Область техники, к которой относится изобретение
Данное изобретение относится к антителам и их антиген-связывающим фрагментам, которые связываются с человеческим белком бета-тирозин фосфатазой (HPTPbeta), и их использованию.
Уровень техники
Ангиогенез, отрастание новых кровеносных сосудов от существующей сосудистой сети, играет важную роль в широком диапазоне физиологических и патологических процессов (Nguyen, L.L. et al., Int. Rev. Cytol., 1-48, (2001)). Ангиогенез представляет собой комплексный процесс, опосредуемый взаимосвязью эндотелиальных клеток, которые выстраивают кровеносные сосуды, с окружающей их средой. На ранних стадиях ангиогенеза клетки ткани или опухолевые клетки продуцируют и секретируют факторы проангиогенного роста в ответ на стимулы окружающей среды, такие как гипоксию. Эти факторы диффундируют к близкорасположенным эндотелиальным клеткам и стимулируют рецепторы, которые ведут к выработке протеаз, которые разрушают окружающий внеклеточный матрикс. Активированные эндотелиальные клетки начинают мигрировать и пролиферировать в окружающую ткань в направлении источника этих факторов роста (Bussolino, F., Trends Biochem. Sci., 22, 251-256, (1997)). Затем эндотелиальные клетки прекращают пролиферировать и дифференцируются в трубчатые структуры, что является первым этапом в формировании стабильных, зрелых кровеносных сосудов. В дальнейшем, периэндотелиальные клетки, такие как перициты и гладкомускульные клетки, становятся материалом для новых образующихся сосудов на следующем этапе формирования зрелых сосудов.
Ангиогенез регулируется балансом природных про- и антиангиогенных факторов. Фактор роста эндотелия сосудов, фактор роста фибробластов и ангиопоэтин представляют собой некоторые факторы из многочисленных потенциальных проангиогенных факторов роста. Эти лиганды связываются с их соответствующими рецепторными тирозин киназами на поверхности эндотелиальных клеток и трансдуцируют сигналы, которые способствуют миграции и пролиферации клеток. Хотя многие регуляторные факторы уже идентифицированы, молекулярные механизмы, которые ведут этот процесс, все еще не поняты полностью.
Существует много болезненных состояний, к которым приводит устойчивый нерегулируемый или неправильно регулируемый ангиогенез. При таких болезненных состояниях нерегулируемый или неправильно регулируемый ангиогенез может вызывать либо конкретную болезнь, либо обострить существующее патологическое состояние. Например, глазная неоваскуляризация вовлечена в наиболее частые случаи слепоты и лежит в основе патологии примерно двадцати глазных заболеваний. При некоторых предварительно существующих состояниях, таких как артрит, вновь образующиеся капиллярные кровеносные сосуды внедряются в суставы и разрушают хрящи. При диабете новые капилляры, образовавшиеся в сетчатке, проникают в тканевую жидкость стекловидного тела, вызывая кровотечение и слепоту.
Как рост, так и метастазирование твердых опухолей также могут быть зависимыми от ангиогенеза, Folkman et al., "Tumor Angiogenesis", Chapter 10, 206-32, in the Molecular Basis of Cancer, Mendelsohn et al., eds., W. B. Saunders, (1995). Показано, что опухоли, имеющие размер более 2 мм в диаметре, должны получать свое собственное кровоснабжение, и получают его путем индуцирования роста новых капиллярных кровеносных сосудов. После этого новые кровеносные сосуды внедряются в опухоль, обеспечивая питание и факторы роста, специфические для роста опухоли, а также средства для введения опухолевых клеток в циркуляцию и метастазирования в отдаленные места, такие как печень, легкое или кость (Weidner, New Eng. J. Med., 324, 1, 1-8 (1991)). Природные ингибиторы ангиогенеза, когда они используются в качестве лекарства для имеющих опухоли животных, могут предотвращать рост небольших опухолей (O'Reilly et al., Cell, 79, 315-28 (1994)). Согласно некоторым протоколам, применение таких ингибиторов ведет к регрессии опухоли и латентности даже после прекращения лечения (O'Reilly et al., Cell, 88, 277-85 (1997)). Более того, применение ингибиторов ангиогенеза к некоторым опухолям может усиливать их ответ на другие терапевтические режимы (см., к примеру, Teischer et al., Int. J. Cancer, 57, 920-25 (1994)).
Хотя многие болезненные состояния управляются устойчивым нерегулируемым или неправильно регулируемым ангиогенезом, некоторые болезненные состояния можно лечить посредством увеличения ангиогенеза. Рост и восстановление тканей являются биологическими событиями, при которых происходят клеточная пролиферация и ангиогенез. Например, важным аспектом заживления ран является реваскуляризация поврежденной ткани посредством ангиогенеза.
Хронические, незаживающие раны являются главной причиной продолжительных болезней в популяции пожилых людей. Это особенно относится к лежачим пациентам или диабетикам, у которых развиваются тяжелые незаживающие раки кожи. Во многих таких случаях задержка в лечении является результатом неадекватного кровоснабжения либо как результата продолжительного давления, либо из-за закупорки сосудов. Слабая капиллярная циркуляция вследствие небольшого артериального атеросклероза или венозного застоя способствует неудаче в восстановлении поврежденной ткани. Такие ткани часто инфицируются микроорганизмами, которые неудержимо размножаются с помощью врожденных защитных систем тела, которым требуется хорошо васкуляризованная ткань для эффективного уничтожения патогенных организмов. Как результат, в большинстве центров терапевтического вмешательства для восстановления кровотока в ишемических тканях разрешается допуск питательных веществ и иммуно-логических факторов в места ран.
Атеросклеротические изменения в крупных сосудах могут вызывать ишемию тканей, которую можно было бы уменьшить модуляцией роста кровеносных сосудов к измененной ткани. Например, атеросклеротические изменения в коронарных артериях могут вызывать стенокардию и инфаркт миокарда, которые можно было бы предотвратить, если восстановить кровоток посредством стимуляции роста коллатеральных артерий. Аналогично, атеросклеротические изменения в больших артериях, которые снабжают кровью ноги, могут вызывать ишемию в скелетных мышцах, что ограничивает подвижность и в некоторых случаях вызывает необходимость ампутации, которую также можно предотвратить улучшением кровотока с помощью ангиогенной терапии.
Другие болезни, такие как диабет и гипертония, характеризуются уменьшением количества и плотности небольших кровеносных сосудов, таких как артериолы и капилляры. Эти небольшие кровеносные сосуды играют важную роль в доставке кислорода и питательных веществ. Уменьшение количества и плотности этих сосудов способствует неблагоприятным последствиям гипертонии и диабета, включая хромоту, ишемические язвы, повышенное давление и почечную недостаточность. Эти обычные нарушения и многие другие, менее обычные недомогания, такие как болезнь Бюргера, могли бы быть уменьшены посредством увеличения количества и плотности небольших кровеносных сосудов с помощью ангиогенной терапии.
Таким образом, имеется постоянная необходимость в идентификации регуляторов ангиогенеза.
В свете отмеченного выше, имеется необходимость идентифицировать биохимические мишени при лечении связанных с ангиогенезом нарушений. Однако ангиогенез включает действие множества факторов роста и родственных им рецепторных тирозин киназ (RTKs) (Yancopoulos et al., Nature, 407, 242-248, 2000). Фактор роста эндотелия сосудов (VEGF), например, важен для дифференциации эндотелиальных клеток в возникающие кровеносные сосуды в сосудистой сети зародыша. Кроме того, VEGF усиливает развитие сосудов в зрелую сосудистую сеть. Введение экзогенного VEGF усиливает развитие коллатеральной сосудистой сети и улучшает кровоток к ишемическим тканям.
На сегодняшний день идентифицировано три VEGF RTKs, VEGFR1 (FLT-1), VEGFR2 (KDR) и VEGF3 (FLT-4). Несмотря на то, что эти три рецептора являются высоко консервативными, что основывается на биохимических характеристиках и биологической активности, каждый имеет специфические и не перекрывающиеся функции. Считается, что из этих трех рецепторов VEGFR2 играет преобладающую роль в опосредовании VEGF-влияния при развитии сосудистой системы и во время ангиогенеза у взрослых. Однако для нормального развития сосудистой системы зародыша требуются также и VEGFR1, и VEGFR3, и, возможно, они имеют важное значение для ангиогенеза в зрелых тканях. После VEGF-связывания и димеризации конформационное изменение в VEGFR2-киназном домене усиливает свою киназную активность, приводя к «ауто-фосфорилированию» другого члена пары на специфических тирозиновых остатках. Эти события аутофосфорилирования служат дальнейшему увеличению киназной активности и обеспечивают якорные точки для соединения интрацеллюлярных сигнальных молекул.
Тем не менее, активация единственного ангиогенного пути может быть недостаточной, чтобы производить устойчивые и функциональные сосуды, которые обеспечивали бы адекватную перфузию к ишемическим тканям. Эти факты, наряду с тем, что многочисленные RTKs вовлечены в сборку сосудистой системы зародыша, указывают, что биохимические мишени, которые модулируют многочисленные ангиогенные пути, будут иметь преимущества над введением единственного фактора роста.
Белковые тирозин фосфатазы (PTPs) содержат большое семейство близкородственных ферментов, которые дефосфорилируют белки, содержащие фосфотирозиновые остатки. Недавние данные позволяют предположить, что одна из функций PTPs заключается в ограничении фосфорилирования и активации RTKs. Например, было показано, что НСРТРА, белковая тирозин фосфатаза с низким молекулярным весом, связывается с VEGFR2 и негативно регулирует его активацию в культивируемых эндотелиальных клетках и его биологическую активность в анализах ангиогенеза (Huang et al., Journal of Biological Chemistry, 274, 38183-38185, 1999).
Помимо VEGFR2, сигнализирующего вход от другой RTK, рецептор Tie-2 также является важным для ангиогенеза (Ang1 и Ang2). Делеция Ang1- либо Tie-2-гена у мыши может привести к летальности зародыша, вторичной ненормальности в развитии сосудистой системы (Yancopoulos et al., Nature, 407, 242-248, 2000). Кроме того, сверхэкспрессия Ang1 в коже увеличивает сосудистость кожи, а введение экзогенного Ang1 увеличивает кровоток к ишемической скелетной мышце (Sun et al., J. Clin. Invest., Science, 282, 468-471, 1998). Более того, ингибирование активации Tie-2 подавляет ангио-генез и ограничивает прогрессирование опухоли на животных моделях рака (Lin et al., J. Clin. Invest., 100, 2072-2078, 1997). Помимо своих ангиогенных активностей активация Tie-2 экзогенным введением Ang1 блокирует опосредуемое VEGF рассеивание сосудов и провоспалительные эффекты, но увеличивает его ангиогенные эффекты (Thurston et al., Nature Medicine, 6, 460-463, 2000). Поэтому биологические мишени, которые модулируют как VEGFR2, так и Tie-2, могут дать превосходную проангиогенную или антиангиогенную терапию.
HPTPbeta (впервые описанную у Kruegar et al., EMBO J., 9 (1990)) предложено использовать для модулирования активности ангиобелка рецепторного типа тирозин киназы Tie-2 (к примеру, WO 00/65088). HPTPbeta также предлагается использовать для регулирования активностей VEGFR2, к примеру, опубликованная заявка на патент США №2004/0077065.
Было бы желательным получить антитела, к примеру, гуманизированное моноклональное антитело, которое селективно регулирует активность HPTPbeta и тем самым усиливает ангиогенное сигнализирование, стимулирует рост кровеносных сосудов (ангиогенез) и (или) увеличивает кровоток к ткани, на которую оказывается воздействие, или уменьшает ангиогенное сигнализирование, уменьшает рост кровеносных сосудов и (или) уменьшает кровоток к ткани, на которую оказывается воздействие. Здесь описываются антитела и их фрагменты, которые связывают HPTPbeta и регулируют ангиогенное сигнализирование клеток, которое, в свою очередь, регулирует ангиогенез.
Сущность изобретения
Настоящее изобретение относится к антителам, которые связывают человеческий белок тирозин фосфатазу бета HPTPbeta и тем самым регулируют ангиогенное клеточное сигнализирование, которое, в свою очередь, регулирует ангиогенез.
В одном варианте осуществления изобретение относится к выделенному антителу или его антиген-связывающему фрагменту, который связывается с человеческой тирозин фосфатазой бета, где упомянутое антитело или его антиген-связывающий фрагмент регулирует ангиогенное клеточное сигнализирование, которое, в свою очередь, регулирует ангиогенез.
В другом варианте осуществления изобретение относится к антителу, которое связывает N-концевую часть человеческого белка тирозин фосфатазы бета.
В другом варианте осуществления изобретение относится к антителу, которое связывает первый FN3-повтор человеческого белка тирозин фосфатазы бета.
В другом варианте осуществления изобретение относится к антителу, которое связывает первый FN3-повтор человеческого белка тирозин фосфатазы бета, где первый FN3-повтор человеческого белка тирозин фосфатазы бета имеет последовательность, как показано в SEQ ID:11, или ее части.
В другом варианте осуществления изобретение относится к антителу, где антитело является моноклональным антителом.
В другом варианте осуществления изобретение относится к антителу, где антитело является моноклональным антителом R15E6 (мышиная гибридома, Balbc-клетки селезенки (В-клетки), депонированные в Американской Коллекции Типовых Культур (АТСС), P.O. Box 1549, Manassas, VA 20108 USA 4 мая 2006, под АТСС № РТА-7580).
В другом варианте осуществления изобретение относится к антителу, имеющему те же самые, или практически те же самые, биологические характеристики, что и R15E6.
В другом варианте осуществления изобретение относится к антителу, где антитело или антиген-связывающий фрагмент является гуманизированным.
В другом варианте осуществления изобретение относится к антителу, где антитело содержит антиген-связывающие остатки области из моноклонального антитела R15E6 и является гуманизированным.
В другом варианте осуществления изобретение относится к антиген-связывающему фрагменту антитела, причем этот фрагмент содержит вариабельные области тяжелой и легкой цепей.
В другом варианте осуществления изобретение относится к антиген-связывающему фрагменту антитела, где антиген-связывающий фрагмент выбран из группы, состоящей из Fv-фрагмента, Fab-фрагмента, Fab'-фрагмента и F(ab')2-фрагмента.
В другом варианте осуществления изобретение относится к способу лечения регулируемого ангиогенезом нарушения у субъекта, содержащему этапы, на которых идентифицируют субъекта, нуждающегося в регулировании ангиогенеза, и вводят субъекту эффективное количество антитела или его антиген-связывающего фрагмента, которые связывают HPTPbeta и регулируют ангиогенез.
В другом варианте осуществления изобретение относится к способу лечения регулируемого ангиогенезом нарушения у субъекта, где регулируемое ангиогенезом нарушение является нарушением с повышенным ангиогенезом и выбрано из группы, состоящей из диабетической ретинопатии, деградации желтого пятна сетчатки глаза, рака, серповидно-клеточной анемии, саркоида, сифилиса, псевдоксантомы эластикум, болезни Педжета, окклюзии вен, окклюзии артерий, обструктивного заболевания сонной артерии, хронического увеита/витрита, микобактериальной инфекции, болезни Лайма, системной волчанки (волчаночный эритематоз), ретинопатии недоношенности, болезни Илза, болезни Бехчета, инфекций, вызванных ретинитом или хориоидитом, предполагаемого глазного гистоплазмоза, болезни Беста, миопии, глазных ямок, болезни Старгардта, pars planitis, хронического отслоения сетчатки, синдрома гипервязкости, токсо-плазмоза, травмы и пост-лазерных осложнений, болезней, связанных с покраснением радужки, и пролиферативной витреоретинопатии.
В другом варианте осуществления изобретение относится к способу лечения регулируемого ангиогенезом нарушения у субъекта, где регулируемое ангиогенезом нарушение является нарушением с повышенным ангиогенезом и выбрано из группы, включающей в себя - но не ограниченной ими - диабетическую ретинопатию, деградацию желтого пятна сетчатки глаза, рак, ревматоидный артрит, гемангиому, болезнь Рандю-Вебера-Ослера, или наследственную геморрагическую телеангиэктазию, и твердые или переносимые кровью опухоли.
В другом варианте осуществления изобретение относится к способу лечения регулируемого ангиогенезом нарушения у субъекта, где регулируемое ангиогенезом нарушение является нарушением с повышенным ангиогенезом и выбрано из группы, состоящей из воспалительных кишечных заболеваний, таких как болезнь Крона и язвенные колиты, псориаза, саркоидоза, ревматоидного артрита, гемангиомы, болезни Рандю-Вебера-Ослера, или наследственной геморрагической телеангиэктазии, твердых или переносимых кровью опухолей и синдрома приобретенного иммунодефицита.
В другом варианте осуществления изобретение относится к способу лечения регулируемого ангиогенезом нарушения у субъекта, где регулируемое ангиогенезом нарушение является нарушением с пониженным ангиогенезом и выбрано из группы, включающей в себя - но не ограниченной ими - ишемию скелетной мышцы или миокарда, паралич, заболевание коронарной артерии, заболевание периферических сосудов, заболевание мозговых сосудов, диабетическую нейропатию и раневые заживления.
В другом варианте осуществления изобретение относится к способу лечения регулируемого ангиогенезом нарушения у субъекта, где регулируемое ангиогенезом нарушение является нарушением с пониженным ангиогенезом и выбрано из группы, состоящей из ишемии скелетной мышцы или миокарда, паралича, заболевания коронарной артерии, заболевания периферических сосудов.
В другом варианте осуществления изобретение относится к способу лечения нарушения с пониженным ангиогенезом у субъекта, где нарушение с пониженным ангиогенезом является заболеванием периферических сосудов.
В другом варианте осуществления изобретение относится к способу лечения нарушения с пониженным ангиогенезом у субъекта, где нарушение с пониженным ангиогенезом является заболеванием коронарной артерии.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей антитело или его фрагмент, который связывается с человеческим белком тирозин фосфатазой бета, и фармацевтически приемлемый носитель.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей антитело или его фрагмент, который связывается с человеческим белком тирозин фосфатазой бета, где антитело является моноклональным антителом R15E6, и фармацевтически приемлемый носитель.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей антитело или его фрагмент, который связывается с человеческим белком тирозин фосфатазой бета, где антитело является моноклональным антителом, имеющим те же самые, или практически те же самые биологические характеристики, как и R15E6, и фармацевтически приемлемый носитель.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей антитело или его фрагмент, который связывается с человеческим белком тирозин фосфатазой бета, где антитело или антиген-связывающий фрагмент является гуманизированным, и фармацевтически приемлемый носитель.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей антитело или его фрагмент, который связывается с человеческим белком тирозин фосфатазой бета, где антитело содержит антиген-связывающую область остатков из моноклонального антитела R15E6 и является гуманизированным, и фармацевтически приемлемый носитель.
Краткое описание чертежей
Фиг.1. Конструкция и получение НРТРβ ECD-белка. (Панель А) Схематическое представление НРТРβ полной длины и НРТРβ внеклеточного домена-6His слитого белка. (Панель В) Серебряное пятно имидазола элюируется с Ni-NTA-колонки, нагруженной супернатантом из HEK293-клеток, трансфецированных вектором, направляющим экспрессию βECD-6His. Обнаружена единственная полоса с высоким молекулярным весом, согласующаяся с внеклеточным доменом-61-tis НРТРβ-белка.
Фиг.2. R15E6 распознает эндогенную НРТРβ в эндотелиальных клетках. (Панель А) Лизаты эндотелиальных клеток осаждают контрольным антителом (Дорожка 1), R15E6 (Дорожка 2) или смесью анти-Tie2 и анти-VEGFR2 антител (Дорожка 3). Иммунопреципитаты растворяют с помощью SDS-PAGE, передают на PVD-мембрану и зондируют посредством western-блота со смесью R15E6, анти-Tie2 и анти-VEGFR2 антител. Единственная главная полоса с высоким молекулярным весом согласующаяся с НРТРβ, видна с R15E6 (Дорожка 2), но не видна для контрольного антитела (Дорожка 1) или смеси анти-Tie2 и анти-VEGFR2 (Дорожка 3). (Панель В) Эндотелиальные клетки подвергают FACS-анализу с помощью R15E6 (белый пик) или контролю без какого-либо первичного антитела (черный пик). Ясный сдвиг по флуоресценции указывает, что R15E6 связывается с НРТРβ на поверхности интактных эндотелиальных клеток.
Фиг.3. R15E6 усиливает Tie2-рецепторную активацию в HUVEC's. Tie2 активацию измеряют в эндотелиальных клетках человека, как описано в Примере 4. В зависимости от дозы R15E6 усиливает как базальную, так и Ang1-индуцированную Tie2-активацию.
Фиг.4. R15E6 увеличивает HUVEC-выживание. Выживание сыворотки голодающих эндотелиальных клеток человека измеряют как описано в Примере 4. В соответствии с его эффектами по Tie2-активации R15E6 в зависимости от дозы увеличивает выживание как базальных, так и Ang1-индуцированных эндотелиальных клеток (Панель А). Кроме того, R15E6 также в зависимости от дозы увеличивает выживание VEGF- и FGF-опосредованных эндотелиальных клеток (Панели В и С). Контрольное антитело не увеличивает выживание эндотелиальных клеток (Панель D).
Фиг.5. R15E6 увеличивает HUVEC-миграцию. Миграцию человеческих эндотелиальных клеток измеряют как описано в Примере 4. В зависимости от дозы R15E6 усиливает как базальную, так и VEGF-индуцированную миграцию эндотелиальных клеток.
Фиг.6. R15E6 увеличивает капиллярный морфогенез в HUVEC/Bead Sprouting анализах. Капиллярный морфогенез человеческих эндотелиальных клеток измеряют в анализе разрастания с помощью бусинок, как описано в Примере 4. R15E6 усиливает капиллярный морфогенез как базальных, так и VEGF-индуцированных эндотелиальных клеток.
Фиг.7. Western-блот анализ локализует R15E6 связывающий эпитоп на N-концевом FN3-повторе внеклеточного домена НРТРβ. (Панель А) Согласно western-анализу R15E6 связывается со всеми мутантами, имеющими С-концевые усечения, демонстрируя, что связывающий эпитоп расположен на N-концевых 2 FN3-повторах. (Панель В) Анализ химерных белков человек/мышь далее локализует R15E6 связывающий эпитоп на N-концевом FN3-повторе НРТРβ.
Фиг.8. MSD-анализ подтверждает локализацию R15E6 связывающего эпитопа на N-концевом FN3-повторе внеклеточного домена НРТРβ. (Панель А) Согласно MSD-анализу R15E6 связывается со всеми мутантами, имеющими С-концевые усечения, подтверждая, что связывающий эпитоп расположен на N-концевых 2 FN3-повторах. (Панель В) Анализ химерных белков человек/мышь далее подтверждает локализацию R15E6 связывающего эпитопа на N-концевом FN3-повторе НРТРβ.
Фиг.9. MSD-анализ показывает, что моновалентный Fab-фрагмент R15E6 также связывается с N-концевым FN3-повтором НРТРβ. (Панель А) Подбно интактному R15E6 антителу Fab-фрагмент R15E6 связывается со всеми мутантами, имеющими С-концевые усечения, подтверждая, что связывающий эпитоп локализуется на N-концевых 2 FN3-повторах. (Панель В) Анализ химерных белков человек/мышь далее локализует связывающий эпитоп Fab-фрагмента R15E6 на N-концевом FN3-повторе НРТРβ.
Фиг.10. Моновалентный Fab-фрагмент R15E6 не усиливает Tie2-активацию и блокирует Tie2-активацию интактным R15E6.
Фиг.11. Fab-фрагмент R15E6 мощно ингибирует выживание эндотелиальных клеток. (Панель А) По сравнению с контрольным Fab-фрагментом Fab-фрагмент R15E6 мощно ингибирует выживание эндотелиальных клеток. (Панель В) Ингибиторное действие Fab-фрагмента R15E6 нейтрализуют посредством конкуренции с интактным R15E6.
Фиг.12. Fab-фрагмент R15E6 ингибирует опосредуемую VEGF миграцию эндотелиальных клеток.
Описание перечня последовательностей
Каждая из нуклеотидных и белковых последовательностей, приведенных в перечне последовательностей, наряду с соответствующим(-и) номером(-ами) доступа в Genbank или Derwent, где это уместно, и видами, из которых она происходит, показана в Таблице I.
Подробное описание изобретения
Настоящее изобретение относится к антителам, которые связывают HPTPbeta, и их применению.
Для получения рекомбинантных ДНК, олигонуклеотидного синтеза, тканевой культуры и трансформации (к примеру, электропорации, липофекции) могут быть использованы стандартные методики. Ферментативные реакции и методики очистки могут выполняться согласно спецификациям производителей или так, как они обычно выполняются в данной области техники, или как описывается здесь. Методики и процедуры обычно выполняются в соответствии с традиционными методами, известными в данной области и как описано в различных общих и более конкретных руководствах, которые цитируются и обсуждаются в описании настоящего изобретения. Если не приводится конкретных определений, для обозначения описываемых здесь лабораторных процедур и методик аналитической химии, химии органического синтеза, медицинской и фармацевтической химии используется терминология, известная и общепринятая в данной области техники. Для химического синтеза, химических анализов, приготовления фармацевтический препаратов, рецептур, методов введения и лечения пациентов могут быть использованы стандартные методики.
Следующие термины, если не указано иначе, должны пониматься в следующих значениях:
«Белок» используется здесь взаимозаменяемо с пептидом и полипептидом. HPTPbeta является человеческим белком тирозин фосфатазой, как определено в перечне последовательностей. В некоторых вариантах осуществления используются различные фрагменты HPTPbeta. Гомологи, ортологи, фрагменты, варианты и мутанты HPTPbeta-белка и гена, как описано ниже, считаются входящими в объем термина «HPTPbeta».
Под «фрагментом» понимается часть нуклеотидной или белковой последовательности. Фрагменты могут сохранять биологическую активность нативного белка. Фрагменты нуклеотидной последовательности полезны также как гибридизационные зонды и праймеры или для регуляции экспрессии гена, к примеру, антисмысловой, siPHK или микроРНК. Биологически активную часть можно получить посредством выделения части одной из нуклеотидных последовательностей согласно изобретению, экспрессирования кодируемой части (к примеру, путем рекомбинантной экспрессии in vitro) и оценки активности кодируемого белка.
Специалист в данной области техники признал бы, что гены и белки из разных видов, иных нежели перечисленные в перечне последовательностей, особенно видов позвоночных, могут быть полезными. Такие виды включают в себя - но не ограничиваются ими - мышей, крыс, морских свинок, кроликов, собак, свиней, коз, коров, обезьян, шимпанзе, овец, хомяков и данио. Специалист в данной области техники также признал бы, что, используя зонды из последовательностей известных видов, можно получить кДНК или геномные последовательности, гомологичные известной последовательности, из тех же самых или альтернативных видов посредством известных методов клонирования. Подразумевается, что такие гомологи и ортологи могут быть полезными в практическом применении изобретения.
Под «вариантами» подразумеваются сходные последовательности. Например, консервативные варианты могут включать в себя такие последовательности, которые, ввиду вырожденности генетического кода, кодируют аминокислотную последовательность одного из полипептидов согласно изобретению. Природно встречающиеся аллельные варианты и сплайсированные варианты могут быть идентифицированы с использованием известных методик, к примеру, с помощью полимеразной цепной реакции (PCR), анализа точечного нуклеотидного полиморфизма (SNP) и гибридизационных методик. Для выделения ортологов и гомологов обычно используют строгие условия гибридизации, диктуемые конкретными последовательностями, длиной последовательности, содержанием гуанин + цитозин (GC) и другие параметры. Варианты нуклеотидных последовательностей также включают в себя синтетически произведенные нуклеотидные последовательности, к примеру, произведенные с использованием сайт-направленного мутагенеза. Варианты могут содержать дополнительные последовательности из одного геномного локуса или в комбинации с другими последовательностями.
Молекулы согласно изобретению также включают усеченные и (или) мутированные белки, где области белка, не требующиеся для лигандного связывания или передачи сигналов, делегированы или модифицированы. Аналогично, они могут быть мутированы для модификации их лигандного связывания или сигнальных активностей. Такие мутации могут включать не-консервативные мутации, делеции или вставки аминокислот или белковых доменов. Варианты белков могут сохранять, а могут не сохранять биологическую активность. Такие варианты могут происходить из, к примеру, генетического полиморфизма или от манипуляции человека.
Здесь также имеются в виду слитые белки. Используя известные методы, специалист в данной области техники смог бы получить слитые белки из белков согласно изобретению, которые, несмотря на отличие от нативной формы, также могут быть полезными. Например, партнером слияния может являться последовательность сигнального (или лидерного) полипептида, которая ко-трансляционно или пост-трансляционно направляет передачу белка из одного участка его синтеза в другой участок (к примеру, лидер дрожжевого α-фактора). Альтернативно, он может быть добавлен для облегчения очистки или идентификации белка согласно изобретению (к примеру, поли-His, Glag-пептид или флюоресцентные белки).
Термин «антиген» относится к молекуле или части молекулы, способной связываться селективным агентом связывания, таким как антитело, и дополнительно способной быть используемой в животных, чтобы производить антитела, способные связываться с эпитопом этого антигена. Антиген может иметь один или более эпитопов.
Термин «эпитоп» включает в себя любую антигенную детерминанту, предпочтительно полипептидную детерминанту, способную к специфическому связыванию с иммуноглобулином или Т-клеточным рецептором. В некоторых вариантах осуществления эпитопные детерминанты включают в себя химически активные поверхностные группы, такие как аминокислоты, сахара, липиды, фосфорильные или сульфонильные группы, и в некоторых вариантах осуществления могут иметь специфические характеристики пространственной структуры, и (или) специфические характеристики заряда. Эпитоп представляет собой область антигена, которая связывается с антителом. В некоторых вариантах осуществления антитело, как говорят, специфически связывается с антигеном, когда распознает предпочтительно свой мишеневый антиген в сложной смеси белков и (или) макромолекул. Антитело также, как говорят, специфически связывается с антигеном, когда он обнаруживает более высокое сродство с антигеном, чем с другими родственными и (или) неродственными молекулами.
Термин «антитело» (Ab), как используется здесь, включает в себя моноклональные антитела, поликлональные антитела, мультиспецифические антитела (к примеру, биспецифические антитела), одноцепочечные антитела, к примеру, антитела из ламы и верблюда, фрагменты антител, к примеру, фрагменты вариабельных областей и (или) константных областей, в той мере, насколько они обнаруживают желательную биологическую активность, к примеру, антиген-связывающую активность. Термин «иммуноглобулин» (Ig) используется здесь взаимозаменяемо с термином «антитело».
«Выделенное антитело» - это антитело, которое идентифицировано и (или) отделено, и (или) восстановлено из его природного окружения.
Основная единица четырехцепочечного антитела является гетеротетрамерным гликобелком, построенным из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей (антитело IgM состоит из 5 основных гетеротетрамерных единиц и дополнительного полипептида, называемого J-цепью, и поэтому содержит 10 сайтов антигенного связывания, в то время как секретируемые антитела IgA могут полимеризоваться с образованием поливалентных образований, содержащих 2-5 основных 4-цепочечных единиц вместе с J-цепью). В случае IgG, четырехцепочечная единица обычно имеет массу около 150 килодальтон (kDa). Каждая L-цепь связана с Н-цепью посредством одной ковалентной дисульфидной связи, в то время как две Н-цепи связаны одна с другой посредством одной или более дисульфидных связей в зависимости от изотипа Н-цепи. Каждая Н- и L-цепь имеет также регулярно размещенные внутри себя дисульфидные мостики. Каждая Н-цепь имеет на N-конце вариабельный домен (VH), за которым следуют три константных домена (CH) для каждой α- и γ-цепей и четыре CH-домена для µ- и ε-изотипов. Каждая L-цепь имеет на N-конце вариабельный домен (VL), за которым следует константный домен (CL) на ее другом конце. VL линейно соединена с VH, a CL линейно соединена с первым константным доменом тяжелой цепи (CH1). Предполагается, что конкретные аминокислотные остатки образуют стык между вариабельными доменами легкой цепи и тяжелой цепи. Спаривание VH и VL образует единичный антиген-связывающий сайт. В отношении структуры и свойств различных классов антител см., к примеру. Basic and Clinical Immunology, 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, 1994, page 71 and Chapter 6.
L-цепь из любого вида позвоночных может быть отнесена к одному из двух ясно различимых типов, называемых каппа и лямбда, на основании аминокислотных последовательностей их константных доменов. В зависимости от аминокислотной последовательности константного домена их тяжелых цепей (CH), иммуноглобулины могут быть отнесены к разным классам или изотипам. Имеется пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, имеющие тяжелые цепи, обозначаемые α, δ, ε, γ и µ, соответственно. γ- и α-классы далее подразделяются на подклассы на основании относительно небольших различий в последовательности и функции CH, к примеру, у человека экспрессируются следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
Члены семейства Camelidae, к примеру, лама, верблюд и дромадеры, содержат уникальный тип антитела, который лишен легких цепей, а также у него отсутствует CH1-домен (Muyldermans, S., Rev. Mol. Biotechnol., 74, 277-302 (2001)). Вариабельную область этих тяжелоцепочечных антител обозначают VHH или VHH, и она составляет самый маленький доступный интактный антиген-связывающий фрагмент (15 кДа), происходящий из функционального иммуноглобулина.
Термин «вариабельный» относится к тому факту, что определенные сегменты вариабельных доменов различаются в широких пределах в последовательностях у антител. V-домен опосредует антигенное связывание и определяет специфичность конкретного антитела к своему антигену. Однако вариабельность неравномерно распределена по 110-аминокислотному диапазону вариабельных доменов. Кроме того, V-области состоят из относительно инвариантных отрезков, называемых обрамляющими областями (FR), из 15-30 аминокислот, разделенных короткими областями вариабельности, называемыми «гипервариабельными областями», каждая из которых имеет длину 5-12 аминокислот. Каждый из вариабельных доменов нативных тяжелой и легкой цепей содержит четыре FGs, в целом имеющих β-складчатую конфигурацию, связанных тремя гипервариабельными областями, которые образуют петлевидные соединения, и в некоторых случаях образующих часть β-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются рядом в тесной близости с помощью FGs, и вместе с гипервариабельными областями другой цепи обеспечивают формирование антиген-связывающего сайта антител. Константные домены непосредственно не включены в связывание антитела с антигеном, но обнаруживают различные эффекторные функции, такие как участие антитела в антител-зависимой клеточной цитотоксичности (ADCC).
Термин «гипервариабельная область», когда он здесь используется, относится к аминокислотным остаткам антитела, которые ответственны за антигенное связывание. Гипервариабельная область обычно содержит аминокислотные остатки из «области, определяющей комплементарность» («CDR») (к примеру, примерно около остатков 24-34 (L1), 50-56 (L2) и 89-97 (L3) в VL и примерно около остатков 1-35 (H1), 50-65 (Р2) и 95-102 (Н3) в VH (Kabat et al., Sequences of Protein of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)), и (или) соответствующие остатки из «гипервариабельной петли».
Термин «моноклональное антитело», как используется здесь, относится к антителу, полученному из популяции существенно гомогенных антител, т.е. индивидуальные антитела, содержащие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в малых количествах. В противоположность препаратам поликлональных антител, которые включают в себя различные антитела, направленные против различных эпитопов, каждое моноклональное антитело направлено против единственного эпитопа, т.е., единственной антигенной детерминанты. Помимо своей специфичности, моноклональные антитела выгодны тем, что они могут быть синтезированы без загрязнения другими антителами. Определение «моноклональное» не истолковывается как требование получения антитела каким-либо конкретным методом. Например, моноклональные антитела, используемые в настоящем изобретении, могут быть приготовлены с помощью гибридомной технологии или могут быть получены с помощью методов рекомбинантных ДНК в бактериальных, эукариотических животных или растительных клетках (см., к примеру, патент США №4.816.567). «Моноклональные антитела» могут быть также выделены из фаговых библиотек антител, используя доступные методики, к примеру, Clackson et al., Nature, 352:624-628 (1991).
Моноклональные антитела включают в себя здесь «химерные» антитела, в которых часть тяжелой и (или) легкой цепи идентична или гомологична соответствующим последовательностям в антителах, происходящих из конкретного вида или принадлежащих к конкретному классу антител или подклассу, в то время как остальная часть цепи(-ей) идентична или гомологична соответствующим последовательностям в антителах, происходящих из другого вида или принадлежащих другому классу антител или подклассу, а также фрагменты таких антител, в той мере, насколько они обнаруживают желательную биологическую активность (см. патент США №4.816.567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81, 6851-6855 (1984)).
«Фрагмент антитела» содержит часть мультимерного антитела, предпочтительно антиген-связывающего антитела или вариабельной области интактного антитела. Примеры фрагментов антитела включают Fab, Fab', F(ab'), димеры и тримеры Fab-конъюгатов, Fv, scFv, минитела, диа-, триа- и тетратела, линейные антитела (см. Hudson et al., Nature Med. 9, 129-134 (2003)).
«Fv» представляет собой минимальный фрагмент антитела, который содержит полный сайт антигенного связывания. Этот фрагмент состоит из димера, образуемого доменами вариабельной области одной тяжелой и вариабельной области одной легкой цепи, находящимися в тесной, нековалентной связи. Из складки этих двух доменов выходят шесть гипервариабельных петель (по 3 петли от Н- и L-цепи), которые дают аминокислотные остатки для антигенного связывания и придают антиген-связывающую специфичность антителу. Однако даже единичный вариабельный домен (или половинка Fv, включающая только три специфичных для антигена CDRs) способен распознавать и связывать антиген и, следовательно, подпадает под определение Fv.
«Одноцепочечные Fv», также обозначаемые сокращенно как «sFv» или «scFv», представляют собой фрагменты антитела, которые содержат VH- и VL-домены антитела, связанные в единичную полипептидную цепочку. Предпочтительно, sFv-полипептид далее содержит полипептидный линкер между VH- и VL-доменами, что позволяет sFv образовывать желательную структуру для антигенного связывания.
Термины «диа-, триа- и тетратела» относятся к маленьким фрагментам антител, полученных конструированием sFv-фрагментов с короткими линкерами (примерно 5-10 остатков) между VH- и VL-доменами, такими что достигается межцепочечное, но не внутрицепочечное спаривание V-доменов, что приводит к получению мультивалентного фрагмента.
Термин «гуманизированное антитело» или «человеческое антитело» относится к антителам, которые содержат последовательности вариабельных областей тяжелой и легкой цепей из вида, не относящегося к человеку (к примеру, из мыши), но в которых по меньшей мере часть VH- и (или) VL-последовательности изменена, чтобы она была «человекоподобной», т.е. более сходной с вариабельными последовательностями, характерными для зародышевой линии человека. Одним из типов гуманизированного антитела является антитело с трансплантированной CDR, в котором человеческие CDR-последовательности введены в не-человеческие VH- и VL-последовательности для замещения соответствующих не-человеческих CDR-последовательностей. Средства для получения химерных антител, антител с CDR-трансплантатами и гуманизированных антител известны специалистам в данной области техники (см., к примеру, патенты США №№4.816.567 и 5.225.539). В одном из методов получения гуманизированных антител используется применение трансгенных животных, таких как трансгенная мышь. Эти трансгенные животные содержат существенную часть генома, продуцирующего человеческие антитела, введенную в их собственный геном, и продуцирование животными их собственных эндогенных антител уменьшается. Методы получения таких трансгенных животных известны в данной области техники. Такие трансгенные животные могут быть получены посредством XenoMouseRTM-технологии или используя «минилокусный» подход. Методы получения XenoMiceRTM описаны в патентах США №№6.162.963, 6.150.584, 6.114.598 и 6.075.181. Методы получения трансгенных животных, используя «минилокусный» подход, описаны в патентах США №№5.545.807 5.545.806 и 5.625.825, а также в заявке WO 93/12227.
Гуманизирование не-человеческого антитела в последние годы стало рутинной процедурой и сейчас входит в объем знаний специалиста в данной области техники. Несколько компаний имеют подразделения по производству гуманизированных антител, к примеру, Xoma, Aries, Medarex, PDL и Cambridge Antibody Technologies. Протоколы гуманизирования широко освещены в технической литературе, к примеру, Kipriyanov and Le Gall, Molecular Biotechnol., Vol.26, pp 39-60 (2004), Humana Press, Totowa, NJ; Lo, Methods Mol. Biol., Vol.248, pp 135-159 (2004), Humana Press, Totowa, NJ; Wu et al., J. Mol. Biol. 294, 151-162 (1999).
В некоторых вариантах осуществления антитела согласно настоящему изобретению можно экспрессировать в клеточных линиях иных нежели гибридомные клеточные линии. Последовательности, кодирующие конкретные антитела, могут быть использованы для трансформации хозяйской клетки подходящего млекопитающего с помощью известных методов введения полинуклеотидов в хозяйскую клетку, в том числе, например, упаковывание полинуклеотида в вирус (или в вирусный вектор) и трансдуцирование хозяйской клетки вирусом, или с помощью трансфекционных процедур, известных в данной области, например, представленных в патентах США №№4.399.216, 4.912.040, 4.740.461 и 4.959.455. Используемая процедура трансформации может зависеть от трансформируемого хозяина. Методы введения гетерологичных полинуклеотидов в клетки млекопитающих известны в данной области и включают в себя - но не ограничены ими - декстран-опосредованную трансфекцию, осаждение фосфатом кальция, полибрен-опосредованную трансфекцию, слияние протопластов, электропорацию, инкапсулирование полинуклеотида(-ов) в липосомы, смешивание нуклеиновой кислоты с положительно заряженными липидами и прямое микроинъецирование ДНК в ядра.
Молекулу нуклеиновой кислоты, кодирующую аминокислотную последовательность константной области тяжелой цепи, вариабельную область тяжелой цепи, константную область легкой цепи или вариабельную область легкой цепи антитела, или его фрагмента, при желании, в подходящей комбинации, вводят в подходящий экспрессионный вектор, используя стандартные методики лигирования. Константная область тяжелой или легкой цепи антитела может быть присоединена к С-концу подходящей вариабельной области и лигирована в экспрессионный вектор. Вектор обычно выбирают из векторов, функциональных в используемых конкретных клетках (т.е. вектор совместим с механизмом функционирования клетки, так что может происходить амплификация гена и (или) экспрессия гена). Обзор по векторам см. в Methods Enzymol. Vol.185 (Goeddel, ed.), 1990, Academic Press.
Антитела и их фрагменты согласно настоящему изобретению связывают HPTPbeta и регулируют ангиогенез. Как определено выше, термин антитело используется для обозначения антиген-связывающего фрагмента. Применения такого антитела и антиген-связывающих фрагментов описывается ниже.
Скрининг-анализы, использующие in vitro- и in vivo-модели ангиогенеза
Антитела согласно изобретению могут быть скринированы анализами ангиогенеза, которые известны в данной области техники. Такие анализы включают в себя in vitro-анализы, в которых измеряются суррогаты роста кровеносных сосудов в культивируемых клетках или образование сосудистых структур из тканевых эксплантов, и in vivo-анализы, в которых прямо или косвенно измеряется рост кровеносных сосудов (Auerbach, R., et al., (2003). Clin Chem 49, 32-40, Vailhe, В., et al. (2001). Lab Invest 81, 439-452).
In vitro-модели ангиогенеза
Большинство этих анализов использует культивируемые эндотелиальные клетки или тканевые экспланты и измеряет эффект агентов по «ангиогенным» клеточным ответам или по образованию капилляроподобных кровеносных структур. Примеры in vitro-анализов ангиогенеза включают в себя - но не ограничиваются ими - анализы миграции и пролиферации эндотелиальных клеток, образования капиллярных трубок, эндотелиального разрастания, анализ аортальных кольцевых эксплантов и анализ дуги аорты цыпленка.
In vivo-модели ангиогенеза
В этих анализах агенты или антитела вводятся локально или систематически в присутствии или отсутствие факторов роста (т.е. VEGF или агниопоэтина 1), и рост новых кровеносных сосудов измеряют прямым наблюдением или измеряя суррогатный маркер, такой как содержание гемоглобина или флюоресцентный индикатор. Примеры ангиогенеза включают в себя - но не ограничиваются ими - хориоаллантисный мембранный анализ цыпленка, анализ роговичного ангиогенеза и MATRIGEL™-plug анализ.
Лечение регулируемых ангиогенезом нарушений
Термин «регулировать» определяется здесь в его словарном значении. Таким образом, термин «регулировать» включает в себя - но не ограничивается им - регулирование против хода или по ходу транскрипции для фиксации, для приведения в порядок или единообразия, чтобы управлять или направлять различные средства. В одном аспекте антитело может использоваться в способе лечения «нарушения с увеличенным ангиогенезом» или «нарушения с уменьшенным ангиогенезом». Как используется здесь, термин «нарушение с увеличенным ангиогенезом» представляет собой нарушение, которое включает в себя нежелательный или увеличенный ангиогенез в биологическом проявлении заболевания, нарушения и (или) состояния; в биологическом каскаде, ведущем к нарушению, или как симптом нарушения. Аналогично, «нарушение с уменьшенным ангиогенезом» представляет собой такое нарушение, которое включает в себя желательный или уменьшенный ангиогенез в биологических проявлениях. Это «включение» в ангиогенез при нарушении с увеличенным/уменьшенным ангиогенезом, без ограничения, может быть следующим:
(1) Ангиогенез как «случай» нарушения или биологического проявления того, увеличен или уменьшен уровень ангиогенеза генетически, инфекцией, аутоиммунно, травмой, биомеханическими причинами, образом жизни или некоторыми другими причинами.
(2) Ангиогенез как часть наблюдаемого проявления заболевания или нарушения. То есть, заболевание или нарушение оценивают в терминах увеличенного или уменьшенного ангиогенеза. С клинической точки зрения, ангиогенез указывает на наличие заболевания. Однако не требуется, чтобы ангиогенез являлся «признаком» заболевания или нарушения.
(3) Ангиогенез является частью биохимического или клеточного каскада, который приводит к заболеванию или нарушению, В этом отношении регуляция ангиогенеза может прервать каскад и может контролировать заболевание. He-ограничивающие примеры регулируемых ангиогенезом нарушений, которые можно лечить, используя настоящее изобретение, описываются здесь ниже.
Антитела согласно настоящему изобретению могут быть использованы для лечения заболеваний, связанных с ретинальной/хороидальной неоваскуляризацией, которые включают в себя - но не ограничиваются ими - диабетическую ретинопатию, деградацию желтого пятна сетчатки глаза, рак, сервовидно-клеточную анемию, pars planitis, хроническое отслоение сетчатки, синдром гипервязкости, токсоплазмоз, травму и пост-лазерные осложнения. Другие заболевания включают в себя - но без ограничения ими - заболевания, связанные с покраснением радужки (неоваскуляризация радужной оболочки глаза) и заболевания, вызванные ненормальной пролиферацией фиброваскулярной или фиброзной ткани, в том числе все формы пролиферативной ретинопатии стекловидного тела, связанной или не связанной с диабетом.
Антитела согласно настоящему изобретению могут быть использованы для лечения заболеваний, связанных с хроническим воспалением. Заболевания с симптомами хронического воспаления включают в себя воспалительные заболевания кишечника, такие как болезнь Крона и язвенный колит, псориаз, саркоидоз и ревматоидный артрит. Ангиогенез является ключевым элементом, который объединяет эти хронические воспалительные заболевания. Хроническое воспаление зависит от постоянного формирования прорастающих капилляров для поддержания притока в воспаленные клетки. Приток и наличие воспалительных клеток производит гранулемы и, таким образом, поддерживает хроническое воспалительное состояние. Ингибирование ангиогенеза композициями и способами согласно настоящему изобретению предотвратило бы образование гранулем и облегчило болезнь.
Болезнь Крона и язвенный колит характеризуются хроническим воспалением и ангиогенезом на различных участках желудочно-кишечного тракта. Болезнь Крона характеризуется хроническим гранулематозным воспалением по всему желудочно-кишечному тракту, состоящим из нескольких прорастающих капилляров, окруженных цилиндром воспалительных клеток. Предотвращение ангиогенеза ингибирует формирование проращивания и предотвращает образование гранулемасс. Болезнь Крона встречается как хроническое трансмуральное воспалительное заболевание, которое весьма общим образом воздействует на дистальные отделы повздошной кишки и ободочной кишки, но может также встретиться в любой части желудочно-кишечного тракта от рта до ануса и окружающей анус площади. У пациентов с болезнью Крона обычно наблюдается хроническая диарея, связанная с болями в животе, лихорадкой, анорексией, потерей веса и вздутием живота. Язвенный колит также представляет собой хроническое, неспецифическое воспалительное и язвенное заболевание, возникающее в слизистой оболочке толстой кишки, и характеризующееся наличием диареи с кровью.
При воспалительных заболеваниях кишечника обнаруживаются также внекишечные проявления, такие как поражения кожи. Такие поражения характеризуются воспалением и ангиогенезом и могут встречаться во многих местах иных, нежели желудочно-кишечный тракт. Антитела согласно изобретению могут быть способны излечивать эти поражения путем предотвращения ангиогенеза за счет уменьшения притока воспалительных клеток и формирования поражения.
Саркоидоз является другим хроническим воспалительным заболеванием, которое характеризуется как мультисистемное гранулематозное нарушение. Гранулемы при этом заболевании могут образовываться в разных местах тела и, следовательно, симптомы зависят от места гранулемы и от того, активно ли идет заболевание. Гранулемы создаются посредством ангиогенного разрастания сосудов, обеспечивающего постоянный запас воспалительных клеток.
Антитела согласно настоящему изобретению могут также излечивать хронические воспалительные состояния, связанные с псориазом. Псориаз, кожное заболевание, является другим хроническим и рецидивирующим заболеванием, которое характеризуется папулами и бляшками разных размеров. Предотвращение образования новых кровеносных сосудов, необходимых для поддержания характерных поражений, ведет к ослаблению симптомов.
Ревматоидный артрит представляет собой хроническое воспалительное заболевание, характеризующееся неспецифическим воспалением периферических суставов. Предполагается, что кровеносные сосуды в синовиальной выстилке суставов подвергаются ангиогенезу. Кроме того, для образования новой кровеносной сети эндотелиальные клетки выделяют факторы и реакционные разновидности кислорода, что ведет к росту паннуса и разрушению хряща. Факторы, включенные в ангиогенез, могут активно способствовать и помогать поддерживать хроническое воспаленное состояние ревматоидного артрита. Другими заболеваниями, которые могут излечиваться согласно настоящему изобретению, являются гемангиомы, болезнь Рандю-Вебера-Ослера или наследственная геморрагическая телеангиэктазия, твердые или переносимые кровью опухоли и синдром приобретенного иммунодефицита.
Антитела согласно настоящему изобретению могут также использоваться для лечение «нарушения с уменьшенным агниогенезом». Как используется здесь, термин «нарушение с уменьшенным агниогенезом» означает такое нарушение, при котором ангиогенез рассматривался бы как преимущество в лечении заболевания, нарушения и (или) состояния. Это нарушение характеризуется тканью, которая страдает от или рискует пострадать от ишемического поражения, инфекции и (или) плохого заживления, которые случаются, когда ткань лишается адекватного запаса насыщенной кислородом крови вследствие неадекватной циркуляции. Как используется здесь, термин «ткань» понимается в широком смысле, включая в себя - но не ограничиваясь ими - следующие: сердечная ткань, такая как миокард и желудочки сердца; кавернозная ткань; скелетная мышца; неврологическая ткань, такая как из мозжечка; внутренние органы, такие как мозг, сердце, поджелудочная железа, печень, селезенка и легкие; или генерализованный участок тела, такой как целые конечности, нога или дистальные отростки, такие как пальцы рук и стоп.
Способы васкуляризации ишемической ткани
В одном объекте антитела могут быть использованы в способе васкуляризации ишемической ткани. Как используется здесь, термин «ишемическая ткань» означает ткань, которая лишена адекватного кровотока. Примеры ишемической ткани включают в себя - но не ограничиваются ими - ткань, в которой отсутствует адекватный запас крови, что происходит из-за миокардиальных и церебральных инфарктов, мезентериальной ишемии или ишемии конечности, или является результатом окклюзии или стеноза. В одном примере прерывание запаса насыщенной кислородом крови может быть вызвано васкулярной окклюзией. Такая васкулярная окклюзия может быть вызвана атеросклерозом, травмой, хирургическим вмешательством, заболеванием и (или) иметь другую этиологию. Для определения того, подвержена ли ткань риску пострадать от ишемического повреждения от нежелательной васкулярной окклюзии, в распоряжении имеются стандартные рутинные методики. Например, при сердечном заболевании эти методы включают в себя множество методик визуализации (к примеру, технологии радиомечения, рентгенография и ЯМР) и физиологические тесты. Таким образом, индукция ангиогенеза является эффективным средством предотвращения и ослабления ишемии в тканях, на которые осуществляется воздействие, или при риске возникновения васкулярной окклюзии. Кроме того, полностью подразумевается излечение скелетно-мышечной и миокардиальной ишемии, инсульта, заболевания коронарной артерии, заболевания периферических сосудов.
Специалист в области использования стандартных методик может измерить васкуляризацию ткани. Не ограничивающие примеры включают в себя измерение васкуляризации у субъекта с использованием SPECT (однофотонная эмиссионная компьютерная томография); PET (позитронная эмиссионная томография); MRI (ЯМР-томография) и их комбинацию, посредством измерения кровотока к ткани до и после лечения. Для оценки макроскопической васкулярности может быть использована ангиография. Для количественного измерения васкулярности при небольшом сосудистом уровне можно использовать гистологическую оценку. Эти и другие методики обсуждаются у Simons, et al., "Clinical trails in coronary angiogenesis", Circulation, 102, 73-86 (2000).
Способы восстановления ткани
В одном объекте антитела могут быть использованы в способе восстановления ткани. Как используется здесь, термин «восстановление ткани» означает способствование восстановлению ткани, регенерацию, рост и (или) поддержание состояния, включая в себя - но не ограничиваясь ими - «починку» ткани или тканевую инженерию. Специалист в данной области техники поймет, что для восстановления ткани требуется образование новых кровеносных сосудов. В свою очередь, ткань может быть повреждена путем, включающим в себя - но не ограничиваясь ими - травматическое повреждение или условия, включающие в себя артрит, остеопороз и другие нарушения скелета, а также ожоги. Ткань может быть также повреждена вследствие хирургических процедур, иррадиации, разрыва, токсических химикатов, вирусной инфекции или бактериальной инфекции, или ожогов. Ткань, нуждающаяся в восстановлении, может также включать в себя незаживающие раны. Примеры незаживающих ран включают в себя незаживающие кожные язвы, происходящие от диабетической патологии или переломов, которые не заживают быстро.
Антитела также могут быть использованы при восстановлении тканей в контексте процедур, сопровождающих тканевую регенерацию (GTR). Такие процедуры в настоящее время используются специалистами в данной области для ускорения заживления ран после инвазивных хирургических процедур.
Антитела могут быть использованы в способе ускорения восстановления ткани, характеризующемся ускоренным ростом ткани во время процесса тканевой инженерии. Как используется здесь, термин «тканевая инженерия» определяется как создание, конструирование и приготовление биологических простетических устройств в комбинации с синтетическими или природными материалами для усиления или замещения тканей тела и органов. Таким образом, способы согласно изобретению могут быть использованы для улучшения конструирования и роста тканей человека вне тела для последующей имплантации при восстановлении или замещении болезненных тканей. Например, антитела могут быть полезными в ускорении роста замещающих кожных трансплантатов, которые используются в качестве терапии при лечении ожогов.
В другом объекте тканевой инженерии антитела согласно настоящему изобретению могут быть включены в содержащие клетки или не содержащие клетки среды, которые индуцируют регенерацию функциональных тканей человека, когда они имплантированы в сайт, который требует регенерации. Как обсуждалось выше, сопровождаемая биоматериалом тканевая регенерацию может быть использована для способствования повторному росту кости, например, при периодонтальном заболевании. Таким образом, антитела могут быть использованы для ускорения роста переконструированных тканей, агрегированных в пространственные конфигурации в сайте поранения или в другой ткани, которая требует восстановления.
В другом объекте тканевой инженерии антитела могут быть включены во внешние или внутренние среды, содержащие ткани человека, сконструированные для замещения функции заболевшей внутренней ткани. Этот подход включает в себя выделение клеток из тела, помещение их на структурные матрицы и имплантацию новой системы в тело или используя систему вне тела. Например, антитела могут быть включены в васкулярный трансплантат клеточной линии, чтобы способствовать росту клеток, содержащихся в трансплантате. Можно предвидеть, что способы согласно изобретению могут быть использованы для подтверждения восстановления тканей, регенерации и конструирования в продуктах, таких как хрящ и кость, ткани центральной нервной системы, мышц, печени и островковых (инсулин-зависимых) клеток поджелудочной железы.
Фармацевтические составы и способы их применения
Антитела согласно изобретению могут вводиться пациентам для лечения или предотвращения заболеваний или нарушений, которые регулируются генами и белками согласно изобретению. Термин «лечение» используется здесь для обозначения того, что введение соединения согласно настоящему изобретению ослабляет заболевание или нарушение у хозяина. Таким образом, термин «лечение» включает в себя предотвращение возникновения нарушения в хозяине, в частности, когда хозяин предрасположен к заболеванию, но ему еще не поставлен диагноз болезни; ингибирование нарушения; и (или) ослабление или обращение нарушения. Поскольку способы согласно настоящему изобретению направлены на предотвращение нарушений, подразумевается, что термин «предотвращает» не требует, чтобы болезненное состояние было полностью прекращено (см. Webster's Ninth Collegegiate Dictionary). Тем не менее, как используется здесь, термин «предотвращение» относится к способности специалиста в данной области техники идентифицировать популяцию, которая подозревается в нарушении, так что введение соединений согласно настоящему изобретению может быть осуществлено до наступления болезни. Данный термин не подразумевает, что болезненного состояния можно полностью избежать. Соединения, идентифицированные посредством методов скрининга согласно настоящему изобретению, могут вводиться в соединении с другими соединениями.
Безопасность и терапевтическая эффективность идентифицированных соединений могут быть определены стандартными процедурами, используя in vitro- или in vivo-технологии. Соединения, которые обнаруживают совпадающие показатели, могут быть предпочтительными, хотя соединения с меньшими терапевтическими индексами могут быть полезными, если уровень побочных эффектов является приемлемым. Данные, полученные от применения in vitro- и in vivo-методик, могут быть использованы для формулирования интервала доз.
Эффективность соединения может также быть оценена либо на животных моделях, либо на клинических испытаниях пациентов с нерегулируемым или неправильно регулируемым ангиогенезом.
Как используется здесь, подразумевается, что термин «фармацевтически приемлемый носитель» включает в себя все растворители, дисперсионные среды, покрытия, антибактериальные и антигрибковые агенты, изотонические или абсорбционные удерживающие агенты и тому подобные, совместимые с фармацевтическим назначением. Использование таких сред и агентов для фармацевтически активных веществ известно в данной области техники. За исключением любых традиционных сред или агентов, которые являются несовместимыми с активным соединением, такие среды могут быть использованы в композициях согласно изобретению. Дополнительные активные соединения также могут быть включены в композиции. Фармацевтическая композиция согласно изобретению составляется так, чтобы быть совместимой с подразумеваемым способом введения. Примеры способов введения включают в себя парентеральное, к примеру, внутривенное, внутрикожное, подкожное, оральное (к примеру, ингаляция), чрескожное (местное), чресслизистое и ректальное введение. Растворы или суспензии, используемые для парентерального, внутрикожного или подкожного применения, могут включать в себя следующие компоненты: стерильный разбавитель, такой как вода для инъекции, солевой раствор, определенные масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензилалкоголь или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатирующие агенты, такие как этилендиаминтетраускусная кислота; буферы, такие как ацетаты, цитраты или фосфаты и агенты для регулирования тонуса, такие как хлорид натрия или декстроза. pH может регулироваться кислотами или основаниями, такими как соляная кислота или гидроксид натрия. Парентеральный препарат может быть заключен в ампулах, размещен в шприцах или многочисленных флаконах, изготовленных из стекла или пластика.
Фармацевтические композиции, подходящие для инъецирования, включают в себя стерильные водные растворы (когда являются водорастворимыми) или дисперсии и стерильные порошки для немедленного приготовления стерильных растворов для инъекций или дисперсии. Для внутривенного введения подходящие носители включают в себя физиологический солевой раствор, бактериостатическую воду, CREMOPHOR EL™ (BASF, Parsippany, Нью Джерси) или солевой фосфатный буфер (PBS). Во всех случаях композиция должна быть стерильной и жидкой до той степени, чтобы легко происходило впрыскивание. Она должна быть стабильной в условиях производства и хранения и должна предохраняться от загрязнения микроорганизмами, такими как бактерии и грибки. Носитель может быть растворяющей или дисперсионной средой, содержащей, например, воду, этанол, полиол (например, глицерол, пропиленгликоль, жидкий полиэтиленгликоль и тому подобное), и подходящей их смесью. Соответствующая текучесть может поддерживаться, например, использованием оболочки, такой как лецитин, поддержанием требуемого размера части в случае дисперсии и использованием поверхностно-активных веществ. Предотвращения действия микроорганизмов можно достичь различными антибактериальными и антигрибковыми агентами, например, парабенами, хлорбутанолом, фенолом, аскорбиновой кислотой, тимерозалом и т.п. Во многих случаях в композицию предпочтительно было бы включить изотонические агенты, например, сахара, полиспирты, такие как маннитол, сорбитол и хлорид натрия. Пролонгированное поглощение инъецируемых композиций могло бы быть обеспечено включением в композицию агента, который задерживает поглощение, например, моностеарата алюминия и желатина.
Стерильные инъецируемые растворы могут быть приготовлены введением активного соединения в требуемом количестве в подходящий растворитель с одним или комбинацией ингредиентов, перечисленных выше, как это необходимо, с последующей фильтрующей стерилизацией. Обычно дисперсии приготовляют введением активного соединения в стерильную среду, которая содержит основную дисперсионную среду и другие необходимые ингредиенты. В случае стерильных порошков для приготовления стерильных растворов для инъецирования предпочтительными методами приготовления являются вакуумная сушка и лиофилизация, которые дают порошок активного ингредиента плюс какой-либо дополнительный желательный ингредиент из его предварительно стерильно отфильтрованного раствора.
Системное введение может также осуществляться трансслизистыми или трансдермальными средствами. Что касается трансслизистого и трансдермального введения, в рецептуре используются пенетранты, способные проникать через перегородки. Такие пенетранты в общем известны в данной области техники и включают в себя, например, для трансслизистого введения, детергенты, желчные соли и производные фусидовой кислоты. Трансслизистое введение может сопровождаться использованием назальных спреев или суппозиториев. Для трансдермального введения активные соединения вводят в помады, мази, гели или кремы, как это обычно известно в данной области.
Соединения могут быть также приготовлены в форме суппозиториев (к примеру, с традиционной суппозиторной основой, такой как масло какао и другими глицеридами) или удерживаемых клизм для ректального введения.
В одном варианте осуществления активные соединения приготовляют с носителями, которые предохраняют соединение от быстрого выведения из тела, такими как составы с контролируемым высвобождением, включая имланты и системы микроинкапсулированной доставки. Могут быть использованы биодеградируемые, биосовместимые полимеры, такие как этиленвиниловый ацетат, полиангидриды, полигликолевые кислоты, коллаген, полиортоэфиры и полимолочная кислота. Методы приготовления таких составов очевидны для специалистов в данной области. Материалы могут быть также получены коммерчески от Alza Corporation и Nova Pharmaceutical, Inc. В качестве фармацевтически приемлемых носителей могут быть также использованы липосомные суспензии (включающие в себя липосомы, нацеленные на инфицированные клетки, с моноклональными антителами к вирусным антигенам). Они могут быть приготовлены в соответствии с методами, известными специалистам в данной области, например, как описано в патенте США №4.522.811.
Особенно выгодно составлять оральные или парентеральные композиции в форме дозировочных единиц для легкого введения и единообразия дозировки. Термин «форма дозировочной единицы», как используется здесь, относится к физически дискретным единицам, пригодным в качестве одноразовых доз для пациента, которого лечат, при этом каждая единица содержит заранее заданное вычисленное количество активного соединения, чтобы произвести желательный терапевтический эффект в соединении с требуемым фармацевтическим носителем. Спецификация форм дозировочных единиц диктуется и прямо зависит от уникальных характеристик активного соединения и конкретного достигаемого терапевтического эффекта, а также ограничений, свойственных для сферы применения химического соединения, такого как активное соединение для лечения индивидуумов.
ПРИМЕРЫ
Пример 1. Получение внеклеточного домена НРТРβ-белка.
Способы: НРТРβ полной длины клонируют из плацентарной библиотеки человека в соответствии с инструкциями производителя (Origene). Этот клон идентичен ранее сообщенному кДНК-клону (Genbank, номер доступа Х54131), за исключением того, что отсутствует FNIII-повтор №5. кДНК, кодирующую полный растворимый внеклеточный домен (ECD) НРТРβ, клонируют с помощью ПЦР от кДНК полной длины (последовательность см. ниже), кодирующей аминокислоты 1-1534 с присоединенным С-концевым His-His-His-His-His-His-Gly (6His-Gly) (SEQ ID NO:1). Результирующую кДНК клонируют в экспрессионные векторы млекопитающих для транзиентной (pShut-tle-CMV) или стабильной (pcDNA3.1(-)) экспрессии в HEK293-клетках. Для получения очищенного ECD НРТРβ (pECD) HEK293-клетки, трансфецированные экспрессирующими pECD векторами, инкубируют в OptiMEM-бессывороточной среде (Gibco) в течение 24 часов при нормальных условиях роста. Затем восстанавливают стандартизованную среду, центрифугируют для удаления дебриса (1000 об/мин × 5 минут) и добавляют по 1 мл промытой Ni-NTA-агарозы (Qiagen)(500 мкл уплотненного материала) к 10 мл очищенной среды и помещают в качалку на всю ночь при 4°С. На следующий день смесь нагружают на колонку и промывают с помощью 20 объемов наполнителя из 50 мМ NaH2PO4, 300 мМ NaCl, 20 мМ имидазола, pH 8. Очищенный внеклеточный домен НРТРβ-белка (SEQ ID NO:2) затем элюируют в шести фракциях с помощью 200 мкл/элюцию в 50 мМ NaH2PO4, 300 мМ NaCl, 250 мМ имидазола, pH 8. Фракции анализируют на содержание белка, используя восстановительно-денатурирующий SDS-полиакриламидный гель-электрофорез, детектируют с помощью серебряного красителя (Invitrogen) и подтверждают масс-спектрометрией.
Результаты: для получения антитела к внеклеточному домену НРТРβ конструируют векторы, направляющие экспрессию тэгированного 6-His внеклеточного домена НРТРβ-белка (Фиг.1, панель А). Далее тэгированный 6-His внеклеточный домен НРТРβ-белка очищают почти до гомогенности (Фиг.1, панель В) из стандартизованной среды HEK293-клеток, трансфецированных экспрессирующим вектором.
Пример 2. Создание моноклональных антител к внеклеточному домену НРТРβ.
Способы: Для получения иммуногена внеклеточного домена НРТР? очищенный внеклеточный домен с 6His НРТРβ-белка конъюгируют со свиным тироглобулином (Sigma), используя химию EDC-объединения (Hockfield, S. et al., (1993) Cold Spring Habor Laboratory Press. Volume 1 pp 111-201, Immunocytochemistry). Полученный НРТРβ-внеклеточный домен-тироглобулиновый конъюгат диализируют против PBS, pH 7,4. Затем взрослых Balb/c-мышей иммунизируют подкожно этим конъюгатом (100-200 мкг) и полным адъювантом Фройнда в смеси 1:1. Спустя 2-3 недели мышей инъецируют внутрибрюшинно или подкожно неполным адъювантом Фройнда и конъюгатом в смеси 1:1. Инъекцию повторяют на 4-6 недели. Сыворотки собирают от мышей спустя 7 дней после третьей инъекции и анализируют на иммунореактивность к антигену внеклеточного домена НРТРβ посредством ELISA и Western-блоттинга. Мышей, которые показывают хороший ответ на антиген, укрепляют с помощью единичной внутриселезеночной инъекции 50 мкл очищенного внеклеточного домена НРТРβ-белка, смешанного 1:1 с гидроксидом алюминия, используя сверхдлинную иглу 31 размера (Goding, J.W., (1996) Monoclonal Antibodies: Principles and Practices. Third Edition, Academic Press Limited. P. 145). Вкратце, мышей анестезируют 2,5%-ным авертином и делают разрез в 1 сантиметр на коже, оставляя на теле косую стенку. Антигенную смесь вводят посредством введения иглы от задней части к передней части селезенки в продольном направлении. На клеточную стенку накладывают швы и кожу закрывают двумя металлическими клипсами. Наблюдают надежность выздоровления мышей. Спустя четыре дня после хирургического вмешательства селезенку мыши удаляют и приготовляют суспензию отдельных клеток для слияния с миеломными клетками мыши для получения гибридомных клеточных линий (Spitz, M., (1986) Methods In Ensymology, Volume 121. Eds. John J., Lagone and Helen Van Vunakis. PP. 33-41 (Academic Press, New York, Нью-Йорк)). Полученные гибридомы культивируют на модифицированной среде Dulbeccos (Gibco), дополненной 15% фетальной телячьей сывороткой (Hyclone) и гипоксантином, аминоптерином и тимидином.
Скрининг на позитивные гибридомы начинают через 8 дней после слияния и продолжают в течение 15 дней. Гибридомы, продуцирующие антитела к НРТРβ-внеклеточному домену, идентифицируют с помощью ELISA на двух наборах 96-луночных планшетов: один покрывают внеклеточным доменом тэгированного гистидином НРТРβ, а другой покрывают белком тэгированного гистидином бактериального MurA в качестве негативного контроля. Вторичное антитело является ослиным антимышиным IgG, меченым пероксидазой хрена (HRP) (Jackson Immunoresearch). Иммунореактивность в лунках наблюдают, используя развитие цвета, инициированного ABTS-таблетками, растворенными в TBS-буфере, рН 7,5. Индивидуальные HRP-реакционные смеси завершают добавлением 100 микролитров 1% SDS и считывают поглощение при 405 нм с помощью спектрофотометра. Гибридомы, продуцирующие антитела, которые взаимодействуют с внеклеточным доменом-6His НРТРβ, но не с murA-6His-белком, используют для дальнейшего анализа. Ограниченные разведения (0,8 клеток на лунку) приготовляют дважды на позитивных клонах в 96-луночных планшетах, с клональностью, определенной как имеющую более 99% лунок с позитивной реактивностью. Изотипы антител определяют с помощью технологии изо-полос (Roche). Чтобы получить очищенное антитело для последующей оценки, аффинно очищают супернатанты тканевых культур, используя белок А- или белок G-колонки.
Результаты: Выделено пять моноклональных антител к внеклеточному домену белка НРТРβ, которым даны следующие номенклатурные наименования - R15E6, R12A7, R3A2, R11C3, R15C2 и R5A8.
Моноклональное антитело R15E6 депонировано в Американской Коллекции Типовых Культур (АТСС), P.O. Box 1549, Manassas, VA 20108 USA 4 мая 2006.
Пример 3. R15E6 связывается с эндогенным НРТРβ на эндотелиальных клетках человека.
А. R15E6 связывает эндогенный НРТРβ, как продемонстрировано иммунопреципитацией и western-блотом.
Материалы: эндотелиальные клетки пупочной вены (HUVECs), EGM-среда и нейтрализующий трипсин раствор от Cambrex; OPTIMEM I (Gibco), бычий сывороточный альбумин (BSA; Santa Cruz), фосфатно-буферный раствор (PBS; Gibco), фактор роста, включающий в себя Ангиопоэтин 1 (Ang I), фактор роста эндотелиальных сосудов (VEGF) и фактор роста фибробластов (FGF) (R&D Systems), моноклональное тело Tie2 (Duke University/P&GP), VEGF-рецептор 2 (VEGFR2) поликлональное антитело (Whitaker et al.), белок-A/G агароза (Santa Cruz), Tris-Глицин заводского изготовления гель-электрофорез/передающая система (6-8%) (Invitrogen), PVDF-мембраны (Invitrogen), лизисный буфер (20 мм Tris-HCl, 137 мм NaCl, 10% глицерол, 1% тритон-Х-100, 2 мМ EDTA, 1 мМ NaOV, 1 мМ NaF, 1 мМ PMSF, 1 мкг/мл лейпептина, 1 мкг/мл пепстатина).
Способ: HUVECs предварительно обрабатывают в течение 30 мин антителом (в OPTIMEM) или только одним OPTIMEM I. После удаления материала предварительной обработки клетки обрабатывают Ang 1 (100 нг/мл) в течение 6 минут в PBS+0,2% BSA и лизируют в лизисном буфере. Лизаты направляют прямо на Tris-Глицин-гель и иммуноосаждают с помощью 2-5 мкг/мл антитела Tie-2 или 10 мкг/мл антитела R15E6 и белок-A/G агарозы. Иммуноосажденные образцы промывают однократно лизисным буфером и кипятят в течение 5 мин в однократном буфере образца. Образцы разрешают на Tris-Глицин-геле, передают на PVDF-мембрану и детектируют western-блотом, используя указанные антитела (pTYR Ab (PY99, Santa Cruz), Tie-2, VEGFR2 и (или) R15E6).
Результаты: При использовании IP/western-блоттинга R15E6 распознает главную, с высоким молекулярным весом полосу, согласующуюся с размером НРТРβ (Фиг.2, панель А, дорожка 2). Менее интенсивные, с меньшим молекулярным весом полосы, вероятно, соответствуют менее гликозилированным формам предшественника НРТР(3. Контрольное иммуноосаждение (IP) не-иммунным IgG не показывает никаких полос в диапазоне молекулярного веса НРТРβ (Фиг.2, панель А, дорожка 1), а комбинированное Tie2/VEGFR2 IP показывает полосы ожидаемого молекулярного веса (Фиг.2, панель А, дорожка 3). Этот результат демонстрирует, что R15E6 распознает и является специфичным для НРТРβ.
В. R15E6 связывает эндогенную НРТРβ, как показывает FACS-анализ.
Материалы: HUVECs, EGM-среда и трипсин-нейтрализущий раствор от Cambrex; Вторичный Alexfluor 488-тэгированное антитело от Molecular Probes; сбалансированный солевой раствор Hanks (Gibco); проточный цитометр FACSCAN и программа CellQuest от Breton Dickenson.
Способ: HUVECs трипсинизируют, обрабатывают трипсин-нейтрализующим раствором, и промывают HBSS. R15E6-антитело (0,6 мкг) добавляют к 250000 клеток в 50 мкл HBSS и инкубируют на льду в течение 20 минут. Клетки промывают 1 мл HBSS, затем добавляют 2 мкг конъюгированного с флуоресцентом вторичного антитела в течение 20 минут на льду. Клетки промывают 1 мл HBSS и ресуспендируют в 1 мл HBSS, затем анализируют на проточном цитометре FACSCAN программой CellQuest. Контрольные клетки обрабатывают только конъюгированным с флуоресцентом вторичным антителом.
Результаты: При использовании FACS-анализа, с интактными HUVECs, R15E6 вызывает сильный сдвиг (>90% клеток) в флуоресцентном сигнале по сравнению с сигналом от одного вторичного антитела (Фиг.2, панель В). Этот результат указывает, что R15E6 связывается с эндогенной НРТРβ, присутствующей на поверхности интактных эндотелиальных клеток.
Пример 4. R15E6 усиливает Tie2-активацию и стимулирует многочисленные ангиогенные ответы (выживание эндотелиальных клеток, миграция и капиллярный морфогенез).
А. R15E6 увеличивает Tie2-фосфорилирование в отсутствие и в присутствии ангиобелка 1 (Ang1), где Tie2 является лигандом.
Способы: HUVECs культивируют в бессывороточной среде, как описано выше, в присутствии или в отсутствие разных концентраций R15E6 и с добавлением или без добавления Ang1. Приготовляют лизаты, проводят иммуносажадение с помощью Tie2-антитела, разрешают посредством электрофореза на полиакриламидном геле и передают на PVDF-мембрану. Связавшиеся с мембраной иммуноосажденные белки затем подвергают сериальному western-блоттированию с помощью антифосфотирозинового антитела, чтобы оценить количественно Tie2-фосфорилирование, после чего с помощью Tie2-антитела квалифицируют общее Tie2. Tie2-фосфорилирование выражают как отношение антифосфотирозинового сигнала к общему Tie2-стгналу.
Результаты: R15E6 увеличивает Tie2-фосфорилирование как в отсутствие, так и в присутствии Ang1 (Фиг.3). Этот результат указывает, что связывание R15E6 с НРТРβ на поверхности эндотелиальных клеток модулирует его биологическую функцию, приводящую к повышенной активации Tie2 в отсутствие или в присутствии лиганда.
В. R15E6 увеличивает выживание эндотелиальных клеток в отсутствие или в присутствии эндотелиальных факторов роста.
Материалы: HUVECs, EGM-среда и трипсин-нейтрализущий раствор от Cambrex; DMEM (Cell Gro), обезжиренный BSA (BD Falcon), набор для титрования клеток Glo ATP (Primega), факторы роста (Ang1, VEGF 165 и FGF) (R&D Systems), многометочный планшетный считыватель Victor V (Perkin Elmer Wallac).
Способы: HUVECs помещают на планшеты в количестве 10 ООО клеток на лунку, сыворотка, требующаяся для среды DMEM/0,2% BSA и обработанная в течение 72 ч в присутствии или в отсутствие фактора роста (Ang1, VEGF или FGF) с или без разных концентраций R15E6-антитела. Спустя 72 часа клетки промывают DMEM и выжившие клетки квалифицируют, измеряя уровни ATP, используя набор для титрования клеток Glo Luminescence в соответствии с инструкциями производителя (Promega).
Результаты: Согласуясь с результатами анализа Tie2-активации, R15E6 увеличивает выживаемость эндотелиальных клеток в отсутствие добавленного фактора роста при концентрациях между 0,5 и 5 нМ (Фиг.4, панель А). Аналогично, R15E6 увеличивает опосредованную Ang1 выживаемость эндотелиальных клеток (Фиг.4, панель А), а также выживаемость клеток, опосредованную VEGF и FGF (Фиг.4, панели В и С). С контрольным моноклональным антителом не наблюдается никакой выживаемости (Фиг.4, панель D). Эти результаты демонстрируют, что связывание R15E6 с НРТРβ на поверхности эндотелиальных клеток увеличивает базисную выживаемость клеток, а также выживаемость клеток, опосредованную многочисленными путями ангиогенеза (Ang1, VEGF или FGF).
С. R15E6 увеличивает миграцию эндотелиальных клеток в отсутствие и в присутствии VEGF.
Материалы: HUVECs, EGM-среда и трипсин-нейтрализущий раствор от Cambrex; ЕВМ без фенола красного (PRF-EMB, Cambrex), обезжиренный BSA (BD Falcon). Система BD Falcon Biocoat миграции эндотелиальных клеток (BD Falcon), Calcein AM (Molecular Probes); факторы роста (VEGF 165) (R&D Systems), многометочный планшетный считыватель Victor V (Perkin Elmer Wallac).
Способы: HUVECs ресуспендируют в PRF-EBM+0,1% BSA и помещают на планшет в количестве 50000 клеток на лунку (BD Bioscience, размер поры 3 мкм). Факторы роста помещают на дно лунки транслуночной камеры и инкубируют 4-22 ч. Клетки, мигрирующие через мембрану, детектируют мечением 4 мкг/мл Calcein AM в течение 90 с.Флюоресценцию измеряют, используя оборудование Victor A (485/535).
Результаты: В согласии с результатами исследования выживаемости, R15E6 увеличивает как базовую, так и VEGF-опосредованную миграцию эндотелиальных клеток (Фиг.5).
D. R15E6 увеличивает разрастание эндотелиальных клеток и капиллярный морфогенез в отсутствие и в присутствии эндотелиальных факторов роста.
Материалы: HUVECs и EGM-среда от Cambrex; бусины Cytodex и коллаген типа I от Sigma; Dulbecco's PBS и М199-среда от Gibco; VEGF от R&D.
Способы: HUVECs пассажа 4 (2×106 клеток) культивируют с 5 мг бусин Cytodex в 10 мл EGM в 100 мм обработанных бактериологических чашках с не-тканевой культурой в течение 48 часов, вращаемых время от времени. Покрытые клетками бусины переносят в 50 мл конические пробирки и ресуспендируют в 380 мкл D-PBS. Коллагеновые гели приготовляют добавлением 71,4 мкл покрытых клетками бусин к 2,8 мл матриксного раствора, состоящего из 3 мг/мл коллагена в М199-среде, дополненной 0,005 NaOH, 20 мМ HEPES и 26 мМ NaHCO3. Триста пятьдесят микролитров бусин распределяют в лунку на 24-луночный планшет тканевой культуры и дают возможность матриксу затвердеть в течение 1 часа при 37°С/5% CO2. Один мл EGM-среды с или без VEGF (10 нг/мл) или R15E6 (7,5 мкг/мл) добавляют на лунку и возвращают в инкубатор. Спустя 48 часов зашторенный наблюдатель визуализирует отростки с помощью микроскопа с инвертированной контрастной фазой и просматривает 50 бусинок на лунку, в трех вариантах лунок, на присутствие отростков эндотелиальных клеток. Результаты выражают как количество отростков на лунку.
Результаты: В согласии с результатами других анализов, R15E6 увеличивает также базисный и VEGF-опосредованный капиллярный морфогенез в анализе эндотелиального разрастания на бусинах (Фиг.6).
Пример 5. Связывающим эпитопом для R15E6 является N-концевой FN3-повтор внеклеточного домена человеческого НРТРβ.
А. Western-блот анализ рекомбинантных мутантов с c-концевым усечением и химерных мышь/человек белков показывает, что R15E6-связывающим эпитопом является N-концевой FN3-повтор внеклеточного домена человеческого НРТРβ.
Способы: HEK293-клетки трансфецируют экспрессионными векторами, кодирующими указанный усеченный мутант НРТРβ или химеру мышь/человек. Затем трансфецированные клетки инкубируют в среде OptiMEM в течение дополнительных 24 часов, после чего стандартизованную среду, содержащую указанный внеклеточный домен НРТРβ, собирают и либо хранят для будущего использования, либо используют немедленно для western-блот- или ECL (см. ниже) исследований. Для western-блот-анализа 20 мкл стандартизованной среды, содержащей указанный НРТРβ-белок или не рекомбинантный белок (Mock, пустой трансфецированный вектор), разрешают с помощью PAGE, переносят на PVDF-мембрану и зондируют с помощью R15E6.
Результаты: В соответствии с western-блот-анализом R15E6 связывает все НРТРβ-мутанты с C-концевыми делециями (Фиг.7), указывая, что связывающий эпитоп находится в пределах первого из двух N-концевых FN3-повторов. R15E6 не связывает внеклеточный домен мышиной НРТРβ (SEQ ID NO:7), демонстрируя специфичность к человеческому белку (Фиг.7В, дорожка 6 против дорожки 2). Замещение первого или первого и второго N-концевых FN3-повторов человеческими последовательностями восстанавливала R15E6-связывание (Фиг.7В, дорожки 3 и 5). Наоборот, замещение только 2-го мышиного FN3-повтора человеческой последовательностью не восстанавливает связывание (Фиг.7В, дорожка 4). Совместное рассмотрение показывает локализацию связывающего эпитопа R15E6 на N-концевом FN3-повторе (~100 аминокислот) человеческого НРТРβ.
В. ECL (электрохемилюминесцетный) анализ мутантов с концевым усечением и химерных белков мышь/человек подтверждает, что R15E6-связывающий эпитоп находится на N-концевом FN3-повторе внеклеточного домена НРТРβ.
Способы: Супернатанты, содержащие указанный HPTPβ-белок наносят на 96-луночный высокосвязывающий планшет MCD (Meso Scaale Discovery), дают возможность высохнуть и блокируют 3% BSA в течение 1 ч. Затем белок инкубируют с R15E6-моноклональным антителом или R15E6 Fab-фрагментом (10 нМ или 1,5 мкг/мл) в течение 1 ч, промывают и инкубируют с козьим анти-мышиным антителом, меченым MSD-Tag ((10 нМ) в течение 1 ч. Избыток антитела смывают и добавляют читаемый MSD-буфер. Эмиссию света измеряют, используя считыватель Sector 2400 (MSD). MSD использует электрохемилюминесцентную детекцию для обнаружения событий связывания в структурированных массивах. Технология Meso Scaale Discovery использует патентованные микроплаты MULTI-ARRAY™ и MULTI-SPOT™ с электродами, интегрированными в дно планшета. MSD-электроды выполняют из углерода, и биологические реагенты могут присоединяться к углероду просто за счет пассивной адсорбции и сохранять высокий уровень биологической активности. В MSD-анализах используются электрохемилюминесцентные метки для сверхчувствительной детекции. Эти электрохемилюминесцентные метки излучают свет, когда возбуждаются электрохимически. Процесс детектирования инициируют на встроенных электродах, расположенных на дне MSD-микроплат, и как только метки вблизи электрода возбуждаются, измеряется излучение на 620 нм.
Результаты: В согласии с western-блот-исследованиями, R15E6 связывается со всеми НРТРβ-белками с С-концевыми усечениями в MSD-анализе (Фиг.8А). Также в согласии с western-блот-анализом R15E6 не связывает внеклеточный домен мышиного НРТРβ, но связывание восстанавливается при замещении мышиного N-концевого FN3-повтора человеческим N-концевым FN3-доменом (Фиг.8В). Эти данные подтверждают, что связывающий эпитоп R15E6 находится на N-концевом FN3-повторе человеческого НРТРβ. Как ожидается, связывающий эпитоп моновалентного Fab-фрагмента R15E6 также мог бы быть картирован на N-конце большей части FN3-повтора человеческого НРТРβ (Фиг.9).
Пример 6. Моновалентный Fab-фрагмент R15E6 блокирует R15E6-опосредованную Tie2-активацию и ингибирует выживание и миграцию эндотелиальных клеток.
Способы: Анализы Tie2-активаци и выживания и миграции эндотелиальных клеток проводили, как описано выше в примере 4. Моновалентные Fab-фрагменты R15E6 приготовляли, как описано выше. Очищенный R15E6 подвергали диализу в 0,1 М Tris-HCl, pH 8,0, содержащем 2 мМ EDTA и 1 мМ дитиотреитола. Папаин (Pierce) в концентрации 1-2 мг/мл активируют в вышеупомянутом буфере в течение 15 минут при 37°С. R15E6 в концентрации 10 мг/мл инкубируют с папаином в том же самом буфере, используя соотношение фермент:субстрат 1:100 в течение 1 ч при 37°С. Расщепление заканчивают добавлением йодоацетамида (финальная концентрация 20 мМ) и держат на льду в течение 1 ч, предохраняя от света. Расщепленный папаиновый материал диализируют в течение ночи в забуференном фосфатом солевом буфере, чтобы удалить йодоацетамид. Степень расщепления отслеживают с помощью SDS-PAGE по исчезновению тяжелой гамма-цепи (MB 55000 кДа) и появлению Fc-фрагмента гамма (MB 27000 кДа) и легких цепей (MB 22000-25000 кДа).
Результаты: В отличие от интактного R15E6-антитела Fab-фрагменты не показывают увеличения Tie2-активации (Фиг.10). Более того, при наличии избытка Fab-фрагмента R15E6-опосредованная Tie2-активация блокируется (Фиг.10). Удивительно, что R15E6 Fab-фрагмент существенно ингибирует выживание эндотелиальных клеток по сравнению с контрольным Fab (Фиг.11А), и этого эффекта можно было бы избежать посредством добавления интактного R15E6 (Фиг.11В). Согласуясь с негативным эффектом выживания эндотелиальных клеток, R15E6 Fab также блокирует VEGF-опосредованную миграцию эндотелиальных клеток (Фиг.12). Эти открытия демонстрируют, что интактное димерное R15E6 необходимо для усиления ангиогенного сигнализирования и что мономерное R15E6 блокирует эти действия и действительно оказывает негативный эффект на ангиогенные ответы эндотелиальных клеток.
Если иначе не оговорено, подразумевается, что все величины, включая количества, процентные содержания, части и соотношения, могут быть модифицированы словом «примерно», и не имеется в виду, что величины указывают точные цифровые значения.
Если иначе не оговорено, артикли «a», «an» и «the» означают «один или несколько».
Все цитируемые документы в релевантной части включены сюда в качестве ссылки; цитирование любого документа не должно пониматься как допущение того, что он является уровнем техники в отношении настоящего изобретения. В той мере, насколько какое-либо значение или определение термина в данном описании противоречит любому значению или определению термина в документе, включенном в качестве ссылки, преимущество имеет значение или определение, даваемое термину в данном описании.
Хотя здесь проиллюстрированы и описаны конкретные варианты осуществления настоящего изобретения, для специалистов в данной области техники было бы очевидным, что могут быть внесены различные другие изменения и модификации без отхода от сущности и объема изобретения. Таким образом, имеется в виду, что в прилагаемой формуле изобретения охватываются все такие изменения и модификации, которые находятся в объеме данного изобретения.























































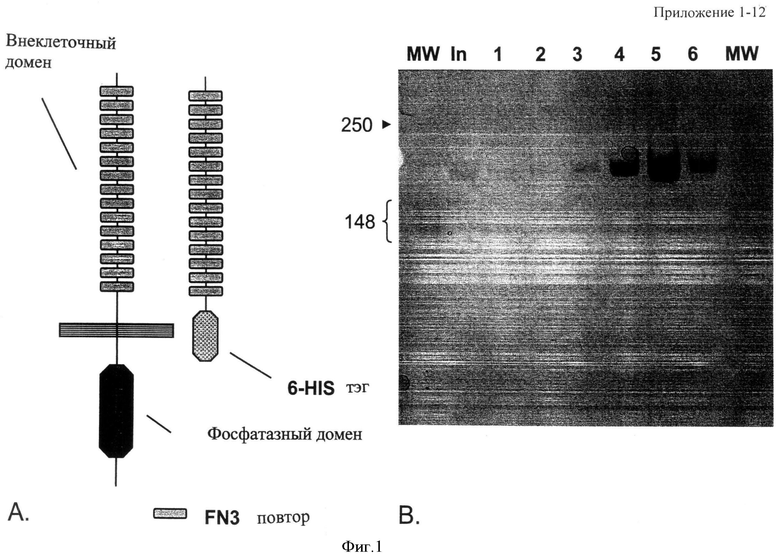

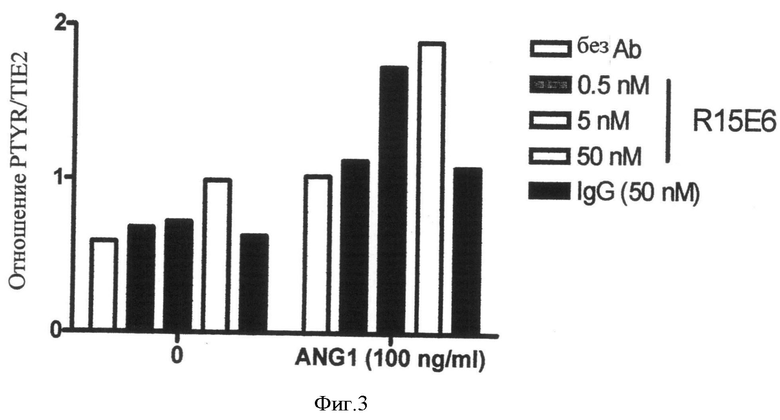
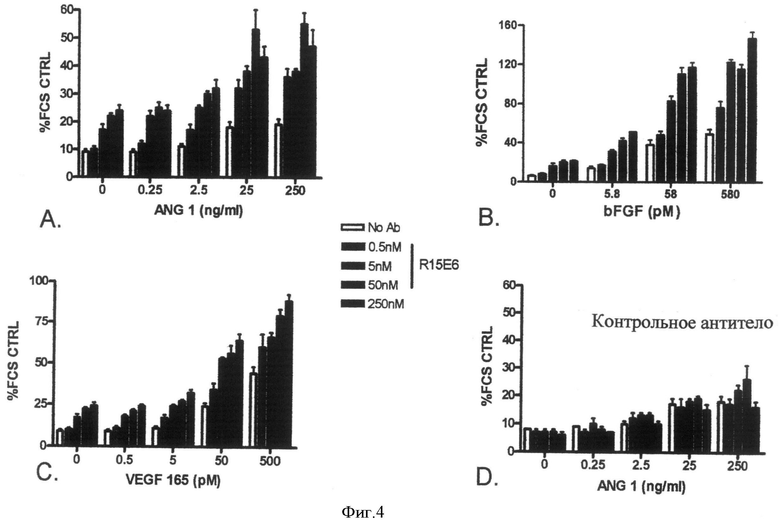

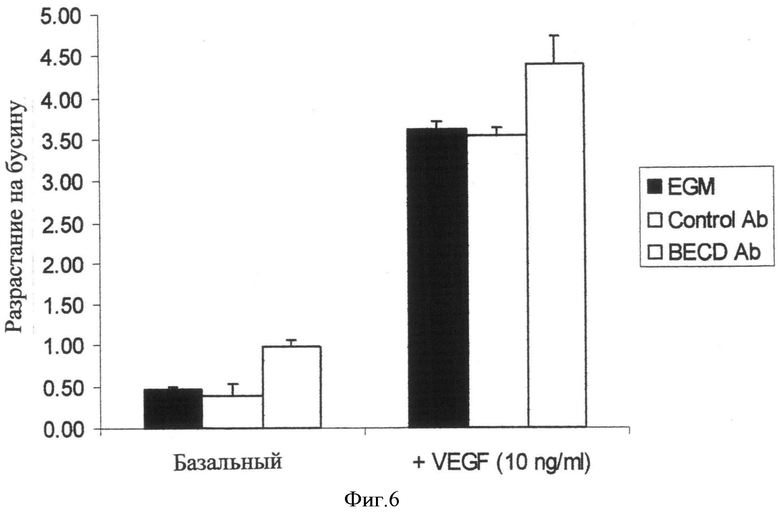

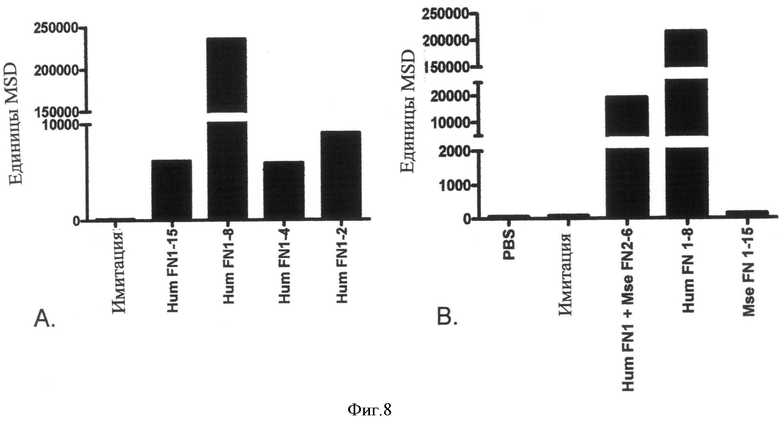



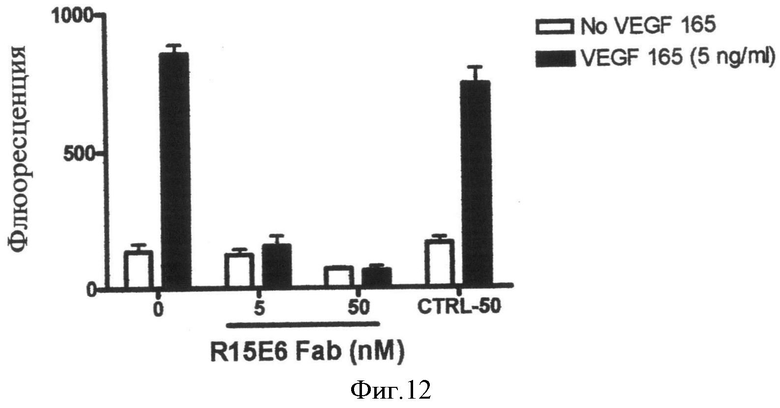
В изобретении раскрыты способы лечения нарушения, регулируемого ангиогенезом, предусматривающие введение субъекту эффективного количества антитела, которое связывается с человеческим белком бета-тирозин фосфатазой (HPTPβ). Связывание антитела с HPTPβ усиливает передачу сигнала Tie-2 и тем самым регулирует (увеличивает или уменьшает) ангиогенез в зависимости от заболевания. Используемое антитело продуцируется гибридомой АТСС №РТА-7580. На основе антитела создана фармацевтическая композиция для лечения нарушения, регулируемого ангиогенезом, которая помимо антитела содержит фармацевтически приемлемый носитель. Способы по изобретению предназначены для лечения нарушений с уменьшенным или увеличенным ангиогенезом, в частности, таких заболеваний, как скелетно-мышечная или миокардиальная ишемия, инсульт, заболевания коронарной артерии и периферических сосудов, серповидно-клеточной анемии, болезнь Педжета, болезнь Лайма и других заболеваний. 3 н. и 4 з.п. ф-лы, 12 ил., 1 табл., 6 пр.
1. Способ лечения нарушения, регулируемого ангиогенезом, у субъекта, содержащий этапы, на которых:
а) идентифицируют субъект, нуждающийся в увеличении ангиогенеза; и
б) вводят этому субъекту эффективное количество изолированного антитела, которое связывается с человеческим белком тирозин фосфатазой бета (HPTPβ), при этом упомянутое антитело произведено гибридомной клеточной линией АТСС № РТА-7580 и при связывании с НРТРβ усиливает передачу сигнала Tie-2 и таким образом увеличивает ангиогенез.
2. Способ по п.1, отличающийся тем, что регулируемое ангиогенезом нарушение представляет собой нарушение с уменьшенным ангиогенезом, выбранное из группы, состоящей из: скелетно-мышечной или миокардиальной ишемии, инсульта, заболевания коронарной артерии и заболевания периферических сосудов.
3. Способ по п.2, отличающийся тем, что нарушение с уменьшенным ангиогенезом является заболеванием периферических сосудов.
4. Способ по п.2, отличающийся тем, что нарушение с уменьшенным ангиогенезом является заболеванием коронарной артерии.
5. Фармацевтическая композиция для лечения нарушения, регулируемого ангиогенезом, содержащая:
a) выделенный антигенсвязывающий фрагмент антитела, который связывается с человеческим белком тирозин фосфатазой бета (HPTPβ), при этом упомянутый фрагмент произведен гибридомной клеточной линией АТСС № РТА-7580 и при связывании с HPTPβ ингибирует передачу сигнала Tie-2 и таким образом уменьшает ангиогенез и
b) фармацевтически приемлемый носитель.
6. Способ лечения нарушения, регулируемого ангиогенезом, при котором лечение обеспечивает уменьшенный ангиогенез, при этом упомянутый способ содержит этап, на котором вводят субъекту фармацевтическую композицию по п.5.
7. Способ по п.6, отличающийся тем, что нарушение выбрано из группы, состоящей из: серповидно-клеточной анемии, саркоида, сифилиса, псевдоксантомы эластикум, болезни Педжета, окклюзии вен, окклюзии артерий, обструктивного заболевания сонной артерии, хронического увеита/витрита, микобактериальной инфекции, болезни Лайма, системной волчанки, ретинопатии недоношенности, болезни Илза, болезни Бехчета, болезни Старгардта, pars planitis, хронического отслоения сетчатки, синдрома гипервязкости, токсоплазмоза, травмы и пост-лазерных осложнений.
| US 20040077065 A1, 22.04.2004, abstr., claims | |||
| US 20040014162 A1, 22.01.2004, abstr., claims | |||
| FOEHR ED et al., "Targeting of the receptor protein tyrosine phosphatase beta with a monocl | |||
| antibody delays tumor growth in a glioblastoma model", Cancer res., 2006, v.66, no.4, p.2271-78 | |||
| РОЙТ и др., «Иммунология», перевод с англ.-М.: Мир, 2000 г., стр.110, 150-158, 537-539. |

