Область техники, к которой относится изобретение
Настоящее изобретение касается антитела, которое специфически связывается с TIE2 (тирозинпротеинкиназным рецептором) и обладает функцией нормализации и стабилизации кровеносных сосудов посредством фосфорилирования рецептора. Более конкретно, настоящее изобретение касается антитела против TIE2, кодирующих его нуклеиновых кислот, векторов, содержащих эти нуклеиновые кислоты, клеток, трансформированных этими векторами, стабилизатора сосудов и содержащих его композиций для лечения связанных с ангиогенезом заболеваний, фармацевтических композиций для лечения опухолей или рака и композиций для комбинированного применения с другими лекарственными средствами, чем антитело, связывающееся с TIE2.
Уровень техники
Ангиогенез - это механизм, посредством которого образуются новые кровеносные сосуды из существующих кровеносных сосудов в результате роста, деления, миграции и т.п.эндотелиальных клеток, который играет ключевую роль в нормальных процессах роста, включая заживление ран и менструальный цикл женщин (Risau, Nature, 386: 671, 1997). Кроме того, известно, что аномально чрезмерный ангиогенез сильно влияет на такие заболевания, как рост опухолей и метастазирование, возрастная дегенерация желтого пятна (ARMD), диабетическая ретинопатия, псориаз, ревматоидный артрит и хроническое воспаление (Carmeliet and Jain, Nature, 407:249, 2000).
Развитие ангиогенеза определяется общим балансом факторов, индуцирующих ангиогенез и ингибирующих ангиогенез, и происходит посредством сложного и последовательного процесса, включающего несколько стадий. Процесс ангиогенеза протекает следующим образом. Сначала различные факторы, индуцирующие ангиогенез, включая фактор роста эндотелия сосудов (VEGF), секретируемые из опухолей или поврежденных тканей, связываются с соответствующими рецепторами у существующих эндотелиальных клеток сосудов, расположенных рядом с ними, активируя эндотелиальные клетки сосудов и тем самым улучшая проницаемость эндотелиальных клеток сосудов, которые секретируют протеолитические ферменты типа матриксной металлопротеиназы (MMP) для разрушения матрикса и внеклеточного матрикса вокруг эндотелиальных клеток сосудов, за счет этого обеспечивая пролиферацию эндотелиальных клеток сосудов и их миграцию из существующих капилляров к тканям, секретирующим факторы, индуцирующие ангиогенез.
Мигрировавшие и пролиферировавшие эндотелиальные клетки сосудов образуют внутрисосудистые структуры, а затем появляются сосудистые перициты, которые являются структурной опорой эндотелиальных клеток сосудов, что приводит к стабильному и зрелому ангиогенезу. В это же время ангиопоэтин 1 (ANG1), секретируемый из эндотелиальных клеток сосудов, связывается со своим рецептором, TIE-2, и тем самым играет ключевую роль в появлении сосудистых перицитов и стабилизации кровеносных сосудов (Suri et al., Cell, 87:1171, 1996).
Между тем, ангиопоэтин 2 (ANG2) гиперэкспрессируется в раковых тканях, в которых активно протекает ангиогенез, или же в плаценте, матке и яичниках, то есть в нормальных тканях, в которых происходит активное ремоделирование кровеносных сосудов (Kong et al., Cancer Res., 61:6248, 2001; Ahmad et al., Cancer, 92:1138, 2001), а когда ANG2 присутствует в количестве, превышающем ANG1, то в результате запускается опухолевый ангиогенез (Linda et al., Cell Research, 13(5), 309-317, 2003). Все это интерпретируется так, что ANG2 действует в качестве антагониста в сигнальном механизме TIE-2, способствуя ангиогенезу, а ANG1 действует в качестве агониста для нормализации структуры сосудов.
Как описано выше, процесс ангиогенеза в основном осуществляется по сигнальному механизму VEGF/VEGFR2, но для преодоления сосудистой утечки, вызванной ангиогенезом, важен сигнальный механизм ANG1-TIE2 с учетом стыковки эндотелиальных клеток сосудов и рекрутирования сосудистых перицитов.
В частности, когда интерстициальное давление возрастает вследствие сосудистой утечки, в новых кровеносных сосудах опухолей возникает гипоксия и тканевой некроз из-за сдавливания сосудов (Folkman J. et al., N. Engl. J. Med. 333:1757-63, 1995). По этой причине, даже если опухоль подвергается обработке терапевтическими средствами, терапевтический эффект снижается из-за снижения поступления препарата внутрь ткани (Gianfranco Baronzio et al., Front. Oncol. 5: 165, 2015).
Таким образом, для нормализации сосудов можно справиться с утечкой из новых кровеносных сосудов в опухоли путем активации сигнального механизма фосфорилирования TIE2. В конечном счете, это должно усилить поступление препарата и тем самым обеспечить противораковое действие, превосходящее таковое при стандартной монотерапии. Кроме того, нормализация сосудов снижает интерстициальное давление в тканях и ослабляет гипоксию, поэтому она должна обладать высоким потенциалом для лечения заболеваний, вызванных сосудистой утечкой, таких как язвы на ступнях, гипертония и диабетическая ретинопатия.
Кроме того, поскольку микросреда в опухоли перепрограммируется из существующего иммуносупрессивного состояния в иммунотолерантное состояние по мере усиления притока иммуноэффекторных Т-клеток при нормализации сосудов после применения сосудистых стабилизаторов (Huang et al., 2013, Cancer Research), то антитела против TIE2 вызывают нормализацию сосудов и тем самым усиливают приток иммуноэффекторных Т-клеток при введении в комбинации с ингибиторами контрольных точек иммунитета и т.п., усиливая иммунную активность, а тем самым и противораковое действие.
На этом техническом фоне авторы настоящего изобретения попытались получить антитело против TIE2. В результате они разработали антитело против TIE2, которое проявляет требуемую способность к связыванию с TIE2, и обнаружили, что это антитело против TIE2 действует в качестве стабилизатора сосудов, уменьшая утечку из сосудов, и предположили, что антитело против TIE2 будет применимо в качестве терапевтического средства для большинства видов рака и для лечения язв на ступнях и диабетической ретинопатии, связанных с аномальным ангиогенезом, наряду с раком. В частности, ожидается, что противораковое антитело к TIE2 будет способно стабилизировать кровеносные сосуды при введении вместе с традиционными терапевтическими средствами, тем самым повышая скорость доставки стандартных терапевтических средств и улучшая иммунную среду в микроокружении опухолей, проявляя при этом отличные противораковые эффекты в сочетании с иммунотерапевтическими средствами. На основе этой разработки и было совершено настоящее изобретение.
Сущность изобретения
Таким образом, одной из задач настоящего изобретения является получение нового антитела к TIE2 либо его антигенсвязывающего фрагмента.
Другой задачей настоящего изобретения является получение нуклеиновой кислоты, кодирующей антитело либо его антигенсвязывающий фрагмент.
Другой задачей настоящего изобретения является получение вектора, включающего нуклеиновую кислоту, клетки, трансформированной вектором, и способа их получения.
Другой задачей настоящего изобретения является получение фармацевтических композиций для профилактики и/или лечения опухолей или заболеваний, связанных с ангиогенезом, содержащих антитело либо его антигенсвязывающий фрагмент, в качестве активного ингредиента.
Другой задачей настоящего изобретения является получение фармацевтических композиций для стабилизации и/или нормализации сосудов, содержащих антитело либо его антигенсвязывающий фрагмент, в качестве активного ингредиента.
Другой задачей настоящего изобретения является получение фармацевтической композиции для монотерапии и/или для комбинированной терапии с другим терапевтическим средством, содержащей антитело либо его антигенсвязывающий фрагмент, в качестве активного ингредиента.
В соответствии с одним аспектом настоящего изобретения, вышеприведенные и другие задачи могут решаться путем получения антитела либо его антигенсвязывающего фрагмента, связывающихся с TIE-2 (тирозинпротеинкиназным рецептором), которые включают:
вариабельную область тяжелой цепи, содержащую участок, определяющий комплементарность связывания с антигеном (CDR1) тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 1, 7, 13, 19, 25 и 31, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 2, 8, 14, 20, 26 и 32, и CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 3, 9, 15, 21, 27 и 33, и
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 4, 10, 16, 22, 28 и 34, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 5, 11, 17, 23, 29 и 35, и CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 6, 12, 18, 24, 30 и 36.
В соответствии с другим аспектом настоящего изобретения предусмотрена нуклеиновая кислота, кодирующая антитело либо его антигенсвязывающий фрагмент.
В соответствии с другим аспектом настоящего изобретения предусмотрен вектор, включающий нуклеиновую кислоту.
В соответствии с другим аспектом настоящего изобретения предусмотрена клетка, трансформированная вектором. В соответствии с другим аспектом настоящего изобретения, предусмотрен способ получения антитела, связывающегося с TIE-2, либо его антигенсвязывающего фрагмента, включающий (a) культивирование клетки и (b) выделение антитела либо его антигенсвязывающего фрагмента из культивируемой клетки.
В соответствии с другим аспектом настоящего изобретения предусмотрен стабилизатор сосудов и/или композиция для лечения связанных с ангиогенезом заболеваний, содержащие антитело либо его антигенсвязывающий фрагмент.
В соответствии с другим аспектом настоящего изобретения предусмотрен стабилизатор сосудов и/или композиция для диагностики связанных с ангиогенезом заболеваний, содержащие антитело либо его антигенсвязывающий фрагмент.
В соответствии с другим аспектом настоящего изобретения предусмотрена композиция для профилактики или лечения опухоли или рака и/или связанных с ангиогенезом заболеваний, содержащая антитело либо его антигенсвязывающий фрагмент.
В соответствии с другим аспектом настоящего изобретения предусмотрена композиция для комбинированного введения с антителом против TIE2.
Краткое описание фигур
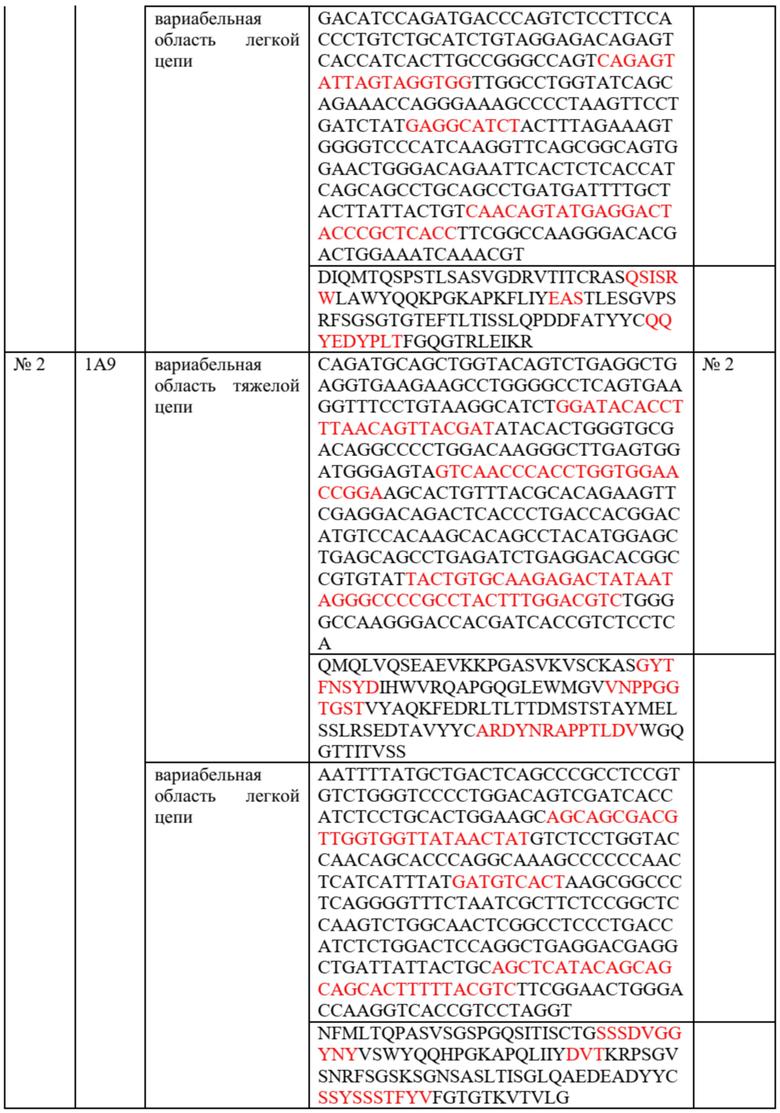
На фиг.1A и 1B представлены результаты проточной цитометрии по оценке связывания антител против TIE2 с клетками, экспрессирующими TIE2 человека и мыши, соответственно.
На фиг.2 представлены результаты ELISA по оценке связывания кратковременно экспрессированных и очищенных антител против TIE2 с TIE2 человека и мыши.
На фиг.3A-3C представлены результаты по оценке действия антител против TIE2 на миграцию клеток с использованием клеток линии HUVEC.
На фиг.4 представлены результаты, показывающие, что отобранное антитело против TIE2 связывается с TIE2 неконкурентно по отношению к ANG1 или ANG2.
На фиг.5A-5C представлены результаты, показывающие, что отобранное антитело против TIE2 может ингибировать индуцированную VEGF проницаемость сосудов.
На фиг.6 представлены результаты, показывающие, что отобранное антитело против TIE2 действует по механизму понижающей регуляции сигналлинга TIE2, подобно ANG1.
На фиг.7 представлены результаты, показывающие, что межклеточные адгезионные контакты на уровне белка в клетках определяют эффект стабилизации сосудов у отобранного антитела против TIE2.
На фиг.8A и 8B представлено улучшение межклеточных адгезионных контактов под действием антитела против TIE2.
На фиг.9 показано, что антитело против TIE2 может нормализировать индуцированную VEGF сосудистую утечку посредством сигналлинга TIE2 в ушах мыши.
На фиг.10A-10C представлены результаты по оценке ингибирующего действия антитела против TIE2 на ангиогенез в сетчатке.
На фиг.11A и 11B представлены результаты по оценке нормализирующего действия антитела против TIE2 на сосуды в опухоли.
На фиг.12A-12D представлены результаты по оценке противоопухолевого действия антитела против TIE2.
На фиг.13A-13C представлены результаты по оценке эффекта ослабления гипоксии у антитела против TIE2.
Раскрытие сущности изобретения
Если не указано иное, все технические и научные термины, используемые здесь, имеют такие же значения, как и те, что приняты специалистами в той области, к которой относится настоящее изобретение. В целом, используемая здесь номенклатура хорошо известна в данной области и обычно она и применяется.
Авторы настоящего изобретения отобрали новое антитело против TIE2. При активации TIE2 антителом или его антигенсвязывающим фрагментом, связывающимися с TIE2, согласно настоящему изобретению, кровеносные сосуды могут нормализироваться, чтобы предотвратить усиление гипоксии внутри опухоли или ракового новообразования, повысить приток крови к опухоли или раковому новообразованию и обеспечить достаточное поступление кислорода, тем самым повышая содержание других доставляемых противораковых препаратов и проникновение их в иммунные клетки.
В одном аспекте настоящего изобретения предусмотрено антитело либо его антигенсвязывающий фрагмент, связывающиеся с TIE-2, включающие вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 1, 7, 13, 19, 25 и 31, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 2, 8, 14, 20, 26 и 32, и CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 3, 9, 15, 21, 27 и 33, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 4, 10, 16, 22, 28 и 34, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 5, 11, 17, 23, 29 и 35, и CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 6, 12, 18, 24, 30 и 36.
В настоящем изобретении термин “антитело” означает такое антитело, которое специфически связывается с TIE-2. В объем настоящего изобретения входят не только полноразмерное антитело, специфически связывающееся с TIE2, но также антигенсвязывающий фрагмент этого антитела.
Термин “полноразмерное антитело” относится к структуре, содержащей две полноразмерные легкие цепи и две полноразмерные тяжелые цепи, причем каждая легкая цепь соединяется с соответствующей тяжелой цепью дисульфидной связью. Константная область тяжелой цепи включает изотипы гамма (γ), мю (μ), альфа (α), дельта (δ) и эпсилон (ε) и подразделяется на гамма-1 (γ1), гамма-2 (γ2), гамма-3 (γ3), гамма-4 (γ4), альфа-1 (α1) и альфа-2 (α2). Константная область легкой цепи включает изотипы каппа (κ) и лямбда (λ).
Антигенсвязывающий фрагмент антитела или фрагмент антитела представляет собой такой фрагмент, который обладает способностью связываться с антигеном и включает Fab, F(ab′), F(ab′)2, Fv и др. Среди фрагментов антитела Fab означает структуру, которая включает вариабельную область каждой из тяжелых цепей и легких цепей, константную область легкой цепи и первый константный домен (CH1) тяжелой цепи, причем каждый Fab-фрагмент содержит один антигенсвязывающий сайт.Fab′ отличается от Fab тем, что он также включает шарнирную область, содержащую по меньшей мере один остаток цистеина на С-конце домена CH1 тяжелой цепи. F(ab′)2 образуется за счет дисульфидной связи между остатками цистеина в шарнирной области Fab′. Fv - минимальный фрагмент антител, содержащий только вариабельную область тяжелой цепи и вариабельную область легкой цепи. Двухцепочечный Fv - это фрагмент, в котором вариабельная область тяжелой цепи и вариабельная область легкой цепи соединяются нековалентной связью, а одноцепочечный Fv (scFv) - это фрагмент, в котором вариабельная область тяжелой цепи и вариабельная область легкой цепи обычно соединяются ковалентной связью через пептидный линкер между ними или же непосредственно на C-конце, образуя структуру в форме димера типа двухцепочечного Fv. Такие фрагменты антител могут быть получены с помощью протеаз (например, Fab может быть получен при расщеплении полноразмерного антитела папаином, а F(ab′)2-фрагмент - при расщеплении полноразмерного антитела пепсином), а также методами генетической рекомбинации.
В одном воплощении антитело по настоящему изобретению представляет собой Fv (к примеру, scFv) или полноразмерное антитело. Кроме того, константная область тяжелой цепи может иметь изотип, выбранный из изотипов гамма (γ), мю (μ), альфа (α), дельта (δ) и эпсилон (ε). Например, константная область может быть гамма-1 (IgG1), гамма-3 (IgG3) или гамма-4 (IgG4), а константная область легкой цепи может иметь изотип каппа или лямбда.
Согласно настоящему изобретению, термин “тяжелая цепь” охватывает как полноразмерную тяжелую цепь, которая включает вариабельный домен (VH), содержащий аминокислотную последовательность, достаточную для придания специфичности к антигену, и три константных домена (CH1, CH2 и CH3), так и ее фрагменты. В настоящем изобретении термин “легкая цепь” охватывает как полноразмерную легкую цепь, которая включает вариабельный домен (VL), содержащий аминокислотную последовательность, достаточную для придания специфичности к антигену, и константный домен (CL), так и ее фрагменты.
Антитела по настоящему изобретению включают, без ограничения, моноклональные антитела, мультиспецифичные антитела, человеческие антитела, гуманизированные антитела, химерные антитела, scFV, одноцепочечные антитела, Fab-фрагменты, F(ab′)-фрагменты, Fvs с дисульфидной связью (sdFVs), антиидиотипические (анти-Id) антитела, эпитопсвязывающие фрагменты таких антител и др.
Термин “моноклональное антитело” относится к идентичным антителам, каждое из которых получают из популяции практически однородных антител, составляющих популяцию, за исключением возможных природных мутаций, которые могут присутствовать в незначительном числе. Моноклональные антитела высокоспецифичны, так как они вырабатываются против одного антигенного сайта. В отличие от обычных (поликлональных) препаратов антител, которые, как правило, включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты антигена.
Термин “эпитоп” означает такую детерминанту белка, с которой антитело может специфически связываться. Эпитопы обычно состоят из группы химически активных поверхностных молекул типа боковых цепей аминокислот или сахаров и обычно имеют не только специфические трехмерные структурные характеристики, но и специфические характеристики заряда. Трехмерные эпитопы отличаются от нетрехмерных эпитопов тем, что связь с первыми разрывается в присутствии денатурирующего растворителя, тогда как связь с последними не разрывается.
Нечеловеческие (например, мышиные) антитела в “гуманизированной” форме - это химерные антитела, включающие одну или несколько аминокислотных последовательностей (например, последовательностей CDR), происходящих из одного или нескольких нечеловеческих антител (донорных или исходных антител), содержащих минимальные последовательности, полученные из нечеловеческих иммуноглобулинов. В большинстве случаев гуманизированное антитело представляет собой иммуноглобулин человека (акцепторное антитело), в котором остатки из гипервариабельного участка реципиента заменены на остатки из гипервариабельного участка другого вида (донорного антитела) типа мыши, крысы, кролика или нечеловекообразного примата, обладающего требуемой специфичностью, аффинностью и активностью. Для гуманизирования можно заменить остатки в одном или нескольких каркасных доменах (FRs) вариабельной области акцепторного антитела человека на соответствующие остатки из донорного антитела другого вида. Это способствует поддержанию правильной трехмерной конфигурации пересаженных участков CDR, тем самым улучшая аффинность и стабильность антитела. Гуманизированные антитела также могут включать и новые остатки, которые отсутствуют в акцепторных или донорных антителах, к примеру, для дальнейшего улучшения характеристик антител.
Согласно настоящему изобретению термин “человеческое антитело” относится к молекулам, происходящим из человеческого иммуноглобулина, в которых все аминокислотные последовательности, составляющие антитело, включая участки, определяющие комплементарность связывания с антигеном, и структурные участки, имеют принадлежность к иммуноглобулинам человека.
Часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующей последовательности антител, происходящих из организма определенного вида или принадлежащих к определенному классу или подклассу антител, а остальная часть (части) “химерного” антитела (иммуноглобулина), идентична или гомологична соответствующим последовательностям антител, происходящих из организма другого вида или принадлежащих к другому классу или подклассу антител, а также их фрагментам, проявляющим требуемую биологическую активность.
В настоящем изобретении термин “вариабельная область антитела” относится к таким областям легкой и тяжелой цепи молекул антител, которые включают аминокислотные последовательности участков, определяющих комплементарность связывания с антигеном (CDR; т.е. CDR1, CDR2 и CDR3) и каркасных участков (FR). VH обозначает вариабельную область тяжелой цепи. VL обозначает вариабельную область легкой цепи.
Термин “участок, определяющий комплементарность связывания с антигеном” (CDR; то есть CDR1, CDR2 и CDR3) относится к таким аминокислотным остаткам вариабельной области антител, которые необходимы для связывания антигена. Каждая вариабельная область обычно содержит три участка CDR, обозначаемых как CDR1, CDR2 и CDR3.
В частности, антитело или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 1, CDR2 тяжелой цепи SEQ ID NO: 2 и CDR3 тяжелой цепи SEQ ID NO: 3, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 4, CDR2 легкой цепи SEQ ID NO: 5 и CDR3 легкой цепи SEQ ID NO: 6;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 7, CDR2 тяжелой цепи SEQ ID NO: 8 и CDR3 тяжелой цепи SEQ ID NO: 9, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 10, CDR2 легкой цепи SEQ ID NO: 11 и CDR3 легкой цепи SEQ ID NO: 12;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 15, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 19, CDR2 тяжелой цепи SEQ ID NO: 20 и CDR3 тяжелой цепи SEQ ID NO: 21, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 22, CDR2 легкой цепи SEQ ID NO: 23 и CDR3 легкой цепи SEQ ID NO: 24;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 25, CDR2 тяжелой цепи SEQ ID NO: 26 и CDR3 тяжелой цепи SEQ ID NO: 27, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 28, CDR2 легкой цепи SEQ ID NO: 29 и CDR3 легкой цепи SEQ ID NO: 30; или
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 31, CDR2 тяжелой цепи SEQ ID NO: 32 и CDR3 тяжелой цепи SEQ ID NO: 33, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 34, CDR2 легкой цепи SEQ ID NO: 35 и CDR3 легкой цепи SEQ ID NO: 36.
Термин “каркасный участок” (FR) обозначает другие остатки вариабельной области антител, чем остатки CDR. Каждая вариабельная обычно содержит четыре FR, обозначаемых как FR1, FR2, FR3 и FR4.
Антитело против TIE2 может быть моновалентным или бивалентным и может включать одинарные или двойные цепи. Функционально сродство связывания антител против TIE2 составляет от 10-5 М до 10-12 М. Например, сродство связывания антител против TIE2 составляет от 10-6 M до 10-12 M, от 10-7 M до 10-12 M, от 10-8 M до 10-12 M, от 10-9 M до 10-12 M, от 10-5 M до 10-11 M, от 10-6 M до 10-11 M, от 10-7 M до 10-11 M, от 10-8 M до 10-11 M, от 10-9 M до 10-11 M, от 10-10 M до 10-11 M, от 10-5 M до 10-10 M, от 10-6 M до 10-10 M, от 10-7 M до 10-10 M, от 10-8 M до 10-10 M, от 10-9 M до 10-10 M, от 10-5 M до 10-9 M, от 10-6 M до 10-9 M, от 10-7 M до 10-9 M, от 10-8 M до 10-9 M, от 10-5 M до 10-8 M, от 10-6 M до 10-8 M, от 10-7 M до 10-8 M, от 10-5 M до 10-7 M, от 10-6 M до 10-7 M или от 10-5 M до 10-6 M.
Связывающееся с TIE2 антитело или его антигенсвязывающий фрагмент может включать вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 38, 42, 46, 50, 54 и 58. Кроме того, связывающееся с TIE2 антитело или его антигенсвязывающий фрагмент может включать вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 40, 44, 48, 52, 56 и 60.
В частности, связывающееся с TIE2 антитело или его антигенсвязывающий фрагмент включает: вариабельную область тяжелой цепи SEQ ID NO: 38 и вариабельную область легкой цепи SEQ ID NO: 40;
вариабельную область тяжелой цепи SEQ ID NO: 42 и вариабельную область легкой цепи SEQ ID NO: 44;
вариабельную область тяжелой цепи SEQ ID NO: 46 и вариабельную область легкой цепи SEQ ID NO: 48;
вариабельную область тяжелой цепи SEQ ID NO: 50 и вариабельную область легкой цепи SEQ ID NO: 52;
вариабельную область тяжелой цепи SEQ ID NO: 54 и вариабельную область легкой цепи SEQ ID NO: 56; или
вариабельную область тяжелой цепи SEQ ID NO: 58 и вариабельную область легкой цепи SEQ ID NO: 60.
“Фаговый дисплей” - это метод экспонирования мутантных полипептидов в виде слитого белка по меньшей мере с частью белка оболочки на поверхности частиц фага, к примеру, нитевидного фага. Полезность фагового дисплея заключается в быстрой и эффективной классификации последовательностей, связывающихся с антигенами- мишенями с высоким сродством в больших библиотеках рандомизированных мутантных белков. Экспонирование пептидных и белковых библиотек на поверхности фагов использовалось для скрининга миллионов полипептидов при идентификации полипептидов со свойствами специфического связывания.
Технология фагового дисплея составила мощный инструмент для получения и скрининга новых белков, связывающихся с определенными лигандами (например, с антигенами). По технологии фагового дисплея можно создавать большие библиотеки мутантных белков и быстро классифицировать последовательности, связывающиеся с высоким сродством с целевыми антигенами. Нуклеиновую кислоту, кодирующую мутантные полипептиды, сливают с последовательностью нуклеиновой кислоты, кодирующей белок оболочки вируса, например, белок гена III или гена VIII. Была разработана монофазная система фагового дисплея, в которой последовательность нуклеиновой кислоты, кодирующая белок или полипептид, сливается с последовательностью нуклеиновой кислоты, кодирующей часть белка гена III. В монофазной системе дисплея слитый ген экспрессируется на низком уровне, а белок гена III дикого типа также экспрессируется, и, таким образом, сохраняется инфекционность частиц.
Для разработки библиотек фагового дисплея антител важно продемонстрировать экспрессию пептидов на поверхности нитевидного фага и экспрессию функциональных фрагментов антител в периферической цитоплазме E.coli. Библиотеки антител или антигенсвязывающих полипептидов получают различными методами, например, методом модификации одного гена путем вставки случайной последовательности ДНК или клонирования последовательности родственного гена. Библиотеки можно подвергать скринингу на предмет экспрессии антител или антигенсвязывающих белков с нужными характеристиками.
Технология фагового дисплея имеет несколько преимуществ по сравнению с обычными гибридомными и рекомбинантными методами получения антител с нужными характеристиками. Этот метод обеспечивает получение больших библиотек антител с различными последовательностями за короткое время без использования животных. Для получения гибридом и получения гуманизированных антител может потребоваться время в несколько месяцев. Кроме того, поскольку иммунитет не требуется, фаговые библиотеки антител могут генерировать антитела против таких антигенов, которые являются токсичными или имеют низкую антигенность. Фаговые библиотеки антител также можно использовать для получения и идентификации новых терапевтических антител.
Можно использовать методы получения человеческих антител из иммуногенных или неиммуногенных последовательностей зародышевой линии человека либо из Ig-репертуаров наивных B-клеток с использованием библиотек фагового дисплея. Можно получать наивные или неиммуногенные антигенсвязывающие библиотеки с использованием различных лимфатических тканей.
Для выделения новых терапевтических антител важны методы идентификации и выделения высокоаффинных антител из библиотек фагового дисплея. Выделение высокоаффинных антител из библиотек зависит от размера библиотеки, эффективности продукции в бактериальных клетках и разнообразия библиотеки. Размер библиотек снижается при неправильной укладке антитела или антигенсвязывающего белка и неэффективной продукции из-за присутствия стоп-кодона. Экспрессия в бактериальных клетках может подавляться при неправильной укладке антитела или антигенсвязывающего домена. Экспрессия может быть улучшена путем поочередной мутации остатков на границе раздела вариабельной/константной области или выбранных остатков CDR. Последовательность каркасных участков является тем элементом, который обеспечивает правильную укладку при получении фаговых библиотек антител в бактериальных клетках.
При выделении высокоаффинных антител важно создавать разнообразные библиотеки антител или антигенсвязывающих белков. Часто было отмечено, что участки CDR3 участвуют в связывании антигена. Поскольку участки CDR3 в тяжелой цепи значительно различаются по размерам, последовательности и структурной/пространственный морфологии, то с их помощью можно получать разнообразные библиотеки.
Также можно создавать разнообразие путем рандомизации участков CDR вариабельных тяжелых и легких цепей, используя все 20 аминокислот в каждом положении. При использовании всех 20 аминокислот образуются последовательности антител с повышенным разнообразием и повышенной вероятностью идентификации новых антител.
Антитела или фрагменты антител по настоящему изобретению могут включать последовательности упомянутых здесь антител против TIE2, а также их биологических эквивалентов, если только они могут специфически распознавать TIE2. Например, в аминокислотную последовательность антител можно вносить дополнительные вариации, чтобы еще больше улучшить сродство связывания и/или другие биологические свойства антител. Такие вариации включают, к примеру, делеции, вставки и/или замены остатков
в аминокислотной последовательности антител. Такие аминокислотные вариации основываются на относительном сходстве заместителей в боковой цепи аминокислот, например, их гидрофобности, гидрофильности, заряда и размера. При анализе размера, формы и типа заместителей в боковой цепи аминокислот видно, что аргинин, лизин и гистидин являются положительно заряженными остатками; аланин, глицин и серин имеют близкие размеры; а фенилаланин, триптофан и тирозин имеют близкие формы. Так, исходя из этих соображений, считается, что аргинин, лизин и гистидин; аланин, глицин и серин; и фенилаланин, триптофан и тирозин образуют соответствующие группы биологически функциональных эквивалентов.
Принимая во внимание варианты, обладающие биологически эквивалентной активностью, аминокислотные последовательности антител или кодирующих их молекул нуклеотидов по настоящему изобретению интерпретируются как включающие последовательности, которые по существу идентичны последовательностям, приведенным под их порядковыми номерами. Термин “по существу идентичны” означает, что последовательности гомологичны по меньшей мере на 90%, наиболее предпочтительно гомологичны по меньшей мере на 95%, на 96%, на 97%, на 98% или на 99% при совмещении последовательности по настоящему изобретению с любой другой последовательностью так, чтобы они соответствовали друг другу настолько близко, насколько это возможно, при анализе совмещенных последовательностей с помощью алгоритмов, обычно используемых в данной области. Методы совмещения для сравнения последовательностей хорошо известны в данной области. Базовый инструмент поиска локального совмещения NCBI (BLAST) доступен через NCBI и т.п.и может использоваться в сочетании с программами анализа последовательностей типа BLASTP, BLASTM, BLASTX, TBLASTN и TBLASTX через Интернет.BLAST доступен на www.ncbi.nlm.nih.gov/BLAST/. Метод сравнения гомологичных последовательностей с помощью этой программы находится на сайте www.ncbi.nlm.nih.gov/BLAST/blast_help.html.
Исходя из этого, антитело либо его антигенсвязывающий фрагмент по настоящему изобретению могут иметь гомологию в 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и более с приведенной здесь последовательностью или же полностью совпадать с ней. Гомология может определяться путем сравнения и/или совмещения последовательностей известными в данной области методами. Например, степень гомологии последовательностей нуклеиновых кислот или белков по настоящему изобретению можно определить с помощью алгоритма сравнения последовательностей (т.е. BLAST или BLAST 2.0), совмещения вручную или визуального осмотра.
В другом аспекте настоящего изобретения предусмотрена нуклеиновая кислота, кодирующая антитело либо его антигенсвязывающий фрагмент.
Нуклеиновая кислота может кодировать вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 37, 41, 45, 49, 53 и 57. С другой стороны, нуклеиновая кислота может кодировать вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 39, 43, 47, 51, 55 и 59.
При выделении нуклеиновой кислоты, кодирующей антитело или его антигенсвязывающий фрагмент по настоящему изобретению, можно получить антитело или его антигенсвязывающий фрагмент рекомбинантным способом. Нуклеиновую кислоту выделяют и вставляют в реплицирующийся вектор с последующим клонированием (амплификацией ДНК) или дальнейшей экспрессией. На основе изложенного в следующем аспекте настоящего изобретения предусмотрены векторы, включающие нуклеиновую кислоту.
Термин “нуклеиновая кислота” используется для обозначения молекул как ДНК (гДНК и кДНК), так и РНК, а нуклеотиды, которые являются основной составляющей нуклеиновых кислот, включают нуклеотиды природного происхождения, а также их аналоги, в которых модифицированы молекулы сахара или основания. Последовательности нуклеиновых кислот, кодирующих вариабельные области тяжелой и легкой цепи по настоящему изобретению, могут варьировать. Такие вариации включают вставки, делеции, либо неконсервативные или консервативные замены нуклеотидов.
ДНК, кодирующую антитело, можно легко выделить или синтезировать по стандартным методикам (например, с помощью олигонуклеотидного зонда, способного специфически связываться с ДНК, кодирующей тяжелую и легкую цепь антитела). Доступны различные векторы. Компоненты вектора обычно включают, без ограничения, один или несколько из следующих компонентов: сигнальные последовательности, точки начала репликации, один или несколько маркерных генов, энхансерные элементы, промоторы и последовательности терминации транскрипции.
В настоящем изобретении термин “вектор” означает средство экспрессии целевых генов в клетках-хозяевах и включает плазмидные векторы, космидные векторы и вирусные векторы типа векторов-бактериофагов, аденовирусных векторов, ретровирусных векторов и аденоассоциированных вирусных векторов. Нуклеиновая кислота, кодирующая антитело, в векторе функционально связана с промотором.
Термин “функционально связана” означает функциональную связь между последовательностью, регулирующей экспрессию нуклеиновой кислоты (например, комплексом сайта связывания промотора, сигнальной последовательности или регулятора транскрипции), и другой последовательностью нуклеиновой кислоты, которая позволяет регулирующей последовательности направлять транскрипцию и/или трансляцию другой последовательности нуклеиновой кислоты.
При использовании прокариотических клеток в качестве хозяев они обычно содержат сильный промотор, способный обеспечивать транскрипцию (например, промотор tac, промотор lac, промотор lacUV5, промотор lpp, промотор pLλ, промотор pRλ, промотор rac5, промотор amp, промотор recA, промотор SP6, промотор trp или промотор Т7), сайт связывания с рибосомой для инициации трансляции и последовательность терминации транскрипции/трансляции. Кроме того, к примеру, при использовании эукариотических клеток в качестве хозяев, они включают промотор, происходящий из генома клеток млекопитающих (например, промотор металлотионеина, промотор β-актина, промотор гемоглобина человека или промотор мышечного креатина человека), или промотор, происходящий из вируса млекопитающих (например, поздний промотор аденовируса, промотор 7.5K вируса коровьей оспы, промотор SV40, промотор цитомегаловируса (CMV), промотор tk HSV, промотор вируса опухолей молочной железы мыши (MMTV), промотор LTR HIV, промотор вируса Молони, промотор вируса Эпштейна-Барр (EBV) или промотор вируса саркомы Рауса (RSV)), и обычно содержит последовательность полиаденилирования в качестве последовательности терминации транскрипции.
Необязательно, вектор может быть слит с другой последовательностью с тем, чтобы облегчить очистку экспрессируемого им антитела. Последовательность для слияния может включать, к примеру, глутатион-S-трансферазу (Pharmacia, USA), мальтозосвязывающий белок (NEB, США), FLAG (IBI, США), 6xHis (гексагистидин; Qiagen, США) и др.
Векторы содержат гены устойчивости к антибиотикам, которые обычно используются в данной области в качестве селективных маркеров, а их примеры включают гены, придающие устойчивость к ампициллину, гентамицину, карбенициллину, хлорамфениколу, стрептомицину, канамицину, генетицину, неомицину и тетрациклину.
В следующем аспекте настоящего изобретения предусмотрена клетка, содержащая вышеприведенные векторы, которые трансформированы указанными выше векторами. Клетка, используемая для получения антитела по настоящему изобретению, может быть прокариотической, дрожжевой или высшей эукариотической клеткой, без ограничения.
Можно использовать прокариотические клетки-хозяева типа Escherichia coli, штаммы рода Bacillus типа Bacillus subtilis и Bacillus thuringiensis, Streptomyces spp., Pseudomonas spp.(к примеру, Pseudomonas putida), Proteus mirabilis и Staphylococcus spp.(к примеру, Staphylococcus carnosus).
Наибольший интерес представляют клетки животных, а примеры полезных линий клеток-хозяев включают, без ограничения, COS-7, BHK, CHO, CHOK1, DXB-11, DG-44, CHO/DHFR, CV1, COS-7, HEK293, BHK, TM4, VERO, HELA, MDCK, BRL 3A, W138, Hep G2, SK-Hep, MMT, TRI, MRC 5, FS4, 3T3, RIN, A549, PC12, K562, PER.C6, SP2/0, NS-0, U20S и HT1080.
В следующем аспекте настоящего изобретения предусмотрен способ получения антитела либо его антигенсвязывающего фрагмента, включающий: (a) культивирование клетки; и (b) выделение антитела либо его антигенсвязывающего фрагмента из культивируемой клетки.
Клетки можно культивировать в различных средах. В качестве культуральной среды можно использовать любые коммерчески доступные среды, без ограничений. Можно вносить в среды и все другие необходимые добавки, хорошо известные специалистам в данной области, в соответствующих концентрациях. Используются условия культивирования при определенных температуре и pH, обычные для клеток- хозяев, выбранных для экспрессии, как должно быть ясно специалистам в данной области.
Выделение антитела либо его антигенсвязывающего фрагмента может осуществляться, к примеру, посредством центрифугирования или ультрафильтрации для удаления примесей и очистки полученного продукта при использовании, скажем, методов аффинной хроматографии. Можно использовать и другие методы очистки, такие, как анионо- или катионообменная хроматография, хроматография гидрофобных взаимодействий и хроматография на гидроксиапатите (ГА).
В следующем аспекте настоящего изобретения предусмотрена композиция для профилактики или лечения связанных с ангиогенезом заболеваний, содержащая антитело либо его антигенсвязывающий фрагмент в качестве активного ингредиента.
Ангиогенез - это явление, при котором образуются или растут новые кровеносные сосуды из уже существующих кровеносных сосудов, а термин “связанные с ангиогенезом заболевания” относится к заболеваниям, связанным с возникновением или прогрессированием ангиогенеза. В число связанных с ангиогенезом заболеваний могут попадать любые заболевания, без ограничения, если только их можно лечить с помощью антитела. Примеры связанных с ангиогенезом заболеваний включают, без ограничения, рак, метастазы рака, диабетическую ретинопатию, ретинопатию недоношенных, отторжение трансплантата роговицы, дегенерацию желтого пятна, неоваскулярную глаукому, эритроз, пролиферативную ретинопатию, псориаз, гемофилический артрит, образование атеросклеротических бляшек в капиллярах, келоиды, грануляцию ран, сосудистые спайки, ревматоидный артрит, остеоартрит, аутоиммунные заболевания, болезнь Крона, рестеноз, атеросклероз, кишечные спайки, болезни от кошачьих царапин, язвы, цирроз печени, нефрит, диабетическую нефропатию, диабетические язвы стопы, хроническую почечную недостаточность, сахарный диабет, воспалительные заболевания, идиопатический фиброз легких, сепсис, острый респираторный дистресс-синдром и нейродегенеративные заболевания.
Кроме того, рак выбирают из группы, состоящей из рака пищевода, рака желудка, рака толстого кишечника, рака прямой кишки, рака полости рта, рака глотки, рака гортани, рака легких, рака толстой кишки, рака груди, рака шейки матки, рака эндометрия, рака яичников, рака простаты, рака яичек, рака мочевого пузыря, рака почек, рака печени, рака поджелудочной железы, рака костей, рака соединительной ткани, рака кожи, рака головного мозга, рака щитовидной железы, лейкемии, Ходжкинской лимфомы, лимфомы и множественного миелоидного рака крови, без ограничения указанным.
В настоящем изобретении термин “предотвращение или профилактика” означает такое действие, которое подавляет или замедляет возникновение данного заболевания посредством введения антитела или композиции по настоящему изобретению. Термин “лечение или терапия” означает такое действие, которое облегчает или положительно изменяет симптомы данного заболевания посредством введения антитела или композиции.
В следующем аспекте настоящего изобретения предусмотрена композиция для стабилизации кровеносных сосудов, содержащая антитело либо его антигенсвязывающий фрагмент в качестве активного ингредиента.
В настоящем изобретении термин “стабилизация сосудов” означает свойство кровеносных сосудов поддерживать свою функцию посредством облегчения или ослабления действия, или устранения факторов, которые могут нарушить целостность кровеносных сосудов, либо замедления, ослабления или устранения повреждений кровеносных сосудов. В частности, “стабилизация сосудов” может включать ингибирование проницаемости кровеносных сосудов, замедление повреждения кровеносных сосудов и/или восстановление целостности поврежденных кровеносных сосудов.
Композиция, содержащая антитело по настоящему изобретению, предпочтительно является фармацевтической композицией и может дополнительно содержать подходящие носители, эксципиенты или разбавители, обычно используемые в данной области.
Фармацевтическая композиция, содержащая фармацевтически приемлемые носители, может представлять собой различные пероральные или парентеральные дозированные формы, как-то таблетки, пилюли, порошки, гранулы, капсулы, суспензии, пероральные растворы, эмульсии, сиропы, стерильные водные растворы, неводные растворы, суспензии, лиофилизаты и свечи. В этом отношении фармацевтическая композиция по настоящему изобретению может быть скомбинирована с такими разбавителями или эксципиентами, как наполнители, загустители, связующие, смачивающие вещества, разрыхлители, поверхностно-активные вещества и др. Твердые препараты для перорального введения могут быть в виде таблеток, пилюль, порошков, гранул, капсул и др. Такие твердые формы могут быть получены путем объединения вещества по настоящему изобретению по меньшей мере с одним эксципиентом типа крахмала, карбоната кальция, сахарозы, лактозы или желатина. Также можно использовать простые эксципиенты, смазывающие вещества типа стеарата магния, талька и др. Жидкие формы для перорального введения могут представлять собой суспензии, пероральные растворы, эмульсии, сиропы и т.п.Жидкие формы могут быть получены путем включения различных эксципиентов типа простых разбавителей, к примеру, воды или вазелинового масла, смачивающих средств, подсластителей, ароматизаторов, консервантов и пр. Кроме того, фармацевтическая композиция по настоящему изобретению может представлять собой парентеральные формы, как-то стерильные водные растворы, неводные растворы, суспензии, эмульсии, лиофилизаты, свечи и т.п.Для инъекций неводных растворов и суспензий можно использовать пропиленгликоль, полиэтиленгликоль, растительные масла типа оливкового масла и сложные эфиры типа этилолеата. Основа свечей может включать витепсол, макрогол, Твин 61, масло какао, лауриновое масло и глицерожелатин.
Фармацевтическую композицию по настоящему изобретению можно вводить в фармацевтически эффективном количестве, причем термин “фармацевтически эффективное количество” означает количество, достаточное для лечения заболеваний при разумном соотношении польза/риск, применимом ко всем медицинским методам лечения. Эффективное количество может варьировать в зависимости от различных факторов, в том числе тяжести подлежащего лечению заболевания, возраста и пола пациента, типа заболевания, активности препарата, чувствительности пациента к препарату, времени введения, способа введения, скорости выведения, продолжительности лечения, совместного введения лекарств и других факторов, хорошо известных в области фармацевтики. Кроме того, композицию по настоящему изобретению можно вводить саму по себе или в сочетании с другими терапевтическими средствами. В этом случае композицию можно вводить последовательно или одновременно с обычными терапевтическими средствами. Кроме того, композицию можно вводить в разовых или множественных дозах. Принимая во внимание эти факторы, следует вводить композицию в минимальном количестве, достаточном для достижения максимальной эффективности без побочных эффектов. Кроме того, количество вводимой композиции может определяться специалистами в данной области. Дозировка фармацевтической композиции по настоящему изобретению ничем особо не ограничивается и изменяется в зависимости от различных факторов, включая состояние здоровья и вес пациента, тяжесть заболевания, тип препарата, способ введения и время введения. Композицию можно вводить млекопитающим, включая мышей, крыс, домашний скот, человека и т.п., в разовых или множественных дозах каждый день, традиционно приемлемым способом, например, перорально, ректально, внутривенно, подкожно, внутриматочно или интрацереброваскулярно.
В следующем аспекте настоящего изобретения предусмотрен способ ингибирования ангиогенеза и способ предотвращения или лечения связанных с ангиогенезом заболеваний, включающий введение антитела или композиции нуждающемуся в этом субъекту.
Способ по настоящему изобретению включает введение фармацевтически эффективного количества фармацевтической композиции субъекту, нуждающемуся в ингибировании ангиогенеза. Субъектами могут быть млекопитающие, как-то собаки, коровы, лошади, кролики, мыши, крысы, куры или люди, без ограничения. Фармацевтическую композицию можно вводить любым подходящим способом, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное или интраназальное введение, в том числе, при необходимости, внутриочаговое введение для местного лечения. Предпочтительные дозировки фармацевтической композиции по настоящему изобретению могут варьировать в зависимости от различных факторов, включая состояние здоровья и вес субъекта, тяжесть заболевания, тип препарата, способ введения и время введения, и они легко определяются специалистами в данной области.
В следующем аспекте настоящего изобретения предусмотрена фармацевтическая композиция для профилактики или лечения рака, содержащая антитело, либо способ профилактики или лечения рака, включающий введение антитела или композиции нуждающемуся в этом субъекту. Термины “антитело”, “профилактика” и “лечение” уже определены выше.
Любой рак может быть мишенью для лечения, без ограничения, если только его можно лечить с помощью антитела. В частности, антитело по настоящему изобретению может предотвращать возникновение или прогрессирование рака путем ингибирования ангиогенеза. Примеры рака включают, без ограничения, рак пищевода, рак желудка, рак толстого кишечника, рак прямой кишки, рак полости рта, рак глотки, рак гортани, рак легких, рак толстой кишки, рак груди, рак шейки матки, рак эндометрия, рак яичников, рак простаты, рак яичек, рак мочевого пузыря, рак почек, рак печени, рак поджелудочной железы, рак костей, рак соединительной ткани, рак кожи, рак головного мозга, рак щитовидной железы, лейкемию, Ходжкинскую лимфому, лимфому и множественный миелоидный рак крови.
Кроме того, антитело по настоящему изобретению может применяться в комбинации с другими антителами или биологически активными средствами или веществами для различных целей. В этом аспекте настоящего изобретения предусмотрена композиция для комбинированного введения с другим терапевтическим средством от ангиогенеза, причем композиция содержат антитело либо его антигенсвязывающий фрагмент.
Другие терапевтические средства от ангиогенеза включают антиангиогенные средства, противовоспалительные средства и/или противоопухолевые средства. Эти терапевтические средства могут преодолевать устойчивость к ним и повышать их эффективность.
При введении композиции по настоящему изобретению в комбинации с другими терапевтическими средствами от связанных с ангиогенезом заболеваний антитело к TIE2 и другие терапевтические средства для лечения связанных с ангиогенезом заболеваний можно вводить последовательно или одновременно. Например, субъекту вводят антиангиогенное средство, противовоспалительное средство и/или противораковое средство, а затем ему вводят композицию, содержащую антитело против TIE2 или его антигенсвязывающий фрагмент в качестве активного ингредиента, или же субъекту вводят композицию, а затем ему вводят антиангиогенное средство, противовоспалительное средство и/или противораковое средство. В некоторых случаях композицию можно вводить субъектам одновременно вместе с антиангиогенным средством, противовоспалительным средством и/или противораковым средством.
В следующем аспекте настоящего изобретения предусмотрена композиция для диагностики связанных с ангиогенезом заболеваний, содержащая антитело либо его антигенсвязывающий фрагмент в качестве активного ингредиента. Настоящим изобретением предусмотрен способ диагностики связанных с ангиогенезом заболеваний у субъектов путем введения им антитела либо его антигенсвязывающего фрагмента.
Связанные с ангиогенезом заболевания можно диагностировать путем измерения уровня экспрессии TIE2 в образцах при помощи антитела либо его антигенсвязывающего фрагмента. Уровень экспрессии можно измерить с помощью антитела стандартным методом иммуноанализа типа радиоиммуноанализа, радиоиммунопреципитации, иммунопреципитации, иммуногистохимического окрашивания, твердофазного иммуноферментного анализа (ELISA), ELISA с захватом антигена, анализа ингибирования или конкуренции, “сэндвич”-анализа, проточной цитометрии, иммунофлуоресцентного окрашивания и иммуноаффинной очистки, без ограничения указанным
Рак можно диагностировать путем анализа интенсивности конечного сигнала в процессе иммуноанализа. А именно, когда маркерный белок, согласно настоящему изобретению, высоко экспрессируется в биологическом образце и генерирует более сильный сигнал, чем в нормальном биологическом образце (например, в нормальной ткани желудка, крови, плазме или сыворотке), то ставится диагноз рака.
В следующем аспекте настоящего изобретения предусмотрен набор для диагностики рака, содержащий композицию для диагностики рака. Набор по настоящему изобретению включает антитело против TIE2 и может диагностировать рак по сигналам, генерируемым во время реакции между образцом и антителом. В этом случае сигнал может включать фермент, связанный с антителом, типа щелочной фосфатазы, β-галактозидазы, пероксидазы хрена, люциферазы или цитохрома P450, без ограничения. При этом, если в качестве фермента используется щелочная фосфатаза, то в качестве субстрата для цветной реакции используется бромхлориндолилфосфат (BCIP), нитросиний тетразолий (NBT), нафтол-AS-B1-фосфат или усилитель хемифлуоресценции (ECF), а если в качестве фермента используется пероксидаза хрена, то в качестве субстрата может использоваться хлорнафтол, аминоэтилкарбазол, диаминобензидин, D-люциферин, люцигенин (бис-N-метилакридиний нитрат), бензиловый эфир резоруфина, люминол, реагент Amplex Red (10-ацетил-3,7-дигидроксифеноксазин), HYR (п-фенилендиамин⋅HCl и пирокатехол), TMB (тетраметилбензидин), ABTS (2,2′-азин-ди[3-этилбензтиазолинсульфонат]), o-фенилендиамин (OPD), нафтол/пиронин, глюкозоксидаза, t-NBT (нитросиний тетразолий) и m-PMS (феназинметосульфат), но настоящее изобретение этим не ограничивается.
Кроме того, набор по настоящему изобретению также может включать и метку для генерирования детектируемого сигнала, причем метка может представлять собой химическое вещество (например, биотин), фермент (щелочную фосфатазу, β-галактозидазу, пероксидазу хрена и цитохром P450), радиоактивное вещество (типа C14, I125, P32 и S35), флуоресцентное вещество (например, флуоресцеин), люминесцентное вещество, хемилюминесцентное вещество и FRET (флуоресцентно-резонансный перенос энергии), без ограничения указанным.
Измерение активности фермента, используемого для диагностики ангиогенеза, или измерение сигнала может проводиться различными способами, известными в данной области. Таким образом, можно качественно или количественно анализировать экспрессию TIE2.
Примеры
Далее настоящее изобретение будет раскрыто более подробно с помощью примеров. Однако специалистам в данной области должно быть ясно, что эти примеры приводятся только для иллюстрации настоящего изобретения и не должны рассматриваться как ограничивающие объем настоящего изобретения.
Пример 1. Скрининг антител, связывающихся с TIE2
Для скрининга антител, связывающихся с TIE2, и получения библиотек использовали библиотеки наивных scFv человека (scFv), раскрытые в Korean Patent Laid-Open Publication No. 10-2008-0109417. В 96-луночный Ni+планшет вносили по 100 мкл на лунку антигена (hTIE2-his: Sino Biological, 10700-H08H) с концентрацией 2 мкг/мл и инкубировали при 4°C в течение ночи. На следующий день покрытый антигеном планшет трижды промывали 0,1% TBST, а затем инкубировали с 200 мкл блокирующего буфера с 2% BSA при комнатной температуре в течение 2 часов. К 2 мл ростовой среды 2xYT-TET (тетрациклин 10 мкг/мл) добавляли 50 мкл исходного раствора XL1-Blue и инкубировали при 37°C в течение 2 ч при 200 об/мин, а затем добавляли еще 13 мл и инкубировали до достижения OD600=0,5. После блокирования в течение 2 часов промывали лунки 3 раза 1xPBS. В каждую промытую лунку вносили порцию фаговой библиотеки, причем фаговую библиотеку смешивали с 4% BSA в равных количествах, а затем вносили по 200 мкл смеси, после чего проводили реакцию при комнатной температуре в течение 2,5 часов. По завершении реакции с фаговой библиотекой отбрасывали супернатанты, а остатки промывали 5 раз 0,1% TBST и 3 раза TBS, добавляли в каждую лунку 100 мкл 100 мМ TEA (триметиламина) и встряхивали полученную смесь при комнатной температуре в течение 10 мин. Через 10 минут в каждую лунку добавляли 50 мкл 1M Триса (pH 7,5), а затем перемешивали. К 10 мл XL1-Blue с OD600=0,5 добавляли супернатанты для индукции инфицирования при 37°C в течение 30 мин. По завершении инфицирования использовали 100 мкл для итогового титра, а остаток центрифугировали при 6000 об/мин в течение 10 мин. Супернатант отбрасывали, а осадок наносили шпателем на большую квадратную чашку (CM 34 мкг/мл+1% глюкозы) и инкубировали при 30°C в течение ночи. Оставленные для итогового титра 100 мкл разводили 1/10, 1/100 и 1/1000, наносили на чашки с CM и инкубировали при 37°C в течение ночи. На следующий день выросшие на квадратной чашке колонии соскребали с помощью петли после добавления туда 50 мл среды 2xYT и центрифугировали при 6000 об/мин в течение 10 мин, после чего супернатант отбрасывали, а осадок использовали для получения первичного материала для пэннинга. В коническую колбу на 500 мл помещали 100 мл культуральной среды 2xYT (ростовая среда: CM 34 мкг/мл+1% глюкозы), туда же добавляли клетки до получения OD600=0,2 и культивировали при 200 об/мин и 37°C до достижения OD600=0,5. После культивирования клеток до достижения OD600=0,5 добавляли фаг-помощник (мутант M13KO7) в количестве, в 20 раз превышающем количество клеток. После заражения фагом-помощником при 37°C в течение 30 мин проводили центрифугирование при 6000 об/мин в течение 10 мин. Супернатант отбрасывали, а клетки инкубировали в течение ночи в 100 мл свежей среды 2xYT (CM 34 мкг/мл+Kan. 70 мкг/мл+1 мМ IPTG+5 мМ MgCl2) при 200 об/мин и 30°C. На следующий день выращенные клетки центрифугировали при 7000 об/мин в течение 10 мин и еще раз таким же образом, как и выше. Собранный супернатант осаждали на льду в присутствии 20% ПЭГ/2,5 М NaCl в количестве 1/5 (об/об) от количества супернатанта в течение 1 часа. После осаждения проводили центрифугирование при 9000 об/мин в течение 1 часа. Супернатант отбрасывали, а осадок суспендировали в 3 мл TBS, фильтровали через фильтр на 0,45 мкм, хранили при 4°C и использовали в следующем раунде пэннинга. Этот процесс повторяли 3-4 раза, а связывание антител с антигеном определяли методом ELISA.
Пример 2. Скрининг несущих моноклональные scFv фагов на специфическое связывание с TIE2 и нейтрализацию связывания с TIE2 (связывающий ELISA)
По завершении процесса пэннинга клетки из последнего раунда разводили до получения 200-500 колоний, наносили шпателем на чашку с агаром CM, а затем инкубировали в течение ночи при 37°C. На следующий день, когда выросли колонии, в 96-луночный глубокий планшет добавляли по 200 мкл среды 2xYT (CM 34 мкг/мл+1% глюкозы), в каждую лунку вносили по одной колонии, а затем инкубировали планшет в течение ночи при 37°C и 3000 об/мин. На следующий день в свежий 96-луночный глубокий планшет добавляли по 200 мкл среды 2xYT (CM 34 мкг/мл+1% глюкозы), в каждую лунку вносили по 20 мкл выращенных накануне клеток и культивировали при 37°C и 3000 об/мин в течение 1 часа 10 минут.Оставшиеся клетки хранили при -70°C в 100 мкл 50% глицерина. Когда клетки подросли, смешивали 1 мкл фага-помощника с 19 мкл среды 2xYT, вносили по 20 мкл полученной смеси в каждую лунку и инкубировали 30 минут при 37°C. После инкубации проводили центрифугирование при 3000 об/мин в течение 10 мин. Супернатанты отбрасывали, а к остаткам добавляли по 200 мкл среды 2xYT (CM 34 мкг/мл+Kan. 70 мкг/мл+1 мМ IPTG+5 мМ MgCl2), а затем инкубировали в MegaGrow при 30°C и 3000 об/мин в течение ночи.
Для отбора фагов, специфически связывающихся с TIE2, сначала в 96-луночный планшет с Ni+вносили 2 мкг/мл антигена (hTIE2-his: Sino Biological, 10700-H08H) по 100 мкл на лунку, а затем инкубировали при 4°C в течение ночи. На следующий день выращенные накануне клетки центрифугировали при 3000 об/мин в течение 10 минут и хранили при 4°C. Покрытый антигеном планшет промывали 3 раза 0,1% TBST, а затем туда же добавляли 200 мкл блокирующего буфера с 2% BSA и инкубировали при 25°C в течение 2 часов. По завершении блокирования планшет отмывали 3 раза 0,1% TBST. В каждой лунке смешивали 50 мкл 4% BSA с 50 мкл фага, хранившегося при пониженной температуре 4°C, и инкубировали со встряхиванием при комнатной температуре в течение 1 часа. По завершении связывания фага продукт реакции трижды промывали 0,1% TBST, добавляли 100 мкл конъюгированного с HRP мышиного антитела против M13 (1:3000, Sino, 11973-MM05) и инкубировали при 25°C в течение 1 часа. По завершении реакции полученный продукт промывали 3 раза 0,1% TBST, добавляли 100 мкл TMB (из набора реагентов для субстрата TMB #BD 555214), проявляли окраску в течение 3-5 мин и добавляли 50 мкл стоп-раствора, а затем проводили анализ на считывающем устройстве для ELISA.
В таблице 1 представлены результаты измерения специфического связывания экспонирующих моноклональные scFv фагов с антигеном TIE2 методом ELISA, а в следующих таблицах 2 и 3 представлены нуклеотидные последовательности отобранных антител.
Таблица 1. Результаты измерения специфического связывания экспонирующих моноклональные scFv фагов с антигеном TIE2 методом ELISA
Таблица 2. Последовательности CDR антител, специфически связывающихся с антигеном TIE2
Таблица 3. Последовательности вариабельной области антител, специфически связывающихся с антигеном TIE2


Пример 3. Отбор антител, способных связываться с клетками, экспрессирующими TIE2
Для того, чтобы определить связывание с клетками, экспрессирующими TIE2, измеряли связывание с гиперэкспрессирующими TIE2 клетками установленных линий CHO-K1 человека/мыши (hTIE2/CHO-K1, mTIE2/CHO-K1) методом проточной цитометрии. Каждую линию клеток поддерживали и культивировали, а клетки hTIE2/CHO-K1 промывали PBS, добавляли 5 мМ EDTA для получения клеточной суспензии и выделяли клетки центрифугированием. Готовили клеточную суспензию в 5×106 клеток/мл в буфере для FACS (PBS, содержащий 2% FBS, 0,05% азида натрия) и в каждую пробирку вносили по 100 мкл суспензии клеток. В каждую пробирку добавляли по 100 мкл клонов при 2 мкг/мл и выдерживали при 4°C в течение 30 мин, индуцируя связывание с клетками. Клетки отмывали 2 мл буфера для FACS, добавляли 100 мкл разведения второго антитела (1/500, конъюгированный с PE Fc-фрагмент поликлонального антитела против IgG человека, #A80-248PE, Bethyl Laboratories Inc., США) и полученную смесь инкубировали при 4°C в течение 20 мин для введения флуоресцентной метки. Клетки отмывали 2 мл буфера FACS, а затем суспендировали в 200 мкл буфера FACS. В качестве положительного контроля на TIE2 использовали помеченное флуоресцентной меткой PE антитело против CD202b (Tie-2, CD202) мыши (124007, Biolegend), антитело с PE против CD202b (Tie2/Tek) человека (334205, Biolegend), конъюгированное с PE поликлональное антитело против IgG мыши (A90-139PE, Bethyl Laboratories Inc.) и конъюгированное с PE поликлональное антитело против IgG человека (A80-248PE, Bethyl Laboratories Inc.). Проводили анализ на установке FACSCalibur фирмы BD Bioscience.
Как показывают результаты исследований, все клоны обладают превосходным связыванием с TIE2 человека, который экспрессирован в клетках линии CHO-K1, а клон 4E2 обладает способностью к связыванию с TIE2 как человека, так и мыши (фиг.1A и 1B).
Пример 4. Экспрессия антител против TIE2
Проводили конверсию scFv, экспрессированных в прошедших скрининг фагах, в молекулы IgG методами молекулярной биологии. Выделяли фагемиды из прошедших скрининг клонов E.coli и амплифицировали вариабельные области методом ПЦР. Амплифицированные вариабельные области тяжелой цепи вставляли в экспрессирующий вектор (Invivogen, pfusess-hchg1), содержащий константную область тяжелой цепи, а амплифицированные вариабельные области легкой цепи вставляли в экспрессирующий вектор (Invivogen, pfuse2ss-hclk), содержащий константную область легкой цепи, завершая клонирование ДНК в форме IgG.
Краткосрочную экспрессию IgG осуществляли с помощью набора для системы экспрессии Expi293F (Thermo Fisher Scientific, США). Клетки Expi293, входящие в набор, культивировали в суспензии на круговой качалке при 125 об/мин при 37°C и 5% CO2 в специальной среде. Каждые 3 дня клетки пассировали до количества 3×105 клеток/мл, а при введении экспрессирующего вектора количество клеток доводили до 3×106 клеток/мл перед использованием. Введение генов проводили с помощью специального реагента ExpiFectamine, причем готовили комплекс липид-ДНК, содержащий 1 мкг ДНК экспрессирующего вектора и 2,7 мкл ExpiFectamine на 1 мл суспензии клеток, который добавляли в суспензию клеток. Через 16-18 часов после введения добавляли 1/2 усилителя для запуска экспрессии. Затем клетки культивировали в тех же условиях, что описаны выше, в течение 3-4 дней и центрифугировали, получая супернатанты, содержащие IgG.
Пример 5. Очистка антител против TIE2
Полученные супернатанты наносили на колонки с протеином A (GE Healthcare) и очищали IgG методом аффинной хроматографии. После уравновешивания колонки смесью 20 мМ Трис-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,0) наносили супернатант, промывали раствором, содержащим 50 мМ Трис-HCl, 500 мМ NaCl, 5 мМ EDTA и 0,2% полисорбата 20 (pH 7,0), элюировали раствором, содержащим 50 мМ NaCl и 0,1 М глицин-HCl (pH 3,5), и нейтрализировали 1 М раствором Триса. Элюированный белок диализовали через мембрану Spectra/Por с отсечением по молекулярной массе 10000 (Spectrum Labs, США), заменяя растворитель на PBS. Затем белок концентрировали до требуемой концентрации с помощью Vivaspin (Sartorius, DE), разливали и хранили при -80°C.
После очистки каждое антитело обрабатывали невосстанавливающим и восстанавливающим LDS буфером для образцов (Thermo Fisher Scientific) и подвергали электрофорезу на установке NuPAGE (Thermo Fisher Scientific). В результате получали IgG с общей молекулярной массой около 150 кДа, содержащие тяжелую цепь 50 кДа и легкую цепь 25 кДа.
Пример 6. Определение перекрестной реактивности антитела против TIE2 к VISTA человека и мыши
Определяли методом ELISA, сохраняется ли у IgG способность scFv фаговых клонов связываться с TIE2. В лунки 96-луночного планшета вносили по 100 мкл на лунку антигена в концентрации 1 мкг/мл (hTIE2-his: Sino Biological, 10700-H08H; mTIE2-his: Sino Biological, 51087-M08H) и инкубировали при 4°C в течение ночи. На следующий день выращенные накануне клетки центрифугировали при 3000 об/мин в течение 10 мин и хранили при 4°C. Покрытый антигеном планшет промывали 3 раза 0,1% TBST и в каждую лунку добавляли по 200 мкл блокирующего буфера с 2% BSA, а затем инкубировали при 25°C в течение 2 часов. По завершении блокирования планшет промывали 3 раза 0,1% TBST. В каждой лунке смешивали 50 мкл 4% BSA с 50 мкл фага, хранившегося при пониженной температуре 4°C, и инкубировали для протекания реакции со встряхиванием при комнатной температуре в течение 1 часа. После связывания фага продукт реакции трижды промывали 0,1% TBST, в каждую лунку добавляли по 100 мкл конъюгированного с HRP антитела козы к антителу человека (1:3000, Sino, 11973-MM05) и инкубировали при 25°C в течение 1 часа. После завершения реакции полученный продукт 3 раза промывали 0,1% TBST, добавляли в каждую лунку по 100 мкл TMB (из набора реагентов для субстрата TMB #BD 555214), выявляли окрашивание в течение 3-5 мин и добавляли 50 мкл стоп-раствора, а затем проводили анализ на считывающем устройстве для ELISA.
В результате видно, что, аналогично scFv, клоны 1A6, 1A9, 2F2, 4E2 и 3D6 связывались с TIE2 человека, в частности, клон 4E2 связывался как с TIE2 мыши, так и с TIE2 человека (фиг.2).
Пример 7. Измерение миграционной активности антитела против TIE2
Для того, чтобы выявить, действующий как наилучший стабилизатор сосудов среди антител против TIE2, проводили скрининг по миграционным характеристикам эндотелиальных клеток сосудов, которые ответственны за раннюю стадию механизма нормализации сосудов. Соответственно, проводили анализ миграции при использовании клеток HUVEC (эндотелиальные клетки пупочной вены человека).
24-луночную поликарбонатную вставку Transwell (размер пор 8,0 мкм, Corning) покрывали 0,1% раствором желатина и высушивали при комнатной температуре в течение примерно 10 мин. Каждый препарат разводили в 600 мкл основной среды и вносили в нижнюю камеру. Затем в нижнюю камеру, содержащую среду, помещали вставочную камеру и инокулировали в нее клетки HUVEC из расчета 1×105 клеток/100 мкл. После инкубации при 37°C в течение 4 часов вставочную камеру трижды промывали дистиллированной водой и окрашивали клетки, прикрепившиеся к вставочной камере, 0,5%-м кристаллическим фиолетовым при комнатной температуре в течение 10 мин. Затем немигрировавшие клетки внутри вставки смывали ватным тампоном, а клетки, мигрировавшие из вставочной камеры наружу, визуализировали при увеличении в 200 раз с помощью оптического микроскопа, а затем проводили количественную оценку с помощью программы Image-J (NIH).
Результаты исследования показывают, что положительный контроль, т.е. группа, полученная при кластеризации 500 нг/мл ANG1 (R&D System, 923-AN-025), проявляла лучшую миграцию, чем некластеризованная группа, а отрицательный контроль, т.е. 500 нг/мл ANG2 (R&D System, 623-AN-025), не проявлял миграции вне зависимости от кластеризации. В частности, видно, что среди опытных групп клоны 4E2 и 2F2 при 30 мкг/мл проявляли усиление миграции. Клоны 1A6, 1A9 и 11F проявляли базовый уровень миграции, а 3D6 проявлял снижение миграции (фиг.3A).
Для того, чтобы установить концентрационную зависимость активности клонов 4E2 и 2F2 и синергический эффект обработки в комбинации с ANG1 на основе результатов исследования миграции, измеряли миграционную активность вышеприведенным способом. Результаты показали, что клоны 4E2 и 2F2 обнаруживают зависимую от концентрации активность и проявляют синергический эффект при обработке в комбинации с ANG1 (фиг.3B и 3C). Таким образом, клоны 4E2 и 2F2 обладают синергическим эффектом в комбинации с ANG1 и быстро вызывают миграцию, происходящую на ранней стадии нормализации сосудов, что свидетельствует об отличной стабилизации сосудов.
Пример 8. Сравнение характеристик связывания между антителами против TIE2 и конкурирующими веществами (ANG1 и ANG2)
Определяли, будут ли 2F2 и 4E2, то есть антитела, отобранные в качестве стабилизаторов сосудов при анализе миграции, конкурировать с ANG1 и ANG2 за связывание с TIE2 и выполнение своих функций, конкурентным методом ELISA. В лунки 96-луночного иммунологического планшета вносили 1 мкг/мл hTIE2 (Sino Biological, 10700-H08H) из расчета 100 мкл на лунку и инкубировали при 4°C в течение ночи. Покрытый hTIE2 планшет два раза промывали PBS, а затем в каждую лунку добавляли по 200 мкл 2% BSA (блокирующего буфера) и инкубировали при комнатной температуре в течение 2 часов. После блокирования клетки дважды промывали 0,1% PBST. Проводили связывание антител-кандидатов против TIE2 и белков ANG1 и ANG2 с TIE2 в 10-кратных серийных разведениях при концентрации 2 мкг/мл и наивысшей концентрации 50 мкг/мл, соответственно. После трехкратного промывания PBST в лунки добавляли по 100 мкл конъюгированного с HRP антитела козы к антителу человека (1:3000, Sino, 11973-MM05), разведенного в отношении 1:2000, и проводили реакцию при комнатной температуре в течение 1 часа для индукции связывания, а полученный продукт 3 раза промывали TBST и выявляли окрашивание с помощью субстратного реагента TBM. Проявление окраски останавливали добавлением 50 мкл 2N H2SO4 и измеряли специфическое поглощение OD450−630 на считывающем устройстве для микропланшетов Sunrise (Tecan, CH).
Как видно из фиг.2, отобранные антитела (2F2 и 4E2) связываются с TIE2, не конкурируя с ANG1 и ANG2 человека (фиг.4).
Пример 9. Оценка связывающей аффинности антител против TIE2
Анализировали аффинность отобранных выбранных антител против TIE2, а именно 4E2, 2F2 и 11F, к TIE2-his человека (Sino Biological, 10700-H08H) и Tie2-his мыши (Sino Biological, 51087-M08H), на установке Octet (ForteBio Inc., США). Антитела против TIE2 фиксировали на биосенсоре и тестировали согласно руководству производителя. Измеряли кинетику связывания при каждой концентрации TIE2 человека и TIE2 мыши и рассчитывали константы скорости связывания (kon), константы скорости диссоциации (koff) и константы связывания (KD) (табл.4).
Было обнаружено, что антитело 11F обладает перекрестной реактивностью между TIE2 человека и TIE22 мыши, а результаты ELISA показали, что антитело 11F обладает очень слабой аффинностью (KD=2,09×10−5 M) к TIE2 мыши. Точно так же и 4E2 связывается с TIE2 и человека, и мыши и обладает высокой аффинностью к TIE2 человека, которое соответствует значению KD=1,0×10−12 M, а также аффинностью к TIE2 мыши (1,38×10−8 M). 2F2 проявляет высокую специфическую аффинность только к TIE2 человека, которая соответствует значению KD=7,94×10−10 M.
Таблица 4
Пример 10. Измерение активности по возмещению сосудистой утечки антитела против TIE2
Для того, чтобы определить активность антитела против TIE2 в качестве стабилизаторов сосудов, проводили тест на возмещение сосудистой утечки на эндотелиальных клетках сосудов. Ангиогенез вызывает разрыв контактов в сосудах вследствие гиперэкспрессии VEGF, что приводит к сосудистой утечке. Для проведения этого теста in vitro осуществляли анализ проницаемости сосудов на клетках линии HUVEC.
24-луночную поликарбонатную вставку Transwell (размер пор 0,4 мкм, Merck Millipore) покрывали 0,1% раствором желатина (Sigma Aldrich, G1890), а затем высушивали при комнатной температуре в течение примерно 10 мин. Затем в нижнюю камеру, содержащую среду, помещали вставочную камеру и инокулировали в нее клетки HUVEC из расчета 6×104 клеток/200 мкл. Затем клетки культивировали в течение 72 часов без замены среды в термостате с 5% CO2 при 37°C. После удаления среды из вставочной камеры обрабатывали основную среду каждым препаратом. Клетки инкубировали при 37°C в течение 30 минут, добавляли VEGF в концентрации 100 нг/мл (R&D System, 293-VE-050) и инкубировали 150 минут.Затем во вставочную камеру добавляли 0,5 мг/мл FITC-декстрана (70 кДа, Sigma) и проводили реакцию при комнатной температуре в течение 30 мин. Собирали 50 мкл FITC-декстрана, просочившегося в нижнюю камеру, и разбавляли 1:20 в PBS. Затем в разведенных образцах измеряли флуоресценцию при длине волны 492 нм/520 нм (возбуждение/испускание) на считывающем устройстве для микропланшетов (Hidex Chameleon).
Результаты исследования показывают, что в группе, получавшей только VEGF, обнаруживалось повышение количества просочившегося FITC-декстрана при разрыве плотных контактов в эндотелиальных клетках сосудов. Однако видно, что количество такого FITC-декстрана снижалось при обработке ANG1, положительным контролем. С другой стороны, ANG2, отрицательный контроль, обнаруживал проницаемость, близкую к таковой в группе, получавшей лишь VEGF, а в группах, получавших 2F2 и 4E2, отмечалось снижение проницаемости. Кроме того, проверяли активность 11F тем же экспериментальным способом, что и выше. Результат показал, что 11F также снижает проницаемость сосудов (фиг.5A и 5B).
Вышеприведенные результаты свидетельствуют о том, что антитело против TIE2 обладает активностью по ослаблению сосудистой утечки, вызванной VEGF. Кроме того, определяли, способно ли антитело против TIE2 возмещать сосудистую утечку даже в условиях культивирования, аналогичных условиям модели заболевания в группе, получавшей и VEGF, и ANG2.
Результаты показывают, что 2F2 снижает проницаемость сосудов до базового уровня, а 4E2 также снижает проницаемость сосудов до базового уровня. Из результатов теста на проницаемость сосудов следует, что клоны 2F2 и 4E2 могут нормализировать сосудистую утечку, вызванную VEGF и ANG2 (фиг.5C).
Пример 11. Исследование механизма нормализации сосудов антител против TIE2
Исследовали механизм понижающей регуляции сигнала в отношении TIE2, чтобы определить, реализуется ли эффект уменьшения сосудистой утечки у отобранных антител против TIE2 по сигнальному механизму нормализации сосудов. Для этого обрабатывали препаратами клетки HUVEC, а затем определяли экспрессию белков, связанных с нормализацией сосудов, методом вестерн-блоттинга.
Для вестерн-блоттинга каждую из линий клеток HUVEC, подвергавшихся взаимодействию с препаратами, дважды промывали PBS и затем инкубировали в буфере для лизиса (Thermo Scientific, буфер RIPA с ингибиторами фосфатаз) на льду в течение 20 мин, получая клеточные экстракты. Затем клетки центрифугировали при 13000 об/мин и 4°C и собирали только супернатанты. После определения белка с помощью BCA наносили лизаты на 10% гель SDS-PAGE и проводили электрофорез. Переносили белки на нитроцеллюлозную мембрану (Bio-Rad) и блокировали 5% обезжиренным молоком в буфере TBST с 0,1% Твин 20 при комнатной температуре в течение 1 часа. Затем разводили 1:1000 первые антитела (p-TIE2, TIE2, p-AKT, AKT, p-ERK, ERK, p-VEGF, p-VE-кадгерин, VE-кадгерин и гистоны; клеточный сигналлинг) и встряхивали в течение ночи при 4°C, а затем проводили реакцию со вторым конъюгированным с пероксидазой антителом (Santa Cruz), разведенным 1:5000, при комнатной температуре в течение 1 часа. Проводили реакцию с раствором ECL (Pierce) и обнаруживали требуемые полосы белков. Важно, что в каждый момент времени наблюдался сигнальный механизм TIE2. Результаты исследования показывают, что антитело 2F2 против TIE2 вызывает фосфорилирование TIE2, подобно ANG1, а также фосфорилирование AKT, сигнала нормализации сосудов. Антитела 4E2 и 11F против TIE2 давали аналогичные результаты. Соответственно, антитела 2F2, 4E2 и 11F, как оказалось, обладают способностью действовать в качестве средств, нормализующих сосуды посредством фосфорилирования TIE2 (фиг.6).
Таким образом, чтобы определить, возмещает ли антитело против TIE2 сосудистую утечку посредством стабилизирующих сосуды сигналов даже в условиях сосудистой утечки, среду, содержащую VEGF, обрабатывали антителом 2F2 против TIE2 и измеряли сигналы фосфорилирования VEGFR и VE-кадгерина, которые отвечают за восстановление плотных контактов в эндотелиальных клетках сосудов, таким же методом вестерн-блоттинга, как и выше (фиг.7).
Результаты исследования показывают, что фосфорилирование VEGFR повышается, и фосфорилирование VE-кадгерина, который составляет межклеточные плотные контакты, повышается в группе VEGF, ответственного за ангиогенез. С другой стороны, ANG1, положительный контроль, ингибировал фосфорилирование VEGFR и VE-кадгерина. Точно так же и антитело 2F2 тоже ингибировало фосфорилирование VEGFR и VE-кадгерина. В этом сигнальном механизме антитело против TIE2 ингибировало активацию VEGFR посредством фосфорилирования TIE2, а ингибированный VEGFR ингибировал фосфорилирование VE-кадгерина, ответственного за межклеточные плотные контакты в клеточной мембране, что приводило к восстановлению межклеточных плотных контактов. Результаты исследования свидетельствуют, что антитело против TIE2 может действовать в качестве стабилизатора сосудов, выполняющего функции возмещения сосудистой утечки и передачи сигналов.
Пример 12. Поддержание межклеточных плотных контактов антителом против TIE2
Чтобы определить, влияют ли антитела против TIE2 на нормализацию сосудов, проводили иммуноцитохимический тест на клетках линии HUVEC. Клетки линии HUVEC высевали по 8×105 клеток на лунку на 4-луночное предметное стекло для ICC. Через 24 часа вносили 100 нг/мл VEGF, ANG1, ANG2, 10 мкг/мл антител против TIE2 (2F2, 4E2) и инкубировали в течение 24 часов. После фиксации в 4% PFA в течение 15 мин пермеабилизировали клеточные мембраны с помощью Тритона X-100 и блокировали 10% козьей сывороткой. После отмывания в PBS проводили связывание с конъюгированным с Alexa-488 антителом против VE-кадгерина (Thermo) в разведении 1:200 в течение ночи и инкубировали с 1 нг/мл DAPI в течение 10 мин. После отмывания PBS препараты фиксировали заливочным раствором и визуализировали под флуоресцентным микроскопом.
Наблюдалась экспрессия VE-кадгерина. Результаты исследования показывают, что экспрессия VE-кадгерина, молекулы межклеточных плотных контактов, снижалась в группе, обработанной и VEGF, и ANG2, причем контакты восстанавливались после обработки ANG1, положительным контролем. После обработки антителом против TIE2 (2F2) экспрессия VE-кадгерина значительно повышалась, как и в случае положительного контроля. При обработке в комбинации с ANG2 экспрессия VE-кадгерина также повышалась. Другое антитело против TIE2 (4E2) также обнаруживало заметное повышение экспрессии VE-кадгерина, как и с положительным контролем, причем 4E2 при обработке в комбинации с ANG2 также повышало экспрессию VE-кадгерина со значением, близким к таковому в группе, обработанной только препаратом (фиг.8). Результаты исследования свидетельствуют, что антитела против TIE2 (2F2, 4E2) усиливают экспрессию VE-кадгерина, приводя к нормализации сосудов.
Пример 13. Тест на нормализацию сосудов in vivo под действием антитела против TIE2
Чтобы определить, проявляется ли функция нормализации сосудов у антитела против TIE2 in vivo, определяли активность антитела против TIE2, используя антитело 4E2, которое обладает способностью к связыванию с TIE2 мыши на модели индуцированной VEGF сосудистой утечки в ушах.
В хвостовую вену 7-недельных самцов мышей Balb/c вводили 1% раствор красителя Evans blue и выжидали 10 минут, пока кровь не окрасится в синий цвет.Затем в кровеносные сосуды правого уха мышей в опытной группе вводили 10 мг/кг 4E2, а в кровеносные сосуды левого уха вводили PBS. Через 30 минут в кровеносные сосуды обоих ушей вводили VEGF (R&D System, 293-VE-050). В контрольной группе вводили PBS в кровеносные сосуды левого уха и VEGF в сосуды правого уха. Через 30 минут уши отрезали и экстрагировали краситель Evans blue этанолом в течение 24 часов. Затем измеряли значения OD на считывающем устройстве для микропланшетов.
В результате отмечалась утечка красителя Evans blue под действием VEGF в группе отрицательного контроля, которая примерно в два раза превышала уровень утечки по сравнению с группой PBS. Среди опытных групп в группе, обработанной 4E2, утечка красителя Evans blue снижалась примерно на 30% по сравнению с группой VEGF. Таким образом, из результатов этого эксперимента следует, что антитело против TIE2 способно в достаточной степени нормализировать кровеносные сосуды, проявляющие сосудистую утечку, вызванную VEGF, за счет передачи сигналов посредством TIE2 (фиг.9).
Пример 14. Оценка действия антитела против TIE2 по ингибированию ангиогенеза в сетчатке
Чтобы определить, обладает ли антитело против TIE2 ингибирующим действием на ангиогенез, проводили тест на его эффективность на модели индуцированной лазером хориоидальной неоваскуляризации (CNV) у мышей. Эффективность действия антитела тестировали в сравнении с коммерческим препаратом афлиберцепт в качестве контроля. После общей анестезии мышей кетамином им в глаза по каплям добавляли анестезирующие капли, чтобы вызвать дополнительную местную анестезию, а также по каплям добавляли мидриатическое средство, чтобы вызвать расширение зрачков. Мышей помещали на операционный стол и разрушали оболочку Бруха, индуцируя лазерный ожог в соответствии с условиями индукции CNV (длина волны 532 нм, диаметр 50 мкм, продолжительность 80 мс, уровень мощности 200 мВт) с помощью Micron-IV. Очаги, в которых не наблюдались пузырьки во время индукции лазерного ожога, классифицировали как неудачные лазерные ожоги и исключали из процесса анализа результатов и статистической обработки на основании критериев исключения по модифицированным критериям, предложенным Gong Y. et al. (Plos One, 1:15, 2015).
Через 24 часа после индукции CNV вводили антитело против TIE2 (20, 40 мкг/глаз) или контрольный препарат афлиберцепт (20 мкг/глаз). Для того, чтобы обнаружить эффект ингибирования ангиогенеза, мышей анестезировали кетамином через 10 дней после индукции CNV, а затем внутрибрюшинно вводили флуоресцентное контрастное вещество. В глаза по каплям вводили анестезирующие капли, чтобы вызвать дополнительную местную анестезию, а также по каплям добавляли мидриатическое средство, чтобы вызвать расширение зрачков. Мышей помещали на операционный стол, фокусировали изображение на глазном дне с помощью камеры для визуализации Micron-IV, в глаза по каплям добавляли смазывающий гель и приводили линзу OCT в контакт с роговицей глаза. Осуществляли запись изображений FFA/OCT, а затем в глаза мышей вводили глазные капли с антибиотиком. Анализ изображений FFA и OCT проводили с помощью программы Image-J. Однако очаги CNV, соответствующие критериям исключения, предложенным Gong Y. et al., исключали из окончательных результатов и процесса статистического анализа.
Чтобы определить эффект восстановления зрительного нерва, проводили электроретинографию (ЭРГ). У мышей индуцировали адаптацию к темноте в темной комнате за 12 часов до проведения ЭРГ. В день проведения ЭРГ (через 11 дней после индукции CNV) мышей подвергали общей анестезии с помощью Rompun® и кетамина и в глаза по каплям вводили Alcaine®, чтобы вызвать дополнительную местную анестезию, а также по каплям добавляли мидриатическое средство, чтобы вызвать расширение зрачков. Мышей помещали на столик для ЭРГ и ставили зонды для ЭРГ на хвост, голову и роговицу. ЭРГ определяли по изменению потенциала сетчатки после однократной импульсной стимуляции (0,9 log cds/m2 (10 ответов/интенсивность)). По завершении ЭРГ в глаза мышей вводили каплю Tobrex. Анализ ЭРграмм проводили с помощью программы LabScribeERG (iWorx DataAcquisition Software). Однако глаза, удовлетворяющие критериям исключения, предложенным Gong Y. et al., исключали из окончательных результатов и процесса статистического анализа.
Результаты оценки действия антитела против TIE2 по снижению ангиогенеза на модели CNV у мышей представлены на фиг.10. Все опытные группы, получавшие антитело против TIE2, проявляли статистически значимое снижение размера очагов CNV по данным FFA по сравнению с CNV в контрольной группе (p<0,0001 для каждой группы) и проявляли гораздо большее снижение даже по сравнению с опытной группой, получавшей афлиберцепт.Как видно из фиг.10B, все опытные группы, получавшие антитело против TIE2, проявляли статистически значимое уменьшение объема очагов по данным OCT по сравнению с CNV в контрольной группе (p<0,0001, p<0,001 для каждой группы). Кроме того, при сравнении потенциала сетчатки по скотопической ЭРГ с контрольной группой CNV на фиг.10C наблюдалось статистически значимое увеличение амплитуды волны B, сравнимое с группой, получавшей афлиберцепт (p<0,0001). Результаты свидетельствуют, что антитело против TIE2 на модели CNV у мышей обладает ингибирующим действием на ангиогенез и сосудистую утечку.
Пример 15. Оценка действия антитела против TIE2 на нормализацию раковых кровеносных сосудов
Для оценки действия антитела против TIE2 на нормализацию раковых сосудов проводили тестирование на модели спонтанной глиобластомы у мышей, используя антитело DC101 против VEGFR в качестве контрольного препарата. Степень индукции опухолей при спонтанной глиобластоме измеряли по визуализации IVIS у 6-7-недельных мышей EGFRviii/viii Luciferasefl/+tdTomatofl/+один раз в неделю. Образование опухолей определяли, как случай, когда яркость сигнала IVIS составляла 3,0×104 фотонов/сек/см2/ср или больше и в головном мозге мышей выявлялся локально концентрированный сигнал. Мышей с опухолями группировали и проводили визуализацию IVIS три раза, в частности, один раз перед введением препарата, один раз во время введения и один раз по окончании введения, каждый раз регистрируя яркость сигнала при визуализации. Препараты антитела против TIE2 (1 мг на мышь) и антитела против VEGFR (1 мг на мышь) вводили внутривенно в общей сложности 4 дозы по два раза в неделю на протяжении 2 недель. Перед введением препаратов мышей тщательно анестезировали путем респираторной анестезии изофлураном. Мышей также тщательно анестезировали путем внутрибрюшинной инъекции смешанного раствора, содержащего 52,5 мкл кетамина, 22,5 мкл Rompun и 75 мкл PBS. Через 10 минут мышей опять анестезировали посредством внутривенной инъекции 200 мкл лектина-649 в ретроорбитальную вену, а затем извлекали мозг.В извлеченном мозге мышей определяли изменения в структуре сосудов и покрытии сосудистых перицитов путем окрашивания на CD31 и PDGFRβ.
По результатам наблюдения сигналов PECAM (CD31), сосредоточенных на канцерогенном участке, где сильно экспрессируется Td-tomato, такие характеристики внешнего вида раковых сосудов, как увеличение диаметра, изогнутая форма и заметное уменьшение ветвей, были ослаблены в группе, получавшей 4E2, по сравнению с группами, получавшими IgG человека и DC101 (фиг.11A). Кроме того, по результатам наблюдения сигналов PDGFRβ оказалось, что большая часть белка PDGFRβ, маркера сосудистых адгезированных клеток, хорошо прикреплялась к участку, окрашенному PECAM, в группе, получавшей антитело против TIE2. Это означает, что антитело против TIE2 обладает свойством нормализации внешнего вида раковых сосудов, ингибируя аномальную пролиферацию эндотелиальных клеток сосудов и улучшая адгезию сосудистых перицитов к эндотелиальным клеткам сосудов (фиг.11B).
Пример 16. Оценка противоопухолевого действия антитела против TIE2
Чтобы определить противоопухолевое действие и эффект усиления внутриопухолевой миграции иммунных клеток за счет нормализации раковых сосудов антителом против TIE2, мышам BALB/c трансплантировали клетки раковой линии CT26 толстой кишки мыши и вводили только антитело против TIE2 или это антитело в комбинации с антителом против PD-1 мыши. У мышей измеряли длину, ширину и толщину трансплантатов колоректальных раковых клеток линии CT26 с помощью штангенциркуля. В то же самое время опухоли взвешивали. Размер опухолей определяли как (длина × ширина × толщина)/2, и мышей с опухолями группировали на основе среднего значения 27,5 мм3. Затем мышам вводили препараты антитела против TIE2 (30 мг/кг) или против PD-1 мыши (10 мг/кг) как сами по себе, так и в комбинации, один раз в неделю и два раза в неделю, соответственно. На 10-й день после введения препаратов всех мышей умерщвляли с помощью газообразного СО2, проводили лапаротомию, проверяли органы визуально на наличие аномалий невооруженным глазом и извлекали опухоли. Каждую извлеченную опухоль взвешивали на химических весах, делали снимки и измеряли иммунные клетки типа Т-клеток CD4+, Т-клеток CD8+и MDSC в опухолях методом проточной цитометрии. Опухоли фиксировали в формалине и проводили окрашивание Т-клеток CD8+и MDSC методами иммуногистохимии.
В результате в группе, получавшей только антитело против TIE2 (10 мг/кг), проявлялся эффект подавления опухолей примерно на 40%, причем снижение было значимым по сравнению с группой, получавшей антитело против PD-1 мыши (10 мг/кг) (фиг.12A и 12B). В частности, совместное введение антитела против TIE2 и антитела против PD-1 мыши вызывало эффект подавления опухолей на величину примерно на 64% (фиг.12A и 12B). При измерении количества иммунных клеток, мигрировавших в опухоли, по данным проточной цитометрии и иммуногистохимии, в группе, получавшей антитело против TIE2, отмечалось повышение примерно в 3 раза содержания Т-клеток CD8+, известных как цитотоксические клетки, по сравнению с группой отрицательного контроля. Даже при совместном введении с антителом к PD-1 проявлялся эквивалентный уровень миграции (фиг.12C и 12D). Таким образом, антитело против TIE2 проявляет противоопухолевый эффект за счет усиления миграции иммунных клеток в опухоли за счет действия, нормализирующего раковые сосуды.
Пример 17. Оценка уменьшения гипоксии под действием антитела против TIE2
Для того, чтобы определить противоопухолевое действие и ослабление внутриопухолевой гипоксии под действием антитела против TIE2 посредством нормализации раковых сосудов, препарат антитела вводили мышам BALB/c, которым были трансплантированы клетки раковой линии CT26 толстой кишки мыши. У мышей измеряли длину, ширину и толщину трансплантатов колоректальных раковых клеток линии CT26 с помощью штангенциркуля. В то же самое время опухоли взвешивали. Размер опухолей определяли, как (длина × ширина × толщина)/2, и мышей с опухолями группировали на основе среднего значения 24,6 мм3. Затем мышам вводили антитело против TIE2 (10 мг/кг) или антитело DC101 против VEGFR (20 мг/кг, контрольный препарат) один раз в неделю или два раза в неделю, соответственно. На 10-й день после введения препаратов двум животным в каждой группе внутривенно (IV) вводили пимонидазол. Через 90 минут всех мышей умерщвляли с помощью газообразного СО2, проводили лапаротомию, проверяли органы на наличие аномалий невооруженным глазом и извлекали опухоли. Каждую извлеченную опухоль взвешивали на химических весах, делали снимки и определяли лимфоциты (Т-клетки CD4+, Т-клетки CD8+) в качестве иммунных клеток в опухолях методом проточной цитометрии. Опухоли со введением пимонидазола фиксировали в 4% PFA и проводили иммуногистохимическое окрашивание с помощью набора Hypoxyprobe.
В группах, получавших антитело против TIE2 или контрольный препарат DC101, обнаруживалось сходное подавление опухолей примерно на 30% (фиг.13A, p<0,01, p<0,001). В группе, получавшей антитело против TIE2, отмечалось примерно 2-кратное повышение количества лимфоцитов, то есть внутриопухолевых иммунных клеток, по сравнению с контрольной группой (фиг.13B, p<0,01). Контрольный препарат, а именно антитело против VEGFR2, не вызывал внутриопухолевую миграцию иммунных клеток, так же, как это наблюдалось в группе отрицательного контроля. Кроме того, результаты по оценке снижения гипоксии методом окрашивания с помощью Hypoxyprobe показали, что интенсивность флуоресцентной окраски в группе, получавшей антитело против TIE2, снижалась примерно в 4 раза по сравнению с группой отрицательного контроля (фиг.13C, p<0,01) и примерно в 2,5 раза по сравнению с группой, получавшей контрольный препарат, антитело против VEGFR (фиг.13C, p<0,01). Таким образом, антитело против TIE2 усиливает миграцию иммунных клеток в опухоли и уменьшает гипоксию за счет действия, нормализующего раковые сосуды.
Промышленная применимость
Антитело против TIE2 либо его антигенсвязывающий фрагмент по настоящему изобретению проявляет желательную способность к связыванию с TIE2 и может применяться для профилактики или лечения раковой/опухолевой мишени, в качестве стабилизатора сосудов и для профилактики или лечения заболеваний, связанных с ангиогенезом. За счет разработки терапевтических средств, мишени которых отличаются от мишеней для стандартных терапевтических средств, нацеленных на TIE2, в соответствии с настоящим изобретением можно обеспечить комбинированную терапию со стандартными терапевтическими средствами и монотерапию для лечения опухолей и связанных с ангиогенезом заболеваний.
Хотя конкретные варианты настоящего изобретения и были описаны подробно, однако специалистам в данной области должно быть ясно, что настоящее описание приводится для изложения предпочтительных воплощений в целях иллюстрации и не должно рассматриваться как ограничивающее объем настоящего изобретения. Таким образом, объем настоящего изобретения, по существу, определяется прилагаемой формулой изобретения и ее эквивалентами.
--->
<130> 2428
<150> KR 19/099,491
<151> 2019-08-14
<160> 60
<170> PatentIn version 3.2
<210> 1
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR1
<400> 1
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 2
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR2
<400> 2
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 3
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR3
<400> 3
Ala Arg Gly Val Asp Ser Ser Met Val Thr Gly Phe Asp His
1 5 10
<210> 4
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR1
<400> 4
Gln Ser Ile Ser Arg Trp
1 5
<210> 5
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR2
<400> 5
Glu Ala Ser
1
<210> 6
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR3
<400> 6
Gln Gln Tyr Glu Asp Tyr Pro Leu Thr
1 5
<210> 7
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR1
<400> 7
Gly Tyr Thr Phe Asn Ser Tyr Asp
1 5
<210> 8
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR2
<400> 8
Val Asn Pro Pro Gly Gly Thr Gly Ser Thr
1 5 10
<210> 9
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR3
<400> 9
Ala Arg Asp Tyr Asn Arg Ala Pro Pro Thr Leu Asp Val
1 5 10
<210> 10
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR1
<400> 10
Ser Ser Ser Asp Val Gly Gly Tyr Asn Tyr
1 5 10
<210> 11
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR2
<400> 11
Asp Val Thr
1
<210> 12
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR3
<400> 12
Ser Ser Tyr Ser Ser Ser Thr Phe Tyr Val
1 5 10
<210> 13
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR1
<400> 13
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 14
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR2
<400> 14
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 15
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR3
<400> 15
Ala Arg Gly Gly Leu His His Gly Phe Asp Ile
1 5 10
<210> 16
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR1
<400> 16
Asn Ile Glu Ser Lys Ser
1 5
<210> 17
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR2
<400> 17
Tyr Asp Asn
1
<210> 18
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR3
<400> 18
Gln Val Trp Asp Thr Tyr Thr Asp Gln Pro Val
1 5 10
<210> 19
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR1
<400> 19
Gly Phe Thr Phe Ser Arg Tyr Asn
1 5
<210> 20
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR2
<400> 20
Ile Ser Ser Ser Gly Arg Phe Ile
1 5
<210> 21
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR3
<400> 21
Ala Arg Asp Ser Pro Thr Gln Gly Pro Tyr Tyr Tyr Tyr Gly Met Asp
1 5 10 15
Val
<210> 22
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR1
<400> 22
Lys Ile Gly Ser Lys Ser
1 5
<210> 23
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR2
<400> 23
Tyr Asp Ser
1
<210> 24
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR3
<400> 24
Gln Val Trp Asp Ser Ser Ser Asp Arg Pro Val
1 5 10
<210> 25
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR1
<400> 25
Gly Phe Thr Phe Ser Asp Tyr Ala
1 5
<210> 26
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR2
<400> 26
Ile Ser Phe Asp Gly Asn Asn Gln
1 5
<210> 27
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR3
<400> 27
Thr Thr Asp Thr Met Ser Gly Tyr Asp Trp Glu Asp Ala Phe Asp Ile
1 5 10 15
<210> 28
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR1
<400> 28
Gln Ser Ile Gly Arg Trp
1 5
<210> 29
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR2
<400> 29
Ala Ser Ser
1
<210> 30
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR3
<400> 30
Gln Gln Ser Tyr Ser Thr Pro Tyr Thr
1 5
<210> 31
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR1
<400> 31
Gly Phe Ser Phe Ser Asp His Ala
1 5
<210> 32
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR2
<400> 32
Val Trp Pro Asp Gly Ser Asn Lys
1 5
<210> 33
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy-chain CDR3
<400> 33
Ala Arg Asp Ser Gly Pro Ile Ser Arg Gly Asp Leu Thr Tyr
1 5 10
<210> 34
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR1
<400> 34
Gly Gly Ser Ile Ala Ser Asn Tyr
1 5
<210> 35
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR2
<400> 35
Glu Asp Asp
1
<210> 36
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Light-chain CDR3
<400> 36
Gln Ser Tyr Asp Asp Thr Asn Val Val
1 5
<210> 37
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> VH
<400> 37
caggtgcagc tggtggagtc cgggggaggc ttggttcagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagtctgag agccgaagac acggccgtgt attactgtgc gagaggcgtg 300
gattcttcta tggtaaccgg atttgatcac tggggccagg gaactttgat caccgtctcc 360
tca 363
<210> 38
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 38
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Val Asp Ser Ser Met Val Thr Gly Phe Asp His Trp Gly
100 105 110
Gln Gly Thr Leu Ile Thr Val Ser Ser
115 120
<210> 39
<211> 324
<212> DNA
<213> Artificial Sequence
<220>
<223> VL
<400> 39
gacatccaga tgacccagtc tccttccacc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gagtattagt aggtggttgg cctggtatca gcagaaacca 120
gggaaagccc ctaagttcct gatctatgag gcatctactt tagaaagtgg ggtcccatca 180
aggttcagcg gcagtggaac tgggacagaa ttcactctca ccatcagcag cctgcagcct 240
gatgattttg ctacttatta ctgtcaacag tatgaggact acccgctcac cttcggccaa 300
gggacacgac tggaaatcaa acgt 324
<210> 40
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Phe Leu Ile
35 40 45
Tyr Glu Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Thr Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Asp Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg
100 105
<210> 41
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> VH
<400> 41
cagatgcagc tggtacagtc tgaggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgtaagg catctggata cacctttaac agttacgata tacactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggagta gtcaacccac ctggtggaac cggaagcact 180
gtttacgcac agaagttcga ggacagactc accctgacca cggacatgtc cacaagcaca 240
gcctacatgg agctgagcag cctgagatct gaggacacgg ccgtgtatta ctgtgcaaga 300
gactataata gggccccgcc tactttggac gtctggggcc aagggaccac gatcaccgtc 360
tcctca 366
<210> 42
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 42
Gln Met Gln Leu Val Gln Ser Glu Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Asn Ser Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Val Asn Pro Pro Gly Gly Thr Gly Ser Thr Val Tyr Ala Gln
50 55 60
Lys Phe Glu Asp Arg Leu Thr Leu Thr Thr Asp Met Ser Thr Ser Thr
65 70 75 80
Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Tyr Asn Arg Ala Pro Pro Thr Leu Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Ile Thr Val Ser Ser
115 120
<210> 43
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> VL
<400> 43
aattttatgc tgactcagcc cgcctccgtg tctgggtccc ctggacagtc gatcaccatc 60
tcctgcactg gaagcagcag cgacgttggt ggttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccca actcatcatt tatgatgtca ctaagcggcc ctcaggggtt 180
tctaatcgct tctccggctc caagtctggc aactcggcct ccctgaccat ctctggactc 240
caggctgagg acgaggctga ttattactgc agctcataca gcagcagcac tttttacgtc 300
ttcggaactg ggaccaaggt caccgtccta ggt 333
<210> 44
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 44
Asn Phe Met Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Ser Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Gln Leu
35 40 45
Ile Ile Tyr Asp Val Thr Lys Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Ser Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ser Ser Ser
85 90 95
Thr Phe Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 45
<211> 354
<212> DNA
<213> Artificial Sequence
<220>
<223> VH
<400> 45
caggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgacctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcacgt atcagtggta gtggtgggag cacaaactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc aagagggggt 300
ctccatcatg gttttgatat ctggggccaa gggacaatgg tcaccgtctc ctca 354
<210> 46
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 46
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Ser Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Leu His His Gly Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 47
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> VL
<400> 47
tcctatgagc tgacacagcc accctcactg tcagtggccc cagggaagac ggccaggatt 60
acatgtgacg gggacaacat tgaaagtaaa agtgtccact ggtaccagca gaagccaggc 120
caggcccctg tgctagtcat ctattatgat aatgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccgga 240
gatgaggccg actattactg tcaggtgtgg gacacttata ccgatcagcc ggtattcggc 300
ggagggacca agctgaccgt cctaggt 327
<210> 48
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 48
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Leu Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Asp Gly Asp Asn Ile Glu Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Asn Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Tyr Thr Asp Gln
85 90 95
Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 49
<211> 372
<212> DNA
<213> Artificial Sequence
<220>
<223> VH
<400> 49
caggtgcagc tggtagagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtatag gctctggctt caccttcagt cgctataaca taaattgggt ccgccaggct 120
ccagggaagg gcctggagtg ggtctcatcc attagtagta gtggaaggtt catccactac 180
gcaggctcag tgaagggccg attcaccgtc tccagagaca acaccaagaa ctcagtgtct 240
ctacaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gagagactct 300
ccaacacagg gcccctacta ctactacggt atggacgtct ggggccaagg gaccacgatc 360
accgtctcct ca 372
<210> 50
<211> 124
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 50
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ile Gly Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Asn Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Gly Arg Phe Ile His Tyr Ala Gly Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Thr Lys Asn Ser Val Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Pro Thr Gln Gly Pro Tyr Tyr Tyr Tyr Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Thr Ile Thr Val Ser Ser
115 120
<210> 51
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> VL
<400> 51
tcctatgagc tgacacagcc accctcagag tcagtggccc caggaaagac ggccacaatt 60
acttgtgggg gaaataaaat tggaagtaaa agtgtgcact ggtaccaaca gaagccaggc 120
caggcccctc taatggtcat ctattatgat agcgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaggtcggg 240
gatgaggccg actattactg tcaggtgtgg gatagtagta gcgatcgtcc ggtgttcggc 300
ggagggacca agctgaccgt cctaggt 327
<210> 52
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 52
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Glu Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Thr Ile Thr Cys Gly Gly Asn Lys Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Leu Met Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Val Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp Arg
85 90 95
Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 53
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> VH
<400> 53
caggtgcagc tggtgcagtc tggaggaggc ttgatccagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt gactatgcta tgcactgggt ccgccaggct 120
cctggcaagg ggctggagtg ggtggcactc atatcttttg atgggaataa tcaatactac 180
gcagactccg tgaagggccg attcaccatc tccagagaca attcgaagaa cacaatatat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt actactgtac cacagatacg 300
atgagtggct acgattggga agatgctttt gatatctggg gccaagggac aatgatcacc 360
gtctcctca 369
<210> 54
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 54
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Ser Phe Asp Gly Asn Asn Gln Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Ile Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Thr Asp Thr Met Ser Gly Tyr Asp Trp Glu Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Ile Thr Val Ser Ser
115 120
<210> 55
<211> 342
<212> DNA
<213> Artificial Sequence
<220>
<223> VL
<400> 55
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctatcggcga cagagtcacc 60
atcacttgcc gggccagtca gagtattggt aggtggttgg cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag agttacagta ccccgtacac ttttggccaa 300
gggaccaagg tggagatcaa acgtggagga gccagcctcg tg 342
<210> 56
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 56
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Ile Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Arg Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105
<210> 57
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> VH
<400> 57
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ccgggaggtc ccttagactc 60
tcctgttcag cgtctggatt ctccttcagt gatcatgcca tgcactgggt ccgccaggct 120
ccaggcaggg gcctagaatg ggtggcaact gtttggcctg atggaagtaa taaatattat 180
gtagattctg tgaacggtcg attcagcatt tccagagaca attccaagaa cacagtgtct 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc acgagattcg 300
ggacctatca gccgcggaga tttgacttac tggggcccgg gagtcctggt caccgtctcc 360
tca 363
<210> 58
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> VH
<400> 58
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Ser Asp His
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Glu Trp Val
35 40 45
Ala Thr Val Trp Pro Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Asn Gly Arg Phe Ser Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Gly Pro Ile Ser Arg Gly Asp Leu Thr Tyr Trp Gly
100 105 110
Pro Gly Val Leu Val Thr Val Ser Ser
115 120
<210> 59
<211> 351
<212> DNA
<213> Artificial Sequence
<220>
<223> VL
<400> 59
aattttatgc tgactcagcc ccactctgtg tcggagtctc cggggaagac tgtaatcatc 60
tcctgcaccg gcagcggtgg cagcattgcc agcaactatg tgcagtggta ccagcagcgc 120
ccgggcagtg cccccaccat tgtgatctat gaagatgatc agagaccctc tggggtccct 180
gatcggtact ctggctccac cgacagctcc tccaactctg cctccctcac catctctgga 240
ctgagggctg aggacgaggc tgactactac tgtcagtctt atgacgatac caatgtggtg 300
ttcggcggag ggaccaagct gaccgtccta ggtggaggag ccagcctcgt g 351
<210> 60
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> VL
<400> 60
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Ile Ile Ser Cys Thr Gly Ser Gly Gly Ser Ile Ala Ser Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ala Pro Thr Ile Val
35 40 45
Ile Tyr Glu Asp Asp Gln Arg Pro Ser Gly Val Pro Asp Arg Tyr Ser
50 55 60
Gly Ser Thr Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Arg Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Asp
85 90 95
Thr Asn Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| АНТИТЕЛО ПРОТИВ ANG2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2788088C1 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| АНТИТЕЛА ПРОТИВ VISTA И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2750723C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |
| АНТИ-DR5-АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2735956C1 |
| АНТИТЕЛА ПРОТИВ ПРЕДСТАВИТЕЛЯ А5 СЕМЕЙСТВА 19 СО СХОДСТВОМ ПОСЛЕДОВАТЕЛЬНОСТЕЙ И СПОСОБ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2785436C2 |
| АНТИТЕЛЬНЫЕ АГЕНТЫ, СПЕЦИФИЧНЫЕ К CD19 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773317C2 |

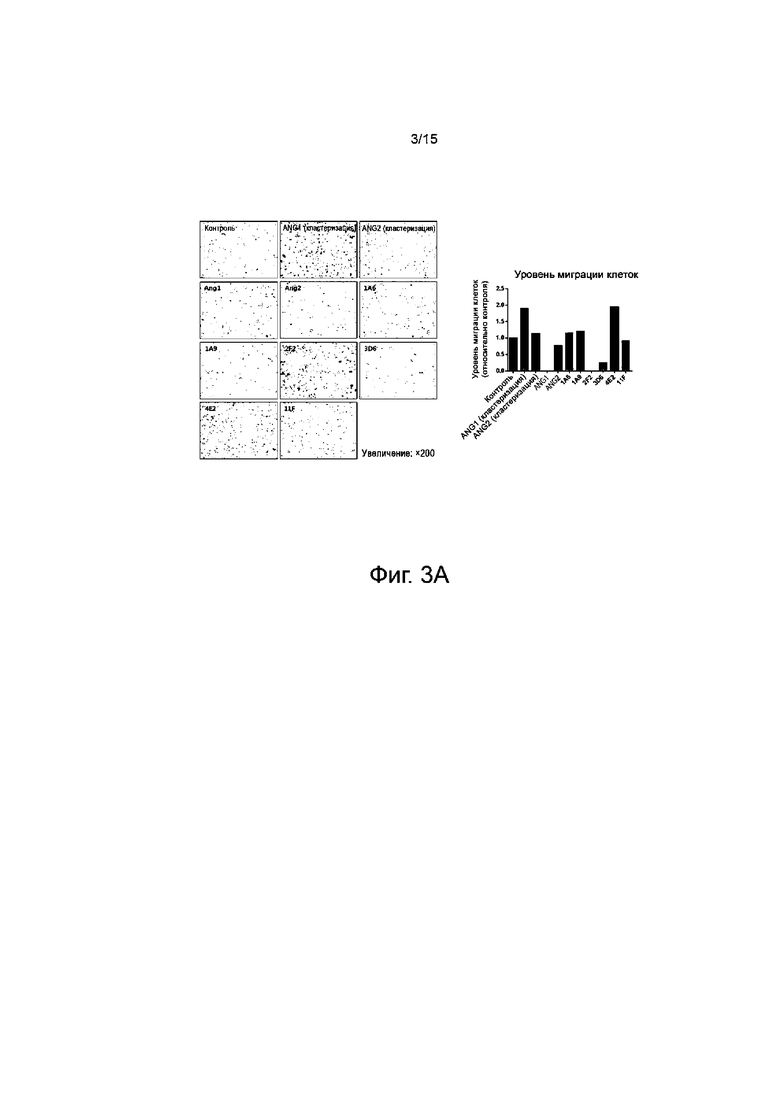

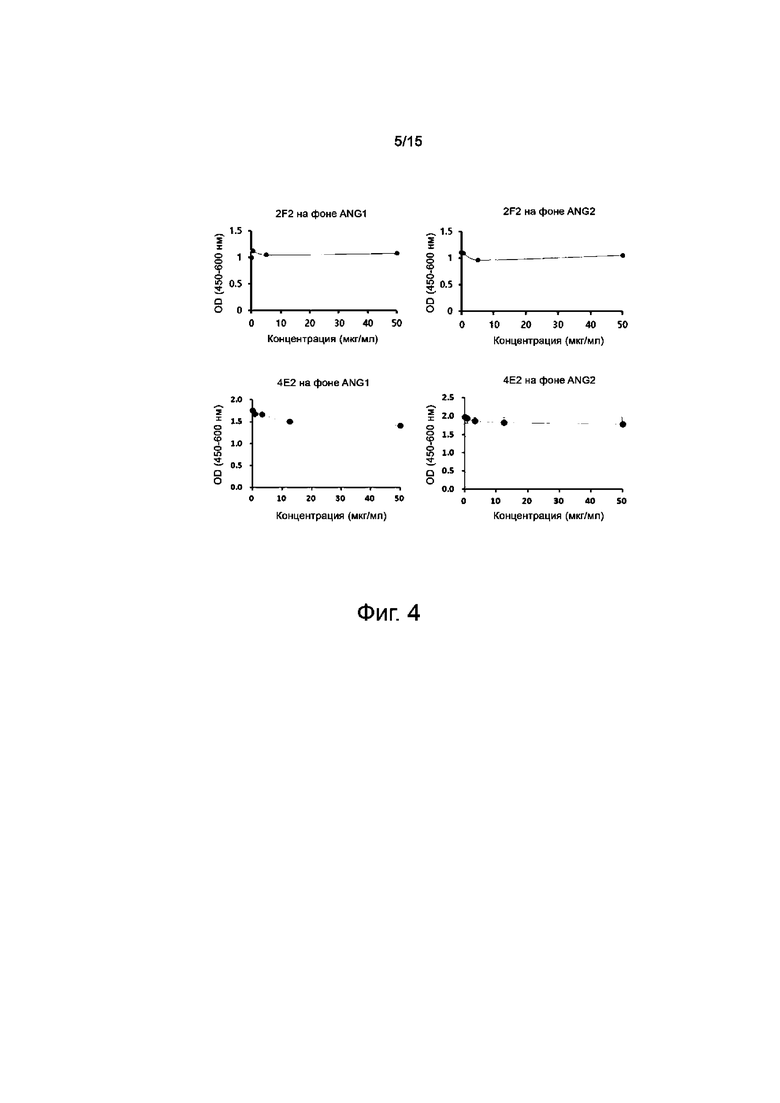
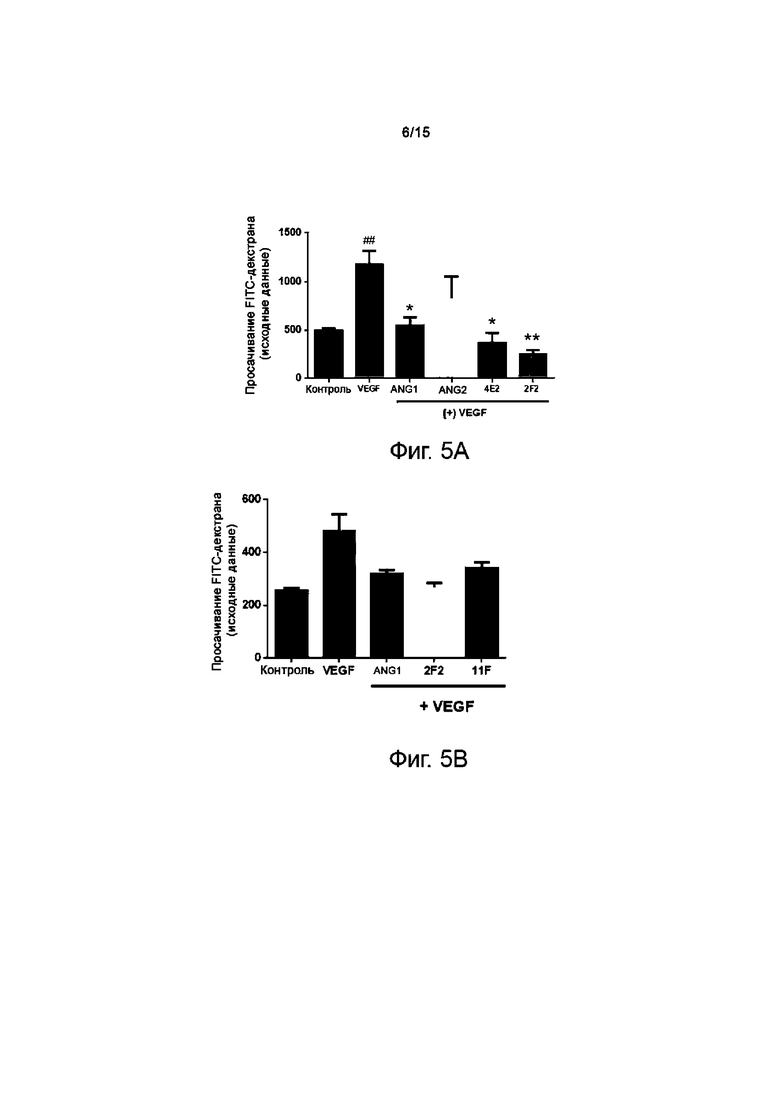
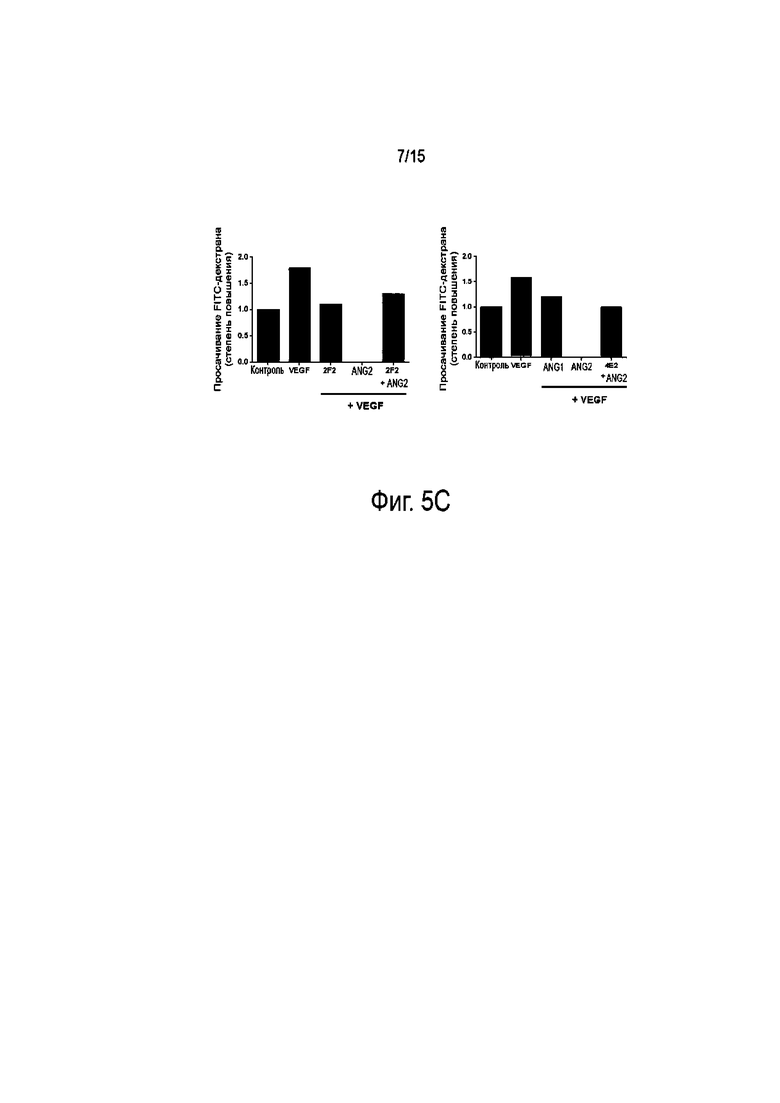
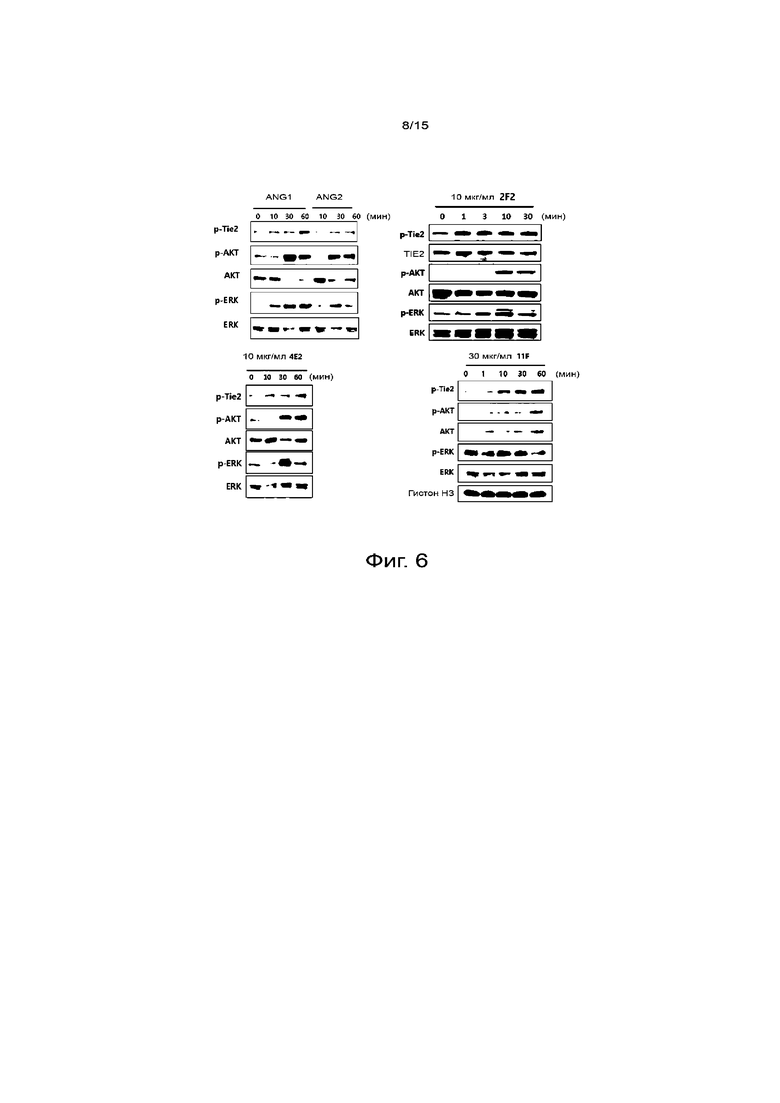
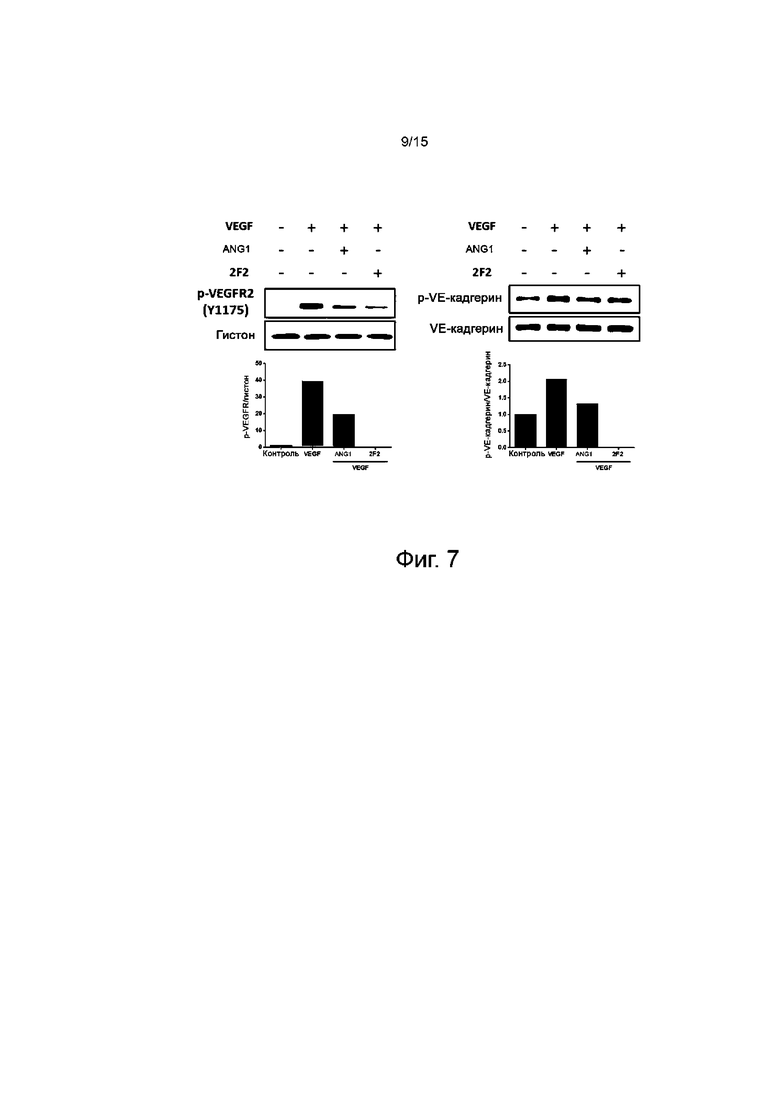



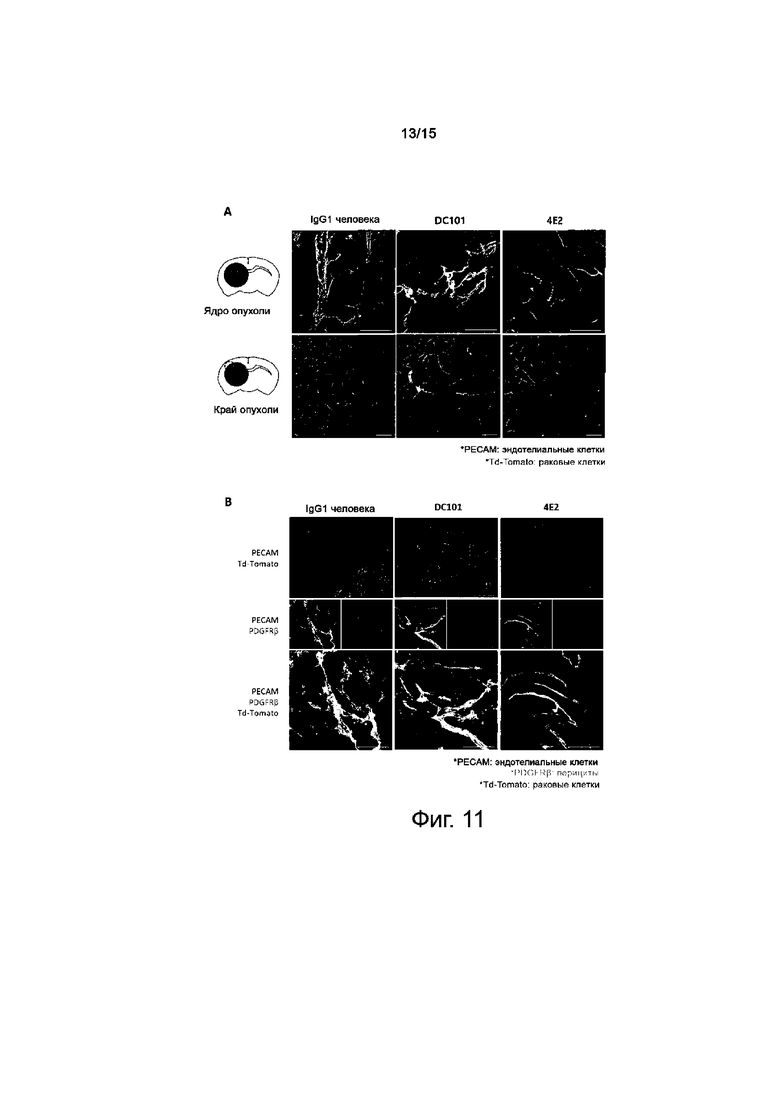
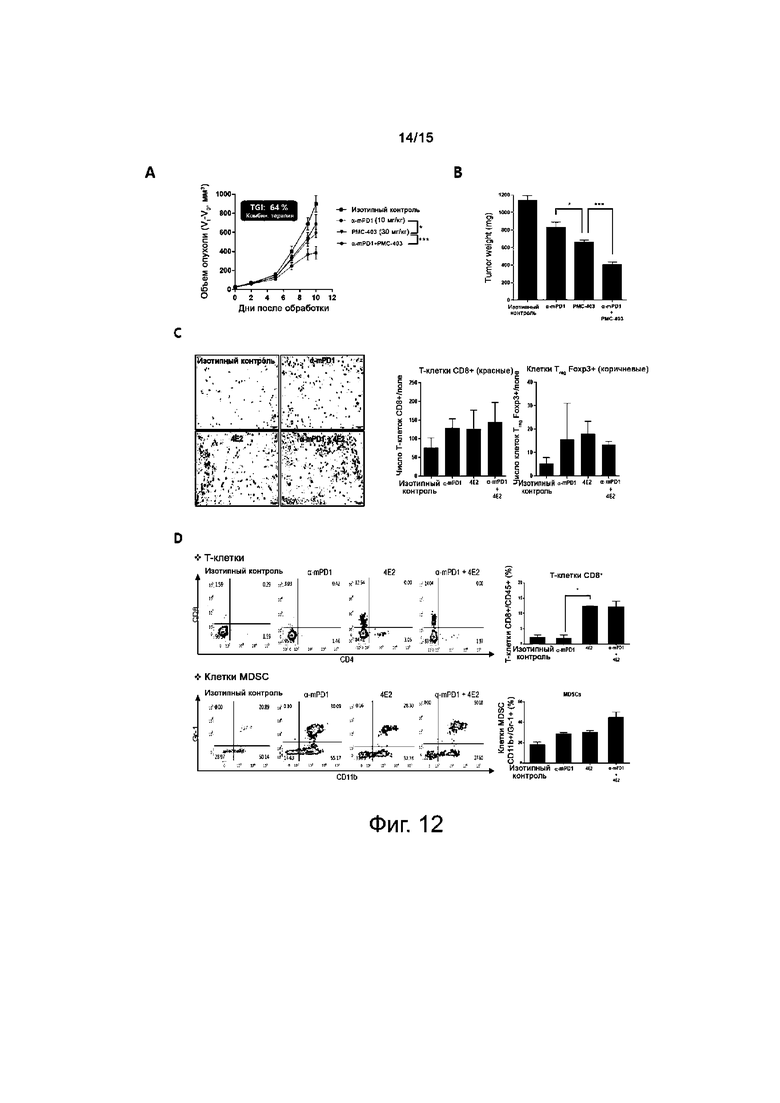
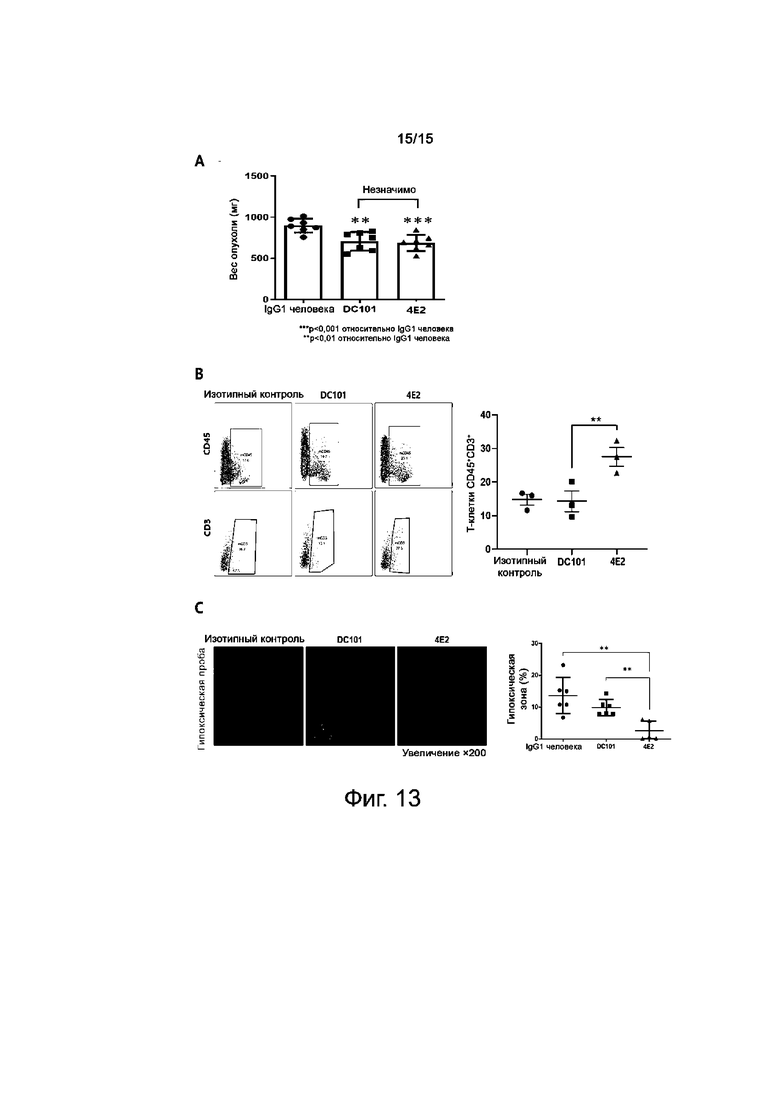
Группа изобретений относится к биотехнологии. Представлены: антитело или его антигенсвязывающий фрагмент, связывающиеся с TIE-2 рецептором тирозин-протеинкиназы, и способ их получения, нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент, экспрессирующий вектор, клетка, трансформированная экспрессирующим вектором, для получения антитела или его антигенсвязывающего фрагмента. Кроме того, представлена композиция для профилактики или лечения связанного с ангиогенезом заболевания, композиция для стабилизации кровеносных сосудов, композиция для диагностики связанного с ангиогенезом заболевания, композиция для профилактики или лечения рака, композиция для профилактики или лечения опухоли, а также применение композиции, содержащей фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента. Изобретение позволяет получить антитело или его антигенсвязывающий фрагмент, связывающиеся с TIE-2 рецептором тирозин-протеинкиназы, а также композиции для профилактики или лечения связанного с ангиогенезом заболевания. 11 н. и 5 з.п. ф-лы, 18 ил., 4 табл., 13 пр.
1. Антитело или его антигенсвязывающий фрагмент, связывающиеся с TIE-2 рецептором тирозин-протеинкиназы, включающие:
вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, имеющий SEQ ID NO: 1, участок CDR2 тяжелой цепи, имеющий SEQ ID NO: 2, и участок CDR3 тяжелой цепи, имеющий SEQ ID NO: 3, и
вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, имеющий SEQ ID NO: 4, участок CDR2 легкой цепи, имеющий SEQ ID NO: 5, и участок CDR3 легкой цепи, имеющий SEQ ID NO: 6;
вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, имеющий SEQ ID NO: 7, участок CDR2 тяжелой цепи, имеющий SEQ ID NO: 8, и участок CDR3 тяжелой цепи, имеющий SEQ ID NO: 9, и
вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, имеющий SEQ ID NO: 10, участок CDR2 легкой цепи, имеющий SEQ ID NO: 11, и участок CDR3 легкой цепи, имеющий SEQ ID NO: 12;
вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, имеющий SEQ ID NO: 13, участок CDR2 тяжелой цепи, имеющий SEQ ID NO: 14, и участок CDR3 тяжелой цепи, имеющий SEQ ID NO:15, и
вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, имеющий SEQ ID NO: 16, участок CDR2 легкой цепи, имеющий SEQ ID NO: 17, и участок CDR3 легкой цепи, имеющий SEQ ID NO: 18;
вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, имеющий SEQ ID NO: 19, участок CDR2 тяжелой цепи, имеющий SEQ ID NO: 20, и участок CDR3 тяжелой цепи, имеющий SEQ ID NO: 21, и
вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, имеющий SEQ ID NO: 22, участок CDR2 легкой цепи, имеющий SEQ ID NO: 23, и участок CDR3 легкой цепи, имеющий SEQ ID NO: 24;
вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, имеющий SEQ ID NO: 25, участок CDR2 тяжелой цепи, имеющий SEQ ID NO: 26, и участок CDR3 тяжелой цепи, имеющий SEQ ID NO: 27, и
вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, имеющий SEQ ID NO: 28, участок CDR2 легкой цепи, имеющий SEQ ID NO: 29, и участок CDR3 легкой цепи, имеющий SEQ ID NO: 30; или
вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, имеющий SEQ ID NO: 31, участок CDR2 тяжелой цепи, имеющий SEQ ID NO: 32, и участок CDR3 тяжелой цепи, имеющий SEQ ID NO: 33, и
вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, имеющий SEQ ID NO: 34, участок CDR2 легкой цепи, имеющий SEQ ID NO: 35, и участок CDR3 легкой цепи, имеющий SEQ ID NO: 36.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, включающие вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 38, 42, 46, 50, 54 и 58.
3. Антитело или его антигенсвязывающий фрагмент по п. 1, включающие вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 40, 44, 48, 52, 56 и 60.
4. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3.
5. Нуклеиновая кислота по п. 4, кодирующая вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 37, 41, 45, 49, 53 и 57.
6. Нуклеиновая кислота по п. 4, кодирующая вариабельную область легкой цепи, выбранная из группы, состоящей из SEQ ID NO: 39, 43, 47, 51, 55 и 59.
7. Экспрессирующий вектор, включающий нуклеиновую кислоту по п. 4.
8. Клетка, трансформированная экспрессирующим вектором по п. 7, для получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3.
9. Способ получения антитела, связывающегося с TIE-2, или его антигенсвязывающего фрагмента, включающий:
(a) культивирование клетки по п. 8; и
(b) выделение антитела или его антигенсвязывающего фрагмента из культивируемой клетки.
10. Композиция для профилактики или лечения связанного с ангиогенезом заболевания, содержащая фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3 в качестве активного ингредиента.
11. Композиция по п. 10, где связанное с ангиогенезом заболевание выбрано из группы, состоящей из опухолей, рака, метастазов опухолей или рака, диабетической ретинопатии, ретинопатии недоношенных, отторжения трансплантата роговицы, дегенерации желтого пятна, неоваскулярной глаукомы, эритроза, пролиферативной ретинопатии, псориаза, гемофилического артрита, образования атеросклеротических бляшек в капиллярах, келоидов, грануляции ран, сосудистых спайков, ревматоидного артрита, остеоартрита, аутоиммунных заболеваний, болезни Крона, рестеноза, атеросклероза, кишечных спаек, болезней от кошачьих царапин, язв, диабетических язв стопы, цирроза печени, нефрита, диабетической нефропатии, хронической почечной недостаточности, сахарного диабета, воспалительных заболеваний, идиопатического фиброза легких, сепсиса, острого респираторного дистресс-синдрома и нейродегенеративных заболеваний.
12. Композиция для стабилизации кровеносных сосудов, содержащая фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3 в качестве активного ингредиента.
13. Композиция для диагностики связанного с ангиогенезом заболевания, содержащая фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3 в качестве активного ингредиента.
14. Композиция для профилактики или лечения рака, содержащая фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3 в качестве активного ингредиента.
15. Композиция для профилактики или лечения опухоли, содержащая фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3 в качестве активного ингредиента.
16. Применение композиции, содержащей фармацевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3 в качестве активного ингредиента, для комбинированного введения вместе с другими терапевтическими средствами для профилактики или лечения связанного с ангиогенезом заболевания.
| WO 2016010014 A1, 21.01.2016 | |||
| HWANG B | |||
| et al., Stimulation of angiogenesis and survival of endothelial cells by human monoclonal Tie2 receptor antibody | |||
| Biomaterials | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Vol | |||
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |
| Pp | |||
| Способ получения камфоры | 1921 |
|
SU119A1 |
| ВЫДЕЛЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩАЯ TIE-2 ЛИГАНД, TIE-2 ЛИГАНД И СПОСОБ ПОЛУЧЕНИЯ, ПЛАЗМИДА (ВАРИАНТЫ), АНТИТЕЛО, КОНЪЮГАТ, ЛИГАНДНОЕ ТЕЛО, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ СТИМУЛИРОВАНИЯ НЕОВАСКУЛЯРИЗАЦИИ, СПОСОБ ПОДДЕРЖАНИЯ КЛЕТКИ, СПОСОБ ИДЕНТИФИКАЦИИ АНТАГОНИСТА TIE-2 РЕЦЕПТОРА | 1995 |
|
RU2232812C2 |
| WO 2013028442 A1, 28.02.2013. | |||

