Техническая область
Настоящее изобретение относится к антителам против NR10 и к фармацевтическим композициям, содержащим антитело против NR10.
Уровень техники
Многие цитокины известны в качестве гуморальных факторов, вовлеченных в рост и дифференцировку различных типов клеток, или в активацию функций дифференцированных зрелых клеток. Стимулированные цитокинами клетки продуцируют различные типы цитокинов, тем самым образуя в организме сети из множества цитокинов. Биологический гомеостаз поддерживается посредством тонкого равновесия взаимной регуляции цитокинов в этих сетях. Полагают, что многие воспалительные заболевания являются следствием нарушений в таких цитокиновых сетях. Таким образом, значительное внимание привлекает анти-цитокиновая терапия на основе моноклональных антител. Например, было показано, что антитела против TNF и антитела против рецептора для IL-6 являются клинически высокоэффективными. С другой стороны, существует множество примеров неуспеха, когда отсутствуют терапевтические эффекты, если блокируют только один цитокин, такой как IL-4, вследствие активации компенсаторных каскадов при конкретных патологических состояниях.
Авторы настоящего изобретения добились успеха в выделении нового рецептора цитокинов NR10, который является высоко гомологичным gp130, рецептору для передачи сигнала IL-6 (патентный документ 1). NR10 образует гетеродимер с рецептором онкостатина M (OSMR) и функционирует в качестве рецептора IL-31 (непатентный документ 1). В отношении IL-31, было описано, что у трансгенных мышей, сверхэкспрессирующих IL-31, самопроизвольно развивается дерматит с зудом (патентный документ 2).
Антитела, которые связываются с NR10 и ингибируют связывание между NR10 и IL-31, могут быть эффективными в отношении лечения воспалительных заболеваний. Для клинического применения требуется, чтобы антитела против NR10 имели низкую иммуногенность. Более того, для достижения высоких терапевтических эффектов, желательными являются антитела с сильной связывающей или нейтрализующей активностью в отношении NR10.
Документы уровня техники для настоящего изобретения представлены ниже.
Патентный документ 1: WO00/75314
Патентный документ 2: WO03/060090
Непатентный документ 1: IL-31 is associated with cutaneous lymphocyte antigen-positive skin homing T cells in patients with atopic dermatitis., J Allergy Clin Immunol. 2006 Feb; 117(2): 418-25.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[Проблемы, решаемые изобретением]
Ввиду обстоятельств, описанных выше, было выполнено настоящее изобретение. Задачей настоящего изобретения является предоставление антитела против NR10, и фармацевтических композиций, содержащих антитело против NR10.
[Средства для решения проблем]
Авторы настоящего изобретения провели специализированные исследования для выполнения задачи, описанной выше. Авторы настоящего изобретения добились успеха в получении антител против NR10, имеющих эффективную нейтрализующую активность против NR10. Более того, авторы настоящего изобретения добились успеха в гуманизации антител при сохранении их активности. Также авторы настоящего изобретения успешно продуцировали антитела с улучшенной фармакокинетикой, усиленной антигенсвязывающей активностью, повышенной стабильностью и/или сниженным риском иммуногенности. Эти антитела пригодны в качестве лекарственных средств от воспалительных заболеваний.
Настоящее изобретение относится к антителам против NR10, и фармацевтическим композициям, содержащим антитело против NR10. Более конкретно, настоящее изобретение включает:
[1] антитело, которое распознает домен 1 NR10;
[2] антитело согласно [1], которое обладает нейтрализующей активностью;
[3] антитело согласно [1] или [2], которое представляет собой гуманизированное антитело;
[4] антитело против NR10, которое представляет собой любое из:
(1) антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 1, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 2, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 3;
(2) антитело, содержащее вариабельную область тяжелой цепи SEQ ID NO: 4;
(3) антитело, содержащее вариабельную область легкой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 5, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 6, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 7;
(4) антитело, содержащее вариабельную область легкой цепи SEQ ID NO: 8;
(5) антитело, содержащее вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитело, содержащее вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (1)-(6), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(6); и
(8) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (1)-(7);
[5] антитело против NR10, которое представляет собой любое из:
(1) антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 9, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 10, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 11;
(2) антитело, содержащее вариабельную область тяжелой цепи SEQ ID NO: 12;
(3) антитело, содержащее вариабельную область легкой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 13, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 14, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 15;
(4) антитело, содержащее вариабельную область легкой цепи SEQ ID NO: 16;
(5) антитело, содержащее вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитело, содержащее вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (1)-(6), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(6); и
(8) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (1)-(7);
[6] антитело против NR10, которое представляет собой любое из:
(1) антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 17, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 18, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 19;
(2) антитело, содержащее вариабельную область тяжелой цепи SEQ ID NO: 20;
(3) антитело, содержащее вариабельную область легкой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 21, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 22, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 23;
(4) антитело, содержащее вариабельную область легкой цепи SEQ ID NO: 24;
(5) антитело, содержащее вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитело, содержащее вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (1)-(6), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(6); и
(8) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (1)-(7);
[7] антитело против NR10, которое представляет собой любое из:
(1) антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 25, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 26, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 27;
(2) антитело, содержащее вариабельную область тяжелой цепи SEQ ID NO: 28;
(3) антитело, содержащее вариабельную область легкой цепи, которая содержит CDR1, содержащую аминокислотную последовательность SEQ ID NO: 29, CDR2, содержащую аминокислотную последовательность SEQ ID NO: 30, и CDR3, содержащую аминокислотную последовательность SEQ ID NO: 31;
(4) антитело, содержащее вариабельную область легкой цепи SEQ ID NO: 32;
(5) антитело, содержащее вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитело, содержащее вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (1)-(6), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(6); и
(8) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (1)-(7);
[8] антитело или вариабельная область антитела, которые представляют собой любое из:
(1) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 196, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 11 (H17);
(2) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 176, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 11 (H19);
(3) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 196, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 184 (H28, H42);
(4) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 9, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 184 (H30, H44);
(5) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 176, CDR2 с SEQ ID NO: 197, CDR3 с SEQ ID NO: 184 (H34, H46);
(6) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 9, CDR2 с SEQ ID NO: 198, и CDR3 с SEQ ID NO: 184 (H57, H78);
(7) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 176, CDR2 с SEQ ID NO: 198, и CDR3 с SEQ ID NO: 184 (H71, H92);
(8) вариабельная область тяжелой цепи, содержащая CDR1 с SEQ ID NO: 9, CDR2 с SEQ ID NO: 199, и CDR3 с SEQ ID NO: 184 (H97, H98);
(9) вариабельная область легкой цепи, содержащая CDR1 с SEQ ID NO: 200, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L11);
(10) вариабельная область легкой цепи, содержащая CDR1 с SEQ ID NO: 201, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L12);
(11) вариабельная область легкой цепи, содержащая CDR1 с SEQ ID NO: 202, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L17);
(12) вариабельная область легкой цепи, содержащая CDR1 с SEQ ID NO: 203, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L50);
(13) антитело, содержащее вариабельную область тяжелой цепи (3) и вариабельную область легкой цепи (11);
(14) антитело, содержащее вариабельную область тяжелой цепи (4) и вариабельную область легкой цепи (11);
(15) антитело, содержащее вариабельную область тяжелой цепи (5) и вариабельную область легкой цепи (11);
(16) антитело, содержащее вариабельную область тяжелой цепи (6) и вариабельную область легкой цепи (11);
(17) антитело, содержащее вариабельную область тяжелой цепи (7) и вариабельную область легкой цепи (11);
(18) антитело, содержащее вариабельную область тяжелой цепи (8) и вариабельную область легкой цепи (12);
(19) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (13)-(18), которое обладает активностью, эквивалентной активности антитела согласно любому из (13)-(18); и
(20) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (13)-(18);
[9] антитело или вариабельная область антитела, которые представляют собой любое из:
(1) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 204 (H17);
(2) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 205 (H19);
(3) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 206 (H28);
(4) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 207 (H30);
(5) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 208 (H34),
(6) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 209 (H42);
(7) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 210 (H44);
(8) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 211 (H46);
(9) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 212 (H57);
(10) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 213 (H71);
(11) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 214 (H78);
(12) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 215 (H92);
(13) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 216 (H97);
(14) вариабельная область тяжелой цепи, содержащая аминокислотную последовательность SEQ ID NO: 217 (H98);
(15) вариабельная область легкой цепи, содержащая аминокислотную последовательность SEQ ID NO: 218 (L11);
(16) вариабельная область легкой цепи, содержащая аминокислотную последовательность SEQ ID NO: 219 (L12);
(17) вариабельная область легкой цепи, содержащая аминокислотную последовательность SEQ ID NO: 220 (L17);
(18) вариабельная область легкой цепи, содержащая аминокислотную последовательность SEQ ID NO: 221 (L50);
(19) антитело, содержащее вариабельную область тяжелой цепи (3) и вариабельную область легкой цепи (17) (H28L17);
(20) антитело, содержащее вариабельную область тяжелой цепи (4) и вариабельную область легкой цепи (17) (H30L17);
(21) антитело, содержащее вариабельную область тяжелой цепи (5) и вариабельную область легкой цепи (17) (H34L17);
(22) антитело, содержащее вариабельную область тяжелой цепи (6) и вариабельную область легкой цепи (17) (H42L17);
(23) антитело, содержащее вариабельную область тяжелой цепи (7) и вариабельную область легкой цепи (17) (H44L17);
(24) антитело, содержащее вариабельную область тяжелой цепи (8) и вариабельную область легкой цепи (17) (H46L17);
(25) антитело, содержащее вариабельную область тяжелой цепи (9) и вариабельную область легкой цепи (17) (H57L17);
(26) антитело, содержащее вариабельную область тяжелой цепи (10) и вариабельную область легкой цепи (17) (H71L17);
(27) антитело, содержащее вариабельную область тяжелой цепи (11) и вариабельную область легкой цепи (17) (H78L17);
(28) антитело, содержащее вариабельную область тяжелой цепи (12) и вариабельную область легкой цепи (17) (H92L17);
(29) антитело, содержащее вариабельную область тяжелой цепи (13) и вариабельную область легкой цепи (18) (H97L50);
(30) антитело, содержащее вариабельную область тяжелой цепи (14) и вариабельную область легкой цепи (18) (H98L50),
(31) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (19)-(30), которое обладает активностью, эквивалентной активности антитела согласно любому из (19)-(30); и
(32) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (19)-(30);
[10] антитело против NR10 согласно любому из [4]-[9], которое представляет собой гуманизированное антитело;
[11] антитело, тяжелая цепь антитела или легкая цепь антитела, которые представляют собой любое из:
(1) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 222 (H17);
(2) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 223 (H19);
(3) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 224 (H28);
(4) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 225 (H30);
(5) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 226 (H34);
(6) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 227 (H42);
(7) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 228 (H44);
(8) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 229 (H46);
(9) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 230 (H57);
(10) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 231 (H71);
(11) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 232 (H78);
(12) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 233 (H92);
(13) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 234 (H97);
(14) тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 235 (H98);
(15) легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 236 (L11);
(16) легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 237 (L12);
(17) легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 238 (L17);
(18) легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 239 (L50);
(19) антитело, содержащее тяжелую цепь (3) и легкую цепь (17) (H28L17);
(20) антитело, содержащее тяжелую цепь (4) и легкую цепь (17) (H30L17);
(21) антитело, содержащее тяжелую цепь (5) и легкую цепь (17) (H34L17);
(22) антитело, содержащее тяжелую цепь (6) и легкую цепь (17) (H42L17);
(23) антитело, содержащее тяжелую цепь (7) и легкую цепь (17) (H44L17);
(24) антитело, содержащее тяжелую цепь (8) и легкую цепь (17) (H46L17);
(25) антитело, содержащее тяжелую цепь (9) и легкую цепь (17) (H57L17);
(26) антитело, содержащее тяжелую цепь (10) и легкую цепь (17) (H71L17);
(27) антитело, содержащее тяжелую цепь (11) и легкую цепь (17) (H78L17);
(28) антитело, содержащее тяжелую цепь (12) и легкую цепь (17) (H92L17);
(29) антитело, содержащее тяжелую цепь (13) и легкую цепь (18) (H97L50);
(30) антитело, содержащее тяжелую цепь (14) и легкую цепь (18) (H98L50);
(31) антитело, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитело согласно любому из (19)-(30), которое обладает активностью, эквивалентной активности антитела согласно любому из (19)-(30); и
(32) антитело, которое связывается с тем же эпитопом, с которым связывается антитело согласно любому из (19)-(30);
[12] фармацевтическая композиция, содержащая антитело согласно любому из [1]-[11];
[13] фармацевтический композиция согласно [12], которая представляет собой средство для лечения воспалительного заболевания;
[14] способ лечения или профилактики воспалительного заболевания, который включает стадию введения антитела согласно любому из [1]-[11]; и
[15] применение антитела согласно любому из [1]-[11] для изготовления лекарственного средства против воспалительного заболевания.
Краткое описание рисунков
На фиг. 1 представлены аминокислотные последовательности вариабельных областей тяжелых цепей антител мыши NS18, NS22, NS23 и NS33.
На фиг. 2 представлены аминокислотные последовательности вариабельных областей легких цепей антител мыши NS18, NS22, NS23 и NS33.
На фиг. 3 представлен график, на котором показано ингибирование роста клеток hNR10/hOSMR/BaF3 супернатантами клеточных культур.
На фиг. 4 представлен график, на котором показано ингибирование роста клеток cynNR10/cynOSMR/BaF3 супернатантами клеточных культур.
На фиг. 5 представлен график, на котором показана оценка активности химерного NS22 (BaF).
На фиг. 6 представлен график, на котором показана оценка активности химерного NS22 (DU-145).
На фиг. 7 представлен график, на котором показана оценка конкуренции химерного NS22 с IL-31.
На фиг. 8 представлен график, на котором показана конкурентная активность антитела против NR10 в отношении связывания NR10.
На фиг. 9 представлен набор графиков, на которых показана оценка конкуренции гуманизированного NS22 (H0L0) с IL-31.
На фиг. 10 представлен эффект константной области гуманизированного антитела против NR10 H0L0 на гетерогенность, оцененный катионообменной хроматографией.
На фиг. 11 представлен набор графиков, на котором показана оценка конкуренции мутантов гуманизированного антитела против NR10, изоэлектрическая точка вариабельных областей которых снижена без существенного снижения связывания NR10, с IL-31.
На фиг. 12 представлен эффект константной области антитела против рецептора IL-6 на гетерогенность, оцененный катионообменной хроматографией.
На фиг. 13 представлен эффект константной области антитела против рецептора IL-6 на пик денатурации, оцененный посредством DSC.
На фиг. 14 представлен эффект новой константной области M14 на гетерогенность в антителе против рецептора IL-6, оцененный катионообменной хроматографией.
На фиг. 15 представлен эффект новой константной области M58 на гетерогенность в антителе против рецептора IL-6, оцененный катионообменной хроматографией.
На фиг. 16 представлен эффект новой константной области M58 на пик денатурации в антителе против рецептора IL-6, оцененный посредством DSC.
На фиг. 17 представлен результат оценки удержания huPM1-IgG1 и huPM1-M58 в плазме трансгенных мышей с FcRn человека.
На фиг. 18 представлена биологическая активность каждого антитела, оцененная с использованием BaF/NR10.
На фиг. 19 представлен анализ катионообменной хроматографией образцов с термическим ускорением (пунктирная линия) и без ускорения (сплошная линия) для каждого модифицированного антитела для сравнения образования продуктов деградации до и после термического ускорения. Стрелками указано положение пика основного компонента, который был изменен.
На фиг. 20 представлен набор графиков, на которых показана оценка (BaF) активности каждого варианта.
На фиг. 21 представлен график, на котором показана оценка (BaF) активности Ha401La402 и H0L0.
На фиг. 22 представлен график, на котором показана оценка (BaF) активности H17L11 и H0L0.
На фиг. 23 представлен график, на котором показана оценка (BaF) активности H19L12 и H0L0.
На фиг. 24 представлен график, на котором показана биологическая активность H0L12 и H0L17, оцененная с использованием BaF/NR10.
На фиг. 25-1 представлен набор графиков, на которых показана оценка (BaF) активности каждого варианта.
На фиг. 25-2 представлено продолжение фиг. 25-1.
На фиг. 26 представлена схематическая диаграмма для NR10-ECD дикого типа человека/мыши и химерного NR10-ECD.
На фиг. 27 представлен набор фотографий, на которых показана детекция связывающего домена вестерн-блоттингом. A представляет собой фотографию, на которой показан результат детекции с использованием гуманизированного антитела против NR10 человека; B представляет собой фотографию, на которой показан результат детекции с использованием антитела мыши против NR10 человека; и C представляет собой фотографию, на которой показан результат детекции с использованием антитела против Myc. В случае антитела против NR10 человека, связывание антигена было выявлено только у hhh, hhm и hmm, но не у mmm, mmh и mhm.
На фиг. 28-1 представлена аминокислотная последовательность каждого варианта H0 (SEQ ID NO: 50).
На фиг. 28-2 представлено продолжение фиг. 28-1.
На фиг. 28-3 представлено продолжение фиг. 28-2.
На фиг. 29-1 представлена аминокислотная последовательность каждого варианта L0 (SEQ ID NO: 52).
На фиг. 29-2 представлено продолжение фиг. 29-1.
Способ осуществления изобретения
NR10
NR10 представляет собой белок, который образует гетеродимер с рецептором онкостатина M (OSMR) и функционирует в качестве рецептора IL-31. NR10 также известен как glm-r (J Biol Chem 277 16831-6, 2002), GPL (J Biol Chem 278, 49850-9, 2003), IL31RA (Nat Immunol 5, 752-60, 2004), и т.п. Таким образом, NR10 в настоящем изобретении также включает белки, имеющие такие названия.
В настоящем изобретении NR10 (также называемый IL31RA, GPL или glm-r) конкретно не ограничен с точки зрения его происхождения, и он включает молекулы, происходящие из человека, мышей, обезьян и других млекопитающих. NR10, происходящий из человека, мышей и обезьян, является предпочтительным, и особенно предпочтительным является NR10, происходящий из человека.
Существует множество известных вариантов по сплайсингу происходящего из человека NR10 (WO 00/075314). Среди указанных выше вариантов по сплайсингу, NR10.1 состоит из 662 аминокислот и содержит трансмембранный домен. NR10.2 представляет собой растворимый подобный рецептору белок, состоящий из 252 аминокислот, без трансмембранного домена. При этом известные варианты по сплайсингу NR10, которые функционируют в качестве трансмембранных рецепторных белков, включают NR10.3 и IL-31RAv3. NR10 человека по настоящему изобретению конкретно не ограничен, при условии, что он образует гетеродимер с рецептором онкостатина M (OSMR) и функционирует в качестве рецептора IL-31. Предпочтительный NR10 включает NR10.3 (также называемый ILRAv4 (Nat Immunol 5, 752-60, 2004)) и IL-31RAv3. NR 10.3 (IL31RAv4) состоит из 662 аминокислот (WO 00/075314; Nat Immunol 5, 752-60, 2004) и IL31RAv3 состоит из 732 аминокислот (регистрационный номер GenBank No: NM_139017). Аминокислотная последовательность IL31RAv4 представлена в SEQ ID NO: 79, и аминокислотная последовательность IL31RAv3 представлена в SEQ ID NO: 80. При этом происходящий из мыши NR10 включает белки, содержащие аминокислотную последовательность SEQ ID NO: 81. Кроме того, происходящий из яванского макака NR10 включает белки, содержащие аминокислотную последовательность SEQ ID NO: 66.
Антитела (последовательности)
Предпочтительные варианты осуществления антитела против NR10 по настоящему изобретению включают антитела против NR10 согласно любому из (1)-(8) в (A)-(D), ниже.
(A)NS18
(1) антитела, имеющие вариабельную область тяжелой цепи, которая содержит CDR1, имеющую аминокислотную последовательность SEQ ID NO: 1 (HCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 2 (HCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 3 (HCDR3);
(2) антитела, имеющие вариабельную область тяжелой цепи SEQ ID NO: 4 (VH);
(3) антитела, имеющие вариабельную область легкой цепи, которая содержит CDR1, имеющую аминокислотную последовательность SEQ ID NO: 5 (LCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 6 (LCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 7 (LCDR3);
(4) антитела, имеющие вариабельную область легкой цепи SEQ ID NO: 8 (VL);
(5) антитела, имеющие вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитела, имеющие вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитела, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитела согласно любому из (1)-(6), которые имеют активность, эквивалентную активности антител согласно любому из (1)-(6); и
(8) антитела, которые связываются с тем же эпитопом, с которым связываются антитела согласно любому из (1)-(7).
(B) NS22
(1) антитела, имеющие вариабельную область тяжелой цепи, которая содержит CDR1, имеющую аминокислотную последовательность SEQ ID NO: 9 (HCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 10 (HCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 11 (HCDR3);
(2) антитела, имеющие вариабельную область тяжелой цепи SEQ ID NO: 12 (VH);
(3) антитела, имеющие вариабельную область легкой цепи, которая содержит CDR1, имеющую аминокислотную последовательность SEQ ID NO: 13 (LCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 14 (LCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 15 (LCDR3);
(4) антитела, имеющие вариабельную область легкой цепи SEQ ID NO: 16 (VL);
(5) антитела, имеющие вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитела, имеющие вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитела, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитела согласно любому из (1)-(6), которые имеют активность, эквивалентную активности антител согласно любому из (1)-(6); и
(8) антитела, которые связываются с тем же эпитопом, с которым связываются антитела согласно любому из (1)-(7).
Конкретные примеры описанных выше замены, делеции, добавления и/или вставки одной или нескольких аминокислот конкретно не ограничены и включают, например, следующие модификации.
Замена Ile в положении 3 в CDR1 тяжелой цепи с SEQ ID NO: 9 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Val.
Замена Met в положении 4 в CDR1 тяжелой цепи с SEQ ID NO: 9 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Ile.
Замена Met в положении 4 в CDR1 тяжелой цепи с SEQ ID NO: 9 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Leu.
Замена Ile в положении 3 в CDR1 тяжелой цепи с SEQ ID NO: 9 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Ala.
Замена Leu в положении 1 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Glu.
Замена Asn в положении 3 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Gln в положении 13 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Lys в положении 14 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Gln.
Замена Lys в положении 16 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Gln.
Замена Gly в положении 17 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Lys и Gly в положениях 16 и 17, соответственно, в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Lys в положении 16 на Gln, и Gly в положении 17 на Asp.
Замена Lys, Lys и Gly в положениях 14, 16 и 17, соответственно, в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Lys в положении 14 на Gln, Lys в положении 16 на Gln, и Gly в положении 17 на Asp.
Замена Gln, Lys, Lys и Gly в положениях 13, 14, 16 и 17, соответственно, в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Gln в положении 13 на Asp, Lys в положении 14 на Gln, Lys в положении 16 на Gln, и Gly в положении 17 на Asp.
Замена Ser в положении 10 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Gln в положении 13 в CDR2 тяжелой цепи с SEQ ID NO: 10 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Pro.
Замена Tyr в положении 3 в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Leu.
Замена Met в положении 10 в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Leu.
Замена Asp в положении 11 в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Glu.
Замена Tyr в положении 12 в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Thr и Ser.
Замена Met, Asp и Tyr в положениях 10, 11 и 12, соответственно, в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Met в положении 10 на Leu, Asp в положении 11 на Glu, и Tyr в положении 12 на Thr.
Замена Asp и Tyr в положениях 11 и 12, соответственно, в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Asp в положении 11 на Glu, и Tyr в положении 12 на Thr.
Замена Tyr, Asp и Tyr в положениях 3, 11 и 12, соответственно, в CDR3 тяжелой цепи с SEQ ID NO: 11 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Tyr в положении 3 на Leu, Asp в положении 11 на Glu, и Tyr в положении 12 на Thr или Ser.
Замена Arg в положении 1 в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Gln.
Замена Asn в положении 5 в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Arg и Asn в положениях 1 и 5, соответственно, в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Arg в положении 1 на Gln, и Asn в положении 5 на Asp.
Замена Ser в положении 8 в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Arg.
Замена Leu в положении 10 в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Val.
Замена Ser и Leu в положениях 8 и 10, соответственно, в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Ser в положении 8 на Arg, и Leu в положении 10 на Val.
Замена Thr в положении 2 в CDR1 легкой цепи с SEQ ID NO: 13 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Ala и Ser.
Замена Asn в положении 1 в CDR2 легкой цепи с SEQ ID NO: 14 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Lys в положении 3 в CDR2 легкой цепи с SEQ ID NO: 14 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Gln.
Замена Leu в положении 5 в CDR2 легкой цепи с SEQ ID NO: 14 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Glu.
Замена Lys в положении 7 в CDR2 легкой цепи с SEQ ID NO: 14 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Gln и Asp.
Замена Lys, Leu и Lys в положениях 3, 5 и 7, соответственно, CDR2 легкой цепи с SEQ ID NO: 14 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но предпочтительные примеры включают замену Lys в положении 3 на Gln, Leu в положении 5 на Glu, и Lys в положении 7 на Gln.
Замена Glu в положении 5 в CDR3 легкой цепи с SEQ ID NO: 15 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Ser в положении 6 в CDR3 легкой цепи с SEQ ID NO: 15 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Asp.
Замена Thr в положении 9 в CDR3 легкой цепи с SEQ ID NO: 15 другой аминокислотой. Аминокислота после замены конкретно не ограничена, но ее предпочтительные замены включают Phe.
Каждую из указанных выше замен можно проводить отдельно, или можно проводить несколько замен в комбинации. Более того, указанные выше замены можно комбинировать с другими заменами. Эти замены могут улучшить фармакокинетику антитела (время удержания в плазме), усилить антигенсвязывающую активность, повысить стабильность и/или снизить риск иммуногенности.
В настоящем изобретении конкретные примеры вариабельных областей, имеющих комбинацию указанных выше замен, включают, например, вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 167, и вариабельные области легкой цепи, имеющие аминокислотную последовательность SEQ ID NO: 168. Более того, примеры антител, имеющих комбинацию указанных выше замен, включают, например, антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 167, и вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 168.
Более того, конкретные примеры вариабельных областей тяжелой или легкой цепи, имеющие комбинацию описанных выше замен, включают, например, следующие вариабельные области:
(a) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 196, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 11 (H17);
(b) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 176, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 11 (H19);
(c) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 196, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 184 (H28, H42);
(d) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 9, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 184 (H30, H44);
(e) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 176, CDR2 с SEQ ID NO: 197, и CDR3 с SEQ ID NO: 184 (H34, H46);
(f) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 9, CDR2 с SEQ ID NO: 198, и CDR3 с SEQ ID NO: 184 (H57, H78);
(g) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 176, CDR2 с SEQ ID NO: 198, и CDR3 с SEQ ID NO: 184 (H71, H92);
(h) вариабельные области тяжелой цепи, которые содержат CDR1 с SEQ ID NO: 9, CDR2 с SEQ ID NO: 199, и CDR3 с SEQ ID NO: 184 (H97, H98);
(i) вариабельные области легкой цепи, которые содержат CDR1 с SEQ ID NO: 200, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L11);
(j) вариабельные области легкой цепи, которые содержат CDR1 с SEQ ID NO: 201, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L12);
(k) вариабельные области легкой цепи, которые содержат CDR1 с SEQ ID NO: 202, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L17); и
(l) вариабельные области легкой цепи, которые содержат CDR1 с SEQ ID NO: 203, CDR2 с SEQ ID NO: 170, и CDR3 с SEQ ID NO: 193 (L50).
Более того, конкретные примеры антител, имеющих комбинацию описанных выше замен, включают, например:
(i) антитела, которые содержат вариабельную область тяжелой цепи (c) и вариабельную область легкой цепи (k);
(ii) антитела, которые содержат вариабельную область тяжелой цепи (d) и вариабельную область легкой цепи (k);
(iii) антитела, которые содержат вариабельную область тяжелой цепи (e) и вариабельную область легкой цепи (k);
(iv) антитела, которые содержат вариабельную область тяжелой цепи (f) и вариабельную область легкой цепи (k);
(v) антитела, которые содержат вариабельную область тяжелой цепи (g) и вариабельную область легкой цепи (k); и
(vi) антитела, которые содержат вариабельную область тяжелой цепи (h) и вариабельную область легкой цепи (l).
(C) NS23
(1) антитела, имеющие вариабельную область тяжелой цепи, которая содержит CDR1, имеющую аминокислотную последовательность SEQ ID NO: 17 (HCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 18 (HCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 19 (HCDR3);
(2) антитела, имеющие вариабельную область тяжелой цепи SEQ ID NO: 20 (VH);
(3) антитела, имеющие вариабельную область легкой цепи, которые содержат CDR1, имеющую аминокислотную последовательность SEQ ID NO: 21 (LCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 22 (LCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 23 (LCDR3);
(4) антитела, имеющие вариабельную область легкой цепи SEQ ID NO: 24 (VL);
(5) антитела, имеющие вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитела, имеющие вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитела, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитела согласно любому из (1)-(6), которые имеют активность, эквивалентную активности антител согласно любому из (1)-(6); и
(8) антитела, которые связываются с тем же эпитопом, с которым связываются антитела согласно любому из (1)-(7).
(D) NS33
(1) антитела, имеющие вариабельную область тяжелой цепи, которые содержат CDR1, имеющую аминокислотную последовательность SEQ ID NO: 25 (HCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 26 (HCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 27 (HCDR3);
(2) антитела, имеющие вариабельную область тяжелой цепи SEQ ID NO: 28 (VH);
(3) антитела, имеющие вариабельную область легкой цепи, которые содержат CDR1, имеющую аминокислотную последовательность SEQ ID NO: 29 (LCDR1), CDR2, имеющую аминокислотную последовательность SEQ ID NO: 30 (LCDR2), и CDR3, имеющую аминокислотную последовательность SEQ ID NO: 31 (LCDR3);
(4) антитела, имеющие вариабельную область легкой цепи SEQ ID NO: 32 (VL);
(5) антитела, имеющие вариабельную область тяжелой цепи (1) и вариабельную область легкой цепи (3);
(6) антитела, имеющие вариабельную область тяжелой цепи (2) и вариабельную область легкой цепи (4);
(7) антитела, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитела согласно любому из (1)-(6), которые имеют активность, эквивалентную активности антител согласно любому из (1)-(6); и
(8) антитела, которые связываются с тем же эпитопом, с которым связываются антитела согласно любому из (1)-(7).
Для описанных выше антител согласно (1) или (3) можно использовать любые каркасные области (FR); однако предпочтительно используют FR человека. Более того, для описанных выше антител согласно (1)-(8) можно использовать любые константные области; однако предпочтительно используют константные области человека. Для антител по настоящему изобретению аминокислотную последовательность исходной FR или константной области можно использовать без модификации или после модификации до отличающейся аминокислотной последовательности посредством замены, делеции, добавления и/или вставки одной или нескольких аминокислот.
Аминокислотная последовательность тяжелой цепи вышеописанного NS18 представлена в SEQ ID NO: 34, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена SEQ ID NO: 33. При этом аминокислотная последовательность легкой цепи представлена в SEQ ID NO: 36, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 35.
Аминокислотная последовательность тяжелой цепи NS22 представлена в SEQ ID NO: 38, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 37. При этом аминокислотная последовательность легкой цепи представлена в SEQ ID NO: 40, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 39.
Аминокислотная последовательность тяжелой цепи NS23 представлена в SEQ ID NO: 42, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 41. При этом аминокислотная последовательность легкой цепи представлена в SEQ ID NO: 44, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 43.
Аминокислотная последовательность тяжелой цепи NS33 представлена в SEQ ID NO: 46, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 45. При этом аминокислотная последовательность легкой цепи представлена в SEQ ID NO: 48, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 47.
В настоящем изобретении "активность, эквивалентная активности антитела согласно любому из (1)-(6)" означает, что активность связывания и/или нейтрализации NR10 (например, NR10 человека) является эквивалентной. В настоящем изобретении термин "эквивалентный" означает, что активность не обязательно является такой же, но что она может быть усилена или снижена, при условии сохранения активности. Антитела со сниженной активностью включают, например, антитела, имеющие активность, которая на 30% или более, предпочтительно 50% или более, и более предпочтительно на 80% или более превышает активность исходного антитела.
Антитела согласно любому из (1)-(6), упомянутые выше, могут иметь замену, делецию, добавление и/или вставку одной или нескольких аминокислот в аминокислотной последовательности вариабельных областей (в последовательностях CDR и/или последовательностях FR), при условии сохранения активности связывания и/или нейтрализации NR10. Хорошо известные специалистам в данной области способы получения аминокислотной последовательности антитела, которая имеет замену, делецию, добавление и/или вставку одной или нескольких аминокислот в аминокислотной последовательности и сохраняет активность связывания и/или нейтрализации NR10, включают способы внесения мутаций в белки. Например, специалисты в данной области могут получить мутанты, функционально эквивалентные антителу, имеющему активность связывания и/или нейтрализации NR10, путем внесения соответствующих мутаций в аминокислотную последовательность антитела, имеющего активность связывания и/или нейтрализации NR10 с использованием сайт-направленного мутагенеза (Hashimoto-Gotoh, T, Mizuno, T, Ogasahara, Y, и Nakagawa, M. (1995) An oligodeoxyribonucleotide-directed dual amber method for site-directed mutagenesis. Gene 152, 271-275, Zoller, MJ, and Smith, M.(1983) Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors. Methods Enzymol. 100, 468-500, Kramer, W, Drutsa, V, Jansen, HW, Kramer, B, Pflugfelder, M, и Fritz, HJ (1984) The gapped duplex DNA approach to oligonucleotide-directed mutation construction. Nucleic Acids Res. 12, 9441-9456, Kramer W, and Fritz HJ (1987) Oligonucleotide-directed construction of mutations via gapped duplex DNA Methods. Enzymol. 154, 350-367, Kunkel, TA (1985) Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc Natl Acad Sci USA. 82, 488-492) или сходные с ними. Таким образом, также к антителу по настоящему изобретению относятся антитела, которые содержат мутации одной или нескольких аминокислот в вариабельных областях и имеют активность связывания и/или нейтрализации NR10.
Когда заменяют аминокислотный остаток, предпочтительно проводят мутацию аминокислоты в отличающуюся аминокислоту(ы), которая сохраняет свойства боковой цепи аминокислоты. Примерами свойств боковой цепи аминокислот являются: гидрофобные аминокислоты (A, I, L, M, F, P, W, Y и V), гидрофильные аминокислоты (R, D, N, C, E, Q, G, H, K, S и T), аминокислоты, содержащие алифатические боковые цепи (G, A, V, L, I и P), аминокислоты, имеющие содержащие гидроксильную группу боковые цепи (S, T и Y), аминокислоты, имеющие содержащие серу боковые цепи (C и M), аминокислоты, имеющие содержащие карбоновую кислоту и амид боковые цепи (D, N, E и Q), аминокислоты, содержащие основные боковые цепи (R, K и H), и аминокислоты, содержащие ароматические боковые цепи (H, F, Y, и W) (аминокислоты представлены в скобках с помощью однобуквенного кода). Аминокислотные замены в пределах каждой группы называют консервативными заменами. Хорошо известно, что полипептид, содержащий модифицированную аминокислотную последовательность, в которой один или несколько аминокислотных остатков данной аминокислотной последовательности удалены, добавлены и/или заменены другими аминокислотами, может сохранять исходную биологическую активность (Mark, D. F. et al., Proc. Natl. Acad. Sci. USA; (1984) 81:5662-6; Zoller, M. J. and Smith, M., Nucleic Acids Res. (1982) 10:6487-500; Wang, A. et al., Science (1984) 224:1431-3; Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79:6409-13). Такие мутанты обладают идентичностью аминокислот, по меньшей мере, 70%, более предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 80%, еще более предпочтительно, по меньшей мере, 85%, еще более предпочтительно, по меньшей мере, 90%, и наиболее предпочтительно, по меньшей мере, 95%, с вариабельными областями (например, последовательностями CDR, последовательностями FR или целыми вариабельными областями) по настоящему изобретению. В настоящем документе идентичность последовательности определяют как процент остатков, идентичных остаткам в исходной аминокислотной последовательности вариабельной области тяжелой цепи или вариабельной области легкой цепи, определенный после выравнивания последовательностей и внесения соответствующим образом пропусков для максимального увеличения идентичности последовательности, при необходимости. Идентичность аминокислотных последовательностей можно определять способом, описанным ниже.
Альтернативно аминокислотные последовательности вариабельных областей, которые имеют замену, делецию, добавление и/или вставку одной или нескольких аминокислот в аминокислотной последовательности вариабельных областей (в последовательностях CDR и/или последовательностях FR) и сохраняют активность связывания и/или нейтрализации NR10, можно получать из нуклеиновых кислот, которые гибридизуются в строгих условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, кодирующей аминокислотную последовательность вариабельных областей. Строгие условия гибридизации для выделения нуклеиновой кислоты, гибридизующейся в строгих условиях с нуклеиновой кислотой, включающей нуклеотидную последовательность, кодирующую аминокислотную последовательность вариабельных областей, включают, например, условия 6M мочевины, 0,4% SDS, 0,5x SSC, и 37ºC, или условия гибридизации со строгостью, эквивалентной этой. При более строгих условиях, например, условиях 6M мочевины, 0,4% SDS, 0,1x SSC и 42ºC, можно ожидать выделения нуклеиновых кислот со значительно более высокой гомологией. Последовательности выделенных нуклеиновых кислот можно определять известными способами, описанными ниже. Общая гомология нуклеотидной последовательности выделенной нуклеиновой кислоты представляет собой, по меньшей мере, 50% идентичность последовательности или более, предпочтительно 70% или более, более предпочтительно 90% или более (например, 95%, 96%, 97%, 98%, 99% или более).
Нуклеиновые кислоты, которые гибридизуются в строгих условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, кодирующей аминокислотную последовательность вариабельных областей, также можно выделять с использованием, вместо описанных выше способов с использованием технологий гибридизации, способов амплификации генов, таких как полимеразная цепная реакция (ПЦР) с использованием праймеров, синтезированных на основе информации нуклеотидной последовательности, кодирующей аминокислотную последовательность вариабельных областей.
Конкретно, идентичность одной нуклеотидной последовательности или аминокислотной последовательности с другой последовательностью можно определять с использованием алгоритма BLAST, Karlin and Altschul (Proc. Natl. Acad. Sci. USA (1993) 90, 5873-7). На основе этого алгоритма были разработаны программы, такие как BLASTN и BLASTX (Altschul et al., J. Mol. Biol. (1990) 215, 403-10). Для анализа нуклеотидных последовательностей согласно BLASTN на основе BLAST, параметры устанавливают, например, как score=100 и wordlength=12. С другой стороны, параметры, используемые для анализа аминокислотных последовательностей с помощью BLASTX на основе BLAST, включают, например, score=50 и wordlength=3. При использовании программ BLAST и Gapped BLAST используют параметры по умолчанию для каждой программы. Конкретные способы для таких анализов известны в данной области (см. web-сайт National Center for Biotechnology Information (NCBI), Basic Local Alignment Search Tool (BLAST); http://www.ncbi.nlm.nih.gov).
Настоящее изобретение также относится к антителам, которые связываются с тем же эпитопом, с которым связываются антитела согласно любому из (1)-(7).
Распознавание антителом того же эпитопа, который распознает другое антитело, можно подтвердить с помощью конкуренции между этими двумя антителами за эпитоп. Конкуренцию между антителами можно оценивать с помощью конкурентных анализов связывания с использованием таких способов, как ELISA, способ переноса энергии флуоресценции (FRET), и технология флуориметрического анализа в микрообъеме (FMAT(R)). Количество антител, связавшихся с антигеном, прямо коррелирует со способностью связывания конкурентных антител-кандидатов (тестируемых антител), которые конкурентно связываются с тем же эпитопом. Иными словами, по мере повышения количества или аффинности тестируемых антител против того же эпитопа, количество антител, связавшихся с антигеном, снижается, и возрастает количество тестируемых антител, связавшихся с антигеном. Конкретно, к антигену одновременно добавляют соответствующим образом меченные антитела и антитела, подлежащие оценке, и проводят детекцию связавшихся таким образом антител с использованием метки. Количество антител, связавшихся с антигеном, можно легко определить предварительным мечением антител. Эта метка конкретно не ограничена, и способ мечения выбирают согласно используемому способу анализа. Способ мечения включает мечение флуоресцентной меткой, мечение радиоактивной меткой, мечение ферментом и т.д.
Например, флуоресцентно меченые антитела и немеченые антитела или тестируемые антитела одновременно добавляют к клеткам животных, экспрессирующим NR10, и детекцию меченых антител проводят с помощью технологии флуориметрического анализа в микрообъеме.
В настоящем документе "антитело, которое распознает тот же эпитоп" относится к антителу, которое может снизить связывание меченого антитела, по меньшей мере, на 50% в концентрации, которая обычно в 100 раз превышает, предпочтительно в 80 раз превышает, более предпочтительно в 50 раз превышает, еще более предпочтительно в 30 раз превышает, и еще более предпочтительно в 10 раз превышает концентрацию, в которой немеченое антитело снижает связывание меченого антитела на 50% (IC50).
Антитела, которые связываются с эпитопом, с которым связываются антитела, указанные в любом из (1)-(7), выше, пригодны, поскольку они имеют особенно высокую нейтрализующую активность.
Антитела, указанные в любом из (1)-(8), выше, предпочтительно представляют собой гуманизированные антитела, но они не ограничиваются конкретно ими.
Более того, настоящее изобретение относится к генам, кодирующим антитела против NR10 согласно любому из (1)-(8) из (A)-(D), выше. Гены по настоящему изобретению могут представлять собой любую форму генов, например, ДНК или РНК.
Антитела (гуманизированные)
Предпочтительные варианты осуществления антител по настоящему изобретению включают гуманизированные антитела, которые связываются с NR10. Гуманизированные антитела можно получать способами, известными специалистам в данной области.
Вариабельная область антитела, как правило, состоит из трех определяющих комплементарность областей (CDR), встроенных между четырьмя каркасами (FR). CDR по существу определяют специфичность связывания антитела. Аминокислотные последовательности CDR являются высоко разнообразными. Напротив, аминокислотные последовательности FR часто проявляют высокую гомологию между антителами, имеющими отличающуюся специфичность связывания. Таким образом, как правило, утверждается, что специфичность связывания антитела можно трансплантировать в отличающееся антитело путем пересадки CDR.
Гуманизированные антитела также называют переформированными антителами человека, и их получают переносом CDR антитела, происходящего из не являющегося человеком млекопитающего, такого как мышь, в CDR антитела человека. Также для их получения известны общие способы генетической рекомбинации (см. публикацию патентной заявки Европы No. 125023 и WO 96/02576).
Конкретно, например, когда CDR происходят из антитела мыши, синтезируют последовательность ДНК, сконструированную так, чтобы CDR антитела мыши были связаны с каркасными областями (FR) антитела человека, с помощью ПЦР с использованием, в качестве праймеров, нескольких олигонуклеотидов, которые имеют участки, охватывающие концы как CDR, так и FR (см. способ, описанный WO 98/13388). Затем полученную ДНК лигируют с ДНК, кодирующей константную область антитела человека, встраивают в экспрессирующий вектор и вводят хозяину для продукции антитела (см. публикацию патентной заявки Европы No. EP 239400 и публикацию международной патентной заявки No. WO 96/02576).
Каркасные области антитела человека, подлежащие связыванию с CDR, выбирают таким образом, чтобы CDR образовывали подходящий антигенсвязывающий участок. Если необходимо, в каркасные области вариабельной области антитела можно вносить аминокислотную замену, делецию, добавление и/или вставку, так чтобы CDR переформированного антитела человека образовывали надлежащий антигенсвязывающий участок. Например, в аминокислотную последовательность FR можно вносить мутации с использованием способа ПЦР, который используют для пересадки CDR мыши в FR человека. Конкретно, мутации можно вносить в часть нуклеотидных последовательностей праймеров, которые подвергаются отжигу с FR. В FR, синтезированные с помощью таких праймеров, вносятся мутации. Антигенсвязывающую активность мутантных антител, имеющих аминокислотные замены, можно определять и оценивать способом, описанным выше, и, тем самым, можно отбирать мутантные последовательности FR, имеющие желательные свойства (Sato, K. et al., Cancer Res. (1993) 53, 851-856).
В качестве константных (C) областей в гуманизированных антителах используют константные области из антител человека. Например, для H-цепей используют Cγ1, Cγ2, Cγ3, Cγ4, Cμ, Cδ, Cα1, Cα2 и Cε; и для L-цепей используют Cκ и Cλ. Аминокислотная последовательность Cκ представлена в SEQ ID NO: 58, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 57. Аминокислотная последовательность Cγ1 представлена в SEQ ID NO: 60, и нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 59. Аминокислотная последовательность Cγ2 представлена в SEQ ID NO: 62, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 61. Аминокислотная последовательность Cγ4 представлена в SEQ ID NO: 64, а нуклеотидная последовательность, кодирующая эту аминокислотную последовательность, представлена в SEQ ID NO: 63. Более того, C-области антитела человека можно модифицировать для повышения стабильности антитела или увеличения продукции антитела. Модифицированные C-области антител человека включают, например, C-области, описанные в настоящем документе ниже. Антитела человека, используемые для гуманизации, могут быть антителами любого изотипа, такого как IgG, IgM, IgA, IgE или IgD; однако предпочтительно в настоящем изобретении используют IgG. IgG, который можно использовать, включает IgG1, IgG2, IgG3, IgG4 и т.п.
Более того, после получения гуманизированного антитела аминокислоты в вариабельной области (например, CDR и FR) и константной области гуманизированного антитела можно удалять, добавлять, встраивать и/или заменять другими аминокислотами. Антитела по настоящему изобретению также включают такие гуманизированные антитела с аминокислотными заменами и т.д.
Источник CDR гуманизированного антитела конкретно не ограничен, и им может быть любое животное. Например, можно использовать последовательности антител мыши, антител крысы, антител кролика, антител верблюда и т.п. Предпочтительными являются последовательности CDR антител мыши.
Как правило, гуманизация антител при сохранении связывания и нейтрализующей активности исходных антител является трудной. Однако настоящее изобретение является успешным с точки зрения получения гуманизированных антител, имеющих связывающую и/или нейтрализующую активность, эквивалентную активности исходных антител мыши. Гуманизированные антитела пригодны для введения человека в терапевтических целях, поскольку они обладают сниженной иммуногенностью в организме человека.
Предпочтительные примеры гуманизированных антител против NR10 по настоящему изобретению включают, например:
(a) гуманизированные антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 50 (H0-VH);
(b) гуманизированные антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 112 (H1-VH);
(c) гуманизированные антитела, которые содержат вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 52 (L0-VL);
(d) гуманизированные антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 50 (H0-VH), и вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 52 (L0-VL); и
(e) гуманизированные антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 112, и вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 52.
Вариабельная область тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 50 (H0-VH), вариабельная область тяжелой цепи, имеющая аминокислотную последовательность SEQ ID NO: 112, и вариабельная область легкой цепи, имеющая аминокислотную последовательность SEQ ID NO: 52 (L0-VL) могут иметь замену, делецию, добавление и/или вставку одной или нескольких аминокислот. Замену, делецию, добавление и/или вставку аминокислот можно проводить в любых или во всех из CDR и FR.
Таким образом, другие предпочтительные варианты осуществления гуманизированного антитела против NR10 по настоящему изобретению включают, например:
(f) антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 50 (H0-VH);
(g) антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 112 (H1-VH);
(h) антитела, которые содержат вариабельную область легкой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 52 (L0-VL);
(i) антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 50 (H0-VH), и вариабельную область легкой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 52 (L0-VL);
(j) антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 112 (H1-VH), и вариабельную область легкой цепи, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 52 (L0-VL);
Без конкретных ограничений, антитела согласно любому из (f)-(j) предпочтительно имеют активность, сходную с активностью антител согласно любому из (a)-(e).
Замена, делеция, добавление и/или вставка аминокислот конкретно не ограничены, но их конкретные примеры включают, например, описанные выше аминокислотные замены.
Более конкретно, например, могут быть включены следующие аминокислотные замены:
Замена Ile в положении 3 CDR1 (SEQ ID NO: 9) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Val (SEQ ID NO: 173). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 9, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 173, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Met в положении 4 CDR1 (SEQ ID NO: 9) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Ile (SEQ ID NO: 174). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 9, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 174, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Met в положении 4 CDR1 (SEQ ID NO: 9) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Leu (SEQ ID NO: 175). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 9 заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 175, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Ile в положении 3 CDR1 (SEQ ID NO: 9) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Ala (SEQ ID NO: 176). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 9, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 176, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Leu в положении 1 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Glu (SEQ ID NO: 113). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 113, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Asn в положении 3 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Asp (SEQ ID NO: 114). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 114, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Gln в положении 13 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Asp (SEQ ID NO: 115). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 115, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Lys в положении 14 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Gln (SEQ ID NO: 116). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 116, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Lys в положении 16 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Gln (SEQ ID NO: 117). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 117, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Gly в положении 17 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Asp (SEQ ID NO: 118). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 118, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Lys в положении 16 и Gly в положении 17 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Gln и Asp, соответственно (SEQ ID NO: 119). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 119, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Lys в положении 14, Lys в положении 16 и Gly в положении 17 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Gln, Gln и Asp, соответственно (SEQ ID NO: 167). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 171, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Gln в положении 13, Lys в положении 14, Lys в положении 16 и Gly в положении 17 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Asp, Gln, Gln и Asp, соответственно (SEQ ID NO: 172). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 172, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Ser в положении 10 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Asp (SEQ ID NO: 177). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 177, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Gln в положении 13 CDR2 (SEQ ID NO: 10) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Pro (SEQ ID NO: 178). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 10, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 178, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Tyr в положении 3 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Leu (SEQ ID NO: 179). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 179, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Met в положении 10 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Leu (SEQ ID NO: 180). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 180, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Asp в положении 11 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Glu (SEQ ID NO: 181). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 181, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Tyr в положении 12 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Thr (SEQ ID NO: 182). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 182, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Tyr в положении 12 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Ser (SEQ ID NO: 183). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 183, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Met в положении 10, Asp в положении 11 и Tyr в положении 12 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Leu, Glu, Thr, соответственно (SEQ ID NO: 184). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 184, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Asp в положении 11 и Tyr в положении 12 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Glu и Thr, соответственно (SEQ ID NO: 185). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 185, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Tyr в положении 3, Asp в положении 11 и Tyr в положении 12 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Leu, Glu и Thr, соответственно (SEQ ID NO: 186). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 186, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Tyr в положении 3, Asp в положении 11 и Tyr в положении 12 CDR3 (SEQ ID NO: 11) в вариабельной области тяжелой цепи SEQ ID NO: 50 или 112 на Leu, Glu и Ser, соответственно (SEQ ID NO: 187). Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 11, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 187, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Замена Arg в положении 1 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Gln (SEQ ID NO: 121). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 121, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Asn в положении 5 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Asp (SEQ ID NO: 122). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 122, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Ser в положении 8 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Arg (SEQ ID NO: 188). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 188, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Leu в положении 10 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Val (SEQ ID NO: 189). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 189, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Ser в положении 8 и Leu в положении 10 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Arg и Val, соответственно (SEQ ID NO: 190). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 190, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Thr в положении 2 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Ala (SEQ ID NO: 191). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 191, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Thr в положении 2 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Ser (SEQ ID NO: 192). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 192, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Asn в положении 1 CDR2 (SEQ ID NO: 14) в вариабельной области легкой цепи SEQ ID NO: 52 на Asp (SEQ ID NO: 123). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 14, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 123, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Lys в положении 3 CDR2 (SEQ ID NO: 14) в вариабельной области легкой цепи SEQ ID NO: 52 на Gln (SEQ ID NO: 124). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 14, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 124, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Leu в положении 5 CDR2 (SEQ ID NO: 14) в вариабельной области легкой цепи SEQ ID NO: 52 на Glu (SEQ ID NO: 125). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 14, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 125, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Lys в положении 7 CDR2 (SEQ ID NO: 14) в вариабельной области легкой цепи SEQ ID NO: 52 на Gln (SEQ ID NO: 126). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 14, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 126, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Lys в положении 7 CDR2 (SEQ ID NO: 14) в вариабельной области легкой цепи SEQ ID NO: 52 на Asp (SEQ ID NO: 127). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 14, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 127, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Arg в положении 1 и Asn в положении 5 CDR1 (SEQ ID NO: 13) в вариабельной области легкой цепи SEQ ID NO: 52 на Gln и Asp, соответственно (SEQ ID NO: 169). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR1, имеющая аминокислотную последовательность SEQ ID NO: 13, заменена CDR1, имеющей аминокислотную последовательность SEQ ID NO: 169, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Lys в положении 3, Leu в положении 5 и Lys в положении 7 CDR2 (SEQ ID NO: 14) в вариабельной области легкой цепи SEQ ID NO: 52 на Gln, Glu и Gln, соответственно (SEQ ID NO: 170). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 14, заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 170, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Glu в положении 5 CDR3 (SEQ ID NO: 15) в вариабельной области легкой цепи SEQ ID NO: 52 на Asp (SEQ ID NO: 193). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 15, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 193, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Ser в положении 6 CDR3 (SEQ ID NO: 15) в вариабельной области легкой цепи SEQ ID NO: 52 на Asp (SEQ ID NO: 194). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 15, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 194, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Замена Thr в положении 9 CDR3 (SEQ ID NO: 15) в вариабельной области легкой цепи SEQ ID NO: 52 на Phe (SEQ ID NO: 195). Таким образом, настоящее изобретение относится к вариабельным областям легкой цепи, в которых CDR3, имеющая аминокислотную последовательность SEQ ID NO: 15, заменена CDR3, имеющей аминокислотную последовательность SEQ ID NO: 195, в вариабельной области легкой цепи, имеющей аминокислотную последовательность SEQ ID NO: 52.
Кроме того, замены, отличные от замен, описанных выше, включают замену Arg в положении 3 FR2 тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 97, другой аминокислотой. Аминокислота после замены конкретно не ограничена; но ее предпочтительные замены включают Gln. Когда Arg в положении 3 в SEQ ID NO: 97 заменен на Gln, Ala в положении 5 может быть заменен на Ser с получением последовательности FR2 человека. Аминокислотная последовательность, в которой Arg и Ala в положениях 3 и 5 в аминокислотной последовательности SEQ ID NO: 97 заменены Gln и Ser, соответственно, представлена в SEQ ID NO: 120. Таким образом, настоящее изобретение относится к вариабельным областям тяжелой цепи, в которых FR2, имеющая аминокислотную последовательность SEQ ID NO: 97, заменена FR2, имеющей аминокислотную последовательность SEQ ID NO: 120, в вариабельной области тяжелой цепи, имеющей аминокислотную последовательность SEQ ID NO: 50 или 112.
Каждую из описанных выше аминокислотных замен можно использовать отдельно или в сочетании с другими аминокислотными заменами, описанными выше. Также их можно комбинировать с аминокислотными заменами, отличными от замен, описанных выше.
Конкретные примеры антител, в которых проведены описанные выше замены, включают, например, антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 167, антитела, которые содержат вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 168, и антитела, которые содержат вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 167, и вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 168.
Более того, конкретные примеры вариабельных областей тяжелой цепи, в которых проведены описанные выше замены, включают, например, следующие вариабельные области тяжелой цепи:
(1) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 204 (H17);
(2) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 205 (H19);
(3) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 206 (H28);
(4) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 207 (H30);
(5) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 208 (H34);
(6) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 209 (H42);
(7) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 210 (H44);
(8) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 211 (H46);
(9) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 212 (H57);
(10) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 213 (H71);
(11) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 214 (H78);
(12) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 215 (H92);
(13) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 216 (H97); и
(14) вариабельные области тяжелой цепи, имеющие аминокислотную последовательность SEQ ID NO: 217 (H98).
При этом конкретные примеры вариабельных областей легкой цепи, в которых проведены описанные выше замены, включают, например, следующие вариабельные области легкой цепи:
(15) вариабельные области легкой цепи, имеющие аминокислотную последовательность SEQ ID NO: 218 (L11);
(16) вариабельные области легкой цепи, имеющие аминокислотную последовательность SEQ ID NO: 219 (L12);
(17) вариабельные области легкой цепи, имеющие аминокислотную последовательность SEQ ID NO: 220 (L17); и
(18) вариабельные области легкой цепи, имеющие аминокислотную последовательность SEQ ID NO: 221 (L50).
Более того, конкретные примеры антител, содержащих описанные выше вариабельные области тяжелой цепи и легкой цепи, включают, например, следующие антитела:
(19) антитела, которые содержат вариабельную область тяжелой цепи (3) и вариабельную область легкой цепи (17) (H28L17);
(20) антитела, которые содержат вариабельную область тяжелой цепи (4) и вариабельную область легкой цепи (17) (H30L17);
(21) антитела, которые содержат вариабельную область тяжелой цепи (5) и вариабельную область легкой цепи (17) (H34L17);
(22) антитела, которые содержат вариабельную область тяжелой цепи (6) и вариабельную область легкой цепи (17) (H42L17);
(23) антитела, которые содержат вариабельную область тяжелой цепи (7) и вариабельную область легкой цепи (17) (H44L17);
(24) антитела, которые содержат вариабельную область тяжелой цепи (8) и вариабельную область легкой цепи (17) (H46L17);
(25) антитела, которые содержат вариабельную область тяжелой цепи (9) и вариабельную область легкой цепи (17) (H57L17);
(26) антитела, которые содержат вариабельную область тяжелой цепи (10) и вариабельную область легкой цепи (17) (H71L17);
(27) антитела, которые содержат вариабельную область тяжелой цепи (11) и вариабельную область легкой цепи (17) (H78L17);
(28) антитела, которые содержат вариабельную область тяжелой цепи (12) и вариабельную область легкой цепи (17) (H92L17);
(29) антитела, которые содержат вариабельную область тяжелой цепи (13) и вариабельную область легкой цепи (18) (H97L50); и
(30) антитела, которые содержат вариабельную область тяжелой цепи (14) и вариабельную область легкой цепи (18) (H98L50).
Константная область, используемая для гуманизированных антител по настоящему изобретению, может представлять собой любую константную область, происходящую из антитела человека. Предпочтительные примеры таких константных областей, происходящих из антител человека, включают, например, константные области, происходящие из IgG1 или IgG2. Более того, также можно использовать константные области, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в константной области, происходящей из антитела человека.
Константные области, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в константной области, происходящей из антитела человека, конкретно не ограничены, и включают, например, следующие константные области:
константные области, имеющие аминокислотную последовательность SEQ ID NO: 128 (M58);
константные области, имеющие аминокислотную последовательность SEQ ID NO: 129 (M14); и
константные области, имеющие аминокислотную последовательность SEQ ID NO: 62 (SKSC).
Конкретные примеры тяжелых цепей или антител, имеющих описанные выше константные области, включают, например:
(1) тяжелые цепи, которые содержат вариабельную область, имеющую аминокислотную последовательность SEQ ID NO: 167, и константную область, имеющую аминокислотную последовательность SEQ ID NO: 128;
(2) тяжелые цепи, в которых CDR2, имеющая аминокислотную последовательность SEQ ID NO: 171, в тяжелых цепях (1) заменена CDR2, имеющей аминокислотную последовательность SEQ ID NO: 172;
(3) антитела, которые содержат тяжелую цепь согласно (1) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 152; и
(4) антитела, которые содержат тяжелую цепь согласно (2) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 152.
Более конкретные примеры гуманизрованных антител против NR10 по настоящему изобретению включают, например, следующие антитела:
(k) антитела, которые содержат тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 54 (H0-VH+константная область);
(l) антитела, которые содержат тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 130 (H1-VH+константная область);
(m) антитела, которые содержат легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 56 (L0-VL+константная область);
(n) антитела, которые содержат тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 54 (H0-VH+константная область), и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 56 (L0-VL+константная область); и
(o) антитела, которые содержат тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 130 (H1-VH+константная область), и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 56 (L0-VL+константная область).
Тяжелая цепь, имеющая аминокислотную последовательность SEQ ID NO: 54 (H0-VH+константная область) и легкая цепь, имеющая аминокислотную последовательность SEQ ID NO: 56 (L0-VL+константная область), могут иметь замену, делецию, добавление и/или вставку одной или нескольких аминокислот. Замену, делецию, добавление и/или вставку аминокислот можно проводить в любых или во всех из вариабельных и константных областей.
Таким образом, настоящее изобретение относится к:
(p) антителам, которые содержат тяжелую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 54 (H0-VH+константная область);
(q) антителам, которые содержат тяжелую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 130 (H1-VH+константная область);
(r) антителам, которые содержат легкую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 56 (L0-VL+константная область);
(s) антителам, которые содержат тяжелую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 54 (H0-VH+константная область), и легкую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 56 (L0-VL+константная область); и
(t) антитела, которые содержат тяжелую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 130 (H1-VH+константная область), и легкую цепь, имеющую аминокислотную последовательность, где одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в аминокислотной последовательности SEQ ID NO: 56 (L0-VL+константная область).
Без конкретных ограничений, антитела согласно любому из (p)-(t) предпочтительно имеют активность, сходную с активностью антител согласно любому из (k)-(o).
Замена, делеция, добавление и/или вставка аминокислот конкретно не ограничены, однако их конкретные примеры включают, например, описанные выше аминокислотные замены.
Более того, нуклеотидная последовательность, кодирующая аминокислотную последовательность описанной выше гуманизированной вариабельной области тяжелой цепи (SEQ ID NO: 50) представлена в SEQ ID NO: 49. Нуклеотидная последовательность, кодирующая аминокислотную последовательность гуманизированной вариабельной области легкой цепи (SEQ ID NO: 52), представлена в SEQ ID NO: 51. Нуклеотидная последовательность, кодирующая аминокислотную последовательность гуманизированной тяжелой цепи (SEQ ID NO: 54), представлена в SEQ ID NO: 53. Нуклеотидная последовательность, кодирующая аминокислотную последовательность гуманизированной легкой цепи (SEQ ID NO: 56) представлена в SEQ ID NO: 55.
Более того, настоящее изобретение относится к антителам, которые распознают тот же эпитоп, который распознают антитела согласно любому из (a)-(t) выше. Связывание с тем же эпитопом уже описано выше.
Более того, настоящее изобретение относится к следующим антителам:
(u) антитела, которые содержат тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 151;
(v) антитела, которые содержат легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 152; и
(w) антитела, которые содержат тяжелую цепь (u) и легкую цепь (v).
Более того, настоящее изобретение относится к следующим тяжелым и легким цепям и антителам:
(1) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 222 (H17);
(2) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 223 (H19);
(3) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 224 (H28);
(4) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 225 (H30);
(5) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 226 (H34);
(6) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 227 (H42);
(7) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 228 (H44);
(8) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 229 (H46);
(9) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 230 (H57);
(10) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 231 (H71);
(11) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 232 (H78);
(12) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 233 (H92);
(13) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 234 (H97);
(14) тяжелые цепи, имеющие аминокислотную последовательность SEQ ID NO: 235 (H98);
(15) легкие цепи, имеющие аминокислотную последовательность SEQ ID NO: 236 (L11)
(16) легкие цепи, имеющие аминокислотную последовательность SEQ ID NO: 237 (L12);
(17) легкие цепи, имеющие аминокислотную последовательность SEQ ID NO: 238 (L17);
(18) легкие цепи, имеющие аминокислотную последовательность SEQ ID NO: 239 (L50);
(19) антитела, которые содержат тяжелую цепь (3) и легкую цепь (17) (H28L17);
(20) антитела, которые содержат тяжелую цепь (4) и легкую цепь (17) (H30L17);
(21) антитела, которые содержат тяжелую цепь (5) и легкую цепь (17) (H34L17);
(22) антитела, которые содержат тяжелую цепь (6) и легкую цепь (17) (H42L17);
(23) антитела, которые содержат тяжелую цепь (7) и легкую цепь (17) (H44L17);
(24) антитела, которые содержат тяжелую цепь (8) и легкую цепь (17) (H46L17);
(25) антитела, которые содержат тяжелую цепь (9) и легкую цепь (17) (H57L17);
(26) антитела, которые содержат тяжелую цепь (10) и легкую цепь (17) (H71L17);
(27) антитела, которые содержат тяжелую цепь (11) и легкую цепь (17) (H78L17);
(28) антитела, которые содержат тяжелую цепь (12) и легкую цепь (17) (H92L17);
(29) антитела, которые содержат тяжелую цепь (13) и легкую цепь (18) (H97L50);
(30) антитела, которые содержат тяжелую цепь (14) и легкую цепь (18) (H98L50);
(31) тяжелые цепи, имеющие аминокислотную последовательность, в которой одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в тяжелые цепи согласно любому из (1)-(14);
(32) легкие цепи, имеющие аминокислотную последовательность, в которой одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в легкие цепи согласно любому из (15)-(18);
(33) антитела, имеющие аминокислотную последовательность, в которой одна или несколько аминокислот заменены, удалены, добавлены и/или вставлены в антитела согласно любому из (19)-(30); и
(34) антитела, которые распознают тот же эпитоп, который распознают антитела согласно любому из (19)-(33).
Замена, делеция, добавление и/или вставка аминокислот являются такими, как описано выше. Антитела, которые распознают тот же эпитоп, который распознает антитело, также описаны выше.
Также настоящее изобретение относится к генам, кодирующим вариабельные области, тяжелые цепи, легкие цепи или антитела по настоящему изобретению.
Также настоящее изобретение относится к векторам, обладающим описанными выше генами.
Также настоящее изобретение относится к клеткам-хозяевам, трансформированным описанными выше векторами.
Также настоящее изобретение относится к способам продукции вариабельных областей тяжелых цепей, легких цепей или антител по настоящему изобретению, которые включают стадию культивирования описанных выше клеток-хозяев.
Векторы, клетки-хозяева и культивирование клеток-хозяев описаны в настоящем документе ниже.
Антитела, которые распознают домены
Предпочтительные варианты осуществления антитела против NR10 по настоящему изобретению включают антитела, которые распознают домен 1 или домен 2. В настоящем изобретении домен 1 относится к области из аминокислот в положениях 21-120 (от LPAKP до LENIA) в аминокислотной последовательности NR10 человека SEQ ID NO: 76, где нумерация аминокислот основана на последовательности, включающей сигнальный пептид. Кроме того, в настоящем изобретении домен 2 относится к области из аминокислот в положениях 121-227 (от KTEPP до EEEAP) в аминокислотной последовательности NR10 человека SEQ ID NO: 76, где нумерация аминокислот основана на последовательности, включающей сигнальный пептид.
Такие антитела конкретно не ограничены; однако, как правило, они обладают нейтрализующей активностью, и предпочтительно они представляют собой гуманизированные антитела.
Примеры предпочтительных антител по настоящему изобретению включают антитела, которые распознают домен 1. Антитела, которые распознают домен 1, имеют сильную нейтрализующую активность, и, таким образом, они являются особенно пригодными в качестве фармацевтических средств.
Антитела (нейтрализующая активность)
Настоящее изобретение также относится к антителам против NR10, имеющим нейтрализующую активность.
В настоящем изобретении нейтрализующая активность против NR10 относится к активности ингибирования связывания между NR10 и его лигандом IL-31, и предпочтительно к активности подавления биологической активности, основанной на NR10.
Антитела, имеющие нейтрализующую активность против NR10, можно отбирать, например, путем добавления антител-кандидатов к IL-31-зависимой клеточной линии и наблюдения их эффекта подавления роста клеточной линии. В этом способе антитела, которые подавляют рост IL-31-зависимой клеточной линии, определяют как антитела, имеющие нейтрализующую активность против NR10.
Антитела (общее)
Антитела по настоящему изобретению не ограничиваются с точки зрения их происхождения, и их источником могут быть любые животные, такие как люди, мыши и крысы. Более того, антитела могут представлять собой рекомбинантные антитела, такие как химерные антитела и гуманизированные антитела. Как описано выше, предпочтительные антитела по настоящему изобретению включают гуманизированные антитела.
Химерные антитела содержат, например, константные области тяжелой и легкой цепи антитела человека, и вариабельные области тяжелой и легкой цепи антитела, не являющегося человеком млекопитающего, такого как мышь. Химерные антитела можно получать известными способами. Например, антитела можно получать клонированием гена антитела из гибридомы, встраиванием его в соответствующий вектор, и введением этой конструкции в хозяев (см., например, Carl, A. K. Borrebaeck, James, W. Larrick, THERAPEUTIC MONOCLONAL ANTIBODIES, Published in the United Kingdom by MACMILLAN PUBLISHERS LTD, 1990). Конкретно, синтезируют кДНК вариабельных областей (V-областей) антитела из мРНК гибридом с использованием обратной транскриптазы. После получения ДНК, кодирующих V-области представляющего интерес антитела, их связывают с ДНК, кодирующими константные области (C-области) требуемого антитела человека. Полученные конструкции встраивают в экспрессирующие векторы. Альтернативно ДНК, кодирующие V-области антитела, можно встраивать в экспрессирующие векторы, содержащие ДНК, кодирующие C-области антитела человека. ДНК встраивают в экспрессирующие векторы, так чтобы они экспрессировались под регуляцией участков регуляции экспрессии, например, энхансеров и промоторов. На следующей стадии клетки-хозяева можно трансформировать экспрессирующими векторами для обеспечения экспрессии химерного антитела.
Способы получения антител человека также известны. Например, желаемые антитела человека с антигенсвязывающей активностью можно получать путем (1) сенсибилизации лимфоцитов человека представляющими интерес антигенами или клетками, экспрессирующими представляющие интерес антигены in vitro; и (2) слияния сенсибилизированных лимфоцитов с клетками миеломы человека, такими как U266 (см. публикацию японской патентной заявки Kokoku No. (JP-B) H01-59878 (рассмотренная, принятая японская патентная заявка, опубликованная для опротестования)). Альтернативно желаемое антитело человека также можно получать иммунизацией трансгенного животного, имеющего полный набор генов антител человека, требуемым антигеном (см. публикации международных патентных заявок No. WO 93/12227, WO 92/03918, WO 94/02602, WO 94/25585, WO 96/34096 и WO 96/33735).
Более того, известны способы получения антител человека сортировкой фаговых библиотек антител человека. Например, с использованием способа фагового дисплея вариабельную область антитела человека экспрессируют в качестве одноцепочечного антитела (scFv) на поверхности фага, и можно отбирать фаги, которые связываются с антигеном. Посредством анализа генов выбранных фагов можно определять последовательности ДНК, кодирующие вариабельные области антител человека, которые связываются с антигеном. Если идентифицируют последовательности ДНК scFv, которые связываются с антигеном, можно конструировать соответствующие экспрессирующие векторы, содержащие эти последовательности, с получением антител человека. Такие способы хорошо известны. В качестве ссылки могут быть приведены WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438, WO 95/15388, и т.п.
Антитела по настоящему изобретению включают не только двухвалентные антитела, которые соответствует IgG, но также одновалентные антитела, поливалентные антитела, которые соответствует IgM и биспецифические антитела, способные связываться с различными антигенами, при условии, что они обладают активностью связывания и/или нейтрализации NR10. Поливалентные антитела по настоящему изобретению включают поливалентные антитела, в которых все антигенсвязывающие участки являются идентичными, и поливалентные антитела, в которых все или некоторые из антигенсвязывающих участков отличаются. Антитела по настоящему изобретению не ограничиваются полноразмерными молекулами антител, но они также могут представлять собой низкомолекулярные антитела или их модифицированные продукты, при условии, что они связываются с белком NR10.
Альтернативно антитела по настоящему изобретению могут представлять собой низкомолекулярные антитела. Такие низкомолекулярные антитела представляют собой антитела, включающие фрагменты антител, лишенные некоторых частей целого антитела (например, целого IgG), и они конкретно не ограничены, при условии, что они сохраняют активность связывания и/или нейтрализации NR10. В настоящем изобретении низкомолекулярные антитела конкретно не ограничены, при условии, что они содержат часть целых антител. Низкомолекулярные антитела предпочтительно содержат вариабельную область тяжелой цепи (VH) или вариабельную область легкой цепи (VL). Особенно предпочтительные низкомолекулярные антитела содержат как VH, так и VL. Кроме того, предпочтительные примеры низкомолекулярных антител по настоящему изобретению включают низкомолекулярные антитела, содержащие CDR антитела. CDR, содержащиеся в низкомолекулярных антителах, могут включать некоторые или все из шести CDR антитела.
Низкомолекулярные антитела по настоящему изобретению предпочтительно имеют меньшую молекулярную массу, чем целые антитела. Однако низкомолекулярные антитела могут образовывать мультимеры, например, димеры, тримеры или тетрамеры, и, таким образом, их молекулярная масса может превышать молекулярную массу целых антител.
Конкретные примеры фрагментов антител включают, например, Fab, Fab', F(ab')2 и Fv. При этом конкретные примеры низкомолекулярных антител включают, например, Fab, Fab', F(ab')2, Fv, scFv (одноцепочечные Fv), антитела-димеры и sc(Fv)2 (одноцепочечные (Fv)2). К низкомолекулярным антителам по настоящему изобретению также относятся мультимеры (например, димеры, тримеры, тетрамеры и полимеры) этих антител.
Фрагменты антител можно получать, например, обработкой антител ферментами с получением фрагментов антител. Ферменты, которые, как известно, образуют фрагменты антител, включают, например, папаин, пепсин и плазмин. Альтернативно можно конструировать ген, кодирующий такой фрагмент антитела, и вводить его в экспрессирующий вектор и экспрессировать в соответствующих клетках-хозяевах (см., например, Co, M.S. et al., J. Immunol. (1994) 152, 2968-2976; Better, M. & Horwitz, A. H. Methods in Enzymology (1989) 178, 476-496; Plueckthun, A. & Skerra, A. Methods in Enzymology (1989) 178, 476-496; Lamoyi, E., Methods in Enzymology (1989) 121, 652-663; Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-669; Bird, R. E. et al., TIBTECH (1991) 9, 132-137).
Расщепляющие ферменты расщепляют конкретный участок фрагмента антитела, с получением фрагментов антител с конкретными структурами, представленными ниже. Можно использовать способы генетической инженерии для удаления произвольной части таких ферментативно полученных фрагментов антител.
Фрагменты антител, полученные с использованием описанных выше расщепляющих ферментов, являются следующими:
Расщепление папаином: F(ab)2 или Fab
Расщепление пепсином: F(ab')2 или Fab'
Расщепление плазмином: Facb
Низкомолекулярные антитела по настоящему изобретению включают фрагменты антител, лишенные произвольной области, при условии, что они обладают активностью связывания и/или активностью нейтрализации NR10.
"Антитело-димер" относится к двухвалентному фрагменту антитела, сконструированному путем слияния генов (Holliger P et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993); EP 404097; WO 93/11161, и т.д.). Антитела-димеры представляют собой димеры, состоящие из двух полипептидных цепей. В каждой из полипептидных цепей, образующих димер, VL и VH обычно связаны линкером в одной цепи. Как правило, линкер в антителе-димере является достаточно коротким, чтобы VL и VH не могли связываться друг с другом. Конкретно, количество аминокислотных остатков, образующих линкер, составляет, например, приблизительно пять остатков. Таким образом, VL и VH, кодируемые на одном полипептиде, не могут образовывать одноцепочечный фрагмент вариабельной области, и образуют димер с другим одноцепочечным фрагментом вариабельной области. В результате антитело-димер имеет два антигенсвязывающих участка.
Антитела ScFv представляют собой одноцепочечные полипептиды, полученные связыванием вариабельной области тяжелой цепи ([VH]) и вариабельной области легкой цепи ([VL]) через линкер, или сходные с ним (Huston, J. S. et al., Proc. Natl. Acad. Sci. U.S.A. (1988) 85, 5879-5883; Pluckthun "The Pharmacology of Monoclonal Antibodies" Vol. 113, eds., Resenburg and Moore, Springer Verlag, New York, pp. 269-315, (1994)). V-область H-цепи и V-область L-цепи scFv могут быть получены из любого антитела, описанного в настоящем документе. Пептидный линкер для связывания V-областей конкретно не ограничен. Например, в качестве линкера можно использовать произвольный одноцепочечный пептид, содержащий приблизительно от трех до 25 остатков. Конкретно, можно использовать пептидные линкеры или сходные с ними, описанные ниже.
V-области обеих цепей можно связывать, например, посредством ПЦР, как описано выше. Сначала в качестве матрицы для связывания V-областей посредством ПЦР используют ДНК, кодирующую полную или желательную неполную аминокислотную последовательность, среди следующих ДНК:
последовательность ДНК, кодирующая H-цепь или V-область H-цепи антитела, и
последовательность ДНК, кодирующая L-цепь или V-область L-цепи антитела.
ДНК, кодирующие V-области H-цепи и L-цепи, амплифицируют посредством ПЦР с использованием пары праймеров, имеющих последовательности, соответствующие последовательностям на обоих концах ДНК, подлежащей амплификации. Затем получают ДНК, кодирующую часть, представляющую собой пептидный линкер. ДНК, кодирующую пептидный линкер, также можно синтезировать посредством ПЦР. В данном случае нуклеотидные последовательности, которые можно лигировать с продуктами амплификации V-областей, синтезированных по отдельности, добавляют к 5'-концу праймеров, подлежащих применению. Затем проводят ПЦР с использованием каждой ДНК [ДНК V-области H-цепи] - [ДНК пептидного линкера] - [ДНК V-области L-цепи], и праймеров для ПЦР со сборкой.
Праймеры для ПЦР со сборкой состоят из комбинации праймера, который подвергается отжигу с 5'-концом [ДНК V-области H-цепи], и праймера, который подвергается отжигу с 3'-концом [ДНК V-области L-цепи]. Иными словами, праймеры для ПЦР со сборкой представляют собой набор праймеров, которые можно использовать для амплификации ДНК, кодирующей полноразмерную последовательность scFv, подлежащего синтезу. При этом нуклеотидные последовательности, которые можно лигировать с ДНК V-областей добавляют к [ДНК пептидного линкера]. Таким образом, эти ДНК связываются, а затем образуется целый scFv в качестве продукта амплификации с помощью праймеров для ПЦР со сборкой. После получения ДНК, кодирующих scFv, можно получать экспрессирующие векторы, обладающие этими ДНК, и рекомбинантные клетки, трансформированные этими экспрессирующими векторами, общепринятыми способами. Более того, scFv можно получать культивированием полученных рекомбинантных клеток для экспрессии ДНК, кодирующих scFv.
Порядок связывания вариабельных областей тяжелой цепи и легкой цепи конкретно не ограничен, и они могут быть расположены в любом порядке. Примеры расположения приведены ниже.
[VH] линкер [VL]
[VL] линкер [VH]
sc(Fv)2 представляет собой одноцепочечное низкомолекулярное антитело, получаемое связыванием двух VH и двух VL с использованием линкеров и сходных с ними (Hudson et al., J Immunol. Methods 1999; 231: 177-189). Например, sc(Fv)2 можно получать связыванием scFv через линкер.
Предпочтительными являются антитела, в которых две VH и две VL расположены в порядке VH-VL-VH-VL ([VH] линкер [VL] линкер [VH] линкер [VL]) от N-конца одноцепочечного полипептида. Однако порядок двух VH и двух VL конкретно не ограничен указанным выше расположением, и они могут быть расположены в любом порядке. Примеры расположения приведены ниже:
[VL] линкер [VH] линкер [VH] линкер [VL]
[VH] линкер [VL] линкер [VL] линкер [VH]
[VH] линкер [VH] линкер [VL] линкер [VL]
[VL] линкер [VL] линкер [VH] линкер [VH]
[VL] линкер [VH] линкер [VL] линкер [VH]
Аминокислотная последовательность вариабельной области тяжелой цепи или вариабельной области легкой цепи в низкомолекулярном антителе может содержать замену, делецию, добавление и/или вставку. Более того, вариабельная область тяжелой цепи и вариабельная область легкой цепи также могут быть лишены некоторых частей или в них могут быть добавлены другие полипептиды, при условии, что они обладают способностью связывать антиген при связывании вместе. Альтернативно, вариабельные области могут быть химеризованными или гуманизированными.
В настоящем изобретении линкеры, которые связывают вариабельные области антитела, включают произвольные пептидные линкеры, которые можно встраивать с использованием способов генетической инженерии, или синтетические линкеры, такие как линкеры, описанные в Protein Engineering, 9(3), 299-305, 1996.
Предпочтительными линкерами в настоящем изобретении являются пептидные линкеры. Длина пептидных линкеров конкретно не ограничена, и специалисты в данной области могут соответствующим образом выбрать длину в зависимости от цели. Типичная длина составляет от одной до 100 аминокислот, предпочтительно от 3 до 50 аминокислот, более предпочтительно от 5 до 30 аминокислот, и особенно предпочтительно от 12 до 18 аминокислот (например, 15 аминокислот).
Аминокислотные последовательности таких пептидных линкеров включают, например:
Ser;
Gly-Ser;
Gly-Gly-Ser;
Ser-Gly-Gly;
Gly-Gly-Gly-Ser (SEQ ID NO: 82);
Ser-Gly-Gly-Gly (SEQ ID NO: 83);
Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 84);
Ser-Gly-Gly-Gly-Gly (SEQ ID NO: 85);
Gly-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 86);
Ser-Gly-Gly-Gly-Gly-Gly (SEQ ID NO: 87);
Gly-Gly-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 88);
Ser-Gly-Gly-Gly-Gly-Gly-Gly (SEQ ID NO: 89);
(Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 84))n; и
(Ser-Gly-Gly-Gly-Gly (SEQ ID NO: 85))n,
где n представляет собой целое число, равное 1 или более.
Аминокислотную последовательность пептидного линкера могут соответствующим образом выбирать специалисты в данной области, в зависимости от цели. Например, вышеуказанный "n", который определяет длину пептидного линкера, обычно составляет от 1 до 5, предпочтительно от 1 до 3, и более предпочтительно 1 или 2.
Синтетические линкеры (химические поперечно-сшивающие агенты) включают поперечно-сшивающие агенты, которые обычно используют для поперечного сшивания пептидов, например, N-гидроксисукцинимид (NHS), дисукцинимидилсуберат (DSS), бис(сульфосукцинимидил)суберат (BS3), дитиобис(сукцинимидилпропионат) (DSP), дитиобис(сульфосукцинимидилпропионат) (DTSSP), этиленгликоль-бис(сукцинимидилсукцинат) (EGS), этиленгликоль-бис(сульфосукцинимидилсукцинат) (сульфо-EGS), дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST), бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES), и бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES). Эти поперечно-сшивающие агенты являются коммерчески доступными.
Когда связывают четыре вариабельных области антитела, обычно требуется три линкера. Такое множество линкеров могут быть одинаковыми или могут отличаться.
Антитела по настоящему изобретению включают антитела, в которых к аминокислотной последовательности антитела по настоящему изобретению добавлены один или несколько аминокислотных остатков. Кроме того, к настоящему изобретению относятся слитые белки, которые получены слиянием одного из описанных выше антител и второго пептида или белка. Эти слитые белки можно получать лигированием полинуклеотида, кодирующего антитело по настоящему изобретению, и полинуклеотида, кодирующего второй пептид или полипептид, в рамке считывания, встраиванием его с экспрессирующий вектор и экспрессией слитой конструкции в хозяине. Для этой цели доступны некоторые способы, известные специалистам в данной области. Пептид- или полипептид-партнер, слитый с антителом по настоящему изобретению, может представлять собой известный пептид, например, FLAG (Hopp, T. P. et al., BioTechnology 6 1204-1210 (1988)), 6x His, состоящий из шести остатков His (гистидина), 10x His, гемагглютинин вируса гриппа (HA), фрагмент c-myc человека, фрагмент VSV-GP, фрагмент p18HIV, T7-метку, HSV-метку, E-метку, фрагмент T-антигена SV40, lck-метку, фрагмент α-тубулина, B-метку, фрагмент белка C. Другие полипептиды-партнеры, подлежащие слиянию с антителами по настоящему изобретению, включают, например, GST (глутатион-S-трансферазу), HA (гемагглютинин вируса гриппа), константную область иммуноглобулина, β-галактозидазу и MBP (связывающий мальтозу белок). Полинуклеотид, кодирующий один из этих коммерчески доступных пептидов или полипептидов, можно подвергать слиянию с полинуклеотидом, кодирующим антитело по настоящему изобретению. Слитный полипептид можно получать путем экспрессии слитой конструкции.
Более того, антитела по настоящему изобретению могут представлять собой конъюгированные антитела, которые связаны с любыми из различных молекул, включающих полимерные вещества, такие как полиэтиленгликоль (PEG) и гиалуроновая кислота, радиоактивные вещества, флуоресцентные вещества, люминесцентные вещества, ферменты и токсины. Такие конъюгированные антитела можно получать путем химической модификации полученных антител. Способы модификации антител являются общепризнанными в данной области (например, US 5057313 и US 5156840). "Антитела" по настоящему изобретению также включают такие конъюгированные антитела.
Более того, антитела, используемые в настоящем изобретении, могут представлять собой биспецифические антитела. Биспецифическое антитело относится к антителу, которое имеет вариабельные области, распознающие различные эпитопы, в одной молекуле антитела. В настоящем изобретении биспецифические антитела могут распознавать различные эпитопы на молекуле NR10, или распознавать NR10 одним антигенсвязывающим центром и отличающуюся молекулу другим антигенсвязывающим центром.
Способы получения биспецифических антител известны. Биспецифические антитела можно получать, например, связыванием двух антител, которые распознают различные антигены. Антитела, подлежащие связыванию, могут представлять собой половины молекул, каждая из которых содержит H-цепь и L-цепь, или четверти молекул, которые состоят только из одной H-цепи. Альтернативно гибридомы, продуцирующие различные моноклональные антитела, можно подвергать слиянию с получением продуцирующей биспецифическое антитело слитой клетки. Более того, биспецифические антитела можно получать способами генетической инженерии.
Антитела по настоящему изобретению могут отличаться по аминокислотной последовательности, молекулярной массе, изоэлектрической точке, наличию/отсутствию цепей сахаров и конформации, в зависимости от клетки или хозяина, продуцирующих антитело, или способа очистки, как описано ниже. Однако полученное антитело относится к настоящему изобретению, при условии, что оно является функционально эквивалентным антителу по настоящему изобретению. Например, когда антитело по настоящему изобретению экспрессируется в прокариотических клетках, например E. coli, на N-конец исходной аминокислотной последовательность антитела добавляют остаток метионина. Такие антитела относятся к настоящему изобретению.
Продукция антител
Антитела по настоящему изобретению могут представлять собой поликлональные или моноклональные антитела. Такие моноклональные антитела, обладающие активностью связывания и/или нейтрализации NR10, можно получать, например, следующим способом: моноклональные антитела против NR10 получают с использованием в качестве антигена NR10 или его фрагмента, происходящих из млекопитающего, такого как человека или мышь, известными способами, а затем из полученных таким образом моноклональных антител против NR10 отбирают антитела, обладающие активностью связывания и/или нейтрализации NR10. В частности, для иммунизации в соответствии с общепринятыми способами иммунизации в качестве сенсибилизирующего антигена используют желаемый антиген или клетки, экспрессирующие желаемый антиген. Моноклональные антитела против NR10 можно получать путем слияния полученных иммунных клеток с известными исходными клетками с использованием общепринятых способов слияния клеток, и скрининга их в отношении продуцирующих моноклональное антитело клеток (гибридом) общепринятыми способами скрининга. Животные, подлежащие иммунизации, включают, например, млекопитающих, таких как мыши, крысы, кролики, овцы, обезьяны, козы, ослы, коровы, лошади и свиньи. Антиген можно получать с использованием известной последовательности гена NR10 в соответствии с известными способами, например, способами с использованием бакуловируса (например, WO 98/46777).
Гибридомы можно получать, например, в соответствии со способом Milstein et al. (Kohler, G. and Milstein, C., Methods Enzymol. (1981) 73: 3-46) или сходными с ним. Когда иммуногенность антигена является низкой, иммунизацию можно проводить после связывания антигена с макромолекулой, обладающей иммуногенностью, такой как альбумин.
Варианты осуществления антител по настоящему изобретению, которые обладают активностью связывания и/или нейтрализации NR10, включают моноклональные антитела, которые обладают активностью связывания и/или нейтрализации NR10 человека. Антигены, используемые для получения моноклональных антител, которые обладают активностью связывания и/или нейтрализации NR10 человека, конкретно не ограничены, при условии, что они обеспечивают получение антител, которые обладают активностью связывания и/или нейтрализации NR10 человека. Например, известно, что существует ряд вариантов NR10 человека, и в качестве иммуногена можно использовать любой вариант, при условии, что он обеспечивает получение антител, которые обладают активностью связывания и/или нейтрализации NR10 человека. Альтернативно в тех же условиях, в качестве иммуногена можно использовать пептидный фрагмент NR10 или белок, в котором в природную последовательность NR10 внесены искусственные мутации. NR10.3 человека является одним из предпочтительных иммуногенов для получения антител, которые обладают активностью связывания и/или нейтрализации NR10, по настоящему изобретению.
Более того, активность связывания и/или нейтрализации антитела против NR10 можно измерять, например, путем выявления эффекта подавления роста IL-31-зависимой клеточной линии, как описано в разделе "Примеры".
При этом моноклональные антитела также можно получать путем иммунизации посредством ДНК. Иммунизация посредством ДНК представляет собой способ, в котором векторную ДНК, сконструированную так, чтобы ген, кодирующий антигенный белок, мог экспрессироваться у животного, подлежащего иммунизации, вводят животному, и в организме животного экспрессируется иммуноген, обеспечивая стимуляцию иммунной системы. По сравнению с общепринятыми способами иммунизации на основе введения белковых антигенов, ожидается, что иммунизация посредством ДНК является преимущественной в том, что:
- она обеспечивает стимуляцию иммунной системы при сохранении структуры мембранного белка; и
- не требуется очистка иммуногена.
С другой стороны, трудно комбинировать иммунизацию посредством ДНК со средством для стимуляции иммунной системы, таким как адъювант.
Для получения моноклонального антитела посредством иммунизации ДНК, животному, подлежащему иммунизации, сначала вводят ДНК, кодирующую NR10. ДНК, кодирующую NR10, можно синтезировать известными способами, такими как ПЦР. Полученную ДНК встраивают в соответствующий экспрессирующий вектор и вводят животному, подлежащему иммунизации. Экспрессирующие векторы, которые можно использовать, включают коммерчески доступные экспрессирующие векторы, такие как pcDNA3.1. Вектор можно вводить в живой организм общепринятыми способами. Например, иммунизмацию ДНК можно проводить путем введения золотых частиц, покрытых экспрессирующим вектором, в клетки с помощью генной пушки. Вспомогательное введение с использованием экспрессирующих NR10 клеток после иммунизации ДНК, представляет собой предпочтительный способ получения моноклонального антитела.
После иммунизации млекопитающего, как описано выше и подтверждения повышения уровня желаемого антитела в сыворотке, собирают иммунные клетки млекопитающего и подвергают их слиянию. Предпочтительными иммунными клетками, в частности, являются клетки селезенки.
Для слияния с указанными выше клетками иммунной системы используют миеломные клетки млекопитающих. Предпочтительно, чтобы миеломные клетки имели соответствующие селективные маркеры для скрининга. Селективный маркер относится к фенотипу, который позволяет (или не позволяет) выживание в конкретных условиях культивирования. Известные селективные маркеры включают дефицит гипоксантингуанинфосфорибозилтрансферазы (в дальнейшем в настоящем документе сокращаемый как "дефицит HGPRT") и дефицит тимидинкиназы (в дальнейшем в настоящем документе сокращаемый как "дефицит TK"). Дефицитные по HGPRT или TK клетки проявляют чувствительность к гипоксантин-аминоптерин-тимидину (в дальнейшем в настоящем документе обозначаемую как "чувствительность к HAT"). В селективной среде HAT чувствительные к HAT клетки не могут синтезировать ДНК и, таким образом, погибают. Однако при слиянии с нормальными клетками они могут продолжать синтезировать ДНК через каскад спасения нормальных клеток и, таким образом, могут расти даже в селективной среде HAT.
Дефицитные по HGPRT или TK клетки можно отбирать с использованием среды, содержащей 6-тиогуанин, 8-азагуанин (в дальнейшем в настоящем документе сокращаемый как "8AG"), или 5'-бромдезоксиуридин. В то время как нормальные клетки погибают вследствие включения этих аналогов пиримидинов в ДНК, клетки, лишенные этих ферментов, могут выживать в селективной среде, поскольку они не могут встраивать эти аналоги пиримидинов. Другой селективный маркер, называемый маркером устойчивости к G418, придает устойчивость к 2-дезоксистрептаминовым антибиотикам (аналогам гентамицина) благодаря гену устойчивости к неомицину. Различные миеломные клетки, пригодные для слияния клеток, известны.
Слияние иммунных клеток и миеломных клеток можно по существу проводить в соответствии с известными способами, например, способом Kohler и Milstein (Kohler, G. and Milstein, C., Methods Enzymol. (1981) 73, 3-46).
Более конкретно, слияние клеток можно проводить, например, в обычной культуральной среде в присутствии средства, обеспечивающего слияние клеток. Средство, обеспечивающее слияние клеток, включает, например, полиэтиленгликоль (PEG) и вирус Сендай (HVJ). При необходимости, также можно добавлять вспомогательное средство, такое как диметилсульфоксид, для повышения эффективности слияния.
Иммунные клетки и миеломные клетки можно использовать в произвольно определенном соотношении. Например, соотношение иммунных клеток и миеломных клеток предпочтительно составляет от 1 до 10. Культуральные среды, используемые для слияния клеток, включают, например, среды, которые пригодны для роста клеток миеломной клеточной линии, такие как RPMI 1640 и MEM, и другие общепринятые культуральные среды, используемые для этого типа клеточной культуры. Кроме того, культуральные среды также можно дополнять сывороткой, такой как эмбриональная телячья сыворотка (FCS).
Заданные количества иммунных клеток и миеломных клеток хорошо перемешивают в культуральной среде, а затем смешивают с раствором PEG, предварительно нагретым до 37°C, с получением слитых клеток (гибридом). В способе слияния клеток, например, PEG со средней молекулярной массой приблизительно 1000-6000 можно добавлять к клеткам, как правило, в концентрации от 30% до 60% (масс./об.). Затем несколько раз проводят последовательное добавление соответствующей культуральной среды, приведенной выше, и удаление супернатанта центрифугированием для устранения средства для слияния клеток и прочих, которые являются неблагоприятными для роста гибридом.
Полученные гибридомы можно подвергать скринингу с использованием селективной среды в соответствии с селективным маркером, которым обладают миеломные клетки, используемые для слияния клеток. Например, дефицитные по HGPRT или TK клетки можно подвергать скринингу путем культивирования их в среде HAT (среде, содержащей гипоксантин, аминоптерин и тимидин). Конкретно, если для слияния клеток используют чувствительные к HAT миеломные клетки, то клетки, успешно слитые с нормальными клетками, можно селективно выращивать в среде HAT. Культивирование клеток с использованием указанной выше среды HAT продолжают в течение периода времени, достаточного для гибели всех клеток (неслитых клеток), за исключением требуемых гибридом. Конкретно, как правило, требуемые гибридомы можно отбирать культивированием клеток в течение от нескольких суток до нескольких недель. Затем можно проводить скрининг и клонирование по отдельности гибридом, которые продуцируют представляющее интерес антитело, проведением обычных способов лимитирующих разведений. Альтернативно антитела, которые распознают NR10, можно получать способом, описанным в WO 03/104453.
Скрининг и отдельное клонирование представляющего интерес антитела можно пригодным образом проводить с помощью известных способов скрининга на основе реакции антиген-антитело. Например, антиген связывают с носителем, таким как гранулы, изготовленные из полистирола или сходных с ним, и коммерчески доступные 96-луночные микропланшеты для титрования, а затем подвергают реакции с культуральным супернатантом гибридомы. Далее, носитель промывают, а затем подвергают реакции с вторичным антителом, меченым ферментом или сходными с ним. Когда культуральный супернатант содержит представляющее интерес антитело, реактивное к сенсибилизирующему антигену, вторичное антитело связывается с носителем через это антитело. Наконец, проводят детекцию вторичного антитела, связанного с носителем, для определения того, содержит ли культуральный супернатант представляющее интерес антитело. Гибридомы, продуцирующие желаемое антитело, способное связываться с антигеном, можно клонировать способом лимитирующих разведений или сходными с ним. Для этой цели в качестве антигена можно использовать не только антиген, используемый для иммунизации, но также белок NR10, по существу эквивалентный ему. Например, в качестве антигена можно использовать клеточную линию, экспрессирующую NR10, внеклеточный домен NR10 или олигопептид, состоящий из неполной аминокислотной последовательности, составляющей домен.
В дополнение к описанному выше способу получения гибридомы с помощью иммунизации не являющегося человеком животного антигеном, представляющие интерес антитела также можно получать сенсибилизацией лимфоцитов человека антигеном. Конкретно, сначала лимфоциты человека сенсибилизируют белком NR10 in vitro. Затем сенсибилизированные лимфоциты подвергают слиянию с соответствующим партнером по слиянию. Например, в качестве партнера по слиянию можно использовать происходящие из человека миеломные клетки со способностью к непрерывному делению (см. публикацию японской патентной заявки Kokoku No. (JP-B) H1-59878 (рассмотренную, принятую японскую патентную заявку, опубликованную для опротестования). Антитела, полученные этим способом, представляют собой антитела человека, обладающие активностью связывания белка NR10.
Нуклеотидную последовательность, кодирующую антитело против NR10, полученную описанным выше способом или сходным с ним, и его аминокислотную последовательность можно получать способами, известными специалистам в данной области. Аминокислоты, содержащиеся в описанных аминокислотных последовательностях согласно настоящему изобретению, могут претерпевать посттрансляционную модификацию (например, модификация N-концевого глутамина в пироглутаминовую кислоту путем пироглутамилирования хорошо известна специалистам в данной области). В природных условиях такие последовательности с посттрансляционно модифицированными аминокислотами также относятся к описанным аминокислотным последовательностям согласно настоящему изобретению.
Исходя из полученной последовательности антитела против NR10, можно получать антитело против NR10, например, способами генетической рекомбинации, известными специалистам в данной области. Конкретно, на основе последовательности распознающего NR10 антитела можно конструировать полинуклеотид, кодирующий антитело, встраивать его в экспрессирующий вектор, а затем экспрессировать в соответствующих клетках-хозяевах (см. например, Co, M. S. et al., J. Immunol. (1994) 152, 2968-2976; Better, M. and Horwitz, A. H., Methods Enzymol. (1989) 178, 476-496; Pluckthun, A. and Skerra, A., Methods Enzymol. (1989) 178, 497-515; Lamoyi, E., Methods Enzymol. (1986) 121, 652-663; Rousseaux, J. et al., Methods Enzymol. (1986) 121, 663-669; Bird, R. E. and Walker, B. W., Trends Biotechnol. (1991) 9, 132-137).
Векторы включают векторы M13, векторы pUC, pBR322, pBluescript и PCR-Script. Альтернативно, когда целью является субклон и вырезанная кДНК, векторы включают, например, pGEM-T, pDIRECT и pT7, в дополнение к векторам, описанным выше. Экспрессирующие векторы особенно пригодны при использовании векторов для продукции антител по настоящему изобретению. Например, когда целью является экспрессия в E. coli, таких как JM109, DH5α, HB101 и XL1-Blue, экспрессирующие векторы не только обладают описанными выше характеристиками, которые позволяют амплификацию вектора в. coli, но большинство из них также обладают промотором, который позволяет эффективную экспрессию в E. coli, например, промотором lacZ (Ward et al., Nature (1989) 341, 544-546; FASEB J. (1992) 6, 2422-2427), промотором araB (Better et al., Science (1988) 240, 1041-1043), промотором T7 или сходными с ним. Такие векторы включают pGEX-5X-1 (Pharmacia), "систему QIAexpress" (Qiagen), pEGFP или pET (в этом случае хозяином предпочтительно являются BL21, которые экспрессируют РНК-полимеразу T7), в дополнение к векторам, описанным выше.
Векторы могут содержать сигнальные последовательности для секреции антитела. В качестве сигнальной последовательности для секреции антитела можно использовать сигнальную последовательность pelB (Lei, S. P. et al J. Bacteriol. (1987) 169, 4379), когда белок секретируется в периплазму E. coli. Вектор можно вводить в клетки-хозяева, например, способами с хлоридом кальция или электропорации.
В дополнение к векторам для E. coli, векторы для продукции антител по настоящему изобретению включают, например, экспрессирующие векторы млекопитающих (например, pcDNA3 (Invitrogen), pEF-BOS (Nucleic Acids Res. 1990, 18(17), p5322), pEF и pCDM8), экспрессирующие векторы из насекомых (например, "бакуловирусную систему Bac-to-BAC" (Gibco-BRL) и pBacPAK8), экспрессирующие векторы из растений (например, pMH1 и pMH2), экспрессирующие векторы из животных (например, pHSV, pMV и pAdexLcw), ретровирусные экспрессирующие векторы (например, pZIPneo), экспрессирующие векторы дрожжей (например, "Pichia Expression Kit" (Invitrogen), pNV11 и SP-Q01) и экспрессирующие векторы Bacillus subtilis (например, pPL608 и pKTH50).
Когда целью является экспрессия в клетках животных, таких как клетки CHO, COS и NIH3T3, векторы должны иметь промотор, необходимый для экспрессии в клетках, например, промотор SV40 (Mulligan et al., Nature (1979) 277, 108), промотор MMLV-LTR, промотор EF1α (Mizushima et al., Nucleic Acids Res. (1990) 18, 5322), и промотор CMV, и более предпочтительно они имеют ген для селекции трансформированных клеток (например, ген устойчивости к лекарственному средству, который позволяет оценку с использованием средства (неомицина, G418 или сходных с ними). Векторы с такими характеристиками включают, например, pMAM, pDR2, pBK-RSV, pBK-CMV, pOPRSV и pOP13.
Кроме того, для стабильной экспрессии генов и амплификации генов в клетках можно использовать следующий способ: в клетки CHO, дефицитные по каскаду синтеза нуклеиновых кислот, вводят вектор (например, pSV2-dhfr (Molecular Cloning 2nd edition, Cold Spring Harbor Laboratory Press, 1989)), который обладает геном DHFR, который компенсирует дефицит, и вектор амплифицируют с использованием метотрексата (MTX). Альтернативно для временной экспрессии гена используют следующий способ: клетки COS с геном, экспрессирующим T-антиген SV40, на их хромосоме, трансформируют вектором (pcD и т.п.) с ориджином репликации SV40. Также можно использовать ориджины репликации, происходящие из вируса полиомы, аденовируса, вируса папилломы крупного рогатого скота (BPV) и т.п. Для амплификации числа копий гена в клетках-хозяевах экспрессирующие векторы могут дополнительно иметь селективные маркеры, такие как ген аминогликозидтрансферазы (APH), ген тимидинкиназы (TK), ген ксантин-гуанин-фосфорибозилтрансферазы E. coli (Ecogpt) и ген дигидрофолатредуктазы (dhfr).
Таким образом, настоящее изобретение относится к способам получения полипептидов по настоящему изобретению, кодируемых генами, кодирующими полипептиды по настоящему изобретению, которые включают стадию культивирования клеток-хозяев, содержащих вектор, в который встроен полинуклеотид, кодирующий полипептид по настоящему изобретению.
Более конкретно, настоящее изобретение относится к способам получения полипептидов по настоящему изобретению, включающим стадии:
(a) культивирования клетки-хозяина, содержащей вектор, в который встроен ген, кодирующий полинуклеотид по настоящему изобретению, и
(b) получение полипептида, кодируемого геном.
Антитела по настоящему изобретению, полученные способами, описанными выше, можно выделять из клеток-хозяев или из окружения клеток (из среды или сходных с ней), и очищать до гомогенного состояния. Антитела можно выделять и очищать способами, обычно используемыми для выделения и очистки антител, и тип этого способа не ограничен. Например, антитела можно выделять и очищать путем надлежащего выбора и комбинирования колоночной хроматографии, фильтрации, ультрафильтрации, высаливания, осаждения растворителем, экстракции растворителем, дистилляции, иммунопреципитации, SDS-полиакриламидного гель-электрофореза, изоэлектрофокусирования, диализа, перекристаллизации и т.п.
Способы хроматографии включают, например, аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрацию, обращенно-фазовую хроматографию и адсорбционную хроматографию (Strategies for Protein Purification and Characterization: A Laboratory Course Manual. Ed Daniel R. Marshak et al., Cold Spring Harbor Laboratory Press, 1996). Способы хроматографии, описанные выше, можно проводить с использованием жидкостной хроматографии, например, ВЭЖХ и FPLC. Колонки, которые можно использовать для аффинной хроматографии, включают колонки с белком A и колонки с белком G. Колонки с использованием белка A включают, например, Hyper D, POROS и Sepharose FF (GE Amersham Biosciences). Настоящее изобретение включает антитела, которые являются в высокой степени очищенными с использованием этих способов очистки.
Активность связывания NR10 полученных антител можно определять способами, известными специалистам в данной области. Способы определения антигенсвязывающей активности антитела включают, например, ELISA (твердофазный иммуноферментный анализ), EIA (иммуноферментный анализ), RIA (радиоиммунный анализ) и способ флуоресцентных антител. Например, при использовании иммуноферментного анализа, содержащие антитело образцы, такие как очищенные антитела и культуральные супернатанты антителопродуцирующих клеток, добавляют в покрытые антигеном планшеты. Добавляют вторичное антитело, меченное ферментом, таким как щелочная фосфатаза, и планшеты инкубируют. После промывания добавляют субстрат фермента, такой как п-нитрофенилфосфат, и измеряют поглощение для оценки антигенсвязывающей активности.
Фармацевтические композиции
Настоящее изобретение также относится к фармацевтическим композициям, содержащим антитело, упомянутое выше, в качестве активного ингредиента. Более того, настоящее изобретение относится к лекарственным средствам от воспалительных заболеваний, которое содержит в качестве активного ингредиента антитело, упомянутое выше.
В настоящем изобретении воспалительное заболевание относится к заболеваниям с патологическими признаками, вовлеченными в цитологические и гистологические реакции, которые возникают в пораженных кровеносных сосудах и соседних тканях в ответ на повреждение или аномальную стимуляцию, вызываемую физическими, химическими или биологическими агентами (Stedman's Medical Dictionary, 5th Ed., MEDICAL VIEW CO., 2005). Как правило, воспалительные заболевания включают дерматит (атопический дерматит, хронический дерматит и т.п.), воспалительные заболевания кишечника (колит и т.п.), астму, артрит (ревматоидный артрит, остеоартрит и т.п.), бронхит, Th2-аутоиммунные заболевания, системную красную волчанку, миастению, хроническую GVHD, болезнь Крона, деформирующий спондилит, боль в пояснице, подагру, воспаление после хирургической операции или травмы, опухание, невралгию, ларингофарингит, цистит, гепатит (неалкогольный стеатогепатит, алкогольный гепатит и т.п.), гепатит B, гепатит C, артериосклероз и зуд.
Предпочтительные примеры воспалительных заболеваний, которые являются объектами настоящего изобретения, включают атопический дерматит, хронический дерматит, ревматизм, остеоартрит, хроническую астму и зуд.
Выражение "включает(ют) антитело против NR10 в качестве активного ингредиента" означает включение антитела против NR10 в качестве, по меньшей мере, одного из активных ингредиентов, и оно не ограничивает долю антитела. Кроме того, лекарственные средства от воспалительных заболеваний по настоящему изобретению также могут содержать, в сочетании с антителом против NR10, упомянутым выше, другие ингредиенты, которые усиливают лечение воспалительных заболеваний.
Лекарственные средства по настоящему изобретению также можно использовать в целях профилактики.
Антитело против NR10 по настоящему изобретению можно изготавливать в качестве составов в соответствии со стандартными способами (см., например, Remington's Pharmaceutical Science, latest edition, Mark Publishing Company, Easton, USA). Кроме того, они могут содержать фармацевтически приемлемые носители и/или добавки, если необходимо. Например, они могут содержать поверхностно-активные вещества (например, PEG и Tween), эксципиенты, антиоксиданты (например, аскорбиновую кислоту), красители, вкусовые добавки, консерванты, стабилизаторы, буферные средства (например, фосфорную кислоту, лимонную кислоту и другие органические кислоты), хелатирующие агенты (например, ЭДТА), суспендирующие вещества, обеспечивающие изотоничность вещества, связующие вещества, дезинтегрирующие вещества, смазывающие вещества, промоторы текучести и модифицирующие вещества. Однако, не ограничиваясь ими, средства для профилактики или лечения воспалительных заболеваний по настоящему изобретению могут содержать другие широко используемые носители. Такие носители конкретно включают легкую безводную кремниевую кислоту, лактозу, кристаллическую целлюлозу, маннит, крахмал, кармелозу кальция, кармелозу натрия, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу, поливинилацетальдиэтиламиноацетат, поливинилпирролидон, желатин, триглицерид жирной кислоты средней длины цепи, полиоксиэтилен-гидрогенизированное касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал и неорганическую соль. Также средства могут содержать другие низкомолекулярные полипептиды, белки, такие как сывороточный альбумин, желатин и иммуноглобулин, и аминокислоты, такие как глицин, глутамин, аспарагин, аргинин и лизин. Когда антитело против NR10 изготавливают в качестве водного раствора для инъекции, антитело против NR10 можно растворять в изотоническом растворе, содержащем, например, физиологический раствор, декстрозу или другие адъюванты. Адъюванты могут включать, например, D-сорбит, D-маннозу, D-маннит и хлорид натрия. Кроме того, вместе с ними можно использовать соответствующие солюбилизирующие вещества, например, спирты (например, этанол), полиспирты (например, пропиленгликоли и PEG) и неионные детергенты (полисорбат 80 и HCO-50).
Если необходимо, антитела против NR10 можно инкапсулировать в микрокапсулы (микрокапсулы, изготовленные из гидроксиметилцеллюлозы, желатина, полиметилметакрилата и т.п.), и преобразовывать в компоненты коллоидных систем для доставки лекарственных средств (липосомы, микросферы с альбумином, микроэмульсии, наночастицы и нанокапсулы) (например, см. "Remington's Pharmaceutical Science 16th edition" &, Oslo Ed. (1980)). Более того, известны способы получения лекарственных средств с замедленным высвобождением, и их можно использовать для антител против NR10 (Langer et al., J. Biomed. Mater. Res. (1981) 15, 167-277; Langer, Chem. Tech. (1982) 12, 98-105; патент США No. 3773919; европейская патентная заявка (EP) No. 58481; Sidman et al., Biopolymers (1983) 22, 547-56; EP 133988).
Фармацевтические композиции по настоящему изобретению можно вводить либо перорально, либо парентерально, но предпочтительно их вводят парентерально. Конкретно, эти средства вводят пациентам путем инъекции или подкожного введения. Инъекции включают, например, внутривенные инъекции, внутримышечные инъекции и подкожные инъекции, для системного или местного введения. Средства можно вводить в области предполагаемого подавления воспаления, или в окружающие их области путем местной инфузии, в частности, внутримышечной инъекции. Способы введения можно надлежащим образом выбирать в соответствии с возрастом и состоянием пациента. Дозу для однократного введения можно выбирать, например, из диапазона от 0,0001 до 100 мг активного ингредиента на кг массы тела. Альтернативно, например, когда средства вводят пациентам-людям, дозу активного ингредиента можно выбирать из диапазона от 0,001 до 1000 мг/кг массы тела. Доза для однократного введения предпочтительно содержит, например, приблизительно от 0,01 до 50 мг/кг массы тела антитела по настоящему изобретению. Однако доза средства для профилактики или лечения воспалительных заболеваний по настоящему изобретению не ограничивается этими примерами.
Все документы уровня техники, цитированные в настоящем описании, включены в настоящий документ в качестве ссылок.
[Примеры]
Ниже настоящее изобретение описано конкретно с помощью примеров, однако его не следует истолковывать, как ограниченное ими.
[Пример 1] Получение гибридом
1.1. Получение плазмид NR10 человека и яванского макака для иммунизации посредством ДНК
1.1.1. Получение экспрессирующих векторов для hNR10 и cynNR10
NR10 человека (нуклеотидная последовательность SEQ ID NO: 75; аминокислотная последовательность SEQ ID NO: 76) встраивали в экспрессирующий вектор pMacII, который экспрессирует белок под контролем промотора β-актина мыши (WO2005/054467), для получения экспрессирующего вектора для hNR10. Аналогичным образом, конструировали экспрессирующий вектор для cynNR10 на основе NR10 яванского макака (нуклеотидная последовательность SEQ ID NO: 65; аминокислотная последовательность SEQ ID NO: 66).
1.1.2. Получение кассеты ДНК
Для применения экспрессирующего вектора для hNR10 или cynNR10, полученного согласно 1.1.1, при иммунизации мышей посредством ДНК, использовали набор Helios Gene Gun Cartridge Kit (BIO-RAD) для получения ДНК-кассеты для каждой ДНК, которая позволяет иммунизацию посредством 1 мкг ДНК за один раз.
1.2. Получение гибридом, продуцирующих антитело против NR10 человека
1.2.1. Получение гибридом с использованием мышей, иммунизированных NR10 человека или яванского макака
Мышей Balb/c (самки; в возрасте шести недель в начале иммунизации; Charles River Laboratories Japan) иммунизировали NR10 человека или яванского макака следующим способом. Для первичной иммунизации мышей иммунизировали ДНК-кассетой, полученной с экспрессирующим вектором для hNR10 с использованием Helios Gene Gun System (BIO-RAD). Через одну неделю проводили вторичную иммунизацию с помощью Helios Gene Gun System (BIO-RAD) с использованием ДНК-кассеты, полученной с экспрессирующим вектором для cynNR10. Третью и последующие иммунизации проводили с интервалами в одну неделю с использованием экспрессирующих векторов для hNR10 и cynNR10 поочередно. После подтверждения повышения титров антител в сыворотке против NR10 человека, внутривенно вводили белок NR10 человека (внеклеточный домен) (справочный пример 4), разбавленный PBS(-), в количестве 10 мкг/мышь в качестве конечной иммунизации. Через четверо суток после конечной иммунизации клетки селезенки мыши подвергали слиянию с клетками миеломы мыши P3X63Ag8U.1 (сокращенно P3U1; ATCC CRL-1597) общепринятым способом с использованием PEG1500 (Roche Diagnostics). Полученные слитые клетки, т.е. гибридомы, культивировали в RPMI1640, дополненной 10% FBS (в дальнейшем в настоящем документе сокращаемой как 10% FBS/RPMI1640).
1.2.2. Отбор гибридом
На следующий день после слияния слитые клетки суспендировали в полутвердой среде (StemCells), и культивировали для селекции, а также колонизации гибридом.
После слияния в течение девяти или десяти суток, колонии гибридомы извлекали, и каждую колонию высевали в каждую лунку 96-луночных планшетов, содержащих селективную среду HAT (10% FBS/RPMI1640, 2% об. 50x концентрата HAT (Dainippon Pharmaceurical), и 5% об. BM-Condimed H1 (Roche Diagnostics)). После культивирования в течение трех суток, культуральные супернатанты собирали из каждой лунки для определения концентрации IgG мыши в супернатанте. Культуральные супернатанты, в которых были выявлены IgG мыши, оценивали с точки зрения активности нейтрализации с использованием зависимой от IL-31 человека клеточной линии (клетки hNR10/hOSMR/BaF3; справочный пример 2), и было получено несколько клонов, имеющих сильную активность нейтрализации NR10 (фиг. 3). Получали клоны, которые подавляют индуцируемый IL-31 человека рост клеток зависимым от концентрации образом и подавляют индуцируемый IL-31 яванского макака рост клеток (клетки cynNR10/cynOSMR/BaF3; справочный пример 2) зависимым от концентрации образом (фиг. 4).
[Пример 2] Получение химерного антитела
Получение экспрессирующих векторов для химерных антител
Из гибридом экстрагировали тотальную РНК с использованием набора RNeasy Mini Kits (QIAGEN), и с нее синтезировали кДНК с использованием набора SMART RACE cDNA Amplification Kit (BD Biosciences). Гены вариабельных областей антитела выделяли с помощью ПЦР с использованием ДНК-полимеразы PrimeSTAR HS (TaKaRa), 10× универсальной смеси праймеров A, прилагаемой к набору SMART RACE cDNA Amplification Kit (BD Biosciences), и праймеров, сконструированных для каждой константной области антитела (H-цепь, mIgG1-rnot; L-цепь, mIgK-rnot). Нуклеотидную последовательность каждого выделенного фрагмента ДНК определяли с помощью секвенатора ДНК ABI PRISM 3730xL или секвенатора ДНК ABI PRISM 3700 (Applied Biosystems), с использованием набора BigDye Terminator Cycle Sequencing Kit (Applied Biosystems) в соответствии со способом, описанным в прилагаемой инструкции. Определенные аминокислотные последовательности вариабельных областей H-цепи и L-цепи в антителах мыши NS18, NS22, NS23 и NS33 представлены на фиг. 1 и 2, соответственно.
Каждый из полученных фрагментов H- и L-цепи повергали ПЦР с использованием ДНК-полимеразы PrimeSTAR HS (TaKaRa) и наборов праймеров, представленных в таблице 1. Полученные амплифицированные фрагменты лигировали с константной областью (γ1 или γ2 человека, и κ человека, соответственно), а затем встраивали в экспрессирующий вектор клеток животных. Нуклеотидную последовательность каждого фрагмента ДНК определяли с помощью ДНК-секвенатора ABI PRISM 3730xL или ДНК-секвенатора ABI PRISM 3700 (Applied Biosystems), с использованием BigDye Terminator Cycle Sequencing Kit (Applied Biosystems) согласно способу, описанному в прилагаемой инструкции.
Получение химерного антитела
Клеточную эмбриональную линию рака почки человека HEK293H (Invitrogen) суспендировали в DMEM (Invitrogen), дополненной 10% эмбриональной телячьей сывороткой (Invitrogen), и 10 мл клеток высевали в чашки для прикрепляющихся клеток (диаметром 10 см; CORNING) с плотностью клеток 6×105 клеток/мл. Клетки инкубировали в инкубаторе с CO2 (37ºC, 5% CO2) в течение одного целого дня и ночи. Затем среду удаляли аспирированием и добавляли 6,9 мл среды CHO-S-SFMII (Invitrogen). К полученной смеси плазмидной ДНК (всего 13,8 мкг) добавляли среду CHO-S-SFMII до объема 700 мкл. Этот раствор смешивали с 20,7 мкл полиэтиленимина (Polysciences Inc.) в концентрации 1 мкг/мл, и позволяли ему стоять при комнатной температуре в течение 10 минут. Раствор добавляли к клеткам в каждой чашке. Клетки инкубировали в инкубаторе с CO2 (37ºC, 5% CO2) в течение от четырех до пяти часов. Затем добавляли 6,9 мл среды CHO-S-SFMII (Invitrogen), и клетки инкубировали в инкубаторе CO2 в течение от трех до четырех суток. Культуральные супернатанты собирали, а затем центрифугировали (приблизительно 2000 g, пять минут, комнатная температура) для удаления клеток. Супернатанты фильтровали через 0,22-мкм фильтр MILLEX®-GV (Millipore). Каждый образец хранили при 4ºC до применения. Антитела очищали из супернатантов с использованием белка G-сефарозы (Amersham Biosciences). Очищенные антитела концентрировали с помощью Amicon Ultra 15 (Millipore), а затем растворитель заменяли PBS(-), содержащим 0,05% NaN3 с использованием колонок для обессоливания PD-10 Desalting columns (Amersham Biosciences). Измеряли поглощение при 280 нм с помощью спектрофотометра ND-1000 (NanoDrop), и концентрации определяли способом Pace et al. (Protein Science (1995) 4: 2411-2423).
Оценка активности химерного NS22
Активность нейтрализации hIL-31 оценивали с использованием клеточной линии hNR10/hOSMR/BaF3, которая растет зависимым от дозы hIL-31 образом, как описано ниже.
Клетки hNR10/hOSMR/BaF3 получали в количестве 1,5×105 клеток/мл с использованием среды RPMI1640 (GIBCO), содержащей 10% FBS (MOREGATE) и 1% пенициллин-стрептомицин (Invitrogen). К аликвотам клеток добавляли hIL-31 (R&D Systems) до конечной концентрации 4 нг/мл (IL-31(+); конечная концентрация: 2 нг/мл). Оставшуюся клеточную суспензию использовали в качестве IL-31(-). Очищенный NS22 доводили до концентрации 2 мкг/мл с использованием среды, и получали восемь серийных разведений с общим отношением разведения, равным 3 (конечная концентрация: 1 мкг/мл или менее). 50 мкл каждого из суспензии клеток и разведения химерного NS22 (γ1, κ человека) добавляли в каждую лунку 96-луночных планшетов с плоским дном (CORNING), и клетки культивировали в инкубаторе с 5% CO2 при 37°C в течение двух суток. После культивирования в каждую лунку добавляли 20 мкл смеси равных количеств набора Cell Counting Kit-8 (Dojindo) и PBS, и измеряли поглощение (450 нм/620 нм) (TECAN, SUNRISE CLASSIC). После того, как реакции позволяли продолжаться в течение двух часов в инкубаторе с 5% CO2 при 37ºC, поглощение вновь измеряли. Нейтрализующую активность NS22 представляли в качестве скорости ингибирования с использованием полученного значения путем вычитания значения для 0 часов из значения для 2 часов. Результат показал, что NS22 подавлял индуцируемый IL-31 рост клеточной линии hNR10/hOSMR/BaF3 зависимым от концентрации образом. Это демонстрирует, что NS22 обладает нейтрализующей активностью против передачи сигнала IL-31 человека (фиг. 5).
Активность нейтрализации IL-31 оценивали, как описано ниже, с использованием клеточной линии DU145 (клеточная линия рака предстательной железы человека), в которой при стимуляции IL-31 индуцируется продукция IL-6.
Клетки DU145 получали в концентрации 2,5×105 клеток/мл в MEM (Invitrogen), содержащей 10% FBS (MOREGATE), 2 ммоль/л L-глутамина (Invitrogen), и 1 ммоль/л пирувата натрия (SIGMA), и 200-мкл аликвоты распределяли в каждую лунку 48-луночных планшетов (CORNING). Клетки инкубировали при 37ºC в 5% CO2 в течение ночи. Очищенный химерный NS22 (γ1, κ человека) разбавляли до 100 мкг/мл посредством MEM, содержащей 10% FBS, 2 ммоль/л L-глутамина и пируват натрия. С использованием этого раствора получали шесть серийных разведений с общим отношением разведения, равным 5. Каждое разведение комбинировали со 100 нг/мл интерлейкина-31 человека (R&D systems) в отношении 1:1, и в каждую лунку добавляли 50-мкл аликвоту. После культивирования в течение двух суток при 37ºC в 5% CO2 определяли концентрацию IL-6 в культуральном супернатанте с использованием набора DuoSet ELISA Development kit (R&D systems). Нейтрализующую активность NS22 оценивали путем определения скорости ингибирования (%). Конкретно, принимая концентрацию IL-6 в отсутствие IL-31 (A) за максимальную ингибиторную активность (100% ингибирование), а концентрацию IL-6 в присутствии IL-31 без NS22 (B) за отсутствие ингибиторной активности (0% ингибирование), определяли концентрацию IL-6 в присутствии IL-31 и NS22 (C) согласно следующей формуле:
Скорость ингибирования (%)=(B-C)/(B-A)×100
Результат показал, что NS22 подавлял индуцируемую IL-31 продукцию IL-6 в клеточной линии DU145 зависимым от концентрации образом, и, таким образом, продемонстрировал, что NS22 обладает активностью нейтрализации передачи сигнала IL-31 человека (фиг. 6).
Оценка конкуренции химерного антитела против NR10 с IL-31
IL-31 человека (R&D Systems) метили FMAT Blue Monofunctional Reactive Dye (Applied Biosystems). 100 мкл hIL-31, приготовленного в концентрации 0,5 мг/мл с использованием 50 мМ фосфатно-натриевого буфера (pH 8,0) смешивали с 5,25 мкл 25 нмоль FMAT Blue, растворенного в DMSO (Junsei). После встряхивания смеси позволяли стоять при комнатной температуре в течение 15 минут. Реакцию конъюгации FMAT Blue с hIL-31 завершали добавлением 5 мкл 1M Tris-HCl (pH 7,4) и 1,1 мкл 10% Tween20, а затем hIL-31, меченного FMAT Blue, и не вступивший в реакцию FMAT Blue отделяли гель-фильтрацией с использованием колонки Superdex 75 (GE Healthcare, 17-0771-01) с 0,1% проявляющим раствором Tween20/PBS.
Антитела оценивали в отношении активности ингибирования связывания IL-31/NR10 с использованием экспрессирующих hNR10 клеток CHO, как описано ниже.
NS22 и NA633 (константная область каждого из которых представляет собой γ1, κ) разбавляли в соответствующей концентрации с использованием буфера для анализа (10 мМ HEPES, 140 мМ NaCl, 2,5 мМ CaCl2, 3 мМ MgCl2, 2% FBS, 0,01% NaN3), а затем получали семь серийных разведений с общим отношением разведения, равным 2. Разведения добавляли в планшеты в концентрации 40 мкл/лунка (96-луночные планшеты FMAT; Applied Biosystems). Затем hIL-31, меченный FMAT Blue, разбавляли в 400 раз буфером для анализа и добавляли в количестве 20 мкл/лунка. Наконец, добавляли суспензии клеток, доведенные до 2,5×105 клеток/мл, с использованием буфера для анализа в количестве 40 мкл/лунка (всего 1×104 клеток/лунка). Через два часа после добавления клеток определяли флуоресценцию (FL1) с использованием системы 8200 Cellular Detection System (Applied Biosystems). Результат показал, что NS22 ингибировало связывание hIL-31/hNR10 зависимым от дозы образом, и продемонстрировало, что его активность превышала активность NA633 (фиг. 7).
[Пример 3] Конкуренция антитела против NR10 в отношении NR10
Антитело NS22, очищенное с супернатантом культуры гибридом, метили FMAT Blue (Applied Biosystems, 4328853). 170 мкл NS22, приготовленного в концентрации 1 мг/мл в PBS, смешивали с 17 мкл 1M раствора NaHCO3 и 3,4 мкл FMAT Blue (17 нмоль), растворенного в DMSO. После встряхивания смеси позволяли стоять при комнатной температуре в течение 30 минут. Реакцию конъюгации FMAT Blue с NS22 завершали добавлением 8 мкл 1M Tris-HCl (pH 7,4) и 1,9 мкл 1% Tween 20, а затем NS22, меченного FMAT Blue (FMAT Blue-NS22) и не вступивший в реакцию FMAT Blue отделяли гель-фильтрацией с использованием колонки Superdex 75 (GE Healthcare, 17-0771-01) с 0,01% проявляющим раствором Tween20/PBS.
Каждое антитело исследовали в отношении ингибирования связывания полученного FMAT Blue-NS22 с экспрессирующими hNR10 клетками CHO (справочный пример 3) с использованием системы 8200 Cellular Detection System (Applied Biosystems, 4342920). Химерные антитела против NR10 (константная область каждого представляет собой γ1, κ) добавляли в различных концентрациях в каждую лунку, содержащую 7500 клеток и 8,8×10-2 мкг/мл FMAT Blue-NS22. Клеткам позволяли стоять в темноте в течение четырех часов, а затем измеряли флуоресцентный сигнал от FMAT Blue, связанного с клетками. Реакцию проводили в 10 мМ Hepes-KOH, содержащем 2,5 мМ CaCl2, 3 мМ MgCl2, 140 мМ NaCl, 2% FBS и 0,01% NaNO3. Результат представлен на фиг. 8. Величина флуоресценции FL1, которая соответствует связыванию FMAT Blue-NS22 с экспрессирующими NR10 клетками, снижалась при увеличении концентрации антитела NS22 или NS23. С другой стороны, величина FL1 практически не снижалась при повышении концентрации антитела NA633 (справочный пример 6) (фиг. 8).
[Пример 4] Гуманизация антитела NS22
Выбор каждой каркасной последовательности
Вариабельные области антитела NS22 мыши сравнивали с эмбриональными последовательностями человека. Последовательности FR, использованные для гуманизации, обобщенно представлены в таблице 2. CDR и FR определяли, исходя из нумерации по Kabat. Гуманизированные последовательности вариабельной области H-цепи, состоящие из FR1, FR2, FR3_1 и FR4, и состоящие из FR1, FR2, FR3_2 и FR4, которые приведены в таблице 2, конструировали в качестве H0-VH (SEQ ID NO: 50) и H1-VH (SEQ ID NO: 112), соответственно. При этом последовательность L-цепи, состоящую из FR1, FR2, FR3 и FR4, конструировали в качестве L0 (SEQ ID NO: 52).
Получение вариабельной области гуманизированного NS22 H0L0
Синтетическую олиго-ДНК конструировали для каждой из H- и L-цепей в целях конструирования вариабельных областей гуманизированного NS22, в которых CDR NS22 пересажены на FR, использованные для гуманизации. Соответствующие синтетические олиго-ДНК смешивали, а затем подвергали ПЦР со сборкой для конструирования гена, кодирующего вариабельную область гуманизированного NS22. ПЦР со сборкой проводили с использованием KOD-Plus (TOYOBO) согласно следующим условиям. Реакционную смесь, содержащую 10 пмоль синтетической олиго-ДНК и прилагаемый буфер для ПЦР, dNTP, MgSO4 и KOD-Plus, нагревали при 94ºC в течение пяти минут. Затем смесь подвергали двум циклам ПЦР, состоящим из: 94ºC в течение двух минут, 55ºC в течение двух минут и 68ºC в течение двух минут. Затем добавляли 10 пмоль каждого праймера, в которые были добавлены участок рестрикции и последовательность Козака на 5'-конце вариабельной области, и праймера, в котором участок рестрикции был добавлен на 3'-конце вариабельной области, и подвергали 35 циклам ПЦР, состоящим из: 94ºC в течение 30 секунд, 55ºC в течение 30 секунд и 68ºC в течение одной минуты, с получением амплифицированного фрагмента. Полученный амплифицированный фрагмент клонировали в вектор TOPO TA Cloning (TOYOBO), и его нуклеотидную последовательность определяли секвенированием. Сконструированные вариабельные области комбинировали с константными областями для получения H0-SKSC (SEQ ID NO: 54) и L0 (SEQ ID NO: 56). Полученную конструкцию встраивали в экспрессирующий вектор, способный экспрессировать встроенный ген в клетках животных. Нуклеотидную последовательность каждого фрагмента ДНК определяли с использованием набора BigDye Terminator Cycle Sequencing Kit (Applied Biosystems) с помощью ДНК-секвенатора ABI PRISM 3730xL или ДНК-секвенатора ABI PRISM 3700 (Applied Biosystems) согласно способу, описанному в прилагаемой инструкции.
Получение вариабельной области для H1 гуманизированного NS22
H1-SKSC (SEQ ID NO: 130) получали путем замены глутамина (E) в положении 73 согласно нумерации Kabat в FR3 H0-SKSC (SEQ ID NO: 54) на лизин (K). Мутант получали с использованием коммерчески доступного набора для сайт-направленного мутагенеза QuikChange Site-Directed Mutagenesis Kit (Stratagene) согласно прилагаемой инструкции.
Экспрессия преобразованного в IgG антитела
Экспрессию антитела проводили способом, описанным ниже. Происходящую из злокачественных клеток почки эмбриона человека клеточную линию HEK293H (Invitrogen) суспендировали в DMEM (Invitrogen), содержащей 10% эмбриональную телячью сыворотку (Invitrogen), и 10 мл клеток с плотностью 5-6×105 клеток/мл высевали в чашки для прикрепления клеток (диаметром 10 см; CORNING). Клетки инкубировали в инкубаторе с CO2 (37ºC, 5% CO2) в течение одного целого дня и ночи. Затем среду удаляли аспирированием, и к клеткам добавляли 6,9 мл среды CHO-S-SFMII (Invitrogen). Полученную смесь плазмидной ДНК (всего 13,8 мкг) смешивали с 20,7 мкл полиэтиленимина в концентрации 1 мкг/мл (Polysciences Inc.) и 690 мкл среды CHO-S-SFMII, и позволяли стоять при комнатной температуре в течение 10 минут. Смесь добавляли к клеткам в каждой чашке, и клетки инкубировали в инкубаторе с CO2 (5% CO2, 37ºC) в течение от четырех до пяти часов. Затем добавляли 6,9 мл среды CHO-S-SFMII (Invitrogen), и клетки инкубировали в инкубаторе с CO2 в течение трех суток. Культуральный супернатант собирали и центрифугировали (приблизительно 2000 g, пять минут, комнатная температура) для удаления клеток. Затем супернатант стерилизовали фильтрацией через 0,22-мкм фильтр MILLEX®-GV (Millipore). Каждый образец хранили при 4ºC до применения.
Очистка преобразованного в IgG антитела
К полученному супернатанту культуры добавляли 50 мкл rProtein A SepharoseTM Fast Flow (Amersham Biosciences), суспендированного в TBS, и перемешивали переворачиванием при 4ºC в течение четырех часов или более. Раствор переносили в 0,22-мкм фильтрационную емкость Ultrafree®-MC (Millipore). После трех промываний 500 мкл TBS, смолу rProtein A SepharoseTM суспендровали в 100 мкл водного раствора 50 мМ ацетата натрия (pH 3,3), и позволяли стоять в течение трех минут для элюирования антитела. Раствор сразу нейтрализовали добавлением 6,7 мкл 1,5M Tris-HCl (pH 7,8). Элюирование проводили два раза и получили 200 мкл очищенного антитела. 2 мкл содержащего антитело раствора подвергали исследованию на спектрофотометре ND-1000 (NanoDrop)(спектрофотометр Thermo Scientific NanoDropTM 1000 (Thermo Scientific)) или 50 мкл подвергали исследованию на спектрофотометре DU-600 (BECKMAN) для измерения поглощения при 280 нм, и вычисляли концентрацию антитела способом Pace et al. (Protein Science (1995) 4: 2411-2423).
Измерение конкуренции с IL-31 с использованием FMAT
Антитела оценивали в отношении активности ингибирования связывания IL-31/NR10 с использованием экспрессирующих hNR10 клеток CHO, как описано ниже. Химерное антитело NS22 и NS22_H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56) разбавляли до соответствующей концентрации с использованием буфера для анализа (10 мМ HEPES, 140 мМ NaCl, 2,5 мМ CaCl2, 3 мМ MgCl2, 2% FBS, 0,01% NaN3, pH7,4), а затем получали восемь серийных разведений при общем отношении разведения, равном 2. Разведения добавляли в планшеты в количестве 40 мкл/лунка (96-луночные планшеты FMAT, Applied Biosystems). Затем hIL-31, меченный FMAT Blue, разбавляли в 400 раз буфером для анализа, и добавляли в количестве 20 мкл/лунка. Наконец, добавляли суспензию клеток, доведенную до концентрации 2,5×105 клеток/мл с использованием буфера для анализа, в количестве 40 мкл/лунка (всего 1×104 клеток/лунка). Через два часа после добавления клеток измеряли флуоресценцию (FL1) с использованием системы 8200 Cellular Detection System (Applied Biosystems).
Результат показал, что, как представлено на фиг. 9, гуманизированные антитела NS22 H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56), и H1L0 (H-цепь, H1-SKSC/SEQ ID NO: 130; L-цепь, L0/SEQ ID NO: 56) проявляли активность конкуренции, сравнимую с активностью конкуренции химерного антитела, указывая на то, что как H0L0, так и H1L0, представляют собой гуманизированные антитела против рецептора IL-31. Кроме того, считается, что FR, использованные для H0L0 и H1L0, можно использовать для гуманизации.
Таким образом, все мутации в CDR, описанные в разделе "Примеры" в дальнейшем в настоящем документе, можно вносить как в H0, так и в H1.
(SEQ ID NO: 96)
(SEQ ID NO: 98)
(SEQ ID NO: 131) (SEQ ID NO: 99)
(SEQ ID NO: 99)
(Регистрационный номер X59315)
(SEQ ID NO: 100)
(Регистрационный номер X59315)
(Регистрационный номер X59315)
(SEQ ID NO: 102)(SEQ ID NO: 103)
[Пример 5] Эффект уменьшения гетерогенности новых константных областей M14 и M58 в гуманизированном антителе против рецептора IL31
Как представлено в справочных примерах 7-9, было показано, что преобразование константной области из IgG2 в M14 или M58 в антителе huPM1, гуманизированном антителе против рецептора IL-6, может снизить гетерогенность, создаваемую шарнирной областью IgG2, без потери стабильности. Таким образом, гуманизированные антитела против рецептора IL-31 также тестировали для оценки того, можно ли снизить гетерогенность путем превращения их константных областей из IgG2 дикого типа в M14 или M58.
H0-M14, H0-M58, H0-IgG1 и H0-IgG2, которые получали комбинированием IgG1 (SEQ ID NO: 60), IgG2 (SEQ ID NO: 132), M14 (SEQ ID NO: 129) и M58 (SEQ ID NO: 128), полученных согласно справочным примерам 8 и 9, с вариабельной областью H-цепи H0 (H0-VH/SEQ ID NO: 50) гуманизированного антитела против рецептора IL-31, полученного согласно примеру 4, использовали в качестве H-цепей, и L0 (L0/SEQ ID NO: 56), полученные согласно примеру 4, использовали в качестве L-цепи, для получения H0L0-IgG1 (H-цепь, H0-IgG1/SEQ ID NO: 133; L-цепь, L0/SEQ ID NO: 56), H0L0-IgG2 (H-цепь, H0-IgG2/SEQ ID NO: 134; L-цепь, L0/SEQ ID NO: 56), H0L0-M14 (H-цепь, H0-M14/SEQ ID NO: 135; L-цепь, L0/SEQ ID NO: 56), и H0L0-M58 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56). Каждое антитело экспрессировали и очищали способом, описанным в примере 4.
Гетерогенность оценивали катионообменной хроматографией. Полученные антитела оценивали в отношении гетерогенности с использованием колонки ProPac WCX-10 (Dionex), 20 мМ ацетата натрия (pH 5,0) в качестве подвижной фазы A, и 20 мМ ацетата натрия/1M NaCl (pH 5,0) в качестве подвижной фазы B, с соответствующей скоростью потока и градиентом. Результат оценки катионообменной хроматографией (IEC) представлен на фиг. 10.
Как представлено на фиг. 10, гетерогенность возрастала при преобразовании константной области из IgG1 в IgG2 в антителе против рецептора IL-31, и гетерогенность можно снижать путем преобразования константной области в M14 или M58 в любом антителе.
[Пример 6] Эффект улучшения фармакокинетики новой константной области M58 антител против рецептора IL-31
Как представлено в справочном примере 9, было выявлено, что преобразование константной области из IgG1 в M58 в антителе против рецептора IL-6 huPM1 улучшает его активность связывания с FcRn человека и фармакокинетику у трансгенных по FcRn человека мышей. Таким образом, антитела против рецептора IL-31 также тестировали для оценки того, улучшит ли преобразование константной области в M58 их фармакокинетику.
H0L0-IgG1 (H-цепь: H0-IgG1/SEQ ID NO: 133; L-цепь: L0/SEQ ID NO: 56) и H0L0-M58 (H-цепь: H0-M58/SEQ ID NO: 136; L-цепь L0/SEQ ID NO: 56), полученные, как описано в примерах 4 и 5, оценивали в отношении активности связывания FcRn человека способом, описанным в справочном примере 9. Результат представлен в таблице 3.
Как представлено в таблице 3, преобразование константной области из IgG1 в M58 также улучшило активность антитела против рецептора IL-31 H0L0 в отношении связывания FcRn, как и в антителе против рецептора IL-6 hPM1. Это указывает на то, что преобразование константной области из IgG1 в M58 может улучшить фармакокинетику антитела против рецептора IL-31 у человека.
[Пример 7] Идентификация участков мутации, снижающих изоэлектрическую точку
Продукция мутантов
Каждый мутант получали способом, описанным в примере 4 или посредством ПЦР со сборкой. В способе с использованием ПЦР со сборкой синтезируют олиго-ДНК на основе прямых и обратных последовательностей, включая измененный участок. Прямую олиго-ДНК, включающую измененный участок, и обратную олиго-ДНК, связывающуюся с вектором, в который встроен ген, подлежащий изменению, объединяли, и объединяли обратную олиго-ДНК, включающую измененный участок, и прямую олиго-ДНК, связывающуюся с вектором, в который встроен ген, подлежащий изменению. ПЦР проводили с использованием PrimeSTAR (Takara) для получения 5'-концевых и 3'-концевых фрагментов, включающих измененный участок. Два фрагмента объединяли с помощью ПЦР со сборкой для получения каждого мутанта. Полученный мутант встраивали в экспрессирующий вектор, способный экспрессировать встроенный ген в клетках животных. Нуклеотидную последовательность полученного экспрессирующего вектора определяли способом, известным специалистам в данной области. Антитела получали и очищали способом, описанным в примере 4.
Идентификация участков мутации
Для улучшения фармакокинетики H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56) оценивали измененные участки, способные снижать изоэлектрическую точку вариабельной области. Скрининг участков мутации в вариабельных областях, предсказанных из модели трехмерной структуры, выявил участки мутации, которые могут снизить изоэлектрическую точку вариабельных областей без существенного снижения их связывания с NR10. Они обобщенно представлены в таблице 4 (Hp5-VH/SEQ ID NO: 137, Hp7-VH/SEQ ID NO: 138, Hp8-VH/SEQ ID NO: 139, Hp6-VH/SEQ ID NO: 140, Hp9-VH/SEQ ID NO: 141, Hp1-VH/SEQ ID NO: 142, Hp13-VH/SEQ ID NO: 143, Lp1-VL/SEQ ID NO: 144, Lp2-VL/SEQ ID NO: 145, Lp3-VL/SEQ ID NO: 146, Lp4-VL/SEQ ID NO: 147, Lp7-VL/SEQ ID NO: 148, Lp5-VL/SEQ ID NO: 149, Lp6-VL/SEQ ID NO: 150). Каждый вариант получали и очищали способом, описанным в примере 4.
Каждый вариант тестировали в отношении активности ингибирования связывания hIL-31/hNR10 с использованием FMAT. Тест проводили согласно способу, описанному в примере 4. Как представлено на фиг. 11, активность конкуренции каждого варианта не снижалась значительно по сравнению с активностью конкуренции H0L0.
(№ по Кабату)
(SEQ ID NO: 97)
(SEQ ID NO: 120)
(SEQ ID NO: 10)
(SEQ ID NO: 113)
(SEQ ID NO: 10)
(SEQ ID NO: 10)
(SEQ ID NO: 10)
(SEQ ID NO: 10)
(SEQ ID NO: 10)
(SEQ ID NO: 13)
(SEQ ID NO: 121)
(SEQ ID NO: 13)
(SEQ ID NO: 122)
(SEQ ID NO: 14)
(SEQ ID NO: 123)
(SEQ ID NO: 14)
(SEQ ID NO: 124)
(SEQ ID NO: 14)
(SEQ ID NO: 125)
(SEQ ID NO: 14)
(SEQ ID NO: 126)
(SEQ ID NO: 14)
(SEQ ID NO: 127)
Примеры гуманизированных антител NS22, изоэлектрическая точка которых снижалась при комбинировании этих изменений, включают Hp3Lp15 (H-цепь: Hp3-SKSC/SEQ ID NO: 151; L-цепь: Lp15/SEQ ID NO: 152). Аффинность к NR10, изоэлектрическую точку и время удержания в плазме у мышей сравнивали между Hp3Lp15 и H0L0.
Измерение аффинности
Аффинность каждого антитела к NR10 определяли способом, описанным в справочном примере 10.
Результат измерения аффинности представлен в таблице 5. Было показано, что аффинность Hp3Lp15 является практически такой же, как и аффинность H0L0.
Определение изоэлектрической точки
Каждое антитело анализировали изоэлектрическим фокусированием для оценки изменений изоэлектрической точки целого антитела вследствие аминокислотных замен в его вариабельной области. Изоэлектрическое фокусирование проводили следующим способом.
Гель Phast-Gel Dry IEF (Amersham Biosciences) подвергали набуханию в кассете Phastsystem Cassette (Amersham Biosciences) в течение приблизительно 30 минут с использованием раствора для набухания, представленного ниже.
Электрофорез проводили в PhastSystem (Amersham Biosciences) с использованием набухшего геля согласно программе, указанной ниже. Образцы наносили на гель на стадии 2. В качестве маркера pI использовали калибровочный набор для pI (Amersham Biosciences).
Стадия 1: 2000 В 2,5 мА 3,5 Вт 15ºC 75 Vh
Стадия 2: 200 В 2,5 мА 3,5 Вт 15ºC 15 Vh
Стадия 3: 2000 В 2,5 мА 3,5 Вт 15ºC 410 Vh
После электрофореза гель фиксировали 20% TCA, а затем подвергали окрашиванию серебром с использованием набора Silver Staining Kit, Protein (Amersham Biosciences), в соответствии с протоколом, прилагаемым к набору. После окрашивания из известных изоэлектрических точек маркеров pI вычисляли изоэлектрическую точку образца (целого антитела).
Результат измерения изоэлектрической точки с помощью изоэлектрического фокусирования показал, что изоэлектрическая точка H0L0 составила приблизительно 7,8, а изоэлектрическая точка Hp3Lp15 составила приблизительно 5,5, указывая на то, что изоэлектрическая точка Hp3Lp15 была снижена приблизительно на 2,3 по сравнению с H0L0. Когда вычисляли теоретическую изоэлектрическую точку вариабельной области VH/VL с помощью GENETYX (GENETYX CORPORATION), теоретические изоэлектрические точки вариабельных областей H0L0 и Hp3Lp15 составили приблизительно 7,76 и 4,63, соответственно. Таким образом, теоретическая изоэлектрическая точка Hp3Lp15 была снижена на 3,13 по сравнению с H0L0.
Оценка фармакокинетики антитела со сниженной изоэлектрической точкой с использованием мышей
Для оценки времени удержания в плазме Hp3Lp15, модифицированного антитела со сниженной изоэлектрической точкой, сравнивали время удержания в плазме H0L0 и Hp3Lp15 у нормальных мышей. Одну дозу H0L0 или Hp3Lp15 внутривенно вводили мышам в количестве 1 мг/кг (C57BL/6J, Charles River Japan, Inc.) для сравнения концентрации в плазме с течением времени. Концентрации в плазме определяли с помощью ELISA. Соответствующие концентрации калибровочного образца и тестируемых образцов плазмы распределяли в иммунологические планшеты (Nunc-Immuno Plate, MaxiSorp (Nalge Nunc International)), покрытые антителом против IgG человека (Fc-специфическим) (Sigma). Образцам позволяли стоять при комнатной температуре в течение одного часа. После реакции с антителом козы против IgG человека-ALP (Sigma) при комнатной температуре в течение одного часа, проводили реакцию проявления цвета с использованием системы BluePhos Microwell Phosphatase Substrates System (Kirkegaard & Perry Laboratories) в качестве субстрата. Поглощение при 650 нм измеряли с помощью устройства для считывания микропланшетов. Концентрации в плазме определяли, исходя из поглощения калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices).
Фармакокинетические параметры (AUC и системное выведение (CL)) вычисляли из данных концентрации в плазме с течением времени с использованием программного обеспечения для фармакокинетического анализа WinNonlin (Pharsight). Параметры представлены в таблице 6. AUC и выведение Hp3Lp15 после внутривенного введения были повышены приблизительно на 14% и снижены приблизительно на 12%, соответственно, по сравнению с H0L0. Таким образом, было определено, что Hp3Lp15, в котором изоэлектрическая точка H0L0 была снижена, обладал улучшенной фармакокинетикой.
[Пример 8] Эффект комбинаций вариабельной области и константной области на биологическую активность
Для оценки эффектов различных константных областей на биологическую активность получали следующие варианты.
SKSC (SEQ ID NO: 62) и M58 (SEQ ID NO: 128), константные области, полученные в справочных примерах 7 и 9, комбинировали с Hp3 (Hp3-VH/SEQ ID NO: 167), вариабельной областью, полученной согласно примеру 7, с получением Hp3-M58 (SEQ ID NO: 240) и Hp3-SKSC (SEQ ID NO: 151) в качестве H-цепей. Полученные H-цепи комбинировали с Lp15 (Lp15/SEQ ID NO: 152), L-цепью, полученной согласно примеру 7, с получением Hp3Lp15-SKSC (H-цепь, Hp3-SKSC/SEQ ID NO: 151; L-цепь, Lp15/SEQ ID NO: 152) и Hp3Lp15-M58 (H-цепь, Hp3-M58/SEQ ID NO: 240; L-цепь, Lp15/SEQ ID NO: 152). Каждое антитело экспрессировали и очищали способом, описанным в примере 4.
Антитела, полученные как описано выше, H0L0-SKSC (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56), полученное с использованием константной области SKSC (SEQ ID NO: 62), описанной в справочном примере 7, и H0L0-M58 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56) и H0L0-IgG2 (H-цепь, H0-IgG2/SEQ ID NO: 134; L-цепь, L0/SEQ ID NO: 56), полученное согласно примеру 5, оценивали в отношении биологической активности способом, описанным в примере 2, с использованием BaF/NR10. Результат обобщенно представлен на фиг. 18.
Как представлено на фиг. 18, не было выявлено значимых отличий в биологической активности между константными областями. Поскольку на биологическую активность не влияло комбинирование двух вариабельных областей H0 и Hp3 с каждой константной областью, комбинирование вариабельных областей, которые будут созданы в будущем, с любой константной областью не приведет к изменению биологической активности.
[Пример 9] Идентификация участков мутации, подавляющих деградацию в исследовании с термическим ускорением
Антитела, используемые для фармацевтических средств, обладают гетерогенностью, даже несмотря на то, что они представляют собой моноклональные антитела, полученные из клонов, происходящих из единичных антителопродуцирующих клеток. Известно, что такая гетерогенность антител является следствием модификации, такой как окисление или дезамидация, и возрастает при длительном хранении или под воздействием в условиях нагрузки, таких как тепловая нагрузка или нагрузка светом (см. "Heterogeneity of Monoclonal Antibodies", Journal of pharmaceutical sciences, vol. 97, No. 7, 2426-2447). Однако когда антитело разрабатывают в качестве фармацевтического средства, физические свойства белка, в частности гомогенность и стабильность, являются в высокой степени важными. Таким образом, желательно, чтобы гетерогенность желаемых/родственных веществ была снижена, и чтобы вещество состояло из одного вещества, насколько это возможно. В этом контексте эксперимент, описанный ниже, проводили для оценки гетерогенности антитела в условиях нагрузки и для снижения гетерогенности.
Для оценки продуктов деградации получали образец H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56) после ускорения способом, описанным ниже. Полученный образец после ускорения и образец без ускорения (исходный) анализировали катионообменной хроматографией с использованием способа, описанного ниже.
- Способ получения образцов после ускорения
Буфер: PBS
Концентрация антитела: от 0,2 до 1,0 мг/мл
Температура ускорения: 60ºC
Период ускорения: одни сутки
- Способ анализа катионообменной хроматографией
Колонка: ProPac WCX-10, 4×250 мм (Dionex)
Подвижная фаза: (A) 25 ммоль/л MES/NaOH, pH 6,1
(B) 25 ммоль/л MES/NaOH, 250 ммоль/л NaCl, pH 6,1
Скорость потока: 0,5 мл/мин
Температура колонки: 40ºC
Градиент: %B 0 до 0 (0-5 мин)→ от 0 до 30 (5-80 мин)
Детекция: 280 нм
Полученные хроматограммы для образцов H0L0 до и после ускорения представлены на фиг. 19. Образец H0L0 после ускорения имел тенденцию к повышению основного пика.
Затем проводили скрининг для снижения этого пика. В результате были выявлены Ha355, Ha356, Ha360 и Ha362. Эти варианты H-цепей комбинировали с L0 с получением Ha355L0 (H-цепь, Ha355-SKSC/SEQ ID NO: 242; L-цепь, L0/SEQ ID NO: 56), Ha356L0 (H-цепь, Ha356-SKSC/SEQ ID NO: 243; L-цепь, L0/SEQ ID NO: 56), Ha360L0 (H-цепь, Ha360-SKSC/SEQ ID NO: 244; L-цепь, L0/SEQ ID NO: 56) и Ha362L0 (H-цепь, Ha362-SKSC/SEQ ID NO: 245; L-цепь, L0/SEQ ID NO: 56). Последовательность каждого варианта представлена в таблице 7.
(№ по Кабату)
(SEQ ID NO: 265)
(SEQ ID NO: 266)
(SEQ ID NO: 265)
(SEQ ID NO: 267)
(SEQ ID NO: 265)
(SEQ ID NO: 268)
(SEQ ID NO: 265)
(SEQ ID NO: 269)
Каждое из идентифицированных антител экспрессировали и очищали способом, описанным в примере 4. Как и в случае H0L0, получали образец каждого антитела после ускорения, и анализировали катионообменной хроматографией. Результат представлен на фиг. 19.
Результат показал, что образование основного пика, увеличенное после ускорения, было снижено в случае модифицированного антитела, содержащего замену аспарагиновой кислоты глутаминовой кислотой в положении 101 в H-цепи, по сравнению с H0L0. Модифицированные антитела оценивали в отношении биологической активности способом, описанным в примере 2, с использованием BaF/NR10. Результат представлен на фиг. 20. Как представлено на фиг. 20, биологическая активность модифицированных антител была сравнимой или более сильной, чем биологическая активность H0L0. Эти данные показали, что модификации Ha355, Ha356, Ha360 и Ha362 подавляют образование продуктов деградации при ускорении, и, таким образом, являются эффективными в отношении повышения стабильности антитела.
[Пример 10] Идентификация участков мутаций, повышающих аффинность
Библиотеку, в которой в последовательности CDR были внесены мутации, конструировали и исследовали в отношении повышения аффинности H0L0 к NR10. В результате скрининга библиотеки, в которой в CDR были внесены мутации, были выявлены мутации, которые повышают аффинность к NR10. Эти мутации представлены в таблице 8. Каждый из вариантов H-цепи Ha101-SKSC (SEQ ID NO: 246), Ha103-SKSC (SEQ ID NO: 247), Ha111-SKSC (SEQ ID NO: 248), Ha204-SKSC (SEQ ID NO: 249) и Ha219-SKSC (SEQ ID NO: 250) комбинировали с L0 (L0/SEQ ID NO: 56); и каждую из модифицированных L-цепей La134 (SEQ ID NO: 251), La130 (SEQ ID NO: 252), La303 (SEQ ID NO: 253) и La328 (SEQ ID NO: 254) комбинировали с H0 (H0-SKSC/SEQ ID NO: 54), для конструирования антитела. Каждый вариант получали и очищали способом, описанным в примере 4.
Аффинность каждого антитела к NR10 оценивали с использованием Biacore. Результат представлен в таблице 9. Анализ проводили с использованием способа, описанного в справочном примере 10. Как представлено в таблице 9, было выявлено, что величина KD для каждого варианта является улучшенной по сравнению с величиной KD H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56).
(SEQ ID NO: 270)
(SEQ ID NO: 272)
(SEQ ID NO: 270)
(SEQ ID NO: 273)
(SEQ ID NO: 270)
(SEQ ID NO: 274)
(SEQ ID NO: 271)
(SEQ ID NO: 275)
(SEQ ID NO: 271)
(SEQ ID NO: 276)
(№ по Кабату)
(SEQ ID NO: 277)
(SEQ ID NO: 279)
(SEQ ID NO: 277)
(SEQ ID NO: 280)
(SEQ ID NO: 278)
(SEQ ID NO: 281)
(SEQ ID NO: 278)
(SEQ ID NO: 282)
(SEQ ID NO: 278)
(SEQ ID NO: 283)
Примеры комбинаций этих повышающих аффинность мутаций с мутациями, снижающими изоэлектрическую точку, полученными согласно примеру 7, включают, например, Ha401La402 (H-цепь, Ha401-SKSC/SEQ ID NO: 255; L-цепь, La402/SEQ ID NO: 256) и H17L11 (H-цепь, H17-M58/SEQ ID NO: 222; L-цепь, L11/SEQ ID NO: 236). Каждый вариант получали и очищали способом, описанным в примере 4.
Ha401La402 (H-цепь, Ha401-SKSC/SEQ ID NO: 255; L-цепь, La402/SEQ ID NO: 256) оценивали в отношении его аффинности в отношении NR10 и его биологической активности способом, описанным в справочном примере 10, и способом с использованием BaF/NR10, как описано в примере 2, соответственно, и их сравнивали с аффинностью и биологической активностью H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56). Результат определения аффинности представлен в таблице 10, и биологическая активность, определенная с использованием BaF/NR10, представлена на фиг. 21. Было выявлено, что как аффинность, так и биологическая активность, являются улучшенными по сравнению с аффинностью и биологической активностью H0L0 (H-цепь, H0-SKSC/SEQ ID NO: 54; L-цепь, L0/SEQ ID NO: 56).
Более того, H17L11 (H-цепь, H17-M58/SEQ ID NO: 222; L-цепь, L11/SEQ ID NO: 236) оценивали в отношении его аффинности к NR10 и его биологической активности способом, описанным в примере 7, и способом с использованием BaF/NR10, как описано в примере 2, соответственно, и их сравнивали с аффинностью и биологической аффинностью H0L0 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56). Результат измерения аффинности представлен в таблице 11, а биологическая активность, определенная с использованием BaF/NR10, представлена на фиг. 22. Было выявлено, что как аффинность, так и биологическая активность являются улучшенными по сравнению с аффинностью и биологической активностью H0L0 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56).
[Пример 11] Идентификация участков мутаций, снижающих риск иммуногенности
Снижение риска иммуногенности в H-цепи CDR1
Эпитопы T-клеток, присутствующие в последовательности вариабельной области H0L0, анализировали с использованием TEPITOPE (Methods 2004 Dec; 34(4): 468-75). В результате было предсказано, что CDR1 H-цепи имеет множество T-клеточных эпитопов, которые связываются с HLA (т.е. имеют последовательности с высоким риском иммуногенности). Затем анализ TEPITOPE проводили для исследования замен, которые могут снизить риск иммуногенности H-цепи CDR1. В результате было выявлено, что риск иммуногенности значительно снижался посредством замены изолейцина в положении 33 согласно нумерации по Kabat аланином (A) (таблица 12). Затем это изменение добавляли к H17, полученной согласно примеру 10, с получением H19 (H19-M58/SEQ ID NO: 223). Полученную H19 комбинировали с L12 с получением H19L12 (H-цепь, H19-M58/SEQ ID NO: 223; L-цепь, L12/SEQ ID NO: 237). Каждый вариант получали и очищали способом, описанным в примере 4.
Антитело оценивали в отношении аффинности к NR10 и биологической активности способом, описанным в справочном примере 10, и способом с использованием BaF/NR10, как описано в примере 2, соответственно, и их сравнивали с аффинностью и биологической активностью H0L0 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56). Результат измерения аффинности представлен в таблице 13, а биологическая активность, определенная с использованием BaF/NR10, представлена на фиг. 23. Было показано, что как аффинность, так и биологическая активность являются практически равными аффинности и биологической активности H0L0.
(SEQ ID NO: 270)
(SEQ ID NO: 284)
Снижение риска иммуногенности в L-цепи CDR1
Треонин (T), находящийся в положении 25 согласно нумерации по Kabat в CDR1 L-цепи, соответствует аланину (A) или серину (S) в эмбриональной последовательности. Таким образом, предсказано, что риск иммуногенности может снизиться путем замены треонина (T) в положении 25 аланином (A) или серином (S) (таблица 14). Таким образом, указанную выше замену добавляли к L12 с получением L17 (SEQ ID NO: 238). Полученную L17 комбинировали с H0 с получением H0L17 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L17/SEQ ID NO: 238). Каждый вариант получали и очищали способом, описанным в примере 4.
Каждый вариант оценивали в отношении аффинности к NR10 и биологической активности способом, описанным в справочном примере 10, и способом с использованием BaF/NR10, как описано в примере 2, соответственно, и их сравнивали с аффинностью и биологической активностью H0L0 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56) и H0L12 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L12/SEQ ID NO: 237). Поскольку L12 содержит последовательность, которая повышает аффинность, она проявляет аффинность, приблизительно в два раза превышающую аффинность H0L0. Результат измерения аффинности представлен в таблице 15, а биологическая активность, определенная с использованием BaF/NR10, представлена на фиг. 24. Было показано, что как аффинность, так и биологическая активность являются практически равными аффинности и биологической активности H0L12.
(SEQ ID NO: 277)
(SEQ ID NO: 285)
(SEQ ID NO: 277)
(SEQ ID NO: 286)
[Пример 12] Получение полностью гуманизированного антитела NS22
Вариабельные области вариантов NS22 получали путем комбинирования нескольких мутаций, которые снижают pI, повышают аффинность, подавляют деградацию H-цепи, и снижают риск иммуногенности, все из которых были выявлены в указанных выше примерах, в H0 (H0-M58/SEQ ID NO: 136), H1 (H1-M58/SEQ ID NO: 257) или L0 (L0/SEQ ID NO: 56), и подвергали различным способам скрининга. В результате были выявлены H28L17 (H-цепь, H28-M58/SEQ ID NO: 224; L-цепь, L17/SEQ ID NO: 238), H30L17 (H-цепь, H30-M58/SEQ ID NO: 225; L-цепь, L17/SEQ ID NO: 238), H34L17 (H-цепь, H34-M58/SEQ ID NO: 226, L-цепь, L17/SEQ ID NO: 238), H42L17 (H-цепь, H42-M58/SEQ ID NO: 227; L-цепь, L17/SEQ ID NO: 238), H44L17 (H-цепь, H44-M58/SEQ ID NO: 228; L-цепь, L17/SEQ ID NO: 238), H46L17 (H-цепь, H46-M58/SEQ ID NO: 229; L-цепь, L17/SEQ ID NO: 238), H57L17 (H-цепь, H57-M58/SEQ ID NO: 230; L-цепь, L17/SEQ ID NO: 238), H71L17 (H-цепь, H71-M58/SEQ ID NO: 231; L-цепь, L17/SEQ ID NO: 238), H78L17 (H-цепь, H78-M58/SEQ ID NO: 232; L-цепь, L17/SEQ ID NO: 238), H92L17 (H-цепь, H92-M58/SEQ ID NO: 233; L-цепь, L17/SEQ ID NO: 238), H97L50 (H-цепь, H97-M58/SEQ ID NO: 234; L-цепь, L50/SEQ ID NO: 239) и H98L50 (H-цепь, H98-M58/SEQ ID NO: 235; L-цепь, L50/SEQ ID NO: 239). Каждый вариант получали и очищали способом, описанным в примере 4.
Каждый вариант оценивали в отношении аффинности к NR10 и биологической активности способом, описанным в справочном примере 10, и способом с использованием BaF/NR10, как описано в примере 2, соответственно, и их сравнивали с аффинностью и биологической активностью H0L0 (H-цепь, H0-M58/SEQ ID NO: 136; L-цепь, L0/SEQ ID NO: 56). Результат измерения аффинности представлен в таблице 16, а биологическая активность, определенная с использованием BaF/NR10, представлена на фиг. 25-1 и 25-2. Было показано, что как аффинность, так и биологическая активность являются практически равными аффинности и биологической активности H0L0.
[Пример 13] Анализ связывающего домен нейтрализующего антитела против NR10
(1) Получение антигенов дикого типа и химерных антигенов человека/мыши
Гены, кодирующие внеклеточные домены дикого типа человека и мыши и химерные внеклеточные домены NR10 (hhh (SEQ ID NO: 258), mmm (SEQ ID NO: 259), hhm (SEQ ID NO: 260), mmh (SEQ ID NO: 261), hmm (SEQ ID NO: 262), mhm (SEQ ID NO: 263) и mhh (SEQ ID NO: 264)), подвергали слиянию с His-меткой и Myc-меткой (HHHHHHEQKLISEEDL/SEQ ID NO: 287) на их C-концах, встраивали в экспрессирующий вектор животных, и временно экспрессировали с использованием системы FreeStyle 293 Expression System (InvitrogenTM). Схематичные диаграммы для NR10-ECD дикого типа человека/мыши и химерных NR10-ECD представлены на фиг. 26.
Антигены дикого типа человека/мыши и химерные антигены (hhh, mmm, hhm, mmh, hmm, mhm и mhh) очищали из культуральных супернатантов колоночной хроматографией Ni-NTA Superflow. Конкретно, 1 мл Ni-NTA Superflow (QIAGEN) помещали в колонку Poly-Prep Empty Column (BioRad), и в нее добавляли 30 мл каждого культурального супернатанта. После промывания D-PBS (фосфатно-солевой буфер Дульбекко), содержащим 150 мМ хлорид натрия и 20 мМ имидазол, колонку элюировали D-PBS, содержащим 150 мМ хлорид натрия и 250 мМ имидазол. В элюированных фракциях заменяли буфер посредством D-PBS и их концентрировали с использованием Amicon-Ultra (Millipore) с пределом молекулярной массы 10K.
(2) Детекция связывания антигена вестерн-блоттингом
Каждый из полученных антигенов человека/мыши и химерных антигенов подвергали электрофорезу в количестве 0,5 мкг/дорожка на трех 4-20% полиакриламидных гелях (Daiichi Pure Chemicals Co.). Белки подвергали электропереносу на мембраны PVDF (Millipore) в устройстве для полусухого блоттинга, и мембраны блокировали посредством TBS, содержащего 5% обезжиренное молоко. Одну мембрану инкубировали с 5 мкг/мл H44M58L17 (система для детекции гуманизированного антитела против NR10 человека); другую - с 5 мкг/мл ND41 (система для детекции антитела мыши против NR10 человека); и третью - с антителом против Myc (SantaCruz, каталожный #sc-789), разбавленным в 500 раз TBS, содержащим 5% обезжиренное молоко (система для детекции Myc-метки) при комнатной температуре в течение одного часа.
После промывания три раза TBS, содержащим 0,05% TweenTM 20, вторичные антитела инкубировали с мембранами. Для детекции гуманизированного антитела против NR10 человека использовали антитело козы против IgGγ человека со щелочной фосфатазой (BIOSOURCE, каталожный #AHI0305); для детекции антитела мыши против NR10 человека использовали антитело козы против IgG мыши со щелочной фосфатазой (SantaCruz, каталожный #sc-2008); и для детекции Myc-метки использовали антитело козы против IgG кролика со щелочной фосфатазой (SantaCruz, каталожный #sc-2057). Реакцию проводили при комнатной температуре в течение одного часа. После промывания четыре раза TBS, содержащим 0,05% TweenTM 20, в течение трех минут, проводили проявление цвета с использованием субстрата BCIP/NBT фосфатазы, 1-компонентной системы (KPL). Используемый здесь TBS (Tris-буферный солевой раствор) получали растворением упаковки порошка TBS (Tris-буферного солевого раствора) (TaKaRa) в 1 л дистиллированной воды. Результат представлен на фиг. 27.
Когда использовали гуманизированное антитело или антитело мыши, связывание выявлялось только для hhh, hhm и hmm, которые представляют собой внеклеточные домены NR10.
[Справочный пример 1] Выделение генов NR10, OSMR и IL-31 яванского макака
Поскольку перекрестную реактивность и нейтрализующую активность у яванских макаков считали важной для оценки безопасности на доклинической стадии, выделяли гены NR10 и OSMR яванского макака. Праймеры конструировали, исходя из опубликованной информации генома макака резус и других, и из кДНК поджелудочной железы яванского макака успешно амплифицировали гены NR10 и OSMR посредством ПЦР. Последовательности выделенных генов NR10, OSMR и IL-31 яванского макака представлены в SEQ ID NO: 65, 69 и 67, соответственно, и аминокислотные последовательности NR10, OSMR и IL-31 яванского макака представлены в SEQ ID NO: 66, 70 и 68, соответственно.
[Справочный пример 2] Получение клеточных линий Ba/F3, экспрессирующих NR10 и OSMR
кДНК полноразмерного NR10 человека (SEQ ID NO: 75) встраивали в экспрессирующий вектор pCOS1 (Biochem. Biophys. Res. Commun. 228, p838-45, 1996), и полученный вектор назвали pCosNR10.3. кДНК рецептора онкостатина M (OSMR, регистрационный номер GenBank No. NM003999) выделяли посредством ПЦР из библиотеки плацент человека, и аналогичным образом конструировали экспрессирующий вектор pCos1-hOSMR. 10 мкг каждого из векторов одновременно вводили в IL-3-зависимую происходящую из про-B-клеток клеточную линию мыши Ba/F3 посредством электропорации (BioRad Gene Pulser, 960 мкФ, 0,33 кВ). После введения добавляли IL-31 человека (R&D Systems), и клетки культивировали с получением клеточной линии (клетки hNR10/hOSMR/BaF3), которая пролиферируют IL-31-зависимым образом. Более того, ген IL-31 яванского макака (SEQ ID NO: 67) встраивали в экспрессирующий вектор клеток млекопитающих и вводили в клеточную линию CHO DG44. Конечный культуральный супернатант получали в качестве IL-31 яванского макака. Как и в случае hNR10/hOSMR/BaF3, гены полноразмерных NR10 и OSMR яванского макака встраивали в экспрессирующий вектор pCOS1 и экспрессировали в клетках Ba/F3, и получали IL-31-зависимую клеточную линию яванского макака (cynNR10/cynOSMR/BaF3 cell) с использованием культурального супернатанта, описанного выше.
[Справочный пример 3] Получение клеточных линий CHO, экспрессирующих NR10
Гены NR10 человека, лишенного цитоплазматического домена (SEQ ID NO: 73), и NR10 яванского макака, лишенного цитоплазматического домена (SEQ ID NO: 71), встраивали в экспрессирующие векторы млекопитающих. Полученные векторы линеаризовывали ферментом рестрикции, а затем вводили в клеточную линию CHO DG44 посредством электропорации (BioRad Gene Pulser, 25 мкФ, 1,5 кВ). После селекции посредством лекарственного средства, отбирали экспрессирующие NR10 клетки и исследовали их в анализе FCM с использованием антитела против NR10 человека. Аминокислотная последовательность, кодируемая нуклеотидной последовательностью гена NR10, лишенного цитоплазматического домена человека(SEQ ID NO: 73), представлена в SEQ ID NO: 74, и аминокислотная последовательность, кодируемая нуклеотидной последовательностью гена NR10 яванского макака, лишенного цитоплазматического домена (SEQ ID NO: 71), представлена в SEQ ID NO: 72.
[Справочный пример 4] Получение белка NR10 (внеклеточный домен)
кДНК NR10 человека использовали в качестве матрицы для амплификации посредством ПЦР только внеклеточного домена. Затем амплифицированную область подвергали слиянию с последовательностью FLAG-метки на C-конце и встраивали в экспрессирующий вектор клеток млекопитающих. Десять мкг линеаризованного вектора вводили в клеточную линию клеток яичника китайского хомяка DG44 посредством электропорации (BioRad Gene PulserII, 25 мкФ, 1,5 кВ). Получили клеточную линию, обладающую высоким уровнем экспрессии. Супернатант клеточной линии, культивированной в большом масштабе, очищали с использованием колонки с антителом против FLAG (Sigma) и гель-фильтрации с получением растворимого NR10. Нуклеотидная последовательность растворимого NR10 представлена в SEQ ID NO: 77, и аминокислотная последовательность представлена в SEQ ID NO: 78.
[Справочный пример 5] Получение антител против NR10 человека
Мышей иммунизировали белком NR10 человека (внеклеточный домен) (описанным в справочном примере 4), и получали гибридомы общепринятым способом. Культуральные супернатанты этих гибридом оценивали в отношении нейтрализующей активности с использованием зависимой от IL-31 человека клеточной линии (клетки hNR10/hOSMR/BaF3), описанной в справочном примере 2, и, тем самым, получили NA633, которое обладает активностью нейтрализации NR10.
Более того, иммунизацию посредством ДНК проводили с помощью He-газоприводной генной пушки с использованием экспрессирующего вектора млекопитающего, обладающего геном полноразмерного NR10 человека (SEQ ID NO: 75), и получали гибридомы общепринятым способом. Культуральные супернатанты этих гибридом оценивали в отношении нейтрализующей активности с использованием зависимой от IL-31 человека клеточной линии (клетки hNR10/hOSMR/BaF3), описанной в справочном примере 2, и, тем самым, получили ND41, который обладает активностью нейтрализации NR10.
[Справочный пример 6] Получение химерного антитела человека
Аминокислотные последовательности вариабельных областей тяжелой цепи и легкой цепи NA633 представлены в SEQ ID NO: 104 и 108, соответственно. Аминокислотные последовательности CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи NA633 представлены в SEQ ID NO: 105, 106 и 107, соответственно, а аминокислотные последовательности CDR1, CDR2 и CDR3 вариабельной области легкой цепи представлены в SEQ ID NO: 109, 110 и 111, соответственно. Более того, получали химерное антитело с этими вариабельными областями мыши и константной областью человека (H-цепь, γ1; L-цепь, κ) общепринятым способом.
[Справочный пример 7] Получение huPM1-SKSC, в котором гетерогенность IgG2 дикого типа снижена без потери стабильности
Поскольку антитело NS22 является нейтрализующим NR10 антителом, его связывание с Fcγ-рецептором может быть неблагоприятным ввиду иммуногенности и побочных эффектов. Возможным способом снижения связывания с Fcγ-рецептором является выбор IgG2 или IgG4 вместо IgG1 в качестве изотипа константной области (Ann Hematol. 1998 Jun; 76(6): 231-48.). С точки зрения Fcγ-рецептора I и времени удержания в плазме, IgG2 считают более желательным, чем IgG4 (Nat Biotechnol. 2007 Dec; 25(12): 1369-72). При этом, когда антитело разрабатывают в качестве фармацевтического средства, свойства белка, в частности гомогенность и стабильность, являются в высокой степени важными. Было описано, что изотип IgG2 имеет очень высокую гетерогенность вследствие дисульфидных связей в шарнирной области (J Biol Chem. 2008 Jun 6; 283(23): 16206-15.). Изготовление его в качестве фармацевтического средства в крупном масштабе является нелегким и более дорогостоящим, при сохранении отличий в гетерогенности желательных/родственных веществ среди продуктов, вследствие причин, указанных выше. Таким образом, желательно, чтобы вещество состояло из одного вещества, насколько это возможно. Таким образом, когда антитела изотипа IgG2 разрабатывают в качестве фармацевтических средств, предпочтительно снижать гетерогенность вследствие дисульфидных связей, без снижения стабильности.
Для снижения гетерогенности IgG2 дикого типа, заменяли остатки цистеина в шарнирной области и CH1-домене IgG2. В результате исследования различных вариантов, SKSC (SEQ ID NO: 62), который представляет собой константную область, полученную заменой цистеина в положении 131 и аргинина в положении 133 согласно нумерации EU (Sequences of proteins of immunological interest, NIH Publication No.91-3242) в CH1-домене H-цепи последовательности константной области IgG2 человека, на серин и лизин, соответственно, и заменой цистеина в положении 219 согласно нумерации EU в верхней шарнирной области H-цепи, на серин, может снизить гетерогенность без снижения стабильности. При этом другими возможными способами снижения гетерогенности являются замена только цистеина в положении 219 согласно нумерации EU в верхней шарнирной области H-цепи на серин, и замена только цистеина в положении 220 согласно нумерации по EU на серин. Таким образом, получили константную область SC (SEQ ID NO: 153), в которой цистеин в положении 219 согласно нумерации EU в IgG2 заменен на серин, и константную область CS (SEQ ID NO: 154), в которой цистеин в положении 220 согласно нумерации в IgG2 заменен на серин.
Для получения каждого антитела, huPM1-SC (SEQ ID NO: 157), huPM1-CS (SEQ ID NO: 158), huPM1-IgG1 (SEQ ID NO: 159), huPM1-IgG2 (SEQ ID NO: 160) и huPM1-SKSC (SEQ ID NO: 161), которые получали комбинированием константных областей, полученных, как описано выше, IgG1 (SEQ ID NO: 60) и IgG2 (SEQ ID NO: 132) с вариабельной областью гуманизированного антитела против рецептора IL-6 (вариабельная область H-цепи, huPM1-VH/SEQ ID NO: 155; вариабельная область L-цепи huPM1-VL/SEQ ID NO: 156)(Cancer Res. 1993 Feb 15; 53(4): 851-6), использовали в качестве H-цепи, а huPM1-L (SEQ ID NO: 162) использовали в качестве L-цепи. Каждое антитело экспрессировали и очищали способом, описанным в примере 4.
Антитела сравнивали друг с другом с точки зрения гетерогенности. Гетерогенность huPM1-IgG1, huPM1-IgG2, huPM1-SC, huPM1-CS и huPM1-SKSC оценивали катионообменной хроматографией. Хроматографию проводили с использованием колонки ProPac WCX-10 (Dionex), 20 мМ ацетата натрия (pH 5,0) в качестве подвижной фазы A, и 20 мМ ацетата натрия/1M NaCl (pH 5,0) в качестве подвижной фазы B, с соответствующими скоростью потока и градиентом. Результат оценки катионообменной хроматографией представлен на фиг. 12.
Как представлено на фиг. 12, преобразование константной области из IgG1 в IgG2 повышало гетерогенность. Напротив, гетерогенность значительно снижалась при преобразовании константной области в SKSC. Хотя константная область SC приводила к значительному снижению гетерогенности, как и SKSC, константная область CS не приводила к достаточному улучшению гетерогенности.
Когда антитело разрабатывают в качестве фармацевтического средства, для продукции стабильных препаратов, как правило, желательно, чтобы антитело обладало высокой стабильностью в дополнение к низкой гетерогенности. Таким образом, для оценки стабильности определяли среднюю температуру денатурации (величину Tm) дифференциальной сканирующей калориметрией (DSC) (VP-DSC; Microcal). Средняя температура денатурации (величина Tm) служит в качестве показателя стабильности. Для получения стабильных препаратов в качестве фармацевтических средств, предпочтительной является более высокая средняя температура денатурации (величина Tm) (J Pharm Sci. 2008 Apr; 97(4): 1414-26.). Таким образом, huPM1-IgG1, huPM1-IgG2, huPM1-SC, huPM1-CS и huPM1-SKSC подвергали диализу против раствора 20 мМ ацетата натрия/150 мМ NaCl (pH 6,0) (EasySEP; TOMY), и проводили измерение DSC с использованием приблизительно 0,1 мг/мл белка при скорости нагревания 1ºC/мин между 40 и 100ºC. Кривые денатурации, полученные с помощью DSC, представлены на фиг. 13. Величины Tm Fab-доменов приведены в таблице 17, ниже.
Величины Tm для huPM1-IgG1 и huPM1-IgG2 были практически одинаковыми, а именно, они составили приблизительно 94ºC (для IgG2 она была ниже приблизительно на 1ºC). При этом величины Tm для huPM1-SC и huPM1-CS составили приблизительно 86ºC, что было значительно ниже, чем у huPM1-IgG1 и huPM1-IgG2. С другой стороны, величина Tm для huPM1-SKSC составила приблизительно 94ºC, и была практически такой же, как и для huPM1-IgG1 и huPM1-IgG2. Поскольку стабильность huPM1-SC и huPM1-CS была значительно ниже, чем стабильность IgG2, huPM1-SKSC, в котором цистеин в домене CH1 также заменен на серин, может быть более предпочтительным для разработки фармацевтических средств. Значительное снижение величины Tm для huPM1-SC и huPM1-CS по сравнению с IgG2 может быть следствием паттерна образования дисульфидных связей huPM1-SC и huPM1-CS, который отличается от IgG2.
Более того, сравнение кривых денатурации DSC показало, что пик денатурации для Fab-домена был острым в huPM1-IgG1 и huPM1-SKSC, между тем как он был более широким в huPM1-SC и huPM1-CS, чем у указанных выше двух антител, а huPM1-IgG2 дало пик с выступом на более низкотемпературной стороне пика денатурации Fab-домена. Пик денатурации в DSC, как правило, становится острым в случае одного компонента, но может становиться шире, когда присутствуют два или более компонентов с отличающимися величинами Tm (а именно, гетерогенностью). Таким образом, было предположено, что huPM1-IgG2, huPM1-SC и huPM1-CS содержат два или более компонентов, и гетерогенность природного IgG2 не снижалась в huPM1-SC и huPM1-CS. Эти данные указывают на то, что остатки цистеина, присутствующие как в шарнирной области, так и домене CH1, вовлечены в гетерогенность природного IgG2, и для снижения гетерогенности на DSC необходимо изменить не только цистеин в шарнирной области, но также цистеин в CH1-домене. Более того, как описано выше, стабильности, эквивалентной стабильности природного IgG2, можно достигнуть только посредством замены не только цистеина в шарнирной области, но также цистеина в CH1-домене.
Как описано выше, в отношении константных областей, в которых гетерогенность вследствие шарнирной области IgG2 является сниженной, было выявлено, что SC и CS, которые представляют собой константные области, в которых заменен только цистеин в шарнирной области на серин, могут быть недостаточными с точки зрения гетерогенности и стабильности, и что значительно снизить гетерогенность при сохранении стабильности, сравнимой с IgG2, можно только посредством дополнительной замены цистеина в положении 131 согласно нумерации по EU в CH1-домене серином. Такие константные области включают SKSC.
[Справочный пример 8] Получение и оценка оптимизированной, не связывающей Fcγ-рецептор константной области M14
В связывающем Fcγ-рецептор домене константной области остатки в положениях 233, 234, 235 и 236 согласно нумерации EU являются остатками не связывающего типа, а остатки в положениях 327, 330 и 331 согласно нумерации EU, отличаются от остатков IgG4, которые являются остатками не связывающего типа. Таким образом, необходимо изменить аминокислоты в положениях 327, 330 и 331 согласно нумерации EU на последовательность IgG4 (G2Δa в Eur J Immunol. 1999 Aug; 29(8):2613-24). Однако поскольку аминокислота в положении 339 согласно нумерации EU представляет собой аланин в IgG4, в то время как она представляет собой треонин в IgG2, замена только аминокислот в положениях 327, 330 и 331 согласно нумерации EU на последовательность IgG4 приведет к образованию новой не встречающейся в природе пептидной последовательности из 9-аминокислот, которая может представлять собой пептид T-клеточного эпитопа, тем самым, обеспечивая риск иммуногенности. Таким образом, было выявлено, что наличие новой пептидной последовательности можно предотвратить путем замены треонина в положении 339 согласно нумерации EU в IgG2 на аланин, в дополнение к заменам, описанным выше. В дополнение к мутациям, описанным выше, метионин в положении 397 согласно нумерации EU заменяли на валин для повышения стабильности IgG2 в кислых условиях. Более того, в SKSC (SEQ ID NO: 62), полученном согласно справочному примеру 7, в котором гетерогенность вследствие дисульфидных связей в шарнирной области снижена, внесение мутаций в положениях 131 и 133 приведет к созданию новой не встречающейся в природе пептидной последовательности из 9-аминокислот, которая может представлять собой пептид T-клеточного эпитопа, тем самым, обеспечивая риск иммуногенности. Таким образом, пептидную последовательность в области положений с 131 по 139 преобразовывали в такую же пептидную последовательность, как и в IgG1, мутацией глутаминовой кислоты в положении 137 согласно нумерации EU на глицин и мутацией серина в положении 138 согласно нумерации EU на глицин. Путем внесения всех указанных выше мутаций получили последовательность константной области M14 (SEQ ID NO: 129).
Экспрессию и очистку huPM1-M14, полученного с использованием huPM1-M14 в качестве H-цепи и huPM1-L (SEQ ID NO: 162) в качестве L-цепи, проводили способом, описанным в справочном примере 7. Полученные huPM1-M14 (SEQ ID NO: 163), huPM1-IgG1 и huPM1-IgG2 оценивали в отношении гетерогенности с использованием катионообменной хроматографии способом, описанным в справочном примере 7.
Как представлено на фиг. 14, гетерогенность huPM1-M14 была снижена так же, как и у huPM1-SKSC.
[Справочный пример 9] Получение huPM1-M58 со сниженной C-концевой гетерогенностью H-цепи и улучшенной фармакокинетикой
Получение молекулы huPM1-M58
huPM1 представляет собой антитело IgG1. Для гетерогенности C-концевой последовательности H-цепи антитела IgG были описаны делеция C-концевого остатка лизина и амидация C-концевой аминогруппы вследствие делеции двух C-концевых аминокислот, глицина и лизина (Anal Biochem. 2007 Jan 1; 360(1): 75-83). Также, в huPM1, хотя основным компонентом является последовательность, в которой C-концевой лизин, кодируемый нуклеотидной последовательностью, удален посредством посттрансляционной модификации, также существует второстепенный компонент, в котором лизин остается, и второстепенный компонент, в котором C-концевая аминогруппа является амидированной вследствие делеции как глицина, так и лизина, что приводит к гетерогенности. Продукция фармацевтического средства в большом масштабе с сохранением отличий в гетерогенности желательных/родственных веществ между продуктами является нелегкой, и даже приводит к повышению затрат, и, таким образом, желательно, чтобы вещество состояло из одного вещества, насколько это возможно. Когда антитело разрабатывают в качестве фармацевтического средства, является желательным снижение гетерогенности. Таким образом, желательно, чтобы при разработке в качестве фармацевтических средств C-конец H-цепи не обладал гетерогенностью. Также является желательным продление времени полужизни антитела для снижения дозы антитела.
Таким образом, вносили изменения, описанные ниже, для получения новой константной области, в которой снижена гетерогенность на C-конце H-цепи, улучшена фармакокинетика по сравнению с huPM1-IgG1, и также снижена гетерогенность, источником которой является IgG2 дикого типа, без потери стабильности.
Конкретно, в huPM1-SKSC, который обладает высокой стабильностью и в котором снижена указанная выше гетерогенность, связанная с антителами с константными областями изотипа IgG2, глутаминовую кислоту в положении 137 согласно нумерации EU заменяли глицином; серин в положении 138 заменяли глицином; гистидин в положении 268 заменяли глутамином; аргинин в положении 355 заменяли глутамином; и глутамин в положении 419 заменяли глутаминовой кислотой. В дополнение к описанным выше заменам, глицин и лизин в положениях 446 и 447 удаляли для снижения гетерогенности C-конца H-цепи, тем самым, получая huPM1-M58 (SEQ ID NO: 164). huPM1-M58, полученный с использованием huPM1-M58 в качестве H-цепи и huPM1-L (SEQ ID NO: 162) в качестве L-цепи, экспрессировали и очищали способом, описанным в примере 4.
huPM1-M58, huPM1-IgG1 и huPM1-IgG2 оценивали в отношении гетерогенности и стабильности способами, описанными в примере 5, с использованием катионообменной хроматографии и DSC, соответственно.
Результат DSC представлен в таблице 18. Как представлено на фиг. 13 и 16, было выявлено, что huPM1-M58 демонстрирует сниженную гетерогенность без утраты стабильности, как в huPM1-SKSC.
Оценка времени удержания huPM1-M58 в плазме
Пролонгированное удержание (медленное выведение) молекулы IgG в плазме является следствием функции FcRn, который известен как рецептор спасения для молекулы IgG (Nat Rev Immunol. 2007 Sep; 7(9): 715-25). При включении в эндосомы путем пиноцитоза, молекулы IgG связываются с FcRn, экспрессированным в эндосомах, в кислых условиях в эндосоме (приблизительно pH 6,0). Хотя молекулы IgG, которые не связываются с FcRn, переносятся в лизосомы и деградируют в них, молекулы IgG, связанные с FcRn, перемещаются на клеточную поверхность, а затем снова высвобождаются из FcRn в плазму при нейтральных условиях в плазме (приблизительно pH 7,4).
Известно, что антитела типа IgG включают изотипы IgG1, IgG2, IgG3 и IgG4. Время полужизни этих изотипов в плазме у человека, согласно сообщениям, составляет приблизительно 36 суток для IgG1 и IgG2; приблизительно 29 суток для IgG3; и 16 суток для IgG4 (Nat. Biotechnol. 2007 Dec; 25(12): 1369-72). Таким образом, полагают, что время удержания IgG1 и IgG2 в плазме является наибольшим. Как правило, изотипы антител, используемых в качестве фармацевтических средств, представляют собой IgG1, IgG2и IgG4. Описанные способы для дальнейшего улучшения фармакокинетики этих IgG-антител включают способы улучшения описанной выше активности связывания с FcRn путем изменения последовательности константной области IgG (J. Biol. Chem. 2007 Jan 19; 282(3): 1709-17; J. Immunol. 2006 Jan 1; 176(1): 346-56).
Существуют видовые отличия между FcRn мыши и FcRn человека (Proc. Natl. Acad. Sci. USA. 2006 Dec 5; 103(49): 18709-14). Таким образом, для предсказания времени удержания IgG-антител, имеющих измененную последовательность константной области, в плазме человека может быть желательной оценка связывания с FcRn человека и времени удержания в плазме у трансгенных по FcRn человека мышей (Int. Immunol. 2006 Dec; 18(12): 1759-69).
Оценка связывания с FcRn человека
FcRn представляет собой комплекс FcRn и β2-микроглобулина. Олиго-ДНК-праймеры получали на основе опубликованной последовательности гена FcRn человека (J. Exp. Med. (1994) 180 (6), 2377-2381). Фрагмент ДНК, кодирующий целый ген, получали посредством ПЦР с использованием кДНК человека (Human Placenta Marathon-Ready cDNA, Clontech) в качестве матрицы и полученных праймеров. С использованием полученного фрагмента ДНК в качестве матрицы, фрагмент ДНК, кодирующий внеклеточный домен, содержащий сигнальную область (Met1-Leu290) амплифицировали посредством ПЦР, и встраивали в экспрессирующий вектор клеток животных (аминокислотная последовательность FcRn человека/SEQ ID NO: 165). Аналогично, олиго-ДНК-праймеры получали на основе опубликованной последовательности гена β2-микроглобулина человека (Proc. Natl. Acad. Sci. U.S.A. 99 (26), 16899-16903 (2002)). Фрагмент ДНК, кодирующий целый ген, получали посредством ПЦР с использованием кДНК человека (Hu-Placenta Marathon-Ready кДНК, CLONTECH) в качестве матрицы и полученных праймеров. С использованием полученного фрагмента ДНК в качестве матрицы амплифицировали посредством ПЦР фрагмент ДНК, кодирующий целую содержащую β2-микроглобулин сигнальную область (Met1-Met119), и встраивали его в экспрессирующий вектор клеток животных (аминокислотная последовательность β2-микроглобулина человека/SEQ ID NO: 166).
Растворимый FcRn человека экспрессировали с помощью следующего способа. Полученные плазмиды для FcRn человека и β2-микроглобулина вводили в происходящую из рака почки эмбриона человека клеточную линию HEK293H (Invitrogen) с использованием 10% эмбриональной телячьей сыворотки (Invitrogen) посредством липофекции. Полученный культуральный супернатант собирали и очищали с использованием IgG Sepharose 6 Fast Flow (Amersham Biosciences) способом, описанным в J. Immunol. 2002 Nov 1; 169(9):5171-80. Затем проводили дальнейшую очистку с использованием HiTrap Q HP (GE Healthcare).
Связывание с FcRn человека оценивали с использованием Biacore 3000. Антитело связывали с белком L или антителом кролика против IgG человека с каппа-цепью, иммобилизованным на сенсорный чип, добавляли FcRn человека в качестве анализируемого образца для взаимодействия с антителом, и из количества связанного FcRn человека вычисляли аффинность (KD). Конкретно, белок L иммобилизовывали на сенсорный чип CM5 (BIACORE) способом присоединения аминов с использованием 50 мМ Na-фосфатного буфера (pH 6,0), содержащего 150 мМ NaCl в качестве буфера для анализа. Затем антитело разбавляли буфером для анализа, содержащим 0,02% Tween20, и инъецировали и позволяли ему связываться с чипом. Затем инъецировали FcRn человека для оценки активности связывания антитела FcRn человека.
Аффинность вычисляли с использованием программного обеспечения BIAevaluation. Полученную сенсограмму использовали для вычисления количества hFcRn, связавшегося с антителом, непосредственно перед завершением инъекции FcRn человека. Ее подгоняли способом аффинности в стационарном режиме для вычисления аффинности FcRn человека к антителу.
Предсказательная оценка времени удержания в плазме huPM1-IgG1 и huPM1-M58 у человека с использованием FcRn человека
Активность связывания huPM1-IgG1 и huPM1-M58 с FcRn человека оценивали с использованием BIAcore. Как представлено в таблице 19, активность связывания huPM1-M58 превышала активность связывания huPM1-IgG1 приблизительно в 1,4 раз.
Оценка времени удержания в плазме у мышей, трансгенных по FcRn человека
Фармакокинетику у трансгенных по FcRn человека мышей (B6.mFcRn-/-.hFcRn Tg line 276 +/+mice; Jackson Laboratories) оценивали следующим способом. Антитело внутривенно вводили мышам один раз в дозе 1 мг/кг, и проводили забор крови в соответствующие моменты времени. Собранную кровь сразу центрифугировали при 15000 об/мин в течение 15 минут при 4ºC для получения плазмы. Отделенную плазму хранили в морозильной камере при -20ºC или ниже до применения. Концентрацию в плазме определяли посредством ELISA.
Предсказательная оценка времени удержания в плазме huPM1-IgG1 и huPM1-M58 у человека с использованием трансгенных по FcRn человека мышей
Оценивали время удержания в плазме huPM1-IgG1 и huPM1-M58 у трансгенных по FcRn человека мышей. Как представлено на фиг.17, результат показал, что фармакокинетика huPM1-M58 была улучшена по сравнению с huPM1-IgG1. Было предположено, что активность связывания FcRn человека у трансгенных по FcRn человека мышей коррелировала со временем удержания в плазме.
[Справочный пример 10] Измерение аффинности реакции антиген-антитело с использованием Biacore
Анализ кинетики реакции антиген-антитело проводили с использованием Biacore T100 (GE Healthcare Biosciences). Взаимодействие антиген-антитело измеряли путем иммобилизации rec-белка A (в дальнейшем в настоящем документе белка A) (ZYMED) на сенсорный чип, улавливания антитела на иммобилизованный белок A, а затем реакцией с антигеном в качестве анализируемого образца. В качестве антигена использовали различные концентрации rhNR10. Параметры кинетики, константу скорости ассоциации ka (1/Мс) и константу скорости диссоциации kd (1/c), вычисляли из сенсограмм, полученных при измерении. Затем определяли KD (M) на основе констант скорости. Каждый параметр определяли с использованием программного обеспечения Biacore T100 Evaluation Software version 1.1 (GE Healthcare Biosciences).
Иммобилизация белка A на сенсорном чипе
Белок A иммобилизовывали на всех проточных ячейках сенсорного чипа CM5 (GE Healthcare Bioscinences) способом присоединения аминов. Эксперимент проводили с использованием HBS-EP+ (10 мМ HEPES, 0,15M NaCl, 3 мМ ЭДТА, 0,05% об./об. поверхностно-активное вещество P20) в качестве буфера для анализа со скоростью потока 10 мкл/мин. Карбоксильные группы карбоксиметилдекстрана на сенсорном чипе активировали 100 мкл смеси 1:1 75 мг/мл EDC (гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиимида) и 11,5 мг/мл NHS (N-гидроксисукцинимида), и позволяли протекать белку A, приготовленному в концентрации 50 мкг/мл с использованием 10 мМ ацетатного буфера (pH 4,5) для реакции. Затем позволяли протекать 100 мкл 1M гидрохлорида этаноламина (pH 8,5) для инактивации не вступивших в реакцию активных групп. В конечном итоге, иммобилизовывали приблизительно от 4000 до 5000 RU. Эксперимент проводили при 25ºC все время.
Измерение аффинности реакции антиген-антитело между rhNR10 и антителом, уловленном с помощью белка A
Используемый буфер для анализа представлял собой HBS-EP+. Каждое антитело приготавливали в концентрации 0,25 мкг/мл, или получали так, чтобы приблизительно 100 RU связывалось с белком A. rhNR10, используемый в качестве анализируемого образца, приготавливали в концентрации 0, 38,5, 77,0 и 154 нМ, или в концентрации 0, 19,25 и 77,01 нМ с использованием HBS-EP+. При измерении сначала антитело из раствора улавливали с помощью белка A, и проводили реакцию анализируемого раствора со скоростью потока 20 мкл/мин в течение трех минут. Затем раствор заменяли на HBS-EP+, и определяли фазу диссоциации в течение пяти минут. После определения фазы диссоциации сенсорный чип подвергали регенерации посредством промывания 10 мМ глицином-HCl (pH 1,5). Полученные сенсограммы подвергали кинетическому анализу с использованием специализированного для Biacore программного обеспечения для анализа данных, Biacore T100 Evaluation Software Version 1.1.
Промышленная применимость
Антитела против NR10, полученные авторами настоящего изобретения, проявляют эффективную нейтрализующую активность против NR10, и они пригодны, например, в лекарственных средствах против воспалительных заболеваний.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ NR10 И ЕГО ПРИМЕНЕНИЕ | 2008 |
|
RU2531521C2 |
| МОЛЕКУЛЫ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2009 |
|
RU2430111C1 |
| ФАРМАЦЕВТИЧЕСКИЙ СОСТАВ, СОДЕРЖАЩИЙ МОЛЕКУЛЫ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2010 |
|
RU2565809C2 |
| АНТИТЕЛА ПРОТИВ ИНТЕРЛЕЙКИНА-13 ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2427589C2 |
| МОДИФИЦИРОВАННАЯ КОНСТАНТНАЯ ОБЛАСТЬ АНТИТЕЛА | 2008 |
|
RU2526512C2 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО, СОДЕРЖАЩЕЕ АНТИТЕЛО К ФОСФОЛИПАЗЕ D4 | 2014 |
|
RU2709741C2 |
| АНТАГОНИСТИЧЕСКИЕ АНТИТЕЛА К IL-17 | 2005 |
|
RU2426741C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ РАКА ПЕЧЕНИ | 2009 |
|
RU2523897C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2812875C1 |
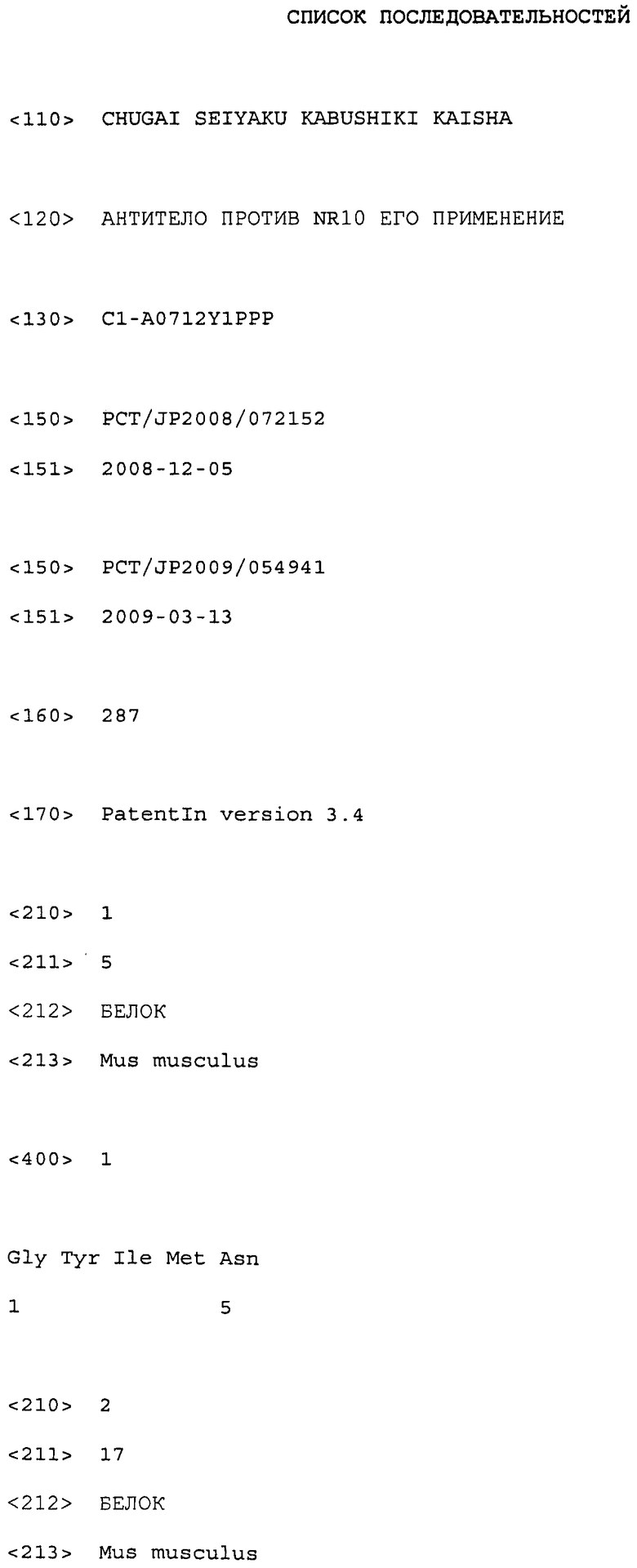

















































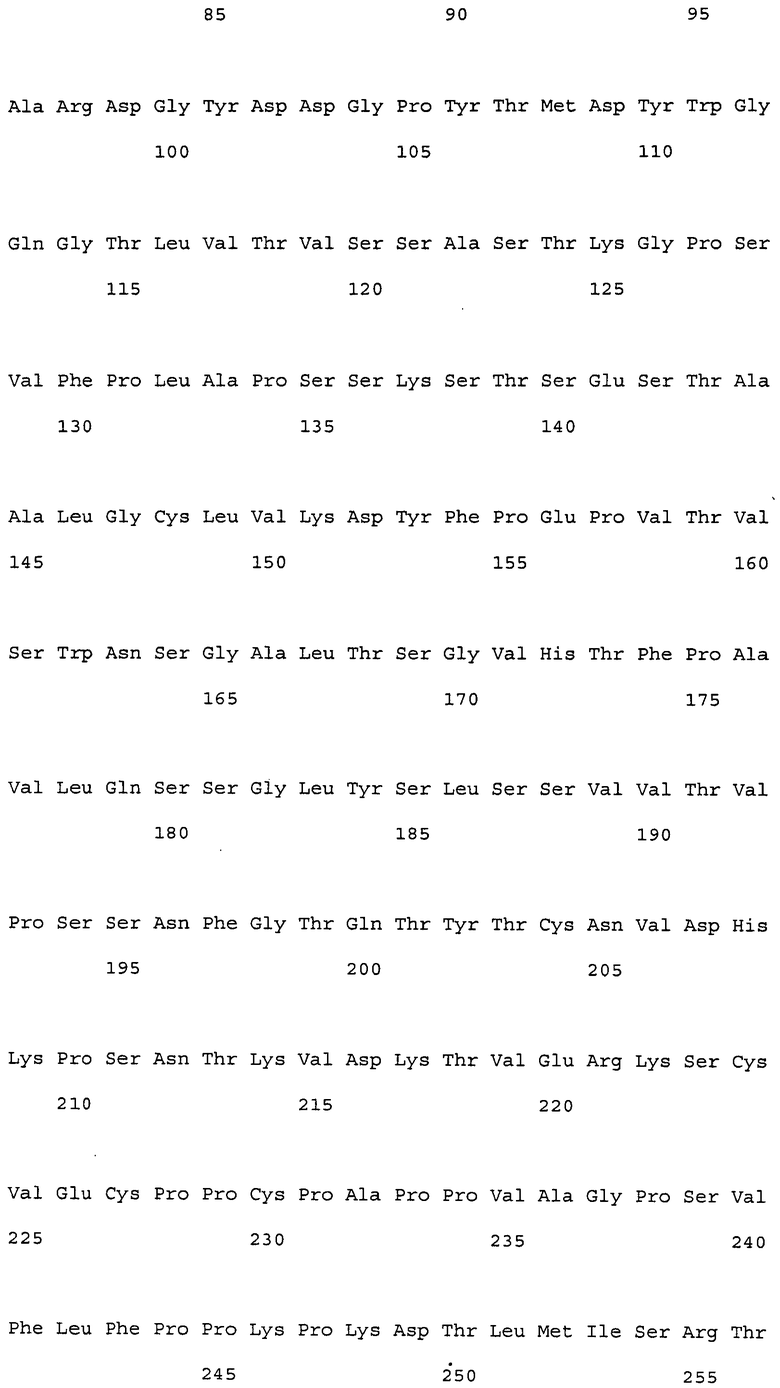
































































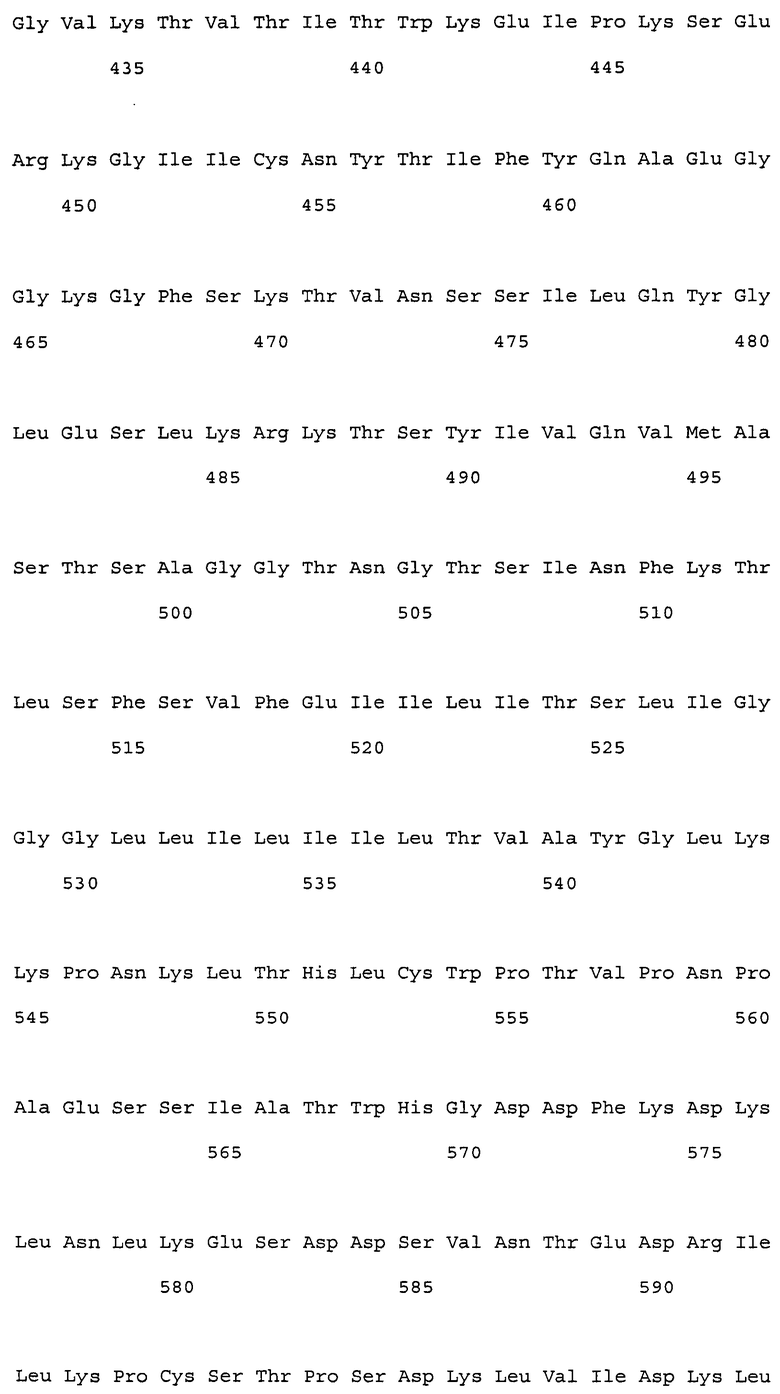


























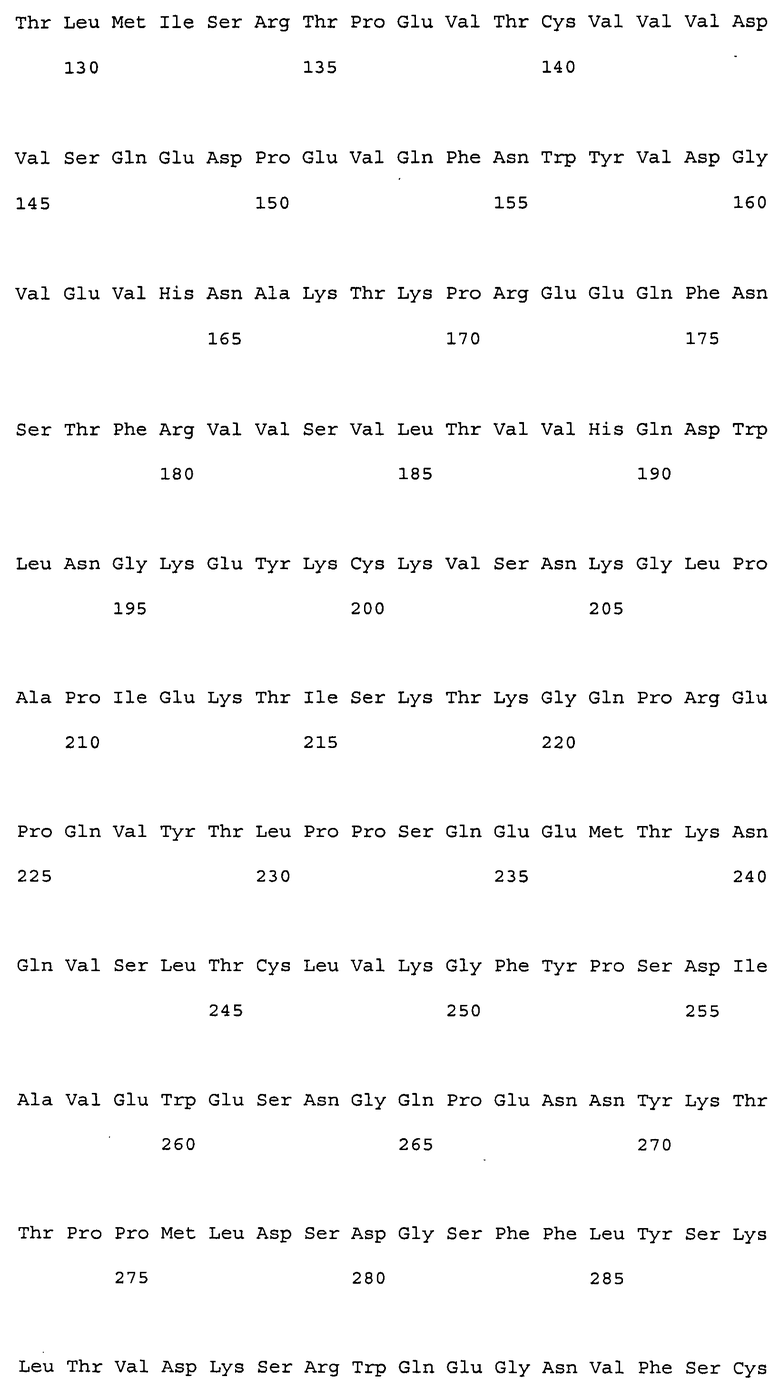




































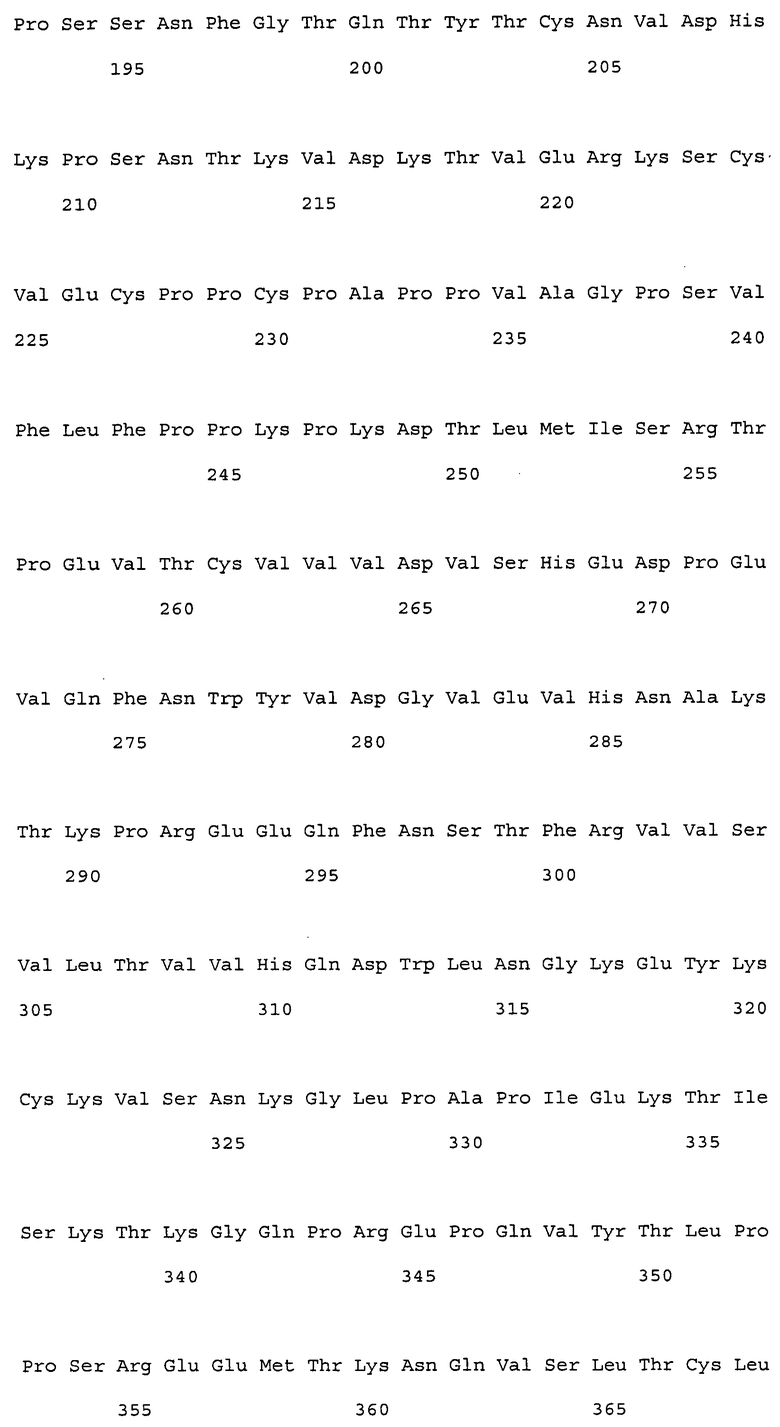















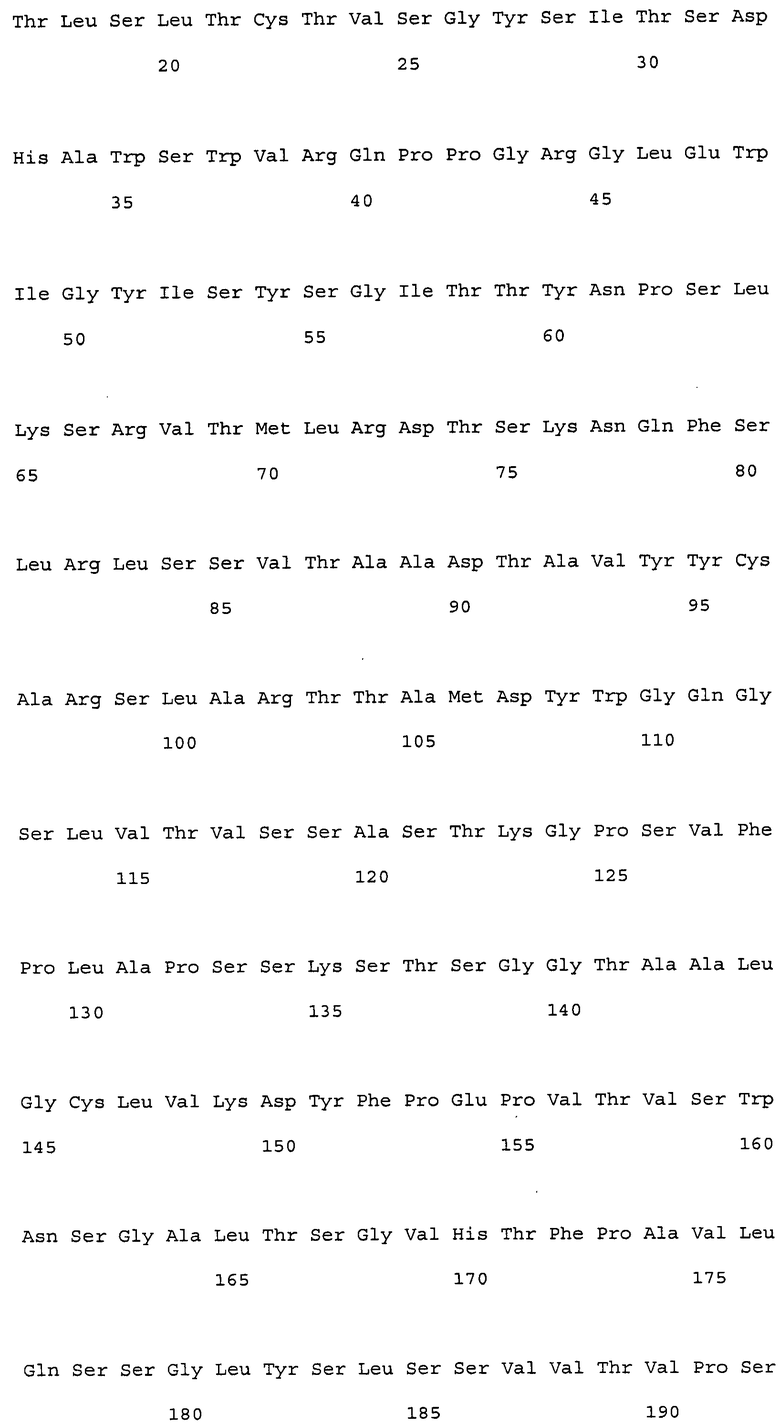
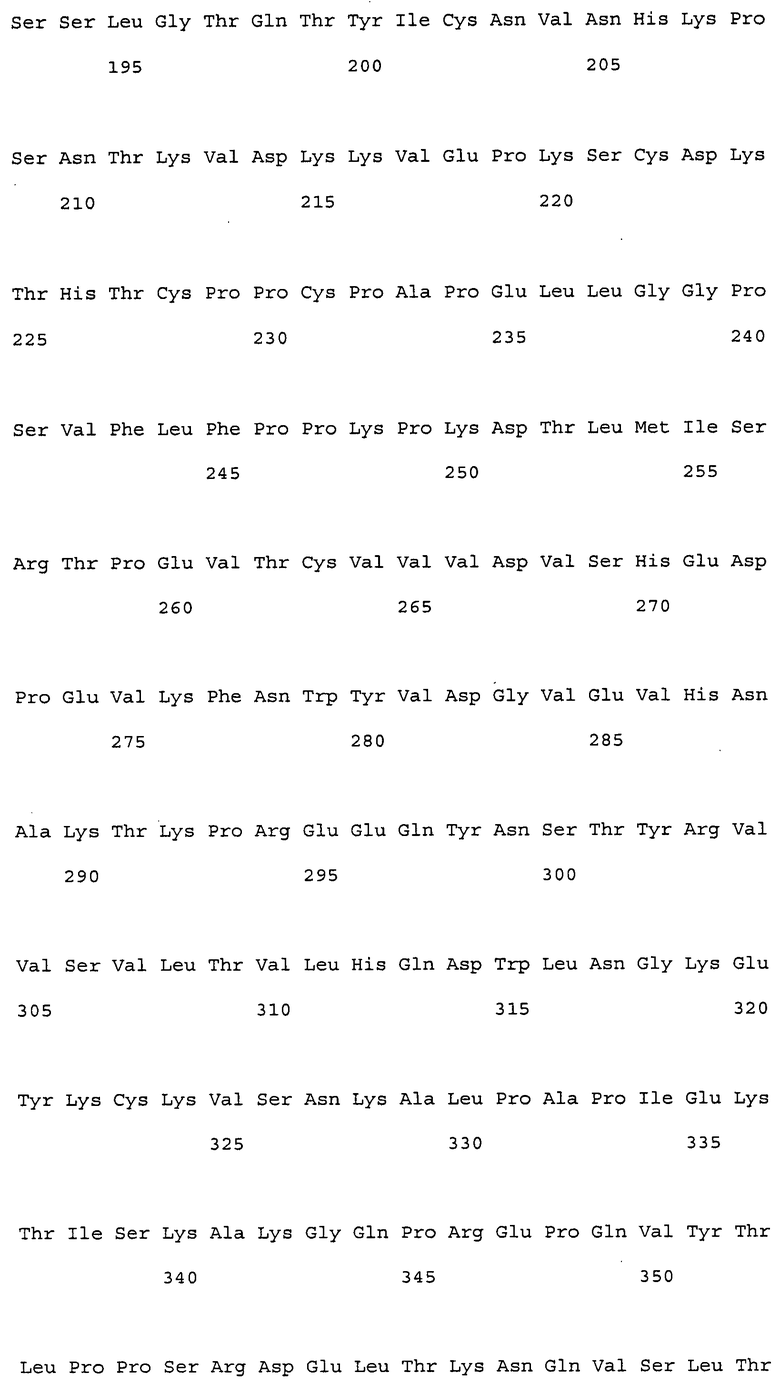


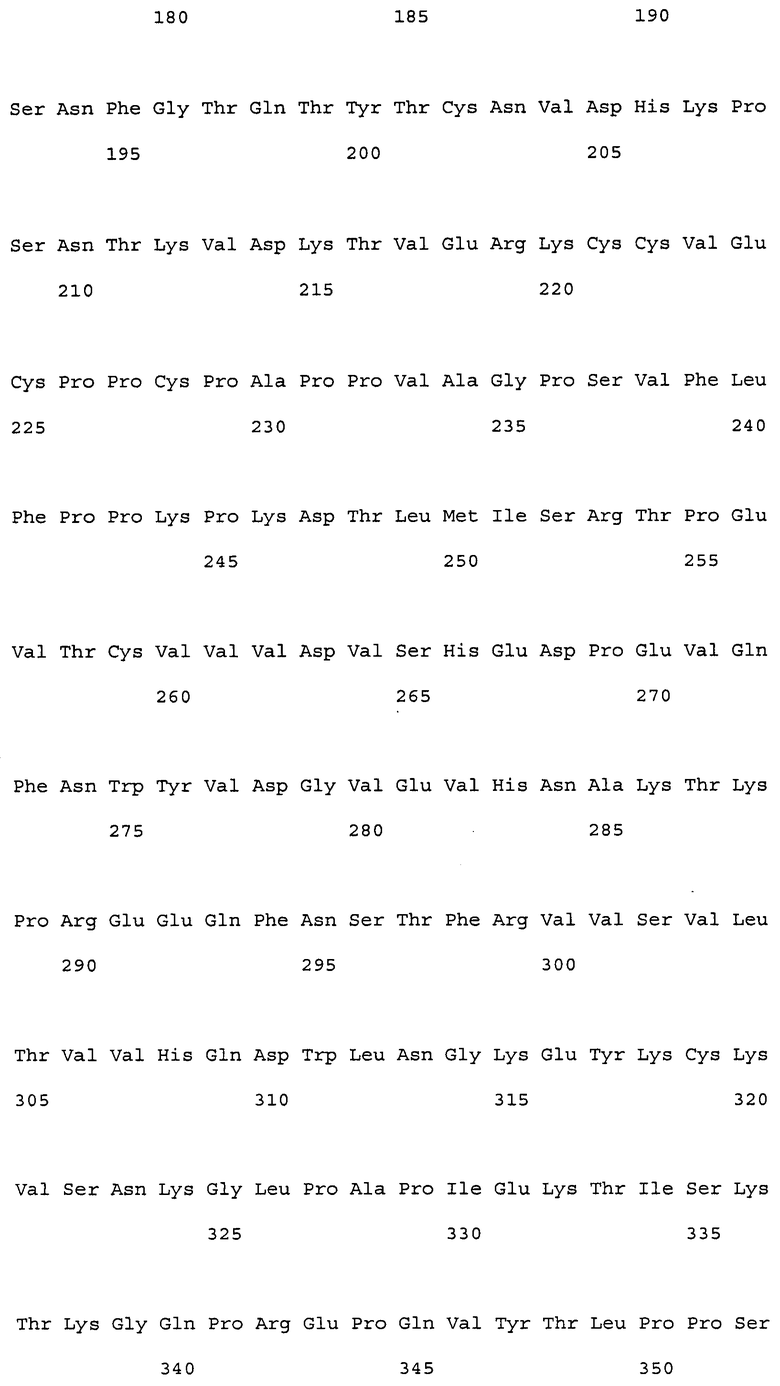


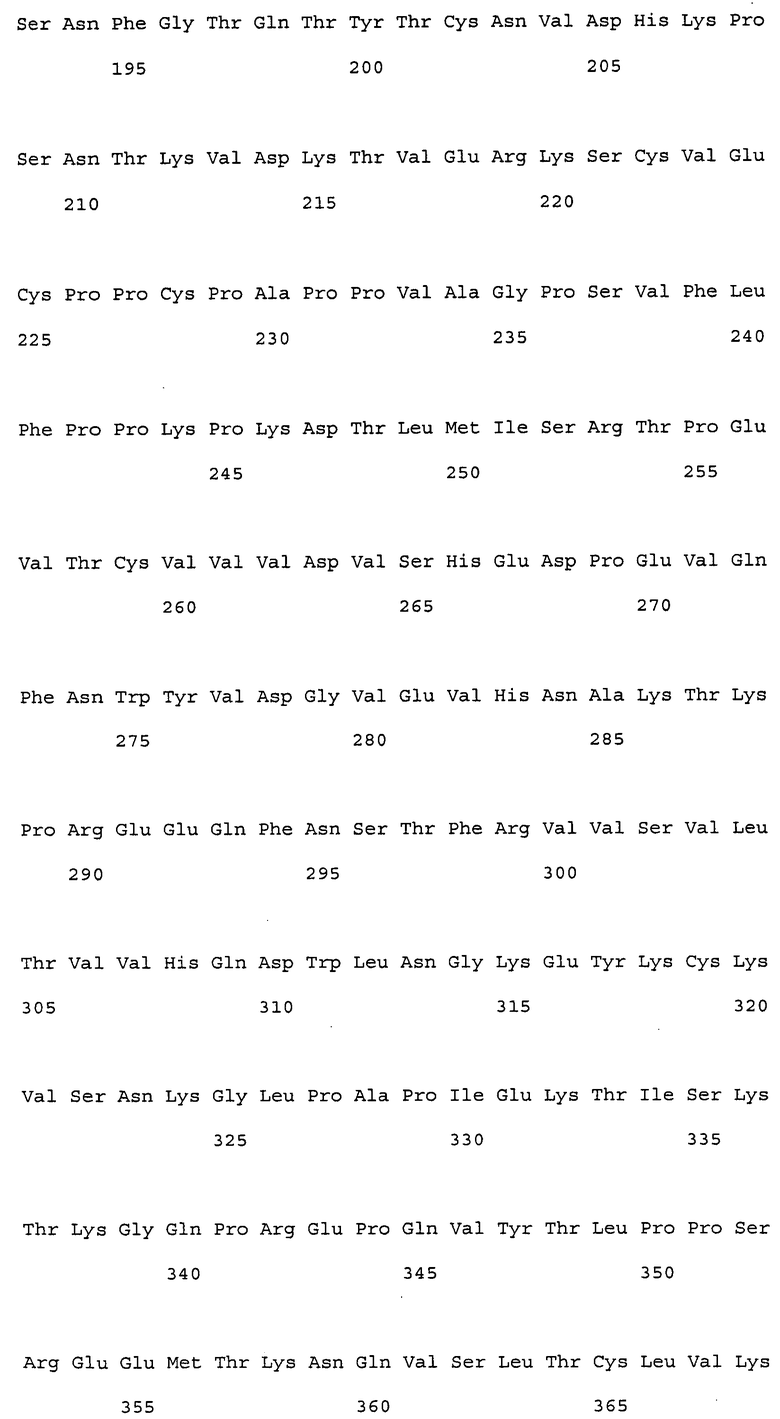

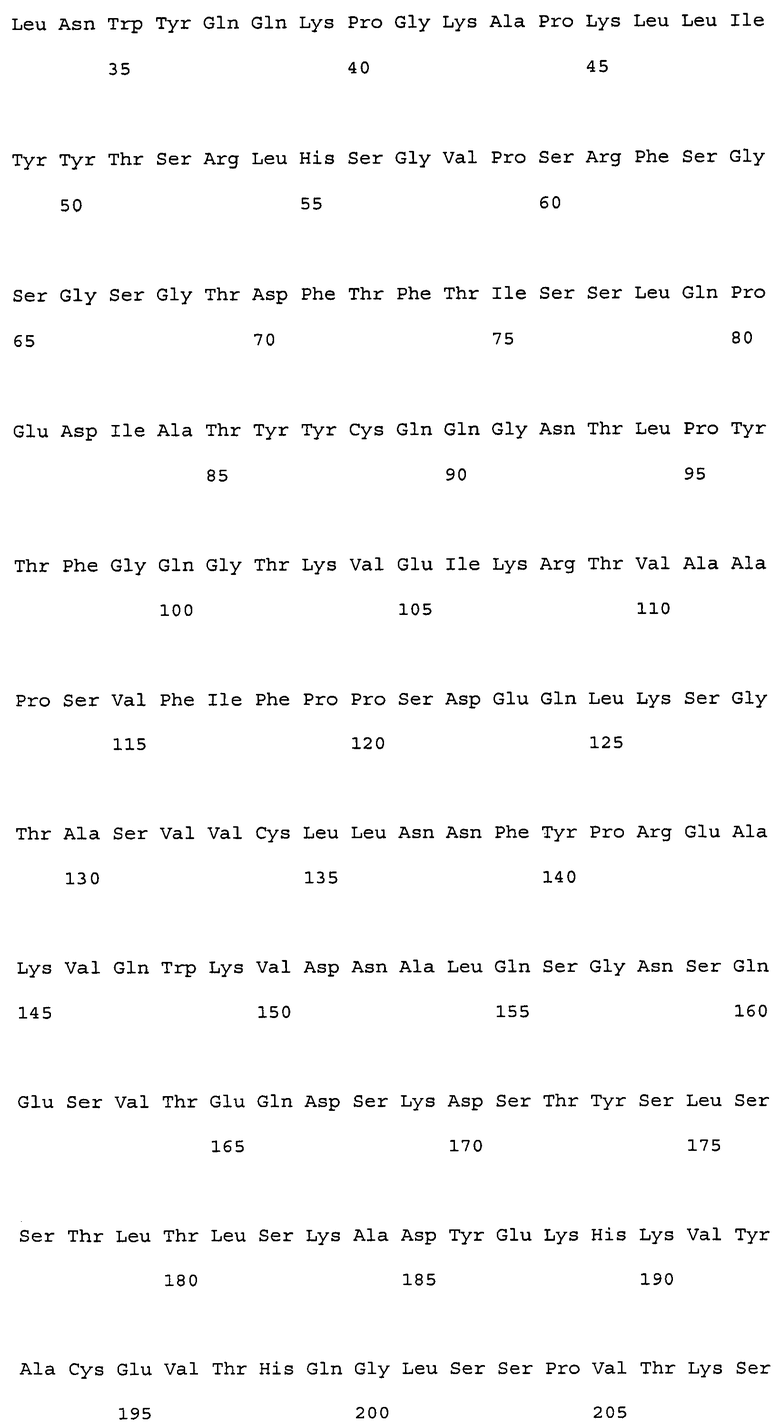




















































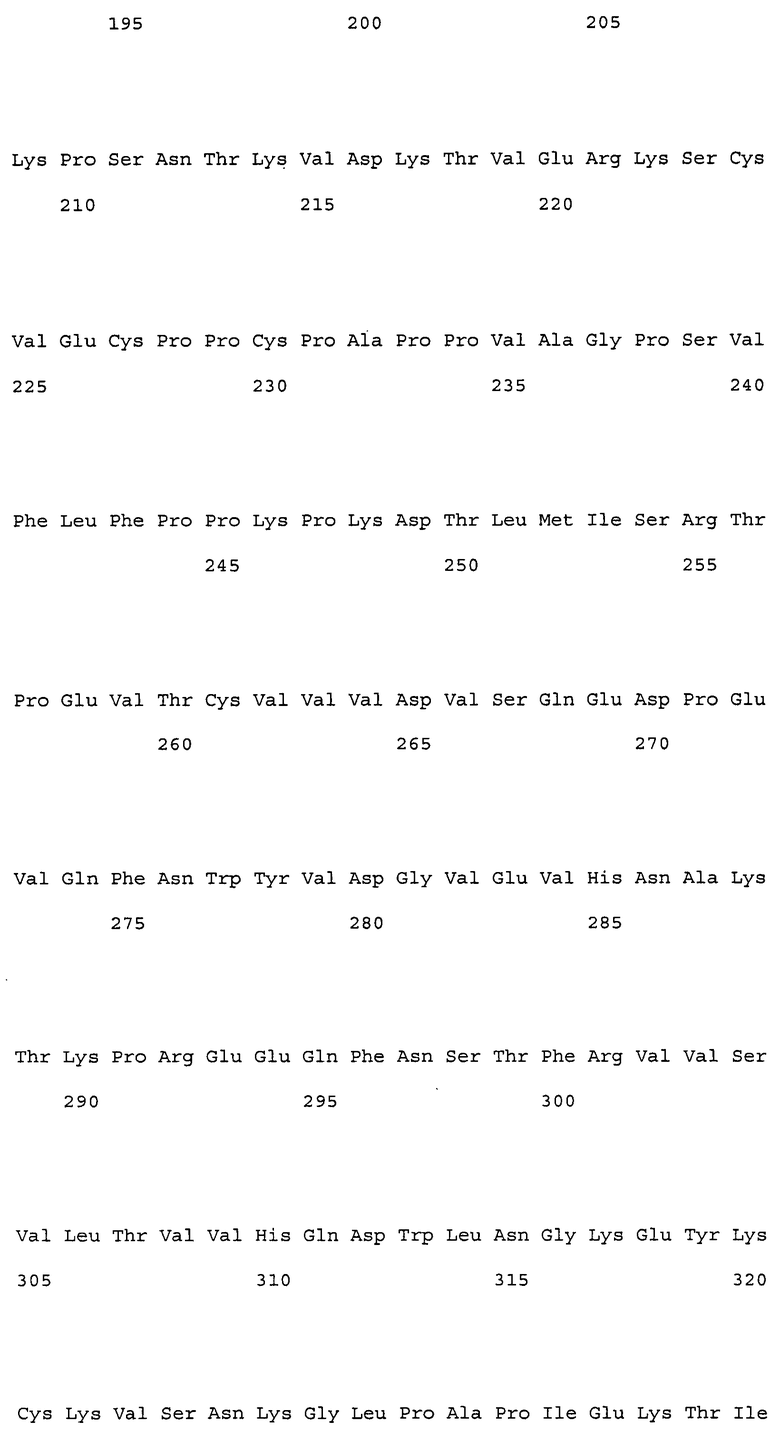
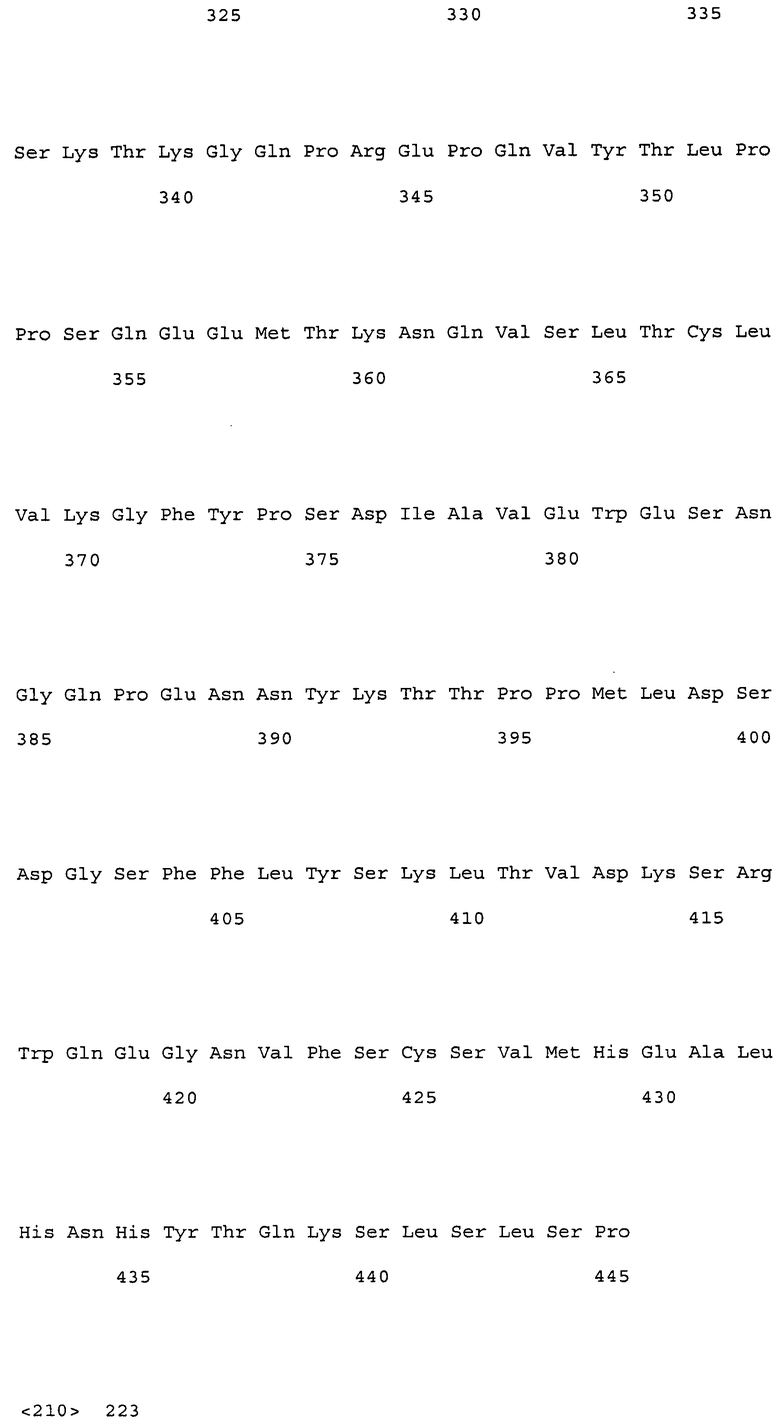









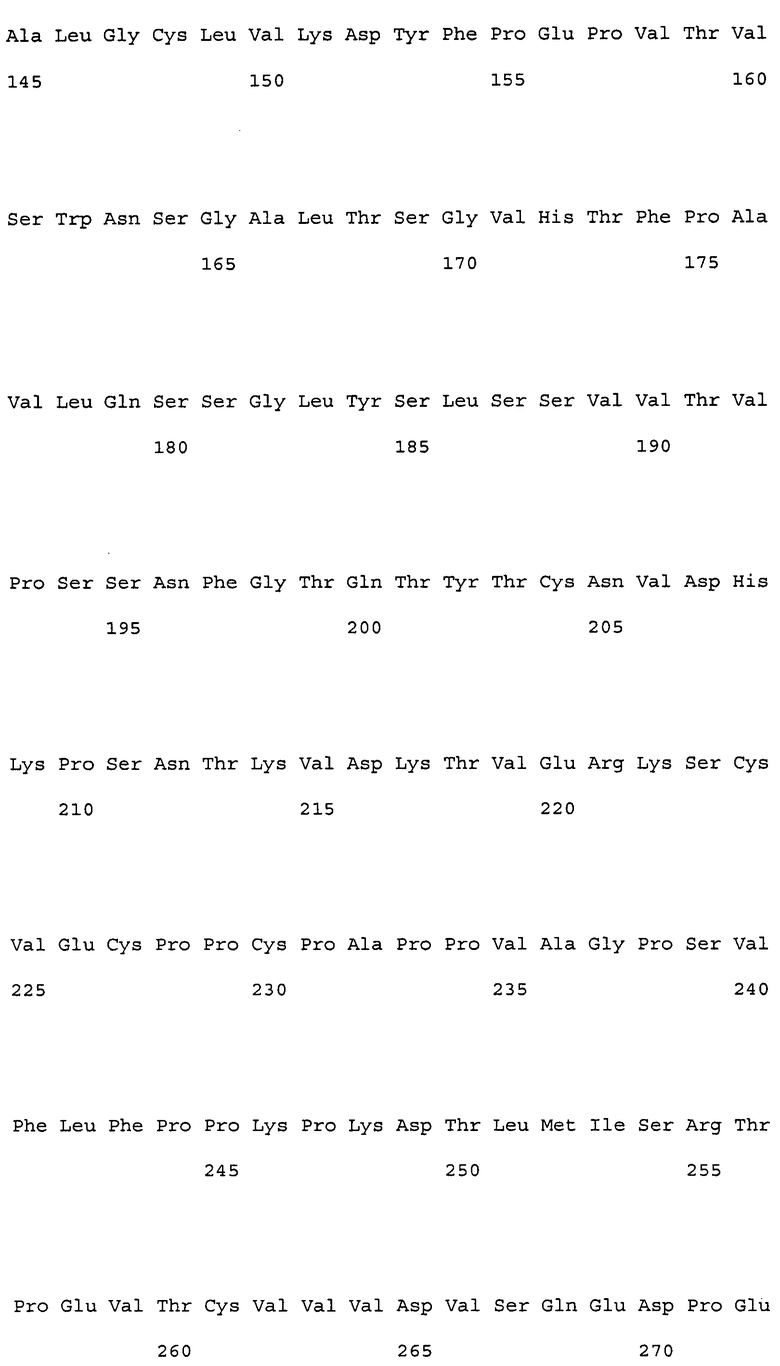






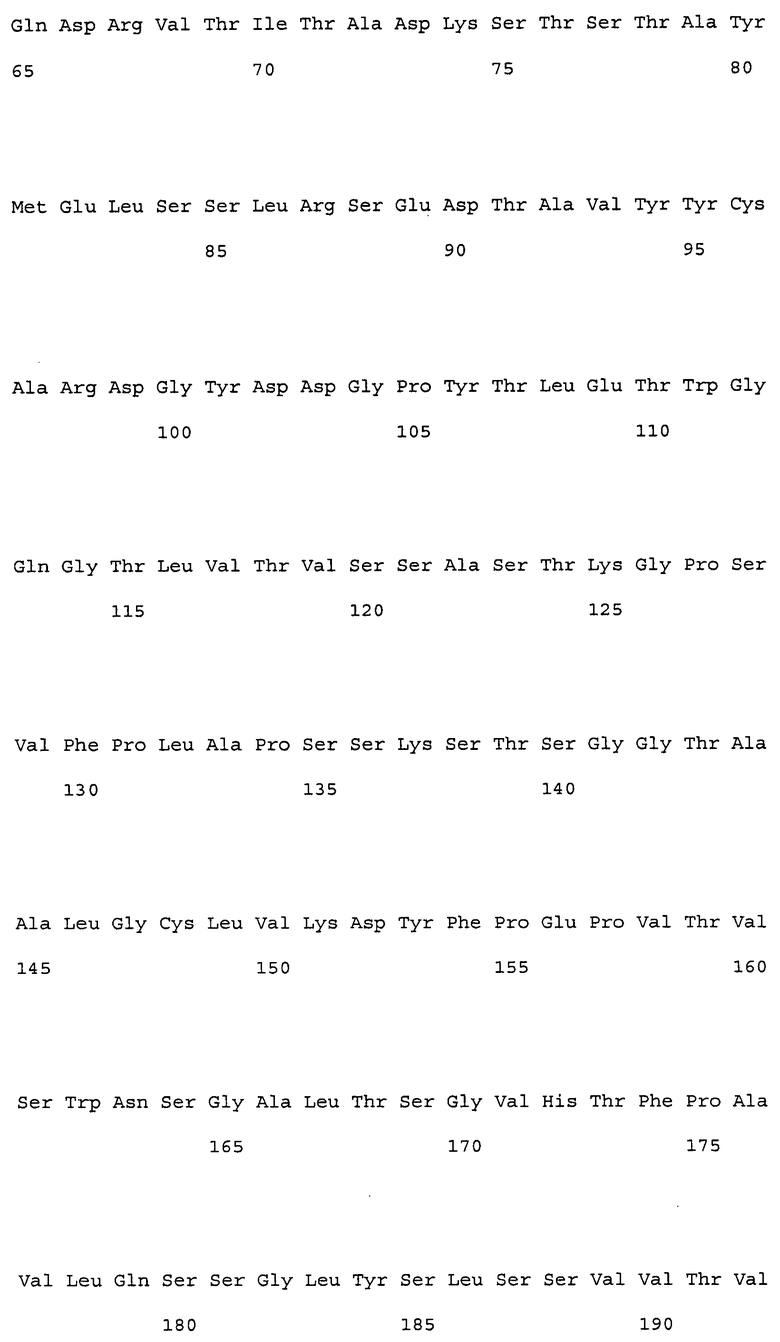

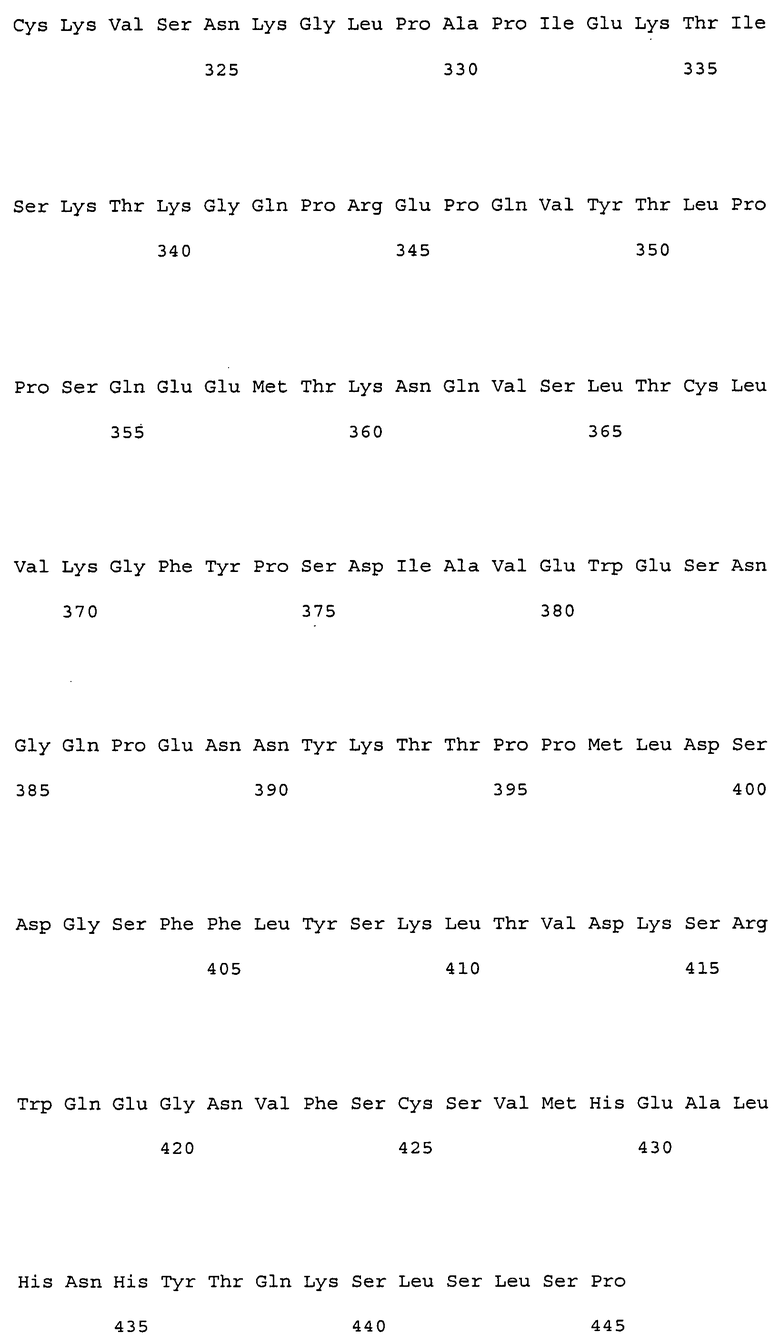

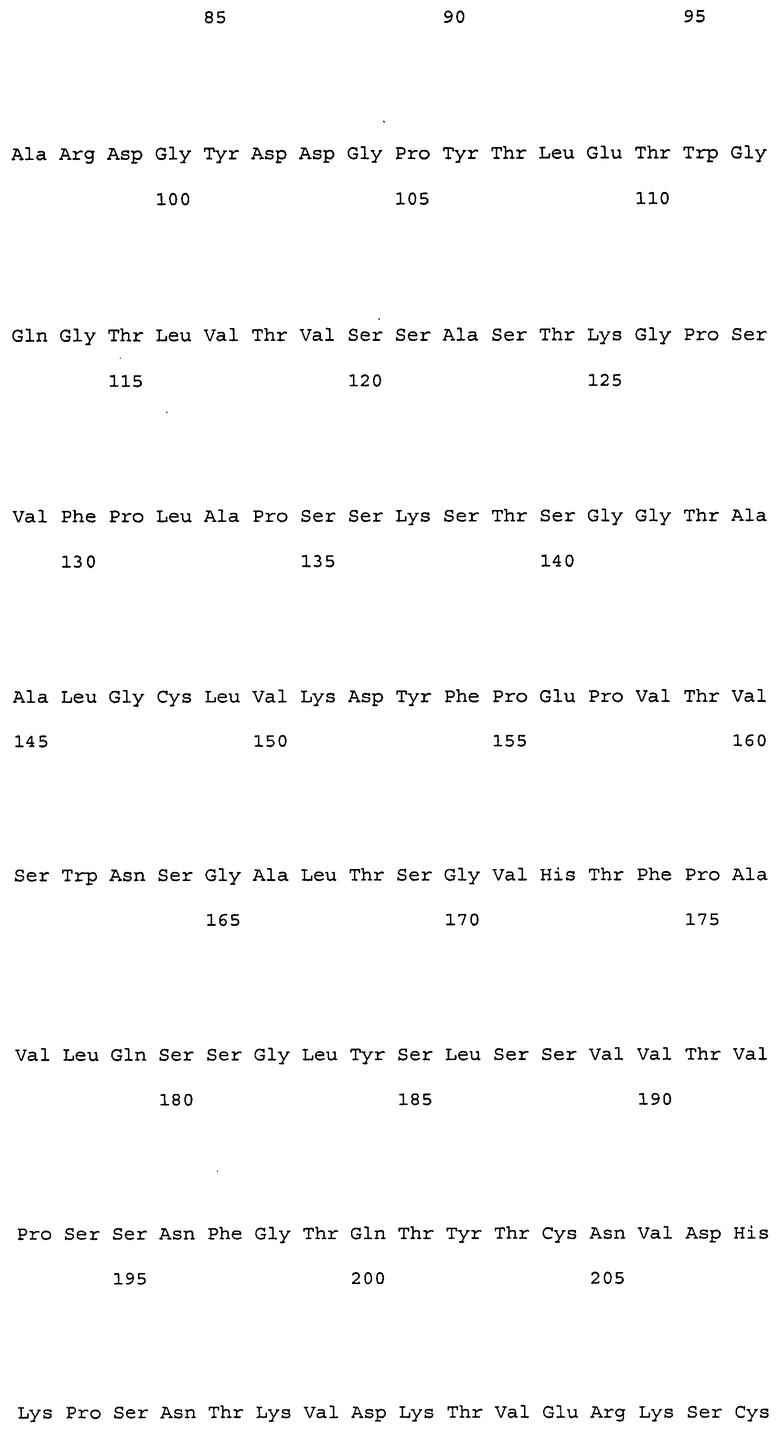

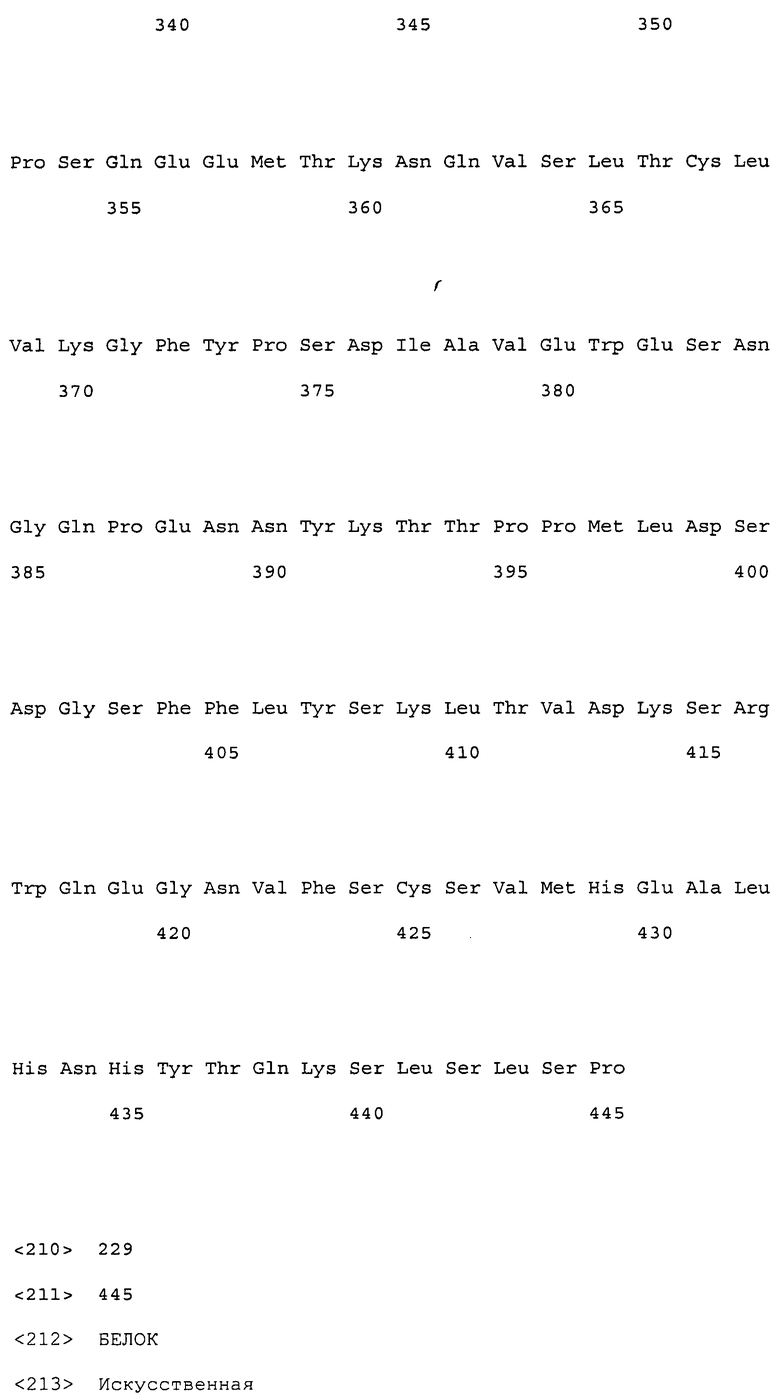


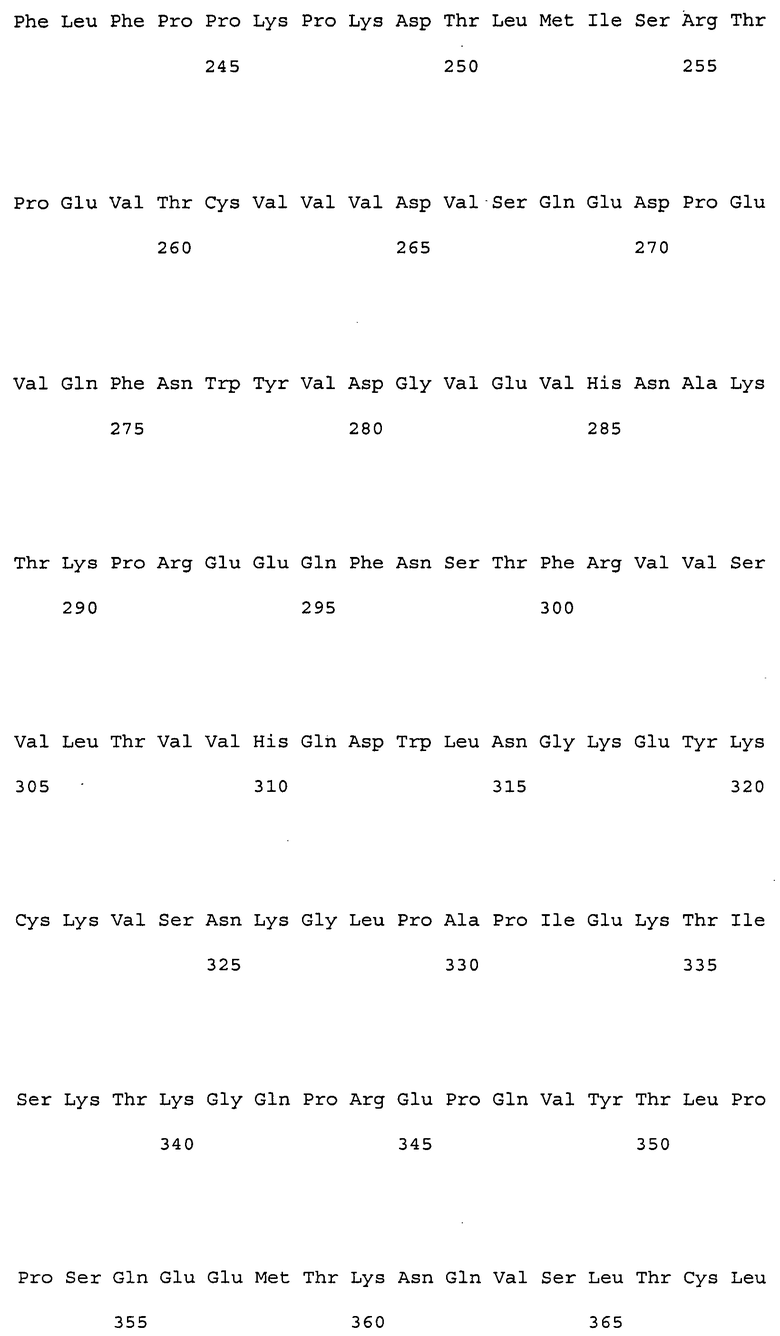











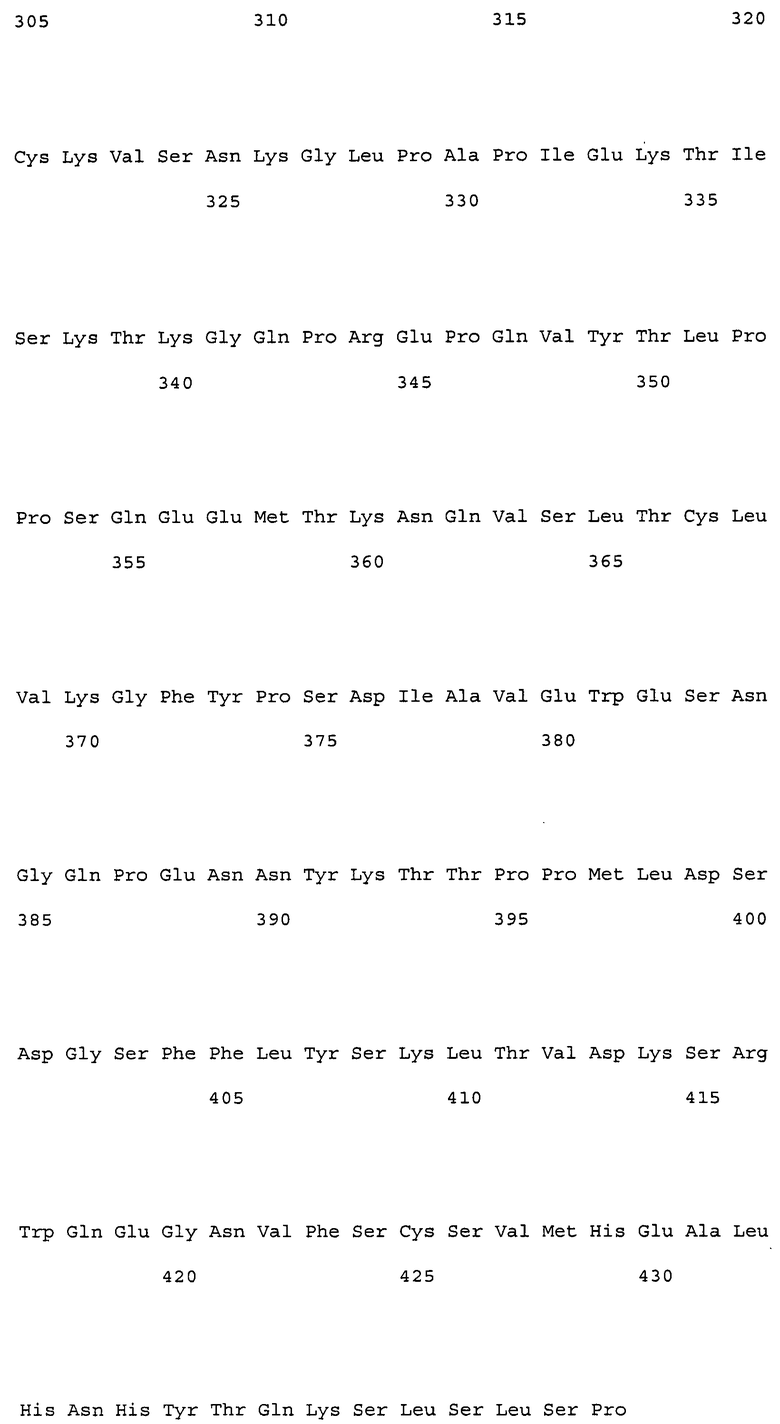


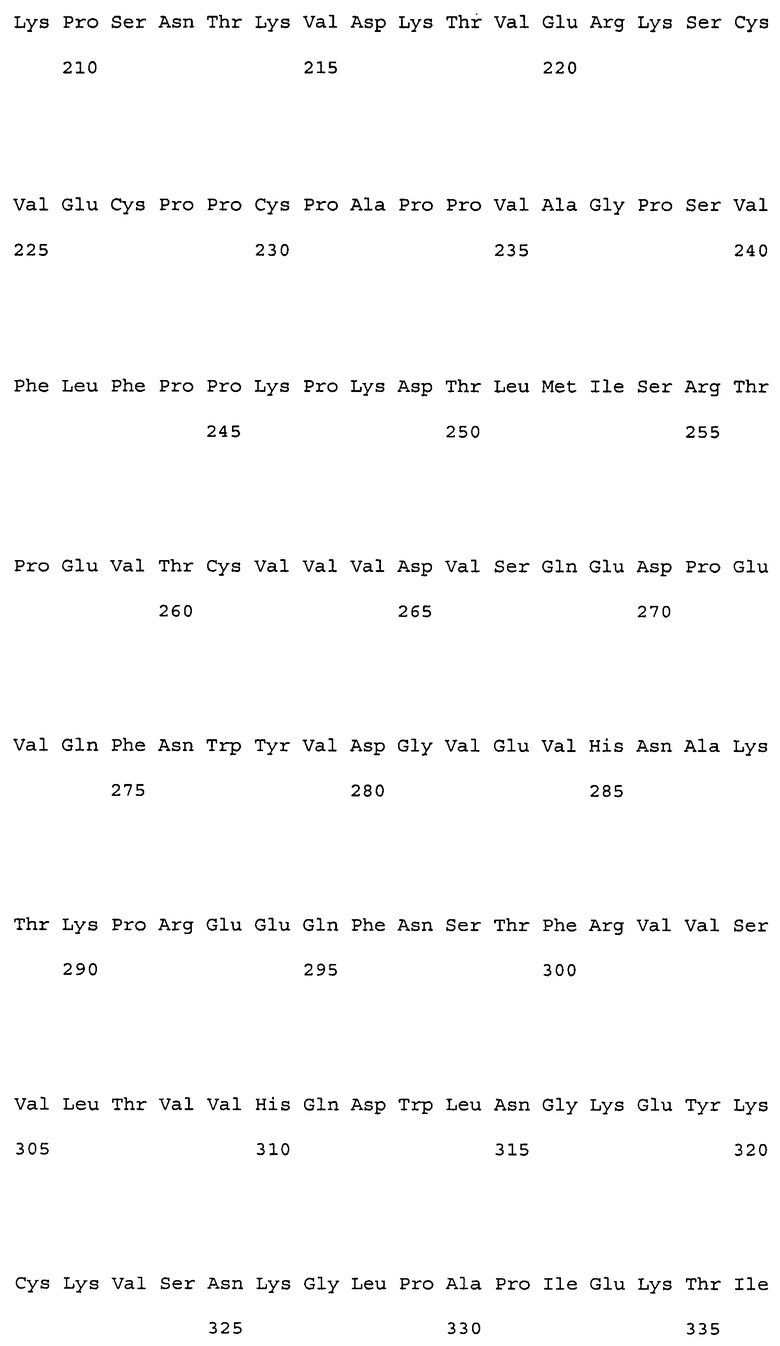

















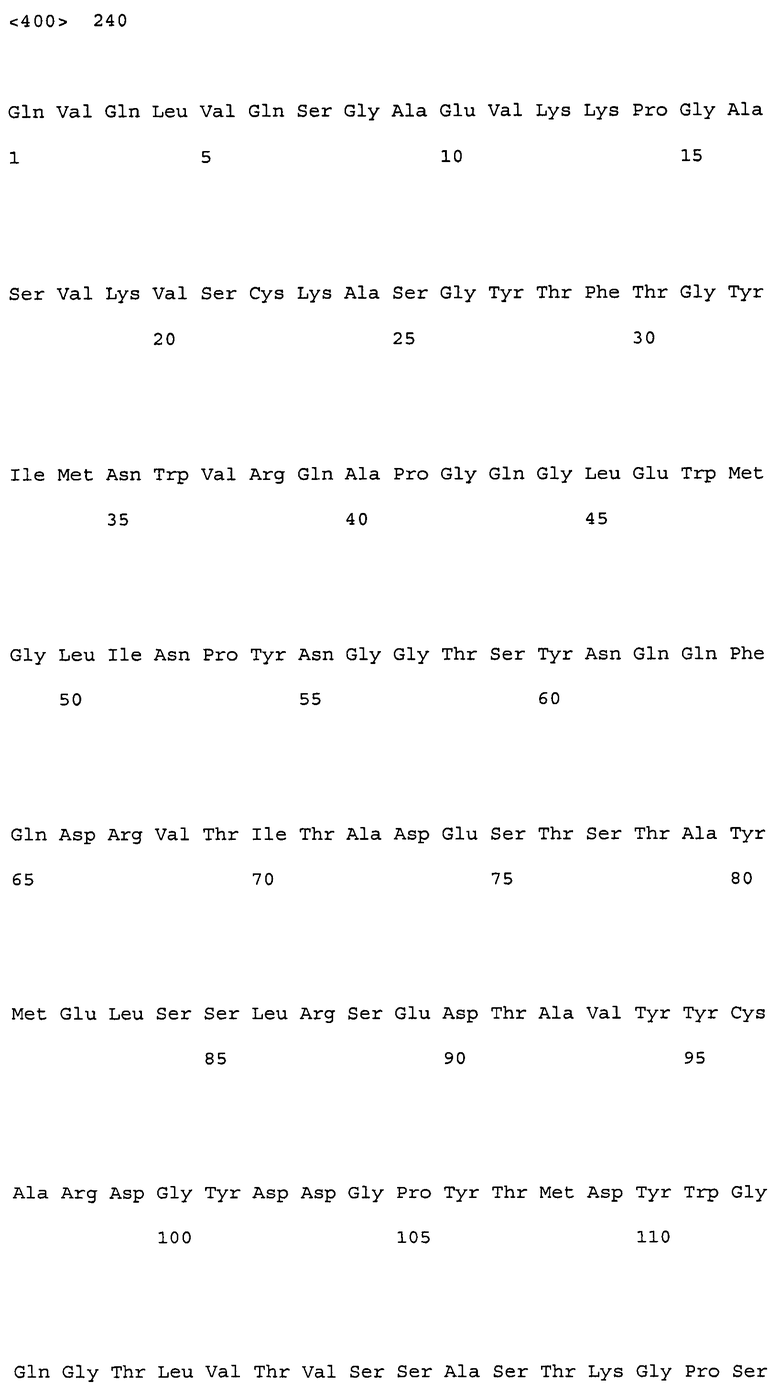




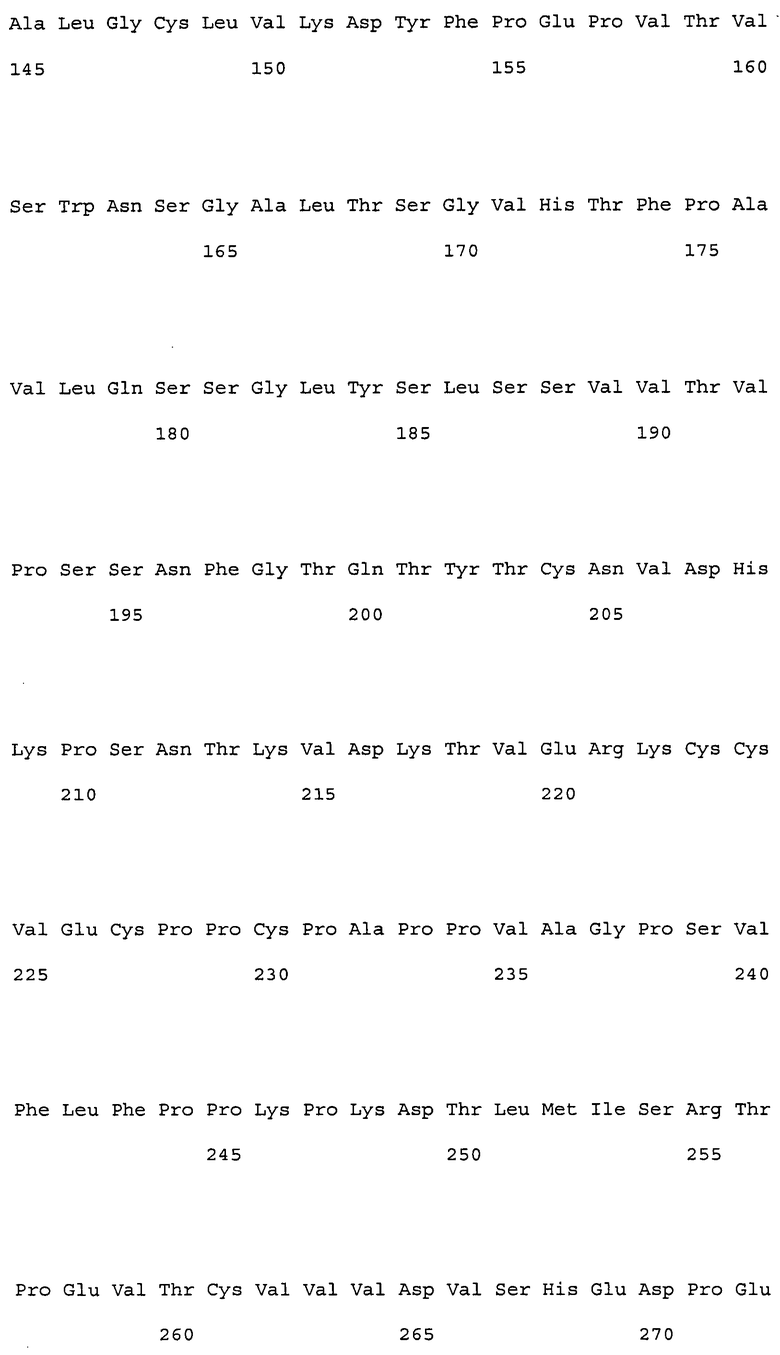



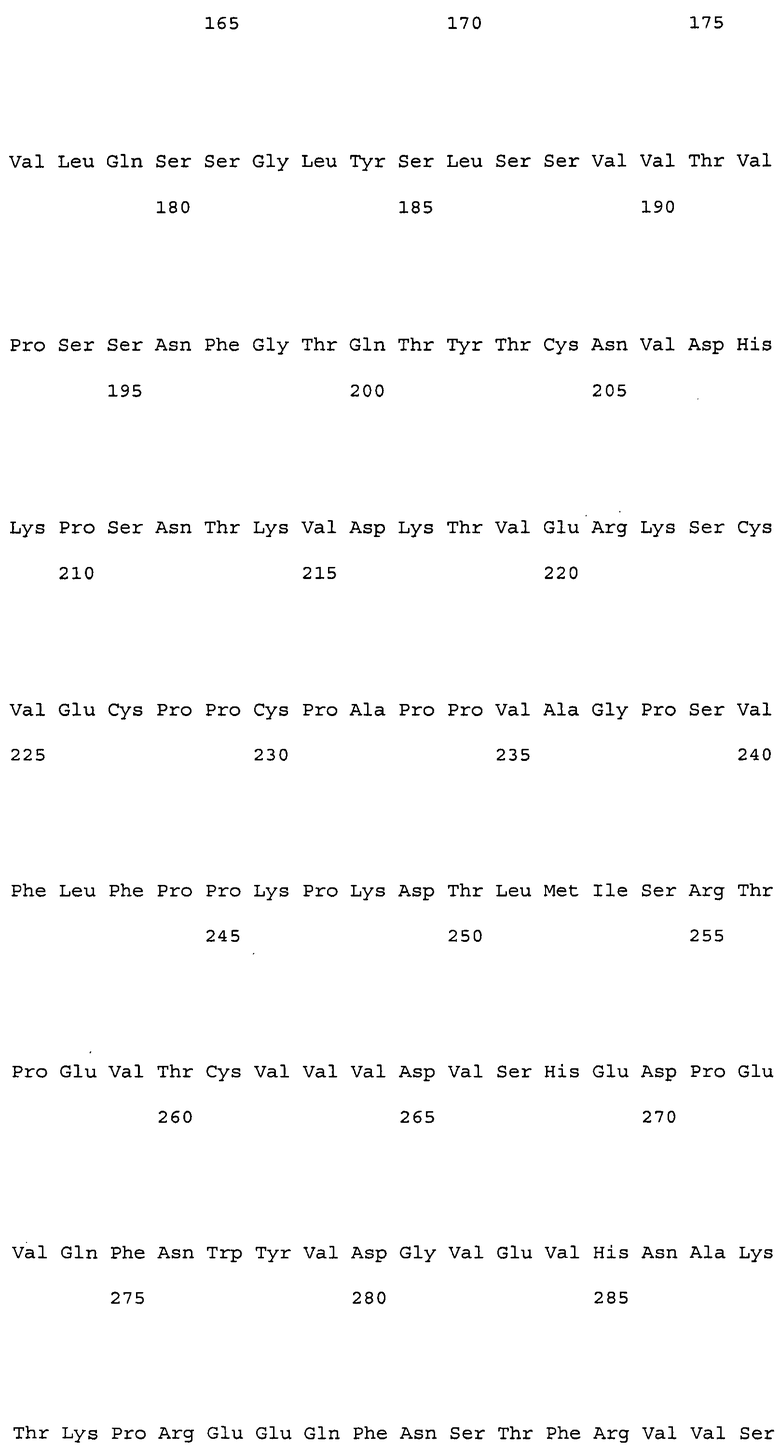










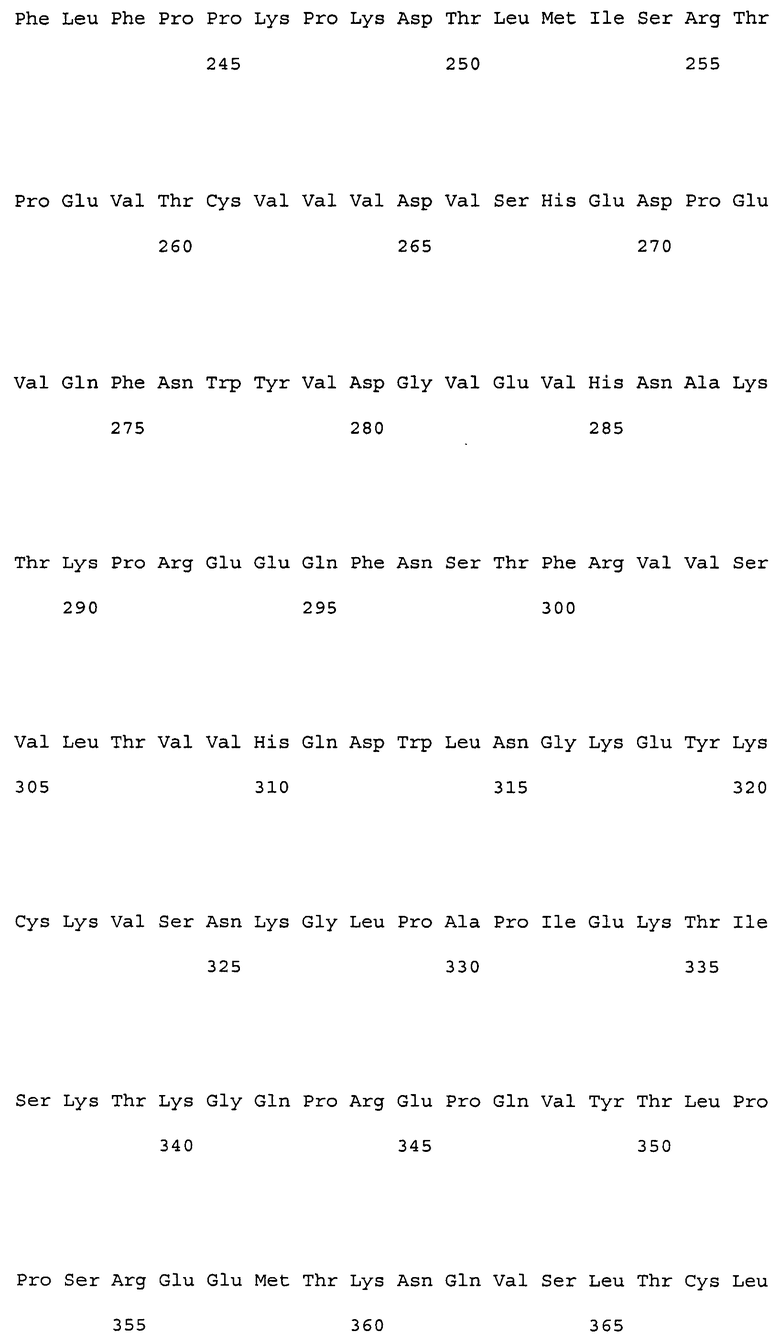















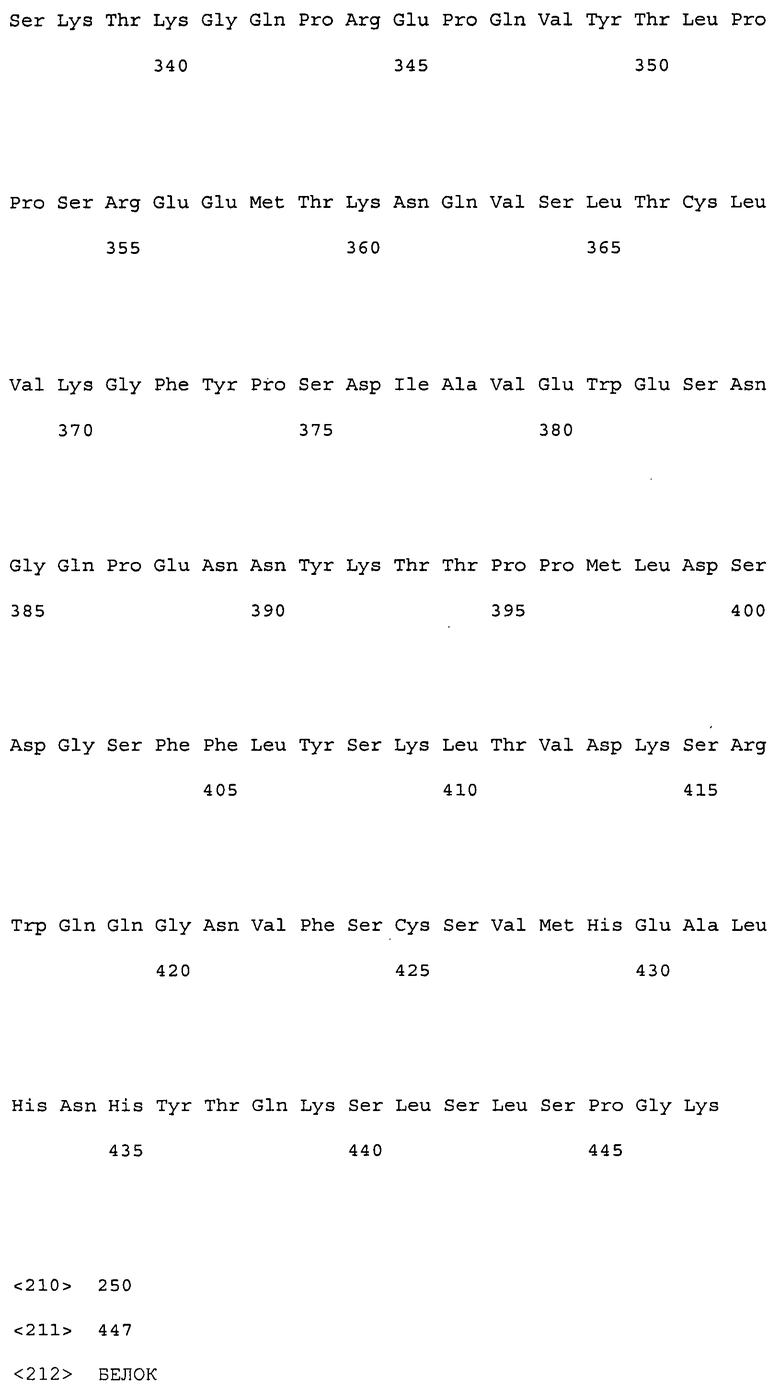
































































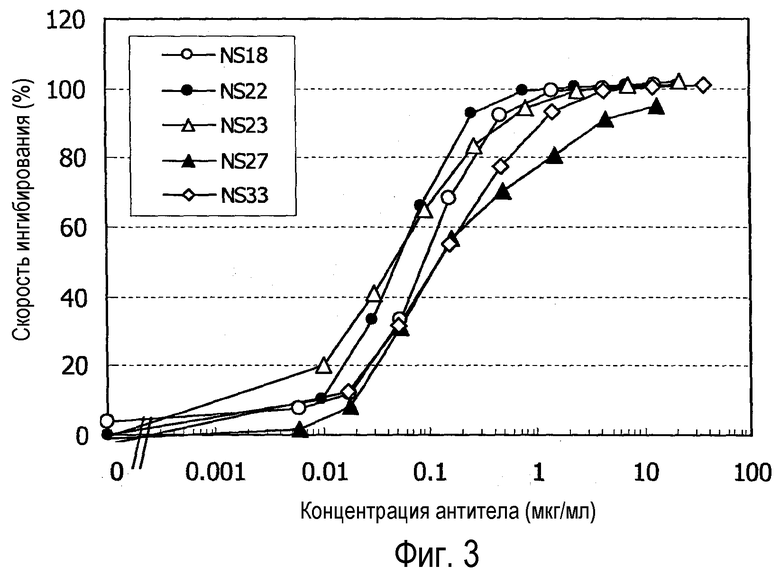
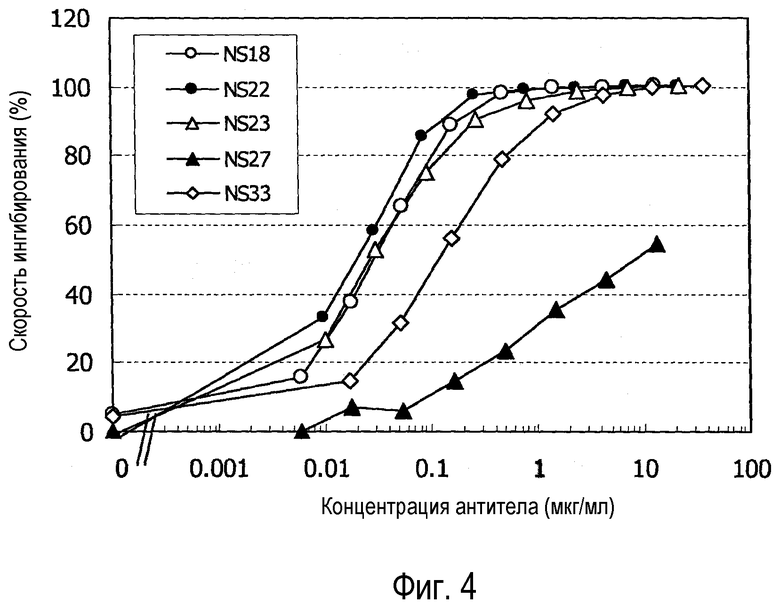
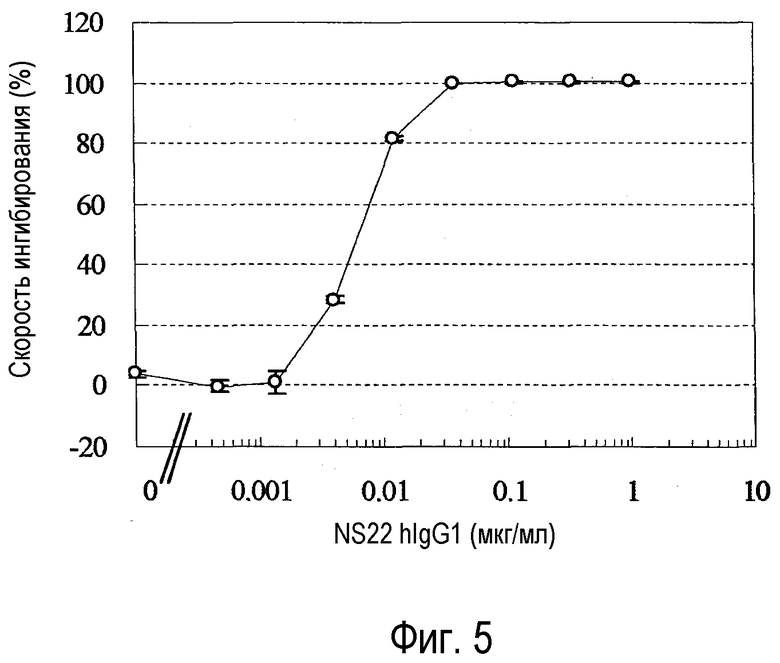
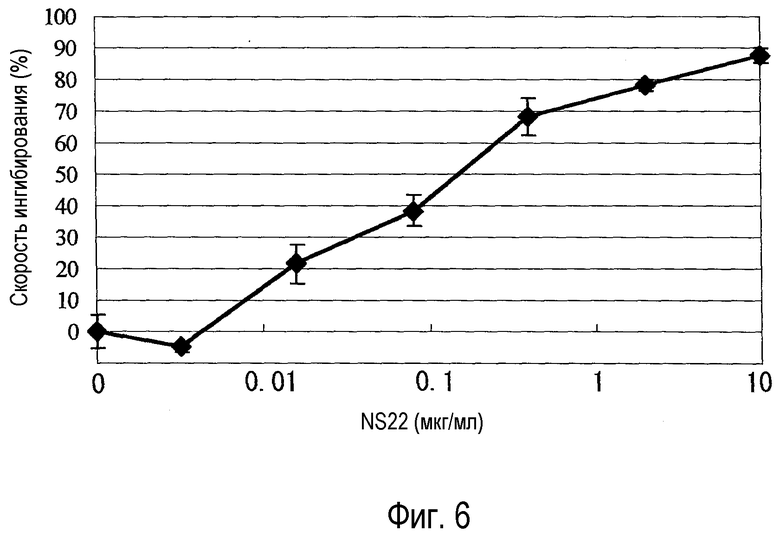
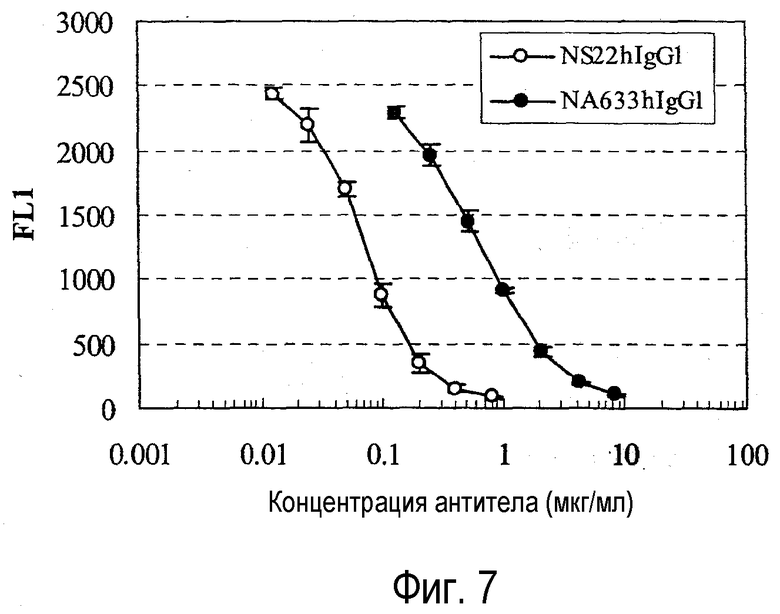
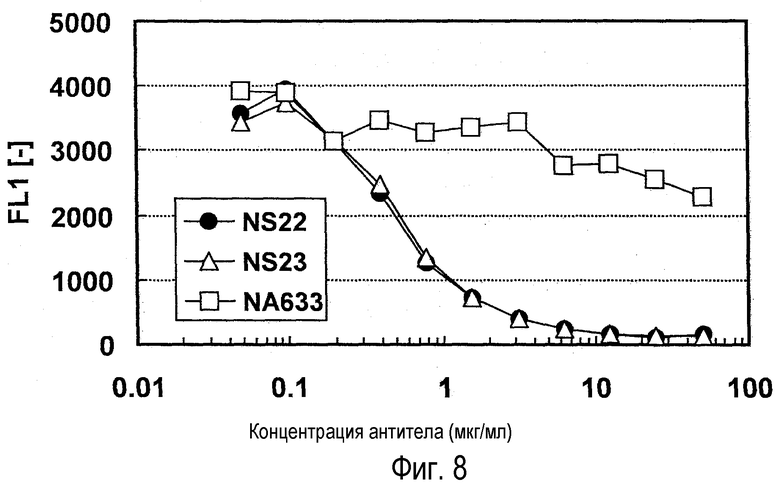
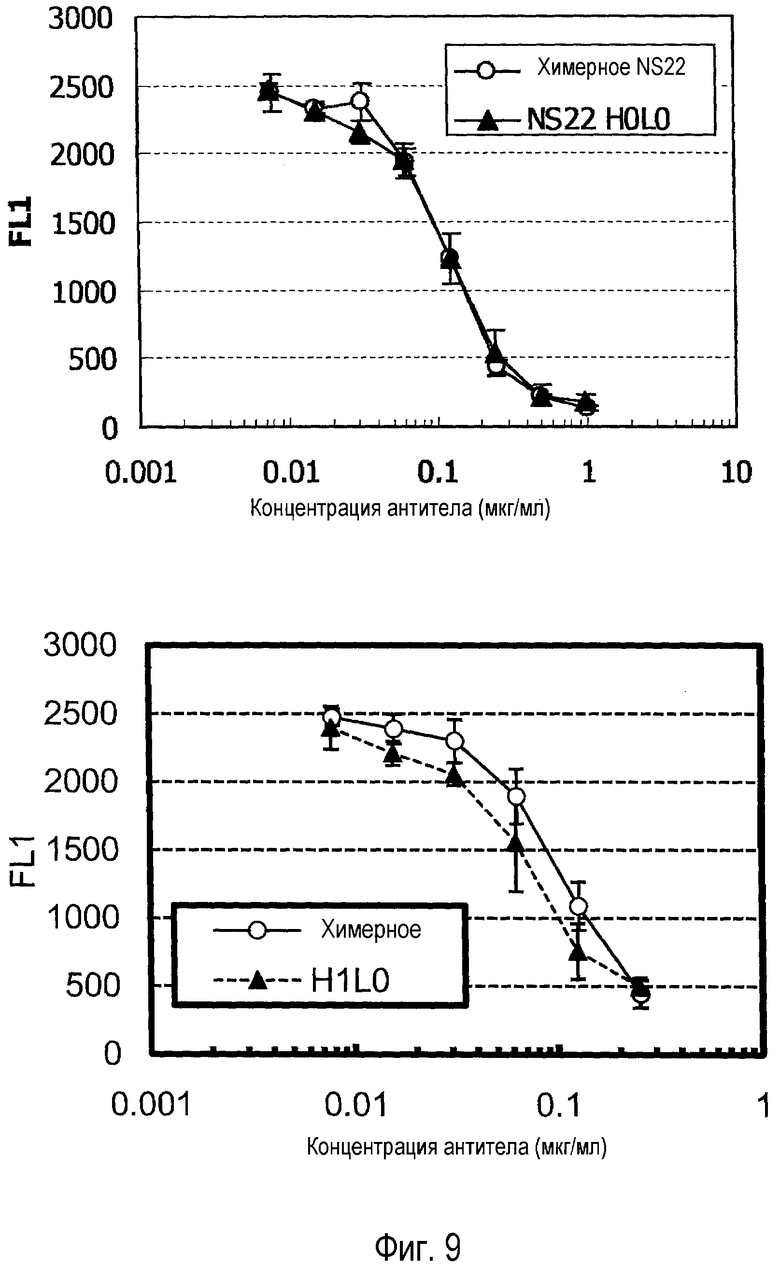
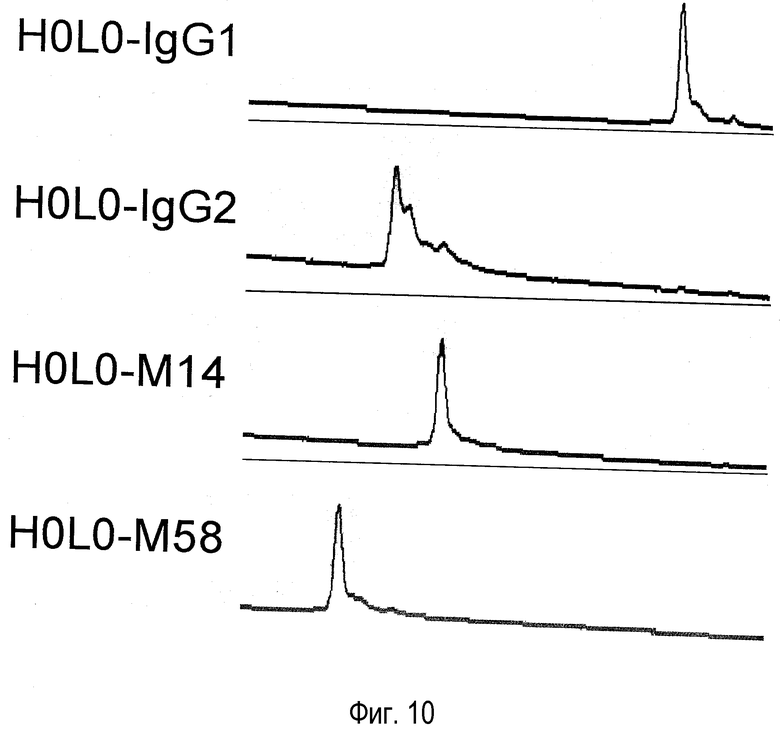
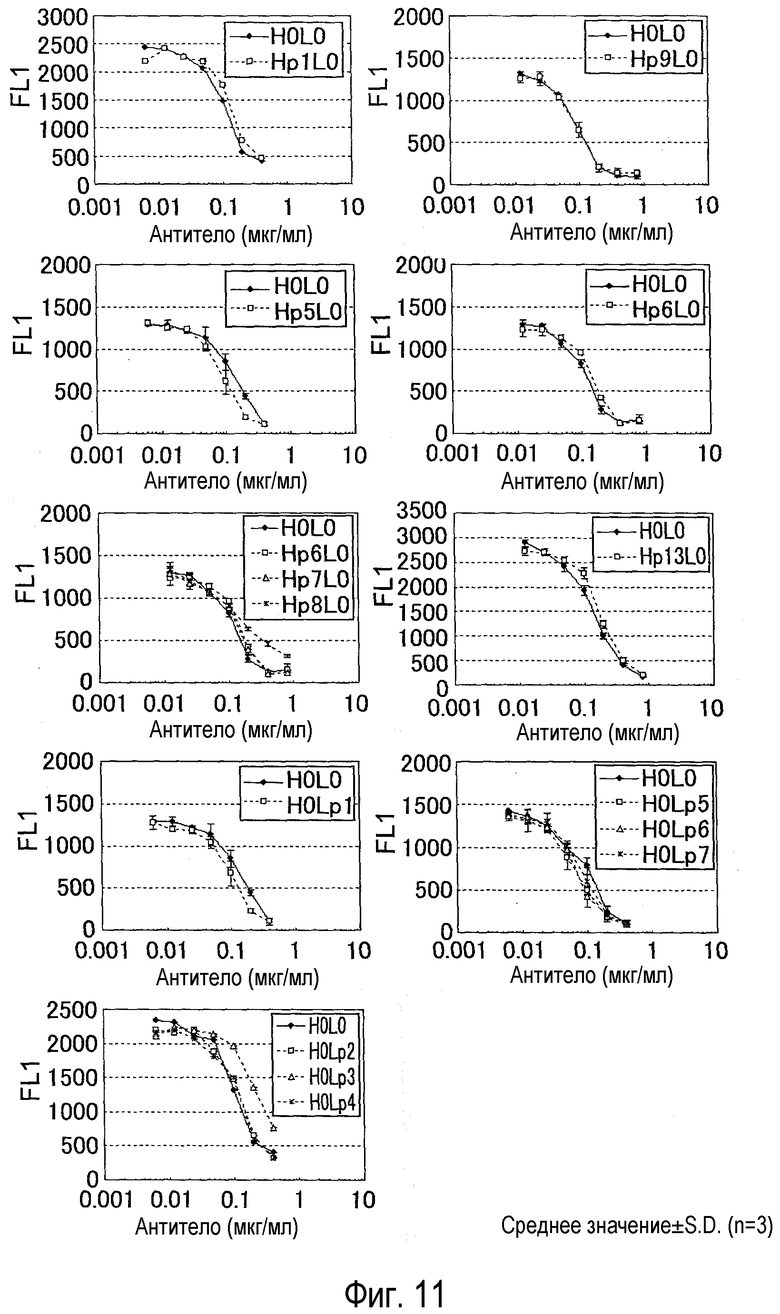
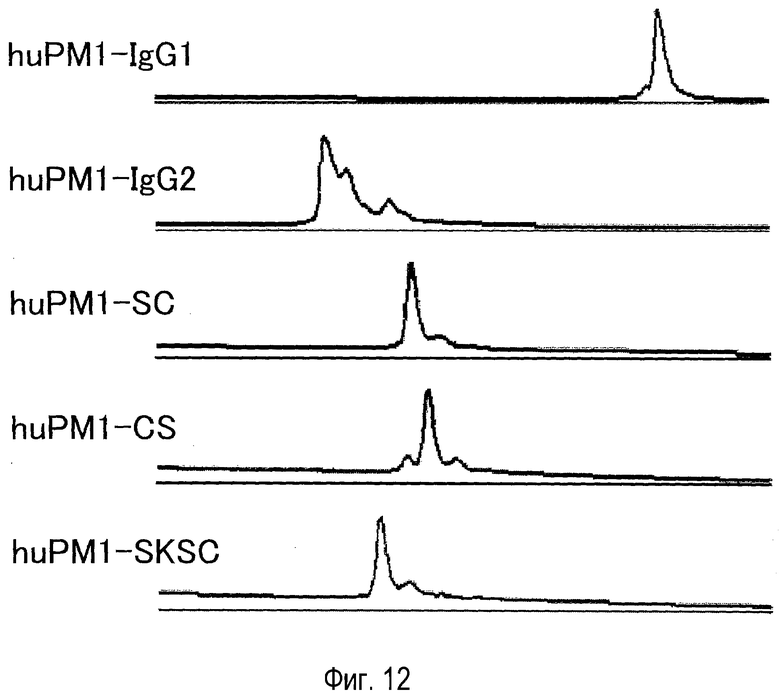
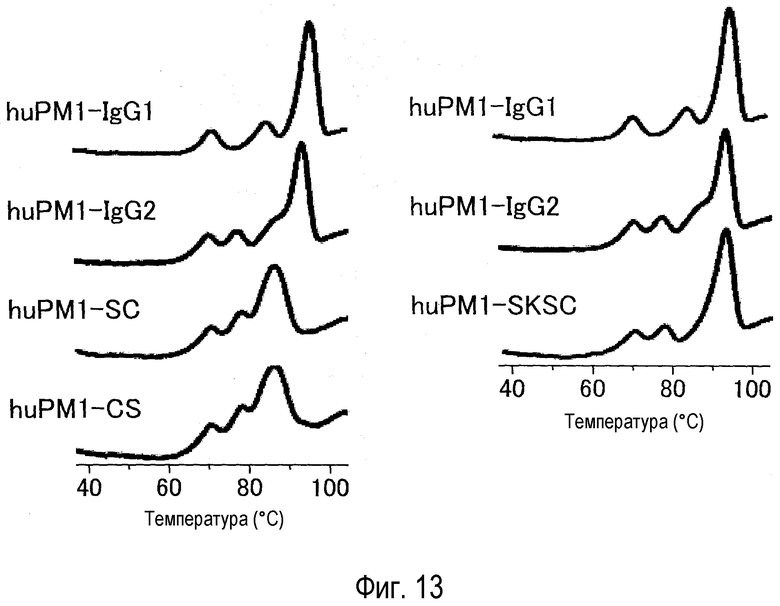
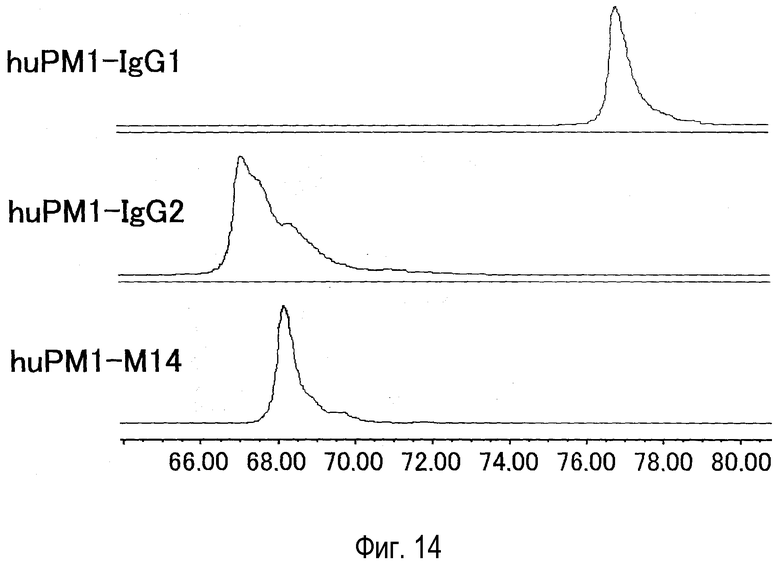
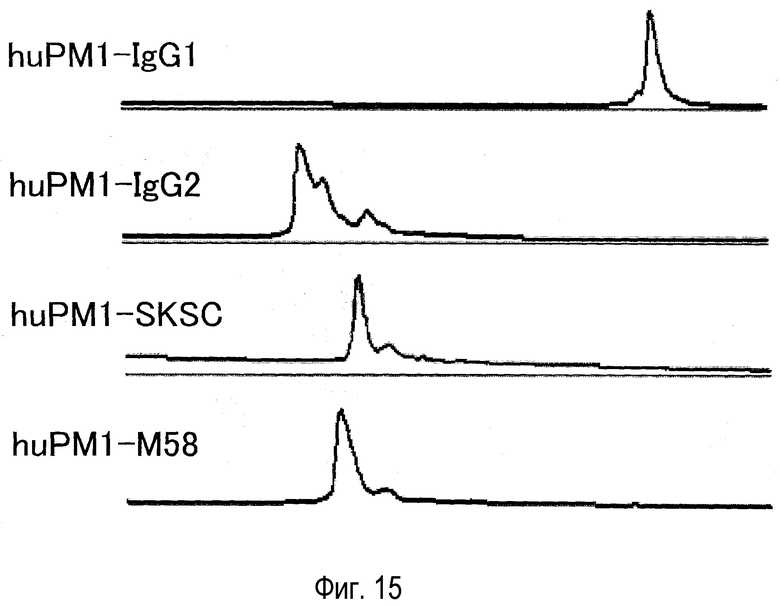
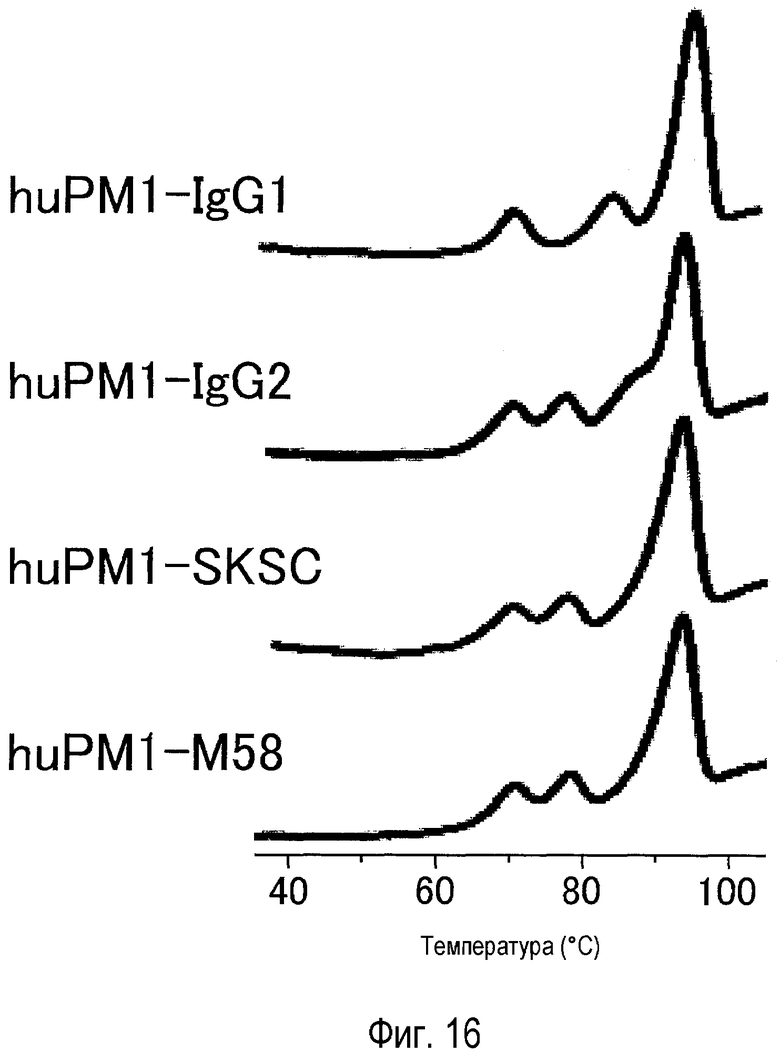
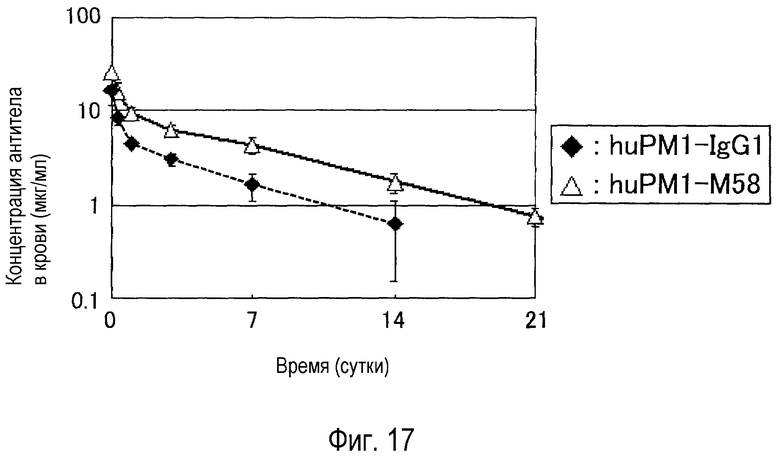
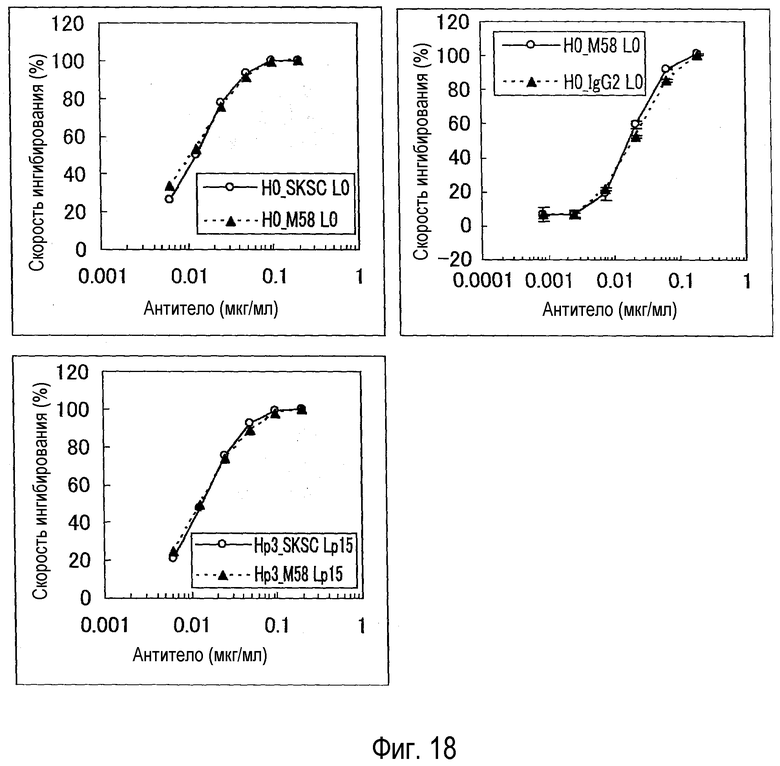
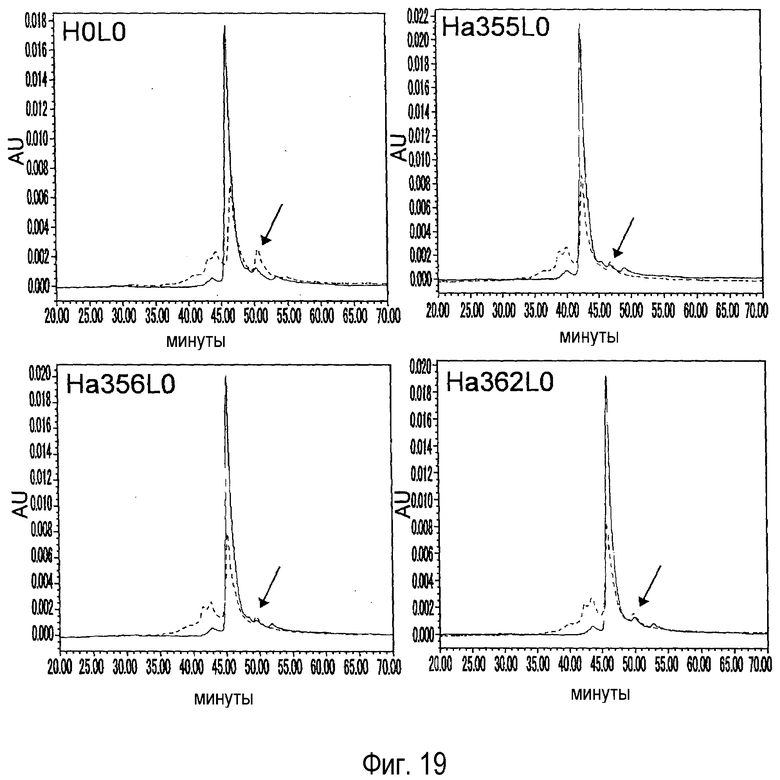
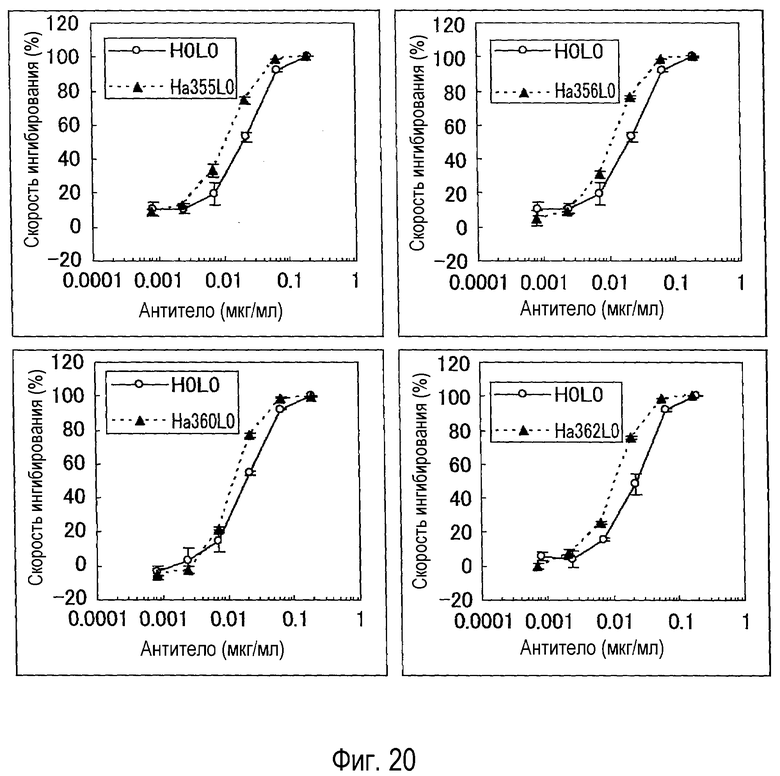
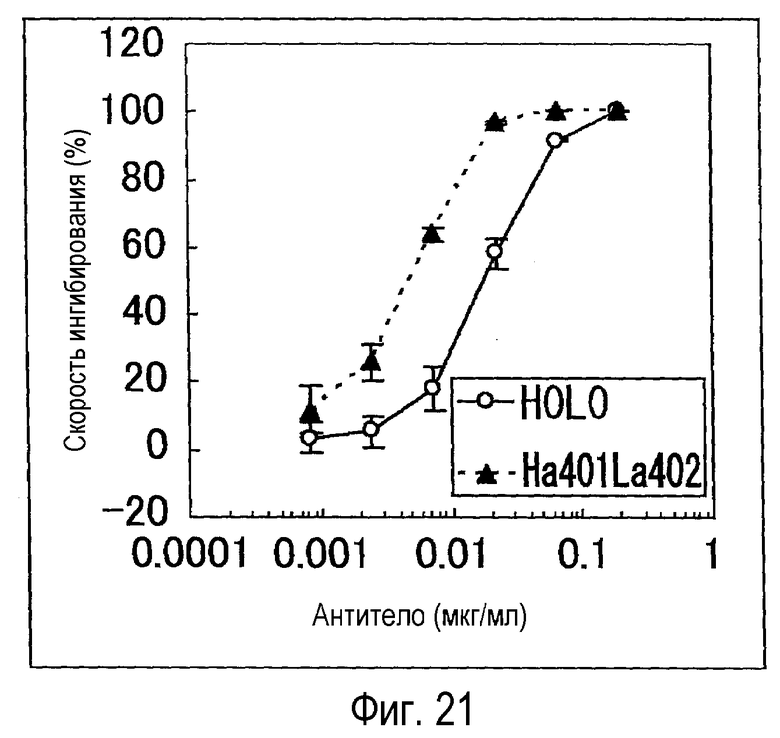
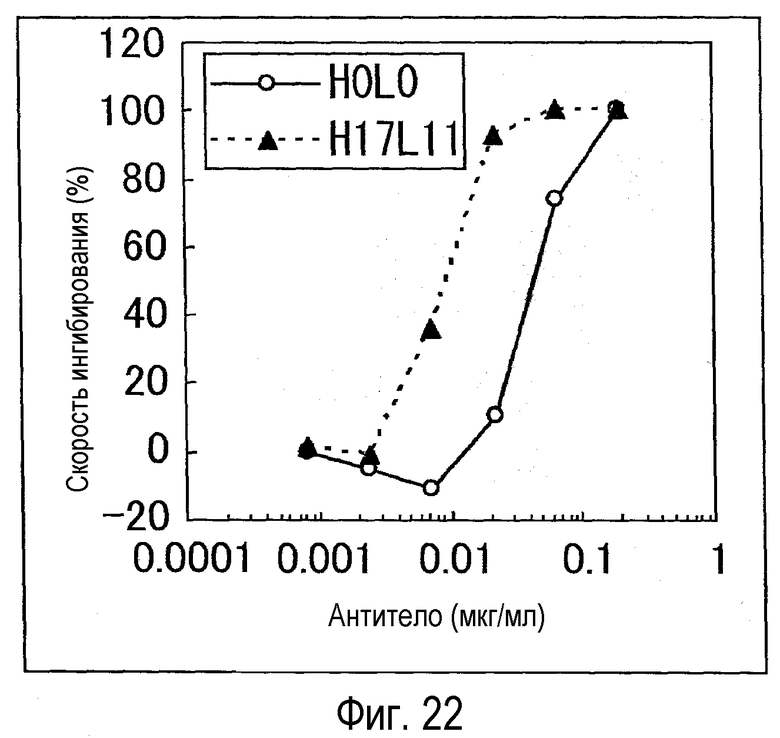

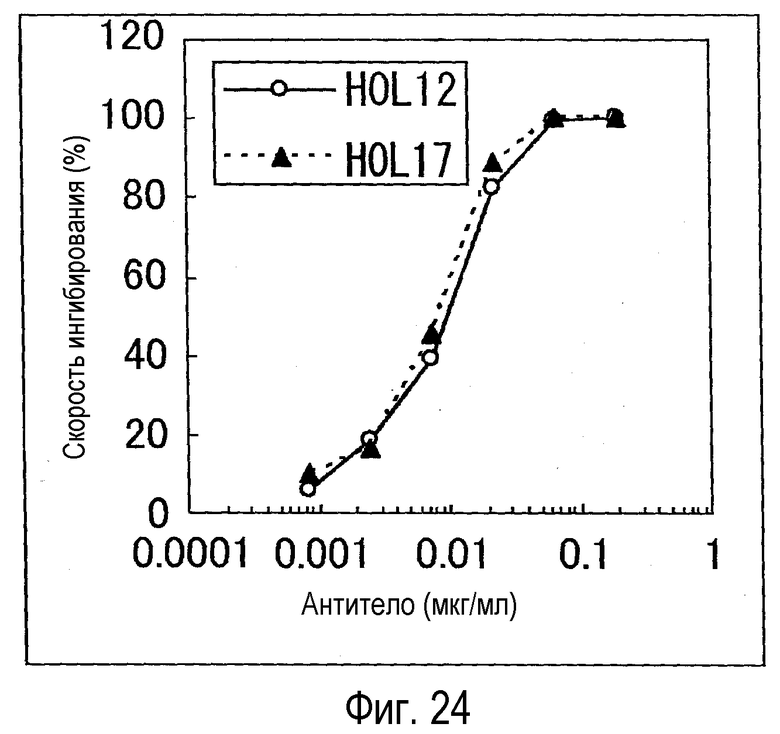
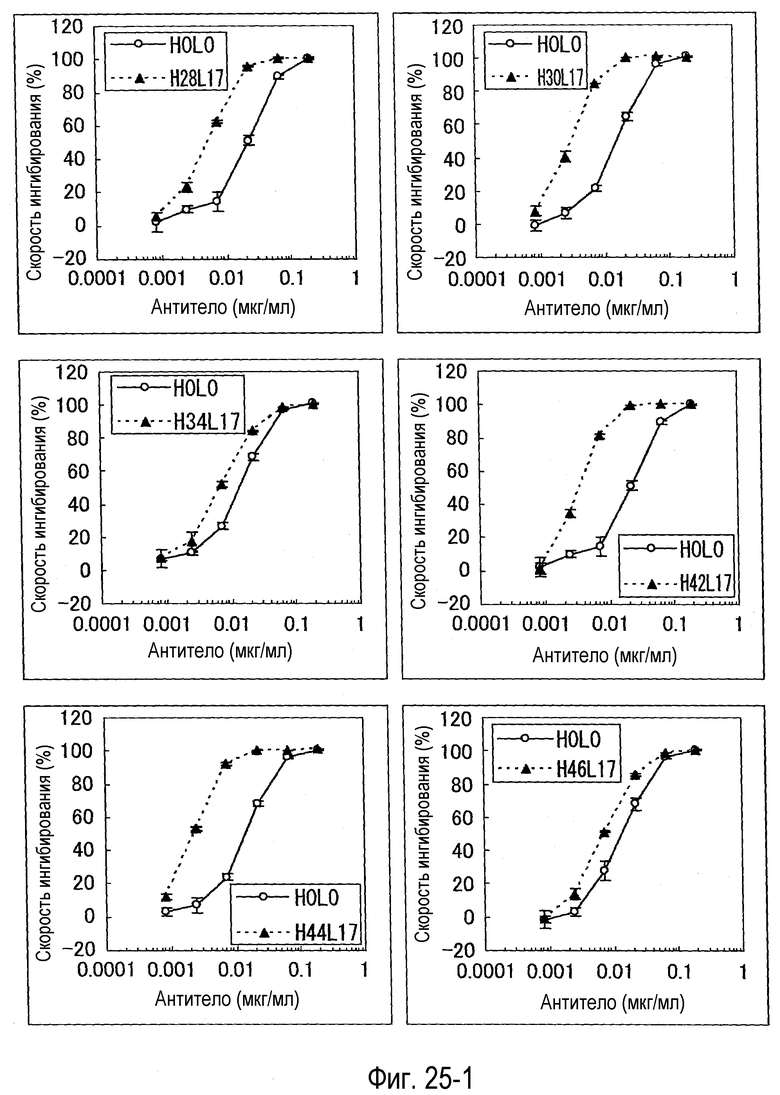
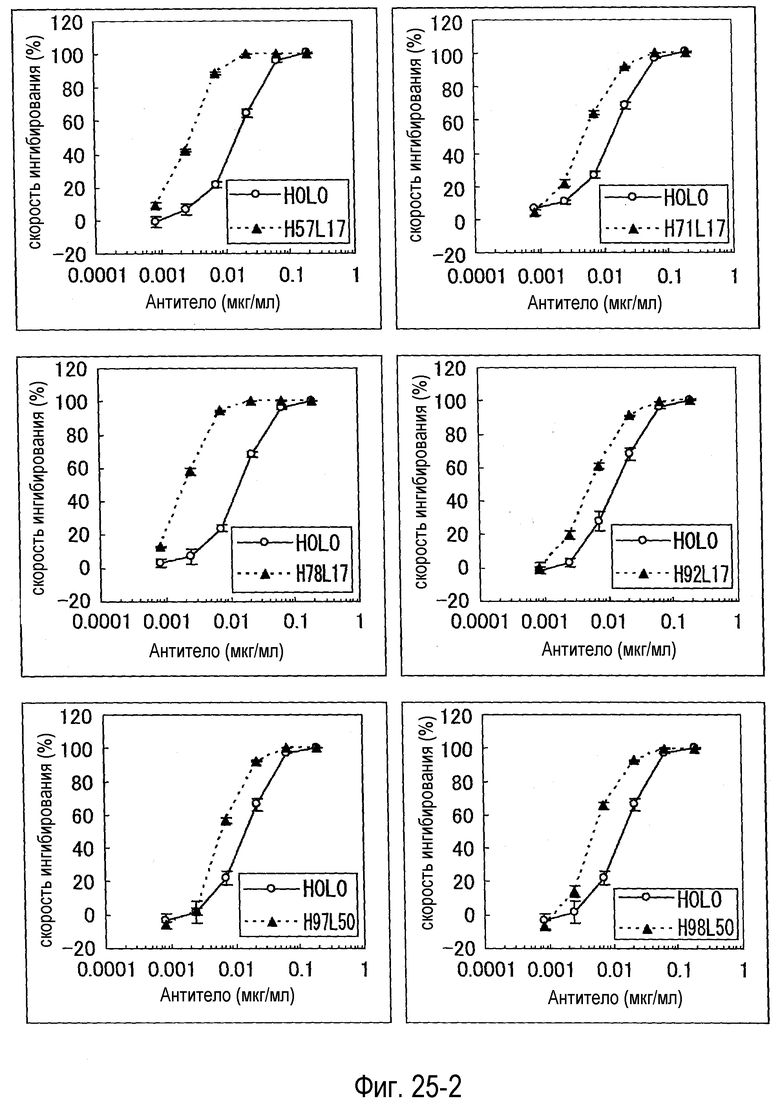

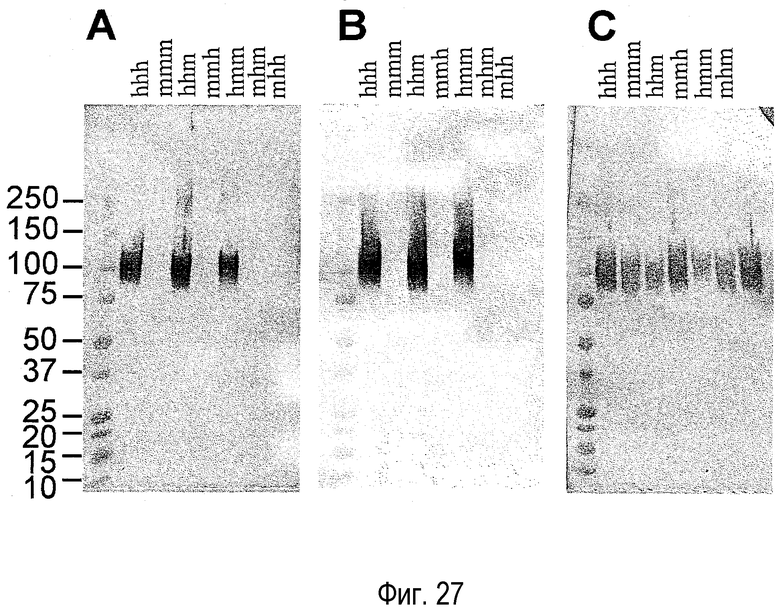
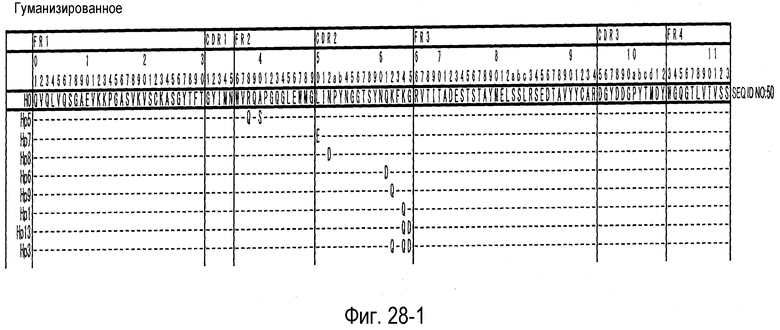
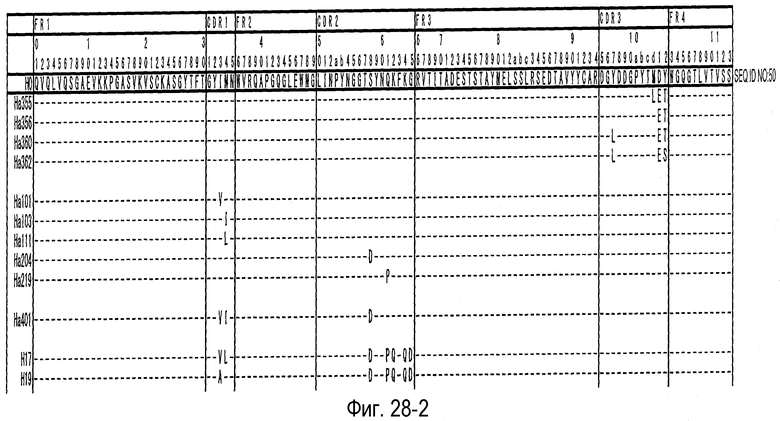
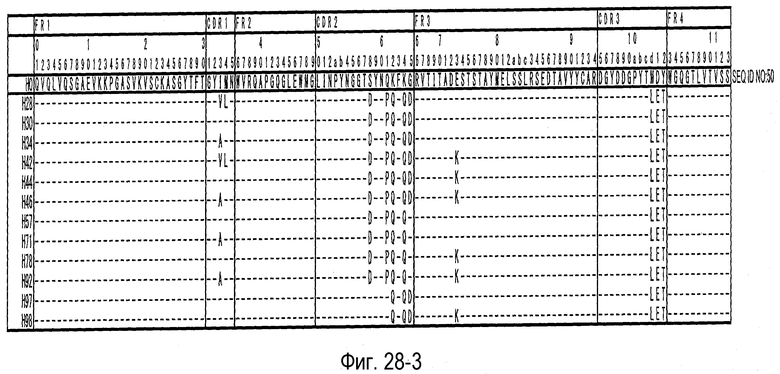
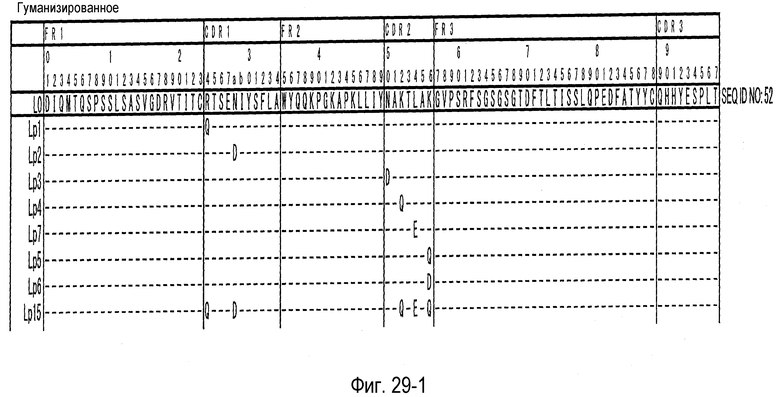

Изобретение относится к области биотехнологии, конкретно к получению антител против NR10, и может быть использовано в медицине. Полученные антитела используют в качестве фармацевтических средств для лечения или профилактики воспалительных заболеваний. Изобретение позволяет получить антитела, обладающие эффективной нейтрализующей активностью против NR10. 6 н. и 1 з.п. ф-лы, 33 ил., 19 табл., 10 пр.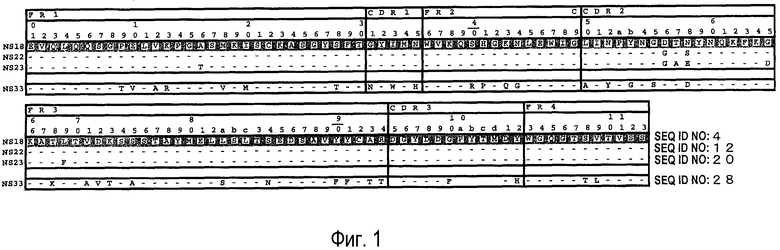
1. Антитело против NR10, которое представляет собой любое из:
(1) антитела, содержащего вариабельную область тяжелой цепи, содержащую CDR1 с SEQ ID NO:196, CDR2 с SEQ ID NO:197 и CDR3 с SEQ ID NO:184 (Н28, Н42), и вариабельную область легкой цепи, содержащую CDR1 с SEQ ID NO:202, CDR2 с SEQ ID NO:170 и CDR3 с SEQ ID NO:193 (L17);
(2) антитела, содержащего вариабельную область тяжелой цепи, содержащую CDR1 с SEQ ID NO:9, CDR2 с SEQ ID NO:197 и CDR3 с SEQ ID NO:184 (Н30, Н44), и вариабельную область легкой цепи, содержащую CDR1 с SEQ ID NO:202, CDR2 с SEQ ID NO:170 и CDR3 с SEQ ID NO:193 (L17);
(3) антитела, содержащего вариабельную область тяжелой цепи, содержащую CDR1 с SEQ ID NO:176, CDR2 с SEQ ID NO:197 и CDR3 с SEQ ID NO: 184 (Н34, Н46), и вариабельную область легкой цепи, содержащую CDR1 с SEQ ID NO:202, CDR2 с SEQ ID NO:170 и CDR3 с SEQ ID NO:193 (L17);
(4) антитела, содержащего вариабельную область тяжелой цепи, содержащую CDR1 с SEQ ID NO:9, CDR2 с SEQ ID NO:198 и CDR3 с SEQ ID NO:184 (Н57, Н78), и вариабельную область легкой цепи, содержащую CDR1 с SEQ ID NO:202, CDR2 с SEQ ID NO:170 и CDR3 с SEQ ID NO:193 (L17);
(5) антитела, содержащего вариабельную область тяжелой цепи, содержащую CDR1 с SEQ ID NO:176, CDR2 с SEQ ID NO:198 и CDR3 с SEQ ID NO:184 (Н71, Н92), и вариабельную область легкой цепи, содержащую CDR1 с SEQ ID NO:202, CDR2 с SEQ ID NO:170 и CDR3 с SEQ ID NO:193 (L17);
(6) антитела, содержащего вариабельную область тяжелой цепи, содержащую CDR1 с SEQ ID NO:9, CDR2 с SEQ ID NO:199 и CDR3 с SEQ ID NO:184 (Н97, Н98), и вариабельную область легкой цепи, содержащую CDR1 с SEQ ID NO:203, CDR2 с SEQ ID NO:170 и CDR3 с SEQ ID NO:193 (L50).
2. Антитело против NR10, которое представляет собой любое из:
(1) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:206 (Н28), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(2) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:207 (Н30), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(3) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:208 (Н34), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(4) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:209 (Н42), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(5) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:210 (Н44), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(6) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:211 (Н46), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(7) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:212 (Н57), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(8) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:213 (Н71), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(9) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:214 (Н78), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(10) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:215 (Н92), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:220 (L17);
(11) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:216 (Н97), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:221 (L50);
(12) антитела, содержащего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:217 (Н98), и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:221 (L50).
3. Антитело против NR10 по п.1 или 2, которое представляет собой гуманизированное антитело против NR10.
4. Антитело против NR10, которое представляет собой любое из:
(1) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:224 (Н28), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(2) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:225 (Н30), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(3) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:226 (Н34), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(4) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:227 (Н42), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(5) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:228 (Н44), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(6) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:229 (Н46), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(7) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:230 (Н57), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(8) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:231 (Н71), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(9) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:232 (Н78), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(10) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:233 (Н92), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:238 (L17);
(11) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:234 (Н97), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:239 (L50);
(12) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:235 (Н98), и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:239 (L50).
5. Фармацевтическая композиция для лечения воспалительного заболевания, которая содержит эффективное количество антитела по любому из пп.1-4 в качестве активного ингредиента и фармацевтически приемлемые добавки.
6. Способ лечения или профилактики воспалительного заболевания, который включает стадию введения антитела по любому из пп.1-4.
7. Применение антитела по любому из пп.1-4 для получения терапевтического средства от воспалительного заболевания.
| DIVEU С.ЕТ AL., GPL, a Novel Cytokine Receptor Related to GP130 and Leukemia Inhibitory Factor Receptor, J | |||
| BIOL | |||
| CHEM., 2003,v.278, n.50, p.49850-49859 | |||
| WO 2007142325 A1, 13.12.2007 | |||
| WO 2006088855 A1, 24.08.2006 | |||
| US 2008219971 A1, 11.09.2008 | |||
| WO 9710354 A1, 20.03.1997 | |||
| WO 9412215 A1, 09.06.1994 | |||
| WO 9623071 A2, 01.08.1996 | |||
| WO 2006119062 A2, |

