Изобретение относится к области генной и белковой инженерии и может быть использовано в анализах различного назначения, основанных на определении пероксидазной активности.
Пероксидаза катализирует окисление люминола в присутствии пероксида водорода с испусканием света. Такая реакция используется в биолюминесцентных методах анализа. В настоящее время наиболее распространенной пероксидазой для таких анализов является пероксидаза из корней хрена (Kricka L.J. (2000) Application of bioluminescence and chemiluminescence in biomedical sciences. Methods Enzymol. 305, 333-345).
Рекомбинантная пероксидаза хрена может производиться в подходящих рекомбинантных растениях (хрен, томаты, кукуруза) в виде гетерогенно-гликозилированной формы за счет суперпродукции. Также известна негликозилированная рекомбинантная пероксидаза хрена, полученная путем экспрессии в клетках Е.соli. В таком случае выход активной пероксидазы составляет порядка 50%.
Техническая задача, решаемая настоящим изобретением, состоит в получении нового белка, а именно мутантной формы рекомбинантной пероксидазы хрена, у которой стабильность в хемилюминесцентной реакции окисления люминола значительно выше, чем у рекомбинантной немутантной формы этого фермента.
Технический результат, получаемый при использовании полученной мутантной формы рекомбинантной пероксидазы хрена, состоит в значительном снижении предела обнаружения пероксидазы хрена в ходе анализа, что обеспечит коммерциализацию его как более стабильного реагента применительно к хемилюминесцентным методам анализа.
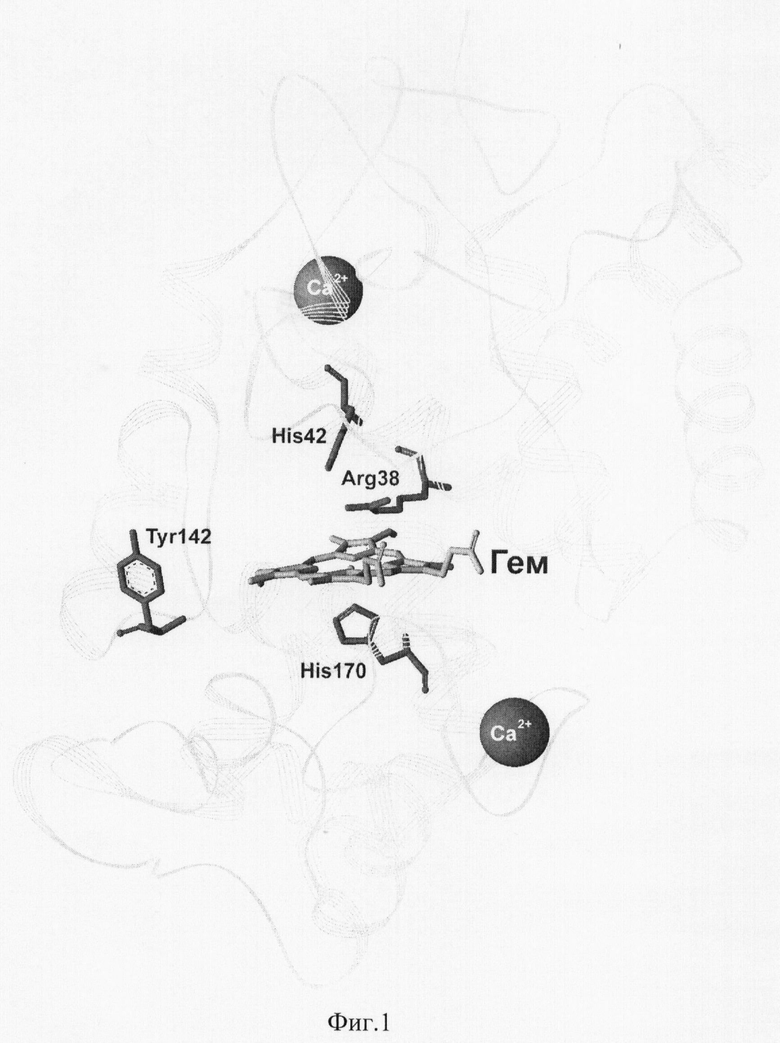
Для достижения указанного технического результата предложено ввести нуклеотидную замену в последовательность кДНК, кодирующую аминокислотную последовательность пероксидазы хрена, в результате чего происходит замена остатка фенилаланина в положении 142 в аминокислотной последовательности фермента на остаток тирозина (фиг.1). В таком случае получаемая мутантная пероксидаза хрена сохраняет исключительно высокую стабильность в реакции хемилюминесцентного окисления люминола, в то время как активность в реакции люминесценции существенно не уменьшается.
Сущность изобретения заключается в следующем. Введение нуклеотидных замен в гене пероксидазы хрена, обеспечивающих замену Phe142Tyr в аминокислотной последовательности фермента, осуществляют с использованием одного из известных специалистам методов направленного мутагенеза. Для последующей трансформации используют полученные плазмиды с введенными нуклеотидными заменами и рекомбинантные штаммы E.coli. Следующим этапом является культивирование трансформированных клеток E.coli до середины логарифмической фазы роста, после чего добавляют индуктор экспрессии гена пероксидазы и продолжают культивирование в течение 3 часов. Для выделения пероксидазы из телец включения полученную биомассу разрушают ультразвуком, центрифугируют, отмывают примесные белки солевым раствором, и солюбилизируют в мочевине. После чего проводят процедуру реактивации солюбилизированной пероксидазы, т.н. рефолдинг. Для этого готовят рефолдинг-среду, содержащую гемин, хлорид кальция и другие компоненты для обеспечения искусственного фолдинга активного фермента. По окончании реактивации рефолдинг-среду концентрируют с использованием ультрафильтрации или любого другого метода, позволяющего уменьшить объем раствора. Полученный препарат очищают с использованием хроматографии, в частности гель-фильтрации. Полученный препарат характеризуется RZ=1,9, удельной активностью по ABTS 1000 Е/мг (табл. 1) и аминокислотной последовательностью, изображенной на фиг. 2.
Сущность разработанного технического решения в дальнейшем будет раскрыта с использованием примера получения и примера использования мутантной формы пероксидазы хрена.
Пример 1.
Направленный мутагенез и получение мутантной пероксидазы хрена с заменой Phe142Thr
Для получения мутантной пероксидазы хрена с заменой Phe142Thr использовали плазмиду pSA 261, содержащую синтетический ген пероксидазы хрена дикого типа. Создание гена и плазмид описано в Европейском патенте ЕР 0299682, 1988 г.
Мутанты пероксидазы хрена получали методом направленного мутагенеза одноцепочечной ДНК. Для проведения сайт-специфического мутагенеза исходная плазмида pSA 261 была рестрицирована HpaI и XbaI, и фрагмент гена пероксидазы размером 0,7kb выделен и клонирован в фаговый вектор М13ВМ20 по сайтам XbaI и HincII.
Для достраивания второй цепи ДНК был применен набор фирмы Amersham для сайт-специфичного мутагенеза и олигонуклеотидные праймеры, несущие точечные мутации. Для получения мутации Phe142Tyr были синтезированы праймеры:
5'-AAATCTACCGGCGCCATATTTCACTCTACCACAAC-3' - прямой и 5'-GTTGTGGTAGAGTGAAATATGGCGCCGGTAGATTT-3' - обратный, имеющие в области мутации сайт рестрикции Bsp119-I, обеспечивающий первичную селекцию мутанта. После достраивания второй цепи ДНК, исходная первая цепь была разрушена, а новая вторая упакована в фаги и высеяна. Отбор фагов осуществлен по появлению или соответственно исчезновению сайтов рестрикции, и ДНК мутантных фагов рестрицирована по NruI и XbaI и клонирована в плазмиду pSA 261, рестрицированную по тем же сайтам.
Отбор клонов с плазмидой, несущей мутацию, осуществляли рестрикционным анализом. Плазмиды были выделены в препаративном количестве и проанализированы рестрикцией BspHI, Bsp119-I и SacI.
Для проведения секвенирования больший SacI фрагмент гена пероксидазы (0,7 kb) клонирован в фаг М13 и считан в двух направлениях. Оба праймера были синтезированы на синтезаторе 380В фирмы Allpied Biosystems, и конечные конструкции были секвенированы на секвенаторе 370А той же фирмы, используя Taq-полимеразу и меченые красителями праймеры. Полученная плазмида, содержащая мутацию, была использована для трансформации штамма E.coli JM 109.
Для получения биомассы клеток с рекомбинантной мутантной пероксидазой хрена единичную колонию трансформированных клеток Е. coli JM 109 помещали в 3 мл среды 2YT (16 г/л бактотриптона, 10 г/л дрожжевого экстракта и 5 г/л хлорида натрия, рН 7,0) в присутствии 40 мкг/мл ампициллина в течение ночи при 37°С и 180 об/мин. Далее из этой пробирки отбирали 1 мл культуральной жидкости и помещали в 200 мл той же среды, находящейся в качалочной колбе объемом 1 л. Культивирование проводили при 37°С и 100 об/мин. Клетки доращивали до начала логарифмической фазы и затем проводили индукцию экспрессию гена пероксидазы добавлением в среду ИПТГ (Изопропил-β-D-1-тиогалактопиранозид) до конечной концентрации 0,2 мМ. Далее температуру понижали до 34°С и культивировали клетки в течение 3 часов. Биомассу собирали центрифугированием при 5000 g на центрифуге фирмы "Eppendorf" 5403. Затем клетки ресуспендировали в 2 М растворе NaCl, содержащем 10 мМ дитиотреитола (ДТТ), и разрушали ультразвуком в течение 10 мин при 22 кГц. Смесь после разрушения оставляли на 1,5 ч при комнатной температуре, и затем повторяли обработку ультразвуком. Надосадочную жидкость удаляли, осадок промывали 10 мл 0,05 М буфера Трис-HCl, рН 8,5, и солюбилизировали в 10 мл 6 М мочевины, содержавшей 1 мМ ДТТ (дитиотреитол). Солюбилизированный апобелок (95% чистоты) приливали по каплям к среде для рефолдинга (100 мл) и инкубировали при 4°. Среда для рефолдинга содержала оптимизированные концентрации мочевины (2 М), окисленного глутатиона (0,7 мМ), ДТТ (0,1 мМ), а также 5 мМ СаСl2, 5 мкМ гемина, 5% глицерина в 50 мМ буфере Трис-HCl, рН 8,5. В процессе инкубации измеряли активность по АБТС. После прекращения роста активности раствор фермента концентрировали в 30 раз на ячейке Amicon объемом 50 мл через мембрану YM-10. Полученный концентрированный раствор фермента наносили порциями по 3 мл на колонку Toyopearl HW 55 уравновешенным 50 мМ буфером Трис-HCl, рН 8,5. Фракции с RZ>2,7 отбирали, а остальные активные фракции подвергали концентрированию и повторной гельфильтрации. Фракции, содержащие активный фермент, объединяли и концентрировали. Чистота полученных препаратов контролировалась значением RZ и электрофоретически. Сравнение данных электрофореограмм очищенных рекомбинантных пероксидазы хрена дикого типа и мутантной формы Phe142Tyr в присутствии белков-маркеров молекулярного веса выявило чистоту порядка 95% и молекулярный вес 36 кДа для обеих форм.
Пример 2.
Свойства мутантной пероксидазы хрена и фермента дикого типа в реакции хемилюминесцентного окисления люминола.
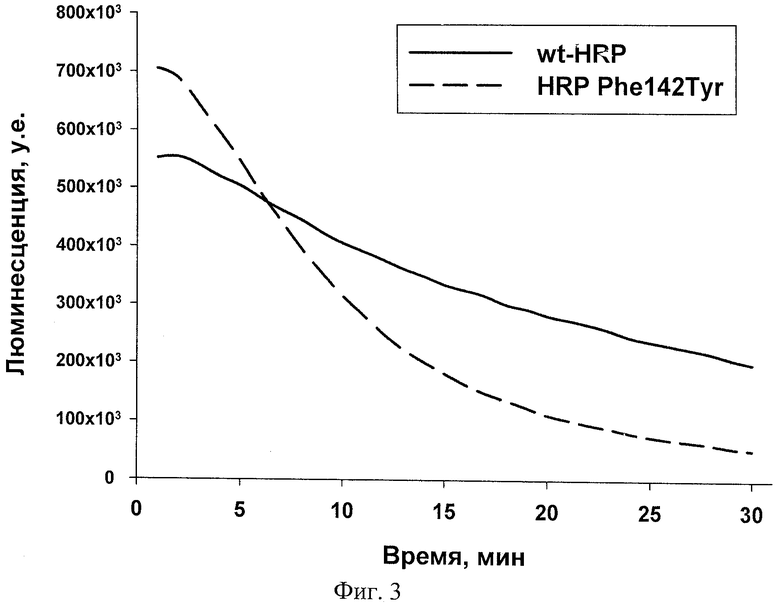
Свойства нового мутанта пероксидазы хрена Phe142Tyr сравнивали с таковыми для рекомбинантной пероксидазы хрена дикого типа. Оптимальные условия катализа окисления люминола не отличаются для обеих форм пероксидазы хрена, а стабильность сигнала в случае мутанта значительно выше (фиг. 3), что обеспечивает более высокую чувствительность определения.
| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ ПЕРОКСИДАЗА ТАБАКА | 2007 |
|
RU2360968C1 |
| ЛЮЦИФЕРАЗЫ | 1995 |
|
RU2192467C2 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ (ВАРИАНТЫ) | 2012 |
|
RU2507262C1 |
| МОДИФИЦИРОВАННАЯ ТЕРМОЛИЗИНПОДОБНАЯ НЕЙТРАЛЬНАЯ МЕТАЛЛОПРОТЕАЗА, СПОСОБ СИНТЕЗА БЕНЗИЛОКСИКАРБОНИЛ-α-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИН МЕТИЛОВОГО СЛОЖНОГО ЭФИРА, СПОСОБ ГИДРОЛИЗА БЕНЗИЛОКСИКАРБОНИЛ-α-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИН МЕТИЛОВОГО СЛОЖНОГО ЭФИРА | 1993 |
|
RU2151186C1 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ | 2009 |
|
RU2412998C1 |
| МУТАНТНАЯ ЛЮЦИФЕРАЗА (ВАРИАНТЫ), ДНК, КОДИРУЮЩАЯ УКАЗАННУЮ ЛЮЦИФЕРАЗУ, И ВЕКТОР ДЛЯ ЭКСПРЕССИИ УКАЗАННОГО БЕЛКА | 1996 |
|
RU2210594C2 |
| МУТАНТНЫЕ ОКСИДАЗЫ D-АМИНОКИСЛОТ | 2007 |
|
RU2362806C2 |
| Мутантная копеподная люцифераза для применения в качестве биолюминесцентного репортера in vitro и in vivo | 2020 |
|
RU2757736C1 |
| МУТАНТНАЯ ГЛЮКОЗОИЗОМЕРАЗА С ИЗМЕНЕННОЙ СУБСТРАТНОЙ СПЕЦИФИЧНОСТЬЮ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 1991 |
|
RU2096457C1 |
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ФОРМИАТДЕГИДРОГЕНАЗА | 2006 |
|
RU2312897C1 |

Изобретение относится к области биохимии. Представлена мутантная пероксидаза хрена, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности пероксидазы хрена дикого типа с заменой остатка фенилаланина в положении 142 на остаток тирозина. Изобретение позволяет снизить предел обнаружения пероксидазы хрена в ходе анализа. 3 ил., 1 табл., 2 пр.
Мутантная пероксидаза хрена, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности пероксидазы хрена дикого типа с заменой остатка фенилаланина в положении 142 на остаток тирозина.
| ЗАХАРОВА Г.С | |||
| и др | |||
| Пероксидаза из корней хрена: модулирование свойств химической модификацией белковой глобулы и гема | |||
| - Успехи биологической химии, 2011, т.51, с.37-3647 | |||
| VEITCH N et al | |||
| Устройство для усиления микрофонного тока с применением самоиндукции | 1920 |
|
SU42A1 |
| Eur | |||
| J | |||
| Biochem., 1995, V.23, n.3, p.p.650-658 | |||
| RU 2008104692 A, 12.02.2008. | |||

