQS (SEQ ID NO: 10); последовательности CDR3 VL, которая включает по меньшей мере аминокислотную последовательность QQTVVAPPL (SEQ ID NO: 11), и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью Jagged, например, Jagged 1 и/или Jagged 2, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH и последовательности CDR3 VH, где по меньшей мере одна из последовательностей CDR1 VH, последовательностей CDR2 VH и последовательностей CDR3 VH выбрана из последовательности CDR1 VH, включающей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SYAMS (SEQ ID NO: 6); последовательности CDR2 VH, включающей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SIDPEGRQTYYADSVKG (SEQ ID NO: 7); последовательности CDR3 VH, включающей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности DIGGRSAFDY (SEQ ID NO: 8), и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью Jagged, например, Jagged 1 и/или Jagged 2, и которые содержат комбинацию последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где по меньшей мере одна из последовательностей CDR1 VL, последовательностей CDR2 VL и последовательностей CDR3 VL выбрана из последовательности CDR1 VL, включающей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RASQSISSY (SEQ ID NO: 9); последовательности CDR2 VL, включающей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности AASSLQS (SEQ ID NO: 10); последовательности CDR3 VL, включающей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности QQTVVAPPL (SEQ ID NO: 11), и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью Jagged, например, Jagged 1 и/или Jagged 2, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH, последовательности CDR3 VH, последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где последовательность CDR1 VH включает по меньшей мере аминокислотную последовательность SYAMS (SEQ ID NO: 6); последовательность CDR2 VH включает по меньшей мере аминокислотную последовательность SIDPEGRQTYYADSVKG (SEQ ID NO: 7); последовательность CDR3 VH включает по меньшей мере аминокислотную последовательность DIGGRSAFDY (SEQ ID NO: 8), последовательность CDR1 VL включает по меньшей мере аминокислотную последовательность RASQSISSY (SEQ ID NO: 9); последовательность CDR2 VL включает по меньшей мере аминокислотную последовательность AASSLQS (SEQ ID NO: 10), а последовательность CDR3 VL включает по меньшей мере аминокислотную последовательность QQTVVAPPL (SEQ ID NO: 11).
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью Jagged, например, Jagged 1 и/или Jagged 2, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH, последовательности CDR3 VH, последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где последовательность CDR1 VH включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SYAMS (SEQ ID NO: 6); последовательность CDR2 VH включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SIDPEGRQTYYADSVKG (SEQ ID NO: 7); последовательность CDR3 VH включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности DIGGRSAFDY (SEQ ID NO: 8); последовательность CDR1 VL включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RASQSISSY (SEQ ID NO: 9); последовательность CDR2 VL включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности AASSLQS (SEQ ID NO: 10); а последовательность CDR3 VL включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности QQTVVAPPL (SEQ ID NO: 11).
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с рецептором эпидермального фактора роста (EGFR), и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH и последовательности CDR3 VH, где по меньшей мере одна из последовательностей CDR1 VH, последовательностей CDR2 VH и последовательностей CDR3 VH выбрана из последовательности CDR1 VH, которая включает по меньшей мере аминокислотную последовательность NYGVH (SEQ ID NO: 12); последовательности CDR2 VH, которая включает по меньшей мере аминокислотную последовательность VIWSGGNTDYNTPFTS (SEQ ID NO: 13); последовательности CDR3 VH, которая включает по меньшей мере аминокислотную последовательность ALTYYDYEFAY (SEQ ID NO: 14); и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с EGFR, и которые содержат комбинацию последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где по меньшей мере одна из последовательностей CDR1 VL, последовательностей CDR2 VL и последовательностей CDR3 VL выбрана из последовательности CDR1 VL, которая включает по меньшей мере аминокислотную последовательность RASQSIGTNIH (SEQ ID NO: 15); последовательности CDR2 VL, которая включает по меньшей мере аминокислотную последовательность KYASESIS (SEQ ID NO: 16); последовательности CDR3 VL, которая включает по меньшей мере аминокислотную последовательность QQNNNWPTT (SEQ ID NO: 17); и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с EGFR, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH и последовательности CDR3 VH, где по меньшей мере одна из последовательностей CDR1 VH, последовательностей CDR2 VH и последовательностей CDR3 VH выбрана из последовательности CDR1 VH, включащей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности NYGVH (SEQ ID NO: 12); последовательности CDR2 VH, включащей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности VIWSGGNTDYNTPFTS (SEQ ID NO: 13); последовательности CDR3 VH, включащей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности ALTYYDYEFAY (SEQ ID NO: 14); и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с EGFR, и которые содержат комбинацию последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где по меньшей мере одна из последовательностей CDR1 VL, последовательностей CDR2 VL и последовательностей CDR3 VL выбрана из последовательности CDR1 VL, включащей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RASQSIGTNIH (SEQ ID NO: 15); последовательности CDR2 VL, включащей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности KYASESIS (SEQ ID NO: 16); последовательности CDR3 VL, включащей последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности QQNNNWPTT (SEQ ID NO: 17); и их комбинаций.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с EGFR, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH, последовательности CDR3 VH, последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где последовательность CDR1 VH включает по меньшей мере аминокислотную последовательность NYGVH (SEQ ID NO: 12); последовательность CDR2 VH включает по меньшей мере аминокислотную последовательность VIWSGGNTDYNTPFTS (SEQ ID NO: 13); последовательность CDR3 VH включает по меньшей мере аминокислотную последовательность ALTYYDYEFAY (SEQ ID NO: 14), последовательность CDR1 VL включает по меньшей мере аминокислотную последовательность RASQSIGTNIH (SEQ ID NO: 15); последовательность CDR2 VL включает по меньшей мере аминокислотную последовательность KYASESIS (SEQ ID NO: 16), а последовательность CDR3 VL включает по меньшей мере аминокислотную последовательность QQNNNWPTT (SEQ ID NO: 17).
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с EGFR, и которые содержат комбинацию последовательности CDR1 VH, последовательности CDR2 VH, последовательности CDR3 VH, последовательности CDR1 VL, последовательности CDR2 VL и последовательности CDR3 VL, где последовательность CDR1 VH включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности NYGVH (SEQ ID NO: 12); последовательность CDR2 VH включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности VIWSGGNTDYNTPFTS (SEQ ID NO: 13); последовательность CDR3 VH включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности ALTYYDYEFAY (SEQ ID NO: 14); последовательность CDR1 VL включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RASQSIGTNIH (SEQ ID NO: 15); последовательность CDR2 VL включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности KYASESIS (SEQ ID NO: 16), а последовательность CDR3 VL включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности QQNNNWPTT (SEQ ID NO: 17).
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере аминокислотная последовательность тяжелой цепи, выбранная из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11. В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере аминокислотная последовательность легкой цепи, выбранная из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11. В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере аминокислотная последовательность тяжелой цепи, выбранная из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11, и аминокислотная последовательность легкой цепи, выбранная из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11.
В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере аминокислотная последовательность тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11. В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере аминокислотная последовательность легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11. В некоторых вариантах осуществления изобретения, описанными здесь мультиспецифическими антителами, мультиспецифическими активируемыми антителами, конъюгированными мультиспецифическими антителами и/или конъюгированными мультиспецифическими активируемыми антителами являются по меньшей мере аминокислотная последовательность тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11, и аминокислотная последовательность легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательностей, представленных в таблице 7 и/или в примере 5, включащем таблицу 11.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело также включает агент, конъюгированный с AB. В некоторых вариантах осуществления изобретения, таким агентом является терапевтический агент. В некоторых вариантах осуществления изобретения, таким агентом является противоопухолевый агент. В некоторых вариантах осуществления изобретения, таким агентом является токсин или его фрагмент. В некоторых вариантах осуществления изобретения, указанный агент конъюгирован с мультиспецифическим антителом посредством линкера. В некоторых вариантах осуществления изобретения, указанным линкером является нерасщепляемый линкер. В некоторых вариантах осуществления изобретения, указанным агентом является ингибитор микротрубочек. В некоторых вариантах осуществления изобретения, указанным агентом является агент, разрушающий нуклеиновую кислоту, такой как ДНК-алкилирующий агент или ДНК-интеркалирующий агент или другой агент, разрушающий ДНК. В некоторых вариантах осуществления изобретения, указанным линкером является расщепляемый линкер. В некоторых вариантах осуществления изобретения, указанным агентом является агент, выбранный из группы агентов, перечисленных в таблице 4. В некоторых вариантах осуществления изобретения, указанным агентом является доластатин. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин Е или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин E (MMAE). В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин D (MMAD). В некоторых вариантах осуществления изобретения, указанным агентом является майтанзиноид или производное майтанзиноида. В некоторых вариантах осуществления изобретения, указанным агентом является DM1 или DM4. В некоторых вариантах осуществления изобретения, указанным агентом является дуокармицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является калихеамицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является пирролобензодиазепин.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело также включает детектируемую группу. В некоторых вариантах осуществления изобретения, указанной детектируемой группой является диагностический агент.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело обычно содержит одну или более дисульфидных связей. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело может быть сконструировано так, чтобы оно содержало одну или более дисульфидных связей.
Настоящее изобретение также относится к выделенной молекуле нуклеиновой кислоты, кодирующей описанное здесь мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, а также к векторами, включающим эти выделенные последовательности нуклеиновой кислоты. Настоящее изобретение относится к способам продуцирования мультиспецифического антитела путем культивирования клеток в условиях, стимулирующих экспрессию антитела, где указанные клетки содержат такую молекулу нуклеиновой кислоты. В некоторых вариантах осуществления изобретения, указанная клетка содержит указанный вектор.
Настоящее изобретение также относится к композициям мультиспецифических активируемых антител и/или мультиспецифических активируемых антител, которые включают первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с первой мишенью или с первым эпитопом, и второе антитело или его антигенсвязывающий фрагмент (AB2), которые связываются со второй мишенью или со вторым эпитопом, где по меньшей мере AB1 связывается или как-либо иначе взаимодействует с маскирующей группой (MM), и такое связывание с MM1 приводит к снижению способности AB1 связываться с мишенью. В некоторых вариантах осуществления изобретения, MM1 связывается с AB1 посредством первой отщепляемой группы (CM1), которая включает последовательность субстрата для протеазы, например, протеазы, которая локализуется вместе с мишенью АВ1 в участке организма индивидуума, подвергаемом лечению или диагностике. Описанные здесь мультиспецифические активируемые антитела являются стабильными в кровотоке и активируются в нужных участках организма, подвергаемых терапии и/или диагностике, но не в здоровых участках организма, например, в здоровой ткани, и в случае активации этих антител, они связываются с мишенью АВ1 на уровне, по меньшей мере сравнимом с уровнем связывания соответствующего немодифицированного мультиспецифического антитела.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело содержит линкерный пептид, расположенный между ММ1 и CM1.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело содержит линкерный пептид, расположенный между CM1 и AB1.
В некоторых вариантах осуществления изобретения, активируемое антитело содержит первый линкерный пептид (LP1) и второй линкерный пептид (LP2), и по меньшей мере часть мультиспецифического активируемого антитела в нерасщепленном состоянии имеет следующую структуру, от N-конца до C-конца: MM1-LP1-CM1-LP2-AB1 или AB1-LP2-CM1-LP1-MM1. В некоторых вариантах осуществления изобретения, два линкерных пептида необязательно должны быть идентичными.
В некоторых вариантах осуществления изобретения, по меньшей мере один из LP1 и LP2 включает аминокислотную последовательность, выбранную из группы, состоящей из (GS)n, (GGS)n, (GSGGS)n (SEQ ID NO: 18) и (GGGS)n (SEQ ID NO: 19), где n равно целому числу по меньшей мере 1. В некоторых вариантах осуществления изобретения, по меньшей мере один из LP1 и LP2 включает аминокислотную последовательность, выбранную из группы, состоящей из GGSG (SEQ ID NO: 20), GGSGG (SEQ ID NO: 21), GSGSG (SEQ ID NO: 22), GSGGG (SEQ ID NO: 23), GGGSG (SEQ ID NO: 24) и GSSSG (SEQ ID NO: 25).
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с первой мишенью или с первым эпитопом, и второе антитело или его антигенсвязывающий фрагмент (AB2), которые специфически связываются со второй мишенью или со вторым эпитопом. В некоторых вариантах осуществления изобретения, каждый AB в мультиспецифическом активируемом антителе независимо выбран из группы, состоящей из моноклонального антитела, однодоменного антитела, одноцепочечного антитела, Fab-фрагмента, F(ab')2-фрагмента, scFv, scAb, dAb, тяжелой цепи однодоменного антитела и легкой цепи однодоменного антитела. В некоторых вариантах осуществления изобретения, каждый AB в мультиспецифическом активируемом антителе представляет собой моноклональное антитело грызунов (например, мышей или крыс), химерное моноклональное антитело, гуманизованное моноклональное антитело или полностью человеческое моноклональное антитело.
В некоторых вариантах осуществления изобретения, каждый AB в мультиспецифическом активируемом антителе имеет константу равновесной диссоциации при связывании с соответствующей мишенью или соответствующим эпитопом, составляющую приблизительно 100 нM или менее.
В некоторых вариантах осуществления изобретения, MM1 имеет константу равновесной диссоциации при связывании с соответствующим AB, которая превышает константу равновесной диссоциации при связывании AB с соответствующей мишенью или соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, MM1 имеет константу равновесной диссоциации при связывании с соответствующим AB, которая не превышает константу равновесной диссоциации при связывании AB с соответствующей мишенью или соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, MM1 не влияет на взаимодействие соответствующего AB с соответствующей мишенью или с соответствующим эпитопом или не конкурирует с соответствующим AB за связывание с соответствующей мишенью или с соответствующим эпитопом, если мультиспецифическое активируемое антитело находится в расщепленном состоянии.
В некоторых вариантах осуществления изобретения, MM1 представляет собой полипептид длиной приблизительно 2-40 аминокислот. В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе представляет собой полипептид длиной не более чем 40 аминокислот.
В некоторых вариантах осуществления изобретения, MM1 имеет полипептидную последовательность, отличающуюся от последовательности мишени для соответствующего AB.
В некоторых вариантах осуществления изобретения, MM1 имеет полипептидную последовательность, которая не более, чем на 50% идентична последовательности любого природного партнера по связыванию с соответствующим AB. В некоторых вариантах осуществления изобретения, MM1 имеет полипептидную последовательность, которая не более чем на 25% идентична последовательности любого природного партнера по связыванию с соответствующим AB. В некоторых вариантах осуществления изобретения, MM1 имеет полипептидную последовательность, которая не более чем на 10% идентична последовательности любого природного партнера по связыванию с соответствующим AB.
В некоторых вариантах осуществления изобретения, связывание MM1 снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM1 с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 20 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание MM1 снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM1 с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 40 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание MM1 снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM1 с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 100 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание MM1 снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM1 с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 1000 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание MM1 снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM1 с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 10000 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, MM1 представляет собой аминокислотную последовательность, выбранную из последовательности MM, представленной в описанных здесь примерах.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает по меньшей мере вторую маскирующую группу (MM2), которая ингибирует связывание AB2 с мишенью, если мультиспецифическое активируемое антитело присутствует в нерасщепленном состоянии, и вторую отщепляемую группу (CM2), связанную с AB2, где CM2 представляет собой полипептид, который служит в качестве субстрата для второй протеазы. В некоторых вариантах осуществления изобретения, CM2 представляет собой полипептид длиной не более, чем 15 аминокислот. В некоторых вариантах осуществления изобретения, вторая протеаза локализуется вместе со второй мишенью или вторым эпитопом в ткани, где вторая протеаза расщепляет CM2 в мультиспецифическом активируемом антителе при его обработке указанной второй протеазой. В некоторых вариантах осуществления изобретения, первая протеаза и вторая протеаза локализуются вместе с первой мишенью или с первым эпитопом и со второй мишенью или со вторым эпитопом в ткани. В некоторых вариантах осуществления изобретения, первая протеаза и вторая протеаза являются одинаковыми. В некоторых вариантах осуществления изобретения, CM1 и CM2 являются различными субстратами для одной и той же протеазы. В некоторых вариантах осуществления изобретения, протеаза выбрана из группы, состоящей из протеаз, представленных в таблице 3. В некоторых вариантах осуществления изобретения, первая протеаза и вторая протеаза являются различными протезами. В некоторых вариантах осуществления изобретения, первая протеаза и вторая протеаза являются различными протезами, выбранными из группы, состоящей из протез, представленных в таблице 3.
В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе, например, MM1 и по меньшей мере MM2, имеет константу равновесной диссоциации при связывании с соответствующим AB, превышающую константу равновесной диссоциации при связывании AB с соответствующией мишенью или соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе имеет константу равновесной диссоциации при связывании с соответствующим AB, которая не превышает константу равновесной диссоциации при связывании AB с соответствующей мишенью или соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе не влияет на взаимодействие соответствующего AB с соответствующей мишенью или с соответствующим эпитопом или не конкурирует с соответствующим AB за связывание с соответствующей мишенью или с соответствующим эпитопом, если мультиспецифическое активируемое антитело находится в расщепленном состоянии.
В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе представляет собой полипептид длиной приблизительно 2-40 аминокислот. В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе представляет собой полипептид длиной не более чем 40 аминокислот.
В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе имеет полипептидную последовательность, отличающуюся от последовательности мишени для соответствующего AB.
В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе имеет полипептидную последовательность, которая не более чем на 50% идентична последовательности любого природного партнера по связыванию с соответствующим AB. В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе имеет полипептидную последовательность, которая не более чем на 25% идентична последовательности любого природного партнера по связыванию с соответствующим AB. В некоторых вариантах осуществления изобретения, каждая MM в мультиспецифическом активируемом антителе имеет полипептидную последовательность, которая не более чем на 10% идентична последовательности любого природного партнера по связыванию с соответствующим AB.
В некоторых вариантах осуществления изобретения, связывание каждой MM снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 20 раз превышать Kd AB в отсутствии связывания MM с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание каждой MM снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 40 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание каждой MM снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 100 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание каждой MM снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 1000 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, связывание каждой MM снижает способность соответствующего AB связываться с мишенью или эпитопом, в результате чего константа диссоциации (Kd) AB, при связывании MM с соответствующей мишенью или с соответствующим эпитопом, будет по меньшей мере в 10000 раз превышать Kd AB в отсутствии связывания MM1 с соответствующей мишенью или с соответствующим эпитопом.
В некоторых вариантах осуществления изобретения, каждая MM представляет собой аминокислотную последовательность, выбранную из последовательности MM, представленной в описанных здесь примерах.
В некоторых вариантах осуществления изобретения, протеаза, которая отщепляет последовательность первой отщепляемой группы (CM1), локализована вместе с мишенью AB1 в мультиспецифическом активируемом антителе, присутствующем в ткани, и эта протеаза отщепляет CM1 в мультиспецифическом активируемом антителе, если такое мультиспецифическое активируемое антитело было обработано протеазой.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает более чем одну последовательность отщепляемой группы, и протеаза, которая отщепляет по меньшей мере одну последовательность отщепляемой группы, локализована вместе с мишенью по меньшей мере одной из областей AB в мультиспецифическом активируемом антителе, присутствующем в ткани, где указанная протеаза отщепляет CM в мультиспецифическом активируемом антителе, если такое мультиспецифическое активируемое антитело было обработано протеазой.
В некоторых вариантах осуществления изобретения, каждая CM, например, CM1 и по меньшей мере CM2, находится в мультиспецифическом активируемом антителе, и если она присутствует в неотщепленном состоянии, то уровень связывания мультиспецифического активируемого антитела с мишенью одной из областей АВ снижается так, что константа равновесной диссоциации по меньшей мере в 20 раз превышает константу равновесной диссоцииции при связывании немодифицированного АВ с мишенью и при связывании АВ в расщепленном состоянии с мишенью.
В некоторых вариантах осуществления изобретения, каждая CM находится в мультиспецифическом активируемом антителе, и если она присутствует в неотщепленном состоянии, то уровень связывания мультиспецифического активируемого антитела с мишенью одной из областей АВ снижается так, что константа равновесной диссоцииции по меньшей мере в 40 раз превышает константу равновесной диссоциации при связывании немодифицированного АВ с мишенью и при связывании АВ в расщепленном состоянии с мишенью.
В некоторых вариантах осуществления изобретения, каждая CM находится в мультиспецифическом активируемом антителе, и если она присутствует в неотщепленном состоянии, то уровень связывания мультиспецифического активируемого антитела с мишенью одной из областей АВ снижается так, что константа равновесной диссоцииции по меньшей мере в 50 раз превышает константу равновесной диссоцииции при связывании немодифицированного АВ с мишенью и при связывании АВ в расщепленном состоянии с мишенью.
В некоторых вариантах осуществления изобретения, каждая CM находится в мультиспецифическом активируемом антителе, и если она присутствует в неотщепленном состоянии, то уровень связывания мультиспецифического активируемого антитела с мишенью одной из областей АВ снижается так, что константа равновесной диссоцииции по меньшей мере в 100 раз превышает константу равновесной диссоцииции при связывании немодифицированного АВ с мишенью и при связывании АВ в расщепленном состоянии с мишенью.
В некоторых вариантах осуществления изобретения, каждая CM находится в мультиспецифическом активируемом антителе, и если она присутствует в неотщепленном состоянии, то уровень связывания мультиспецифического активируемого антитела с мишенью одной из областей АВ снижается так, что константа равновесной диссоцииции по меньшей мере в 200 раз превышает константу равновесной диссоцииции при связывании немодифицированного АВ с мишенью и при связывании АВ в расщепленном состоянии с мишенью.
В некоторых вариантах осуществления изобретения, каждая CM в мультиспецифическом активируемом антителе представляет собой полипептид длиной до 15 аминокислот.
В некоторых вариантах осуществления изобретения, по меньшей мере одна CM в мультиспецифическом активируемом антителе включает аминокислотную последовательность LSGRSDNH (SEQ ID NO: 26). В некоторых вариантах осуществления изобретения, по меньшей мере одна отщепляемая группа была обработана специфической протеазой, например, протеазой, которая, как известно, локализуется вместе по меньшей мере с одной мишенью мультиспецифического активируемого антитела. Так, например, подходящие отщепляемые группы, присутствующие в мультиспецифических активируемых антителах согласно изобретению, отщепляются по меньшей мере такой протеазой как урокиназа, легумаин и/или матриптаза (также обозначаемая здесь MT-SP1 или MTSP1). В некоторых вариантах осуществления изобретения, подходящая отщепляемая группа включает по меньшей мере одну из следующих последовательностей: TGRGPSWV (SEQ ID NO: 27); SARGPSRW (SEQ ID NO: 28); TARGPSFK (SEQ ID NO: 29); LSGRSDNH (SEQ ID NO: 26); GGWHTGRN (SEQ ID NO: 30); HTGRSGAL (SEQ ID NO: 31); PLTGRSGG (SEQ ID NO: 32); AARGPAIH (SEQ ID NO: 33); RGPAFNPM (SEQ ID NO: 34); SSRGPAYL (SEQ ID NO: 35); RGPATPIM (SEQ ID NO: 36); RGPA (SEQ ID NO: 37); GGQPSGMWGW (SEQ ID NO: 38); FPRPLGITGL (SEQ ID NO: 39); VHMPLGFLGP (SEQ ID NO: 40); SPLTGRSG (SEQ ID NO: 41); SAGFSLPA (SEQ ID NO: 42); LAPLGLQRR (SEQ ID NO: 43); SGGPLGVR (SEQ ID NO: 44) и/или PLGL (SEQ ID NO: 45).
В некоторых вариантах осуществления изобретения, каждая CM в мультиспецифическом активируемом антителе служит субстратом для протеазы, выбранной из группы, состоящей из протеаз, представленных в таблице 3. В некоторых вариантах осуществления изобретения, протеаза выбрана из группы, состоящей из uPA, легумаина, MT-SP1, ADAM17, BMP-1, TMPRSS3, TMPRSS4, эластазы нейтрофилов, MMP-7, MMP-9, MMP-12, MMP-13 и MMP-14. В некоторых вариантах осуществления изобретения, протеазой является катепсин, такой как, но не ограничивающийся им, катепсин S. В некоторых вариантах осуществления изобретения, каждая CM в мультиспецифическом активируемом антителе служит субстратом для протеазы, выбранной из группы, состоящей из uPA (урокиназного активатора плазминогена), легумаина и MT-SP1 (матриптазы). В некоторых вариантах осуществления изобретения, протеазой является uPA. В некоторых вариантах осуществления изобретения, протеазой является легумаин. В некоторых вариантах осуществления изобретения, протеазой является MT-SP1. В некоторых вариантах осуществления изобретения, протеазой является металлопротеиназа матрикса (MMP).
В некоторых вариантах осуществления изобретения, по меньшей мере одна CM в мультиспецифическом активируемом антителе служит субстратом по меньшей мере для двух протеаз. В некоторых вариантах осуществления изобретения, каждая протеаза выбрана из группы, состоящей из протеаз, представленных в таблице 3. В некоторых вариантах осуществления изобретения, по меньшей мере одна CM в мультиспецифическом активируемом антителе служит субстратом по меньшей мере для двух протеаз, где одна из протеаз выбрана из группы, состоящей из uPA, легумаина и MT-SP1, а другая протеаза выбрана из группы, состоящей из протеаз, представленных в таблице 3. В некоторых вариантах осуществления изобретения, по меньшей мере одна CM в мультиспецифическом активируемом антителе служит субстратом по меньшей мере для двух протеаз, выбранных из группы, состоящей из uPA, легумаина и MT-SP1.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает по меньшей мере первую CM (CM1) и вторую CM (CM2). В некоторых вариантах осуществления изобретения, CM1 и CM2 представляют собой часть одного расщепляемого линкера, который соединяет MM с AB. В некоторых вариантах осуществления изобретения, CM1 представляет собой часть расщепляемого линкера, который соединяет MM1 с AB1, а CM2 представляет собой часть отдельного расщепляемого линкера, который соединяет MM2 с AB2. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело содержит более чем две CM. В некоторых вариантах осуществления изобретения, такое мультиспецифическое активируемое антитело содержит более чем две CM и более, чем две MM. В некоторых вариантах осуществления изобретения, CM1 и CM2 представляют собой полипептиды длиной не более чем 15 аминокислот. В некоторых вариантах осуществления изобретения, по меньшей мере одна из первой CM и второй CM представляет собой полипептид, который служит субстратом для протеазы, выбранной из группы, состоящей из протеаз, перечисленных в таблице 3. В некоторых вариантах осуществления изобретения, по меньшей мере одна из первой CM и второй CM представляет собой полипептид, который служит субстратом для протеазы, выбранной из группы, состоящей из uPA, легумаина и MT-SP1. В некоторых вариантах осуществления изобретения, первая CM отщепляется под действием первого расщепляющего агента, выбранного из группы, состоящей из uPA, легумаина и MT-SP1, в ткани-мишени, а вторая CM отщепляется под действием первого расщепляющего агента в ткани-мишени. В некоторых вариантах осуществления изобретения, другая протеаза выбрана из группы, состоящей из протеаз, представленных в таблице 3. В некоторых вариантах осуществления изобретения, первый расщепляющий агент и второй расщепляющий агент представляют собой одну и ту же протеазу, выбранную из группы, состоящей из протеаз, перечисленных в таблице 3, а первая СМ и вторая СМ представляют собой различные субстраты фермента. В некоторых вариантах осуществления изобретения, первый расщепляющий агент и второй расщепляющий агент представляют собой одну и ту же протеазу, выбранную из группы, состоящей из uPA, легумаина и MT-SP1, а первая СМ и вторая СМ представляют собой различные субстраты фермента. В некоторых вариантах осуществления изобретения, первый расщепляющий агент и второй расщепляющий агент представляют собой одну и ту же протеазу, выбранную из группы, состоящей из протеаз, перечисленных в таблице 3, а первая СМ и вторая СМ представляют собой один и тот же субстрат. В некоторых вариантах осуществления изобретения, первый расщепляющий агент и второй расщепляющий агент представляют собой различные протеазы. В некоторых вариантах осуществления изобретения, первый расщепляющий агент и второй расщепляющий агент представляют собой различные протеазы, выбранные из группы, состоящей из протеаз, представленных в таблице 3. В некоторых вариантах осуществления изобретения, первый расщепляющий агент и второй расщепляющий агент находятся в одной ткани-мишени. В некоторых вариантах осуществления изобретения, первая СМ и вторая СМ отщепляются по меньшей мере одним расщепляющим агентом в ткани-мишени.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело обрабатывают и подвергают расщеплению протеазой так, чтобы это активированное мультиспецифическое активируемое антитело в активированном или расщепленном состоянии включало аминокислотную последовательность легкой цепи, содержащую по меньшей мере часть последовательности LP2 и/или CM после отщепления CM протеазой.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело также включает сигнальный пептид. В некоторых вариантах осуществления изобретения, сигнальный пептид конъюгирован с мультиспецифическим активируемым антителом посредством спейсера. В некоторых вариантах осуществления изобретения, спейсер конъюгирован с мультиспецифическим активируемым антителом в отсутствии сигнального пептида. В некоторых вариантах осуществления изобретения, спейсер непосредственно присоединен по меньшей мере к одной из MM мультиспецифического активируемого антитела.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело в нерасщепленном состоянии содержит спейсер, который непосредственно присоединен к первой MM и имеет следующую структуру, от N-конца до С-конца: спейсер-MM1-CM-AB1. В некоторых вариантах осуществления изобретения, спейсер содержит по меньшей мере аминокислотную последовательность QGQSGQ (SEQ ID NO: 46).
В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке превышает время полужизни соответствующего мультиспецифического антитела, например, pK мультиспецифического активируемого антитела в сыворотке превышает pK соответствующего мультиспецифического антитела. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке аналогично времени полужизни соответствующего мультиспецифического антитела. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 15 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 12 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 11 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 10 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 9 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 8 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 7 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 6 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 5 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 4 дня. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 3 дня. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 2 дня. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 24 часа. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 20 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 18 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 16 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 14 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 12 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 10 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 8 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 6 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 4 часа. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 3 часа.
Настоящее изобретение также относится к композициям, включающим мультиспецифическое активируемое антитело, и к способам, включающим применение мультиспецифического активируемого антитела, где указанное мультиспецифическое активируемое антитело включает по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью, и второе антитело или его фрагмент (AB2), где по меньшей мере первое AB в мультиспецифическом активируемом антителе связывается с маскирующей группой (MM1), что приводит к снижению способности AB1 связываться с мишенью. В некоторых вариантах осуществления изобретения, каждый AB связывается с MM, что приводит к снижению способности соответствующего АВ связываться с каждой мишенью. Так, например, в вариантах, относящихся к биспецифическому активируемому антителу, AB1 связывается с первой маскирующей группой (MM1), что приводит к снижению способности AB1 связываться с мишенью, а AB2 связывается со второй маскирующей группой (MM1), что приводит к снижению способности AB2 связываться с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело содержит более, чем две области AB; и в таких вариантах осуществления изобретения, AB1 связывается с первой маскирующей группой (MM1), что приводит к снижению способности AB1 связываться с мишенью, AB2 связывается со второй маскирующей группой (MM1), что приводит к снижению способности AB2 связываться с мишенью, AB3 связывается с третьей маскирующей группой (MM3), что приводит к снижению способности AB3 связываться с мишенью, и так далее, для каждого AB в мультиспецифическом активируемом антителе.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело также содержит по меньшей мере одну отщепляемую группу (CM), которая служит субстратом для протеазы, где CM соединяет MM с AB. Так, например, в некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает по меньшей мере первое антитело или его фрагмент (AB1), которые специфически связываются с мишенью, и второе антитело или его фрагмент (AB2), где по меньшей мере первое AB в мультиспецифическом активируемом антителе связывается с маскирующей группой (MM1) посредством первой отщепляемой группы (CM1), что приводит к снижению способности AB1 связываться с мишенью. В некоторых вариантах, относящихся к биспецифическому активируемому антителу, AB1 связывается с MM1 посредством СМ1, а AB2 связывается со второй маскирующей группой (MM2) посредством второй отщепляемой группы (СМ2), что приводит к снижению способности AB2 связываться с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело содержит более чем две области AB; и в некоторых этих вариантах осуществления изобретения, AB1 связывается с MM1 посредством СМ1, AB2 связывается с MM2 посредством СМ2, а AB3 связывается с третьей маскирующей группой (ММ3) посредством третьей отщепляемой группы (СМ3), что приводит к снижению способности AB3 связываться с мишенью, и так далее, для каждого AB в мультиспецифическом активируемом антителе.
Описанные здесь композиции и способы позволяют присоединять один или более агентов к одному или более цистеиновым остаткам в любой из областей АВ, причем, такое присоединение не оказывает негативного влияния на активность (например, маскирующую, активирующую или связывающую активность) мультиспецифического активируемого антитела. В некоторых вариантах осуществления изобретения, описанные здесь композиции и способы позволяют осуществлять присоединение одного или более агентов к одному или более цистеиновым остаткам в любой из областей AB без восстановления или какого-либо разрыва одной или более дисульфидных связей в любой MM. Описанные здесь композиции и способы позволяют получить мультиспецифическое активируемое антитело, конъюгированное с одним или более агентами, например, с любыми различными терапевтическими, диагностическими и/или профилактическими агентами, а предпочтительно, без использования какого-либо (каких-либо) агента(ов), конъюгированного(ых) с ММ мультиспецифического активируемого антитела. Описанные здесь композиции и способы позволяют получить конъюгированные мультиспецифические активируемые антитела, в которых каждая MM сохраняет способность к достаточно эффективной маскировке соответствующей АВ мультиспецифического активируемого антитела в расщепленном состоянии. Описанные здесь композиции и способы позволяют получить конъюгаты мультиспецифических активируемых антител, в которых активируемое антитело все еще остается активированным, то есть расщепляется в присутствии протеазы, которая может отщеплять CM.
Мультиспецифические активируемые антитела имеют по меньшей мере один сайт для конъюгирования агента, но в описанных здесь способах и композициях, не все сайты из множества возможных сайтов конъюгирования являются доступными для конъюгирования с агентом. В некоторых вариантах осуществления изобретения, один или более сайтов конъюгирования представляют собой атомы серы, участвующие в образовании дисульфидных связей. В некоторых вариантах осуществления изобретения, один или более сайтов конъюгирования представляют собой атомы серы, участвующие в образовании межцепьевых сульфидных связей. В некоторых вариантах осуществления изобретения, один или более сайтов конъюгирования представляют собой атомы серы, участвующие в образовании межцепьевых дисульфидных связей, но при этом, не все атомы серы участвуют в образовании внутрицепьевых дисульфидных связей. В некоторых вариантах осуществления изобретения, один или более сайтов конъюгирования представляют собой атомы серы цистеина или других аминокислотных остатков, содержащих атом серы. Такие остатки могут присутствовать в структуре природных антител, либо они могут быть введены в антитело посредством сайт-направленного мутагенеза, химического превращения или ошибочного включения не-природных аминокислот.
Настоящее изобретение также относится к способам получения конъюгата мультиспецифического активируемого антитела, имеющего одну или более межцепьевых дисульфидных связей в одном или более АВ и одну или более внутрицепьевых дисульфидных связей в соответствуюей MM, и содержащего лекарственное средство, реагирующее со свободными тиолами. Указанный способ, по существу, включает частичное восстановление межцепьевых дисульфидных связей в активируемом антителе под действием восстановителя, такого как, например, TCEP; и конъюгирование лекарственного средства, реагирующего со свободными тиолами, с частично восстановленным активируемым антителом. Используемый здесь термин «частичное восстановление» означает ситуацию, в которой при контактировании мультиспецифического активируемого антитела с восстановителем, не все дисульфидные связи, например, не все возможные сайты конъюгирования являются восстановленными. В некоторых вариантах осуществления изобретения, восстановленными сайтами конъюгирования являются менее чем 99%, 98%, 97%, 96%, 95%, 90%, 85%, 80%, 75%, 70%, 65%, 60%, 55%, 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10% или менее чем 5% из всех возможных сайтов конъюгирования.
В других своих вариантах, настоящее изобретение относится к способу восстановления и конъюгирования агента, например, лекарственного средства с мультиспецифическим активируемым антителом, в целях селективной локализации данного агента. В общих чертах, этот способ включает частичное восстановление мультиспецифического активируемого антитела восстановителем так, чтобы какие-либо сайты конъюгирования в любых маскирующих группах или в другой не-АВ-части активируемого антитела оставались невосстановленными; и конъюгирование агента с межцепьевыми тиоалми в одной или более областях AB мультиспецифического активируемого антитела. Сайт(ы) конъюгирования выбирают так, чтобы он(и) представлял(и) собой желаемое положение, в котором конъюгирование данного агента происходит в желаемом сайте. Восстановителем является, например, TCEP. Условия реакции восстановления, такие как, например, отношение восстановителя к активируемому антителу, время инкубирования, температура в процессе инкубирования, pH реакционного раствора для восстановления и т.п., определяют, например, путем идентификации условий, позволяющих продуцировать конъюгированное активируемое антитело, при которых ММ сохраняет способность результативно и эффективно маскировать AB активируемого антитела в нерасщепленном состоянии. Отношение восстановителя к мультиспецифическому активируемому антителу варьируется в зависимости от типа активируемого антитела. В некоторых вариантах осуществления изобретения, отношение восстановителя к мультиспецифическому активируемому антителу составляет в пределах приблизительно от 20:1 до 1:1, приблизительно от 10:1 до 1:1, приблизительно от 9:1 до 1:1, приблизительно от 8:1 до 1:1, приблизительно от 7:1 до 1:1, приблизительно от 6:1 до 1:1, приблизительно от 5:1 до 1:1, приблизительно от 4:1 до 1:1, приблизительно от 3:1 до 1:1, приблизительно от 2:1 до 1:1, приблизительно от 20:1 до 1:1,5, приблизительно от 10:1 до 1:1,5, приблизительно от 9:1 до 1:1,5, приблизительно от 8:1 до 1:1,5, приблизительно от 7:1 до 1:1,5, приблизительно от 6:1 до 1:1,5, приблизительно от 5:1 до 1:1,5, приблизительно от 4:1 до 1:1,5, приблизительно от 3:1 до 1:1,5, приблизительно от 2:1 до 1:1,5, приблизительно от 1,5:1 до 1:1,5 или приблизительно от 1:1 до 1:1,5. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 5:1 до 1:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 5:1 до 1,5:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 4:1 до 1:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 4:1 до 1,5:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 8:1 до 1:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 2,5:1 до 1:1.
В некоторых своих вариантах, настоящее изобретение относится к способу восстановления межцепьевых дисульфидных связей в одной или более областях AB мультиспецифического активируемого антитела и к способу конъюгирования агента, например, тиол-содержащего агента, такого как лекарственное средство, с полученными межцепьевыми тиолами, для селективной локализации агента(ов) на AB. Этот способ, в общих чертах, включает частичное восстановление одной или более областей AB восстановителем с образованием по меньшей мере двух межцепьевых тиолов, но без образования всех возможных межцепьевых тиолов в активируемом антителе, и конъюгирование агента с межцепьевыми тиолами частично восстановленного АВ. Так, например, частичное восстанавление одной или более областей AB мультиспецифического активируемого антитела осуществляют приблизительно в течение 1 часа приблизительно при 37°C в желаемом отношении «восстановитель:активируемое антитело». В некоторых вариантах осуществления изобретения, отношение восстановителя к активируемому антителу составляет в пределах приблизительно от 20:1 до 1:1, приблизительно от 10:1 до 1:1, приблизительно от 9:1 до 1:1, приблизительно от 8:1 до 1:1, приблизительно от 7:1 до 1:1, приблизительно от 6:1 до 1:1, приблизительно от 5:1 до 1:1, приблизительно от 4:1 до 1:1, приблизительно от 3:1 до 1:1, приблизительно от 2:1 до 1:1, приблизительно от 20:1 до 1:1,5, приблизительно от 10:1 до 1:1,5, приблизительно от 9:1 до 1:1,5, приблизительно от 8:1 до 1:1,5, приблизительно от 7:1 до 1:1,5, приблизительно от 6:1 до 1:1,5, приблизительно от 5:1 до 1:1,5, приблизительно от 4:1 до 1:1,5, приблизительно от 3:1 до 1:1,5, приблизительно от 2:1 до 1:1,5, приблизительно от 1,5:1 до 1:1,5 или приблизительно от 1:1 до 1:1,5. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 5:1 до 1:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 5:1 до 1,5:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 4:1 до 1:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 4:1 до 1,5:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 8:1 до 1:1. В некоторых вариантах осуществления изобретения, такое отношение составляет в пределах приблизительно от 2,5:1 до 1:1.
Тиол-содержащим реагентом может быть, например, цистеин или N-ацетилцистеин. Восстановителем может быть, например, TCEP. В некоторых вариантах осуществления изобретения, восстановленное активируемое антитело может быть очищено перед конъюгированием, например, с помощью колоночной хроматографии, диализа или диафильтрации. Альтернативно, восстановленное антитело не очищают после частичного восстановления и перед конъюгированием.
Настоящее изобретение также относится к частично восстановленным мультиспецифическим активируемым антителам, в которых по меньшей мере одна межцепьевая дисульфидная связь была восстановлена под действием восстановителя без разрыва каких-либо внутрицепьевых дисульфидных связей в мультиспецифическом активируемом антителе, где мультиспецифическое активируемое антитело включает по меньшей мере первое антитело или его антигенсвязывающий фрагмент (AB1), которые специфически связываются с мишенью; первую маскирующую группу (MM1), которая ингибирует связывание AB1 мультиспецифического активируемого антитела в нерасщепленном состоянии с мишенью; первую отщепляемую группу (CM1), связанную с AB1, где CM1 представляет собой полипептид, который служит в качестве субстрата для протеазы, и второе антитело или его антигенсвязывающий фрагмент (AB2), которые специфически связываются со второй мишенью. В некоторых вариантах осуществления изобретения, MM1 связывается с AB1 посредством CM1. В некоторых вариантах осуществления изобретения, одна или более внутрицепьевых дисульфидных связей мультиспецифического активируемого антитела не разрываются под действием восстановителя. В некоторых вариантах осуществления изобретения, одна или более внутрицепьевых дисульфидных связей ММ1 мультиспецифического активируемого антитела не разрываются под действием восстановителя. В некоторых вариантах осуществления изобретения, восстановителем является TCEP.
В некоторых вариантах осуществления изобретения, описанные здесь мультиспецифические антитела и/или мультиспецифические активируемые антитела используют в сочетании с одним или более дополнительными агентами или комбинацией дополнительных агентов. Подходящими дополнительными агентами являются современные фармацевтические средства и/или средства, используемые в хирургии, например, для удаления раковой опухоли. Так, например, мультиспецифические антитела и/или мультиспецифические активируемые антитела могут быть использованы в комбинации с дополнительным химиотерапевтическим или противоопухолевым агентом.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент приготавливают в виде одной терапевтической композиции, и такое мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент вводят одновременно. Альтернативно, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент вводят отдельно друг от друга, например, каждый из них приготавливают в виде отдельной терапевтической композиции, и мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент вводят одновременно, либо мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент вводят в различные периоды времени в течение всего курса лечения. Так, например, либо мультиспецифическое антитело и/или мультиспецифическое активируемое антитело вводят перед введением дополнительного агента, либо мультиспецифическое антитело и/или мультиспецифическое активируемое антитело вводят после введения дополнительного агента, либо мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент вводят поочередно. Как описано в настоящей заявке, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и дополнительный агент вводят в разовых дозах или в дробных дозах.
Настоящее изобретение также относится к выделенной молекуле нуклеиновой кислоты, кодирующей описанное здесь мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, а также к векторам, содержащим эти выделенные последовательности нуклеиновой кислоты. Настоящее изобретение относится к способам продуцирования мультиспецифического антитела и/или мультиспецифического активируемого антитела путем культивирования клетки в условиях, стимулирующих экспрессию мультиспецифического антитела и/или мультиспецифического активируемого антитела, где указанная клетка содержит указанную молекулу нуклеиновой кислоты. В некоторых вариантах осуществления изобретения, указанная клетка содержит этот вектор.
Настоящее изобретение также относится к способу получения мультиспецифических антител и/или мультиспецифических активируемых антител согласно изобретению путем (a) культивирования клетки, содержащей конструкцию нуклеиновой кислоты, кодирующую мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, в условиях, стимулирующих экспрессию мультиспецифического антитела и/или мультиспецифического активируемого антитела, и (b) выделения мультиспецифического антитела и/или мультиспецифического активируемого антитела.
Настоящее изобретение также относится к способам лечения, предупредупреждения, замедления прогрессирования или какого-либо другого ослабления симптома одной или более патологий или ослабления симптома, ассоциированного с такими патологиями, путем введения мультиспецифического антитела и/или мультиспецифического активируемого антитела согласно изобретению индивидууму, нуждающемуся в таком лечении или предупреждении заболевания. Индивидуумом, подвергаемым лечению, является, например, человек или другое млекопитающее. В некоторых вариантах осуществления изобретения, указанным индивидуумом является любое млекопитающее, кроме человека, такое как примат, не являющийся человеком, животное-компаньон (например, кошка, собака, лошадь), сельскохозяйственное животное, рабочий скот или животное, содержащееся в зоопарке. В некоторых вариантах осуществления изобретения, указанным животным является грызун.
Настоящее изобретение также относится к способам индуцирования мишень-зависимой активации Т-клеток и к цитолизу клеток-мишеней путем введения мультиспецифического активируемого антитела согласно изобретению индивидууму, нуждающемуся в таком индуцировании, где в случае, если мультиспецифическое активируемое антитело присутствует в расщепленном состоянии, например, если каждая маскирующая группа в мультиспецифическом активируемом антителе больше не связана или как-либо иначе не ассоциирована с соответствующим доменом АВ, то это указывает на мишень-зависимую активацию Т-клеток и цитолиз клеток-мишеней, и в случае, если мультиспецифическое активируемое антитело присутствует в нерасщепленном состоянии, например, если по меньшей мере одна маскирующая группа в мультиспецифическом активируемом антителе связана или как-либо иначе ассоциирована с соответствующим доменом АВ, то это указывает на снижение или какое-либо другое ингибирование мишень-зависимой активации Т-клеток и цитолиза клеток-мишеней. В этих способах могут применяться любые описанные здесь мультиспецифические активируемые антитела. Индивидуумом, подвергаемым лечению, является, например, человек или другое млекопитающее. В некоторых вариантах осуществления изобретения, указанным индивидуумом является любое млекопитающее, кроме человека, такое как примат, не являющийся человеком, животное-компаньон (например, кошка, собака, лошадь), сельскохозяйственное животное, рабочий скот или животное, содержащееся в зоопарке. В некоторых вариантах осуществления изобретения, указанным животным является грызун.
Мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, используемое в любом из вариантов этих способов и применений, может быть введено на любой стадии заболевания и/или на любой стадии, в которой активация Т-клеток и цитолиз клеток-мишеней являются желательными. Так, например, такое мультиспецифическое антитело и/или мультиспецифическое активируемое антитело может быть введено пациенту, страдающему раком на любой стадии, то есть, на ранней стадии или на стадии появления метастазов. Используемые здесь термины «индивидуум» и «пациент» являются синонимами. Мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, используемое в любом из вариантов этих способов и применений, может быть введено для проведения курса лечения в соответствии со схемой неоадъювантной терапии. Мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, используемое в любом из вариантов этих способов и применений, может быть введено отдельно или в комбинации с одним или более дополнительными агентами, используемыми в терапии, включая терапию на основе низкомолекулярных ингибиторов, других антител, полипептидов или пептидов, нуклеиновых кислот и/или других биологических соединений.
Настоящее изобретение также относится к способам и наборам, в которых используются мультиспецифические антитела и/или мультиспецифические активируемые антитела в различных диагностических и/или профилактических целях. Так, например, настоящее изобретение относится к способам и наборам для детектирования присутствия или отсутствия расщепляющего агента и представляющей интерес мишени в организме индивидуума или в образце посредством (i) контактирования индивидуума или образца с мультиспецифическим активируемым антителом, которое включает по меньшей мере первую маскирующую группу (MM1), первую отщепляемую группу (CM1), которая отщепляется под действием расщепляющего агента, и по меньшей мере первый антигенсвязывающий домен или его фрагмент (AB1), которые специфически связываются с представляющей интерес мишенью, и второй антигенсвязывающий домен или его фрагмент (AB2), которые специфически связываются со второй мишенью и/или со вторым эпитопом, где (a) MM1 представляет собой пептид, который ингибирует связывание AB1 с мишенью, и где MM1 не имеет аминокислотной последовательности природного партнера по связыванию с AB1 и не является модифицированной формой природного партнера по связыванию с AB1; и где (b) MM1, если она присутствует в нерасщепленном и неактивированном состоянии, негативно влияет на специфическое связывание AB1 с мишенью, а если она присутствует в отщепленном и активированном состоянии, то она не влияет на специфическое связывание AB1 с мишенью или не конкурирует с AB1 за специфическое связывание с мишенью; и (ii) измерения уровня активированного мультиспецифического активируемого антитела в организме индивидуума или в образце, где детектируемый уровень активированного мультиспецифического активируемого антитела у индивидуума или в образце указывает на присутствие расщепляющего агента и мишени у индивидуума или в образце, и где отсутствие такого детектируемого уровня активированного мультиспецифического активируемого антитела у индивидуума или в образце указывает на отсутствие и/или на недостаточное присутствие расщепляющего агента, мишени или расщепляющего агента и мишени у индивидуума или в образце.
В некоторых вариантах осуществления изобретения, активированное мультиспецифическое активируемое антитело представляет собой активированное мультиспецифическое активируемое антитело, конъюгированное с терапевтическим агентом. В некоторых вариантах осуществления изобретения, активированное мультиспецифическое активируемое антитело не конъюгировано с агентом. В некоторых вариантах осуществления изобретения, активированное мультиспецифическое активируемое антитело содержит детектируемую метку. В некоторых вариантах осуществления изобретения, такая детектируемая метка находистся на AB1. В некоторых вариантах осуществления изобретения, измерение уровня активированного мультиспецифического активируемого антитела у индивидуума или в образце осуществляют с использованием второго реагента, который специфически связывается с активированным мультиспецифическим активируемым антителом, где указанный реагент содержит детектируемую метку. В некоторых вариантах осуществления изобретения, указанным вторым реагентом является антитело, содержащее детектируемую метку.
В некоторых вариантах этих способов и наборов, активированное мультиспецифическое активируемое антитело включает детектируемую метку. В некоторых вариантах этих способов и наборов, детектируемой меткой является визуализирующий агент, агент для контрастного окрашивания, фермент, флуоресцентная метка, хромофор, краситель, один или более ионов металла или метка на основе лиганда. В некоторых вариантах этих способов и наборов, визуализирующий агент включает радиоизотоп. В некоторых вариантах этих способов и наборов, радиоизотопом является индий или технеций. В некоторых вариантах этих способов и наборов, агент для контрастного окрашивания включает йод, гадолиний или оксид железа. В некоторых вариантах этих способов и наборов, фермент представляет собой пероксидазу хрена, щелочную фосфатазу или β-галактозидазу. В некоторых вариантах этих способов и наборов, флуоресцентная метка включает белок, флуоресцирующий в желтом диапазоне спектра (YFP); белок, флуоресцирующий в сине-зеленом диапазоне спектра (CFP); белок, флуоресцирующий в зеленом диапазоне спектра (GFP); модифицированный белок, флуоресцирующий в красном диапазоне спектра (mRFP); t-димер 2 белка, флуоресцирующего в красном диапазоне спектра (RFP tdimer2), HCRED или производное европия. В некоторых вариантах этих способов и наборов, люминесцентная метка включает производное N-метилакридиния. В некоторых вариантах этих способов, указанной меткой является метка Alexa Fluor®, такая как Alex Fluor® 680 или Alexa Fluor® 750. В некоторых вариантах этих способов и наборов, метка на основе лиганда включает биотин, авидин, стрептавидин или один или более гаптенов.
В некоторых вариантах этих способов и наборов, указанным индивидуумом является млекопитающее. В некоторых вариантах этих способов и наборов, указанным индивидуумом является человек. В некоторых вариантах осуществления изобретения, указанным индивидуумом является любое млекопитающее, кроме человека, такое как примат, не являющийся человеком, животное-компаньон (например, кошка, собака, лошадь), сельскохозяйственное животное, рабочий скот или животное, содержащееся в зоопарке. В некоторых вариантах осуществления изобретения, указанным животным является грызун.
В некоторых вариантах этих способов, указанным способом является способ in vivo. В некоторых вариантах этих способов, указанным способом является способ in situ. В некоторых вариантах этих способов, указанным способом является способ ex vivo. В некоторых вариантах этих способов, указанным способом является способ in vitro.
В некоторых вариантах этих способов и наборов, указанный способ или набор применяется для идентификации или какого-либо другого определения группы пациентов, подходящих для введения им мультиспецифического активируемого антитела согласно изобретению. Так, например, пациенты с положительным тестом на мишень и протеазу, которая расщепляет субстрат в первой отщепляемой группе (CM1) мультиспецифического активируемого антитела, тестируемого этими способами, были идентифицированы как подходящие кандидаты на проведение лечения с использованием указанного мультиспецифического активируемого антитела, содержащего такую группу CM1. Аналогичнным образом, пациенты с отрицательным тестом на мишень и протеазу, которая расщепляет субстрат в CM1 мультиспецифического активируемого антитела, тестируемого этими способами, могут быть идентифицированы как подходящие кандидаты на проведение другой формы терапии.
В некоторых вариантах осуществления изобретения, указанный способ или набор применяется для идентификации или какого-либо другого определения группы пациентов, подходящих для введения им мультиспецифического активируемого антитела и/или конъюгированного мультиспецифического активируемого антитела (например, мультиспецифического активируемого антитела, конъюгированного с терапевтическим агентом) согласно изобретению, и проведения последующего лечения путем введения индивидууму, нуждающемуся в этом, мультиспецифического антитела и/или мультиспецифического активируемого антитела и/или конъюгированного мультиспецифического активируемого антитела. Так, например, пациенты с положительным тестом на мишень и протеазу, которая расщепляет субстрат в первой отщепляемой группе (CM1) мультиспецифического активируемого антитела и/или конъюгированного мультиспецифического активируемого антитела, тестируемого этими способами, могут быть идентифицированы как подходящие кандидаты на проведение лечения с использованием указанного мультиспецифического активируемого антитела, содержащего указанную группу CM1, и/или конъюгированного мультиспецифического активируемого антитела, содержащего указанную группу CM1, и этим пациентам затем может быть введено терапевтически эффективное количество мультиспецифического активируемого антитела и/или конъюгированного мультиспецифического активируемого антитела, которое было протестировано ранее. Аналогичнным образом, пациенты с отрицательным тестом на мишень или протеазу или на мишень и протеазу, которая расщепляет субстрат в CM1 мультиспецифического активируемого антитела и/или конъюгированного мультиспецифического активируемого антитела, тестируемого этими способами, могут быть идентифицированы как подходящие кандидаты на проведение другой формы терапии.
В некоторых вариантах осуществления изобретения, такие пациенты могут быть обследованы на возможность введения им других мультиспецифических антител и/или мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител до тех пор, пока не будут идентифицированы мультиспецифическое активируемое антитело и/или конъюгированное мультиспецифическое активируемое антитело, подходящее для лечения, например, мультиспецифическое активируемое антитело и/или конъюгированное мультиспецифическое активируемое антитело, содержащее CM, которая отщепляется в пораженном участке организма пациента. В некоторых вариантах осуществления изобретения, затем, пациенту с положительным тестом вводят терапевтически эффективное количество мультиспецифического активируемого антитела и/или конъюгированного мультиспецифического активируемого антитела.
Фармацевтические композиции согласно изобретению могут включать мультиспецифическое антитело и/или мультиспецифическое активируемое антитело согласно изобретению и носитель. Эти фармацевтические композиции могут быть включены в наборы, например, в диагностические наборы.
Для специалиста в данной области очевидно, что антитела согласно изобретению могут быть применены в различных целях. Так, например, белки согласно изобретению могут быть использованы в качестве терапевтических агентов для лечения различных расстройств. Антитела согласно изобретению могут быть также использованы в качестве реагентов в диагностических наборах или в качестве диагностических средств, либо эти антитела могут быть использованы в анализах на конкурентное связывание для получения терапевтических реагентов.
Краткое описание чертежей
На фигуре 1 проиллюстрированы биспецифические антитела в различных форматах (адаптированные по Chan and Carter, Nat. Rev. Immunol. 2010).
На фигурах 2A-2C представлена серия изображений, иллюстрирующих мультиспецифические антитела в различных форматах, которые могут быть использованы в композициях и способах согласно изобретению. В биспецифическом антителе, scFv может быть присоединен к карбокси-концу тяжелой цепи (Hc) антитела IgG (фигура 2A); к карбокси-концу легкой цепи (Lc) (фигура 2B) или к карбокси-концу тяжелой и легкой цепей (фигура 2C). На фигуре 2D представлена диаграмма одной генетической конструкции, используемой для экспрессии гибридных мультиспецифических антител.
На фигуре 3A представлена серия изображений, иллюстрирующих мультиспецифические активируемые антитела в различных форматах, которые могут быть использованы в композициях и способах согласно изобретению. На фигуре 3B представлена диаграмма генетических конструкций, используемых для экспрессии гибридных мультиспецифических активируемых антител.
На фигуре 4, панелях A-J, схематически представлена серия диаграмм для выбранного ряда возможных пермутаций в мультиспецифических антителах согласно изобретению. В серых заштрихованных рамках представлены конфигурации, проиллюстрированные на фигуре 2 и приведенные здесь для полноты сравнения.
На фигуре 5, панелях A-J, схематически представлена серия диаграмм для выбранного ряда возможных пермутаций в мультиспецифических антителах согласно изобретению. В серых заштрихованных рамках представлены конфигурации, проиллюстрированные на фигуре 3 и приведенные здесь для полноты сравнения. В частности, на этой фигуре проиллюстрированы мультиспецифические активируемые антитела, в которых первый антигенсвязывающий сайт является маскированным (то есть, активируемым), а дополнительный(е) антигенсвязывающий(ие) домен(ы) является (являются) немаскированным(и).
На фигуре 6, панелях A-J, схематически представлена серия диаграмм, где показан массив мультиспецифических активируемых антител, в которых все антигенсвязывающие домены являются маскированными.
На фигуре 7, панелях A-J, схематически представлена серия диаграмм, где показан массив мультиспецифических активируемых антител, в которых второй антигенсвязывающий домен является маскированным, а дополнительный(е) антигенсвязывающий(ие) домен(ы) является (являются) немаскированным(и).
На фигуре 8, панелях A-J, схематически представлена серия диаграмм, где показан массив мультиспецифических активируемых антител, в которых большинство, но не все антигенсвязывающие домены является маскированными, а по меньшей мере один дополнительный антигенсвязывающий домен является немаскированным.
На фигуре 9, панелях A-D, схематически представлена серия диаграмм, где показан массив мультиспецифических активируемых антител, в которых первый антигенсвязывающий домен и второй антигенсвязывающий домен являются маскированными, а остальные антигенсвязывающие домены являются немаскированными.
На фигуре 10 представлена фотография, иллюстрирующая анализ мультиспецифических антител и мультиспецифических активируемых антител с помощью электрофореза в ПААГ, который проводили для сравнения молекулярных масс гибридных тяжелых цепей scFv в образцах 1-8 и молекулярных масс соответствующих моноспецифических антител или активируемых антител в образцах 9-11.
На фигуре 11 представлен график, на котором проиллюстрирована способность мультиспецифических антител специфически связываться с их когнатными антигенами.
На фигурах 12A-12B представлена серия графиков, на которых проиллюстрирована способность мультиспецифических антител и мультиспецифических активируемых антител, включающих scFv против человеческого CTLA-4, специфически связываться (A) с мышиным и (B) с человеческим CTLA-4.
На фигуре 13 представлен график, на котором проиллюстрирована способность OKT3-содержащих мультиспецифических активируемых антител, называемых здесь мультиспецифическим активируемым анти-EGFR антителом 3954-1204-C225v5—OKT3 и мультиспецифическим активируемым анти-Jagged антителом 5342-1204-4D11v2—OKT3, специфически связываться с человеческим CD3ε в экспериментах на связывание, проводимых с помощью ELISA.
На фигуре 14 представлена серия фотографий, на которых проиллюстрирован анализ гидролизатов uPA, проводимый с помощью электрофореза в ПААГ, который указывал на активацию мультиспецифических активируемых антител без расщепления гибридных тяжелых цепей scFv.
На фигуре 15 представлена серия графиков, на которых проиллюстрирована способность активированных мультиспецифических активируемых антител связываться с различными мишенями.
На фигуре 16A представлен график, иллюстрирующий связывание мультиспецифических антител согласно изобретению с CD3ε-позитивными Т-клетками Jurkat.
На фигуре 16B представлен график, иллюстрирующий связывание мультиспецифических антител согласно изобретению с CD3ε-позитивными Т-клетками Jurkat и EGFR.
На фигуре 17 представлена серия графиков, иллюстрирующих EGFR-зависимую активацию Т-клеток Jurkat мультиспецифическими антителами согласно изобретению.
На фигуре 18A представлен график, иллюстрирующий EGFR-зависимую активацию первичных CD8+-Т-клеток мультиспецифическими антителами согласно изобретению.
На фигуре 18B представлен график, иллюстрирующий EGFR-зависимый цитолиз клеток-мишеней мультиспецифическими антителами согласно изобретению.
На фигуре 18C представлена серия графиков, иллюстрирующих CD69-активацию первичных Т-клеток мультиспецифическим антителом C225v5-OKT3m-H-N в EGFR-экспрессирующих клетках SW480 и минимальную активацию в EGFR-негативных клетках U266.
На фигуре 18D представлена серия графиков, иллюстрирующих EGFR-зависимый цитолиз клеток-мишеней мультиспецифическим антителом C225v5-OKT3m-H-N в EGFR-экспрессирующих клетках SW480 и почти полное отсутствие цитолиза в EGFR-негативных клетках U266.
На фигуре 19 представлена серия графиков, иллюстрирующих цитолиз первичных Т-клеток в панели EGFR-экспрессирующих клеточных линий под действием мультиспецифического антитела C225v5-OKT3m-H-N и отсутствие цитолиза EGFR-негативной клеточной линии U266.
На фигуре 20A и на фигуре 20B представлена серия графиков, иллюстрирующих снижение уровня связывания мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N с EGFR-экспрессирующими клетками по сравнению с уровнем связывания мультиспецифического антитела C225v5-OKT3m-H-N с EGFR. Кроме того, на фигурах 20A и 20B показано, что активация мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N под действием протеазы матриптазы (также обозначаемой здесь MTSP1 или MT-SP1) приводит к восстановлению уровня связывания с EGFR до уровня, эквивалентного уровню связывания, наблюдаемому для мультиспецифического антитела C225v5-OKT3m-H-N. На фигурах 20A и 20B также показано, что такое мультиспецифическое активируемое антитело и мультиспецифическое антитело связывается с EGFR по механизму, аналогичному механизму связывания соответствующего моноспецифического активируемого антитела и моноспецифического антитела.
На фигуре 21A и на фигуре 21B представлена серия графиков, иллюстрирующих связывание мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N и мультиспецифического антитела C225v5-OKT3m-H-N с CD3ε-позитивными Т-клетками Jurkat.
На фигуре 22 представлен график, иллюстрирующий снижение EGFR-зависимой активации Т-клеток Jurkat под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N по сравнению с активацией под действием мультиспецифического антитела C225v5-OKT3m-H-N. Кроме того, на этой фигуре показано, что активация мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N под действием протеазы матриптазы приводит к восстановлению уровня EGFR-зависимой активации до уровня, эквивалентного уровню активации, наблюдаемому для мультиспецифического антитела C225v5-OKT3m-H-N.
На фигуре 23A представлен график, иллюстрирующий снижение EGFR-зависимой активации первичных CD8+-Т-клеток под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N по сравнению с активацией под действием мультиспецифического антитела C225v5-OKT3m-H-N. Кроме того, на этой фигуре показано, что активация мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N под действием протеазы матриптазы приводит к восстановлению уровня EGFR-зависимой активации до уровня, эквивалентного уровню активации, наблюдаемому для мультиспецифического антитела C225v5-OKT3m-H-N.
На фигуре 23B представлен график, иллюстрирующий снижение EGFR-зависимого цитолиза клеток-мишеней под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N по сравнению с цитолизом под действием мультиспецифического антитела C225v5-OKT3m-H-N. Кроме того, на этой фигуре показано, что активация мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N под действием протеазы матриптазы приводит к восстановлению уровня EGFR-зависимого цитолиза клеток-мишеней до уровня, эквивалентного уровню цитолиза, наблюдаемому для мультиспецифического антитела C225v5-OKT3m-H-N.
Подробное описание изобретения
Настоящее изобретение относится к мультиспецифическим антителам и/или к мультиспецифическим активируемым антителам. Описанное здесь мультиспецифическое антитело представляют собой антитело, которое распознает два или более различных антигенов или эпитопов, а мультиспецифическое активируемое антитело представляет собой мультиспецифическое антитело, которое включает по меньшей мере одну маскирующую группу (ММ), связанную по меньшей мере с одним антиген- или эпитоп-связывающим доменом мультиспецифического антитела, и такое связывание с ММ приводит к снижению способности антиген- или эпитоп-связывающего домена связываться с мишенью. Описанные здесь активируемые мультиспецифические антитела являются стабильными в кровотоке, активируются в нужных участках, подвергаемых лечению и/или диагностике, но не в нормальной, то есть здоровой ткани, и в случае такой активации, эти антитела обладают способностью связываться с мишенью на уровне, по меньшей мере сравнимом с уровнем связывания соответствующего немодифицированного мультиспецифического антитела.
Неограничивающими примерами мультиспецифических антител являются биспецифические антитела, триспецифические антитела, тетраспецифические антитела и другие мультиспецифические антитела. Описанные здесь мультиспецифические антитела также являются поливалентными, где используемый здесь термин «поливалентность» означает общее число сайтов связывания на антителе, независимо от того, распознают ли эти сайты связывания одинаковые или различные антигены или эпитопы. Неограничивающими примерами мультиспецифических активируемых антител являются биспецифические активируемые антитела, триспецифические активируемые антитела, тетраспецифические активируемые антитела и другие мультиспецифические активируемые антитела. Описанные здесь мультиспецифические активируемые антитела также являются поливалентными.
В некоторых вариантах осуществления изобретения, мультиспецифические антитела или их фрагменты и/или мультиспецифические активируемые антитела или их фрагменты конструируют так, чтобы они связывались с Т-клетками и/или с другими иммунными эффекторными клетками. Мультиспецифические активируемые антитела или их фрагменты, связывающиеся с Т-клетками, также называются здесь мультиспецифическими антителами или их фрагментами, связывающимися с Т-клетками, и/или мультиспецифическими активируемыми антителами или их фрагментами, связывающимися с Т-клетками. Мультиспецифические активируемые антитела или их фрагменты, связывающиеся с иммунными эффекторными клетками, также называются здесь мультиспецифическими антителами или их фрагментами, связывающимися с иммунными эффекторными клетками, и/или мультиспецифическими активируемыми антителами или их фрагментами, связывающимися с иммунными эффекторными клетками. В некоторых вариантах осуществления изобретения, мультиспецифические антитела или их фрагменты и/или мультиспецифические активируемые антитела или их фрагменты конструируют так, чтобы они связывались или как-либо иначе взаимодействовали с более, чем одной мишенью и/или с более, чем одним эпитопом, и такие антитела также называются здесь антителами или их фрагментами, нацеленными на множество антигенов, и/или активируемыми антителами или их фрагментами, нацеленными на множество антигенов.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент включает домен IgG и домен scFv. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент включает вариабельный домен IgG и домен scFv. В некоторых вариантах осуществления изобретения, один домен мультиспецифического антитела или его фрагмента обладает специфичностью к антигену-мишени, а другой домен этого антитела обладает специфичностью к антигену Т-клеточной поверхности. В некоторых вариантах осуществления изобретения, один домен мультиспецифического антитела или его фрагмента обладает специфичностью к одному антигену-мишени, а другой домен этого антитела обладает специфичностью к другому антигену-мишени. В некоторых вариантах осуществления изобретения, один домен мультиспецифического антитела или его фрагмента обладает специфичностью к одному эпитопу антигена-мишени, а другой домен этого антитела обладает специфичностью к другому эпитопу того же самого антигена-мишени.
Различные варианты мультиспецифических активируемых антител или их фрагментов согласно изобретению представлены на фигурах 3A и 5-9. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент, содержащие IgG, имеют маскированные вариабельные домены IgG. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент, содержащие scFv, имеют маскированные домены scFv. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент имеют вариабельные домены IgG и домены scFv, где по меньшей мере один из доменов IgG связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент имеют вариабельные домены IgG и домены scFv, где по меньшей мере один из доменов scFv связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент имеют вариабельные домены IgG и домены scFv, где по меньшей мере один из вариабельных доменов IgG связан с маскирующей группой и по меньшей мере один из вариабельных доменов scFv связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент имеют вариабельные домены IgG и домены scFv, где каждый вариабельный домен IgG и домен scFv связан со своей собственной маскирующей группой. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела или его фрагмента обладает специфичностью к антигену-мишени, а другой домен этого антитела обладает специфичностью к антигену Т-клеточной поверхности. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела или его фрагмента обладает специфичностью к антигену-мишени, а другой домен этого антитела обладает специфичностью к другому антигену-мишени. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела или его фрагмента обладает специфичностью к эпитопу антигена-мишени, а другой домен этого антитела обладает специфичностью к другому эпитопу того же самого антигена-мишени.
В некоторых вариантах осуществления изобретения, мультиспецифическими антителами или их фрагментами согласно изобретению являются по меньшей мере (i) антитело или его фрагмент, которые связываются с T-клетками и включают первое антитело или его антигенсвязывающий фрагмент (AB1), связывающиеся с первой мишенью, которая связывается с Т-клетками и представляет собой поверхностный антиген на Т-клетках, и (ii) второе антитело или его фрагмент, которые включают второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело включает антитела или их фрагменты, связывающиеся с Т-клетками и присоединенные к комбинации одного или более N-концов и/или одного или более С-концов мультиспецифического антитела. Настоящее изобретение также относится к мультиспецифическим антителам, содержащим антитело или его фрагмент, связывающиеся с другими иммуными эффекторными клетками, например, с поверхностным антигеном природной клетки-киллера (NK), мононуклеарной клетки, такой как миелоидная мононуклеарная клетка, макрофага и/или других иммунных эффекторных клеток.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают по меньшей мере два антитела или их фрагмента, связывающиеся с Т-клетками, где указанные антитела или их фрагменты связываются с одним или более рецепторами, активирующими Т-клетки, где указанными фрагментами являются scFv-фрагменты, связывающиеся с Т-клетками, включая, но неограничиваясь ими, OX40/GITR, CD137/GITR, CD137/OX40, CD27/NKG2D, и с дополнительной комбинацией активирующих рецепторов; и одно связывающееся с мишенью антитело, где указанное антитело представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают два антитела или их фрагмента, связывающихся с Т-клетками, а именно, с одним или более Т-клеточными ингибирующими рецепторами, и одно связывающееся с мишенью антитело, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с одной и той же мишенью, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными мишенями, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными эпитопами на одной и той же мишени, связывающейся с Т-клетками.
В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) первую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с Т-клетками и включающего первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клетками, и (ii) вторую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с мишенью и включающего второе антитело или его антигенсвязывающий фрагмент (AB2), которые связываются со второй мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент включают (iii) по меньшей мере третий антигенсвязывающий фрагмент антитела, связывающегося с мишенью и включающего третье антитело или его антигенсвязывающий фрагмент (AB3), которые связываются с третьей мишенью. В некоторых вариантах осуществления изобретения, вторая, третья или более мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, вторая, третья или более мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, вторая, третья или более мишени представляют собой различные эпитопы на одном и том же антигене.
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела или их фрагменты согласно изобретению включают по меньшей мере (i) антитело или его фрагмент, которые связываются с T-клетками и включают первое антитело или его антигенсвязывающий фрагмент (AB1), связывающиеся с первой мишенью, которая связывается с Т-клетками и представляет собой поверхностный антиген на Т-клетках, где АВ1 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ1 связываться с первой мишенью; и (ii) второе антитело или его фрагмент, которые включают второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает антитела или их фрагменты, связывающиеся с Т-клетками и присоединенные к комбинации одного или более N-концов и/или одного или более С-концов мультиспецифического активируемого антитела. Настоящее изобретение также относится к мультиспецифическим активируемым антителам, содержащим другое антитело или его фрагмент, связывающиеся с другими иммуными эффекторными клетками, например, с поверхностным антигеном природной клетки-киллера (NK), мононуклеарной клетки, такой как миелоидная мононуклеарная клетка, макрофага и/или других иммунных эффекторных клеток.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) два антитела или их фрагмента, связывающиеся с Т-клетками, где указанные антитела или их фрагменты связываются с одним или более рецепторами, активирующими Т-клетки, где указанными фрагментами являются scFv-фрагменты, связывающиеся с Т-клетками, включая, но не ограничиваясь ими, OX40/GITR, CD137/GITR, CD137/OX40, CD27/NKG2D, и с дополнительной комбинацией активирующих рецепторов; где АВ1 одного из фрагментов антитела, связывающегося с Т-клетками, связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ1 связываться с соответствующей мишенью, которая связывается с Т-клетками; и (ii) одно связывающееся с мишенью антитело, где указанное антитело представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают два антитела или их фрагмента, связывающихся с Т-клетками, а именно, с одним или более Т-клеточными ингибирующими рецепторами, и одно связывающеся с мишенью антитело, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с одной и той же мишенью, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными мишенями, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными эпитопами на одной и той же мишени, связывающейся с Т-клетками.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) два антитела или их фрагмента, связывающиеся с Т-клетками, где указанные антитела или их фрагменты связываются с одним или более рецепторами, активирующими Т-клетки, где указанными фрагментами являются scFv-фрагменты, связывающиеся с Т-клетками, включая, но неограничиваясь ими, OX40/GITR, CD137/GITR, CD137/OX40, CD27/NKG2D, и с дополнительной комбинацией активирующих рецепторов; где каждая АВ1 фрагментов антитела, связывающегося с Т-клетками, связывается с их собственной маскирующей группой (ММ), и такое связывание с каждой ММ приводит к снижению способности соответствующего АВ1 связываться с соответствующей мишенью, которая связывается с Т-клетками; и (ii) одно связывающееся с мишенью антитело, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают два антитела или их фрагмента, связывающиеся с Т-клетками, а именно, с одним или более Т-клеточными ингибирующими рецепторами, и одно связывающееся с мишенью антитело, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с одной и той же мишенью, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными мишенями, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными эпитопами на одной и той же мишени, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) первую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с Т-клетками и включающего первое антитело или антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клетками, где АВ1 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ1 связываться с первой мишенью; и (ii) вторую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с мишенью и включающего второе антитело или его антигенсвязывающий фрагмент (AB2), которые связываются со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела или их фрагменты согласно изобретению включают по меньшей мере (i) антитело или его фрагмент, которые связываются с T-клетками и включают первое антитело или его антигенсвязывающий фрагмент (AB1), связывающиеся с первой мишенью, которая связывается с Т-клетками и представляет собой поверхностный антиген на Т-клетках, и (ii) второе антитело или его фрагмент, которые включают второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где АВ2 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает антитела или их фрагменты, связывающиеся с Т-клетками и присоединенные к комбинации одного или более N-концов и/или одного или более С-концов мультиспецифического активируемого антитела. Настоящее изобретение также относится к мультиспецифическим активируемым антителам, содержащим другое антитело или его фрагмент, связывающиеся с другими иммуными эффекторными клетками, например, с поверхностным антигеном природной клетки-киллера (NK), мононуклеарной клетки, такой как миелоидная мононуклеарная клетка, макрофага и/или других иммунных эффекторных клеток.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) два антитела или их фрагмента, связывающиеся с Т-клетками, где указанные антитела или их фрагменты связываются с одним или более рецепторами, активирующими Т-клетки, где указанными фрагментами являются scFv-фрагменты, связывающиеся с Т-клетками, включая, но неограничиваясь ими, OX40/GITR, CD137/GITR, CD137/OX40, CD27/NKG2D, и с дополнительной комбинацией активирующих рецепторов; и (ii) одно связывающееся с мишенью антитело, которое представляет собой второе антитело или его антигенсвязывающий фрагмент (АВ2), которые связываются со второй мишенью, где АВ2 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ2 связываться со второй мишенью, где scFv-фрагменты, связывающиеся с Т-клетками, присоединены к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают два антитела или их фрагмента, связывающихся с Т-клетками, а именно, с одним или более Т-клеточными ингибирующими рецепторами, и одно антитело, связывающееся с мишенью, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с одной и той же мишенью, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными мишенями, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными эпитопами на одной и той же мишени, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) первую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с Т-клетками, и (ii) вторую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с мишенью и включающего второе антитело или его антигенсвязывающий фрагмент (AB2), которые связываются со второй мишенью, где АВ2 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела или их фрагменты согласно изобретению включают по меньшей мере (i) антитело или его фрагмент, которые связываются с T-клетками и включают первое антитело или его антигенсвязывающий фрагмент (AB1), связывающиеся с первой мишенью, которая связывается с Т-клетками и представляет собой поверхностный антиген на Т-клетках, где АВ1 связывается с маскирующей группой (ММ1), и такое связывание с ММ1 приводит к снижению способности АВ1 связываться с первой мишенью, и (ii) второе антитело или его фрагмент, которые включают второе антитело или его антигенсвязывающий фрагмент (AB2), связывающиеся со второй мишенью, где АВ2 связывается с маскирующей группой (ММ2), и такое связывание с ММ2 приводит к снижению способности АВ2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу тяжелой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к N-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, антитело или его фрагмент, связывающиеся с Т-клетками, присоединены к С-концу легкой цепи мишень-специфического антитела. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело включает антитела или их фрагменты, связывающиеся с Т-клетками и присоединенные к комбинации одного или более N-концов и/или одного или более С-концов мультиспецифического активируемого антитела. Настоящее изобретение также относится к мультиспецифическим активируемым антителам, содержащим другое антитело или его фрагмент, связывающиеся с другими иммуными эффекторными клетками, например, с поверхностным антигеном природной клетки-киллера (NK), мононуклеарной клетки, такой как миелоидная мононуклеарная клетка, макрофага и/или других иммунных эффекторных клеток.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) два антитела или их фрагмента, связывающиеся с Т-клетками, где указанные антитела или их фрагменты связываются с одним или более рецепторами, активирующими Т-клетки, где указанными фрагментами являются scFv-фрагменты, связывающиеся с Т-клетками, включая, но неограничиваясь ими, OX40/GITR, CD137/GITR, CD137/OX40, CD27/NKG2D, и с дополнительной комбинацией активирующих рецепторов, где АВ1 одного из фрагментов антитела, связывающегося с Т-клетками, связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ1 связываться с его соответствующей мишенью, связывающейся с Т-клетками, и (ii) одно связывающееся с мишенью антитело, которое представляет собой второе антитело или его антигенсвязывающий фрагмент (АВ2), которые связываются со второй мишенью, где АВ2 связывается с маскирующей группой (ММ2), и такое связывание с ММ2 приводит к снижению способности АВ2 связываться со второй мишенью, где scFv-фрагменты, связывающиеся с Т-клетками, присоединены к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают два антитела или их фрагмента, связывающиеся с Т-клетками, а именно, с одним или более Т-клеточными ингибирующими рецепторами, и одно антитело, связывающееся с мишенью, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с одной и той же мишенью, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными мишенями, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными эпитопами на одной и той же мишени, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включают по меньшей мере (i) два антитела или их фрагмента, связывающиеся с Т-клетками, где указанные антитела или их фрагменты связываются с одним или более рецепторами, активирующими Т-клетки, где указанными фрагментами являются scFv-фрагменты, связывающиеся с Т-клетками, включая, но не ограничиваясь ими, OX40/GITR, CD137/GITR, CD137/OX40, CD27/NKG2D, и с дополнительной комбинацией активирующих рецепторов, где каждое АВ1 фрагментов антитела, связывающегося с Т-клетками, связывается со своей собственной маскирующей группой (ММ1), и такое связывание каждой ММ1 с соответствующим АВ1 приводит к снижению способности АВ1 связываться с его соответствующей мишенью, связывающейся с Т-клетками, и (ii) одно связывающееся с мишенью антитело, которое представляет собой второе антитело или его антигенсвязывающий фрагмент (АВ2), которые связываются со второй мишенью, где АВ2 связывается с маскирующей группой (ММ2), и такое связывание с ММ2 приводит к снижению способности АВ2 связываться со второй мишенью, где scFv-фрагменты, связывающиеся с Т-клетками, присоединены к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, мультиспецифическое антитело или его фрагмент согласно изобретению включают два антитела или их фрагмента, связывающиеся с Т-клетками, а именно, с одним или более Т-клеточными ингибирующими рецепторами, и одно антитело, связывающееся с мишенью, которое представляет собой scFv-фрагменты, связывающиеся с Т-клетками и присоединенные к обеим ветвям антитела, связывающегося с мишенью. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с одной и той же мишенью, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными мишенями, связывающимися с Т-клетками. В некоторых вариантах осуществления изобретения, два фрагмента антитела, связывающихся с Т-клетками, связываются с различными эпитопами на одной и той же мишени, связывающейся с Т-клетками. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело или его фрагмент согласно изобретению включает по меньшей мере (i) первую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с Т-клетками и включающего первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клетками, где АВ1 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ1 связываться с первой мишенью; и (ii) вторую ветвь, содержащую антигенсвязывающий фрагмент антитела, связывающегося с мишенью и включающего второе антитело или антигенсвязывающий фрагмент (AB2), которые связываются со второй мишенью, где АВ2 связывается с маскирующей группой (ММ), и такое связывание с ММ приводит к снижению способности АВ2 связываться со второй мишенью. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой один и тот же антиген. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные антигены. В некоторых вариантах осуществления изобретения, первая и вторая мишени представляют собой различные эпитопы на одном и том же антигене. В некоторых вариантах осуществления изобретения, антитела или их фрагменты, связывающиеся с Т-клетками, связываются не с одним или более Т-клеточными активирующими рецепторами, а с одним или более Т-клеточными ингибирующими рецепторами.
В некоторых вариантах осуществления изобретения, антигеном-мишенью является антиген, который в высокой степени экспрессируется в нормальной, здоровой и пораженной ткани. В некоторых вариантах осуществления изобретения, антигеном-мишенью является антиген, представленный в таблице 1, где указанный антиген в высокой степени экспрессируется в нормальной, здоровой и пораженной ткани.
В некоторых вариантах осуществления изобретения, антигеном-мишенью является антиген, который в высокой степени экспрессируется в пораженной ткани, но не в нормальной здоровой ткани. В некоторых вариантах осуществления изобретения, антигеном-мишенью является антиген, представленный в таблице 1, где указанный антиген в высокой степени экспрессируется в пораженной ткани, но не в нормальной здоровой ткани. При этом, указанный антиген-мишень может экспрессироваться в нормальной здоровой ткани, но не может сверхэкспрессироваться или экспрессироваться в высокой степени в этой ткани.
В некоторых вариантах осуществления изобретения, максирующая группа (MM) связана с антителом или с его антигенсвязывающим фрагментом (AB) посредством отщепляемой группы (CM), которая служит в качестве субстрата для протеазы. Подходящие протеазы, которые могут быть использованы в мультиспецифических активируемых антителах согласно изобретению, могут быть определены по экспрессии этих протеаз в предполагаемом участке поражения и/или в диагностируемом участке. В некоторых вариантах осуществления изобретения, протеазой являются активатор плазминогена u-типа (uPA, также называемый урокиназой), легумаин и/или матриптаза (также обозначаемая MT-SP1 или MTSP1). В некоторых вариантах осуществления изобретения, протеазой является металлопротеаза матрикса (MMP).
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела конструируют так, чтобы они включали маскурующую группу (MM), связанную с антителом или его антигенсвязывающим фрагментом (AB) посредством нерасщепляемого линкера. Так, например, в некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом является мультиспецифическое активируемое антитело, связывающееся с Т-клетками и включающее нацеливающее антитело или его антигенсвязывающий фрагмент, и антитело, связывающееся с Т-клетками или его антигенсвязывающую часть, где антитело, связывающееся с Т-клетками, или его антигенсвязывающий фрагмент представляют собой первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клетками, где AB1 связывается с маскирующей группой (MM1) посредством нерасщепляемого линкера, и такое связывание с ММ приводит к снижению способности AB1 связываются с первой мишенью; и где указанное нацеливающее антитело или его антигенсвязывающий фрагмент являются немаскированными.
В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом является мультиспецифическое активируемое антитело, связывающееся с Т-клетками, где указанное антитело включает нацеливающее антитело или его антигенсвязывающий фрагмент и антитело, связывающееся с Т-клетками, или его антигенсвязывающую часть, где антитело, связывающееся с Т-клетками, или его антигенсвязывающий фрагмент включают первое антитело или его антигенсвязывающий фрагмент (AB1), которые связываются с первой мишенью, связывающейся с Т-клетками, где AB1 связывается с маскирующей группой (MM1) посредством нерасщепляемого линкера, и такое связывание с MM приводит к снижению способности AB1 связываться с первой мишенью, а нацеливающее антитело или его антигенсвязывающий фрагмент включают второе антитело или его антигенсвязывающий фрагмент (AB2), которые связываются со второй мишенью, где AB2 связывается с маскирующей группой (MM1) посредством нерасщепляемого линкера, и такое связывание с MM приводит к снижению способности AB2 связываться со второй мишенью.
Общая концепция биспецифических антител была впервые разработана по меньшей мере 50 лет назад (работа Nisonoff, A. and Mandy, W.J., Nature 194, 355-359 (1962), цитируемая Chan, A.C. and Carter, P.J., Nature Reviews Immunol. 10, 301-316 (2010)).
В литературе был описан ряд основополагающих идей создания биспецифических антител (см., например, фигуру 1 настоящей заявки; Liu, M.A., et al., Proc. Natl. Acad. Sci. USA 82, 8648-8652 (1985); обзорную статью Kroesen, B.J., et al. Adv. Drug Delivery Rev. 31, 105-129 (1998); Marvin, J.S. and Zhu, Z., Acta Pharm. Sinica 26, 649-658 (2005); Chan and Carter, Nat. Rev. Immunol. 2010; Fitzgerald and Lugovsky, MAbs. 3(3):299-309 (2011); Riethmuller, G., Cancer Immunity 12, 12-18 (2012)). Общая концепция конструирования биспецифических антител заключается в связывании белок-связывающих доменов, обычно на основе множества доменов иммуноглобулина, используемых для конструирования молекул, способных связываться с двумя или более антигенами-мишенями и обладающих нужными функиями, такими как IgG-подобное физиологическое распределение, фармакологические свойства и эффекторные функции. Эффекторные функции могут включать антителозависимую цитотоксичность (ADCC), комплемент-зависимую цитотоксичность (CDC), рекрутинг T-клеток (BiTEs™) (см. например, Baeuerle, P.A. and Reinhardt, C., Cancer Res. 69, 4941-4944 (2009) и TandAbs™ (см. Cochlovius et al., Cancer Res. 60, 4336-4341 (2000)) и/или доставку цитотоксической полезной нагрузки в форме химически конъюгированной молекулы, такой как ингибитор микротрубочек, ДНК-алкилирующий агент или другой токсин в форме конъюгата «антитело-лекарственное средство» (ADC). Было проведено исследование биспецифических антител во множестве форматов, причем, стандартные гетеродимерные IgG (BiTEs™), несмотря на их продуцирование на субоптимальном уровне и на их субоптимальные фармакологические свойства, являются особенно перспективными с точки зрения их применения в медицине (см., Chan and Carter, Nat. Rev. Immunol. 2010). Биспецифические антитела могут быть использованы в различных целях, исходя из специфических признаков конкретных форматов и выбранных антигенов-мишеней, причем, не все форматы биспецифических антител могут быть применены в тех или иных целях. Так, например, Fab-формат «два в одном» (Bostrom, J., et al., Science 323, 1610-1614 (2009)) состоит из одного связывающего домена, связывающегося с антигенами A и B по тому или иному механизму, и таким образом, биспецифическое антитело этого типа может связываться с двумя копиями антигена А, с двумя копиями антигена В или с одной копией каждого антигена. С другой стороны, DVD-формат (Wu, C., et al., Nature Biotechnol. 25, 1290-1297 (2007)) обеспечивает связывание с двумя копиями антигена А, а также с двумя копиями антигена B. Пространственная и относительная ориентация каждого связывающего домена может также играть важную роль в связывании с множеством антигенов по механизму, сообщающему нужную активность, например, посредством перекрестно-связывающихся рецепторов (Jackman, J., et al., J. Biol. Chem. 285, 20850-20859 (2010)), а там, где необходимо связывание одной копии каждого рецептора, может потребоваться моновалентное взаимодействие с каждым из двух отдельных антигенов.
Механистически отличающаяся область применения биспецифического антитела включает рекрутинг иммунных эффекторных клеток (таких как цитотоксические Т-лимфоциты, NK-клетки и/или миелоидные эффекторные клетки) для атаки на опухолевые клетки, осуществляемый путем конструирования биспецифического антитела, связывающегося с антигеном, присутствующим на поверхности цитотоксических клеток-мишеней (например, опухолевых клеток), и антигеном, присутствующим на поверхности цитотоксических иммунных клеток (например, Т-клеток). Катумаксомаб представляет собой репрезентативное клинически апробированне биспецифическое антитело, нацеленное на опухолевый антиген EpCAM, присутствующий на раковых клетках, и стимулирующее рекрутинг цитотоксических Т-клеток посредством связывания с CD3 на поверхности Т-клеток (Linke, R., Klein, A., and Seimetz, D., mAbs 2, 129-136 (2010)); однако, применение такого препарата имеет некоторые ограничения и требует внутрибрюшинного введения. Антитела BiTE™ представляют собой биспецифические антитела, которые позволяют осуществлять рекрутинг Т-клеток в другом формате и обладают другими фармакологическими свойствами (Baeuerle, P.A. and Reinhardt, C., Анти-Cancer Res. 69, 4941-4944 (2009)), а в частности, антитела BiTE состоят из двух одноцепочечных вариабельных фрагментов (scFvs) и имеют очень короткое фармакологическое время полужизни in vivo, а поэтому они должны быть введены путем непрерывного вливания. Антитело аналогичного формата, называемое TandAb™, представляет собой четырехвалентную конструкцию биспецифического диантитела, в которой два вариабельных домена связаны с антигеном-мишенью, и два вариабельных домена связаны с СD3 на поверхности Т-клеток (Cochlovius et al., Cancer Res. 60, 4336-4341 (2000)). Было подтверждено, что связывание двухвалентной конструкции такого формата с антигеном-мишенью и антигеном Т-клеточной поверхности может быть применено для сообщения способности связываться с Т-клетками и для сообщения антиген-направленной цитотоксичности. Однако антитело формата TandAb также имеет очень короткое время полужизни in vivo.
Общее ограничение применения биспецифических антител, связывающихся с Т-клетками, заключается в том, что цитотоксичность Т-клеток может быть очень сильной и может активироваться относительно низкими уровнями антигена-мишени, присутствующего на поверхности клеток. Поэтому даже умеренные уровни экспрессии антигена-мишени, такого как EGFR, на здоровых тканях могут стимулировать достаточно высокий уровень токсичности, что ограничивает или даже не допускает клинического применения такого терапевтического антитела (Lutterbuese, R. et al., Proc. Natl. Acad. Sci. USA 107, 12605-12610 (2010)). Таким образом, необходимость в получении эффективных терапевтических средств на основе антител, которые были бы более безопасными, например, обладали бы пониженной токсичностью, остается актуальной.
Мультиспецифические активируемые антитела согласно изобретению являются более безопасными, чем мультиспецифические (например, биспецифические) антитела, поскольку активность мультиспецифического активируемого антитела ограничена окружением локального участка поражения. В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом является мультиспецифическое активируемое антитело, связывающееся с иммунными эффекторными клетками. В некоторых вариантах осуществления изобретения, мультиспецифическим активируемым антителом является мультиспецифическое активируемое антитело, связывающееся с Т-клетками. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело распознает две или более мишеней. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет формат IgG-scFv, сообщающий антителу IgG продолжительное время полужизни. В некоторых вариантах осуществления изобретения, такое мультиспецифическое активируемое антитело дополнительно модулируют путем введения Fc-мутаций в сайт связывания с FcRn (Petkova, S. B. et al., Intl. Immunol. 18, 1759-1769 (2006)); Deng, R. et al., mAbs 4, 101-109 (2012)); Olafson, T Methods Mol. Biol. 907, 537-556 (2012)). В некоторых вариантах осуществления изобретения, такое мультиспецифическое активируемое антитело включает мутации, введенные в Fc-домен, такие как мутация N297A (Lund, J. et al., Mol. Immunol. 29, 53-39 (1992)), которая приводит к снижению эффекторных функций IgG (ADCC and CDC) и, тем самым, к снижению нежелательных токсических эффектов.
Мультиспецифические активируемые антитела стимулируют активацию протеазной активности, которая является широко известным признаком развития множества патологических состояний, и такие антитела позволяют специфически доставлять терапевтические средства в пораженную ткань. Это активируемое антитело было создано на основе антитела IgG или его фрагмента, такого как scFv-область, Fab-область и один VH- или VL-домен, которые были модифицированы так, чтобы они включали маскирующую группу (MM), связанную с антителом посредством отщепляемой группы (CM), такой как группа, отщепляемая протеазой (см., например, Международную заявку PCT WO 2009/025846, опубликованную 26 февраля 2009; Международную заявку PCT WO 2010/081173, опубликованную 15 июля 2010). Альтернативно, для сообщения одной или более специфичностей связывания может быть использован домен белка, не являющегося антителом (см., например, Международную заявку PCT WO 2010/096838, опубликованную 26 августа 2010; Boersma, Y.L. et al., J. Biol. Chem. 286, 41273-41285 (2011)). Мультиспецифическое антитело, подобно стандартному IgG, может быть использовано в качестве исходного материала для продуцирования мультиспецифического активируемого антитела. Такое мультиспецифическое активируемое антитело обеспечивает высокоаффинное нацеливание на все антигены, распознаваемые родительским мультиспецифическим антителом, которе обладает тканеспецифичностью, определяемой селективно активированным расщепляемым линкером. MM и CM, в зависимости от формата используемого мультиспецифического антитела, могут присутствовать у N- или С-конца соответствующего домена.
Мультиспецифическое активируемое антитело включает по меньшей мере один домен, происходящий от IgG и обладающий специфичностью к первому антигену, а также синтетически или биосинтетически присоединенный к другому домену (который может происходить от IgG или другого белка), обладающему специфичностью связывания со вторым антигеном. Дополнительная специфичность может быть сообщена посредством связывания такого мультиспецифического активируемого антитела с одним или более дополнительными доменами, ответственными за дополнительную специфичность связывания с первым антигеном, со вторым антигеном или с дополнительными антигенами. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет один или более из указанных доменов, маскированных соответствующей маскирующей группой (MM). Каждое из антител такого мультиспецифического формата, описанных на фигуре 1, может быть преобразовано в мультиспецифическое активируемое антитело посредством маскировки одного или более антигенсвязывающих молекул одного или более доменов иммуноглобулина. Примеры подходящих форматов мультиспецифических активируемых антител представлены на фигурах 3A и 5-9.
Использование scFv-домена, сообщающего специфичность связывания с конкретным антигеном, позволяет осуществлять модульное конструирование мультиспецифических антител и мультиспецифических активируемых антител. Использование scFv-домена(ов), присоединенного(ых) к концу тяжелой или легкой цепи IgG в целях конструирования биспецифических антител, описано в литературе (см., например, Orcutt, K.D. et al., Prot. Eng. Design Select. 23, 221-228 (2010)); Dong et al., (2011)). Этот формат («IgG-scFv») позволяет превращать стандартный IgG в биспецифическое антитело, где первая специфичность кодируется в вариабельных доменах IgG, а вторая специфичность кодируется в доменах scFv, связанных посредством гибкой линкерной области. Варианты такого формата включают присоединение scFv-доменов у N- или C-концов тяжелой или легкой цепи, где scFv могут обладать одной и той же или различными специфичностями связывания с антигеном (Spangler, J. B. et al., J. Mol. Biol. 422, 532-544 (2012)). Кроме того, scFv с различными специфичностями могут быть присоединены к N- или C-концу каждой тяжелой цепи посредством гетеролимеров тяжелой цепи (например, имеющих конструкцию типа «узлы в дырках» или аналогичные конструкции).
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет маскированные вариабельные домены IgG. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет маскированные вариабельные домены scFv. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет маскированные вариабельные домены IgG и маскированные домены scFv. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к антигену-мишени, а другой домен этого антитела обладает специфичностью к антигену Т-клеточной поверхности. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к одному антигену-мишени, а другой домен этого антитела обладает специфичностью к другому антигену-мишени. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к одному эпитопу антигена-мишени, а другой домен этого антитела обладает специфичностью к другому эпитопу антигена-мишени.
В настояшем описании представлены примеры структур мультиспецифических активируемых антител, которыми являются, но не ограничиваются ими, следующие структуры: (VL-CL)2:(VH-CH1-CH2-CH3-L4-VH*-L3-VL*-L2-CM-L1-MM)2; (VL-CL)2:(VH-CH1-CH2-CH3-L4-VL*-L3-VH*-L2-CM-L1-MM)2; (MM-L1-CM-L2-VL-CL)2:(VH-CH1-CH2-CH3-L4-VH*-L3-VL*)2; (MM-L1-CM-L2-VL-CL)2:(VH-CH1-CH2-CH3-L4-VL*-L3-VH*)2; (VL-CL)2:(MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL)2:(MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL-CL)2:(VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL-CL)2:(VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL*-L3-VH*-L4-VL-CL)2:(VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VH*-L3-VL*-L4-VL-CL)2:(VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*)2: (MM-L1-CM-L2-VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*)2: (MM-L1-CM-L2-VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2: (VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (VL*-L3-VH*-L4-VH-CH1-CH2-CH3)2; или (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2: (VH*-L3-VL*-L4-VH-CH1-CH2-CH3)2,
где: VL и VH представляют собой вариабельные домены легкой и тяжелой цепей первой специфичности, содержащиеся в IgG; VL* и VH* представляют собой вариабельные домены второй специфичности, содержащиеся в scFv; L1 представляет собой линкерный пептид, соединяющий маскирующую группу (MM) и отщепляемую группу (CM); L2 представляет собой линкерный пептид, соединяющий отщепляемую группу (CM) и антитело; L3 представляет собой линкерный пептид, соединяющий вариабельные домены scFv; L4 представляет собой линкерный пептид, соединяющий антитело первой специфичности и антитело второй специфичности; CL представляет собой константный домен легкой цепи, а CH1, CH2, CH3 представляют собой константные домены тяжелой цепи. Первая и вторая специфичности могут относиться к любому антигену или эпитопу. Дополнительными структурами являются, но не ограничиваются ими, следующие структуры: (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL-CL-L4-VH*-L3-VL*)2:(VH-CH1-CH2-CH3)2; (MM-L1-CM-L2-VL-CL--L4-VL*-L3-VH*)2:(VH-CH1-CH2-CH3)2; (VL-CL-L4-VH*-L3-VL*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3-L4-VH*-L3-VL*-L2-CM-L1-MM)2; (VL-CL-L4-VL*-L3-VH*-L2-CM-L1-MM)2:(VH-CH1-CH2-CH3-L4-VL*-L3-VH*-L2-CM-L1-MM)2; (MM-L1-CM-L2-VL-CL-L4-VH*-L3-VL*)2:(VH-CH1-CH2-CH3)2; или (MM-L1-CM-L2-VL-CL--L4-VL*-L3-VH*)2:(VH-CH1-CH2-CH3)2.
В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело, включающее IgG, имеет маскированные вариабельные домены IgG. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело, включающее scFv, имеет маскированные домены scFv. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где по меньшей мере один из вариабельных доменов IgG связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где по меньшей мере один из доменов scFv связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где по меньшей мере один из вариабельных доменов IgG связан с маскирующей группой, и по меньшей мере один из доменов scFv связан с маскирующей группой. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет вариабельные домены IgG и домены scFv, где каждый вариабельный домен IgG и домен scFv связан с их собственной маскирующей группой. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к антигену мишени, а другой домен этого антитела обладает специфичностью к антигену Т-клеточной поверхности. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к одному антигену мишени, а другой домен этого антитела обладает специфичностью к другому антигену мишени. В некоторых вариантах осуществления изобретения, один домен мультиспецифического активируемого антитела обладает специфичностью к одному эпитопу антигена мишени, а другой домен этого антитела обладает специфичностью к другому эпитопу антигена мишени.
В некоторых вариантах изобретения, относящихся к мультиспецифическому активируемому антителу, связывающемуся с иммунной эффекторной клеткой, такому как мультиспецифическое активируемое антитело, связывающееся с T-клеткой, антигеном обычно является антиген, присутствующий на поверхности опухолевой клетки или клетки другого типа, ассоциированной с заболеванием, и такими антигенами являются, но не ограничиваются ими, любые мишени, перечисленные в таблице 1, такие как, но не ограничивающиеся ими, EGFR, erbB2, EpCAM, Jagged, PD-L1, B7H3 или CD71 (рецептор трансферрина), а другим антигеном обычно являются стимулирующий или ингибирующий антиген, присутствующий на поверхности Т-клетки, природной клетки-киллера (NK), миелоидной мононуклеарной клетки, макрофага и/или других иммунных эффекторных клеток, где таким антигеном являются, но не ограничиваются ими, B7-H4, BTLA, CD3, CD4, CD8, CD16a, CD25, CD27, CD28, CD32, CD56, CD137, CTLA-4, GITR, HVEM, ICOS, LAG3, NKG2D, OX40, PD-1, TIGIT, TIM3 или VISTA. Домен антитела, сообщающий специфичность к антигену Т-клеточной поверхности, может быть также заменен лигандом или доменом лиганда, который связывается с рецептором Т-клеток, рецептором NK-клеток, рецептором макрофагов и/или рецептором других иммунных эффекторных клеток, и такими лигандами являются, но не ограничиваются ими, B7-1, B7-2, B7H3, PD-L1, PD-L2 или TNFSF9. В некоторых вариантах осуществления изобретения, относящихся к активируемому антителу, нацеленному на множество антигенов, один антиген выбран из группы мишеней, перечисленных в таблице 1, и другой антиген выбран из группы мишеней, перечисленных в таблице 1.
В некоторых вариантах осуществления изобретения, одноцепочечный вариабельный домен, специфически связывающийся с мишенью Т-клеточной поверхности, присоединен к карбоксильному концу полностью человеческого IgG1 (нацеливающего антитела), которое связывается с антигеном клеточной поверхности. scFv может быть присоединен к карбоксильному концу тяжелой цепи, к карбоксильному концу легкой цепи или к обеим цепям (фигура 2). В некоторых вариантах осуществления изобретения, одноцепочечный вариабельный домен, специфически связывающийся с мишенью Т-клеточной поверхности, присоединен к амино-концу полностью человеческого IgG1 (нацеливающего антитела), которое связывается с антигеном клеточной поверхности. scFv может быть присоединен к амино-концу тяжелой цепи, к амино-концу легкой цепи или к обеим цепям. Такие гибриды получают в виде одной генетической конструкции и экспрессируют в клеточной культуре. Нацеливающее антитело может специфически связываться с одним или более антигенами опухолевой поверхности или с любыми клетками, на удаление которых направлено это антитело. scFv может обладать специфичностью к одним и тем же или к различным антигенам.
В некоторых вариантах осуществления изобретения, нацеливающим антителом является анти-EGFR антитело. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225v5, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225v4, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является C225v6, которое специфически связывается с EGFR. В некоторых вариантах осуществления изобретения, нацеливающим антителом является анти-Jagged антитело. В некоторых вариантах осуществления изобретения, нацеливающим антителом является 4D11, которое специфически связывается с человеческим и мышиным Jagged 1 и Jagged 2. В некоторых вариантах осуществления изобретения, нацеливающим антителом является 4D11v2, которое специфически связывается с человеческим и мышиным Jagged 1 и Jagged 2.
В некоторых вариантах осуществления изобретения, нацеливающее антитело может присутствовать в форме активируемого антитела. В некоторых вариантах осуществления изобретения, scFv может (могут) присутствовать в форме Pro-scFv (см., например, WO 2009/025846, WO 2010/081173).
В некоторых вариантах осуществления изобретения, scFv специфически связывается с CD3ε, например, с OKT3. В некоторых вариантах осуществления изобретения, scFv специфически связывается с CTLA-4 (также называемым здесь CTLA и CTLA4).
В некоторых вариантах осуществления изобретения, scFv специфически связывается с одной или более Т-клетками, одной или более NK-клетками и/или с одним или более макрофагами. В некоторых вариантах осуществления изобретения, scFv специфически связывается с мишенью, выбранной из группы, состоящей из B7-H4, BTLA, CD3, CD4, CD8, CD16a, CD25, CD27, CD28, CD32, CD56, CD137, CTLA-4, GITR, HVEM, ICOS, LAG3, NKG2D, OX40, PD-1, TIGIT, TIM3, или VISTA и их комбинаций.
Определения
Если это не оговорено особо, то все технические и научные термины, используемые в настоящей заявке, имеют значения, в основном, понятные среднему специалисту в области, к которой относится настоящее изобретение. Кроме того, если это не оговорено особо, термины, употребляемые в единственном числе, могут относиться и к существительным во множественном числе, а термины, употребляемые во множественном числе, могут относиться к существительным в единственном числе. Вообще говоря, номенклатура, используемая в приведенных здесь описаниях методов культивирования клеток и тканей, методов молекулярной биологии и химии белков и олиго- или полинуклеотидов, а также методов гибридизации, является хорошо известной и широко применяется специалистами. Для конструирования рекомбинантных ДНК, синтеза олигонуклеотидов, культивирования и трансформации тканей (например, путем электропорации и липофекции) применяются стандартные методы. Ферментативные реакции и методы очистки осуществляют в соответствии с инструкциями производителей или общеизвестными методами, описанными в литературе или в настоящей заявке. Вышеупомянутые методы и процедуры обычно осуществляют стандартными способами, хорошо известными специалистам и описанными в различных общих и специальных руководствах, цитируемых и обсуждаемых в описании настоящей заявки. См., например, руководство Sambrook et al. Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)). Номенклатура, используемая здесь в описаниях лабораторных процедур и методов аналитической химии, химии органического синтеза и медицинской и фармацевтической химии, хорошо известна и широко применяется специалистами. Химический синтез, химический анализ, приготовление фармацевтических препаратов, составление рецептуры таких препаратов и их доставку, а также лечение пациентов осуществляют стандартными методами.
Термины, используемые в описании настоящего изобретения, имеют значения, указанные ниже, если это не оговорено особо.
Используемый здесь термин «антитело» означает молекулы иммуноглобулина и иммунологически активные части молекул иммуноглобулина (Ig), то есть молекул, которые содержат антигенсвязывающий сайт, специфически связывающийся (подвергающийся иммунной реакции) с антигеном. Термины «специфически связывается», или «подвергается иммунной реакции», или «иммуноспецифически связывается» означают, что антитело реагирует с одной или более антигенными детерминантами нужного антигена и не реагирует с другими полипептидами или связывается с ними с гораздо более низкой аффинностью (Kd>10-6). Антителами являются, но не ограничиваются ими, поликлональное антитело, моноклональное антитело, химерное антитело, доменное антитело, одноцепочечное антитело, Fab- и F(ab')2-фрагменты, scFv и Fab-экспрессирующая библиотека.
Известно, что основной структурной единицей антитела является тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, где каждая пара имеет одну «легкую цепь» (приблизительно 25 кДа) и одну «тяжелую цепь» (приблизительно 50-70 кДа). Амино-концевая часть каждой цепи включает вариабельную область длиной приблизительно 100-110 или более аминокислот, ответственную, главным образом, за распознавание антигена. Карбокси-концевую часть каждой цепи определяют как константную область, ответственную, главным образом, за эффекторную функцию. В общих чертах, молекулы человеческого антитела принадлежат к любому из классов IgG, IgM, IgA, IgE и IgD, которые отличаются друг от друга природой тяжелой цепи, присутствующей в молекуле антитела. Некоторые классы также подразделяются на подклассы, такие как IgG1, IgG2, IgG3, IgG4 и другие. Кроме того, у человека, легкая цепь может представлять собой цепь каппа или лямбда.
Используемые здесь термины «моноклональное антитело» (mAb) или «композиция моноклонального антитела» означают популяцию молекул антитела, которая содержит молекулы антитела только одного вида, состоящие из уникального генного продукта легкой цепи и уникального генного продукта тяжелой цепи. В частности, комплементарность-определяющие области (CDR) моноклонального антитела идентичны во всех молекулах данной популяции. MAb содержат антигенсвязывающий сайт, способный подвергаться иммунной реакции с конкретным эпитопом антигена и характеризующийся уникальной аффинностью связывания с этим эпитопом.
Термин «антигенсвязывающий сайт» или «связывающая часть» означает часть молекулы иммуноглобулина, участвующая в связывании с антигеном. Антигенсвязывающий сайт образован аминокислотными остатками N-концевых вариабельных («V») областей тяжелой цепи («H») и легкой цепи («L»). Три в высокой степени дивергентных фрагмента в V-областях тяжелой и легкой цепей, называемых «гипервариабельными областями», расположены между более консервативными фланкирующими фрагментами, известными как «каркасные области» или «FR». Таким образом, термин «FR» означает аминокислотные последовательности, которые обычно присутствуют между гипервариабельными областями иммуноглобулинов и являются смежными с этими областями. В молекуле антитела, три гипервариабельных области легкой цепи и три гипервариабельных области тяжелой цепи имеют пространственное расположение относительно друг друга и образуют антигенсвязывающую поверхность. Антигенсвязывающая поверхность комплементарна трехмерной поверхности связанного антигена, и три гипервариабельных области каждой тяжелой и легкой цепей называются «комплементарность-определяющими областями» или «CDR». Приписывание аминокислот к каждому домену осуществляется в соответствии с нумерацией по Кэбату (Kabat Sequences of Proteins of Immunological Interest) (National Institutes of Health, Bethesda, Md. (1987 and 1991)) или по Чотию (Chothia & Lesk J. Mol. Biol. 196:901-917 (1987), Chothia et al., Nature 342:878-883 (1989).
Используемый здесь термин «эпитоп» означает любую белковую детерминанту, способную специфически связываться с иммуноглобулином, scFv или T-клеточным рецептором. Термин «эпитоп» включает любую белковую детерминанту, способную специфически связываться с иммуноглобулином или с T-клеточным рецептором. Антигенные детерминанты (эпитопы) обычно состоят из химически активных групп, расположенных на поверхности молекул, таких как аминокислотные или сахарные боковые цепи, и обычно имеют специфические трехмерные структуры, а также обладают специфическими зарядовыми свойствами. Так, например, антитела могут быть направлены против N-концевых или C-концевых пептидов полипептида. Считается, что антитело специфически связывается с антигеном, если константа диссоциации указанного антигена составляет ≤1 мкM, в некоторых вариантах осуществления изобретения, ≤100 нM, а в некоторых вариантах осуществления изобретения, ≤10 нM.
Используемые здесь термины «специфическое связывание», «иммунологическое связывание» и «иммунологические связывающие свойства» означают нековалентное взаимодействие определенного типа, которое происходит между молекулой иммуноглобулина и антигеном, для которого данный иммуноглобулин является специфичным. Сила или аффинность иммунологических взаимодействий типа связывания могут быть определены терминами «константа диссоциации (Kd)» реакции взаимодействия, причем, чем меньше Kd, тем выше аффинность взаимодействия. Иммунологические связывающие свойства выбранных полипептидов могут быть количественно оценены хорошо известными методами. Один из таких методов позволяет определять скорости образования комплекса «антигенсвязывающий сайт/антиген» и диссоциации этого комплекса, где указанные скорости зависят от концентраций партнеров комплекса, аффинности взаимодействия и геометрических параметров, которые в равной степени влияют на скорость в обоих направлениях. Таким образом, «константа скорости ассоциации» (Kon) и «константа скорости диссоциации» (Koff) могут быть определены путем вычисления концентраций и фактических скоростей ассоциации и диссоциации. (См. Nature 361:186-87 (1993)). Отношение Koff/Kon позволяет не учитывать все параметры, не относящиеся к аффинности, и это отношение равно константе диссоциации Kd. (Общий обзор см. Davies et al. (1990) Annual Rev. Biochem. 59:439-473). Говорят, что антитело согласно изобретению специфически связывается с EGFR, если константа равновесной диссоциации при связывании (Kd) составляет ≤1 мкM, например, в некоторых вариантах осуществления изобретения, ≤100 нM, в некоторых вариантах осуществления изобретения, ≤10 нM, а в некоторых вариантах осуществления изобретения, от ≤100 пM и приблизительно до 1 пM, как было определено в анализах, таких как анализы на связывание с радиоактивным лигандом или аналогичные анализы, известные специалистам.
Используемый здесь термин «выделенный полинуклеотид» означает полинуклеотид геномной ДНК, кДНК или синтезированной ДНК или некоторых их комбинаций, где указанный «выделенный полинуклеотид», по своей природе, (1) не ассоциируется с полноразмерным полинуклеотидом или с его частью, в которой этот «выделенный полинуклеотид» является природным, (2) функционально присоединен к полинуклеотиду, с которым он не связан по своей природе, или (3) не присутствует в природе как часть более крупной последовательности. Полинуклеотиды согласно изобретению включают молекулы нуклеиновой кислоты, кодирующие описанные здесь молекулы тяжелой цепи иммуноглобулина, и молекулы нуклеиновой кислоты, кодирующие описанные здесь молекулы легкой цепи иммуноглобулина.
Используемый здесь термин «выделенный белок» означает белок, экспрессируемый кДНК или рекомбинантной РНК, или синтезированный белок или некоторые их комбинации, где указанный «выделенный белок», по своему происхождению или источнику, от которого он происходит, (1) не ассоциируется с природными белками, (2) не содержит других белков, происходящих от того же источника, (3) экспрессируется клетками других видов или (4) не существует в природе.
Термин «полипептид», используемый здесь как общий термин, означает нативный белок, фрагменты или аналоги полипептидной последовательности. Следовательно, фрагменты нативного белка и его аналоги принадлежат к роду полипептидов. Полипептиды согласно изобретению включают описанные здесь молекулы тяжелой цепи иммуноглобулина и молекулы легкой цепи иммуноглобулина, а также молекулы антител, образованные комбинациями, включающими молекулы тяжелой цепи иммуноглобулина и молекулы легкой цепи иммуноглобулина, такие как молекулы легкой цепи каппа иммуноглобулина и наоборот, а также их фрагменты и аналоги.
Используемый здесь термин «природный» относится к объекту, который, фактически, может существовать в природе. Так, например, полипептидная или полинуклеотидная последовательность, которая присутствует в организме (включая вирусы) и может быть выделена из природного источника, и которая не была специально модифицирована человеком в лаборатории или как-либо иначе, является природной.
Используемый здесь термин «функционально присоединенный» относится к положениям описанных здесь компонентов, которые, находясь в данных положениях и взаимодействуя друг с другом, выполняют соответствующие функции. Регуляторная последовательность, «функционально присоединенная» к кодирующей последовательности, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с экспрессией регуляторных последовательностей.
Используемый здесь термин «регуляторная последовательность» означает полинуклеотидные последовательности, необходимые для экспрессии и процессинга кодирующих последовательностей, с которыми они лигированы. Природа таких регуляторных последовательностей зависит от типа организма-хозяина, то есть, от того, является ли этот организм-хозяин прокариотом или эукариотом, и такие регуляторные последовательности обычно включают промотор, сайт связывания с рибосомой и последовательность терминации транскрипции, а чаще всего они включают промоторы и последовательность терминации транскрипции. Термин «регуляторные последовательности» включает, как минимум, все компоненты, присутствие которых играет важную роль в экспрессии и процессинге, и такие последовательности могут также включать дополнительные компоненты, присутствие которых является предпочтительным, например, лидерные последовательности и последовательности партнера по связыванию. Используемый здесь термин «полинуклеотид» означает нуклеотиды длиной по меньшей мере 10 оснований, и представляет собой либо рибонуклеотиды, либо дезоксинуклеотиды, либо модифицированную форму нуклеотида любого типа. Этот термин включает одноцепочечные и двухцепочечные формы ДНК.
Используемый здесь термин «олигонуклеотид» означает природные и модифицированные нуклеотиды, связанные друг с другом природными и неприродными олигонуклеотидными связями. Олигонуклеотиды представляют собой субсерию полинуклеотидов, обычно имеющих длину 200 оснований или менее. В некоторых вариантах осуществления изобретения, олигонуклеотиды имеют длину от 10 до 60 оснований, например, в некоторых вариантах осуществления изобретения, олигонуклеотиды имеют длину 12, 13, 14, 15, 16, 17, 18, 19 или 20-40 оснований. Олигонуклеотиды обычно являются одноцепочечными, например, если они используются в качестве зондов, однако, они могут быть также двухцепочечными, например, если они используются для конструирования генного мутанта. Олигонуклеотидами согласно изобретению являются смысловые или антисмысловые олигонуклеотиды.
Используемый здесь термин «природные нуклеотиды» означает дезоксирибонуклеотиды и рибонуклеотиды. Используемый здесь термин «модифицированные нуклеотиды» означает нуклеотиды с модифицированными или замененными сахарными группами и т.п. Используемый здесь термин «олигонуклеотидные связи» означает такие олигонуклеотидные связи, как фосфортиоат, фосфордитиоат, фосфорселеноат, фосфордиселеноат, фосфоранилотиоат, фосфораниладат, фосфорамидат и т.п. См., например, LaPlanche et al. Nucl. Acids Res. 14:9081 (1986); Stec et al. J. Am. Chem. Soc. 106:6077 (1984), Stein et al. Nucl. Acids Res. 16:3209 (1988), Zon et al. Anti Cancer Drug Design 6:539 (1991); Zon et al. Oligonucleotides and Analogues: A Practical Approach, pp. 87-108 (F. Eckstein, Ed., Oxford University Press, Oxford England (1991)); Stec et al. патент США No. 5151510; Uhlmann and Peyman Chemical Reviews 90:543 (1990). Олигонуклеотид может включать, если это необходимо, метку для детектирования.
Используемые здесь термины «двадцать стандартных аминокислот» и их аббревиатуры употребляются в своем общепринятом значении. См., Immunology-A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderland7 Mass. (1991)). Стереоизомеры (например, D-аминокислот) двадцати стандартных аминокислот; неприродных аминокислот, таких как α,α-дизамещенные аминокислоты; N-алкиламинокислот, молочной кислоты и других нестандартных аминокислот могут также представлять собой подходящие компоненты полипептидов согласно изобретению. Примерами нестандартных аминокислот являются: 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие аналогичные аминокислоты и иминокислоты (например, 4-гидроксипролин). В используемых здесь определениях полипептидов, направление слева направо означает направление от амино-конца, а направление справа налево означает направление от карбокси-конца, и такие обозначения являются стандартными и общепринятыми.
Аналогичным образом, если это не оговорено особо, то направление от левого конца одноцепочечных полинуклеотидных последовательностей означает направление от 5'-конца, то есть, от левого конца двухцепочечных полинуклеотидных последовательностей, и указывается как 5'-направление. Присоединение растущей цепи РНК-транскриптов в направлении 5'→3' называется направлением транскрипции, и области последовательности на цепи ДНК, которые имеют такую же последовательность, как и РНК, и которые находятся со стороны 5'-конца по отношению к 5'-концу РНК-транскрипта, называются «вышерасположенными последовательностями», а области последовательности на цепи ДНК, которые имеют такую же последовательность, как и РНК, и которые находятся со стороны 3'-конца по отношению к 3'-концу РНК-транскрипта, называются «нижерасположенными последовательностями».
Термин «по существу, идендичный», если он относится к полипептидам, означает, что две пептидных последовательности, при их оптимальном выравнивании, например, с помощью программ GAP или BESTFIT, где используются «веса» за удаление и вставку символов по умолчанию, являются идентичными по меньшей мере на 80 процентов, например, в некоторых вариантах осуществления изобретения, две пептидных последовательности являются идентичными по меньшей мере на 90 процентов, например, в некоторых вариантах осуществления изобретения, по меньшей мере на 95 процентов, а в некоторых вариантах осуществления изобретения, по меньшей мере на 99 процентов.
В некоторых вариантах осуществления изобретения, положения остатков, которые не являются идентичными, отличаются консервативными аминокислотными заменами.
Как обсуждается в настоящей заявке, небольшие изменения в аминокислотных последовательностях молекул антител или иммуноглобулинов рассматриваются как изменения, входящие в объем настоящего изобретения, при условии, что в случае внесения изменений в аминокислотную последовательность, идентичность сравниваемой аминокислотной последовательности с эталонной аминокислотной последовательностью будет сохраняться по меньшей мере на 75%, например, в некоторых вариантах осуществления изобретения, по меньшей мере на 80%, 90%, 95%, 99%, а в некоторых вариантах осуществления изобретения, на 99%. В частности, рассматриваются также консервативные аминокислотные замены. Консервативными заменами являются замены аминокислотами, принадлежащими к семейству аминокислот, которые являются родственными по своим боковым цепям. Кодируемые генами аминокислоты, по существу, можно разделить на следующие семейства: (1) кислотные аминокислоты, которыми являются аспартат, глутамат; (2) основные аминокислоты, которыми являются лизин, аргинин, гистидин; (3) неполярные аминокислоты, которыми являются аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные аминокислоты, которыми являются глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Гидрофильными аминокислотами являются аргинин, аспарагин, аспартат, глутамин, глутамат, гистидин, лизин, серин и треонин. Гидрофобными аминокислотами являются аланин, цистеин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан, тирозин и валин. Другие семейства аминокислот включают (i) серин и треонин, которые принадлежат к семейству алифатических аминокислот, содержащих гидроксигруппу; (ii) аспарагин и глутамин, которые принадлежат к семейству аминокислот, содержащих амид; (iii) аланин, валин, лейцин и изолейцин, которые принадлежат к семейству алифатических аминокислот; и (iv) фенилаланин, триптофан и тирозин, которые принадлежат к семейству ароматических аминокислот. Так, например, считается, что отдельная замена лейцина на изолейцин или валин, замена аспартата на глутамат, замена треонина на серин или аналогичная замена одной аминокислоты на другую структурно родственную аминокислоту не будут оказывать значительного влияния на связывание или другие свойства полученной молекулы, например, в случае, если такая замена не осуществляется в гипервариабельной области. Влияние аминокислотной замены на свойства функционального пептида можно легко определить путем анализа удельной активности производного полипептида. Такие анализы подробно описаны в настоящей заявке. Фрагменты или аналоги молекул антител или иммуноглобулинов могут быть легко получены специалистами в данной области. В некоторых вариантах осуществления изобретения, амино- и карбокси-концы фрагментов или аналогов находятся на границах функциональных доменов. Структурные и функциональные домены могут быть идентифицированы путем сравнения данных о нуклеотидных и/или аминокислотных последовательностях с данными, имеющимися в общедоступных базах данные или патентованных базах данных последовательностей. Для идентификации мотивов последовательностей или предсказанных конформационных доменов белка, которые присутствуют в других белках, имеющих известную структуру и/или функцию, применяются компьютеризированные методы сравнения. Методы идентификации белковых последовательностей, которые имеют укладку в виде известной трехмерной структуры, известны специалистам. См. Bowie et al., Science 253:164 (1991). Таким образом, в вышеупомянутых примерах продемонстрировано, что специалист в данной области может самостоятельно определить мотивы последовательности и их структурные конформации, которые могут быть использованы для идентификации структурных и функциональных доменов согласно изобретению.
В некоторых вариантах осуществления изобретения, аминокислотными заменами являются замены, которые: (1) снижают чувствительность к протеолизу, (2) снижают чувствительность к окислению, (3) изменяют аффинность связывания для образования белковых комплексов, (4) изменяют аффинности связывания и (5) сообщают таким аналогам другие физико-химические или функциональные свойства или изменяют эти свойства. Аналоги могут включать различные мутеины с последовательностью, которая отличается от природных пептидных последовательностей. Так, например, одна или несколько аминокислотных замен (например, консервативных аминокислотных замен) могут быть введены в природную последовательность (например, в часть полипептида, расположенную за пределами домена(ов), образующего(их) межмолекулярные контакты). Консервативная аминокислотная замена, по существу, не должна изменять структурные свойства родительской последовательности (например, аминокислотная замена не должна приводить к разрыву спирали, присутствующей в родительской последовательности, или нарушать вторичную структуру других типов, характерную для родительской последовательности). Примеры известных вторичных и третичных структур полипептидов описаны в публикациях «Proteins, Structures and Molecular Principles» (Creighton, Ed., W. H. Freeman and Company, New York (1984)); «Introduction to Protein Structure» (C. Branden and J. Tooze, eds., Garland Publishing, New York, N.Y. (1991)); и Thornton et at. Nature 354:105 (1991).
Используемый здесь термин «полипептидный фрагмент» означает полипептид, который имеет амино-концевую и/или карбокси-концевую делецию и/или одну или более внутренних делеций, но в котором остальная аминокислотная последовательность идентична соответствующим частям природной последовательности, кодируемой, например, полноразмерной последовательностью кДНК. Фрагменты обычно имеют длину по меньшей мере 5, 6, 8 или 10 аминокислот, например, в некоторых вариантах осуществления изобретения, по меньшей мере 14 аминокислот, в некоторых вариантах осуществления изобретения, по меньшей мере 20 аминокислот, а обычно 50 аминокислот, и в некоторых вариантах осуществления изобретения, по меньшей мере 70 аминокислот. Используемый здесь термин «аналог» означает полипептиды, состоящие из сегмента длиной по меньшей мере 25 аминокислот, который, по существу, идентичен части кодированной аминокислотной последовательности, и который специфически связывается с EGFR в подходящих условиях связывания. Обычно, полипептидные аналоги, в отличие от природной последовательности, имеют консервативную аминокислотную замену (или добавление или делецию). Аналоги обычно имеют длину по меньшей мере в 20 аминокислот, например, в некоторых вариантах осуществления изобретения, по меньшей мере в 50 аминокислот или более, а чаще всего, они имеют полноразмерную последовательность природного полипептида.
Используемый здесь термин «агент» означает химическое соединение, смесь химических соединений, биологическую макромолекулу или экстракт, выделенный из биологических материалов.
Используемые здесь термины «метка» или «меченный» относятся к введению детектируемого маркера, например, путем включения радиоактивно меченой аминокислоты или присоединения к полипептиду биотинильных групп, которые могут быть детектированы меченным авидином (например, стрептавидином, содержащим флуоресцентный маркер или обладающим ферментативной активностью, которая может быть детектированы оптическими или калориметрическими методами). В некоторых случаях, метка или маркер также могут представлять собой терапевтический агент. При этом могут быть применены различные методы мечения полипептидов и гликопротеинов, известные специалистам. Примерами меток для полипептидов, являются, но не ограничиваются ими, радиоизотопы или радионуклиды (например, 3H, 14C, 15N, 35S, 90Y, 99Tc, 111In, 125I, 131I), флуоресцентные метки (например, флуорофор, родамин, флуоресцирующие вещества лантанидной группы), ферментные метки (например, пероксидаза хрена, п-галактозидаза, люцифераза, щелочная фосфатаза), хемилюминесцентные метки, биотинильные группы, предварительно определенные полипептидные эпитопы, распознаваемые вторичным репортером (например, последовательности пар «лейциновых молний», сайты связывания для «вторых» антител, металл-связывающие домены, эпитопные метки). В некоторых вариантах осуществления изобретения, метки присоединяют посредством спейсерных групп различной длины для снижения степени потенциальных стерических затруднений. Используемый здесь термин «фармацевтический агент или лекарственное средство» означает химическое соединение или композицию, способные индуцировать желаемый терапевтический эффект при их соответствующем введении пациенту.
Используемый здесь термин «по существу, чистый» относится к объекту определенного вида, который преимущественно присутствует в данном веществе (то есть этот объект, исходя из его молярной концентрации, преобладает по сравнению с любыми отдельными компонентами другого вида, присутствующими в данной композиции), и такая, по существу, очищенная фракция представляет собой композицию, в которой данный объект составляет по меньшей мере приблизительно 50 процентов (по своей молярной концентрации) от всех присутствующих макромолекулярных компонентов одного вида.
Вообще говоря, по существу, чистая композиция содержит приблизительно более чем 80 процентов от всех макромолекулярных компонентов, присутствующих в этой композиции, а в некоторых вариантах осуществления изобретения, приблизительно более, чем 85%, 90%, 95% и 99%. В некоторых вариантах осуществления изобретения, данный объект очищен почти до гомогенности (это означает, что примеси, имеющиеся в данной композиции, не могут быть детектированы стандартными методами детектирования), где указанная композиция состоит, в основном, из макромолекул одного вида.
Термин «пациент» включает человека и животное, находящееся под наблюдением ветеринара.
Другие используемые здесь химические термины употребляются в своем общепринятом смысле, как было определено в Словаре химических терминов [McGraw-Hill Dictionary of Chemical Terms (Parker, S., Ed., McGraw-Hill, San Francisco (1985)].
Мультиспецифические антитела и мультиспецифические активируемые антитела
Репрезентативными мультиспецифическими антителами и/или мультиспецифическими активируемыми антителами согласно изобретению являются, например, антитела, описанные в настоящей заявке, в разделе «Примеры», и их варианты.
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ в мультиспецифическом антителе является партнером по связыванию с любой мишенью, представленной в таблице 1.
Репрезентативные мишени
интегрин
интегрин
CA-125)
(IL-2RG)
(Lewis a)
Notch
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ мультиспецифического антитела представляет собой последовательность или происходит от последовательности, представленной в таблице 7 в разделе «Примеры» настоящей заявки.
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ мультиспецифического антитела представляет собой антитело или происходит от антитела, представленного в таблице 2.
Репрезентативные источники Ab
В настоящем изобретении также описаны мультиспецифические антитела и/или мультиспецифические активируемые антитела, связывающиеся с тем же самым эпитопом, с которым связываются описанные здесь мультиспецифические антитела и/или мультиспецифические активируемые антитела.
Специалист в данной области без излишнего экспериментирования может определить, имеет ли мультиспецифическое антитело и/или мультиспецифическое активируемое антитело такую же специфичность, как и мультиспецифическое антитело и/или мультиспецифическое активируемое антитело согласно изобретению, или аналогичную специфичность, и для этого необходимо установить, может ли первое антитело блокировать связывание второго антитела с мишенью. Если протестированное мультиспецифическое антитело и/или мультиспецифическое активируемое антитело конкурирует с мультиспецифическим антителом и/или с мультиспецифическим активируеым антителом согласно изобретению, на что указывает снижение уровня связывания мультиспецифического антитела и/или мультиспецифического активируемого антитела согласно изобретению, то это означает, что два мультиспецифических антитела и/или мультиспецифических активируемых антитела связываются с одним и тем же или с близкородственным эпитопом.
Одним из вариантов определения того факта, имеет ли мультиспецифическое антитело и/или мультиспецифическое активируемое антитело такую же специфичность, как и мультиспецифическое антитело и/или мультиспецифическое активируемое антитело согласно изобретению, или аналогичную специфичность, является предварительное инкубирование мультиспецифического антитела и/или мультиспецифического активируемого антитела согласно изобретению с растворимой мишенью, с которой оно обычно реагирует, и последующее добавление тестируемого мультиспецифического антитела и/или мультиспецифического активируемого антитела для того, чтобы определить, может ли тестируемое мультиспецифическое антитело и/или мультиспецифическое активируемое антитело ингибировать способность связываться с мишенью. Если тестируемое мультиспецифическое антитело и/или мультиспецифическое активируемое антитело ингибируется, то, по всей вероятности, это означает, что оно имеет такую же специфичность к эпитопу, как и специфичность мультиспецифического антитела и/или мультиспецифического активируемого антитела согласно изобретению, или функционально эквивалентную специфичность.
Мультиспецифическое антитело и/или мультиспецифическое активируемое антитело получают, например, методами, описанными ниже в примерах. Мультиспецифическое антитело и/или мультиспецифическое активируемое антитело может быть также получено любыми хорошо известными методами продуцирования и/или очистки антител.
Фрагменты антител, такие как Fv, F(ab’)2 и Fab, используемые в мультиспецифическом антителе и/или мультиспецифическом активируемом антителе, могут быть получены путем расщепления интактного белка, например, посредством расщепления протеазой или химического расщепления. Альтернативно, может быть сконструирован усеченный ген. Так, например, химерный ген, кодирующий часть F(ab’)2-фрагмента, может включать последовательности ДНК, кодирующие домен CH1 и шарнирную область Н-цепи, за которыми следует кодон терминации трансляции, в результате чего может быть получена усеченная молекула.
Экспрессионными векторами являются плазмиды, ретровирусы, YAC, эписомы, происходящие от EBV, и т.п. Стандартным вектором является вектор, кодирующий функционально полноценную последовательность CH или CL человеческого иммуноглобулина, причем, соответствующие рестрикционные сайты конструируют так, чтобы любая последовательность VH или VL могла быть легко встроена и экспрессирована. В таких векторах, сплайсинг обычно осуществляется между сайтом-донором сплайсинга во встроенной J-области, и сайтом-акцептором сплайсинга, за которым следует человеческая С-область, и кроме того, такой сплайсинг также осуществляется в областях сплайсинга, присутствующих в человеческих экзонах СН. Полиаденилирование и терминация транскрипции осуществляется в нативных хромосомных сайтах, расположенных ниже кодирующих областей. Полученное антитело может быть присоединено к любому сильному промотору, включая ретровирусные LTR, например, LTR раннего промотора SV-40 (Okayama et al. Mol. Cell. Bio. 3:280 (1983)), LTR вирус саркомы Рауса (Gorman et al. P.N.A.S. 79:6777 (1982)), и LTR вируса мышиного лейкоза Молони (Grosschedl et al. Cell 41:885 (1985)). Следует отметить, что могут быть также использованы нативные промоторы Ig и т.п.
Кроме того, мультиспецифические антитела и/или мультиспецифические активируемые антитела могут быть получены методами представления, включая, но не ограничиваясь ими, методы фагового представления, ретровирусного представления, рибосомного представления и т.п., и другими методами, хорошо известными специалистам, и полученные таким образом молекулы могут быть подвергнуты дополнительному созреванию, такому как аффинное созревание, хорошо известное специалистам. См. Wright et al. Crit, Reviews in Immunol. 12125-168 (1992), Hanes and Plückthun PNAS USA 94:4937-4942 (1997) (рибосомное представление), Parmley and Smith Gene 73:305-318 (1988) (фаговое представление), Scott, TIBS, vol. 17:241-245 (1992), Cwirla et al. PNAS USA 87:6378-6382 (1990), Russel et al. Nucl. Acids Research 21:1081-1085 (1993), Hoganboom et al. Immunol. Reviews 130:43-68 (1992), Chiswell and McCafferty TIBTECH; 10:80-8A (1992) и патент США No. 5733743.
Может оказаться желательной модификация мультиспецифического антитела и/или мультиспецифического активируемого антитела согласно изобретению, а именно, модификация эффекторной функции в целях усиления или ослабления такой функции для повышения эффективности антитела при лечении заболеваний и расстройств. Так, например, в Fc-область может (могут) быть введен(ы) цистеиновый(е) остаток (остатки), что будет приводить к образованию межцепьевой дисульфидной связи в этой области. Полученное таким образом гомодимерное антитело может иметь повышенную способность к интернализации и/или повышенную способность к комплемент-опосредуемому цитолизу клеток и повышенную антитело-зависимую клеточную цитотоксичность (ADCC). (см., Caron et al., J. Exp Med., 176: 1191-1195 (1992) and Shopes, J. Immunol., 148: 2918-2922 (1992)). Альтернативно, может быть сконструировано антитело, которое имеет две Fc-области, а поэтому, оно обладает повышенной способностью к лизису комплемента и повышенной ADCC-активностью (см. Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989)). В некоторых вариантах осуществления изобретения могут быть введены Fc-мутации для удаления сайтов гликозилирования и тем самым для снижения функции Fc.
Мультиспецифические активируемые антитела
Описанные здесь мультиспецифические активируемые антитела и композиции мультиспецифических активируемых антител содержат по меньшей мере первое антитело или его фрагмент (имеющие в настоящем описании общее название AB1), которые специфически связываются с первой мишенью и/или с первым эпитопом, и второе антитело или его фрагмент (имеющие в настоящем описании общее название AB2), которые специфически связываются со второй мишенью и/или со вторым эпитопом, где по меньшей мере одно AB модифицировано маскирующей группой (MM). В некоторых вариантах осуществления изобретения, каждое AB в мультиспецифическом активируемом антителе модифицировано своей собственной маскирующей группой.
Если по меньшей мере одно AB в мультиспецифическом активируемом антителе модифицировано MM и находится в присутствии его мишени, то специфическое связывание АВ с его мишенью снижается или ингибируется по сравнению со специфическим связыванием АВ, не модифицированного группой ММ, с мишенью или по сравнению со специфическим связыванием родительского АВ с мишенью.
Kd при связывании MM-модифицированного AB с мишенью, по меньшей мере в 5, 10, 20, 25, 40, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 50000, 100000, 500000, 1000000, 5000000, 10000000, 50000000 или более или в пределах 5-10, 10-100, 10-1000, 10-10000, 10-100000, 10-1000000, 10-10000000, 100-1000, 100-10000, 100-100000, 100-1000000, 100-10000000, 1000-10000, 1000-100000, 1000-1000000, 1000-10000000, 10000-100000, 10000-1000000, 10000-10000000, 100000-1000000 или 100000-10000000 раз превышает Kd при связывании AB, не модифицированного группой ММ, с мишенью или родительского АВ с мишенью. И наоборот, аффинность связывания MM-модифицированного AB с мишенью по меньшей мере в 5, 10, 20, 25, 40, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 50000, 100000, 500000, 1000000, 5000000, 10000000, 50000000 или более или в пределах 5-10, 10-100, 10-1000, 10-10000, 10-100000, 10-1000000, 10-10000000, 100-1000, 100-10000, 100-100000, 100-1000000, 100-10000000, 1000-10000, 1000-100000, 1000-1000000, 1000-10000000, 10000-100000, 10000-1000000, 10000-10000000, 100000-1000000 или 100000-10000000 раз ниже, чем аффинность связывания AB, не модифицированного ММ, с мишенью или родительского АВ с мишенью.
Константа диссоциации (Kd) при связывании по меньшей мере одного MM-модифицированного AB в мультиспецифическом активируемом антителе с мишенью обычно превышает Kd при связывании AB с мишенью. Kd при связывания MM-модифицированного AB с мишенью по меньшей мере в 5, 10, 25, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 100000, 1000000 или даже в 10000000 раз выше, чем Kd при связывании AB с мишенью. И наоборот, аффинность связывания MM-модифицированного AB с мишенью обычно ниже, чем аффинность связывания АВ с мишенью. Аффинность связывания MM-модифицированного AB с мишенью может быть по меньшей мере в 5, 10, 25, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 100000, 1000000 или даже в 10000000 раз ниже, чем аффинность связывания AB с мишенью.
Если по меньшей мере одно AB в мультиспецифическом активируемом антителе модифицировано MM и находится в присутствии его мишени, то специфическое связывание АВ с его мишенью снижается или ингибируется по сравнению со специфическим связыванием АВ, не модифицированного группой ММ, с мишенью или по сравнению со специфическим связыванием родительского АВ с мишенью. В отличие от связывания AB, не модифицированного группой ММ, с мишенью, или связывания родительского АВ мишенью, способность АВ связываться с мишенью в том случае, если оно модифицировано ММ, может быть снижена по меньшей мере на 50%, 60%, 70%, 80%, 90%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и даже на 100% по меньшей мере через 2, 4, 6, 8, 12, 28, 24, 30, 36, 48, 60, 72, 84 или 96 часов, или 5, 10, 15, 30, 45, 60, 90, 120, 150 или 180 дней, или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев или более, как было определено в анализе in vivo или в анализе in vitro.
MM ингибирует связывание по меньшей мере одного AB мультиспецифического активируемого антитела с его мишенью. MM связывается с антигенсвязывающим доменом АВ и ингибирует связывание АВ с его мишенью. The MM может стерически ингибировать связывание АВ с мишенью. MM может аллостерически ингибировать связывание АВ с мишенью. В этих вариантах осуществления изобретения, если AB модифицировано ММ или связано с MM и находится в присутствии мишени, то в этом случае, какого-либо связывания или какого-либо детектируемого связывания АВ с мишенью не происходит, либо уровень связывания АВ с мишенью не превышает 0,001%, 0,01%, 0,1%, 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40% или 50% по сравнению с уровнем связывания AB, не модифицированного ММ, с мишенью, связывания родительского АВ с мишенью или связывания АВ, не присоединенного к ММ, с мишенью, по меньшей мере через 2, 4, 6, 8, 12, 28, 24, 30, 36, 48, 60, 72, 84 или 96 часов, или 5, 10, 15, 30, 45, 60, 90, 120, 150 или 180 дней, или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев или более, как было определено в анализе in vivo или в анализе in vitro.
Если по меньшей мере одно АВ в мультиспецифическом активируемом антителе связано с ММ или модифицировано ММ, то такая ММ «маскирует», снижает или как-либо иначе ингибирует специфическое связывание АВ с мишенью. Если по меньшей мере одно АВ в мультиспецифическом активируемом антителе связано с ММ или модифицировано ММ, то такое связывание или такая модификация могут приводить к структурным изменениям, которые, в свою очередь, будут снижать или ингибировать способность АВ специфически связываться с мишенью.
В мультиспецифическом активируемом антителе, если по меньшей мере одно АВ в мультиспецифическом активируемом антителе связано с ММ или модифицировано ММ, то по меньшей мере часть мультиспецифического активируемого антитела может быть представлена нижеследующими формулами (в направлении от амино(N)-концевой области до карбокси(C)-концевой области:
(MM)-(AB)
(AB)-(MM)
(MM)-L-(AB)
(AB)-L-(MM),
где: MM означает маскирующую группу, AB означает антитело или его фрагмент, а L означает линкер. Во многих вариантах осуществления изобретения, в композицию, для придания ей гибкости, желательно ввести один или более линкеров, например, гибких линкеров.
В некоторых вариантах осуществления изобретения, MM не является природным партнером по связыванию с АВ. В некоторых вариантах осуществления изобретения, MM не содержит какого-либо природного партнера по связыванию с АВ или, по существу, не гомологична этому партнеру. В других вариантах осуществления изобретения, MM не более чем на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 80% аналогична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 80% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 25% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 50% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 20% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 10% идентична любому природному партнеру по связыванию с AB.
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела включают AB, которое было модифицировано MM, а также одну или более отщепляемых групп (CM). Такие мультиспецифические активируемые антитела обладают способностью к активируемому/переключаемому связыванию с мишенью АВ. Мультиспецифические активируемые антитела, по существу, представляют собой по меньшей мере одно антитело или его фрагмент (AB), модифицированные маскирующей группой или связанные с маскирующей группой (MM) и содержащие модифицируемую или отщепляемую группу (CM). В некоторых вариантах осуществления изобретения, CM содержит аминокислотную последовательность, которая служит в качестве субстрата для представляющей интерес протеазы.
Элементы мультиспецифических активируемых антител могут быть расположены таким образом, что если каждая из MM и CM находится в расщепленном (или в относительно активном) состоянии и в присутствии мишени, то соответствующее AB связывается с мишенью, а если они находятся в нерасщепленном (или в относительно неактивном) состоянии и в присутствии мишени, то специфическое связывание АВ с мишенью снижается или ингибируется. Специфическое связывание АВ с мишенью пожет подавляться посредством ингибирования или маскировки способности АВ специфически связываться с мишенью посредством MM.
Kd каждого AB, модифицированного MM и CM, при его связывании с мишенью, по меньшей мере в 5, 10, 20, 25, 40, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 50000, 100000, 500000, 1000000, 5000000, 10000000, 50000000 или более или в пределах 5-10, 10-100, 10-1000, 10-10000, 10-100000, 10-1000000, 10-10000000, 100-1000, 100-10000, 100-100000, 100-1000000, 100-10000000, 1000-10000, 1000-100000, 1000-1000000, 1000-10000000, 10000-100000, 10000-1000000, 10000-10000000, 100000-1000000 или 100000-10000000 раз превышает Kd при связывании AB, не модифицированного ММ и СМ, с мишенью или родительского АВ с мишенью. И наоборот, аффинность связывания каждого MM- и СМ-модифицированного AB с мишенью по меньшей мере в 5, 10, 20, 25, 40, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 50000, 100000, 500000, 1000000, 5000000, 10000000, 50000000 или более или в пределах 5-10, 10-100, 10-1000, 10-10000, 10-100000, 10-1000000, 10-10000000, 100-1000, 100-10000, 100-100000, 100-1000000, 100-10000000, 1000-10000, 1000-100000, 1000-1000000, 1000-10000000, 10000-100000, 10000-1000000, 10000-10000000, 100000-1000000 или 100000-10000000 раз ниже, чем аффинность связывания AB, не модифицированного ММ и СМ, с мишенью или родительского АВ с мишенью.
Если по меньшей мере одно AB модифицировано MM и СМ и находится в присутствии его мишени, но не в присутствии модифицирующего агента (например, протеазы), то специфическое связывание АВ с мишенью снижается или ингибируется по сравнению со специфическим связыванием АВ, не модифицированного ММ и СМ, с мишенью или по сравнению со специфическим связыванием родительского АВ с мишенью. В отличие от связывания родительского АВ с мишенью или связывания AB, не модифицированного ММ и СМ, с мишенью, способность АВ связываться с мишенью в том случае, если оно модифицировано ММ и СМ, может быть снижена по меньшей мере на 50%, 60%, 70%, 80%, 90%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и даже на 100% по меньшей мере через 2, 4, 6, 8, 12, 28, 24, 30, 36, 48, 60, 72, 84 или 96 часов, или 5, 10, 15, 30, 45, 60, 90, 120, 150 или 180 дней, или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев или более, как было определено в анализе in vivo или в анализе in vitro.
Используемый здесь термин «расщепленное состояние» означает состояние мультиспецифических активируемых антител после модификации СМ под действием протеазы. Используемый здесь термин «нерасщепленное состояние» означает состояние мультиспецифических активируемых антител в отсутствии отщепления СМ протеазой. Как обсуждалось выше, используемый здесь термин «мультиспецифические активируемые антитела» означает мультиспецифическое активируемое антитело, присутствующее в нерасщепленном (нативном) состоянии, а также в расщепленном состоянии. Для специалиста в данной области очевидно, что в некоторых вариантах осуществления изобретения, расщепленное мультиспецифическое активируемое антитело может не содержать ММ, что обусловлено отщеплением группы СМ протеазой, которое приводит к высвобождению по меньшей мере MM (например, в случае, если MM не присоединена к мультиспецифическим активируемым антителам посредством ковалентной связи (например, дисульфидной связи между цистеиновыми остатками)).
Термин «активируемое или переключаемое связывание» означает, что мультиспецифическое активируемое антитело находится на первом уровне связывания с мишенью, если оно присутствует в ингибированном, маскированном или нерасщепленном состоянии (то есть, в первой конформации), и на втором уровне связывания с мишенью, если оно присутствует в неингибированном, немаскированном и/или отщепленном состоянии (то есть во второй конформации), где второй уровень связывания с мишенью превышает первый уровень связывания. Вообще говоря, доступность мишени для соответствующего AB мультиспецифического активируемого антитела повышается в присутствии расщепляющего агента, способного отщеплять СМ, в отличие от ситуации, когда такой агент отсуствует. Таким образом, если мультиспецифическое активируемое антитело присутствует в нерасщепленном состоянии, то связывание по меньшей мере одного АВ с мишенью ингибируется и может быть замаскировано (то есть, в первой конформации, АВ не может связываться с мишенью), а если оно присутствует в расщепленном состоянии, то связывание АВ с мишенью не ингибируется или не маскируется.
CM и AB мультиспецифических активируемых антител выбирают так, чтобы первое АВ представляло собой группу, связывающуюся с первой мишенью и/или с первым эпитопом, а СМ представляла собой субстрат для протеазы, который локализуется вместе с мишенью на участке, подвергаемом лечению или диагностике в организме индивидуума. Описанные здесь мультиспецифические активируемые антитела, в частности, применяются в том случае, когда, например, протеаза, способная расщеплять сайт в CM, присутствует на относительно более высоких уровнях в мишень-содержащей ткани на участке, подвергаемом лечению или диагностике, чем в ткани непораженных участков (например, в здоровой ткани).
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела обладают пониженной токсичностью и/или дают меньше побочных эффектов, чем это происходит при связывании первого АВ в непораженных участках, если связывание АВ с мишенью не маскируется или как-либо иначе не ингибируется.
В общих чертах, мультиспецифическое активируемое антитело может быть сконструировано путем отбора первого представляющего интерес АВ и остальной части активируемого антитела так, чтобы ММ, при ее конформационном ограничении, обеспечивала маскировку связывания АВ с мишенью или снижение уровня связывания АВ с мишенью. Для обеспечения таких функциональных признаков следует руководствоваться структурными критериями конструирования.
Настоящее изобретение относится к мультиспецифическим активируемым антителам, имеющим переключаемый фенотип в желаемом динамическом интервале связывания с мишенью в ингибированной конформации в отличие от фенотипа в неингибированной конформации. Термин «динамический интервал», по существу, означает, отношение (a) максимального детектированного уровня параметра в первом ряде условий к (b) минимальному детектированному уровню параметра во втором ряде условий. Так, например, в случае мультиспецифического активируемого антитела, «динамический интервал» означает, отношение (a) максимального детектированного уровня связывания белка-мишени с мультиспецифическим активируемым антителом в присутствии протеазы, способной отщеплять СМ от активируемых антител, к (b) минимальному детектированному уровню связывания белка-мишени с мультиспецифическим активируемым антителом в отсутствии протеазы. Динамический интервал для мультиспецифического активируемого антитела может быть вычислен как отношение константы равновесной диссоциации мультиспецифического активируемого антитела, обработанного расщепляющим агентом (например, ферментом) к константе равновесной диссоциации активируемых антител, обработанных расщепляющим агентом. Чем выше динамический интервал для мультиспецифического активируемого антитела, тем лучше переключаемый фенотип активируемого антитела. Активируемые антитела с относительно более высокой величиной динамического интервала (например, выше, чем 1) имеют более желательный переключающий фенотип, при котором связывание белка мишени с активируемыми антителами происходит на более высоком уровне (например, на доминантном уровне) в присутствии расщепляющего агента (например, фермента), способного отщеплять CM от активируемых антител, чем в отсутствии расщепляющего агента.
Мультиспецифические активируемые антитела могут иметь различные структурные конфигурации. Репрезентативные формулы по меньшей мере части мультиспецифического активируемого антитела представлены ниже. В частности, в активируемом антителе, порядок расположения элементов от N-конца до С-конца, а именно: первое АВ, соответствующие MM и CM, может быть изменен на обратный. Также может быть рассмотрен случай, когда CM и MM в аминокислотной последовательности могут перекрываться, например, СМ может находиться в ММ.
Так, например, по меньшей мере часть мультиспецифических активируемых антител может быть представлена нижеследующими формулами (в направлении от амино(N)-концевой области до карбокси(C)-концевой области:
(MM)-(CM)-(AB)
(AB)-(CM)-(MM),
где: MM означает маскирующую группу, СМ означает отщепляемую группу, а AB означает первое антитело или его фрагмент. Следует отметить, что, хотя MM и CM указаны в вышеприведенных формулах как отдельные компоненты, однако, во всех репрезентативных вариантах осуществления изобретения (включая формулы), аминокислотные последовательности MM и CM могут перекрываться, например, CM может полностью или частично содержаться в MM. Кроме того, в вышеуказанных формулах, дополнительные аминокислотные последовательности могут быть расположены у N-конца или C-конца по отношению к элементам активируемых антител.
В некоторых вариантах осуществления изобретения, MM не является природным партнером по связыванию с АВ. В некоторых вариантах осуществления изобретения, MM не содержит какого-либо природного партнера по связыванию с АВ или, по существу, не гомологична этому партнеру. В других вариантах осуществления изобретения, MM не более, чем на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 80% аналогична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 80% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более, чем на 25% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 50% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 20% идентична любому природному партнеру по связыванию с AB. В некоторых вариантах осуществления изобретения, MM не более чем на 10% идентична любому природному партнеру по связыванию с AB.
Во многих вариантах осуществления изобретения, может оказаться желательным введение в конструкцию мультиспецифического активируемого антитела одного или более линкеров, например, гибких линкеров, для придания одному или более MM-CM-стыкам, CM-AB-стыкам или обоим стыкам большей гибкости. Так, например, AB, MM, и/или CM могут не содержать достаточного количества остатков (например, Gly, Ser, Asp, Asn, а в частности, Gly и Ser, а конкретно Gly), что будет обеспечивать желаемую гибкость. По существу, введение одной или более аминокислот для обеспечения гибкости линкера может оказаться полезным для сообщения указанным конструкциям мультиспецифического активируемого антитела переключаемого фенотипа. Кроме того, как описано ниже, если мультиспецифическое активируемое антитело было получено в виде конструкции с конформационными ограничениями, то в нее может быть введен гибкий линкер для облегчения образования и сохранения циклической структуры нерасщепленного мультиспецифического активируемого антитела.
Так, например, в некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело имеет одну из нижеследующих структурных формул (где указанная ниже формула представляет собой аминокислотную последовательность, расположенную либо в направлении от N-конца до C-конца, либо от С-конца до N-конца):
(MM)-L1-(CM)-(AB)
(MM)-(CM)-L2-(AB)
(MM)-L1-(CM)-L2-(AB),
где MM, CM и AB определены выше; и где каждый из L1 и L2, независимо друг от друга и необязательно, присутствует или отсутствуют, и оба они представляют собой одинаковые или различные гибкие линкеры, включающие по меньшей мере 1 гибкую аминокислоту (например, Gly). Кроме того, в вышеуказанных формулах, дополнительные аминокислотные последовательности могут быть расположены у N-конца или C-конца по отношению к элементам активируемых антител. В качестве примеров служат, но не ограничиваются ими, нацеливающие молекулы (например, лиганд для клеточного рецептора, присутствующего в ткани-мишени) и молекулы, увеличивающие время полужизни в сыворотке (например, полипептиды, связывающиеся с сывороточными белками, такие как иммуноглобулин (например, IgG) или сывороточный альбумин (например, альбумин человеческой сыворотки (HAS)).
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно AB в мультиспецифическом активируемом антителе является партнером по связыванию с любой мишенью, представленной в таблице 1.
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ в мультиспецифическом активируемом антителе представляет собой последовательность или происходит от последовательности, представленной в таблице 7 в разделе «Примеры» настоящей заявки.
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ в мультиспецифическом активируемом антителе представляет собой последовательность или происходит от последовательности, представленной в примере 5 в разделе «Примеры» настоящей заявки. В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ в мультиспецифическом активируемом антителе представляет собой последовательность или происходит от последовательности, представленной в таблице 11 в разделе «Примеры» настоящей заявки.
В некоторых неограничивающих вариантах осуществления изобретения, по меньшей мере одно АВ в мультиспецифическом активируемом антителе представляет собой антитело или происходит от антитела, представленного в таблице 2.
В некоторых вариантах осуществления изобретения выбирают маскирующую группу, подходящую для ее использования в специфическом антителе или в его фрагменте. Так, например, маскирующими группами, подходящими для их использования в антителах, связывающихся с EGFR, являются MM, которые имеют последовательность CISPRG (SEQ ID NO: 75). Неограничивающими примерами MM могут быть такие последовательности, как CISPRGC (SEQ ID NO: 339), CISPRGCG (SEQ ID NO: 76); CISPRGCPDGPYVMY (SEQ ID NO: 77); CISPRGCPDGPYVM (SEQ ID NO: 78), CISPRGCEPGTYVPT (SEQ ID NO: 79) and CISPRGCPGQIWHPP (SEQ ID NO: 80). Другими подходящими маскирующими группами являются любые EGFR-специфические маскирующеи группы, описанные в публикации заявки PCT No. WO 2010/081173, такие как, но не ограничивающиеся ими, например, GSHCLIPINMGAPSC (SEQ ID NO: 81); CISPRGCGGSSASQSGQGSHCLIPINMGAPSC (SEQ ID NO: 82); CNHHYFYTCGCISPRGCPG (SEQ ID NO: 83); ADHVFWGSYGCISPRGCPG (SEQ ID NO: 84); CHHVYWGHCGCISPRGCPG (SEQ ID NO: 85); CPHFTTTSCGCISPRGCPG (SEQ ID NO: 86); CNHHYHYYCGCISPRGCPG (SEQ ID NO: 87); CPHVSFGSCGCISPRGCPG (SEQ ID NO: 88); CPYYTLSYCGCISPRGCPG (SEQ ID NO: 89); CNHVYFGTCGCISPRGCPG (SEQ ID NO: 90); CNHFTLTTCGCISPRGCPG (SEQ ID NO: 91); CHHFTLTTCGCISPRGCPG (SEQ ID NO: 92); YNPCATPMCCISPRGCPG (SEQ ID NO: 93); CNHHYFYTCGCISPRGCG (SEQ ID NO: 94); CNHHYHYYCGCISPRGCG (SEQ ID NO: 95); CNHVYFGTCGCISPRGCG (SEQ ID NO: 96); CHHVYWGHCGCISPRGCG (SEQ ID NO: 97); CPHFTTTSCGCISPRGCG (SEQ ID NO: 98); CNHFTLTTCGCISPRGCG (SEQ ID NO: 99); CHHFTLTTCGCISPRGCG (SEQ ID NO: 100); CPYYTLSYCGCISPRGCG (SEQ ID NO: 101); CPHVSFGSCGCISPRGCG (SEQ ID NO: 102); ADHVFWGSYGCISPRGCG (SEQ ID NO: 103); YNPCATPMCCISPRGCG (SEQ ID NO: 104); CHHVYWGHCGCISPRGCG (SEQ ID NO: 105); C(N/P)H(H/V/F)(Y/T)(F/W/T/L)(Y/G/T/S)(T/S/Y/H)CGCISPRGCG (SEQ ID NO: 106); CISPRGCGQPIPSVK (SEQ ID NO: 107); CISPRGCTQPYHVSR (SEQ ID NO: 108); и/или CISPRGCNAVSGLGS (SEQ ID NO: 109).
Маскирующими группами, подходящими для их использования в антителах, связывающихся с мишенью Jagged, например, с Jagged 1 и/или Jagged 2, являются, но не ограничиваются ими, например, маскирующие группы, имеющие такие последовательности, как QGQSGQCNIWLVGGDCRGWQG (SEQ ID NO: 338); QGQSGQGQQQWCNIWINGGDCRGWNG (SEQ ID NO: 110); PWCMQRQDFLRCPQP (SEQ ID NO: 111); QLGLPAYMCTFECLR (SEQ ID NO: 112); CNLWVSGGDCGGLQG (SEQ ID NO: 113); SCSLWTSGSCLPHSP (SEQ ID NO: 114); YCLQLPHYMQAMCGR (SEQ ID NO: 115); CFLYSCTDVSYWNNT (SEQ ID NO: 116); PWCMQRQDYLRCPQP (SEQ ID NO: 117); CNLWISGGDCRGLAG (SEQ ID NO: 118); CNLWVSGGDCRGVQG (SEQ ID NO: 119); CNLWVSGGDCRGLRG (SEQ ID NO: 120); CNLWISGGDCRGLPG (SEQ ID NO: 121); CNLWVSGGDCRDAPW (SEQ ID NO: 122); CNLWVSGGDCRDLLG (SEQ ID NO: 123); CNLWVSGGDCRGLQG (SEQ ID NO: 124); CNLWLHGGDCRGWQG (SEQ ID NO: 125); CNIWLVGGDCRGWQG (SEQ ID NO: 126); CTTWFCGGDCGVMRG (SEQ ID NO: 127); CNIWGPSVDCGALLG (SEQ ID NO: 128); CNIWVNGGDCRSFEG (SEQ ID NO: 129); YCLNLPRYMQDMCWA (SEQ ID NO: 130); YCLALPHYMQADCAR (SEQ ID NO: 131); CFLYSCGDVSYWGSA (SEQ ID NO: 132); CYLYSCTDSAFWNNR (SEQ ID NO: 133); CYLYSCNDVSYWSNT (SEQ ID NO: 134); CFLYSCTDVSYW (SEQ ID NO: 135); CFLYSCTDVAYWNSA (SEQ ID NO: 136); CFLYSCTDVSYWGDT (SEQ ID NO: 137); CFLYSCTDVSYWGNS (SEQ ID NO: 138); CFLYSCTDVAYWNNT (SEQ ID NO: 139); CFLYSCGDVSYWGNPGLS (SEQ ID NO: 140); CFLYSCTDVAYWSGL (SEQ ID NO: 141); CYLYSCTDGSYWNST (SEQ ID NO: 142); CFLYSCSDVSYWGNI (SEQ ID NO: 143); CFLYSCTDVAYW (SEQ ID NO: 144); CFLYSCTDVSYWGST (SEQ ID NO: 145); CFLYSCTDVAYWGDT (SEQ ID NO: 146); GCNIWLNGGDCRGWVDPLQG (SEQ ID NO: 147); GCNIWLVGGDCRGWIGDTNG (SEQ ID NO: 148); GCNIWLVGGDCRGWIEDSNG (SEQ ID NO: 149); GCNIWANGGDCRGWIDNIDG (SEQ ID NO: 150); GCNIWLVGGDCRGWLGEAVG (SEQ ID NO: 151); GCNIWLVGGDCRGWLEEAVG (SEQ ID NO: 152); GGPALCNIWLNGGDCRGWSG (SEQ ID NO: 153); GAPVFCNIWLNGGDCRGWMG (SEQ ID NO: 154); GQQQWCNIWINGGDCRGWNG (SEQ ID NO: 155); GKSEFCNIWLNGGDCRGWIG (SEQ ID NO: 156); GTPGGCNIWANGGDCRGWEG (SEQ ID NO: 157); GASQYCNLWINGGDCRGWRG (SEQ ID NO: 158); GCNIWLVGGDCRPWVEGG (SEQ ID NO: 159); GCNIWAVGGDCRPFVDGG (SEQ ID NO: 160); GCNIWLNGGDCRAWVDTG (SEQ ID NO: 161); GCNIWIVGGDCRPFINDG (SEQ ID NO: 162); GCNIWLNGGDCRPVVFGG (SEQ ID NO: 163); GCNIWLSGGDCRMFMNEG (SEQ ID NO: 164); GCNIWVNGGDCRSFVYSG (SEQ ID NO: 165); GCNIWLNGGDCRGWEASG (SEQ ID NO: 166); GCNIWAHGGDCRGFIEPG (SEQ ID NO: 167); GCNIWLNGGDCRTFVASG (SEQ ID NO: 168); GCNIWAHGGDCRGFIEPG (SEQ ID NO: 169); GFLENCNIWLNGGDCRTG (SEQ ID NO: 170); GIYENCNIWLNGGDCRMG (SEQ ID NO: 171); и/или GIPDNCNIWINGGDCRYG (SEQ ID NO: 172).
Маскирующими группами, подходящими для их использования в антителах, связывающихся с мишенью, такой как интерлейкин 6, например, рецептор интерлейкина 6 (IL-6R), являются, но не ограничиваются ими, например, маскирующие группы, имеющие такие последовательности, как QGQSGQYGSCSWNYVHIFMDC (SEQ ID NO: 174); QGQSGQGDFDIPFPAHWVPIT (SEQ ID NO: 175); QGQSGQMGVPAGCVWNYAHIFMDC (SEQ ID NO: 176); YRSCNWNYVSIFLDC (SEQ ID NO: 177); PGAFDIPFPAHWVPNT (SEQ ID NO: 178); ESSCVWNYVHIYMDC (SEQ ID NO: 179); YPGCKWNYDRIFLDC (SEQ ID NO: 180); YRTCSWNYVGIFLDC (SEQ ID NO: 181); YGSCSWNYVHIFMDC (SEQ ID NO: 182); YGSCSWNYVHIFLDC (SEQ ID NO: 183); YGSCNWNYVHIFLDC (SEQ ID NO: 184); YTSCNWNYVHIFMDC (SEQ ID NO: 185); YPGCKWNYDRIFLDC (SEQ ID NO: 186); WRSCNWNYAHIFLDC (SEQ ID NO: 187); WSNCHWNYVHIFLDC (SEQ ID NO: 188); DRSCTWNYVRISYDC (SEQ ID NO: 189); SGSCKWDYVHIFLDC (SEQ ID NO: 190); SRSCIWNYAHIHLDC (SEQ ID NO: 191); SMSCYWQYERIFLDC (SEQ ID NO: 192); YRSCNWNYVSIFLDC (SEQ ID NO: 193); YGSCSWNYVHIFMDC (SEQ ID NO: 194); SGSCKWDYVHIFLDC (SEQ ID NO: 195); YKSCHWDYVHIFLDC (SEQ ID NO: 196); YGSCTWNYVHIFMEC (SEQ ID NO: 197); FSSCNWNYVHIFLDC (SEQ ID NO: 198); WRSCNWNYAHIFLDC (SEQ ID NO: 199); YGSCQWNYVHIFLDC (SEQ ID NO: 200); YRSCNWNYVHIFLDC (SEQ ID NO: 201); NMSCHWDYVHIFLDC (SEQ ID NO: 202); FGPCTWNYARISWDC (SEQ ID NO: 203); XXsCXWXYvhIfXdC (SEQ ID NO: 204); MGVPAGCVWNYAHIFMDC (SEQ ID NO: 205); RDTGGQCRWDYVHIFMDC (SEQ ID NO: 206); AGVPAGCTWNYVHIFMEC (SEQ ID NO: 207); VGVPNGCVWNYAHIFMEC (SEQ ID NO: 208); DGGPAGCSWNYVHIFMEC (SEQ ID NO: 209); AVGPAGCWWNYVHIFMEC (SEQ ID NO: 210); CTWNYVHIFMDCGEGEGP (SEQ ID NO: 211); GGVPEGCTWNYAHIFMEC (SEQ ID NO: 212); AEVPAGCWWNYVHIFMEC (SEQ ID NO: 213); AGVPAGCTWNYVHIFMEC (SEQ ID NO: 214); SGASGGCKWNYVHIFMDC (SEQ ID NO: 215); MGVPAGCVWNYAHIFMDC (SEQ ID NO: 216); TPGCRWNYVHIFMECEAL (SEQ ID NO: 217); VGVPNGCVWNYAHIFMEC (SEQ ID NO: 218); PGAFDIPFPAHWVPNT (SEQ ID NO: 219); RGACDIPFPAHWIPNT (SEQ ID NO: 220); QGDFDIPFPAHWVPIT (SEQ ID NO: 221); XGafDIPFPAHWvPnT (SEQ ID NO: 222); RGDGNDSDIPFPAHWVPRT (SEQ ID NO: 223); SGVGRDRDIPFPAHWVPRT (SEQ ID NO: 224); WAGGNDCDIPFPAHWIPNT (SEQ ID NO: 225); WGDGMDVDIPFPAHWVPVT (SEQ ID NO: 226); AGSGNDSDIPFPAHWVPRT (SEQ ID NO: 227); ESRSGYADIPFPAHWVPRT (SEQ ID NO: 228); и/или RECGRCGDIPFPAHWVPRT (SEQ ID NO: 173).
В некоторых вариантах осуществления изобретения, отщепляемая группа (CM) мультиспецифического активируемого антитела включает аминокислотную последовательность, которая может служить в качестве субстрата для протеазы, а обычно, внеклеточной протеазы. CM может быть выбрана в зависимости от типа протеазы, которая локализуется в ткани вместе с нужной мишенью по меньшей мере для одного АВ мультиспецифического активируемого антитела. Существуют различные условия, при которых представляющая интерес мишень локализуется вместе с протеазой, где субстрат для этой протеазы известен специалистам. В случае рака, тканью-мишенью может быть раковая ткань, а в частности, раковая ткань солидной опухоли. В литературе имеются сообщения о повышенных уровнях протеаз, имеющих известные субстраты в ряде раковых опухолей, например, солидных опухолей. См., например, La Rocca et al., (2004) British J. of Cancer 90(7): 1414-1421. Неограничивающими примрами заболеваний являются: раковые заболевания всех типов (рак молочной железы, рак легких, рак прямой и ободочной кишки, рак предстательной железы, меланома, рак головы и шеи, рак поджелудочной железы и т.п.), ревматоидный артрит, болезнь Крона, СКВ, сердечно-сосудистые расстройства, ишемия и т.п. Так, например, показаниями являются лейкоз, включая Т-клеточный острый лимфобластный лейкоз (T-ОЛЛ), лимфобластные заболевания, включая множественную миелому, и солидные опухоли, включая рак легких, рак прямой и ободочной кишки, рак предстательной железы, рак поджелудочной железы и рак молочной железы, включая рак молочной железы, негативный по триаде симптомов. Так, например, показаниями являются раковые заболевания костей или метастазы в кость, независимо от происхождения первичной опухоли; рак молочной железы, включая, но не ограничиваясь ими, например, ER/PR+-рак молочной железы, Her2+-рак молочной железы, рак молочной железы, негативный по триаде симптомов; рак прямой и ободочной кишки, рак эндометрия; рак желудка; глиобластома; рак головы и шеи, такой как рак пищевода; рак легких, такой как, но не ограничивающийся им, например, немелкоклеточный рак легких; множественная миелома; рак яичника; рак поджелудочной железы; рак предстательной железы; саркома, такая как остеосаркома; рак почек, такой как, но не ограничивающийся им, почечно-клеточная карцинома; и/или рак кожи, такой как, но не ограничивающийся ими, плоскоклеточный рак, базальноклеточная карцинома и/или меланома. В некоторых вариантах осуществления изобретения, раковым заболеванием является плоскоклеточный рак. В некоторых вариантах осуществления изобретения, раковым заболеванием является плоскоклеточная карцинома кожи. В некоторых вариантах осуществления изобретения, раковым заболеванием является плоскоклеточная карцинома пищевода. В некоторых вариантах осуществления изобретения, раковым заболеванием является плоскоклеточная карцинома головы и шеи. В некоторых вариантах осуществления изобретения, раковым заболеванием является плоскоклеточная карцинома легких.
CM специфически отщепляется ферментом со скоростью приблизительно 0,001-1500x104 М-1с-1 или по меньшей мере 0,001, 0,005, 0,01, 0,05, 0,1, 0,5, 1, 2,5, 5, 7,5, 10, 15, 20, 25, 50, 75, 100, 125, 150, 200, 250, 500, 750, 1000, 1250 или 1500x104 М-1с-1.
Для специфического расщепления ферментом, этот фермент подвергают контактированию с CM. Если мультиспецифическое активируемое антитело, содержащее по меньшей мере первое АВ, связанное с ММ и СМ, находится в присутствии мишени и достаточного количества фермента, то СМ может отщепляться. Термин «достаточная ферментативная активность» может означать способность фермента контактировать с СМ и отщеплять эту группу. Нетрудно понять, что фермент может присутствовать поблизости от CM, но не отщеплять эту группу, поскольку этот фермент может быть модифицирован другими клеточными факторами или белками.
Репрезентативными субстратами являются, но не ограничиваются ими, субстраты, расщепляемые одним или более из нижеследующих ферментов или протеаз, перечисленных в таблице 3.
Репрезентативные протеазы и/или ферменты
Так, например, в некоторых вариантах осуществления изобретения, субстрат расщепляется одним или более нижеследующими ферментами или протеазами: uPA, легумаином, MT-SP1, ADAM17, BMP-1, TMPRSS3, TMPRSS4, MMP-9, MMP-12, MMP-13 и/или MMP-14. В некоторых вариантах осуществления изобретения, протеазу выбирают из группы, состоящей из uPA, легумаина и MT-SP1. В некоторых вариантах осуществления изобретения, протеазой является металлопротеиназа матрикса. В некоторых вариантах осуществления изобретения, протеазой является uPA. В некоторых вариантах осуществления изобретения, протеазой является легумаин. В некоторых вариантах осуществления изобретения, протеазой является MT-SP1.
В некоторых вариантах осуществления изобретения, CM выбирают для ее использования вместе со специфической протеазой. В некоторых вариантах осуществления изобретения, CM является субстратом по меньшей мере для одной протеазы, выбранной из группы, состоящей из ADAM 17, BMP-1, цистеиновой протеазы, такой как катепсин, HtrA1, легумаина, матриптазы (MT-SP1), металлопротеазы матрикса (MMP), эластазы нейтрофилов, TMPRSS, такой как TMPRSS3 или TMPRSS4, тромбина и активатора плазминогена u-типа (uPA, также называемого урокиназой).
В некоторых вариантах осуществления изобретения, CM является субстратом для ADAM17. В некоторых вариантах осуществления изобретения, CM является субстратом для BMP-1. В некоторых вариантах осуществления изобретения, CM является субстратом для катепсина. В некоторых вариантах осуществления изобретения, CM является субстратом для цистеиновой протеазы. В некоторых вариантах осуществления изобретения, CM является субстратом для HtrA1. В некоторых вариантах осуществления изобретения, CM является субстратом для легумаина. В некоторых вариантах осуществления изобретения, CM является субстратом для MT-SP1. В некоторых вариантах осуществления изобретения, CM является субстратом для MMP. В некоторых вариантах осуществления изобретения, CM является субстратом для эластазы нейтрофилов. В некоторых вариантах осуществления изобретения, CM является субстратом для тромбина. В некоторых вариантах осуществления изобретения, CM является субстратом для TMPRSS. В некоторых вариантах осуществления изобретения, CM является субстратом для TMPRSS3. В некоторых вариантах осуществления изобретения, CM является субстратом для TMPRSS4. В некоторых вариантах осуществления изобретения, CM является субстратом для uPA.
В некоторых вариантах осуществления изобретения, отщепляемую группу выбирают для ее использования вместе со специфической протеазой, например, протеазой, которая, как известно, локализуется вместе с мишенью активируемого антитела. Так, например, подходящие отщепляемые группы, используемые в активируемых антителах согласно изобретению, имеют последовательности TGRGPSWV (SEQ ID NO: 27); SARGPSRW (SEQ ID NO: 28); TARGPSFK (SEQ ID NO: 29); LSGRSDNH (SEQ ID NO: 26); GGWHTGRN (SEQ ID NO: 30); HTGRSGAL (SEQ ID NO: 31); PLTGRSGG (SEQ ID NO: 32); AARGPAIH (SEQ ID NO: 33); RGPAFNPM (SEQ ID NO: 34); SSRGPAYL (SEQ ID NO: 35); RGPATPIM (SEQ ID NO: 36); RGPA (SEQ ID NO: 37); GGQPSGMWGW (SEQ ID NO: 38); FPRPLGITGL (SEQ ID NO: 39); VHMPLGFLGP (SEQ ID NO: 40); SPLTGRSG (SEQ ID NO: 41); SAGFSLPA (SEQ ID NO: 42); LAPLGLQRR (SEQ ID NO: 43); SGGPLGVR (SEQ ID NO: 44); и/или PLGL (SEQ ID NO: 45).
В некоторых вариантах осуществления изобретения, CM является субстратом по меньшей мере для одной металлопротеазы матрикса (MMP). Примерами MMP являются MMP1; MMP2; MMP3; MMP7; MMP8; MMP9; MMP10; MMP11; MMP12; MMP13; MMP14; MMP15; MMP16; MMP17; MMP19; MMP20; MMP23; MMP24; MMP26 и MMP27. В некоторых вариантах осуществления изобретения, CM является субстратом для MMP9, MMP14, MMP1, MMP3, MMP13, MMP17, MMP11 и MMP19. В некоторых вариантах осуществления изобретения, CM является субстратом для MMP7. В некоторых вариантах осуществления изобретения CM является субстратом для MMP9. В некоторых вариантах осуществления изобретения, CM является субстратом для MMP14. В некоторых вариантах осуществления изобретения, CM является субстратом для двух или более MMP. В некоторых вариантах осуществления изобретения, CM является субстратом по меньшей мере для MMP9 и MMP14. В некоторых вариантах осуществления изобретения, CM содержит субстрат для двух или более MMP. В некоторых вариантах осуществления изобретения, CM содержит субстрат по меньшей мере для двух или более MMP9. В некоторых вариантах осуществления изобретения, CM содержит субстрат по меньшей мере для двух или более MMP14.
В некоторых вариантах осуществления изобретения, CM является субстратом для MMP и включает последовательности ISSGLLSS (SEQ ID NO: 316); QNQALRMA (SEQ ID NO: 317); AQNLLGMV (SEQ ID NO: 318); STFPFGMF (SEQ ID NO: 319); PVGYTSSL (SEQ ID NO: 320); DWLYWPGI (SEQ ID NO: 321); MIAPVAYR (SEQ ID NO: 322); RPSPMWAY (SEQ ID NO: 21); WATPRPMR (SEQ ID NO: 323); FRLLDWQW (SEQ ID NO: 324); LKAAPRWA (SEQ ID NO: 325); GPSHLVLT (SEQ ID NO: 326); LPGGLSPW (SEQ ID NO: 327); MGLFSEAG (SEQ ID NO: 328); SPLPLRVP (SEQ ID NO: 329); RMHLRSLG (SEQ ID NO: 330); LAAPLGLL (SEQ ID NO: 331); AVGLLAPP (SEQ ID NO: 332); LLAPSHRA (SEQ ID NO: 333); PAGLWLDP (SEQ ID NO: 334); и/или ISSGLSS (SEQ ID NO: 335).
В некоторых вариантах осуществления изобретения, мультиспецифические антитела и/или мультиспецифические активируемые антитела согласно изобретению могут быть получены путем биосинтеза методами рекомбинантных ДНК и экспрессии в эукариотах или прокариотах. В случае мультиспецифических активируемых антител, кДНК, кодирующие маскирующую группу, линкерная последовательность (которая может включать отщепляемую группу (CM) и цепь антитела (тяжелую или легкую цепь)) может быть присоединена к последовательности в направлении 5’→3’ (от N- до C-конца в транслированном продукте) с образованием конструкции нуклеиновой кислоты, которая экспрессируется как белок мультиспецифического активируемого антитела по стандартному механизму экспрессии антител. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело может быть полусинтетическим и может быть продуцировано посредством экспрессии CM-антитела с последующим химическим присоединением маскирующей группы к N-концу белка или поблизости от N-конца белка. В некоторых вариантах осуществления изобретения, мультиспецифическое активируемое антитело может быть продуцировано посредством экспрессии антитела с последующим химическим присоединением маскирующей группы и СМ к N-концу белка или поблизости от N-конца белка так, чтобы мультиспецифическое активируемое антитело в нерасщепленном состоянии имело следующую структуру, от N-конца до C-конца: MM-CM-AB или AB-CM-MM.
Линкерами, подходящиими для их использования в описанных здесь композициях, являются, в основном, линкеры, сообщающие модифицированному АВ или мультиспецифическим активируемым антителам, гибкость, достаточную для ингибирования связывания по меньшей мере первого АВ с мишенью. Такие линкеры обычно называются гибкими линкерами. Подходящие линкеры могут легко выбраны специалистом и могут иметь любую подходящую длину, например, от 1 аминокислоты (например Gly) до 20 аминокислот, от 2 аминокислот до 15 аминокислот, от 3 аминокислот до 12 аминокислот, включая длину от 4 аминокислот до 10 аминокислот, от 5 аминокислот до 9 аминокислот, от 6 аминокислот до 8 аминокислот, или от 7 аминокислот до 8 аминокислот, и могут иметь длину 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот.
Репрезентативными гибкими линкерами являются полимеры глицина (G)n, полимеры глицина-серина (включая, например, (GS)n, (GSGGS)n (SEQ ID NO: 18) и (GGGS)n (SEQ ID NO: 19), где n равно целому числу по меньшей мере 1), полимеры глицина-аланина, полимеры аланина-серина и другие гибкие линкеры, известные специалистам. Полимеры глицина и полимеры глицина-серина являются относительно неструктурированными, а поэтому они могут служить в качестве нейтральной связи между компонентами. Глицин занимает значительно большее фи-пси-пространство, чем даже аланин, и имеет гораздо меньше структурных ограничений, чем остатки с более длинными боковыми цепями (см. Scheraga, Rev. Computational Chem. 11173-142 (1992)). Репрезентативными гибкими линкерами являются, но не ограничиваются ими, Gly-Gly-Ser-Gly (SEQ ID NO: 20), Gly-Gly-Ser-Gly-Gly (SEQ ID NO: 21), Gly-Ser-Gly-Ser-Gly (SEQ ID NO: 22), Gly-Ser-Gly-Gly-Gly (SEQ ID NO: 23), Gly-Gly-Gly-Ser-Gly (SEQ ID NO: 24), Gly-Ser-Ser-Ser-Gly (SEQ ID NO: 25) и т.п. Для среднего специалиста очевидно, что конструкция активируемых антител может включать линкеры, которые являются полностью или частично гибкими, и такие линкеры могут представлять собой гибкий линкер, а также одну или более его частей, которые придают структуре нужных мультиспецифических активируемых антител меньшую гибкость.
Мультиспецифические активируемые антитела могут содержать, помимо вышеописанных элементов, дополнительные элементы, такие как, например, аминокислотные последовательности, расположенные со стороны N- или C-конца мультиспецифических активируемых антител. Так, например, мультиспецифические активируемые антитела могут включать нацеливающую группу, облегчающую доставку этих антител в представляющие интерес клетки или ткани. Мультиспецифические активируемые антитела могут быть конъюгированы с агентом, таким как терапевтический агент, противоопухолевый агент, токсин или его фрагмент, детектируемая группа или диагностический агент. Примеры таких агентов описаны в настоящей заявке.
Мультиспецифические активируемые антитела могут также включать любые конъюгированные агенты, линкеры и другие описанные здесь компоненты в комбинации с мультиспецифическим антителом согласно изобретению, включая, но не ограничиваясь ими, например, любые агенты, перечисленные в таблице 4 и/или любые линкеры, перечисленные в таблице 5 и/или в таблице 6.
Конъюгированные мультиспецифические антитела и конъюгированные мультиспецифические активируемые антитела
Настоящее изобретение также относится к иммуноконъюгатам, содержащим мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, конъюгированное с цитотоксическим агентом, таким как токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (то есть радиоконъюгат). Подходящими цитотоксическими агентами являются, например, доластатины и его производные (например, ауристатин Е, AFP, MMAD, MMAF, MMAE). Так, например, цитотоксическим агентом является монометилауристатин E (MMAE). В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин D (MMAD). В некоторых вариантах осуществления изобретения, указанным агентом является агент, выбранные из группы агентов, перечисленных в таблице 4. В некоторых вариантах осуществления изобретения, указанным агентом является доластатин. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин Е или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин E (MMAE). В некоторых вариантах осуществления изобретения, указанным агентом является майтанзиноид или производное майтанзиноида. В некоторых вариантах осуществления изобретения, указанным агентом является DM1 или DM4. В некоторых вариантах осуществления изобретения, указанным агентом является дуокармицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является калихеамицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является пирролобензодиазепин.
Ферментативно активными токсинами и их фрагментами, которые могут быть использованы, являются А-цепь дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, А-цепь экзотоксина (от Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки фитолакки американской (Phytolaca americana) (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Для продуцирования радиоактивных конъюгированных антител могут быть использованы различные радионуклиды. В качестве примеров могут служить 212Bi, 64Cu, 125I, 131I, 131In, 99mTc, 90Y, 186Re и 89Zr.
Конъюгаты антитела и цитотоксического агента получают с использованием различных бифункциональных белок-связывающих агентов, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат-HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азодобензоил) гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Так, например, иммунотоксин рицин может быть получен как описано в публикации Vitetta et al., Science 238: 1098 (1987). Репрезентативным хелатообразующим агентом, используемым для конъюгирования радионуклида с антителом, является 14С-меченная 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) (см. WO94/11026).
В таблице 4 перечислены некоторые репрезентативные фармацевтические агенты, которые могут быть использованы в настоящем изобретении, но этот список не является исчерпывающим.
Репрезентативные фармацевтические агенты для конъюгирования
Специалистам в данной области известно, что к полученным мультиспецифическим антителам и/или мультиспецифическим активируемым антителам согласно изобретению может быть присоединено большое число различных молекул (см., например, публикацию «Conjugate Vaccines», Contributions to Microbiology and Immunology, J. M. Cruse and R. E. Lewis, Jr (eds), Carger Press, New York, (1989), содержание которой во всей своей полноте вводится в настоящее описание посредством ссылки).
Присоединение может быть осуществлено посредством любой химической реакции связывания двух молекул, при условии, что антитело и другая молекула будут сохранять соответствующую активность. Такое присоединение может быть осуществлено по многим химическим механизмам, таким как, например, ковалентное связывание, аффинное связывание, интеркалирование, координационное связывание и комплексообразование. Однако в некоторых вариантах осуществления изобретения предпочтительным связыванием является ковалентное связывание. Ковалентное связывание может быть осуществлено либо посредством прямой конденсации имеющихся боковых цепей, либо посредством включения внешних мостиковых молекул. Многие двухвалентные или поливалентные линкерные агенты могут быть использованы для связывания белковых молекул, таких как антитела согласно изобретению, с другими молекулами. Так, например, репрезентативными связывающими агентами могут быть органические соединения, такие как сложные тиоэфиры, карбодиимиды, сукцинимидоэфиры, диизоцианаты, глутаральдегид, диазобензолы и гексаметилендиамины. Этот список не является исчерпывающим и не охватывает все известные связывающие агенты различных классов, а всего лишь включает наиболее распространенные связывающие агенты (см. Killen and Lindstrom, Jour. Immun. 133:1335-2549 (1984); Jansen et al., Immunological Reviews 62:185-216 (1982); и Vitetta et al., Science 238:1098 (1987).
В некоторых своих вариантах, настоящее изобретение, помимо описанных здесь композиций и способов, относится к конъюгированному активируемому антителу, которое может быть также модифицировано для сайт-специфического конъюгирования посредством модифицированных аминокислотных последовательностей, встроенных или как-либо иначе введенных в последовательность активируемого антитела. Эти модифицированные аминокислотные последовательности конструируют так, чтобы можно было регулировать место введения и/или количество конъюгированого агента в конъюгированном активируемом антителе. Так, например, активируемое антитело может быть сконструировано так, чтобы оно имело цистеиновые замены в положениях легкой и тяжелой цепей, а именно, реакционноспособные тиоловые группы, и чтобы такие замены не оказывали негативного влияния на укладку и сборку белка и на связывание с антигеном. В некоторых вариантах осуществления изобретения, активируемое антитело может быть сконструировано так, чтобы оно включало или как-либо иначе приобретало один или более не-природных аминокислотных остатков, обеспечивающих подходящие сайты конъюгирования. В некоторых вариантах осуществления изобретения, активируемое антитело может быть сконструировано так, чтобы в его последовательность были включены или как-либо иначе введены ферментативно активируемые пептидные последовательности.
Подходящие линкеры описаны в литературе (см., например, публикацию Ramakrishnan, S. et al., Cancer Res. 44:201-208 (1984), где описано использование MBS (M-малеимидобензоил-N-гидроксисукцинимидоэфира). См. также патент США No. 5030719, в котором описано использование галогенированного производного ацетилгидразида, связанного с антителом посредством олигопептидного линкера. Особенно подходящими линкерами являются: (i) SMPT (4-сукцинимидилоксикарбонил-альфа-метил-альфа-(2-пиридилдитио)толуол (Pierce Chem. Co., Cat. (21558G); (ii) SPDP (сукцинимидил-6-[3-(2-пиридилдитио)пропионамидо]гексаноат (Pierce Chem. Co., Cat #21651G); и (iii) сульфо-LC-SPDP (сульфосукцинимидил-6-[3-(2-пиридилдитио)пропионамид]гексаноат (Pierce Chem. Co. Cat. #2165-G). Дополнительными линкерами являются, но не ограничиваются ими, SMCC, сульфо-SMCC, SPDB или сульфо-SPDB.
Вышеописанные линкеры содержат компоненты, имеющие различные характерные признаки, что позволяет получать конъюгаты с различными физико-химическими свойствами. Так, например, линкер SMPT содержит стерически затрудненную дисульфидную связь и может образовывать конъюгаты с повышенной стабильностью. Дисульфидные связи являются, в основном, менее стабильными, чем другие связи, поскольку дисульфидная связь расщепляется in vitro, что приводит к образованию менее доступного конъюгата.
Реагент EDC (гидрохлорид 1-этил-3-(3-диметиламинопропил)карбодиимида) может быть применен для получения карбоксамида с использованием карбоновой кислоты и первичного или вторичного амина в качестве исходных соединений. Таким образом, EDC может быть использован для присоединения лизиновых остатков в антителе посредством карбоновой кислоты, присутствующей в линкере или токсине, или для присоединения аспартатных или глутаматных остатков в антителе посредством амина, присутствующего в линкере или токсине. Скорость таких реакций конъюгирования, проводимых с использованием EDC, может быть увеличена путем добавления NHS (N-гидроксисукцинимида) или сульфо-NHS (N-гидрокси-3-оксисульфонилсукцинимида). Добавление NHS или сульфо-NHS в такие реакционные смеси для конъюгирования может приводить к увеличению скорости, выхода, селективности и/или репродуцируемости реакций конъюгирования.
В некоторых вариантах осуществления изобретения, линкеры являются расщепляемыми. В некоторых вариантах осуществления изобретения, линкеры являются нерасщепляемыми. В некоторых вариантах осуществления изобретения используются два или более линкеров. Эти два или более линкеров могут быть одинаковыми, например, расщепляемыми или нерасщепляемыми, либо они могут быть различными, например, по меньшей мере один линкер может быть расщепляемым, а другой нерасщепляемым.
Настоящее изобретение относится к некоторым способам присоединения агентов к АВ мультиспецифических антител и/или мультиспецифических активируемых антител, где указанные способы включают: (a) присоединение к углеводным группам AB, или (b) присоединение к сульфгидрильным группам AB, или (c) присоединение к аминогруппам AB, или (d) присоединение к карбоксилатным группам AB. В соответствии с настоящим изобретением, AB могут быть ковалентно связаны с агентом посредством промежуточного линкера, имеющего по меньшей мере две реакционноспособных группы, одна из которых реагирует с АВ, а другая реагирует с агентом. Линкер, который может включать любое совместимое органическое соединение, может быть выбран так, чтобы реакция взаимодействия с АВ (или агентом) не оказывала какого-либо негативного воздействия на реактивность и селективность АВ. Кроме того, связывание линкера с агентом может не оказывать негативного влияния на активность агента. Подходящими линкерами, используемыми для проведения реакции с окисленными антителами или окисленными фрагментами антител, являются линкеры, содержащие амин, выбранный из группы, состоящей из первичного амина, вторичного амина, гидразина, гидразида, гидроксиламина, фенилгидразина, полукарбазида и тиосемикарбазида. Такие реакционноспособные функциональные группы могут присутствовать как часть структуры линкера, либо они могут быть введены подходящими методами химической модификации линкеров, не содержащих таких групп.
В соответствии с настоящим изобретением, подходящие линкеры, используемые для связывания с восстановленными АВ мультиспецифических антител и/или мультиспецифических активируемых антител, включают некоторые реакционноспособные группы, способные реагировать с сульфгидрильной группой восстановленного антитела или его фрагмента. Такими реакционноспособными группами являются, но не ограничиваются ими: реакционноспособные галогеналкильные группы (включая, например, галогенацетильные группы), группы п-бензоата ртути и группы, подходящие для проведения реакций присоединения по типу реакции Михаэля (включая, например, малеимиды и группы, описанные в публикации Mitra and Lawton, 1979, J. Amer. Chem. Soc. 101: 3097-3110).
В соответствии с настоящим изобретением, подходящие линкеры, используемые для присоединения к неокисленным и невосстановленным АВ мультиспецифических антител и/или мультиспецифических активируемых антител, включают некоторые функциональные группы, способные реагировать с первичными аминогруппами, присутствующими в немодифицированных лизиновых остатках в АВ. Такми реакционноспособными группами являются, но не ограничиваются ими: NHS-эфиры карбоновой или угольной кислоты, сульфо-NHS-эфиры карбоновой или угольной кислоты, 4-нитрофениловые эфиры карбоновой или угольной кислоты, пентафторфениловые эфиры карбоновой или угольной кислоты, ацилимидазолы, изоцианаты и изотиоцианаты.
В соответствии с настоящим изобретением, подходящие линкеры, используемые для присоединения к неокисленным и невосстановленным АВ, включают некоторые функциональные группы, способные реагировать с группами карбоновой кислоты, присутствующими в аспартатных или глутаматных остатках в АВ, которые были активированы подходящими реагентами. Подходящими активирующими реагентами являются EDC, содержащие или не содержащие NHS или сульфо-NHS, и другие дегидратирующие агенты, используемые для образования карбоксамида. В этих случаях, функциональными группами, присутствующими в подходящих линкерах, являются первичные и вторичные амины, гидразины, гидроксиламины и гидразиды.
Агент может быть присоединен к линкеру до или после присоединения линкера к АВ. В некоторых случаях может оказаться желательным сначала получение промежуточного АВ-линкера, где указанный линкер не содержит связанного с ним агента. В зависимости от конкретных целей, специфический агент может быть затем ковалентно присоединен к данному линкеру. В других вариантах осуществления изобретения, AB сначала присоединяют к MM, CM и к связанным с ними линкерам, а затем его присоединяют к линкеру для конъюгирования.
Разветвленные линкеры: В конкретных вариантах осуществления изобретения используют разветвленные линкеры, имеющие множество сайтов для присоединения агентов. В случае многосайтовых линкеров, одно ковалентное связывание с AB может приводить к образованию промежуточного AB-линкера, способного связываться с агентом в различных сайтах. Такими сайтами могут быть альдегидные или сульфгидрильные группы или любой химический сайт, к которому могут быть присоединены агенты.
Альтернативно, более высокая удельная активность (или более высокое отношение агентов к АВ) может быть достигнута путем присоединения односайтового линкера во множество сайтов на АВ. Такое множество сайтов может быть введено в АВ любыми двумя методами. В первом методе, может быть создано множество альдегидных групп и/или сульфгидрильных групп на одном и том же АВ. Во втором методе, к альдегиду или сульфгидрилу АВ может быть присоединен «разветвленный линкер», имеющий множество функциональных сайтов для последующего присоединения к линкерам. Функциональными сайтами разветвленного линкера или многосайтового линкера могут быть альдегидные или сульфгидрильные группы, либо любой химический сайт, к которому могут быть присоединены линкеры. Еще более высокие удельные активности могут быть достигнуты путем объединения этих двух методов, то есть, путем присоединения многосайтовых линкеров в несколько сайтов на АВ.
Расщепляемые линкеры: В одном из вариантов осуществления изобретения могут быть использованы пептидные линкеры, которые могут расщепляться под действием ферментов системы комплемента, таких как, но не ограничивающихся ими, урокиназа, тканевый активатор плазминогена, трипсин, плазмин или другой фермент, обладающий протеолитической активностью. В соответствии с одним из способов согласно изобретению, агент присоединяют посредством линкера, который может расщепляться под действием комплемента. Антитело выбирают из антител определенного класса, которые могут активировать комплемент. Таким образом, конъюгат «антитело- агент» активирует каскад реакции комплемента и высвобождает агент в сайт-мишень. В соответствии с другим способом согласно изобретению, агент присоединяют посредством линкера, который может расщепляться под действием ферментов, обладающих протеолитической активностью, таких как урокиназа, тканевый активатор плазминогена, плазмин или трипсин. Эти расщепляемые линкеры могут быть использованы в конъюгированных активируемых антителах, которые включают внеклеточный токсин, неограничивающим примером которого являются любые внеклеточные токсины, представленные в таблице 4.
Неограничивающие примеры последовательностей расщепляемых линкеров представлены в таблице 5.
Репрезентативные линкерные последовательности для конъюгирования
(цепь α1(I))
(цепь α2(I)))
(цепь α1(II))
печени (цепь α1(III))
Кроме того, агенты могут быть присоединены к АВ посредством дисульфидных связей (например, дисульфидных связей на молекуле цистеина). Поскольку многие опухоли по своей природе высвобождают высокие уровни глутатиона (восстановителя), то это может приводить к восстановлению дисульфидных связей и к последующему высвобождению агента в область доставки. В некоторых конкретных вариантах осуществления изобретения, восстановитель, который может модифицировать CM, может также модифицировать и линкер конъюгированного активируемого антитела.
Спейсерные элементы и расщепляемые элементы: В некоторых вариантах осуществления изобретения может оказаться необходимым сконструировать линкер для оптимизации пространства между агентом и АВ активируемого антитела. Это может быть осуществлено с использованием линкера, имеющего общую структурную формулу:
W – (CH2)n – Q,
где:
W представляет собой --NH--CH2-- или --CH2--;
Q представляет собой аминокислоту, пептид; и
n равно целому числу от 0 до 20.
В других вариантах осуществления изобретения, линкер может содержать спейсерный элемент и расщепляемый элемент. Спейсерный элемент служит для введения расщепляемого элемента в положение, удаленное от сердцевины AB так, чтобы расщепляемый элемент был более доступным для фермента, ответственного за расщепление. Некоторые вышеописанные разветвленные линкеры могут служит в качестве спейсерных элементов.
Как обсуждалось в настоящей заявке, очевидно, что присоединение линкера к агенту (или присоединение спейсерного элемента к расщепляемому элементу или расщепляемого элемента к агенту) необязательно должно быть осуществлено каким-либо конкретным методом присоединения или посредством какой-либо конкретной реакции. При этом может быть осуществлена любая реакция, в результате которой образуется продукт, имеющий подходящую стабильность и биологическую совместимость.
Сывороточный комплемент и отбор линкеров: В соответствии с одним из способов согласно изобретению, если желательно высвобождение агента, то используется АВ, которое представляет собой антитело определенного класса, способное активировать комплемент. Полученный конъюгат сохраняет способность связываться с антигеном и активировать каскад комплемента. Таким образом, в соответствии с этим вариантом осуществления изобретения, агент присоединяют к одному концу расщепляемого линкера или расщепляемого элемента, а другой конец линкерной группы присоединяют к специфическому сайту на АВ. Так, например, если агент имеет гидроксигруппу или аминогурппу, то этот агент может быть присоединен к карбокси-концу пептида, аминокислоты или другого соответствующим образом выбранного линкера посредством сложного эфира или амидной связи, соответственно. Так, например, такие агенты могут быть присоединены к линкерному пептиду посредством карбодиимидной реакции. Если агент содержит функциональную группу, негативно влияющую на присоединение агента к линкеру, то такие нежелательные функциональные группы могут быть блокированы до присоединения, а затем с них может быть снята защита после получения конъюгата или промежуточного продукта. Противоположный конец или амино-конец линкера затем используют непосредственно для связывания с АВ или после модификации, проводимой для связывания с АВ, которое может активировать комплемент.
Линкеры (или спейсерные элементы линкеров) могут иметь любую желаемую длину, и один конец этого линкера может быть ковалентно присоединен к специфическим сайтам на АВ активируемого антитела. Другой конец линкера или спейсерного элемента может быть присоединен к аминокислотному или пептидному линкеру.
Таким образом, если эти конъюгаты связываются с антигеном в присутствии комплемента, то амидная или эфирная связь, посредством которой агент связывается с линкером, может расщепляться, что будет приводить к высвобождению агента в активной форме. Эти конъюгаты, при их введении индивидууму, будут доставлять и высвобождать агент в сайт-мишень, а поэтому они являются особенно эффективными для in vivo доставки фармацевтических агентов, антибиотиков, антиметаболитов, антипролиферирующих агентов и т.п., неограничивающие примеры которых представлены в таблице 4.
Линкеры для высвобождения без активации комплемента: При другом применении целевой доставки может оказаться желательным высвобождение агента без активации комплемента, поскольку активация каскада комплемента будет, в конечном счете, приводить к лизису клеток-мишеней. Следовательно, такой подход является полезным в том случае, если доставка и высвобождение агента должны быть осуществлены без уничтожения клеток-мишеней. Такая цель может быть поставлена, если желательна доставка клеточных медиаторов, таких как гормоны, ферменты, кортикостероиды, нейромедиаторы, гены или ферменты в клетки-мишени. Указанные конъюгаты могут быть получены путем присоединения к АВ агента, который не способен активировать комплемент посредством линкера, плохо поддающегося расщеплению сывороточными протеазами. При введении этого конъюгата индивидууму происходит быстрое образование комплексов «антиген-антитело», а расщепление агента осуществляется медленно, что приводит к высвобождению соединения в сайт-мишень.
Биохимические перекрестносшивающие линкеры: В других вариантах осуществления изобретения, активируемое антитело может быть конъюгировано с одним или более терапевтическими агентами посредством некоторых биохимических перекрестносшивающих линкеров. Перекрестносшивающие реагенты образуют молекулярные мостики, которые связывают функциональные группы двух различных молекул. Для постадийного связывания двух различных белков могут быть использованы гетеробифункциональные перекрестносшивающие линкеры, препятствующие образованию нежелательных гомополимеров.
Могут быть также использованы пептидильные линкеры, например, Val-Cit, Val-Ala или другие дипептиды, расщепляемые лизосомными протеазами. Кроме того, могут быть использованы линкеры, чувствительные к действию кислот, например, бис-сиалилэфирные линкеры, расщепляемые лизосомами при низком pH. Другими подходящими линкерами являются субстраты, чувствительные к действию катепсина, а в частности, линкеры, имеющие оптимальную функцию при кислотном рН.
Репрезентативные примеры гетеробифункциональных перекрестносшивающих линкеров представлены в таблице 6.
Репрезентативные гетеробифункциональные перекрестносшивающие линкеры 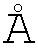
Нерасщепляемые линкеры или прямое присоединение: В других вариантах осуществления изобретения, конъюгат может быть сконструирован так, чтобы агент доставлялся к мишени, но не высвобождался. Это может быть осуществлено путем присоединения агента к АВ либо непосредственно, либо посредством нерасщепляемого линкера.
Такие нерасщепляемые линкеры могут включать аминокислоты, пептиды, D-аминокислоты или другие органические соединения, которые могут быть модифицированы путем введения в них функциональных групп, используемых для последующего связывания с АВ описанными здесь методами. Такой органический линкер может иметь следующую общую формулу:
W – (CH2)n – Q,
где:
W представляет собой --NH--CH2-- или --CH2--;
Q представляет собой аминокислоту, пептид; и
n равно целому числу от 0 до 20.
Нерасщепляемые конъюгаты: Альтернативно, соединение может быть присоединено к АВ, которое не активирует комплемент. При использовании АВ, которые не обладают способностью активировать комплемент, такое присоединение может быть осуществлено с использованием линкеров, чувствительных к расщеплению активированным комплементом, или с использованием линкеров, не чувствительных к расщеплению активированным комплементом.
Описанные здесь антитела могут быть также приготовлены в виде иммунолипосом. Липосомы, содержащие антитело, получают известными методами, например, методами, описанными в публикациях Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al., Proc. Natl Acad. Sci. USA, 77: 4030 (1980); и в патентах США NN. 4485045 и 4544545. Липосомы, имеющие более длительное время полужизни в кровотоке, описаны в патенте США No. 5013556.
Особенно подходящие липосомы могут быть получены методом обращенно-фазового выпаривания из липидной композиции, содержащей фосфотидилхолин, холестерин и ПЭГ-дериватизированный фосфатидилэтаноламин (ПЭГ-ФЭ). Липосомы экструдируют через фильтры с определенным размером пор, в результате чего получают липосомы нужного диаметра. Fab’-фрагмент антитела согласно изобретению могут быть конъюгирован с липосомами, как описано в публикации Martin et al., J. Biol. Chem., 257: 286-288 (1982), посредством реакции дисульфидного обмена.
Мультиспецифические активируемые антитела, содержащие несвязывающиеся стерические группы или партнеры по связыванию с несвязывающимися стерическими группами
Настоящее изобретение также относится к мультиспецифическим активируемым антителам, которые включают несвязывающиеся стерические группы (NB) или партнеры по связыванию (BP) с несвязывающимися стерическими группами, где BP стимулируют рекрутинг или как-либо иначе притагивают NB к мультиспецифическому активируемому антителу. Описанными здесь мультиспецифическими активируемыми антителами являются, например, мультиспецифическое активируемое антитело, которое включает несвязывающуюся стерическую группу (NB), расщепляемый линкер (CL) и по меньшей мере первое антитело или его фрагмент (AB1), которые связываются с первой мишенью или с первым эпитопом; мультиспецифическое активируемое антитело, которое включает партнера по связыванию с несвязывающейся стерической группой (BP), CL и AB1; и мультиспецифическое активируемое антитело, включающее BP, к которому притягивается NB; CL и AB1, которые связываются с первой мишенью или первым эпитопом. Мультиспецифические активируемые антитела, в которых NB ковалентно связана с CL и AB1 или ассоциирована посредством взаимодействия с BP, который ковалентно связан с CL и AB1, называются здесь «NB-содержащими мультиспецифическими активируемыми антителами». Термин «активируемое или переключаемое связывание» означает, что активируемое антитело находится на первом уровне связывания с мишенью, если оно присутствует в ингибированном, маскированном или нерасщепленном состоянии (то есть, в первой конформации), и на втором уровне связывания с мишенью, если оно присутствует в неингибированном, немаскированном и/или отщепленном состоянии (то есть, во второй конформации, то есть в активированной форме), где второй уровень связывания с мишенью превышает первый уровень связывания. Композиции мультиспецифических активируемых антител могут иметь повышенную биологическую доступность и более благоприятное биологическое распределение по сравнению со стандартными терапевтическими антителами.
В некоторых вариантах осуществления изобретения, мультиспецифические активируемые антитела обладают пониженной токсичностью и/или дают меньше побочных эффектов, чем это происходит при связывании мультиспецифического активируемого антитела в непораженных участках и/или в недиагностируемых участках, если связывание мультиспецифического активируемого антитела с мишенью в этом участке не маскируется или как-либо иначе не ингибируется.
В одном из вариантов осуществления изобретения, мультиспецифическое активируемое антитело включает несвязывающуюся стерическую группу (NB), расщепляемый линкер (CL) и по меньшей мере первое антитело или его фрагмент (AB1), которые специфически связываются с первой мишенью или с первым эпитопом, где NB представляет собой полипептид, который специфически не связывается с AB1; CL представляет собой полипептид, который включает субстрат (S) для фермента; CL расположен так, что, если он находится в нерасщепленном состоянии, то NB блокирует связывание АВ1 с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ1 с мишенью, а NB не ингибирует расщепление CL ферментом. Используемый здесь термин «полипептид» означает любой полипептид, имеющий по меньшей мере два аминокислотных остатка, включая более крупные полипептиды, полноразмерные белки и их фрагменты, причем, в понятие «полипептид» входят не только одноцепочечные полипептиды, но также и полипептиды, имеющие множество звеньев, например, множество цепей. В тех случаях, когда полипептид имеет более короткую длину, например, всего менее 50 аминокислот, то используемые здесь термины «пептид» и «полипептид» являются синонимами, а в тех случаях, когда полипептид является более длинным, например, имеет длину 50 аминокислот или более, то используемые здесь термины «полипептид» и «белок» являются синонимами.
В одном из вариантов осуществления изобретения, мультиспецифическое активируемое антитело включает несвязывающуюся стерическую группу (NB); расщепляемый линкер (CL) и по меньшей мере первое антитело или его фрагмент (AB1), которые специфически связываются с первой мишенью или с первым эпитопом, где (i) NB представляет собой полипептид, который специфически не связывается с AB1; (ii) CL представляет собой полипептид, который имеет длину до 50 аминокислот и включает субстрат (S) для фермента; (iii) CL расположен так, что, если он находится в нерасщепленном состоянии, то NB блокирует связывание АВ1 с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ1 с мишенью; и (iv) NB не ингибирует расщепление CL ферментом. Так, например, CL имеет длину до 15 аминокислот, до 20 аминокислот, до 25 аминокислот, до 30 аминокислот, до 35 аминокислот, до 40 аминокислот, до 45 аминокислот, до 50 аминокислот, в пределах 10-50 аминокислот, в пределах 15-50 аминокислот, в пределах 20-50 аминокислот, в пределах 25-50 аминокислот, в пределах 30-50 аминокислот, в пределах 35-50 аминокислот, в пределах 40-50 аминокислот, в пределах 45-50 аминокислот, в пределах 10-40 аминокислот, в пределах 15-40 аминокислот, в пределах 20-40 аминокислот, в пределах 25-40 аминокислот, в пределах 30-40 аминокислот, в пределах 35-40 аминокислот, в пределах 10-30 аминокислот, в пределах 15-30 аминокислот, в пределах 20-30 аминокислот, в пределах 25-30 аминокислот, в пределах 10-20 аминокислот или в пределах 10-15 аминокислот.
В одном из вариантов осуществления изобретения, мультиспецифическое активируемое антитело включает несвязывающуюся стерическую группу (NB); расщепляемый линкер (CL) и по меньшей мере первое антитело или его фрагмент (AB1), которые специфически связываются с первой мишенью или с первым эпитопом, где (i) NB представляет собой полипептид, который специфически не связывается с AB1; (ii) CL представляет собой полипептид, который включает субстрат (S) для фермента; (iii) CL расположен так, что, если он находится в нерасщепленном состоянии, то NB блокирует связывание АВ1 с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ1 с мишенью; (iv) NB не ингибирует расщепление CL ферментом; и (v) по меньшей мере часть мультиспецифического активируемого антитела, если она находится в нерасщепленном состоянии, имеет следующую структуру, от N-конца до C-конца: NB-CL-AB1 или AB1-CL-NB.
В одном из вариантов осуществления изобретения, мультиспецифическое активируемое антитело включает несвязывающуюся стерическую группу (NB); расщепляемый линкер (CL) и по меньшей мере первое антитело или его фрагмент (AB1), которые специфически связываются с первой мишенью или с первым эпитопом, где (i) NB представляет собой полипептид, который специфически не связывается с AB1; (ii) CL представляет собой полипептид, который включает субстрат (S) для фермента; (iii) CL расположен так, что, если он находится в нерасщепленном состоянии, то NB блокирует связывание АВ1 с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ1 с мишенью; и где NB в нерасшепленном активируемом антителе снижает способность АВ1 связываться с мишенью по меньшей мере на 50%, например, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или по меньшей мере на 100% по сравнению со способностью расщепленного AB1 связываться с мишенью; и (iv) NB не ингибирует расщепление CL ферментом. Снижение способности AB связываться с мишенью определяют, например, с помощью анализа, описанного в настоящей заявке, или in vitro анализа на смещение мишени, например, анализа, описанного в публикациях заявок PCT WO 2009/025846 и WO 2010/081173.
В одном из вариантов осуществления изобретения, мультиспецифическое активируемое антитело включает партнера по связыванию (ВР) с несвязывающейся стерической группой (NB); расщепляемый линкер (CL) и по меньшей мере первое антитело или его фрагмент (AB1), которые специфически связываются с первой мишенью и/или с первым эпитопом, где ВР представляет собой полипептид, который связывается с NB при его обработке этой группой; NB специфически не связывается с AB1; CL представляет собой полипептид, который включает субстрат (S) для фермента; CL расположен так, что, если он находится в нерасщепленном состоянии в присутствии NB, то NB блокирует связывание АВ1 с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ1 с мишенью, а ВР не блокирует связывание АВ с мишенью; и NB и ВР не ингибируют расщепление CL ферментом. В некоторых примерах этого варианта, BP активируемого антитела связывается, но необязательно, с NB. В одном из вариантов осуществления изобретения, BP активируемого антитела осуществляет рекрутинг NB in vivo.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело приготавливают в виде композиции. В некоторых этих вариантах, композиция также включает NB, которую вводят в мультиспецифическое активируемое антитело, включающее ВР, CL и АВ. В некоторых примерах этого варианта, BP выбирают из группы, состоящей из альбумин-связывающего пептида, фибриноген-связывающего пептида, фибронектин-связывающего пептида, гемоглобин-связывающего пептида, трансферрин-связывающего пептида, пептида, связывающегося с доменом иммуноглобулина, и пептидов, связывающихся с другими сывороточными белками.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, NB представляет собой растворимый глобулярный белок. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, NB представляет собой белок, который циркулирует в кровотоке. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, NB выбирают из группы, состоящей из альбумина, фибриногена, фибронектина, гемоглобина, трансферрина, домена иммуноглобулина и других сывороточных белков.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, CL представляет собой полипептид, который включает субстрат (S) для протеазы. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, протеаза локализуется вместе с мишенью в ткани и расщепляет CL в мультиспецифическом активируемом антителе при обработке этого антитела протеазой. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, CL представляет собой полипептид длиной до 50 аминокислот. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, CL представляет собой полипептид, который включает субстрат (S), имеющий длину до 15 аминокислот, например, до 3 аминокислот, до 4 аминокислот, до 5 аминокислот, до 6 аминокислот, до 7 аминокислот, до 8 аминокислот, до аминокислот, до 10 аминокислот, до 11 аминокислот, до 12 аминокислот, до 13 аминокислот, до 14 аминокислот или до 15 аминокислот.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, по меньшей мере часть мультиспецифического активируемого антитела в нерасщепленном состоянии имеет следующую структуру, от N-конца до C-конца: NB-CL-AB, AB-CL-NB, BP-CL-AB или AB-CL-BP. В тех вариантах, в которых мультиспецифическое активируемое антитело включает BP и находится в присутствии соответствующего NB, по меньшей мере часть мультиспецифического активируемого антитела в нерасщепленном состоянии имеет следующую структуру, от N-конца до C-конца: NB:BP-CM-AB или AB-CM-BP:NB, где «:» означает взаимодействие, например, связывание NB и BP.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело включает антитело или его антигенсвязывающий фрагмент, которые специфически связываются с мишенью, и таким антителом является моноклональное антитело, однодоменное антитело, одноцепочечное антитело, Fab-фрагмент, F(ab')2-фрагмент, scFv, scab, dAb, однодоменное антитело с тяжелой цепью и однодоменное антитело с легкой цепью. В некоторых вариантах осуществления изобретения, такое антитело или его иммунологически активный фрагмент, которые связываются с мишенью, представляют собой мышиное, химерное, гуманизованное или полностью человеческое моноклональное антитело.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело также включает агент, конъюгированный с АВ. В некоторых вариантах осуществления изобретения, указанным агентом является терапевтический агент. В некоторых вариантах осуществления изобретения, указанным агентом является противоопухолевый агент. В некоторых вариантах осуществления изобретения, указанным агентом является токсин или его фрагмент. В некоторых вариантах осуществления изобретения, указанный агент конъюгирован с АВ посредством линкера. В некоторых вариантах осуществления изобретения, указанным линкером является расщепляемй линкер. В некоторых вариантах осуществления изобретения, указанным агентом является агент, выбранный из группы, состоящей из агентов, перечисоленных в таблице 4. В некоторых вариантах осуществления изобретения, указанным агентом является доластатин. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин E или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин E (MMAE). В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин D (MMAD). В некоторых вариантах осуществления изобретения, указанным агентом является майтанзиноид или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является DM1 или DM4. В некоторых вариантах осуществления изобретения, указанным агентом является дуокармицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является калихеамицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является пирролобензодиазепин.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело также включает детектируемую группу. В некоторых вариантах осуществления изобретения, указанной детектируемой группой является диагностический агент.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело также включает спейсер. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело также включает сигнальный пептид. В некоторых вариантах осуществления изобретения, указанный сигнальный пептид конъюгирован с мультиспецифическим активируемым антителом посредством спейсера. В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, спейсер непосредственно присоединен к MM мультиспецифического активируемого антитела.
В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке превышает время полужизни соответствующего мультиспецифического антитела, например, pK мультиспецифического активируемого антитела в сыворотке превышает pK соответствующего мультиспецифического антитела. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке аналогично времени полужизни соответствующего мультиспецифического антитела. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 15 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 12 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 11 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 10 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 9 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 8 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 7 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 6 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 5 дней. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 4 дня. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 3 дня. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 2 дня. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 24 часа. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 20 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 18 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 16 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 14 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 12 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 10 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 8 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 6 часов. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 4 часа. В некоторых вариантах осуществления изобретения, время полужизни мультиспецифического активируемого антитела в сыворотке после его введения в организм составляет по меньшей мере 3 часа.
Настоящее изобретение также относится к выделенной молекуле нуклеиновой кислоты, кодирующей любое из указанных мультиспецифических активируемых антител, а также к векторам, содержащим эти выделенные последовательности нуклеиновой кислоты. Настоящее изобретение относится к способам продуцирования мультиспецифического активируемого антитела путем культивирования клетки в условиях, стимулирующих экспрессию мультиспецифического активируемого антитела, где указанная клетка содержит указанную молекулу нуклеиновой кислоты. В некоторых вариантах осуществления изобретения, указанная клетка содержит этот вектор.
Константа диссоциации (Kd) при связывании NB-содержащего мультиспецифического активируемого антитела с мишенью превышает Kd при связывании AB с мишенью, если АВ не ассоциируется с NB или NB:BP. Константа диссоциации (Kd) при связывании NB-содержащего мультиспецифического активируемого антитела с мишенью превышает Kd при связывании родительского AB с мишенью. Так, например, Kd при связывании NB-содержащего мультиспецифического активируемого антитела с мишенью по меньшей мере в 5, 10, 20, 25, 40, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 50000, 100000, 500000, 1000000, 5000000, 10000000, 50000000 или более или в пределах 5-10, 10-100, 10-1000, 10-10000, 10-100000, 10-1000000, 10-10000000, 100-1000, 100-10000, 100-100000, 100-1000000, 100-10000000, 1000-10000, 1000-100000, 1000-1000000, 1000-10000000, 10000-100000, 10000-1000000, 10000-10000000, 100000-1000000 или 100000-10000000 раз превышает Kd при связывании AB, не ассоциированного с NB или NB:BP, или Kd при связывании родительского АВ с мишенью. И наоборот, аффинность связывания NB-содержащего мультиспецифического активируемого антитела с мишенью ниже, чем аффинность связывания AB, не ассоциированного с NB или NB:BP, или ниже, чем аффинность связывания родительского АВ с мишенью. Так, например, аффинность связывания NB-содержащего мультиспецифического активируемого антитела с мишенью по меньшей мере в 5, 10, 20, 25, 40, 50, 100, 250, 500, 1000, 2500, 5000, 10000, 50000, 100000, 500000, 1000000, 5000000, 10000000, 50000000 или более или в пределах 5-10, 10-100, 10-1000, 10-10000, 10-100000, 10-1000000, 10-10000000, 100-1000, 100-10000, 100-100000, 100-1000000, 100-10000000, 1000-10000, 1000-100000, 1000-1000000, 1000-10000000, 10000-100000, 10000-1000000, 10000-10000000, 100000-1000000 или 100000-10000000 раз ниже, чем аффинность связывания AB, не ассоциированного с NB или NB:BP, или ниже чем аффинность связывания родительского АВ с мишенью.
Если NB-содержащее мультиспецифическое активируемое антитело находится в присутствии мишени, то специфическое связывание АВ с мишенью снижается или ингибируется по сравнению со специфическим связыванием АВ, не ассоциированного с NB или NB:BP. Если NB-содержащее мультиспецифическое активируемое антитело находится в присутствии мишени, то специфическое связывание АВ с мишенью снижается или ингибируется по сравнению со специфическим связыванием родительского АВ с мишенью. В отличие от связывания АВ, не ассоциированного с NB или NB:BP, или связывания родительского АВ с мишенью, способность NB-содержащего мультиспецифического активируемого антитела связываться с мишенью снижается, например, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и даже на 100% по меньшей мере через 2, 4, 6, 8, 12, 28, 24, 30, 36, 48, 60, 72, 84 или 96 часов, или 5, 10, 15, 30, 45, 60, 90, 120, 150 или 180 дней, или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев или более, как было определено в анализе in vitro и/или in vivo.
Если NB-содержащее мультиспецифическое активируемое антитело находится в присутствии мишени, но не в присутствии модифицирующего агента (например, протеазы или другого фермента), то специфическое связывание АВ с мишенью снижается или ингибируется по сравнению со специфическим связыванием АВ, не ассоциированного с NB или NB:BP. Если NB-содержащее мультиспецифическое активируемое антитело находится в присутствии мишени, но не в присутствии модифицирующего агента (например, протеазы, другого фермента, восстановителя или светового излучения), то специфическое связывание АВ с мишенью снижается или ингибируется по сравнению со специфическим связыванием родительского АВ с мишенью. В отличие от связывания АВ, не ассоциированного с NB или NB:BP, или связывания родительского АВ с мишенью, способность NB-содержащего мультиспецифического активируемого антитела связываться с мишенью снижается, например, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и даже на 100% по меньшей мере через 2, 4, 6, 8, 12, 28, 24, 30, 36, 48, 60, 72, 84 или 96 часов, или 5, 10, 15, 30, 45, 60, 90, 120, 150 или 180 дней, или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев или более, как было определено в анализе in vitro и/или in vivo.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифическое активируемое антитело также включает агент, который был конъюгирован с АВ с получением конъюгата мультиспецифического активируемого антитела. В некоторых вариантах конъюгата мультиспецифического активируемого антитела, указанным агентом является терапевтический агент. В некоторых вариантах осуществления изобретения, указанным агентом является дмагностический агент. В некоторых вариантах осуществления изобретения, указанным агентом является детектируемый маркер. В некоторых вариантах конъюгата мультиспецифического активируемого антитела, указанным агентом является противоопухолевый агент. В некоторых вариантах конъюгата мультиспецифического активируемого антитела, указанным агентом является токсин или его фрагмент. В некоторых вариантах конъюгата мультиспецифического активируемого антитела, указанный агент конъюгирован с АВ посредством линкера. В некоторых вариантах конъюгата мультиспецифического активируемого антитела, указанным линкером является расщепляемй линкер. В некоторых вариантах осуществления изобретения, указанным агентом является агент, выбранный из группы, состоящей из агентов, перечисленных в таблице 4. В некоторых вариантах осуществления изобретения, указанным агентом является доластатин. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является ауристатин E или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин E (MMAE). В некоторых вариантах осуществления изобретения, указанным агентом является монометилауристатин D (MMAD). В некоторых вариантах осуществления изобретения, указанным агентом является майтанзиноид или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является DM1 или DM4. В некоторых вариантах осуществления изобретения, указанным агентом является дуокармицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является калихеамицин или его производное. В некоторых вариантах осуществления изобретения, указанным агентом является пирролобензодиазепин.
В некоторых примерах, относящихся к любым вариантам этих мультиспецифических активируемых антител, мультиспецифические активируемые антитела представляют собой мультиспецифические активируемые антитела, связывающиеся с двумя мишенями. Такие мультиспецифические активируемые антитела, связывающиеся с двумя мишенями, содержат два Ab, которые могут связываться с одной и той же мишенью или с различными мишенями. В конкретных вариантах осуществления изобретения, мультиспецифические активируемые антитела, нацеленные на две мишени, включают биспецифические антитела или их фрагменты.
Мультиспецифические активируемые антитела, связывающиеся с двумя мишенями, конструируют так, чтобы они содержали CL, расщепляемый под действием расщепляющего агента, который локализуется в ткани-мишени вместе с одной или обеими мишенями, способными связываться с АВ мультиспецифических активируемых антител. Мультиспецифические активируемые антитела, связывающиеся с двумя мишенями и имеющие более чем одно АВ против одной и той же мишени или различных мишеней, могут быть сконструированы так, чтобы они имели более чем один CL, где первый CL может отщепляться под действием расщепляющего агента в первой ткани-мишени, а второй CL может отщепляться под действием расщепляющего агента во второй ткани-мишени, причем одна или более мишеней могут связываться с AB мультиспецифических активируемых антител. В одном из вариантов осуществления изобретения, первая и вторая ткани-мишени расположены на расстоянии друг от друга, например, они находятся в различных участках организма. В одном из вариантов осуществления изобретения, первая и вторая ткани-мишени представляют собой одни и те же ткани, которые являются мишенями в различные периоды времени, например, они представляют собой одни и те же ткани, которые являются мишенями в два различных периода времени, например, первым периодом времени является ранняя стадия развития опухолевой ткани, а вторым периодом времени является поздняя стадия развития опухолевой ткани.
Настоящее изобретение также относится к молекулам нуклеиновой кислоты, кодирующим описанные здесь мультиспецифические активируемые антитела. Настоящее изобретение также относится к векторам, содержащим эти нуклеиновые кислоты. Описанные здесь мультиспецифические активируемые антитела получают путем культивирования клетки в условиях, стимулирующих экспрессию мультиспецифического активируемого антитела, где указанная клетка включает эти молекулы нуклеиновой кислоты или векторы.
Настоящее изобретение также относится к способам получения мультиспецифических активируемых антител. В одном из вариантов осуществления изобретения, указанный способ включает стадии (а) культивирования клетки, включающей конструкцию нуклеиновой кислоты, кодирующей мультиспецифическое активируемое антитело, в условиях, стимулирующих экспрессию мультиспецифического активируемого антитела, где указанное мультиспецифическое активируемое антитело включает (i) несвязывающуюся стерическую группу (NB); (ii) расщепляемый линкер (CL); и (iii) антитело или его антигенсвязывающий фрагмент (AB), которые специфически связываются с мишенью, где (1) NB специфически не связывается с AB; (2) CL представляет собой полипептид, который включает субстрат (S) для фермента; (3) CL расположен так, что, если он находится в нерасщепленном состоянии, то NB блокирует связывание АВ с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ с мишенью; и (4) NB не ингибирует расщепление CL ферментом; и (b) выделения мультиспецифического активируемого антитела.
В другом варианте осуществления изобретения, указанный способ включает стадии (а) культивирования клетки, включающей конструкцию нуклеиновой кислоты, кодирующей мультиспецифическое активируемое антитело, в условиях, стимулирующих экспрессию мультиспецифического активируемого антитела, где указанное мультиспецифическое активируемое антитело включает (i) партнера по связыванию (ВР) с несвязывающейся стерической группой (NB); (ii) расщепляемый линкер (CL); и (iii) антитело или его антигенсвязывающий фрагмент (AB), которые специфически связываются с мишенью, где (1) NB специфически не связывается с AB; (2) CL представляет собой полипептид, который включает субстрат (S) для фермента; (3) CL расположен так, что, если он находится в нерасщепленном состоянии в присутствии NB, то NB блокирует связывание АВ с мишенью, а если он находится в расщепленном состоянии, то NB не блокирует связывание АВ с мишенью и ВР не блокирует связывание АВ с мишенью; и (4) NB и ВР не ингибируют расщепление CL ферментом; и (b) выделения мультиспецифического активируемого антитела. В некоторых примерах этого варианта, ВР мультиспецифического активируемого антитела связывается с NB.
Применение мультиспецифических антител и мультиспецифических активируемых антител
Следует отметить, что терапевтические средства согласно изобретению могут быть введены в комбинации с подходящими носителями, наполнителями и другими агентами, включенными в композиции для облегчения переноса и доставки, а также для улучшения переносимости лекарственных средств и т.п. Множество соответствующих препаратов можно найти в реестре всех лекарственных средств, известных химикам-фармацевтам, см. руководство Remington's Pharmaceutical Sciences (15th ed., Mack Publishing Company, Easton, PA (1975)), а в частности, главу 87, Blaug, Seymour. Такими препаратами являются, например, порошки, пасты, мази, гели, воски, масла, липиды, липид (катионный или анионный)-содержащие везикулы (такие как Lipofectin™), ДНК-конъюгаты, безводные абсорбирующие пасты, эмульсии типа «масло в воде» и «вода в масле», эмульсии карбовакс (Сarbowax) (полиэтиленгликоли с различными молекулярными массами), полутвердые гели и полутвердые смеси, содержащие карбовакс. Любая из вышеупомянутых смесей может быть использована для лечения и терапии способами согласно изобретению, при условии, что активный ингредиент в данном препарате не был инактивирован при приготовлении такого препарата, и что такой препарат является физиологически совместимым и подходящим для данного способа введения. См. также публикации Baldrick P. «Pharmaceutical excipient development: the need for preclinical guidance». Regul. Toxicol Pharmacol. 32(2):210-8 (2000), Wang W. «Lyophilization and development of solid protein pharmaceuticals». Int. J. Pharm. 203(1-2):1-60 (2000), Charman WN «Lipids, lipophilic drugs, and oral drug delivery-some emerging concepts». J. Pharm. Sci.89(8):967-78 (2000), Powell et al. «Compendium of excipients for parenteral formulations» PDA J. Pharm. Sci. Technol. 52:238-311 (1998), и цитируемые в них работы, в которых приводится дополнительная информация о препаратах, наполнителях и носителях, хорошо известных химикам-фармацевтам.
В одном из вариантов осуществления изобретения, мультиспецифические антитело и/или мультиспецифическое активируемое антитело согласно изобретению могут быть использованы в качестве терапевтических агентов. Такие агенты обычно используются для диагностики, прогноза, мониторинга, лечения, ослабления симптомов и/или предупреждения заболевания или патологии у индивидуума. Курс терапии проводят после выявления у пациента, например, у человека или другого млекопитающего, расстройства (или риска развития такого расстройства) стандартными методами. Так, например, в некоторых вариантах осуществления изобретения, препарат мультиспецифического антитела и/или мультиспецифического активируемого антитела, обладающего высокой специфичностью и высокой аффинностью по отношению к двум или более антигенам мишени, вводят индивидууму, и этот препарат действует, в основном, посредством его связывания с мишенями. Введение мультиспецифического антитела и/или мультиспецифического активируемого антитела может блокировать, ингибировать или предотвращать передачу сигнала одной или более мишенями. Введение мультиспецифического антитела и/или мультиспецифического активируемого антитела может блокировать, ингибировать или предотвращать связывание одной или более мишеней с эндогенным лигандом, с которым они связываются в природе.
Вообще говоря, ослабление симптомов или лечение заболевания или расстройства включает ослабление одного или более симптомов или патологических состояний, ассоциированных с таким заболеванием или расстройством. Так, например, в случае рака, терапевтически эффективное количество лекарственного средства может давать один эффект или комбинацию нижеследующих эффектов, а именно: снижение числа раковых клеток; уменьшение размера опухоли; ингибирование (то есть снижение до определенной степени и/или блокирование) инфильтрации раковых клеток в периферические органы; ингибирование метастазов опухоли; ингибирование, до некоторой степени, роста опухоли и/или ослабление, до некоторой степени, одного или более симптомов, ассоциированных с раковым заболеванием. В некоторых вариантах осуществления изобретения, композиция согласно изобретению может быть использована для предупреждения начала развития или рецидива заболевания или расстройства у индивидуума, например, у человека или другого млекопитающего, такого как примат, не являющийся человеком, животное-компаньон (например, кошка, собака, лошадь), сельскохозяйственное животное, рабочий скот или животное, содержащееся в зоопарке. Используемые здесь термины «индивидуум» и «пациент» являются синонимами.
Термин «терапевтически эффективное количество» мультиспецифического антитела и/или мультиспецифического активируемого антитела согласно изобретению, по существу, означает количество, необходимое для достижения терапевтического эффекта. Как указывалось выше, такой эффект может представлять собой связывание мультиспецифического антитела и/или мультиспецифического активируемого антитела с антигенами мишени, где указанное связывание, в некоторых случаях, может блокировать функцию мишеней. Кроме того, количество антитела, необходимое для введения, зависит от аффинности связывания мультиспецифического антитела и/или мультиспецифического активируемого антитела со специфическим антигеном, а также от скорости, с которой указанное мультиспецифическое антитело и/или мультиспецифическое активируемое антитело выводится в свободном виде у другого индивидуума, которому было введено это антитело. Обычно, интервалы терапевтически эффективнх доз мультиспецифического антитела и/или его фрагмента и/или мультиспецифического активируемого антитела согласно изобретению составляют, например, но не ограничиваются ими, приблизительно от 0,1 мг/кг массы тела до 50 мг/кг массы тела. Частота введения доз может составлять, например, от двух раз в день до одного раза в неделю.
Эффективность лечения определяют любым известным методом диагностики или лечения конкретного расстройства. Методами скрининга мультиспецифических антител и/или мультиспецифических активируемых антител, которые обладают нужной специфичностью, являются, но не ограничиваются ими, твердофазный иммуноферментный анализ (ELISA) и другие иммунологически опосредованные методы, известные специалистам.
В другом варианте осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, направленное на две или более мишеней, используют в известных методах определения локализации и/или количественной оценки мишеней (например, для измерения уровней одной или более мишеней в соответствующих физиологических образцах, для диагностики, для визуализации белка и т.п.). В данном варианте осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, направленное на две или более мишеней, или его производное, фрагмент, аналог или гомолог, содержащие антигенсвязывающий домен этого антитела, используют в качестве фармакологически активных соединений (называемых далле «терапевтическими средствами»).
В другом варианте осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, направленное на две или более мишеней, используют для выделения одной или более мишеней стандартными методами, такими как иммуноаффинная хроматография, хроматография или иммунопреципитация. Мультиспецифическое антитело и/или мультиспецифическое активируемое антитело (или его фрагмент), направленное на две или более мишеней, используют в диагностических целях для мониторинга уровенй белка в ткани при проведении клинического анализа, например, для определения эффективности данного курса лечения. Детектирование может быть упрощено путем связывания (то есть, физического связывания) антитела с детектируемым веществом. Примерами детектируемых веществ являются различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества и радиоактивные вещества. Примерами подходящих ферментов являются пероксидаза хрена, щелочная фосфатаза, β-галактозидаза или ацетилхолинэстераза; примерами подходящих комплексов простетических групп являются стрептавидин/биотин и авидин/биотин; примерами подходящих флуоресцентных веществ являются умбеллиферон, флуоресцеин, флуоресцеинизотиоцианат, родамин, дихлортриазиниламин-флуоресцеин, данзилхлорид или фикоэритрин; примером люминесцентного вещества является люминол; примерами биолюминесцентных веществ являются люцифераза, люциферин и экворин, а примерами подходящих радиоактивных веществ являются 125I, 131I, 35S или 3H.
В другом варианте осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело, направленное на две или более мишеней, могут быть использованы в качестве агента для детектирования присутствия одной или более мишеней (или их фрагментов) в образце. В некоторых вариантах осуществления изобретения, антитело содержит детектируемую метку. Антителами являются поликлональные антитела или, в некоторых вариантах осуществления изобретения, моноклональные антитела. При этом может быть использовано интактное антитело или его фрагмент (например, Fab, scFv или F(ab)2). Термин «меченный», если он относится к зонду или антителу, включает прямое мечение зонда или антитела путем связывания (то есть физического связывания) детектируемого вещества с зондом или антителом, а также непрямое мечение зонда или антитела посредством реакции взаимодействия с другим реагентом, который является непосредственно меченным. Примерами непрямого мечения являются детектирование «первого» антитела с использованием флуоресцентно меченного «второго» антитела и концевое мечение антитела биотином, где такое антитело может быть детектировано с использованием флуоресцентно меченного стрептавидина. Термин «биологический образец» включает ткани, клетки и биологические жидкости, выделенные у индивидуума, а также ткани, клетки и жидкости, присутствующие у индивидуума. В определение термина «биологический образец» также входят кровь и фракция или компонент крови, включая сыворотку крови, плазму крови или лимфу. Таким образом, способ детектирования согласно изобретению может быть применен для детектирования белка в биологическом образце in vitro и in vivo. Так, например, in vitro методами детектирования белка-аналита являются твердофазные иммуноферментные анализы (ELISA), вестерн-блот-анализы, иммунопреципитация и иммунофлуоресценция. Процедуры проведения иммуноанализов описаны, например, в публикациях “ELISA: Theory and Practice: Methods in Molecular Biology”, Vol. 42, J. R. Crowther (Ed.) Human Press, Totowa, N.J., 1995; “Immunoassay”, E. Diamandis and T. Christopoulus, Academic Press, Inc., San Diego, CA, 1996; и “Practice and Theory of Enzyme Immunoassays”, P. Tijssen, Elsevier Science Publishers, Amsterdam, 1985. Кроме того, in vivo методы детектирования белка-аналита включают введение индивидууму меченного антитела против белка-аналита. Так, например, антитело может быть помечено радиоактивным маркером, присутствие и локализация которого у индивидуума могут быть детектированы стандартными методами визуализации.
Для приготовления различных диагностических и профилактических препаратов используют мультиспецифические антитела и/или мультиспецифические активируемые антитела согласно изобретению. В одном из вариантов осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело вводят пациентам с риском развития у них одного или более вышеупомянутых расстройств. Предрасположенность организма пациента или какого-либо его органа к развитию одного или более вышеупомянутых расстройств может быть определена с использованием генотипических, серологических или биохимических маркеров.
В другом варианте осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело вводят человеку, у которого была диагностирована клиническая патология, ассоциированная с одним или более вышеупомянутыми расстройствами. После установления диагноза, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело вводят для ослабления или устранения симптомов такой клинической патологии.
Мультиспецифические антитела и/или мультиспецифические активируемые антитела могут быть также использованы для детектирования одной или более мишени в образцах, взятых у пациента, и соответственно, для установления диагноза заболевания. Так, например, мультиспецифические антитела и/или мультиспецифические активируемые антитела согласно изобретению могут быть использованы в анализах in vitro, например, ELISA, для детектирования уровней одной или более мишеней в образце, взятом у пациента.
В одном из вариантов осуществления изобретения, мультиспецифическое антитело и/или мультиспецифическое активируемое антитело иммобилизуют на твердом носителе (например, на лунке(ах) микротитрационного планшета). Иммобилизованное антитело и/или активируемое антитело служит в качестве антитела для захвата любой(ых) мишени(ей), которая(ые) может (могут) присутствовать в тестируемом образце. Перед контактированием иммобилизованного мультиспецифического антитела и/или иммобилизованного мультиспецифического активируемого антитела с образцом, взятым у пациента, твердый носитель промывают и обрабатывают блокирующим агентом, таким как молочный белок или альбумин, для предупреждения неспецифической адсорбции аналита.
Затем лунки обрабатывают тестируемым образцом, предположительно, содержащим антиген, или раствором, содержащим стандартное количество антигена. Таким образцом является, например, проба сыворотки, взятая у индивидуума с подозрением на наличие определенных уровней антигена в кровотоке, которые могут служить показателем для диагностики данной патологии. После промывки тестируемого образца или стандарта, твердый носитель обрабатывают «вторым» антителом, которое является детектируемо меченным. Меченное «второе» антитело служит в качестве детектирующего антитела. Затем измеряют уровень детектируемой метки и определяют концентрацию антигена(ов) мишени в тестируемом образце путем сравнения по стандартной кривой, построенной для стандартных образцов.
Следует отметить, что исходя из результатов, полученных для мультиспецифического антитела и/или мультиспецифического активируемого антитела в диагностическом анализе in vitro, можно определить стадию развития заболевания у индивидуума по уровням экспрессии антигена(ов) мишени. Для мониторига течения данного заболевания, у пациентов с диагностированным заболеванием берут пробы крови на различных стадиях прогрессирования заболевания и/или в различные периоды времени проведения терапевтического лечения данного заболевания. С использованием набора образцов, которые будет обеспечивать статистически значимые результаты для каждой стадии прогрессирования или лечения заболевания, определяют интервал концентраций антигена, которые могут рассматриваться как характерные для каждой стадии заболевания.
Мультиспецифические антитела и/или мультиспецифические активируемые антитела могут быть также использованы в методах диагностики и/или визуализации. В некоторых вариантах осуществления изобретения, такими методами являются методы in vitro. В некоторых вариантах осуществления изобретения, такими методами являются методы in vivo. В некоторых вариантах осуществления изобретения, такими методами являются методы in situ. В некоторых вариантах осуществления изобретения, такими методами являются методы ex vivo. Так, например, мультиспецифические антитела и/или мультиспецифические активируемые антитела, имеющее ферментативно отщепляемую СМ, могут быть использованы для детектирования присутствия или отсутствия фермента, способного отщеплять СМ. Такие мультиспецифические антитела и/или мультиспецифические активируемые антитела могут быть использованы в диагностике, которая может включать in vivo детектирование (например, качественное или количественнное) ферментативной активности (или, в некоторых вариантах осуществления изобретения, определение условий повышения потенциала восстановления, таких как условия, способствующие восстановлению дисульфидной связи) путем измерения уровня аккумуляции активированных мультиспецифических антител (то есть, антител, образующихся в результате расщепления мультиспецифического активируемого антитела) в данной клетке или ткани указанного организма хозяина. Такая аккумуляция активированных мультиспецифических антител указывает не только на то, что ткань экспрессирует ферментативную активность (или имеет повышенный потенциал восстановления в зависимости от природы CM), но также и на то, что эта ткань экспрессирует по меньшей мере одну мишень, с которой связывается активированное антитело.
Так, например, может быть выбрана такая CM, которая является субстратом для протеазы, присутствующей на участке опухоли, на участке вирусной или бактериальной инфекции в биологически изолированном участке (например, таком как абсцесс, орган и т.п.) и т.п. По меньшей мере одно AB может представлять собой антитело, которое связывается с антигеном мишени. Детектируемая метка (например, флуоресцентная метка или радиоактивная метка или радиоактивный индикатор) может быть конъюгирована, если это необходимо, с AB или другой областью мультиспецифического антитела и/или мультиспецифического активируемого антитела с применением методов, известных специалистам. Подходящие детектируемые метки обсуждаются в описании вышеупомянутых методов скрининга и в дополнительных конкретных примерах, представленных ниже. При использовании по меньшей мере одного AB, специфичного к белку или пептиду, находящемуся на участке поражения вместе с протеазой, активность которой повышается в представляющей интерес пораженной ткани, эти активируемые антитела будут обнаруживать повышенный уровень связывания с пораженной тканью, по сравнению с тканями, в которых CM-специфический фермент либо не присутствует на детектиремом уровне, либо присутствует на более низком уровне, чем в пораженной ткани, либо является неактивным (например, присутствует в форме зимогена или в комплексе с ингибитором). Поскольку небольшие белки и пептиды быстро выводятся из кровотока благодаря системе почечной фильтрации, и поскольку фермент, специфичный к CM, не присутствует на детектиремом уровне (или присутствует на более низких уровнях в нормальных тканях или присутствует в неактивной конформации), аккумуляция активированных мультиспецифических антител в пораженной ткани повышается в отличие от аккумуляции этих антител в непораженных тканях.
В другом примере, активируемые мультиспецифические антитела могут быть использованы для детектирования присутствия или отсутствия расщепляющего агента в образце. Так, например, если мультиспецифические активируемые антитела содержат СМ, чувствительную к отщеплению ферментом, то такие мультиспецифические активируемые антитела могут быть использованы для детектирования (качественного или количественнного) присутствия фермента в образце. В другом примере, если мультиспецифические активируемые антитела содержат СМ, чувствительную к отщеплению восстановителем, то такие мультиспецифические активируемые антитела могут быть использованы для детектирования (качественного или количественнного) присутствия восстановителя в образце. Для облегчения проведения анализа этими методами, мультиспецифические активируемые антитела могут быть детектируемо помечены и могут быть связаны с носителем (например, с твердым носителем, таким как предметное стекло или сферы). Детектируемая метка может присутствовать на фрагменте активируемого антитела, которое не высвобождается после расщепления, например, такой детектируемой меткой может быть гашеная флуоресцентная метка или другая метка, которая не детектируется до ее отщепления. Этот анализ может быть проведен, например, путем контактирования иммобилизованных детектируемо меченных мультиспецифических активируемых антител с образцом, предположительно содержащим фермент и/или восстановитель, в течение периода времени, достаточного для расщепления, и путем последующего удаления избыточного количества образца и примесей. Затем, присутствие или отсутствие расщепляющего агента (например, фермента или восстановителя) в образце оценивают по изменению детектируемого сигнала мультиспецифических активируемых антител до их контактирования с образцом, например, проводят анализ на присутствие и/или усиление детектируемого сигнала, обусловленное расщеплением активируемого антитела расщепляющим агентом в образце.
Такие методы детектирования могут быть также адаптированы для анализа на присутствие или отсутствие мишени, способной связываться по меньшей мере с одним АВ мультиспецифических активируемых антител в расщепленной форме. Таким образом, эти анализы могут быть адаптированы для оценки присутствия или отсутствия расщепляющего агента и присутствия или отсутствия представляющей интерес мишени. Присутствие или отсутствие расщепляющего агента может быть детектировано по присутствию и/или увеличению детектируемого уровня мультиспецифических активируемых антител как описано выше, а присутствие или отсутствие мишени может быть детектировано по обнаружению комплекса «мишень-АВ», например, с использованием детектируемо меченного антитела против мишени.
Мультиспецифические активируемые антитела могут быть также использованы в методе визуализации in situ для подтверждения активации активируемого антитела, например, по расщеплению протеазой и связыванию с конкретной мишенью. Визуализация in situ представляет собой метод, позволяющий определить локализацию протеолитической активности и мишени в биологических образцах, таких как клеточные культуры или срезы тканей. С применением этого метода можно подтвердить связывание с указанной мишенью и наличие протеолитической активности по присутствию детектируемой метки (например, флуоресцентной метки).
Эти методы могут быть проведены с использованием любых замороженных клеток или тканей, взятых из пораженного участка (например, ткани опухоли), или из здоровых тканей. Эти методы могут быть также проведены с использованием образцов свежих клеток или тканей.
В этих методах, активируемое антитело было помечено детектируемой меткой. Такой детектируемой меткой могут быть флуоресцентный краситель (например, флуорофор, флуоресцеин-изотиоцианат (FITC), родамин-изотиоцианат (TRITC), метка Alexa Fluor®, краситель, флуоресцирующий в области ближнего инфракрасного излучения (NIR) (например, нанокристаллы Qdot®), коллоидный металл, гаптен, радиоактивный маркер, биотин и реагент для амплификации, такой как стрептавидин или фермент (например, пероксидаза хрена или щелочная фосфатаза).
Детектирование метки в образце, который был инкубирован с меченным мультиспецифическим активируемым антителом, указывает на то, что этот образец содержит мишень и протеазу, специфичную к CM мультиспецифического активируемого антитела. В некоторых вариантах осуществления изобретения, присутствие протеазы может быть подтверждено с использованием описанных здесь ингибиторов протеазы широкого спектра и/или с использованием агента, специфичного к протеазе, например, антитела, такого как A11, которое является специфичным к матриктазе (MT-SP1) и ингибирует протеолитическую активность MT-SP1; см., например, публикацию Международной заявки WO 2010/129609, опубликованную 11 ноября 2010. Тот же самый подход с использованием описанных здесь ингибиторов протезаы широкого спектра и/или с использованием более селективного ингибирующего агента может быть применен для идентификации протеазы или класса протеаз, специфичных к CM активируемого антитела. В некоторых вариантах осуществления изобретения, присутствие мишени может быть подтверждено с использованием агента, специфичного к мишени, или детектируемой метки, которая может конкурировать с немеченной мишенью. В некоторых вариантах осуществления изобретения может быть использовано немеченное активируемое антитело, но в этом случае, детектирование осуществляют с использованием меченного «второго» антитела или более сложной системы детектирования.
Аналогичные методы могут быть также применены для визуализации in vivo, где детектирование флуоресцентного сигнала у индивидуума, например, у млекопитающего, включая человека, указывает на то, что пораженный участок содержит мишень и протеазу, специфичную к CM мультиспецифического активируемого антитела.
Эти методы могут быть также применены с использованием наборов и/или реагентов для детектирования, идентификации или характеризации протеазной активности в различных клетках, тканях и организмах, где указанные методы были разработаны на основе протеазо-специфической CM в мультиспецифическом активируемом антителе.
Терапевтическое применение и приготовление мультиспецифических антител и/или мультиспецифических активируемых антител
Следует отметить, что терапевтические средства согласно изобретению могут быть введены в комбинации с подходящими носителями, наполнителями и другими агентами, включенными в композиции для облегчения переноса и доставки, а также для улучшения переносимости лекарственных средств и т.п. Множество соответствующих препаратов можно найти в реестре всех лекарственных средств, известных химикам-фармацевтам, см. руководство Remington's Pharmaceutical Sciences (15th ed., Mack Publishing Company, Easton, PA (1975)), а в частности, главу 87, Blaug, Seymour. Такими препаратами являются, например, порошки, пасты, мази, гели, воски, масла, липиды, липид (катионный или анионный)-содержащие везикулы (такие как Lipofectin™), ДНК-конъюгаты, безводные абсорбирующие пасты, эмульсии типа «масло в воде» и «вода в масле», эмульсии карбовакс (Сarbowax) (полиэтиленгликоли с различными молекулярными массами), полутвердые гели и полутвердые смеси, содержащие карбовакс. Любая из вышеупомянутых смесей может быть использована для лечения и терапии способами согласно изобретению, при условии, что активный ингредиент в данном препарате не был инактивирован при приготовлении такого препарата, и что такой препарат является физиологически совместимым и подходящим для данного способа введения. См. также публикации Baldrick P. «Pharmaceutical excipient development: the need for preclinical guidance». Regul. Toxicol Pharmacol. 32(2):210-8 (2000), Wang W. «Lyophilization and development of solid protein pharmaceuticals». Int. J. Pharm. 203(1-2):1-60 (2000), Charman WN «Lipids, lipophilic drugs, and oral drug delivery-some emerging concepts». J. Pharm. Sci.89(8):967-78 (2000), Powell et al. «Compendium of excipients for parenteral formulations» PDA J. Pharm. Sci. Technol. 52:238-311 (1998), и цитируемые в них работы, в которых приводится дополнительная информация о препаратах, наполнителях и носителях, хорошо известных химикам-фармацевтам.
В некоторых вариантах осуществления изобретения, композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител вводят в сочетании с одним или более дополнительными агентами или комбинацией дополнительных агентов. Подходящими дополнительными агентами являются современные фармацевтические средства и/или средства, используемые в хирургии. Так, например, композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител могут быть использованы в комбинации с дополнительными химиотерапевтическими или противоопухолевыми агентами. Так, например, композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент приготавливают в виде одной терапевтической композиции, и указанные композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент вводят одновременно. Альтернативно, композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент вводят отдельно друг от друга, например, каждый из них приготавливают в виде отдельной терапевтической композиции, и указанные композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент вводят одновременно, либо композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент вводят в различные периоды времени в течение всего курса лечения. Так, например, композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител вводят перед введением дополнительного агента, либо указанные композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител вводят после введения дополнительного агента, либо композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент вводят поочередно. Как описано в настоящей заявке, композиции мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител и дополнительный агент вводят в разовых дозах или в дробных дозах.
В некоторых вариантах осуществления изобретения, дополнительный агент связывают с композициями мультиспецифических антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител, или как-либо иначе присоединяют к этим композициям.
Подходящие дополнительные агенты выбирают в соответствии с целями их применения (то есть, в целях подавления или предупреждения пролиферации клеток, для гормонотерапии или генотерапии). Такими агентами могут быть, но не ограничиваются ими, например, фармацевтические агенты, токсины, фрагменты токсинов, алкилирующие агенты, ферменты, антибиотики, антиметаболиты, антипролиферативные средства, гормоны, нейромедиаторы, ДНК, РНК, киРНК, олигонуклеотиды, антисмысловые РНК, аптамеры, диагностические средства, рентгеноконстрастные красители, радиоактивные изотопы, флуорогенные соединения, магнитные метки, наночастицы, маркерные соединения, лектины, соединения, модифицирующие проницаемось клеточной мембраны, фотохимические соединения, низкомолекулярные соединения, липосомы, мицеллы, векторы для генотерапии, вирусные векторы и т.п. И наконец, могут быть использованы комбинации этих агентов или комбинации агентов различных классов.
Композиции мультиспецифические антител, мультиспецифических активируемых антител и/или конъюгированных мультиспецифических активируемых антител согласно изобретению (также называемые здесь «активными соединениями») и их производные, фрагменты, аналоги и гомологи могут быть включены в фармацевтические композиции, подходящие для введения. Принципы и аспекты, лежащие в основе приготовления таких композиций, а также инструкции по выбору компонентов этих композиций изложены, например, в руководствах Remington: The Science And Practice Of Pharmacy» 19th ed. (Alfonso R. Gennaro, et al., editors) Mack Pub. Co., Easton, Pa.: 1995; «Drug Absorption Enhancement: Concepts, Possibilities, Limitations, And Trends, Harwood Academic Publishers, Langhorne, Pa., 1994; and Peptide And Protein Drug Delivery (Advances In Parenteral Sciences, Vol. 4), 1991, M. Dekker, New York.
Такие композиции обычно содержат мультиспецифическое антитело и/или мультиспецифическое активируемое антитело и фармацевтически приемлемый носитель. Если мультиспецифическое антитело и/или мультиспецифическое активируемое антитело включает фрагмент домена AB, то может быть использован наименьший фрагмент АВ, который специфически связывается со связывающимся доменом белка мишени. Так, например, на основе последовательностей вариабельной области антитела могут быть сконструированы пептидные молекулы, сохраняющие способность АВ связываться с последовательностью белка мишени. Такие пептиды могут быть получены методами химического синтеза и/или методами рекомбинантных ДНК (см. например, Marasco et al., Proc. Natl. Acad. Sci. USA, 90: 7889-7893 (1993)).
Используемый здесь термин «фармацевтически приемлемый носитель» включает любые и все растворители; дисперсионные среды; материалы для покрытий; антибактериальные и противогрибковые агенты; агенты, придающие изотоничность; агенты, замедляющие абсорбцию и т.п., совместимые со способом введения фармацевтической композиции. Подходящие носители описаны в самом последнем издании классического руководства Remington's Pharmaceutical Sciences, в соответствующем разделе, который вводится в настоящее описание посредством ссылки. Предпочтительными примерами таких носителей или разбавителей являются, но не ограничиваются ими, вода, физиологический раствор, растворы Рингера, раствор декстрозы и 5% альбумин человеческой сыворотки. При этом, могут быть также использованы липосомы и безводные носители, такие как жирные масла. Применение таких сред и агентов для приготовления фармацевтически активных веществ хорошо известно специалистам. В описанных здесь композициях могут быть использованы любые стандартные среды или агенты, при условии, что они будут совместимыми с активным соединением.
Препараты, используемые для введения in vivo, должны быть стерильными. Это может быть достигнуто путем фильтрации через стерильные фильтровальные мембраны.
Фармацевтическую композицию согласно изобретению приготавливают так, чтобы она была совместима с выбранным способом введения. Примерами таких способов введения являются парентеральное введение, например, внутривенное введение, интрадермальное введение, подкожное введение, пероральное введение (например, введение путем ингаляции), чрезкожное введение (то есть, местное введение), введение через слизистую и ректальное введение. Растворы или суспензии для парентерального, интрадермального или подкожного введения могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, жирные масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатообразующие агенты, такие как этилендиаминтетрауксусная кислота (EDTA); буферы, такие как ацетаты, цитраты или фосфаты; и агенты, придающие тоничность, такие как хлорид натрия или декстроза. pH может быть скорректирован путем добавления кислот или оснований, таких как соляная кислота или гидроксид натрия. Препарат для парентерального введения может быть заключен в ампулы, одноразовые шприцы или сосуды для введения дробных доз, изготовленные из стекла или пластика.
Фармацевтическими композициями, подходящими для инъекций, являются стерильные водные растворы (то есть, водорастворимые композиции) или дисперсии и стерильные порошки для немедленного приготовления препаратов в виде стерильных инъецируемых растворов или дисперсий. Для внутривенного введения, подходящими носителями являются физиологический раствор, бактериостатическая вода, Cremophor EL™ (BASF, Parsippany, N.J.) или забуференный фосфатом физиологический раствор (PBS). Во всех случаях, такая композиция должна быть стерильной и должна обладать текучестью, достаточной для легкого наполнения шприца. Эта композиция должна быть стабильной в условиях ее изготовления и хранения, а также должна быть защищена от контаминирующего действия микроорганизмов, таких как бактерии и грибы. Таким носителем может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Нужная текучесть может поддерживаться, например, с использованием вещества для нанесения покрытия, такого как лецитин, благодаря сохранению нужного размера частиц, в случае дисперсии, и с использованием поверхностно-активных веществ. Предупреждение воздействия микроорганизмов может быть достигнуто с использованием различных антибактериальных и противогрибковых агентов, таких как, например, парабены, хлорбутанол, фенол, аскорбиновая кислота, тимерозал и т.п. Во многих случаях может оказаться предпочтительным включение в композицию агентов, придающих изотоничность, таких как, например, сахара, многоатомные спирты, такие как маннит и сорбит, и хлорид натрия. Пролонгированное всасывание инъецируемых композиций может быть достигнуто путем включения в композицию агента, замедляющего всасывание, например, моностеарата алюминия и желатина.
Стерильные растворы для инъекций могут быть приготовлены путем включения активного соединения в необходимом количестве в соответствующем растворителе вместе с одним из ингредиентов или, если это необходимо, в комбинации с ингредиентами, перечисленными выше, и последующей стерилизации на фильтре. Дисперсии обычно приготавливают путем включения активного соединения в стерильный наполнитель, который содержит основную дисперсионную среду и другие необходимые ингредиенты, перечисленные выше. Методы получения стерильных порошков в целях приготовления стерильных растворов для инъекций включают вакуумную сушку и лиофилизацию с получением порошка, содержащего активный ингредиент и любой дополнительный нужный ингредиент, полученный из предварительно отфильтрованного стерильного раствора.
Композиции для перорального введения обычно включают инертный разбавитель или пищевой носитель. Эти композиции могут быть заключены в желатиновые капсулы или спрессованы в таблетки. Активное соединение, предназначенное для терапевтического перорального введения, может быть введено вместе с наполнителями и приготовлено в форме таблеток, пастилок или капсул. Композиции для перорального введения могут быть также приготовлены с использованием жидкого носителя, применяемого как жидкость для полоскания рта, где указанное соединение в жидком носителе приготавливают в виде препарата для перорального введения, в виде препарата для полоскания, в виде отхаркивающющего средства или в виде препарата для проглатывания. Фармацевтически совместимые связывающие агенты и/или адъюванты могут быть включены как часть композиции. Таблетки, драже, капсулы, пастилки и т.п. могут содержать любой из нижеследующих ингредиентов или любые соединения аналогичной природы, а именно: связывающее вещество, такое как микрокристаллическая целлюлоза; трагакантовая камедь или желатин; наполнитель, такой как крахмал или лактоза; дезинтегрирующий агент, такой как альгиновая кислота, примогель или кукурузный крахмал; замасливатель, такой как стеарат магния или Sterotes; вещество, повышающее скольжение, такое как коллоидная двуокись кремния; подсластитель, такой как сахароза или сахарин; или ароматизатор, такой как перечная мята, метилсалицилат или апельсиновая отдушка.
Соединения, вводимые путем ингаляции, доставляют в форме аэрозольного спрея из контейнера под давлением или дозирующего устройства, содержащего подходящий пропеллент, например, газ, такой как диоксид углерода, или из аэрозольного распылителя.
Системное введение может быть также осуществлено через слизистую или чрезкожно. В препаратах для введения через слизистую или чрезкожного введения используются подходящие пенетранты, проникающие через барьер. Пенетранты, применяемые для введения через слизистую, по существу, известны специалистам, и ими являются, например, детергенты, соли желчных кислот и производные фузидиновой кислоты. Введение через слизистую может быть осуществлено с помощью интраназальных спреев или суппозиториев. Для трансдермального введения, активные соединения приготавливают в виде твердых мазей, жидких мазей, гелей или кремов, по существу, известных специалистам.
Соединения могут быть также получены в форме суппозиториев (например, содержащих стандартные основы для суппозиториев, такие как масло какао и другие глицериды) или удерживающих клизм для ректального введения.
В одном из вариантов осуществления изобретения, активные соединения приготавливают вместе с носителями, которые защищают соединение от его быстрого выведения из организма, в результате чего получают препараты замедленного пролонгированного высвобождения, включая имплантаты и микроинкапсулированные системы доставки. При этом могут быть использованы биологически разлагаемые и биологически совместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Методы получения таких препаратов известны специалистам.
Так, например, активные ингредиенты могут быть также заключены в микрокапсулы, полученные, например, методами коацервации или межфазной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и в полиметилметакрилатные микрокапсулы, соответственно, в коллоидные системы доставки лекарственных средств (например, в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии.
Могут быть получены препараты пролонгированного высвобождения. Подходящими примерами препаратов прологнированного высвобождения являются полупроницаемые матрицы, состоящие из твердых гидрофобных полимеров, содержащих антитело, где указанные матрицы получают в виде сформованных изделий, например, пленок или микрокапсул. Примерами матриц прологнированного высвобождения являются сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат) или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида) и поли-D-(-)-3-гидроксимасляная кислота. Полимеры, такие как этиленвинилацетат и сополимер молочной кислоты и гликолевой кислоты, высвобождают молекулы более чем за 100 дней, а некоторые гидрогели высвобождают белки за более короткие промежутки времени.
Материалы могут быть также получены у таких компаний, как Alza Corporation и Nova Pharmaceuticals, Inc. Липосомные суспензии (включая липосомы, доставляемые в инфицированные клетки вместе с моноклональными антителами против вирусных антигенов) могут быть также использованы в виде фармацевтически приемлемых носителей. Эти суспензии могут быть получены методами, известными специалистам, например, методами, описанными в патенте США No. 4522811.
Для простоты введения и равномерного распределения доз, особенно предпочтительно, чтобы композиции для перорального или парентерального введения были приготовлены в виде унифицированной лекарственной формы. Используемый здесь термин «унифицированная лекарственная форма» означает физически дискретные формы, подходящие для их введения индивидууму, подвергаемому лечению, в виде унифицированных доз, где каждую унифицированную дозу, содержащую предварительно определенное количество активного соединения в комбинации с необходимым фармацевтическим носителем, вычисляют так, чтобы эта доза давала желаемый терапевтический эффект. Спецификация для унифицированных лекарственных форм согласно изобретению может быть составлена исходя из нижеследующих факторов и непосредственно зависит от этих факторов, а именно: уникальных свойств активного соединения и конкретного терапевтического эффекта, а также ограничений, известных специалистам в области составления рецептуры лекарственных средств, таких как активное соединение для лечения индивидуумов.
Фармацевтические композиции могут быть включены в контейнер, упаковку или дозирующее устройство, содержащие инструкции по введению таких фармацевтических композиций.
Препарат может также содержать более чем одно активное соединение, необходимое для лечения конкретного заболевания, например, соединения с дополнительными активностями, не оказывающие негативного воздействия друг на друга. Альтернативно или дополнительно, такая композиция может содержать агент, улучшающий ее функцию, такой как, например, цитотоксический агент, цитокин, химиотерапевтический агент или рост-ингибирующий агент. Такие молекулы обычно присутствуют в комбинации в количестве, эффективном для достижения нужной цели.
В одном из вариантов осуществления изобретения, для проведения комбинированной терапии, активные соединения вводят в комбинации с другими агентами, например, терапевтическими агентами, подходящими для лечения патологических состояний или расстройств, таких как аутоимунные расстройства и воспалительные заболевания. В этом контексте, термин «в комбинации» означает, что указанные агенты, в основном, вводят совместно, то есть либо одновременно, либо последовательно. При последовательном введении, в начале введения второго соединения, первое соединение еще детектируется в эффективной концентрации на участке введения.
Так, например, комбинированная терапия может включать применение одного или более антител согласно изобретению, приготовленных и/или вводимых вместе с одним или более дополнительными терапевтическими агентами, например, с одним или более соединениями, такими как цитокин и ингибиторы фактора роста, иммунодепрессанты, противовоспалительные агенты, ингибиторы метаболизма, ингибиторы ферментов и/или цитотоксические или цитостатические агенты, более подробно описанные ниже. Кроме того, описанные здесь одно или более антител могут быть использованы в комбинации с двумя или более описанными здесь терапевтическими агентами. В такой комбинированной терапии могут быть преимущественно использованы более низкие дозы вводимых терапевтических агентов, что позволит избежать возможного продуцирования токсических эффектов или осложнений, ассоциированных с проведением различных курсов монотерапии.
В других вариантах осуществления изобретения, одно или более антител согласно изобретению могут быть приготовлены и/или введены вместе с одним или более противовоспалительными средствами, иммунодепрессантами или ингибиторами метаболизма или ингибиторами ферментов. Неограничивающими примерами лекарственных средств или ингибиторов, которые могут быть использованы в комбинации с описанными здесь антителами, являются, но не ограничиваются ими, одно или более из нижеследующих лекарственных средств, таких как нестероидное(ые) противовоспалительное(ые) лекарственное(ые) средство(а) (НСПВС), например, ибупрофен, тенидап, напроксин, мелоксикам, пироксикам, диклофенак и индометацин; сульфасалазин; кортикостероиды, такие как преднизолон; противовоспалительное(ые) лекарственное(ые) средство(а), ингибирующее(ие) действие цитокинов (CSAID); ингибиторы биосинтеза нуклеотидов, например, ингибиторы биосинтеза пурина; антагонисты фолата (например, метотрексат, (N-[4-[[(2,4-диамино-6-птеридинил)метил]метиламино]бензоил]-L-глутаминовая кислота); и ингибиторы биосинтеза пиримидина, например, ингибиторы дигидрооротат-дегидрогеназы (DHODH). Подходящими терапевтическими агентами, используемыми в комбинации с антителами согласно изобретению, являются НСПВС, CSAID, ингибиторы DHODH (например, лефлуномид) и антагонисты фолата (например, метотрексат).
Примерами дополнительных ингибиторов являются одно или более из нижеследующих соединений, таких как кортикостероиды (для перорального введения, введения путем ингаляции и местного введения); иммунодепрессанты, например, циклоспорин, такролимус (FK-506); и ингибиторы mTOR, например, сиролимус (рапамицин-RAPAMUNETM или производное рапамицина, например, растворимые производные рапамицина (например, производные сложного эфира рапамицина, например, CCI-779); агенты, блокирующие передачу сигнала провоспалительными цитокинами, такими как TNFα или IL-1 (например, IRAK, NIK, IKK, p38 или ингибиторы киназы MAP); ингибиторы COX2, например, целекоксиб, рофекоксиб и их варианты; ингибиторы фосфодиэстеразы, например, R973401 (ингибитор фосфодиэстеразы типа IV); ингибиторы фосфолипазы, например, ингибиторы цитозольной фосфолипазы 2 (cPLA2) (например, аналоги трифторметилкетона); ингибиторы васкулярного эндотелиального фактора клеточного роста или рецептора фактора роста, например, ингибитор VEGF и/или ингибитор VEGF-R; и ингибиторы ангиогенеза. Терапевтическими агентами, подходящими для использования в комбинации с антителами согласно изобретению, являются иммунодепрессанты, например, циклоспорин, такролимус (FK-506); и ингибиторы mTOR, например, сиролимус (рапамицин) или производные рапамицина, например, растворимые производные рапамицина (например, производные сложного эфира рапамицина, например, CCI-779); ингибиторы COX2, например, целекоксиб и его варианты, и ингибиторы фосфолипазы, например, ингибиторы цитозольной фосфолипазы 2 (cPLA2), например, аналоги трифторметилкетона.
Дополнительными примерами терапевтических агентов, которые могут быть использованы в комбинации с антителом согласно изобретению, являются одно или более соединений, таких как 6-меркаптопурины (6-MP); азатиоприн; сульфасалазин; мезалазин; олсалазин; хлорохин/гидроксихлорохин (PLAQUENIL®); пеницилламин; ауротиомалат (для внутримышечного и перорального введения); азатиопин; колхицин; агонисты адренорецептора бета-2 (сальбутамол, тербуталин, сальметерал); ксантины (теофиллин, аминофеллин); кромогликат; недокромил; кетотифен; ипратропий и окситропий; микофенолят мофетил; агонисты аденозина; антитромботические агенты; ингибиторы комплемента и адренергические агенты.
Все цитируемые здесь публикации и патентные документы во всей своей полноте и во всех целях вводятся в настоящее описание посредством ссылки так, как если бы каждая отдельная публикация или отдельный документ были конкретно и отдельно введены во всей своей полноте и во всех целях в описание настоящего изобретения. Цитирование любых публикаций и патентных документов, приведенных для их раскрытия до даты подачи, не должно быть истолковано как признание того, что авторы изобретения не имеют право датировать такие публикации и патентные документы задним числом на основании предшествующего изобретения. Описанное здесь изобретение представлено в виде текстового материала, и для специалиста в данной области очевидно, что настоящее изобретение может быть практически осуществлено в различных вариантах, и что приведенное выше описание и нижеследующие примеры представлены лишь в иллюстративных целях и не должны рассматриваться как ограничение объема изобретения, сформулированного в прилагаемой формуле изобретения.
Примеры
В приведенных здесь примерах используются последовательности, представленные ниже в таблице 7:
Таблица 7: Репрезентативные последовательности
Антитело scFv против человеческого CTLA-4:
ggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagctagc (SEQ ID NO: 229)
GGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 230)
Антитело scFv OKT3 против человеческого CD3 эпсилон:
ggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 231)
GGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 232)
Антитело 4D11v2 против человеческого антигена Jagged: тяжелая цепь
gaggtgcacctgttggagtctgggggaggcttggtacagcctggggggtccctgagactctcctgtgcagcctctggattcacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtgtcaagtattgacccggaaggtcggcagacatattacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaagacatcggcggcaggtcggcctttgactactggggccagggaaccctggtcaccgtctcctcagctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaa (SEQ ID NO:233)
EVHLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSSIDPEGRQTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDIGGRSAFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO:234)
Антитело 4D11v2 против человеческого антигена Jagged: легкая цепь
Gacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctatgcggcatccagtttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactactgtcaacagacggttgtggcgcctccgttattcggccaagggaccaaggtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgancaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 235)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQTVVAPPLFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLXKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 236)
Антитело 4D11 против человеческого антигена Jagged: тяжелая цепь
gaggtgcagctgttggagtctgggggaggcttggtacagcctggggggtccctgagactctcctgtgcagcctctggattcacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtgtcaagtattgacccggaaggtcggcagacatattacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaagacatcggcggcaggtcggcctttgactactggggccagggaaccctggtcaccgtctcctcagctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaa (SEQ ID NO: 237)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSSIDPEGRQTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDIGGRSAFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 238)
Активируемое анти-Jagged антитело 5342-1204-4D11v2: легкая цепь
caaggccagtctggccagtgcaatatttggctcgtaggtggtgattgcaggggctggcaggggggctcgagcggtggcagcggtggctctggtggtctgagcggccgttccgataatcatggcggcggttctgacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctatgcggcatccagtttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactactgtcaacagacggttgtggcgcctccgttattcggccaagggaccaaggtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 239)
QGQSGQCNIWLVGGDCRGWQGGSSGGSGGSGGLSGRSDNHGGGSDIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQTVVAPPLFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLXKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 240)
Мультиспецифическое анти-Jagged антитело 4D11v2 – анти-CD3 антитело OKT3: тяжелая цепь
GAGGTGCACCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTGTCAAGTATTGACCCGGAAGGTCGGCAGACATATTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGACATCGGCGGCAGGTCGGCCTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAAggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 241)
EVHLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSSIDPEGRQTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDIGGRSAFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 242)
Мультиспецифическое анти-Jagged антитело 4D11v2 – анти-CD3 антитело OKT3: легкая цепь
gacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctatgcggcatccagtttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactactgtcaacagacggttgtggcgcctccgttattcggccaagggaccaaggtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgancaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 243)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQTVVAPPLFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLXKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 244)
Мультиспецифическое активируемое анти-Jagged—анти-CD3 антитело 5342-1204-4D11v2—CD3 OKT3: легкая цепь
Caaggccagtctggccagtgcaatatttggctcgtaggtggtgattgcaggggctggcaggggggctcgagcggtggcagcggtggctctggtggtctgagcggccgttccgataatcatggcggcggttctgacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctatgcggcatccagtttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactactgtcaacagacggttgtggcgcctccgttattcggccaagggaccaaggtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgancaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 245)
QGQSGQCNIWLVGGDCRGWQGGSSGGSGGSGGLSGRSDNHGGGSDIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQTVVAPPLFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLXKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 246)
Мультиспецифическое анти-Jagged антитело 4D11v2 — анти-CTLA-4 антитело: тяжелая цепь
GAGGTGCACCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTGTCAAGTATTGACCCGGAAGGTCGGCAGACATATTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGACATCGGCGGCAGGTCGGCCTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAAggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagc (SEQ ID NO: 247)
EVHLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSSIDPEGRQTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDIGGRSAFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 248)
Мультиспецифическое анти-Jagged антитело 4D11v2 — анти-CTLA-4 антитело: легкая цепь
gacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctatgcggcatccagtttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactactgtcaacagacggttgtggcgcctccgttattcggccaagggaccaaggtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgancaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagctagc (SEQ ID NO: 249)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQTVVAPPLFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLXKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 250)
Мультиспецифическое активируемое анти-Jagged—анти-CTLA-4 антитело 5342-1204-4D11v2—CTLA-4: легкая цепь
Caaggccagtctggccagtgcaatatttggctcgtaggtggtgattgcaggggctggcaggggggctcgagcggtggcagcggtggctctggtggtctgagcggccgttccgataatcatggcggcggttctgacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctatgcggcatccagtttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactactgtcaacagacggttgtggcgcctccgttattcggccaagggaccaaggtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgancaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagctagc (SEQ ID NO: 251)
QGQSGQCNIWLVGGDCRGWQGGSSGGSGGSGGLSGRSDNHGGGSDIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQTVVAPPLFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLXKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 252)
Анти-EGFR антитело C225v5: тяжелая цепь
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgaactgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga (SEQ ID NO: 253)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSQDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 254)
Анти-EGFR антитело C225v5: легкая цепь
cagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 255)
QILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 256)
Анти-EGFR антитело C225v5: тяжелая цепь caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagcaacgataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagcgcgttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgaactgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaa (SEQ ID NO: 257)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSNDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 258)
Анти-EGFR антитело C225v5: легкая цепь
Ggcggtacccagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagcg (SEQ ID NO: 259)
DILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 260)
Анти-EGFR антитело C225v4: тяжелая цепь
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagcaacgataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgaactgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga (SEQ ID NO: 261)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSNDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 262)
Анти-EGFR антитело C225v5: легкая цепь
Cagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgttag (SEQ ID NO: 263)
QILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 264)
Анти-EGFR антитело C225v6: тяжелая цепь
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacgccagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgaactgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga (SEQ ID NO: 265)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSQDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 266)
Анти-EGFR антитело C225v6: легкая цепь
Cagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgttag (SEQ ID NO: 267)
QILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 268)
Активируемое анти-EGFR антитело 3954-1204-C225v5: легкая цепь:
[Спейсер (SEQ ID NO: 283)][маскирующая группа (SEQ ID NO: 284)][линкер 1 (SEQ ID NO: 285)][субстрат 1204 (SEQ ID NO: 286)][линкер 2 (SEQ ID NO: 287)][легкая цепь C225v5 (SEQ ID NO: 255)]

[Спейсер (SEQ ID NO: 46)][ маскирующая группа (SEQ ID NO: 77)][ линкер 1 (SEQ ID NO: 288)][субстрат 1204(SEQ ID NO: 26)][линкер 2 (SEQ ID NO: 289)][C225v5 (SEQ ID NO: 256)]

Мультиспецифическое анти-EGFR антитело C225v5 – анти-cd3 антитело OKT3: тяжелая цепь
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgaactgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaaggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 271)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSQDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 272)
Мультиспецифическое анти-EGFR антитело C225v5 – анти-cd3 антитело OKT3: легкая цепь
cagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 273)
QILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 274)
Мультиспецифическое активируемое анти-EGFR антитело C225v5 – анти-cd3 антитело 3954-1204-C225v5-OKT3: легкая цепь
caaggccagtctggccagtgcatctcacctcgtggttgtccggacggcccatacgtcatgtacggctcgagcggtggcagcggtggctctggtggatccggtctgagcggccgttccgataatcatggcagtagcggtacccagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactattgcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 275)
QGQSGQCISPRGCPDGPYVMYGSSGGSGGSGGSGLSGRSDNHGSSGTQILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINR (SEQ ID NO: 276)
Мультиспецифическое анти-EGFR антитело C225v5 – анти-CTLA-4 антитело: тяжелая цепь
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgaactgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaaggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagctagc (SEQ ID NO: 277)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSQDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 278)
Мультиспецифическое анти-EGFR антитело C225v5 – анти-CTLA-4 антитело: легкая цепь
cagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagctagc (SEQ ID NO: 279)
QILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 280)
Мультиспецифическое активируемое анти-EGFR антитело – анти- CTLA-4 антитело 3954-1204-C225v5-CTLA-4: легкая цепь
caaggccagtctggccagtgcatctcacctcgtggttgtccggacggcccatacgtcatgtacggctcgagcggtggcagcggtggctctggtggatccggtctgagcggccgttccgataatcatggcagtagcggtacccagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtgaaattgtgttgacacagtctccaggcaccctgtctttgtctccaggggaaagagccaccctctcctgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaacctggccaggctcccaggctcctcatctatggtgcatccagcagggccactggcatcccagacaggttcagtggcagtgggtctgggacagacttcactctcaccatcagcagactggagcctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctcactttcggcggagggaccaaggtggaaatcaaacgttccggagggtcgaccataacttcgtataatgtatactatacgaagttatcctcgagcggtacccaggtgcagctggtgcagactgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggatccacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgacaaactccctttactggtacttcgatctctggggccgtggcaccctggtcactgtctcttcagctagc (SEQ ID NO: 281)
QGQSGQCISPRGCPDGPYVMYGSSGGSGGSGGSGLSGRSDNHGSSGTQILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGECGGGSGGGGSGSGGGSGGGGSGGGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPLTFGGGTKVEIKRSGGSTITSYNVYYTKLSSSGTQVQLVQTGGGVVQPGRSLRLSCAASGSTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCATNSLYWYFDLWGRGTLVTVSSAS (SEQ ID NO: 282)
Пример 1: Получение мультиспецифических антител
В данном примере продемонстрированы конструирование, экспрессия и очистка мультиспецифических антитело против Jagged–CD3, против Jagged–CTLA-4, против EGFR–CD3 и против EGFR-CTLA-4.
Были использованы векторы для экспрессии тяжелой цепи анти-Jagged антитела (4D11v2), легкой цепи антитела 5342-1204-4D11v2, тяжелой цепи анти-EGFR антитела C225v5 и легкой цепи антитела 3954-1204-C225v5, последовательности которых указаны выше. Такие векторы описаны в одновременно рассматриваемых заявках: PCT/US2013/038540, поданной 26 апреля 2013 (озаглавленной «Activatable Antibodies That Bind Epidermal Growth Factor Receptor And Methods Of Use Thereof») и PCT/US2013/047109, поданной 21 июня, 2013 (озаглавленной «Анти-Jagged Antibodies, Activatable Анти-Jagged Antibodies and Methods of Use Thereof»), содержания которых во всей своей полноте вводятся в настоящее описание посредством ссылки.
Векторы были гидролизованы рестриктирующими ферментами NheI и NotI, и фрагмент вектора был выделен с помощью гель-электрофорезы. Вставки получали следующим образом. Фрагмент CH2CH3 человеческого IgG амплифицировали в реакции 1 из Pop Hygro 4D11v2 с использованием праймера HCForNhe (таблица 8, SEQ ID NO: 3) и праймера HCRevOL (таблица 8, SEQ ID NO: 4), анти-CTLA-4 scFv амплифицировали в реакции 2 из кДНК анти-CTLA-4 scFv (SEQ ID NO: 229) с использованием праймера CTRevNot (таблица 8, SEQ ID NO: 1) и праймера CTForOL (таблица 8, SEQ ID NO: 2), scFv OKT3 амплифицировали в реакции 3 из кДНК scFv OKT3 (SEQ ID NO: 231) с использованием праймера OKRevNot (Table 8, SEQ ID NO: 5) и праймера CTForOL (таблица 8, SEQ ID NO: 2). Гибриды человеческого IgG, CH2CH3/CTLA-4 scFv, получали путем объединения 10% продукта реакции 1, 10% продукта реакции 2 и амплификации с использованием праймеров HCForNhe и CTRevNot.
Последовательности праймеров
После амплификации, полученную ДНК гидролизовали рестриктирующими ферментами NheI и NotI, и гибридную ДНК CH2CH3/CTLA-4 scFv выделяли с помощью гель-электрофореза. Гибриды человеческого IgG, CH2CH3/CTLA-4 scFv, получали путем объединения 10% продукта реакции 1, 10% продукта реакции 2 и амплификации с использованием праймеров HCForNhe и CTRevNot. После амплификации, полученную ДНК гидролизовали рестриктирующими ферментами NheI и NotI, и гибридную ДНК CH2CH3/CTLA-4 scFv выделяли с помощью гель-электрофореза.
Для вставки фрагментов CH2CH3/scFv в экспрессионные векторы, нижеследующие комбинации, представленные в таблице 9, лигировали в течение ночи с ДНК-лигазой T4 (Invitrogen Inc., Carlsbad, CA). После лигирования, ДНК переносили в штамм MC106 E. coli и отбирали на резистентность к ампициллину. В результате секвенирвоания ДНК были идентифицированы клоны, содержащие ДНК-вставки, кодирующие нужные мультиспецифические антитела или мультиспецифические активируемые антитела мультиспецифические активируемые антитела, а затем получали ДНК для ее трансфекции в клетки млекопитающих.
Реакции лигирования
Пример 2: Продуцирование мультиспецифических антител и мультиспецифических активируемых антител
Полностью человеческие IgG экспрессировали из транзиентно трансфецированных клеток HEK-293. Ко-трансфекция отдельными векторами, экспрессирующими тяжелую и легкую цепи, как показано в таблице 10, позволяла экспрессировать мультиспецифические активируемые антитела.
Мультиспецифические антитела и мультиспецифические активируемые антитела, экспрессируемые в клетках HEK-293, очищали с помощью хроматографии на белке A.
Анализы на связывание: Как показано на фигуре 11, ELISA-эксперименты на связывание продемонстрировали, что мультиспецифические антитела и мультиспецифические активируемые антитела против Jagged—CTLA-4 и Jagged—OKT3 связывались с человеческим Jagged 1, а мультиспецифические антитела и мультиспецифические активируемые антитела против EGFR—CTLA-4 и против EGFR—OKT3 специфически связывались с человеческим EGFR.
Человеческий Jagged 1-Fc (R&D Systems; Cat # 1277-JG-050) и человеческий EGFR (R&D Systems, Cat # 344-ER-050) адсорбировали на различных лунках 96-луночного ELISA-планшета. Очищенные анти-Jagged—CTLA, анти-Jagged—OKT3, анти-EGFR—CTLA или анти-EGFR—OKT3 антитела наносили на планшет и оставляли для связывания. Связанное антитело визуализировали с использованием конъюгата антитела против человеческого IgG-ПХ (Fab-специфического, Sigma, St Louis, MO; Cat # A0293-1ML) и проявляли с использованием хромогенного субстрата TMB.
Как показано на фигурах 12A и 12B, ELISA-эксперименты на связывание продемонстрировали, что анти-Jagged—CTLA-4 и анти-EGFR—CTLA-4 антитела специфически связывались с человеческим и мышиным CTLA-4 (антитело против человеческого CTLA-4 перекрестно реагировало с мышиным и человеческим CTLA-4): Человеческий CTLA-4 (R&D Systems; Cat # 325-CT-200/CF) или мышиный CTLA-4 (R&D Systems, Cat # 434-CT-200/CF) адсорбировали на лунки отдельных 96-луночных ELISA-планшетов. Очищенные анти-Jagged антитело 4D11v2—CTLA-4, активируемое анти-Jagged антитело 5342-1204-4D11v2—CTLA-4, анти-EGFR антитело C225v5—CTLA-4 или активируемое антитело 3954-1204-C225v5—CTLA-4 наносили на планшет и оставляли для связывания. Связанное антитело визуализировали с использованием конъюгата антитела против человеческого IgG-ПХ (Fab-специфического, Sigma, St Louis, MO; Cat # A0293-1ML) и проявляли с использованием хромогенного субстрата TMB. На специфическое связывание указывала неспособность гибрида антитело-OKT3 специфически связываться с человеческим CD3ε.
Пример 3: Связывание OKT3 с CD3ε
Как показано на фигуре 13, ELISA-эксперименты на связывание продемонстрировали, что мультиспецифическое активируемое анти-EGFR антитело 3954-1204-C225v5—OKT3 и мультиспецифическое активируемое анти-Jagged антитело 5342-1204-4D11v2—OKT3 специфически связывались с человеческим CD3ε. Человеческий CD3ε (NovoProtein, Cat# C578) адсорбировали на лунки 96-луночного ELISA-планшета. Очищенные мультиспецифическое активируемое анти-EGFR антитело 3954-1204-C225v5—CTLA-4, мультиспецифическое активируемое анти-EGFR антитело 3954-1204-C225v5—OKT3, мультиспецифическое активируемое анти-Jagged антитело 5342-1204-4D11v2—CTLA-4 или мультиспецифическое активируемое анти-Jagged антитело 5342-1204-4D11v2—OKT3 наносили на планшет и оставляли для связывания. Связанное антитело визуализировали с использованием конъюгата антитела против человеческого IgG-ПХ (Fab-специфического, Sigma, St Louis, MO; Cat # A0293-1ML) и проявляли с использованием хромогенного субстрата TMB.
Пример 4: Активация мультиспецифических активируемых антител протеазой
Активация
Мультиспецифические антитела и/или мультиспецифические активируемые антитела разводили в PBS до конечной концентрации 0,8 мг/мл. Рекомбинантн человеческий uPA (R&D Systems, Catalog 1310-SE) добавляли до конечной концентрации 700 нM и инкубировали при 37°C в течение ~20 часов. Затем брали аликвоты гидролизата и prepared for SDS-PAGE analysis как описано ниже, as described below, и разводили до 100 нM в PBS; 0,05% Твин 20 и 10 мг/мл альбумина бычьей сыворотки для проведения ELISA-анализов на связывание.
Образцы для электрофореза в ПААГ денатурировали при 70°C в течение 10 минут в 1x буфере для образцов LDS, и перед проведением электрофореза добавляли TCEP до конечной концентрации 40 мМ. 6 мкг антитела загружали в 10% бис-трис-гель NuPAGE (Invitrogen), и белки разделяли по размеру с использованием буфера для электрофореза MOPS. После проведения электрофореза, гель окрашивали кумасси синим и полученные результаты были представлены на фигуре 14. Изменение подвижности легких цепей активируемого анти-EGFR антитела 3954-1204-C225v5, мультиспецифического активируемого анти-EGFR антитела 3954-1204-C225v5—CTLA-4, активируемого анти-Jagged антитела 5342-1204-4D11v2 и мультиспецифического активируемого анти-Jagged антитела 5342-1204-4D11v2—CTLA-4 в присутствии uPA указывало на протеолитическую активацию мультиспецифических активируемых антител. Отсутствие какого-либо изменения подвижности гибридов тяжелых цепей указывало на резистентность к расщеплению протеазой.
Анализы на связывание
На панели A фигуры 15 показано, что в ELISA-анализе, мультиспецифическое активируемое анти-EGFR антитело 3954-1204-C225v5—CTLA-4 связывалось с EGFR с меньшей аффинностью (Kd=12,8 нM) по сравнению с аффинностью связывания с C225v5 (3 нМ) или с C225v5—CTLA-4 (0,33 нМ). Однако, после uPA-активации, мультиспецифическое активируемое анти-EGFR антитело 3954-1204-C225v5—CTLA-4 связывалось с EGFR с такой же аффинностью (0,45 нМ) как и C225v5 (0,21 нМ) и C225v5—CTLA4 (0,33 нМ). Важно отметить, что uPA не расщеплял анти-CTLA4 scFv, присоединенный к карбокси-концу тяжелой цепи. Как проиллюстрировано на панели В фигуры 15, uPA-обработка не влияла на связывание с человеческим CTLA4.
Аналогичным образом, ELISA-анализ показал, что мультиспецифическое активируемое анти-Jagged антитело 5342-1204-4D11v2—CTLA-4 связывалось с человеческим антигеном Jagged 1 с меньшей аффинностью (15 нМ) по сравнению с аффинностью связывания с 4D11v2 (0,44 нМ) или 4D11v2—CTLA4 (0,77 нМ). После активации, мультиспецифическое активируемое анти-Jagged антитело 5342-1204-4D11v2—CTLA-4 связывалось с человеческим антигеном Jagged 1 с такой же аффинностью, как и 4D11v2 (0,54 нМ) и 4D11v2—CTLA4 (0,92 нМ) (фигура 15, панель C). uPA-обработка не влияла на связывание с CTLA4 (фигура 15, панель D). ELISA-анализы на связывание осуществляли следующим образом. Человеческий Jagged 1-Fc (R&D Systems; Cat # 1277-JG-050), человеческий EGFR (R&D Systems, Cat#344-ER-050) или человеческий CTLA4 (R&D Systems; Cat # 325-CT-200/CF) адсорбировали на лунки 96-луночного ELISA-планшета. Трехкратные разведения, начиная со 100 нМ, необработанных и uPA-обработанных антител, мультиспецифических антител или мультиспецифических активируемых антител наносили на планшет и оставляли на 1 час для ассоциации с антигеном, связанным с планшетом. После связывания, связанное антитело визуализировали с использованием конъюгата антитела против человеческого IgG-ПХ (Fab-специфического, Sigma, St Louis, MO; Cat # A0293-1ML) и проявляли с использованием хромогенного субстрата TMB.
Пример 5: Последовательности дополнительных мультиспецифических антител и мультиспецифических активируемых антител
В этом примере описаны последовательности дополнительных мультиспецифических антител и мультиспецифических активируемых антител.
Антитело OKT3m scFv
Аминокислотная последовательность
qvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr (SEQ ID NO: 306)
нуклеотидная последовательность
caggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 307)
Антитело OKT3m scFv Lv
Аминокислотная последовательность
qivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr (SEQ ID NO: 308)
нуклеотидная последовательность
cagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgt (SEQ ID NO: 309)
Антитело OKT3m scFv Hv
Аминокислотная последовательность
qvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvss (SEQ ID NO: 310)
нуклеотидная последовательность
caggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagc (SEQ ID NO: 311)
Антитело OKT3m scFv с длинным линкером присоединяли к C-концу легкой цепи (HC) или легкой цепи (LC) антитела или активируемого антитела для продуцирования мультиспецифического антитела или активируемого антитела, соответственно
Аминокислотная последовательность
Gggsggggsgsgggsggggsgggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr* (SEQ ID NO: 290)
нуклеотидная последовательность
ggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttga (SEQ ID NO: 291)
Антитело OKT3m scFv с коротким линкером присоединяли к N-концу HC антитела или активируемого антитела для продуцирования мультиспецифического антитела или активируемого антитела, соответственно
Аминокислотная последовательность
qvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinrggggs (SEQ ID NO: 292)
нуклеотидная последовательность
caggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgtggaggtggtggatcc (SEQ ID NO: 293)
Тяжелая цепь мультиспецифического антитела C225v5-OKT3m-H-N (антитела OKT3m, присоединенного к N-концу антитела C225v5 HC)
Аминокислотная последовательность
qvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinrggggsqvqlkqsgpglvqpsqslsitctvsgfsltnygvhwvrqspgkglewlgviwsggntdyntpftsrlsinkdnsksqvffkmnslqsqdtaiyycaraltyydyefaywgqgtlvtvsaastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk* (SEQ ID NO: 294)
нуклеотидная последовательность
caggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgtggaggtggtggatcccaggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga (SEQ ID NO: 295)
Тяжелая цепь мультиспецифического антитела C225v5-OKT3m-H-С (антитела OKT3m, присоединенного к N-концу антитела C225v5 HC)
Аминокислотная последовательность
qvqlkqsgpglvqpsqslsitctvsgfsltnygvhwvrqspgkglewlgviwsggntdyntpftsrlsinkdnsksqvffkmnslqsqdtaiyycaraltyydyefaywgqgtlvtvsaastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkgggsggggsgsgggsggggsgggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr* (SEQ ID NO: 296)
нуклеотидная последовательность
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaaggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttga (SEQ ID NO: 297)
Легкая цепь мультиспецифического антитела C225v5-OKT3m-L-С (антитела OKT3m, присоединенного к С-концу антитела C225v5 LC)
Аминокислотная последовательность
Dilltqspvilsvspgervsfscrasqsigtnihwyqqrtngsprllikyasesisgipsrfsgsgsgtdftlsinsvesediadyycqqnnnwpttfgagtklelkrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgecgggsggggsgsgggsggggsgggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr* (SEQ ID NO: 298)
нуклеотидная последовательность
GACatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttag (SEQ ID NO: 299)
Тяжелая цепь мультиспецифического антитела C225v5(N297Q)-OKT3m-H-N
Аминокислотная последовательность
qvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinrggggsqvqlkqsgpglvqpsqslsitctvsgfsltnygvhwvrqspgkglewlgviwsggntdyntpftsrlsinkdnsksqvffkmnslqsqdtaiyycaraltyydyefaywgqgtlvtvsaastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqyqstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk* (SEQ ID NO: 300)
нуклеотидная последовательность
caggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgtggaggtggtggatcccaggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtaccagagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga (SEQ ID NO: 301)
Тяжелая цепь мультиспецифического антитела C225v5(N297Q)-OKT3m-H-С
Аминокислотная последовательность
qvqlkqsgpglvqpsqslsitctvsgfsltnygvhwvrqspgkglewlgviwsggntdyntpftsrlsinkdnsksqvffkmnslqsqdtaiyycaraltyydyefaywgqgtlvtvsaastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqyqstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkgggsggggsgsgggsggggsgggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr* (SEQ ID NO: 302)
нуклеотидная последовательность
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtaccagagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaaggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttga (SEQ ID NO: 303)
Легкая цепь мультиспецифического антитела C225v5(N297Q)-OKT3m-L-C
Аминокислотная последовательность
Dilltqspvilsvspgervsfscrasqsigtnihwyqqrtngsprllikyasesisgipsrfsgsgsgtdftlsinsvesediadyycqqnnnwpttfgagtklelkrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgecgggsggggsgsgggsggggsgggqvqlqqsgaelarpgasvkmsckasgytftrytmhwvkqrpgqglewigyinpsrgytnynqkfkdkatlttdkssstaymqlssltsedsavyycaryyddhysldywgqgttltvssggggsggggsggggsqivltqspaimsaspgekvtmtcsasssvsymnwyqqksgtspkrwiydtsklasgvpahfrgsgsgtsysltisgmeaedaatyycqqwssnpftfgsgtkleinr* (SEQ ID NO: 298)
нуклеотидная последовательность
GACatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttag (SEQ ID NO: 299)
Тяжелая цепь антитела C225v5(N297Q)
Аминокислотная последовательность
qvqlkqsgpglvqpsqslsitctvsgfsltnygvhwvrqspgkglewlgviwsggntdyntpftsrlsinkdnsksqvffkmnslqsqdtaiyycaraltyydyefaywgqgtlvtvsaastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqyqstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk* (SEQ ID NO: 336)
нуклеотидная последовательность
caggtgcagctgaaacagagcggcccgggcctggtgcagccgagccagagcctgagcattacctgcaccgtgagcggctttagcctgaccaactatggcgtgcattgggtgcgccagagcccgggcaaaggcctggaatggctgggcgtgatttggagcggcggcaacaccgattataacaccccgtttaccagccgcctgagcattaacaaagataacagcaaaagccaggtgttttttaaaatgaacagcctgcaaagccaggataccgcgatttattattgcgcgcgcgcgctgacctattatgattatgaatttgcgtattggggccagggcaccctggtgaccgtgagcgcggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtaccagagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga (SEQ ID NO: 337)
Легкая цепь активируемого антитела 3954-1204-C225v5
Аминокислотная последовательность
qgqsgqcisprgcpdgpyvmygssggsggsggsglsgrsdnhgssgtqilltqspvilsvspgervsfscrasqsigtnihwyqqrtngsprllikyasesisgipsrfsgsgsgtdftlsinsvesediadyycqqnnnwpttfgagtklelkrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec* (SEQ ID NO: 304)
нуклеотидная последовательность
caaggccagtctggccagtgcatctcacctcgtggttgtccggacggcccatacgtcatgtacggctcgagcggtggcagcggtggctctggtggatccggtctgagcggccgttccgataatcatggcagtagcggtacccagatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttag (SEQ ID NO: 305)
Мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N, которое включает легкую цепь, кодируемую плазмидой 3954-1204-C225, и тяжелую цепь, кодируемую плазмидой C225v5-OKT3m-H-N.
Легкая цепь антитела C225v5
Аминокислотная последовательность
Dilltqspvilsvspgervsfscrasqsigtnihwyqqrtngsprllikyasesisgipsrfsgsgsgtdftlsinsvesediadyycqqnnnwpttfgagtklelkrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec* (SEQ ID NO: 314)
нуклеотидная последовательность
GACatcttgctgacccagagcccggtgattctgagcgtgagcccgggcgaacgtgtgagctttagctgccgcgcgagccagagcattggcaccaacattcattggtatcagcagcgcaccaacggcagcccgcgcctgctgattaaatatgcgagcgaaagcattagcggcattccgagccgctttagcggcagcggcagcggcaccgattttaccctgagcattaacagcgtggaaagcgaagatattgcggattattattgccagcagaacaacaactggccgaccacctttggcgcgggcaccaaactggaactgaaacgtacggtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtggaggtggatctggaggtggcggttcaggctctggcggaggctcaggtggtggaggatcaggcggaggtcaggttcagctgcagcagagcggtgcagaactggcacgtccgggtgcaagcgttaaaatgagctgtaaagcaagcggttatacctttacccgttataccatgcattgggttaaacagcgtccgggtcagggtctggaatggattggttatatcaatccgagccgtggttataccaactacaaccagaaattcaaagataaagcaaccctgaccaccgataaaagcagcagcaccgcctatatgcagctgagcagcctgacctcagaggatagcgcagtttattactgtgcacgctattatgatgatcactatagcctggattattggggtcagggcaccaccctgaccgttagcagcggtggtggtggtagtggtggcggtggttcaggcggtggcggtagccagattgttctgacccagagtccggcaattatgagcgcaagtccgggtgaaaaagttaccatgacctgtagcgccagcagcagcgttagctatatgaattggtatcagcagaaaagcggcaccagcccgaaacgttggatttatgataccagcaaactggcaagcggtgttccggcacattttcgtggtagcggtagcggcacctcatatagcctgaccattagcggtatggaagcagaagatgcagcaacctattattgtcagcagtggtcaagcaatccgtttacctttggtagtggcaccaaactggaaattaatcgttag (SEQ ID NO: 315).
В таблице 11 представлены примеры объединения последовательностей тяжелой цепи (HC) и легкой цепи (LC) для получения мультиспецифического антитела или мультиспецифического активируемого антитела согласно изобретению. Следует отметить, что при употреблении названий легких цепей, легкие цепи антител, содержащие C225v5 также называются здесь легкими цепями, содержащими C225.
Мультиспецифические антитела и мультиспецифические активируемые антитела согласно изобретению
LC C225
LC C225
LC C225-OKT3m-L-C
LC C225-3954-1204
LC C225-3954-1204
LC C225
LC C225
LC C225-OKT3m-L-C
LC C225-3954-1204
LC C225-3954-1204
LC C225-3954-1204-OKT3m-L-C
Пример 6: Связывание мультиспецифических антител с CD3ε на Т-клетках Jurkat
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению связываться с Т-клетками.
Были протестированы три формата мультиспецифических антител: (1) C225v5-OKT3m-H-N, мультиспецифическое антитело, в котором scFv OKT3m (которое связывается с CD3ε (также называемое здесь CD3e и CD3) на Т-клетках) был присоединен к N-концу тяжелой цепи анти-EGFR антитела C225v5; (2) C225v5-OKT3m-H-C, мультиспецифическое антитело, в котором scFv OKT3m был присоединен к С-концу тяжелой цепи анти-EGFR антитела C225v5; и (3) C225v5-OKT3m-L-C, мультиспецифическое антитело, в котором scFv OKT3m был присоединен к С-концу легкой цепи анти-EGFR антитела C225v5. Аминокислотные последовательности этих мультиспецифических антител согласно изобретению представляют собой аминокислотные последовательности антител C225v5 и OKT3. Было также протестировано антитело контрольного изотипа, а именно, человеческий IgG1 контрольного изотипа, Enzo, Catalog ALX-804-133-C100.
Для того, чтобы определить, могут ли мультиспецифические антитела этих трех форматов связываться с CD3ε-позитивными Т-клетками Jurkat (также называемыми здесь клетками Jurkat и Jurkats), проводили анализ на связывание с помощью проточной цитометрии. Т-клетки Jurkat (клон E6-1, ATCC, TIB-152) культивировали в среде RPMI-1640, содержащий GlutaMAX™ (Life Technologies, Catalog 72400-120), 10% термоинактивированную фетальную бычью сыворотку (HI-FBS, Life Technologies, Catalog 10438-026), 100 ед./мл пенициллина и 100 мкг/мл стрептомицина (Life Technologies, Catalog 15140-122) (также называемой здесь полной средой) в соответствии с инструкциями ATCC. Клетки собирали путем центрифугирования (200xg, 4°C, 5 минут) и ресуспендировали в PBS с 2% HI-FBS (FACS-буфер). Приблизительно 250000 клеток Jurkat на лунку переносили в 96-луночный планшет с U-образным дном, собирали и ресуспендировали в 50 микролитрах (также сокращенно обозначаемых здесь мкл) тестируемых антител. Исходная концентрация мультиспецифических антител составляла 100 нМ, а концентрация антител контрольного изотипа составляла 166,7 нМ, и каждое антитело затем подвергали 5-кратному разведению с получением всего 8 концентраций.
Клетки и антитела инкубировали при 4°C со встряхиванием приблизительно в течение 1 часа, собирали и три раза промывали 200 мкл FACS-буфера. Полученные клетки Jurkat ресуспендировали в 50 мкл AlexaFluor® 647-конъюгированного антитела против человеческого IgG (H+L) (Jackson ImmunoResearch, Catalog 709-606-149) и инкубировали при 4°C со встряхиванием приблизительно в течение 30 минут. Полученные клетки Jurkat собирали, 3 раза промывали 200 мкл FACS-буфера и ресуспендировали в конечном объеме 150 мкл FACS-буфера с 2,5 мкг/мл 7-AAD (BD Pharmigen, Catalog 559925). Образцы анализировали на проточном цитометре BD Accuri C6 (BD Biosciences) и среднюю интенсивность флуоресценции (MFI) жизнеспособных клеток вычисляли с помощью FlowJo V10 (Treestar). 7-AAD-окрашивание показало, что боковое окно прямого рассеяния было достаточным для идентификации жизнеспособных клеток. Величины EC50 вычисляли с помощью GraphPad Prism 6 путем построения кривой зависимости логарифмических данных log (агонист) - ответ (по трем параметрам).
На фигуре 16A прнодемонстрировано, что эти мультиспецифические антитела всех трех форматов связывались с Т-клетками Jurkat при концентрациях EC50, составляющих в пределах от однозначных целых величин нМ до дробных величин нМ.
Пример 7: Мультиспецифические антитела связываются с CD3ε-экспрессирующими T-клетами и рекомбинантным человеческим EGFR
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению связываться с Т-клетками и клетками, содержащими антигены мишени (эти антитела также называются здесь антителами, связывающимися с двумя мишенями).
Для того, чтобы определить, может ли мультиспецифическое антитело C225v5-OKT3m-H-N, C225v5-OKT3m-H-C или C225v5-OKT3m-L-C связываться с CD3ε-позитивными Т-клетками Jurkat и EGFR, проводили проточный цитометрический анализ на связывание с двумя мишенями. Клетки Jurkat предварительно обрабатывали насыщающей концентрацией каждого из мультиспецифических антител, а затем титровали с использованием биотинилированного EGFR и детектировали с использованием конъюгата стрептавидина и R-фикоэритрина (SAPE). Было также протестировано антитело контрольного изотипа, а именно, человеческий IgG1 контрольного изотипа, Enzo, Catalog ALX-804-133-C100.
Клетки Jurkat культивировали и собирали как описано выше. Приблизительно 500000 клеток Jurkat на лунку переносили в планшет с U-образным дном, собирали и ресуспендировали в 50 мкл 40 нМ мультиспецифического антитела или 67 нМ антитела контрольного изотипа. Клетки инкубировали при 4°C со встряхиванием приблизительно в течение 1 часа, собирали и 3 раза промывали 200 мкл FACS-буфера. Полученные клетки Jurkat (связывающиеся с мультиспецифическим антителом согласно изобретению) ресуспендировали в 50 мкл биотинилированного рекомбинантого человеческого белка EGFR (Abcam, Catalog ab168702), начиная приблизительно с концентрации 290 нМ, и каждое антитело затем подвергали 5-кратному разведению с получением всего 8 концентраций. Клетки инкубировали при 4°C со встряхиванием приблизительно в течение 1 часа, собирали и 3 раза промывали 200 мкл FACS-буфера. Полученные клетки Jurkat ресуспендировали в 50 мкл 10 мкг/мл конъюгата стрептавидина и R-фикоэритрина (Life Technologies, S866) и инкубировали при 4°C со встряхиванием приблизительно в течение 1 часа. Клетки собирали, один раз промывали 200 мкл FACS-буфера и ресуспендировали в 150 мкл FACS-буфера. Образцы анализировали на BD Accuri C6 и MFI вычисляли как описано выше. Величины EC50 вычисляли с помощью GraphPad Prism 6, как описано выше.
На фигуре 16В прнодемонстрировано, что эти мультиспецифические антитела всех трех форматов связывались с EGFR в зависимости от концентрации и одновременно связывались с CD3ε на Т-клетках. Концентрации EC50 составляли в пределах от дробных величин нМ до однозначных целых величин нМ.
Пример 8: Мишень-зависимая активация Т-клеток мультиспецифическими антителами
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению активировать Т-клетки по мишень-зависимому механизму.
Для того, чтобы определить, может ли связывание мультиспецифических антител C225v5-OKT3m-H-N, C225v5-OKT3m-H-C или C225v5-OKT3m-L-C с Т-клетками Jurkat и с EGFR-позитивными клетками-мишенями индуцировать активацию клеток Jurkat, был проведен проточный цитометрический анализ. Клетки Jurkat и CFSE-меченные клетки-мишени совместно культивировали в течение приблизительно 15 часов, и активацию Т-клеток оценивали по окрашиванию на экспрессию маркера ранней активации Т-клеток, присутствующиего на поверхности Т-клеток, CD69.
EGFR-позитивные клетки SW480, также называемые здесь клетками SW480 (ATCC, Catalog CCL-228), и EGFR-негативные клетки U266, также называемые здесь клетками U266 (ATCC, Catalog TIB-196), были культивированы в среде RPMI-1640, содержащей GlutaMAX™, 10% HI-FBS, 100 ед./мл пенициллина и 100 мкг/мл стрептомицина (называемой здесь полной средой) в соответствии с инструкциями ATCC. Эти клетки-мишени собирали, один раз промывали PBS и ресуспендировали в PBS при плотности 2x106 клеток/мл. 5 мМ маточного раствора CFSE stock (Life Technologies, CellTrace™ CFSE Cell Proliferation Kit, Catalog C34554) приготавливали в ДМСО, а затем разводили в PBS до 30 нМ. Этот рабочий раствор PBS/CFSE использовали сразу после его приготовления. Равные объемы клеток-мишеней и маточного раствора CFSE объединяли до конечной концентрации CFSE 15 нМ и конечной плотности 1×106 клеток/мл. Клетки инкубировали в течение 15 минут при 37°C. Реакцию мечения гасили добавлением равного объема HI-FBS. Инкубированные клетки-мишени собирали, один раз промывали полной средой и ресуспендировали в полной среде при 5×105 клеток/мл. 50 мкл клеточной суспензии на лунку добавляли в 96-луночный плоскодонный планшет и получали всего 25000 клеток-мишеней на лунку.
Т-клетки Jurkat культивировали, как описано выше. Клетки Jurkat собирали, ресуспендировали в полной среде при плотности 1×106 клеток/мл, а затем в плоскодонный планшет, содержащий клетки-мишени, добавляли 50 мкл клеточной суспензии на лунку и получали всего приблизительно 50000 клеток Jurkat на лунку. Отношение эффекторных Т-клеток к клеткам-мишеням составяло 2:1.
Трехкратно концентрированные маточные растворы антитела получали в полной среде. Самая высокая из используемых концентраций составляла 1,2 нМ для мультиспецифических антител каждого формата и 1,5 нМ для антитела OKT3 (BioLegend, Catalog 317304), антитела C225v5 (аминокислотная последовательность которого приводится в данном описании) и антител контрольного изотипа. Затем каждое антитело подвергали 5-кратному разведению в полной среде с получением всего 8 концентраций. 50 мкл антитела на лунку добавляли в плоскодонный планшет, содержащий Т-клетки и клетки-мишени, и все маточные растворы 3-кратно разводили, начиная с концентрации 400 пM для мультиспецифического антитела каждого формата или 500 пM для антител OKT3, C225v5 и контрольного изотипа.
Приблизительно через 15 часов, клетки переносили в планшет с U-образным дном и собирали. Для максимизации выхода, планшет для культивирования клеток промывали 150 мкл FACS-буфера и промывку переносили в планшет с U-образным дном. Клетки собирали, ресуспендировали в 50 мкл ФЭ-конъюгированного анти-CD69 антитела (BD Biosciences, Catalog 555531, используемого в концентрации, рекомендованной производителями) и окрашивали в течение 1 часа при 4°C со встряхиванием. Клетки один раз промывали 200 мкл FACS-буфера, а затем ресуспендировали в конечном объеме 150 мкл. Монохроматический контроль использовали для серийной компенсации на BD Accuri C6. 10000 клеток собирали в CFSE-негативном боковом окне прямого рассеяния и вычисляли MFI с использованием FlowJo. Величины EC50 вычисляли с помощью GraphPad Prism 6 как описано в настоящей заявке.
На фигуре 17 продемонстрировано, что наиболее сильная CD69-активация, наблюдаемая начиная с суб-пМ-концентраций, зависела от формата каждого мультиспецифического антитела, культивированного в присутствии Т-клеток Jurkat и EGFR-позитивных клеток SW480. В противоположность этому, активация T-клеток, индуцированная при совместном культивировании с EGFR-негативными клетками U266, была значительно менее эффективной, то есть, выражалась в суб-нМ-величинах EC50, и такое снижение составляло 75% от максимального CD69-индуцирования по сравнению с активацией, индуцированной EGFR-позитивными клетками. Незвасисимая от EGFR активация мультиспецифических антител, наблюдаемая при использовании клеток U266, была аналогична активации антитела OKT3, а в случае антител C225v5 и антител контрольного изотипа, CD69-окрашивание было минимальным.
Пример 9: Мишень-зависимая активация первичных CD8+-T-клеток под действием мультиспецифических антител
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению активировать первичные CD8-позитивные (CD8+) Т-клетки.
Для того, чтобы определить, может ли мультиспецифическое антитело C225v5-OKT3m-H-N, C225v5-OKT3m-H-C или C225v5-OKT3m-L-C опосредовать EGFR-зависимую активацию первичных человеческих CD8+-T-клеток, был проведен проточный цитометрический анализ. CD8+-T-клетки, происходящие от человеческих мононуклеарных клеток периферической крови (МКПК), и DDAO-SE-меченные клетки-мишени совместно культивировали в течение ночи, а затем оценивали активацию по окрашиванию на маркер ранней активации, CD69.
EGFR-позитивные клетки SW480 культивировали и метили как описано выше, за исключением того, что: (1) клетки-мишени SW480 метили CellTrace™ FarRed DDAO-SE (Life Technologies, Catalog C34553) и (2) в полную среду добавляли 25 ед./мл IL-2 (R&D Systems, Catalog 202-IL-050/CF). 50 мкл суспензии клеток-мишеней (при плотности 5×105 клеток/мл) на лунку добавляли в 96-луночный плоскодонный планшет и получали всего 25000 клеток-мишеней на лунку.
Свежие цитотоксические CD8+-T-клетки нормальной периферической крови (AllCells, Catalog PB009-3) получали от AllCells (Alameda, CA), собирали путем центрифугирования (200xg, при комнатной температуре, в течение 15 минут) и ресуспендировали в полной среде, в которую было добавлено 250 ед./мл IL-2, при плотности 1,5×106 клеток/мл. 50 мкл клеточной суспензии на лунку добавляли в плоскодонный планшет, содержащий клетки-мишени, и получали всего приблизительно 75000 цтиттоксических CD8+-T-клеток, где отношение T-клеток к клеткам-мишеням составляло 3:1.
Трехкратно концентрированные маточные растворы антител получали в полной среде, в которую было добавлено 250 ед/мл IL-2. Самая высокая из используемых концентраций составляла 600 пМ, и каждое антитело подвергали 5-кратному серийному разведению с получением всего 8 концентраций. 50 мкл антитела на лунку добавляли в плоскодонный планшет, содержащий CD8+-клетки и клетки-мишени, и все маточные растворы 3-кратно разводили, начиная с концентрации 200 пM.
После инкубирования в течение ночи, плоскодонный планшет центрифугировали, и брали 100 мкл супернатанта для проведения описанного здесь люминесцентного анализа на цитотоксичность. Оставшийся супернатант переносили в планшет с U-образным дном, и клетки, находящиеся в плоскодонном планшете, отделяли с использованием 0,25% трипсина (Life Technologies, Catalog 25200-056). Трипсиновую активность гасили добавлением 3 объемов FACS-буфера, и клеточную суспензию переносили в планшет с U-образным дном. После сбора, клетки ресуспендировали в 50 мкл смеси ФЭ-конъюгированного анти-CD69 антитела/ФИТЦ-конъюгированного анти-CD8 антитела (anti-CD8 FITC, BD Biosciences, Catalog 561948), ФИТЦ-конъюгированного антитела контрольного изотипа (BD BioSciences, Catalog 340755) или ФЭ-конъюгированного антитела контрольного изотипа (BD BioSciences, Catalog 340761). Все антитела использовали в концентрациях, рекомендованных производителями. Клетки окрашивали в течение 1 часа при 4°C со встряхиванием, собирали и ресуспендировали в конечном объеме 150 мкл FACS-буфера с 2,5 мкг/мл 7-AAD. Монохроматический контроль использовали для серийной компенсации на BD Accuri C6, и собирали клеточную суспензию в фиксированном объеме. Дискриминационное окно для CD8+-клеток было достаточным для дифференциации клеток-мишеней и жизнеспособных T-клеток. Активацию количественно оценивали как выраженное в процентах увеличение числа Т-клеток, экспрессирующих анти-CD69 антитело, по сравнению с клетками, экспрессирующими ФЭ-конъюгированое антитело контрольного изотипа.
На фигуре 18A продемонстрировано, что эти мультиспецифические антитела всех трех форматов индуцировали зависимую от концентрации активацию первичных CD8+-T-клеток при величнах EC50 в пределах однозначных значений пМ. Обработка антителами OKT3, C225v5 или человеческим IgG1 контрольного изотипа (Enzo) приводила к минимальному CD69-индуцированию.
Пример 10: Мишень-зависимое уничтожение клеток-мишеней под действием мультиспецифических антител
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению индуцировать опосредуемый Т-клетками мишень-зависимый лизис клеток.
Для того, чтобы определить, может ли мультиспецифическое антитело C225v5-OKT3m-H-N, C225v5-OKT3m-H-C или C225v5-OKT3m-L-C индуцировать опосредуемый Т-клетками мишень-зависимый лизис клеток, клетки SW480 культивировали вместе с CD8+-T-клетками в отношении эффектор:клетка-мишень=3:1, как описано в настоящей заявке. После инкубирования в течение ночи, 100 мкл супернатанта помещали в 96-луночные планшеты с белыми стенками (Greiner Bio One Catalog 655098) для анализа на явную протеазную активность, ассоциированную с цитотоксичностью, в соответствии с протоколом производителей (CytoTox-Glo™ Cytotoxicity Assay, Catalog G9292, Promega), в котором используется люминогенный пептидный субстрат для измерения активности протеаз, высвобождаемых клетками, имеющими мембрану с нарушенной целостностью, а поэтому, подверженными цитолизу. Цитотоксичность клеток-мишеней, индуцируемую мультиспецифическими антителами, выражали как интенсивность люминесценции после вычитания фоновых необработанных величин, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
На фигуре 18B продемонстрировано, что эти мультиспецифические антитела всех трех форматов индуцировали опосредуемый Т-клетками лизис клеток SW480. Как и предполагалось, одно антитело OKT3 не индуцировало какого-либо детектируемого лизиса клеток SW480, что позволяет предположить, что для обеспечения эффективной цитотоксичности под действием мультиспецифических антител необходимо связывание с CD3 и с опухолевой мишенью.
Пример 11: Мишень-зависимая активация Т-клеток и лизис клеток-мишеней под действием мультиспецифических антител
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению индуцировать опосредуемую Т-клетками мишень-зависимую активацию CD8+-Т-клеток и лизис клеток-мишеней.
Для того, чтобы определить, зависит ли активация Т-клеток от экспрессии EGFR клетками-мишенями, CFSE-меченные EGFR-позитивные клетки SW480 или EGFR-негативные клетки U266 совместно культивировали с человеческими CD8+-T-клетками в присутствии мультиспецифического антитела C225v5-OKT3m-H-N или OKT3, каждое из которых подвергали 5-кратному серийному разведению, начиная с 200 пM, как описано выше. CFSE-мечение клеток-мишеней (30 нМ) проводили для дифференциации клеток-мишеней от Т-клеток описанным здесь методом. Замороженные цитотоксические CD8+-T-клетки нормальной периферической крови (AllCells, Catalog PB009-3F) оттаивали в соответствии с инструкциями производителей. После инкубирования в течение ночи, Т-клетки (CFSE-негативные клетки) оценивали на экспрессию маркера ранней активации Т-клеток, присутствующиего на поверхности Т-клеток, CD69, как описано в настоящей заявке. Вкратце, клетки удаляли из аналитического планшета, а прилипшие клетки удаляли с использованием трипсина (Life Technologies), и эти клетки один раз промывали FACS-буфером. Клетки окрашивали в течение 1 часа с использованием ФЭ-конъюгированного анти-CD69-антитела (BD Bioscience). Клетки промывали и анализировали на экспрессию CD69 клеточной поверхности с помощью BD Accuri. Величины MFI вычисляли, как описано выше. Результаты выражали в процентах как увеличение числа Т-клеток, экспрессирующих анти-CD69 антитело, по сравнению с клетками, экспрессирующими антитело контрольного изотипа, с помощью аналитической компьютерной программы FCS Express, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
Как показано на фигуре 18C, мультиспецифическое антитело C225v5-OKT3m-H-N индуцировало сильную активацию Т-клеток в суб-пM-концентрациях в присутствии клеток SW480 и лишь минимальную активацию Т-клеток в присутствии клеток U266 даже при концентрации, которая более чем в 3 log выше, чем концентрация в случае использования клеток SW480. Антитело OKT3 индуцировало минимальную активацию Т-клеток. Эти результатаы показали, что эффективная активация Т-клеток зависит от экспрессии EGFR клетками-мишенями.
Для того, чтобы определить, зависит ли лизис клеток-мишеней от экспрессии EGFR, 100 мкл супернатанта T-клеточной культуры, в которой присутствовали клетки SW480 или U266, оценивали на протеазную активность, ассоциированную с цитотоксичностью (CytoTox-Glo, Promega). Результаты выражали как интенсивность люминесценции без вычитания фона для выявления U266-ответа, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
На фигуре 18D показано, что мультиспецифическое антитело C225v5-OKT3m-H-N индуцирует цитотоксичность EGFR-экспрессирующих клеток SW480 в суб-пM-концентрациях, но при этом, какого-либо заметного цитолиза EGFR-негативных клеток U266 не детектировалось. Антитело OKT3 также не индуцировало какого-либо заметного цитолиза клеток SW480 или клеток U266.
Пример 12: Способность мультиспецифических антител связываться с Т-клетками и уничтожать популяцию EGFR-экспрессирующих клеточных линий
В этом примере продемонстрирована способность мультиспецифических антител согласно изобретению связываться с Т-клетками и уничтожать популяцию EGFR-экспрессирующих клеточных линий.
Для того, чтобы определить, может ли мультиспецифическое антитело C225v5-OKT3m-H-N индуцировать цитотоксичность дополнительных EGFR-экспрессирующих клеточных линий, человеческие CD8+-T-клетки культивировали в среде RPMI-1640, содержащей 2% человеческую сыворотку, в 96-луночных планшетах с белыми лунками, вместе с этими дополнительными EGFR-экспрессирующими клеточными линиями в отношении эффектор:мишень = 5:1 (за исключением клеток U266, которые использовали в отношении 3:1 и культивировали в 96-луночных плоскодонных планшетах) и проводили титрование мультиспецифического антитела C225v5-OKT3m-H-N в присутствии HEK-293, HCT-15, HCT 116, Hs 766T, HT-29, NCI-H2405, SW480, SK-OV-3 и EGFR-негативной клеточной линии, U266 (все клеточные линии были взяты из ATCC). Анти-EGFR антитело C225v5 (в концентрации 200 пM) использовали в качестве негативного контроля. После инкубирования в течение ночи, люминогенный пептидный субстрат, используемый в анализе на цитотоксичность CytoTox-Glo™ (Promega), непосредственно добавляли в планшеты (со 150 мкл супернатанта) для определения высвобождаемой протеазной активности, за исключением образцов U266, где для оценки протеазной активности использовали 100 мкл супернатанта U266. Результаты выражали как интенсивность люминесценции после вычитания фоновых необработанных величин, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
На фигуре 19 показано, что все EGFR-экспрессирующие клеточные линии, обработанные мультиспецифическим антителом C225v5-OKT3m-H-N в присутствии CD8+-T-клеток, обладали дозозависимой цитотоксичностью. В противоположность этому, EGFR-негативные клетки U266 не подвергались воздействию мультиспецифического антитела C225v5-OKT3m-H-N. Ни одна из этих клеточных линий, обработанных анти-EGFR антителом C225v5 при 200 пM, не обнаруживала какой-либо заметной цитотоксичности независимо от того, экспрессировала она EGFR или нет.
Пример 13: Снижение уровня связывания EGFR под действием мультиспецифических активируемых антител
В этом примере продемонстрировано, что уровень связывания с EGFR под действием мультиспецифического активируемого антитела согласно изобретению был ниже, чем уровень связывания с EGFR под действием мультиспецифического антитела согласно изобретению. В этом примере также показано, что связывание с EGFR под действием мультиспецифического активируемого антитела, которое включает группу, расщепляемую протеазой, восстанавливается после расщепления мультиспецифического активируемого антитела такой протеазой.
Мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N, активированное мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N и мультиспецифическое антитело C225v5-OKT3m-H-N тестировали на их способность связываться с EGFR-экспрессирующими клетками SW480.
Активацию мультиспецифического активируемого антитела осуществляли следующим образом: 825 мкг мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N в PBS расщепляли путем добавления оттитрованной по активному центру матриптазы (также обозначаемой здесь MT-SP1 и MTSP1 и поставляемой от R&D Systems, Catalog 3946-SE-010) до конечной концентрации 100 нМ. Гидролизат инкубировали при 37°C в течение ночи, и расщепление подтверждали путем взятия аликвот для анализа методом капиллярного электрофореза (GX-II Capillary Electrophoresis, Perkin Elmer). Затем протеазу и отщепленную маскирующую группу удаляли путем очистки на белке А. Вкратце, гидролизованный образец разводили в PBS до 2 мл и загружали в уравновешенные сферы MabSelect SuRe™ (GE Healthcare Life Sciences, Product 11-0026-01 AD). Эти сферы промывали 5 колоночными объемами (CV) 1xPBS, а затем 5 колоночными объемами (CV) 5xPBS, в который был добавлен 5% изопропиловый спирт (IPA), и наконец 5 CV 1xPBS. Антитело элюировали 10 колоночными объемами 0,1 M глицина, pH 3,0, и фракции нейтрализовали 1 M Триса, pH 8,0, объединяли, концентрировали и буфер заменяли на PBS.
EGFR-экспрессирующие клетки SW480 отделяли с использованием буфера для диссоциации клеток (Sigma, Catalog C5789), промывали и инкубировали в течение 1 часа с 5-кратным разведением 1000 нМ мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, мультиспецифического антитела C225v5-OKT3m-H-N, активируемого антитела 3954-1204-C225v5 или антитела C225v5 в FACS-буфере на льду. Клетки 3 раза промывали FACS-буфером и инкубировали со «вторым» антителом, а именно, антителом AF488, специфичным к человеческому Fc-гамма (Jackson ImmunoResearch Catalog 109-546-098), в FACS-буфере в отношении 1:400 на льду. Клетки 3 раза промывали FACS-буфером и MFI AF488 считывали на проточном цитометре BD Accuri (BD Biosciences). MFI образца, содержащего только «второе» контрольное антитело, вычитали из экспериментального MFI, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
На фигуре 20A показано, что уровень связывания с EGFR под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N был ниже, чем уровень связывания EGFR под действием мультиспецифического антитела C225v5-OKT3m-H-N, однако, уровень связывания с EGFR под действием мультиспецифического активируемого антитела полностью восстанавливался после расщепления мультиспецифического активируемого антитела протеазой, а именно, матриптазой. На фигуре 20В показано, что уровень связывания с EGFR под действием антитела C225v5 и активируемого антитела 3954-1204-C225v5 был сравним с уровенм связывания с EGFR под действием мультиспецифического антитела C225v5-OKT3m-H-N и мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, соответственно, что указывало на то, что молекула анти-CD3ε антитела, присутствующая в мультиспецифическом антителе, не влияла на профиль связывания с EGFR под действием мультиспецифического антитела или мультиспецифического активируемого антитела.
Пример 14: Способность мультиспецифических активируемых антител связываться с CD3+-T-клетками
В этом примере продемонстрировано, что мультиспецифическое активируемое антитело, активированное мультиспецифическое активируемое антитело и мультиспецифическое антитело всех вариантов настоящего изобретения связываются с CD3+-T-клетками.
Для того, чтобы определить, влияет ли маскировка EGFR-связывающего сайта мультиспецифического активируемого анти-EGFR антитела на связывание с CD3ε, был проведен описанный здесь анализ на связывание с Т-клетками Jurkat с использованием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N и мультиспецифического антитела C225v5-OKT3m-H-N. Клетки Jurkat инкубировали с 5-кратным разведением 1000 нМ мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного матриптазой мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N или мультиспецифического антитела C225v5-OKT3m-H-N в FACS-буфере в течение 1 часа при 4°C. Клетки 3 раза промывали и инкубировали со «вторым» антителом, а именно, антителом AF488, специфичным к человеческому Fc-гамма (Jackson ImmunoResearch) в отношении 1:400. Клетки 3 раза промывали, и MFI AF488 считывали на проточном цитометре BD Accuri (BD Biosciences). MFI образца, содержащего только «второе» контрольное антитело, вычитали из экспериментального MFI, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
На фигуре 21A показано, что мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N и мультиспецифическое антитело C225v5-OKT3m-H-N обнаруживали эквивалентные уровни связывания с Т-клетками Jurkat. На фигуре 21В показано, что мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N и активированное мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N обнаруживали эквивалентные уровни связывания с Т-клетками Jurkat. Эти результаты показали, что маскировка EGFR-связывающей молекулы мультиспецифического активируемого антитела не влияет на способность мультиспецифического активируемого антитела связываться с Т-клетками.
Пример 15: Мишень-зависимая активация Т-клеток под действием мультиспецифических активируемых антител
В этом примере продемонстрировано, что мишень-зависимая активация Т-клеток под действием мультиспецифического активируемого антитела согласно изобретению снижалась по сравнению с активацией под действием мультиспецифического антитела согласно изобретению. В этом примере также показано, что мишень-зависимая активация Т-клеток под действием мультиспецифического активируемого антитела, которое включает группу, расщепляемую протеазой, восстанавливается после расщепления мультиспецифического активируемого антитела такой протеазой.
Для того, чтобы определить, может ли маскировка EGFR-связывающего сайта мультиспецифического активируемого анти-EGFR антитела приводить к снижению мишень-зависимой активации Т-клеток, и для того, чтобы определить, может ли активация мультиспецифического активируемого антитела протеазой приводить к восстановлению активации Т-клеток, был проведен анализ на активацию клеток Jurkat, описанный в настоящей заявке, где указанный анализ осуществляли с использованием тестируемого мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, мультиспецифического антитела C225v5-OKT3m-H-N, анти-EGFR антитела C225v5 и антитела контрольного изотипа Synagis (Medimmune), описанных в настоящей заявке.
На фигуре 22 показано, что EGFR-зависимая активация, определяемая по CD69-индуцированию Т-клеток Jurkat, культивированных вместе с EGFR-экспрессирующими клетками SW480, под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, была ниже, чем EGFR-зависимая активация под действием мультиспецифического антитела C225v5-OKT3m-H-N. На этой фигуре также показано, что EGFR-зависимая активация под действием мультиспецифического активируемого антитела была полностью восстановлена после расщепления мультиспецифического активируемого антитела протеазой, а именно матриптазой. Ни анти-EGFR антитело C225v5, ни антитело контрольного изотипа не индуцировали активацию Т-клеток.
Пример 16: Мишень-зависимая активация Т-клеток и лизис клеток-мишеней под действием мультиспецифических активируемых антител
В этом примере продемонстрировано, что мишень-зависимая активация Т-клеток и лизис клеток-мишеней под действием мультиспецифического активируемого антитела согласно изобретению снижались по сравнению с активацией и лизисом под действием мультиспецифического антитела согласно изобретению. В этом примере также показано, что мишень-зависимая активация Т-клеток и лизис клеток-мишеней под действием мультиспецифического активируемого антитела, которое включает группу, расщепляемую протеазой, восстанавливались после расщепления мультиспецифического активируемого антитела такой протеазой.
Для того, чтобы определить, может ли маскировка EGFR-связывающего сайта мультиспецифического активируемого анти-EGFR антитела приводить к снижению мишень-зависимой активации, и для того, чтобы определить, может ли активация мультиспецифического активируемого антитела протеазой восстанавливать такую активацию, был проведен анализ на активацию клеток Jurkat, описанный в настоящей заявке, где указанный анализ осуществляли с использованием тестируемого мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, мультиспецифического антитела C225v5-OKT3m-H-N, анти-CD3ε антитела OKT3, анти-EGFR антитела C225v5 и антитела контрольного изотипа, описанных в настоящей заявке.
Для того, чтобы определить, влияет ли маскировка EGFR-связывающего сайта мультиспецифического активируемого анти-EGFR антитела на цитотоксичность и способность активированного протеазой мультиспецифического активируемого антитела восстанавливать цитотоксическую активность, EGFR-экспрессирующие клетки SW480 культивировали вместе с Т-клетками в отношении эффектор:мишень=5:1 с 5-кратным серийным разведением, начиная с 200 пM, мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, активированного мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N или мультиспецифического антитела C225v5-OKT3m-H-N. 200 пM OKT3, C225v5 и антитела IgG1 контрольного изотипа Synagis (Medimmune) использовали в качестве контроля. После инкубирования в течение ночи, 100 мкл супернатанта помещали в 96-луночные планшеты с белыми стенками для анализа на явную протеазную активность, ассоциированную с цитотоксичностью (CytoTox-Glo, Promega). Результаты выражали как интенсивность люминесценции после вычитания фоновых необработанных величин, и с помощью программы Prism строили кривую зависимости логарифмических данных log (агонист)-ответ (по трем параметрам).
На фигуре 23А показано, что EGFR-зависимая активация, определяемая по CD69-индуцированию первичных Т-клеток, культивированных вместе с EGFR-экспрессирующими клетками SW480, под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N, была ниже, чем EGFR-зависимая активация под действием мультиспецифического антитела C225v5-OKT3m-H-N. На этой фигуре также показано, что EGFR-зависимая активация под действием мультиспецифического активируемого антитела была полностью восстановлена после расщепления мультиспецифического активируемого антитела протеазой, а именно, матриптазой. OKT3, анти-EGFR антитело C225v5 и антитело IgG1 контрольного изотипа Synagis индуцировали активацию Т-клеток на минимальном уровне.
На фигуре 23В показано, что EGFR-зависимый лизис клеток SW480 под действием мультиспецифического активируемого антитела 3954-1204-C225v5-OKT3m-H-N был ниже, чем EGFR-зависимая цитотоксичность, индуцируемая мультиспецифическим антителом C225v5-OKT3m-H-N. На этой фигуре также показано, что EGFR-зависимая цитотоксичность, индуцируемая мультиспецифическим активируемым антителом, была полностью восстановлена после расщепления мультиспецифического активируемого антитела протеазой, а именно, матриптазой. C225v5, OKT3, и антитело IgG1 контрольного изотипа Synagis индуцировали цитотоксичность на минимальном уровне.
Другие варианты осуществления изобретения
Хотя в настоящей заявке дано подробное описание настоящего изобретения, однако, оно приводится лишь в целях иллюстрации и не должно рассматриваться как ограничение объема изобретения, который определен в прилагаемой формуле изобретения. Другие аспекты, преимущества и модификации также входят в объем настоящего изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CytomX Therapeutics, Inc.
Lowman, Henry Bernard
West, James William
La Porte, Sherry Lynn
Irving, Bryan Allen
Hostetter, Daniel Robert
Wong, Chihunt
<120> Мультиспецифические антитела, мультиспецифические активируемые антитела и их применение
<130> CYTM-025/001WO 322001-2133
<140> PCT/US14/48289
<141> 2014-07-25
<150> US 61/858,402
<151> 2013-07-25
<160> 339
<170> PatentIn version 3.5
<210> 1
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер CTRevNot
<400> 1
tcgagcggcc gctcaactag ctgaagagac agtg 34
<210> 2
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CTForOL
<400> 2
gccctctaga ctcgatctag ctagctgaag agacagtgac cagg 44
<210> 3
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер HCForNhe
<400> 3
ctcagctagc accagggccc atcggtc 27
<210> 4
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер HCRevOL
<400> 4
ctttacccgg agacagggag aggctcttct gc 32
<210> 5
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер OKREVNot
<400> 5
ctcgagcggc cgctcaacga ttaatttcca gtttg 35
<210> 6
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-Jagged VH CDR1
<400> 6
Ser Tyr Ala Met Ser
1 5
<210> 7
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-Jagged VH CDR2
<400> 7
Ser Ile Asp Pro Glu Gly Arg Gln Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 8
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-Jagged VH CDR3
<400> 8
Asp Ile Gly Gly Arg Ser Ala Phe Asp Tyr
1 5 10
<210> 9
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-Jagged VL CDR1
<400> 9
Arg Ala Ser Gln Ser Ile Ser Ser Tyr
1 5
<210> 10
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-Jagged VL CDR2
<400> 10
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 11
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-Jagged VL CDR3
<400> 11
Gln Gln Thr Val Val Ala Pro Pro Leu
1 5
<210> 12
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-EGFR VH CDR1
<400> 12
Asn Tyr Gly Val His
1 5
<210> 13
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-EGFR VH CDR2
<400> 13
Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr Ser
1 5 10 15
<210> 14
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-EGFR VH CDR3
<400> 14
Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr
1 5 10
<210> 15
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-EGFR VL CDR1
<400> 15
Arg Ala Ser Gln Ser Ile Gly Thr Asn Ile His
1 5 10
<210> 16
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-EGFR VL CDR2
<400> 16
Lys Tyr Ala Ser Glu Ser Ile Ser
1 5
<210> 17
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> анти-EGFR VL CDR3
<400> 17
Gln Gln Asn Asn Asn Trp Pro Thr Thr
1 5
<210> 18
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<220>
<221> Повтор
<222> (1)..(5)
<223> Может быть повторен
<400> 18
Gly Ser Gly Gly Ser
1 5
<210> 19
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<220>
<221> Повтор
<222> (1)..(4)
<223> Может быть повторен
<400> 19
Gly Gly Gly Ser
1
<210> 20
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<400> 20
Gly Gly Ser Gly
1
<210> 21
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<400> 21
Gly Gly Ser Gly Gly
1 5
<210> 22
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<400> 22
Gly Ser Gly Ser Gly
1 5
<210> 23
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<400> 23
Gly Ser Gly Gly Gly
1 5
<210> 24
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<400> 24
Gly Gly Gly Ser Gly
1 5
<210> 25
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкерный пептид
<400> 25
Gly Ser Ser Ser Gly
1 5
<210> 26
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 26
Leu Ser Gly Arg Ser Asp Asn His
1 5
<210> 27
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 27
Thr Gly Arg Gly Pro Ser Trp Val
1 5
<210> 28
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 28
Ser Ala Arg Gly Pro Ser Arg Trp
1 5
<210> 29
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 29
Thr Ala Arg Gly Pro Ser Phe Lys
1 5
<210> 30
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 30
Gly Gly Trp His Thr Gly Arg Asn
1 5
<210> 31
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 31
His Thr Gly Arg Ser Gly Ala Leu
1 5
<210> 32
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 32
Pro Leu Thr Gly Arg Ser Gly Gly
1 5
<210> 33
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 33
Ala Ala Arg Gly Pro Ala Ile His
1 5
<210> 34
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 34
Arg Gly Pro Ala Phe Asn Pro Met
1 5
<210> 35
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 35
Ser Ser Arg Gly Pro Ala Tyr Leu
1 5
<210> 36
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 36
Arg Gly Pro Ala Thr Pro Ile Met
1 5
<210> 37
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 37
Arg Gly Pro Ala
1
<210> 38
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 38
Gly Gly Gln Pro Ser Gly Met Trp Gly Trp
1 5 10
<210> 39
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 39
Phe Pro Arg Pro Leu Gly Ile Thr Gly Leu
1 5 10
<210> 40
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 40
Val His Met Pro Leu Gly Phe Leu Gly Pro
1 5 10
<210> 41
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 41
Ser Pro Leu Thr Gly Arg Ser Gly
1 5
<210> 42
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 42
Ser Ala Gly Phe Ser Leu Pro Ala
1 5
<210> 43
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 43
Leu Ala Pro Leu Gly Leu Gln Arg Arg
1 5
<210> 44
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 44
Ser Gly Gly Pro Leu Gly Val Arg
1 5
<210> 45
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 45
Pro Leu Gly Leu
1
<210> 46
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> спейсерный пептид
<400> 46
Gln Gly Gln Ser Gly Gln
1 5
<210> 47
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 47
Pro Arg Phe Lys Ile Ile Gly Gly
1 5
<210> 48
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 48
Pro Arg Phe Arg Ile Ile Gly Gly
1 5
<210> 49
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 49
Ser Ser Arg His Arg Arg Ala Leu Asp
1 5
<210> 50
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 50
Arg Lys Ser Ser Ile Ile Ile Arg Met Arg Asp Val Val Leu
1 5 10
<210> 51
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 51
Ser Ser Ser Phe Asp Lys Gly Lys Tyr Lys Lys Gly Asp Asp Ala
1 5 10 15
<210> 52
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 52
Ser Ser Ser Phe Asp Lys Gly Lys Tyr Lys Arg Gly Asp Asp Ala
1 5 10 15
<210> 53
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 53
Ile Glu Gly Arg
1
<210> 54
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 54
Ile Asp Gly Arg
1
<210> 55
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 55
Gly Gly Ser Ile Asp Gly Arg
1 5
<210> 56
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 56
Pro Leu Gly Leu Trp Ala
1 5
<210> 57
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 57
Gly Pro Gln Gly Ile Ala Gly Gln
1 5
<210> 58
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 58
Gly Pro Gln Gly Leu Leu Gly Ala
1 5
<210> 59
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 59
Gly Ile Ala Gly Gln
1 5
<210> 60
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 60
Gly Pro Leu Gly Ile Ala Gly Ile
1 5
<210> 61
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 61
Gly Pro Glu Gly Leu Arg Val Gly
1 5
<210> 62
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 62
Tyr Gly Ala Gly Leu Gly Val Val
1 5
<210> 63
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 63
Ala Gly Leu Gly Val Val Glu Arg
1 5
<210> 64
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 64
Ala Gly Leu Gly Ile Ser Ser Thr
1 5
<210> 65
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 65
Glu Pro Gln Ala Leu Ala Met Ser
1 5
<210> 66
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 66
Gln Ala Leu Ala Met Ser Ala Ile
1 5
<210> 67
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 67
Ala Ala Tyr His Leu Val Ser Gln
1 5
<210> 68
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 68
Met Asp Ala Phe Leu Glu Ser Ser
1 5
<210> 69
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 69
Glu Ser Leu Pro Val Val Ala Val
1 5
<210> 70
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 70
Ser Ala Pro Ala Val Glu Ser Glu
1 5
<210> 71
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 71
Asp Val Ala Gln Phe Val Leu Thr
1 5
<210> 72
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 72
Val Ala Gln Phe Val Leu Thr Glu
1 5
<210> 73
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 73
Ala Gln Phe Val Leu Thr Glu Gly
1 5
<210> 74
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> расщепляемая линкерная последовательность
<400> 74
Pro Val Gln Pro Ile Gly Pro Gln
1 5
<210> 75
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 75
Cys Ile Ser Pro Arg Gly
1 5
<210> 76
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 76
Cys Ile Ser Pro Arg Gly Cys Gly
1 5
<210> 77
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 77
Cys Ile Ser Pro Arg Gly Cys Pro Asp Gly Pro Tyr Val Met Tyr
1 5 10 15
<210> 78
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 78
Cys Ile Ser Pro Arg Gly Cys Pro Asp Gly Pro Tyr Val Met
1 5 10
<210> 79
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 79
Cys Ile Ser Pro Arg Gly Cys Glu Pro Gly Thr Tyr Val Pro Thr
1 5 10 15
<210> 80
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 80
Cys Ile Ser Pro Arg Gly Cys Pro Gly Gln Ile Trp His Pro Pro
1 5 10 15
<210> 81
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 81
Gly Ser His Cys Leu Ile Pro Ile Asn Met Gly Ala Pro Ser Cys
1 5 10 15
<210> 82
<211> 32
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 82
Cys Ile Ser Pro Arg Gly Cys Gly Gly Ser Ser Ala Ser Gln Ser Gly
1 5 10 15
Gln Gly Ser His Cys Leu Ile Pro Ile Asn Met Gly Ala Pro Ser Cys
20 25 30
<210> 83
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 83
Cys Asn His His Tyr Phe Tyr Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 84
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 84
Ala Asp His Val Phe Trp Gly Ser Tyr Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 85
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 85
Cys His His Val Tyr Trp Gly His Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 86
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 86
Cys Pro His Phe Thr Thr Thr Ser Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 87
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 87
Cys Asn His His Tyr His Tyr Tyr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 88
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 88
Cys Pro His Val Ser Phe Gly Ser Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 89
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 89
Cys Pro Tyr Tyr Thr Leu Ser Tyr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 90
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 90
Cys Asn His Val Tyr Phe Gly Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 91
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 91
Cys Asn His Phe Thr Leu Thr Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 92
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 92
Cys His His Phe Thr Leu Thr Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Pro Gly
<210> 93
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 93
Tyr Asn Pro Cys Ala Thr Pro Met Cys Cys Ile Ser Pro Arg Gly Cys
1 5 10 15
Pro Gly
<210> 94
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 94
Cys Asn His His Tyr Phe Tyr Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 95
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 95
Cys Asn His His Tyr His Tyr Tyr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 96
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 96
Cys Asn His Val Tyr Phe Gly Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 97
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 97
Cys His His Val Tyr Trp Gly His Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 98
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 98
Cys Pro His Phe Thr Thr Thr Ser Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 99
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 99
Cys Asn His Phe Thr Leu Thr Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 100
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 100
Cys His His Phe Thr Leu Thr Thr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 101
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 101
Cys Pro Tyr Tyr Thr Leu Ser Tyr Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 102
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 102
Cys Pro His Val Ser Phe Gly Ser Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 103
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 103
Ala Asp His Val Phe Trp Gly Ser Tyr Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 104
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 104
Tyr Asn Pro Cys Ala Thr Pro Met Cys Cys Ile Ser Pro Arg Gly Cys
1 5 10 15
Gly
<210> 105
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 105
Cys His His Val Tyr Trp Gly His Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 106
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa may be Asn or Pro
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa may be His, Val or Phe
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa may be Tyr or Thr
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa may be Phe, Trp, Thr or Leu
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa may be Tyr, Gly, Thr or Ser
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> Xaa may be Thr, Ser, Tyr or His
<400> 106
Cys Xaa His Xaa Xaa Xaa Xaa Xaa Cys Gly Cys Ile Ser Pro Arg Gly
1 5 10 15
Cys Gly
<210> 107
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 107
Cys Ile Ser Pro Arg Gly Cys Gly Gln Pro Ile Pro Ser Val Lys
1 5 10 15
<210> 108
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 108
Cys Ile Ser Pro Arg Gly Cys Thr Gln Pro Tyr His Val Ser Arg
1 5 10 15
<210> 109
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 109
Cys Ile Ser Pro Arg Gly Cys Asn Ala Val Ser Gly Leu Gly Ser
1 5 10 15
<210> 110
<211> 26
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 110
Gln Gly Gln Ser Gly Gln Gly Gln Gln Gln Trp Cys Asn Ile Trp Ile
1 5 10 15
Asn Gly Gly Asp Cys Arg Gly Trp Asn Gly
20 25
<210> 111
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 111
Pro Trp Cys Met Gln Arg Gln Asp Phe Leu Arg Cys Pro Gln Pro
1 5 10 15
<210> 112
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 112
Gln Leu Gly Leu Pro Ala Tyr Met Cys Thr Phe Glu Cys Leu Arg
1 5 10 15
<210> 113
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 113
Cys Asn Leu Trp Val Ser Gly Gly Asp Cys Gly Gly Leu Gln Gly
1 5 10 15
<210> 114
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 114
Ser Cys Ser Leu Trp Thr Ser Gly Ser Cys Leu Pro His Ser Pro
1 5 10 15
<210> 115
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 115
Tyr Cys Leu Gln Leu Pro His Tyr Met Gln Ala Met Cys Gly Arg
1 5 10 15
<210> 116
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 116
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ser Tyr Trp Asn Asn Thr
1 5 10 15
<210> 117
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 117
Pro Trp Cys Met Gln Arg Gln Asp Tyr Leu Arg Cys Pro Gln Pro
1 5 10 15
<210> 118
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 118
Cys Asn Leu Trp Ile Ser Gly Gly Asp Cys Arg Gly Leu Ala Gly
1 5 10 15
<210> 119
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 119
Cys Asn Leu Trp Val Ser Gly Gly Asp Cys Arg Gly Val Gln Gly
1 5 10 15
<210> 120
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 120
Cys Asn Leu Trp Val Ser Gly Gly Asp Cys Arg Gly Leu Arg Gly
1 5 10 15
<210> 121
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 121
Cys Asn Leu Trp Ile Ser Gly Gly Asp Cys Arg Gly Leu Pro Gly
1 5 10 15
<210> 122
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 122
Cys Asn Leu Trp Val Ser Gly Gly Asp Cys Arg Asp Ala Pro Trp
1 5 10 15
<210> 123
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 123
Cys Asn Leu Trp Val Ser Gly Gly Asp Cys Arg Asp Leu Leu Gly
1 5 10 15
<210> 124
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 124
Cys Asn Leu Trp Val Ser Gly Gly Asp Cys Arg Gly Leu Gln Gly
1 5 10 15
<210> 125
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 125
Cys Asn Leu Trp Leu His Gly Gly Asp Cys Arg Gly Trp Gln Gly
1 5 10 15
<210> 126
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 126
Cys Asn Ile Trp Leu Val Gly Gly Asp Cys Arg Gly Trp Gln Gly
1 5 10 15
<210> 127
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 127
Cys Thr Thr Trp Phe Cys Gly Gly Asp Cys Gly Val Met Arg Gly
1 5 10 15
<210> 128
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 128
Cys Asn Ile Trp Gly Pro Ser Val Asp Cys Gly Ala Leu Leu Gly
1 5 10 15
<210> 129
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 129
Cys Asn Ile Trp Val Asn Gly Gly Asp Cys Arg Ser Phe Glu Gly
1 5 10 15
<210> 130
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 130
Tyr Cys Leu Asn Leu Pro Arg Tyr Met Gln Asp Met Cys Trp Ala
1 5 10 15
<210> 131
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 131
Tyr Cys Leu Ala Leu Pro His Tyr Met Gln Ala Asp Cys Ala Arg
1 5 10 15
<210> 132
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 132
Cys Phe Leu Tyr Ser Cys Gly Asp Val Ser Tyr Trp Gly Ser Ala
1 5 10 15
<210> 133
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 133
Cys Tyr Leu Tyr Ser Cys Thr Asp Ser Ala Phe Trp Asn Asn Arg
1 5 10 15
<210> 134
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 134
Cys Tyr Leu Tyr Ser Cys Asn Asp Val Ser Tyr Trp Ser Asn Thr
1 5 10 15
<210> 135
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 135
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ser Tyr Trp
1 5 10
<210> 136
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 136
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ala Tyr Trp Asn Ser Ala
1 5 10 15
<210> 137
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 137
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ser Tyr Trp Gly Asp Thr
1 5 10 15
<210> 138
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 138
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ser Tyr Trp Gly Asn Ser
1 5 10 15
<210> 139
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 139
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ala Tyr Trp Asn Asn Thr
1 5 10 15
<210> 140
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 140
Cys Phe Leu Tyr Ser Cys Gly Asp Val Ser Tyr Trp Gly Asn Pro Gly
1 5 10 15
Leu Ser
<210> 141
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 141
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ala Tyr Trp Ser Gly Leu
1 5 10 15
<210> 142
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 142
Cys Tyr Leu Tyr Ser Cys Thr Asp Gly Ser Tyr Trp Asn Ser Thr
1 5 10 15
<210> 143
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 143
Cys Phe Leu Tyr Ser Cys Ser Asp Val Ser Tyr Trp Gly Asn Ile
1 5 10 15
<210> 144
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 144
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ala Tyr Trp
1 5 10
<210> 145
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 145
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ser Tyr Trp Gly Ser Thr
1 5 10 15
<210> 146
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 146
Cys Phe Leu Tyr Ser Cys Thr Asp Val Ala Tyr Trp Gly Asp Thr
1 5 10 15
<210> 147
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 147
Gly Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg Gly Trp Val Asp
1 5 10 15
Pro Leu Gln Gly
20
<210> 148
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 148
Gly Cys Asn Ile Trp Leu Val Gly Gly Asp Cys Arg Gly Trp Ile Gly
1 5 10 15
Asp Thr Asn Gly
20
<210> 149
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 149
Gly Cys Asn Ile Trp Leu Val Gly Gly Asp Cys Arg Gly Trp Ile Glu
1 5 10 15
Asp Ser Asn Gly
20
<210> 150
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 150
Gly Cys Asn Ile Trp Ala Asn Gly Gly Asp Cys Arg Gly Trp Ile Asp
1 5 10 15
Asn Ile Asp Gly
20
<210> 151
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 151
Gly Cys Asn Ile Trp Leu Val Gly Gly Asp Cys Arg Gly Trp Leu Gly
1 5 10 15
Glu Ala Val Gly
20
<210> 152
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 152
Gly Cys Asn Ile Trp Leu Val Gly Gly Asp Cys Arg Gly Trp Leu Glu
1 5 10 15
Glu Ala Val Gly
20
<210> 153
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 153
Gly Gly Pro Ala Leu Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg
1 5 10 15
Gly Trp Ser Gly
20
<210> 154
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 154
Gly Ala Pro Val Phe Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg
1 5 10 15
Gly Trp Met Gly
20
<210> 155
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 155
Gly Gln Gln Gln Trp Cys Asn Ile Trp Ile Asn Gly Gly Asp Cys Arg
1 5 10 15
Gly Trp Asn Gly
20
<210> 156
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 156
Gly Lys Ser Glu Phe Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg
1 5 10 15
Gly Trp Ile Gly
20
<210> 157
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 157
Gly Thr Pro Gly Gly Cys Asn Ile Trp Ala Asn Gly Gly Asp Cys Arg
1 5 10 15
Gly Trp Glu Gly
20
<210> 158
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 158
Gly Ala Ser Gln Tyr Cys Asn Leu Trp Ile Asn Gly Gly Asp Cys Arg
1 5 10 15
Gly Trp Arg Gly
20
<210> 159
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 159
Gly Cys Asn Ile Trp Leu Val Gly Gly Asp Cys Arg Pro Trp Val Glu
1 5 10 15
Gly Gly
<210> 160
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 160
Gly Cys Asn Ile Trp Ala Val Gly Gly Asp Cys Arg Pro Phe Val Asp
1 5 10 15
Gly Gly
<210> 161
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 161
Gly Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg Ala Trp Val Asp
1 5 10 15
Thr Gly
<210> 162
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 162
Gly Cys Asn Ile Trp Ile Val Gly Gly Asp Cys Arg Pro Phe Ile Asn
1 5 10 15
Asp Gly
<210> 163
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 163
Gly Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg Pro Val Val Phe
1 5 10 15
Gly Gly
<210> 164
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 164
Gly Cys Asn Ile Trp Leu Ser Gly Gly Asp Cys Arg Met Phe Met Asn
1 5 10 15
Glu Gly
<210> 165
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 165
Gly Cys Asn Ile Trp Val Asn Gly Gly Asp Cys Arg Ser Phe Val Tyr
1 5 10 15
Ser Gly
<210> 166
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 166
Gly Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg Gly Trp Glu Ala
1 5 10 15
Ser Gly
<210> 167
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 167
Gly Cys Asn Ile Trp Ala His Gly Gly Asp Cys Arg Gly Phe Ile Glu
1 5 10 15
Pro Gly
<210> 168
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 168
Gly Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg Thr Phe Val Ala
1 5 10 15
Ser Gly
<210> 169
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 169
Gly Cys Asn Ile Trp Ala His Gly Gly Asp Cys Arg Gly Phe Ile Glu
1 5 10 15
Pro Gly
<210> 170
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 170
Gly Phe Leu Glu Asn Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg
1 5 10 15
Thr Gly
<210> 171
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 171
Gly Ile Tyr Glu Asn Cys Asn Ile Trp Leu Asn Gly Gly Asp Cys Arg
1 5 10 15
Met Gly
<210> 172
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 172
Gly Ile Pro Asp Asn Cys Asn Ile Trp Ile Asn Gly Gly Asp Cys Arg
1 5 10 15
Tyr Gly
<210> 173
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 173
Arg Glu Cys Gly Arg Cys Gly Asp Ile Pro Phe Pro Ala His Trp Val
1 5 10 15
Pro Arg Thr
<210> 174
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 174
Gln Gly Gln Ser Gly Gln Tyr Gly Ser Cys Ser Trp Asn Tyr Val His
1 5 10 15
Ile Phe Met Asp Cys
20
<210> 175
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 175
Gln Gly Gln Ser Gly Gln Gly Asp Phe Asp Ile Pro Phe Pro Ala His
1 5 10 15
Trp Val Pro Ile Thr
20
<210> 176
<211> 24
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 176
Gln Gly Gln Ser Gly Gln Met Gly Val Pro Ala Gly Cys Val Trp Asn
1 5 10 15
Tyr Ala His Ile Phe Met Asp Cys
20
<210> 177
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 177
Tyr Arg Ser Cys Asn Trp Asn Tyr Val Ser Ile Phe Leu Asp Cys
1 5 10 15
<210> 178
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 178
Pro Gly Ala Phe Asp Ile Pro Phe Pro Ala His Trp Val Pro Asn Thr
1 5 10 15
<210> 179
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 179
Glu Ser Ser Cys Val Trp Asn Tyr Val His Ile Tyr Met Asp Cys
1 5 10 15
<210> 180
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 180
Tyr Pro Gly Cys Lys Trp Asn Tyr Asp Arg Ile Phe Leu Asp Cys
1 5 10 15
<210> 181
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 181
Tyr Arg Thr Cys Ser Trp Asn Tyr Val Gly Ile Phe Leu Asp Cys
1 5 10 15
<210> 182
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 182
Tyr Gly Ser Cys Ser Trp Asn Tyr Val His Ile Phe Met Asp Cys
1 5 10 15
<210> 183
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 183
Tyr Gly Ser Cys Ser Trp Asn Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 184
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 184
Tyr Gly Ser Cys Asn Trp Asn Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 185
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 185
Tyr Thr Ser Cys Asn Trp Asn Tyr Val His Ile Phe Met Asp Cys
1 5 10 15
<210> 186
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 186
Tyr Pro Gly Cys Lys Trp Asn Tyr Asp Arg Ile Phe Leu Asp Cys
1 5 10 15
<210> 187
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 187
Trp Arg Ser Cys Asn Trp Asn Tyr Ala His Ile Phe Leu Asp Cys
1 5 10 15
<210> 188
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 188
Trp Ser Asn Cys His Trp Asn Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 189
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 189
Asp Arg Ser Cys Thr Trp Asn Tyr Val Arg Ile Ser Tyr Asp Cys
1 5 10 15
<210> 190
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 190
Ser Gly Ser Cys Lys Trp Asp Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 191
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 191
Ser Arg Ser Cys Ile Trp Asn Tyr Ala His Ile His Leu Asp Cys
1 5 10 15
<210> 192
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 192
Ser Met Ser Cys Tyr Trp Gln Tyr Glu Arg Ile Phe Leu Asp Cys
1 5 10 15
<210> 193
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 193
Tyr Arg Ser Cys Asn Trp Asn Tyr Val Ser Ile Phe Leu Asp Cys
1 5 10 15
<210> 194
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 194
Tyr Gly Ser Cys Ser Trp Asn Tyr Val His Ile Phe Met Asp Cys
1 5 10 15
<210> 195
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 195
Ser Gly Ser Cys Lys Trp Asp Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 196
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 196
Tyr Lys Ser Cys His Trp Asp Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 197
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 197
Tyr Gly Ser Cys Thr Trp Asn Tyr Val His Ile Phe Met Glu Cys
1 5 10 15
<210> 198
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 198
Phe Ser Ser Cys Asn Trp Asn Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 199
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 199
Trp Arg Ser Cys Asn Trp Asn Tyr Ala His Ile Phe Leu Asp Cys
1 5 10 15
<210> 200
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 200
Tyr Gly Ser Cys Gln Trp Asn Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 201
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 201
Tyr Arg Ser Cys Asn Trp Asn Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 202
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 202
Asn Met Ser Cys His Trp Asp Tyr Val His Ile Phe Leu Asp Cys
1 5 10 15
<210> 203
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 203
Phe Gly Pro Cys Thr Trp Asn Tyr Ala Arg Ile Ser Trp Asp Cys
1 5 10 15
<210> 204
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<220>
<221> misc_feature
<222> (1)..(2)
<223> Xaa может быть любой аминокислотой
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa может быть любой аминокислотой
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может быть любой аминокислотой
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa может быть любой аминокислотой
<400> 204
Xaa Xaa Ser Cys Xaa Trp Xaa Tyr Val His Ile Phe Xaa Asp Cys
1 5 10 15
<210> 205
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 205
Met Gly Val Pro Ala Gly Cys Val Trp Asn Tyr Ala His Ile Phe Met
1 5 10 15
Asp Cys
<210> 206
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 206
Arg Asp Thr Gly Gly Gln Cys Arg Trp Asp Tyr Val His Ile Phe Met
1 5 10 15
Asp Cys
<210> 207
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 207
Ala Gly Val Pro Ala Gly Cys Thr Trp Asn Tyr Val His Ile Phe Met
1 5 10 15
Glu Cys
<210> 208
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 208
Val Gly Val Pro Asn Gly Cys Val Trp Asn Tyr Ala His Ile Phe Met
1 5 10 15
Glu Cys
<210> 209
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 209
Asp Gly Gly Pro Ala Gly Cys Ser Trp Asn Tyr Val His Ile Phe Met
1 5 10 15
Glu Cys
<210> 210
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 210
Ala Val Gly Pro Ala Gly Cys Trp Trp Asn Tyr Val His Ile Phe Met
1 5 10 15
Glu Cys
<210> 211
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 211
Cys Thr Trp Asn Tyr Val His Ile Phe Met Asp Cys Gly Glu Gly Glu
1 5 10 15
Gly Pro
<210> 212
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 212
Gly Gly Val Pro Glu Gly Cys Thr Trp Asn Tyr Ala His Ile Phe Met
1 5 10 15
Glu Cys
<210> 213
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 213
Ala Glu Val Pro Ala Gly Cys Trp Trp Asn Tyr Val His Ile Phe Met
1 5 10 15
Glu Cys
<210> 214
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 214
Ala Gly Val Pro Ala Gly Cys Thr Trp Asn Tyr Val His Ile Phe Met
1 5 10 15
Glu Cys
<210> 215
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 215
Ser Gly Ala Ser Gly Gly Cys Lys Trp Asn Tyr Val His Ile Phe Met
1 5 10 15
Asp Cys
<210> 216
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 216
Met Gly Val Pro Ala Gly Cys Val Trp Asn Tyr Ala His Ile Phe Met
1 5 10 15
Asp Cys
<210> 217
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 217
Thr Pro Gly Cys Arg Trp Asn Tyr Val His Ile Phe Met Glu Cys Glu
1 5 10 15
Ala Leu
<210> 218
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 218
Val Gly Val Pro Asn Gly Cys Val Trp Asn Tyr Ala His Ile Phe Met
1 5 10 15
Glu Cys
<210> 219
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 219
Pro Gly Ala Phe Asp Ile Pro Phe Pro Ala His Trp Val Pro Asn Thr
1 5 10 15
<210> 220
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 220
Arg Gly Ala Cys Asp Ile Pro Phe Pro Ala His Trp Ile Pro Asn Thr
1 5 10 15
<210> 221
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 221
Gln Gly Asp Phe Asp Ile Pro Phe Pro Ala His Trp Val Pro Ile Thr
1 5 10 15
<210> 222
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<220>
<221> misc_feature
<222> (1)..(1)
<223> Xaa может быть любой аминокислотой
<400> 222
Xaa Gly Ala Phe Asp Ile Pro Phe Pro Ala His Trp Val Pro Asn Thr
1 5 10 15
<210> 223
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 223
Arg Gly Asp Gly Asn Asp Ser Asp Ile Pro Phe Pro Ala His Trp Val
1 5 10 15
Pro Arg Thr
<210> 224
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 224
Ser Gly Val Gly Arg Asp Arg Asp Ile Pro Phe Pro Ala His Trp Val
1 5 10 15
Pro Arg Thr
<210> 225
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 225
Trp Ala Gly Gly Asn Asp Cys Asp Ile Pro Phe Pro Ala His Trp Ile
1 5 10 15
Pro Asn Thr
<210> 226
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 226
Trp Gly Asp Gly Met Asp Val Asp Ile Pro Phe Pro Ala His Trp Val
1 5 10 15
Pro Val Thr
<210> 227
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 227
Ala Gly Ser Gly Asn Asp Ser Asp Ile Pro Phe Pro Ala His Trp Val
1 5 10 15
Pro Arg Thr
<210> 228
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 228
Glu Ser Arg Ser Gly Tyr Ala Asp Ile Pro Phe Pro Ala His Trp Val
1 5 10 15
Pro Arg Thr
<210> 229
<211> 819
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv антитела против человеческого CTLA-4
<400> 229
ggaggtggat ctggaggtgg cggttcaggc tctggcggag gctcaggtgg tggaggatca 60
ggcggaggtg aaattgtgtt gacacagtct ccaggcaccc tgtctttgtc tccaggggaa 120
agagccaccc tctcctgcag ggccagtcag agtgttagca gcagctactt agcctggtac 180
cagcagaaac ctggccaggc tcccaggctc ctcatctatg gtgcatccag cagggccact 240
ggcatcccag acaggttcag tggcagtggg tctgggacag acttcactct caccatcagc 300
agactggagc ctgaagattt tgcagtgtat tactgtcagc agtatggtag ctcaccgctc 360
actttcggcg gagggaccaa ggtggaaatc aaacgttccg gagggtcgac cataacttcg 420
tataatgtat actatacgaa gttatcctcg agcggtaccc aggtgcagct ggtgcagact 480
gggggaggcg tggtccagcc tgggaggtcc ctgagactct cctgtgcagc ctctggatcc 540
acctttagca gctatgccat gagctgggtc cgccaggctc cagggaaggg gctggagtgg 600
gtctcagcta ttagtggtag tggtggtagc acatactacg cagactccgt gaagggccgg 660
ttcaccatct ccagagacaa ttccaagaac acgctgtatc tgcaaatgaa cagcctgaga 720
gccgaggaca cggccgtata ttactgtgcg acaaactccc tttactggta cttcgatctc 780
tggggccgtg gcaccctggt cactgtctct tcagctagc 819
<210> 230
<211> 273
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv антитела против человеческого CTLA-4
<400> 230
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Glu Ile Val Leu Thr Gln Ser Pro Gly
20 25 30
Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
35 40 45
Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro
50 55 60
Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr
65 70 75 80
Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys
100 105 110
Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile Thr Ser Tyr Asn Val Tyr
130 135 140
Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln Val Gln Leu Val Gln Thr
145 150 155 160
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
165 170 175
Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala Met Ser Trp Val Arg Gln
180 185 190
Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser Gly Ser Gly
195 200 205
Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
210 215 220
Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
225 230 235 240
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Thr Asn Ser Leu Tyr Trp
245 250 255
Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala
260 265 270
Ser
<210> 231
<211> 792
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3 против человеческого CD3 эпсилон
<400> 231
ggaggtggat ctggaggtgg cggttcaggc tctggcggag gctcaggtgg tggaggatca 60
ggcggaggtc aggttcagct gcagcagagc ggtgcagaac tggcacgtcc gggtgcaagc 120
gttaaaatga gctgtaaagc aagcggttat acctttaccc gttataccat gcattgggtt 180
aaacagcgtc cgggtcaggg tctggaatgg attggttata tcaatccgag ccgtggttat 240
accaactaca accagaaatt caaagataaa gcaaccctga ccaccgataa aagcagcagc 300
accgcctata tgcagctgag cagcctgacc tcagaggata gcgcagttta ttactgtgca 360
cgctattatg atgatcacta ttgcctggat tattggggtc agggcaccac cctgaccgtt 420
agcagcggtg gtggtggtag tggtggcggt ggttcaggcg gtggcggtag ccagattgtt 480
ctgacccaga gtccggcaat tatgagcgca agtccgggtg aaaaagttac catgacctgt 540
agcgccagca gcagcgttag ctatatgaat tggtatcagc agaaaagcgg caccagcccg 600
aaacgttgga tttatgatac cagcaaactg gcaagcggtg ttccggcaca ttttcgtggt 660
agcggtagcg gcacctcata tagcctgacc attagcggta tggaagcaga agatgcagca 720
acctattatt gtcagcagtg gtcaagcaat ccgtttacct ttggtagtgg caccaaactg 780
gaaattaatc gt 792
<210> 232
<211> 264
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3 против человеческого CD3 эпсилон
<400> 232
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser Gly Ala
20 25 30
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
50 55 60
Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
65 70 75 80
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
85 90 95
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
100 105 110
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys
115 120 125
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Val
145 150 155 160
Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val
165 170 175
Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr
180 185 190
Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser
195 200 205
Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser Gly
210 215 220
Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala Ala
225 230 235 240
Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Ser
245 250 255
Gly Thr Lys Leu Glu Ile Asn Arg
260
<210> 233
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь антитела 4D11v2 против человеческого Jagged
<400> 233
gaggtgcacc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtgtcaagt attgacccgg aaggtcggca gacatattac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagacatc 300
ggcggcaggt cggcctttga ctactggggc cagggaaccc tggtcaccgt ctcctcagct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaa 1347
<210> 234
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь антитела 4D11v2 против человеческого Jagged
<400> 234
Glu Val His Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Pro Glu Gly Arg Gln Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Ile Gly Gly Arg Ser Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 235
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь антитела 4D11v2 против человеческого Jagged
<220>
<221> misc_feature
<222> (545)..(545)
<223> n может быть любым нуклеотидом
<400> 235
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattagc agctatttaa attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgcg gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag acggttgtgg cgcctccgtt attcggccaa 300
gggaccaagg tggaaatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgancaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 236
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь антитела 4D11v2 против человеческого Jagged
<220>
<221> misc_feature
<222> (182)..(182)
<223> Xaa может быть любой аминокислотой
<400> 236
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Val Val Ala Pro Pro
85 90 95
Leu Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Xaa Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 237
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь антитела 4D11 против человеческого Jagged
<400> 237
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtgtcaagt attgacccgg aaggtcggca gacatattac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagacatc 300
ggcggcaggt cggcctttga ctactggggc cagggaaccc tggtcaccgt ctcctcagct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaa 1347
<210> 238
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь антитела 4D11 против человеческого Jagged
<400> 238
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Pro Glu Gly Arg Gln Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Ile Gly Gly Arg Ser Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 239
<211> 774
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь активируемого анти-Jagged антитела 5342-1204-4D11v2
<400> 239
caaggccagt ctggccagtg caatatttgg ctcgtaggtg gtgattgcag gggctggcag 60
gggggctcga gcggtggcag cggtggctct ggtggtctga gcggccgttc cgataatcat 120
ggcggcggtt ctgacatcca gatgacccag tctccatcct ccctgtctgc atctgtagga 180
gacagagtca ccatcacttg ccgggcaagt cagagcatta gcagctattt aaattggtat 240
cagcagaaac cagggaaagc ccctaagctc ctgatctatg cggcatccag tttgcaaagt 300
ggggtcccat caaggttcag tggcagtgga tctgggacag atttcactct caccatcagc 360
agtctgcaac ctgaagattt tgcaacttac tactgtcaac agacggttgt ggcgcctccg 420
ttattcggcc aagggaccaa ggtggaaatc aaacgtacgg tggctgcacc atctgtcttc 480
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 540
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 600
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 660
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 720
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt 774
<210> 240
<211> 258
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь активируемого анти-Jagged антитела 5342-1204-4D11v2
<220>
<221> misc_feature
<222> (226)..(226)
<223> Xaa может быть любой аминокислотой
<400> 240
Gln Gly Gln Ser Gly Gln Cys Asn Ile Trp Leu Val Gly Gly Asp Cys
1 5 10 15
Arg Gly Trp Gln Gly Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Leu Ser Gly Arg Ser Asp Asn His Gly Gly Gly Ser Asp Ile Gln Met
35 40 45
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
50 55 60
Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn Trp Tyr
65 70 75 80
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser
85 90 95
Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
100 105 110
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
115 120 125
Thr Tyr Tyr Cys Gln Gln Thr Val Val Ala Pro Pro Leu Phe Gly Gln
130 135 140
Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe
145 150 155 160
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
165 170 175
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
180 185 190
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
195 200 205
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
210 215 220
Leu Xaa Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
225 230 235 240
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
245 250 255
Glu Cys
<210> 241
<211> 2139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-Jagged антитела 4D11v2 - анти-CD3 антитела OKT3
<400> 241
gaggtgcacc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtgtcaagt attgacccgg aaggtcggca gacatattac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagacatc 300
ggcggcaggt cggcctttga ctactggggc cagggaaccc tggtcaccgt ctcctcagct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaagga ggtggatctg gaggtggcgg ttcaggctct 1380
ggcggaggct caggtggtgg aggatcaggc ggaggtcagg ttcagctgca gcagagcggt 1440
gcagaactgg cacgtccggg tgcaagcgtt aaaatgagct gtaaagcaag cggttatacc 1500
tttacccgtt ataccatgca ttgggttaaa cagcgtccgg gtcagggtct ggaatggatt 1560
ggttatatca atccgagccg tggttatacc aactacaacc agaaattcaa agataaagca 1620
accctgacca ccgataaaag cagcagcacc gcctatatgc agctgagcag cctgacctca 1680
gaggatagcg cagtttatta ctgtgcacgc tattatgatg atcactattg cctggattat 1740
tggggtcagg gcaccaccct gaccgttagc agcggtggtg gtggtagtgg tggcggtggt 1800
tcaggcggtg gcggtagcca gattgttctg acccagagtc cggcaattat gagcgcaagt 1860
ccgggtgaaa aagttaccat gacctgtagc gccagcagca gcgttagcta tatgaattgg 1920
tatcagcaga aaagcggcac cagcccgaaa cgttggattt atgataccag caaactggca 1980
agcggtgttc cggcacattt tcgtggtagc ggtagcggca cctcatatag cctgaccatt 2040
agcggtatgg aagcagaaga tgcagcaacc tattattgtc agcagtggtc aagcaatccg 2100
tttacctttg gtagtggcac caaactggaa attaatcgt 2139
<210> 242
<211> 713
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-Jagged антитела 4D11v2 - анти-CD3 антитела OKT3
<400> 242
Glu Val His Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Pro Glu Gly Arg Gln Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Ile Gly Gly Arg Ser Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser Gly
465 470 475 480
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala
485 490 495
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
500 505 510
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
515 520 525
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
530 535 540
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
545 550 555 560
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
565 570 575
Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
580 585 590
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile
595 600 605
Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
610 615 620
Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
625 630 635 640
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
645 650 655
Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser
660 665 670
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala
675 680 685
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly
690 695 700
Ser Gly Thr Lys Leu Glu Ile Asn Arg
705 710
<210> 243
<211> 1434
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-Jagged антитела 4D11v2 - анти-CD3 антитела OKT3
<220>
<221> misc_feature
<222> (545)..(545)
<223> n может быть любым нуклеотидом
<400> 243
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattagc agctatttaa attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgcg gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag acggttgtgg cgcctccgtt attcggccaa 300
gggaccaagg tggaaatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgancaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggaggtgg atctggaggt 660
ggcggttcag gctctggcgg aggctcaggt ggtggaggat caggcggagg tcaggttcag 720
ctgcagcaga gcggtgcaga actggcacgt ccgggtgcaa gcgttaaaat gagctgtaaa 780
gcaagcggtt atacctttac ccgttatacc atgcattggg ttaaacagcg tccgggtcag 840
ggtctggaat ggattggtta tatcaatccg agccgtggtt ataccaacta caaccagaaa 900
ttcaaagata aagcaaccct gaccaccgat aaaagcagca gcaccgccta tatgcagctg 960
agcagcctga cctcagagga tagcgcagtt tattactgtg cacgctatta tgatgatcac 1020
tattgcctgg attattgggg tcagggcacc accctgaccg ttagcagcgg tggtggtggt 1080
agtggtggcg gtggttcagg cggtggcggt agccagattg ttctgaccca gagtccggca 1140
attatgagcg caagtccggg tgaaaaagtt accatgacct gtagcgccag cagcagcgtt 1200
agctatatga attggtatca gcagaaaagc ggcaccagcc cgaaacgttg gatttatgat 1260
accagcaaac tggcaagcgg tgttccggca cattttcgtg gtagcggtag cggcacctca 1320
tatagcctga ccattagcgg tatggaagca gaagatgcag caacctatta ttgtcagcag 1380
tggtcaagca atccgtttac ctttggtagt ggcaccaaac tggaaattaa tcgt 1434
<210> 244
<211> 478
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-Jagged антитела 4D11v2 - анти-CD3 антитела OKT3
<220>
<221> misc_feature
<222> (182)..(182)
<223> Xaa может быть любой аминокислотой
<400> 244
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Val Val Ala Pro Pro
85 90 95
Leu Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Xaa Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln
225 230 235 240
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
245 250 255
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His
260 265 270
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile
275 280 285
Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
290 295 300
Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
305 310 315 320
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr
325 330 335
Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
340 345 350
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
355 360 365
Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala
370 375 380
Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val
385 390 395 400
Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg
405 410 415
Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe
420 425 430
Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met
435 440 445
Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn
450 455 460
Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
465 470 475
<210> 245
<211> 1566
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-Jagged-анти-CD3 антитела 5342-1204-4D11v2-CD3 OKT3 Light Chain
<220>
<221> misc_feature
<222> (677)..(677)
<223> n может быть любым нуклеотидом
<400> 245
caaggccagt ctggccagtg caatatttgg ctcgtaggtg gtgattgcag gggctggcag 60
gggggctcga gcggtggcag cggtggctct ggtggtctga gcggccgttc cgataatcat 120
ggcggcggtt ctgacatcca gatgacccag tctccatcct ccctgtctgc atctgtagga 180
gacagagtca ccatcacttg ccgggcaagt cagagcatta gcagctattt aaattggtat 240
cagcagaaac cagggaaagc ccctaagctc ctgatctatg cggcatccag tttgcaaagt 300
ggggtcccat caaggttcag tggcagtgga tctgggacag atttcactct caccatcagc 360
agtctgcaac ctgaagattt tgcaacttac tactgtcaac agacggttgt ggcgcctccg 420
ttattcggcc aagggaccaa ggtggaaatc aaacgtacgg tggctgcacc atctgtcttc 480
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 540
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 600
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 660
agcaccctga cgctgancaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 720
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgtggaggt 780
ggatctggag gtggcggttc aggctctggc ggaggctcag gtggtggagg atcaggcgga 840
ggtcaggttc agctgcagca gagcggtgca gaactggcac gtccgggtgc aagcgttaaa 900
atgagctgta aagcaagcgg ttataccttt acccgttata ccatgcattg ggttaaacag 960
cgtccgggtc agggtctgga atggattggt tatatcaatc cgagccgtgg ttataccaac 1020
tacaaccaga aattcaaaga taaagcaacc ctgaccaccg ataaaagcag cagcaccgcc 1080
tatatgcagc tgagcagcct gacctcagag gatagcgcag tttattactg tgcacgctat 1140
tatgatgatc actattgcct ggattattgg ggtcagggca ccaccctgac cgttagcagc 1200
ggtggtggtg gtagtggtgg cggtggttca ggcggtggcg gtagccagat tgttctgacc 1260
cagagtccgg caattatgag cgcaagtccg ggtgaaaaag ttaccatgac ctgtagcgcc 1320
agcagcagcg ttagctatat gaattggtat cagcagaaaa gcggcaccag cccgaaacgt 1380
tggatttatg ataccagcaa actggcaagc ggtgttccgg cacattttcg tggtagcggt 1440
agcggcacct catatagcct gaccattagc ggtatggaag cagaagatgc agcaacctat 1500
tattgtcagc agtggtcaag caatccgttt acctttggta gtggcaccaa actggaaatt 1560
aatcgt 1566
<210> 246
<211> 522
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-Jagged-анти-CD3 антитела 5342-1204-4D11v2-CD3 OKT3 Light Chain
<220>
<221> misc_feature
<222> (226)..(226)
<223> Xaa может быть любой аминокислотой
<400> 246
Gln Gly Gln Ser Gly Gln Cys Asn Ile Trp Leu Val Gly Gly Asp Cys
1 5 10 15
Arg Gly Trp Gln Gly Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Leu Ser Gly Arg Ser Asp Asn His Gly Gly Gly Ser Asp Ile Gln Met
35 40 45
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
50 55 60
Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn Trp Tyr
65 70 75 80
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser
85 90 95
Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
100 105 110
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
115 120 125
Thr Tyr Tyr Cys Gln Gln Thr Val Val Ala Pro Pro Leu Phe Gly Gln
130 135 140
Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe
145 150 155 160
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
165 170 175
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
180 185 190
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
195 200 205
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
210 215 220
Leu Xaa Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
225 230 235 240
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
245 250 255
Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly
260 265 270
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe
500 505 510
Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
515 520
<210> 247
<211> 2162
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-Jagged 4D11v2-анти-CTLA-4 антитела
<400> 247
gaggtgcacc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtgtcaagt attgacccgg aaggtcggca gacatattac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagacatc 300
ggcggcaggt cggcctttga ctactggggc cagggaaccc tggtcaccgt ctcctcagct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaagga ggtggatctg gaggtggcgg ttcaggctct 1380
ggcggaggct caggtggtgg aggatcaggc ggaggtgaaa ttgtgttgac acagtctcca 1440
ggcaccctgt ctttgtctcc aggggaaaga gccaccctct cctgcagggc cagtcagagt 1500
gttagcagca gctacttagc ctggtaccag cagaaacctg gccaggctcc caggctcctc 1560
atctatggtg catccagcag ggccactggc atcccagaca ggttcagtgg cagtgggtct 1620
gggacagact tcactctcac catcagcaga ctggagcctg aagattttgc agtgtattac 1680
tgtcagcagt atggtagctc accgctcact ttcggcggag ggaccaaggt ggaaatcaaa 1740
cgttccggag ggtcgaccat aacttcgtat aatgtatact atacgaagtt atcctcgagc 1800
ggtacccagg tgcagctggt gcagactggg ggaggcgtgg tccagcctgg gaggtccctg 1860
agactctcct gtgcagcctc tggatccacc tttagcagct atgccatgag ctgggtccgc 1920
caggctccag ggaaggggct ggagtgggtc tcagctatta gtggtagtgg tggtagcaca 1980
tactacgcag actccgtgaa gggccggttc accatctcca gagacaattc caagaacacg 2040
ctgtatctgc aaatgaacag cctgagagcc gaggacacgg ccgtatatta ctgtgcgaca 2100
aactcccttt actggtactt cgatctctgg ggccgtggca ccctggtcac tgtctcttca 2160
gc 2162
<210> 248
<211> 722
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-Jagged 4D11v2-анти-CTLA-4 антитела
<400> 248
Glu Val His Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Pro Glu Gly Arg Gln Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Ile Gly Gly Arg Ser Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Glu Ile Val Leu Thr Gln Ser Pro
465 470 475 480
Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg
485 490 495
Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala
515 520 525
Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe
530 535 540
Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile Thr Ser Tyr Asn Val
580 585 590
Tyr Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln Val Gln Leu Val Gln
595 600 605
Thr Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
610 615 620
Ala Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala Met Ser Trp Val Arg
625 630 635 640
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser Gly Ser
645 650 655
Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
660 665 670
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
675 680 685
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Thr Asn Ser Leu Tyr
690 695 700
Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
705 710 715 720
Ala Ser
<210> 249
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-Jagged 4D11v2-анти-CTLA-4 антитела
<220>
<221> misc_feature
<222> (545)..(545)
<223> n может быть любым нуклеотидом
<400> 249
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattagc agctatttaa attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgcg gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag acggttgtgg cgcctccgtt attcggccaa 300
gggaccaagg tggaaatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgancaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggaggtgg atctggaggt 660
ggcggttcag gctctggcgg aggctcaggt ggtggaggat caggcggagg tgaaattgtg 720
ttgacacagt ctccaggcac cctgtctttg tctccagggg aaagagccac cctctcctgc 780
agggccagtc agagtgttag cagcagctac ttagcctggt accagcagaa acctggccag 840
gctcccaggc tcctcatcta tggtgcatcc agcagggcca ctggcatccc agacaggttc 900
agtggcagtg ggtctgggac agacttcact ctcaccatca gcagactgga gcctgaagat 960
tttgcagtgt attactgtca gcagtatggt agctcaccgc tcactttcgg cggagggacc 1020
aaggtggaaa tcaaacgttc cggagggtcg accataactt cgtataatgt atactatacg 1080
aagttatcct cgagcggtac ccaggtgcag ctggtgcaga ctgggggagg cgtggtccag 1140
cctgggaggt ccctgagact ctcctgtgca gcctctggat ccacctttag cagctatgcc 1200
atgagctggg tccgccaggc tccagggaag gggctggagt gggtctcagc tattagtggt 1260
agtggtggta gcacatacta cgcagactcc gtgaagggcc ggttcaccat ctccagagac 1320
aattccaaga acacgctgta tctgcaaatg aacagcctga gagccgagga cacggccgta 1380
tattactgtg cgacaaactc cctttactgg tacttcgatc tctggggccg tggcaccctg 1440
gtcactgtct cttcagctag c 1461
<210> 250
<211> 487
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-Jagged 4D11v2-анти-CTLA-4 антитела
<220>
<221> misc_feature
<222> (182)..(182)
<223> Xaa может быть любой аминокислотой
<400> 250
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Val Val Ala Pro Pro
85 90 95
Leu Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Xaa Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Glu Ile Val
225 230 235 240
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
245 250 255
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
260 265 270
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly
275 280 285
Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
290 295 300
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
305 310 315 320
Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe
325 330 335
Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile
340 345 350
Thr Ser Tyr Asn Val Tyr Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln
355 360 365
Val Gln Leu Val Gln Thr Gly Gly Gly Val Val Gln Pro Gly Arg Ser
370 375 380
Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala
385 390 395 400
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
405 410 415
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
420 425 430
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
435 440 445
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
450 455 460
Thr Asn Ser Leu Tyr Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu
465 470 475 480
Val Thr Val Ser Ser Ala Ser
485
<210> 251
<211> 1593
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-Jagged-анти-CTLA-4 антитела 5342-1204-4D11v2-CTLA-4
<220>
<221> misc_feature
<222> (677)..(677)
<223> n может быть любым нуклеотидом
<400> 251
caaggccagt ctggccagtg caatatttgg ctcgtaggtg gtgattgcag gggctggcag 60
gggggctcga gcggtggcag cggtggctct ggtggtctga gcggccgttc cgataatcat 120
ggcggcggtt ctgacatcca gatgacccag tctccatcct ccctgtctgc atctgtagga 180
gacagagtca ccatcacttg ccgggcaagt cagagcatta gcagctattt aaattggtat 240
cagcagaaac cagggaaagc ccctaagctc ctgatctatg cggcatccag tttgcaaagt 300
ggggtcccat caaggttcag tggcagtgga tctgggacag atttcactct caccatcagc 360
agtctgcaac ctgaagattt tgcaacttac tactgtcaac agacggttgt ggcgcctccg 420
ttattcggcc aagggaccaa ggtggaaatc aaacgtacgg tggctgcacc atctgtcttc 480
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 540
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 600
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 660
agcaccctga cgctgancaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 720
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgtggaggt 780
ggatctggag gtggcggttc aggctctggc ggaggctcag gtggtggagg atcaggcgga 840
ggtgaaattg tgttgacaca gtctccaggc accctgtctt tgtctccagg ggaaagagcc 900
accctctcct gcagggccag tcagagtgtt agcagcagct acttagcctg gtaccagcag 960
aaacctggcc aggctcccag gctcctcatc tatggtgcat ccagcagggc cactggcatc 1020
ccagacaggt tcagtggcag tgggtctggg acagacttca ctctcaccat cagcagactg 1080
gagcctgaag attttgcagt gtattactgt cagcagtatg gtagctcacc gctcactttc 1140
ggcggaggga ccaaggtgga aatcaaacgt tccggagggt cgaccataac ttcgtataat 1200
gtatactata cgaagttatc ctcgagcggt acccaggtgc agctggtgca gactggggga 1260
ggcgtggtcc agcctgggag gtccctgaga ctctcctgtg cagcctctgg atccaccttt 1320
agcagctatg ccatgagctg ggtccgccag gctccaggga aggggctgga gtgggtctca 1380
gctattagtg gtagtggtgg tagcacatac tacgcagact ccgtgaaggg ccggttcacc 1440
atctccagag acaattccaa gaacacgctg tatctgcaaa tgaacagcct gagagccgag 1500
gacacggccg tatattactg tgcgacaaac tccctttact ggtacttcga tctctggggc 1560
cgtggcaccc tggtcactgt ctcttcagct agc 1593
<210> 252
<211> 531
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-Jagged-анти-CTLA-4 антитела 5342-1204-4D11v2-CTLA-4
<220>
<221> misc_feature
<222> (226)..(226)
<223> Xaa может быть любой аминокислотой
<400> 252
Gln Gly Gln Ser Gly Gln Cys Asn Ile Trp Leu Val Gly Gly Asp Cys
1 5 10 15
Arg Gly Trp Gln Gly Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Leu Ser Gly Arg Ser Asp Asn His Gly Gly Gly Ser Asp Ile Gln Met
35 40 45
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
50 55 60
Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn Trp Tyr
65 70 75 80
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser
85 90 95
Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
100 105 110
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
115 120 125
Thr Tyr Tyr Cys Gln Gln Thr Val Val Ala Pro Pro Leu Phe Gly Gln
130 135 140
Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe
145 150 155 160
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
165 170 175
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
180 185 190
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
195 200 205
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
210 215 220
Leu Xaa Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
225 230 235 240
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
245 250 255
Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly
260 265 270
Ser Gly Gly Gly Gly Ser Gly Gly Gly Glu Ile Val Leu Thr Gln Ser
275 280 285
Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys
290 295 300
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln
305 310 315 320
Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg
325 330 335
Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
340 345 350
Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr
355 360 365
Tyr Cys Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe Gly Gly Gly Thr
370 375 380
Lys Val Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile Thr Ser Tyr Asn
385 390 395 400
Val Tyr Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln Val Gln Leu Val
405 410 415
Gln Thr Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser
420 425 430
Cys Ala Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala Met Ser Trp Val
435 440 445
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser Gly
450 455 460
Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
465 470 475 480
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
485 490 495
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Thr Asn Ser Leu
500 505 510
Tyr Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser
515 520 525
Ser Ala Ser
530
<210> 253
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v5
<400> 253
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgaactg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaatga 1350
<210> 254
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v5
<400> 254
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 255
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v5
<400> 255
cagatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 256
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v5
<400> 256
Gln Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 257
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v5
<400> 257
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag caacgatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagcgcgt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgaactg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaa 1347
<210> 258
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v5
<400> 258
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 259
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v5
<400> 259
ggcggtaccc agatcttgct gacccagagc ccggtgattc tgagcgtgag cccgggcgaa 60
cgtgtgagct ttagctgccg cgcgagccag agcattggca ccaacattca ttggtatcag 120
cagcgcacca acggcagccc gcgcctgctg attaaatatg cgagcgaaag cattagcggc 180
attccgagcc gctttagcgg cagcggcagc ggcaccgatt ttaccctgag cattaacagc 240
gtggaaagcg aagatattgc ggattattat tgccagcaga acaacaactg gccgaccacc 300
tttggcgcgg gcaccaaact ggaactgaaa cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagcg 648
<210> 260
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v5
<400> 260
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 261
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v4
<400> 261
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag caacgatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgaactg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaatga 1350
<210> 262
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v4
<400> 262
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 263
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v4
<400> 263
cagatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 264
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v4
<400> 264
Gln Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 265
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v6
<400> 265
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacgccagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgaactg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaatga 1350
<210> 266
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь анти-EGFR антитела C225v6
<400> 266
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 267
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v6
<400> 267
cagatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 268
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь анти-EGFR антитела C225v6
<400> 268
Gln Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 269
<211> 786
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь активируемого анти-EGFR антитела 3954-1204-C225v5
<400> 269
caaggccagt ctggccagtg catctcacct cgtggttgtc cggacggccc atacgtcatg 60
tacggctcga gcggtggcag cggtggctct ggtggatccg gtctgagcgg ccgttccgat 120
aatcatggca gtagcggtac ccagatcttg ctgacccaga gcccggtgat tctgagcgtg 180
agcccgggcg aacgtgtgag ctttagctgc cgcgcgagcc agagcattgg caccaacatt 240
cattggtatc agcagcgcac caacggcagc ccgcgcctgc tgattaaata tgcgagcgaa 300
agcattagcg gcattccgag ccgctttagc ggcagcggca gcggcaccga ttttaccctg 360
agcattaaca gcgtggaaag cgaagatatt gcggattatt attgccagca gaacaacaac 420
tggccgacca cctttggcgc gggcaccaaa ctggaactga aacgtacggt ggctgcacca 480
tctgtcttca tcttcccgcc atctgatgag cagttgaaat ctggaactgc ctctgttgtg 540
tgcctgctga ataacttcta tcccagagag gccaaagtac agtggaaggt ggataacgcc 600
ctccaatcgg gtaactccca ggagagtgtc acagagcagg acagcaagga cagcacctac 660
agcctcagca gcaccctgac gctgagcaaa gcagactacg agaaacacaa agtctacgcc 720
tgcgaagtca cccatcaggg cctgagctcg cccgtcacaa agagcttcaa caggggagag 780
tgttag 786
<210> 270
<211> 261
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь активируемого анти-EGFR антитела 3954-1204-C225v5
<400> 270
Gln Gly Gln Ser Gly Gln Cys Ile Ser Pro Arg Gly Cys Pro Asp Gly
1 5 10 15
Pro Tyr Val Met Tyr Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Ser Gly Leu Ser Gly Arg Ser Asp Asn His Gly Ser Ser Gly Thr Gln
35 40 45
Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly Glu
50 55 60
Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn Ile
65 70 75 80
His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile Lys
85 90 95
Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly Ser
100 105 110
Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser Glu
115 120 125
Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr Thr
130 135 140
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
145 150 155 160
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
165 170 175
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
180 185 190
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
195 200 205
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
210 215 220
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
225 230 235 240
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
245 250 255
Asn Arg Gly Glu Cys
260
<210> 271
<211> 2139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-EGFR антитела C225v5 - анти-CD3 антитела OKT3
<400> 271
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgaactg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaagga ggtggatctg gaggtggcgg ttcaggctct 1380
ggcggaggct caggtggtgg aggatcaggc ggaggtcagg ttcagctgca gcagagcggt 1440
gcagaactgg cacgtccggg tgcaagcgtt aaaatgagct gtaaagcaag cggttatacc 1500
tttacccgtt ataccatgca ttgggttaaa cagcgtccgg gtcagggtct ggaatggatt 1560
ggttatatca atccgagccg tggttatacc aactacaacc agaaattcaa agataaagca 1620
accctgacca ccgataaaag cagcagcacc gcctatatgc agctgagcag cctgacctca 1680
gaggatagcg cagtttatta ctgtgcacgc tattatgatg atcactattg cctggattat 1740
tggggtcagg gcaccaccct gaccgttagc agcggtggtg gtggtagtgg tggcggtggt 1800
tcaggcggtg gcggtagcca gattgttctg acccagagtc cggcaattat gagcgcaagt 1860
ccgggtgaaa aagttaccat gacctgtagc gccagcagca gcgttagcta tatgaattgg 1920
tatcagcaga aaagcggcac cagcccgaaa cgttggattt atgataccag caaactggca 1980
agcggtgttc cggcacattt tcgtggtagc ggtagcggca cctcatatag cctgaccatt 2040
agcggtatgg aagcagaaga tgcagcaacc tattattgtc agcagtggtc aagcaatccg 2100
tttacctttg gtagtggcac caaactggaa attaatcgt 2139
<210> 272
<211> 713
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-EGFR антитела C225v5 - анти-CD3 антитела OKT3
<400> 272
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser Gly
465 470 475 480
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala
485 490 495
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
500 505 510
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
515 520 525
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
530 535 540
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
545 550 555 560
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
565 570 575
Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
580 585 590
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile
595 600 605
Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
610 615 620
Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
625 630 635 640
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
645 650 655
Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser
660 665 670
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala
675 680 685
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly
690 695 700
Ser Gly Thr Lys Leu Glu Ile Asn Arg
705 710
<210> 273
<211> 1434
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-EGFR антитела C225v5 - анти-CD3 антитела OKT3
<400> 273
cagatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggaggtgg atctggaggt 660
ggcggttcag gctctggcgg aggctcaggt ggtggaggat caggcggagg tcaggttcag 720
ctgcagcaga gcggtgcaga actggcacgt ccgggtgcaa gcgttaaaat gagctgtaaa 780
gcaagcggtt atacctttac ccgttatacc atgcattggg ttaaacagcg tccgggtcag 840
ggtctggaat ggattggtta tatcaatccg agccgtggtt ataccaacta caaccagaaa 900
ttcaaagata aagcaaccct gaccaccgat aaaagcagca gcaccgccta tatgcagctg 960
agcagcctga cctcagagga tagcgcagtt tattactgtg cacgctatta tgatgatcac 1020
tattgcctgg attattgggg tcagggcacc accctgaccg ttagcagcgg tggtggtggt 1080
agtggtggcg gtggttcagg cggtggcggt agccagattg ttctgaccca gagtccggca 1140
attatgagcg caagtccggg tgaaaaagtt accatgacct gtagcgccag cagcagcgtt 1200
agctatatga attggtatca gcagaaaagc ggcaccagcc cgaaacgttg gatttatgat 1260
accagcaaac tggcaagcgg tgttccggca cattttcgtg gtagcggtag cggcacctca 1320
tatagcctga ccattagcgg tatggaagca gaagatgcag caacctatta ttgtcagcag 1380
tggtcaagca atccgtttac ctttggtagt ggcaccaaac tggaaattaa tcgt 1434
<210> 274
<211> 478
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-EGFR антитела C225v5 - анти-CD3 антитела OKT3
<400> 274
Gln Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln
225 230 235 240
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
245 250 255
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His
260 265 270
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile
275 280 285
Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
290 295 300
Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
305 310 315 320
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr
325 330 335
Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
340 345 350
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
355 360 365
Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala
370 375 380
Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val
385 390 395 400
Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg
405 410 415
Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe
420 425 430
Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met
435 440 445
Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn
450 455 460
Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
465 470 475
<210> 275
<211> 1575
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-EGFR - анти-CD3 антитела 3954-1204-C225v5-OKT3
<400> 275
caaggccagt ctggccagtg catctcacct cgtggttgtc cggacggccc atacgtcatg 60
tacggctcga gcggtggcag cggtggctct ggtggatccg gtctgagcgg ccgttccgat 120
aatcatggca gtagcggtac ccagatcttg ctgacccaga gcccggtgat tctgagcgtg 180
agcccgggcg aacgtgtgag ctttagctgc cgcgcgagcc agagcattgg caccaacatt 240
cattggtatc agcagcgcac caacggcagc ccgcgcctgc tgattaaata tgcgagcgaa 300
agcattagcg gcattccgag ccgctttagc ggcagcggca gcggcaccga ttttaccctg 360
agcattaaca gcgtggaaag cgaagatatt gcggattatt attgccagca gaacaacaac 420
tggccgacca cctttggcgc gggcaccaaa ctggaactga aacgtacggt ggctgcacca 480
tctgtcttca tcttcccgcc atctgatgag cagttgaaat ctggaactgc ctctgttgtg 540
tgcctgctga ataacttcta tcccagagag gccaaagtac agtggaaggt ggataacgcc 600
ctccaatcgg gtaactccca ggagagtgtc acagagcagg acagcaagga cagcacctac 660
agcctcagca gcaccctgac gctgagcaaa gcagactacg agaaacacaa agtctacgcc 720
tgcgaagtca cccatcaggg cctgagctcg cccgtcacaa agagcttcaa caggggagag 780
tgtggaggtg gatctggagg tggcggttca ggctctggcg gaggctcagg tggtggagga 840
tcaggcggag gtcaggttca gctgcagcag agcggtgcag aactggcacg tccgggtgca 900
agcgttaaaa tgagctgtaa agcaagcggt tataccttta cccgttatac catgcattgg 960
gttaaacagc gtccgggtca gggtctggaa tggattggtt atatcaatcc gagccgtggt 1020
tataccaact acaaccagaa attcaaagat aaagcaaccc tgaccaccga taaaagcagc 1080
agcaccgcct atatgcagct gagcagcctg acctcagagg atagcgcagt ttattactgt 1140
gcacgctatt atgatgatca ctattgcctg gattattggg gtcagggcac caccctgacc 1200
gttagcagcg gtggtggtgg tagtggtggc ggtggttcag gcggtggcgg tagccagatt 1260
gttctgaccc agagtccggc aattatgagc gcaagtccgg gtgaaaaagt taccatgacc 1320
tgtagcgcca gcagcagcgt tagctatatg aattggtatc agcagaaaag cggcaccagc 1380
ccgaaacgtt ggatttatga taccagcaaa ctggcaagcg gtgttccggc acattttcgt 1440
ggtagcggta gcggcacctc atatagcctg accattagcg gtatggaagc agaagatgca 1500
gcaacctatt attgtcagca gtggtcaagc aatccgttta cctttggtag tggcaccaaa 1560
ctggaaatta atcgt 1575
<210> 276
<211> 525
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-EGFR - анти-CD3 антитела 3954-1204-C225v5-OKT3
<400> 276
Gln Gly Gln Ser Gly Gln Cys Ile Ser Pro Arg Gly Cys Pro Asp Gly
1 5 10 15
Pro Tyr Val Met Tyr Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Ser Gly Leu Ser Gly Arg Ser Asp Asn His Gly Ser Ser Gly Thr Gln
35 40 45
Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly Glu
50 55 60
Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn Ile
65 70 75 80
His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile Lys
85 90 95
Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly Ser
100 105 110
Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser Glu
115 120 125
Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr Thr
130 135 140
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
145 150 155 160
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
165 170 175
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
180 185 190
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
195 200 205
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
210 215 220
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
225 230 235 240
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
245 250 255
Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser
260 265 270
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu
275 280 285
Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met
290 295 300
Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp
305 310 315 320
Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn
325 330 335
Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala
340 345 350
Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser
355 360 365
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr
370 375 380
Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
385 390 395 400
Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
405 410 415
Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser
420 425 430
Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser
435 440 445
Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp
450 455 460
Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg
465 470 475 480
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu
485 490 495
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro
500 505 510
Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
515 520 525
<210> 277
<211> 2166
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-EGFR C225v5-анти-CTLA-4 антитела
<400> 277
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgaactg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaagga ggtggatctg gaggtggcgg ttcaggctct 1380
ggcggaggct caggtggtgg aggatcaggc ggaggtgaaa ttgtgttgac acagtctcca 1440
ggcaccctgt ctttgtctcc aggggaaaga gccaccctct cctgcagggc cagtcagagt 1500
gttagcagca gctacttagc ctggtaccag cagaaacctg gccaggctcc caggctcctc 1560
atctatggtg catccagcag ggccactggc atcccagaca ggttcagtgg cagtgggtct 1620
gggacagact tcactctcac catcagcaga ctggagcctg aagattttgc agtgtattac 1680
tgtcagcagt atggtagctc accgctcact ttcggcggag ggaccaaggt ggaaatcaaa 1740
cgttccggag ggtcgaccat aacttcgtat aatgtatact atacgaagtt atcctcgagc 1800
ggtacccagg tgcagctggt gcagactggg ggaggcgtgg tccagcctgg gaggtccctg 1860
agactctcct gtgcagcctc tggatccacc tttagcagct atgccatgag ctgggtccgc 1920
caggctccag ggaaggggct ggagtgggtc tcagctatta gtggtagtgg tggtagcaca 1980
tactacgcag actccgtgaa gggccggttc accatctcca gagacaattc caagaacacg 2040
ctgtatctgc aaatgaacag cctgagagcc gaggacacgg ccgtatatta ctgtgcgaca 2100
aactcccttt actggtactt cgatctctgg ggccgtggca ccctggtcac tgtctcttca 2160
gctagc 2166
<210> 278
<211> 722
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического анти-EGFR C225v5-анти-CTLA-4 антитела
<400> 278
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Glu Ile Val Leu Thr Gln Ser Pro
465 470 475 480
Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg
485 490 495
Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala
515 520 525
Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe
530 535 540
Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile Thr Ser Tyr Asn Val
580 585 590
Tyr Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln Val Gln Leu Val Gln
595 600 605
Thr Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
610 615 620
Ala Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala Met Ser Trp Val Arg
625 630 635 640
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser Gly Ser
645 650 655
Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
660 665 670
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
675 680 685
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Thr Asn Ser Leu Tyr
690 695 700
Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
705 710 715 720
Ala Ser
<210> 279
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-EGFR C225v5-анти-CTLA-4 антитела
<400> 279
cagatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggaggtgg atctggaggt 660
ggcggttcag gctctggcgg aggctcaggt ggtggaggat caggcggagg tgaaattgtg 720
ttgacacagt ctccaggcac cctgtctttg tctccagggg aaagagccac cctctcctgc 780
agggccagtc agagtgttag cagcagctac ttagcctggt accagcagaa acctggccag 840
gctcccaggc tcctcatcta tggtgcatcc agcagggcca ctggcatccc agacaggttc 900
agtggcagtg ggtctgggac agacttcact ctcaccatca gcagactgga gcctgaagat 960
tttgcagtgt attactgtca gcagtatggt agctcaccgc tcactttcgg cggagggacc 1020
aaggtggaaa tcaaacgttc cggagggtcg accataactt cgtataatgt atactatacg 1080
aagttatcct cgagcggtac ccaggtgcag ctggtgcaga ctgggggagg cgtggtccag 1140
cctgggaggt ccctgagact ctcctgtgca gcctctggat ccacctttag cagctatgcc 1200
atgagctggg tccgccaggc tccagggaag gggctggagt gggtctcagc tattagtggt 1260
agtggtggta gcacatacta cgcagactcc gtgaagggcc ggttcaccat ctccagagac 1320
aattccaaga acacgctgta tctgcaaatg aacagcctga gagccgagga cacggccgta 1380
tattactgtg cgacaaactc cctttactgg tacttcgatc tctggggccg tggcaccctg 1440
gtcactgtct cttcagctag c 1461
<210> 280
<211> 487
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического анти-EGFR C225v5-анти-CTLA-4 антитела
<400> 280
Gln Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Glu Ile Val
225 230 235 240
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
245 250 255
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
260 265 270
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly
275 280 285
Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
290 295 300
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
305 310 315 320
Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe
325 330 335
Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile
340 345 350
Thr Ser Tyr Asn Val Tyr Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln
355 360 365
Val Gln Leu Val Gln Thr Gly Gly Gly Val Val Gln Pro Gly Arg Ser
370 375 380
Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala
385 390 395 400
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
405 410 415
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
420 425 430
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
435 440 445
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
450 455 460
Thr Asn Ser Leu Tyr Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu
465 470 475 480
Val Thr Val Ser Ser Ala Ser
485
<210> 281
<211> 1602
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-EGFR - анти-CTLA-4 антитела 3954-1204-C225v5-CTLA-4
<400> 281
caaggccagt ctggccagtg catctcacct cgtggttgtc cggacggccc atacgtcatg 60
tacggctcga gcggtggcag cggtggctct ggtggatccg gtctgagcgg ccgttccgat 120
aatcatggca gtagcggtac ccagatcttg ctgacccaga gcccggtgat tctgagcgtg 180
agcccgggcg aacgtgtgag ctttagctgc cgcgcgagcc agagcattgg caccaacatt 240
cattggtatc agcagcgcac caacggcagc ccgcgcctgc tgattaaata tgcgagcgaa 300
agcattagcg gcattccgag ccgctttagc ggcagcggca gcggcaccga ttttaccctg 360
agcattaaca gcgtggaaag cgaagatatt gcggattatt attgccagca gaacaacaac 420
tggccgacca cctttggcgc gggcaccaaa ctggaactga aacgtacggt ggctgcacca 480
tctgtcttca tcttcccgcc atctgatgag cagttgaaat ctggaactgc ctctgttgtg 540
tgcctgctga ataacttcta tcccagagag gccaaagtac agtggaaggt ggataacgcc 600
ctccaatcgg gtaactccca ggagagtgtc acagagcagg acagcaagga cagcacctac 660
agcctcagca gcaccctgac gctgagcaaa gcagactacg agaaacacaa agtctacgcc 720
tgcgaagtca cccatcaggg cctgagctcg cccgtcacaa agagcttcaa caggggagag 780
tgtggaggtg gatctggagg tggcggttca ggctctggcg gaggctcagg tggtggagga 840
tcaggcggag gtgaaattgt gttgacacag tctccaggca ccctgtcttt gtctccaggg 900
gaaagagcca ccctctcctg cagggccagt cagagtgtta gcagcagcta cttagcctgg 960
taccagcaga aacctggcca ggctcccagg ctcctcatct atggtgcatc cagcagggcc 1020
actggcatcc cagacaggtt cagtggcagt gggtctggga cagacttcac tctcaccatc 1080
agcagactgg agcctgaaga ttttgcagtg tattactgtc agcagtatgg tagctcaccg 1140
ctcactttcg gcggagggac caaggtggaa atcaaacgtt ccggagggtc gaccataact 1200
tcgtataatg tatactatac gaagttatcc tcgagcggta cccaggtgca gctggtgcag 1260
actgggggag gcgtggtcca gcctgggagg tccctgagac tctcctgtgc agcctctgga 1320
tccaccttta gcagctatgc catgagctgg gtccgccagg ctccagggaa ggggctggag 1380
tgggtctcag ctattagtgg tagtggtggt agcacatact acgcagactc cgtgaagggc 1440
cggttcacca tctccagaga caattccaag aacacgctgt atctgcaaat gaacagcctg 1500
agagccgagg acacggccgt atattactgt gcgacaaact ccctttactg gtacttcgat 1560
ctctggggcc gtggcaccct ggtcactgtc tcttcagcta gc 1602
<210> 282
<211> 534
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического активируемого анти-EGFR - анти-CTLA-4 антитела 3954-1204-C225v5-CTLA-4
<400> 282
Gln Gly Gln Ser Gly Gln Cys Ile Ser Pro Arg Gly Cys Pro Asp Gly
1 5 10 15
Pro Tyr Val Met Tyr Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Ser Gly Leu Ser Gly Arg Ser Asp Asn His Gly Ser Ser Gly Thr Gln
35 40 45
Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly Glu
50 55 60
Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn Ile
65 70 75 80
His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile Lys
85 90 95
Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly Ser
100 105 110
Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser Glu
115 120 125
Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr Thr
130 135 140
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
145 150 155 160
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
165 170 175
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
180 185 190
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
195 200 205
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
210 215 220
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
225 230 235 240
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
245 250 255
Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser
260 265 270
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Glu Ile Val Leu
275 280 285
Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr
290 295 300
Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp
305 310 315 320
Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala
325 330 335
Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser
340 345 350
Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe
355 360 365
Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro Leu Thr Phe Gly
370 375 380
Gly Gly Thr Lys Val Glu Ile Lys Arg Ser Gly Gly Ser Thr Ile Thr
385 390 395 400
Ser Tyr Asn Val Tyr Tyr Thr Lys Leu Ser Ser Ser Gly Thr Gln Val
405 410 415
Gln Leu Val Gln Thr Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu
420 425 430
Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Phe Ser Ser Tyr Ala Met
435 440 445
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
450 455 460
Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
465 470 475 480
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
485 490 495
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Thr
500 505 510
Asn Ser Leu Tyr Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val
515 520 525
Thr Val Ser Ser Ala Ser
530
<210> 283
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, кодирующая спейсерный пептид
<400> 283
caaggccagt ctggccag 18
<210> 284
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, кодирующая маскирующий пептид
<400> 284
tgcatctcac ctcgtggttg tccggacggc ccatacgtca tgtac 45
<210> 285
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, кодирующая линкерный пептид
<400> 285
ggctcgagcg gtggcagcgg tggctctggt ggatccggt 39
<210> 286
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 1204 Substrate coding sequence
<400> 286
ctgagcggcc gttccgataa tcat 24
<210> 287
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, кодирующая линкерный пептид
<400> 287
ggcagtagcg gtacc 15
<210> 288
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкерный пептид
<400> 288
Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
1 5 10
<210> 289
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкерный пептид
<400> 289
Gly Ser Ser Gly Thr
1 5
<210> 290
<211> 264
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3m с длинным линкером
<400> 290
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser Gly Ala
20 25 30
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
50 55 60
Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
65 70 75 80
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
85 90 95
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
100 105 110
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
115 120 125
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Val
145 150 155 160
Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val
165 170 175
Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr
180 185 190
Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser
195 200 205
Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser Gly
210 215 220
Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala Ala
225 230 235 240
Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Ser
245 250 255
Gly Thr Lys Leu Glu Ile Asn Arg
260
<210> 291
<211> 795
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3m с длинным линкером
<400> 291
ggaggtggat ctggaggtgg cggttcaggc tctggcggag gctcaggtgg tggaggatca 60
ggcggaggtc aggttcagct gcagcagagc ggtgcagaac tggcacgtcc gggtgcaagc 120
gttaaaatga gctgtaaagc aagcggttat acctttaccc gttataccat gcattgggtt 180
aaacagcgtc cgggtcaggg tctggaatgg attggttata tcaatccgag ccgtggttat 240
accaactaca accagaaatt caaagataaa gcaaccctga ccaccgataa aagcagcagc 300
accgcctata tgcagctgag cagcctgacc tcagaggata gcgcagttta ttactgtgca 360
cgctattatg atgatcacta tagcctggat tattggggtc agggcaccac cctgaccgtt 420
agcagcggtg gtggtggtag tggtggcggt ggttcaggcg gtggcggtag ccagattgtt 480
ctgacccaga gtccggcaat tatgagcgca agtccgggtg aaaaagttac catgacctgt 540
agcgccagca gcagcgttag ctatatgaat tggtatcagc agaaaagcgg caccagcccg 600
aaacgttgga tttatgatac cagcaaactg gcaagcggtg ttccggcaca ttttcgtggt 660
agcggtagcg gcacctcata tagcctgacc attagcggta tggaagcaga agatgcagca 720
acctattatt gtcagcagtg gtcaagcaat ccgtttacct ttggtagtgg caccaaactg 780
gaaattaatc gttga 795
<210> 292
<211> 246
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3m с коротким линкером
<400> 292
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
130 135 140
Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser
145 150 155 160
Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser
165 170 175
Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
180 185 190
Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
195 200 205
Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
210 215 220
Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
225 230 235 240
Arg Gly Gly Gly Gly Ser
245
<210> 293
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3m с коротким линкером
<400> 293
caggttcagc tgcagcagag cggtgcagaa ctggcacgtc cgggtgcaag cgttaaaatg 60
agctgtaaag caagcggtta tacctttacc cgttatacca tgcattgggt taaacagcgt 120
ccgggtcagg gtctggaatg gattggttat atcaatccga gccgtggtta taccaactac 180
aaccagaaat tcaaagataa agcaaccctg accaccgata aaagcagcag caccgcctat 240
atgcagctga gcagcctgac ctcagaggat agcgcagttt attactgtgc acgctattat 300
gatgatcact atagcctgga ttattggggt cagggcacca ccctgaccgt tagcagcggt 360
ggtggtggta gtggtggcgg tggttcaggc ggtggcggta gccagattgt tctgacccag 420
agtccggcaa ttatgagcgc aagtccgggt gaaaaagtta ccatgacctg tagcgccagc 480
agcagcgtta gctatatgaa ttggtatcag cagaaaagcg gcaccagccc gaaacgttgg 540
atttatgata ccagcaaact ggcaagcggt gttccggcac attttcgtgg tagcggtagc 600
ggcacctcat atagcctgac cattagcggt atggaagcag aagatgcagc aacctattat 660
tgtcagcagt ggtcaagcaa tccgtttacc tttggtagtg gcaccaaact ggaaattaat 720
cgtggaggtg gtggatcc 738
<210> 294
<211> 695
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5-OKT3m-H-N
<400> 294
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
130 135 140
Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser
145 150 155 160
Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser
165 170 175
Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
180 185 190
Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
195 200 205
Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
210 215 220
Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
225 230 235 240
Arg Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly
245 250 255
Leu Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly
260 265 270
Phe Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly
275 280 285
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp
290 295 300
Tyr Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser
305 310 315 320
Lys Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Gln Asp Thr
325 330 335
Ala Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe
340 345 350
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Ala Ser Thr
355 360 365
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
370 375 380
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
385 390 395 400
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
405 410 415
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
420 425 430
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
435 440 445
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
450 455 460
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
465 470 475 480
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
485 490 495
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
500 505 510
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
515 520 525
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
530 535 540
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
545 550 555 560
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
565 570 575
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
580 585 590
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
595 600 605
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
610 615 620
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
625 630 635 640
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
645 650 655
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
660 665 670
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
675 680 685
Leu Ser Leu Ser Pro Gly Lys
690 695
<210> 295
<211> 2088
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5-OKT3m-H-N
<400> 295
caggttcagc tgcagcagag cggtgcagaa ctggcacgtc cgggtgcaag cgttaaaatg 60
agctgtaaag caagcggtta tacctttacc cgttatacca tgcattgggt taaacagcgt 120
ccgggtcagg gtctggaatg gattggttat atcaatccga gccgtggtta taccaactac 180
aaccagaaat tcaaagataa agcaaccctg accaccgata aaagcagcag caccgcctat 240
atgcagctga gcagcctgac ctcagaggat agcgcagttt attactgtgc acgctattat 300
gatgatcact atagcctgga ttattggggt cagggcacca ccctgaccgt tagcagcggt 360
ggtggtggta gtggtggcgg tggttcaggc ggtggcggta gccagattgt tctgacccag 420
agtccggcaa ttatgagcgc aagtccgggt gaaaaagtta ccatgacctg tagcgccagc 480
agcagcgtta gctatatgaa ttggtatcag cagaaaagcg gcaccagccc gaaacgttgg 540
atttatgata ccagcaaact ggcaagcggt gttccggcac attttcgtgg tagcggtagc 600
ggcacctcat atagcctgac cattagcggt atggaagcag aagatgcagc aacctattat 660
tgtcagcagt ggtcaagcaa tccgtttacc tttggtagtg gcaccaaact ggaaattaat 720
cgtggaggtg gtggatccca ggtgcagctg aaacagagcg gcccgggcct ggtgcagccg 780
agccagagcc tgagcattac ctgcaccgtg agcggcttta gcctgaccaa ctatggcgtg 840
cattgggtgc gccagagccc gggcaaaggc ctggaatggc tgggcgtgat ttggagcggc 900
ggcaacaccg attataacac cccgtttacc agccgcctga gcattaacaa agataacagc 960
aaaagccagg tgttttttaa aatgaacagc ctgcaaagcc aggataccgc gatttattat 1020
tgcgcgcgcg cgctgaccta ttatgattat gaatttgcgt attggggcca gggcaccctg 1080
gtgaccgtga gcgcggctag caccaagggc ccatcggtct tccccctggc accctcctcc 1140
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 1200
ccggtgacgg tgtcgtggaa ctcaggcgcc ctgaccagcg gcgtgcacac cttcccggct 1260
gtcctacagt cctcaggact ctactccctc agcagcgtgg tgaccgtgcc ctccagcagc 1320
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtggac 1380
aagaaagttg agcccaaatc ttgtgacaaa actcacacat gcccaccgtg cccagcacct 1440
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 1500
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 1560
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 1620
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 1680
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 1740
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 1800
ccatcccggg aggagatgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 1860
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 1920
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 1980
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 2040
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gtaaatga 2088
<210> 296
<211> 713
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5-OKT3m-H-С
<400> 296
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser Gly
465 470 475 480
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala
485 490 495
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
500 505 510
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
515 520 525
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
530 535 540
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
545 550 555 560
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
565 570 575
Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
580 585 590
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile
595 600 605
Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
610 615 620
Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
625 630 635 640
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
645 650 655
Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser
660 665 670
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala
675 680 685
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly
690 695 700
Ser Gly Thr Lys Leu Glu Ile Asn Arg
705 710
<210> 297
<211> 2142
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5-OKT3m-H-С
<400> 297
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaagga ggtggatctg gaggtggcgg ttcaggctct 1380
ggcggaggct caggtggtgg aggatcaggc ggaggtcagg ttcagctgca gcagagcggt 1440
gcagaactgg cacgtccggg tgcaagcgtt aaaatgagct gtaaagcaag cggttatacc 1500
tttacccgtt ataccatgca ttgggttaaa cagcgtccgg gtcagggtct ggaatggatt 1560
ggttatatca atccgagccg tggttatacc aactacaacc agaaattcaa agataaagca 1620
accctgacca ccgataaaag cagcagcacc gcctatatgc agctgagcag cctgacctca 1680
gaggatagcg cagtttatta ctgtgcacgc tattatgatg atcactatag cctggattat 1740
tggggtcagg gcaccaccct gaccgttagc agcggtggtg gtggtagtgg tggcggtggt 1800
tcaggcggtg gcggtagcca gattgttctg acccagagtc cggcaattat gagcgcaagt 1860
ccgggtgaaa aagttaccat gacctgtagc gccagcagca gcgttagcta tatgaattgg 1920
tatcagcaga aaagcggcac cagcccgaaa cgttggattt atgataccag caaactggca 1980
agcggtgttc cggcacattt tcgtggtagc ggtagcggca cctcatatag cctgaccatt 2040
agcggtatgg aagcagaaga tgcagcaacc tattattgtc agcagtggtc aagcaatccg 2100
tttacctttg gtagtggcac caaactggaa attaatcgtt ga 2142
<210> 298
<211> 478
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического антитела C225v5-OKT3m-L-C
<400> 298
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln
225 230 235 240
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
245 250 255
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His
260 265 270
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile
275 280 285
Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
290 295 300
Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
305 310 315 320
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr
325 330 335
Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
340 345 350
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
355 360 365
Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala
370 375 380
Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val
385 390 395 400
Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg
405 410 415
Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe
420 425 430
Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met
435 440 445
Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn
450 455 460
Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
465 470 475
<210> 299
<211> 1437
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мультиспецифического антитела C225v5-OKT3m-L-C
<400> 299
gacatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggaggtgg atctggaggt 660
ggcggttcag gctctggcgg aggctcaggt ggtggaggat caggcggagg tcaggttcag 720
ctgcagcaga gcggtgcaga actggcacgt ccgggtgcaa gcgttaaaat gagctgtaaa 780
gcaagcggtt atacctttac ccgttatacc atgcattggg ttaaacagcg tccgggtcag 840
ggtctggaat ggattggtta tatcaatccg agccgtggtt ataccaacta caaccagaaa 900
ttcaaagata aagcaaccct gaccaccgat aaaagcagca gcaccgccta tatgcagctg 960
agcagcctga cctcagagga tagcgcagtt tattactgtg cacgctatta tgatgatcac 1020
tatagcctgg attattgggg tcagggcacc accctgaccg ttagcagcgg tggtggtggt 1080
agtggtggcg gtggttcagg cggtggcggt agccagattg ttctgaccca gagtccggca 1140
attatgagcg caagtccggg tgaaaaagtt accatgacct gtagcgccag cagcagcgtt 1200
agctatatga attggtatca gcagaaaagc ggcaccagcc cgaaacgttg gatttatgat 1260
accagcaaac tggcaagcgg tgttccggca cattttcgtg gtagcggtag cggcacctca 1320
tatagcctga ccattagcgg tatggaagca gaagatgcag caacctatta ttgtcagcag 1380
tggtcaagca atccgtttac ctttggtagt ggcaccaaac tggaaattaa tcgttag 1437
<210> 300
<211> 695
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела Multispecific C225v5 N297Q - OKT3m-H-N
<400> 300
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
130 135 140
Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser
145 150 155 160
Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser
165 170 175
Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
180 185 190
Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
195 200 205
Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
210 215 220
Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
225 230 235 240
Arg Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly
245 250 255
Leu Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly
260 265 270
Phe Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly
275 280 285
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp
290 295 300
Tyr Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser
305 310 315 320
Lys Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Gln Asp Thr
325 330 335
Ala Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe
340 345 350
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Ala Ser Thr
355 360 365
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
370 375 380
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
385 390 395 400
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
405 410 415
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
420 425 430
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
435 440 445
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
450 455 460
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
465 470 475 480
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
485 490 495
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
500 505 510
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
515 520 525
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
530 535 540
Gln Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
545 550 555 560
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
565 570 575
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
580 585 590
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
595 600 605
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
610 615 620
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
625 630 635 640
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
645 650 655
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
660 665 670
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
675 680 685
Leu Ser Leu Ser Pro Gly Lys
690 695
<210> 301
<211> 2088
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5 N297Q - OKT3m-H-N
<400> 301
caggttcagc tgcagcagag cggtgcagaa ctggcacgtc cgggtgcaag cgttaaaatg 60
agctgtaaag caagcggtta tacctttacc cgttatacca tgcattgggt taaacagcgt 120
ccgggtcagg gtctggaatg gattggttat atcaatccga gccgtggtta taccaactac 180
aaccagaaat tcaaagataa agcaaccctg accaccgata aaagcagcag caccgcctat 240
atgcagctga gcagcctgac ctcagaggat agcgcagttt attactgtgc acgctattat 300
gatgatcact atagcctgga ttattggggt cagggcacca ccctgaccgt tagcagcggt 360
ggtggtggta gtggtggcgg tggttcaggc ggtggcggta gccagattgt tctgacccag 420
agtccggcaa ttatgagcgc aagtccgggt gaaaaagtta ccatgacctg tagcgccagc 480
agcagcgtta gctatatgaa ttggtatcag cagaaaagcg gcaccagccc gaaacgttgg 540
atttatgata ccagcaaact ggcaagcggt gttccggcac attttcgtgg tagcggtagc 600
ggcacctcat atagcctgac cattagcggt atggaagcag aagatgcagc aacctattat 660
tgtcagcagt ggtcaagcaa tccgtttacc tttggtagtg gcaccaaact ggaaattaat 720
cgtggaggtg gtggatccca ggtgcagctg aaacagagcg gcccgggcct ggtgcagccg 780
agccagagcc tgagcattac ctgcaccgtg agcggcttta gcctgaccaa ctatggcgtg 840
cattgggtgc gccagagccc gggcaaaggc ctggaatggc tgggcgtgat ttggagcggc 900
ggcaacaccg attataacac cccgtttacc agccgcctga gcattaacaa agataacagc 960
aaaagccagg tgttttttaa aatgaacagc ctgcaaagcc aggataccgc gatttattat 1020
tgcgcgcgcg cgctgaccta ttatgattat gaatttgcgt attggggcca gggcaccctg 1080
gtgaccgtga gcgcggctag caccaagggc ccatcggtct tccccctggc accctcctcc 1140
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 1200
ccggtgacgg tgtcgtggaa ctcaggcgcc ctgaccagcg gcgtgcacac cttcccggct 1260
gtcctacagt cctcaggact ctactccctc agcagcgtgg tgaccgtgcc ctccagcagc 1320
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtggac 1380
aagaaagttg agcccaaatc ttgtgacaaa actcacacat gcccaccgtg cccagcacct 1440
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 1500
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 1560
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 1620
gaggagcagt accagagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 1680
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 1740
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 1800
ccatcccggg aggagatgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 1860
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 1920
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 1980
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 2040
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gtaaatga 2088
<210> 302
<211> 713
<212> PRT
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5 N297Q - OKT3m-H-С
<400> 302
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gln Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gln Val Gln Leu Gln Gln Ser Gly
465 470 475 480
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala
485 490 495
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
500 505 510
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
515 520 525
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
530 535 540
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
545 550 555 560
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
565 570 575
Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
580 585 590
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile
595 600 605
Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
610 615 620
Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
625 630 635 640
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
645 650 655
Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser
660 665 670
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala
675 680 685
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly
690 695 700
Ser Gly Thr Lys Leu Glu Ile Asn Arg
705 710
<210> 303
<211> 2142
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мультиспецифического антитела C225v5 N297Q - OKT3m-H-С
<400> 303
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtaccagagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaagga ggtggatctg gaggtggcgg ttcaggctct 1380
ggcggaggct caggtggtgg aggatcaggc ggaggtcagg ttcagctgca gcagagcggt 1440
gcagaactgg cacgtccggg tgcaagcgtt aaaatgagct gtaaagcaag cggttatacc 1500
tttacccgtt ataccatgca ttgggttaaa cagcgtccgg gtcagggtct ggaatggatt 1560
ggttatatca atccgagccg tggttatacc aactacaacc agaaattcaa agataaagca 1620
accctgacca ccgataaaag cagcagcacc gcctatatgc agctgagcag cctgacctca 1680
gaggatagcg cagtttatta ctgtgcacgc tattatgatg atcactatag cctggattat 1740
tggggtcagg gcaccaccct gaccgttagc agcggtggtg gtggtagtgg tggcggtggt 1800
tcaggcggtg gcggtagcca gattgttctg acccagagtc cggcaattat gagcgcaagt 1860
ccgggtgaaa aagttaccat gacctgtagc gccagcagca gcgttagcta tatgaattgg 1920
tatcagcaga aaagcggcac cagcccgaaa cgttggattt atgataccag caaactggca 1980
agcggtgttc cggcacattt tcgtggtagc ggtagcggca cctcatatag cctgaccatt 2040
agcggtatgg aagcagaaga tgcagcaacc tattattgtc agcagtggtc aagcaatccg 2100
tttacctttg gtagtggcac caaactggaa attaatcgtt ga 2142
<210> 304
<211> 261
<212> PRT
<213> Искусственная последовательность
<220>
<223> Легкая цепь активируемого антитела 3954-1204-C225v5
<400> 304
Gln Gly Gln Ser Gly Gln Cys Ile Ser Pro Arg Gly Cys Pro Asp Gly
1 5 10 15
Pro Tyr Val Met Tyr Gly Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
20 25 30
Ser Gly Leu Ser Gly Arg Ser Asp Asn His Gly Ser Ser Gly Thr Gln
35 40 45
Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly Glu
50 55 60
Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn Ile
65 70 75 80
His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile Lys
85 90 95
Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly Ser
100 105 110
Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser Glu
115 120 125
Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr Thr
130 135 140
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
145 150 155 160
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
165 170 175
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
180 185 190
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
195 200 205
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
210 215 220
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
225 230 235 240
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
245 250 255
Asn Arg Gly Glu Cys
260
<210> 305
<211> 1578
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь активируемого антитела 3954-1204-C225v5
<400> 305
caaggccagt ctggccagtg catctcacct cgtggttgtc cggacggccc atacgtcatg 60
tacggctcga gcggtggcag cggtggctct ggtggatccg gtctgagcgg ccgttccgat 120
aatcatggca gtagcggtac ccagatcttg ctgacccaga gcccggtgat tctgagcgtg 180
agcccgggcg aacgtgtgag ctttagctgc cgcgcgagcc agagcattgg caccaacatt 240
cattggtatc agcagcgcac caacggcagc ccgcgcctgc tgattaaata tgcgagcgaa 300
agcattagcg gcattccgag ccgctttagc ggcagcggca gcggcaccga ttttaccctg 360
agcattaaca gcgtggaaag cgaagatatt gcggattatt attgccagca gaacaacaac 420
tggccgacca cctttggcgc gggcaccaaa ctggaactga aacgtacggt ggctgcacca 480
tctgtcttca tcttcccgcc atctgatgag cagttgaaat ctggaactgc ctctgttgtg 540
tgcctgctga ataacttcta tcccagagag gccaaagtac agtggaaggt ggataacgcc 600
ctccaatcgg gtaactccca ggagagtgtc acagagcagg acagcaagga cagcacctac 660
agcctcagca gcaccctgac gctgagcaaa gcagactacg agaaacacaa agtctacgcc 720
tgcgaagtca cccatcaggg cctgagctcg cccgtcacaa agagcttcaa caggggagag 780
tgtggaggtg gatctggagg tggcggttca ggctctggcg gaggctcagg tggtggagga 840
tcaggcggag gtcaggttca gctgcagcag agcggtgcag aactggcacg tccgggtgca 900
agcgttaaaa tgagctgtaa agcaagcggt tataccttta cccgttatac catgcattgg 960
gttaaacagc gtccgggtca gggtctggaa tggattggtt atatcaatcc gagccgtggt 1020
tataccaact acaaccagaa attcaaagat aaagcaaccc tgaccaccga taaaagcagc 1080
agcaccgcct atatgcagct gagcagcctg acctcagagg atagcgcagt ttattactgt 1140
gcacgctatt atgatgatca ctatagcctg gattattggg gtcagggcac caccctgacc 1200
gttagcagcg gtggtggtgg tagtggtggc ggtggttcag gcggtggcgg tagccagatt 1260
gttctgaccc agagtccggc aattatgagc gcaagtccgg gtgaaaaagt taccatgacc 1320
tgtagcgcca gcagcagcgt tagctatatg aattggtatc agcagaaaag cggcaccagc 1380
ccgaaacgtt ggatttatga taccagcaaa ctggcaagcg gtgttccggc acattttcgt 1440
ggtagcggta gcggcacctc atatagcctg accattagcg gtatggaagc agaagatgca 1500
gcaacctatt attgtcagca gtggtcaagc aatccgttta cctttggtag tggcaccaaa 1560
ctggaaatta atcgttag 1578
<210> 306
<211> 241
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3m
<400> 306
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
130 135 140
Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser
145 150 155 160
Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser
165 170 175
Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
180 185 190
Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
195 200 205
Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
210 215 220
Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
225 230 235 240
Arg
<210> 307
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv антитела OKT3m
<400> 307
caggttcagc tgcagcagag cggtgcagaa ctggcacgtc cgggtgcaag cgttaaaatg 60
agctgtaaag caagcggtta tacctttacc cgttatacca tgcattgggt taaacagcgt 120
ccgggtcagg gtctggaatg gattggttat atcaatccga gccgtggtta taccaactac 180
aaccagaaat tcaaagataa agcaaccctg accaccgata aaagcagcag caccgcctat 240
atgcagctga gcagcctgac ctcagaggat agcgcagttt attactgtgc acgctattat 300
gatgatcact atagcctgga ttattggggt cagggcacca ccctgaccgt tagcagcggt 360
ggtggtggta gtggtggcgg tggttcaggc ggtggcggta gccagattgt tctgacccag 420
agtccggcaa ttatgagcgc aagtccgggt gaaaaagtta ccatgacctg tagcgccagc 480
agcagcgtta gctatatgaa ttggtatcag cagaaaagcg gcaccagccc gaaacgttgg 540
atttatgata ccagcaaact ggcaagcggt gttccggcac attttcgtgg tagcggtagc 600
ggcacctcat atagcctgac cattagcggt atggaagcag aagatgcagc aacctattat 660
tgtcagcagt ggtcaagcaa tccgtttacc tttggtagtg gcaccaaact ggaaattaat 720
cgt 723
<210> 308
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv LV антитела OKT3m
<400> 308
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Arg
100 105
<210> 309
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv LV антитела OKT3m
<400> 309
cagattgttc tgacccagag tccggcaatt atgagcgcaa gtccgggtga aaaagttacc 60
atgacctgta gcgccagcag cagcgttagc tatatgaatt ggtatcagca gaaaagcggc 120
accagcccga aacgttggat ttatgatacc agcaaactgg caagcggtgt tccggcacat 180
tttcgtggta gcggtagcgg cacctcatat agcctgacca ttagcggtat ggaagcagaa 240
gatgcagcaa cctattattg tcagcagtgg tcaagcaatc cgtttacctt tggtagtggc 300
accaaactgg aaattaatcg t 321
<210> 310
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> scFv Hv антитела OKT3m
<400> 310
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 311
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv Hv антитела OKT3m
<400> 311
caggttcagc tgcagcagag cggtgcagaa ctggcacgtc cgggtgcaag cgttaaaatg 60
agctgtaaag caagcggtta tacctttacc cgttatacca tgcattgggt taaacagcgt 120
ccgggtcagg gtctggaatg gattggttat atcaatccga gccgtggtta taccaactac 180
aaccagaaat tcaaagataa agcaaccctg accaccgata aaagcagcag caccgcctat 240
atgcagctga gcagcctgac ctcagaggat agcgcagttt attactgtgc acgctattat 300
gatgatcact atagcctgga ttattggggt cagggcacca ccctgaccgt tagcagc 357
<210> 312
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 312
Arg Pro Ser Pro Met Trp Ala Tyr
1 5
<210> 313
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь C225v5
<400> 313
cagatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 314
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> Мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N
<400> 314
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 315
<211> 1437
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Мультиспецифическое активируемое антитело 3954-1204-C225v5-OKT3m-H-N
<400> 315
gacatcttgc tgacccagag cccggtgatt ctgagcgtga gcccgggcga acgtgtgagc 60
tttagctgcc gcgcgagcca gagcattggc accaacattc attggtatca gcagcgcacc 120
aacggcagcc cgcgcctgct gattaaatat gcgagcgaaa gcattagcgg cattccgagc 180
cgctttagcg gcagcggcag cggcaccgat tttaccctga gcattaacag cgtggaaagc 240
gaagatattg cggattatta ttgccagcag aacaacaact ggccgaccac ctttggcgcg 300
ggcaccaaac tggaactgaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggaggtgg atctggaggt 660
ggcggttcag gctctggcgg aggctcaggt ggtggaggat caggcggagg tcaggttcag 720
ctgcagcaga gcggtgcaga actggcacgt ccgggtgcaa gcgttaaaat gagctgtaaa 780
gcaagcggtt atacctttac ccgttatacc atgcattggg ttaaacagcg tccgggtcag 840
ggtctggaat ggattggtta tatcaatccg agccgtggtt ataccaacta caaccagaaa 900
ttcaaagata aagcaaccct gaccaccgat aaaagcagca gcaccgccta tatgcagctg 960
agcagcctga cctcagagga tagcgcagtt tattactgtg cacgctatta tgatgatcac 1020
tatagcctgg attattgggg tcagggcacc accctgaccg ttagcagcgg tggtggtggt 1080
agtggtggcg gtggttcagg cggtggcggt agccagattg ttctgaccca gagtccggca 1140
attatgagcg caagtccggg tgaaaaagtt accatgacct gtagcgccag cagcagcgtt 1200
agctatatga attggtatca gcagaaaagc ggcaccagcc cgaaacgttg gatttatgat 1260
accagcaaac tggcaagcgg tgttccggca cattttcgtg gtagcggtag cggcacctca 1320
tatagcctga ccattagcgg tatggaagca gaagatgcag caacctatta ttgtcagcag 1380
tggtcaagca atccgtttac ctttggtagt ggcaccaaac tggaaattaa tcgttag 1437
<210> 316
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 316
Ile Ser Ser Gly Leu Leu Ser Ser
1 5
<210> 317
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 317
Gln Asn Gln Ala Leu Arg Met Ala
1 5
<210> 318
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 318
Ala Gln Asn Leu Leu Gly Met Val
1 5
<210> 319
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 319
Ser Thr Phe Pro Phe Gly Met Phe
1 5
<210> 320
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 320
Pro Val Gly Tyr Thr Ser Ser Leu
1 5
<210> 321
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 321
Asp Trp Leu Tyr Trp Pro Gly Ile
1 5
<210> 322
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 322
Met Ile Ala Pro Val Ala Tyr Arg
1 5
<210> 323
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 323
Trp Ala Thr Pro Arg Pro Met Arg
1 5
<210> 324
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 324
Phe Arg Leu Leu Asp Trp Gln Trp
1 5
<210> 325
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 325
Leu Lys Ala Ala Pro Arg Trp Ala
1 5
<210> 326
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 326
Gly Pro Ser His Leu Val Leu Thr
1 5
<210> 327
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 327
Leu Pro Gly Gly Leu Ser Pro Trp
1 5
<210> 328
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 328
Met Gly Leu Phe Ser Glu Ala Gly
1 5
<210> 329
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 329
Ser Pro Leu Pro Leu Arg Val Pro
1 5
<210> 330
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 330
Arg Met His Leu Arg Ser Leu Gly
1 5
<210> 331
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 331
Leu Ala Ala Pro Leu Gly Leu Leu
1 5
<210> 332
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 332
Ala Val Gly Leu Leu Ala Pro Pro
1 5
<210> 333
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 333
Leu Leu Ala Pro Ser His Arg Ala
1 5
<210> 334
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 334
Pro Ala Gly Leu Trp Leu Asp Pro
1 5
<210> 335
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> отщепляемая группа
<400> 335
Ile Ser Ser Gly Leu Ser Ser
1 5
<210> 336
<211> 449
<212> PRT
<213> Искусственная последовательность
<220>
<223> antibody C225v5 N297Q heavy chain
<400> 336
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Gln Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gln Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 337
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь антитела C225v5 N297Q
<400> 337
caggtgcagc tgaaacagag cggcccgggc ctggtgcagc cgagccagag cctgagcatt 60
acctgcaccg tgagcggctt tagcctgacc aactatggcg tgcattgggt gcgccagagc 120
ccgggcaaag gcctggaatg gctgggcgtg atttggagcg gcggcaacac cgattataac 180
accccgttta ccagccgcct gagcattaac aaagataaca gcaaaagcca ggtgtttttt 240
aaaatgaaca gcctgcaaag ccaggatacc gcgatttatt attgcgcgcg cgcgctgacc 300
tattatgatt atgaatttgc gtattggggc cagggcaccc tggtgaccgt gagcgcggct 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtaccagagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaatga 1350
<210> 338
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 338
Gln Gly Gln Ser Gly Gln Cys Asn Ile Trp Leu Val Gly Gly Asp Cys
1 5 10 15
Arg Gly Trp Gln Gly
20
<210> 339
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> маскирующая группа
<400> 339
Cys Ile Ser Pro Arg Gly Cys
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2815683C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2837770C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| ПОЛУЧЕНИЕ МУЛЬТИСПЕЦИФИЧЕСКИХ СМЕСЕЙ АНТИТЕЛ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2815826C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| МУЛЬТИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО СО СПЕЦИФИЧНОСТЬЮ СВЯЗЫВАНИЯ IL-13 И IL-17 ЧЕЛОВЕКА | 2020 |
|
RU2827946C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| АНТИТЕЛА К ХЕЛАТНЫМ РАДИОНУКЛИДАМ | 2019 |
|
RU2833233C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |


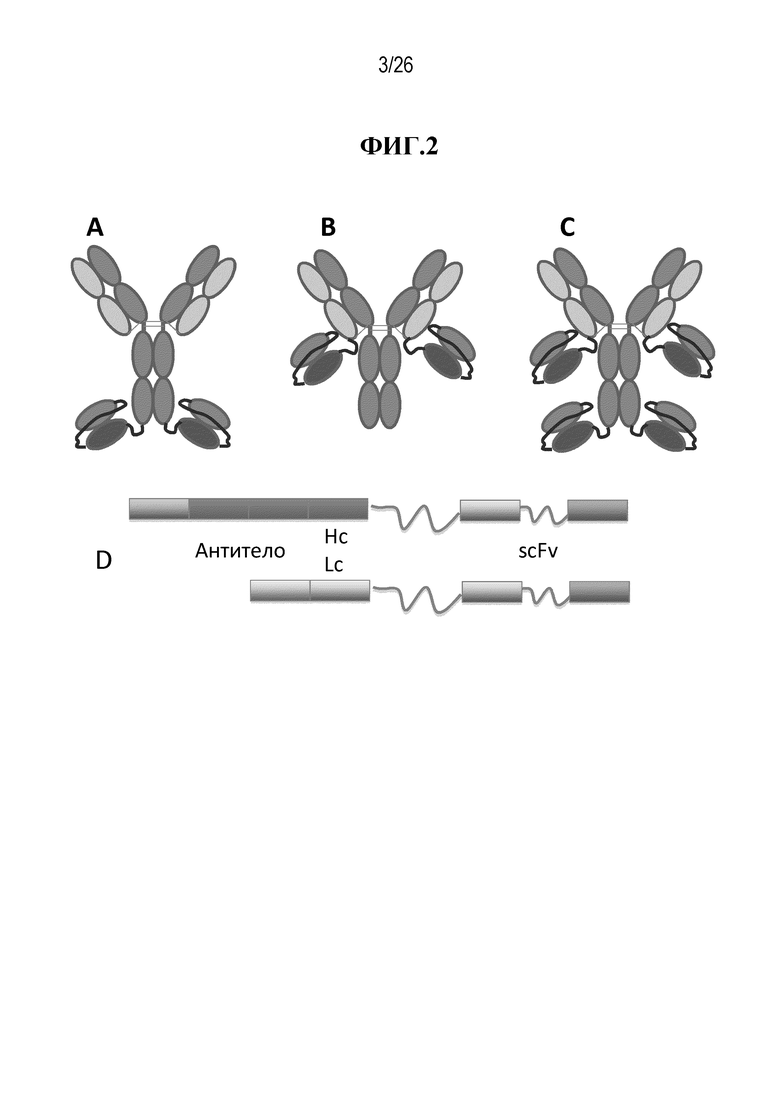
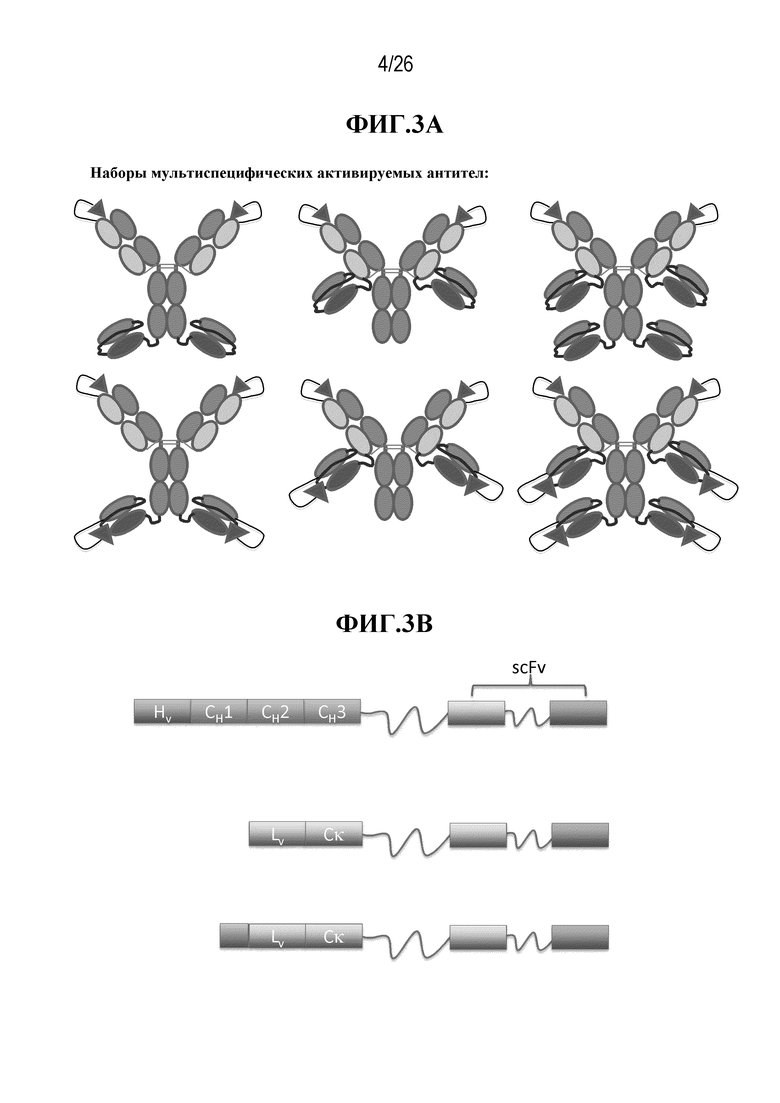





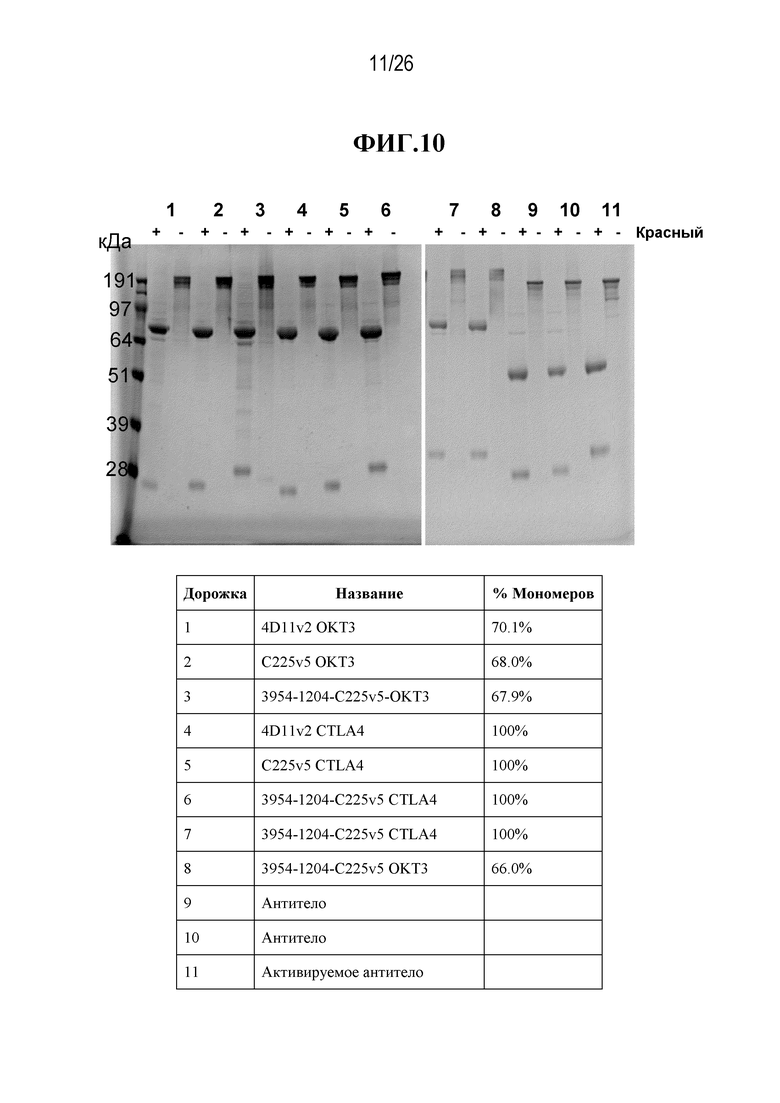
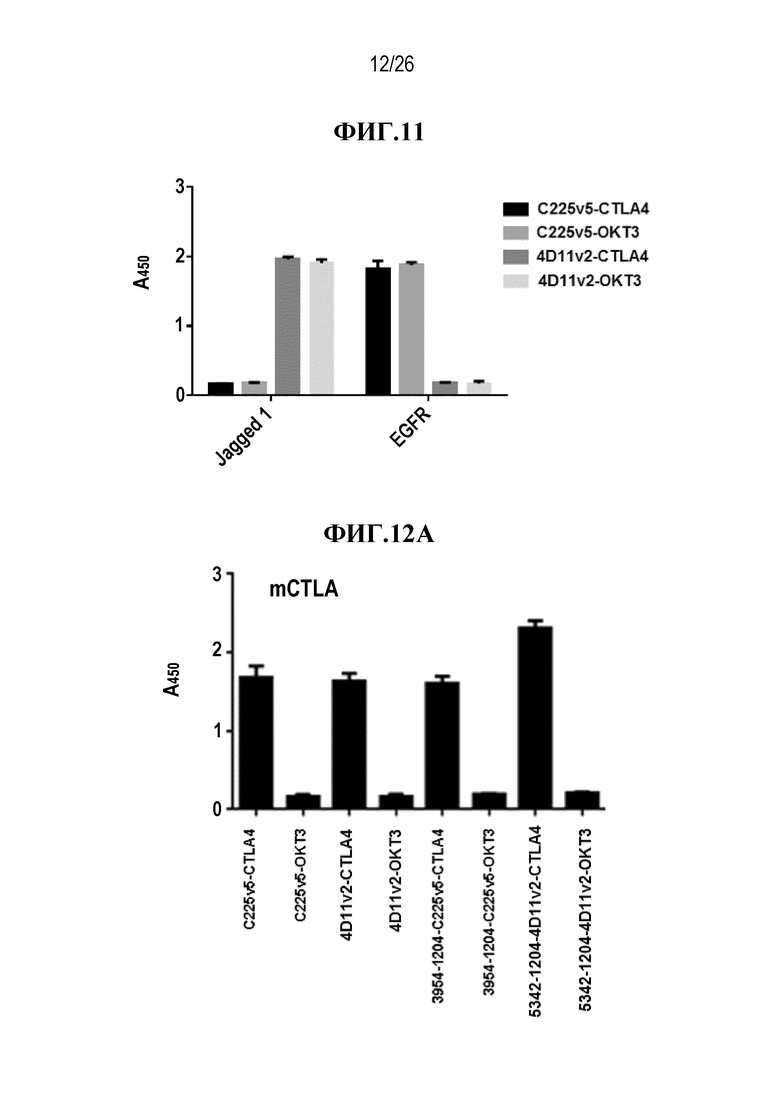
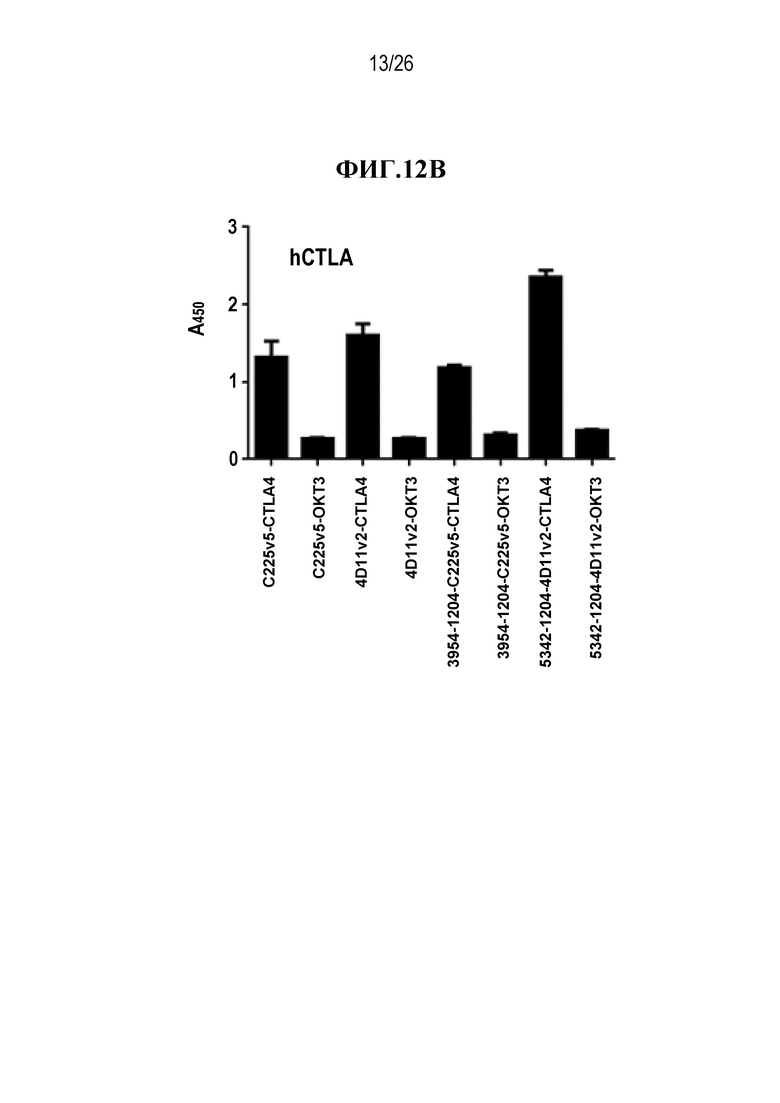
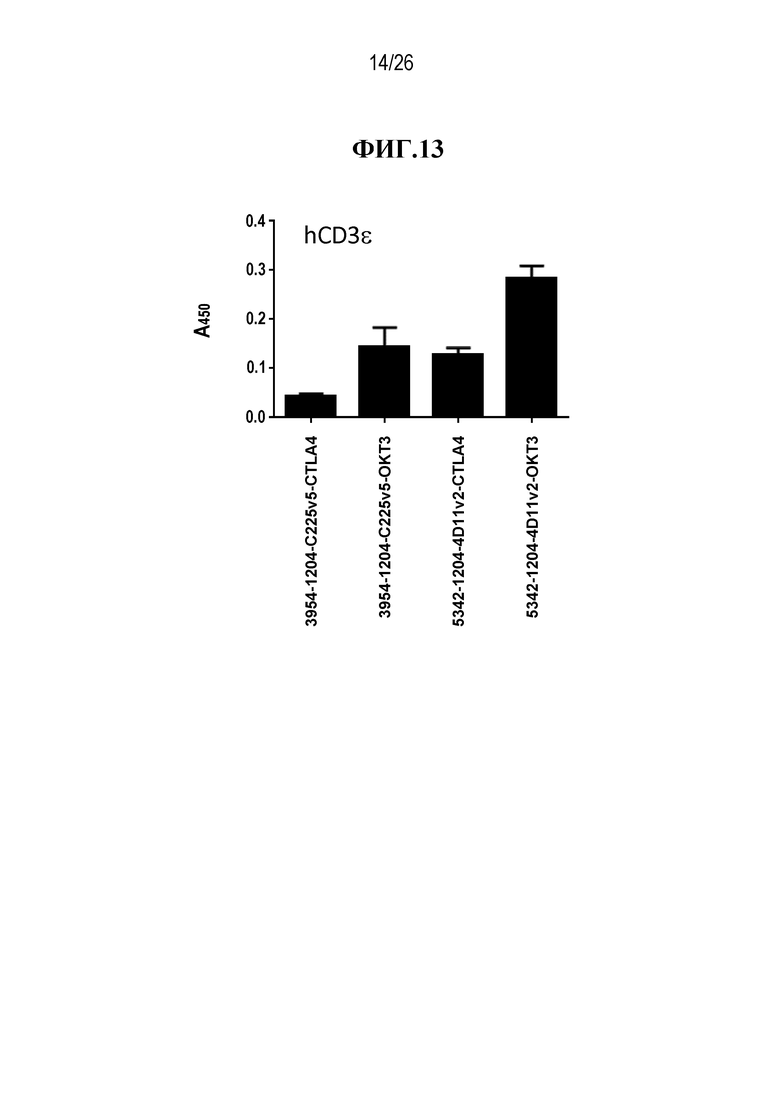
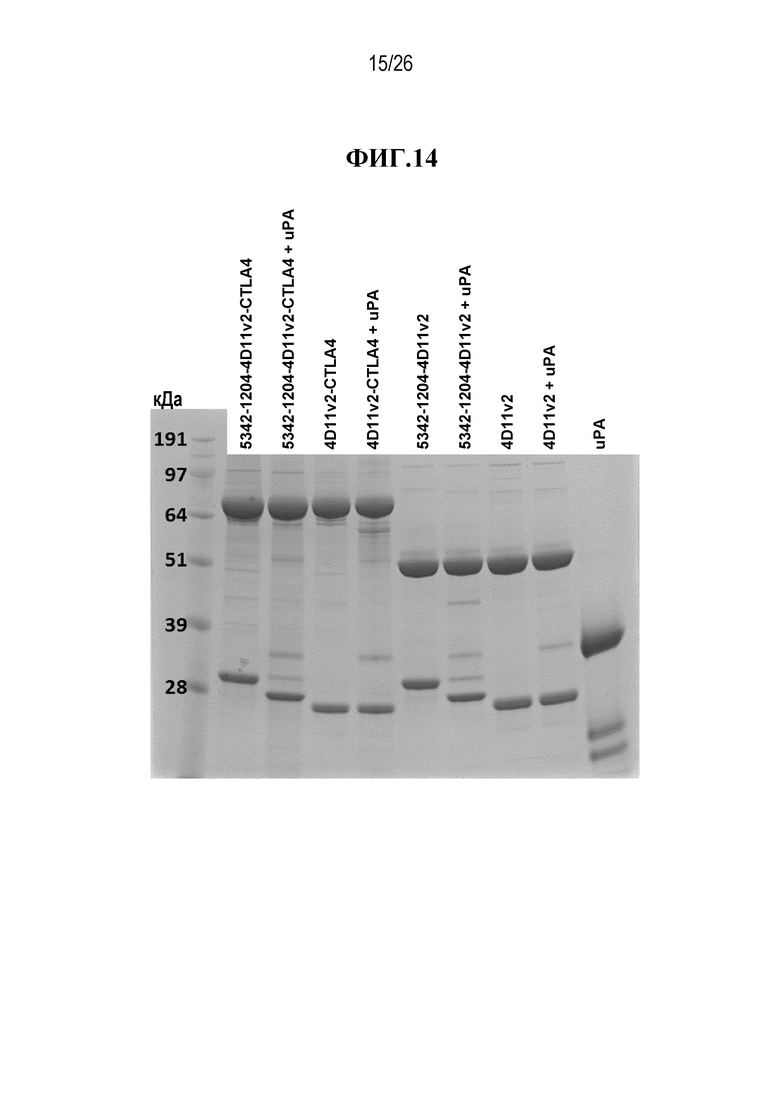
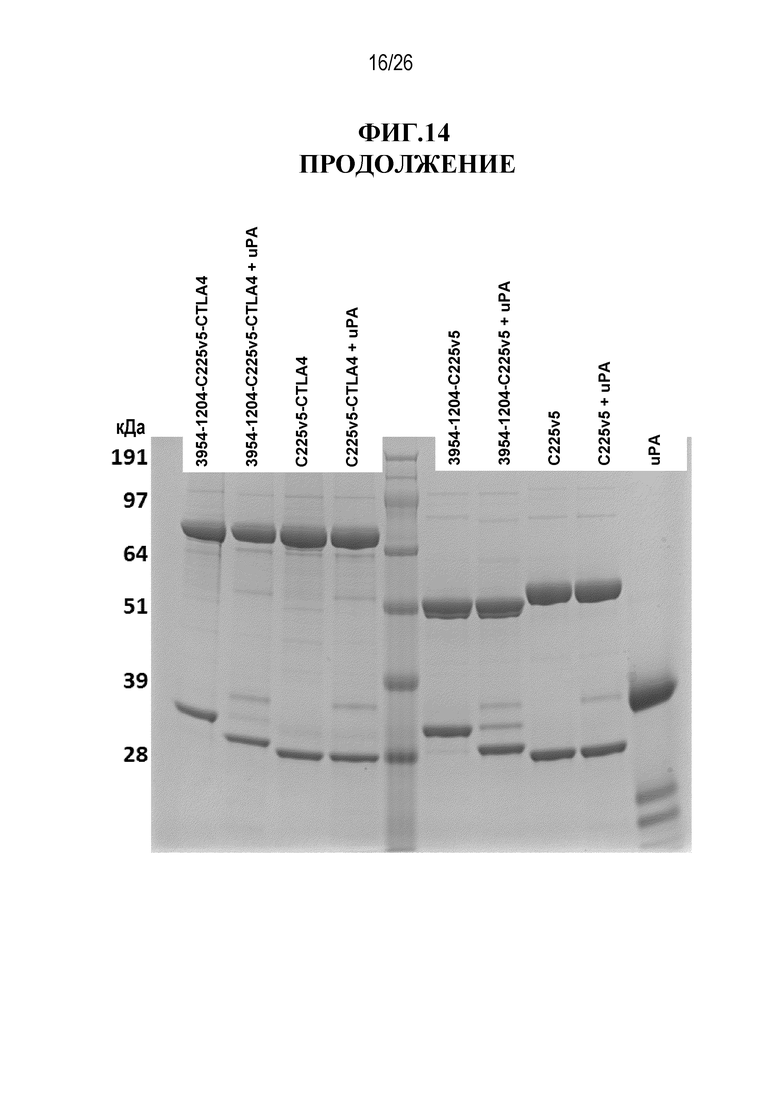
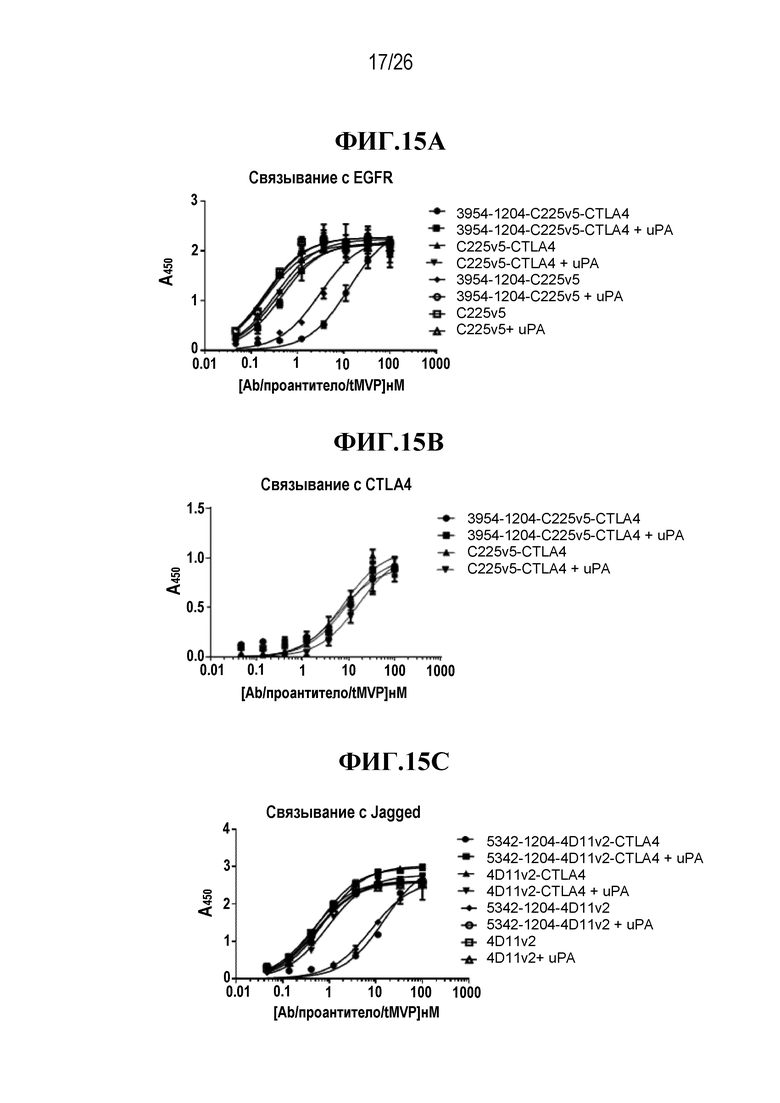
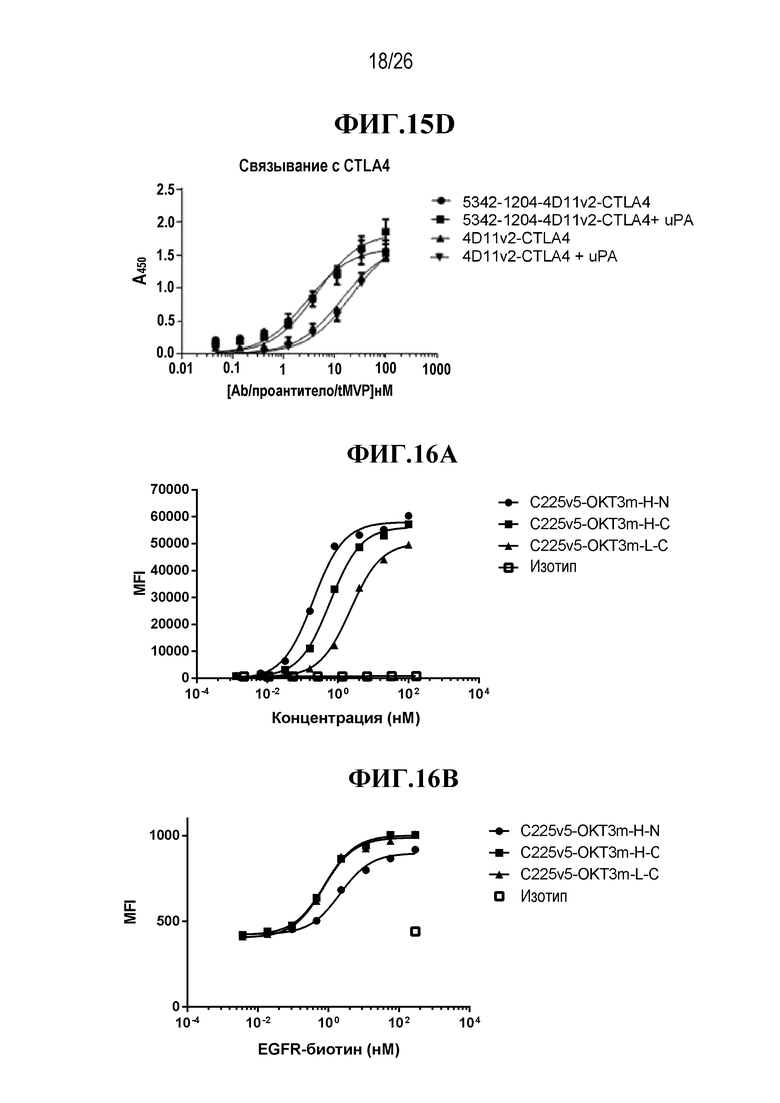
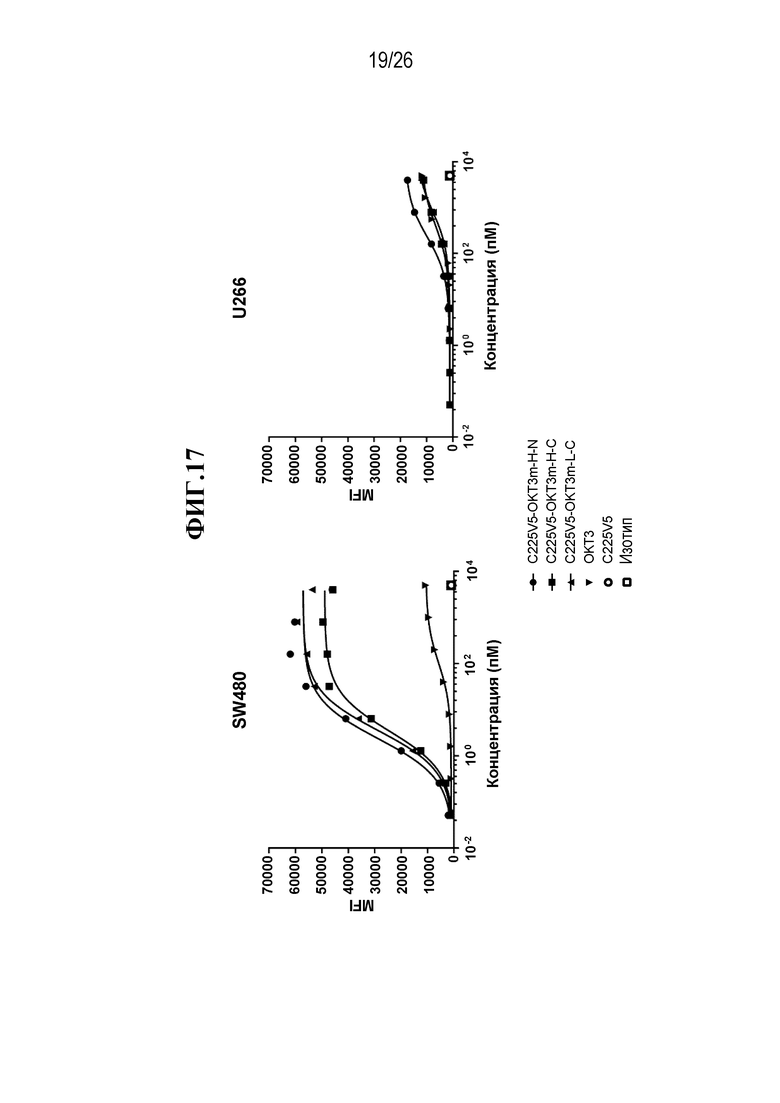
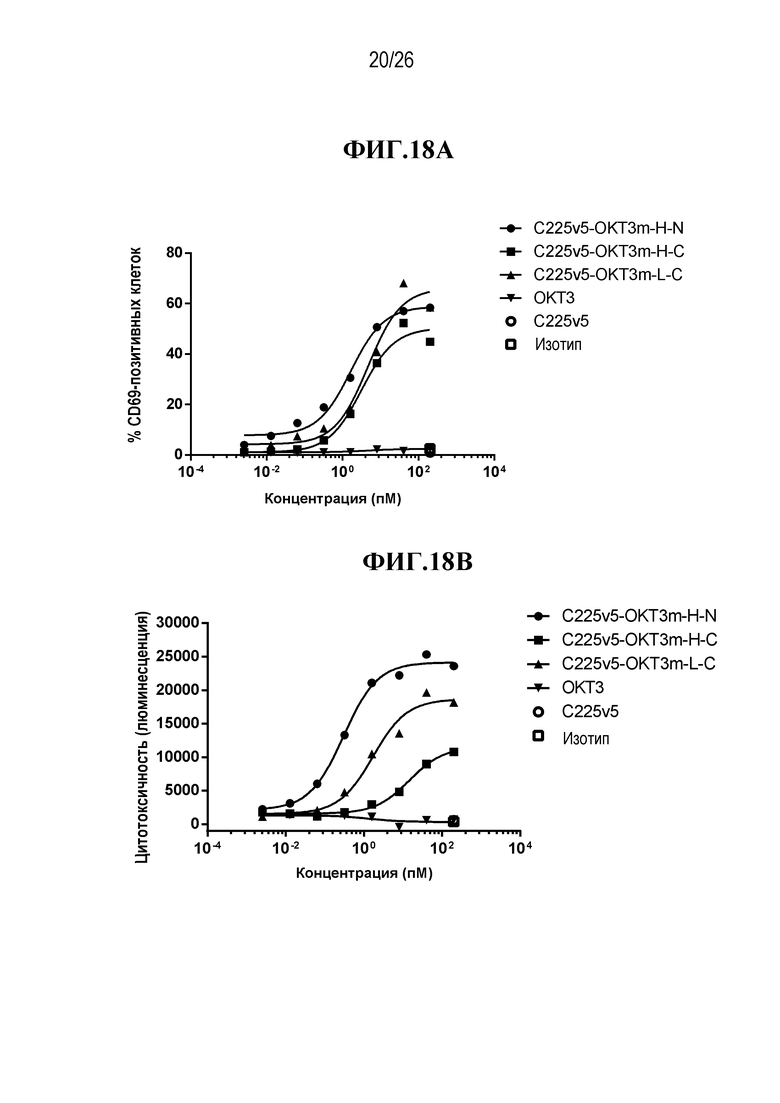
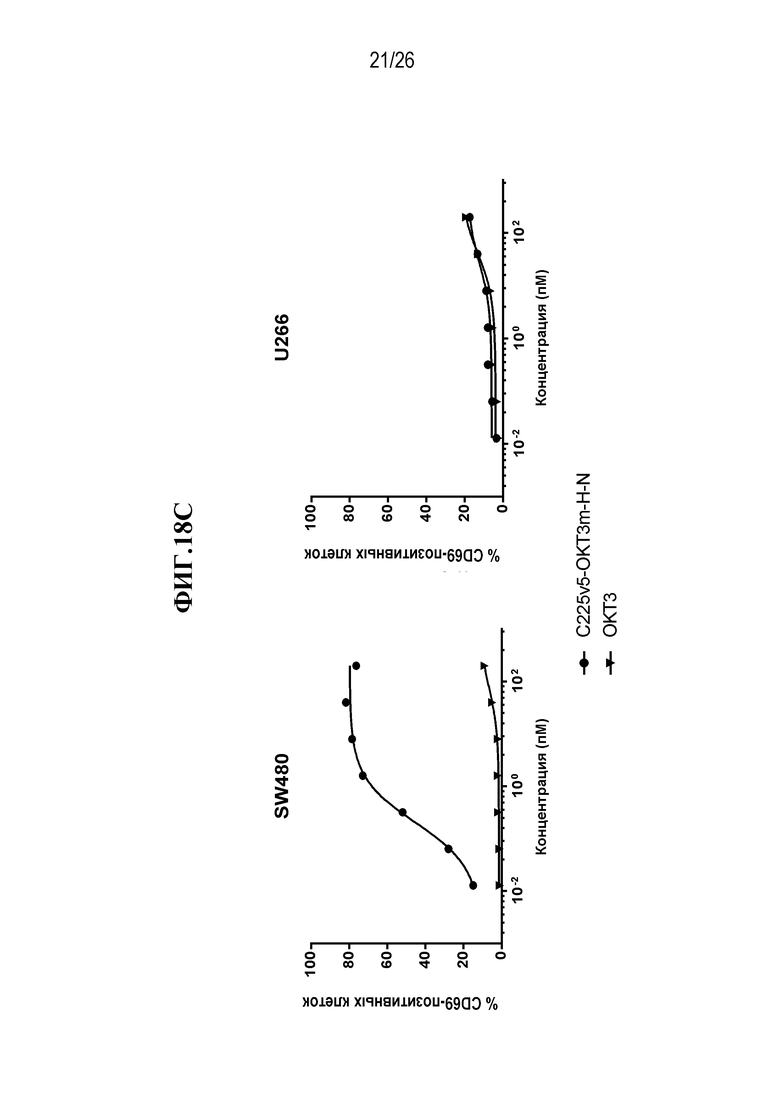
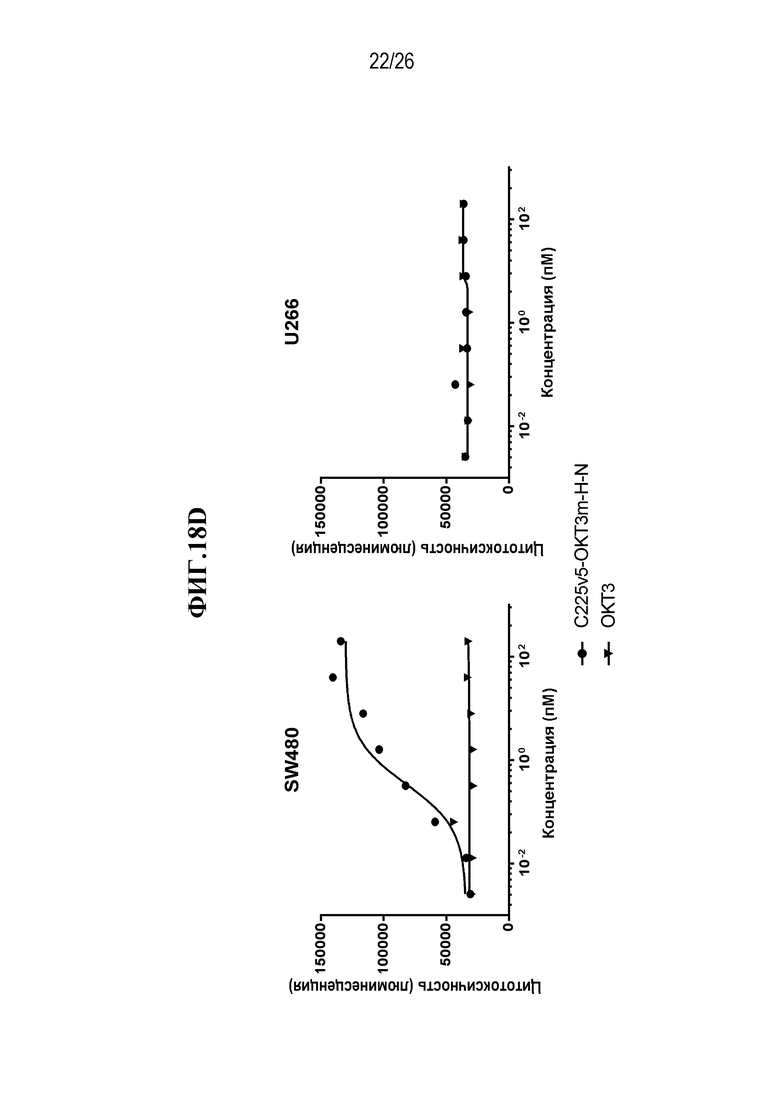
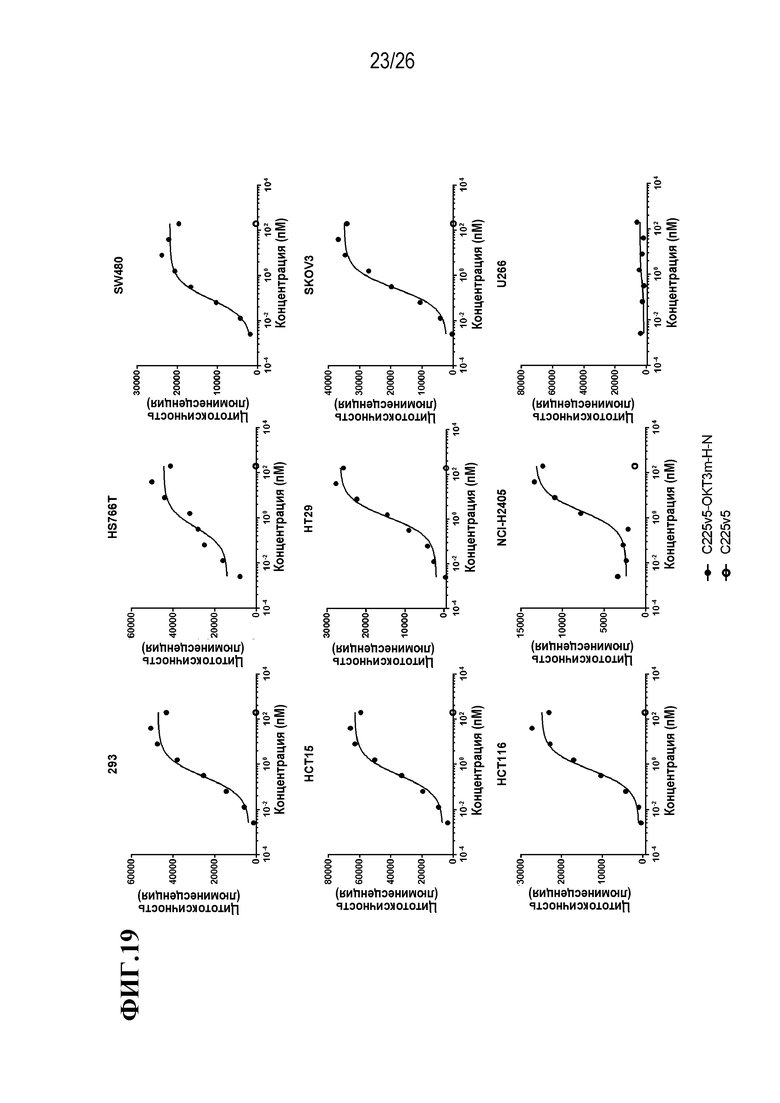
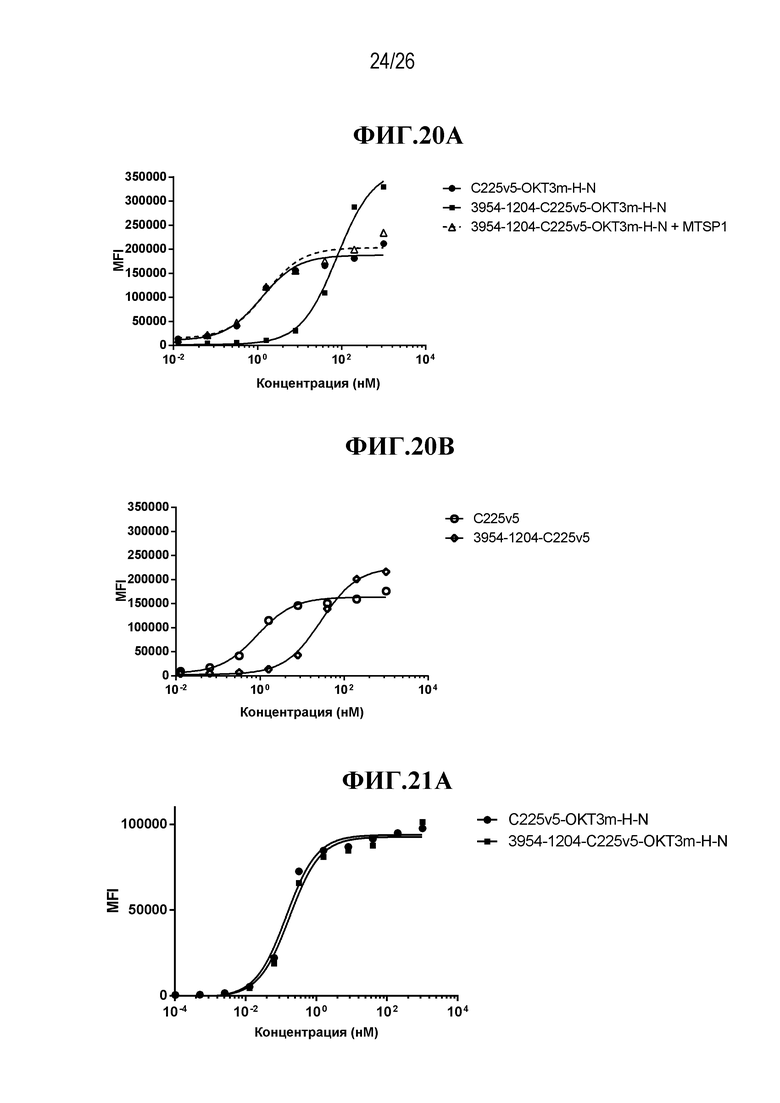
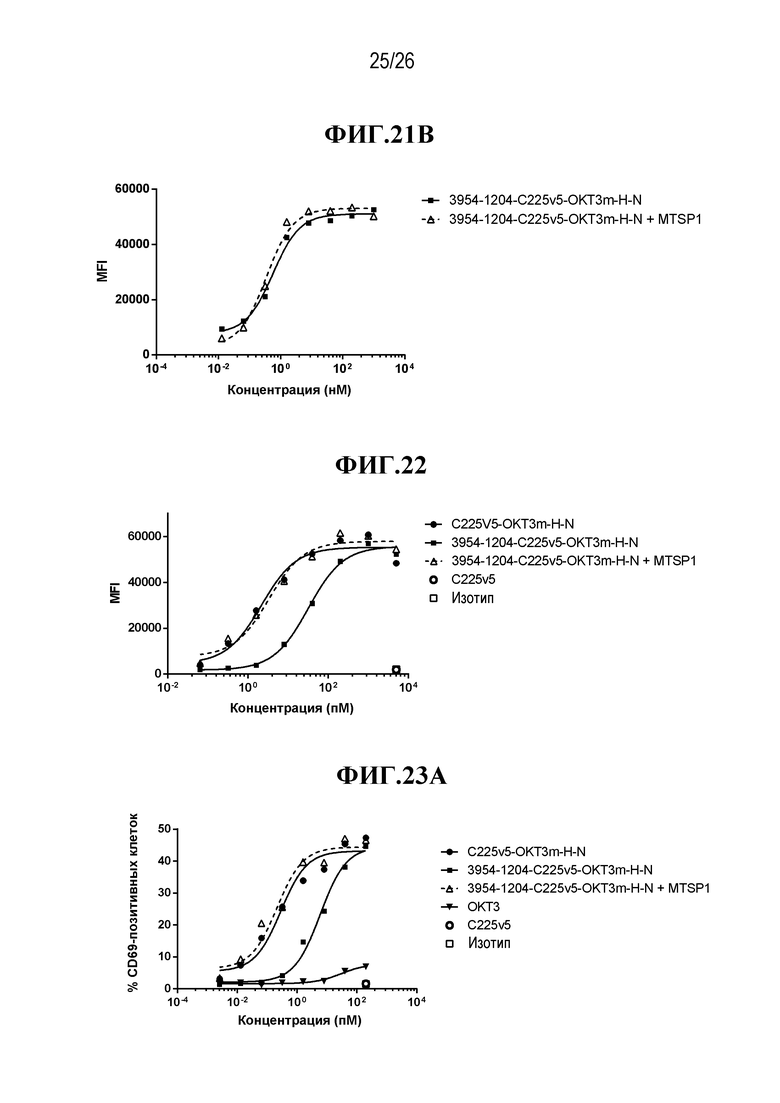
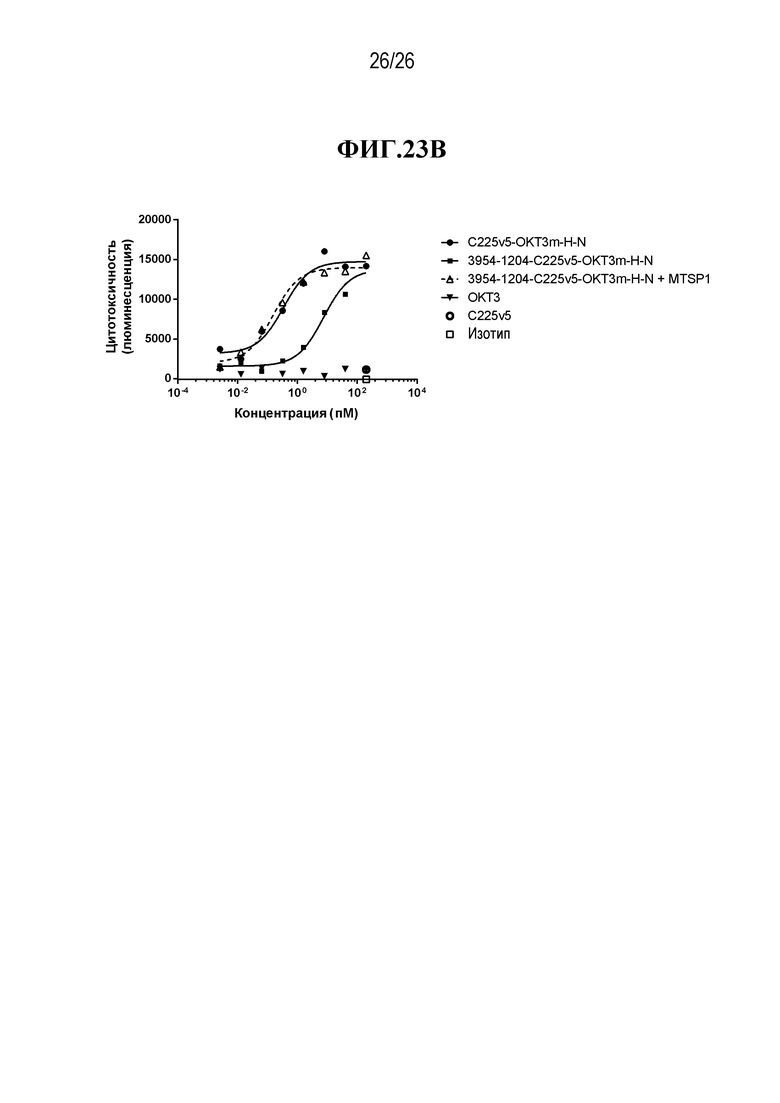
Настоящая группа изобретений относится к иммунологии. Предложена выделенная молекула нуклеиновой кислоты, кодирующая мультиспецифичное активируемое антитело, где антитело связано с расщепляемыми пептидными группами, которые являются субстратом для протеаз, и маскирующими группами. Также представлен вектор экспрессии, способы получения мультиспецифического антитела и применение антитела для облегчения симптома клинического заболевания больного. Данная группа изобретений позволяет получать и применять мультиспецифичные антитела, обладающие повышенной селективностью. 5 н. и 14 з.п. ф-лы, 11 табл., 16 пр., 23 ил.
1. Выделенная молекула нуклеиновой кислоты, кодирующая мультиспецифичное активируемое антитело, содержащее следующую структуру:
(a) антитело (АВ1), нацеленное на злокачественную опухоль, которое специфически связывается с мишенью или эпитопом злокачественной опухоли,
где АВ1 содержит полноразмерное антитело IgG, содержащее две тяжелые цепи или две легкие цепи; и
где АВ1 связано с:
i. двумя первыми маскирующими пептидными группами (MM1); и
ii. двумя расщепляемыми пептидными группами (CM1), каждая CM1 является субстратом для первой протеазы;
где каждая ММ1 связана в направлении от N- к C-концу с CM1, образуя два пептида MM1-CM1, и
где карбоксильный конец каждого пептида MM1-CM1 связан с амино-концом каждой легкой цепи AB1; и
(b) два scFv (AB2), связывающиеся с иммунными эффекторными клетками, каждый AB2 специфично связывается с мишенью или эпитопом иммунной эффекторной клетки,
где каждый AB2 содержит домен вариабельной области легкой цепи антитела и домен тяжелой цепи вариабельной области антитела; и
где каждый AB2 связан:
i. со второй маскирующей пептидной группой (MM2); и
ii. со второй расщепляемой пептидной группой (CM2), которая является субстратом для второй протеазы;
где MM2 связан в направлении от N- к C-концу с CM2, образуя пептид MM2-CM2;
где карбоксильный конец пептида ММ2-СМ2 связан с амино-концом AB2;
где карбоксильный конец каждого AB2 связан с амино-концом каждой тяжелой цепи AB1.
2. Выделенная молекула нуклеиновой кислоты по п.1, где MM1 кодируемого мультиспецифичного активируемого антитела при связывании с AB1 имеет константу равновесной диссоциации, превышающую константу равновесной диссоциации при связывании AB1 с его мишенью или эпитопом,
где СМ1 расположена в активируемом антителе таким образом, что в нерасщепленном состоянии MM1 препятствует специфичному связыванию AB1 с мишенью или эпитопом злокачественной опухоли,
где MM1 не влияет на связывание с мишенью злокачественной опухоли или эпитопом или не конкурирует за связывание с мишенью злокачественной опухоли или эпитопом с AB1, если мультиспецифичное активируемое антитело находится в расщепленном состоянии,
где полипептидная последовательность ММ1 отличается от полипептидной последовательности мишени злокачественной опухоли,
где полипептидная последовательность ММ1 не более чем на 50% идентична любому природному партнеру связывания АВ1, и/или
где ММ1 представляет собой полипептид, длина которого составляет не более 40 аминокислот.
3. Выделенная молекула нуклеиновой кислоты по п.1 или 2, где MM2 кодируемого мультиспецифичного активируемого антитела при связывании с AB2 имеет константу равновесной диссоциации, превышающую константу равновесной диссоциации при связывании AB2 с его мишенью или эпитопом,
где СМ2 расположена в активируемом антителе таким образом, что в нерасщепленном состоянии MM2 препятствует специфичному связыванию AB2 с мишенью или эпитопом иммунной эффекторной клетки,
где MM2 не влияет на связывание мишени иммунной эффекторной клетки или не конкурирует за связывание мишени иммунной эффекторной клетки с АВ2, если мультиспецифичное активируемое антитело находится в расщепленном состоянии,
где полипептидная последовательность ММ2 отличается от полипептидной последовательности мишени иммунной эффекторной клетки,
где полипептидная последовательность ММ2 не более чем на 50% идентична любому природному партнеру связывания АВ2, и/или
где ММ2 представляет собой полипептид, длина которого составляет не более 40 аминокислот.
4. Выделенная молекула нуклеиновой кислоты по любому из пп.1-3, где первая протеаза, которая расщепляет СМ1 кодируемого мультиспецифичного активируемого антитела, расположена рядом с мишенью или эпитопом злокачественной опухоли в ткани, и
где первая протеаза расщепляет СМ1 в кодируемом мультиспецифичном активируемом антителе, если мультиспецифичное активированное антитело подвергается воздействию первой протеазы.
5. Выделенная молекула нуклеиновой кислоты по любому из пп.1-4, где вторая протеаза, которая расщепляет CM2 кодируемого мультиспецифичного активируемого антитела, расположена рядом с мишенью или эпитопом иммунной эффекторной клетки в ткани, и
где вторая протеаза расщепляет СМ2 в кодируемом мультиспецифичном активируемом антител, если мультиспецифичное активированное антитело подвергается воздействию второй протеазы.
6. Выделенная молекула нуклеиновой кислоты по любому из пп.1-5, где по меньшей мере часть кодируемого мультиспецифичного активируемого антитела в нерасщепленном состоянии имеет следующее расположение структурных элементов в направлении от N-конца к C-концу: MM1-CM1-AB1.
7. Выделенная молекула нуклеиновой кислоты по любому из пп.1-6, где кодируемое мультиспецифичное активируемое антитело содержит:
связывающий пептид между ММ1 и СМ1; или
связывающий пептид между СМ1 и АВ1; или
первый связывающий пептид (LP1) и второй связывающий пептид (LP2), где мультиспецифическое активируемое антитело в нерасщепленном состоянии имеет следующее расположение структурных элементов в направлении от N-конца к C-концу: MM1-LP1-CM1-LP2-AB1,
необязательно, где два связывающих пептида не должны быть идентичны друг другу,
необязательно, где каждый LP1 и LP2 представляет собой пептид с длиной примерно от 1 до 20 аминокислот.
8. Выделенная молекула нуклеиновой кислоты по любому из пп.1-7, где по меньше мере часть кодируемого мультиспецифичного активируемого антитела в нерасщепленном состоянии имеет следующее расположение структурных элементов в направлении от N-конца к C-концу: MM2-CM2-AB2.
9. Выделенная молекула нуклеиновой кислоты по любому из пп.1-8, где кодируемое мультиспецифичное активируемое антитело содержит:
связывающий пептид между ММ2 и СМ2; или
связывающий пептид между СМ2 и АВ2; или
первый связывающий пептид (LP1) и второй связывающий пептид (LP2), где мультиспецифическое активируемое антитело в нерасщепленном состоянии имеет следующее расположение структурных элементов в направлении от N-конца к C-концу: MM2-LP1-CM2-LP2-AB2,
необязательно, где два связывающих пептида не должны быть идентичны друг другу,
необязательно, где каждый LP1 и LP2 представляет собой пептид с длиной примерно от 1 до 20 аминокислот.
10. Выделенная молекула нуклеиновой кислоты по любому из пп.1-9, где СМ1 кодируемого мультиспецифичного активируемого антитела представляет собой субстрат для первой протеазы, выбранной из протеаз ADAMS, ADAMTS, ADAM8, ADAM9, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAMDEC1, ADAMTS1, ADAMTS4, ADAMTS5, аспартат-протеаз, BACE, ренина, аспарагиновая кислота – катепсинов, катепсина D, катепсина E, каспаз, каспазы 1, каспазы 2, каспазы 3, каспазы 4, каспазы 5, каспазы 6, каспазы 7, каспазы 8, каспазы 9, каспазы 10, каспазы 14, цистеин-катепсинов, катепсина B, катепсина C, катепсина K, катепсина L, катепсина S, катепсина V/L2, катепсина X/Z/P, цистеиновых протеиназ, крузипаина, легумаина, отубаина-2, KLKs, KLK4, KLK5, KLK6, KLK7, KLK8, KLK10, KLK11, KLK13, KLK14, металлопротеиназ, меприна, неприлизина, PSMA, BMP-1, MMP, MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP10, MMP11, MMP12, MMP13, MMP14, MMP15, MMP16, MMP17, MMP19, MMP20, MMP23, MMP24, MMP26, MMP27, сериновых протеаз, активированного белка C, катепсина A, катепсина G, химазы, протеаз фактора свертывания крови, FVIIa, FIXa, FXa, FXIa, FXIIa, эластазы, гранзима В, гуанидинобензоатазы, HtrA1, эластазы человеческих нейтрофилов, лактоферрина, марапсина, NS3/4A, PACE4, плазмина, PSA, tPA, тромбина, триптазы, uPA, трансмембранных сериновых протеаз типа II (TTSP), DESC1, DPP-4, FAP, гепсина, матриптазы-2, MT-SP1/матриптазы, TMPRSS2, TMPRSS3, TMPRSS4, и/или
где СМ1 представляет собой пептид длиной до 15 аминокислот.
11. Выделенная молекула нуклеиновой кислоты по любому из пп.1-10, где СМ2 кодируемого мультиспецифичного активируемого антитела представляет собой субстрат для второй протеазы, выбранной из протеаз ADAMS, ADAMTS, ADAM8, ADAM9, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAMDEC1, ADAMTS1, ADAMTS4, ADAMTS5, аспартат-протеаз, BACE, ренина, аспарагиновая кислота–катепсинов, катепсина D, катепсина E, каспаз, каспазы 1, каспазы 2, каспазы 3, каспазы 4, каспазы 5, каспазы 6, каспазы 7, каспазы 8, каспазы 9, каспазы 10, каспазы 14, цистеин-катепсинов, катепсина B, катепсина C, катепсина K, катепсина L, катепсина S, катепсина V/L2, катепсина X/Z/P, цистеиновых протеиназ, крузипаина, легумаина, отубаина-2, KLKs, KLK4, KLK5, KLK6, KLK7, KLK8, KLK10, KLK11, KLK13, KLK14, металлопротеиназ, меприна, неприлизина, PSMA, BMP-1, MMP, MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP10, MMP11, MMP12, MMP13, MMP14, MMP15, MMP16, MMP17, MMP19, MMP20, MMP23, MMP24, MMP26, MMP27, сериновых протеаз, активированного белка C, катепсина A, катепсина G, химазы, Протеаз фактора свертывания крови, FVIIa, FIXa, FXa, FXIa, FXIIa, эластазы, гранзима В, гуанидинобензоатазы, HtrA1, эластазы человеческих нейтрофилов, лактоферрина, марапсина, NS3/4A, PACE4, плазмина, PSA, tPA, тромбина, триптазы, uPA, трансмембранных сериновых протеаз типа II (TTSP), DESC1, DPP-4, FAP, гепсина, матриптазы-2, MT-SP1/матриптазы, TMPRSS2, TMPRSS3, TMPRSS4, и/или
где СМ2 представляет собой пептид длиной не более 15 аминокислот.
12. Выделенная молекула нуклеиновой кислоты по любому из пп.1-11:
где первая протеаза и вторая протеаза расположены вместе со связанной со злокачественной опухолью мишенью или эпитопом и со связывающейся с иммунной эффекторной клеткой мишенью или эпитопом в ткани; и/или
где первая протеаза и вторая протеаза являются одинаковыми протеазами,
необязательно, где СМ1 и СМ2 представляют собой разные субстраты одной протеазы,
необязательно, где протеаза выбрана из группы, состоящей из протеаз ADAMS, ADAMTS, ADAM8, ADAM9, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAMDEC1, ADAMTS1, ADAMTS4, ADAMTS5, аспартат-протеаз, BACE, ренина, аспарагиновая кислота–катепсинов, катепсина D, катепсина E, каспаз, каспазы 1, каспазы 2, каспазы 3, каспазы 4, каспазы 5, каспазы 6, каспазы 7, каспазы 8, каспазы 9, каспазы 10, каспазы 14, цистеин-катепсинов, катепсина B, катепсина C, катепсина K, катепсина L, катепсина S, катепсина V/L2, катепсина X/Z/P, цистеиновых протеиназ, крузипаина, легумаина, отубаина-2, KLKs, KLK4, KLK5, KLK6, KLK7, KLK8, KLK10, KLK11, KLK13, KLK14, металлопротеиназ, меприна, неприлизина, PSMA, BMP-1, MMP, MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP10, MMP11, MMP12, MMP13, MMP14, MMP15, MMP16, MMP17, MMP19, MMP20, MMP23, MMP24, MMP26, MMP27, сериновых протеаз, активированного белка C, катепсина A, катепсина G, химазы, протеаз фактора свертывания крови, FVIIa, FIXa, FXa, FXIa, FXIIa, эластазы, гранзима В, гуанидинобензоатазы, HtrA1, эластазы человеческих нейтрофилов, лактоферрина, марапсина, NS3/4A, PACE4, плазмина, PSA, tPA, тромбина, триптазы, uPA, трансмембранных сериновых протеаз типа II (TTSP), DESC1, DPP-4, FAP, гепсина, матриптаза-2, MT-SP1/матриптазы, TMPRSS2, TMPRSS3, TMPRSS4, или
где первая протеаза и вторая протеаза являются разными протеазами, выбранными из протеаз ADAMS, ADAMTS, ADAM8, ADAM9, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAMDEC1, ADAMTS1, ADAMTS4, ADAMTS5, аспартат-протеаз, BACE, ренина, аспарагиновая кислота–катепсинов, катепсина D, катепсина E, каспаз, каспазы 1, каспазы 2, каспазы 3, каспазы 4, каспазы 5, каспазы 6, каспазы 7, каспазы 8, каспазы 9, каспазы 10, каспазы 14, цистеин-катепсинов, катепсина B, катепсина C, катепсина K, катепсина L, катепсина S, катепсина V/L2, катепсина X/Z/P, цистеиновых протеиназ, крузипаина, легумаина, отубаина-2, KLKs, KLK4, KLK5, KLK6, KLK7, KLK8, KLK10, KLK11, KLK13, KLK14, металлопротеиназ, меприна, неприлизина, PSMA, BMP-1, MMP, MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP10, MMP11, MMP12, MMP13, MMP14, MMP15, MMP16, MMP17, MMP19, MMP20, MMP23, MMP24, MMP26, MMP27, сериновых протеаз, активированного белка C, катепсина A, катепсина G, химазы, протеаз фактора свертывания крови, FVIIa, FIXa, FXa, FXIa, FXIIa, эластазы, гранзима В, гуанидинобензоатазы, HtrA1, эластазы человеческих нейтрофилов, лактоферрина, марапсина, NS3/4A, PACE4, плазмина, PSA, tPA, тромбина, триптазы, uPA, трансмембранных сериновых протеаз типа II (TTSP), DESC1, DPP-4, FAP, гепсина, матриптазы-2, MT-SP1/матриптазы, TMPRSS2, TMPRSS3, TMPRSS4.
13. Выделенная молекула нуклеиновой кислоты по любому из пп.1-12, где АВ2 кодируемого мультиспецифичного активируемого антитела представляет собой:
scFv, связывающийся с иммунной эффекторной клеткой;
scFv, связывающийся с лейкоцитом;
scFv, связывающийся с Т-клеткой;
scFv, связывающийся с NK-клеткой;
scFv, связывающийся с макрофагом; или
scFv, связывающийся с мононуклеарной клеткой.
14. Выделенная молекула нуклеиновой кислоты по любому из пп.1-13, где AB2 кодируемого мультиспецифичного активируемого антитела представляет собой анти-CD3 эпсилон scFv или анти-CTLA-4 scFv, необязательно, где анти-CD3 эпсилон scFv получен из OKT3.
15. Выделенная молекула нуклеиновой кислоты по любому из пп.1-14, где АВ1 кодируемого мультиспецифичного активируемого антитела представляет собой или получено из антитела, которое связывается с мишенью выбранной из группы, состоящей из 1-92-LFA-3, Альфа-4 интегрина, Альфа-V интегрина, Альфа4бета1 интегрина, Альфа4бета7 интегрина, AGR2Анти-Lewis-Y антитела, рецептора апелина J, APRIL, B7-H4, BAFF, BTLA, комплемента C5, C-242, CA9, CA19-9, Lewis a, карбоновой ангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD125, CD132 (IL-2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA-90), CLAUDIN-3, CLAUDIN-4, cMet, коллагена, Cripto, CSFR, CSFR-1, CTLA-4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL4, DPP-4, DSG1, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, белка F RSV, FAP, FGF-2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, рецептора фолата, GAL3ST1, G-CSF, G-CSFR, GD2, GITR, GLUT1, GLUT4, GM-CSF, GM-CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFN-альфа, IFN-бета, IFN-гамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL-12R, IL-12Rбета1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL-31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, лигандов Jagged, Jagged 1, Jagged 2, LAG-3, LIF-R, Lewis X, LIGHT, LRP4, LRRC26, MCSP, мезотелина, MRP4, MUC1, муцина-16 (MUC16, CA-125), Na/K АТФазы, эластазы нейтрофила, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM-R, OX-40, PAR2, PDGF-AA, PDGF-BB, PDGFR-альфа, PDGFR-бета, PD-1, PD-L1, PD-L2, фосфатидил-серина, P1GF, PSCA, PSMA, RAAG12, RAGE, SLC44A4, сфингозин 1 фосфата, STEAP1, STEAP2, TAG-72, TAPA1, TGF-бета, TIGIT, TIM-3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNF-альфа, TNFR, TNFRS12A, TRAIL-R1, TRAIL-R2, трансферрина, рецептора трансферрина, TRK-A, TRK-B, uPAR, VAP1, VCAM-1, VEGF, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP-1, WISP-2, WISP-3, или
где АВ1 кодируемого мультиспецифичного активируемого антитела представляет собой анти-Jagged антитело или анти-EGFR антитело, или
где AB1 кодируемого мультиспецифичного активируемого антитела содержит по меньшей мере часть аминокислотой последовательности, кодируемой последовательностями нуклеиновых кислот SEQ ID NO: 229, 231, 233, 235, 237, 239, 241, 253, 255, 257, 259, 261, 263, 265, 267, 269, или
где АВ1 кодируемого мультиспецифичного активируемого антитела содержит по меньшей мере часть аминокислотной последовательности SEQ ID NO: 230, 232, 234, 236, 238, 240, 254, 256, 258, 260, 262, 264, 266, 268, 270, или
где АВ1 кодируемого мультиспецифичного активируемого антитела представляет собой анти-Jagged антитело или анти-EGFR антитело и где АВ2 кодируемого мультиспецифичного активируемого антитела представляет собой анти-CD3 эпсилон scFc или анти-CTLA-4 scFv, или
где кодируемое мультиспецифичное активируемое антитело содержит по меньшей мере часть аминокислотной последовательности SEQ ID NO: 292, 294, 300, 304, 306, 308, 310, 336 или любой их комбинации.
16. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по любому из пп.1-15.
17. Способ получения мультиспецифичного активируемого антитела путем культивирования клетки в условиях, приводящих к экспрессии мультиспецифичного активируемого антитела, где клетка содержит молекулу нуклеиновой кислоты по любому из пп.1-15 или вектор по п.16.
18. Способ получения мультиспецифического активируемого антитела, включающий:
(a) культивирование клетки в условиях, приводящих к экспрессии мультиспецифичного активируемого антитела, где клетка содержит вектор по п.16; и
(b) выделение мультиспецифического активируемого антитела.
19. Применение мультиспецифичного активируемого антитела, кодируемого молекулой нуклеиновой кислоты по любому из пп.1-15 или вектором по п.16, для облегчения симптома клинического заболевания больного, где антитело вводят больному в количестве, достаточном для облегчения симптома клинического заболевания, где мультиспецифичное активируемое антитело активируется опухоле-ассоциированными протеазами, где заболевание представляет собой злокачественное заболевание, необязательно, где больным является человек.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| DONALDSON J.M | |||
| et al., "Design and development of masked therapeutic antibodies to limit off-target effects: application to anti-EGFR antibodies." Cancer biology & therapy, 2009; 8(22): 2147-2152 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |

