Область техники
Изобретение относится к области биотехнологии, а именно к технологии получения биологически активных веществ (БАВ) методами генной инженерии, точнее к методам получения ферментативно активного мутеина [C112S] легкой цепи энтерокиназы человека с высоким выходом.
Предшествующий уровень техники.
Энтерокиназа (синоним: энтеропептидаза) - это сериновая протеиназа (шифр Международного Классификатора Ферментов ЕС 3.4.21.9), физиологический активатор трипсина с высокой избирательностью гидролиза пептидной связи Lys-X в сайте узнавания -Asp-Asp-Asp-Asp-Lys-X-, где Х - любая аминокислота, кроме пролина (Light A. and Janska H., The amino-terminal sequence of the catalytic subunit of bovine enterokinase. J. Protein Chem., 1991, 10(5): 475-480). Энтерокиназа человека - мембранный белок, продуцируемый преимущественно энтероцитами двенадцатиперстной кишки (Grant, D.A.W.; Hermon-Taylor, J.; Enterokinase. Methods Enzym. Anal., 3rd Ed. (Bergmeyer, H.U., ed.) 5, 143-155 (1984)) и состоящий в зрелой форме из двух цепей, соединенными дисульфидной связью - тяжелой цепи (120 кДа) и легкой цепи (47 кДа). Протеолитическая активность энтерокиназы полностью обуславливается ее легкой цепью, удаление тяжелой цепи белка также практически не влияет на специфичность фермента в отношении искуственных субстратов, но понижает каталитическую эффективность энтерокиназы в отношении ее природного субстрата - трипсиногена (В.В.Лихарева, Б.В.Васьковецкий, H.Э.Шепель, С.К.Гаранин, А.Г.Михайлова, Л.Д.Румш. Новые субстраты энтеропептидазы 1. Биологически активные гепта-нонапептиды. Биоорганическая химия, 2003, том 29, №2, с.129-134).
В силу того, что высокая субстратная специфичность энтерокиназы имеет важное физиологическое значение, последовательность узнавания энтерокиназы -Asp-Asp-Asp-Asp-Lys- встречается только в составе активационных пептидов трипсиногенов. Это обстоятельство делает энтерокиназу чрезвычайно удобной специфической протеазой для получения биотехнологических лекарственных средств. Введение последовательности узнавания энтерокиназы непосредственно перед первым аминокислотным остатком зрелой формы целевого белка позволяет специфически разделять слитой белок или отделять от целевого белка N-концевой пептид и, таким образом, получать рекомбинантный белок без дополнительных аминокислотных остатков.
Получение слитых белков или белков-предшественников с отделяемым N-концевым пептидом является предпочтительным способом биосинтеза гетерологичных белков для медицинского применения в бактериальной системе экспрессии Е.coli, поскольку способность Е.coli секретировать белки с лидерными пептидами довольно ограничена, а цитоплазматическая экспрессия неизбежно приводит к появлению продуктов с дополнительным остатком метионина на N-конце белка, что приводит к его иммуногенности и падению клинической эффективности.
Очевидным источником для выделения энтерокиназы является внутренняя выстилка двенадцатиперстной кишки крупного рогатого скота или свиней (Anderson LE, Walsh KA, Neurath H. Biochemistry, 1977, 16(15):3354-3360), при этом выделяемая энтерокиназа будет загрязнена сходными с ней протеолитическими ферментами и потенциально может содержать вирусы или прионы млекопитающих.
Более предпочтительной для использования в биотехнологическом производстве является рекомбинантная энтерокиназа, точнее рекомбинантная легкая цепь энтерокиназы человека. Использование рекомбинантной легкой цепи энтерокиназы человека вместо более распространенных вариантов энтерокиназы крупного рогатого скота позволяет полностью избежать загрязнения препаратов целевых белков следами потенциально иммуногенной для человека энтерокиназы крупного рогатого скота. Следует отметить, что выявление следовых количеств энтерокиназы, и, в особенности, следовых количеств ферментативно неактивных протеолитических фрагментов энтерокиназы в препаратах очищенных рекомбинантных белков для медицинского применения практически невозможно.
Рекомбинантная легкая цепь энтерокиназы была успешно получена в большинстве распространенных систем экспрессии гетерологичных генов, в том числе Esherichia coli (патент США 6746859), дрожжах Saccharomyces cerevisae (Choi SI, Song HW, Moon JW, Seong BL. Recombinant enterokinase light chain with affinity tag: expression from Saccharomyces cerevisiae and its utilities in fusion protein technology. Biotechnol Bioeng. 2001; 75(6):718-24) и Pichia pastoris (Vozza LA, Wittwer L, Higgins DR, Purcell TJ, Bergseid M, Collins-Racie LA, LaVallie ER, Hoeffler JP. Production of a recombinant bovine enterokinase catalytic subunit in the methylotrophic yeast Pichia pastoris. Biotechnology (N Y). 1996; 14(1):77-81), мицелиальных грибах Aspergillus (Svetina M, Krasevec N, Gaberc-Porekar V, Komel R. Expression of catalytic subunit of bovine enterokinase in the filamentous fungus Aspergillus niger. J Biotechnol. 2000; 76(2-3):245-51) и культивируемых клетках млекопитающих COS-1 (LaVallie ER, Rehemtulla A, Racie LA, DiBlasio EA, Ferenz C, Grant KL, Light A, McCoy JM. Cloning and functional expression of a cDNA encoding the catalytic subunit of bovine enterokinase. J Biol Chem. 1993; 268(31):23311-7).
Предпочтительной системой экспрессии энтерокиназы являются бактерии Е.coli, поскольку получаемый белок или его предшественник не подвергается гликозилированию и соответственно будет лишен антигенных детерминант олигосахаридов дрожжей. Удельная ферментативная активность гликозилированной и негликозилированной форм легкой цепи энтерокиназы практически не различается (патент США 6746859).
Протеолитическая активность энтерокиназы полностью исчезает при добавлении к N-концу легкой цепи любых аминокислотных остатков, вследствие этого прямая цитоплазматическая экспрессия гена легкой цепи энтерокиназы в Е.coli не приводит к появлению активного продукта (Song HW, Choi SI, Seong BL, Engineered recombinant enteropeptidase catalytic subunit: effect of N-terminal modification. Arch Biochem Biophys. 2002; 400(1):1-6). Для получения в Е.coli легкой цепи энтерокиназы без дополнительных аминокислотных остатков на N-конце могут быть применены два известных основных подхода - добавление к последовательности ДНК, кодирующей легкую цепь энтерокиназы, распознаваемого бактериями лидерного пептида или слияние последовательности ДНК, кодирующей легкую цепь энтерокиназы, в рамке с последовательностями, кодирующими белки-партнеры, например, тиоредоксин I, дисульфидизомеразу А, глутатион-S-трансферазу или мальтозо-связывающий белок. Домены белка-партнера и легкой цепи энтерокиназы при этом могут быть соединены линкерным участком, включающим сайт узнавания энтерокиназы -Asp-Asp-Asp-Asp-Lys-, непосредственно предшествующий первому аминокислотному остатку Ile легкой цепи энтерокиназы. В таком случае ферментативное разделение белка-партнера и интактной легкой цепи энтерокиназы может быть проведено аутокаталитически после добавления небольшого стартового количество ферментативно активной энтерокиназы.
Второй подход к получению рекомбинантной легкой цепи энтерокиназы с интактным N-концом является более распространенным, с его применением был получен ряд препаратов энтерокиназы с значительным выходом продукта и высокой удельной активностью. Так, в работе (Huang L, Ruan H, Gu W, Xu Z, Cen P, Fan L. Functional expression and purification of bovine enterokinase light chain in recombinant Escherichia coli. Prep Biochem Biotechnol. 2007; 37(3):205-17) при использовании белка-партнера дисульфидизомеразы A (DsbA) была достигнута конечная продуктивность 6,8 мг активной энтерокиназы быка на 1 литр бактериальной культуры. В случае белка-партнера тиоредоксина I была достигнута продуктивность 43 мг активной энтерокиназы быка на 1 л культуры (Yuan LD, Hua ZC. Expression, purification, and characterization of a biologically active bovine enterokinase catalytic subunit in Escherichia coli. Protein Expr Purif. 2002; 2 5(2):300-4), при этом отделение энтерокиназы от белка-партнера происходило аутокаталитически in vivo, свободную энтерокиназу извлекали из фракции растворимых цитоплазматических белков и подвергали многостадийной хроматографической очистке. Особенно высокий выход активной легкой цепи энтерокиназы быка - 106 мг на 1 л культуры - был достигнут в работе (Tan H, Wang J, Zhao ZK. Purification and refolding optimization of recombinant bovine enterokinase light chain overexpressed in Escherichia coli. Protein Expr Purif. 2007; 56(1):40-7. Epub 2007 Jul 18) при использовании глутатион-S-трансферазы в качестве белка-партнера.
Препараты рекомбинантной легкой цепи энтерокиназы человека были получены по аналогичной схеме с использованием тиоредоксина в качестве белка-партнера (Yuan LD, Hua ZC. Expression, purification, and characterization of a biologically active bovine enterokinase catalytic subunit in Escherichia coli. Protein Expr Purif. 2002 Jul; 25(2):300-4), при этом слитой белок накапливался в нерастворимой форме, и выход конечного продукта был достаточно низок. В независимой работе (Yi JH, Zhang YX. [Refolding of the fusion protein of recombinant enterokinase light chain rEKL]. [Article in Chinese] Sheng Wu Gong Cheng Xue Bao. 2006 Sep; 22(5):811-5) был применен белок-партнер DsbA и выход конечного продукта увеличен до 60 мг/л культуры.
Все перечисленные выше примеры получения рекомбинантной легкой цепи энтерокиназы обладают основным неустранимым недостатком - неизбежным присутствием в препарате очищенной энтерокиназы остаточных количеств слитого белка, включающего иммуногенный для человека домен белка-партнера, как правило, бактериального происхождения. Чувствительные методы контроля остаточного уровня белка-партнера в препаратах рекомбинантной энтерокиназы являются весьма дорогостоящими и требуют использования антител к белку-партнеру. Вследствие этого получение рекомбинантной негликозилированной легкой цепи энтерокиназы человека с высоким выходом и без использования белков-партнеров представляет большой интерес для фармацевтической промышленности.
Другим существенным недостатком описанных выше примеров является наличие непарного остатка цистеина в положении 112 легкой цепи энтерокиназы, который образует дисульфидную связь с остатком цистеина тяжелой цепи в природном белке. При выделении легкой цепи энтерокиназы из природных источников данный остаток цистеина обычно остается связанным с цистеином короткого остаточного пептида тяжелой цепи, в то время как у рекомбинантных вариантов легкой цепи энтерокиназы он должен находиться в восстановленном состоянии, что уменьшает стабильность фермента и потенциально может приводить к дисульфидному обмену с остатками цистеина расщепляемых энтерокиназой целевых белков и образованию ковалентных мультимеров.
Краткое описание настоящего изобретения
Технической задачей, решаемой авторами, являлось создание технологии получения рекомбинантной негликозилированной легкой цепи энтерокиназы человека с более высоким выходом и без применения белков-партнеров.
Технический результат достигался путем создания технологии, включающей в себя новую экспрессионную плазмидную ДНК р6Е-hEK-6, кодирующую мутеин [C112S] легкой цепи энтерокиназы человека, создание штамма-продуцента E.coli на ее основе и технологию выделения рекомбинантной легкой цепи энтерокиназы человека.
В основе данного решения лежат разработанная авторами плазмидная ДНК р6Е-hEK-6 длиной 6031 п.о., кодирующая отщепляемый N-концевой лидерный пептид длиной 19 аминокислот, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, слитую с ним в рамке последовательность, кодирующую мутеин [C112S] легкой цепи энтерокиназы человека и последовательность, кодирующую неотщепляемый С-концевой гексагистидиновый кластер. Указанный фрагмент содержит оптимальные для Е.coli кодоны, позволяющие увеличить уровень экспрессии гетерологичного белка за счет эффективной трансляции всех аминокислот полипептида. Наличие короткого N-концевого пептида, практически не содержащего антигенных детерминант, позволяет проводить очистку ферментативно неактивного белка-предшественника в денатурирующих условиях при помощи металлохелатной хроматографии и проводить рефолдинг для очищенного белка-предшественника, что обычно приводит к значительному увеличению выхода ренатурированного целевого белка. Наличие неотщепляемого С-концевого гексагистидинового кластера позволяет удалять энтерокиназу из реакционной смеси при ее применении для расщепления гибридных белков и белков-предшественников.
Мутация [C112S] позволяет ренатурировать легкую цепь энтерокиназы человека с более высоким выходом, так как отсутствие непарного цистеина, который в природном белке участвует в образовании дисульфидной связи с тяжелой цепью, снижет вероятность установления межмолекулярных дисульфидных связей, т.е. образования ковалентных мультимеров.
Целью настоящего изобретения является предоставление экспрессионной плазмиды, содержащей фрагмент ДНК, кодирующий мутеин [C112S] легкой цепи энтерокиназы человека, включающий последовательность, кодирующую отщепляемый N-концевой лидер, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей мутеин [C112S] легкой цепи энтерокиназы человека, и последовательность, кодирующую неотщепляемый С-концевой гексагистидиновый кластер, под контролем промотора, функционирующего в бактериальной клетке.
Также целью настоящего изобретения является предоставление описанной выше экспрессионной плазмиды, где указанная плазмида представлена плазмидой p6E-hEK-6.
Также целью настоящего изобретения является предоставление бактерии, принадлежащей к роду Escherichia, трансформированной описанной выше плазмидой, - продуцента мутеина [C112S] легкой цепи энтерокиназы человека.
Также целью настоящего изобретения является предоставление описанной выше бактерии, где указанная бактерия представлена штаммом Е.coli BL21[DE3]/p6E-hEK-6.
Также целью настоящего изобретения является предоставление предшественника рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека, содержащего отщепляемый N-концевой пептид, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, а также неотщепляемый С-концевой пептид, содержащий гексагистидиновый кластер.
Также целью настоящего изобретения является предоставление способа получения рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека, включающий культивирование описанной выше бактерии в питательной среде, выделение телец включения, солюбилизацию белка-предшественника, металлохелатную хроматографию в денатурирующих условиях, рефолдинг белка-предшественника, аутокаталитическое получение зрелого белка и выделение зрелого белка.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором культивируют штамм Е.coli BL21[DE3]/p6E-hEK-6.
Также целью настоящего изобретения является предоставление рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека, включающего неотщепляемый С-концевой пептид, содержащий гексагистидиновый кластер, полученного описанным выше способом.
Подробное описание настоящего изобретения
Для реализации настоящего изобретения главной технической задачей явилось создание способа получения рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека с использованием бактерии, трансформированной экспрессионной плазмидой, содержащей фрагмент ДНК, кодирующий предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека, включающий последовательность, кодирующую отщепляемый N-концевой дополнительный пептид, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, и слитую с ним в рамке последовательность, кодирующую рекомбинантный мутеин [C112S] легкой цепи энтерокиназы человека и неотщепляемый С-концевой гексагистидиновый кластер, под контролем промотора, функционирующего в бактериальной клетке.
Термин «экспрессионная плазмида» означает плазмидную ДНК, содержащую все необходимые генетические элементы для экспрессии внедренного в него гена, например, такие как промотор, терминатор. Конкретным примером генетических элементов, необходимых для экспрессии предшественника рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека в составе экспрессионной кассеты согласно настоящему изобретению, является, но не ограничивается им, промотор РНК-полимеразы бактериофага Т7.
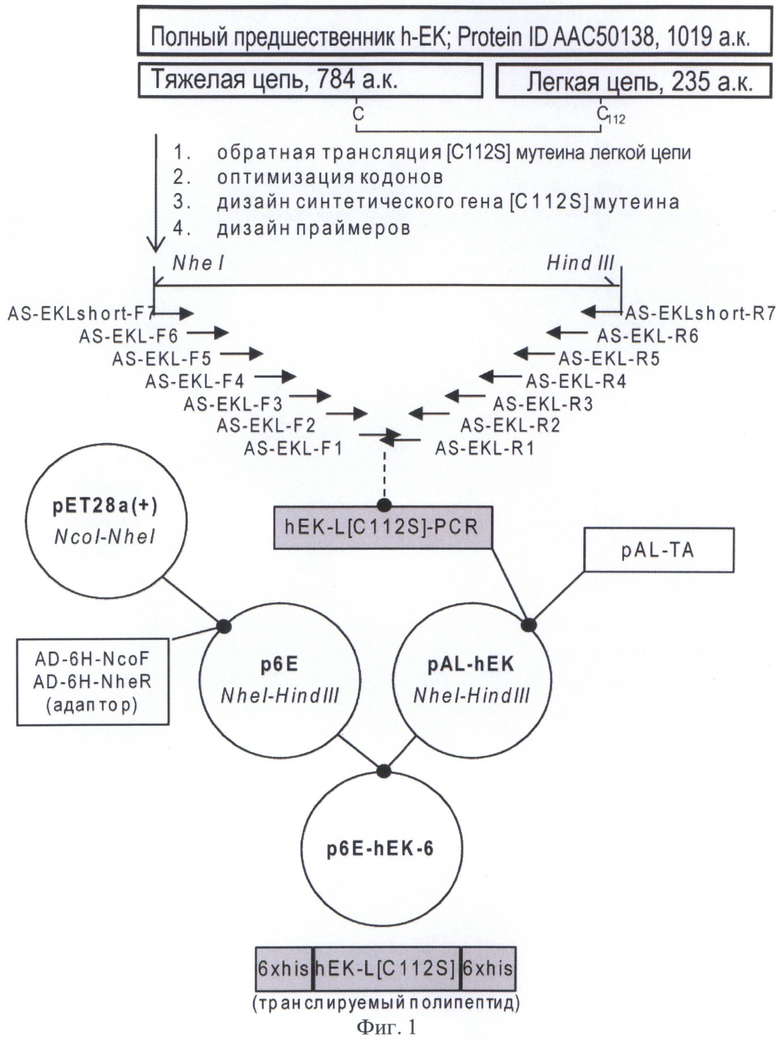
Фрагментом ДНК, кодирующим предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека согласно настоящему изобретению, является, например, синтетический ген, кодирующий предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека, включающий последовательность, кодирующую отщепляемый N-концевой дополнительный пептид, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей рекомбинантный мутеин [C112S] легкой цепи энтерокиназы человека и неотщепляемый С-концевой гексагистидиновый кластер. Указанный фрагмент ДНК может быть получен, например, методом ПНР (см. Пример 1, Фиг.1). Также указанный фрагмент ДНК может быть получен с использованием технологии клонирования фирмы Sloning BioTechnology, описанной в заявке РСТ WO2005071077.
Чтобы обеспечить эффективную трансляцию клонированного гена в Е.coli, предпочтительно, чтобы в последовательности, кодирующей предшественник рекомбинаного мутеина [C112S] легкой цепи энтерокиназы человека, все редкие кодоны были заменены синонимичными часто встречающимися в активно транслируемых генах Е.coli кодонами.
Последовательность гена, кодирующего предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека согласно настоящему изобретению, представлена в Перечне последовательностей под номером SEQ ID NO:1. Аминокислотная последовательность предшественника рекомбинантной легкой цепи энтерокиназы человека согласно настоящему изобретению представлена в Перечне последовательностей под номером SEQ ID NO:2. Аминокислотная последовательность зрелого рекомбинаного мутеина [C112S] легкой цепи энтерокиназы человека представляет собой последовательность под номером SEQ ID NO:2 без 19 первых аминокислот.
Фрагменты ДНК, которые кодируют по существу тот же белок, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК (SEQ ID NO:1), кодирующей предшественник рекомбинаного мутеина [C112S] легкой цепи энтерокиназы человека, например, посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации. Фрагменты ДНК, которые кодируют по существу тот же белок, могут быть выявлены путем экспрессии фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке и установления активности экспрессируемого продукта.
Энтерокиназа (синоним: энтеропептидаза) - это сериновая протеаза (шифр Международного Классификатора Ферментов ЕС 3.4.21.9), которая катализирует расщепление пептидной связи преимущественно после остатка лизина, более преимущественно между последовательностью аминокислот -Asp-Asp-Asp-Asp-Lys- и любой следующей аминокислотой, кроме пролина. Энтерокиназа человека - это мембранный гликозилированный белок, в зрелой форме состоящий из двух цепей, соединенных дисульфидными связями. Тяжелая цепь энтерокиназы имеет массу около 120 кДа, а легкая цепь - около 47 кДа. В организме Энтерокиназа осуществляет превращение трипсиногена в трипсин.
В силу того, что сайт узнавания энтерокиназы (-Asp-Asp-Asp-Asp-Lys-) относительно редко встречается в белках, энтеропептидаза широко используется в биотехнологии для расщепления рекомбинантных гибридных белков.
Легкая цепь энтерокиназы является химотрипсиноподобной сериновой протеазой, обладает практически той же ферментативной активностью и специфичностью, что и полный белок.
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами рекомбинантной легкой цепи энтерокиназы человека, определяются по его протеолитической активности (его способности гидролизовать пептидную связь). Так, например, активность рекомбинантной легкой цепи энтерокиназы человека можно детектировать по гидролизу синтетического субстрата как описано в Примере 5, либо по гидролизу слитых белков, содержащих сайт узнавания энтерокиназы, как описано в Примере 6. Считается, что вариант белка обладает свойствами рекомбинантной легкой цепи энтерокиназы человека при условии, что активность указанного варианта составляет не ниже 1% активности нативной легкой цепи энтерокиназы человека.
Экспрессионная плазмида согласно настоящему изобретению содержит фрагмент ДНК, кодирующий предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека, включающий последовательность, кодирующую отщепляемый N-концевой дополнительный пептид, включающий гексагистидиновый кластер, и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей рекомбинантный мутеин [C112S] легкой цепи энтерокиназы человека и неотщепляемый С-концевой гексагистидиновый кластер, под контролем промотора, функционирующего в бактериальной клетке.
В качестве рекомбинантной плазмиды согласно настоящему изобретению могут использоваться различные плазмиды, обладающие способностью к экспрессии в клетке-реципиенте, такие как плазмиды pBR322, pMW119, pUC19, pET22b, pET28b и подобные им, но список плазмид не ограничивается ими.
Конкретным вариантом реализации настоящего изобретения являются плазмида, которая состоит из:
1) фрагмента NheI-NcoI длиной 29 п.о., представляющего собой синтетический адаптер, кодирующий гексагистидиновый кластер;
2) фрагмента NcoI-HmdIII вектора рЕТ28а(+) длиной 5246 п.о., содержащего область начала репликации плазмиды pBR322, ген РНК-организующего белка Rop, участок инициации репликации бактериофага f1, последовательность, кодирующую аминогликозид-3'-фосфотрансферазу, промотор РНК-полимеразы бактериофага Т7; участок терминации транскрипции; последовательность, кодирующую репрессор лактозного оперона; последовательность, кодирующую С-концевой гексагистидиновый кластер;
3) фрагмента NheI-HindIII длиной 756 п.о., кодирующего фрагмент рекомбинантной легкой цепи энтерокиназы человека и слитую в рамке последовательность узнавания энтерокиназой.
Указанная плазмида содержат уникальные сайты узнавания эндонуклеазами рестрикции: PsiI (371), PciI (2142), ApaI (4040), XbaI (5031), NheI (5099), PstI (5562), HindIII (5855).
Структура плазмиды p6E-hEK-6 приведена на Фиг.2.
При помощи созданной плазмиды можно трансформировать бактериальную клетку, предпочтительно бактерию, принадлежащей к роду Escherichia, восприимчивую к подобной трансформации указанной плазмидой. Выбор конкретной клетки не является критическим, поскольку методология и приемы трансформации хорошо известны специалисту в данной области техники. И хотя в зависимости от вида клетки и условий культивирования полученного трансформанта уровень экспрессии предшественника мутеина легкой цепи энтерокиназы человека может варьироваться, факт экспрессии целевого белка будет иметь место при условии успешной трансформации клетки - реципиента.
«Трансформация клетки плазмидой» означает введение плазмиды в клетку с помощью методов, хорошо известных специалисту в данной области техники. Трансформация этой плазмидой приводит к экспрессии гена, кодирующего белок согласно настоящему изобретению, и к синтезу белка в бактериальной клетке. Методы трансформации включают любые стандартные методы, известные специалисту в данной области техники, например метод, описанный в Jac A. Nickoloff, Electroporation Protocols for Microorganisms (Methods in Molecular Biology) // Humana Press; 1st edition (August 15, 1995).
Согласно настоящему изобретению, «бактериальная клетка - продуцент предшественника мутеина легкой цепи энтерокиназы человека» означает бактериальную клетку, обладающую способностью к продукции и накоплению предшественника мутеина легкой цепи энтерокиназы человека согласно настоящему изобретению, когда бактериальная клетка согласно настоящему изобретению выращивается в указанной питательной среде. Используемый здесь термин «бактериальная клетка - продуцент предшественника мутеина рекомбинантной легкой цепи энтерокиназы человека» также означает клетку, которая способна накапливать продукт предшественника мутеина легкой цепи энтерокиназы человека в количестве не менее чем 1 мг/л, более предпочтительно, не менее чем 40 мг/л. Указанный предшественник мутеина легкой цепи энтерокиназы человека накапливается в указанной клетке предпочтительно в виде телец включения.
Предпочтительно использование бактерии, принадлежащей к роду Escherichia, для трансформации рекомбинантной плазмидой, содержащей фрагмент ДНК, кодирующий предшественник мутеина легкой цепи энтерокиназы человека.
Термин «бактерия, принадлежащая к роду Escherichia» может означать, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, может быть упомянута бактерия Escherichia coli (Е.coli).
Круг бактерий, принадлежащих к роду Escherichia, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть приведены в качестве примеров.
Конкретным примером штамма-реципиента для получения продуцента предшественника мутеина легкой цепи энтерокиназы человека согласно настоящему изобретению, является, но не ограничиваются им, штамм Escherichia coli BL21[DE3].
Штамм Escherichia coli BL21[DE3] характеризуется следующими культурально-морфологическими, физиолого-биохимическими признаками и генетическими признаками.
Культурально-морфологические особенности штамма: грамотрицательные палочки, образуют нити; на агаризованной среде - беловатые крупные колонии с неровным краем. Активность штамма определяется методом денситометрия электрофореграммы. Штамм хранится в следующих условиях: среда Лурье-Бертрана, 1% глюкозы, 10% глицерина. Штамм размножается в следующих условиях - среда Лурье-Бертрана, 1% глюкозы, канамицина сульфат 30 мкг/мл.
Генетические особенности штамма. Генотип штамма - F- ompT gal dcm Ion hsdSB(rB - mB -)λ(DE3 [lacI lacUV5-T7 gene 1 ind1 sam7 nin5])
Трансформация штамма Escherichia coli BL21[DE3] плазмидой p6E-hEK-6 приводит к получению штамма-продуцента BL21[DE3]/p6E-hEK-6, который обеспечивает синтез рекомбинантного белка-предшественника мутеина легкой цепи энтерокиназы человека в количестве 30-70% от суммарного содержания белка клеток.
Штамм Escherichia coli BL21[DE3]/p6E-hEK-6 кодирует белок-предшественник 6Е-hEK-6, состоящий из аминокислотной последовательности мутеина [C112S] легкой цепи энтерокиназы человека, слитого с ним в рамке N-концевого пептида длиной 19 аминокислот, содержащего гексагистидиновый кластер, сайт расщепления энтерокиназой, находящийся непосредственно перед первой аминокислотой мутеина легкой цепи энтерокиназы человека, и неотщепляемый С-концевой гексагистидиновый кластер.
Способ получения мутеина [C112S] легкой цепи энтерокиназы человека согласно настоящему изобретению включает культивирование описанной выше бактерии в питательной среде, подходящей для выращивания указанных прокариотических клеток, индукцию промотора гена предшественника мутеина [C112S] легкой цепи энтерокиназы человека, выделение телец включения, солюбилизацию белка-предшественника, металлохелатную хроматографию в денатурирующих условиях, рефолдинг белка-предшественника, аутокаталитическое получение зрелого белка, инициируемое добавлением ферментативно активной энтерокиназы, и выделение указанного зрелого белка рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека.
Особенности плазмиды и результаты ее практического применения приведены на следующих чертежах.
Краткое описание чертежей
На Фигуре 1 показана схема сборки синтетического гена мутеина [C112S] легкой цепи энтерокиназы человека из олигонуклеотидных праймеров и схема получения экспрессионной плазмиды p6E-hEK-6. Используются следующие обозначения: Полный предшественник h-EK; Protein ID AAC50138, 1019 а.к. - продукт экспрессии природного гена энтерокиназы человека; hEK-L[C112S]-PCR - продукт полимеразной цепной реакции, кодирующий мутеин [C112S] легкой цепи энтерокиназы человека; 6xhis-hEK-L[C112S]-6xhis - продукт экспрессии плазмиды p6E-hEK-6. Пунктирная линия обозначает полимеразную цепную реакцию, сплошная - рестрикцию и лигирование фрагментов ДНК.
На Фигуре 2 показана карта экспрессионной плазмиды p6E-hEK-6. Используются следующие обозначения: «pBR322ori» область начала репликации плазмиды pBR322; «ROP» - ген РНК-организующего белка Rop; «f1 ori» - участок инициации репликации бактериофага f1; «KanR2» - последовательность, кодирующая аминогликозид-3'-фосфотрансферазу, обеспечивающую устойчивость бактерий к канамицину; «Т7 prom» - промотор РНК-полимеразы бактериофага Т7; «T7term» - участок терминации транскрипции; «lacI» - последовательность, кодирующая репрессор лактозного оперона; «p6E-hEK-6» - открытая рамка считывания (ОРС) полипептида белка-предшественника мутеина [C112S] легкой цепи энтерокиназы человека, включающего отделяемый N-концевой дополнительный пептид, содержащий гексагистидиновый кластер «6xhis», сайт узнавания энтерокиназой «EK-PRO», неотделяемый С-концевой гексагистидиновый кластер «6xhis»». Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
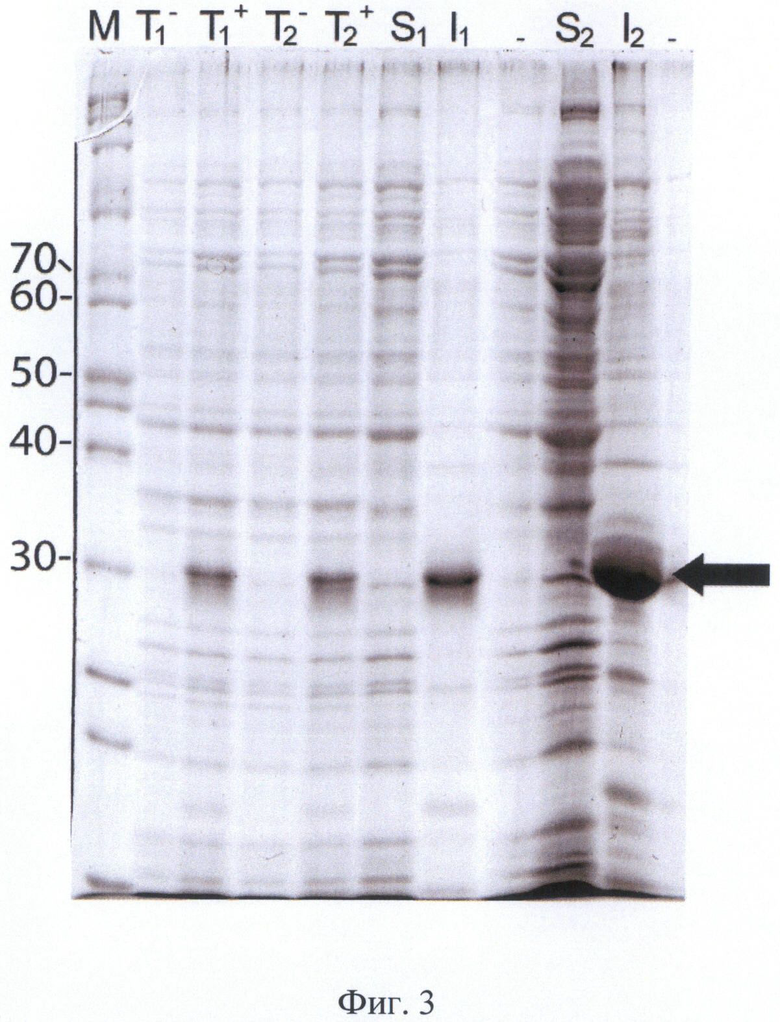
На Фигуре 3 показана электрофореграмма тотального белка, фракций растворимого и нерастворимого белка клеток штамма-продуцента BL21[DE3]/p6E-hEK-6 при индукции для двух случайно выбранных клонов. Культивация во встряхиваемой колбе, среда 2xYT, индукция 1 мМ ИПТГ, 37°С, 20 часов. Обозначения: T1 + и T2 + - тотальный белок после индукции, T1 - и T2 - - тотальный белок до индукции, S1 и S2 - фракция растворимых белков, I1 и I2 - фракция нерастворимых белков (тельца включения) для клонов 1 и 2 соответственно. М - маркер молекулярных масс. Молекулярные массы полос маркера указаны в кДа. Положение целевого белка указано стрелкой.
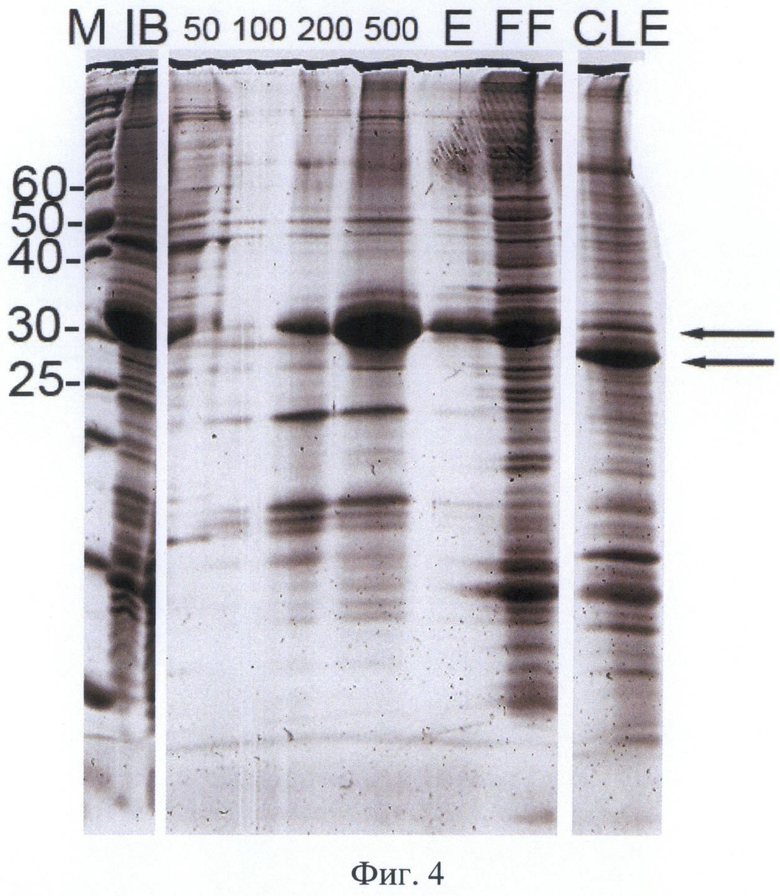
На Фигуре 4 приведена электрофореграмма фракций белка-предшественника 6Е-hEK-6 при очистке металлохелатной хроматографией в денатурирующих условиях, рефолдинга и аутокаталитического процессинга. Восстанавливающие условия. Обозначения: "IB" - солюбилизированные тельца включения; "FF" - фракция проскока, "50-500" - фракции элюций соответствующими концентрациями имидазола, в мМ; "Е" - фракция элюата раствором ЭДТА-Na; "CLE" - зрелый белок hEK-6 после аутокаталитического процессинга N-концевого дополнительного пептида; "М" - маркер молекулярных масс. Молекулярные массы полос маркера указаны в кДа. Положение целевого белка указано стрелкой.
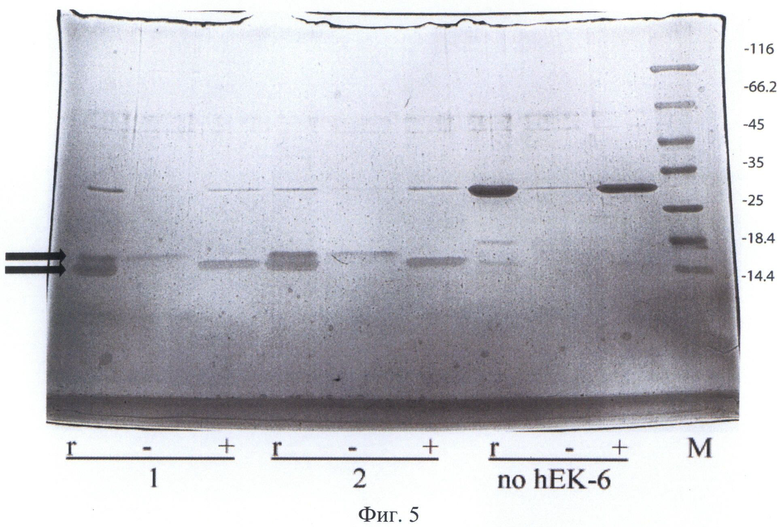
На Фигуре 5 показана электрофореграмма расщепления гибридного белка Trx-MOG очищенной энтерокиназой hEK-6. Невосстанавливающие условия. Обозначения: группа дорожек "1" - гидролиз гибридного белка при помощи hEK-6 в соотношении 1:200; дорожка "r" - реакционная смесь, дорожка "-" - фракция не связавшихся с металлохелатным сорбентом белков (MOG), дорожка "+" - фракция связавшихся с металлохелатным сорбентом белков, элюированных ЭДТА-Na (Trx, hEK-6). Группа дорожек "2" - гидролиз при помощи рекомбинантной энтерокиназы быка (Sigma, США) в соотношении 1:200; обозначения дорожек аналогично "I". Группа дорожек "no hEK-6" - контрольная инкубация гибридного белка без добавления энтерокиназ, обозначения дорожек аналогично "I". М - маркер молекулярных масс, массы приведены в кДа. Положения специфических продуктов распада гибридного белка Trx и MOG указаны стрелками.
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Получение плазмидной ДНК pAL-hEK
Для аминокислотной последовательности, включающей легкую цепь энтерокиназы человека с добавленным 19-аминокислотным N-концевым пептидом (SEQ ID NO:3), содержащим последовательность узнавания энтерокиназой, была проведена обратная трансляция в последовательность нуклеотидов ДНК. При этом были использованы кодоны, оптимальные для экспрессии этого гена в Е.coli класса В, а также была проведена оптимизация структуры гена по вторичной структуре мРНК, GC составу, обеспечено отсутствие нежелательных регуляторных элементов (например, отсутствие внутренних сайтов связывания рибосом), а также отсутствие протяженных повторов, палиндромов.
Индекс CAI (Codon Adaptation Index), отражающий эффективность экспрессии гена в данном организме, для полученной последовательности составил 0,7, что является хорошим прогностическим показателем для промышленной пригодности полученного на его основе штамма-продуцента. Полученная нуклеотидная последовательность приведена в SEQ ID NO:4.
Синтетический ген рекомбинантной легкой цепи энтерокиназы человека собирали методом полимеразной цепной реакции (ПЦР) с праймерами AS-EKL-F1 - AS-EKL-F7 и AS-EKL-R1 - AS-EKL-R7 (SEQ ID NO:5-18), последовательности которых приведены в Таблице 1.
ПЦР проводили на приборе Терцик МС2 («ДНК-технология», Россия). Препаративные реакции проводили в объеме 50 мкл. Готовили инкубационную смесь следующего состава: 1х буфер для термостабильной ДНК-полимеразы; 10 пМ каждого праймерного олигонуклеотида; 2 мМ каждого дезоксирибонуклеотидтрифосфата; 2 ед. термостабильной ДНК-полимеразы. Поверх этой смеси наслаивали 50 мкл минерального масла и вели амплификацию по схеме: 1 цикл - денатурация - 94°С, 3 мин; 25 циклов - денатурация 94°С, 30 сек, отжиг 62°С 30 сек, наращивание цепи 72°С, 120 сек; 1 цикл - наращивание цепи 72°С, 10 мин.
Продукты ПЦР выделяли из 1% агарозного геля, используя набор реактивов «Wizard SV Gel and PCR Clean-Up System» («Promega», США) по протоколу производителя, после чего лигировали в Т-вектор pGEM-T («Promega», США) с использованием ДНК-лигазы фага Т4 и стандартного буферного раствора («Fermentas», Литва). Лигирование вели в объеме 10 мкл при молярном соотношении вектора и вставки 1:10, в течение 2-20 часов при комнатной температуре. Полученными лигазными смесями трансформировали клетки Е.coli штамма DH5α, с генотипом F-φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17 (rk-, mk+) phoA supE44 λ- thi-1 gyrA96 relA1. Для этого к 200 мкл замороженной суспензии клеток Е.coli добавляли 5 мкл лигазной смеси, инкубировали на льду 20 минут для сорбции плазмидной ДНК, нагревали до 42°С в течение 45 секунд и инкубировали на льду в течение 5 минут. После чего добавляли 800 мкл питательного бульона LB и инкубировали при 37°С в течение 60 минут. После инкубации переносили суспензию на чашку Петри с твердой агаризованной средой, содержащей ампициллин в концентрации 100 мкг/1 мл агара, и помещали в термостат на 18 часов при 37°С.
Колонии Е.coli, отобранные в результате бело-голубого скрининга, анализировали методом ПЦР с клонов, с использованием праймеров к последовательностям реципиентной плазмиды T7prom (SEQ ID NO:19) и SP6 (SEQ ID NO:20). Отобранные клоны наращивали в 5 мл питательного бульона LB и проводили выделение плазмидной ДНК набором реактивов «Wizard Plus SV Minipreps» («Promega», США) по протоколу производителя, первичную нуклеотидную последовательность конструкции pAL-hEK подтверждали методом ПЦР-секвенирования с использованием праймеров T7prom (SEQ ID NO:19) и SP6 (SEQ ID NO:20).
После этого проводили препаративную рестрикцию плазмиды pAL-hEK эндонуклеазами рестрикции NheI и HindIII. Продукты реакции разделяли в 1% агарозном геле и выделяли с помощью набора реактивов Wizard SV Gel&PCR Clean-Up System no методике производителя донорный фрагмент для получения экспрессионной плазмиды р6Е-hEK-6.
Пример 2. Получение векторной плазмиды р6Е.
Последовательность, кодирующую фрагмент N-концевого пептида, включающего гексагистидиновый кластер, получали методом отжига двух частично комплиментарных олигонуклеотидов AD-6H-NcoF (SEQ ID NO:21) и AD-6H-NheR (SEQ ID NO:22) с образованием выступающих «липких» 5'-концов. Для этого вносили в пробирку по 100 пм каждого олигонуклеотида, нагревали до 95°С и медленно охлаждали до комнатной температуры.
Реципиентную плазмиду рЕТ28а (+) (Novagen) расщепляли последовательно каждой из эндонуклеаз NcoI и NheI. Сначала аликвоты плазмиды инкубировали в течение 2 часов при 37°C с каждой из рестриктаз, затем контролировали электрофорезом в агарозном геле полную линеаризацию плазмид (в пределах чувствительности метода), после чего добавляли вторую рестриктазу и инкубировали в течение еще 2 часов. Затем пробы объединяли, рестриктазы инактивировали прогреванием при 65°С в течение 20 минут. Рестрицированную плазмиду переосаждали 3 объемами этанола, центрифугировали 10 минут на скорости 13200 об/мин при комнатной температуре, промывали осадок 70% спиртом, растворяли в воде и использовали для постановки реакции лигирования в рабочей концентрации 10-2 мкг/мкл. Выделенные фрагменты лигировали с использованием Т4 ДНК лигазы («Fermentas», Литва) по методике производителя. После этого проводили трансформацию клеток Е.coli штамма DH5α, как описано выше. Колонии Е.coli анализировали методом ПЦР с клонов, с использованием олигонуклеотидов AD-6H-NcoF (SEQ ID NO:21) и T7t (SEQ ID NO:23). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, выделяли плазмидную ДНК набором «GeneJET Plasmid Miniprep Kit» («Fermentas», Литва) по протоколу производителя. Для полученных генетических конструкций р10Е определяли нуклеотидную последовательность ДНК методом ПЦР-секвенирования с праймера T7t (SEQ ID NO:23).
Пример 3. Получение экспрессионной плазмидной ДНК р6Е-hEK-6.
Реципиентную плазмиду р6Е расщепляли последовательно каждой из эндонуклеаз NheI и HindIII. Сначала аликвоты плазмиды инкубировали 2 часа при 37°C с каждой из рестриктаз, контролировали электрофорезом в агарозном геле полную линеаризацию плазмид, затем добавляли вторую рестриктазу и инкубировали в течение еще 2 часов. Затем пробы объединяли, ферменты инактивировали прогреванием при 65°С в течение 20 минут и проводили дефосфорилирование щелочной фосфатазой («Fermentas», Литва) по протоколу производителя. Щелочную фосфатазу инактивировали прогреванием до 85°С в течение 20 минут. Рестрицированную и дефосфорилированную плазмиду переосаждали 3 объемами этанола, центрифугировали 10 минут на скорости 13200 об/мин при комнатной температуре, промывали осадок 70% спиртом, растворяли в воде и использовали для постановки реакции лигирования в рабочей концентрации 10-2 мкг/мкл.
Реакцию лигирования очищенного фрагмента, соответствующего минигену энтерокиназы и реципиентной плазмиды, проводили, как описано выше. После этого проводили трансформацию клеток Е.coli штамма DH5α, как описано выше.
Колонии Е.coli анализировали методом ПЦР с клонов с праймеров T7prom (SEQ ID NO:19) и T7t (SEQ ID NO:23). Отобранные клоны наращивали в 5 мл питательного бульона 2xYT-Kan, проводили выделение плазмидной ДНК с набором «Wizard Plus SV Minipreps» («Promega», США) по протоколу производителя. При помощи ПЦР-секвенирования для конструкции р6Е-hEK-6 определяли нуклеотидную последовательность обеих комплементарных цепей ДНК для области вставки с использованием праймеров T7prom (SEQ ID NO:19) и T7t (SEQ ID NO:23). В результате секвенирования установили, что в полученном препарате плазмиды не содержатся мутации в области вставки, то есть кодируется корректная последовательность гена мутеина легкой цепи энтерокиназы человека. Карта конструкции приведена на Фиг.2.
Пример 4. Получение штамма-продуцента Е.coli BL21[DE3]/p6E-hEK-6, оценка продуктивности штамма-продуцента и локализация целевого белка.
Для получения штамма-продуцента мутеина [C112S] легкой цепи энтерокиназы человека конструкцию, полученную по примеру 3, использовали для трансформации компетентных клеток Escherichia coli BL21(DE3) (с генотипом F- ompT hsdSa (r-m-) gal dcm (DE3)) и проводили отбор клонов, сохраняющих уровень биосинтеза рекомбинантного полипептида после индукции не ниже 30% от суммарного клеточного белка в течение, по крайней мере, четырех последовательных пассажей. Для получения штамма Е.coli BL21[DE3]/p6E-hEK-6 - продуцента белка-предшественника мутеина [C112S] легкой цепи энтерокиназы человека клетки штамма Е.coli BL21[DE3] трансформировали экспрессионной плазмидой р6Е-hEK-6. Трансформанты Е.coli BL21[DE3] высевали на агаризованную среду 2xYT с добавлением канамицина до 30 мкг/мл и глюкозы до 2%, проводили индукцию экспрессии целевого гена для пяти случайно выбранных колоний трансформантов с типичным фенотипом колоний. Клоны подращивали в питательном бульоне с добавлением канамицина до 30 мкг/мл и раствора глюкозы до 2% в течение 6-7 часов, инокулировали новую порцию питательной среды в соотношении 1:100, растили культуру до достижения оптической плотности 2 О.Е., индуцировали изопропилтио-β-D-галактозидом (ИПТГ) и культивировали в течение еще 16 ч. После окончания культивации осадок клеток отделяли центрифугированием, ресуспендировали клетки в растворе 10 мМ Трис-HCl, 2 мМ ЭДТА-Na, 0.1% Тритона-X100, 10 мкг/мл лизоцима в соотношении 10 мл раствора на 1 г клеточной пасты, выдерживали суспензию в течение 30 мин на льду и проводили разрушение клеток ультразвуковым диспергатором до исчезновения видимой вязкости суспензии. Отбирали образцы для электрофоретического анализа, разделяли в них растворимую и нерастворимую фракцию белков центрифугированием в микроцентрифуге, дополнительно ресуспендировали осадок в том же растворе и осаждали центрифугированием. Результаты электрофоретического анализа тотального белка и фракций растворимых и нерасторимых белков для двух колоний штамма BL21[DE3]/p6E-hEK-6 приведены на Фиг.3.
Электрофоретическая подвижность целевого белка соответствует расчетному значению. По данным гель-электрофореза белковых фракций целевой белок практически полностью локализован в нерастворимой фракции белков, т.е. находится в форме «телец включения».
Пример 5. Выделение, очистка, ренатурация и процессинг рекомбинантной легкой цепи энтерокиназы человека.
Штамм-продуцент BL21[DE3]/p6E-hEK-6 высевали из музея петлей истощающим штрихом на чашку Петри с агаризованной средой LB, содержащей 30 мкг/мл канамицина и 1% глюкозы, растили 14 часов при 37°С. Одну отдельную колонию штамма переносили в 5 мл жидкой среды LB, содержащей 30 мкг/мл канамицина и 1% глюкозы, и растили на качалке в течение 14 часов при 37°С. Содержимым инокулировали 250 мл среды 2xYT, содержащей 30 мкг/мл канамицина и 0,1% глюкозы, растили на качалке в течение 3,5 часов при 37°С, отбирали образцы бактериальной суспензии для анализа, добавляли ИПТГ до конечной концентрации 1 мМ и растили в течение еще 3-15 ч.
Осадок клеток отделяли центрифугированием, ресуспендировали в 20 мл раствора А (50 мМ Трис рН=7,4, 2 мМ ЭДТА), добавили лизоцим до 0,1 мг/мл и Тритон X100 до 0,1%, инкубировали в течение 30 мин на льду. Проводили разрушение клеток и геномной ДНК при помощи ультразвукового диспергатора пульсами по 10 с до исчезновения повышенной вязкости суспензии. Отделяли осадок центрифугированием в течение 10 мин при 20000 об/мин. Осадок ресуспендировали в растворе А, добавляли детергент NP-40 до 1%, суспензию обрабатывали ультразвуковым диспергатором до исчезновения частиц осадка крупнее 1 мм, отделяли осадок центрифугированием в течение 10 мин при 20000 об/мин. Полученный осадок ресуспендировали в растворе А, добавляли NaCl до 500 мМ, суспензию обрабатывали ультразвуковым диспергатором до исчезновения частиц осадка крупнее 1 мм, отделяли осадок центрифугированием в течение 10 мин при 20000 об/мин. Полученный осадок ресуспендировали в растворе 50 мМ Трис рН=7,4 суспензию обрабатывали ультразвуковым диспергатором до исчезновения частиц осадка крупнее 1 мм, отделяли осадок центрифугированием в течение 10 мин при 20000 об/мин. Полученный осадок обогащенных телец включения хранили при -70°С.
Для проведения солюбилизации целевого белка к осадку телец включения добавляли раствор Б (8 М мочевины, 50 мМ Трис-HCl, 50 мМ бета-меркаптоэтанола, рН=9,5) в соотношении 10 мл раствора на 1 г осадка. Суспензию инкубировали при перемешивании 2 часа при +37°С, отделяли нерастворившийся клеточный дебрис центрифугированием 10 мин при 20000 об/мин. Супернатант разбавляли в 5 раз раствором В (8М мочевины, 50 мМ фосфата натрия, 500 мМ хлорида натрия, рН=8,0) для уменьшения конечной концентрации бета-меркаптоэтанола, отделяли выпавший осадок центрифугированием в течение 10 мин при 20000 об/мин и наносили супернатант на колонку с сорбентом Chelating Sepharose Fast Flow («GE Healthcare»,CUIA), содержащим хелатированные ионы никеля и уравновешенным раствором В. Последовательно промывали колонку раствором Г (8 М мочевины, 50 мМ фосфата натрия, 500 мМ хлорида натрия, рН 7.0) и раствором Д (8 М мочевины, 50 мМ фосфата натрия, 500 мМ хлорида натрия, 50 мМ имидазола рН=7,0) до стабилизации базовой линии. Элюировали очищенный белок растворами с концентрацией имидазола 100, 200 и 500 мМ и раствором Е (8 М мочевины, 50 мМ фосфата натрия, 500 мМ хлорида натрия, 50 мМ ЭДТА-Na, рН=7,0). Элюат, содержащий очищенный белок-предшественник 6E-hEK-6, концентрировали ультрафильтрацией до конечной концентрации общего белка 20 мг/мл и обессоливали, полученный раствор хранили в замороженном виде. Полипептид был очищен в денатурирующих условиях до видимой гомогенности (более 95% чистоты по денситометрии электрофореграммы, Фиг.4).
Для рефолдинга в растворе к размороженному раствору 6E-hEK-6 добавляли ДТТ до концентрации 10 мМ и инкубировали при 37°С в течение 2 часов. Затем белок разбавляли буферным раствором для рефолдинга (50 мМ ТРИС, рН=8,8, 2 мМ ЭДТА, 2 мМ восстановленного глутатиона, 0,2 мМ окисленного глутатиона; 2 М мочевины) в соотношении 1:40 и инкубировали при комнатной температуре в течение 12-16 ч. После этого белок вновь концентрировали до 2 мг/мл ультрафильтрацией, вносили CaCl2 до 5 мМ, добавляли активную энтерокиназу ("Sigma", США) в молярном соотношении 1:1000 и вели аутокаталитическое отщепление N-концевого пептида в течение 16 ч при комнатной температуре. Полученный препарат зрелого белка hEK-6 обладал 50-90% удельной протеолитической активности в сравнении со стандартом рекомбинантной энтерокиназы ("Sigma", США). Ферментативную активность определяли при помощи синтетического субстрата GDDDDK-β-нафтиламид ("Sigma", США) по методике производителя субстрата.
Пример 6. Расщепление модельного полипептида.
Для проверки ферментативной активности препарата hEK-6 проводили расщепление модельного гибридного белка Trx-MOG, состоящего из N-концевого домена тиоредоксина I (Trx), линкерного участка, содержащего гексагистидиновый кластер и сайт расщепления энтерокиназы, и непосредственно следующий за линкером внеклеточный домен миелинового олигодендроцитарного гликопротеина человека (MOG).
При подборе условий расщепления было установлено, что оптимальными для расщепления Trx-MOG являются условия соотношение фермент:субстрат 1:20 и реакционный буфер следующего состава: 25 мМ NaCl, 20 мМ Tris-HCl pH 7.4, 10 мМ CaCl2, 1 М мочевины. Данный гибридный белок относительно резистентен к воздействию энтерокиназы.
Был проведен гидролиз двух порций Trx-MOG одинаковыми количествами белка hEK-6 и коммерчески доступной рекомбинантной легкой цепью энтерокиназы быка (Sigma, США). Результаты ДСН-ПААГ анализа реакционных смесей приведены на Фигуре 5. Полученные реакционные смеси (r) подвергались очистке при помощи 10 мкл аликвот металлохелатного сорбента Chelating Sepharose Fast Flow, заряженного ионами никеля. Исходный гибридный белок, отщепляемый N-концевой слитой белок-партнер, белок hEK-6 и, предположительно, препарат рекомбинантной энтерокиназы быка содержат гексагистидиновый кластер, поэтому все продукты реакции и исходные белки, кроме свободного продукта реакции MOG, связывались с сорбентом и видны на дорожках «+». Несвязавшийся с сорбентом белок на дорожках «-» представлял собой полностью очищенный свободный MOG.
Приведенные результаты показали, что включение в последовательность синтезируемого целевого белка короткого дополнительного N-концевого пептида, кодируемого оптимальными для Е.coli кодонами, позволяет проводить хроматографическую очистку экспрессируемого белка в денатурирующих условиях методом металлохелатной хроматографии и проводить процедуру рефолдинга для очищенного белка-предшественника с высоким выходом. Такой порядок стадий позволяет получать полностью очищенный и ренатурированный продукт при помощи одной хроматографической стадии очистки. N-концевой пептид отделялся в ходе активации фермента, которая инициировалась внесением малых количеств активной энтерокиназы ("Sigma", США), а затем продолжалась автокаталитически.
В случае использования для расщепления слитых белков полученный активный фермент энтерокиназы человека hEK-6 может быть удален металлохелатной хроматографией вместе с белком-партнером и остатками нерасщепленного слитого белка.
Преимущества предлагаемого штамма Е.coli BL21[DE3] заключаются в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого de novo мутеина легкой цепи энтерокиназы человека и контаминации выделяемого белка наиболее активными протеазами Е.coli. Встроенный в геном штамма-реципиента ген РНК полимеразы бактериофага Т7 под контролем lacUV5 промотора при использовании Т7-lac промотора и Т7 терминатора в плазмидах приводит к быстрой и эффективной продукции белка.
Еще одним общим преимуществом использованного штамма, экспрессионного вектора и стратегии биосинтеза является возможность проводить индукцию без изменения температуры культивирования.
Еще одним преимуществом является возможность получения полностью активной легкой цепи энтерокиназы человека, не содержащей загрязнений белками-партнерами, без дополнительной хроматографической очистки.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.







Изобретение относится к биотехнологии и предоставляет способ получения рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека с использованием бактерии, принадлежащей к роду Escherichia, трансформированной экспрессионной плазмидой, содержащей фрагмент ДНК, кодирующий неактивный предшественник мутеина [C112S] легкой цепи энтерокиназы человека, включающий последовательность, кодирующую отщепляемый N-концевой пептид, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, и слитую с ним в рамке последовательность, кодирующую предшественник мутеина [C112S] легкой цепи энтерокиназы человека с неотщепляемым С-концевым гексагистидиновым кластером под контролем промотора, функционирующего в бактериальной клетке. Предложенное изобретение позволяет получать рекомбинантный мутеин [C112S] легкой цепи энтерокиназы человека с высоким выходом. 5 н. и 6 з.п. ф-лы, 5 ил., 1 табл., 6 пр.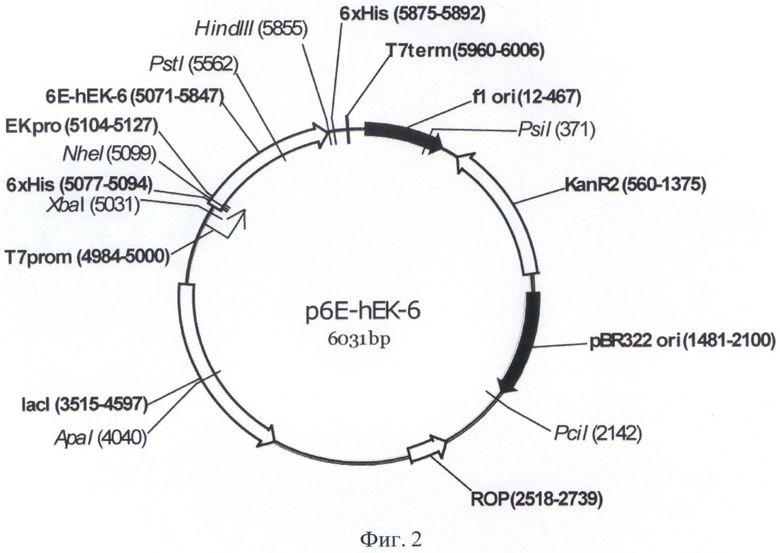
1. Плазмида для экспрессии в клетках бактерии рода Escherichia неактивного предшественника мутеина [C112S] легкой цепи энтерокиназы человека, содержащего отщепляемый N-концевой пептид, включающий гексагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, и неотщепляемый С-концевой гексагистидиновый кластер, под контролем промотора, функционирующего в указанной клетке бактерии, в следующей последовательности по существу содержащая:
- фрагмент вектора рЕТ28а(+), содержащий участок инициации репликации бактериофага f1, последовательность, кодирующую аминогликозид-3'-фосфотрансферазу, область начала репликации плазмиды pBR322, ген РНК-организующего белка Rop;
- последовательность, кодирующую репрессор лактозного оперона;
- фрагмент ДНК, содержащий промотор, функционирующий в бактериальной клетке, и синтетический адаптер, кодирующий отщепляемый N-концевой лидер, включающий гексагистидиновый кластер и последовательность узнавания энтерокиназы, слитую в рамке с последовательностью, кодирующей предшественник мутеина [C112S] легкой цепи энтерокиназы человека;
- фрагмент, кодирующий предшественник мутеина [C112S] легкой цепи энтерокиназы человека с неотщепляемым С-концевым гексагистидиновым кластером; и
- участок терминации транскрипции.
2. Плазмида по п.1, отличающаяся тем, что указанным фрагментом ДНК, кодирующим предшественник мутеина [C112S] легкой цепи энтерокиназы человека с отщепляемым N-концевым пептидом, включающим гексагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, и неотщепляемым С-концевым гексагистидиновым кластером является фрагмент ДНК, представленный в Перечне последовательностей под номером SEQ ID:1, или его вариант.
3. Плазмида по п.1, отличающаяся тем, что указанным промотором является промотор РНК-полимеразы бактериофага Т7.
4. Плазмида по п.1, отличающаяся тем, что указанной плазмидой является плазмида p6E-hEK-6.
5. Бактерия, принадлежащая к роду Escherichia, трансформированная плазмидой по п.1, - продуцент предшественника рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека с отщепляемым N-концевым пептидом, включающим гексагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, и неотщепляемым С-концевым гексагистидиновым кластером.
6. Бактерия по п.5, отличающаяся тем, что указанной бактерией является бактерия Е.Coli BL21[DE3]/p6E-hEK-6.
7. Предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека с отщепляемым N-концевым пептидом, включающим гексагистидиновый кластер и слитую в рамке последовательность узнавания энтерокиназой, и неотщепляемым С-концевым гексагистидиновым кластером, полученный путем культивирования бактерии по п.5 в питательной среде.
8. Предшественник рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека по п.7, содержащий последовательность аминокислот, представленную в Перечне последовательностей под номером SEQ ID NO:2, или ее функциональный вариант.
9. Способ получения рекомбинантного мутеина [C112S] легкой цепи энтерокиназы человека по п.7, включающий культивирование бактерии по п.5 в питательной среде, выделение телец включения, солюбилизацию белка-предшественника, металлохелатную хроматографию в денатурирующих условиях, рефолдинг белка-предшественника, получение зрелого белка обработкой энтерокиназой и выделение указанного зрелого белка мутеина [C112S] легкой цепи энтерокиназы человека, содержащего неотщепляемый С-концевой гексагистидиновый кластер.
10. Способ по п.9, отличающийся тем, что культивируют штамм Е.coli BL21[DE3]/p6E-hEK-6.
11. Рекомбинантный мутеин [C112S] легкой цепи энтерокиназы человека, содержащий неотщепляемый С-концевой гексагистидиновый кластер, полученный способом по п.9.
| RU 2006110371 А, 10.10.2007 | |||
| ПРЕПАРАТ ЧЕЛОВЕЧЕСКОГО ЭНДОСТАТИНА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2004 |
|
RU2278688C1 |
| МОДИФИЦИРОВАННЫЕ ИНГИБИТОРЫ ПРОТЕИНАЗЫ | 1995 |
|
RU2182598C2 |
| KITAMOTO Y | |||
| ET AL, Enterokinase, the initiator of intestinal digestion, is a mosaic protease composed of a distinctive assortment of domains, Proc Natl Acad Sci U S A, 1994, v.91, no.16, pp.7588-7592 | |||
| US 0006746859 B1, 08.06.2004 | |||
| KR 20030097036 A, 31.12.2003 | |||
| CN | |||

