Область техники, к которой относится изобретение: Изобретение относится к области биотехнологии, в частности к разработке способа подготовки клеточного материала для проведения протеомного и фосфопротеомного анализа с использованием J-агрегатов.
Уровень техники: В настоящее время для молекулярного изучения протеома клетки активно используются протеомные и фосфопротеомные технологии, благодаря которым становится возможным оценить статус сигнального пути клетки, в том числе подвергшейся действию вирусной инфекции, и идентифицировать сигнальные белки, задействованные в ответе на любую вирусную инфекцию вне зависимости от таксономической принадлежности вируса. Такие белки являются наиболее перспективной клеточной мишенью для разработки средств антивирусной терапии, однако для проведения анализа методами протеомики и фосфопротеомики необходима отработка методик предобработки клеточных гомогенатов, применяемых для данного анализа.
Как известно, N- и O-связанные олигосахариды являются основными структурными компонентами многих белков, при этом разнообразие олигосахаридных структур, а именно изменение структуры и разная степень насыщенности сайтов гликозилирования в гликопротеине вносят свой вклад в неоднородность по массе, что обуславливает существенные трудности при анализе белков с использованием различных протеомных и фосфопротеомных методов. Поскольку O-связанные олигосахариды обычно меньше по массе, чем N-связанные гликаны, но при этом являются более многочисленными и более гетерогенными по структуре, подготовка биологического материала к протеомному и фосфопротеомному анализу должна включать дегликозилирование O-гликопротеинов. Методы дегликозилирования можно разделить на две группы: ферментативные и химические. Для ферментативного дегликозилирования используют эндо- и экзогликозидазы, которые гидролизуют O-гликозидную связь между моносахаридными остатками олигосахаридов, а также способны расщеплять связь между углеводной частью и серином/треонином белковой части молекулы. В отличие от дегликозилирования N-гликопротеинов, которое проводят с использованием определенной N-гликаназы, и при этом реакция проходит полностью, в случае O-гликозидной связи олигосахаридная цепь может быть удалена только смесью гликозидаз, каждая из которых характеризуется узкой специфичностью. Подобный метод используется в наборе «prO-LINK Extender™ Kit for Complex O-Linked Glycans» фирмы ProZyme Inc., США (http://www.prozyme.com). Также существуют наборы ферментов для одновременного дегликозилирования N- и O-связанных гликопротеинов: набор «GlycoPro™ Enzymatic Deglycosylation Kit for N-Linked & Simple O-Linked Glycans», фирмы ProZyme Inc., США и набор «Enzymatic Protein Deglycosylation Kit» фирмы Sigma-Aldrich Co. LLC, США (http://www.sigmaaldrich.com). В патенте US 20060269980 предлагается протокол ферментативного дегликозилирования N- и O-связанных с олигосахаридами гликопротеинов с использованием набора гликозидаз (ПНГаза F, Эндо H, Эндо F, O-гликозидазы и нейраминидазы) с последующей детекцией продуктов гидролиза масс-спектрометрическими методами, в том числе методом SELDI масс-спектрометрии на аффинных чипах [1].
В патенте US 20100190146 для проведения дегликозилирования N- и O-связанных гликопротеинов и последующего анализа профиля отщепленных гликаганов предлагается микрофлюидный прибор. В конструктив прибора входит дегликозилирующая колонка с твердой фазой на инертном носителе, к которой присоединены дегликозилирующие ферменты для специфичного удаления олигосахаридов с поверхности гликопротеинов. Оценка прохождения реакции и идентификация продуктов дегликозилирования возможна с использованием методов масс-спектрометрии и белковых микрочипов [2].
Основными достоинствами ферментативных методов являются более мягкие условия и отсутствие неспецифических побочных реакций. К недостаткам данной группы методов можно отнести высокую стоимость анализа, необходимость соблюдения строгих условий хранения набора, возможность получения низкого выхода желаемых продуктов из-за стерических затруднений, а также трудоемкость идентификации дегликозилированных белков ввиду полного отщепления олигосахаридных цепей от гликопротеинов.
Для химического дегликозилирования гликопротеинов используют модифицированный метод периодатного окисления, β-элиминирование, а также методы с безводной трифторметансульфокислотой (TFMS) и безводным гидразином. При периодатном окислении происходит разрыв C-C связи в гликолях с образованием альдегидных групп. Существуют две модификации периодатного окисления: метод Смита (заключается в восстановлении окисленного олигосахарида боргидридом натрия с последующим гидролизом полученного полиола; возможно проведение ступенчатой деградации олигосахарида) и метод Бэрри (заключается в избирательном расщеплении окисленного олигосахарида под действием фенилгидразина; происходит расщепление как ацетальных, так и гликозидных связей). К преимуществам периодатного окисления можно отнести количественное расщепление гликольных группировок и протекание реакций в микрообъемах, тогда как к недостаткам - окисление некоторых аминокислот, что создает дополнительные сложности для дальнейшего анализа свойств белка [3].
Гидразинолиз гликопротеинов, несмотря на возможность одновременного удаления N- и O-связанных олигосахаридов, приводит к полной деградации белковой части ввиду неселективной атаки амидных связей гидразином и образования гидразидов соответствующих аминокислот, и поэтому неприменим в тех случаях, когда требуется сохранение структуры белка для проведения дальнейшего анализа. Подобный метод используется в наборе «GlycoRelease™ Glycan Hydrazinolysis Kit» фирмы ProZyme Inc., США.
Метод с использованием трифторметансульфоновой кислоты (TFMS) в смеси с анизолом позволяет неспецифически разрушать N- и O-связанные олигосахариды гликопротеинов без заметной деградации белковой компоненты. Тем не менее даже в оптимизированном варианте метод с TFMS является очень трудоемким, и при этом приводит к неполному удалению сахара: могут оставаться связанные с гликопротеином моносахариды, для удаления которых требуется раскрытие пиранозного кольца посредством обработки периодатом натрия и повторное проведение реакции с TFMS или β-элиминирования для получения полностью дегликозилированного гликопротеина [4]. Подобный метод используется в наборах «GlycoFree™ Chemical Deglycosylation Kit» фирмы ProZyme Inc., США и «GlycoProfile™ IV Chemical Deglycosylation Kit TFMS Deglycosylation System» фирмы Sigma-Aldrich Co. LLC, США.
Дегликозилирование по механизму β-элиминирования посредством обработки гликопротеинов в мягких щелочных условиях позволяет проводить преимущественное разрушение O-связанных олигосахаридов, а именно отщеплять O-гликаны в виде олигозилальдитолов. Проведение реакции β-элиминирования в присутствии восстанавливающего агента позволяет избежать деградации щелочелабильных связей, что делает данный метод практически без недостатков. Подобный метод используется в наборе «GlycoProfile™ β-Elimination Kit» фирмы Sigma-Aldrich Co. LLC, США.
После проведения дегликозилирования необходимо определить паттерн белков, в том числе выяснить, какие из этих белков были первично гликозилированы, поскольку подобная информация позволит более полно охарактеризовать изучаемый объект. В зависимости от выбранного метода дегликозилирования возможны различные варианты идентификации паттерна дегликозилированных и изначально негликозилированных белков. Так, например, после окисления периодатом натрия происходит раскрытие пиранозного кольца, связанного с гликопротеином моносахарида, и образуются альдегидные группы, которые можно идентифицировать фотоколориметрически методом, основанным на восстановлении хлорида 2,3,5-трифенилтетразолия альдегидными группами с образованием красного красителя формазана. Интенсивность окраски полученного раствора определяют на фотоэлектрическом колориметре с зеленым светофильтром либо на спектрофотометре при длине волны 546 нм. Метод позволяет определить суммарное количество гликопротеинов, подвергшихся окислению периодатом натрия, посредством оценки содержания альдегидных групп по градуировочному графику с использованием контрольных растворов гликопротеинов. Подобный метод детекции используется в наборе (Glycoprotein Carbohydrate Estimation Kit» фирмы Thermo Fisher Scientific Inc., США (http://www.thermoscientific.com)). К преимуществам данного метода можно отнести достаточно точные и хорошо воспроизводимые результаты и возможность определения малых количеств альдегидных групп, при этом основным недостатком является возможность определения степени дегликозилирования только в целом, а не применительно к каждому конкретному белку.
Для визуализации паттерна частично дегликозилированных гликопротеинов с моносахаридными остатками распространены реакции с флуоресцентно мечеными производными аминов, гидразина и гидроксиламина в присутствии восстановителя. Среди наиболее широко используемых реагентов можно выделить дансилгидразин и дансилэтилендиамин [5], различные гидразиды биотина [6], а также различные ароматические амины (например, 2-аминопиридин [7]).
В заявке на патент WO 0228841 также предложен новый ряд модифицирующих красителей для флуоресцентного маркирования биомолекул, содержащих функциональные альдегидные группы, которые, в том числе, подходят для флуоресцентного мечения альдегидов, образовавшихся после окисления периодатом натрия моносахаридных остатков гликопротеинов [8].
После проведения реакции образовавшийся аддукт стабилизируют посредством обработки боргидридом или цианоборгидридом натрия и далее методами разделительной хроматографии, электрофореза или преципитации проводят прямую или опосредованную детекцию меченых продуктов [9]. В случае биотин-пероксидазного метода детекцию проводят после иммобилизации на нитроцеллюлозной мембране путем проявления биотинилированных моносахаридных остатков гликопротеинов в системе стрептавидин-щелочная фосфатаза.
После проведения дегликозилирования по механизму β-элиминирования олигосахарид отщепляется от белкового компонента вместе с водородом углеродного атома серина/треонина, что также позволяет проводить дальнейшую модификацию для идентификации паттерна дегликозилированных гликопротеинов. Для маркирования дегликозилированных подобным образом гликопротеинов чаще всего используют дитиотреитол (DTT) и биотин пентиламин (BAP), электрофильное присоединение которых происходит по двойной связи, при этом образуется ковалентная связь между протоном и одним из атомов углерода двойной связи [10]. В случае обработки дегликозилированного гликопротеина DTT образуются стабильные сульфидные аддукты, которые могут быть идентифицированы методами масс-спектрометрии [11]. В свою очередь, использование биотин пентиламина позволяет проводить селективное биотинилирование дегликозилированных гликопротеинов с последующим обогащением с помощью аффинной хроматографии и идентификацией методами жидкостной хроматографии и масс-спектрометрии. Также возможна визуализация биотинилированных моносахаридных остатков гликопротеинов после переноса на мембрану с использованием стрептавидина и щелочной фосфатазы [10].
Техническим результатом изобретения, достигаемым при осуществлении способа согласно формуле, является идентификация белков, меняющих состав или степень своего O-гликозилирования в результате какого-либо физиологического воздействия на клетку. Идентификация осуществляется визуально в форме выбора пятен на двумерных электрофореграммах, окрашенных азотнокислым серебром, которые далее могут быть экстрагированы и использованы для идентификации белка с помощью MALDI-TOF, фосфопротеомного анализа или другим высокочувствительным методом определения последовательности белка.
Раскрытие изобретения: Сущностью изобретения является способ подготовки биологического материала к протеомному и фосфопротеомному анализу посредством дегликозилирования гликопротеинов с последующим введением ковалентной метки и визуализацией паттерна изначально негликозилированных и дегликозилированных белков в предобработанном клеточном гомогенате с использованием J-агрегатов.
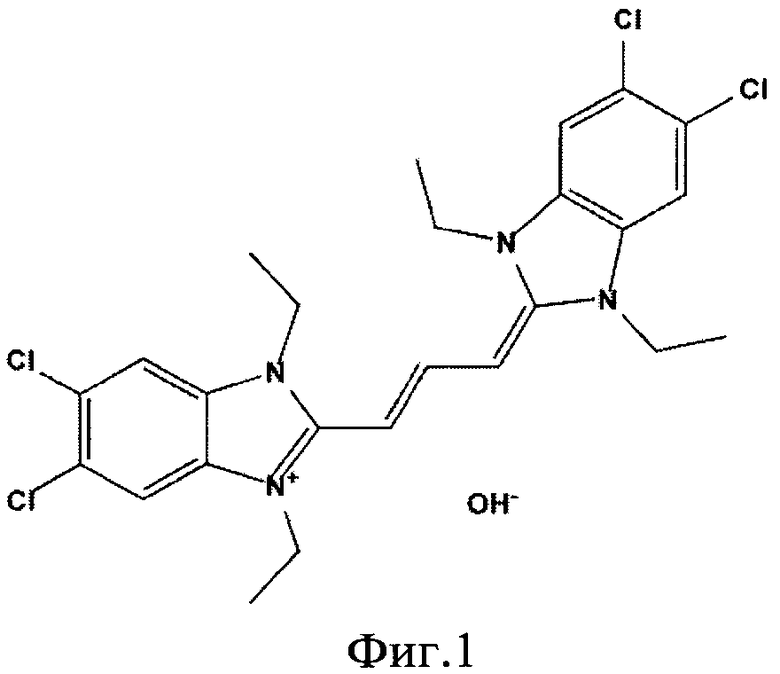
Структурная формула красителя JC-1, используемого в качестве ковалентной метки и базиса для образования J-агрегатов, приведена на Фиг.1. Постадийная схема проведения дегликозилирования белков с введением ковалентной модификации и визуализацией паттерна негликозилированных и дегликозилированных белков в предобработанном клеточном гомогенате с использованием J-агрегатов приведена на Фиг.2.
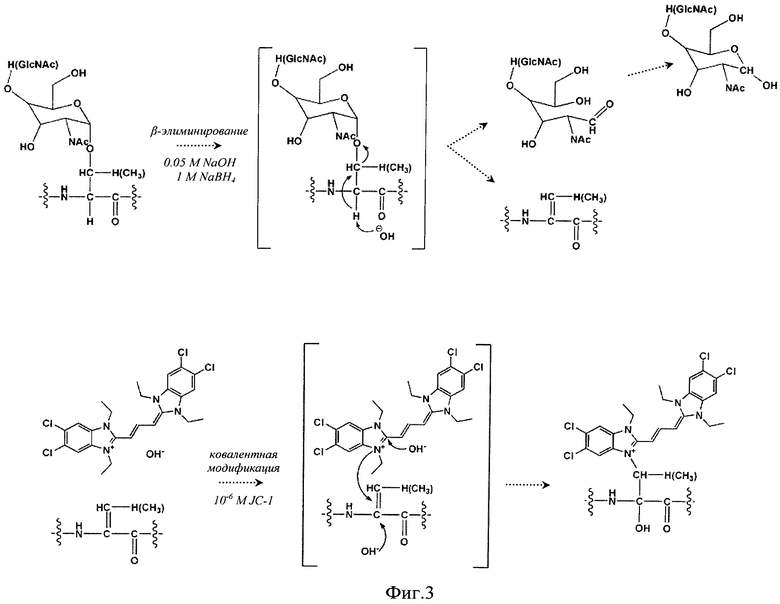
Основным аспектом данного изобретения является применение цианиновых красителей, в частности красителя JC-1, для ковалентной модификации O-дегликозилированных по механизму β-элиминирования гликопротеинов и получения меченого продукта. Схема реакций дегликозилирования гликопротеинов и последующего маркирования дегликозилированного белкового продукта цианиновым красителем с указанием возможных механизмов образования ковалентных связей приведена на Фиг.3.
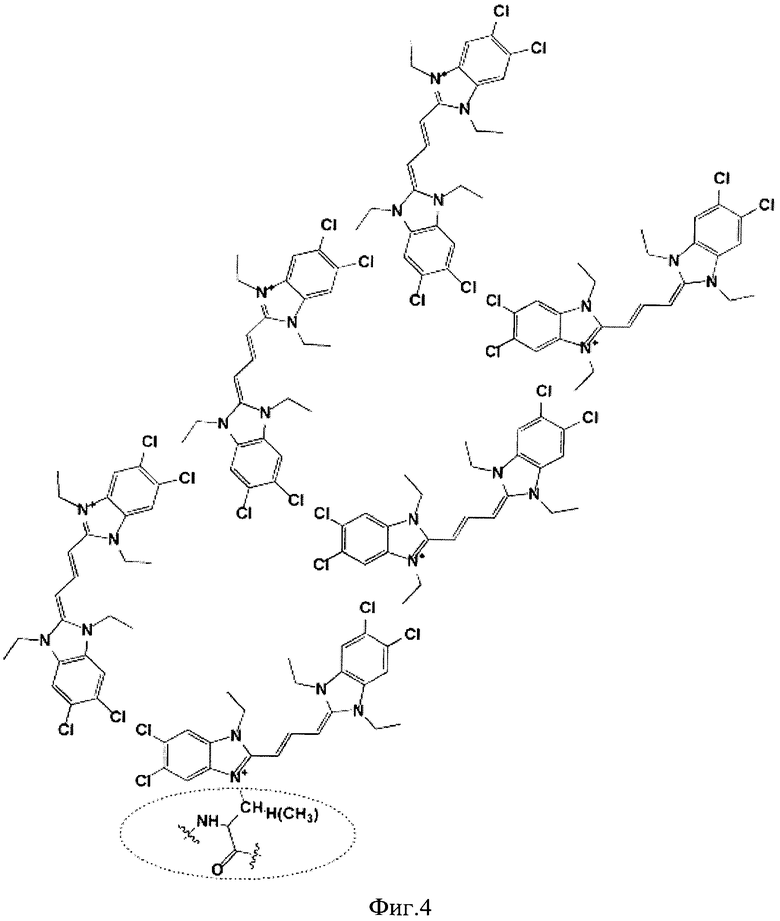
Другим аспектом данного изобретения является нековалентное мечение первично модифицированных красителем дегликозилированных белков, в частности красителем JC-1, высокими концентрациями того же красителя посредством формирования супрамолекулярных J-агрегатов. Модель упаковки J-агрегатов, сформированных на базе ковалентно-модифицированных красителем дегликозилированных гликопротеинов, при высокой концентрации цианинового красителя JC-1 приведена на Фиг.4.
Метод основан на способности цианиновых красителей к образованию супрамолекулярных структур, а именно сложных полимолекулярных ассоциатов - J-агрегатов. Формирование J-агрегатов происходит в результате адсорбции краски на поверхности и реализуется путем самосборки в буферном растворе при высокой концентрации красителя в присутствии ионов металлов или полимеров [12]. Известно, что образование J-агрегатов тиокарбоцианиновых красителей индуцируется добавлением желатина и других биологически активных молекул с положительно заряженными группами, с которыми посредством электростатического взаимодействия связываются анионы красителя и происходит дальнейшая самосборка J-агрегатов [13]. При использовании тиокарбоцианиновых красителей для эффективного формирования J-агрегатов необходимым условием является присутствие моновалентных катионов, добавление которых обуславливает снижение электростатического отталкивания молекул красителя и, таким образом, способствует ускорению процессов агрегации [14]. В отличие от тиокарбоцианиновых красителей JC-1 краситель представляет собой липофильный катион 5,5', 6,6'-тетрахлоро-1,1,3,3'-тетраэтилбензимидазолил-карбоцианин, который также широко используется в качестве люминесцентного зонда для биомедицинского применения, в том числе для оценки мембранного потенциала митохондрий, благодаря практически однозначному изменению спектральных характеристик в зависимости от концентрации красителя и сформированной структуры [12, 15]. При низкой концентрации красителя JC-1 в спектре люминесценции (λвозб=490 нм) наблюдается интенсивная полоса мономеров при длине волны 530 нм, тогда как увеличение концентрации JC-1 приводит к смещению пика в длинноволновую область спектра до 595 нм, при этом по мере увеличения концентрации красителя пик также сужается, что свидетельствует не только о формировании и увеличении размеров J-агрегатов JC-1, но и об улучшении их структуры [12, 15]. Так, например, после проведения электрофоретического анализа паттерна всех белков и окрашивания пластины ПААГ азотнокислым серебром вторичное окрашивание модифицированных красителем дегликозилированных гликопротеинов приводит к образованию J-агрегатов, формирование которых определяется методом флуоресцентной детекции с использованием трансиллюминатора синего цвета со светофильтром янтарного цвета Safe Imager™ 2.0 фирмы Life Technologies, США (http://www.lifetechnologies.com) или UVIblue фирмы Uvitec Cambridge, Великобритания (www.uvitec.со.uk) и визуально проявляется в виде светящихся в темноте полос соответствующих дегликозилированных гликопротеинов. Гель-документацию сформированных молекулярных цепочек J-агрегатов на базе ковалентно-модифицированных красителем JC-1 дегликозилированных гликопротеинов проводят с помощью цифровой фотокамеры Panasonic Lumix DMC-GF2C в темной комнате. Подобная простая визуальная детекция паттерна изначально негликозилированных белков и ковалентно-модифицированных дегликозилированных гликопротеинов делает разработанный способ анализа применимым для подготовки биологического материала к протеомному и фосфопротеомному анализу.
Примеры анализа результатов, полученных после проведения двумерного электрофореза по О'Фареллу негликозилированных и дегликозилированных белков в гомогенате клеток, зараженных модельным штаммом вируса клещевого энцефалита ЭК-328 или вариантом М штамма ЭК-328, до проведения дегликозилирования гликопротеинов, а также после дегликозилирования и ковалентной модификации красителем JC-1 без обработки пластины ПААГ и с обработкой раствором того же красителя в высокой концентрации с образованием J-агрегатов приведены на Фиг.5.
Краткое описание графических изображений:
Фиг.1. Структурная формула красителя JC-1, используемого в качестве ковалентной метки и как базиса для образования J-агрегатов.
Фиг.2. Постадийная схема преобработки гомогенатов клеток, зараженных модельными штаммами вирусов, посредством проведения дегликозилирования гликопротеинов с последующим введением ковалентной метки и визуализацией паттерна негликозилированных и дегликозилированных белков в предобработанном клеточном гомогенате с использованием J-агрегатов.
Фиг.3. Схема реакций дегликозилирования гликопротеинов по механизму β-элиминирования и последующего маркирования дегликозилированного белкового продукта цианиновым красителем JC-1 с указанием возможных механизмов образования ковалентных связей.
Фиг.4. Модель упаковки J-агрегатов, сформированных на базе ковалентно-модифицированных красителем дегликозилированных гликопротеинов, при высокой концентрации цианинового красителя JC-1.
Фиг.5. Электрофоретический анализ паттерна негликозилированных и дегликозилированных белков в гомогенате клеток СПЭВ, зараженных модельным штаммом вируса клещевого энцефалита ЭК-328 (1-3) или вариантом М штамма ЭК-328 (4-6): до проведения дегликозилирования гликопротеинов (1, 4), после дегликозилирования и ковалентной модификации гликопротеинов красителем JC-1 без обработки пластины ПААГ (2, 5) и с обработкой раствором цианинового красителя JC-1 с образованием упорядоченных молекулярных цепочек J-агрегатов (3, 6). Белки разделены методом 20-электрофореза по О'Фареллу и окрашены на общий белок азотнокислым серебром с использованием набора «PageSilver™ Silver Staining Kit». Стрелками указаны полосы, соответствующие дегликозилированным гликопротеинам.
Осуществление изобретения: На основе метода O-дегликозилирования гликопротеинов по механизму β-элиминирования [10, 11] был разработан и оптимизирован способ идентификации паттерна первично негликозилированных и дегликозилированных белков в предобработанном клеточном гомогенате посредством ковалентного связывания дегликозилированных гликопротеинов с цианиновым красителем JC-1 [12], электрофильное присоединение которого, как и в случае с DTT и ВАР, происходит по двойной связи [10]. До и после дегликозилирования гликопротеинов по механизму β-элиминирования с ковалентной модификацией и мечением дегликозилированных белков цианиновым красителем JC-1 был проведен 2D-электрофорез по О'Фареллу с последующей визуализацией паттерна изначально негликозилированных и модифицированных красителем дегликозилированных белков посредством окрашивания гелей азотнокислым серебром с использованием набора «PageSilver™ Silver Staining Kit» фирмы Thermo Fisher Scientific Inc., США (http://www.fermentas.com). После идентификации паттерна всех белков посредством оценки гель-документированных изображений двумерных электрофореграмм методом сканирования на сканере Epson expression 1680 или с помощью цифровой фотокамеры Panasonic Lumix DMC-GF2C с целью подтверждения проявления на пластине ПААГ полос, соответствующих именно дегликозилированным гликопротеинам, гель был помещен в горизонтальную электрофорезную камеру, и было проведено окрашивание первично меченых дегликозилированных белков посредством добавления цианинового красителя JC-1 в высокой концентрации с образованием упорядоченных молекулярных цепочек J-агрегатов.
Ковалентная модификация дегликозилированных белков может быть осуществлена с использованием других цианиновых красителей, в том числе окса- и тиацианиновых красителей [16, 17], способных к образованию супрамолекулярных J-агрегатов.
Повторная идентификация с использованием трансиллюминатора синего цвета со светофильтром янтарного цвета Safe Imager™ 2.0 («Life Technologies), США) или UVIblue («Uvitec Cambridge), Великобритания) и цифровой фотокамеры Panasonic Lumix DMC-GF2C в темной комнате с последующим наложением полученных гель-документированных изображений позволяет быстро и с удобным способом визуальной детекции определить паттерн изначально негликозилированных белков и ковалентно-модифицированных дегликозилированных белков, что делает данный метод оптимальным для использования при подготовке биологического материала к протеомному и фосфопротеомному анализу.
Далее изобретение иллюстрируется примерами, которые показывают применение предлагаемого способа подготовки биологического материала к протеомному и фосфопротеомному анализу, основанного на предобработке гомогенатов клеток, в том числе зараженных модельными штаммами вирусов, а именно дегликозилировании содержащихся в них гликопротеинов и последующем введении ковалентной метки с визуализацией паттерна негликозилированных и дегликозилированных белков с использованием J-агрегатов JC-1. Следует понимать, что приводимые примеры служат исключительно для иллюстрации и не предназначены для ограничения объема притязаний, выраженных в формуле изобретения. На основании настоящего описания специалист в данной области сможет легко предложить свои варианты и модификации осуществления изобретения, не отходя от общей концепции настоящего изобретения и без привлечения собственной изобретательской деятельности, так что должно быть понятно, что такие варианты и модификации также будут входить в объем притязаний настоящего изобретения.
Пример 1. Приготовление лизата клеток, зараженных модельным штаммом вируса клещевого энцефалита ЭК-328 или вариантом М, полученным из штамма ЭК-328.
Суспензию клеток почки эмбриона свиньи СПЭВ после заражения штаммом вируса клещевого энцефалита ЭК-328 или вариантом М штамма ЭК-328, и обработки тщательно перемешивают на ротаторе и отбирают аликвоты следующего объема:
При титре 1:512-0,25 мл,
При титре 1:256-0,5 мл,
При титре 1:128-1 мл.
К клеточным суспензиям в среде DMEM добавляют 100 мг стеклянных шариков диаметром 0,8 мм, 10 мкл на мл коктейля ингибиторов протеолиза «Halt Protease Inhibitor Cocktail» («Thermo Fisher Scientific Inc.», США) и подвергают интенсивной обработке на встряхивателе ВП1-2: 10 серий по 30 сек для каждого препарата, поддерживая температуру препарата около 0°C путем инкубации на ледяной бане. Затем к суспензиям добавляют детергент Triton Х-100 до конечной концентрации 1% и инкубируют на льду при периодическом встряхивании в течение 3 часов.
Суспензии декантируют, осветляют посредством центрифугирования при 2000 g в течение 30 мин и замораживают в жидком азоте для дальнейшего использования.
Пример 2. Предобработка клеточных гомогенатов: проведение O-дегликозилирования гликопротеинов по механизму β-элиминирования с последующей ковалентной модификацией дегликозилированных белков цианиновым красителем JC-1.
В качестве буферного раствора для клеточного гомогената используют буферный раствор со слабощелочным pH 7,2 на основе 100 мМ Tris-HCl с добавлением 100 мМ NaCl. Образец объемом 400 мл предварительно концентрируют на ячейке для тангенциальной ультрафильтрации («Amicon», США). Первичный концентрат диализуют против буферного раствора Tris-HCl 100 мМ, NaCl 100 мМ в течение суток при +4°C: 75 мл образца наносят в 3 приема по 25 мл на гель-фильтрационную колонку с Sepharose 6 FastFlow (буфер элюции содержит 100мМ Tris-HCl и 100мМ NaCl, pH 7,2). Условия подобраны таким образом, что большинство примесей белковой и небелковой природы не способны связываться с сорбентом. Вирусный белок выходит из колонки в свободном объеме, равном ~ 66% от общего объема колонки. Для дальнейшей работы используют 3 фракции первого пика.
К фракции первого пика вирусного материала объемом 0,5 мл добавляют 0,5 мл 0,1 М раствора NaOH и 38 мг NaBH4. Смесь тщательно перемешивают и нагревают 16 часов при +45°C. После этого к смеси добавляют 10 мкл 10-4 М цианинового красителя JC-1, растворенного в бинарном растворе ацетон-Tris-HCl (C=0,05 М, pH=8,5), и инкубируют 15 мин при комнатной температуре (краситель JC-1 предоставлен лабораторией процессов фотосенсибилизации Института биохимической физики имени Н.М. Эмануэля РАН (ИБХФ РАН)). После завершения инкубации вирусный препарат подвергают окончательной очистке методом катионообменной хроматографии на Sepharose 6 Fast Flow при pH 7,2; очищенный белок элюируют из колонки буферным раствором с содержанием 100 мМ Tris-HCl и 100 MM NaCl.
Пример 5. Анализ белков из первичного и предобработанного препаратов вирусного гомогената с помощью 20-электрофореза.
Для изучения белкового состава препаратов, полученных до и после проведения O-дегликозилирования гликопротеинов по механизму β-элиминирования с последующим мечением дегликозилированных белков цианиновым красителем JC-1, используют 2D-электрофорез по О'Фареллу. Для этого аликвоты хроматографических фракций объемом 500 мкл, с содержанием общего белка около 100 мкг, осаждают добавлением 100% ацетона (1:1) в течение 30 мин при +4°C и последующим центрифугированием на настольной центрифуге при 14000 об/мин в течение 15 мин. Образовавшийся осадок обессоливают, промывают 1 раз 100 мкл 50% водного ацетона, и солюбилизируют, прогревая при 99°C в течение 10 мин в 50 мкл буфера на основе мочевины 9 М, содержащем в качестве восстановителя 1 М тиомочевину, для денатурации образцов.
Полученный денатурированный образец центрифугируют на настольной центрифуге при максимальных оборотах в течение 10 мин, после чего осветленный супернатант используют для проведения изофокусировки в трубке. Фракционирование в первом направлении представляет собой изоэлектрофокусирование (ИЭФ) в стеклянных трубках (2,4×180 мм), заполненных 4% ПААГ, приготовленном на 9М растворе мочевины, содержащем 2% тритона Х-100 и 2% смеси амфолинов (pH 5-7, 5-8 и 3,5-10 в соотношении 4/1). Белковый экстракт (от 100 до 150 мкл) наносят на «кислотный» край геля и проводят ИЭФ в приборе фирмы BioRad (США), до достижения 2400 В/час суммарно на каждую колонку ПААГ. Затем колонки ПААГ с разделенными в ходе ИЭФ белками используют в качестве стартовой зоны при фракционировании во втором направлении, которое проводят электрофорезом в пластинах ПААГ (200×200×1 мм) с линейным градиентом концентрации акриламида 7,5-25% и в присутствии 0,1% SDS на приборе для вертикального электрофореза фирмы Helicon (Россия). С края каждой пластины формируют карман для нанесения белков-маркеров. Для визуализации белков пластины ПААГ окрашивают азотнокислым серебром с использованием набора «PageSilver™ Silver Staining Kit» в соответствии с инструкцией производителя («Тhеrmо Fisher Scientific Inc.», США).
Гель-документацию полученных двумерных электрофореграмм проводят методом сканирования на сканере «Epson expression 1680» или с помощью цифровой фотокамеры Panasonic Lumix DMC-GF2C. Наложение гель-документированных изображений, полученных после проведения электрофоретического анализа паттерна первично негликозилированных белков (Фиг.5 (1, 4)) и паттерна всех белков (Фиг.5 (2, 5)) позволяет определить возможный паттерн изначально негликозилированных и ковалентно-модифицированных дегликозилированных белков без возможности подтверждения или опровержения артефактности проявившихся на пластине ПААГ полос, предположительно соответствующих именно дегликозилированным гликопротеинам.
Пример 4. Идентификация паттерна негликозилированных и дегликозилированных белков из очищенного предобработанного гомогената клеток, зараженных модельным штаммом вируса клещевого энцефалита ЭК-328 или вариантом М, полученным из штамма ЭК-328.
Для однозначной идентификации паттерна изначально негликозилированных и дегликозилированных белков из предобработанного препарата вирусного гомогената после проведения электрофоретического анализа всех белков и окрашивания геля азотнокислым серебром проводят вторичное окрашивание модифицированных красителем дегликозилированных гликопротеинов. Для этого окрашенную азотнокислым серебром пластину ПААГ помещают в камеру для горизонтального электрофореза multiSUB MAXI («Сleaver Scientific Ltd», Великобритания, www.cleaverscientific.com), добавляют 1200 мл 10-5 M цианинового красителя JC-1 в бинарном растворе ацетон-Tris-НСl (C=0,05 М, pH=8,5), содержащем также предварительно растворенный ПАВ цетилпиридиний бромид в концентрации 10-3 М, добавление которого приводит к образованию оболочки из молекул ПАВ вокруг J-агрегатов, обуславливая более эффективную структуризацию, и выдерживают 10 мин при комнатной температуре. После завершения инкубации раствор для окрашивания сливают, гель промывают охлажденным до +4°C раствором PBS и документируют формирование молекулярных цепочек J-агрегатов на базе ковалентно-модифицированных красителем дегликозилированных гликопротеинов с помощью трансиллюминатора синего цвета со светофильтром янтарного цвета Safe Imager™ 2.0 («Life Technologies», США) или UVIblue («Uvitec Cambridge», Великобритания) и цифровой фотокамеры Panasonic Lumix DMC-GF2C в темной комнате (Фиг.5 (3, 6)).
Наложение гель-документированных изображений, полученных после проведения электрофоретического анализа паттерна первично негликозилированных белков (Фиг.5 (1, 4)) и паттерна всех белков (Фиг.5 (2, 5)) после окрашивания геля азотнокислым серебром, а также паттерна модифицированных красителем дегликозилированных гликопротеинов после вторичного окрашивания (Фиг.5 (3, 6)), которое приводит к образованию J-агрегатов, позволяет однозначно определить паттерн изначально негликозилированных и ковалентно-модифицированных дегликозилированных белков.
Список использованных источников
1. Gibbs B.F. Complete chemical and enzymatic treatment of phosphorylated and glycosylated proteins on protein chip arrays. Дата приоритета 11.05.2005. Патент US 2006269980 (A1).
2. Bynum M.A., Grimm R., Killeen K.P., Robotti K.M. Microfluidic Glycan Analysis. Дата приоритета 29.01.2009. Патент US 20100190146 (A1).
3. Виноградов А.А., Ямсков И.А. Дегликозилирование гликопротеинов. Биоорганическая химия. - 1998. - Т. 24. - №11. - С.803-815.
4. Edge A.S. Deglycosylation of glycoproteins with trifluoromethanesulphonic acid: elucidation of molecular structure and function. - 2003. Biochemical Journal. - V. 376. - P. 339-350.
5. Ingham K.C., Brew S.A. Fluorescent labeling of the carbohydrate moieties of human chorionic gonadotropin and alpha 1-acid glycoprotein. Biochimica et Biophysica Acta. - 1981.- V. 670. -№2. - P. 181-189.
6. De Bank P.A., Kellam В., Kendall D.A., Shakesheff K.M. Surface engineering of living myoblasts via selective periodate oxidation. Biotechnology and Bioengineering. - 2003. - V. 81. - №7. - P. 800-808.
7. Shilova N.V., Galanina O.E., Rubina A.Y., Butvilovskaya V.I., Huflejt M.E., Chambers J., Roucoux A., Bovin N.V. 2-Aminopyridine-a label for bridging of oligosaccharides HPLC profiling and glycoarray printing. Glycoconjugate Journal. - 2008. - V. 25. - №1. - P. 11-14.
8. Haugland R.P., Steinberg Т.Н., Patton W.P., Zhenjun D. Reagents for labeling biomolecules having aldehyde or ketone moieties. Дата приоритета 02.10.2000. Заявка на патент WO 0228841 A2.
9. Sinkeldam R.W., Greco N.J., Tor Y. Fluorescent analogs of biomolecular building blocks: design, properties, and applications. Chemical Reviews. - 2010. - V. 110. - №5. - P. 2579-2619.
10. Wells L., Vosseller K., Cole R.N., Cronshaw J.M., Matunis M.J., Hart G.W. Mapping sites of O-GlcNAc modification using affinity tags for serine and threonine post-translational modifications. Molecular & Cellular Proteomics. - 2002. - V. 1. - №10. - P. 791-804.
11. Murrey H.E., Hsieh-Wilson L.C. The chemical neurobiology of carbohydrates. Chemical Reviews. - 2008. - V. 108. - №5. - P. 1708-1731.
12. Сорокин А.В. Оптические свойства и структура J-агрегатов красителя JC-1 в растворах. Биофизический вестник. - 2008. - Т. 21. - №2. - С.115-121.
13. Slavnova T.D., Gorner Н., Chibisov А.К. J-aggregation of anionic ethyl meso-thiacarbocyanine dyes induced by binding to proteins. Journal of Physical Chemistry. - 2007. - V. 111. - №33. - P. 10023-10031.
14. Yao H., Isohashi Т., Kimura K. Electrolyte-induced mesoscopic aggregation of thiacarbocyanine dye in aqueous solution: counterion size specificity. Journal of Physical Chemistry. - 2007. - V. 111. - №25. - P. 7176-7183.
15. Reers M., Smith T.W., Chen L.B. J-aggregate formation of a carbocyanine as a quantitative fluorescent indicator of membrane potential. Biochemistry. - 1991. - V. 30. - №18. - P. 4480-4486.
16. Yamaguchi A., Kometani N., Yonezawa Y. Luminescence properties of the mixed J-aggregate of oxacyanine dye and thiacyanine dye. Formation of a persistence-type aggregate. Journal of Physical Chemistry. - 2005. - V. 109. - №4. - P. 1408-1414.
17. Achyuthan K.E., Lu L., Lopez G.P., Whitten D.G. Supramolecular photochemical self-assemblies for fluorescence "turn on" and "turn off assays for chem-bio-helices. Photochemical & Photobiological Sciences. - 2006.- V. 5. - №10.- P. 931-937.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ФЕРМЕНТАТИВНО-МЕЧЕННОГО АНТИГЕНА G2 ХАНТАВИРУСА В КЛЕТКАХ E.coli С ЦЕЛЬЮ ЕГО ПРИМЕНЕНИЯ В ИММУНОФЕРМЕНТНОМ АНАЛИЗЕ ПРИ ДИАГНОСТИКЕ ГЛПС | 2012 |
|
RU2539836C2 |
| ПОЛУЧЕНИЕ IN VIVO N-ДЕГЛИКОЗИЛИРОВАННЫХ РЕКОМБИНАНТНЫХ БЕЛКОВ ПОСРЕДСТВОМ СОВМЕСТНОЙ ЭКСПРЕССИИ С ENDO H | 2015 |
|
RU2741347C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ВИРУСНЫХ ЧАСТИЦ С УПРОЩЕННЫМ ГЛИКОЗИЛИРОВАНИЕМ ПОВЕРХНОСТНЫХ БЕЛКОВ | 2011 |
|
RU2607452C2 |
| МЕТОД ПОЛУЧЕНИЯ БИБЛИОТЕК СЕРИЙНЫХ ДВУХСТОРОННИХ ДЕЛЕЦИЙ С ПОМОЩЬЮ ПЦР С ВЫРОЖДЕННЫМ ПРАЙМЕРОМ | 2012 |
|
RU2511424C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИГЕНА G2 ХАНТАВИРУСА ДОБРАВА В КЛЕТКАХ E.coli | 2012 |
|
RU2509805C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИГЕНА G2 ХАНТАВИРУСА ДОБРАВА В КЛЕТКАХ E. coli | 2011 |
|
RU2495938C2 |
| СПОСОБ ПЕРСОНИФИЦИРОВАННОГО ПОДБОРА АНТИАГРЕГАНТНЫХ ПРЕПАРАТОВ БОЛЬНЫМ, НУЖДАЮЩИМСЯ В ПОДОБНОМ ЛЕЧЕНИИ | 2012 |
|
RU2499261C1 |
| Рекомбинантный фрагмент ДНК, кодирующий альфа-фетопротеин (АФП) человека, содержащий "медленные" кодоны, кодирующие лейцин, экспрессионная плазмида, содержащая указанный фрагмент, клетка Saccharomyces cerevisiae, трансформированная указанной плазмидой, и способ получения рекомбинантного АФП человека | 2015 |
|
RU2617943C2 |
| СПОСОБ ДИАГНОСТИКИ СИФИЛИСА С ИСПОЛЬЗОВАНИЕМ МЕТОДА ПРЯМОГО ПРОТЕОМНОГО ПРОФИЛИРОВАНИЯ СЫВОРОТКИ КРОВИ | 2008 |
|
RU2381505C1 |
| ФРАГМЕНТ ГЕНА ENV HTLV-1, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PATLE 495, ШТАММ БАКТЕРИЙ ЕSСHERICHIA COLI HB101/PATLE 495-ПРОДУЦЕНТ ПОЛИПЕПТИДА Е 495 | 1997 |
|
RU2152955C2 |

Изобретение относится к области биотехнологии и касается способа выявления O-гликозилированных белков в составе клеточных гомогенатов, подготавливаемых к протеомному и фосфопротеомному анализу. Предложенное изобретение может быть использовано для проведения протеомного и фосфопротеомного анализа. Способ включает проведение двумерного электрофореза с последующей идентификацией пятен методами спектроскопии MALDI-TOF или фосфопротеомики. Проводят обессоливание клеточных гомогенатов методом гель-хроматографии или диализа клеточных гомогенатов. Подвергают клеточные гомогенаты дегликозилированию по принципу β-элиминирования в растворе 0,05 М NaOH, который содержит 38 мг/мл NaBH4, в течение 16 часов при +45°C с последующим добавлением цианинового красителя JC-1 в концентрации 10-6 М. Инкубируют клеточные гомогенаты в течение 15 мин при комнатной температуре. Концентрируют гомогенаты осаждением 50% ацетоном, подвергают двумерному электрофорезу с образованием электрофореграмм, анализируемых на флуоресценцию при облучении на трансиллюминаторе синего цвета со светофильтром янтарного цвета, визуально проявляющуюся в виде светящихся в темноте полос. Данные полосы экстрагируют из геля и используют для проведения протеомного или фосфопротеомного анализа. Дополнительный анализ интенсивности и расположения экстрагируемых полос выполняют за счет сравнения окрашенных азотнокислым серебром электрофореграмм гомогенатов до и после процедуры дегликозилирования. Предложенное изобретение позволяет идентифицировать белки, меняющие состав или степень своего O-гликозилирования в результате какого-либо физиологического воздействия на клетку. 5 ил.
Способ выявления O-гликозилированных белков в составе клеточных гомогенатов, подготавливаемых к протеомному и фосфопротеомному анализу методом двумерного электрофореза с последующей идентификацией пятен методами спектроскопии MALDI-TOF или фосфопротеомики, отличающийся тем, что тщательно обессоленные методом гель-хроматографии или диализа клеточные гомогенаты подвергаются дегликозилированию по принципу β-элиминирования в растворе 0,05 М NaOH, содержащего 38 мг/мл NaBH4, в течение 16 часов при +45°C с последующим добавлением цианинового красителя JC-1 в концентрации 10-6 М и инкубацией в течение 15 мин при комнатной температуре, концентрируются осаждением 50% ацетоном, подвергаются двумерному электрофорезу с образованием электрофореграмм, анализируемых на флуоресценцию при облучении на трансиллюминаторе синего цвета со светофильтром янтарного цвета, визуально проявляющуюся в виде светящихся в темноте полос, которые могут быть экстрагированы из геля и использованы для проведения протеомного или фосфопротеомного анализа, при этом дополнительный анализ интенсивности и расположения экстрагируемых полос может быть выполнен за счет сравнения окрашенных азотнокислым серебром электрофореграмм гомогенатов до и после процедуры дегликозилирования.
| US 20100190146 A1, 29.07.2010 | |||
| MURREY H.E | |||
| ET AL., The chemical neurobiology of carbohydrates | |||
| Chemical Reviews, 2008, v.108, no.5, p.1708-1731 | |||
| WELLS L | |||
| ET AL., Mapping sites of O-GlcNAc modifi cation using affinity tags for serine and threonine posttranslational modifications, Molecular & Cellular Proteomics, 2002, v.1, no.10, p.791-804. |

