Область техники, к которой относится изобретение: Изобретение относится к области биотехнологии, в частности, к получению вакцины для профилактики вирусных инфекций, а именно инфекции геморрагической лихорадки с почечным синдромом.
Уровень техники: геморрагическая лихорадка с почечным синдромом (ГЛПС) - вирусный нетрансмиссивный зооноз, занимает в России ведущее место по числу заболевших среди природно-очаговых инфекций.
Наиболее эффективным методом борьбы с ГЛПС является специфическая профилактика, то есть вакцинация населения эндемичных регионов [1]. Инактивированная вакцина Hantavax [2] была разработана для применения в республике Корея, где доминируют виды ханавирусов Ханаан и Сеул. В настоящее время вакцина под маркой Hantavax производится промышленно и коммерчески доступна. Она разрешена для клинического применения в республике Корея.
Наряду с Hantavax в Китае и в Южной Корее разработаны, лицензированы и активно используются для профилактики ГЛПС другие варианты убитых вакцин против ГЛПС:
1) В Китае производится инактивированная формалином вакцина на основе вируса Сеул, выращенного на первичной культуре клеток почек сирийского хомячка; для инактивации вируса при производстве этой вакцины используется β-пропиолактон.
2) Также в Китае производится вакцина на основе вируса Хантаан, выращенного на первичной культуре клеток почек монгольской песчанки, инактивированная формалином.
3) Очищенная вакцина на основе вируса Хантаан, полученного из мозга экспериментально зараженных крыс-сосунков [3].
4) В настоящее время в Китае также разработана и проходит клинические испытания бивалентная вакцина на основе вирусов Хантаан и Сеул, полученных на перевиваемых культурах клеток Vero-Е6 (инактивация β-пропиолактоном). По мнению разработчиков, данный препарат должен заменить используемые инактивированные вакцины, поскольку последние в силу низкой степени очистки вызывают серьезные побочные эффекты, не обеспечивая при этом максимального уровня синтеза нейтрализующих антител.
Тем не менее, ни одна из этих вакцин не может применяться в европейских регионах России, так как не обеспечивает защиты от вирусов Пуумала и Добрава - возбудителей ГЛПС в этих регионах [4-10].
Для изготовления и контроля вакцины против вирусов Пуумала и Добрава необходимо решить проблему разработки эффективных серологических методов на основе белков внешней оболочки - энвелопа хантавирусов. Однако до настоящего времени задача получения полноразмерных коммерческих препаратов белков энвелопа G1 и G2 хантавирусов не решена нигде в мире, что обусловлено их токсичностью по отношению к бактериальным клеткам.
Ранее был получен рекомбинантный антиген G2 хантавирусов НТ-Δ12 [11]. Однако выход этого белка не превышал 20 мг на литр культуры, что в совокупности с потерями во время очистки препарата не позволяет использовать данный антиген в иммунологических тестах.
Целью нашей работы является разработка способа увеличения количества накопления продукта в штамме продуцента за счет изменения структуры целевого белка.
Раскрытие изобретения: сущностью изобретения является способ повышения выхода рекомбинантного антигена G2 хантавирусов в клетках E.coli за счет оптимизации структуры антигена до трехфункционального производного пептида НТ.
На основе конструкции рЕТ-НТ-Δ12 [11], кодирующей пептид НТ в изолированном состоянии, была спроектирована и получена конструкция, где этот пептид находился в составе трифункционального слитого белка. При этом N-концевое положение занимал зеленый флуоресцентный белок GFP, центральное - пептид НТ, а С-концевое - легкая цепь двуцепочечного белкового ингибитора Кунитца из клубней картофеля (PKPI-B1). При сборке конструкций вместо вектора рЕТ23 на основе промотора Т7 был использован pQE30, содержащий менее мощный промотор Т5. Это решение было продиктовано стремлением улучшить условия фолдинга продукта за счет снижения интенсивности его накопления. С той же целью на границы доменов GFP, НТ и PKPI-B1 конструируемого трифункционального слитого белка были введены искусственные гибкие пептидные линкеры, обогащенные Gly, Ser и Ala, кодируемые синтетическими олигонуклеотидами. Кроме того, при конструировании новой плазмиды ген пептида НТ был укорочен на 40 п.н. с 3'-конца, что не сказывается на иммунохимических свойствах белка, но позволяет сократить в нем число дисульфидных связей и гидрофобных участков, что способствует улучшению растворимости продукта и ослабляет его негативное влияние на жизнеспособность клеток продуцента.
Краткое описание графических изображений:
Рис.1. Плазмидная конструкция рНК6, принципиальная схема.
Рис.2. Последовательность трифункционального слитого гена, кодирующего производное пептида НТ, в составе конструкции рНК6.
• Последовательность гена GFP подчеркнута волнистой линией;
• Последовательность пептида НТ из состава кДНК вируса Добрава 
• легкая цепь ингибитора Кунитца из клубней картофеля 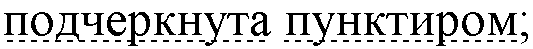
• искусственно введенные линкерные последовательности, обеспечивающие гибкость сочленения глобулярных доменов различного происхождения, выделены курсивом.
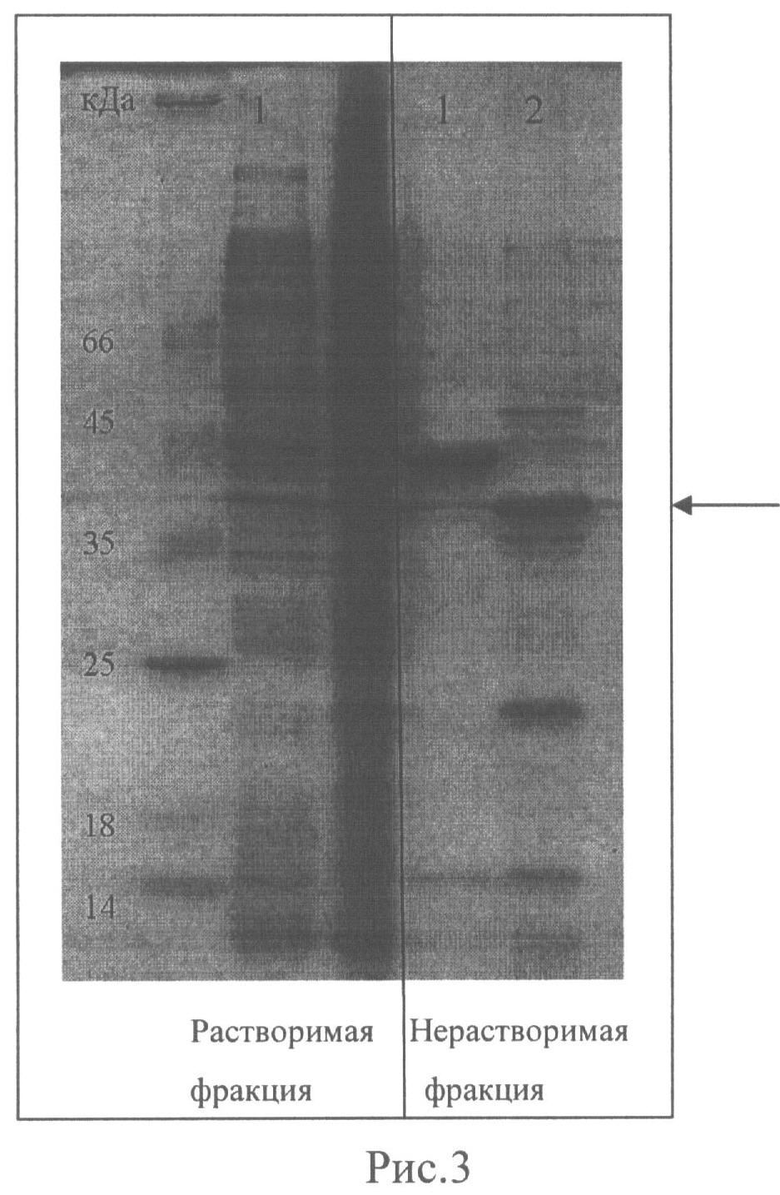
Рис.3. Электрофоретический анализ продуктов экспрессии конструкции рНК6 в растворимой и нерастворимой фракции лизата клеток Е.coli TG1, затрансформированных: (1) Вектором pQE30, (2) Конструкцией рНК6. Белки разделены в денатурирующем 15% ПААГ и окрашены на общий белок Coomassie Blue R-250. Стрелкой указано расположение белка НК6 (расчетная масса 42 кДа).
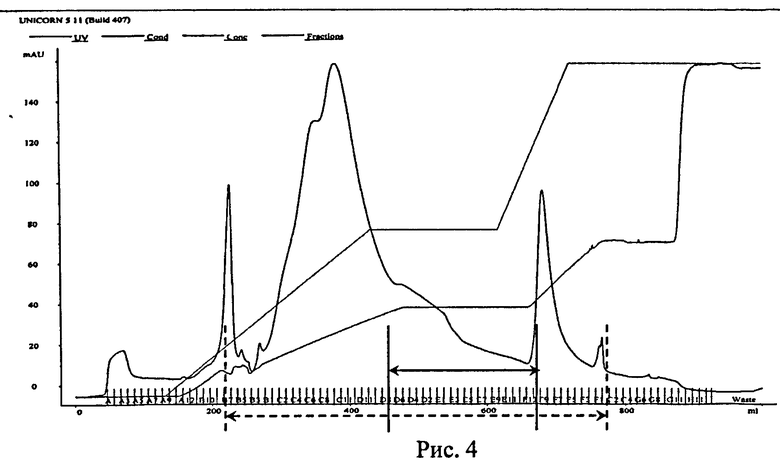
Рис.4. Анионообменная хроматография белка НК6 на колонке Sepharose-Q в присутствии 4М мочевины. Тонкими серыми линиями обозначены концентрация NaCl в элюирующем буфере и электропроводность раствора, черной линией - оптическая плотность при λ=280 нм. Штриховыми линиями обозначен диапазон фракций, отобранных для электрофоретического анализа, черными линями - фракции, отобранные для дальнейшей работы.
Рис.5. Электрофоретический анализ фракций белка НК6 после ионообменной хроматографии в денатурирующих условиях. Белки разделены в денатурирующем 15% ПААГ и окрашены на общий белок Coomassie Blue R-250. Положение целевого белка массой 42 кДа обозначено пунктирным овалом. Для дальнейшей работы были отобраны фракции D6-F6.
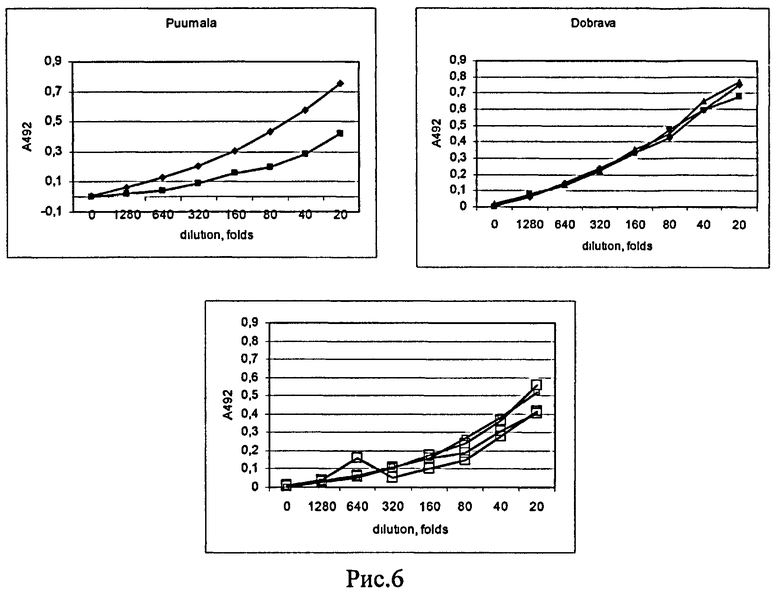
Рис.6. Изучение антигенной активности белка НК6 методом твердофазного иммуноферментного анализа. Планшеты активированы белком-антигеном НК6 и заблокированы нормальной сывороткой кролика в разведении 100 раз. Сыворотки больных ГЛПС (вирусы Добрава, Пуумала) и здоровых доноров раститровывались в серии разведении от 20 до 1280 раз с шагом 2 раза. На оси ординат указана величина А492 каждой точки за вычетом фона (контроль без добавления сыворотки человека).
Осуществление изобретения:
Методология исследования:
1) получение конструкции
2) экспрессия и анализ выхода белка
3) подготовка образцов к хроматографии
4) ионообменная хроматография.
5) проведение иммунохимических тестов.
1) Получение конструкции
Получение конструкции проводили по следующей схеме:
1. С использованием матрицы конструкции pET-Ht-Δ12 и праймеров Li4 (GGAGATCTCTAGCGGGGGCGCCGGAGGAATT) и НТ12 (GGACTAGTATCAAAGTCAAGTGCCTT) проведена ПЦР. Продукт обработан рестриктазой BglII.
2. На матрице плазмиды pQE-GFP проведена ПЦР с использованием праймеров ECFP1 (GGGGATCCATGGTGAGCAAGGGCGAGGA) и GFP10 (GGAGATCTCTTGTACAGCTCGTCCATGCC). Продукт обработан рестриктазой BglII.
3. Продукты стадий 1 и 2 объединены и подвергнуты лигированию.
4. На матрице продукта лигирования стадии 3 проведена ПЦР с праймерами ECFP1 и НТ12. Продукт ПЦР очищен выделением из геля и подвергнут расщеплению рестриктазами BamHI и SpeI.
5. Конструкция pQ-Knl, представляющая собой ген PKPI-B1 (S.tuberosum сорт Истринский, номер доступа NCBI AY692479) в составе вектора pQE30, обработана рестриктазами SpeI и BamHI.
6. Продукты реакции рестрикции со стадий 4 и 5 объединены, подвергнуты лигированию и введены в клетки Е. coli TG1 путем трансформации.
7. Полученную промежуточную конструкцию 6 на базе вектора pQE30 расщепляли рестриктазой SpeI и лигировали с синтетическим ДНК-дуплексом, полученным путем смешения заранее фосфорилированных олигонуклеотидов CTAGCGGGGGCGCCGGAGGAATT и CTAGTTCCGGCGCCCCCG с целью введения линкерной последовательности на границе областей НТ и легкой С-концевой цепи белкового ингибитора PKPI-B1. Полученная экспрессионная конструкция на базе вектора pQE30 получила обозначение рНК6 (Рис.1 и Рис.2).
2) Экспрессия и анализ выхода белка
Конструкцию рНК6 вводили в клетки штамма Е. coli TG1, отбирая ампициллин-устойчивые колонии. Полученный продуцент культивировали при 30°С в жидкой среде (0,5% дрожжевой экстракт, 1% пептон, 0,5% NaCl) с добавлением 100 мкг/мл ампициллина в колбах Эрленмейера объемом 750 мл (30 мл среды на колбу) в течение 14-18 часов. Посевной материал представлял собой смыв культуры клеток с чашек с агаризованной средой (0,5% дрожжевой экстракт, 1% пептон, 1,5% агар, 0,5% NaCl), полученных высевом первичных трансформантов. Возраст трансформантов - не более 40 часов с момента окончания трансформации, температура культивирования - 30°С. Доза засева - 5×108 клеток на колбу. Индукцию промотора в клетках продуцента не проводили.
Оценку выхода целевого продукта осуществляли с помощью электрофоретического анализа суммарных белков рекомбинантного продуцента по Лэммли. Для этого из грубого клеточного лизата каждой культуры отбирали по 100 мкл. Лизат подвергали центрифугированию при 14000 G. в течение 15 мин и разделяли растворимую и нерастворимую клеточную фракции. Белки солюбилизировали в 30 мкл буфере Лэммли и анализировали состав белков с помощью денатурирующего SDS-электрофореза в 15% ПААГ (Рис.3).
Проведенный анализ показал, что трифункциональный слитой белок на основе пептида НТ - продукт конструкции рНК6, как и свободный пептид НТ, накапливается в нерастворимой клеточной фракции, с выходом около 40 мг/л культуры.
3) Подготовка образцов к хроматографии
Грубый клеточный лизат культуры TGl(pHK6) подвергали центрифугированию при 8000 G. в течение 1 часа и разделяли растворимую и нерастворимую клеточную фракции. Объединенный осадок нерастворимой клеточной фракций трижды промывали: в 25 мл 4 М NaCl с добавлением 1% тритона, в 25 мл физиологического раствора с 1% тритоном и в 25 мл воды. Осадок суспендировали в 8 мл буфера (Tris-HCl, 10 мМ, рН=8,6), содержащего 8 М мочевину и 0,1% β-меркаптоэтанол. Раствор кипятили на водяной бане в течение 10 минут и центрифугировали. Полученный осветленный супернатант наносили на гельфильтрационную хроматографическую колонку с Sephadex G-25 fine со скоростью 5 мл/мин, используя жидкостную хроматографическую систему низкого давления "AKTA-Purifile" (GE Healthcare, США). Разделение проводили в буфере (Tris-HCl, 10 мМ, рН=8,6), содержащего 4 М мочевину, с целью удаления солей, избытка восстановителя и низкомолекулярных клеточных примесей. Солюбилизированный в денатурирующих условиях белок подвергали очистке с помощью анионообменной хроматографии.
Выделение и очистку целевых белков из полученного супернатанта проводили с использованием жидкостной хроматографической системы "AKTA-Purifile" (GE Healthcare, США).
4) Ионообменная хроматография
Основной целью анионнобменной хроматографии белков - производных пептида НТ в денатурирующих условиях, являлось удаление примесей нуклеиновых кислот, формирующих коллоид с участием белков и препятствующих эффективному применению гель-фильтрации и других методов хроматографии. При этом удаление из препарата примесей клеточных белков Е.coli рассматривалась лишь как побочная задача.
Полученный после гель-фильтрации раствор денатурированного белка НК6 в объеме 15 мл наносили на анионообменную колонку Sepharose-Q (XL) (GE Healthcare, США, высота 24 см, объем 48 мл), уравновешенную буфером А. Элюцию проводили линейным градиентом NaCl от 0 до 0,5 М в хроматографическом буфере В (Tris-HCl, 10 мМ, рН=8,6, 4 М мочевина, 1 М NaCl) со скоростью 8 мл/мин (Рис.4).
Собранные 20 фракций объемом 10 мл каждая анализировали с помощью денатурирующего SDS-электрофореза в 15% ПААГ. Для электрофореза отбиралось по 50-100 мкл от каждой фракции. Белки подвергали осаждению в 1 мл смеси (500 мкл 100% ацетона, 100 мкл 70% ТХУ на 1 мл). Полученные осадки солюбилизировали в денатурирующих условиях в 15 мкл буфера Лэммли (с добавлением мочевины) (Рис.5).
Материал фракций белка НК6, полученных в результате анионообменной хроматографии в денатурирующих условиях, оказался иммунохимически чистым и электрофоретически гомогенным, что позволило непосредственно использовать его для проведения ренатурации и иммунохимических тестов (Рис.6).
5) Проведение иммунохимических тестов
В качестве формата проведения анализа был выбран вариант тИФА с непосредственной сорбцией рекомбинантного белка-антигена НК6 на поверхность иммунологического планшета. В эксперименте использовались хроматографически очищенный белок НК6 в денатурированной форме. В процессе сорбции белковый препарат, хранящийся в виде концентрированных растворов в присутствии 4 М мочевины, разводили в 20-50 раз карбонат-бикарбонатным буфером (КГБ), доводя концентрацию общего белка в растворе до 10 мкг/мл. Полученный раствор вносили в иммунологические планшеты на 1 сутки, после чего проводили блокирование неспецифического связывания на подложке с помощью 1% БСА в буфере КГБ. Необходимым этапом проведения анализа оказалось блокирование связывания нормальных антител, присутствующих в сыворотке крови здоровых доноров, за счет конкуренции. С этой целью планшеты, подвергшиеся блокированию 1% БСА, дополнительно обрабатывали, внося в каждую лунку нормальную кроличью сыворотку, разбавленную буфером PBS в соотношении 1:100.
На следующей стадии проведения анализа проводилось серийное титрование анализируемой сыворотки больных ГЛПС (или здоровых доноров в группе сравнения). При этом с целью исследования специфичности реакции в выборки сравнения включали а) больных ГЛПС, инфицированных вирусом Добрава, b) больных ГЛПС, инфицированных вирусом Пуумала, с) здоровых доноров. В качестве контрольного образца использовали лунки планшета, в которые вместо разведенной сыворотки человека вносили буфер PBS («конъюгатный контроль»).
В заключение все лунки планшета обрабатывали антивидовым конюгатом против IgG человека, меченным пероксидазой, и субстратом ТМВ в присутствии пероксида водорода. Сигнал мерили на планшетном сканере-спектрофотометре при λ=492 нм. Результаты определений представлены на Рис.6.
Результаты показывают высокую реакционную способность антител положительных сывороток в отношении антигена НК6. При этом сигналы положительных сывороток больных в 2-2,5 раза превышают сигнал сывороток здоровых доноров в том же разведении, что позволяет достоверно диагносцировать наличие в крови людей специфических антител против вирусов Добрава и Пуумала.
Список использованных источников
1. Maes P., Clement J., Van Ranst M. Recent approaches in hantavirus vaccine development. - Expert Rev Vaccines. - 2009 - V.8, №1, P.67-76.
2. Cho H.W., Howard C.R., Lee H.W. Review of an inactivated vaccine against hantaviruses. - Intervirology. - 2002 - V.45, №4-6, P.328-333.
3. Song G. Epidemiological progresses of hemorrhagic fever with renal syndrome in China. - Chin Med J (Engi). - 1999 - V.112, №5, P.472-477.
4. Oya A. Japanese encephalitis vaccine. - Acta Paediatr Jpn. - 1988 - V.30, №2, Р.175-184.
5. Choi Y. Ahn C.J., Seong K.M., Jung M.Y., Ahn B.Y. Inactivated Hantaan virus vaccine derived from suspension culture of Vero cells. - Vaccine. - 2003 - V.21, №17-18, P.1867-1873.
6. Lee H.W., Chu Y.K., Woo Y.D. Immune responses after two or three doses of Hantavax vaccination against Hantaan virus // Proc. fifth international conference on hemorrhagic fever with renal syndrome, hantavirus pulmonary syndrome and hantaviruses. Lion, Franch - 2001. - P.234-242.
7. Ruan Y., Xu X., Liu W., Deng X, Weng S, Zhou W, Wang Q, Chen L, Fang L, Xu Z, Yan Q, Liu W, Dong G, Gu H, Yu Y, Xu Z. A study on immunogenicity and safety of bivalent inactivated vaccine against hemorrhagic fever with renal syndrome. - Zhonghua Yu Fang Yi Xue Za Zhi. - 1999 - V.33, №6, P.340-342.
8. Hooper J.W., Kamrud K.I., Eigh F., Custer D., Schmaljohn C.S. DNA vaccination with hantavirus M segment elicits neutralizing antibodies and protects against seoul virus infection. - Virology. - 1999 - V.255, №2, P.269-278.
9. Kallio-Kokko H., Leveelahti R., Brummer-Korvenkontio M., Lundkvist A., Vaheri A., Vapalahti O. Human immune response to Puumala virus glycoproteins and nucleocapsid protein expressed in mammalian cells. - J Med Virol. - 2001 - V.65, №3, P.605-613.
10. Huang H., Li X., Zehua Z. Genetic immunization with Hantavirus vaccine combining expression of G2 glycoprotein and fused interleukin-2. - Genetic Vaccines and Therapy. - 2008 - V.6. - P.15-21.
11. А.Б.Шевелев, М.С.Барботько, М.В.Баловнева, Е.Ю.Эпова, О.А.Леонович, Т.К.Дзагурова, Е.А.Ткаченко. Способ получения рекомбинантного антигена G2 хантавирусов в клетках Е.coli. Заявка на патент. Вход. №060029. Peг. №2010141819. Дата поступления 13.10.2010.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИГЕНА G2 ХАНТАВИРУСА ДОБРАВА В КЛЕТКАХ E.coli | 2012 |
|
RU2509805C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ФЕРМЕНТАТИВНО-МЕЧЕННОГО АНТИГЕНА G2 ХАНТАВИРУСА В КЛЕТКАХ E.coli С ЦЕЛЬЮ ЕГО ПРИМЕНЕНИЯ В ИММУНОФЕРМЕНТНОМ АНАЛИЗЕ ПРИ ДИАГНОСТИКЕ ГЛПС | 2012 |
|
RU2539836C2 |
| Рекомбинантная плазмида pET21-GST-ND, обеспечивающая синтез и секрецию эктодомена Gn гликопротеина вируса Хантаан, и рекомбинантный штамм Escherichia coli BL21(DE3)pLysE pET21-GST-ND - продуцент белка Gn - эктодомена гликопротеина вируса Хантаан | 2023 |
|
RU2809199C1 |
| Рекомбинантная плазмида pET21-GST-CD, обеспечивающая синтез и секрецию эктодомена Gc гликопротеина вируса Хантаан, и рекомбинантный штамм Escherichia coli BL21(DE3)pLysE pET21-GST-CD - продуцент белка Gc - эктодомена гликопротеина вируса Хантаан | 2023 |
|
RU2807520C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТЕСТ-СИСТЕМЫ "ХАНТА-N" ДЛЯ ОПРЕДЕЛЕНИЯ СПЕЦИФИЧЕСКОЙ АКТИВНОСТИ ВАКЦИНЫ ПРОТИВ ГЕМОРРАГИЧЕСКОЙ ЛИХОРАДКИ С ПОЧЕЧНЫМ СИНДРОМОМ | 2012 |
|
RU2590606C2 |
| ШТАММ ВИРУСА - ВОЗБУДИТЕЛЯ ГЕМОРРАГИЧЕСКОЙ ЛИХОРАДКИ С ПОЧЕЧНЫМ СИНДРОМОМ ДЛЯ ИЗГОТОВЛЕНИЯ ВАКЦИННЫХ ПРЕПАРАТОВ (ВАРИАНТЫ) | 2009 |
|
RU2423520C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОФЕРМЕНТНОЙ ТЕСТ-СИСТЕМЫ ДЛЯ ОПРЕДЕЛЕНИЯ ЭПИТОПОВ ОБОЛОЧЕЧНОГО БЕЛКА ВИРУСА ПУУМАЛА ПРОТЕКТИВНОЙ НАПРАВЛЕННОСТИ В ВАКЦИННЫХ ПРЕПАРАТАХ ПРОТИВ ГЕМОРРАГИЧЕСКОЙ ЛИХОРАДКИ С ПОЧЕЧНЫМ СИНДРОМОМ | 2013 |
|
RU2565543C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОМБИНИРОВАННОЙ БИВАЛЕНТНОЙ, КУЛЬТУРАЛЬНОЙ, ИНАКТИВИРОВАННОЙ, КОНЦЕНТРИРОВАННОЙ, ОЧИЩЕННОЙ ВАКЦИНЫ ДЛЯ ПРОФИЛАКТИКИ ГЕМОРРАГИЧЕСКОЙ ЛИХОРАДКИ С ПОЧЕЧНЫМ СИНДРОМОМ | 2009 |
|
RU2445117C2 |
| Рекомбинантная плазмидная ДНК pMBP-PUUV_N, обеспечивающая синтез и секрецию слитого белка MBP-PUUV_N в клетках E. coli, штамм Escherichia coli KRX/pMBP-PUUV_N, продуцирующий указанный белок, используемый для получения рекомбинантного белка PUUV_N, применяемого для диагностики геморрагической лихорадки с почечным синдромом | 2023 |
|
RU2838919C1 |
| СПОСОБ ПОЛУЧЕНИЯ "ДИАГНОСТИКУМА ГЕМОРРАГИЧЕСКОЙ ЛИХОРАДКИ С ПОЧЕЧНЫМ СИНДРОМОМ КУЛЬТУРАЛЬНОГО, ПОЛИВАЛЕНТНОГО ДЛЯ НЕПРЯМОГО МЕТОДА ИММУНОФЛЮОРЕСЦЕНЦИИ" | 2010 |
|
RU2431148C1 |

Изобретение относится к области биотехнологии, а именно к способу получения рекомбинантного антигена G2 хантавируса Добрава в клетках Е.coli. Способ характеризуется тем, что ДНК конструкции рНК6, указанной на фиг.1, кодирующая слитой белок из трех частей, где N-концевое положение занимает зеленый флуоресцентный белок GFP, центральное - пептид длиной 73 а.о. с последовательностью аминокислот SRKKCNFATTPICEYDGNMVSGYKKVMATIDSFQAFNTSYIHYTDEQIEW KDPDGMLKDHLNILVTKDIDFDT, а С-концевое - легкая цепь двуцепочечного белкового ингибитора типа Кунитца из клубней картофеля (PKPI-BI), вводят в клетки Е.coli. Культивируют трансформированные данной конструкцией клетки, лизируют биомассу, отделяют нерастворимую фракцию лизата центрифугированием. Продукт экспрессии в форме телец включения солюбилизируют с помощью денатуранта. Проводят хроматографию в денатурирующих условиях. Используют полученный продукт для выявления специфических антител в сыворотке больных геморрагической лихорадкой с почечным синдромом. Предложенное изобретение позволяет получать рекомбинантный антиген G2 хантавируса Добрава в клетках Е.coli с увеличенным выходом. 6 ил., 1 пр.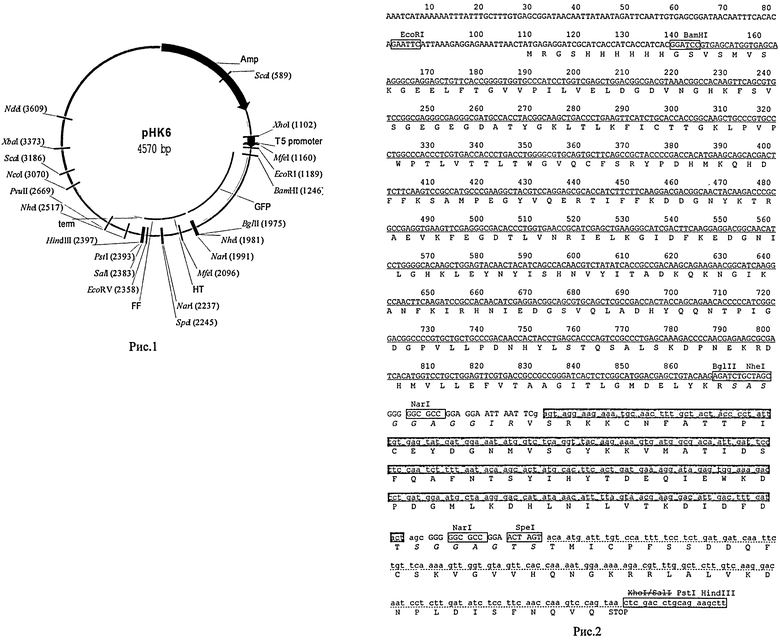
Способ получения рекомбинантного антигена G2 хантавируса Добрава в клетках Е.coli, характеризующийся тем, что ДНК конструкции рНК6, указанной на фиг.1, кодирующая слитой белок из трех частей, где N - концевое положение занимает зеленый флуоресцентный белок GFP, центральное - пептид длиной 73 а.о. с последовательностью аминокислот SRKKCNFATTPICEYDGNMVSGYKKVMATIDSFQAFNTSYIHYTDEQIEWKD PDGMLKDHLNILVTKDIDFDT, а С-концевое - легкая цепь двуцепочечного белкового ингибитора типа Кунитца из клубней картофеля (PKPI-BI), вводят в клетки Е.coli, культивируют трансформированные данной конструкцией клетки, лизируют биомассу, отделяют нерастворимую фракцию лизата центрифугированием, продукт экспрессии в форме телец включения солюбилизируют с помощью денатуранта, проводят хроматографию в денатурирующих условиях и используют полученный продукт для выявления специфических антител в сыворотке больных геморрагической лихорадкой с почечным синдромом.
| KOCH | |||
| J | |||
| ЕТ AL., Human recombinant neutralizing antibodies against hantaan virus G2 protein, Virology, 2003, v.308, n.1, pp.64-73 | |||
| US 20040053216 A1, 18.03.2004 | |||
| US 20040053216 A1, 18.03.2004 | |||
| OKUNO Y | |||
| ET AL., Haemagglutination-inhibition test for hemorrhagic fever with renal syndrome using virus antigen prepared from infected tissue culture fluid, |

