Изобретение относится к областям молекулярной и клеточной биотехнологии и, в частности, к люциферазам. Изобретение представляет собой синтетическую копеподную люциферазу ThermoLuc, полученную направленной молекулярной эволюцией и множественным случайным мутагенезом на основе MLuc7 изоформы люциферазы Metridia longa и демонстрирующую улучшенные свойства в сравнение с люциферазой дикого типа, для использования в качестве биолюминесцентного репортера для визуализации молекулярных процессов в живых клетках in vivo, мечения белков и клеток, и в различных люминометрических анализах in vitro.
Копеподные люциферазы - семейство небольших секреторных белков массой 18,4-24,3 кДа, с учетом отщепляемого сигнального пептида для секреции, - ответственны за яркую секретируемую биолюминесценцию некоторых морских веслоногих рачков. Эти люциферазы катализируют простую реакцию окисления низкомолекулярного субстрата целентеразина без каких-либо дополнительных кофакторов, сопровождающуюся излучением голубого света. Простота реакции, отсутствие ее токсичности для живой клетки, неинвазивность и легкость регистрации квантов в широком динамическом диапазоне сопоставимо с точностью радиоизотопных меток, а также отличная проницаемость клеточных мембран для субстрата целентеразина позволяет широко использовать целентеразин-зависимые люциферазы как репортеры молекулярных процессов в живых клетках и организмах. Копеподные люциферазы не имеют никакого сходства в аминокислотных последовательностях или пространственных структурах с другими идентифицированными биолюминесцентными белками, включая люциферазы Renilla и Oplophorus, также использующие целентеразин в качестве субстрата. Небольшой размер, высокая активность и стабильность, в том числе термостабильность, делают секретируемые люциферазы копепод очень привлекательными кандидатами на роль репортерных белков в биомедицинских исследованиях, которые особенно полезны для различных неразрушающих репортерных анализов, в том числе в высокопроизводительном формате.
Наиболее известными и исследованными представителями копеподных люцифераз являются первые клонированные люциферазы GpLuc (или GLuc) и MLuc из рачков Gaussia princeps и Metridia longa соответственно. Сразу после клонирования эти гомологичные люциферазы были успешно применены в качестве биолюминесцентных репортеров in vivo и in vitro, и с тех пор сфера их применения продолжает расширяться [Markova S.V. et. al, Photochem. Photobiol., 2019, 95: 705-721]. Но природные белки, как правило, нуждаются в улучшении свойств для использования в качестве репортеров.
Известен M43I мутант люциферазы из Gaussia princeps (GLuc, ~18.2 кДа) [патент US 8980603 В2, МПК C12N 9/00, опубл. 17.03.2015], который демонстрировал более медленную кинетику светоизлучения в сравнении с диким типом, подходящую для высокопроизводительных скрининговых исследований, но только в присутствии 0,1% детергента Тритон Х-100. Данный подход не подходит для анализа живых клеток, так как Тритон Х-100 их разрушает.
Другие известные мутанты люциферазы из Gaussia princeps, заявленные как суперлюминесцентные мутанты со значительно повышенной активностью, стабильностью светового сигнала и измененным спектром световой эмиссии [патент ЕР 2420573 А1, МПК C12N 15/09, опубл. 22.02.2012], не нашли впоследствии полного подтверждения своих заявленных характеристик [Wu Т. et. al, 2015, BBA-Proteins Proteom., 1854:1392-1399; Hunt E. A. et. al, Sci. Rep.8:26814]. Недостатком этих мутантов и других известных на сегодняшний день мутантов GLuc люциферазы Gaussia, а также укороченного мутанта люциферазы ML-Green изоформы MLuc 164 из Metridia longa [RU 2009130108 A, C12N 9/02, опубл. 20.10.2013] является то, что они не были охарактеризованы в отношении температурных оптимумов биолюминесцентной реакции и их эффективности при работе в условиях повышенной температуры до 37°С.
Наиболее близким аналогом, принятым за прототип, рассматривается изоформа MLuc7 люциферазы Metridia longa, которая с размером в 152 аминокислотных остатка и 16,5 кДа в зрелом виде является наименьшей природной люциферазой из известных [WO 2008095622 A3, C12Q 1/66, опубл. 02.10.2008]. Высокая активность, малый размер и способность сохранять свои свойства в составе гибридных белков обуславливают ее привлекательность как биолюминесцентного репортера, обеспечивая наименьшую метаболическую нагрузку на клетку при использовании в качестве генетически-кодируемого репортера, и наименьшую структурную интерференцию с присоединяемым доменом при использовании в качестве метки в гибридных белках и других соединениях. Однако, успешное использование MLuc7 как репортера в живых клетках млекопитающих ограничивается низким температурным оптимумом люциферазной реакции, лежащим в диапазоне 10-17°С, а при температуре 37°С, являющейся оптимальной для роста клеток млекопитающих, остаточная активность составляет около 10% (Фиг. 2). Похожая картина наблюдается и для другой популярной копеподной люциферазы GpLuc (-18.2 кДа) из Gaussia princeps с оптимумом в 14-20°С и остаточной активностью при 37°С около 20% [Markova S.V. et. al, Photochem. Photobiol., 2019, 95: 705-721]. Таким образом, получение мутантных форм люциферазы с расширенным или смещенным в сторону более высоких температур оптимумом реакции повышает чувствительность репортера за счет увеличения его активности в диапазоне рабочих температур от комнатной в 23°С до температуры 37°С, оптимальной для клеток млекопитающих.
Кроме того, кинетика биолюминесцентной реакции MLuc7, как и других копеподных люцифераз, в оптимальных условиях с большим избытком субстрата характеризуется эмиссией типа «вспышка», когда световой сигнал достигает максимума в течение нескольких секунд после добавления субстрата, а затем быстро снижается. Такая кинетика реакции репортерного белка подразумевает мгновенную регистрацию сигнала каждого образца после добавления субстрата, что требует люминометров с функцией впрыска, увеличивает время измерения множественных образцов в планшетном формате и существенно затрудняет использование таких репортеров для высокопоточных анализов. Соответственно, увеличение стабильности светового сигнала очень желательно для биолюминесцентного репортера, так как это ведет к упрощению проведения измерений и к расширению сфер применения маркерного белка.
Основной задачей, на решение которой направлено заявляемое изобретение, является расширение арсенала маркировочных средств, используемых в молекулярной и клеточной биологии, медицинской диагностике и биотехнологии.
Техническим результатом изобретения является получение нового синтетического биолюминесцентного репортера - люциферазы ThermoLuc на основе высокоактивной 16,5 кДа MLuc7 изоформы люциферазы Metridia longa, с повышенной чувствительностью и большей стабильностью светового сигнала.
Технический результат достигается тем, что мутантная копеподная люцифераза ThermoLuc для применения в качестве биолюминесцентного репортера in vitro и in vivo получена путем направленной молекулярной эволюции из MLuc7 изоформы люциферазы Metridia longa, новым является наличие аминокислотных замен Y72W, 173 L, G81E, G90A и D146V в последовательности, характеризуется увеличенной активностью, температурным оптимумом реакции в области 31-38°С и повышенной стабильностью люминесцентного сигнала.
Сущность изобретения заключается в том, что путем введения множественных точечных аминокислотных замен в природную последовательность MLuc7 изоформы люциферазы Metridia longa размером -16 кДа в положениях 72, 73, 81, 90 и 146 (нумерация по полноразмерной последовательности, включающей отщепляемый природный сигнальный пептид для секреции - Seq. l), 4 из них находятся в двух высококонсервативных повторах, формирующих молекулу люциферазы (Фиг. 1). При этом в природной последовательности в положении 72 тирозиновый остаток заменен на триптофановый (Y72W), в положении 73 изолейциновый остаток заменен на лейциновый (173 L), в положении 81 глициновый остаток заменен на остаток глутаминовой кислоты (G81E), в положении 74 глициновый остаток заменен на аланиновый (G90A) и в положении 146 остаток аспарагиновой кислоты заменен на валиновый (D146V). Эти ключевые для изменения свойств остатки были идентифицированы по отдельности путем направленной молекулярной эволюции: множественным случайным мутагенезом MLuc7 изоформы люциферазы Metridia longa с последующим отбором мутантных вариантов, выявляющих большую активность при 37°С и большую стабильность биолюминесцентного сигнала. Первичную селекцию мутантных люцифераз проводили прямо на бактериальных колониях, экспрессирующих люциферазу в состав внешней клеточной мембраны. После опрыскивания колоний субстратом целентеразином, свечение регистрировали высокочувствительной ПЗС-камерой с охлаждением. Наилучшие варианты единичных мутаций, ведущие к улучшению репортерных свойств, были объединены на одной матрице методом олигонуклеотид-направленного мутагенеза. Окончательный вариант комплекта мутаций для ThermoLuc люциферазы был выбран после характеризации очищенных препаратов активных мутантных рекомбинантных белков, полученных путем рефолдинга и хроматографической очистки из телец включения при экспрессии в Е. coli.
В заявляемом изобретении посредством случайного и сайт-направленного мутагенеза получен белок люциферазы ThermoLuc, молекула которого содержит аминокислотные замены, приводящие к повышению активности, сдвигу температурного оптимума реакции в диапазон 31-38°С и увеличению стабильности светового сигнала, что как по отдельности, так и в сочетании обуславливает общее увеличение чувствительности биолюминесцентного репортера и удобства его измерения как для живых клеток и тканей in vivo, так и для применения в аналитике in vitro.
Основными отличиями предлагаемой синтетической люциферазы ThermoLuc - 16.5 кДа для использования в качестве биолюминесцентного репортера от выбранного прототипа являются другая аминокислотная последовательность, содержащая улучшающие репортерные свойства мутации, а также повышение активности в сравнение с диким типом, наличие температурного оптимума реакции в диапазоне 31-38°С и увеличение стабильности светового сигнала.
Таким образом, перечисленные выше отличительные от прототипа признаки позволяют сделать вывод о соответствии заявляемого технического решения критерию «новизна».
Совокупность признаков, отличающие заявляемое техническое решение от прототипа, не выявлены в других технических решениях и, следовательно, обеспечивают заявляемому решению соответствие критерию «изобретательский уровень».
Сущность изобретения поясняется с помощью графических материалов:
На Фиг. 1 представлено сравнение аминокислотных последовательностей (элаймент): ThermoLuc для секреторной экспрессии с наличием сигнального пептида и для экспрессии в виде зрелого белка в сравнение с исходной изоформой MLuc7 из Metridia longa. Мутации, отличающие ThermoLuc, выделены серыми (сходные по свойствам аминокислотные остатки) и черными (различающиеся по свойствам аминокислотные остатки) квадратами, мутации также отмечены звездочками внизу.
На Фиг. 2 представлена зависимость пиковой эмиссии биолюминесцентного сигнала от температуры для белка изобретения ThermoLuc (черная линия) в сравнении с природный люциферазой дикого типа MLuc7 из Metridia longa (серая линия).
На Фиг. 3 представлены нормированные кинетические кривые биолюминесцентного сигнала белка изобретения ThermoLuc в сравнении с природной люциферазой дикого типа MLuc7 из Metridia longa, демонстрирующие замедленное падения уровня светового сигнала ThermoLuc со временем.
На Фиг. 4 представлена зависимость активности люциферазы ThermoLuc от температуры инкубации в течение часа (кривая термоинактивации) в сравнении с природной люциферазой дикого типа MLuc7.
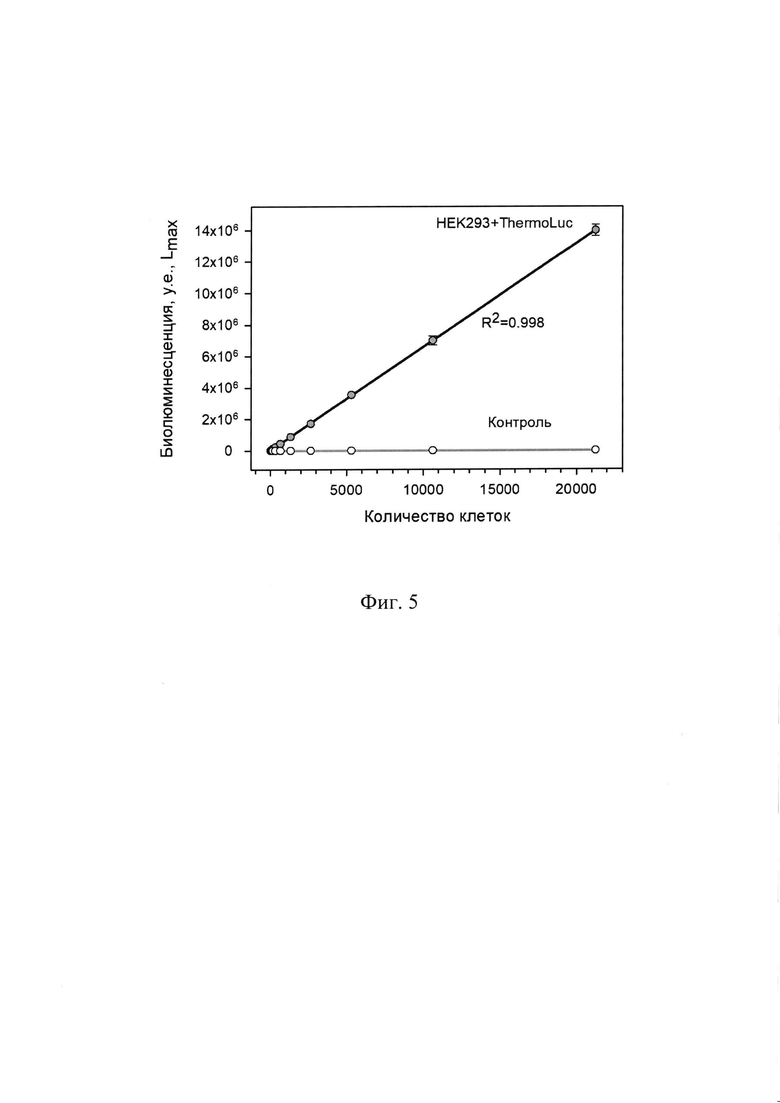
На Фиг. 5 представлена зависимость интенсивности биолюминесцентного сигнала в культуральной среде от количества экспрессирующих ThermoLuc жизнеспособных клеток НЕК293 в лунке 96-луночного планшета. Культуральная среда была проанализирована через 24 ч после рассева клеток с серийным разведением 2.
Каждая точка - среднее значение четырех независимых измерений. Минимальное количество - 5 клеток в образце.
Описание представленных аминокислотных последовательностей (ДНК последовательности для одного и того же белка ThermoLuc могут быть разными в зависимости от выбранной клетки-хозяина для экспрессии вследствие вырожденности генетического кода):
Последовательность №1 (Seq. 1) представляет полноразмерную последовательность белка изобретения ThermoLuc, включающую отщепляемый природный сигнальный пептид для секреции для варианта секреторной эукариотической экспрессии и бактериальной экспрессии в периплазматическое пространство.
Последовательность №2 (Seq. 2) представляет зрелую последовательность ThermoLuc без отщепляемого природного сигнального пептида для секреции.
Последовательность №3 (Seq. 3) представляет зрелую последовательность ThermoLuc без отщепляемого природного сигнального пептида, дополненная метионином, инициирующим трансляцию. Данная последовательность используется для цитоплазматической экспрессии и бактериального синтеза люциферазы для определения биолюминесцентных свойств и использования в аналитике in vitro.
Последовательность №4 (Seq. 4) представляет пример гена, кодирующего полноразмерную последовательность ThermoLuc с отщепляемым природным сигнальным пептидом для секреции и двумя стоп-кодонами для успешной экспрессии в эукариотических клетках.
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение последовательности синтетической люциферазы ThermoLuc с улучшенными репортерными свойствами
Получение последовательности люциферазы ThermoLuc. Все генно-инженерные манипуляции, если не оговорено особо, были выполнены с помощью методов, хорошо известных для специалистов в данной области. Синтетическая люцифераза ThermoLuc с улучшенными свойствами была получена направленной молекулярной эволюцией и множественным случайным мутагенезом на основе высокоактивной MLuc7 изоформы люциферазы Metridia longa, обладающей наименьшими размерами среди природных люцифераз. Для этого последовательность MLuc7 (Gene ID KP242023) была клонирована в вектор pET22b + в виде фьюжина с белком YebF (Gene ID 946363) внешней мембраны Е. coli [Zhang G. et. al., Nat. Biotechnol., 2006, 24:100-104] в векторе pET22-YebF-MLuc7, что обеспечило отличное созревание и доставку люциферазы в состав внешней мембраны бактериальной клетки. Наличие люциферазы в составе внешней мембраны клетки стерически обеспечило максимальный биолюминесцентный ответ на добавление субстрата путем опрыскивания колоний на чашках и позволило проводить селекцию мутантов прямо на колониях. Библиотека мутантов люциферазы MLuc7 была создана путем клонирования в конструкцию pET22-YebF набора мутантных молекул, синтезированных при использовании набора для случайного мутагенеза «Gene Morph II Random Mutagenesis Kit» (Agilent). Условия синтеза были подобраны так, чтобы в среднем на одну последовательность люциферазы приходилось до 6 нуклеотидных мутаций.
Финальная библиотека может быть получена методом электропорации и составляет обычно около 100 тысяч независимых клонов Е. coli. Клоны были рассеяны на чашки Петри по ~1-3 тысячи клонов на чашку для последующего функционального скрининга. Чашки опрыскивали из пульверизатора раствором субстрата целентеразина и немедленно фиксировали свечение при помощи высокочувствительной имиджинг-камеры BFI-System, разработанной ИПФ СО РАН (Нижний Новгород). Данный способ позволял производить отбор колоний по интенсивности их свечения после обработки субстратом в различных условиях. Кроме того, фиксировалась длительность свечения колоний. Лучшие варианты были идентифицированы секвенированием, далее эти улучшающие репортерные свойства мутации были собраны на единой матрице люциферазы путем олигонуклеотид-направленного мутагенеза при использовании набора QuikChange® Site-Directed Mutagenesis Kit (Agilent) согласно руководству производителя. Полученная финальная аминокислотная последовательность показана в Seq. 2.
Пример 2. Получение препаратов белка ThermoLuc
Белок синтетической люциферазы ThermoLuc в достаточных количествах с целью характеризации свойств и использования в аналитических целях in vitro может быть легко получен как минимум в двух системах экспрессии: 1) его природная форма напрямую может быть получена очисткой из клеточной среды при использовании Вас-to-Bac бакуловирусной системы экспрессии (Invitrogen) в клетках насекомых, например Sf9; 2) путем окислительного рефолдинга из телец включения при экспрессии в клетках E.coli, например, в экспрессионной плазмиде pET22b + (Novagen).
Для эукариотической бакуловирусной экспрессии в клетках насекомых ген, кодирующий полноразмерную люциферазу ThermoLuc (Seq. 1) клонировали во фрагмент бинарного бакуловирусного вектора плазмиду pFastBacl (Invitrogen). В конец гена через сайт TEV протеазы (ENLIF/G) вводили отщепляемый семичленный полигистидиновый таг His7 для аффинной очистки. Далее рекомбинантный вектор трансформировали в клетки Е. coli DH10Bac, где формировалась бакмида с геном люциферазы. Далее бакмиду трансфецировали в насекомные клетки Sf9 с использованием трансфецирующего реагента Cellfectin (Gibco), амплифицировали вирус, заражали клетки Sf9 и препаративно нарабатывали белок согласно руководству Bac-to-Bac Baculovirus expression system (Invitrogen). Эффективность трансфекции и наработку вируса можно отслеживать по активности люциферазы в среде.
Очистку нативного белка ThermoLuc проводили из бессывороточной культуральной среды насекомных клеток через 72 часа после заражения в несколько этапов. После осаждения клеток центрифугированием люциферазу концентрировали из среды с помощью дифференциального осаждения сульфатом аммония 40-65% (вес/объем). Затем все растворяли в Ni-связывающем буфере (0,3 М NaCl, 20 мМ имидазол, 50 мМ фосфатный буфер, рН 7,1). Смесь пропускали через 5 мл колонку HisTrap (GE Healthcare) и элюировали градиентом имидазола (0-0,5 М) в том же буфере. Пик люциферазы концентрировали на ячейке Amicon (EMD Millipore) с одновременной заменой буфера для расщепления протеазой TEV (0,3 М NaCl, 0,02 мМ DTT, 20 мМ Трис-HCl, рН 7,5). His7-фрагмент отщепляли TEV-протеазой в соотношении 50:1 (вес/вес) путем инкубации в течение ночи образца люциферазы при 4°С. Затем образец повторно пропускали через колонку Ni-NTA для сорбционного удаления TEV-протеазы, также содержащей полигистидиновый фрагмент. Прошедшее через колонку концентрировали с заменяющим буфером (0,3 М NaCl, 1 мМ ЭДТА, 20 мМ Трис-HCl, рН 7,5) и пропускали через гель-фильтрационную колонку Superdex 75 (GE Healthcare), уравновешенную тем же буфером. На выходе образец люциферазы был гомогенным в соответствии с SDS-ПААГ. Выход люциферазы составлял 3-5 мг/л культуры клеток насекомых. Для хранения в концентрированный образец ThermoLuc добавляли 0,01% NP-40. Для длительного хранения добавляли еще и 50% глицерин и хранили при -20°С. В этих условиях активность люциферазы ThermoLuc полностью сохраняется как минимум 6 месяцев. Лиофильно высушенный образец ThermoLuc (без глицерина) полностью восстанавливает свою активность как минимум через 1 год хранения при -20°С.
Для бактериальной экспрессии использовали вариант последовательности ThermoLuc Seq. 3 без сигнального пептида и прямую цитоплазматическую экспрессию при использовании плазмиды pET22b + (Novagen) в штамме Е. coli BL21-CodonPlus (DE3)-RIPL (Agilent). Клетки E.coli BL21 с экспрессионной плазмидой pET22-ThermoLuc, наращивали в среде LB, содержащей ампициллин (200 мкг/мл), при 37°С до достижения оптической плотности OD600=1.0-1.2. Синтез ThermoLuc индуцировали добавлением ИПТГ до конечной концентрации 1 мМ. После индукции культивирование продолжали при 37°С в течение 1 часа так как клетки переставали расти после начала синтеза люциферазы. При экспрессии в клетках Е. coli в векторе pET22b+(Novagen), рекомбинантная люцифераза Metridia longa и ее мутанты образуют нерастворимые тельца включения, которые использовались для последующей очистки мутантной люциферазы ThermoLuc с ее одновременным рефолдингом. Клетки осаждали центрифугированием при 4°С, ресуспендировали (1:20, вес/объем) в буфере 1 мМ ЭДТА, 20 мМ Трис-HCl, рН 8,0 и разрушали ультразвуком (10 с × 6) при 0°С, а затем центрифугировали при 14000 g в течение 20 минут.Осадок телец включения последовательно промывали следующими растворами: (а) 0,9% NaCl, 1 мМ ЭДТА, 20 мМ Трис-HCl, рН 8,0; (б) 1% Тритон Х-100, 1 мМ ЭДТА, 20 мМ Трис-HCl, рН 8,0 для удаления примесных компонентов.
Для получения нативной структуры ThermoLuc (фолдинга) тельца включения, полученные из 100-200 мл культуры растворяли в 1 мл раствора, содержащего 4 М гуанидин-HCl, 5 мМ ЭДТА, в течение 1 часа. Затем смесь центрифугировали и к супернатанту добавляли 40 мМ Трис-HCl рН 8,0 и 50 мМ DTT для полного восстановления цистеиновых остатков в течение 2-3 ч. Для корректного фолдинга белка 1 мл экстракта ThermoLuc по каплям добавляли к 30 мл буфера при 4°С, содержащего 20 мМ Трис-HCl рН 7,0, 150 мМ NaCl, 0,3 М аргинин, 10 мМ MgCl2, 5 мМ GSH (глутатион восстановленный), 0,5 мМ GSSG (глутатион окисленный), 0,01% NP-40. Инкубировали с перемешиванием 1 час, затем добавляли 5 мкл 1 М CuCl2 и оставляли на ~16 часов при 4°С при постоянном перемешивании. Полученный раствор концентрировали центрифугированием на ячейках Amicon-Ultra (EMD Millipore) до объема 0.5-1 мл и обессоливали образец пропусканием через колонку 5 ml HiTrap desalting (GE Healthcare) в буфере 20 мМ Bis-Tris-пропан рН 9.3, 50 мМ NaCl, 1 мМ ЕДТА, 0.01% NP-40, 5% глицерин, 20 мкМ ДТТ. Далее весь собранный объем, имеющий биолюминесцентную активность, пропускали через анионообменную колонку 5 ml HiTrap Q HP в том же буфере для сорбции загрязняющих кислых белков Е. coli. Прошедший объем с люциферазной активностью концентрировали до не менее чем 500 мкл и подвергали гель-фильтрации на колонке Sephadex75, уравновешенной буфером 20 мМ Tris-HCl рН 7,5, 150 мМ NaCl, 1 мМ ЕДТА, 0,01% NP-40, 5% глицерин, 20 мкМ ДТТ. Чистоту полученного белка контролировали с помощью электрофореза в ПААГ. Содержание целевого белка составило не менее 99%. Мономерная фракция соответствовала по своим свойствам нативно сложенному белку, полученному из насекомных клеток, и может быть использована для тестирования физико-химических свойств и для использования в аналитических приложениях in vitro. Концентрацию белка определяли биуретовым методом с использованием DC™ Protein Assay kit (Bio-Rad).
Пример 3. Основные биолюминесцентные свойства синтетической люциферазы ThermoLuc
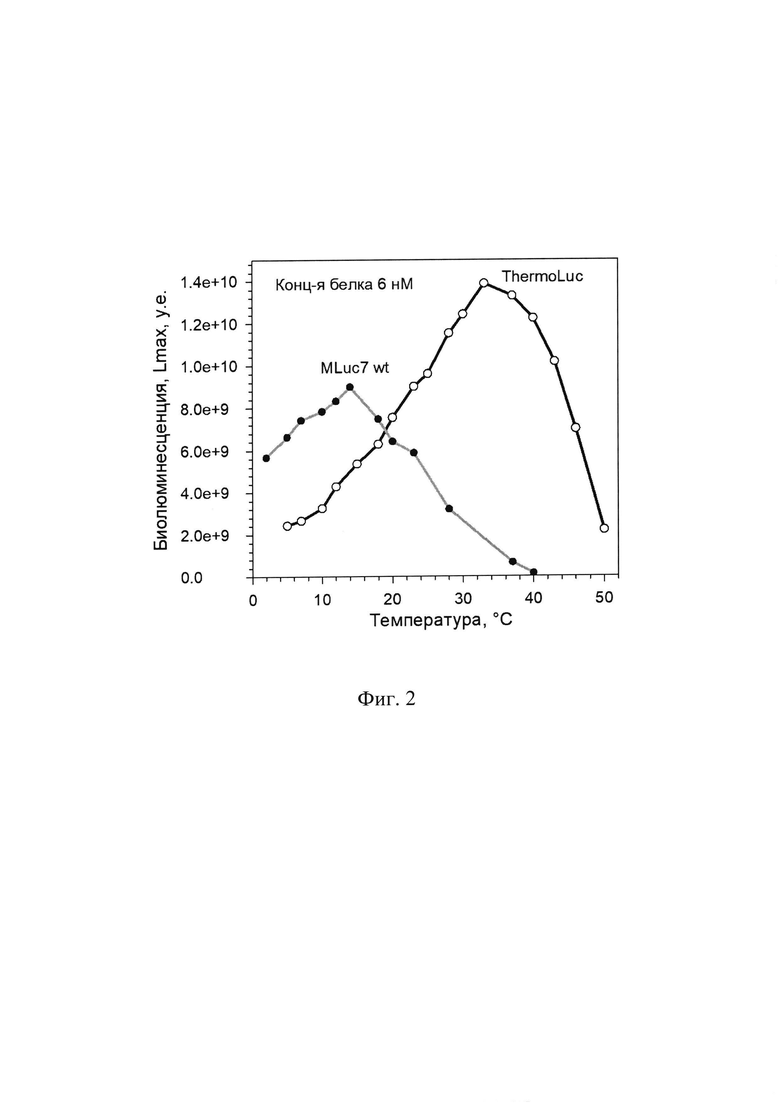
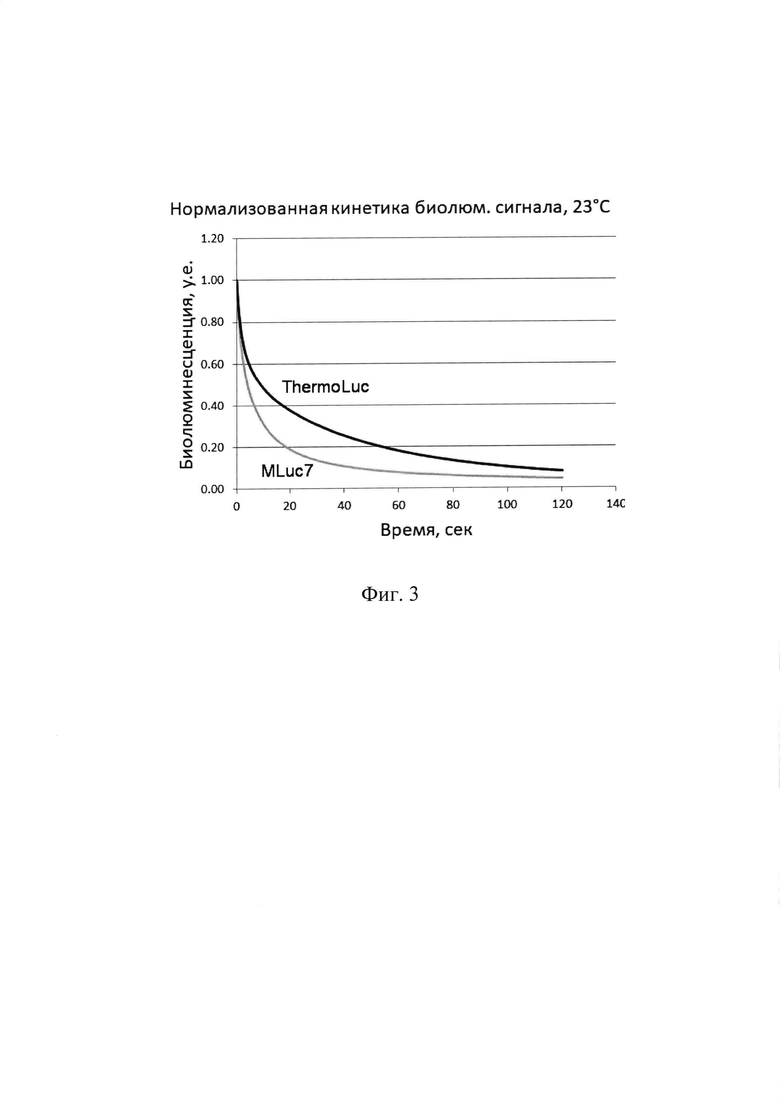
Полученные препараты синтетической люциферазы ThermoLuc анализировали по спектрам биолюминесценции, уровню биолюминесцентной активности (Таблица 1), кинетике биолюминесцентной реакции (Фиг. 3), термостабильности (Фиг. 4) и определяли зависимость активности от температуры для определения температурного оптимума реакции в сравнении с исходной люциферазой дикого типа MLuc7 из копепод Metridia longa (Фиг. 2).
Спектр биолюминесценции. Спектр биолюминесценции измеряли на спектрофлуориметре Сагу Eclipse (Varian Сагу, Австралия) при температуре 23°С. Спектр записывали в ML-буфере (0,5 М NaCl, 50 мМ Трис-HCl рН 7,5, 0,01% желатин) содержащем 0,01% NP-40. Образцы выравнивали по концентрации белка, которая в кювете составляла 0,3 мкг/мл. Биолюминесцентную реакцию запускали добавлением 10-кратного избытка целентеразина, спектры записывали при минимальном падении интенсивности сигнала, не требующего коррекции спектра на падение интенсивности сигнала. Коррекция спектров биолюминесценции на спектральную чувствительность ФЭУ была проведена с помощью программного обеспечения прибора. По итогам измерений спектры ThermoLuc и люциферазы дикого типа MLuc7 выявляли несущественные различия.
Уровень биолюминесцентной активности и кинетика биолюминесцентного сигнала. Биолюминесцентную активность определяли, впрыскивая 5 мкл спиртового 0.1 мМ раствора целентеразина в кювету биолюминометра БЛМ-003 с температурной стабилизацией (СКТБ «Наука», Красноярск, РФ), содержащую 500 мкл ML-буфера с образцом белка. Финальная концентрация целентеразина в кювете составляла 1 мкМ. Запись биолюминесценции проводили до 10 мин с интеграцией сигнала каждые 0,1 сек, по кривой падения сигнала математически с использованием программного обеспечения SigmaPlot 12 определяли кинетические параметры и тотальный сигнал. Для построения графиков использовали максимальные значения биолюминесцентной активности Lmax 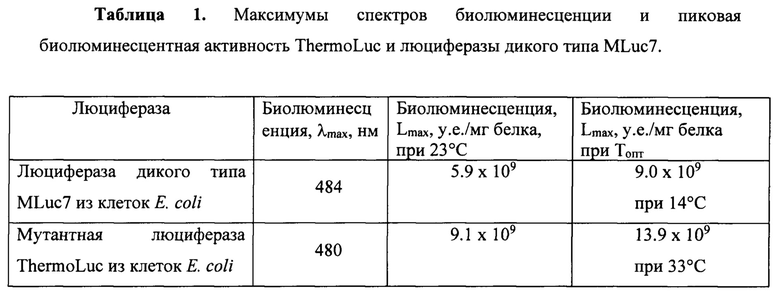
Как можно видеть из таблицы, мутантная люцифераза по изобретению ThermoLuc имеет увеличенную активность пиковой эмиссии в сравнении с люциферазой дикого типа почти в 1,5 раза даже при комнатной температуре. При температурных оптимумах (Топт) активность ThermoLuc превышала активность люциферазы дикого типа приблизительно в 2,4 раза, при 37°С такая разница составляла приблизительно 19 раз (Фиг. 2).
Хотя кинетика биолюминесцентной реакции сильно зависит от состава буфера и других факторов, падение уровня биолюминесцентного сигнала у ThermoLuc происходило значительнее медленнее (Фиг. 3) в сравнении с люциферазой дикого типа MLuc7 во всех исследованных условиях, что еще больше увеличивает разницу в уровне сигнала в пользу ThermoLuc при накоплении сигнала за фиксированный временной промежуток. Такая стабилизация биолюминесцентного сигнала также обеспечивает увеличение чувствительности репортерного белка.
Зависимость люциферазной активности от температуры.
Измерения биолюминесцентной активности приводили при помощи биолюминометра БЛМ-003 с температурной стабилизацией (СКТБ «Наука», Красноярск, РФ) как описано выше, но при различных температурах. Кюветы с измерительным буфером заранее выдерживали при температуре измерения. Концентрация белка в кювете составляла 6 нМ. Как видно из графиков зависимости уровня пиковой биолюминесценции от температуры, температурный оптимум активности мутантной люциферазы по изобретению ThermoLuc оказался сдвинутым с 10-17°С для MLuc7 в область 31-38°С, что является благоприятным для использования ThermoLuc в качестве биолюминесцентного репортера в живых клетках млекопитающих, растущих при 37°С.
Термостабильность. Для исследования термоинактивации люцифераз при разных температурах, аликвоты, содержащие по 100 мкл раствора белка с концентрацией 0,01 мг/мл в буфере для хранения белка (20 мМ Трис-HCl рН 7,5, 0,3 М NaCl, 1 мМ ЭДТА, 0,01% NP-40) инкубировали 1 час при различных температурах. Температурную инкубацию прерывали охлаждением во льду 5 мин и сразу же измеряли активность образцов при 23°С. Термостабильность мутантной люциферазы ThermoLuc не отличалась от высокотермостабильной MLuc7 в диапазоне до 60°С, далее в диапазоне до 100°С после часовой инкубации активность ThermoLuc падала немного быстрее. Но все равно ее термостабильность оставалась очень высокой - даже 1-часовая инкубация при 100°С приводила к потере только около 40% активности ThermoLuc - Фиг. 4.
Пример 4. Применение мутантной люциферазы ThermoLuc как секретируемого биолюминесцентного репортера для мониторинга жизнеспособности клеток человека
Для тестирования мутантной люциферазы ThermoLuc как биолюминесцентного репортера могут быть использованы, например, клетки почки эмбриона человека линии НЕК293, транзиентно или постоянно трансфецированные, например, векторной плазмидой pcDNA3.1 + (Invitrogene), в которую под конститутивный цитомегаловирусный (CMV) промотор вставлен ген Seq. 4, кодирующий аминокислотную последовательность Seq. 1 - полноразмерную мутантную люциферазу ThermoLuc. Плазмида pcDNA3.1 является вектором для экспрессии целевого белка в клетках млекопитающих под сильным конститутивным CMV промотором. В данном случае продукт экспрессии - мутантная люцифераза ThermoLuc будет секретироваться в культуральную среду и ее уровень в среде будет отражать количество клеток НЕК293 и уровень их метаболической активности, поскольку конститутивный промотор обеспечивает постоянный синтез контролируемого белка, уровень которого будет зависеть от физиологического состояния клетки.
Секретируемый репортер позволяет мониторить внутриклеточные события без разрушения клеток путем простого неоднократного отбирания аликвот культуральной среды. Для ThermoLuc размер отбираемых аликвот составлял 1-3 мкл. Полноразмерный ген (Seq. 4), кодирующий мутантную люциферазу ThermoLuc (Seq. 1) с отщепляемым природным сигнальным пептидом для секреции, был полученный в ходе примера 1. Для эукариотической экспрессии ген (Seq. 4), кодирующий полноразмерную мутантную люциферазу ThermoLuc с сигнальным пептидом, был синтезирован в ходе ПЦР со специфическими праймерами, содержащими краевые сайты рестрикции KpnI и XhoI, затем вставлен по соответствующим сайтам в плазмиду pcDNA3.1+.
Трансфекцию плазмиды в клетки человека НЕК293 в достаточном для последующих экспериментов количестве проводили с помощью трансфецирующего агента Lipofectamine 3000 (Thermo Fisher Scientific) согласно рекомендациям производителя. Уровень транзиентной трансфекции в эксперименте составлял около 90% (по визуальной оценке) клеток, трансфецированных контрольной плазмидой, содержащей вместо люциферазы ген зеленого флуоресцентного белка. Далее клетки рассевали в различных разведениях в 96-луночном планшете, инкубировали в СО2-инкубаторе и через 24 ч определяли биолюминесцентную активность среды в отобранной аликвоте (2 мкл) с использованием планшетного люминометра Mithras LB 940 Multimode Reader (Berthold, Германия). В лунки вносили по 100 мкл ML буфера и 2 мкл образца. Сигнал измеряли сразу после впрыска 50 мкл свежеприготовленного субстрата целентеразина (Prolume, США) в том же ML-буфере, хранившегося на льду, в том числе, в процессе измерения. Финальная концентрация целентеразина в лунке была 1 мкМ. Все измерения проводили при комнатной температуре во множественных повторах, для построения кривой использовали усредненные результаты.
Результаты представлены на графике зависимости величины биолюминесцентного сигнала в культуральной среде от количества трансфецированных клеток человека НЕК293 в лунке (Фиг. 5). В среде контрольных клеток, трансфецированных только векторной плазмидой, практически не наблюдалось биолюминесцентной активности. В исследованном диапазоне от 5 до ~22000 клеток, экспрессирующих ThermoLuc, наблюдалась строгая линейная зависимость величины биолюминесцентного сигнала культуральной среды от количества жизнеспособных клеток в лунке (Фиг. 5). Коэффициент линейной корреляции составлял R2=0,998. При этом сигнал в среде от минимального образца с 5 клетками более чем в 5 раз превышал фоновый сигнала от среды нетрансфецированных контрольных клеток и клеток, трансфецированных векторной плазмидой без вставки. В данном эксперименте измерения биолюминесцентной активности репортера ThermoLuc позволили идентифицировать количество живых клеток НЕК293 в линейном диапазоне, перекрывающем 3,5 порядка.
Мутантная люцифераза ThermoLuc является новой биолюминесцентной репортерной молекулой для использования в качестве биолюминесцентного репортера для визуализации молекулярных процессов в живых клетках in vivo, мечения белков и клеток, и в различных люминометрических анализах in vitro, что расширяет арсенал маркерных белков в молекулярной биологии и биотехнологии, а также увеличивает чувствительность измерений.


Изобретение относится к областям молекулярной и клеточной биотехнологии и, в частности, к люциферазам. Мутантная копеподная люцифераза для применения в качестве биолюминесцентного репортера in vitro и in vivo имеет аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:1, 2, 3, и получена путем направленной молекулярной эволюции из MLuc7 изоформы люциферазы Metridia longa, отличается от природной аминокислотной последовательности наличием аминокислотных замен Y72W, I73L, G81E, G90A и D146V с нумерацией по SEQ ID NO:1. Полученная люцифераза характеризуется увеличенной активностью, температурным оптимумом реакции в области 31-38°С и повышенной стабильностью люминесцентного сигнала. 5 ил., 1 табл., 4 пр.
Мутантная копеподная люцифераза для применения в качестве биолюминесцентного репортера in vitro и in vivo, имеющая аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:1, 2, 3, и полученная путем направленной молекулярной эволюции из MLuc7 изоформы люциферазы Metridia longa, отличающаяся от природной аминокислотной последовательности наличием аминокислотных замен Y72W, I73L, G81E, G90A и D146V с нумерацией по SEQ ID NO:1, характеризующаяся увеличенной активностью, температурным оптимумом реакции в области 31-38°С и повышенной стабильностью люминесцентного сигнала.
| RU 2009130108 A, 10.02.2011 | |||
| RU 97113723 A, 20.06.1999 | |||
| УКОРОЧЕННАЯ МУТАНТНАЯ ЛЮЦИФЕРАЗА ИЗ Metridia longa ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ БИОЛЮМИНЕСЦЕНТНОГО РЕПОРТЕРА В ЖИВЫХ КЛЕТКАХ | 2012 |
|
RU2495929C1 |
| WO 2008095622 A2, 14.08.2008. | |||

