ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение в целом относится к выделенному моноклональному антителу, в частности мышиному, гибридному или гуманизированному моноклональному антителу, которое специфично связывается с PD-1 человека с высокой аффинностью и функциональной активностью. Также предложена кодирующая указанное антитело молекула нуклеиновой кислоты, вектор экспрессии, клетка-хозяин и способ экспрессии указанного антитела. Настоящее изобретение дополнительно обеспечивает иммуноконъюгат, биспецифичную молекулу, онколитический вирус и фармацевтическую композицию, содержащую антитело, а также способ диагностики и лечения с использованием антитела против PD-1 согласно изобретению.
УРОВЕНЬ ТЕХНИКИ
[0002] Белок программируемой клеточной гибели 1, также известный как PD-1 или CD279, представляет собой член семейства T-клеточных регуляторов CD28 и экспрессируется на активированных B-клетках, T-клетках и миелоидных клетках (Agata et al., (1996) Int Immunol 8:765-72; Okazaki et al., (2002) Curr. Opin. Immunol. 14: 391779-82; Bennett et al., (2003) J Immunol 170:711-8). Он содержит расположенный близко к мембране иммунорецепторный тирозиновый ингибиторный мотив (ITIM) и удаленный от мембраны тирозиновый переключающий мотив (ITSM) (Thomas, M. L. (1995) J Exp Med 181:1953-6; Vivier, E and Daeron, M (1997) Immunol Today 18:286-91). Были идентифицированы два лиганда PD-1 - PD-L1 и PD-L2, - оба из которых являются гомологами B7, которые связываются с PD-1, но не связываются с другими членами семейства CD28.
[0003] Некоторые данные дают основание полагать, что PD-1 и его лиганды обеспечивают отрицательную регуляцию иммунных ответов. Например, было обнаружено, что PD-1 присутствует в большом количестве в различных раковых опухолях человека (Dong et al., (2002) Nat. Med. 8:787-9). Также сообщалось, что взаимодействие между PD-1 и PD-L1 приводит к снижению уровня инфильтрирующих опухоль лимфоцитов и опосредованной T-клеточным рецептором пролиферации, а также ускользанию раковых клеток от иммунологического надзора (Dong et al., (2003) J. Mol. Med. 81:281-7; Blank et al., (2005) Cancer Immunol. Immunother. 54:307-314; Konishi et al., (2004) Clin. Cancer Res. 10:5094-100). Исследования также показали, что ингибирование локального взаимодействия PD-1 с PD-L1 может устранять подавление иммунитета, и указанный эффект усиливается при блокировании взаимодействия также между PD-1 и PD-L2 (Iwai et al., (2002) Proc. Nat'l. Acad. Sci. USA 99:12293-7; Brown et al., (2003) J. Immunol. 170:1257-66).
[0004] У животных с недостаточностью PD-1 могут развиваться различные аутоиммунные фенотипы, включая аутоиммунную кардиомиопатию и волчаночный синдром с артритом и нефритом (Nishimura et al., (1999) Immunity 11:141-51; Nishimura et al., (2001) Science 291:319-22). Кроме того, было обнаружено, что PD-1 принимает участие в аутоиммунном энцефаломиелите, системной красной волчанке, реакции «трансплантат против хозяина» (РТПХ), диабете I типа и ревматоидном артрите (Salama et al., (2003) J Exp Med 198:71-78; Prokunina and Alarcon-Riquelme (2004) Hum Mol Genet 13:R143; Nielsen et al., (2004) Lupus 13:510). На мышиной опухолевой линии B-клеток было показано, что ITSM-мотив PD-1 является важным для блокирования BCR-опосредованного Ca2+-тока и тирозинового фосфорилирования нижележащих эффекторных молекул (Okazaki et al., (2001) PNAS 98:13866-71).
[0005] Ряд агентов для противораковой иммунотерапии, которые нацелены на PD-1, были разработаны для лечения заболевания. Одно такое антитело против PD-1 представляет собой ниволумаб (продается под торговым названием OPDIVO® компанией Bristol Myers Squibb), который вызывал полные или частичные ответы в случае немелкоклеточного рака легких, меланомы и почечно-клеточного рака в клиническом исследовании, включающем в общем 296 пациентов (Topalian SL et al., (2012) The New England Journal of Medicine. 366 (26): 2443-54). Он был одобрен в Японии в 2014 и Управлением по контролю качества пищевых продуктов и лекарственных средств США (FDA) в 2014 для лечения метастатической меланомы. Другое антитело против PD-1, пемролизумаб (KEYTRUDA™, MK-3475, Merck), направленное на PD-1, было также одобрено Управлением по контролю качества пищевых продуктов и лекарственных средств США (FDA) в 2014 для лечения метастатической меланомы. Он используется в клинических исследованиях в США для лечения рака легких, лимфомы и мезотелиомы.
[0006] Несмотря на наличие уже разработанных и одобренных антител против PD-1, существует необходимость в дополнительных моноклональных антителах, обладающих повышенной аффинностью связывания с PD-1 и другими желаемыми фармацевтическими характеристиками.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0007] Настоящее изобретение обеспечивает выделенное моноклональное антитело, например, мышиное, человеческое, гибридное или гуманизированное моноклональное антитело, которое связывается с PD-1 (например, PD-1 человека и PD-1 обезьяны) и обладает повышенной аффинностью в отношении PD-1 и сравнимым или лучшим противоопухолевым эффектом по сравнению с существующими антителами против PD-1, такими как ниволумаб.
[0008] Антитело согласно изобретению можно использовать в различных способах применения, включая выявление белка PD-1 и лечение и предотвращение связанных с PD-1 заболеваний, таких как раковые заболевания, аутоиммунная кардиомиопатия, аутоиммунный энцефаломиелит, системная красная волчанка, реакция «трансплантат против хозяина» (РТПХ), диабет I типа и ревматоидный артрит.
[0009] Соответственно, согласно одному аспекту, изобретение относится к выделенному моноклональному антителу (например, мышиному, гибридному или гуманизированному антителу) или его антиген-связывающей части, которая связывается с PD-1 и содержит вариабельный участок тяжелой цепи, содержащий участок CDR1, участок CDR2 и участок CDR3, где указанные участки CDR1, CDR2 и CDR3 содержат аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям (1) SEQ ID NO: 1, 2 и 3 соответственно; (2) SEQ ID NO: 4, 5 и 6 соответственно; (3) SEQ ID NO: 7, 8 и 9 соответственно; (4) SEQ ID NO: 10, 11 и 12 соответственно; (5) SEQ ID NO: 13, 14 и 15 соответственно; (6) SEQ ID NO: 16, 17 и 18 соответственно; (7) SEQ ID NO: 19, 20 и 21 соответственно; (8) SEQ ID NO: 22, 23 и 24 соответственно; (9) SEQ ID NO: 25, 26 и 27 соответственно; (10) SEQ ID NO: 28, 29 и 30 соответственно или (11) SEQ ID NO: 31, 32 и 33 соответственно, где указанное антитело или его антиген-связывающий фрагмент связывается с PD-1.
[0010] Согласно одному аспекту, выделенное моноклональное антитело или его антиген-связывающая часть согласно настоящему изобретению содержит вариабельный участок тяжелой цепи, содержащий аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичную последовательности SEQ ID NO: 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78 или 79, где указанное антитело или его антиген-связывающий фрагмент связывается с PD-1. Последовательности SEQ ID NO: 67 и 69-79 могут кодироваться последовательностями нуклеиновых кислот SEQ ID NO: 102-113 соответственно.
[0011] Согласно одному аспекту, выделенное моноклональное антитело или его антиген-связывающая часть согласно настоящему изобретению содержит вариабельный участок легкой цепи, который содержит участок CDR1, участок CDR2 и участок CDR3, где указанные участки CDR1, CDR2 и CDR3 содержат аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям (1) SEQ ID NO: 34, 35 и 36 соответственно; (2) SEQ ID NO: 37, 38 и 39 соответственно; (3) SEQ ID NO: 40, 41 и 42 соответственно; (4) SEQ ID NO: 43, 44 и 45 соответственно; (5) SEQ ID NO: 46, 47 и 48 соответственно; (6) SEQ ID NO: 49, 50 и 51 соответственно; (7) SEQ ID NO: 52, 53 и 54 соответственно; (8) SEQ ID NO: 55, 56 и 57 соответственно; (9) SEQ ID NO: 58, 59 и 60 соответственно; (10) SEQ ID NO: 61, 62 и 63 соответственно или (11) SEQ ID NO: 64, 65 и 66 соответственно, где указанное антитело или его антиген-связывающий фрагмент связывается с PD-1.
[0012] Согласно одному аспекту, выделенное моноклональное антитело или его антиген-связывающая часть согласно настоящему изобретению содержит вариабельный участок легкой цепи, содержащий аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичную последовательности SEQ ID NO: 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95 или 96, где указанное антитело или его антиген-связывающий фрагмент связывается с PD-1. SEQ ID NO: 80 и 86-96 могут кодироваться последовательностями нуклеиновых кислот SEQ ID NO: 114-125 соответственно.
[0013] Согласно одному аспекту, выделенное моноклональное антитело или его антиген-связывающая часть согласно настоящему изобретению содержит вариабельный участок тяжелой цепи и вариабельный участок легкой цепи, каждый из которых содержит участок CDR1, участок CDR2 и участок CDR3, где вариабельные участки тяжелой цепи CDR1, CDR2 и CDR3 и вариабельные участки легкой цепи CDR1, CDR2 и CDR3 содержат аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям (1) SEQ ID NO: 1, 2, 3, 34, 35 и 36 соответственно; (2) SEQ ID NO: 4, 5, 6, 37, 38 и 39 соответственно; (3) SEQ ID NO: 7, 8, 9, 40, 41 и 42 соответственно; (4) SEQ ID NO: 10, 11, 12, 43, 44 и 45 соответственно; (5) SEQ ID NO: 13, 14, 15, 46, 47 и 48 соответственно; (6) SEQ ID NO: 16, 17, 18, 49, 50 и 51 соответственно; (7) SEQ ID NO: 19, 20, 21, 52, 53 и 54 соответственно; (8) SEQ ID NO: 22, 23, 24, 55, 56 и 57 соответственно; (9) SEQ ID NO: 25, 26, 27, 58, 59 и 60 соответственно; (10) SEQ ID NO: 28, 29, 30, 61, 62 и 63 соответственно или (11) SEQ ID NO: 31, 32, 32, 64, 65 и 66 соответственно, где антитело или его антиген-связывающий фрагмент связывается с PD-1.
[0014] Согласно одному варианту реализации изобретения, выделенное моноклональное антитело или его антиген-связывающая часть согласно настоящему изобретению содержит вариабельный участок тяжелой цепи и вариабельный участок легкой цепи, где указанные вариабельный участок тяжелой цепи и вариабельный участок легкой цепи содержат аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям (1) SEQ ID NO: 67 и 80 соответственно; (2) SEQ ID NO: 68 и 81 соответственно; (3) SEQ ID NO: 69 и 81 соответственно; (4) SEQ ID NO: 68 и 82 соответственно; (5) SEQ ID NO: 68 и 83 соответственно; (6) SEQ ID NO: 68 и 84 соответственно; (7) SEQ ID NO: 68 и 85 соответственно; (8) SEQ ID NO: 68 и 86 соответственно; (9) SEQ ID NO: 69 и 86 соответственно; (10) SEQ ID NO: 70 и 87 соответственно; (11) SEQ ID NO: 71 и 88 соответственно; (12) SEQ ID NO: 72 и 89 соответственно; (13) SEQ ID NO: 73 и 90 соответственно; (14) SEQ ID NO: 74 и 91 соответственно; (15) SEQ ID NO: 75 и 92 соответственно; (16) SEQ ID NO: 76 и 93 соответственно; (17) SEQ ID NO: 77 и 94 соответственно; (18) SEQ ID NO: 78 и 95 соответственно; или (19) SEQ ID NO: 79 и 96 соответственно, где антитело или его антиген-связывающий фрагмент связывается с PD-1.
[0015] Согласно одному варианту реализации изобретения, выделенное моноклональное антитело или его антиген-связывающая часть согласно настоящему изобретению содержит тяжелую цепь и легкую цепь, где указанная тяжелая цепь содержит вариабельный участок тяжелой цепи и константный участок тяжелой цепи, а указанная легкая цепь содержит вариабельный участок легкой цепи и константный участок легкой цепи, где указанный константный участок тяжелой цепи содержит аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям SEQ ID No: 97, 99 или 129, и указанный константный участок легкой цепи содержит аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям SEQ ID No: 98 или 130, и указанные вариабельный участок тяжелой цепи и вариабельный участок легкой цепи содержат аминокислотные последовательности, описанные выше, где указанное антитело или его антиген-связывающий фрагмент связывается с PD-1. Указанные аминокислотные последовательности SEQ ID NO: 97 и 99 могут кодироваться последовательностями нуклеиновых кислот SEQ ID NO: 126 и 128 соответственно. SEQ ID NO:98 может кодироваться последовательностью SEQ ID NO: 127.
[0016] Антитело согласно настоящему изобретению в соответствии с некоторыми вариантами реализации настоящего изобретения содержит или состоит из двух тяжелых цепей и двух легких цепей, где каждая указанная тяжелая цепь содержит константный участок тяжелой цепи, вариабельный участок или CDR-последовательности тяжелой цепи, перечисленные выше, и каждая легкая цепь содержит константный участок легкой цепи, вариабельный участок или CDR-последовательности легкой цепи, перечисленные выше, где указанное антитело связывается с PD-1. Антитело согласно изобретению может представлять собой полноразмерное антитело, например, изотип IgG1, IgG2 или IgG4. Антитело согласно настоящему изобретению в соответствии с другими вариантами реализации изобретения может представлять собой одноцепочечное антитело или фрагменты антитела, такие как Fab- или Fab′2-фрагменты.
[0017] Антитело или его антиген-связывающая часть согласно настоящему изобретению обладает повышенной аффинностью связывания с PD-1 человека по сравнению с антителами против PD-1 предшествующего уровня техники, такими как ниволумаб, связывающийся с PD-1 человека с KD, составляющей 6,36×10-9M или менее, и ингибирующим связывание PD-L1 с PD-1. Антитело или его антиген-связывающая часть согласно изобретению также обеспечивает сравнимый или лучший противоопухолевый эффект по сравнению с существующими антителами против PD-1, такими как ниволумаб.
[0018] Изобретение также обеспечивает иммуноконъюгат, содержащий антитело согласно изобретению или его антиген-связывающую часть, связанную с терапевтическим агентом, таким как цитотоксин. Изобретение также обеспечивает биспецифичную молекулу, содержащую антитело или его антиген-связывающую часть согласно изобретению, связанную со вторым функциональным фрагментом (например, вторым антителом), обладающим специфичностью связывания, отличной от специфичности указанного антитела или его антиген-связывающей части. Согласно другому аспекту, антитело или его антиген-связывающие части согласно настоящему изобретению могут быть созданы как часть гибридного антигенного рецептора (chimeric antigen receptor, CAR). Антитело или его антиген-связывающие части согласно настоящему изобретению также могут кодироваться онколитическим вирусом или использоваться совместно с онколитическим вирусом.
[0019] Также предложены композиции, содержащие антитело или его антиген-связывающую часть или иммуноконъюгат, биспецифичную молекулу, онколитический вирус или CAR согласно изобретению и фармацевтически приемлемый носитель.
[0020] Изобретение также включает молекулы нуклеиновой кислоты, кодирующие антитела или их антиген-связывающие части согласно изобретению, а также векторы экспрессии, содержащие такие нуклеиновые кислоты, и клетки-хозяева, содержащие указанные векторы экспрессии. Также предложен способ получения антитела против PD-1 с использованием клетки-хозяина, содержащей вектор экспрессии, включающий этапы (i) экспрессии антитела в клетке-хозяине и (ii) выделения указанного антитела из клетки-хозяина или ее клеточной линии.
[0021] Согласно другому аспекту, изобретение обеспечивает способ модулирования иммунного ответа у субъекта, включающий введение указанному субъекту антитела или его антиген-связывающей части согласно изобретению таким образом, что достигается модулирование иммунного ответа у указанного субъекта. Предпочтительно, антитело согласно изобретению усиливает, стимулирует или положительно регулирует иммунный ответ у субъекта. Согласно некоторым вариантам реализации настоящего изобретения, способ включает введение композиции, биспецифичной молекулы, иммуноконъюгата, CAR-T-клетки или кодирующего антитело или содержащего антитело онколитического вируса согласно изобретению или, альтернативно, молекулы нуклеиновой кислоты, способной обеспечивать экспрессию антитела у указанного субъекта.
[0022] Согласно дополнительному аспекту, изобретение обеспечивает способ подавления опухолевого роста у субъекта, включающий введение указанному субъекту терапевтически эффективного количества антитела или его антиген-связывающей части согласно настоящему изобретению. Опухоль может представлять собой солидную или не солидную опухоль, включая, но не ограничиваясь указанными, лимфому, лейкоз, множественную миелому, меланому, аденокарциному толстой кишки, рак поджелудочной железы, рак толстой кишки, желудочно-кишечный рак, рак предстательной железы, рак мочевого пузыря, рак почки, рак яичника, рак шейки матки, рак молочной железы, рак легких, почечно-клеточный рак и рак носоглотки. Согласно некоторым вариантам реализации настоящего изобретения, способ включает введение композиции, биспецифичной молекулы, иммуноконъюгата, CAR-T-клетки или кодирующего антитело или содержащего антитело онколитического вируса согласно изобретению или, альтернативно, молекулы нуклеиновой кислоты, способной обеспечивать экспрессию антитела у указанного субъекта.
[0023] Согласно другому аспекту, изобретение обеспечивает способ лечения инфекционного заболевания у субъекта, включающий введение указанному субъекту терапевтически эффективного количества антитела или его антиген-связывающей части согласно настоящему изобретению. Согласно некоторым вариантам реализации настоящего изобретения, способ включает введение композиции, биспецифичной молекулы, иммуноконъюгата, CAR-T-клетки или кодирующего антитело или содержащего антитело онколитического вируса согласно изобретению или, альтернативно, молекулы нуклеиновой кислоты, способной обеспечивать экспрессию антитела у указанного субъекта.
[0024] Изобретение также обеспечивает способ усиления иммунного ответа на антиген у субъекта, включающий введение указанному субъекту: (i) антигена и (ii) антитела или его антиген-связывающей части таким образом, что происходит усиление иммунного ответа на указанный антиген у субъекта. Антиген может представлять собой, например, опухолевый антиген, вирусный антиген, бактериальный антиген или антиген патогена.
[0025] Антитела согласно изобретению можно использовать в комбинации по меньшей мере с одним дополнительным агентом, таким как иммуностимулирующее антитело (например, антитело против PD-L1 и/или антитело против CTLA-4), цитокин (например, IL-2 и/или IL-21) или костимулирующее антитело (например, антитело против CD137 и/или антитело против GITR).
[0026] Другие признаки и преимущества настоящего изобретения будут очевидны из следующего далее подробного описания и примеров, которые не ограничивают настоящее изобретение. Содержание всех ссылок, входных данных базы Genbank, патентов и опубликованных патентных заявок, цитированных в настоящей заявке, специально включено в настоящую заявку посредством ссылки.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0027] На Фигуре 1 показана кривая плавления гуманизированного антитела против PD-1 huC1E1-V10.
[0028] На Фигуре 2 показан результат эксклюзионной по размеру хроматографии гуманизированного антитела против PD-1 huC1E1-V10.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0029] Для более легкого понимания настоящего изобретения сначала будет приведено описание конкретных терминов. Дополнительные определения изложены в подробном описании.
[0030] Термин «PD-1» относится к белку программированной клеточной гибели 1. Термин «PD-1» включает варианты, изоформы, гомологи, ортологи и паралоги. Например, антитело, специфичное в отношении белка PD-1 человека, может в определенных случаях перекрестно реагировать с белком PD-1 из видов, отличных от человека, таких как обезьяна. Согласно другим вариантам реализации изобретения, антитело, специфичное в отношении белка PD-1 человека, может быть абсолютно специфичным в отношении белка PD-1 человека и не проявлять перекрестной реакционной активности с другими видами или другими типами или может перекрестно реагировать с PD-1 из конкретных других видов, но не всех других видов.
[0031] Термин «PD-1 человека» относится к белку PD-1, имеющему человеческую аминокислотную последовательность, такую как аминокислотная последовательность PD-1 человека, имеющая номер доступа в базе данных Genbank NP_005009,2. Термины «PD-1 обезьяны или макаки-резус» и «PD-1 мыши» относятся к последовательностям PD-1 обезьяны и мыши соответственно, например, последовательностями, имеющими аминокислотные последовательности, имеющие номер доступа в базе данных Genbank NP_001107830 и CAA48113 соответственно.
[0032] Термин «иммунный ответ» относится к действию, например, лимфоцитов, антиген-презентирующих клеток, фагоцитов, гранулоцитов и растворимых макромолекул, продуцируемых указанными выше клетками или печенью (включая антитела, цитокины и компоненты комплемента), которое приводит к селективному повреждению, разрушению или устранению из тела человека инвазирующих патогенов, инфицированных патогенами клеток или тканей, раковых клеток или нормальных клеток или тканей человека в случаях аутоиммунной реакции или патологического воспаления.
[0033] Термин «антиген-специфичный T-клеточный ответ» относится к ответу T-клетки, который является результатом стимуляции указанной T-клетки антигеном, в отношении которого указанная T-клетка является специфичной. Неограничивающие примеры ответов, осуществляемых T-клеткой при антиген-специфичной стимуляции, включают пролиферацию и продукцию цитокинов (например, продукцию IL-2).
[0034] Термин «антитело», как описано в настоящей заявке, включает полноразмерные антитела и любой антиген-связывающий фрагмент (т.е. «антиген-связывающую часть») или одну цепь антитела. Полноразмерные антитела представляют собой гликопротеины, содержащие две тяжелые (H) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельного участка тяжелой цепи (сокращенное название в настоящей заявке - VH) и константного участка тяжелой цепи. Константный участок тяжелой цепи состоит из трех доменов - CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельного участка легкой цепи (сокращенное название в настоящей заявке - VL) и константного участка легкой цепи. Константный участок легкой цепи состоит из одного домена - CL. Участки VH и VL можно дополнительно разделить на участки гипервариабельности, называемые определяющими комплементарность участками (complementarity determining regions, CDR), чередующиеся с участками, которые являются более консервативными и называются каркасными участками (framework regions, FR). Каждый VH и V L состоит из трех CDR и четырех FR, организованных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные участки тяжелой и легкой цепи содержат домен связывания, который взаимодействует с антигеном. Константные участки антитела могут опосредовать связывание иммуноглобулина с тканями и факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
[0035] Термин «антиген-связывающая часть» антитела (или просто «часть антитела») при употреблении в настоящей заявке относится к одному или более фрагментам антитела, которые сохраняют способность специфично связываться с антигеном (например, белком PD-1). Было показано, что антиген-связывающая функция антитела может осуществляться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, включенных в термин «антиген-связывающая часть» антитела, включают (i) Fab-фрагмент - моновалентный фрагмент, состоящий из доменов VL, VH, CL и C H1; (ii) F(ab')2-фрагмент - бивлентный фрагмент, содержащий два Fab-фрагмента, связанных с помощью дисульфидного мостика в шарнирном участке; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одной цепи антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; (vi) выделенный определяющий комплементарность участок (CDR) и (viii) нанотело - вариабельный участок тяжелой цепи, содержащий один вариабельный домен и два константных домена. Более того, несмотря на то что два домена Fv-фрагмента - VL и VH - кодируются отдельными генами, их можно соединять с помощью рекомбинантных методов с использованием синтетического линкера, который обеспечивает их получение в виде одной белковой цепи, в которой участки VL и VH соединяются с образованием моновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, Bird et al., (1988) Science 242:423-426; и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие одноцепочечные антитела также включены в термин «антиген-связывающая часть» антитела. Такие фрагменты антител получают с использованием стандартных способов, известных специалистам в данной области техники, и скрининг фрагментов на предмет их применимости проводят таким же образом, как и скрининг интактных антител.
[0036] Предполагается, что «выделенное антитело» при употреблении в настоящей заявке относится к антителу, которое по существу не содержит другие антитела, имеющие отличающуюся антигенную специфичность (например, выделенное антитело, которое специфично связывается с белком PD-1, по существу не содержит антитела, которые специфично связываются с антигенами, отличными от белков PD-1). Выделенное антитело, которое специфично связывается с белком PD-1 человека, может однако обладать перекрестной реакционной активностью с другими антигенами, такими как белки PD-1 из других видов. Более того, выделенное антитело может по существу не содержать другой клеточный материал и/или химические вещества.
[0037] Термины «моноклональное антитело» или «композиция моноклонального антитела» при употреблении в настоящей заявке относятся к препарату молекул антитела одномолекулярного состава. Композиция на основе моноклонального антитела проявляет единственную специфичность связывания и аффинность в отношении конкретного эпитопа.
[0038] Предполагается, что термин «мышиное антитело» при употреблении в настоящей заявке включает антитела, содержащие вариабельные участки, в которых как каркасные, так и CDR-участки происходят из последовательностей иммуноглобулинов мышиной зародышевой линии. Кроме того, если антитело содержит константный участок, то указанный константный участок также происходит из последовательностей иммуноглобулинов мышиной зародышевой линии. Мышиные антитела согласно изобретению могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов мышиной зародышевой линии (например, содержащие мутации, введенные путем рандомного или сайт-специфично мутагенеза in vitro или путем соматической мутации in vivo). Однако предполагается, что термин «мышиное антитело» при употреблении в настоящей заявке не включает антитела, в которых последовательности CDR, происходящие из зародышевой линии других видов млекопитающих, были привиты на мышиные каркасные последовательности.
[0039] Термин «гибридное антитело» относится к антителу, созданному путем объединения генетического материала из нечеловеческого источника с генетическим материалом человека. Более обобщенно, гибридное антитело представляет собой антитело, содержащее генетический материал из конкретных видов с генетическим материалом из других видов.
[0040] Термин «гуманизированное антитело» при употреблении в настоящей заявке относится к антителу из видов, не представляющих собой человека, белковые последовательности которых были модифицированы для повышения степени подобия вариантам природных антител, которые продуцируются у людей.
[0041] Термин «изотип» относится к классу антител (например, IgM или IgG1), которые кодируются генами константных участков тяжелой цепи.
[0042] Фразы «антитело, распознающее антиген», и «антитело, специфичное в отношении антигена», используются в настоящей заявке взаимозаменяемо с термином «антитело, которое специфично связывается с антигеном».
[0043] Предполагается, что при употреблении в настоящей заявке антитело, которое «специфично связывается с PD-1 человека», относится к антителу, которое связывается с белком PD-1 человека (и, возможно, белком PD-1 из одного или более видов, не представляющих собой человека), но по существу не связывается с белками, не представляющими собой PD-1. Предпочтительно, антитело связывается с белком PD-1 человека с «высокой аффинностью», а именно со значением KD, составляющим 5,0 x10-8 M или менее, более предпочтительно, 1,0 x10-8 M или менее, и более предпочтительно, 7,0×10-9 M или менее.
[0044] Термин «по существу не связывается» с белком или клетками при употреблении в настоящей заявке означает отсутствие связывания или отсутствие связывания с высокой аффинностью с указанным белком или клетками, т.е. относится к связыванию с белком или клетками со значением KD, составляющим 1,0×10-6 M или более, более предпочтительно, 1,0×10-5 M или более, более предпочтительно, 1,0×10-4 M или более, более предпочтительно, 1,0×10-3 M или более, более предпочтительно, 1,0×10-2 M или более.
[0045] Термин «высокая аффинность», относящийся к антителу IgG, относится к антителу, имеющему значение KD для антигена-мишени, составляющее 1,0×10-6 M или менее, более предпочтительно, 5,0×10-8 M или менее, более предпочтительно, 1,0×10-8 M или менее, более предпочтительно, 7,0×10-9 M или менее, более предпочтительно, 1,0×10-9 M или менее. Однако «высокая аффинность» связывания может варьировать для других изотипов антитела. Например, «высокая аффинность» связывания для изотипа IgM относится к антителу, имеющему значение KD, составляющее 10-6 M или менее, более предпочтительно, 10-7 M или менее, более предпочтительно, 10-8 M или менее.
[0046] Предполагается, что термин «Kassoc» или «Ka» при употреблении в настоящей заявке относится к скорости ассоциации для конкретного взаимодействия антитело-антиген, тогда как термин «Kdis» или «Kd» при употреблении в настоящей заявке относится к скорости диссоциации для конкретного взаимодействия антитело-антиген. Предполагается, что термин «KD» при употреблении в настоящей заявке относится к константе диссоциации, которую получают из отношения Kd к Ka (т.е., Kd/Ka) и выражают как молярную концентрацию (M). Значения KD для антител можно определять с использованием способов, общепризнанных в данной области техники. Предпочтительный способ определения KD антитела представляет собой использование поверхностного плазмонного резонанса, предпочтительно, с использованием биосенсорной системы, такой как система Biacore™.
[0047] Термин «EC50», также известный как полумаксимальная эффективная концентрация, относится к концентрации антитела, которая вызывает ответ, интенсивность которого находится посередине между исходным уровнем и максимальным уровнем после определенного времени воздействия.
[0048] Термин «IC50», также известный как полумаксимальная ингибирующая концентрация, относится к концентрации антитела, которая приводит к подавлению специфической биологической или биохимической функции на 50% по отношению к подавлению в отсутствии антитела.
[0049] Термин «субъект» включает любое животное, представляющее или не представляющее собой человека. Термин «не представляющее собой человека животное» включает всех позвоночных, например, млекопитающих и не млекопитающих, таких как не представляющие собой человека приматы, овцы, собаки, кошки, коровы, лошади, куры, амфибии и рептилии, при этом млекопитающие являются предпочтительными, такие как не представляющие собой человека приматы, овцы, собаки, кошки, коровы и лошади.
[0050] Термин «терапевтически эффективное количество» означает количество антитела согласно настоящему изобретению, достаточное для предотвращения или ослабления симптомов, связанных с заболеванием или состоянием (таким как рак) и/или снижения тяжести указанного заболевания или состояния. Предполагается, что терапевтически эффективное количество рассматривается в контексте состояния, подлежащего лечению, при этом специалисты в данной области техники легко определят реальное эффективное количество.
[0051] Различные аспекты настоящего изобретению описаны более подробно в следующих подразделах.
[0052] Антитела против PD-1, обладающие повышенной аффинностью связывания с PD-1 человека и лучшим противоопухолевым эффектом
[0053] Антитело или его антиген-связывающая часть согласно изобретению специфично связывается с PD-1 человека и обладает повышенной аффинностью связывания, а также сравнимым или лучшим противоопухолевым эффектом по сравнению с ранее описанными антителами против PD-1, в частности, по сравнению с ниволумабом.
[0054] Антитело или его антиген-связывающая часть согласно изобретению предпочтительно связывается с белком PD-1 человека со значением KD, составляющим 7,0×10-9 M или менее, более предпочтительно, со значением KD, составляющим 5,0×10-10 M или менее. Антитела согласно изобретению также связываются с PD-1 макаков-крабоедов со значением KD, составляющим от примерно 1,0×10-7 M до 1,0×10-10 M.
[0055] Дополнительные функциональные свойства включают способность блокировать взаимодействие PD-1/PD-L1. Антитела согласно настоящему изобретению, согласно одному варианту реализации, могут ингибировать связывание PD-1 и PD-L1 в сходной с ниволумабом концентрации.
[0056] Другие функциональные свойства включают способность антитела стимулировать иммунный ответ, такой как антиген-специфичный T-клеточный ответ. Указанную способность можно проверить, например, путем оценки способности антитела стимулировать продукцию интерлейкина-2 (IL-2) при антиген-специфичном T-клеточном ответе. Согласно конкретным вариантам реализации изобретения, антитело связывается с PD-1 человека и стимулирует антиген-специфичный T-клеточный ответ. Согласно другим вариантам реализации изобретения, антитело связывается с PD-1 человека, но не стимулирует антиген-специфичный T-клеточный ответ. Другие способы оценки способности антитела стимулировать иммунный ответ включают исследование его способности подавлять опухолевый рост, например, на in vivo модели опухолевого трансплантата, или способности стимулировать аутоиммунный ответ, например, способности обеспечивать развитие аутоиммунного заболевания на аутоиммунной модели, например, способности обеспечивать развитие диабета на NOD-мышиной модели.
[0057] Предпочтительные антитела согласно изобретению представляют собой человеческие моноклональные антитела. Дополнительно или альтернативно, антитела могут, например, представлять собой гибридные или гуманизированные моноклональные антитела.
[0058] Моноклональное антитело против PD-1
[0059] Предпочтительное антитело согласно изобретению представляет собой моноклональное антитело, имеющее структурные и химические характеристики, описанные ниже и в следующих далее Примерах. Аминокислотная последовательность VH антитела против PD-1 представлена в последовательности SEQ ID NO: 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78 или 79. Аминокислотная последовательность VL антитела против PD-1 показана в последовательности SEQ ID NO: 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95 или 96. Идентификационные номера (ID) аминокислотных последовательностей вариабельных участков тяжелых/легких цепей антител суммированы в Таблице 1, ниже, при этом некоторые клоны содержат одинаковый VH- или VL-участок. Константный участок тяжелой цепи для всех клонов может представлять собой константный участок тяжелой цепи IgG, имеющий аминокислотную последовательность, представленную, например, в последовательности SEQ ID NO: 97 или 129, и константный участок легкой цепи для всех клонов может представлять собой константный участок каппа-цепи, имеющий аминокислотную последовательность, представленную, например, в SEQ ID NO: 98 или 130. Fab-антитело может содержать вариабельный участок тяжелой/легкой цепи, CH1-участок тяжелой цепи (такой как участок, представленный в последовательности SEQ ID NO: 99) и константный участок легкой цепи.
[0060] Вариабельные участки CDR тяжелых цепей и вариабельные участки CDR легких цепей из Таблицы 1 были определены с помощью системы нумерации IMGT и системы нумерации Кабата соответственно. Однако, как хорошо известно в данной области техники, участки CDR также можно определять с помощью других систем, таких как система нумерации Чотиа и CCG-система/метод, на основе последовательностей вариабельных участков тяжелой/легкой цепи.
Таблица 1. ID номера аминокислотных последовательностей вариабельных участков тяжелой/легкой цепи
[0060] Последовательности VH и VL (или последовательности CDR) других антител против PD-1, которые связываются с PD-1 человека, можно «сочетать и комбинировать» с последовательностями VH и VL (или последовательностями CDR) антитела против PD-1 согласно настоящему изобретению. Предпочтительно, когда сочетают и комбинируют цепи VH и VL (или CDR в пределах таких цепей), последовательность VH из конкретной пары VH/VL замещают на структурно сходную последовательность VH. Подобным образом, последовательность VL из конкретной пары VH/VL предпочтительно замещают на структурно сходную последовательность VL.
[0062] Соответственно, согласно одному варианту реализации изобретения, антитело согласно изобретению или его антиген-связывающая часть содержит:
(a) вариабельный участок тяжелой цепи, содержащий аминокислотную последовательность, перечисленную выше в Таблице 1; и
(b) вариабельный участок легкой цепи, содержащий аминокислотную последовательность, перечисленную выше в Таблице 1, или VL другого антитела против PD-1, где указанное антитело специфично связывается с PD-1 человека.
[0063] Согласно другому варианту реализации изобретения, антитело согласно изобретению или его антиген-связывающая часть содержит:
(a) участки CDR1, CDR2 и CDR3 вариабельного участка тяжелой цепи, перечисленные выше в Таблице 1; и
(b) участки CDR1, CDR2 и CDR3 вариабельного участка легкой цепи, перечисленные выше в Таблице 1, или участки CDR другого антитела против PD-1, где указанное антитело специфично связывается с PD-1 человека.
[0064] Согласно другому варианту реализации изобретения, антитело или его антиген-связывающая часть содержит вариабельный участок тяжелой цепи CDR2 антитела против PD-1, объединенный с участками CDR других антител, которые связываются с PD-1 человека, например, CDR1 и/или CDR3 из вариабельного участка тяжелой цепи и/или CDR1, CDR2 и/или CDR3 из вариабельного участка легкой цепи другого антитела против PD-1.
Кроме того, в данной области техники хорошо известно, что домен CDR3 независимо от домена (доменов) CDR1 и/или CDR2 отдельно может определять специфичность связывания антитела с когнатным антигеном и что представляется возможным создание множества антител на основе общей последовательности CDR3, обладающих одинаковой специфичностью связывания. См., например, Klimka et al.,, British J. of Cancer 83(2):252-260 (2000); Beiboer et al.,, J. Mol. Biol. 296:833-849 (2000); Rader et al.,, Proc. Natl. Acad. Sci. U.S.A. 95:8910-8915 (1998); Barbas et al.,, J. Am. Chem. Soc. 116:2161-2162 (1994); Barbas et al.,, Proc. Natl. Acad. Sci. U.S.A. 92:2529-2533 (1995); Ditzel et al.,, J. Immunol. 157:739-749 (1996); Berezov et al.,, BIAjournal 8: Scientific Review 8 (2001); Igarashi et al.,, J. Biochem (Tokyo) 117:452-7 (1995); Bourgeois et al.,, J. Virol 72:807-10 (1998); Levi et al.,, Proc. Natl. Acad. Sci. U.S.A. 90:4374-8 (1993); Polymenis and Stoller, J. Immunol. 152:5218-5329 (1994) and Xu and Davis, Immunity 13:37-45 (2000). См. также, патенты США № 6 951 646; 6 914 128; 6 090 382; 6 818 216; 6 156 313; 6 827 925; 5 833 943; 5 762 905 и 5 760 185. Каждая из указанных ссылок полностью включена в настоящую заявку посредством ссылки.
[0066] Соответственно, согласно другому варианту реализации изобретения, антитела согласно изобретению содержат CDR2 вариабельного участка тяжелой цепи антитела против PD-1 и по меньшей мере CDR3 вариабельного участка тяжелой и/или легкой цепи антитела против PD-1 или CDR3 вариабельного участка тяжелой и/или легкой цепи другого антитела против PD-1, где указанное антитело способно специфично связываться с PD-1 человека. Указанные антитела предпочтительно (a) конкурируют за связывание с PD-1; (b) сохраняют функциональные характеристики; (c) связываются с одним и тем же эпитопом и/или (d) обладают сходной аффинностью связывания по сравнению с антителом против PD-1 согласно настоящему изобретению. Согласно другому варианту реализации изобретения, антитела дополнительно могут содержать CDR2 вариабельного участка легкой цепи антитела против PD-1 или CDR2 вариабельного участка легкой цепи другого антитела против PD-1, где указанное антитело способно специфично связываться с PD-1 человека. Согласно другому варианту реализации изобретения, антитела согласно изобретению могут содержать CDR1 вариабельного участка тяжелой и/или легкой цепи антитела против PD-1 или CDR1 вариабельного участка тяжелой и/или легкой цепи другого антитела против PD-1, где указанное антитело способно специфично связываться с PD-1 человека.
[0067] Консервативные модификации
[0068] Согласно другому варианту реализации я, антитело согласно изобретению содержит последовательности вариабельного участка CDR1, CDR2 и CDR3 тяжелой и/или легкой цепи, которые отличаются от последовательностей антитела против PD-1 согласно настоящему изобретению по одной или более консервативным модификациям. Следует понимать в данной области техники, что возможны определенные консервативные модификации последовательности, которые не устраняют способность связывания антигена. См., например, Brummell et al., (1993) Biochem 32:1180-8; de Wildt et al., (1997) Prot. Eng. 10:835-41; Komissarov et al., (1997) J. Biol. Chem. 272:26864-26870; Hall et al., (1992) J. Immunol. 149:1605-12; Kelley and O'Connell (1993) Biochem,32:6862-35; Adib-Conquy et al., (1998) Int. Immunol,10:341-6 и Beers et al., (2000) Clin. Can. Res. 6:2835-43.
[0069] Соответственно, согласно одному варианту реализации изобретения, антитело содержит вариабельный участок тяжелой цепи, содержащий последовательности CDR1, CDR2 и CDR3, и/или вариабельный участок легкой цепи, содержащий последовательности CDR1, CDR2 и CDR3, где:
(a) последовательность вариабельного участка тяжелой цепи CDR1 содержит последовательность, перечисленную в Таблице 1, выше, и/или ее консервативные модификации; и/или
(b) последовательность вариабельного участка тяжелой цепи CDR2 содержит последовательность, перечисленную в Таблице 1, выше, и/или ее консервативные модификации; и/или
(c) последовательность вариабельного участка тяжелой цепи CDR3 содержит последовательность, перечисленную в Таблице 1, выше, и ее консервативные модификации; и/или
(d) последовательности вариабельного участка легкой цепи CDR1 и/или CDR2 и/или CDR3 содержат последовательность (последовательности), перечисленную в Таблице 1, выше; и/или ее консервативные модификации; и
(e) антитело специфично связывается с PD-1 человека.
[0070] Антитело согласно настоящему изобретению обладает одним или более из следующих функциональных свойств, описанных выше, таких как высокая аффинность связывания с PD-1 человека и способность вызывать антитело-зависимую клеточную цитотоксичность (АЗКЦ) или комплемент-зависимую цитотоксичность (КЗЦ) в отношении PD-1-эксперссирующих клеток.
[0071] Согласно различным вариантам реализации изобретения, антитело может представлять собой, например, мышиное, человеческое, гуманизированное или гибридное антитело.
[0072] Предполагается, что при употреблении в настоящей заявке термин «консервативные модификации последовательности» относится к аминокислотным модификациям, которые значительно не влияют или не изменяют характеристики связывания антитела, содержащего указанную аминокислотную последовательность. Такие консервативные модификации включают аминокислотные замены, добавления и делеции. Модификации можно вводить в антитело согласно изобретению с помощью стандартных методов, известных в данной области техники, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены представляют собой замены, при которых аминокислотный остаток замещают на аминокислотный остаток, имеющий сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в данной области техники. Указанные семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), аминокислоты с кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), аминокислоты с незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), аминокислоты с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейин, пролин, фенилаланин, метионин), аминокислоты с бета-разветвленными боковыми цепи (например, треонин, валин, изолейцин) и аминокислоты с ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или более аминокислотных остатков в пределах CDR-участков антитела согласно изобретению могут быть замещены на другие аминокислотные остатки из того же семейства по боковым цепям, и сохранение функции измененного антитела (т.е. функций, изложенных выше) можно исследовать с использованием функциональных анализов, описанных в настоящей заявке.
[0073] Сконструированные и модифицированные антитела
[0074] Антитела согласно изобретению могут быть получены с использованием антитела, содержащего одну или более из последовательностей VH/VL антитела против PD-1 согласно настоящему изобретению, в качестве исходного материала для конструирования модифицированного антитела. Антитело может быть сконструировано путем модифицирования одного или более остатков в пределах одного или обоих вариабельных участков (т.е. VH и/или VL), например, в пределах одного или более CDR-участков и/или в пределах одного или более каркасных участков. Дополнительно или альтернативно, антитело может быть сконструировано путем модифицирования остатков в пределах константного участка (участков), например, для изменения эффекторной функции (функций) указанного антитела.
[0075] Согласно конкретным вариантам реализации изобретения, прививку CDR можно использовать для конструирования вариабельных участков антител. Антитела взаимодействуют с антигенами-мишенями главным образом через аминокислотные остатки, которые расположены в шести определяющих комплементарность участках (CDR) тяжелой и легкой цепи. По этой причине аминокислотные последовательности в пределах CDR-участков различаются сильнее между отдельными антителами, чем последовательности вне CDR. Поскольку CDR-последовательности отвечают за большинство взаимодействий антитело-антиген, возможно экспрессировать рекомбинантные антитела, которые имитируют свойства специфичных природных антител, путем конструирования векторов экспрессии, которые содержат CDR-последовательности из специфичного природного антитела, привитые на каркасные последовательности из другого антитела с отличающимися свойствами (см., например, Riechmann et al., (1998) Nature 332:323-327; Jones et al., (1986) Nature 321:522-525; Queen et al., (1989) Proc. Natl. Acad. См. также заявки на патенты США 86:10029-10033; патенты США № 5 225 539; 5 530 101; 5 585 089; 5 693 762 и 6 180 370).
[0076] Соответственно, другой вариант реализации изобретения относится к выделенному моноклональному антителу или его антиген-связывающей части, содержащей вариабельный участок тяжелой цепи, который содержит последовательности CDR1, CDR2 и CDR3, содержащие последовательности согласно настоящему изобретению, как описано выше, и/или вариабельный участок легкой цепи, содержащий последовательности CDR1, CDR2 и CDR3, которые содержат последовательности согласно настоящему изобретению, как описано выше. Тогда как указанные антитела содержат CDR-последовательности VH и VL моноклонального антитела согласно настоящему изобретению, они могут содержать различные каркасные последовательности.
[0077] Такие каркасные последовательности могут быть получены из общедоступных баз данных ДНК или опубликованных ссылок, которые включают последовательности генов антител зародышевой линии. Например, последовательности ДНК зародышевой линии для генов человеческих вариабельных участков тяжелой и легкой цепи можно найти в базе данных последовательностей зародышевой линии человека «VBase» (доступна в интернете по адресу www.mrc-cpe.cam.ac.uk/vbase), а также в источнике Kabat et al., (1991), цитированном выше; Tomlinson et al., (1992) J. Mol. Biol. 227:776-798; и Cox et al., (1994) Eur. J. Immunol. 24:827-836; содержание каждого из которых специально включено в настоящую заявку посредством ссылки. В качестве другого примера, последовательности ДНК зародышевой линии для генов человеческого вариабельного участка тяжелой и легкой цепи можно найти в базе данных Genbank. Например, следующие последовательности тяжелой цепи зародышевой линии, встречающиеся в мышином HCo7 HuMAb, доступны по прилагаемым номерам доступа в базе данных Genbank: 1-69 (NG--0010109, NT--024637 & BC070333), 3-33 (NG--0010109 & NT--024637) и 3-7 (NG--0010109 & NT--024637). В качестве другого примера, следующие последовательности тяжелой цепи зародышевой линии, встречающиеся в мышином HCo12 HuMAb, доступны по прилагаемым номерам доступа в базе данных Genbank: 1-69 (NG--0010109, NT--024637 & BC070333), 5-51 (NG--0010109 & NT--024637), 4-34 (NG--0010109 & NT--024637), 3-30,3 (CAJ556644) & 3-23 (AJ406678).
[0078] Белковые последовательности антитела сравнивают с базой данных компилированных белковых последовательностей с использованием одного из способов поиска подобия по последовательностям, называемого Gapped BLAST (Altschul et al., (1997), выше), который хорошо известен специалисту в данной области техники.
[0079] Предпочтительные каркасные последовательности для применения в антителах согласно изобретению представляют собой последовательности, которые структурно сходны с каркасными последовательностями, используемыми в антителах согласно изобретению. Последовательности CDR1, CDR2 и CDR3 VH можно прививать на каркасные участки, которые имеют последовательность, идентичную последовательности, встречающейся в гене иммуноглобулина зародышевой линии, из которой происходит каркасная последовательность, или CDR-последовательности можно прививать на каркасные участки, которые содержат одну или более мутаций по сравнению с последовательностями зародышевой линии. Например, было обнаружено, что в конкретных примерах благоприятным является введение мутаций в остатки в пределах каркасных участков для поддержания или усиления способности антитела связывать антиген (см., например, патенты США № 5 530 101; 5 585 089; 5 693 762 и 6 180 370).
[0080] Другой тип модификации вариабельного участка представляет собой мутацию аминокислотных остатков в пределах участков CDR1, CDR2 и/или CDR3 VH и/или VL для улучшения таким образом одного или более свойств связывания (например, аффинности) интересующего антитела. Для введения мутации (мутаций) можно осуществлять сайт-направленный мутагенез или ПЦР-опосредованный мутагенез и воздействие на связывание антитела или другое интересующее функциональное свойство можно оценивать с помощью in vitro или in vivo анализов, как известно в данной области техники. Предпочтительно вводят консервативные модификации (как известно в данной области техники). Мутации могут представлять собой аминокислотные замены, добавления или делеции, но предпочтительно представляют собой замены. Более того, как правило, изменяют не более чем один, два, три, четыре или пять остатков в пределах CDR-участка.
[0081] Соответственно, согласно другому варианту реализации, настоящее изобретение обеспечивает выделенные моноклональные антитела против PD-1 или их антиген-связывающие части, содержащие вариабельный участок тяжелой цепи, содержащий: (a) участок CDR1 VH, содержащий последовательность согласно настоящему изобретению или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений; (b) участок CDR2 VH, содержащий последовательность согласно настоящему изобретению или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений; (c) участок CDR3 VH, содержащий последовательность согласно настоящему изобретению или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений; (d) участок CDR1 VL, содержащий последовательность согласно настоящему изобретению или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений; (e) участок CDR2 VL, содержащий последовательность согласно настоящему изобретению или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений; и (f) участок CDR3 VL, содержащий последовательность согласно настоящему изобретению или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений.
[0082] Сконструированные антитела согласно изобретению включают антитела, в которые были внесены модификации в каркасные остатки в пределах VH и/или VL, например для улучшения свойств указанного антитела. Как правило, такие модификации каркасных остатков осуществляют для снижения иммуногенности антитела. Например, один подход представляет собой «мутирование к первоначальному виду» одного или более каркасных остатков к соответствующей последовательности зародышевой линии. Более конкретно, антитело, которое прошло соматическую мутацию, может содержать каркасные остатки, которые отличаются от последовательности зародышевой линии, из которой произошло антитело. Такие остатки можно идентифицировать путем сравнения каркасных последовательностей антитела с последовательностями зародышевой линии, из которой произошло антитело.
[0083] Другой тип каркасной модификации включает мутирование одного или более остатков в пределах каркасного участка или даже в пределах одного или более CDR-участков для удаления T-клеточных эпитопов для снижения, таким образом, потенциальной иммуногенности антитела. Этот поход также называется «деиммунизацией» и описан более подробно в публикации патента США № 20030153043.
[0084] В качестве дополнения или альтернативы модификациям, сделанным в пределах каркасных или CDR-участков, антитела согласно изобретению могут быть сконструированы таким образом, что включают модификации в пределах Fc-участка, как правило, для изменения одного или более функциональных свойств указанного антитела, таких как время полужизни в сыворотке крови, связывание комплемента, связывание Fc-рецептора и/или антиген-зависимая клеточная цитотоксичность. Более того, антитело согласно изобретению может быть химически модифицировано (например, один или более химических фрагментов могут быть присоединены к антителу) или модифицировано с изменением степени его гликозилирования также для изменения одного или более функциональных свойств указанного антитела.
[0085] Согласно одному варианту реализации изобретения, шарнирный участок CH1 модифицирован таким образом, что количество цистеиновых остатков в указанном шарнирном участке изменено, например, повышено или понижено. Этот подход описан дополнительно в патенте США № 5 677 425. Количество цистеиновых остатков в шарнирном участке CH1 изменяют, например, для облегчения сборки легких и тяжелых цепей или для повышения или снижения стабильности антитела.
[0086] Согласно другому варианту реализации изобретения, в Fc-шарнирный участок антитела вводят мутации для снижения времени биологической полужизни антитела. Более конкретно, одну или более аминокислотных мутаций вводят в пограничную область домена CH2-CH3 Fc-шарнирного фрагмента таким образом, что связывание антитела с белком А стафилококка (SpA) нарушается по сравнению с SpA-связыванием нативного Fc-шарнирного домена. Этот подход описан более подробно в патенте США № 6 165 745.
[0087] Согласно другому варианту реализации изобретения, модифицируют степень гликозилирования антитела. Например, может быть получено гликозилированное антитело (т.е. если антитело не гликозилировано). Степень гликозилирования можно изменять, например, для повышения аффинности антитела в отношении антигена. Такие углеводные модификации можно осуществлять, например, путем изменения одного или более сайтов гликозилирования в пределах последовательности антитела. Например, могут быть сделаны одна или более аминокислотных замен, которые приводят к устранению одного или более сайтов гликозилирования в каркасе вариабельного участка для устранения таким образом гликозилирования в этом сайте. Такое агликозилирование может приводить к повышению аффинности антитела в отношении антигена. См., например, патент США № 5 714 350 и 6 350 861.
[0088] Дополнительно или альтернативно, может быть получено антитело с измененным типом гликозилирования, такое как гипофукозилированное антитело, содержащее пониженное количество фукозильных остатков, или антитело, имеющее повышенное содержание структур с остатком GlcNac в точке ветвления. Было продемонстрировано, что такие измененные паттерны гликозилирования повышают способность антитела к антитело-зависимой клеточной цитотоксичности (АЗКЦ). Такие углеводные модификации можно осуществлять путем, например, экспрессии антитела в клетке-хозяине с измененной системой гликозилирования. Клетки с измененной системой гликозилирования были описаны в данной области техники и могут использоваться в качестве клеток-хозяев для экспрессии в них рекомбинантного антитела согласно изобретению с получением таким образом антитела с измененным гликозилированием. Например, клеточные линии Ms704, Ms705 и Ms709 не содержат ген фукозилтрансферазы, FUT8 (α(1,6)-фукозилтрансфераза), и таким образом антитела, экспрессируемые в клеточных линиях Ms704, Ms705 и Ms709, не содержат фукозу на улеводах. Клеточные линии Ms704, Ms705 и Ms709 FUT8-/- были созданы с помощью направленного разрушения гена FUT8 в клетках CHO/DG44 с использованием двух векторов замещения (см. публикацию патента США № 20040110704 и Yamane-Ohnuki et al., (2004) Biotechnol Bioeng 87:614-22). В качестве другого примера, европейский патент EP 1 176 195 описывает клеточную линию с функционально разрушенным геном FUT8, который кодирует фукозилтрансферазу, таким образом, что антитела, экспрессируемые в такой клеточной линии, являются гипофукозилированными из-за снижения или удаления связанного с α-1,6-связью фермента. Европейский патент EP 1 176 195 также описывает клеточные линии, которые обладают низкой ферментативной активностью для добавления фукозы на N-ацетилглюкозамин, который связывается с Fc-участком антитела, или не обладает ферментативной активностью, например, крысиная клеточная линия миеломы YB2/0 (ATCC CRL 1662). Публикация международной заявки согласно PCT WO 03/035835 описывает вариант клеточной линии CHO - клетки Lec13 - со сниженной способностью присоединять фукозу к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых в указанной клетке-хозяине (см. также Shields et al., (2002) J. Biol. Chem. 277:26733-26740). Антитела с модифицированным профилем гликозилирования также могут быть получены в куриных яйцах, как описано в публикации международной заявки согласно PCT WO 06/089231. Альтернативно, антитела с модифицированным профилем гликозилирования могут быть получены в растительных клетках, таких как Lemna. Способы получения антител в растительной системе описаны в заявке на патент США, соответствующей заявке Alston & Bird LLP, номер патентного реестра 040989/314911, поданной 11 августа 2006 года. Публикация международной заявки согласно PCT WO 99/54342 описывает клеточные линии, сконструированные для экспрессии модифицирующих гликопротеины гликозилтрансфераз (например, β(1,4)-N-ацетилглюкозаминилтранферазы III (GnTIII)) таким образом, что антитела, экспрессируемые в указанных сконструированных клеточных линиях, содержат повышенное количество структур с остатком GlcNac в точке ветвления, что приводит к повышенной АЗКЦ активности антител (см. также Umana et al., (1999) Nat. Biotech. 17:176-180). Альтернативно, фукозные остатки антитела можно отщеплять с использованием фермента фукозидазы; например, фукозидаза α-L-фукозидаза удаляет фукозильные остатки от антитела (Tarentino et al., (1975) Biochem. 14:5516-23).
[0089] Другая модификация антитела, описанного в настоящей заявке, которая рассматривается согласно настоящему изобретению, представляет собой пэгилирование. Антитело может быть пэгилировано, например, для повышения времени биологической полужизни (например, полужизни в сыворотке) указанного антитела. Для пэгилирования антитело или его фрагмент, как правило, подвергают реагированию с полиэтиленгликолем (ПЭГ), таким как реакционно активный сложный эфир или альдегидное производное ПЭГ, в условиях, при которых одна или более ПЭГ-групп присоединяется к антителу или фрагменту антитела. Предпочтительно, пэгилирование осуществляют посредством реакции ацилирования или реакции алкилирования с реакционно активной ПЭГ-молекулой (или аналогичным реакционно активным водорастворимым полимером). Предполагается, что при употреблении в настоящей заявке термин «полиэтиленгликоль» включает любые из форм ПЭГ, которые использовались для дериватизации других белков, такие как моно (C1-C10)алкокси- или арилокси-полиэтиленгликоль или полиэтиленгликоль-малеимид. Согласно конкретным вариантам реализации изобретения, антитело, подлежащее пэгилированию, представляет собой агликозилированное антитело. Способы пэгилирования белков известны в данной области техники и могут применяться для антител согласно изобретению. См., например, европейские патенты EP 0 154 316 и EP 0 401 384.
[0090] Физические свойства антитела
[0091] Антитела согласно изобретению могут быть охарактеризованы по различным физическим свойствам для выявления и/или определения различий между разными классами.
[0092] Например, антитела могут содержать один или более сайтов гликозилирования в вариабельном участке легкой или тяжелой цепи. Такие сайты гликозилирования могут приводить к повышенной иммуногенности антитела или изменению его pK благодаря измененному связыванию антигена (Marshall et al (1972) Annu Rev Biochem 41:673-702; Gala and Morrison (2004) J Immunol 172:5489-94; Wallick et al., (1988) J Exp Med 168:1099-109; Spiro (2002) Glycobiology 12:43R-56R; Parekh et al (1985) Nature 316:452-7; Mimura et al., (2000) Mol Immunol 37:697-706). Известно, что гликозилирование происходит в мотивах, содержащий последовательность N-X-S/T. В некоторых примерах предпочтительным является антитело против PD-1 без гликозилирования вариабельного участка, что может быть достигнуто либо путем выбора антител, которые не содержат мотив гликозилирования в вариабельном участке, либо путем мутирования остатков в области гликозилирования.
[0093] Согласно предпочтительному варианту реализации изобретения, антитела не содержат сайтов изомерии аспарагина. Дезамидирование аспарагина может происходить на последовательностях N-G или D-G и приводить к образованию остатка изоаспарагиновой кислоты, который вводит связь в полипептидную цепь и снижает ее стабильность (эффект изоаспарагиновой кислоты).
[0094] Каждое антитело имеет уникальную изоэлектрическую точку (pI), которая в целом находится в пределах диапазона pH между 6 и 9,5. Значение pI для антитела IgG1, как правило, находится в пределах диапазона pH 7-9,5, и значение pI для антитела IgG4, как правило, находится в пределах диапазона pH 6-8. Существует предположение, что антитела, значение pI которых находится за пределами нормального диапазона, могут характеризоваться некоторой степенью разворачивания и нестабильности в условиях in vivo. Таким образом, предпочтительным является антитело против PD-1, значение pI которого находится в пределах нормального диапазона, что может быть достигнуто путем выбора антител, имеющих pI в пределах нормального диапазона, или путем мутирования заряженных поверхностных остатков.
[0095] Молекулы нуклеиновой кислоты, кодирующие антитела согласно изобретению
[0096] Согласно другому аспекту, изобретение обеспечивает молекулы нуклеиновой кислоты, которые кодируют вариабельные участки или CDR-участки тяжелых и/или легких цепей антител согласно изобретению. Нуклеиновые кислоты могут быть представлены в целых клетках, в клеточном лизате или в частично очищенной или по существу чистой форме. Нуклеиновая кислота является «выделенной» или «приведенной по существу в чистое состояние», когда она очищена от других клеточных компонентов или других примесей, например, других клеточных нуклеиновых кислот или белков, с помощью стандартных методов. Нуклеиновая кислота согласно изобретению может представлять собой, например, ДНК или РНК и может содержать или не содержать интронные последовательности. Согласно предпочтительному варианту реализации изобретения, нуклеиновая кислота представляет собой молекулу кДНК.
[0097] Нуклеиновые кислоты согласно изобретению могут быть получены с использованием стандартных методов молекулярной биологии. В случае антител, экспрессированных с помощью гибридом (например, гибридом, полученных их трансгенных мышей, содержащих гены человеческого иммуноглобулина, как описано дополнительно ниже), молекулы кДНК, кодирующие легкие и тяжелые цепи указанного антитела, полученного с помощью гибридомы, можно получать с помощью стандартных методов ПЦР-амплификации или кДНК-клонирования. В случае антител, полученных из библиотеки генов иммуноглобулинов (например, с использованием методов фагового дисплея), нуклеиновая кислота, кодирующая такие антитела, может быть получена из генной библиотеки.
[0098] Предпочтительные молекулы нуклеиновых кислот согласно изобретению включают нуклеиновые кислоты, кодирующие последовательности VH и VL моноклонального антитела против PD-1 или участки CDR. После получения фрагментов ДНК, кодирующих сегменты VH и VL, указанные фрагменты можно подвергать дополнительным манипуляциям с помощью стандартных методов рекомбинантной ДНК, например, для превращения генов вариабельного участка в гены цепей полноразмерного антитела, в гены Fab-фрагмента или ген scFv. В указанных процедурах VL- или VH-кодирующий фрагмент ДНК функционально связан с другим фрагментом ДНК, кодирующим другой белок, такой как константный участок или подвижный линкер антитела. Предполагается, что термин «функционально связан» при использовании в данном контексте означает, что два фрагмента ДНК соединены таким образом, что аминокислотные последовательности, кодирующиеся двумя фрагментами ДНК, остаются в одной рамке считывания.
[0099] Выделенную ДНК, кодирующую VH-участок, можно превращать в ген полноразмерной тяжелой цепи путем функционального связывания кодирующей VH ДНК с другой молекулой ДНК, кодирующей константные участки тяжелой цепи (CH1, CH2 и CH3). Последовательности генов человеческих константных участков тяжелой цепи известны в данной области техники, и ДНК фрагменты, включающие эти участки, могут быть получены с помощью стандартной ПЦР-амплификации. Константный участок тяжелой цепи может представлять собой константный участок IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно представляет собой константный участок IgG1 или IgG4. Для гена Fab-фрагмента тяжелой цепи кодирующая VH ДНК может быть функционально связана с другой молекулой ДНК, кодирующей только константный участок CH1 тяжелой цепи.
[00100] Выделенную ДНК, кодирующую VL-участок, можно превращать в ген полноразмерной легкой цепи (а также ген Fab легкой цепи) путем функционального связывания кодирующей VL ДНК с другой молекулой ДНК, кодирующей константный участок легкой цепи CL. Последовательности генов человеческого константного участка легкой цепи известны в данной области техники, и ДНК фрагменты, включающие указанные участки, могут быть получены с помощью стандартной ПЦР-амплификации. Согласно предпочтительным вариантам реализации, константный участок легкой цепи может представлять собой константный участок каппа- или лямбда-цепи.
[00101] Для создания гена scFv фрагменты ДНК, кодирующей VH и VL, функционально связывают с другим фрагментом, кодирующим подвижный линкер, например, кодирующим аминокислотную последовательность (Gly4-Ser)3, таким образом, что последовательности VH и VL могут экспрессироваться в виде непрерывного одноцепочечного белка, участки VL и VH которого соединены подвижным линкером (см. например, Bird et al., (1988) Science 242:423-426; Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., (1990) Nature 348:552-554).
[00102] Получение моноклонального антитела согласно изобретению
[00103] Моноклональные антитела (mAb) согласно настоящему изобретению могут быть получены с использованием хорошо известного способа гибридизации соматических клеток (гибридомы) согласно источнику Kohler and Milstein (1975) Nature 256: 495. Другие варианты реализации для продукции моноклональных антител включают вирусную или онкогенную трансформацию B-лимфоцитов и методы фагового дисплея. Гибридные или гуманизированные антитела также хорошо известны в данной области техники. См., например, патенты США № 4 816 567; 5 225 539; 5 530 101; 5 585 089; 5 693 762 и 6 180 370, содержание которых специально полностью включено в настоящую заявку посредством ссылки.
[00104] Создание трансфектов, продуцирующих моноклональные антитела согласно изобретению
[00105] Антитела согласно изобретению также могут быть получены в трансфектомной клетке-хозяине с использованием, например, комбинации методов рекомбинантной ДНК и генной трансфекции, как хорошо известно в данной области техники (см, например, Morrison, S. (1985) Science 229:1202). Согласно одному варианту реализации изобретения, ДНК, кодирующую части или полноразмерные легкие и тяжелые цепи, полученную с помощью стандартных методов молекулярной биологии, встраивают в один или более векторов экспрессии таким образом, что гены функционально связывают с последовательностями регуляции транскрипции или трансляции. Предполагается, что в данном контексте термин «функционально связанные» означает, что ген антитела лигирован в вектор таким образом, что последовательности контроля транскрипции и трансляции в пределах указанного вектора выполняют свою предполагаемую функцию регуляции транскрипции и трансляции гена антитела.
[00106] Предполагается, что термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования), которые контролируют транскрипцию и трансляцию генов антитела. Такие регуляторные последовательности описаны, например, в источнике Goeddel, Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, Calif. (1990). Предпочтительные регуляторные последовательности для экспрессии в хозяйских клетках млекопитающих включают вирусные элементы, которые направляют белковую экспрессию на высоком уровне в указанных клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовируса, например, главный поздний промотор аденовируса (AdMLP) и полиом. Альтернативно, можно использовать невирусные регуляторные последовательности, такие как убиквитиновый промотор или промотор β-глобина. Кроме того, существуют регуляторные элементы, состоящие из последовательностей из различных источников, такие как система промоторов SRα, которая содержит последовательности из раннего просмотра SV40 и длинный концевой повтор вируса T-клеточного лейкоза человека 1 типа (Takebe et al., (1988) Mol. Cell. Biol. 8:466-472). Вектор экспрессии и последовательности контроля экспрессии выбирают таким образом, чтобы они были совместимы с системой экспрессии используемой клетки-хозяина.
[00107] Ген легкой цепи антитела и ген тяжелой цепи антитела можно встраивать в один или раздельные векторы экспрессии. Согласно предпочтительным вариантам реализации, вариабельные участки используются для создания генов полноразмерного антитела любого изотипа путем встраивания их в векторы экспрессии, уже кодирующие константные участки тяжелой цепи и легкой цепи желаемого изотипа, таким образом, чтобы VH-сегмент был функционально связан с CH-сегментом (сегментами) в пределах вектора и VL-сегмент был функционально связан с CL-сегментом в пределах вектора. Дополнительно или альтернативно, рекомбинантный вектор экспрессии может кодировать сигнальный пептид, который способствует секреции цепи антитела из клетки-хозяина. Ген цепи антитела можно клонировать в вектор таким образом, чтобы сигнальный пептид был связан в рамке считывания с амино-концом гена цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (т.е. сигнальный пептид из белка, не представляющего собой иммуноглобулин).
[00108] Помимо генов цепи антитела и регуляторных последовательностей рекомбинантные векторы экспрессии согласно изобретению могут содержать дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, ориджины репликации), и селектируемые маркерные гены. Селектируемый маркерный ген способствует селекции клеток-хозяев, в которые был встроен вектор (см., например, патенты США № 4 399 216; 4 634 665 и 5 179 017). Например, селектируемый маркерный ген, как правило, предоставляет клетке-хозяину, в которую был введен вектор, устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат. Предпочтительные селектируемые маркерные гены включают ген дигидрофолатредуктазы (DHFR) (для применения в dhfr-клетках-хозяевах с селекцией/амплификацией по метатрексату) и нео-ген (для селекции по G418).
[00109] Для экспрессии легких и тяжелых цепей вектор (векторы) экспрессии, кодирующий указанные тяжелые и легкие цепи, трансфицируют в клетку-хозяина с помощью стандартных методов. Предполагается, что различные формы термина «трансфекция» включают широкое разнообразие методов, обычно используемых для введения экзогенной ДНК в прокариотическую ии эукариотическую клетку-хозяина, например, электропорацию, преципитацию фосфатом кальция, ДЭАЭ-декстрановую трансфекцию и т.п. Несмотря на теоретическую возможность экспрессии антител согласно изобретению как в прокариотических, так и в эукариотических клетках-хозяевах, наиболее предпочтительной является экспрессия антител в эукариотических клетках, наиболее предпочтительно, хозяйских клетках млекопитающих, поскольку в таких эукариотических клетках, в частности, клетках млекопитающих, с большей вероятностью происходит сборка и секреция правильно свернутого и иммунологически активного антитела по сравнению с прокариотическими клетками.
[00110] Предпочтительные хозяйские клетки млекопитающих для экспрессии рекомбинантных антител согласно изобретению включают клетки яичника китайского хомячка (CHO) (включая клетки dhfr-CHO, описанные в источнике Urlaub and Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, используемые с DHFR-селектируемым маркером, например, как описано в источнике R. J. Kaufman and P. A. Sharp (1982) J. Mol. Biol. 159:601-621), клетки миеломы NSO, клетки COS и клетки SP2. В частности, для применения с клетками миеломы NSO другая предпочтительная система экспрессии представляет собой систему экспрессии гена GS, описанную в публикации международной заявки WO 87/04462, публикации международной заявки WO 89/01036 и европейском патенте EP 338 841. Когда рекомбинантные векторы экспрессии, кодирующие гены антитела, вводят в хозяйскую клетку млекопитающего, продукцию антител обеспечивают путем культивирования указанных клеток-хозяев в течение периода времени, достаточного для обеспечения экспрессии антитела в указанных клетках-хозяевах, или, более предпочтительно, секреции антитела в культуральную среду, в которой растят указанную клетку-хозяина. Антитела можно восстанавливать из культуральной среды с использованием стандартных способов белковой очистки.
[00111] Иммуноконъюгаты
[00112] Антитела согласно изобретению можно конъюгировать с терапевтическим агентом с образованием иммуноконъюгата, такого как конъюгат антитело-лекарственное средство (КАЛС). Подходящие терапевтические агенты включают цитотоксины, алкилирующие агенты, связывающиеся с малой бороздкой ДНК вещества, ДНК-интеркаляторы, ДНК-кросслинкеры, ингибиторы гистондеацетилазы, ингибиторы ядерного экспорта, протеасомные ингибиторы, ингибиторы топоизомеразы I или II, ингибиторы белков теплового шока, ингибиторы тирозинкиназы, антибиотики и антимитотические агенты. В КАЛС антитело и терапевтический агент предпочтительно конъюгированы с помощью расщепляемого линкера, такого как пептидильный, дифульфидный или гидразоновый линкер. Более предпочтительно, линкер представляет собой пептидильный линкер, такой как Val-Cit, Ala-Val, Val-Ala-Val, Lys-Lys, Pro-Val-Gly-Val-Val, Ala-Asn-Val, Val-Leu-Lys, Ala-Ala-Asn, Cit-Cit, Val-Lys, Lys, Cit, Ser или Glu. КАЛС могут быть получены, как описано в патентах США № 7 087 600; 6 989 452 и 7 129 261; публикации международной заявки согласно PCT WO 02/096910; публикации международной заявки WO 07/038 658; публикации международной заявки WO 07/051 081; публикации международной заявки WO 07/059 404; публикации международной заявки WO 08/083 312 и публикации международной заявки WO 08/103 693; публикациях патентов США 20060024317; 20060004081 и 20060247295; описание которых включено в настоящую заявку посредством ссылки.
[00113] Биспецифичные молекулы
[00114] Согласно другому аспекту, настоящее изобретение обеспечивает биспецифичные молекулы, содержащие одно или более антител согласно изобретению, связанных по меньшей мере с одной другой функциональной молекулой, например, другим пептидом или белком (например, другим антителом или лигандом рецептора) для создания биспецифичной молекулы, которая связывается по меньшей мере с двумя различными сайтами связывания или молекулами-мишенями. Таким образом, при употреблении в настоящей заявке термин «биспецифичная молекула» включает молекулы, которые обладают тремя или более специфичностями.
[00115] Согласно варианту реализации изобретения, биспецифичная молекула помимо специфичности связывания с Fc и специфичности связывания с PD-1 обладает третьей специфичностью. Указанная третья специфичность может представлять собой специфичность в отношении усиливающего фактора (enhancement factor, EF), например, молекулу, которая связывается с поверхностным белком, вовлеченным в цитотоксическую активность и, таким образом, повышает иммунный ответ в отношении клетки-мишени. Например, молекула, направленная на усиливающий фактор, может связываться с цитотоксической T-клеткой (например, через CD2, CD3, CD8, CD28, CD4, PD-1 или ICAM-1) или другой иммунной клеткой, приводя к повышенному иммунному ответу против клетки-мишени.
[00116] Биспецифичные молекулы могут иметь множество различных форм и размеров. Биспецифичная молекула одного конца размерного ряда сохраняет традиционную форму антитела, за исключением того, что вместо двух групп связывания, обладающих идентичной специфичностью, она содержит две группы связывания с разной специфичностью. Биспецифичные молекулы противоположного конца размерного ряда состоят из двух одноцепочечных фрагментов антитела (scFv's), связанных с помощью пептидной цепи, - так называемый конструкт Bs(scFv)2. Биспецифичные молекулы промежуточного размера включают два различных F(ab)-фрагмента, связанных с помощью пептидильного линкера. Биспецифичные молекулы указанных и других форм могут быть получены с помощью генной инженерии, соматической гибридизации или химических методов. См., например, источник Kufer et al, цитированный выше; Cao and Suresh, Bioconjugate Chemistry, 9 (6), 635-644 (1998); и van Spriel et al., Immunology Today, 21 (8), 391-397 (2000), и цитированные в них ссылки.
[00117] Онколитический вирус, кодирующий антитело и содержащий антитело
[00118] Онколитический вирус предпочтительно инфицирует и убивает раковые клетки. Антитела согласно настоящему изобретению можно использовать вместе с онколитическими вирусами. Альтернативно, онколитические вирусы, кодирующие антитела согласно настоящему изобретению, можно вводить в тело человека.
[00119] Фармацевтические композиции
[00120] Согласно другому аспекту, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую одно или более антител согласно настоящему изобретению, представленных вместе с фармацевтически приемлемым носителем. Антитела могут быть дозированы раздельно, если композиция содержит более одного антитела. Композиция может необязательно содержать один или более дополнительных фармацевтически активных ингредиентов, таких как другое антитело или лекарственное средство, такое как противоопухолевое лекарственное средство.
[00121] Фармацевтическая композиция может содержать любое количество вспомогательных веществ. Вспомогательные вещества, которые можно использовать, включают носители, поверхностно-активные агенты, загустители или эмульгирующие агенты, твердые связующие вещества, дисперсионные или суспензионные добавки, солюбилизаторы, красители, ароматизаторы, покрытия, дезинтегрирующие агенты, скользящие вещества, подсластители, консерванты, изотонические агенты и их комбинации. Выбор и применение подходящих вспомогательных веществ обсуждаются в источнике Gennaro, ed., Remington: The Science and Practice of Pharmacy, 20th Ed. (Lippincott Williams & Wilkins 2003), описание которого включено в настоящую заявку посредством ссылки.
[00122] Предпочтительно, фармацевтическая композиция является подходящей для внутривенного, внутримышечного, подкожного, парентерального, спинномозгового или эпидермального введения (например, путем инъекция или инфузии). В зависимости от способа введения активный ингредиент может быть покрыт материалом для защиты от действия кислот и других природных условий, которые могут приводить к его инактивации. Фраза «парентеральное введение» при употреблении в настоящей заявке относится к способам введения, отличным от кишечного и местного введения, осуществляемым, как правило, путем инъекции, и включает, без ограничений, внутривенную, внутримышечную, интраартериальную, интратекальную, внутрисуставную, внутриглазничную, внутрисердечную, внтурикожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, интраартикулярную, субкапсулярную, субарахноидальную, внутриспинномозговую, эпидуральную и интрастернальную инъекцию и инфузию. Альтернативно, антитело согласно изобретению можно вводить с помощью непарентерального способа, такого как местное, эпидермальное или слизистое введение, например, интраназально, перорально, вагинально, ректально, подъязычно или местно.
[00123] Фармацевтические композиции могут быть представлены в виде стерильных водных растворов или дисперсий. Они также могут быть представлены в виде микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для введения высокой концентрации лекарственного средства.
[00124] Количество активного ингредиента, которое можно комбинировать с материалом носителя для получения единичной формы дозирования, варьирует в зависимости от субъекта, подлежащего лечению, и конкретного способа введения и в целом представляет собой такое количество композиции, которое производит терапевтический эффект. В целом, из ста процентов указанное количество варьирует от примерно 0,01% до примерно девяносто девяти процентов активного ингредиента, предпочтительно, от примерно 0,1% до примерно 70%, наиболее предпочтительно, от примерно 1% до примерно 30% активного ингредиента в комбинации с фармацевтически приемлемым носителем.
[00125] Режимы дозирования адаптируют с достижением оптимального желаемого ответа (например, терапевтического ответа). Например, можно вводить одну болюсную дозу, можно вводить несколько раздельных доз в течение периода времени или же дозу можно пропорционально снижать или повышать в соответствии с нуждами терапевтической ситуации. Особенно предпочтительным является предоставление композиции для парентерального введения в виде стандартной лекарственной формы для простоты введения и единообразия дозы. Стандартная лекарственная форма при употреблении в настоящей заявке относится к физически дискретным единицам, адаптированным в качестве единичных доз для субъектов, подлежащих лечению; каждая единица содержит заранее определенное количество активного ингредиента, рассчитанное для получения желаемого терапевтического эффекта, вместе с требуемым фармацевтическим носителем. Альтернативно, антитело можно вводить в виде лекарственной формы с замедленным высвобождением, в случае чего требуется меньшая частота введений.
[00126] Для введения антитела доза может варьировать от примерно 0,0001 до 100 мг/кг, обычно от 0,01 до 5 мг/кг массы тела хозяина. Например, дозы могут составлять 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела или в пределах диапазона 1-10 мг/кг. Пример режима лечения включает введение один раз в неделю, один раз в две недели, один раз в три недели, один раз в четыре недели, один раз в месяц, один раз в каждые 3 месяца или один раз в от трех до 6 месяцев. Предпочтительные режимы дозирования для антитела против PD-1 согласно изобретению включают 1 мг/кг массы тела или 3 мг/кг массы тела посредством внутривенного введение, где указанное антитело вводится с использованием одного из следующих протоколов дозирования: (i) каждые четыре недели для шести доз, затем каждые три месяца; (ii) каждые три недели; (iii) один раз доза 3 мг/кг массы тела с последующей дозой 1 мг/кг массы тела каждые три недели. В некоторых способах дозу адаптируют для достижения концентрации антитела в плазме крови, составляющей примерно 1-1000 мкг/мл, и в некоторых способах - примерно 25-300 мкг/мл.
[00127] «Терапевтически эффективная доза» антитела против PD-1 согласно изобретению предпочтительно приводит к снижению тяжести симптомов заболевания, повышению частоты и продолжительности свободных от симптомов заболевания периодов времени или предотвращению нарушения или недостаточности, вызванной поражением заболеванием. Например, для лечения субъектов, имеющих опухоль, «терапевтически эффективная доза» предпочтительно подавляет опухолевый рост по меньшей мере примерно на 20%, более предпочтительно, по меньшей мере примерно на 40%, более предпочтительно, по меньшей мере примерно на 60%, и еще более предпочтительно, по меньшей мере примерно на 80% по отношению к субъектам, не получающим лечение. Терапевтически эффективное количество терапевтического антитела может приводить к снижению размер опухоли или ослаблять другим образом симптомы у субъекта, который, как правило, представляет собой человека или может представлять собой другое млекопитающее.
[00128] Фармацевтическая композиция может представлять собой лекарственную форму с контролируемым высвобождением, включая импланты, трансдермальные пластыри и микроинкапсулирваонные системы доставки. Можно использовать биодеградируемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. См., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
[00129] Терапевтические композиции можно вводить с помощью медицинских устройств, таких как (1) устройства для безыгольной подкожной инъекции (см., например, патенты США № 5 399 163; 5 383 851; 5 312 335; 5 064 413; 4 941 880; 4 790 824; и 4 596 556); (2) насосы для микроинфузии (патент США № 4 487 603); (3) трансдермальные устройства (патент США № 4 486 194); (4) инфузионные приборы (патенты США № 4 447 233 и 4 447 224) и (5) осмотические устройства (патенты США № 4 439 196 и 4 475 196); описание которых включено в настоящую заявку посредством ссылки.
[00130] Согласно конкретным вариантам реализации изобретения, моноклональные антитела согласно изобретению могут быть представлены в форме для обеспечения соответствующего распределения in vivo. Например, для обеспечения прохождения терапевтического антитела согласно изобретению через гематоэнцефалический барьер указанное антитело может быть представлено в виде липосом, которые могут дополнительно содержать нацеливающие фрагменты для усиления селективного транспорта к конкретным клеткам или органам. См., например патенты США № 4 522 811; 5 374 548; 5 416 016; и 5 399 331; V. V. Ranade (1989) J. Clin.Pharmacol.29:685; Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038; Bloeman et al., (1995) FEBS Lett.357:140; M. Owais et al., (1995) Antimicrob. Agents Chemother. 39:180; Briscoe et al., (1995) Am. J. Physiol. 1233:134; Schreier et al., (1994) J. Biol. Chem. 269:9090; Keinanen and Laukkanen (1994) FEBS Lett. 346:123; and Killion and Fidler (1994) Immunomethods 4:273.
[00131] Применение и способы согласно изобретению
[00132] Антитела (композиции, биспецифичные антитела, иммуноконъюгаты и другие формы, включая онколитические вирусы) согласно настоящему изобретению имеют множество вариантов in vitro и in vivo применения, включая, например, усиление иммунных ответов путем блокирования PD-1. Антитела можно вводить в клетки в культуре, in vitro или ex vivo или субъектам, представляющим собой людей, например, in vivo, для усиления иммунитета в различных ситуациях. Соответственно, согласно одному аспекту, изобретение обеспечивает способ модифицирования иммунного ответа у субъекта, включающий введение указанному субъекту антитела или его антиген-связывающей части согласно изобретению таким образом, что происходит модифицирование иммунного ответ у указанного субъекта. Предпочтительно, происходит усиление, стимуляция или положительная регуляция иммунного ответа.
[00133] Предпочтительные субъекты включают пациентов, представляющих собой людей, нуждающихся в усилении иммунного ответа. Способы, в частности, подходят для лечения пациентов, представляющих собой людей, страдающих расстройством, которое можно лечить путем усиления иммунного ответа (например, опосредованного T-клетками иммунного ответа). Согласно конкретному варианту реализации изобретения, способы, в частности, подходят для лечения рака in vivo. Для достижения антиген-специфичного усиления иммунитета антитела против PD-1 можно вводить вместе с интересующим антигеном, или же указанный антиген уже может присутствовать у субъекта, подлежащего лечению (например, у субъекта, имеющего опухоль или вирус). Когда антитела против PD-1 вводят вместе с другими агентами, их можно вводить в любом порядке или одновременно.
134Учитывая способность антител против PD-1 согласно изобретению ингибировать связывание PD-1 с молекулами PD-L1 и/или PD-L2 и стимулировать антиген-специфичные T-клеточные ответы, изобретение также обеспечивает in vitro и in vivo способы применения антител для стимуляции, усиления или положительной регуляции антиген-специфичных T-клеточных ответов. Например, изобретение обеспечивает способ стимуляции антиген-специфичного T-клеточного ответа, включающий взаимодействие указанной T-клетки с антителом согласно изобретению таким образом, что происходит стимуляция антиген-специфичного T-клеточного ответа. Любой подходящий показатель антиген-специфичного T-клеточного ответа можно использовать для его измерения.
[00135] Неограничивающие примеры таких подходящих показателей включают повышенную пролиферацию T-клеток в присутствии антитела и/или повышенную продукцию цитокинов в присутствии антитела. Согласно предпочтительному варианту реализации изобретения, происходит стимуляция продукции интерлейкина-2 антиген-специфичной T-клеткой.
[00136] Изобретение также обеспечивает способ стимуляции иммунного ответа (например, антиген-специфичного T-клеточного ответа) у субъекта, включающий введение антитела согласно изобретению указанному субъекту таким образом, что происходит стимуляция иммунного ответа (например, антиген-специфичного T-клеточного ответа) у субъекта. Согласно предпочтительному варианту реализации изобретения, субъект представляет собой имеющего опухоль субъекта, и происходит стимуляция иммунного противоопухолевого ответа. Согласно другому предпочтительному варианту реализации изобретения, субъект представляет собой имеющего вирус субъекта, и происходит стимуляция иммунного противовирусного ответа.
[00137] Согласно другому варианту реализации, изобретение обеспечивает способы подавления роста опухолевых клеток у субъекта, включающие введение указанному субъекту антитела согласно изобретению таким образом, что у указанного субъекта происходит подавление роста опухоли. Согласно другому варианту реализации, изобретение обеспечивает способы лечения вирусной инфекции у субъекта, включающие введение указанному субъекту антитела согласно изобретению таким образом, что обеспечивается лечение вирусной инфекции у указанного субъекта.
[00138] Эти и другие способы согласно изобретению обсуждаются более подробно ниже.
[00139] Рак
[00140] Блокирование PD-1 антителами может приводить к усилению иммунного ответа против раковых клеток у пациента. Согласно одному аспекту, настоящее изобретение относится к лечению субъекта in vivo с использованием антитела против PD-1 таким образом, что происходит подавление роста раковых опухолей. Антитело против PD-1 можно использовать отдельно для подавления роста раковых опухолей. Альтернативно, антитело против PD-1 можно использовать вместе с другими иммуногенными агентами, используемыми в способах лечения рака, такими как онколитические вирусы или другие антитела, как описано ниже.
[00141] Соответственно, согласно одному варианту реализации, изобретение обеспечивает способ подавления роста опухолевых клеток у субъекта, включающий введение указанному субъекту терапевтически эффективного количества антитела против PD-1 или его антиген-связывающей части. Предпочтительно, антитело представляет собой мышиное, гибридное или гуманизированное антитело против PD-1.
[00142] Предпочтительные раковые опухоли, рост которых можно подавлять с использованием антител согласно изобретению, включают раковые опухоли, как правило, чувствительные к иммунотерапии. Неограничивающие примеры предпочтительных раковых опухолей для лечения включают меланому (например, метастатическую злокачественную меланому), почечный рак (например, светлоклеточную карциному), рак предстательной железы (например, гормонально-рефрактерную аденокарциному предстательной железы), рак молочной железы, рак толстой кишки и рак легких (например, немелкоклеточный рак легких), первичный или метастатический. Дополнительно, изобретение включает рефрактерные или рецидивирующие злокачественные опухоли, рост которых можно подавлять с использованием антител согласно изобретению.
[00143] Примеры других раковых опухолей, которые можно лечить с использованием способов согласно изобретению, включают рак кости, рак поджелудочной железы, рак кожи, рак головы и шеи, кожную или интраокулярную злокачественную меланому, рак матки, рак яичников, ректальный рак, рак анальной области, рак желудка, рак яичка, карциному фаллопиевых труб, карциному эндометрия, карциному шейки матки, вагинальную карциному, карциному вульвы, ходжкинскую болезнь, неходжкинские лимфомы, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркому мягких тканей, рак мочеточника, пениальный рак, хронические или острые лейкозы, включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфобластный лейкоз, солидные детские опухоли, лимфоцитарные лимфомы, рак мочевого пузыря, рак почки или мочеточника, карциному почечной лоханки, неоплазму центральной нервной системы (ЦНС), первичные лимфомы ЦНС, опухолевый ангиогенез, спинномозговую опухоль, глиому ствола мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, T-клеточные лимфомы, вызванные окружающими условиями раковые опухоли, включая опухоли, вызванные асбестом, и комбинации указанных раковых опухолей. Настоящее изобретение также применимо для лечения метастатических раковых опухолей, в особенности, метастатических раковых опухолей, экспрессирующих PD-1 (Iwai et al., (2005) Int. Immunol. 17:133-144).
[00144] Необязательно антитела против PD-1 можно объединять с иммуногенными агентами, такими как раковые клетки, очищенные опухолевые антигены (включая рекомбинантные белки, пептиды и молекулы углеводов) и клетки, трансфицированные генами, кодирующими иммуностимулирующие цитокины (He et al., (2004) J. Immunol. 173:4919-28). Неограничивающие примеры опухолевых вакцин, которые можно использовать, включают пептиды антигенов меланом, такие как пептиды gp100, MAGE-антигены, Trp-2, MART1, и/или тирозиназу или опухолевые клетки, трансфицированные для экспрессии цитокина ГМ-КСФ.
[00145] Блокирование PD-1, вероятно, является более эффективным при комбинировании с протоколом вакцинации. Было разработано много экспериментальных подходов для противоопухолевой вакцинации (см. Rosenberg, S., 2000, Development of Cancer Vaccines, ASCO Educational Book Spring: 60-62; Logothetis, C., 2000, ASCO Educational Book Spring: 300-302; Khayat, D. 2000, ASCO Educational Book Spring: 414-428; Foon, K. 2000, ASCO Educational Book Spring: 730-738; см. также Restifo, N. and Sznol, M., Cancer Vaccines, Ch. 61, pp. 3023-3043 in DeVita et al. (eds.), 1997, Cancer: Principles and Practice of Oncology, Fifth Edition). В одной из указанных стратегий вакцину готовят с использованием аутологичных или аллогенных опухолевых клеток. Было показано, что указанные клеточные вакцины являются наиболее эффективными, когда опухолевые клетки тарнсдуцированы для экспрессии ГМ-КСФ. Было показано, что ГМ-КСФ является сильным активатором презентирования антигена для опухолевой вакцинации (Dranoff et al. (1993) Proc. Natl. Acad. Sci. U.S.A. 90: 3539-43).
[00146] Исследование паттернов генной экспрессии и широкомасштабной генной экспрессии в различных опухолях привело к определению так называемых опухоль-специфичных антигенов (Rosenberg, SA (1999) Immunity 10: 281-7). Во многих случаях указанные опухоль-специфичные антигены являются дифференцировочными антигенами, экспрессируемыми в опухолях и в клетках, из которых возникла опухоль, например, антигены меланоцитов gp100, MAGE-антигены и Trp-2. Более важно, может быть показано, что многие из указанных антигенов могут являться мишенями опухоль-специфичных T-клеток, встречающихся в организме хозяина. Блокирование PD-1 можно использовать вместе с набором рекомбинантных белков и/или пептидов, экспрессируемых в опухоли, для запуска иммунного ответа на указанные белки. В норме иммунная система воспринимает указанные белки как собственные антигены и, таким образом, толерантна к ним. Опухолевый антиген может включать белок теломеразу, которая требуется для синтеза теломер хромосом и которая экспрессируется в более чем 85% раковых опухолей человека и только в ограниченном количестве соматических тканей (Kim et al. (1994) Science 266: 2011-2013). Указанные соматические ткани можно защищать от иммунной атаки различными способами. Опухолевый антиген также может представлять собой «нео-антигены», экспрессируемые в раковых клетках в результате соматических мутаций, которые приводят к изменению белковой последовательности или созданию белков слияния между двумя несвязанными последовательностями (т.е. bcr-abl в хромосоме Philadelphia), или идиотип из B-клеточных опухолей.
[00147] Другие опухолевые вакцины могут включать белки из вирусов, вовлеченных в раковые заболевания человека, таких как вирусы папилломы человека (HPV), вирусы гепатита (HBV и HCV) и вирус герпеса, ассоциированный с саркомой Капоши (KHSV). Другая форма опухоль-специфичного антигена, который можно использовать вместе с блокирваонием PD-1, представляет собой очищенные белки теплового шока (HSP), выделенные из самой опухолевой ткани. Указанные белки теплового шока содержат фрагменты белков из опухолевых клеток и являются высоко эффективными в доставке к антиген-презентирующим клеткам для индукции противоопухолевого иммунного ответа (Suot & Srivastava (1995) Science 269:1585-1588; Tamura et al. (1997) Science 278:117-120).
[00148] Дендритные клетки (ДК) представляют собой активные антиген-презентирующие клетки, которые можно использовать для запуска антиген-специфичных ответов. ДК могут быть получены ex vivo и нагружены различными белковыми и пептидными антигенами, а также опухолевыми клеточными экстрактами (Nestle et al. (1998) Nature Medicine 4: 328-332). ДК также можно трансдуцировать с помощью генетических способов для экспрессии указанных опухолевых антигенов. ДК также сливали непосредственно с опухолевыми клетками для иммунизации (Kugler et al., (2000) Nature Medicine 6:332-336). В качестве способа вакцинации ДК-иммунизацию можно эффективно объединять с блокированием PD-1 для активации более сильных противоопухолевых ответов.
[00149] Блокирование PD-1 также можно комбинировать со стандартными способами лечениями рака. Блокирование PD-1 можно эффективно комбинировать с химиотерапевтическими схемами лечения. Указанные примеры могут позволять снизить дозу вводимого химиотерапевтического реагента (Mokyr et al., (1998) Cancer Research 58: 5301-5304). Пример такой комбинации представляет собой антитело против PD-1 в комбинации с декабазином для лечения меланомы. Другой пример такой комбинации представляет собой антитело против PD-1 в комбинации с интерлейкином-2 (IL-2) для лечения меланомы. Научным обоснованием комбинированного применения блокирования PD-1 и химиотерапии является то, что клеточная гибель, являющаяся последствием цитотоксического действия большинства химиотерапевтических соединений, должна приводить к повышенному уровню опухолевого антигена в пути антигенной презентации. Другие комбинированные терапии, которые вместе с блокированием PD-1 могут приводить в синергическому эффекту, опосредованному клеточной гибелью, представляют собой облучение, хирургическое лечение и выключение эндокринной функции. Каждый из указанных протоколов создает источник опухолевого антигена в организме-хозяине. Ингибиторы ангиогенеза также можно комбинировать с блокированием PD-1. Ингибирование ангиогенеза приводит к гибели опухолевых клеток, обеспечивающей опухолевый антиген в пути антигенной презентации хозяина.
[00150] Блокирующие PD-1 антитела также можно использовать в комбинации с биспецифичными антителами, которые нацеливают экспрессирующие Fcα- или Fcγ-рецептор эффекторные клетки на опухолевые клетки (см., например, патенты США № 5 922 845 и 5 837 243). Биспецифичные антитела можно использовать для нацеливания на два отдельных антигена. Например, биспецифичные антитела против Fc-рецептора/противоопухолевого антигена (например, Her-2/neu) использовались для нацеливания макрофагов на опухолевые участки. Такое нацеливание может вызывать более эффективную активацию специфичных в отношении опухоли ответы. T-клеточное звено указанных ответов будет усиливаться в результате блокирования PD-1. Альтернативно, антиген может быть доставлен непосредственно к ДК путем применения биспецифичных антител, которые связываются с опухолевым антигеном и специфичным для дендритных клеток маркером клеточной поверхности.
[00151] Опухоли ускользают от иммунного надзора с помощью большого разнообразия механизмов. Многие из указанных механизмов можно преодолевать путем инактивации белков, которые экспрессируются опухолями и которые являются иммуносупрессорными. Указанные белки включают среди прочих TGF-β (Kehrl et al., (1986) J. Exp. Med. 163: 1037-1050), IL-10 (Howard & O'Garra (1992) Immunology Today 13: 198-200) и Fas-лиганд (Hahne et al. (1996) Science 274: 1363-1365). Антитела к каждому из указанных факторов можно использовать в комбинации с антителами против PD-1 для противодействия эффектам иммуносупрессорного агента и содействия противоопухолевым иммунным ответам хозяина.
[00152] Другие антитела, которые активируют иммунный ответ хозяина, можно использовать в комбинации с антителами против PD-1. Указанные антитела включают молекулы на поверхности дендритных клеток (ДК), которые активируют функцию ДК и презентацию антигенов. Антитела против CD40 способны эффективно возмещать активность T-хелперных клеток (Ridge et al. (1998) Nature 393: 474-478) и могут использоваться вместе с антителами против PD-1 (Ito et al. (2000) Immunobiology 201 (5) 527-40). Активирующие антитела против T-клеточных костимулирующих молекул, таких как CTLA-4 (например, патент США № 5 811 097), OX-40 (Weinberg et al. (2000) Immunol 164: 2160-2169), 4-1BB (Melero et al. (1997) Nature Medicine 3: 682-685 (1997) и ICOS (Hutloff et al. (1999) Nature 397: 262-266), также могут обеспечивать повышенный уровень T-клеточной активации.
[00153] Также существует несколько экспериментальных лечебных протоколов, которые включают ex vivo активацию и размножение антиген-специфичных T-клеток и адоптивный перенос указанных клеток реципиенту для стимуляции противоопухолевого эффекта антиген-специфичных T-клеток (Greenberg & Riddell (1999) Science 285: 546-51). Указанные способы также можно использовать для активации T-клеточных ответов на инфекционные агенты, такие как CMV. Ex vivo активация в присутствии антитела против PD-1 может повысить количество и активность адаптивно перенесенных T-клеток.
[00154] Инфекционные заболевания
[00155] Другие способы согласно изобретению используются для лечения пациентов, которые были подвержены воздействию конкретных токсинов или патогенов. Соответственно, другой аспект настоящего изобретения обеспечивает способ лечения инфекционного заболевания у субъекта, включающий введение указанному субъекту антитела против PD-1 или его антиген-связывающей части таким образом, что обеспечивается лечение субъекта от указанного инфекционного заболевания. Предпочтительно, антитело представляет собой гибридное или гуманизированное антитело.
[00156] Подобно применению опосредованного антителом блокирования PD-1 в отношении опухолей, как обсуждается выше, его можно использовать отдельно или в качестве адъюванта в комбинации с вакцинами для стимуляции иммунного ответа на патогены, токсины и собственные антигены. Примеры патогенов, для которых указанный терапевтический подход может быть, в частности, полезен, включают патогены, для которых на сегодняшний день не существует эффективной вакцины, или патогены, для которых стандартные вакцины не полностью эффективны. Указанные патогены включают, но не ограничиваются указанными, ВИЧ, вирус гепатита (A, B и C), вирус гриппа, вирус герпеса, Giardia, Malaria, Leishmania, Staphylococcus aureus, Pseudomonas aeruginosa. Блокирование PD-1, в частности, является полезным для борьбы с установленными инфекциями, вызванными агентами, такими как ВИЧ, презентирующими измененные антигены во время инфекции. Указанные новые эпитопы распознаются как чужеродные во время введения антитела против PD-1 человека, вызывая таким образом сильный T-клеточный ответ, который не подавляется отрицательными сигналами, опосредованными PD-1.
[00157] Некоторые примеры патогенных вирусов, вызывающих инфекции, поддающиеся лечению с помощью способов согласно изобретению, включают ВИЧ, гепатит (A, B или C), вирус герпеса (например, VZV, HSV-1, HAV-6, HSV-II и CMV, вирус Эпштейна-Барра), аденовирус, вирус гриппа, флавивирусы, эховирусы, риновирусы, вирус Каксаки, коронавирус, респираторный синцитиальный вирус, вирус свинки, ротавирус, вирус кори, вирус краснухи, парвовирус, вирус коровьей оспы, Т-лимофцитарный вирус человека (HTLV), вирус денге, папилломавирус, вирус моллюска, полиовирус, вирус бешенства, вирус Джона Каннингема (JC) и арбовирусный вирус энцефалита.
[00158] Некоторые примеры патогенных бактерий, вызывающих инфекции, поддающиеся лечению с помощью способов согласно изобретению, включают хламидии, рикетсии, микобактерии, стафилококки, стрептококки, пневмококки, менингококки и гонококки, клебсиеллу, протеус, серацию, псевдомонаду, легионеллу, бактерии, вызывающие дифтерию, сальмонеллу, палочковидную бактерию, бактерию, вызывающую холеру, столбняк, ботулизм, сибирскую язву, чуму, лептоспироз и болезнь Лайма.
[00159] Некоторые примеры патогенных грибов, вызывающих инфекции, поддающиеся лечению с помощью способов согласно изобретению, включают грибы Candida (albicans, krusei, glabrata, tropicalis, и т.д.), Cryptococcus neoformans, Aspergillus (fumigatus, niger, и т.д.), Genus Mucorales (mucor, absidia, rhizopus), Sporothrix schenkii, Blastomyces dermatitidis, Paracoccidioides brasiliensis, Coccidioides immitis и Histoplasma capsulatum.
[00160] Некоторые примеры патогенных паразитов, вызывающих инфекции, поддающиеся лечению с помощью способов согласно изобретению, включают Entamoeba histolytica, Balantidium coli, Naegleriafowleri, Acanthamoeba sp., Giardia lambia, Cryptosporidium sp., Pneumocystis carinii, Plasmodium vivax, Babesia microti, Trypanosoma brucei, Trypanosoma cruzi, Leishmania donovani, Toxoplasma gondii, Nippostrongylus brasiliensis.
[00161] Во всех из указанных выше способах блокирование PD-1 можно комбинировать с другими формами иммунотерапии, такими как лечение цитокинами (например, интерферонами, ГМ-КСФ, Г-КСФ, IL-2) или терапия биспецифичным антителом, которое обеспечивает усиленную презентацию опухолевых антигенов (см., например, Holliger (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak (1994) Structure 2:1121-1123).
[00162] Аутоиммунные реакции
[00163] Антитела против PD-1 могут вызывать и усиливать аутоиммунные ответы. В самом деле, индукция противоопухолевых ответов с помощью опухолевой клетки и пептидных вакцин показывает, что многие противоопухолевые ответы вовлекают аутореактивность (van Elsas et al. (2001) J. Exp. Med. 194:481-489; Overwijk, et al., (1999) Proc. Natl. Acad. Sci. U.S.A. 96: 2982-2987; Hurwitz, (2000), выше; Rosenberg & White (1996) J. Immunother Emphasis Опухоль Immunol 19 (1): 81-4). Таким образом, возможно рассматривать использование блокирования PD-1 вместе с различными собственными белками для разработки протоколов вакцинации для эффективной стимуляции иммунных ответов против указанных собственных белков для лечения заболевания.
[00164] Другие собственные белки также могут использоваться в качестве мишеней, например, IgE для лечения аллергии и астмы и TNFα для лечения ревматоидного артрита. Наконец, ответы антитела на различные гормоны могут быть вызваны применением антитела против PD-1. Ответы нейтрализующих антител на половые гормоны можно использовать для контрацепции. Ответ нейтрализующего антитела на гормоны и другие растворимые факторы, которые требуются для роста конкретных опухолей, также можно рассматривать как возможные мишени для вакцинации.
[00165] Аналогичные способы, как описано выше, для применения антитела против PD-1 можно использовать для индукции терапевтических аутоиммунных ответов для лечения пациентов, страдающих нарушенным накоплением других собственных антигенов, таких как цитокины, такие как TNFα и IgE.
[00166] Комбинированная терапия
[00167] Согласно другому аспекту, изобретение обеспечивает способы комбинированной терапии, в которых антитело против PD-1 (или его антиген-связывающую часть) согласно настоящему изобретению вводят совместно с одним или более дополнительными антителами, которые являются эффективными в стимулировании иммунных ответов, таким образом, для дополнительного усиления, стимуляции или положительной регуляции иммунных ответов у субъекта. Согласно одному варианту реализации, изобретение обеспечивает способ стимуляции иммунного ответа у субъекта, включающий введение указанному субъекту антитела против PD-1 и одного или более дополнительных иммуностимулирующих антител, таких как антитело против LAG-3, антитело против PD-L1 и/или антитело против CTLA-4, таким образом, что происходит стимуляция иммунного ответа у субъекта, например, для ингибирования опухолевого роста или для стимуляции противовирусного ответа. Согласно другому варианту реализации изобретения, субъекту вводят антитело против PD-1 и антитело против LAG-3. Согласно другому варианту реализации изобретения, субъекту вводят антитело против PD-1 и антитело против PD-L1. Согласно другому варианту реализации изобретения, субъекту вводят антитело против PD-1 и антитело против CTLA-4. Согласно другому варианту реализации изобретения, по меньшей мере одно дополнительное иммуностимулирующее антитело (например, антитело против PD-1, антитело против PD-L1 и/или антитело против CTLA-4) представляет собой человеческое антитело. Альтернативно, по меньшей мере одно дополнительное иммуностимулирующее антитело может представлять собой, например, гибридное или гуманизированное антитело (например, полученное из мыши антитело против LAG-3, антитело против PD-L1 и/или антитело против CTLA-4).
[00168] Согласно другому варианту реализации изобретения, изобретение обеспечивает способ лечения гиперпролиферативного заболевания (например, рака), включающий введение антитела против PD-1 и антитела против CTLA-4 субъекту. Согласно дополнительным вариантам реализации изобретения, антитело против PD-1 вводят в субтерапевтической дозе, антитело против CTLA-4 вводят в субтерапевтической дозе или оба указанных антитела вводят в субтерапевтической дозе. Согласно другому варианту реализации изобретения, настоящее изобретение обеспечивает способ изменения неблагоприятного явления, связанного с лечением гиперпролиферативного заболевания, с помощью иммуностимулирующего агента, включающий введение антитело против PD-1 и субтерапевтической дозы антитела против CTLA-4 субъекту. Согласно конкретным вариантам реализации изобретения, субъект представляет собой человека. Согласно другим вариантам реализации изобретения, антитело против CTLA-4 представляет собой человеческую последовательность моноклонального антитела 10D1 (описанного в публикации международной заявки согласно PCT WO 01/14424), и антитело против PD-1 представляет собой мышиную последовательность моноклонального антитела, такого как антитело против PD-1 C1H5, описанного в настоящей заявке. Другие антитела против CTLA-4, включенные согласно способам настоящего изобретения, включают, например, антитела, описанные в следующих источниках: публикация международной заявки WO 98/42752; публикация международной заявки WO 00/37504; патент США № 6 207 156; Hurwitz et al. (1998) Proc. Natl. Acad. Sci. USA 95(17):10067-10071; Camacho et al., (2004) J. Clin. Oncology 22(145): Abstract No. 2505 (antibody CP-675206); and Mokyr et al., (1998) Cancer Res. 58:5301-5304. Согласно конкретным вариантам реализации изобретения, антитело против CTLA-4 связывается с человеческим CTLA-4 со значением KD, составляющим 5x10-8 M или менее, связывается с человеческим CTLA-4 со значением KD, составляющим 1x10-8 M или менее, связывается с человеческим CTLA-4 со значением KD, составляющим 5x10-9 M или менее или связывается с человеческим CTLA-4 со значением KD, составляющим между 1x10-8 M и 1x10-10 M или менее.
[00169] Согласно другому варианту реализации изобретения, настоящее изобретение обеспечивает способ лечения гиперпролиферативного заболевания (например, рака), включающий введение антитела против PD-1 и антитела против LAG-3 субъекту.
[00170] Согласно другому варианту реализации изобретения, настоящее изобретение обеспечивает способ лечения гиперпролиферативного заболевания (например, рака), включающий введение антитела против PD-1 и антитела против PD-L1 субъекту.
[00171] Блокирование PD-1 и одного или более вторых антигенов-мишеней, таких как CTLA-4 и/или LAG-3 и/или PD-L1, антителами может приводить к усилению иммунного ответа на раковые клетки у пациента. Раковые опухоли, рост которых может подавляться при использовании антител согласно настоящему изобретению, включают раковые опухоли, как правило, чувствительные к иммунотерапии. Типичные примеры раковых опухолей для лечения с помощью комбинированной терапии согласно настоящему изобретению включают такие раковые опухоли, которые конкретно перечислены выше в обсуждении монотерапии антителами против PD-1.
[00172] Согласно конкретным вариантам реализации изобретения, комбинацию терапевтических антител, обсуждаемую в настоящей заявке, можно вводить одновременно в виде одной композиции в фармацевтически приемлемом носителе или одновременно в виде раздельных композиций, где каждое антитело представлено в фармацевтически приемлемом носителе. Согласно другому варианту реализации изобретения, комбинацию терапевтических антител можно вводить последовательно.
[00173] Более того, если комбинированная терапия предполагает последовательное введение более чем одной дозы, порядок указанного последовательного введения можно изменять или сохранять в каждый момент введения. Последовательные введения можно комбинировать с одновременными введениями или любой их комбинацией.
[00174] Опухоли ускользают от хозяйского иммунологического надзора с помощью большого разнообразия механизмов. Многие из указанных механизмов можно преодолевать с помощью инактивации белков, которые экспрессируются опухолями и которые являются иммуносупрессорными. Указанные механизмы включают среди прочих TGF-β (Kehrl et al., (1986) J. Exp. Med. 163: 1037-1050), IL-10 (Howard & O'Garra (1992) Immunology Today 13: 198-200), и Fas-лиганд (Hahne et al., (1996) Science 274: 1363-1365). В другом примере антитела к каждому из указанных элементов можно дополнительно комбинировать с комбинацией антител против PD-1 и против CTLA-4 и/или против LAG-3 и/или против PD-L1 для противодействия эффектам иммуносупрессорных агентов и содействия хозяйским противоопухолевым иммунным ответам.
[00175] Другие антитела, которые можно использовать для активации хозяйских иммунологических реакций можно дополнительно использовать вместе с комбинацией антител против PD-1 и против CTLA-4 и/или против LAG-3 и/или против PD-L1. Указанные антитела включают антитела против молекул на поверхности дендритных клеток, которые активируют функцию ДК и презентацию антигена. Антитела против CD40 (Ridge et al., выше) можно использовать вместе с комбинацией антител против PD-1 и против CTLA-4 и/или антител против LAG-3 и/или против PD-L1 (Ito et al., выше). Другие активирующие антитела к T-клеточным костимулирующим молекулам (Weinberg et al., выше, Melero et al., выше, Hutloff et al., выше) могут также обеспечивать повышенный уровень T-клеточной активации.
[00176] Как обсуждается выше, трансплантация костного мозга в настоящее время используется для лечения различных гематопоэтических опухолей. Комбинированное блокирование PD-1 и CTLA-4 и/или LAG-3 и/или PD-L1 можно использовать для повышения эффективности специфичных в отношении донорной трансплантированной опухоли T-клеток.
[00177] Некоторые экспериментaльные лечебные протоколы включают ex vivo активацию и размножение антиген-специфичных T-клеток и адоптивный перенос указанных клеток реципиентам антиген-специфичных T-клеток против опухоли (Greenberg & Riddell, выше). Указанные способы также можно применять для активации T-клеточных ответов на инфекционные агенты, такие как CMV. Ожидается, что ex vivo активация в присутствии антител против PD-1 и против CTLA-4 и/или против LAG-3 и/или против PD-L1 может повышать количество и активность адаптивно перенесенных T-клеток.
[00178] Согласно конкретным вариантам реализации, настоящее изобретение обеспечивает способ изменения неблагоприятного явления, связанного с лечением гиперпролиферативного заболевания (например, рака), с помощью иммуностимулирующего агента, включающий введение антитела против PD-1 и субтерапевтической дозы антитела против CTLA-4 и/или против LAG-3 и/или против PD-L1 субъекту. Например, способы согласно настоящему изобретению обеспечивают способ снижения встречаемости вызванного иммуностимулирующим терапевтическим антителом колита или диареи путем введения невсасывающегося стероида пациенту. Поскольку любой пациент, который получает иммуностимулирующее терапевтическое антитело, имеет риск развития колита или диареи, вызванных таким антителом, вся популяция указанных пациентов подходит для терапии в соответствии со способами согласно настоящему изобретению. Несмотря на то что введение стероидов использовалось для лечения воспалительного заболевания кишечника (ВЗК) и предотвращения обострений указанного ВЗК, оно не использовалось для предотвращения (снижения частоты встречаемости) ВЗК у пациентов, у которых не было диагностировано ВЗК. Значительные побочные эффекты, связанные со стероидами (даже невсасывающимися стероидами) ограничивали их профилактическое применение.
[00179] Согласно дополнительным вариантам реализации изобретения, комбинированное блокирование PD-1 и CTLA-4 и/или LAG-3 и/или PD-L1 (т.е. использование иммуностимулирующих терапевтических антител против PD-1 и CTLA-4 и/или антител против LAG-3 и/или против PD-L1) можно дополнительно комбинировать с применением любого невсасывающегося стероида. При употреблении в настоящей заявке «невсасывающийся стероид» представляет собой глюкокортикостероид, который подвергается активному метаболизму первого прохождения таким образом, что после его метаболизма в печени наблюдается низкая биодоступность указанного стероида, т.е. составляющая менее чем примерно 20%. Согласно одному варианту реализации изобретения, невсасывающийся стероид представляет собой будесонид. Будесонид представляет собой глюкокортикостероид местного действия, который активно подвергается метаболизму, главным образом, в печени, после перорального введения. ENTOCORT EC™ (Astra-Zeneca) представляет собой зависимую от pH и времени пероральную лекарственную форму будесонида, разработанную для оптимизации доставки лекарственного средства в подвздошную кишку на всю длину ободочной кишки. ENTOCORT EC™ одобрен в США для лечения болезни Крона от легкой до умеренной степени тяжести, затрагивающей подвздошную кишку и/или восходящую ободочную кишку. Обычная пероральная доза ENTOCORT EC™ для лечения болезни Крона составляет от 6 до 9 мг/день. ENTOCORT EC™ высвобождается в кишечнике до всасывания и удерживания в слизистой оболочке кишечника. После прохождения через кишечную слизистую ткань-мишень ENTOCORT EC™ подвергается активному метаболизму с помощью системы цитохрома P450 в печени до образования метаболитов, обладающих незначительной глюкокортикостероидной активностью. Таким образом, его биодоступность является низкой (примерно 10%). Низкая биодоступность будесонида приводит к улучшенному терапевтическому отношению по сравнению с другими глюкокортикостероидами, подвергающимися менее интенсивному метаболизму первого прохождения. Будесонид вызывает меньшие неблагоприятные эффекты, включая менее выраженное подавление гипоталамо-гипофизарной системы, по сравнению с кортикостероидами системного действия. Однако длительное введение ENTOCORT EC™ может приводить к системным эффектам глюкокортикостероидов, таким как гиперкортицизм и угнетение функции надпочечников. СМ. PDR 58th ed. 2004; 608-610.
[00180] Согласно другим дополнительным вариантам реализации изобретения, комбинированное блокирование PD-1 и CTLA-4 и/или LAG-3 и/или PD-L1 (т.е. использование иммуностимулирующих терапевтических антител против PD-1 и против CTLA-4 и/или антител против LAG-3 и/или против PD-L1) совместно с использованием невсасывающегося стероида можно дополнительно комбинировать с применением салицилата. Салицилаты включают 5-ASA агенты такие как, например: сульфасалазин (AZULFIDINE™, Pharmacia & UpJohn); ольсалазин (DIPENTUM™, Pharmacia & UpJohn); балсалазид (COLAZAL™, Salix Pharmaceuticals, Inc.); и месаламин (ASACOL™, Procter & Gamble Pharmaceuticals; PENTASA™, Shire US; CANASA™, Axcan Scandipharm, Inc.; ROWASA™, Solvay).
[00181] В соответствии со способами согласно настоящему изобретению, салицилат, вводимый в комбинации с антителами против PD-1 и против CTLA-4 и/или антителами против LAG-3 и/или против PD-L1 и невсасывающимся стероидом, может включать любое перекрывающееся или последовательное введение салицилата и невсасывающегося стероида для снижения частоты встречаемости колита, вызванного иммуностимулирующим антителом. Таким образом, например, способы снижения частоты встречаемости колита, вызванного иммуностимулирующим антителом в соответствии с настоящим изобретением, включают одновременное или последовательное введение салицилата и невсасывающегося стероида (например, салицилат вводят через 6 часов после введения невсасывающегося стероида) или любую их комбинацию. Дополнительно, в соответствии с настоящим изобретением, салицилат и невсасывающийся стероид можно вводить с помощью одного и того же способа (например, оба вещества вводят перорально) или с помощью различных способов (например, салицилат вводят перорально и невсасывающийся стероид вводят ректально), которые могут отличаться от способа (способов), используемых для введения комбинации антител против PD-1 и против CTLA-4 и/или антител против LAG-3 и/или против PD-L1.
[00182] Настоящее изобретение дополнительно проиллюстрировано с помощью следующих далее примеров, которые, как предполагается, дополнительно не ограничивают настоящее изобретение. Содержание всех фигур и всех ссылок, последовательностей согласно базе данных Genbank, патентов и опубликованных патентных заявок, цитированных на протяжении настоящей заявки, специально включено в настоящую заявку посредством ссылки.
[00183] Примеры
[00184] Пример 1. Создание мышиных моноклональных антител против PD-1 с использованием гибридомной технологии
[00185] Иммунизация
[00186] Мышей иммунизировали в соответствии со способом, как описано в источнике E Harlow, D. Lane, Antibody: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1998. Рекомбинантный человеческий белок PD-1 с меткой человеческого IgG1-Fc на C-конце (Acro biosystems, #PD-1-H5257, содержащий внеклеточный домен, AA Leu 25 - Gln 167) использовали в качестве иммуногена. Человеческий белок PD-1-his (Sino biological, #10377-H08H) применяли для определения антисывороточного титра и для скрининга гибридом, секретирующих антиген-специфичные антитела.
[00187] В частности, каждому животному инъецировали 25 мкг человеческого белка PD1-Fc в полном адъюванте Фрейнда (Sigma, Сент-Луис, Миссури, США) и затем проводили стимуляцию от 2 до 3 раз путем инъекции 25 мкг человеческого белка PD1-Fc в неполном адъюванте Фрейнда (Sigma, Сент-Луис, Миссури, США) в зависимости от антисывороточного титра. Антисывороточный титр измеряли с помощью скрининга на основе ИФА с использованием рекомбинантного человеческого белка PD1-his. Вкратце, разведенную сыворотку (60 мкл) добавляли в каждую лунку и инкубировали при 37°C в течение 40 минут. Планшеты затем промывали 4 раза, HRP-конъюгированные козьи антитела против IgG мыши (Jackson Immuno research, Cat#115-036-071) использовали для выявления и оптическую плотность (OD), характеризующую связывание, наблюдали при 450 нм. Животным, имеющим высокие титры, вводили конечную дополнительную дозу путем внутрибрюшинной инъекции до слияния гибридомы.
[00188] Слияние гибридомы и скрининг
[00189] Клетки мышиной клеточной линии миеломы (SP2/0-Ag14, ATCC#CRL-1581) культивировали до достижения фазы логарифмического роста непосредственно перед слиянием. Клетки селезенки из иммунизированных мышей получали в стерильных условиях и сливали с клетками миеломы в соответствии с методом, описанным в источнике Kohler G, and Milstein C, "Continuous cultures of fused cells secreting antibody of predefined specificity", Nature, 256: 495-497(1975). Слитые «гибридные клетки» затем распределяли в 96-луночные планшеты в среде DMEM/20% ЭБС/HAT. Выжившие колонии гибридом наблюдали под микроскопом через семь-десять дней после слияния. Через две недели супернатант из каждой лунки подвергали скринингу на основе ИФА с использованием рекомбинантного белка PD1-his человека. Вкратце, ИФА-планшеты покрывали 60 мкл человеческого PD1-his (Sino biological, #10377-H08H, 2,0 мкг/мл в PBS) в течение ночи при 4°C. Планшеты промывали 4 раза PBS-T и блокировали 200 мкл блокирующего буфера (5% обезжиренное молоко в PBS-T). Разведенный супернатант гибридомы (60 мкл) добавляли в каждую лунку и инкубировали при 37°C в течение 40 минут. Планшеты затем промывали 4 раза, конъюгированные с HRP козьи антитела против IgG мыши (Jackson Immuno research, Cat#115-036-071) использовали для выявления и OD, характеризующую связывание, наблюдали при 450 нм. Гибридому с положительным результатом, секретирующую антитело, которое связывается с человеческим PD1-his, затем выбирали и переносили в 24-луночные планшеты. Клоны гибридомы, продуцирующие антитела, которые показали высокое специфичное связывание и блокирующую PD1/PDL1 активность, субклонировали и антитела, продуцированные указанными субклонами, очищали с помощью аффинной хроматографии на белке А. Вкратце, колонку, содержащую белок A-сефарозу (от компании Bestchrom (Shanghai) Biosciences, Cat#AA0273), промывали с использованием буфера PBS в 5-10 колоночных объемах. Клеточные супернатанты пропускали через колонки и затем колонки промывали с использованием буфера PBS до тех пор, пока абсорбция белка не достигала базового уровня. Элюирование колонки проводили элюирующим буфером (0,1 М глицин-HCl, pH 2,7), который сразу собирали в пробирки на 1,5 мл, содержащие нейтрализующий буфер (1 M Трис-HCl, pH 9,0). Фракции, содержащие IgG, объединяли и диализовали в PBS в течение ночи при 4°C.
[00190] Пример 2. Определение аффинности мышиных моноклональных антител против PD-1 с использованием метода поверхностного плазмонного резонанса BIACORE
[00191] Осуществляли характеристику очищенных мышиных моноклональных антител (mAb) против PD-1, созданных в Примере 1, содержащих последовательности вариабельных участков тяжелой/легкой цепи, как описано в Таблице 1, выше, и последовательности константных участков тяжелой/легкой цепи SEQ ID NO: 129 и 130 соответственно, на предмет аффинности и кинетики связывания с помощью системы Biacore T200 (GE healthcare, Питтсбург, Пенсильвания, США).
[00192] Вкратце, козьи антитела против IgG мыши (GE healthcare, Cat#BR100839, набор для захвата человеческих антител) ковалентно связывали с чипом CM5 (покрытым карбоксиметилдекстраном чипом) через первичные амины с использованием стандартного набора для сопряжения аминов (GE healthcare, Питтсбург, Пенсильвания, США), предложенного Biacore. Непрореагировавшие фрагменты на поверхности биосенсора блокировали этаноламином. Затем очищенные антитела против PD-1 и ниволумаб (OPDIVO®) в концентрации 66,7 нМ наносили на чип при скорости потока 10 мкл/мин. Затем рекомбинантный человеческий белок PD-1-his (Sino biological, #10377-H08H) или белок PD-1-his макака-крабоеда (Acro biosystems, #PD-1-C5223) в буфере HBS EP (предложенном Biacore) наносили на чип при скорости потока 30 мкл/мин. За кинетикой ассоциации антиген-антитело наблюдали в течение 2 минут и за кинетикой диссоциации наблюдали в течение 10 минут. Кривые ассоциации и диссоциации подводили к 1:1 модели связывания Ленгмюра с использованием программы оценки BIA.
[00193] Значения ka, kd и KD были определены и показаны в Таблице 2, ниже.
Таблица 2. Кинетика связывания мышиных моноклональных антител против PD-1 с PD-1 человека или макака-крабоеда по результатам анализа Biacore
[00194] Антитела согласно настоящему изобретению специфично связывались с PD-1 человека при более низком значении KD по сравнению с ниволумабом, что указывает на их более высокую аффинность в отношении PD-1 человека.
[00195] Пример 3. Активность связывания мышиных моноклональных антител против PD-1
[00196] 96-луночные микропланшеты покрывали козьими антителами против Fcγ-фрагментов IgG мыши в концентрации 2 мкг/мл (Jackson Immuno Research, #115-006-071,100 мкл/лунка) в PBS и инкубировали в течение ночи при 4°C. Планшеты промывали 4 раза промывочным буфером (PBS+0,05% Tween-20, PBS-T) и затем блокировали блокирующим буфером в концентрации 200 мкл/лунка (5% (по массе к объему) обезжиренное молоко в PBS-T ) в течение 2 часов при 37°C. Планшеты снова промывали и инкубировали с очищенными антителами против PD-1 из Примера 1 в концентрации 100 мкл/лунка и ниволумабом (0,004-66,7 нМ, 5-кратные серийные разведения в 2,5%-ом обезжиренном молоке в PBS-T) в течение 40 минут при 37°C и затем снова промывали 4 раза. Планшеты, содержащие захваченные антитела против PD-1, инкубировали с меченным биотином белком PD-1-Fc человека (SEQ ID NO: 100, 60 нМ в 2,5%-ом обезжиренном молоке в PBS-T, 100 мкл/лунка) в течение 40 минут при 37°C, промывали 4 раза и инкубировали с конъюгированной со стрептавидином пероксидазой хреннна (HRP, 1:10000 разведение в PBS-T, Jackson Immuno Research, #016-030-084, 100 мкл/лунка) в течение 40 минут при 37°C. После конечной промывки планшеты инкубировали в присутствии ™B-субстрата для ИФА (100 мкл/лунка, Innoreagents). Реакцию останавливали через 15 минут при 25°C с помощью 1М H2S04 в концентрации 50 мкл/лунка, и абсорбцию считывали при 450 нм. Данные анализировали с использованием программы Graphpad Prism и регситировали значения EC50.
[00197] Результаты суммированы в Таблице 3, ниже.
Таблица 3. Активность связывания антител против PD-1 с PD-1 человека
[00198] Результат показал, что антитела согласно настоящему изобретению специфично связывались с PD-1 человека, при этом несколько клонов имели более низкие значения EC50 по сравнению с ниволумабом.
[00199] Пример 4. Функциональные анализы блокирования с использованием ИФА и репортерных анализов
[00200] 4,1 ИФА-анализ блокирования лиганда
[00201] Способность антитела против PD-1 согласно настоящему изобретению блокировать взаимодействие PD-1-PD-L1 измеряли с использованием конкурентного ИФА-анализа. Вкратце, PD-L1-Fc белки человека (SEQ ID NO:101) наносили на 96-луночные микропланшеты в концентрации 2 мкг/мл PBS и инкубировали в течение ночи при 4°C. На следующий день планшеты промывали промывочным буфером (PBS+0,05% Tween-20, PBS-T) и блокировали 5% обезжиренным молоком в PBS-T в течение 2 часов при 37°C. Планшеты затем снова промывали с использованием промывочного буфера.
[00202] Готовили разведения антител против PD-1 согласно настоящему изобретению или ниволумаба (начиная с концентрации 100 нМ с 4-кратными серийными разведениями) в растворе меченного биотином PD1-Fc человека (SEQ ID NO: 100, 10 нМ в 2,5% обезжиренном молоке в PBS-T) и инкубировали при комнатной температуре в течение 40 минут, затем смеси антитела/PD-1-Fc-биотин (100 мкл/лунка) добавляли в покрытые PD-L1 планшеты. После инкубирования при 37°C в течение 40 минут планшеты промывали 4 раза с использованием промывочного буфера. Затем добавляли конъюгированную со стрептавидином HRP в концентрации 100 мкл/лунка и инкубировали в течение 40 минут при 37°C для выявления меченного биотином PD-1 человека, связанного с PD-L1. Планшеты снова промывали с использованием промывочного буфера. Наконец, добавляли ™B-субстрат и реакцию останавливали с использованием 1М H2SO4 и абсорбцию считывали при 450 нм. Данные анализировали с использованием программы Graphpad Prisme и регистрировали значения IC50.
[00203] 4,2 ИФА-анализ блокирования стандартного образца
[00204] Способность антител против PD-1 согласно настоящему изобретению блокировать стандартный образец измеряли связывание ниволумаб-PD-1 человека с использованием конкурентного ИФА-анализа. Вкратце, ниволумаб наносили на 96-луночные микропланшеты в концентрации 2 мкг/мл в PBS и инкубировали в течение ночи при температуре 4°C. На следующий день планшеты промывали промывочным буфером и блокировали 5%-ым обезжиренным молоком в PBS-T в течение 2 часов при 37°C. Во время блокирования меченный биотином PD-1-Fc человека (SEQ ID NO:100, 10 нМ в 2,5%-ом обезжиренном молоке в PBS-T) смешивали с каждым из тестируемых антител (137 пкМ-100 нМ, 3-кратные серийные разведения) и инкубировали в течение 40 минут при 25°C. После промывки смеси PD-1/антитело (100 мкл /лунка) добавляли в планшеты, покрытые ниволумабом, и инкубировали в течение 40 минут при 37°C. Планшеты снова промывали промывочным буфером и затем добавляли стрептавидин (SA)-HRP в концентрации 100 мкл/лунка и инкубировали в течение 40 минут при 37°C для выявления меченного биотином PD-1 человека, связанного с Opdivo®. Планшеты в конечном итоге промывали с использованием промывочного буфера. Добавляли ™B-субстрат и реакцию останавливали с использованием 1М H2SO4 и абсорбцию считывали при 450 нм. Данные анализировали с использованием программы Graphpad Prism и регистрировали значения IC50.
[00205] 4,3 Клеточные функциональные анализы
[00206] Активность блокирования антителами взаимодействия PD-1/PD-L1 на клеточной мембране оценивали с использованием клеточного репортерного анализа. Указанный анализ включал две генноинженерные клеточных линии - PD-1-эффекторную клеточную линию (Genscript, GS-J2/PD-1), стабильно экспрессирующую PD-1 человека и репортерный ген люциферазы, запускаемый репортерным элементом NFAT (NFAT-RE), и PD-L1-клеточную линию (Genscript, GS-C2/PD-L1, клетки APC), стабильно экспрессирующую человеческий PD-L1 и сконструированный белок клеточной поверхности-антигенный пептид/главный комплекс гистосовместимости (major histocompatibility complex, MHC). Когда указанные две клеточные линии сокультивировали, происходиол ингибирование опосредованной T-клеточным рецептором (T-cell receptor, TCR) экспрессии люциферазы в PD-1-эффекторных клетках (через путь NFAT) в результате взаимодействия PD-1/PD-L1.
[00207] Клеточный функциональный анализ проводили, как следует далее. Вкратце, клетки PD-L1 в фазе логарифмического роста высевали в 384-луночные планшеты для клеточных культур в плотности 5*105/мл. На следующий день готовили разведение антител против PD-1 согласно настоящему изобретению или ниволумаба (начиная с 333,3 нМ, 5-кратные серийные разведения) в аналитическом буфере (RPMI 1640+1%ЭБС). Тем временем среду от PD-L1-клеток в 384-луночных планшетах сливали и затем в 384-луночные планшеты для клеточных культур добавляли антитела против PD-1 в разведениях (20 мкл/лунка) и PD-1-эффекторные клетки (в плотности 6,25*105/мл, 20 мкл/лунка). После сокультивирования при 37°C в течение шести часов планшеты доставали из инкубатора и люминесценцию для каждой лунки считывали в соответствии с инструкциями производителя с использованием системы люциферазного анализа One-Glo (Promega, #E6120). Кривые зависимости эффекта от дозы анализировали с использованием программы Graphpad Prism и регистрировали значения EC50.
[00208] Результаты трех анализов суммированы в Таблице 4, ниже.
Таблица 4. Способность антител против PD-1 блокировать взаимодействие PD-1/PD-L1
[00209] Можно видеть, что антитела согласно настоящему изобретению были способны блокировать взаимодействие PD-1 человека/PD-L1 человека, при этом несколько клонов имело более низкие значения EC50 или IC50 по сравнению с ниволумабом.
[00210] Данные также показали, что антитела согласно настоящему изобретению были способны блокировать взаимодействие PD-1 человека/ниволумаб, где антитела из клона 2G2 обеспечивали частичное блокирование, указывая на то, что они связывались с тем же или сходным эпитопом, что и ниволумаб.
[00211] Пример 5. Создание и характеристика гибридных антител
[00212] Вариабельные домены тяжелой и легкой цепи мышиного mAb против PD1 C1E1 клонировали в рамке считывания с константными участками тяжелой цепи IgGl человека и легкой каппа-цепи человека соответственно. Вариабельный участок тяжелой цепи и вариабельный участок легкой цепи имели аминокислотные последовательности, указанные в SEQ ID NO: 67 и 80 соответственно, тогда как аминокислотные последовательности константных участков тяжелой цепи IgGl человека и легкой каппа-цепи человека представлены в SEQ ID NO: 97 и 98 соответственно. Активность полученных гибридных антител подтверждали с помощью ИФА-анализа связывания с захватом, конкурентного ИФА и клеточного функционального репортерного анализа в соответствии с протоколами, описанными в предшествующих примерах. Данные показали, что гибридное антитело C1E1 обладало активностями, сравнимыми с активностями стандартного образца (OPDIVO®), как показано в Таблице 5, ниже.
Таблица 5. Связывание и функциональная активность гибридных антител
[00213] Пример 6. Гуманизирование мышиного моноклонального антитела против PD-1 C1E1
[00214] Мышиное антитело против PD1 C1E1 выбирали для гуманизирования и дополнительных исследований. Гуманизирование мышиного антитела проводили с использованием хорошо установленного метода прививки CDR, как описано подробно ниже.
[00215] Для выбора акцепторных каркасов для гуманизирования мышиного антитела C1E1 проводили исследование последовательности вариабельного участка легкой и тяжелой цепи мышиного C1E1 с помощью BLAST против базы данных генов человеческих иммуноглобулинов. Человеческие зародышевые линии IGVH и IGVK с максимальной гомологией к мышиному C1E1 были выбраны в качестве акцепторных каркасов для гуманизирования. CDR-участки вариабельного участка тяжелой/легкой цепи мышиного антитела встраивали в выбранные каркасы и остаток (остатки) в указанных каркасах дополнительно подвергали мутированию для получения большего количества вариабельных участков тяжелой цепи/легкой цепи, являющихся кандидатами.
[00216] Векторы, содержащие нуклеотидные последовательности, кодирующие вариабельные участки тяжелой цепи/легкой цепи гуманизированного C1E1 и константные участки тяжелой цепи IgGl человека и легкой каппа-цепи человека временно трансфицировали в 50 мл суспензионной клеточной культуры 293F с отношением конструктов, содержащих легкую цепь, к конструктам, содержащим тяжелую цепь, составляющим 60%:40%, в присутствии 1,2 мг/мл PEI. Клеточные супернатанты собирали через шесть дней во встряхиваемые колбы, откручивали для осаждения клеток и фильтровали через фильтр с размером пор 0,22 мкм до отделения IgG. Антитела очищали с помощью аффинной хроматографии на основе белка А. Вкратце, колонку, заполненную белок А-сефарозой (от компании Bestchrom (Shanghai) Biosciences, Cat#AA0273) промывали с использованием буфера PBS в 5-10 колоночных объемах. Клеточные супернатанты пропускали через колонки и затем колонки промывали с использованием буфера PBS до тех пор, пока абсорбция белка не достигала базового уровня. Элюирование колонок проводили с помощью элюирующего буфера (0,1 М глицин-HCl, pH 2,7), который сразу собирали в пробирки на 1,5 мл, содержавшие нейтрализующий буфер (1 M Трис-HCl, pH 9,0). Фракции, содержавшие IgG, объединяли и диализовали в PBS в течение ночи при 4°C.
[00217] В целом были получены 10 гуманизированных антител, 8 из которых были дополнительно охарактеризованы, а именно huC1E1-V1 - huC1E1-V7 и huC1E1-V10. Аминокислотные последовательности вариабельных участков тяжелой цепи/легкой цепи 8 указанных антител суммированы в Таблице 1, выше, и последовательности константных участков тяжелой цепи IgGl человека и легкой каппа-цепи человека указаны в SEQ ID NO: 97 и 98 соответственно.
[00218] Аффинность связывания huC1E1-V1 - huC1E1-V7 с PD1 человека оценивали с помощью метода BIACORE, как описано в Примере 2, и значения KD анализа аффинности суммированы в Таблице 6, ниже. Активность связывания антитела huC1E1-V10 с PD1 человека оценивали с помощью ИФА-анализа связывания с захватом, как описано в Примере 3, в котором для захвата IgG использовали козье захватывающие антитело против IgG-Fab человека (Jackson ImmunoResearch, Cat#109-005-097), и значения EC50 анализа связывания суммированы в Таблице 7. Все 8 гуманизированных антител обладали аффинностью, сравнимой с гибридным антителом C1E1.
Таблица 6. Аффинность гуманизированных mAb C1E1
Таблица 7. Активность связывания гуманизированного mAb C1E1
(EC50, нМ)
[00219] Гуманизированное антитело huC1E1-V10 затем исследовали на предмет аффинности в отношении PD1 человека и макака-крабоеда с помощью анализа Biacore и с помощью ИФА-анализа связывания с захватом, а также исследовали на предмет функциональной активности с помощью конкурентного ИФА и клеточного репортерного анализа в соответствии с протоколами, описанными в Примерах 2-4. Как показано в Таблице 8, huC1E1-V10 показал сравнимые с гибридным антителом C1E1 значения in vitro активности.
Таблица 8. Связывание и функциональная активность гуманизированного mAb C1E1
[00220] Пример 7. Физико-химические свойства гуманизированного антитела C1E1
[00221] Физико-химические свойства гуманизированного антитело против PD1 huC1E1-V10 проверяли с использованием белкового анализа теплового сдвига, метода cIEF, метода SEC и метода замораживания-оттаивания, как описано ниже.
[00222] Белковый анализ теплового сдвига для определения Tm
[00223] Термостабильность гуманизированного антитела против PD1 huC1E1-V10 измеряли с использованием набора для анализа белковой стабильности на основе теплового сдвига GloMelt™ (Biotium, Cat#33022-T, lot#: 181214). Вкратце, красителю GloMelt™ позволяли оттаивать и достигать комнатной температуры. Ампулу, содержащую краситель, вортексировали и центрифугировали. 10-кратный раствор красителя готовили путем добавления 5 мкл 200-кратного раствора красителя к 95 мкл PBS. 2 мкл 10-кратного раствора красителя и 10 мкг антитела добавляли в общей реакционный объем 20 мкл. Устанавливали параметры для запуска программы анализа кривой плавления, подробно описанные в Таблице 9. Пробирки быстро центрифугировали и помещали в амплификатор для ПЦР в реальном времени (Roche, LightCycler 480 II). Результаты анализировали с использованием программы Microsoft Excel 2010.
Таблица 9. Параметры для программы анализа кривой плавления
[00224] Метод cIEF для определения pI
[00225] Капиллярное изоэлектрическое фокусирование (cIEF) использовали для определения pI и гетерогенности заряда гуманизированного антитела против PD1 huC1E1-V10. Вкратце, 35 мкл 0,1% MC, 4 мкл pH3-10 Pharmlyte, 2 мкл Arg в концентрации 0,5 моль/л, 1 мкл Mark-7,05 и 1 мкл Mark-9,99 добавляли в пробирку, содержащую 20 мкг антитела в PBS, затем добавляли ddH2O до 100 мкл. Программа электрофореза состояла из двух фаз: изначальное разделение в течение 1 минуты при 1500 В с последующими разделением в течение 6 минут при 3000 В. Через 5 мин экспозиции для выявления данные анализировали с использованием программы Maurice™Compass™ (Proteinsimple Inc., США).
[00226] Метод SEC для определения агрегирования
[00227] Процент мономеров оценивали с помощью эксклюзионной по размеру хроматографии (size exclusion chromatography, SEC) (Agilent Technologies, 1260 Infinity II). Антитела, которые содержались в водной подвижной фазе, пропускали через пористую смолу, представляющую собой неподвижную фазу, упакованную в колонку. Время удерживания в колонке представляло собой функцию гидродинамического размера белка и размера пор в упакованном слое смолы. Меньшие по размеру молекулы могут проникать в более маленькие поры в смоле и удерживаются дольше по сравнению с молекулами большего размера. При элюировании из колонки белки выявляли с помощью УФ-абсорбции. Подвижная фаза представляла собой фосфатно-солевой буферный раствор. За абсорбцией наблюдали при 280 нм. Скорость потока составляла 0,8 мл/мин. Вводимый объем составлял 40 мкл образца в концентрации 1 мг/мл. Температура колонки представляла собой комнатную температуру. Температура автодозатора составляла 2-8°C. Общее время прогона составляло 25 минут.
[00228] Метод замораживания-оттаивания для определения стабильности
[00229] Растворы антитела в концентрации 1 мг/мл в лекарственной форме (формах), представляющей собой 1xPBS (содержащий 137 ммоль/л NaCl, 2,7 ммоль/л KCl, 10 ммоль/л Na2HPO4,12H2O, 2 ммоль/L KH2PO4), замораживали при -80°C по меньшей мере на 24 часа и затем оттаивали при комнатной температуре в течение 30-60 минут. Цикл замораживания и оттаивания повторяли 5 раз для каждого образца. После конкретных циклов замораживания-оттаивания, например, второго, третьего и четвертого, часть раствора отбирали для анализа с помощью SEC до повторного замораживания.
[00230] Нестабильность, приводящую к денатурации, оценивали с помощью набора для измерения белковой стабильности на основе теплового сдвига GloMelt™. Как показано на Фигуре 1, для гуманизированного антитела против PD1 huC1E1-V10 наблюдалась высокая температура плавления (Tm), как правило, 78°C, также наблюдался минорный пик при 69,5°C. Анализ huC1E1-V10 с помощью cIEF показал, что % содержание главной изоформы составляло примерно 81,5%, % содержание кислотных видов составляло примерно 11,0%, и % содержание основных видов составляло примерно 7,5%. Значения pI рассчитывали с использованием программы Maurice™Compass™. Значение pI для антитела huC1E1-V10 составляло 8,36. Значение pI гуманизированного антитела huC1E1-V10 значительно превышало 7, предполагая, таким образом, что указанное антитело, будет иметь значительный положительный заряд при нейтральном pH. SEC-анализ huC1E1-V10 показал, что % содержание мономеров составляло 100,0% (см. Фиг. 2). Устойчивость к замораживанию и оттаиванию оценивали с помощью эксклюзионной по размеру хроматографии (SEC). Для антитела huC1E1-V10 не было показано очевидного повышения уровня агрегирования с течением времени, также не наблюдалось осаждения или помутнения после 5 повторных замораживаний и оттаиваний.
[00231] Пример 8. In Vivo противоопухолевая эффективность мышиного антитела C1E1
[00232] In vivo противоопухолевую активность мышиного антитела C1E1 оценивали на мышах NCG. Вкратце, мышам NCG подкожно инъецировали 4x106 клеток меланомы человека A375, смешанных с 5x105 реактивированных опухолью мононуклеарных клеток периферической крови (МКПК) в правую подмышечную область на 1 день. Объем опухолей измеряли два раза в неделю в течение трех недель с использованием электронного штангенциркуля и рассчитывали как (длина × ширина2)/2. Двадцать четыре мыши, имеющие опухоли, выбирали и рандомизировали в четыре группы на 14 день. Животным вводили внутрибрюшинно наполнитель (5% раствор глюкозы), мышиное антитело C1E1, мышиное антитело C1E1-Fab и ниволумаб соответственно в дозе 3 мг/кг на 14, 21 и 28 день. Мышиное антитело C1E1 содержало мышиный вариабельный участок тяжелой цепи и человеческий константный участок IgG1, а также мышиный вариабельный участок легкой цепи и человеческий константный участок Ck, имеющие последовательности, представленные в SEQ ID NO: 67, 97, 80 и 98 соответственно, тогда как мышиный C1E1-Fab содержал мышиный вариабельный домен тяжелой цепь и человеческий константный домен CH1 IgG1, а также вариабельный домен легкой цепи и человеческий константный домен Ck1, имеющие последовательности, представленные в SEQ ID NO: 67, 99, 80 и 98 соответственно.
[00233] Мышей усыпляли на 35 день и опухоли собирали и взвешивали. Рассчитывали подавление опухолевого роста (%TGI = (1-средний объем опухоли в каждой лечебной группе/средний объем опухоли в контрольной группе) × 100%).
[00234] Все варианты лечения хорошо переносились содержащими опухоли животными. Как можно видеть из Таблицы 10, лечение мышиным C1E1 в концентрации 3 мг/кг и лечение мышиным C1E1-Fab в концентрации 3 мг/кг приводило к значительно лучшей эффективности по сравнению с ниволумабом в концентрации 3 мг/кг на модели ксенотрансплантата меланомы человека A375.
Таблица 10. Противоопухолевая эффективность мышиного антитела C1E1 на модели ксенотрансплантата клеток меланомы человека A375
a По сравнению с контрольной группой, получавшей наполнитель, по t-критерию Стьюдента,*P<0,05.
[00235] Пример 9. In Vivo противоопухолевая эффективность вируса герпеса T3011 со встроенными последовательностями, кодирующими мышиный C1E1
[00236] Противоопухолевая эффективность вируса герпеса T3011 со встроенными последовательностями, кодирующими мышиное антитело C1E1, оценивали на модели ксенотрансплантата меланомы B16F10-hPD-L1.
[00237] Вирус герпеса T3011 представляет собой генетически вирус простого герпеса 1 типа (HSV-1), модифицированный путем удаления области инвертированного повтора и замещения его на ген человеческого IL-12 (см., например, PCT/CN2016/080025) помимо инсерции последовательности Fab-фрагмента C1E1 в геномную область между UL3 и UL4. Последовательности Fab C1E1 содержат мышиный вариабельный домен тяжелой цепи и человеческий константный домен CH1 IgG1, а также мышиный вариабельный домен легкой цепи и человеческий константный домен Cкаппа1, имеющие последовательности, представленные в SEQ ID NO: 67, 99, 80 и 98 соответственно.
[00238] Вкратце, самкам гуманизированных мышей B-hPD-1 подкожно инъецировали в переднюю часть правого бока клетки меланомы B16F10-hPD-L1 в концентрации 1x105. Когда объем опухолей достигал примерно 90 мм3, мышей рандомизировали на пять групп, по 8 мышей в каждой группе. Указанным мышам вводили внутрь опухолей наполнитель (5% раствор глюкозы), T3011 (5×106 PFU/мышь), T3011 (1×107 PFU/мышь), T3011 (3×107 PFU/мышь) и NV1020 (3×107 PFU/мышь, рекомбинантный онколитический вирус на основе HSV, разработанный компанией MediGene (ранее NeuroVir) для потенциального лечения рака (см., например, Phase I/II study of oncolytic herpes simplex virus NV1020 in patients with extensively pretreated refractory colorectal cancer metastatic to the liver. Hum Gene Ther. 2010 Sep;21(9):1119-28) соответственно. Объем опухолей измеряли два раза в неделю.
[00239] Через двенадцать дней после введения лекарственного средства мышей усыпляли и опухоли собирали, взвешивали и фотографировали. Рассчитывали относительный объем опухоли (RTV = TVn/TV0, где TVn представляет собой объем опухоли на день n и TV0 представляет собой объем опухоли на день 0), коэффициент подавления опухолевого роста (TGI% = 1-средний объем опухоли в каждой лечебной группе/средний объем опухоли в контрольной группе) × 100%) и коэффициент снижения массы опухоли (IRTW = (1-средняя масса опухоли в каждой лечебной группе/средняя масса опухоли в контрольной группе) × 100%).
[00240] Как показано в Таблице 11, ниже, T3011 со встроенными последовательностями, кодирующими мышиное антитело C1E1, обеспечивал очевидную противоопухолевую дозозависимую активность. Введение T3011 показало лучшую противоопухолевую эффективность по сравнению с введением NV1020 в дозе 3×107 PFU/мышь.
Таблица 11. Противоопухолевая эффективность вируса герпеса T3011 со встроенными последовательностями, кодирующими мышиный C1E1, на модели ксенотрансплантата меланомы B16F10-hPD-L1
a По сравнению с контрольной группой, получавшей наполнитель, по t-критерию Стьюдента, *P<0,05 и **P<0,01.
[00241] Пример 10. In Vivo Противоопухолевая эффективность гуманизированного антитела C1E1
[00242] Эффект гуманизированного антитела C1E1 на опухолевый рост оценивали на модели ксенотрансплантата MC38. Вкратце, самкам мышей B-hPD-1 подкожно инъецировали в заднюю часть правого бока 5×105 клеток. Когда объемы опухолей достигали примерно 100-150 мм3, мышей рандомизировали на 7 групп, по 8 мышей на группу. Животным внутрибрюшинно вводили наполнитель (PBS), huC1E1-V10 и OPDIVO® соответственно в дозе 1 мг/кг, 3 мг/кг или 10 мг/кг два раза в неделю в течение трех недель. Гуманизированное антитело huC1E1-V10 содержало вариабельный участок и константный участок тяжелой цепи, а также вариабельный участок и константный участок легкой цепи, имеющие последовательности, представленные в SEQ ID NO: 69, 97, 86 и 98 соответственно. Объем опухолей измеряли во время исследования.
[00243] Через двадцать три дня после начала лечения мышей усыпляли и опухоли собирали, взвешивали и фотографировали. Рассчитывали коэффициент подавления роста объема опухоли (TGITV), данные показаны в Таблице 12.
[00244] Как показано в Таблице 12, ниже, все варианты лечения хорошо переносились содержащими опухоли животными. Лечение HuC1E1-V10 приводило к дозозависимой противоопухолевой активности на модели ксенотрансплантата MC38, что было сравнимо с OPDIVO®.
Таблица 12. Противоопухолевая эффективность huC1E1-V10 на модели ксенотрансплантата MC38
(мм3)
a По сравнению с контрольной группой, получавшей наполнитель, по t-критерию Стьюдента, *P<0,05 и **P<0,01.
[00245] Следует понимать, что несмотря на то, что изобретение было описано выше в связи с одним или более вариантами реализации, предполагается, что настоящее изобретение не ограничивается указанными вариантами реализации и что описание включает все альтернативы, модификации и эквиваленты, которые могут быть включены в объем и сущность прилагаемой формулы изобретения. Все ссылки, цитированные в настоящей заявке, также полностью включены посредством ссылки.
[00246] Последовательности, описанные в настоящей заявке, суммированы ниже.
последовательность/SEQ ID NO
GFTFSSYL (SEQ ID NO: 1)
ISGGGGDT (SEQ ID NO: 2)
VRFGGAGYYWYFD (SEQ ID NO: 3)
GFTFSSYT (SEQ ID NO: 4)
ISGGGSNI (SEQ ID NO: 5)
VLNYAYAMDYWGQ (SEQ ID NO: 6)
GFAFSSYD (SEQ ID NO: 7)
ITGGGSSS (SEQ ID NO: 8)
ASPYLSYFDYWGQ (SEQ ID NO: 9)
GFTFSNYA (SEQ ID NO: 10)
ISGGGGNI (SEQ ID NO: 11)
ASPYANYVWYLDV (SEQ ID NO: 12)
GFTFSSNT (SEQ ID NO: 13)
ISGGGVNT (SEQ ID NO: 14)
ARHGNYNYYGMDY (SEQ ID NO: 15)
GYTFTNYW (SEQ ID NO: 16)
IYPGGGYT (SEQ ID NO: 17)
ARGYGTNYWYFDV (SEQ ID NO: 18)
GFSLSTSGMG (SEQ ID NO: 19)
IWWDDDK (SEQ ID NO: 20)
ARTGGFITTGYWY (SEQ ID NO: 21)
GYKFTDYA (SEQ ID NO: 22)
ISTYSGDV (SEQ ID NO: 23)
SRLGITAGFAYWG (SEQ ID NO: 24)
GFTFSSNT (SEQ ID NO: 25)
ISGGGVDT (SEQ ID NO: 26)
ARHGNYNYYGMDY (SEQ ID NO: 27)
GFTFSYYG (SEQ ID NO: 28)
ISSGSSFT (SEQ ID NO: 29)
TRREGIYDASWDY (SEQ ID NO: 30)
GYTFTNYG (SEQ ID NO: 31)
INTYSGEP (SEQ ID NO: 32)
VRQGDFDYEDAMD (SEQ ID NO: 33)
RASKSVDDSGISFMH (SEQ ID NO: 34)
AASNQGS (SEQ ID NO: 35)
HQTKEVPWT (SEQ ID NO: 36)
RASQDISNFLN (SEQ ID NO: 37)
YTSRLQS (SEQ ID NO: 38)
QQGSSLPWT (SEQ ID NO: 39)
RASQSISNNLH (SEQ ID NO: 40)
GSQSMS (SEQ ID NO: 41)
QQSNSWPLT (SEQ ID NO: 42)
RASQDISNYLN (SEQ ID NO: 43)
YTSRLHS (SEQ ID NO: 44)
QQSNALPWT (SEQ ID NO: 45)
RASESVDNSGISFMN (SEQ ID NO: 46)
TASNQGS (SEQ ID NO: 47)
QQSYEVPWT (SEQ ID NO: 48)
KASQSVSNDVA (SEQ ID NO: 49)
YAFHRYT (SEQ ID NO: 50)
QQDYSSPYT(SEQ ID NO: 51)
RASQDISNYLI (SEQ ID NO: 52)
YTSRLHS (SEQ ID NO: 53)
QQHKTLPWT (SEQ ID NO: 54)
KASQNVRTAVA (SEQ ID NO: 55)
LASNRHT (SEQ ID NO: 56)
LQHWNYPYT (SEQ ID NO: 57)
RASESVDNSGISFMN (SEQ ID NO: 58)
IASNHGS (SEQ ID NO: 59)
QQSYEVPWT (SEQ ID NO: 60)
RSSQSIIRSNGNTYLE (SEQ ID NO: 61)
KVSNRFS (SEQ ID NO: 62)
FQGSHVPWT (SEQ ID NO: 63)
RSSQSIVHSNGHIYLE (SEQ ID NO: 64)
KVSKRFS (SEQ ID NO: 65)
FQGSHGT (SEQ ID NO: 66)
EVMLVESGGGLVKPGGSLKLSCAASGFTFSSYLMSWVRQTPEKRLEWVATISGGGGDTYFPDSVKGRFTISRDNVKNNLYLQMSSLRSEDTALYYCVRFGGAGYYWYFDVWGAGTTVTVSS (SEQ ID NO: 67)
GAAGTGATGCTGGTGGAGTCTGGGGGAGGGTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCACGTTCAGTAGTTATCTTATGTCTTGGGTTCGCCAGACTCCGGAGAAGAGGCTGGAGTGGGTCGCAACCATTAGTGGTGGTGGTGGTGACACCTACTTTCCAGACAGTGTGAAGGGTCGATTCACCATCTCCAGAGACAATGTCAAGAACAACCTGTACCTGCAAATGAGCAGTCTTAGGTCTGAGGACACGGCCTTGTATTACTGTGTAAGATTTGGGGGCGCTGGTTACTACTGGTATTTCGATGTCTGGGGCGCAGGGACCACGGTCACCGTCTCCTCA (SEQ ID NO: 102)
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYLMSWVRQAPGKGLEWVATISGGGGDTYFPDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCVRFGGAGYYWYFDVWGAGTLVTVSS (SEQ ID NO: 68)
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYLMSWVRQAPGKGLEWVATISGGGGDTYFPDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCVRFGGAGYYWYFDVWGQGTLVTVSS (SEQ ID NO: 69)
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTAGCTATCTTATGTCCTGGGTCCGCCAGGCTCCAGGCAAGGGGCTAGAGTGGGTGGCAACTATATCAGGTGGTGGAGGTGACACATACTTCCCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCTGAGGACACGGCTGTGTATTACTGTGTGAGATTCGGTGGTGCTGGTTACTACTGGTACTTTGACGTCTGGGGCCAAGGAACCCTGGTCACCGTCTCGAGT (SEQ ID NO: 103)
EVMLVESGGGLVKPGGSLKLSCAASGFTFSSY™SWVRQTPEKRLEWVATISGGGSNIYYPDSVEGRFTVSRDNARNTLYLHMSSLRSEDTALYYCVLNYAYAMDYWGQGTSVTVSS (SEQ ID NO: 70)
GAAGTGATGCTGGTGGAGTCTGGGGGAGGCTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCACTTTCAGTAGCTATACCATGTCTTGGGTTCGCCAGACTCCGGAGAAGAGGCTGGAGTGGGTCGCAACCATTAGTGGTGGTGGTAGTAACATCTACTATCCAGACAGTGTGGAGGGTCGATTCACCGTCTCCAGAGACAATGCCAGGAACACCCTGTACCTGCATATGAGCAGTCTGAGGTCTGAGGACACGGCCTTATATTACTGTGTCCTTAACTACGCCTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO: 104)
EVQLVESGGGLVKPGGSLKLSCAASGFAFSSYDMSWVRQTPEKRLEWVAYITGGGSSSFYPDTVKGRFTISRDNSKNTLYLQMTSLRSEDTAMYYCASPYLSYFDYWGQGTTLTVSS (SEQ ID NO: 71)
GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCGCTTTCAGTAGCTATGACATGTCTTGGGTTCGCCAGACTCCGGAGAAGAGGCTGGAGTGGGTCGCATACATTACTGGTGGTGGTAGTAGTTCCTTCTATCCAGACACTGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACCCTGTATTTGCAAATGACCAGTCTGAGGTCTGAGGACACAGCCATGTATTACTGTGCAAGCCCCTACTTATCCTACTTTGACTACTGGGGCCAAGGCACCACTCTCACAGTCTCCTCA (SEQ ID NO: 105)
EVMLVESGGGLVKPGGSLKLSCAASGFTFSNYAMSWVRQTPEKRLEWVATISGGGGNIYYPDSVKGRFTISRDNARNTLYLQMSSLRSEDTALYYCASPYANYVWYLDVWGAGTTVTVSS (SEQ ID NO: 72)
GAAGTGATGCTGGTGGAGTCTGGGGGAGGCTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCACTTTCAGTAACTATGCCATGTCTTGGGTTCGCCAGACTCCGGAGAAGAGGCTGGAGTGGGTCGCAACCATTAGTGGTGGTGGTGGTAACATCTACTATCCAGACAGTGTGAAGGGTCGATTCACCATCTCCAGAGACAATGCCAGGAACACCCTGTACCTGCAAATGAGCAGTCTGAGGTCTGAGGACACGGCCTTGTATTATTGTGCAAGCCCGTATGCTAACTACGTATGGTACCTCGATGTCTGGGGCGCAGGGACCACGGTCACCGTCTCCTCA (SEQ ID NO: 106)
EVMLVESGGGLVKPGGSLKLSCAASGFTFSSN™SWVRQTPEKRLEWVAAISGGGVNTYYPDSVKGRFTISRDNARNTLYLQMSSLRSEDTALYYCARHGNYNYYGMDYWGQGTSVTVSS (SEQ ID NO: 73)
GAAGTGATGCTGGTGGAGTCTGGGGGAGGCTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCACTTTCAGTAGCAATACCATGTCTTGGGTTCGCCAGACTCCGGAGAAGAGGCTGGAGTGGGTCGCAGCCATTAGTGGTGGTGGTGTTAACACCTACTATCCAGACAGTGTGAAGGGTCGATTCACCATCTCCAGAGACAATGCCAGGAACACCCTGTACCTGCAAATGAGCAGTCTGAGGTCTGAGGACACGGCCCTGTATTACTGTGCAAGACATGGTAACTACAATTACTATGGTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO: 107)
QVQLQQSGAELVRPGTSVKISCKASGYTFTNYWLGWVKQRPGHGLEWIGDIYPGGGYTNYNENFKGKATLTADTSSSTAYMQLSSLTSEDSAVFFCARGYGTNYWYFDVWGAGTTVTVSS (SEQ ID NO: 74)
CAGGTCCAGTTGCAGCAGTCTGGAGCTGAGCTGGTAAGGCCTGGGACTTCAGTGAAGATATCCTGCAAGGCTTCTGGCTACACCTTCACTAACTACTGGCTAGGTTGGGTAAAACAGAGGCCTGGACATGGACTTGAGTGGATTGGAGATATTTACCCTGGAGGTGGTTATACTAACTACAATGAGAACTTCAAGGGCAAGGCCACACTGACTGCAGACACATCCTCCAGCACTGCCTACATGCAGCTCAGTAGCCTGACATCTGAGGACTCTGCTGTCTTTTTCTGTGCAAGAGGCTACGGTACTAATTACTGGTACTTCGATGTCTGGGGCGCAGGGACCACGGTCACCGTCTCCTCA (SEQ ID NO: 108)
QVTLKESGPGILKPSQTLSLTCSFSGFSLSTSGMGVGWIRQPSGKGLEWLAHIWWDDDKFYNPSLKSQLTISKDTSRNQVFLKITSVDTADTATYYCARTGGFITTGYWYFDVWGAGTTVTVSS (SEQ ID NO: 75)
CAAGTTACTCTAAAAGAGTCTGGCCCTGGGATATTGAAGCCCTCACAGACCCTCAGTCTGACTTGTTCTTTCTCTGGGTTTTCACTGAGCACTTCTGGTATGGGTGTAGGCTGGATTCGTCAGCCTTCAGGGAAGGGTCTGGAGTGGCTGGCACACATTTGGTGGGATGATGATAAGTTCTATAACCCATCCCTGAAGAGCCAGCTCACAATCTCCAAGGATACCTCCAGAAACCAGGTTTTCCTCAAGATCACCAGTGTGGACACTGCAGATACTGCCACTTACTACTGTGCTCGAACCGGGGGGTTTATTACTACGGGCTACTGGTACTTCGATGTCTGGGGCGCAGGGACCACGGTCACCGTCTCCTCA (SEQ ID NO: 109)
QVQLQQSGAELVRPGVSVKISCKGSGYKFTDYAMHWVRQSHAKSLEWIGIISTYSGDVSFNQNFKGKA™TVDKSSSTAYMELARLTSEDSAIYYCSRLGITAGFAYWGQGTLVTVSA (SEQ ID NO: 76)
CAGGTCCAGCTGCAACAGTCTGGGGCTGAGCTGGTGAGGCCTGGGGTCTCAGTGAAGATTTCCTGCAAGGGTTCTGGCTACAAATTCACTGATTATGCTATGCACTGGGTGAGGCAGAGTCATGCAAAGAGTCTAGAGTGGATTGGAATTATTAGTACTTACTCTGGTGACGTTAGTTTCAACCAGAACTTCAAGGGCAAGGCCACAATGACTGTAGACAAATCCTCCAGCACAGCCTATATGGAACTTGCCAGACTGACATCTGAGGATTCTGCCATCTATTACTGTTCAAGACTGGGGATTACGGCGGGGTTTGCTTACTGGGGCCAAGGGACTCTGGTCACTGTCTCTGCA (SEQ ID NO: 110)
EVMLVESGGGLVKPGGSLKLSCAASGFTFSSN™SWVRQTPEKRLEWVAAISGGGVDTYYPDSVKGRFTISRDNARNTLYLQMSSLRSEDTALYYCARHGNYNYYGMDYWGQGTSVTVSS (SEQ ID NO: 77)
GAAGTGATGCTGGTGGAGTCTGGGGGAGGCTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCACTTTCAGTAGCAATACCATGTCTTGGGTTCGCCAGACTCCGGAGAAGAGGCTGGAGTGGGTCGCAGCCATTAGTGGTGGTGGTGTTGACACCTACTATCCAGACAGTGTGAAGGGTCGATTCACCATCTCCAGAGACAATGCCAGGAACACCCTGTACCTGCAAATGAGCAGTCTGAGGTCTGAGGACACGGCCCTGTATTACTGTGCAAGACATGGTAACTACAATTACTATGGTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO: 111)
EVQLVESGGDLVKPGGSLKLSCTASGFTFSYYGMSWVRQTPDKRLEWVATISSGSSFTYSPDSVKGRFTISRDNAKNTLYLQMNSLKSEDTAIYYCTRREGIYDASWDYSMDYWGQGTSVTVSS (SEQ ID NO: 78)
GAGGTGCAACTGGTGGAGTCTGGGGGAGACTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTACAGCCTCTGGATTCACTTTCAGTTACTATGGCATGTCTTGGGTTCGCCAGACTCCAGACAAGAGGCTGGAATGGGTCGCAACCATTAGTAGTGGTAGTAGTTTCACCTACTCTCCAGACAGTGTGAAGGGGCGATTCACCATCTCCAGAGACAATGCCAAGAACACCCTGTACCTACAAATGAACAGTCTGAAGTCTGAGGACACAGCCATTTATTACTGTACAAGACGAGAGGGGATCTATGATGCTTCCTGGGATTATTCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO: 112)
QIQLVQSGPELKKPGETVKISCKASGYTFTNYGINWVKQAPGKGLKWMGWINTYSGEPTYSDDFKGRFAFSLETSASTAYLQINNLKNEDTSTYFCVRQGDFDYEDAMDYWGQGTSVTVSS (SEQ ID NO: 79)
CAGATCCAGTTGGTGCAGTCAGGACCTGAGCTGAAGAAGCCTGGAGAGACAGTCAAGATCTCCTGCAAGGCTTCTGGGTATACCTTCACAAACTATGGAATAAACTGGGTGAAGCAGGCTCCAGGAAAGGGTTTAAAGTGGATGGGCTGGATAAACACCTATAGTGGAGAGCCAACATATTCTGATGACTTCAAGGGACGCTTTGCCTTCTCTTTGGAAACCTCTGCCAGCACTGCCTATTTGCAGATCAACAACCTCAAAAATGAGGACACGTCTACATATTTCTGTGTAAGACAGGGGGACTTTGATTACGAGGATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO: 113)
DIVLTQSPASLAVSLGQRATISCRASKSVDDSGISFMHWFQQKPGQPPKLLIYAASNQGSGVPARFRGSGSGTDFSLNIHPMEEDDTAMYFCHQTKEVPWTFGGGTKLEIK (SEQ ID NO: 80)
GACATTGTGCTGACCCAATCTCCAGCTTCTTTGGCTGTGTCTCTAGGGCAGAGGGCCACCATCTCCTGCAGAGCCAGCAAAAGTGTTGATGATTCTGGCATTAGTTTTATGCACTGGTTCCAACAGAAACCAGGACAGCCACCCAAACTCCTCATCTATGCTGCATCCAACCAAGGATCCGGGGTCCCTGCCAGGTTTCGTGGCAGTGGGTCTGGGACAGACTTCAGCCTCAACATCCATCCTATGGAGGAGGATGATACTGCAATGTATTTCTGTCACCAAACTAAGGAGGTTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 114)
EIVLTQSPATLSLSPGERATLSCRASKSVDDSGISFMHWFQQKPGQPPRLLIYAASNQGSGVPARFSGSGSGTDFTLTISSLEPEDFAVYFCHQTKEVPWTFGQGTKVEIK (SEQ ID NO: 81)
EIVLTQSPATLSLSPGERATLSCRASKSVDDSGISFMHWYQQKPGQPPRLLIYAASNQGSGVPARFSGSGSGTDFTLTISSLEPEDFAVYFCHQTKEVPWTFGQGTKVEIK (SEQ ID NO: 82)
EIVLTQSPATLSLSPGERATLSCRASKSVDDSGISFMHWFQQKPGQAPRLLIYAASNQGSGVPARFSGSGSGTDFTLTISSLEPEDFAVYFCHQTKEVPWTFGQGTKVEIK (SEQ ID NO: 83)
EIVLTQSPATLSLSPGERATLSCRASKSVDDSGISFMHWFQQKPGQPPRLLIYAASNQGSGIPARFSGSGSGTDFTLTISSLEPEDFAVYFCHQTKEVPWTFGQGTKVEIK (SEQ ID NO: 84)
EIVLTQSPATLSLSPGERATLSCRASKSVDDSGISFMHWFQQKPGQPPRLLIYAASNQGSGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCHQTKEVPWTFGQGTKVEIK (SEQ ID NO: 85)
EIVLTQSPATLSLSPGERATLSCRASKSVDDSGISFMHWYQQKPGQAPRLLIYAASNQGSGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCHQTKEVPWTFGQGTKVEIK (SEQ ID NO: 86)
GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTAAGTCTGTTGACGACAGTGGTATCAGCTTCATGCACTGGTATCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGCTGCATCCAACCAGGGCTCTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCATCAGACTAAGGAGGTGCCTTGGACGTTCGGCCAAGGGACCAAGGTGGAGATCAAA (SEQ ID NO: 115)
DIQMTQTTSSLSASLGDRVTISCRASQDISNFLNWYQQKPDGTVKLLIYYTSRLQSGVPSRFSGTGSGTDYSLTISNLEQEDLATYFCQQGSSLPWTFGGGTKLEIK (SEQ ID NO: 87)
GATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCCAGTCAGGACATTAGCAATTTTTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCTACTACACATCAAGATTACAGTCAGGAGTCCCATCAAGGTTCAGTGGCACTGGGTCTGGGACAGATTATTCTCTCACCATTAGCAACCTGGAACAAGAAGATCTTGCCACTTACTTTTGCCAACAGGGTAGTTCGCTTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 116)
DIVLTQSPATLSVTPGDSVSLSCRASQSISNNLHWYQQKSHESPRLLIKYGSQSMSGIPSRFSGSGSGTDFTLVINSVETEDFGMYFCQQSNSWPLTFGAGTKLELK (SEQ ID NO: 88)
GATATTGTGCTAACTCAGTCTCCAGCCACCCTGTCTGTGACTCCAGGAGATAGCGTCAGTCTTTCCTGCAGGGCCAGCCAAAGTATTAGCAACAACCTACACTGGTATCAACAAAAATCACATGAGTCTCCAAGGCTTCTCATCAAGTATGGTTCCCAGTCCATGTCTGGGATCCCCTCCAGGTTCAGTGGCAGTGGATCAGGGACAGATTTCACTCTCGTTATCAACAGTGTGGAGACTGAAGATTTTGGAATGTATTTCTGTCAACAGAGTAACAGCTGGCCTCTCACGTTCGGTGCTGGGACCAAGCTGGAGCTGAAA (SEQ ID NO: 117)
DIQMTQTTSSLSASLGDRVTISCRASQDISNYLNWYQQKPDGTVKLLIYYTSRLHSGVPSRFSGSGSGTDFSLTISNLEEEDIATYFCQQSNALPWTFGGGTKLEIK (SEQ ID NO: 89)
GATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTAGCAATTATTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCTACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTTTTCTCTCACCATTAGCAACCTGGAAGAAGAAGATATTGCCACTTACTTTTGCCAACAGAGTAATGCGCTTCCGTGGACGTTCGGTGGAGGCACCAAACTGGAAATCAAA (SEQ ID NO: 118)
DIVLTQSPASLVVSLGQRATISCRASESVDNSGISFMNWFQQKPGQPPKLLIYTASNQGSGVPARFSGSGSGTDFSLNIHPMEEDDSAMYFCQQSYEVPWTFGGGTKLEIK (SEQ ID NO: 90)
GACATTGTGCTGACCCAATCTCCAGCTTCTTTGGTTGTGTCTCTAGGGCAGAGGGCCACCATCTCCTGCAGAGCCAGCGAAAGTGTTGATAATTCTGGCATTAGTTTTATGAACTGGTTCCAACAGAAACCAGGACAGCCACCCAAACTCCTCATCTATACTGCATCCAACCAAGGATCCGGGGTCCCTGCCAGGTTTAGTGGCAGTGGGTCTGGGACAGACTTCAGCCTCAACATCCATCCTATGGAGGAGGATGATTCTGCAATGTATTTCTGTCAGCAAAGTTATGAGGTTCCTTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 119)
SIVMTQTPKFLLVSAGDRVTITCKASQSVSNDVAWYQQKPGQSPKLLIYYAFHRYTGVPDRFTGSGYGTDFTFTISTVQAEDLAVYFCQQDYSSPYTFGGGTKLEIK (SEQ ID NO: 91)
AGTATTGTGATGACCCAGACTCCCAAATTCCTGCTTGTATCAGCAGGAGACAGGGTTACCATAACCTGCAAGGCCAGTCAGAGTGTGAGTAATGATGTAGCTTGGTACCAACAGAAGCCAGGGCAGTCTCCTAAACTGCTGATATACTATGCATTTCATCGCTACACTGGAGTCCCTGATCGCTTCACTGGCAGTGGATATGGGACGGATTTCACTTTCACCATCAGCACTGTGCAGGCTGAAGACCTGGCAGTTTATTTCTGTCAGCAGGATTATAGCTCTCCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATAAAA (SEQ ID NO: 120)
DIQMTQTTSSLSASLGDRVTISCRASQDISNYLIWYQQKTDGTLKLLIYYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQHKTLPWTFGGGTKLEIK (SEQ ID NO: 92)
GATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTAGCAATTATTTAATCTGGTATCAGCAGAAAACAGATGGAACTCTTAAACTCCTGATCTACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTATTCTCTCACCATTAGCAACCTGGAGCAAGAAGATATTGCCACTTACTTTTGCCAGCAGCATAAAACGCTTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 121)
DIVMTQSQKFMSTSVGDRVTITCKASQNVRTAVAWYQQKPGQSPKALIYLASNRHTGVPDRFTGSGSGTDFTLTISNVQSKDLADYFCLQHWNYPYTFGGGTKLEIK (SEQ ID NO: 93)
GACATTGTGATGACCCAGTCTCAAAAATTCATGTCCACATCAGTAGGAGACAGGGTCACCATCACCTGCAAGGCCAGTCAGAATGTTCGTACTGCTGTAGCCTGGTATCAACAGAAACCAGGGCAGTCTCCTAAAGCACTGATTTACTTGGCATCCAACCGGCACACTGGAGTCCCTGATCGCTTCACAGGCAGTGGATCTGGGACAGATTTCACTCTCACCATTAGCAATGTGCAATCTAAAGACCTGGCAGATTATTTCTGTCTGCAACATTGGAATTATCCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATAAAA (SEQ ID NO: 122)
DIVLTQSPASLAVSLGQRATISCRASESVDNSGISFMNWFQQKPGQSPKLLIYIASNHGSGVPARFSGSGSGTDFSLNIHPMEEDDSAMYFCQQSYEVPWTFGGGTKLEIK (SEQ ID NO: 94)
GACATTGTGCTGACCCAATCTCCAGCTTCTTTGGCTGTGTCTCTAGGGCAGAGGGCCACCATCTCCTGCAGAGCCAGCGAAAGTGTTGATAATTCTGGCATTAGTTTTATGAACTGGTTCCAACAGAAACCAGGACAGTCACCCAAACTCCTCATCTATATTGCATCCAACCACGGATCCGGGGTCCCTGCCAGGTTTAGTGGCAGTGGGTCTGGGACAGACTTCAGCCTCAACATCCATCCTATGGAGGAGGATGATTCTGCAATGTATTTCTGTCAGCAAAGTTATGAGGTTCCTTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 123)
DVLMTQTPLSLPVSLGDQASISCRSSQSIIRSNGNTYLEWYLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEADDLGLYYCFQGSHVPWTFGGGTKLEIK (SEQ ID NO: 95)
GATGTTTTGATGACCCAAACTCCACTCTCCCTGCCTGTCAGTCTTGGAGATCAAGCCTCCATCTCTTGCAGATCTAGTCAGAGCATTATACGTAGTAATGGAAACACCTATTTAGAATGGTACCTGCAGAAACCAGGCCAGTCTCCAAAGCTCCTGATCTACAAAGTTTCCAACCGATTTTCTGGGGTCCCAGACAGGTTCAGTGGCAGTGGATCAGGGACAGATTTCACACTCAAGATCAGCAGAGTGGAGGCTGACGATCTGGGACTTTATTACTGCTTTCAAGGTTCACATGTTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 124)
DVLMTQSPLSLPVSLGDQASISCRSSQSIVHSNGHIYLEWYLQKPGQSPKLLIYKVSKRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYYCFQGSHGTFGGGTKLEIK (SEQ ID NO: 96)
GATGTTTTGATGACCCAAAGTCCACTCTCCCTGCCTGTCAGTCTTGGAGATCAAGCCTCCATCTCTTGCAGATCTAGTCAGAGTATTGTACATAGTAATGGACACATCTATTTAGAATGGTACCTGCAGAAACCAGGCCAGTCTCCAAAGCTCCTGATCTACAAAGTTTCCAAGCGATTTTCTGGGGTCCCAGACAGGTTCAGTGGCAGTGGATCAGGGACAGATTTCACACTCAAGATCAGCAGAGTGGAGGCTGAGGATCTGGGAGTTTATTACTGCTTTCAAGGTTCACATGGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA (SEQ ID NO: 125)
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 97)
GCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA (SEQ ID NO: 126)
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 98)
CGTACGGTGGCGGCGCCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGT (SEQ ID NO: 127)
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKV (SEQ ID NO:99)
Gcctccaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagtt (SEQ ID NO: 128)
LDSPDRPWNPPTFSPALLVVTEGDNATFTCSFSNTSESFVLNWYRMSPSNQTDKLAAFPEDRSQPGQDCRFRVTQLPNGRDFHMSVVRARRNDSGTYLCGAISLAPKAQIKESLRAELRVTERRAEVPTAHPSPSPRPAGQFQEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSL (SEQ ID NO:100)
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERTEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 101)
AKTTPPSVYPLAPGSAAQTNSMVTLGCLVKGYFPEPVTVTWNSGSLSSGVHTFPAVLQSDLYTLSSSVTVPSSTWPSETVTCNVAHPASSTKVDKKIVPRDCGCKPCICTVPEVSSVFIFPPKPKDVLTITLTPKVTCVVVDISKDDPEVQFSWFVDDVEVHTAQTQPREEQFNSTFRSVSELPIMHQDWLNGKEFKCRVNSAAFPAPIEKTISKTKGRPKAPQVYTIPPPKEQMAKDKVSLTCMITDFFPEDITVEWQWNGQPAENYKNTQPIMDTDGSYFVYSKLNVQKSNWEAGNTFTCSVLHEGLHNHHTEKSLSHSPGK (SEQ ID NO: 129)
RADAAPTVSIFPPSSEQLTSGGASVVCFLNNFYPKDINVKWKIDGSERQNGVLNSWTDQDSKDSTYSMSSTLTLTKDEYERHNSYTCEATHKTSTSPIVKSFNRGEC (SEQ ID NO: 130)
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Immvira Co., Limited
<120> СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> 55532 00004
<150> US62/657927
<151> 2018-04-15
<160> 130
<170> PatentIn version 3.5
<210> 1
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для C1E1, huC1E1-V1 - huC1E1-V7 и huC1E1-V10
<400> 1
Gly Phe Thr Phe Ser Ser Tyr Leu
1 5
<210> 2
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для C1E1, huC1E1-V1 - huC1E1-V7 и huC1E1-V10
<400> 2
Ile Ser Gly Gly Gly Gly Asp Thr
1 5
<210> 3
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для C1E1, huC1E1-V1 - huC1E1-V7 и huC1E1-V10
<400> 3
Val Arg Phe Gly Gly Ala Gly Tyr Tyr Trp Tyr Phe Asp
1 5 10
<210> 4
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для D1F2
<400> 4
Gly Phe Thr Phe Ser Ser Tyr Thr
1 5
<210> 5
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для D1F2
<400> 5
Ile Ser Gly Gly Gly Ser Asn Ile
1 5
<210> 6
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для D1F2
<400> 6
Val Leu Asn Tyr Ala Tyr Ala Met Asp Tyr Trp Gly Gln
1 5 10
<210> 7
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для C1F5
<400> 7
Gly Phe Ala Phe Ser Ser Tyr Asp
1 5
<210> 8
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для C1F5
<400> 8
Ile Thr Gly Gly Gly Ser Ser Ser
1 5
<210> 9
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для C1F5
<400> 9
Ala Ser Pro Tyr Leu Ser Tyr Phe Asp Tyr Trp Gly Gln
1 5 10
<210> 10
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для D1A1
<400> 10
Gly Phe Thr Phe Ser Asn Tyr Ala
1 5
<210> 11
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для D1A1
<400> 11
Ile Ser Gly Gly Gly Gly Asn Ile
1 5
<210> 12
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для D1A1
<400> 12
Ala Ser Pro Tyr Ala Asn Tyr Val Trp Tyr Leu Asp Val
1 5 10
<210> 13
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для D1F1
<400> 13
Gly Phe Thr Phe Ser Ser Asn Thr
1 5
<210> 14
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для D1F1
<400> 14
Ile Ser Gly Gly Gly Val Asn Thr
1 5
<210> 15
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для D1F1
<400> 15
Ala Arg His Gly Asn Tyr Asn Tyr Tyr Gly Met Asp Tyr
1 5 10
<210> 16
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для C1E2
<400> 16
Gly Tyr Thr Phe Thr Asn Tyr Trp
1 5
<210> 17
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для C1E2
<400> 17
Ile Tyr Pro Gly Gly Gly Tyr Thr
1 5
<210> 18
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для C1E2
<400> 18
Ala Arg Gly Tyr Gly Thr Asn Tyr Trp Tyr Phe Asp Val
1 5 10
<210> 19
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для C1A1
<400> 19
Gly Phe Ser Leu Ser Thr Ser Gly Met Gly
1 5 10
<210> 20
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для C1A1
<400> 20
Ile Trp Trp Asp Asp Asp Lys
1 5
<210> 21
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для C1A1
<400> 21
Ala Arg Thr Gly Gly Phe Ile Thr Thr Gly Tyr Trp Tyr
1 5 10
<210> 22
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для C1F4
<400> 22
Gly Tyr Lys Phe Thr Asp Tyr Ala
1 5
<210> 23
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для C1F4
<400> 23
Ile Ser Thr Tyr Ser Gly Asp Val
1 5
<210> 24
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для C1F4
<400> 24
Ser Arg Leu Gly Ile Thr Ala Gly Phe Ala Tyr Trp Gly
1 5 10
<210> 25
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для D2C2
<400> 25
Gly Phe Thr Phe Ser Ser Asn Thr
1 5
<210> 26
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для D2C2
<400> 26
Ile Ser Gly Gly Gly Val Asp Thr
1 5
<210> 27
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для D2C2
<400> 27
Ala Arg His Gly Asn Tyr Asn Tyr Tyr Gly Met Asp Tyr
1 5 10
<210> 28
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для 2G2
<400> 28
Gly Phe Thr Phe Ser Tyr Tyr Gly
1 5
<210> 29
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для 2G2
<400> 29
Ile Ser Ser Gly Ser Ser Phe Thr
1 5
<210> 30
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для 2G2
<400> 30
Thr Arg Arg Glu Gly Ile Tyr Asp Ala Ser Trp Asp Tyr
1 5 10
<210> 31
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR1 для C1C5
<400> 31
Gly Tyr Thr Phe Thr Asn Tyr Gly
1 5
<210> 32
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR2 для C1C5
<400> 32
Ile Asn Thr Tyr Ser Gly Glu Pro
1 5
<210> 33
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-CDR3 для C1C5
<400> 33
Val Arg Gln Gly Asp Phe Asp Tyr Glu Asp Ala Met Asp
1 5 10
<210> 34
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для C1E1, huC1E1-V1 - huC1E1-V7 и huC1E1-V10
<400> 34
Arg Ala Ser Lys Ser Val Asp Asp Ser Gly Ile Ser Phe Met His
1 5 10 15
<210> 35
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для C1E1, huC1E1-V1 - huC1E1-V7 и huC1E1-V10
<400> 35
Ala Ala Ser Asn Gln Gly Ser
1 5
<210> 36
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для C1E1, huC1E1-V1 - huC1E1-V7 и huC1E1-V10
<400> 36
His Gln Thr Lys Glu Val Pro Trp Thr
1 5
<210> 37
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для D1F2
<400> 37
Arg Ala Ser Gln Asp Ile Ser Asn Phe Leu Asn
1 5 10
<210> 38
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для D1F2
<400> 38
Tyr Thr Ser Arg Leu Gln Ser
1 5
<210> 39
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для D1F2
<400> 39
Gln Gln Gly Ser Ser Leu Pro Trp Thr
1 5
<210> 40
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для C1F5
<400> 40
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1 5 10
<210> 41
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для C1F5
<400> 41
Gly Ser Gln Ser Met Ser
1 5
<210> 42
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для C1F5
<400> 42
Gln Gln Ser Asn Ser Trp Pro Leu Thr
1 5
<210> 43
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для D1A1
<400> 43
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 44
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для D1A1
<400> 44
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 45
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для D1A1
<400> 45
Gln Gln Ser Asn Ala Leu Pro Trp Thr
1 5
<210> 46
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для D1F1
<400> 46
Arg Ala Ser Glu Ser Val Asp Asn Ser Gly Ile Ser Phe Met Asn
1 5 10 15
<210> 47
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для D1F1
<400> 47
Thr Ala Ser Asn Gln Gly Ser
1 5
<210> 48
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для D1F1
<400> 48
Gln Gln Ser Tyr Glu Val Pro Trp Thr
1 5
<210> 49
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для C1E2
<400> 49
Lys Ala Ser Gln Ser Val Ser Asn Asp Val Ala
1 5 10
<210> 50
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для C1E2
<400> 50
Tyr Ala Phe His Arg Tyr Thr
1 5
<210> 51
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для C1E2
<400> 51
Gln Gln Asp Tyr Ser Ser Pro Tyr Thr
1 5
<210> 52
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для C1A1
<400> 52
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Ile
1 5 10
<210> 53
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для C1A1
<400> 53
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 54
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для C1A1
<400> 54
Gln Gln His Lys Thr Leu Pro Trp Thr
1 5
<210> 55
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для C1F4
<400> 55
Lys Ala Ser Gln Asn Val Arg Thr Ala Val Ala
1 5 10
<210> 56
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для C1F4
<400> 56
Leu Ala Ser Asn Arg His Thr
1 5
<210> 57
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для C1F4
<400> 57
Leu Gln His Trp Asn Tyr Pro Tyr Thr
1 5
<210> 58
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для D2C2
<400> 58
Arg Ala Ser Glu Ser Val Asp Asn Ser Gly Ile Ser Phe Met Asn
1 5 10 15
<210> 59
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для D2C2
<400> 59
Ile Ala Ser Asn His Gly Ser
1 5
<210> 60
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для D2C2
<400> 60
Gln Gln Ser Tyr Glu Val Pro Trp Thr
1 5
<210> 61
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для 2G2
<400> 61
Arg Ser Ser Gln Ser Ile Ile Arg Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 62
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для 2G2
<400> 62
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 63
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для 2G2
<400> 63
Phe Gln Gly Ser His Val Pro Trp Thr
1 5
<210> 64
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR1 для C1C5
<400> 64
Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly His Ile Tyr Leu Glu
1 5 10 15
<210> 65
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR2 для C1C5
<400> 65
Lys Val Ser Lys Arg Phe Ser
1 5
<210> 66
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-CDR3 для C1C5
<400> 66
Phe Gln Gly Ser His Gly Thr
1 5
<210> 67
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для C1E1
<400> 67
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Leu Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Gly Asp Thr Tyr Phe Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Phe Gly Gly Ala Gly Tyr Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 68
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для huC1E1-V1, huC1E1-V3 - huC1E1-V7
<400> 68
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Leu Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Gly Asp Thr Tyr Phe Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Phe Gly Gly Ala Gly Tyr Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 69
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для huC1E1-V2 и huC1E1-V10
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Leu Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Gly Asp Thr Tyr Phe Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Phe Gly Gly Ala Gly Tyr Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 70
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для D1F2
<400> 70
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Ser Asn Ile Tyr Tyr Pro Asp Ser Val
50 55 60
Glu Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Arg Asn Thr Leu Tyr
65 70 75 80
Leu His Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Leu Asn Tyr Ala Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 71
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для C1F5
<400> 71
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Ala Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Tyr Ile Thr Gly Gly Gly Ser Ser Ser Phe Tyr Pro Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Leu Ser Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 72
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для D1A1
<400> 72
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Gly Asn Ile Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Ala Asn Tyr Val Trp Tyr Leu Asp Val Trp Gly Ala
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 73
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для D1F1
<400> 73
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Asn
20 25 30
Thr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Ala Ile Ser Gly Gly Gly Val Asn Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg His Gly Asn Tyr Asn Tyr Tyr Gly Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 74
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для C1E2
<400> 74
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Leu Gly Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Gly Gly Gly Tyr Thr Asn Tyr Asn Glu Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Phe Phe Cys
85 90 95
Ala Arg Gly Tyr Gly Thr Asn Tyr Trp Tyr Phe Asp Val Trp Gly Ala
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 75
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для C1A1
<400> 75
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Phe Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Gln Leu Thr Ile Ser Lys Asp Thr Ser Arg Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Thr Ser Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Thr Gly Gly Phe Ile Thr Thr Gly Tyr Trp Tyr Phe Asp
100 105 110
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 76
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для C1F4
<400> 76
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Val
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Gly Ser Gly Tyr Lys Phe Thr Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ser His Ala Lys Ser Leu Glu Trp Ile
35 40 45
Gly Ile Ile Ser Thr Tyr Ser Gly Asp Val Ser Phe Asn Gln Asn Phe
50 55 60
Lys Gly Lys Ala Thr Met Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ala Arg Leu Thr Ser Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ser Arg Leu Gly Ile Thr Ala Gly Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala
115
<210> 77
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для D2C2
<400> 77
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Asn
20 25 30
Thr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Ala Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg His Gly Asn Tyr Asn Tyr Tyr Gly Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 78
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для 2G2
<400> 78
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Ser Ser Phe Thr Tyr Ser Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Thr Arg Arg Glu Gly Ile Tyr Asp Ala Ser Trp Asp Tyr Ser Met Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 79
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для C1C5
<400> 79
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Ile Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Ser Gly Glu Pro Thr Tyr Ser Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ser Thr Tyr Phe Cys
85 90 95
Val Arg Gln Gly Asp Phe Asp Tyr Glu Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 80
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для C1E1
<400> 80
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Arg Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 81
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V1 и huC1E1-V2
<400> 81
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 82
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V3
<400> 82
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 83
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V4
<400> 83
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 84
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V5
<400> 84
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 85
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V6
<400> 85
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 86
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V7 и huC1E1-V10
<400> 86
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Asp Asp Ser
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Thr Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 87
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для D1F2
<400> 87
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Thr Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Leu Ala Thr Tyr Phe Cys Gln Gln Gly Ser Ser Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 88
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для C1F5
<400> 88
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1 5 10 15
Asp Ser Val Ser Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Gly Ser Gln Ser Met Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Val Ile Asn Ser Val Glu Thr
65 70 75 80
Glu Asp Phe Gly Met Tyr Phe Cys Gln Gln Ser Asn Ser Trp Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 89
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для D1A1
<400> 89
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr Ile Ser Asn Leu Glu Glu
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Ser Asn Ala Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 90
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для D1F1
<400> 90
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Val Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Ser
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Ser Ala Met Tyr Phe Cys Gln Gln Ser Tyr
85 90 95
Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 91
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для C1E2
<400> 91
Ser Ile Val Met Thr Gln Thr Pro Lys Phe Leu Leu Val Ser Ala Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Ser Val Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ala Phe His Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Phe Thr Ile Ser Thr Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Phe Cys Gln Gln Asp Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 92
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для C1A1
<400> 92
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Ile Trp Tyr Gln Gln Lys Thr Asp Gly Thr Leu Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln His Lys Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 93
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для C1F4
<400> 93
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Arg Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Ala Leu Ile
35 40 45
Tyr Leu Ala Ser Asn Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Lys Asp Leu Ala Asp Tyr Phe Cys Leu Gln His Trp Asn Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 94
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для D2C2
<400> 94
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Ser
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro
35 40 45
Lys Leu Leu Ile Tyr Ile Ala Ser Asn His Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Ser Ala Met Tyr Phe Cys Gln Gln Ser Tyr
85 90 95
Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 95
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для 2G2
<400> 95
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Ile Arg Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Asp Asp Leu Gly Leu Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 96
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для C1C5
<400> 96
Asp Val Leu Met Thr Gln Ser Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly His Ile Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Gly Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 97
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный участок тяжелой цепи IgG1 человека
<400> 97
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 98
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный участок легкой каппа-цепи человека
<400> 98
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 99
<211> 98
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CH1-участок тяжелой цепи IgG1 человека
<400> 99
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 100
<211> 371
<212> БЕЛОК
<213> Homo sapiens
<400> 100
Leu Asp Ser Pro Asp Arg Pro Trp Asn Pro Pro Thr Phe Ser Pro Ala
1 5 10 15
Leu Leu Val Val Thr Glu Gly Asp Asn Ala Thr Phe Thr Cys Ser Phe
20 25 30
Ser Asn Thr Ser Glu Ser Phe Val Leu Asn Trp Tyr Arg Met Ser Pro
35 40 45
Ser Asn Gln Thr Asp Lys Leu Ala Ala Phe Pro Glu Asp Arg Ser Gln
50 55 60
Pro Gly Gln Asp Cys Arg Phe Arg Val Thr Gln Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Val Val Arg Ala Arg Arg Asn Asp Ser Gly Thr
85 90 95
Tyr Leu Cys Gly Ala Ile Ser Leu Ala Pro Lys Ala Gln Ile Lys Glu
100 105 110
Ser Leu Arg Ala Glu Leu Arg Val Thr Glu Arg Arg Ala Glu Val Pro
115 120 125
Thr Ala His Pro Ser Pro Ser Pro Arg Pro Ala Gly Gln Phe Gln Glu
130 135 140
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
145 150 155 160
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
165 170 175
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
180 185 190
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
195 200 205
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
210 215 220
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
225 230 235 240
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
245 250 255
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
260 265 270
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
275 280 285
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
290 295 300
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
305 310 315 320
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
325 330 335
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
340 345 350
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
355 360 365
Leu Ser Leu
370
<210> 101
<211> 453
<212> БЕЛОК
<213> Homo sapiens
<400> 101
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser Tyr Arg
50 55 60
Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 102
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для C1E1
<400> 102
gaagtgatgc tggtggagtc tgggggaggg ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtgcag cctctggatt cacgttcagt agttatctta tgtcttgggt tcgccagact
120
ccggagaaga ggctggagtg ggtcgcaacc attagtggtg gtggtggtga cacctacttt
180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgtcaagaa caacctgtac
240
ctgcaaatga gcagtcttag gtctgaggac acggccttgt attactgtgt aagatttggg
300
ggcgctggtt actactggta tttcgatgtc tggggcgcag ggaccacggt caccgtctcc
360
tca
363
<210> 103
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для huC1E1-V2 и huC1E1-V10
<400> 103
gaggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc
60
tcctgtgcag cctctggatt caccttcagt agctatctta tgtcctgggt ccgccaggct
120
ccaggcaagg ggctagagtg ggtggcaact atatcaggtg gtggaggtga cacatacttc
180
ccagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat
240
ctgcaaatga acagcctgag agctgaggac acggctgtgt attactgtgt gagattcggt
300
ggtgctggtt actactggta ctttgacgtc tggggccaag gaaccctggt caccgtctcg
360
agt
363
<210> 104
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для D1F2
<400> 104
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtgcag cctctggatt cactttcagt agctatacca tgtcttgggt tcgccagact
120
ccggagaaga ggctggagtg ggtcgcaacc attagtggtg gtggtagtaa catctactat
180
ccagacagtg tggagggtcg attcaccgtc tccagagaca atgccaggaa caccctgtac
240
ctgcatatga gcagtctgag gtctgaggac acggccttat attactgtgt ccttaactac
300
gcctatgcta tggactactg gggtcaagga acctcagtca ccgtctcctc a
351
<210> 105
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для C1F5
<400> 105
gaagtgcagc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtgcag cctctggatt cgctttcagt agctatgaca tgtcttgggt tcgccagact
ccggagaaga ggctggagtg ggtcgcatac attactggtg gtggtagtag ttccttctat
180
ccagacactg tgaagggccg attcaccatc tccagagaca attccaagaa caccctgtat
240
ttgcaaatga ccagtctgag gtctgaggac acagccatgt attactgtgc aagcccctac
300
ttatcctact ttgactactg gggccaaggc accactctca cagtctcctc a
351
<210> 106
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для D1A1
<400> 106
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtgcag cctctggatt cactttcagt aactatgcca tgtcttgggt tcgccagact
120
ccggagaaga ggctggagtg ggtcgcaacc attagtggtg gtggtggtaa catctactat
180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaggaa caccctgtac
240
ctgcaaatga gcagtctgag gtctgaggac acggccttgt attattgtgc aagcccgtat
300
gctaactacg tatggtacct cgatgtctgg ggcgcaggga ccacggtcac cgtctcctca
360
<210> 107
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для D1F1
<400> 107
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtgcag cctctggatt cactttcagt agcaatacca tgtcttgggt tcgccagact
120
ccggagaaga ggctggagtg ggtcgcagcc attagtggtg gtggtgttaa cacctactat
180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaggaa caccctgtac
240
ctgcaaatga gcagtctgag gtctgaggac acggccctgt attactgtgc aagacatggt
300
aactacaatt actatggtat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca
360
<210> 108
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для C1E2
<400> 108
caggtccagt tgcagcagtc tggagctgag ctggtaaggc ctgggacttc agtgaagata
60
tcctgcaagg cttctggcta caccttcact aactactggc taggttgggt aaaacagagg
120
cctggacatg gacttgagtg gattggagat atttaccctg gaggtggtta tactaactac
180
aatgagaact tcaagggcaa ggccacactg actgcagaca catcctccag cactgcctac
240
atgcagctca gtagcctgac atctgaggac tctgctgtct ttttctgtgc aagaggctac
300
ggtactaatt actggtactt cgatgtctgg ggcgcaggga ccacggtcac cgtctcctca
360
<210> 109
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для C1A1
<400> 109
caagttactc taaaagagtc tggccctggg atattgaagc cctcacagac cctcagtctg
60
acttgttctt tctctgggtt ttcactgagc acttctggta tgggtgtagg ctggattcgt
120
cagccttcag ggaagggtct ggagtggctg gcacacattt ggtgggatga tgataagttc
180
tataacccat ccctgaagag ccagctcaca atctccaagg atacctccag aaaccaggtt
240
ttcctcaaga tcaccagtgt ggacactgca gatactgcca cttactactg tgctcgaacc
300
ggggggttta ttactacggg ctactggtac ttcgatgtct ggggcgcagg gaccacggtc
360
accgtctcct ca
372
<210> 110
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для C1F4
<400> 110
caggtccagc tgcaacagtc tggggctgag ctggtgaggc ctggggtctc agtgaagatt
tcctgcaagg gttctggcta caaattcact gattatgcta tgcactgggt gaggcagagt
120
catgcaaaga gtctagagtg gattggaatt attagtactt actctggtga cgttagtttc
180
aaccagaact tcaagggcaa ggccacaatg actgtagaca aatcctccag cacagcctat
240
atggaacttg ccagactgac atctgaggat tctgccatct attactgttc aagactgggg
300
attacggcgg ggtttgctta ctggggccaa gggactctgg tcactgtctc tgca
354
<210> 111
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для D2C2
<400> 111
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtgcag cctctggatt cactttcagt agcaatacca tgtcttgggt tcgccagact
120
ccggagaaga ggctggagtg ggtcgcagcc attagtggtg gtggtgttga cacctactat
180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaggaa caccctgtac
240
ctgcaaatga gcagtctgag gtctgaggac acggccctgt attactgtgc aagacatggt
300
aactacaatt actatggtat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca
360
<210> 112
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для 2G2
<400> 112
gaggtgcaac tggtggagtc tgggggagac ttagtgaagc ctggagggtc cctgaaactc
60
tcctgtacag cctctggatt cactttcagt tactatggca tgtcttgggt tcgccagact
120
ccagacaaga ggctggaatg ggtcgcaacc attagtagtg gtagtagttt cacctactct
180
ccagacagtg tgaaggggcg attcaccatc tccagagaca atgccaagaa caccctgtac
240
ctacaaatga acagtctgaa gtctgaggac acagccattt attactgtac aagacgagag
300
gggatctatg atgcttcctg ggattattct atggactact ggggtcaagg aacctcagtc
360
accgtctcct ca
372
<210> 113
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для C1C5
<400> 113
cagatccagt tggtgcagtc aggacctgag ctgaagaagc ctggagagac agtcaagatc
60
tcctgcaagg cttctgggta taccttcaca aactatggaa taaactgggt gaagcaggct
120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct atagtggaga gccaacatat
180
tctgatgact tcaagggacg ctttgccttc tctttggaaa cctctgccag cactgcctat
240
ttgcagatca acaacctcaa aaatgaggac acgtctacat atttctgtgt aagacagggg
300
gactttgatt acgaggatgc tatggactac tggggtcaag gaacctcagt caccgtctcc
360
tca
363
<210> 114
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для C1E1
<400> 114
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc
60
atctcctgca gagccagcaa aagtgttgat gattctggca ttagttttat gcactggttc
120
caacagaaac caggacagcc acccaaactc ctcatctatg ctgcatccaa ccaaggatcc
180
ggggtccctg ccaggtttcg tggcagtggg tctgggacag acttcagcct caacatccat
240
cctatggagg aggatgatac tgcaatgtat ttctgtcacc aaactaagga ggttccgtgg
300
acgttcggtg gaggcaccaa gctggaaatc aaa
333
<210> 115
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для huC1E1-V7 и huC1E1-V10
<400> 115
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc
60
ctctcctgca gggccagtaa gtctgttgac gacagtggta tcagcttcat gcactggtat
120
caacagaaac ctggccaggc tcccaggctc ctcatctatg ctgcatccaa ccagggctct
180
ggcatcccag ccaggttcag tggcagtggg tctgggacag acttcactct caccatcagc
240
agcctagagc ctgaagattt tgcagtttat tactgtcatc agactaagga ggtgccttgg
300
acgttcggcc aagggaccaa ggtggagatc aaa
333
<210> 116
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для D1F2
<400> 116
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc
60
atcagttgca gggccagtca ggacattagc aattttttaa actggtatca gcagaaacca
120
gatggaactg ttaaactcct gatctactac acatcaagat tacagtcagg agtcccatca
180
aggttcagtg gcactgggtc tgggacagat tattctctca ccattagcaa cctggaacaa
240
gaagatcttg ccacttactt ttgccaacag ggtagttcgc ttccgtggac gttcggtgga
300
ggcaccaagc tggaaatcaa a
321
<210> 117
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для C1F5
<400> 117
gatattgtgc taactcagtc tccagccacc ctgtctgtga ctccaggaga tagcgtcagt
60
ctttcctgca gggccagcca aagtattagc aacaacctac actggtatca acaaaaatca
120
catgagtctc caaggcttct catcaagtat ggttcccagt ccatgtctgg gatcccctcc
180
aggttcagtg gcagtggatc agggacagat ttcactctcg ttatcaacag tgtggagact
240
gaagattttg gaatgtattt ctgtcaacag agtaacagct ggcctctcac gttcggtgct
300
gggaccaagc tggagctgaa a
321
<210> 118
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для D1A1
<400> 118
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc
60
atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca
120
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca
180
aggttcagtg gcagtgggtc tggaacagat ttttctctca ccattagcaa cctggaagaa
240
gaagatattg ccacttactt ttgccaacag agtaatgcgc ttccgtggac gttcggtgga
300
ggcaccaaac tggaaatcaa a
321
<210> 119
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для D1F1
<400> 119
gacattgtgc tgacccaatc tccagcttct ttggttgtgt ctctagggca gagggccacc
60
atctcctgca gagccagcga aagtgttgat aattctggca ttagttttat gaactggttc
120
caacagaaac caggacagcc acccaaactc ctcatctata ctgcatccaa ccaaggatcc
180
ggggtccctg ccaggtttag tggcagtggg tctgggacag acttcagcct caacatccat
240
cctatggagg aggatgattc tgcaatgtat ttctgtcagc aaagttatga ggttccttgg
300
acgttcggtg gaggcaccaa gctggaaatc aaa
333
<210> 120
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для C1E2
<400> 120
agtattgtga tgacccagac tcccaaattc ctgcttgtat cagcaggaga cagggttacc
60
ataacctgca aggccagtca gagtgtgagt aatgatgtag cttggtacca acagaagcca
120
gggcagtctc ctaaactgct gatatactat gcatttcatc gctacactgg agtccctgat
180
cgcttcactg gcagtggata tgggacggat ttcactttca ccatcagcac tgtgcaggct
240
gaagacctgg cagtttattt ctgtcagcag gattatagct ctccgtacac gttcggaggg
300
gggaccaagc tggaaataaa a
321
<210> 121
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для C1A1
<400> 121
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc
60
atcagttgca gggcaagtca ggacattagc aattatttaa tctggtatca gcagaaaaca
120
gatggaactc ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca
180
aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa
240
gaagatattg ccacttactt ttgccagcag cataaaacgc ttccgtggac gttcggtgga
300
ggcaccaagc tggaaatcaa a
321
<210> 122
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для C1F4
<400> 122
gacattgtga tgacccagtc tcaaaaattc atgtccacat cagtaggaga cagggtcacc
60
atcacctgca aggccagtca gaatgttcgt actgctgtag cctggtatca acagaaacca
120
gggcagtctc ctaaagcact gatttacttg gcatccaacc ggcacactgg agtccctgat
180
cgcttcacag gcagtggatc tgggacagat ttcactctca ccattagcaa tgtgcaatct
240
aaagacctgg cagattattt ctgtctgcaa cattggaatt atccgtacac gttcggaggg
300
gggaccaagc tggaaataaa a
321
<210> 123
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для D2C2
<400> 123
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc
60
atctcctgca gagccagcga aagtgttgat aattctggca ttagttttat gaactggttc
120
caacagaaac caggacagtc acccaaactc ctcatctata ttgcatccaa ccacggatcc
180
ggggtccctg ccaggtttag tggcagtggg tctgggacag acttcagcct caacatccat
240
cctatggagg aggatgattc tgcaatgtat ttctgtcagc aaagttatga ggttccttgg
300
acgttcggtg gaggcaccaa gctggaaatc aaa
333
<210> 124
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для 2G2
<400> 124
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc
60
atctcttgca gatctagtca gagcattata cgtagtaatg gaaacaccta tttagaatgg
120
tacctgcaga aaccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgacga tctgggactt tattactgct ttcaaggttc acatgttccg
300
tggacgttcg gtggaggcac caagctggaa atcaaa
336
<210> 125
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для C1C5
<400> 125
gatgttttga tgacccaaag tccactctcc ctgcctgtca gtcttggaga tcaagcctcc
60
atctcttgca gatctagtca gagtattgta catagtaatg gacacatcta tttagaatgg
120
tacctgcaga aaccaggcca gtctccaaag ctcctgatct acaaagtttc caagcgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgggacg
300
ttcggtggag gcaccaagct ggaaatcaaa
330
<210> 126
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константный участок тяжелой цепи IgG1 человека
<400> 126
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg
60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg
120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca
180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc
240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc
300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga
360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct
420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg
480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac
540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag
600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc
660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag
720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc
780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg
840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg
900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg
960
cagaagagcc tctccctgtc tccgggtaaa
990
<210> 127
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константный участок легкой каппа-цепи человека
<400> 127
cgtacggtgg cggcgccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct
60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag
120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac
180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag
240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag
300
agcttcaaca ggggagagtg t
321
<210> 128
<211> 294
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CH1-участок тяжелой цепи IgG1 человека
<400> 128
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg
60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg
120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca
180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc
240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agtt
294
<210> 129
<211> 324
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи для мышиных антител
<400> 129
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro
100 105 110
Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 130
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи для мышиных антител
<400> 130
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| СПОСОБЫ КОМБИНИРОВАННОГО ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ, ВКЛЮЧАЮЩИЕ СРЕДСТВА, БЛОКИРУЮЩИЕ CTLA-4 И PD-1 | 2020 |
|
RU2833523C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| АНТИТЕЛА К PD-1 И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2761640C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| НОВЫЕ КОМПОЗИЦИИ АНТИТЕЛ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2019 |
|
RU2804490C2 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2020 |
|
RU2814713C2 |
| ИММУНОГЛОБУЛИН С ТАНДЕМНЫМИ FAB-ФРАГМЕНТАМИ И ВАРИАНТЫ ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2767329C2 |
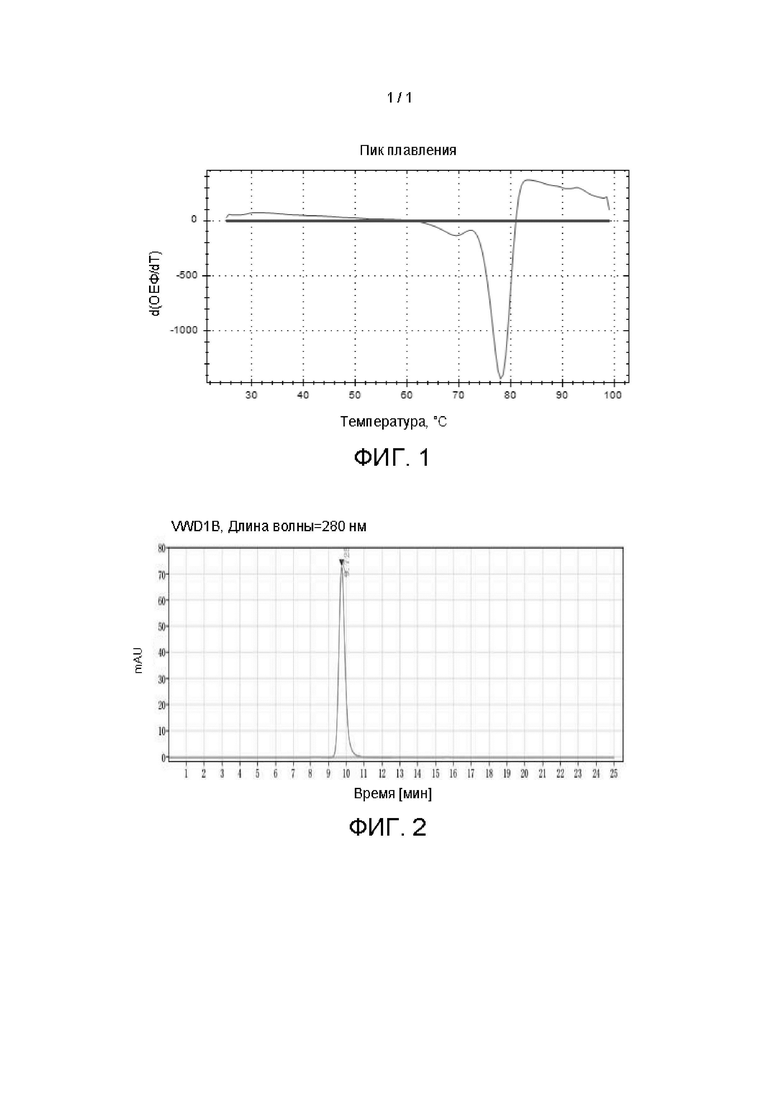
Настоящее изобретение относится к области биотехнологии, в частности к новому антителу, и может быть использовано в медицине. Изобретение позволяет получить выделенное моноклональное антитело, способное связываться с PD-1 с повышенной аффинностью, и раскрывает возможность его применения, в том числе и в составе фармацевтической композиции, для подавления роста опухолевых клеток при терапии различных раковых заболеваний. 3 н. и 8 з.п. ф-лы, 12 табл., 2 ил., 10 пр.
1. Выделенное моноклональное антитело или его антиген-связывающая часть, которое связывается с PD-1 и содержит:
вариабельную область тяжелой цепи, содержащую участок тяжелой цепи CDR1, участок тяжелой цепи CDR2 и участок тяжелой цепи CDR3, и
вариабельную область легкой цепи, содержащую участок легкой цепи CDR1, участок легкой цепи CDR2 и участок легкой цепи CDR3,
где указанные участок тяжелой цепи CDR1, участок тяжелой цепи CDR2, участок тяжелой цепи CDR3, участок легкой цепи CDR1, участок легкой цепи CDR2 и участок легкой цепи CDR3 содержат аминокислотные последовательности, представленные в последовательностях
(1) SEQ ID NO: 1, 2, 3, 34, 35, и 36, соответственно;
(2) SEQ ID NO: 4, 5, 6, 37, 38 и 39, соответственно;
(3) SEQ ID NO: 7, 8, 9, 40, 41 и 42, соответственно;
(4) SEQ ID NO: 10, 11, 12, 43, 44 и 45, соответственно;
(5) SEQ ID NO: 13, 14, 15, 46, 47 и 48, соответственно;
(6) SEQ ID NO: 16, 17, 18, 49, 50 и 51, соответственно;
(7) SEQ ID NO: 19, 20, 21, 52, 53 и 54, соответственно;
(8) SEQ ID NO: 22, 23, 24, 55, 56 и 57, соответственно;
(9) SEQ ID NO: 25, 26, 27, 58, 59 и 60, соответственно;
(10) SEQ ID NO: 28, 29, 30, 61, 62 и 63, соответственно; или
(11) SEQ ID NO: 31, 32, 33, 64, 65 и 66, соответственно.
2. Выделенное моноклональное антитело или его антиген-связывающая часть по п. 1, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где указанные вариабельные области тяжелой цепи и легкой цепи содержат аминокислотные последовательности, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичные последовательностям
(1) SEQ ID NO: 67 и 80 соответственно;
(2) SEQ ID NO: 68 и 81 соответственно;
(3) SEQ ID NO: 69 и 81 соответственно;
(4) SEQ ID NO: 68 и 82 соответственно;
(5) SEQ ID NO: 68 и 83 соответственно;
(6) SEQ ID NO: 68 и 84 соответственно;
(7) SEQ ID NO: 68 и 85 соответственно;
(8) SEQ ID NO: 68 и 86 соответственно;
(9) SEQ ID NO: 69 и 86 соответственно;
(10) SEQ ID NO: 70 и 87 соответственно;
(11) SEQ ID NO: 71 и 88 соответственно;
(12) SEQ ID NO: 72 и 89 соответственно;
(13) SEQ ID NO: 73 и 90 соответственно;
(14) SEQ ID NO: 74 и 91 соответственно;
(15) SEQ ID NO: 75 и 92 соответственно;
(16) SEQ ID NO: 76 и 93 соответственно;
(17) SEQ ID NO: 77 и 94 соответственно;
(18) SEQ ID NO: 78 и 95 соответственно или
(19) SEQ ID NO: 79 и 96 соответственно.
3. Выделенное моноклональное антитело или его антиген-связывающая часть по п. 1, содержащее константную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичную последовательности SEQ ID NO: 97, 99 или 129, и/или константную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичную последовательности SEQ ID NO: 98 или 130.
4. Антитело или его антиген-связывающая часть по п. 1, которое (a) связывается с PD-1 человека; (b) связывается с PD-1 обезьяны; (c) ингибирует связывание PD-L1 с PD-1; (d) повышает пролиферацию T-клеток; (e) стимулирует иммунный ответ и/или (f) стимулирует антиген-специфичный T-клеточный ответ.
5. Антитело или его антиген-связывающая часть по п. 1, представляющие собой мышиное, человеческое, гибридное или гуманизированное антитело.
6. Антитело или его антиген-связывающая часть по п. 1, представляющие собой изотип IgG1, IgG2 или IgG4.
7. Фармацевтическая композиция для лечения рака, содержащая антитело или его антиген-связывающую часть по п. 1 и фармацевтически приемлемый носитель.
8. Фармацевтическая композиция по п. 7, дополнительно содержащая противоопухолевый агент.
9. Способ подавления опухолевого роста у субъекта, включающий введение указанному субъекту терапевтически эффективного количества фармацевтической композиции по п. 7.
10. Способ по п. 9, отличающийся тем, что опухоль представляет собой солидную или не солидную опухоль.
11. Способ по п. 9, отличающийся тем, что опухоль выбрана из группы, состоящей из лимфомы, лейкоза, множественной миеломы, меланомы, аденокарциномы толстой кишки, рака поджелудочной железы, рака толстой кишки, желудочно-кишечного рака, рака предстательной железы, рака мочевого пузыря, рака почки, рака яичника, рака шейки матки, рака молочной железы, рака легких, почечно-клеточного рака и рака носоглотки.
| WO 2018053709 A1, 29.03.2018 | |||
| US 20180011114 A1, 11.01.2018 | |||
| US 20160039921 A1, 11.02.2016 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2013 |
|
RU2599417C2 |

