Область изобретения
Настоящее изобретение относится к новому варианту белка, обладающему активностью экспорта инозин-5'-монофосфата (IMP), к содержащему его микроорганизму и к способу получения IMP, и способу увеличения экспорта IMP с его использованием.
Предшествующий уровень техники
Инозин-5'-монофосфат (далее IMP), являющийся веществом на основе нуклеиновой кислоты, представляет собой промежуточное соединение в метаболическом пути нуклеиновой кислоты и используется во множестве областей, таких как продукты питания, лекарственные средства, различные медицинские применения и т.п. В частности, IMP широко используется в качестве добавки для пищевых приправ или пищевых продуктов наряду с гуанин-5'-монофосфатом (далее GMP). Хотя известно, что IMP сам по себе обеспечивает вкус мяса, также известно, что он усиливает вкус и запах мононатриевой соли глутаминовой кислоты (MSG) и таким образом привлекает внимание в качестве усиливающей вкус приправы на основе нуклеиновой кислоты.
Примеры способов получения IMP включают способ ферментативного расщепления рибонуклеиновой кислоты, экстрагированной из клеток дрожжей (публикация патента Японии №1614/1957), способ химического фосфорилирования инозина, продуцируемого путем ферментации (Agri. Biol. Chem., 36, 1511 и т.п.), способ культивирования микроорганизмов, которые могут непосредственно продуцировать IMP и выделять IMP в культуральный бульон и т.п.Среди этих способов наиболее часто используемым в настоящее время способом является способ с использованием микроорганизмов, способных непосредственно продуцировать IMP.
В то же самое время, поскольку ферменты не всегда демонстрируют оптимальные свойства, по природе относящиеся к активности, стабильности, субстратной специфичности в отношении оптических изомеров и т.п., которые требуются для промышленного применения, предпринимались различные попытки улучшить ферменты для того, чтобы соответствовать предполагаемому применению путем модификации их аминокислотных последовательностей и т.п. Среди этих попыток, несмотря на то, что рациональное конструирование и сайт-направленный мутагенез ферментов применялись для улучшения функции фермента, было показано, что во многих случаях эти попытки были неудачными, поскольку информация о структуре целевых ферментов не является достаточной или корреляция между структурой и функцией не является ясной, что препятствует их эффективному применению. Кроме того, ранее сообщали о способе улучшения активности фермента за счет попытки улучшить ферменты посредством направленной эволюции, предназначенной для отбора ферментов с желаемыми характеристиками из библиотеки модифицированных ферментов, сконструированных путем случайного мутагенеза генов ферментов.
Раскрытие изобретения
Техническая проблема
Для получения IMP с высоким выходом с использованием способа непосредственного продуцирования IMP путем микробной ферментации, IMP должен беспрепятственно экспортироваться. Для выполнения этой задачи авторы настоящего изобретения открыли белок, вовлеченный в активность экспорта IMP, и также приложили множество усилий для увеличения продуцирования IMP. В результате они открыли варианты белка, обладающие активностью экспорта IMP, завершив тем самым настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить вариант белка, обладающий активностью экспорта IMP.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить полинуклеотид, кодирующий вариант белка по настоящему изобретению.
Еще одна задача описания настоящего изобретения заключается в том, чтобы предложить вектор, включающий полинуклеотид по настоящему изобретению.
Еще одна задача описания настоящего изобретения заключается в том, чтобы предложить микроорганизм, продуцирующий IMP, который содержит вариант белка и вектор по настоящему изобретению.
Еще одна задача описания настоящего изобретения заключается в том, чтобы предложить способ получения IMP, включающий культивирование микроорганизма по настоящему изобретению в среде.
Еще одна задача описания настоящего изобретения заключается в том, чтобы предложить способ увеличения экспорта IMP, включающий усиление активности варианта белка по настоящему изобретению, который обладает активностью экспорта IMP.
Полезные эффекты изобретения
IMP может быть получен с высоким выходом путем культивирования микроорганизма рода Corynebacterium, продуцирующего IMP с использованием варианта белка по настоящему изобретению, который способен экспортировать IMP.
Лучший вариант осуществления изобретения
Настоящее изобретение будет подробно описано далее. В то же самое время, каждое пояснение и типичное воплощение может применяться в отношении других соответствующих пояснений и типичных воплощений. То есть, все комбинации раскрытых здесь разных факторов относятся к объему настоящего изобретения. Кроме того, объем настоящего изобретения не должен ограничиваться конкретным раскрытием, приведенным далее.
Для достижения вышеприведенных задач в аспекте настоящего изобретения предложен вариант белка, обладающий активностью экспорта IMP.
Используемый в настоящем описании термин "белок, который экспортирует инозин-5'-монофосфат (IMP)" относится к белку, вовлеченному во внеклеточный экспорт IMP. В описании настоящего изобретения этот термин может быть использован взаимозаменяемо с белком, обладающим активностью экспорта IMP, белком экспорта IMP, белком, обладающим активностью экспорта инозин-5'-монофосфата, экспортирующим инозин-5'-монофосфат белком, и т.п.; в частности, белок может быть выражен как ImpE, и, более конкретно, может быть выражен как ImpE1 или ImpE2, но не ограничивается ими. Кроме того, белок может быть получен из микроорганизма рода Corynebacterium, и в частности Corynebacterium stationis, но микроорганизм не ограничивается ими.
Белок, например, может состоять из аминокислотной последовательности, представленной SEQ ID NO: 1 или SEQ ID NO: 2, но любая последовательность, обладающая той же самой активностью как белок, может быть включена без ограничения, и специалист в данной области техники может получить информацию о последовательности в GenBank NCBI (Национальный центр биотехнологической информации), представляющей собой хорошо известную базу данных. Кроме того, белок может включать аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2, или аминокислотную последовательность, имеющую гомологию или идентичность с последовательностью SEQ ID NO: 1 или SEQ ID NO: 2 по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99%. Кроме того, очевидно, что любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой в часть последовательности, также может быть включен в объем настоящего изобретения, равно как и аминокислотная последовательность, имеющая описанную выше гомологию или идентичность и обладающая действием, соответствующим действию белка.
То есть, хотя в описании настоящего изобретения белок описан как "белок, имеющий аминокислотную последовательность конкретной SEQ ID NO" или "белок, состоящий из аминокислотной последовательности конкретной SEQ ID NO", белок может обладать активностью, которая идентична или соответствует активности белка, состоящего из аминокислотной последовательности, соответствующей SEQ ID NO. В таком случае, очевидно, что любые белки, имеющие аминокислотную последовательность с делецией, модификацией, заменой, консервативной заменой или вставкой в часть последовательности, также могут быть использованы в настоящем изобретении. Например, в том случае, когда белок обладает активностью, которая является такой же или соответствует активности модифицированного белка, не исключается вставка последовательности выше или ниже относительно аминокислотной последовательности, которая не изменяет функцию белка, мутация, которая может возникать в природе, его молчащая мутация или консервативная замена, и даже тогда, когда присутствует вставка последовательности или мутация, они очевидно относятся к объему настоящего изобретения.
Используемый здесь термин "гомология" или "идентичность" относится к степени совпадения двух заданных аминокислотных последовательностей или нуклеотидных последовательностей, и может выражаться в процентах.
Термины "гомология" и "идентичность" часто могут использоваться взаимозаменяемо друг с другом.
Гомология или идентичность последовательности консервативных полинуклеотидных или полипептидных последовательностей может быть определена при помощи стандартных алгоритмов выравнивания и могут быть использованы с штрафом за пропуск, устанавливаемым по умолчанию в используемой программе. В общем, ожидается, что по существу гомологичные или идентичные последовательности гибридизуются в условиях умеренной или высокой жесткости вдоль всей длины или по меньшей мере приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80% или приблизительно 90% от всей длины последовательностей. Также рассматриваются полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов в гибридизующихся полипептидах.
Обладают ли любые две полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, может быть определено с использованием известного компьютерного алгоритма, такого как программа "FASTA" (Pearson et al., (1988) [Proc. Natl. Acad. Sci. USA 85]: 2444: с использованием параметров по умолчанию в 2444). Альтернативно, они могут быть определены с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), который реализован в программе Needleman в пакете EMBOSS ((EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или более позние версии) (пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL, J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, и [CARILLO ETA/.] (1988) SIAM J Applied Math 48: 1073). Например, гомология, сходство или идентичность могут быть определены с использованием BLAST или ClustalW от National Center for Biotechnology Information (NCBI).
Гомология, сходство или идентичность полинуклеотидных или полипептидных последовательностей могут быть определены путем сравнения информации о последовательностях с использованием, например компьютерной программы GAP (например, Needleman et at, (1970), J Mol Biol. 48:443), как опубликовано (например, Smith and Waterman, Adv. Appl. Math (1981) 2:482). В общем, программа GAP определяет гомологию, сходство или идентичность в виде значения, получаемого путем деления количества одинаково выравненных символов (т.е. нуклеотидов или аминокислот) на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать (1) однокомпонентную матрицу сравнения (содержащую значение 1 для идентичности и 0 для отсутствия идентичности) и взвешенную матрицу сравнения в соответствии с Gribskov et al. (1986), Nucl. Acids Res. 14:6745, как раскрыто в Schwartz and Dayhoff, eds., Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp. 353-358, 1979; (2) штраф 3,0 для каждого пропуска и дополнительно штраф 0,10 для каждого символа в каждом пропуске (или штраф за внесение пропуска в выравнивание 10 и штраф за удлинение пропуска 0,5); и (3) отсутствие штрафа за концевые пропуски. Соответственно, используемый здесь термин "гомология" или "идентичность" относится к соответствию между последовательностями. Конкретно, вариант белка по настоящему изобретению, обладающий активностью экспорта IMP, может представлять собой вариант, в котором по меньшей мере одна аминокислота, выбранная из группы, состоящий из 164-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1, 222-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1, 2-й аминокислоты в аминокислотной последовательности SEQ ID NO: 2 и 64-й аминокислоты в аминокислотной последовательности SEQ ID NO: 2, заменена на другую аминокислоту, но не ограничивается ими.
Например, в варианте белка, обладающем активностью экспорта IMP, 164-я аминокислота в аминокислотной последовательности SEQ ID NO: 1 заменена на лизин, аргинин, аспарагин, глицин, треонин или пролин; 2-я аминокислота в аминокислотной последовательности SEQ ID NO: 2 заменена на изолейцин, фенилаланин, метионин, глутаминовую кислоту, гистидин или аспарагин; или 64-я аминокислота в аминокислотной последовательности с SEQ ID NO: 2 заменена на аспарагиновую кислоту, глутаминовую кислоту, аспарагин, цистеин, изолейцин или фенилаланин, но не ограничивается ими.
В качестве конкретного примера варианта белка, обладающего активностью экспорта IMP, может представлять собой белок, имеющий аминокислотную последовательность, состоящую из SEQ ID NO: 141, 142, 145, 147, 149 или 151, белок, имеющий аминокислотную последовательность, кодируемую полинуклеотидом с SEQ ID NO: 153 или 154, или белок, имеющий аминокислотную последовательность, имеющую гомологию с ними по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99%. Кроме того, понятно, что белок имеющий делецию, модификацию, замену или добавление некоторой последовательности, может быть использован как белок по настоящему изобретению при условии, что он представляет собой белок, имеющий аминокислотную последовательность с вышеприведенной гомологией и демонстрирующий действие, соответствующее действию белка по изобретению.
В еще одном аспекте настоящего изобретения предложен полинуклеотид, кодирующий вариант белка, или вектор, включающий этот полинуклеотид.
Используемый здесь термин "полинуклеотид" относится к полимеру из нуклеотидов, которые связаны в длинной цепи при помощи ковалентных связей, и имеет длину цепи ДНК или цепи РНК больше определенной, и более конкретно, относится к фрагменту полинуклеотид а, кодирующему вариант белка.
Понятно, что полинуклеотид, который может быть транслирован за счет вырожденности кодона в белок, состоящий из аминокислотной последовательности SEQ ID NO: 141, 142, 145, 147, 149 или 151, в белок, состоящий из аминокислотной последовательности, кодируемой полинуклеотидом с SEQ ID NO: 153 или 154, или в белок, обладающий гомологией с ними, также может быть включен в качестве полинуклеотид а по настоящему изобретению. Например, полинуклеотид по настоящему изобретению может представлять собой полинуклеотид, имеющий нуклеотидную последовательность SEQ ID NO: 143, 144, 146, 148, 150, 152, 153 или 154, и, более конкретно, может представлять собой полинуклеотид, состоящий из нуклеотидной последовательности SEQ ID NO: 143, 144, 146, 148, 150, 152, 153 или 154. Кроме того, полинуклеотидная последовательность, которая кодирует белок, обладающий активностью белка, состоящего из аминокислотной последовательности SEQ ID NO: 141, 142, 145, 147, 149 или 151, или аминокислотной последовательности, кодируемой полинуклеотидом с SEQ ID NO: 153 или 154, определенная путем гибридизации в жестких условиях с зондом, который может быть получен из известной последовательности гена, например последовательности, комплементарной всей или части нуклеотидной последовательности, может быть включена без ограничений.
Термин "жесткие условия" относится к условиям, при которых становится возможной гибридизация между полинуклеотидами. Такие условия в частности описаны в источниках (например, J. Sambrook et al., ранее). Например, эти условия могут включать осуществление гибридизации между генами, обладающими высокой гомологией, гомологией 40% или выше, в частности 90% или выше, конкретней 95% или выше, еще конкретней 97% или выше, и, в частности, 99% или выше, при отсутствии гибридизации между генами, имеющими гомологию меньше чем вышеприведенные гомологии; или для осуществления гибридизации один раз, в частности два или три раза, в обычных условиях отмывания для Саузерн гибридизации, составляющих 60°С, 1× SSC (раствор хлорида и цитрата натрия) и 0,1% SDS (додецилсульфат натрия), в частности при концентрации соли и температуре, соответствующей 60°С, 0,1× SSC и 0,1% SDS, и более конкретно при 68°С, 0,1× SSC и 0,1% SDS.
Для гибридизации требуется, чтобы две нуклеиновые кислоты имели комплементарную последовательность, хотя ошибки спаривания между основаниями могут быть возможны в зависимости от жесткости гибридизации. Термин "комплементарный" используется для описания взаимосвязи между попарно гибридизуемыми нуклеотидными основаниями. Например, в отношении ДНК аденозин комплементарен тимину, и цитозин комплементарен гуанину. Соответственно, описание настоящего изобретения также может включать выделенные фрагменты нуклеиновых кислот, комплементарные полноразмерной последовательности, а также по существу похожие последовательности нуклеиновых кислот.
В частности, полинуклеотиды, имеющие гомологию, могут быть обнаружены при величине Tm (температуры плавления) 55°С с использованием условий гибридизации, которые включают стадию гибридизации и использование описанных выше условий. Кроме того, величина Tm может составлять 60°С, 63°С или 65°С, но не ограничивается ими, и может быть подходящим образом скорректирована специалистом в данной области техники в соответствии с предполагаемой задачей.
Жесткость, подходящая для гибридизации полинуклеотидов, зависит от длины и комплементарности полинуклеотидов и связанных переменных, хорошо известных в области техники (смотри, Sambrook et al., ранее, 9.50-9.51 и 11.7-11.8).
Используемый в настоящем описании термин "вектор" относится к конструкции ДНК, включающей нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, в которой целевой белок функционально связан с подходящей контрольной последовательностью, таким образом, что целевой белок может экспрессироваться в подходящем хозяине. Контрольная последовательность может включать промотор, способный инициировать транскрипцию, любую операторную последовательность для контроля транскрипции, последовательность, кодирующую подходящий домен мРНК для связывания с рибосомой, и последовательность, контролирующую окончание транскрипции и трансляции. Этот вектор после трансформации в подходящую клетку-хозяин может реплицироваться или функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Вектор, используемый в настоящем описании, может быть не ограничен конкретным образом при условии, что этот вектор является реплицируемым в клетке-хозяине, и он может быть сконструирован с использованием любого вектора, известного в области техники. Примеры вектора могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора может быть использован pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.п; и в качестве плазмидного вектора может быть использован вектор на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и т.п. В частности, могут быть использованы векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.п.
В одном воплощении полинуклеотид, кодирующий целевой белок, может быть заменен на модифицированный полинуклеотид в хромосоме с использованием вектора для встраивания в хромосому в клетке. Встраивание полинуклеотида в хромосому может быть осуществлено с использованием способа, известного в области техники, например путем гомологичной рекомбинации, но не ограничивается им. В частности, маркер селекции может быть дополнительно включен для подтверждения встраивания в хромосому. Маркер селекции используется для отбора трансформированной клетки, т.е. для подтверждения того, осуществлено ли встраивание целевой нуклеиновой кислоты, и могут быть использованы маркеры, способные обеспечить способные к отбору фенотипы, такие как устойчивость к лекарственным средствам, потребность в питательных веществах, устойчивость к цитотоксическим агентам и экспрессия поверхностных белков. В условиях, когда обрабатывают агентами селекции, только клетки, способные экспрессировать маркеры селекции, могут выживать или проявлять другие фенотипические характеристики, и, таким образом, легко могут быть отобраны трансформированные клетки.
В еще одном аспекте настоящего изобретения предложен микроорганизм, продуцирующий IMP, который содержит вариант белка по настоящему изобретению, полинуклеотид по настоящему изобретению, кодирующий указанный вариант белка, или вектор по настоящему изобретению. В частности, микроорганизм, содержащий вариант белка и/или полинуклеотид, кодирующий этот вариант белка, может представлять собой микроорганизм, полученный путем трансформации с использованием вектора, содержащего полинуклеотид, кодирующий указанный вариант белка, однако микроорганизм не ограничивается ими.
Используемый в описании настоящего изобретения термин "трансформация" относится к процессу внедрения вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, за счет этого обеспечивая экспрессию белка, кодируемую этим полинуклеотидом, в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения, встроен ли он в хромосому клетки-хозяина и находится в ней или находится вне хромосомы, при условии, что этот трансформированный полинуклеотид может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и РНК, которые кодируют целевой белок. Полинуклеотид может быть встроен в любой форме при условии, что он может быть внедрен в клетку хозяина и экспрессироваться в ней. Например, полинуклеотид может быть внедрен в клетку хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все существенные элементы, требующиеся для самоэкспрессии. Экспрессионная кассета обычно может включать промотор, функционально связанный с полинуклеотидом, сигнал прекращения транскрипции, домен связывания с рибосомой и сигнал прекращения трансляции. Экспрессионная кассета может находиться в форме самореплицируемого экспрессионного вектора. Кроме того, полинуклеотид может быть внедрен в клетку-хозяина сам по себе и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине, но не ограничивается этим.
Кроме того, используемый в настоящем описании термин "функционально связанный" относится к функциональной связи между промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок, т.е. конъюгата в соответствии с настоящим описанием, и вышеуказанной генной последовательностью.
Используемый в настоящем описании термин "микроорганизм, продуцирующий IMP" относится к микроорганизму, который в естественных условиях способен продуцировать IMP; или к микроорганизму, в который внедрена способность продуцировать или экспортировать IMP, родительский штамм которого в природе не способен продуцировать и/или экспортировать IMP в соответствии с настоящим описанием, причем микроорганизм, продуцирующий IMP, может быть использован взаимозаменяемо с микроорганизмом, обладающим активностью экспорта IMP.
Микроорганизм, продуцирующий IMP, представляет собой клетку или микроорганизм, который содержит вариант белка, обладающий активностью экспорта IMP, или полинуклеотид, кодирующий этот вариант белка, или который трансформирован вектором, содержащим полинуклеотид, кодирующий этот вариант белка, и за счет этого способным экспрессировать этот вариант белка. Для целей в соответствии с настоящим описанием клетка-хозяин микроорганизма, продуцирующего IMP, или микроорганизм может представлять собой любой микроорганизм, содержащий вариант белка, за счет которого он способен продуцировать IMP. Например, микроорганизм может представлять собой микроорганизм рода Escherichia, микроорганизм рода Serratia, микроорганизм рода Erwinia, микроорганизм рода Enterobacteria, микроорганизм рода Salmonella, микроорганизм рода Streptomyces, микроорганизм рода Pseudomonas, микроорганизм рода Brevibacterium, микроорганизм рода Corynebacterium и т.п, и, в особенности, микроорганизм рода Corynebacterium.
Используемый в описании настоящего изобретения термин "микроорганизм, продуцирующий IMP, рода Corynebacterium" относится к микроорганизму рода Corynebacterium, который в естественных условиях способен продуцировать IMP, или способен продуцировать IMP за счет модификации. В частности, используемый в настоящем описании микроорганизм рода Corynebacterium, способный продуцировать IMP, относится к нативному штамму микроорганизма рода Corynebacterium, способному продуцировать IMP; или микроорганизму рода Corynebacterium, обладающему усиленной способностью продуцировать IMP, полученному посредством встраивания гена, ассоциированного с продукцией IMP, или путем усиления или ослабления эндогенного гена, ассоциированного с продукцией IMP. Более конкретно, в настоящем описании микроорганизм рода Corynebacterium, способный продуцировать IMP, относится к микроорганизму рода Corynebacterium, который обладает улучшенной способностью продуцировать IMP за счет включения варианта белка, обладающего активностью экспорта IMP, или полинуклеотида, кодирующего этот вариант белка, или за счет трансформации вектором, содержащим полинуклеотид, кодирующий этот вариант белка. Термин "микроорганизм рода Corynebacterium с усиленной способностью продуцировать IMP" относится к микроорганизму рода Corynebacterium с улучшенной способностью продуцировать IMP по сравнению со способностью его родительского штамма до трансформации или способностью немодифицированного микроорганизма рода Corynebacterium. Термин "немодифицированный микроорганизм рода Corynebacterium" относится к нативному типу микроорганизма рода Corynebacterium, микроорганизму рода Corynebacterium, который не содержит вариант белка, способный экспортировать IMP, или микроорганизму рода Corynebacterium, который не трансформирован вектором, содержащим полинуклеотид, кодирующий вариант белка, способный экспортировать IMP.
В воплощении настоящего изобретения микроорганизм по настоящему изобретению может представлять собой микроорганизм рода Corynebacterium, у которого активность аденилосукцинатсинтетазы и/или IMP-дегидрогеназы дополнительно ослаблена.
В настоящем описании термин "микроорганизм рода Corynebacterium" в частности относится к Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens, Corynebacterium stationis и т.п., но микроорганизм не обязательно ограничен ими.
В еще одном аспекте описания настоящего изобретения предложен способ получения IMP, включающий культивирование микроорганизма рода Corynebacterium в среде.
В частности, способ в соответствии с описанием настоящего изобретения может дополнительно включать стадию выделения IMP из микроорганизма или среды.
В вышеприведенном способе культивирование микроорганизма может быть осуществлено в периодическом процессе, непрерывном процессе, периодическом процессе с подпиткой и т.п., которые известны из уровня техники, но способ культивирования конкретно не ограничивается ими. В частности, в отношении условий культивирования, рН культуры может быть доведен до подходящего рН (например, рН 5-9, в частности рН 6-8, и, более конкретно, при помощи подходящего основного соединения (например, гидроксида натрия, гидроксида калия или аммиака) или кислотного соединения (например, фосфорной кислоты или серной кислоты), и аэробное состояние культуры может поддерживаться путем введения в культуру кислорода или содержащей кислород газовой смеси. Температура культивирования, как правило, может находиться в диапазоне от 20°С до 45°С, и, в частности, от 25°С до 40°С, в течение, приблизительно от 10 до 160 часов, однако условия культивирования не ограничены ими. IMP, продуцируемый при помощи вышеприведенного культивирования, может быть секретирован в культуру или может сохраняться в клетках.
Кроме того, примеры источников углерода, используемых в культуральной среде, могут включать сахара и углеводы (например, глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу); масла и жиры (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло); жирные кислоты (например, пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту); спирты (например, глицерин и этанол) и органические кислоты (например, уксусную кислоту), но не ограничены ими. Эти источники углерода могут быть использованы по отдельности или в комбинации, но не ограничены этим. Примеры источников азота, используемых в культуральной среде, могут включать азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной сок, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) и т.п. Эти источники азота могут быть использованы по отдельности или в комбинации, но не ограничены этим. Примеры источников фосфора, используемых в культуральной среде, могут включать дигидрофосфат калия, дикалия гидрофосфат, соответствующие натрийсодержащие соли и т.п., но не ограничены ими. Кроме того, в среде могут содержаться соли металлов (например, сульфат магния или сульфат железа), аминокислоты, витамины и т.п., которые представляют собой важные вещества, способствующие росту.
В описании настоящего изобретения способ выделения IMP, продуцируемого на стадии культивирования, может быть осуществлен путем сбора IMP из культурального бульона с использованием подходящего метода, известного в области техники. Например, могут быть использованы такие методы, как центрифугирование, фильтрация, анионообменная хроматография, кристаллизация, HPLC (высокоэффективная жидкостная хроматография) и т.п., и требуемый IMP может быть выделен из культуры или культивируемого микроорганизма с использованием подходящего метода, известного в области техники.
Кроме того, выделение может включать процесс очистки и может быть осуществлено с использованием подходящего метода, известного в области техники. Таким образом, выделяемый IMP может быть представлен в очищенной форме или в форме ферментативного бульона микроорганизма, содержащего IMP.
В еще одном аспекте настоящего изобретения предложена композиция для продуцирования IMP, содержащая вариант белка по настоящему изобретению, который обладает активностью экспорта IMP, или кодирующий его полинуклеотид.
Композиция по настоящему изобретению может дополнительно включать, без ограничения, структуру, способную делать полинуклеотид функциональным. В композиции по настоящему изобретению полинуклеотид может находиться в форме, включенной в вектор для экспрессии функционально связанного гена в клетке хозяине, в которую он внедрен.
Кроме того, композиция дополнительно может включать любые подходящие эксципиенты, обычно используемые в композиции для продукции IMP. Такие эксципиенты могут представлять собой, например, консерванты, увлажнители, суспендирующие агенты, буферы, стабилизирующие агенты или изотонические агенты, но не ограничены ими.
В еще одном аспекте настоящего изобретения предложено применение белка по настоящему изобретению для увеличения продуцирования IMP в микроорганизмах рода Corynebacterium.
В еще одном аспекте настоящего изобретения предложен способ увеличения экспорта IMP, включающий усиление активности варианта белка, который обладает активностью экспорта IMP, в микроорганизме рода Corynebacterium.
Термины "белок, обладающий активностью экспорта IMP", "усиление" и "микроорганизм рода Corynebacterium" являются такими, как описано выше.
В еще одном аспекте настоящего изобретения предложено применение белка по настоящему изобретению для увеличения экспорта IMP в микроорганизме рода Corynebacterium.
Подробное описание воплощения
Далее настоящее изобретение будет подробно описано с помощью типичных воплощений. Однако для специалиста в данной области техники должно быть очевидно, что эти типичные воплощения приведены исключительно с иллюстративной целью и не предполагается, что они ограничивают объем описания настоящего изобретения.
Пример 1: Обнаружение белков экспорта IMP
Библиотеку геномной ДНК Corynebacterium stationis АТСС6872 получали для идентификации мембранных белков Corynebacterium, вовлеченных в экспорт IMP. Затем, поскольку штамм дикого типа Corynebacterium не может продуцировать IMP, или даже если он продуцирует IMP, то продуцирует лишь небольшое его количество, то получали происходящий из штамма АТСС6872 штамм, названный CJI0323, который способен продуцировать IMP, для идентификации способности продуцировать IMP. Полученный штамм CJI0323 подвергали скринингу мембранных белков, вовлеченных в экспорт IMP, с использованием библиотеки геномной ДНК штамма АТСС6872. Конкретная подробная информация об эксперименте является следующей.
Пример 1-1: Отбор продуцирующего IMP штамма CJI0323
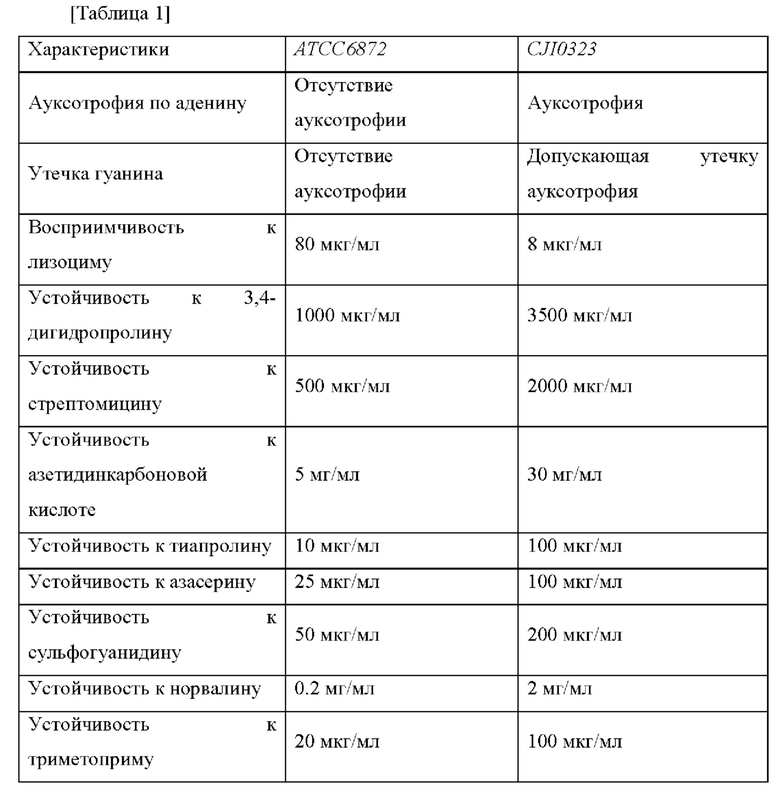
Клетки АТСС6872 суспендировали в фосфатном буфере (рН 7,0) или в цитратном буфере (рН 5,5) в концентрации от 107 клеток/мл до 108 клеток/мл для получения продуцирующего IMP штамма, являющегося производным АТСС6872, и клетки подвергали УФ (ультрафиолетовой) обработке для того, чтобы индуцировать мутацию. Полученные в результате клетки дважды промывали 0,85% физиологическим раствором, и затем разбавляли и высевали на среду, которую получали путем добавления обеспечивающего устойчивость вещества в подходящей концентрации в минимальную среду, содержащую 1,7% агар, и после этого получали колонии. Каждую колонию культивировали в питательной среде и культивировали в среде для посева в течение 24 часов. После культивирования колоний в течение от 3 до 4 суток в ферментационной среде отбирали колонию, обладающую самыми высокими способностями к продуцированию IMP, накапливаемому в культуральной среде. В процессе приготовления штамма, способного продуцировать IMP в высокой концентрации, для того, чтобы обеспечить ауксотрофию по аденину, утечку гуанина, восприимчивость к лизоциму, устойчивость к 3,4-дигидропролину, устойчивость к стрептомицину, устойчивость к азетидинкарбоновой кислоте, устойчивость к тиапролину, устойчивость к азасерину, устойчивость к сульфагуанидину, устойчивость к норвалину и устойчивость к триметоприму, вышеуказанные процедуры осуществляли последовательно для каждого вещества. Наконец, в результате отобрали CJI0323, который демонстрировал устойчивость к вышеприведенным веществам и превосходные способности продуцировать IMP. Степень устойчивости сравнивали между АТСС6872 и CJI0323, и результаты представлены в таблице 1 ниже.
- Минимальная среда: 2% глюкоза, 0,3% сульфат натрия, 0,1% KH2SO4, 0,3% K2HPO4, 0,3% сульфат магния, хлорид кальция (10 мг/л), сульфат железа (10 мг/л), сульфат цинка (1 мг/л), хлорид марганца (3,6 мг/л), L-цистеин (20 мг/л), пантотенат кальция (10 мг/л), тиамина хлоргидрат (5 мг/л), биотин (30 мкг/л), аденин (20 мг/л), гуанин (20 мг/л), рН 7,3
- Питательная среда: 1% петон, 1% мясной сок, 0,25% хлорин натрия, 1% дрожжевой экстракт, 2% агар, рН 7,2
- Среда для посева: 1% глюкоза, 1% пептон, 1% мясной сок, 1% дрожжевой экстракт, 0,25% хлорид натрия, аденин (100 мг/л), гуанин (100 мг/л), рН 7,5
- Ферментационная среда: 0,1% глутамат натрия, 1% хлорид аммония, 1,2% сульфат магния, 0,01% хлорид кальция, сульфат железа (20 мг/л), сульфат марганца (20 мг/л), сульфат цинка (20 мг/л), сульфат меди (5 мг/л), L-цистеин (23 мг/л), аланин (24 мг/л), никотиновая кислота (8 мг/л), биотин (45 мкг/л), тиамина хлоргидрат (5 мг/л), аденин (30 мг/л), 1,9% фосфорная кислота (85%), 2,55% глюкоза, 1,45% фруктоза.
Пример 1-2: Эксперименты по титру ферментации CJI0323
Среду для посева (2 мл) разливали в пробирки для тестирования (диаметр: 18 мм), которые затем автоклавировали и каждую из которых инокулировали АТСС6872 и CJI0323. Затем полученные в результате культуры культивировали при встряхивании при 30°С в течение 24 часов и затем использовали в качестве раствора посевной культуры. Ферментационную среду (29 мл) разливали в колбы Эрленмейера (250 мл) для встряхивания, автоклавировали при 121°С в течение 15 минут, и в нее инокулировали раствор посевной культуры (2 мл) и культивировали в течение 3 суток. Условия культивирования устанавливались следующими: 170 об/мин, 30°С и рН 7,5.
После завершения культивирования количество продуцируемого IMP измеряли при помощи HPLC (SHIMAZDU LC20A), и результаты культивирования представлены в таблице 2 ниже.

Штамм CJI0323 был назван Corynebacterium stationis CN01-0323. Этот штамм был депонирован в соответствии с Будапештским договором в Korean Culture Center of Microorganisms (Корейском центре культур микроорганизмов) (KCCM) 7 ноября 2017 года. Кроме того, этому штамму был присвоен номер доступа KCCM12151P.
Пример 1-3: Обнаружение белков экспорта
Условия скрининга, демонстрирующие ингибирование роста штамма CJI0323, определяли путем дополнительного добавления IMP в минимальную среду, содержащую 1,7% агар. Плазмиды геномной библиотеки штамма АТСС6872 трансформировали в штамм CJI0323 путем электропорации (van der Rest et al. 1999), и отбирали те колонии, в которых ингибирование роста запускалось в условиях среды, дополненной избыточным количеством IMP. Из отобранных колоний получали плазмиды, и их анализировали при помощи метода секвенирования. В результате идентифицировали один тип мембранного белка, вовлеченного в запуск ингибирования роста в условиях, при которых добавляли избыточное количество IMP.
Этот один тип мембранного белка Corynebacterium идентифицировали на основании аминокислотной последовательности SEQ ID NO: 2 и нуклеотидной последовательности SEQ ID NO: 4 (NCBI GenBank: NZ_CP014279, WP_066795121, MFS транспортер). Этот мембранный белок известен как MFS транспортер, но его специфическая функция не была подтверждена, и кроме того, его функция в отношении экспорта IMP все еще неизвестна. В настоящем описании этот мембранный белок назван ImpE2(WT).
Пример 2: Идентификация ImpE1 и ImpE2
Пример 2-1: Подтверждение impE1 и impE2
Для исследования функций мембранного белка ImpE2 структуру гена с SEQ ID NO: 4 подтверждали в NCBI (NCBI GenBank: NZ_СР014279, WP_066795121, MFS транспортер). В результате было подтверждено, что исходный фрагмент длиной 7 п.о. в ORF (открытая рамка считывания) в SEQ ID NO: 4 (impE2) перекрывается на 7 п.о. с другим геном (NCBI GenBank: NZ_СР014279, WP_066795119, регулятор транскрипции), который расположен выше относительно impE2. Поскольку функция гена, расположенного выше относительно impE2, и белка, кодируемого этим геном, еще не подтверждена, в настоящем описании, этот белок был назван ImpE1(WT) (аминокислотная последовательность SEQ ID NO: 1 и нуклеотидная последовательность SEQ ID NO: 3).
Пример 2-2: Получение вектора, дефектного по impE1 или impE2
Для подтверждения того, может ли делеция ImpE1 или ImpE2, которые вовлечены в запуск ингибирования роста, которое вызвано IMP, как установлено в примерах 1 и 2-1, в продуцирующем IMP штамме уменьшить его способность экспортировать IMP, были предприняты попытки получить векторы, дефектные по каждому из этих генов.
Генные фрагменты для получения векторов получали при помощи ПЦР с использованием геномной ДНК штамма АТСС6872 в качестве матрицы.
Более конкретно, ПЦР для impE1 осуществляли с использованием праймеров с SEQ ID NO: 5 и 6 и праймеров с SEQ ID NO: 7 и 8; и ПЦР для impE2 осуществляли с использованием праймеров с SEQ ID NO: 9 и 10 и праймеров с SEQ ID NO: 11 и 12 (таблица 3).

В частности, используемые праймеры получали на основе информации о гене Corynebacterium stationis (АТСС6872) (NCBI Genbank: NZ_СР014279), зарегистрированном в NIH GenBank, и соседних с ним нуклеотидных последовательностях.
ПЦР осуществляли посредством исходной денатурации при 94°С в течение 5 минут; 25 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 52°С в течение 30 минут и полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут.
ПЦР с перекрывающимися праймерами осуществляли с использованием двух фрагментов гена impE1, которые амплифицировали с использованием праймеров с SEQ ID NO: 5 и 6 и праймеров с SEQ ID NO: 7 и 8, в качестве матриц, и в результате получали полинуклеотидную матрицу (1,8 т.п.о. (тысяч пар оснований)). Полученный генный фрагмент клонировали в линеаризованный вектор pDZ (Корейский патент №10-0924065 и Международная заявка на патент №2008-033001), который расщепляли ферментом рестрикции (XbaI) и лигировали с использованием лигазы Т4, и, таким образом, получали вектор pDZ-ΔimpE1. Кроме того, полимеразную цепную реакцию с перекрывающимися праймерами осуществляли с использованием фрагмента гена impE2, амплифицированного с использованием праймеров с SEQ ID NO: 9 и 10, и двух фрагментов гена impE2, амплифицированных с использованием праймеров с SEQ ID NO: 11 и 12, в качестве матриц, и в результате получали полинуклеотидную матрицу (1,7 т.п.о.). Полученный генный фрагмент расщепляли ферментами рестрикции XbaI и SpeI. Генный фрагмент клонировали с использованием лигазы Т4 в линеаризованный вектор pDZ, который уже был расщеплен ферментом рестрикции (XbaI), и, таким образом получали вектор pDZ-ΔimpE2.
Пример 2-3: Получение векторов, дефектных по интегрированию impE1 и impE2
Поскольку гены impE1 и impE2, которые кодируют белки, вовлеченные в запуск ингибирования роста, которое вызвано IMP, перекрываются, существует потребность в одновременной регуляции обоих генов. Поэтому были осуществлены попытки получить вектор, дефектный как по impE1, так и по impE2.
Для ПЦР генов impE1 и impE2 использовали праймеры с SEQ ID NO: 5 и 13 и праймеры с SEQ ID NO: 14 и 12. Используемые праймеры получали на основе информации о гене Corynebacterium stationis (АТСС6872) (NCBI Genbank: NZ_CP014279), зарегистрированной в NIH GenBank, и соседних с ним нуклеотидных последовательностях. ПЦР с перекрывающимися праймерами осуществляли с использованием фрагмента гена impE1, амплифицированного с использованием праймеров с SEQ ID NO: 5 и 13, и двух фрагментов гена impE2, амплифицированных с использованием праймеров с SEQ ID NO: 14 и 12, в качестве матриц, и в результате получали полинуклеотидную матрицу (2,0 т.п.о.). Полученные генные фрагменты расщепляли XbaI и SpeI, соответственно. Генные фрагменты клонировали с использованием лигазы Т4 в линеаризованный вектор pDZ, который уже был расщеплен ферментом рестрикции (XbaI), и, таким образом получали вектор pDZ-ΔimpE1E2.
Пример 2-4: Получение штаммов, дефектных по impEl и impE2
Каждый из двух видов плазмид, полученных в примере 2-2, и один вид плазмиды, полученный в примере 2-3, трансформировали в штамм CJI0323 путем электропорации (с использованием метода трансформации, раскрытого в Appl. Microbiol. Biotechnol. (1999) 52: 541-545). Штаммы, в которых вектор встраивался в хромосому посредством рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Генетический дефект в окончательно трансформированных штаммах подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NO: 5 и 8, SEQ ID NO: 9 и 12 и SEQ ID NO: 5 и 12.
Отобранные штаммы были названы CJI0323_ΔimpE1, CJI0323_ΔimpE2 и CJI0323_ΔimpE1E2. Кроме того, оценивали способности этих штаммов продуцировать IMP.
Среду для посева (2 мл) разливали в пробирки для тестирования (диаметр: 18 мм), которые затем автоклавировали, каждую из которых инокулировали CJI0323, CJI0323_ΔimpE1, CJI0323_ΔimpE2 и CJI0323_ΔimpE1E2, культивировали при перемешивании при 30°С в течение 24 часов и использовали в качестве растворов посевных культур. Ферментационную среду (29 мл) разливали в колбы Эрленмейера (250 мл) для встряхивания и автоклавировали при 121°С в течение 15 минут. Затем раствор посевной культуры (2 мл) инокулировали в нее, и полученную в результате культуру культивировали в течение 3 суток. Условия культивирования устанавливались следующими: 170 об/мин, 30°С и рН 7,5.
После завершения культивирования количество продуцируемого IMP измеряли при помощи HPLC, и результаты для культуры представлены в таблице 4 ниже.

Количество IMP, накопленное в каждом штамме, сравнивали с количеством IMP для родительского штамма Corynebacterium stationis CJI0323. В результате обнаружено, что, как показано выше в таблице 4 выше, концентрации IMP в штаммах CJI0323_ΔimpE1, CJI0323_ΔimpE2 и CJI0323_ΔimpE1E2 были снижены по сравнению с родительским штаммом на приблизительно 8 г/л в тех же самых условиях, что подтверждает то, что ImpE1 и ImpE2 представляют собой белки, вовлеченные в экспорт IMP.
Пример 3: Подтверждение нуклеотидных последовательностей impE1 и impE2 в продуцирующем IMP штамме CJI0323
В случае штамма CJI0323, продуцирующего IMP в высокой концентрации в примере 1, возможно, что этот штамм обладает улучшенной способностью экспортировать IMP, таким образом, чтобы продуцировать IMP в высокой концентрации. Соответственно, была предпринята попытка подтвердить присутствие какой-либо мутации в impE1 и impE2 в штамме CJI0323.
Хромосомную ДНК штамма CJI0323 амплифицировали при помощи полимеразной цепной реакции (далее "ПЦР"). Более конкретно, сначала ПЦР осуществляли путем повторения 28 циклов, состоящих из денатурации при 94°С в течение 1 минуты, отжига при 58°С в течение 30 секунд и полимеризации при 72°С в течение 2 минут с использованием хромосомной ДНК штамма CJI0323 в качестве матрицы вместе с праймерами с SEQ ID NO: 15 и 16 (таблица 5), и таким образом амплифицировали фрагмент из приблизительно 2,8 т.п.о.
При анализе нуклеотидной последовательности с использованием тех же самых праймеров подтвердили, что 490-й нуклеотид гена impE1 gene (т.е. g) заменен на "а" по сравнению с нуклеотидной последовательностью штамма дикого типа АТСС6872. Эта замена свидетельствует о существовании модификации, при которой 164я аминокислота в белке ImpE1 (т.е. глутаминовая кислота) заменена на лизин.
Кроме того, подтвердили, что 4-й нуклеотид в гене impE2 (т.е. g) заменен на "а" (это означает, что 666-й нуклеотид в гене impE1 (т.е. g) заменен на "а") и 191-й нуклеотид в гене impE1 (т.е. g) заменен на "а". Эти замены свидетельствуют о модификациях, при которых 2-я аминокислота белка ImpE2 (т.е. валин), которая соответствует 222-й аминокислоте белка ImpE1, заменена на изолейцин; и 64-я аминокислота белка ImpE2 (т.е. глицин) заменена на глутаминовую кислоту.
Нуклеотид impE1 штамма CJI0323 назван impE1 CJI0323 (SEQ ID NO: 143), и соответствующий ему белок назван ImpE1 CJI0323 (SEQ ID NO: 141), тогда как нуклеотид impE2 штамма CJI0323 назван impE2_CJI0323 (SEQ ID NO: 144), и соответствующий ему белок назван ImpE2_CJI0323 (SEQ ID NO: 142).
Пример 4: Восстановление модификаций в impE1 и impE2
Пример 4-1: Получение векторов для восстановления модификаций в impE1 или impE2
В примере 3 исследовали наличие какой-либо модификации в impE1 и impE2 в продуцирующем IMP штамме CJI0323. В результате подтвердили, что impE1 имеет одну модификацию и impE2 имеет две модификации. Поскольку штамм CJI0323 продуцирует IMP в высокой концентрации, весьма вероятно, что эта модификация представляет собой такую модификацию, которая может улучшать способность экспортировать IMP. Соответственно, после восстановления мутантных impE1 и impE2 до нативного дикого типа ImpE без модификации осуществили следующие эксперименты для подтверждения того, действительно ли каждая модификация обеспечивает способность экспортировать IMP.
Для получения восстанавливающего вектора осуществляли ПЦР с использованием Corynebacterium stationis АТСС6872 в качестве матрицы.
Фрагмент гена impE1impE2, амплифицированного с использованием праймеров в соответствии с SEQ ID NOS: 17 и 18, обрабатывали ферментом рестрикции XbaI и клонировали в сайт рестрикции XbaI на векторе pDZ, и, таким образом получали pDZ-impE1E2(WT).
Пример 4-2: Получение векторов с единичной модификацией в impE1 или impE2
Вектор с единичной модификацией E164K в гене ImpE1 получали с использованием нативного штамма дикого типа Corynebacterium stationis АТСС6872 в качестве матрицы вместе с праймерами с SEQ ID NO: 19 и 20 и праймерами в соответствии с SEQ ID N: 21 и 22. ПЦР с перекрывающимися праймерами осуществляли с использованием фрагмента гена E164K-1, амплифицированного с использованием праймеров в соответствии с SEQ ID NOS: 19 и 20 и двух фрагментов гена E164K-2, амплифицированных с использованием праймеров с SEQ ID NO: 21 и 22, и, таким образом получали матрицу с полинуклеотидом в 1,8 т.п.о. Полученные генные фрагменты расщепляли при помощи XbaI и клонировали в линеаризованный вектор pDZ, который уже был расщеплен при помощи XbaI, с использованием лигазы Т4, и, таким образом получали вектор pDZ-impE1(E164K).
Вектор с единичной модификацией V2I в гене ImpE2 получали с использованием штамма АТСС6872 в качестве матрицы вместе с праймерами с SEQ ID NO: 19 и 23 и праймерами с SEQ ID NO: 24 и 22. ПЦР с перекрывающимися праймерами осуществляли с использованием фрагмента гена V2I-1, амплифицированного с использованием праймеров с SEQ ID NO: 19 и 23, и двух фрагментов гена V2I-2, амплифицированных с использованием праймеров с SEQ ID NO: 24 и 22, и, таким образом, получали матрицу с полинуклеотидом в 1,8 т.п.о. Полученные генные фрагменты расщепляли при помощи XbaI и клонировали в линеаризованный вектор pDZ, который уже был расщеплен при помощи XbaI, с использованием лигазы Т4, и, таким образом получали вектор pDZ-impE2(V2I).
Вектор с единичной модификацией G64E в гене ImpE2 получали с использованием штамма АТСС6872 в качестве матрицы вместе с праймерами с SEQ ID NO: 19 и 25 и праймерами с SEQ ID NO: 26 и 22. ПЦР с перекрывающимися праймерами осуществляли с использованием фрагмента гена G64E-1, амплифицированного с использованием праймеров с SEQ ID NO: 19 и 25, и двух фрагментов генов G64E-2, амплифицированных с использованием праймеров с SEQ ID NO: 26 и 22, и, таким образом, получали матрицу с полинуклеотидом в 1,8 т.п.о. Полученные генные фрагменты расщепляли при помощи XbaI и клонировали в линеаризованный вектор pDZ, который уже был расщеплен при помощи XbaI, с использованием лигазы Т4, и, таким образом получали вектор pDZ-impE2(G64E).
Пример 4-3: Восстановление модификаций impE1, impE2 и получение штаммов с единичной модификацией
Плазмиду, полученную в примере 4-1, трансформировали в штамм CJI0323 путем электропорации (с использованием метода трансформации, раскрытого в Appl. Microbiol. Biotechnol. (1999) 52: 541-545). Штаммы, в которых вектор был встроен в хромосому путем рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Восстановление модификации в окончательно трансформированных штаммах подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NO: 15 и 16 с последующим анализом путем секвенирования нуклеотидов. Полученный штамм был назван CJI0323_impE1E2(WT).
Каждый из трех типов плазмид, полученных в примере 4-2, трансформировали в штамм CJI0323_impE1E2(WT) путем электропорации (с использованием метода трансформации, раскрытого ъАрр1. Microbiol. Biotechnol. (1999) 52: 541-545). Штаммы, в которых вектор был встроен в хромосому путем рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Внедрение модификации в окончательно трансформированные штаммы подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NO: 15 и 16 с последующим анализом путем секвенирования нуклеотидов. Отобранные штаммы были названы CJI0323_impE1(E164K), CJI0323_impE2(V2I) и CJI0323_impE2(G64E).
Штаммы Corynebacterium stationis CJI0323_impE1(E164K), Corynebacterium stationis CJI0323_impE2(V2I) и Corynebacterium stationis CJI0323_impE2(G64E) были депонированы в соответствии с Будапештским договором в Корейский центр культур микроорганизмов (Korean Culture Center of Microorganisms; KKCCM) 2 ноября 2018 года. Кроме того, этим штаммам были присвоены номера KCCM12359P, KCCM12360P и KCCM12361P, соответственно.
Пример 4-4: Получение штаммов, модифицированных путем интеграции impE1 и impE2
Плазмиды pDZ-impE2(V2I) и pDZ-impE2(G64E), полученные в примере 4-2, трансформировали в штамм CJI0323_impE1(E164K) путем электропорации (с использованием метода трансформации, раскрытого в Appl. Microbiol. Biotechnol. (1999) 52: 541-545). Штаммы, в которых векторы были встроены в хромосому путем рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Внедрение модификации в окончательно трансформированные штаммы подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NO: 15 и 16 с последующим анализом путем секвенирования нуклеотидов. Полученные штаммы были названы CJI0323_impE1(E164K)_impE2(V2I) и CJI0323_impE1(164K)_impE2(G64E).
Плазмиду pDZ-impE2(G64E) трансформировали в штамм CJI0323_impE2(V2I) путем электропорации (с использованием метода трансформации, раскрытого в Appl. Microbiol. Biotechnol. (1999) 52: 541-545). Штаммы, в которых вектор был встроен в хромосому путем рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Внедрение модификации в окончательно трансформированные штаммы подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NO: 15 и 16 с последующим анализом путем секвенирования нуклеотидов. Отобранный штамм был назван CJI0323_impE2(V2I)(G64E).
Пример 4-5: Оценка штаммов с модификациями impE1, impE2
Среду для посева (2 мл) разливали в пробирки для тестирования (диаметр: 18 мм), которые затем автоклавировали, каждую из которых инокулировали CJI0323_impE1E2(WT), CJI0323_impE1(E164K), CJI0323_impE2(V2I), CJI0323_impE2(G64E), CJI0323_impE1(E164K)_impE2(V2I), CJI0323_impE1(E164K)_impE2(G64E) и CJI0323_impE2(V2I)(G64E), культивировали при перемешивании при 30°С в течение 24 часов и использовали в качестве растворов посевных культур. Ферментационную среду (29 мл) разливали в колбы Эрленмейера (250 мл) для встряхивания и автоклавировали при 121°С в течение 15 минут. Затем ее инокулировали растворами посевных культур (2 мл) и полученные в результате культуры выращивали в течение 3 суток. Условия выращивания устанавливались следующими: 170 об./мин, 30°С и рН 7,5.
После завершения выращивания количество продуцируемого IMP измеряли при помощи HPLC, и результаты для культуры представлены в таблице 7 ниже.
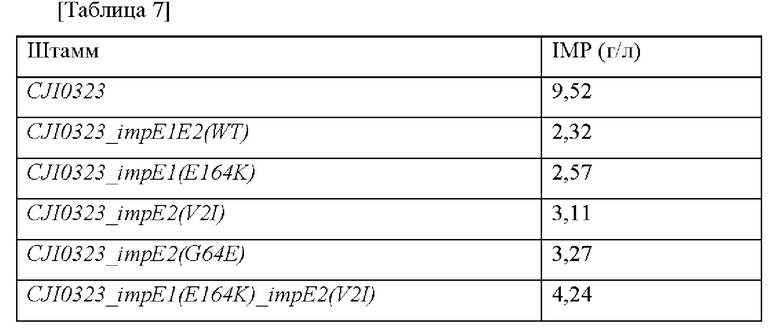
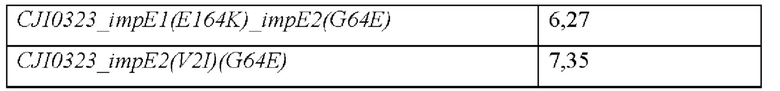
Как показано выше, было подтверждено в отношении каждой позиции модификации, что как один тип модификации, так и интегрирование двух типов модификаций, а также интегрирование трех типов модификаций, вовлечены в экспорт IMP. Соответственно, в микроорганизме рода Corynebacterium, который не продуцирует IMP или продуцирует лишь небольшое его количество, увеличение количественного пордуцирования IMP вследствие модификаций белка ImpE в соответствии с описанием настоящего изобретения может быть интерпретировано как весьма значительное.
Пример 5: Замена аминокислот в модификациях impE1, impE2 на другие аминокислоты
Пример 5-1: Получение векторов для замещающей вставки аминокислот в модификациях impE1, impE2
Для того, чтобы подтвердить позиционную важность, получали типичные три вида модификаций (т.е. impE1(E164K), impE2(V2I) и impE2(G64E)), обладающие усиленными способностями продуцировать IMP, как установлено согласно вышеприведенным результатам, и вектор для внедрения модификаций (например, модификацию, заменяющую 164-ю аминокислоту в аминокислотной последовательности impE1, 2-ю аминокислоту в аминокислотной последовательности impE2 и 64-ю аминокислоту в аминокислотной последовательности impE2 на другую аминокислоту).
Во-первых, процедура получения вектора для внедрения модификации ImpE1(E164K) является следующей.
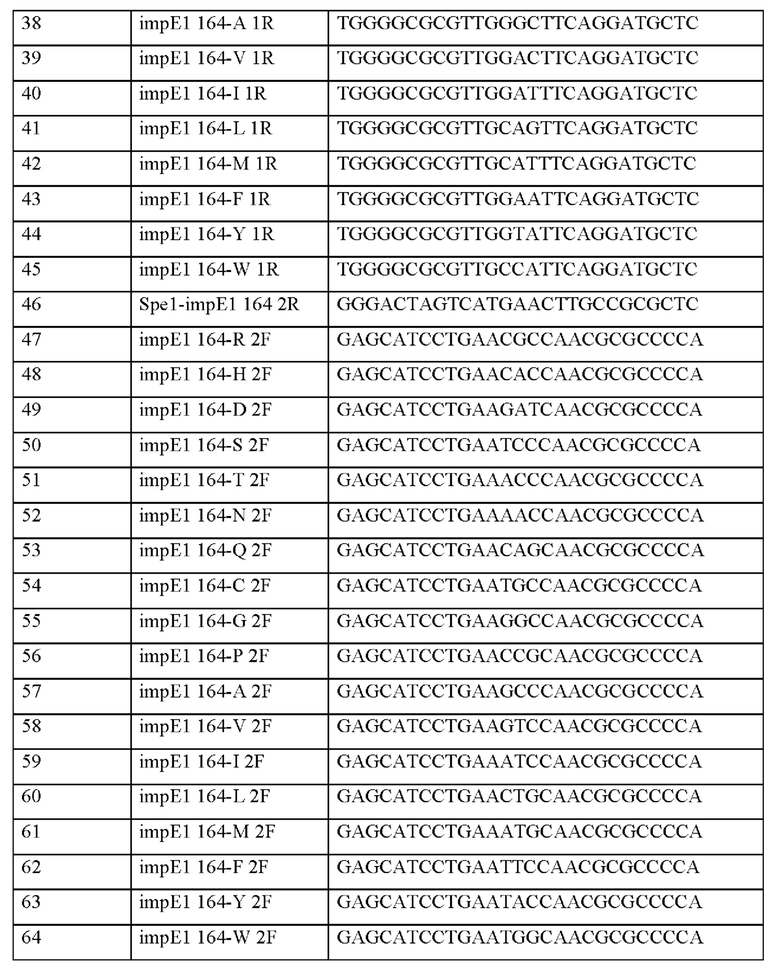
На основе приведенных полинуклеотидных последовательностей выделяли хромосомные гены Corynebacterium stationis CJI0323, и генные фрагменты получали путем осуществления ПЦР с использованием хромосомной ДНК Corynebacterium stationis CJI0323 в качестве матрицы вместе с парами праймеров из праймера с SEQ ID NO: 27 и каждым из праймеров с SEQ ID NO: 28-45. ПЦР осуществляли путем исходной денатурации при 94°С в течение 5 минут; 20 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут. В результате получали 18 типов полинуклеотидов с 0,7 т.п.о. Затем выделяли хромосомные гены Corynebacterium stationis CJI0323, и генные фрагменты получали путем осуществления ПЦР с использованием хромосомной ДНК Corynebacterium stationis CJI0323 в качестве матрицы вместе с парами праймеров из праймером с SEQ ID NO: 46 и каждым из праймеров с SEQ ID NO: 47-64. ПЦР осуществляли путем исходной денатурации при 94°С в течение 5 минут; 20 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут. В результате получали 18 типов полинуклеотидов с 0,7 т.п.о.
ПЦР с перекрывающимися праймерами осуществляли с использованием двух фрагментов, полученных в соответствии с вышеуказанными результатами, в качестве матрицы, и, таким образом получали 18 типов полинуклеотидов с 1,4 т.п.о, используемых в качестве матриц. Полученные фрагменты генов расщепляли при помощи фермента рестрикции SpeI, лигировали с линеаризованным вектором pDZ, который уже был расщеплен ферментом рестрикции XbaI, трансформировали в Е. coli DH5a, и трансформанты высевали на твердую среду LB, содержащую канамицин (25 мг/л).
Информация о последовательности праймеров, используемых для получения вектора, представлена в таблице 8 ниже.

После отбора при помощи ПЦР колоний, трансформированных вектором, в который встроен целевой ген, получали плазмиды с использованием общеизвестного метода плазмидной экстракции. Информация о полученных плазмидах представлена в таблице 9 ниже.
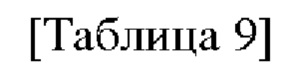

Во вторых, способ получения вектора для внедрения ImpE2(V2I) является следующим.
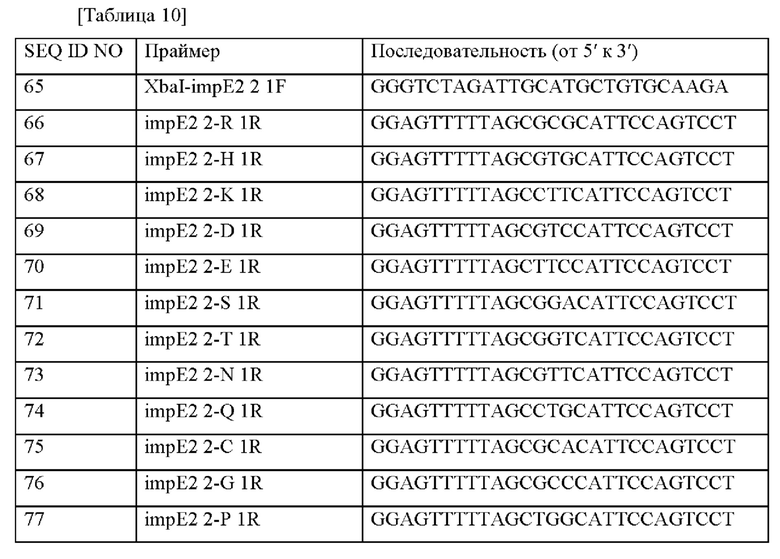
На основе приведенных полинуклеотидных последовательностей выделяли хромосомные гены Corynebacterium stationis CJI0323, и генные фрагменты получали путем осуществления ПЦР с использованием хромосомной ДНК Corynebacterium stationis CJI0323 в качестве матрицы вместе с парами праймеров из праймера с SEQ ID NO: 65 и каждым из праймеров с SEQ ID NO: 66-83. ПЦР осуществляли путем исходной денатурации при 94°С в течение 5 минут; 20 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут. В результате получали 18 видов полинуклеотидов с 0,7 т.п.о. Затем выделяли хромосомные гены Corynebacterium stationis CJI0323, и генные фрагменты получали путем осуществления ПЦР с использованием хромосомной ДНК Corynebacterium stationis CJI0323 в качестве матрицы вместе с парами праймеров из праймера с SEQ ID NO: 84 и каждым из праймеров с SEQ ID NO: 85-102. ПЦР осуществляли путем исходной денатурации при 94°С в течение 5 минут; 20 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут. В результате получали 18 видов полинуклеотидов с 0.7 т.п.о.
ПЦР с перекрывающимися праймерами осуществляли с использованием двух фрагментов, полученных в соответствии с вышеприведенными результатами в качестве матрицы, и, таким образом получали 18 типов полинуклеотидов с 1,4 т.п.о, используемых в качестве матриц. Полученные фрагменты генов расщепляли при помощи фермента рестрикции XbaI, лигировали с линеаризованным pDZ вектором, который уже был расщеплен ферментом рестрикции XbaI, трансформировали в Е. coli DH5a, и трансформанты высевали на твердую среду LB, содержащую канамицин (25 мг/л).
Информация о последовательности праймеров, используемых для получения вектора, представлена в таблице 10 ниже.
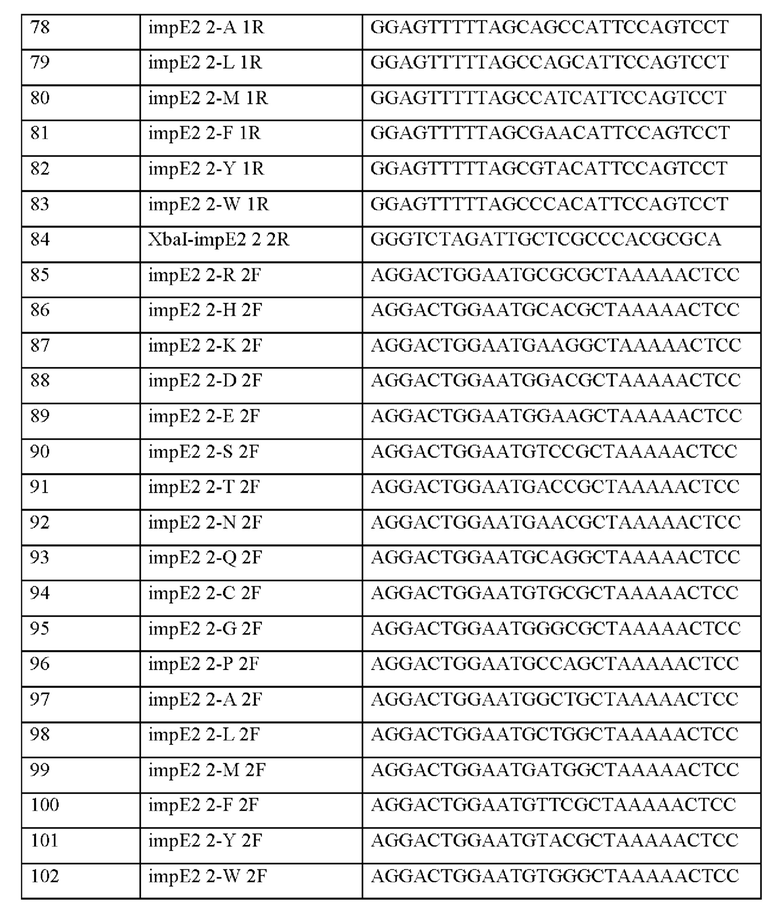
После отбора при помощи ПЦР колоний, трансформированных вектором, в который встроен целевой ген, получали плазмиды с использованием общеизвестного метода плазмидной экстракции. Информация о полученных плазмидах представлена в таблице 11 ниже.

В заключение, способ получения вектора для внедрения ImpE2(G64E) является следующим.
На основе приведенных полинуклеотидных последовательностей выделяли хромосомные гены Corynebacterium stationis CJI0323, и генные фрагменты получали путем осуществления ПЦР с использованием хромосомной ДНК Corynebacterium stationis CJI0323 в качестве матрицы с денатурацией при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации вместе с парами праймеров из праймера с SEQ ID NO: 103 и каждым из праймеров с SEQ ID NO: 104-121. ПЦР осуществляли путем исходной денатурации при 94°С в течение 5 минут; 20 циклов, состоящих из денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут. В результате получали 18 типов полинуклеотидов с 1 т.п.о. Затем выделяли хромосомные гены Corynebacterium stationis CJI0323, и генные фрагменты получали путем осуществления ПЦР с использованием хромосомной ДНК Corynebacterium stationis CJI0323 в качестве матрицы вместе с парами праймеров из праймера с SEQ ID NO: 84 и каждым из праймеров с SEQ ID NO: 85-102. ПЦР осуществляли путем исходной денатурации при 94°С в течение 5 минут; 20 циклов, состоящих из полимеризации при 72°С в течение 1 минуты; и окончательной полимеризации при 72°С в течение 5 минут. В результате получали 18 типов полинуклеотидов с 1 т.п.о.
ПЦР с перекрывающимися праймерами осуществляли с использованием двух фрагментов, полученных в соответствии с вышеприведенными результатами в качестве матрицы, и, таким образом получали 18 типов полинуклеотидов с 2 т.п.о, используемых в качестве матриц. Полученные фрагменты генов расщепляли при помощи фермента рестрикции XbaI, лигировали с линеаризованным вектором pDZ, который уже был расщеплен ферментом рестрикции XbaI, трансформировали в Е. coli DH5a, и трансформанты высевали на твердую среду LB, содержащую канамицин (25 мг/л).
Информация о последовательности праймеров, используемых для получения вектора, представлена в таблице 12 ниже.
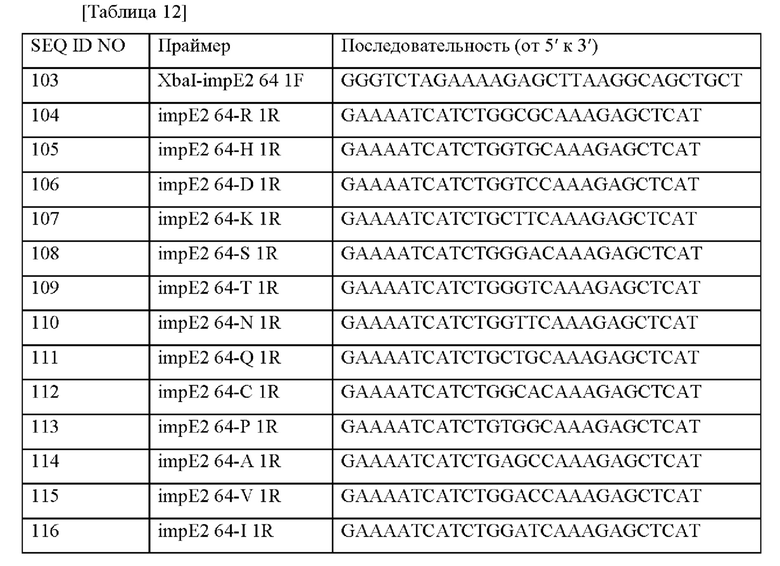
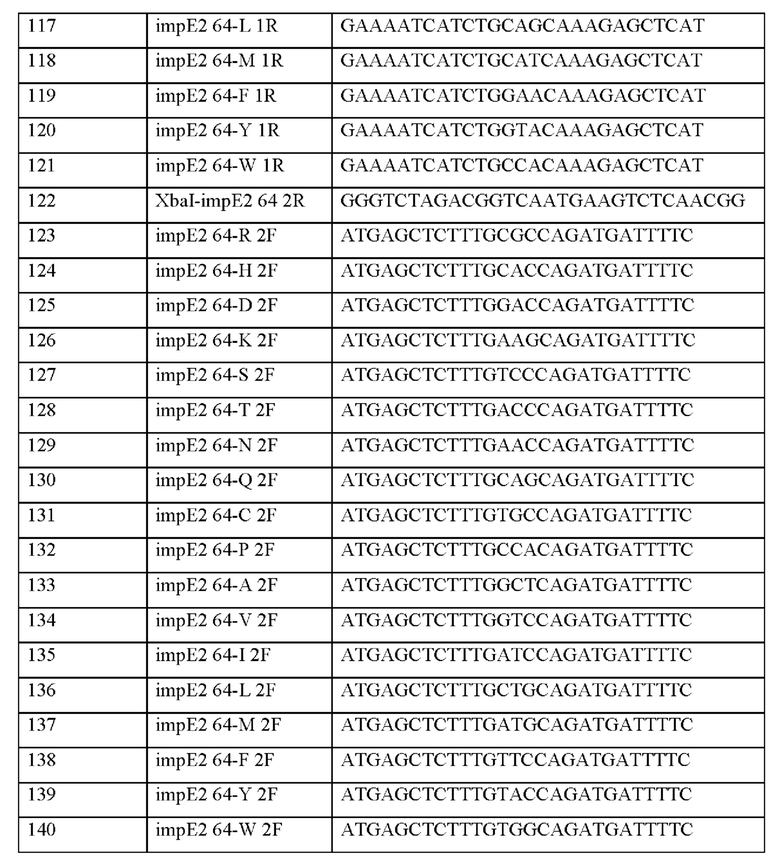
После отбора при помощи ПЦР колоний, трансформированных вектором, в который встроен целевой ген, получали плазмиды с использованием общеизвестного метода плазмидной экстракции. Информация о полученных плазмидах представлена в таблице 13 ниже.


Пример 5-2: Получение штаммов, в которых аминокислоты в позициях модифицированных продуктов (ImpE1, ImpE2) заменены на другие аминокислоты, и сравнение способности продуцировать IMP
54 типа плазмид, полученных в примере 5-1, трансформировали в штамм CJI0323. Штаммы, в которых вектор был встроен в хромосому путем рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Внедрение модификации в окончательно трансформированные штаммы подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NOS: 15 и 16 с последующим анализом путем секвенирования нуклеотидов. Наименования штаммов в соответствии с внедренными модификациями представлены в таблице 14 ниже.



Культивирование осуществляли как в примере 1, и анализировали концентрацию IMP, продуцируемую ими (Таблица 15).


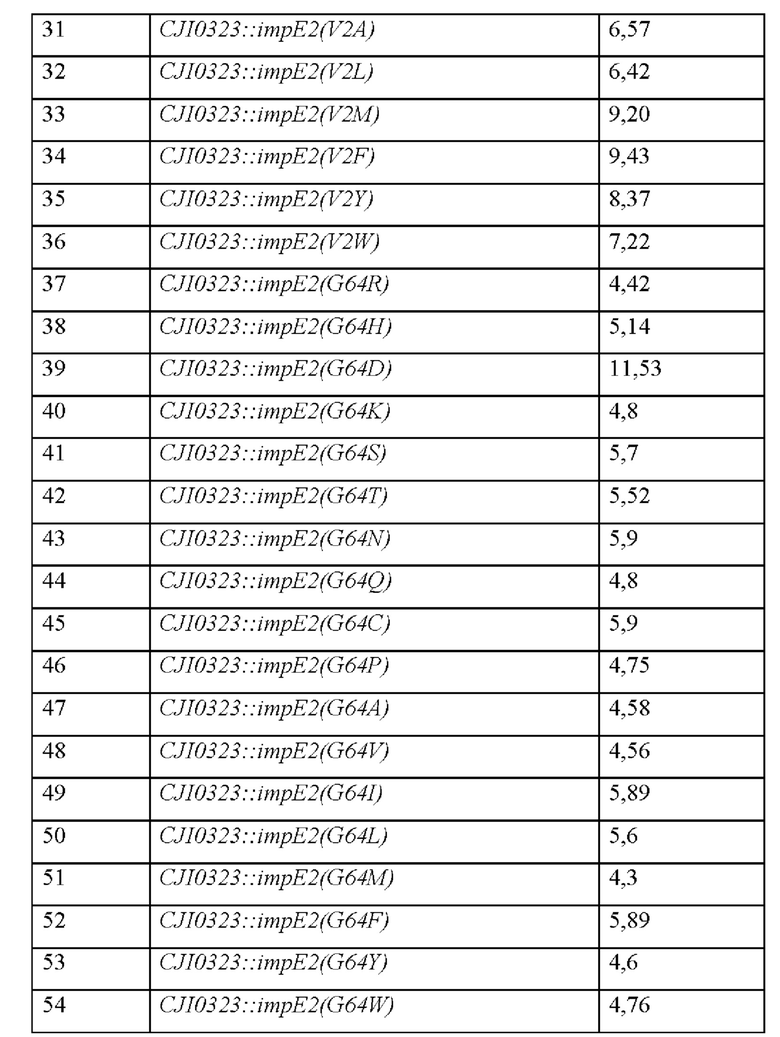
Как показано выше, все модифицированные штаммы демонстрировали увеличение способности продуцировать IMP по сравнению с каждым из контрольных штаммов, и, таким образом подтверждено, что три позиции модификации представляют собой важные сайты, которые оказывают существенное влияние на увеличение способности белка ImpE в отношении экспорта IMP.
Пример 6: Внедрение модификаций impE1, impE2, основанных на штаммах, продуцирующих IMP
Пример 6-1: Получение штаммов с модификациями impE1, impE2, основанных на штаммах, продуцирующих IMP
Для подтверждения влияния внедрения модификаций impE1 и impE2 получали штамм, продуцирующий IMP, в котором были ослаблены активности аденилосукцинатсинтетазы и IMP-дегидрогеназы, соответствующие метаболическому пути разрушения IMP в штамме АТСС6872. Инициирующий кодон был изменен путем замены первого основания с 'а' на 't' в каждой нуклеотидной последовательности двух генов purA и guaB, которые кодируют эти два фермента. Штамм, в котором экспрессия двух генов была ослаблена в штамме АТСС6872, был назван CJI9088. Векторы pDZ-impE1(E164K), pDZ-impE2(V2I) и pDZ-impE2(G64E), полученные в примере 4-2, трансформировали в штамм CJI9088 путем электропорации, и вектор pDZ-impE2(G64D), полученный в примере 5-1, трансформировали в штамм CJI9088_impE1(E164K)_impE2(V2I) путем электропорации. Штаммы, в которых векторы были внедрены в хромосому путем рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин (25 мг/л). Отобранные первичные штаммы подвергали второму кроссинговеру. Внедрение модификации в окончательно трансформированные штаммы подтверждали путем осуществления ПЦР с использованием пары праймеров с SEQ ID NO: 15 и 16 с последующим анализом путем секвенирования нуклеотидов.
Оценивали способность полученных штаммов (т.е. CJI9088, CJI9088_impE1(E164K), CJI9088_impE2(V2I), CJI9088_impE2(G64E) и CJI9088_impE1(E164K)_impE2(V2I)(G64D)) продуцировать IMP. После завершения культивирования количественную продукцию IMP измеряли при помощи HPLC, и результаты представлены в таблице 16 ниже.

После подтверждения количества IMP, накопленного в культуральной среде, было подтверждено, что эти штаммы демонстрируют увеличение продукции IMP по меньшей мере на 61% с максимальным увеличением на 727% по сравнению с родительским штаммом CJ9088. Соответственно, количественное увеличение продукции IMP вследствие модификаций белка ImpE в соответствии с описанием настоящего изобретения может быть интерпретировано как весьма значительное.
Исходя из вышеизложенного, специалист в данной области техники, к которой относится настоящее изобретение, будет способен понять, что настоящее изобретение может быть воплощено в других конкретных формах без модификации технических принципов или существенных характеристик настоящего изобретения. В этой связи раскрытые в настоящем описании типичные воплощения приведены только для иллюстративных целей и их не следует рассматривать, как ограничивающие объем настоящего изобретения. Наоборот, предполагается, что настоящее изобретение охватывает не только эти типичные воплощения, но также и различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящего изобретения, как определено в прилагаемой формуле изобретения.
Изобретение относится к биотехнологии, в частности к новому полипептиду, вовлеченному во внеклеточный экспорт инозин-5’-монофосфата, содержащему его микроорганизму, способу получения инозин-5'-монофосфата с его использованием и способу увеличения экспорта инозин-5'-монофосфата. Изобретение позволяет увеличить получение инозин-5’-монофосфата. 5 н. и 3 з.п. ф-лы, 16 табл., 6 пр.
1. Вариант белка, вовлеченный во внеклеточный экспорт инозин-5’-монофосфата, имеющий аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2, где
(A) 164-я аминокислота в аминокислотной последовательности SEQ ID NO: 1 заменена на другую аминокислоту; или
(B) (i) 2-я аминокислота, (ii) 64-я аминокислота или (iii) 2-я аминокислота и 64-я аминокислота аминокислотной последовательности SEQ ID NO: 2 заменена на другую аминокислоту.
2. Вариант белка по п. 1, где
(A) 164-я аминокислота в аминокислотной последовательности SEQ ID NO: 1 заменена на аминокислоту, выбранную из группы, состоящей из лизина, аргинина, аспарагина, глицина, треонина и пролина;
или (B) (i) 2-я аминокислота в аминокислотной последовательности SEQ ID NO: 2 заменена на аминокислоту, выбранную из группы, состоящей из изолейцина, фенилаланина, метионина, глутаминовой кислоты, гистидина и аспарагина; (ii) 64-я аминокислота в аминокислотной последовательности SEQ ID NO: 2 заменена на аминокислоту, выбранную из группы, состоящей из аспартата, глутаминовой кислоты, аспарагина, цистеина, изолейцина и фенилаланина; или (iii) каждая из 2-й аминокислоты и 64-й аминокислоты в аминокислотной последовательности SEQ ID NO: 2 заменена на аминокислоту, выбранную из группы, состоящей из метионина, глутаминовой кислоты, гистидина, аспарагина, аспартата, цистеина, изолейцина и фенилаланина.
3. Полинуклеотид, кодирующий вариант белка по п. 1 или 2.
4. Экспрессионный вектор, содержащий полинуклеотид по п. 3.
5. Микроорганизм рода Corynebacterium, продуцирующий инозин-5’-монофосфат, который содержит вариант белка по п. 1 или 2 либо вектор, содержащий полинуклеотид, кодирующий вариант белка по п. 1 или 2.
6. Микроорганизм рода Corynebacterium по п. 5, где микроорганизм рода Corynebacterium представляет собой Corynebacterium stationis.
7. Способ получения инозин-5’-монофосфата, включающий культивирование микроорганизма рода Corynebacterium по п. 5 в среде и выделение инозин-5’-монофосфата из микроорганизма или среды.
8. Способ по п. 7, где микроорганизм рода Corynebacterium представляет собой Corynebacterium stationis.
| Штамм СоRYNевастеRIUм аммоNIаGеNеS - продуцент инозин-5 @ -монофосфата | 1991 |
|
SU1806199A3 |
| СПОСОБ ПОЛУЧЕНИЯ ИНОЗИНА И ИНОЗИН 5'-МОНОФОСФАТА, ШТАММ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ BACILLUS - ПРОДУЦЕНТ ИНОЗИНА (ВАРИАНТЫ) | 2002 |
|
RU2260040C2 |
| NCBI Reference Sequence: WP_066795119.1, 26.08.2016, найдено в Интернете по адресу https://www.ncbi.nlm.nih.gov/protein/1055045151?sat=46&satkey=154081494. | |||

