Предпосылки создания изобретения
1. Область техники, к которой относится изобретение
Настоящее изобретение относится к микроорганизмам, относящимся к роду Corynebacterium, которые продуцируют 5'-инозиновую кислоту, где экспрессия генов, кодирующих ферменты, связанные с биосинтезом пуринов, повышается более чем в случае их природной экспрессии, а также к способу получения 5'-инозиновой кислоты, включающему культивирование указанного микроорганизма из рода Corynebacterium с повышенной продукцией 5'-инозиновой кислоты.
2. Описание уровня техники
Одно из нуклеотидных соединений, 5'-инозиновая кислота, представляющее собой промежуточное соединение в метаболическом пути биосинтеза нуклеотидов, используется во многих областях, относящихся, в частности, к пищевым продуктам, медицинским препаратам и различным другим направлениям медицинского применения, и играет важную роль в физиологии животных и растений. В частности, 5'-инозиновая кислота используется как приправа нуклеотидной природы, которая привлекла большое внимание в качестве острой приправы, в связи с ее синергическим действием при совместном использовании с глютаматом мононатрия (МНГ (MSG)).
Известные к настоящему времени способы получения 5'-инозиновой кислоты включают способ ферментативного разложения рибонуклеиновой кислоты, экстрагированной из дрожжевых клеток (опубликованная рассмотренная заявка на патент Японии № 1614/1957), способ химического фосфорилирования инозина, получаемого путем ферментации (Agric. Biol. Chem., 36, 1511 (1972) и т.п.), и способ культивирования микроорганизма, способного к продукции 5'-инозиновой кислоты, с последующим восстановлением инозинмонофосфата (ИМФ (IMF)), накапливающегося в среде. В настоящее время способы получения 5'-инозиновой кислоты с помощью микроорганизмов применяются чаще других. Штаммы рода Corynebacterium широко использовались в качестве микроорганизма, продуцирующего 5'-инозиновую кислоту, и описан, например, способ получения 5'-инозиновой кислоты при культвивировании Corynebacterium ammoniagenes (публикация патента Кореи № 2003-0042972).
Для повышения выхода продуцируемой 5'-инозиновой кислоты микроорганизмом были проведены исследования, направленные на разработку штаммов за счет повышения или снижения активности или экспрессии ферментов, вовлекаемых в биосинтез или разложение 5'-инозиновой кислоты. В патенте Кореи № 785248 описывается микроорганизм, в котором ген purC, кодирующий фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, подвергается сверхэкспрессии в пути биосинтеза пуринов, и описывается также способ получения 5'-инозиновой кислоты с его использованием. Кроме того, в патенте Кореи № 857379 описывается штамм Corynebacterium ammoniagenes, в котором кодируемая геном purKE фосфорибозиламиноимидазол-карбоксилаза подвергается сверхэкпрессии, и описывается способ получения высоко концентрированного ИМФ с высоким выходом при использовании указанного штамма.
Однако все еще остается потребность в разработке штамма, способного продуцировать более высокие количества 5'-инозиновой кислоты, и, соответственно, в способе получения 5'-инозиновой кислоты с его использованием.
В этой связи, авторы настоящего изобретения провели исследования в направлении разработки штамма, способного к образованию 5'-инозиновой кислоты с высокой продуктивностью. В результате, они обнаружили, что продукция 5'-инозиновой кислоты может быть повышена при одновременном повышении активности основных ферментов, вовлекаемых в путь биосинтеза пуринов, до уровня, превосходящего их природную активность, что и составило суть настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Одним объектом настоящего изобретения является получение микроорганизма из рода Corynebacterium, обладающего улучшенной продуктивностью в отношении 5'-инозиновой кислоты.
Другим объектом настоящего изобретения является разработка способа получения 5'-инозиновой кислоты с использованием микроорганизма из рода Corynebacterium, обладающего повышенной продуктивностью в отношении 5'-инозиновой кислоты.
КРАТКОЕ ОПИСАНИЕ ФИГУР
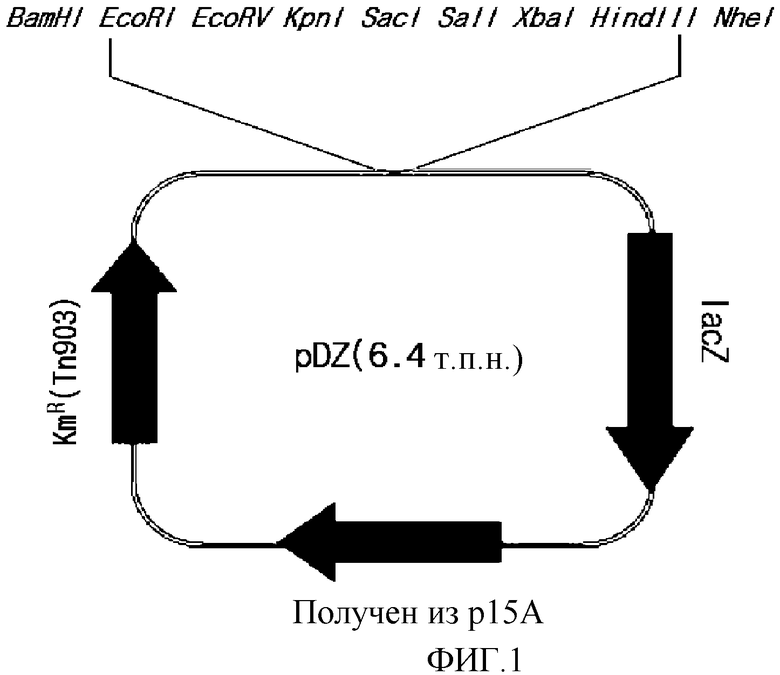
на фиг. 1 показан вектор pDZ для вставки в хромосому микроорганизма из рода Corynebacterium;
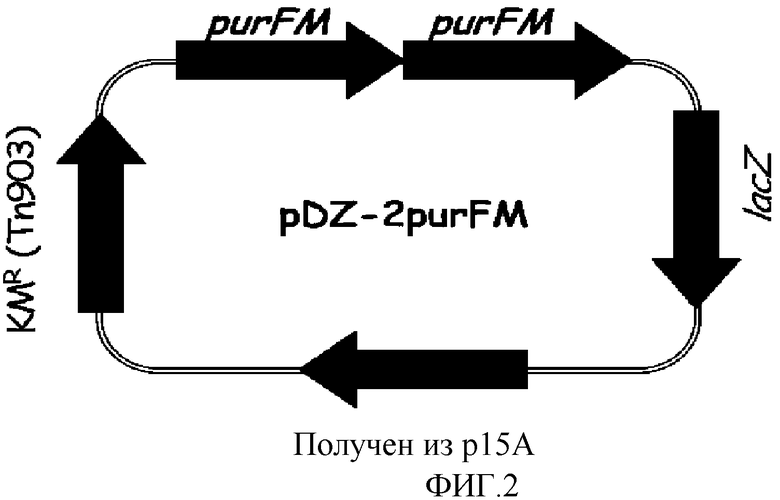
на фиг. 2 показан вектор pDZ-2purFM для вставки в хромосому микроорганизма из рода Corynebacterium;
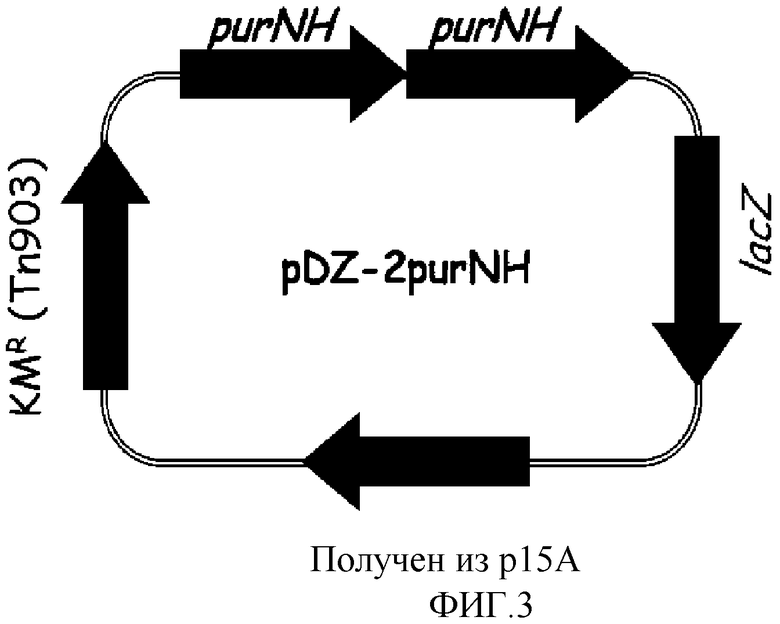
на фиг. 3 показан вектор pDZ-2purNH для вставки в хромосому микроорганизма из рода Corynebacterium;
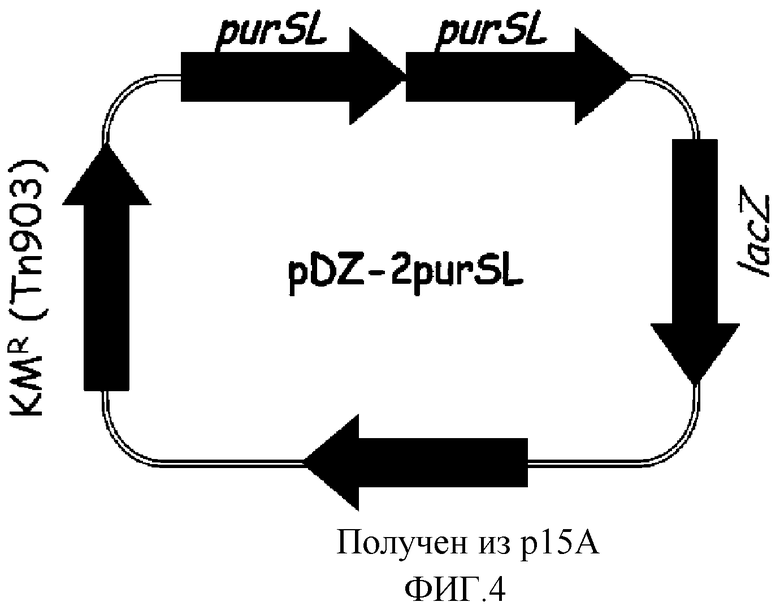
на фиг. 4 показан вектор pDZ-2purSL для вставки в хромосому микроорганизма из рода Corynebacterium;
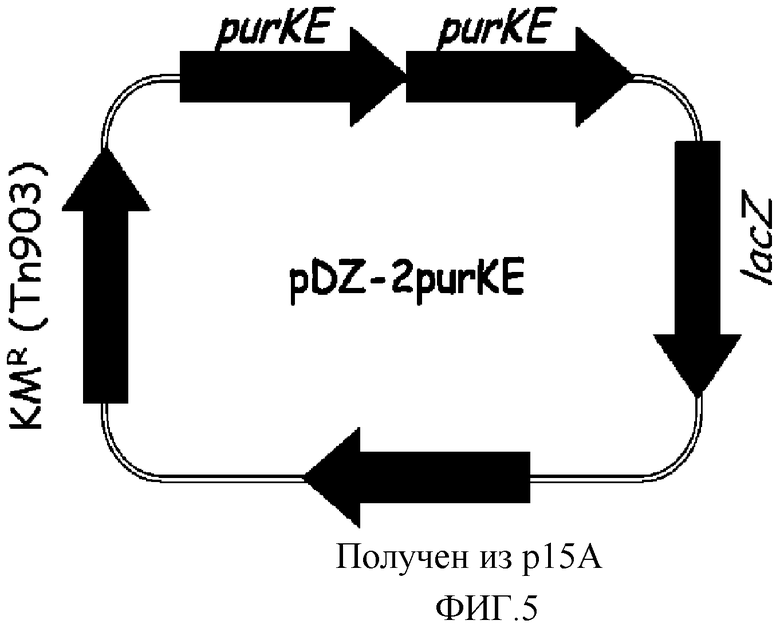
на фиг. 5 показан вектор pDZ-2purKE для вставки в хромосому микроорганизма из рода Corynebacterium;
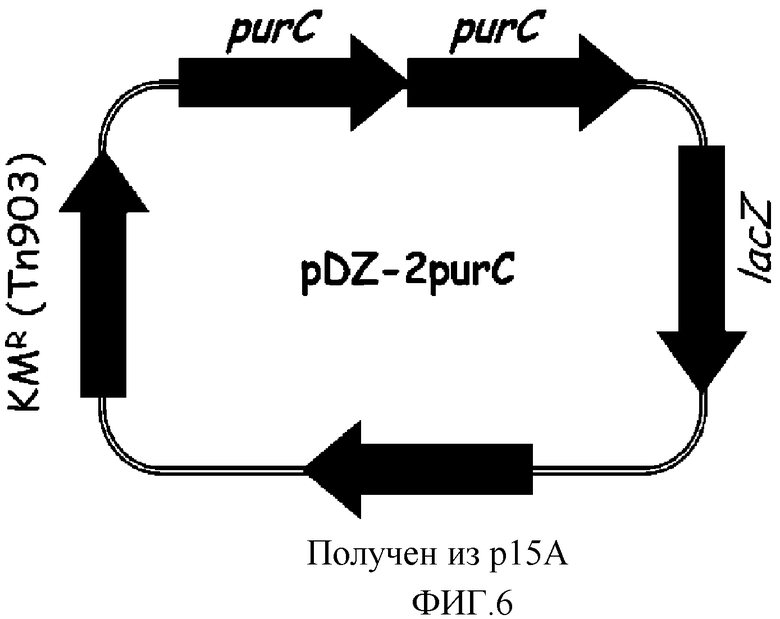
на фиг. 6 показан вектор pDZ-2purC для вставки в хромосому микроорганизма из рода Corynebacterium;
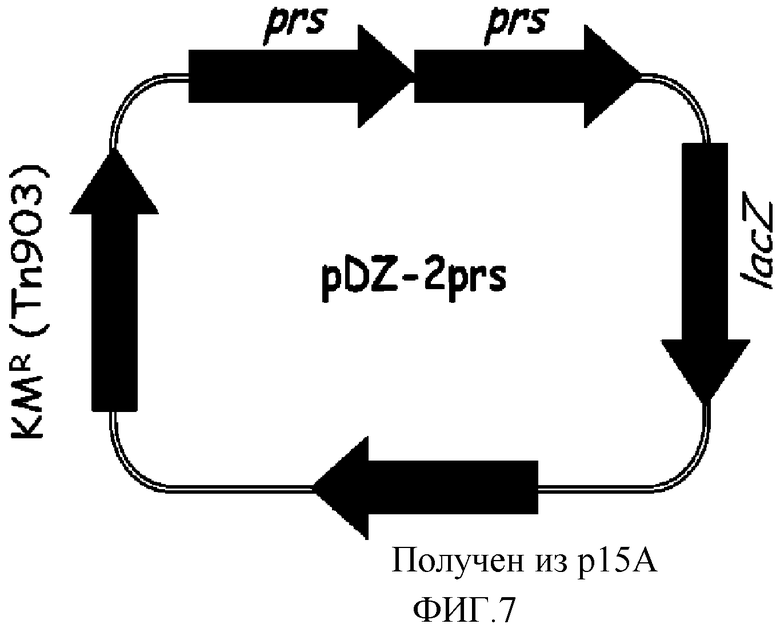
на фиг. 7 показан вектор pDZ-2prs для вставки в хромосому микроорганизма из рода Corynebacterium.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Соответственно, настоящее изобретение относится к микроорганизму, относящемуся к роду Corynebacterium, который продуцирует 5'-инозиновую кислоту, где экспрессия генов, кодирующих ферменты пуринового биосинтеза, повышается больше, чем в случае их природной экспрессии, что позволяет решить указанные выше задачи.
Микроорганизм рода Corynebacterium по настоящему изобретению характеризуется более высокой продукцией 5'-инозиновой кислоты, которая превышает способность родительского штамма, поскольку экспрессия генов, кодирующих ферменты пуринового биосинтеза, повышается больше, чем в случае их экспрессии на уровне, соответствующем природному состоянию.
В контексте настоящего описания термин «фермент биосинтеза пуринов» обозначает фермент, который катализирует реакцию, вовлекаемую в путь биосинтеза пуринов с образованием пуринового основания в качестве конечного продукта, и включает фосфорибозилпирофосфат-амидотрансферазу, фосфорибозилглицинамид-формилтрансферазу, фосфорибозилформилглицинамидин-синтетазу, фосфорибозилформилглицинамидин-синтетазу II, фосфорибозиламиноимидазол-синтетазу, фосфорибозиламиноимидазол-карбоксилазу, фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, циклогидролазу инозиновой кислоты, рибозофосфат-пирофосфокиназу или другие.
В конкретном варианте осуществления настоящего изобретения, ферменты биосинтеза пуринов могут представлять собой сочетание с одним другим или несколькими ферментами, выбранными из группы, состоящей из фосфорибозилпирофосфат-амидотрансферазы, фосфорибозилглицинамид-формилтрансферазы, фосфорибозилформилглицинамидин-синтетазы, фосфорибозилформилглицинамидин-синтетазы II, фосфорибозиламиноимидазол-синтетазы, фосфорибозиламиноимидазол-карбоксилазы, фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазы, циклогидролазы инозиновой кислоты или рибозофосфат-пирофосфокиназы.
В конкретном варианте осуществления настоящего изобретения, ген, кодирующий ферменты биосинтеза пуринов, экспрессия которых повышается до уровня, превосходящего их природную экспрессию, представляет собой сочетание гена purN с последовательностью SEQ ID NO: 36, который кодирует фосфорибозилглицинамид-формилтрансферазу, гена purS с последовательностью SEQ ID NO: 37, который кодирует фосфорибозилформилглицинамидин-синтетазу, гена purL с последовательностью SEQ ID NO: 38, который кодирует фосфорибозилформилглицинамидин-синтетазу II, гена purKE с последовательностью SEQ ID NO: 40, который кодирует фосфорибозиламиноимидазол-карбоксилазу, гена purC с последовательностью SEQ ID NO: 41, который кодирует фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, гена purH с последовательностью SEQ ID NO: 42, который кодирует циклогидролазу инозиновой кислоты, и гена prs с последовательностью SEQ ID NO: 43, который кодирует рибозофосфат-пирофосфокиназу.
В конкретном варианте осуществления настоящего изобретения, ген, кодирующий ферменты биосинтеза пуринов, экспрессия которых повышается до уровня, превосходящего их природную экспрессию, представляет собой сочетание гена purF с последовательностью SEQ ID NO: 35, который кодирует фосфорибозилпирофосфат-амидотрансферазу, гена purN с последовательностью SEQ ID NO: 36, который кодирует фосфорибозилглицинамид-формилтрансферазу, гена purS с последовательностью SEQ ID NO: 37, который кодирует фосфорибозилформилглицинамидин-синтетазу, гена purL с последовательностью SEQ ID NO: 38, который кодирует фосфорибозилформилглицинамидин-синтетазу II, гена purM с последовательностью SEQ ID NO: 39, который кодирует фосфорибозиламиноимидазол-синтетазу, гена purKE с последовательностью SEQ ID NO: 40, который кодирует фосфорибозиламиноимидазол-карбоксилазу, гена purC с последовательностью SEQ ID NO: 41, который кодирует фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, гена purH с последовательностью SEQ ID NO: 42, который кодирует циклогидролазу инозиновой кислоты, и гена prs с последовательностью SEQ ID NO: 43, который кодирует рибозофосфат-пирофосфокиназу.
В контексте настоящего описания, используемый применительно к экспрессии термин «повышенный до уровня, превосходящего природный» означает, что уровень экспрессии гена выше, чем при естественной экспрессии в микроорганизме, или выше, чем в варианте его экспрессии в родительском штамме, и включает повышение количества (числа копий) генов, кодирующих соответствующие ферменты, и, соответственно, повышение уровня экспрессии, или повышение уровня экспрессии за счет мутации, или же повышение уровня экспрессии как результат действия обоих этих механизмов.
В конкретном варианте осуществления настоящего изобретения, повышение уровня экспрессии гена, кодирующего фермент биосинтеза пуринов, включает повышение количества копий гена путем дополнительного введения в штамм соответствующего чужеродного гена или путем амплификации естественного гена, или повышение эффективности транскрипции или эффективности трансляции путем мутации последовательности, регулирующей транскрипцию или трансляцию, но не ограничивается приведенным перечнем. Амплификация естественного гена может быть легко осуществлена по известному в данной области методу, например, путем культивирования в условиях подходящего селекционного давления.
В конкретном варианте осуществления настоящего изобретения, уровень экспрессии гена, кодирующего фермент биосинтеза пуринов, может быть повышен за счет дополнительного введения в клетку гена, кодирующего фермент биосинтеза пуринов, или путем амплификации естественного гена, кодирующего фермент биосинтеза пуринов.
В конкретном варианте осуществления настоящего изобретения, ген, кодирующий ферменты биосинтеза пуринов, экспрессия которых повышается до уровня, превосходящего их природную экспрессию, может существовать в виде двух или более копий в микроорганизме рода Corynebacterium, обладающего повышенной продуктивной способностью в отношении 5'-инозиновой кислоты, за счет введения в клетку одной или нескольких копий гена, в дополнение к уже имеющемуся гену.
В конкретном варианте осуществления настоящего изобретения, ген, кодирующий ферменты биосинтеза пуринов, вводят в микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, посредством трансформации с использованием рекомбинантного вектора, содержащего две копии соответствующего гена, который последовательно организованы.
В конкретном варианте осуществления настоящего изобретения, рекомбинантный вектор, используемый для получения микроорганизма рода Corynebacterium, обладающего повышенной способностью к продукции 5'-инозиновой кислоты, может быть выбран из группы, состоящей из рекомбинантных векторов pDZ-2purFM, pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC и pDZ-2prs, которые характеризуются картами расщепления, показанными на фиг. 2-7, соответственно, в зависимости от природы введенного гена.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, может быть выбран из микроорганизмов рода Corynebacterium, способных к повышенной продукции 5'-инозиновой кислоты. Например, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты по настоящему изобретению, может быть получен из Corynebacterium ammoniagenes ATCC6872, Corynebacterium thermoaminogenes FERM BP-1539, Corynebacterium glutamicum ATCC13032, Brevibacterium flavum ATCC14067, Brevibacterium lactofermentum ATCC13869 и штаммы, полученные на их основе.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, может включать две или более копии гена, кодирующего фермент биосинтеза пуринов.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, может представлять собой Corynebacterium ammoniagenes, и более предпочтительно, Corynebacterium ammoniagenes, в котором активность сочетания prs гена и одного или нескольких генов, выбранных из группы, состоящей из purF, purN, purS, purL, purM, purKE, purC и purH, повышена и приводит к образованию высокой концентрации 5'-инозиновой кислоты.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, может представлять собой штамм, где указанный продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 (KCCM-10610) содержит введенные, в последовательном порядке или в сочетании, каждый из рекомбинантных векторов pDZ-2purFM, pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC и pDZ-2prs, характеризующихся сайтами расщепления, показанными на фиг. 2, 3, 4, 5, 6 и 7, и одна из двух копии введенных генов purF, purN, purS, purL, purM, purKE, purC, purH и prs замещают естественные гены в результате проведенной гомологичной рекомбинации, так что по две копии генов purF, purN, purS, purL, purM, purKE, purC и purH вводятся в данный штамм.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, может представлять собой Corynebacterium ammoniagenes, содержащий две копии генов, кодирующих ферменты биосинтеза пуринов, которые представляют собой сочетание гена purN с последовательностью SEQ ID NO: 36, который кодирует фосфорибозилглицинамид-формилтрансферазу, гена purS с последовательностью SEQ ID NO: 37, который кодирует фосфорибозилформилглицинамидин-синтетазу, гена purL с последовательностью SEQ ID NO: 38, который кодирует фосфорибозилформилглицинамидин-синтетазу II, гена purKE с последовательностью SEQ ID NO: 40, который кодирует фосфорибозиламиноимидазол-карбоксилазу, гена purC с последовательностью SEQ ID NO: 41, который кодирует фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, гена purH с последовательностью SEQ ID NO: 42, который кодирует циклогидролазу инозиновой кислоты, и гена prs с последовательностью SEQ ID NO: 43, который кодирует рибозофосфат-пирофосфокиназу, и предпочтительно представляет собой Corynebacterium ammoniagenes CN01-0120.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, обладающий повышенной способностью к продукции 5'-инозиновой кислоты, может представлять собой Corynebacterium ammoniagenes, содержащий две копии генов, кодирующих ферменты биосинтеза пуринов, которые представляют собой сочетание гена purF с последовательностью SEQ ID NO: 35, который кодирует фосфорибозилпирофосфат-амидотрансферазу, гена purN с последовательностью SEQ ID NO: 36, который кодирует фосфорибозилглицинамид-формилтрансферазу, гена purS с последовательностью SEQ ID NO: 37, который кодирует фосфорибозилформилглицинамидин-синтетазу, гена purL с последовательностью SEQ ID NO: 38, который кодирует фосфорибозилформилглицинамидин-синтетазу II, гена purM с последовательностью SEQ ID NO: 39, который кодирует фосфорибозиламиноимидазол-синтетазу, гена purKE с последовательностью SEQ ID NO: 40, который кодирует фосфорибозиламиноимидазол-карбоксилазу, гена purC с последовательностью SEQ ID NO: 41, который кодирует фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, гена purH с последовательностью SEQ ID NO: 42, который кодирует циклогидролазу инозиновой кислоты, и гена prs с последовательностью SEQ ID NO: 43, который кодирует рибозофосфат-пирофосфокиназу, и предпочтительно представляет собой Corynebacterium ammoniagenes CN01-0316 (КССМ 10992Р).
Кроме того, настоящее изобретение относится к способу получения 5'-инозиновой кислоты, включающему стадии культивирования микроорганизма рода Corynebacterium, продуцирующего 5'-инозиновую кислоту, в котором экспрессия гена, кодирующего фермент биосинтеза пуринов, повышена относительно естественного уровня его экспрессии, и затем выделения 5'-инозиновой кислоты из культуральной среды.
В способе получения 5'-инозиновой кислоты по настоящему изобретению, среда и другие условия, используемые при культивировании микроорганизма рода Corynebacterium, могут быть такими же, как это обычно применяется при культивировании микроорганизма рода Corynebacterium, и любой специалист в данной области может легко подобрать и соответствующим образом их откорректировать. Кроме того, указанное культивирование может проводиться по любому известному в данной области способу культивирования, такому как, например, периодическое культивирование, непрерывное культивирование и периодическое культивирование с добавлением субстрата, но не ограничивается указанными способами.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, продуцирующий 5'-инозиновую кислоту, может представлять собой Corynebacterium ammoniagenes.
В конкретном варианте осуществления настоящего изобретения, микроорганизм рода Corynebacterium, продуцирующий 5'-инозиновую кислоту, может представлять собой Corynebacterium ammoniagenes CN01-0120 или Corynebacterium ammoniagenes CN01-0316 (KCCM 10992P).
В конкретном варианте осуществления настоящего изобретения, культивирование микроорганизма рода Corynebacterium проводят в стандартной среде, содержащей подходящие источники углерода, источники азота, аминокислоты, витамины или т.п., в аэробных условиях, при соответствующих значениях температуры, рН или т.п.
В качестве источника углерода могут использоваться углеводы, такие как глюкоза и фруктоза. В качестве источника азота могут использоваться различные неорганические источники азота, такие как аммиак, хлорид аммония и сульфат аммония, а также органические источники азота, такие как пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, кукурузный экстракт, гидролизат казеина, рыбное сырье или рыбная кормовая мука, а также обезжиренный соевый жмых или обезжиренная соевая мука. Примеры неорганических соединений включают моногидрофосфат калия, дигидрофосфат калия, сульфат магния, сульфат железа, сульфат марганца и карбонат кальция. При необходимости, могут использоваться витамины и ауксотрофные основания.
Культивирование проводят в аэробных условиях, например при встряхивании культуры или при перемешивании культуры, предпочтительно при температуре от 28°С до 36°С. В ходе культивирования рН предпочтительно поддерживают в диапазоне значений от 6 до 8. Культивирование может проводиться в течение периода времени, составляющего от 4 до 6 дней.
Ниже, настоящее изобретение описывается более подробно с использованием примеров. Однако следует понимать, что указанные примеры даны лишь с целью пояснения и никоим образом не ограничивают настоящее изобретение.
Пример 1. Встраивание генов, кодирующих ферменты биосинтеза пуринов, с использованием вектора (pDZ) для встраивания в хромосому и разработка на этой основе штамма, продуцирующего высокие количества 5'-инозиновой кислоты.
Для встраивания чужеродного гена в хромосому штамма Corynebacterium ammoniagenes использовали рекомбинантный вектор на основе pDZ, содержащий две последовательно организованные копии соответствующего гена. Вектор pDZ представляет собой вектор, который используют для встраивания в хромосому микроорганизма рода Corynebacterium и который был получен по способу, описанному в публикации по патенту Кореи № 2008-0025355, включенному в настоящее описание в качестве ссылки. На фиг. 1 приведена схематическая диаграмма, показывающая структуру pDZ вектора.
В приведенных ниже пунктах (1)-(6) описывается получение рекомбинантного вектора, где функция рекомбинантного вектора состоит во встраивании гена, кодирующего фермент биосинтеза пуринов, в хромосому микроорганизма рода Corynebacterium с получением двух копий каждого гена. Трансформация каждым рекомбинантным вектором и селекция трансформантов проводятся по описанной ниже процедуре.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 (KCCM-10610) трансформировали по методу электропорации рекомбинантным вектором pDZ, содержащим желательный ген, кодирующий фермент биосинтеза пуринов, и затем штаммы, в которых ген, перенесенный вектором, встраивался в их хромосому в результате гомологичной рекомбинации, отбирали на селективной среде, содержащей 25 мг/л канамицина. Встраивание вектора в хромосому оценивали при осмотре цвета колоний на твердой среде (1% мясной экстракт, 1% дрожжевой экстракт, 1% пептон, 0,25% хлорид натрия, 1% аденин, 1% гуанин, 1,5% агароза), содержащей X-gal (5-бром-4-хлор-3-индолил-β-D-галактозид). Так, при этом шел отбор синих колоний в качестве трансформантов, где вектор был встроен в хромосому. Штамм, в котором вектор был встроен в хромосому микроорганизма за счет первого кроссинговера, культивировали на качалке (30°С, 8 часов) в питательной среде (1% глюкоза, 1% мясной экстракт, 1% дрожжевой экстракт, 1% пептон, 0,25% хлорид натрия, 1% аденин, 1% гуанин). Затем полученный при культивировании штамм подвергали серийному разведению от 10-4 до 10-10 и разбавленную культуру вносили на твердую среду, содержащую X-gal. Большая часть колоний имела синий цвет, хотя в небольшом количестве присутствовали и белые колонии. При селекции белых колоний были отобраны штаммы, в которых последовательность вектора была удалена из хромосомы в результате второго кроссинговера. Выбранный штамм был идентифицирован как искомый, по результатам теста на чувствительность к канамицину и данным ПЦР-анализа генной последовательности.
(1) Клонирование purFM гена и конструирование рекомбинантного вектора (pDZ-2purFM)
Гены purF и purM расположены близко друг к другу в хромосоме микроорганизма рода Corynebacterium, благодаря этому был сконструирован вектор purFM, содержащий оба этих гена и промоторный участок, для целей одновременной экспрессии обоих указанных генов.
Хромосому выделяли из штамма Corynebacterium ammoniagenes CJIP2401, продуцирующего 5'-инозиновую кислоту, и затем была проведена полимеразная цепная реакция (ПЦР) с использованием указанной хромосомы в качестве матрицы для получения purFM, а именно: фрагмента, содержащего последовательно объединенные гены purF и purM. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltraTM (PfuUltraTM High-Fidelity (Stratagene)), и полимеразную цепную реакцию проводили путем денатурации в течение 30 циклов при температуре 96°С в течение 30 секунд, с последующими отжигом при температуре 53°С в течение 30 секунд и полимеризацией при температуре 72°С в течение 2 минут. В результате, получали два purFM гена, содержащих промоторный участок (purFM-A, purFM-B). Далее, purFM-A амплифицировали, используя праймеры с последовательностями SEQ ID NO: 1 и 2, и purFM-B амплифицировали, используя праймеры с последовательностями SEQ ID NO: 3 и 4. Продукты амплификации клонировали в векторе E.coli pCR2.1 с использованием набора для клонирования Topo (Topo Cloning Kit (Invitrogen)) с тем, чтобы получить векторы pCR-purFM-A и pCR-purFM-B соответственно. Векторы pCR обрабатывали рестриктазами, содержащимися на каждом конце purFM-A и purFM-B (purFM-A: EcoRI+XbaI, purFM-B: XbaI+HindIII), и каждый purFM ген отделяли от pCR векторов. После этого вектор pDZ обрабатывали рестриктазами EcoRI и HindIII и клонировали проводя 3-компонентное лигирование с тем, чтобы в итоге получить конструкцию рекомбинантного вектора pDZ-2purFM, в котором оба purFM гена будут последовательно объединены. На фиг.2 показан вектор pDZ-2purFM, используемый для встраивания в хромосому Corynebacterium.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 трансформировали вектором pDZ-2purFM путем электропорации, и один ген purFM дополнительно встраивали рядом с естественным геном purFM на хромосоме через второй кроссинговер, так что в итоге был получен штамм, содержащий уже обе копии. Указанные последовательно встроенные гены purFM были идентифицированы по процедуре ПЦР с использованием праймеров SEQ ID NO: 5 и 6, которые позволяют амплифицировать участки, охватывающие два соединенных purFM гена.
(2) Клонирование purNH гена и конструирование рекомбинантного вектора (pDZ-2purNH), получение purNH-содержащего штамма.
Гены purN и purH расположены близко друг к другу в хромосоме микроорганизма рода Corynebacterium, благодаря этому был сконструирован вектор purNH, содержащий промоторный участок, для целей одновременной экспрессии обоих указанных генов.
Хромосому выделяли из штамма Corynebacterium ammoniagenes CJIP2401, продуцирующего 5'-инозиновую кислоту, и затем была проведена полимеразная цепная реакция (ПЦР) с использованием указанной хромосомы в качестве матрицы для получения purNH, а именно: фрагмента, содержащего последовательно объединенные гены purN и purH. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltraTM (PfuUltraTM High-Fidelity (Stratagene)) и полимеразную цепную реакцию проводили путем денатурации в течение 30 циклов при температуре 96°С в течение 30 секунд, с последующими отжигом при температуре 53°С в течение 30 секунд и полимеризацией при температуре 72°С в течение 2 минут. В результате, получали два purNH гена, содержащих промоторный участок (purNH-A, purNH-B). Далее, purNH-A амплифицировали, используя праймеры с последовательностями SEQ ID NO: 7 и 8, и purNH-B амплифицировали, используя праймеры с последовательностями SEQ ID NO: 8 и 9. Продукты амплификации клонировали в векторе E.coli pCR2.1 с использованием набора для клонирования Topo (Topo Cloning Kit (Invitrogen)) с тем, чтобы получить векторы pCR-purNH-A и pCR-purNH-B соответственно. Векторы pCR обрабатывали рестриктазами, содержащимися на каждом конце purNH-A и purNH-B (purNH-A: BamHI+SalI, purNH-B: SalI), и каждый purNH ген отделяли от pCR векторов. После этого вектор pDZ обрабатывали рестриктазами BamHI и SalI и клонировали проводя 3-компонентное лигирование с тем, чтобы в итоге получить конструкцию рекомбинантного вектора pDZ-2purNH, в котором оба purNH гена будут последовательно объединены. На фиг. 3 показан вектор pDZ-2purNH, используемый для встраивания в хромосому Corynebacterium.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 трансформировали вектором pDZ-2purNH путем электропорации, и один ген purNH дополнительно встраивали рядом с естественным геном purNH на хромосоме через второй кроссинговер так, что в итоге был получен штамм, содержащий уже обе копии. Указанные последовательно встроенные гены purNH были идентифицированы по процедуре ПЦР с использованием праймеров SEQ ID NO: 10 и 11, которые позволяют амплифицировать участки, охватывающие два соединенных purNH гена.
(3) Клонирование purSL гена и конструирование рекомбинантного вектора (pDZ-2purSL), получение purSL-содержащего штамма.
Гены purS и purL расположены близко друг к другу в хромосоме микроорганизма рода Corynebacterium, благодаря этому был сконструирован вектор purSL, содержащий промоторный участок, для целей одновременной экспрессии обоих указанных генов.
Хромосому выделяли из штамма Corynebacterium ammoniagenes CJIP2401, продуцирующего 5'-инозиновую кислоту, и затем была проведена полимеразная цепная реакция (ПЦР) с использованием указанной хромосомы в качестве матрицы для получения purSL, а именно фрагмента, содержащего последовательно объединенные гены purS и purL. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltraTM (PfuUltraTM High-Fidelity (Stratagene)), и полимеразную цепную реакцию проводили путем денатурации в течение 30 циклов при температуре 96°С в течение 30 секунд, с последующими отжигом при температуре 53°С в течение 30 секунд и полимеризацией при температуре 72°С в течение 2 минут. В результате получали два purSL гена, содержащих промоторный участок (purSL-A, purSL-B). Далее, purSL-A амплифицировали, используя праймеры с последовательностями SEQ ID NO: 12 и 13, и purSL-B амплифицировали, используя праймеры с последовательностями SEQ ID NO: 14 и 15. Продукты амплификации клонировали в векторе E.coli pCR2.1 с использованием набора для клонирования Topo (Topo Cloning Kit (Invitrogen)) с тем, чтобы получить векторы pCR-purSL-A и pCR-purSL-B соответственно. Векторы pCR обрабатывали рестриктазами, содержащимися на каждом конце purSL-A и purSL-B (purSL-A: BamHI+SalI, purSL-B: SalI+ BamHI), и каждый purSL ген отделяли от pCR векторов. После этого вектор pDZ обрабатывали рестриктазой BamHI и клонировали проводя 3-компонентное лигирование с тем, чтобы в итоге получить конструкцию рекомбинантного вектора pDZ-2purSL, в котором оба purSL гена будут последовательно объединены. На фиг. 4 показан вектор pDZ-2purSL, используемый для встраивания в хромосому Corynebacterium.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 трансформировали вектором pDZ-2purSL путем электропорации, и один ген purSL дополнительно встраивали рядом с естественным геном purSL на хромосоме через второй кроссинговер так, что в итоге был получен штамм, содержащий уже обе копии. Указанные последовательно встроенные гены purSL были идентифицированы по процедуре ПЦР с использованием праймеров SEQ ID NO: 16 и 17, которые позволяют амплифицировать участки, охватывающие два соединенных purSL гена.
(4) Клонирование purKE гена и конструирование рекомбинантного вектора (pDZ-2purKE), получение purKE-содержащего штамма.
Гены purK и purE расположены близко друг к другу в хромосоме микроорганизма рода Corynebacterium, благодаря этому был сконструирован вектор purKE, содержащий промоторный участок, для целей одновременной экспрессии обоих указанных генов.
Хромосому выделяли из штамма Corynebacterium ammoniagenes CJIP2401, продуцирующего 5'-инозиновую кислоту, и затем была проведена полимеразная цепная реакция (ПЦР) с использованием указанной хромосомы в качестве матрицы для получения purKE, а именно фрагмента, содержащего последовательно объединенные гены purK и purE. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltraTM (PfuUltraTM High-Fidelity (Stratagene)), и полимеразную цепную реакцию проводили путем денатурации в течение 30 циклов при температуре 96°С в течение 30 секунд, с последующими отжигом при температуре 53°С в течение 30 секунд и полимеризацией при температуре 72°С в течение 2 минут. В результате получали два purKE гена, содержащих промоторный участок (purKE-A, purKE-B). Далее, purKE-A амплифицировали, используя праймеры с последовательностями SEQ ID NO: 18 и 19, и purKE-B амплифицировали, используя праймеры с последовательностями SEQ ID NO: 20 и 21. Продукты амплификации клонировали в векторе E.coli pCR2.1 с использованием набора для клонирования Topo (Topo Cloning Kit (Invitrogen)) с тем, чтобы получить векторы pCR-purKE-A и pCR-purKE-B соответственно. Векторы pCR обрабатывали рестриктазами, содержащимися на каждом конце purKE-A и purKE-B (purKE-A: BamHI+KpnI, purKE-B: KpnI+XbaI), и каждый purKE ген отделяли от pCR векторов. После этого вектор pDZ обрабатывали рестриктазами BamHI и XbaI и клонировали проводя 3-компонентное лигирование с тем, чтобы в итоге получить конструкцию рекомбинантного вектора pDZ-2purKE, в котором оба purKE гена будут последовательно объединены. На фиг. 5 показан вектор pDZ-2purKE, используемый для встраивания в хромосому Corynebacterium.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 трансформировали вектором pDZ-2purKE путем электропорации, и один ген purKE дополнительно встраивали рядом с естественным геном purKE на хромосоме через второй кроссинговер так, что в итоге был получен штамм, содержащий уже обе копии. Указанные последовательно встроенные гены purKE были идентифицированы по процедуре ПЦР с использованием праймеров SEQ ID NO: 22 и 23, которые позволяют амплифицировать участки, охватывающие два соединенных purKE гена.
(5) Клонирование purC гена и конструирование рекомбинантного вектора (pDZ-2purC), получение purC-содержащего штамма.
Хромосому выделяли из штамма Corynebacterium ammoniagenes CJIP2401 и проводили полимеразную цепную реакцию (ПЦР) с использованием указанной хромосомы в качестве матрицы для получения purC. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltraTM (PfuUltraTM High-Fidelity (Stratagene)) и полимеразную цепную реакцию проводили путем денатурации в течение 30 циклов при температуре 96°С в течение 30 секунд, с последующими отжигом при температуре 53°С в течение 30 секунд и полимеризацией при температуре 72°С в течение 2 минут. В результате, получали два purC гена, содержащих промоторный участок (purC-A, purC-B). Далее, purC-A амплифицировали, используя праймеры с последовательностями SEQ ID NO: 24 и 25, и purC-B амплифицировали, используя праймеры с последовательностями SEQ ID NO: 25 и 26. Продукты амплификации клонировали в векторе E.coli pCR2.1 с использованием набора для клонирования Topo (Topo Cloning Kit (Invitrogen)) с тем, чтобы получить векторы pCR-purC-A и pCR-purC-B соответственно. Векторы pCR обрабатывали рестриктазами, содержащимися на каждом конце purC-A и purC-B (purC-A: BamHI+SalI, purC-B: SalI), и каждый purC ген отделяли от pCR векторов. После этого вектор pDZ обрабатывали рестриктазами BamHI и SalI и клонировали проводя 3-компонентное лигирование с тем, чтобы в итоге получить конструкцию рекомбинантного вектора pDZ-2purC, в котором оба purC гена будут последовательно объединены. На фиг. 6 показан вектор pDZ-2purC, используемый для встраивания в хромосому Corynebacterium.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 трансформировали вектором pDZ-2purC путем электропорации, и один ген purC дополнительно встраивали рядом с естественным геном purC на хромосоме через второй кроссинговер так, что в итоге был получен штамм, содержащий уже обе копии. Указанные последовательно встроенные гены purC были идентифицированы по процедуре ПЦР с использованием праймеров SEQ ID NO: 27 и 28, которые позволяют амплифицировать участки, охватывающие два соединенных purC гена.
(6) Клонирование prs гена и конструирование рекомбинантного вектора (pDZ-2prs), получение prs-содержащего штамма.
Хромосому выделяли из штамма Corynebacterium ammoniagenes CJIP2401 и проводили полимеразную цепную реакцию (ПЦР) с использованием указанной хромосомы в качестве матрицы для получения prs. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltraTM (PfuUltraTM High-Fidelity (Stratagene)), и полимеразную цепную реакцию проводили путем денатурации в течение 30 циклов при температуре 96°С в течение 30 секунд, с последующими отжигом при температуре 53°С в течение 30 секунд и полимеризацией при температуре 72°С в течение 2 минут. В результате получали два prs гена, содержащих промоторный участок (prs-A, prs-B). Далее, prs-A амплифицировали, используя праймеры с последовательностями SEQ ID NO: 29 и 30, и prs-B амплифицировали, используя праймеры с последовательностями SEQ ID NO: 31 и 32. Продукты амплификации клонировали в векторе E.coli pCR2.1 с использованием набора для клонирования Topo (Topo Cloning Kit (Invitrogen)) с тем, чтобы получить векторы pCR-prs-A и pCR-prs-B соответственно. Векторы pCR обрабатывали рестриктазами, содержащимися на каждом конце prs-A и prs-B (prs-A: BamHI+SpeI, prs-B: SpeI+PstI), и каждый prs ген отделяли от pCR векторов. После этого вектор pDZ обрабатывали рестриктазами BamHI и PstI и клонировали проводя 3-компонентное лигирование с тем, чтобы в итоге получить конструкцию рекомбинантного вектора pDZ-2prs, в котором оба prs гена будут последовательно объединены. На фиг. 7 показан вектор pDZ-2prs, используемый для встраивания в хромосому Corynebacterium.
Продуцирующий 5'-инозиновую кислоту штамм Corynebacterium ammoniagenes CJIP2401 трансформировали вектором pDZ-2prs путем электропорации, и один ген prs дополнительно встраивали рядом с естественным геном prs на хромосоме через второй кроссинговер так, что в итоге был получен штамм, содержащий уже обе копии. Указанные последовательно встроенные гены prs были идентифицированы по процедуре ПЦР с использованием праймеров SEQ ID NO: 33 и 34, которые позволяют амплифицировать участки, охватывающие два соединенных prs гена.
(7) Разработка штамма, продуцирующего высокие количества 5'-инозиновой кислоты за счет усиления биосинтеза пуринов.
Сочетания векторов pDZ-2purFM, pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC и pDZ-2prs, сконструированных согласно пп. (1) - (6), вводили в штамм Corynebacterium ammoniagenes CJIP2401, продуцирующий 5'-инозиновую кислоту. Порядок введения векторов соответствовал неорганизованному режиму, а метод введения и идентификация осуществлялись по описанной выше процедуре.
Использовали в качестве родительского штамма Corynebacterium ammoniagenes CJIP2401 и трансформировали его сочетанием векторов pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC и pDZ-2prs и сочетанием векторов pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC, pDZ-2purFM pDZ-2prs с получением Corynebacterium ammoniagenes CN01-0120 (2purNH + 2purSL + 2purKE + 2purC + 2prs) и Corynebacterium ammoniagenes CN01-0316 (2purNH + 2purSL + 2purKE + 2purC + 2purFM + 2prs), которые содержат по две копии генов, кодирующих основные ферменты, участвующие в пути биосинтеза пуринов.
Пример 2. Тест по определению титра при ферментации рекомбинантного Corynebacterium ammoniagenes.
Вносили по 3 посевной среды указанного ниже состава в опытные пробирки диаметром 18 мм и стерилизовали под давлением. Затем инокулировали родительский штамм Corynebacterium ammoniagenes CJIP2401 и полученные согласно процедурам примера 1 штаммы Corynebacterium ammoniagenes CN01-0120 и Corynebacterium ammoniagenes CN01-0316, культивировали на качалке при температуре 30°C в течение 24 часов, для получения посевной культуры. Вносили по 27 мл ферментационной среды указанного ниже состава в эрленмейеровские качалочные колбы по 500 мл и стерилизовали под давлением при температуре 120°C в течение 10 минут, затем инокулировали в них по 3 мл посевной культуры и далее культивировали на качалке в течение 5-6 дней. Культивирование проводили при качании со скоростью 200 об/мин, при температуре 32°C и при pH 7,2.
Посевная среда и ферментационная среда имели следующий состав.
Посевная среда: 1% глюкоза, 1% пептон, 1% мясной экстракт, 1% дрожжевой экстракт, 0,25% хлорид натрия, 100 мг/л аденин, 100 мг/л гуанин, pH 7,2.
Ферментационная среда для колбы: 0,1% глютамат натрия, 1% хлорид аммония, 1,2% сульфат магния, 0,01% хлорид кальция, 20 мг/л сульфат железа, 20 мг/л сульфат марганца, 20 мг/л сульфат цинка, 5 мг/л сульфат меди, 23 мш/л L-цистеин, 24 мг/л аланин, 8 мг/л никотиновая кислота, 45 мкг/л биотин, 5 мг/л тиамина гидрохлорид, 30 мг/л аденин, 1,9% фосфорная кислота (85%), 4,2% глюкоза и 2,4% неочищенный сахар. По завершению культивирования уровень образованной 5'-инозиновой кислоты определяли по методу ВЭЖХ и полученные данные по накоплению 5'-инозиновой кислоты в культуральной среде показаны в приведенной ниже таблице.
(после 5 дней культивирования)
(г/л/час)
(после 5 дней культивирования)
(CJIP2401)
Количество накопленной в культуральной среде 5'-инозиновой кислоты сравнивали с ее количеством в родительском штамме, Corynebacterium ammoniagenes CJIP2401. В результате было показано, что продуктивность штаммов Corynebacterium ammoniagenes CN01-0120 и Corynebacterium ammoniagenes CN01-0316 по 5'-инозиновой кислоте в час возрастала на 10,9-11,4% в одних и тех же условиях, в сравнении с родительским штаммом Corynebacterium ammoniagenes CJIP2401.
Штамм Corynebacterium ammoniagenes CN01-0316, обладающий повышенной продуктивностью по 5'-инозиновой кислоте за счет повышения активности ферментов биосинтеза пуринов, был депонирован в Корейском Центре Культур микроорганизмов (KCCM), расположенном в Сеуле по адресу: Hongje 1-dong, Seodaemun-gu, Seoul, с номером доступа No. KCCM 10992P, 19 февраля 2009 года, в соответствии с Будапештским Договором.
ЭФФЕКТ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Микроорганизм рода Corynebacterium, продуцирующий 5'-инозиновую кислоту, по настоящему изобретению, в котором экспрессия гена, кодирующего ферменты биосинтеза пуринов, повышен до уровня, превосходящего его природную экспрессию, может быть использован для получения 5'-инозиновой кислоты с высокой концентрацией, что соответственно снижает производственные расходы.















Изобретение относится к области генной инженерии и биотехнологии и может быть использовано в пищевой и фармацевтической промышленностях. Микроорганизму рода Corynebacterium, продуцирующему 5'-инозиновую кислоту, придана способность сверхэкспрессировать гены, кодирующие ферменты пути биосинтеза пуринов. Указанные ферменты включают рибозофосфат-пирофосфокиназу, фосфорибозилглицинамид-формилтрансферазу, фосфорибозилформилглицинамидин-синтетазу, фосфорибозилформилглицинамидин-синтетазу II, фосфорибозиламиноимидазол-карбоксилазу, фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, циклогидролазу инозиновой кислоты, фосфорибозилпирофосфат-амидотрансферазу и фосфорибозиламиноимидазол-синтетазу. Способ получения 5'-инозиновой кислоты предусматривает культивирование указанного микроорганизма и выделение целевого продукта из культуральной среды. Применение изобретения обеспечивает высокий выход 5'-инозиновой кислоты. 2 н. и 10 з.п. ф-лы, 7 ил., 1 табл., 2 пр.
1. Микроорганизм, принадлежащий к роду Corynebacterium, продуцирующий 5'-инозиновую кислоту, в котором уровень экспрессии генов, кодирующих ферменты пути биосинтеза пуринов, превосходит соответствующий уровень природной экспрессии, где указанные ферменты пути биосинтеза пуринов включают рибозофосфат-пирофосфокиназу, фосфорибозилглицинамид-формилтрансферазу, фосфорибозилформилглицинамидин-синтетазу, фосфорибозилформилглицинамидин-синтетазу II, фосфорибозиламиноимидазол-карбоксилазу, фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу и циклогидролазу инозиновой кислоты.
2. Микроорганизм по п.1, где ферменты пути биосинтеза пуринов дополнительно включают фосфорибозилпирофосфат-амидотрансферазу и фосфорибозиламиноимидазол-синтетазу.
3. Микроорганизм по п.1, где указанные гены, кодирующие ферменты пути биосинтеза пуринов, включают ген purN с последовательностью SEQ ID NO: 36, который кодирует фосфорибозилглицинамид-формилтрансферазу, ген purS с последовательностью SEQ ID NO: 37, который кодирует фосфорибозилформилглицинамидин-синтетазу, ген purL с последовательностью SEQ ID NO: 38, который кодирует фосфорибозилформилглицинамидин-синтетазу II, гены ригКЕ с последовательностью SEQ ID NO: 40, которые кодируют фосфорибозиламиноимидазол-карбоксилазу, ген purC с последовательностью SEQ ID NO: 41, который кодирует фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, ген purH с последовательностью SEQ ID NO: 42, который кодирует циклогидролазу инозиновой кислоты, и ген prs с последовательностью SEQ ID NO: 43, который кодирует рибозофосфат-пирофосфокиназу.
4. Микроорганизм п.2, где указанные гены, кодирующие ферменты пути биосинтеза пуринов, включают ген purF с последовательностью SEQ ID NO: 35, который кодирует фосфорибозилпирофосфат-амидотрансферазу, ген purN с последовательностью SEQ ID NO: 36, который кодирует фосфорибозилглицинамид-формилтрансферазу, ген purS с последовательностью SEQ ID NO: 37, который кодирует фосфорибозилформилглицинамидин-синтетазу, ген purL с последовательностью SEQ ID NO: 38, который кодирует фосфорибозилформилглицинамидин-синтетазу II, ген purM с последовательностью SEQ ID NO: 39, который кодирует фофсфорибозиламиноимидазол-синтетазу, гены purKE с последовательностью SEQ ID NO: 40, которые кодируют фосфорибозиламиноимидазол-карбоксилазу, ген purC с последовательностью SEQ ID NO: 41, который кодирует фосфорибозиламиноимидазол-сукцинокарбоксамид-синтетазу, ген purH с последовательностью SEQ ID NO: 42, который кодирует циклогидролазу инозиновой кислоты, и ген prs с последовательностью SEQ ID NO: 43, который кодирует рибозофосфат-пирофосфокиназу.
5. Микроорганизм по п.1 или 2, в котором сверхэкспрессируются гены, кодирующие ферменты пути биосинтеза пуринов, за счет дополнительного введения в клетку генов, кодирующих ферменты пути биосинтеза пуринов, или за счет амплификации естественных генов, кодирующих ферменты пути биосинтеза пуринов.
6. Микроорганизм по п.5, в котором отдельный ген, кодирующий фермент пути биосинтеза пуринов, представлен в виде двух или более копий, в результате введения в клетку одной или нескольких копий, в дополнение к соответствующему естественному гену.
7. Микроорганизм по п.6, в котором отдельный ген, кодирующий фермент пути биосинтеза пуринов, введен в клетку путем трансформации с использованием рекомбинантного вектора, содержащего две копии соответствующего гена, которые последовательно организованы.
8. Микроорганизм по п.7, где указанный рекомбинантный вектор для введения указанного гена в микроорганизм по п.1 выбран из группы, состоящей из pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC и pDZ-2prs, и указанный рекомбинантный вектор для введения указанного гена в микроорганизм по п.2 выбран из группы, состоящей из pDZ-2purFM, pDZ-2purNH, pDZ-2purSL, pDZ-2purKE, pDZ-2purC и pDZ-2prs, причем векторы характеризуются картами, показанными на фиг.2-7 соответственно.
9. Микроорганизм по п.1 или 2, где указанный микроорганизм из рода Corynebacterium представляет собой Corynebacterium ammoniagenes.
10. Микроорганизм по п.3, где указанный микроорганизм представляет собой Corynebacterium ammoniagenes CN01-0120 (2purNH+2purSL+2purKE+2purC+2prs), полученный из Corynebacterium ammoniagenes CJIP2401.
11. Микроорганизм по п.4, где указанный микроорганизм представляет собой Corynebacterium ammoniagenes CN01-0316, депонированный как КССМ 10992Р.
12. Способ получения 5'-инозиновой кислоты, включающий культивирование микроорганизма, принадлежащего к роду Corynebacterium, по любому из пп.1-11, и выделение 5'-инозиновой кислоты из культуральной среды.
| WO 2008088156 A1, 24.07.2008 | |||
| MEGUMI SHIMAOKA et al | |||
| Effect of amplification of desensitized purF and prs on inosine accumulation in Escherichia coli | |||
| Journal of Bioscience and Bioengineering, 2005, v.103, №3, p.255-261 | |||
| US 6586214 B1, 01.07.2003 | |||
| KR 20060098340 A, 18.09.2006 | |||
| ШТАММЫ БАКТЕРИЙ Bacillus subtilis И Bacillus amyloliquefaciens-ПРОДУЦЕНТЫ ИНОЗИНА И СПОСОБ ПРОДУКЦИИ ИНОЗИНА С ИХ ИСПОЛЬЗОВАНИЕМ | 2005 |
|
RU2333949C2 |

