ОБЛАСТЬ ТЕХНИКИ
Настоящий документ относится к материалам и способам получения рекомбинантных белков, представляющих интерес, в растениях в не N-гликозилированной (не гликозилированной по аспартату) форме. Была разработана стратегия получения целевых белков в растениях в не N-гликозилированной форме, но без замены аминокислот в сайте NxS/T получающихся дегликозилированных белков, которая может обеспечить получение не N-гликозилированных рекомбинантных белков в растениях или других эукариотических систем экспрессии, имеющих третичную структуру, подобную природной. Также описываются материалы и способы in vivo дегликозилирования рекомбинантных N-гликозилированных белков посредством совместной экспрессии с Эндо-β-N-ацетилглюкозаминидазой Н (Endo Н) в растениях с использованием транзиентной системы экспрессии. Также обеспечивается способ экспрессии активного Endo Н в растениях.
КРАТКОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Растения стали альтернативной системой экспрессии и во все большей степени стали использоваться в промышленности и научных организациях для получения целевых белков. Однако способность растений гликозилировать белки может стать существенных ограничением для таких белков, которые не нуждаются в N-гликозилировании. Например, белки Plasmodium falciparum или А цепь фактора XHI человека не содержат N-присоединенные гликаны, или защитный антиген (РА) из Bacillus anthracis не является гликопротеином; однако эти белки содержат сайты потенциального N-опосредованного гликозилирования, которые могут быть ошибочно гликозилированы в процессе экспрессии в дрожжах, млекопитающих или растительных системах, что потенциально может привести к пониженной функциональности и иммуногенности ввиду неправильного / измененного свертывания и/или маскирования эпитопов. Для того, чтобы разрешить указанную проблему, мы недавно разработали стратегию ферментативного дегликозилирования белков in vivo посредством совместной экспрессии с бактериальной PNGазой F (Пептид: N-гликозидаза F) с использованием транзиентной экспрессии в растениях (международная патентная заявка WO/2012/170678), которая позволила осуществить получение кандидата вакцины против малярии Pfs48/45, который может обеспечить высокую активность блокирования передачи (ТВ) (Mamedov и др., 2012). В дополнение, другие дегликозилированные антигены индуцировали существенно более высокие уровни ответов антител, нейтрализующих токсины, в мышах по сравнению с гликозилированными формами (Mamedov и др., статья была подана). Хотя обработка PNGазой F (in vivo дегликозилирование) целиком удаляет указанный олигосахарид, но она приводит к замене аминокислот в указанном дегликозилированном белке вследствие деаминирования аспарагина (N) в указанном сайте NxS/T (последовательность) с образованием аспартата (D). В настоящем исследовании была разработана стратегия получения целевых белков в растениях в не N-гликозилированной форме, но без замены аминокислоты в указанном сайте NxS/T получаемых дегликозилированных белков, что может обеспечить получение не N-гликозилированных рекомбинантных белков в растениях или других эукариотических системах со свертыванием, подобным нативному. Таким образом, в настоящем изобретении раскрыты материалы и способы in vivo дегликозилирования рекомбинантных N-гликозилированных белков посредством совместной экспрессии с Эндо-β-N-ацетилглюкозаминидазой Н (Endo Н) в растениях с использованием транзиентной системы экспрессии. Также обеспечивается способ экспрессии активного Endo Н в растениях.
УРОВЕНЬ ТЕХНИКИ
N-гликозилирование является критической пост-трансляционной модификацией для правильного свертывания и стабильности множества белков, и биологическая активность множества рекомбинантных белков, полученных в гетерологических системах экспрессии зависит от их статуса гликозилирования. Однако некоторые эукариотические, а также бактериальные белки не содержат N-гликаны в природном хозяине, но содержат множество потенциальных сайтов N-гликозилирования, которые могут быть неправильно гликозилированы, когда эти белки экспрессируются в гетерологических эукариотических системах экспрессии, что потенциально может привести к снижению функциональности и иммуногенности ввиду неправильного/измененного сворачивания или маскировки эпитопов. Например, белок Pfs48/45 из Plasmodium falciparum или А цепь фактора XHI человека нес одержит N-связанных гликанов, и указанный защитный антиген (РА) из Bacillus anthracis не является гликопротеином; однако, эти белки содержат потенциальные сайты N-связанного гликозилирования, которые могут быть ошибочно гликозилированы в ходе экспрессии в дрожжах, млекопитающих или растительных системах. Растения становятся альтернативной системой экспрессии и все чаще используются в индустрии и академических институтах для получения целевых белков. Однако, способность растений гликозилировать белки также может быть существенным ограничением при применении систем экспрессии, основанных на растениях. В нашем предыдущем исследовании мы разработали стратегию ферментативного дегликозилирования белков in vivo посредством совместной экспрессии с бактериальной PNGазой F (Peptide: N-glycosidase F, Пептид: N-гликозидазой F) с использованием транзиентной экспрессии в растениях (патентная заявка РСТ WO/2012/170678). С использованием стратегии дегликозилирования in vivo был получен белок Pfs48/45 в N. benthamiana в не-N-гликозилированной форме и четыре мАт, полученных против различных эпитопов белка Pfs48/45, два из которых являются конформационно специфическими, узнававшими дегликозилированную форму Pfs48/45 в от 2-х до 6-ти раз лучше, чем они узнавали гликозилированную форму того же белка (Mamedov и др., 2012). В дополнение, самое сильное связывание и максимальная степень ингибирования сигнала наблюдалась при использовании мАт III in vivo только с PNGазой F-дегликозилированным Pfs48/45, в то время как in vitro дегликозилированный, гликозилированный Pfs48/45 (Mamedov и др., 2011) был эквивалентен по его способности ингибировать сигнал мАт III. В дополнение, его также проверяли с другими мишенями и результаты продемонстрировали, что только in vivo дегликозилированная форма имела намного более сильное связывание с антителами по сравнению с in vitro дегликозилированной и гликозилированной формами, что позволяет предположить, что ошибочное, неправильное гликозилирование может приводить к маскированию важных эпитопов или может вызывать неправильное/измененное складывание белка Pfs48/45 (Mamedov и др., 2012).
Эндо-β-N-ацетилглюкозаминидаза Н (Endo Н, ЕС3.2.1.96) является гликогидролазой, которая секретируется Streptomyces plicatus и несколькими другими видами Streptomyces (Tarentino и др., 1976). Она расщепляет р-1,4-гликозидную связь N-ацетилглюкозаминового остова олигосахаридов и оставляет одну N-ацетилхитобиозу присоединенной к аспарагиновому остатку указанного гликопротеина (Trimble и др., 1978; Muramatsu 1971). Ген Endo Н из S. plicatus имеет длину 939 п. о. (номер доступа в GenBank - ААА26738.1) и кодирует белок весом 28,9 кДа. Недавно Endo Н из Streptomyces plicatus экспрессировали в Pichia pastoris и дегликозилирующую активность Endo Н, полученной из P. pastoris, продемонстрировали in vitro как при совмещенной с ферментацией обработкой, так и при обработке после ферментации (Wang и др., 2015). Однако, условия N-дегликозилирования белков in vivo посредством фермента Endo Н не были достигнуты. В настоящем исследовании описано и предлагается in vivo дегликозилирование рекомбинантных N-гликозилированных белков посредством совместной экспрессии с Endo Н в растениях с использованием транзиентной системы экспрессии.
ЦЕЛЬ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
В нашем предыдущем исследовании мы продемонстрировали дегликозилирование целевого белка in vivo посредством совместной экспрессии целевых белков с бактериальной PNGазой F (Mamedov Т. WO/2012/170678, 2012; Mamedov и др., 2012). Хотя дегликозилирование с помощью PNGазы F (in vivo или in vitro) целиком удаляет олигосахариды, но оно вызывает замену аминокислоты в дегликозилированном белке вследствие деаминирования аспарагина (N) в сайте NxS/T (последовательность) с образованием аспартата (D). В данный момент, другие дегликозилирующие ферменты, такие как Эндо-Р-N-ацетилглюкозаминидаза Н (Endo Н), катализирует расщепление между двумя остатками GlcNAc хитобиозного ядра N-связанных гликанов, оставляя единичный остаток GlcNAc, присоединенный к аспарагинам (Фиг. 1), в противоположность PNGазе F, а дегликозилирование посредством Endo Н не приводит к изменению последовательности аминокислот в сайте NxS/T получающихся дегликозилированных белков. Поскольку обработка Endo Н не приводит к изменению аминокислоты в получающихся дегликозилированных белках, я предположил, что дегликозилированные белки, полученные с помощью Endo Н, могут иметь третичную структуру, похожую на нативную, и, как следстви,е лучшую функциональную активность (иммуногенность, связывание с рецепторами, взаимодействие белок-антитело, активность фермента и т.п.) по сравнению с дегликозилированной формой тех же белков, полученных с помощью PNGазы F. Я также предположил, что поскольку Endo Н расщепляет, оставляя один остаток GlcNAc, присоединенный к аспарагинам, остающийся моносахарид привносит заряд, что может повышать растворимость и стабильность дегликозилированных белков.
ПРОМЫШЛЕННОЕ ПРИМЕНЕНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Как описано выше, обработка PNGазой F (дегликозилирование in vivo) удаляет олигосахарид полностью, но приводит к замене аминокислоты в указанном дегликозилированном белке вследствие деаминирования аспарагина в указанном сайте NxS/T (последовательности) с получением аспартата. Обработка с помощью Endo Н не приводит к замене аминокислоты в указанном сайте NxS/T получаемых дегликозилированных белков. В данный момент предполагается, что дегликозилированные белки, полученные с помощью Endo Н, могут иметь третичную структуру, более похожую на природную, и как следствие лучшую функциональную активность (иммуногенность, связывание с рецепторами, взаимодействие белок-антитело, активность фермента и т.п.) по сравнению с дегликозилированной формой тех же белков, полученных с помощью PNGазы F. Также предполагается, что поскольку Endo Н расщепляет, оставляя один остаток GlcNAc, присоединенный к аспарагинам, остающийся моносахарид привносит заряд, что может повышать растворимость и стабильность дегликозилированных белков. В этом исследовании я впервые демонстрирую экспрессию дегликозилирующего бактериального фермента Endo Н в растениях. Я также впервые демонстрирую дегликозилирование целевых белов посредством Endo Н in vivo в растении N. benthamiana. Мы демонстрируем, что указанный рекомбинантный Endo Н, полученный в растениях, является полностью активным in vivo и успешно отщепляет N-присоединенные гликаны от белков РА83 и Pfs48/45. В дополнение, одновременная экспрессия РА83 с Endo Н приводит к накоплению РА83, имеющего такой же размер, как и молекула, дегликозилированная in vivo с помощью бактериальной PNGазы F. Важно отметить, что результаты нашего предварительного анализа стабильности демонстрируют, что РА83, дегликозилированный in vivo посредством Endo Н, оказался более стабильным по сравнению с гликозилированной или дегликозилированной с помощью PNGазы F (in vivo) формами того же белка, в особенности при повышенной температуре. В заключение, все эти результаты свидетельствуют о том, что стратегия одновременной экспрессии Endo Н может быть использована для получения не-гликозилированных рекомбинантных белков в указанной транзиентной системе экспрессии на основе N. benthamiana в формах, подобных нативным формам. В дополнение, результаты этого исследования выявили потенциал РА83, дегликозилированного Endo Н, в качестве вакцины против сибирской язвы и Pfs48/45 в качестве вакцины против малярии, и подтвердили их дальнейшие характеристики для разработки перед клиническими испытаниями. Эти результаты демонстрируют, что Endo Н успешно отщепляет N-связанные гликаны от всех протестированных гликопротеинов и позволяют предположить, что указанная стратегия одновременной экспрессии с Endo Н может быть использована для получения негликозилированных антигенов для вакцин, терапевтических белков, антител и бактериальных белков (в особенности кандидатов антигенов для вакцин) - ферментов. В дополнение к этому, настоящее изобретение может быть использовано для получения промышленных ферментов, в особенности ферментов бактериального происхождения, в любых эукариотических системах для повышения биоэнергетического/биотопливного выхода, а также для улучшения качества пищевых продуктов, в особенности при получении натуральных пищевых добавок.
ОПИСАНИЕ РИСУНКОВ
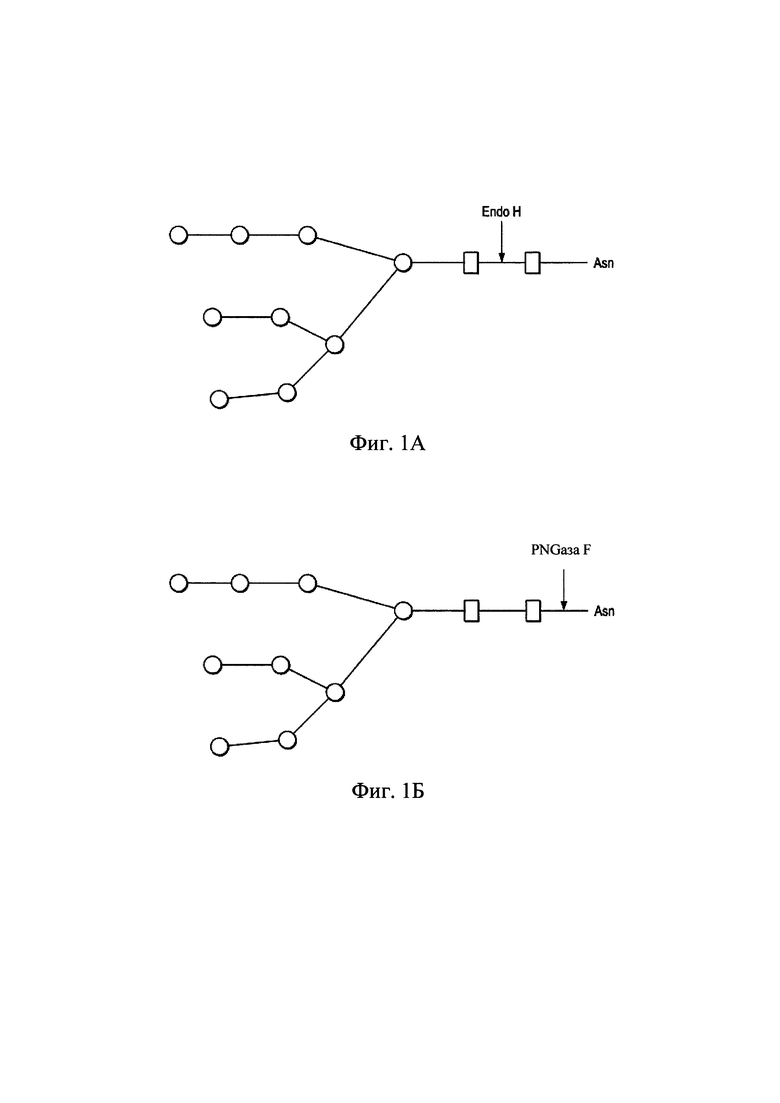
На Фиг. 1А изображено как Endo Н расщепляет связь между двумя остатками N-ацетилглюкозамина (GlcNAc) в диацетилхитобиозном ядре олигосахарида с образованием укороченной молекулы сахара с один остатком GlcNAc, оставшимся на аспарагинах (Asn).
На Фиг. 1Б изображено как Пептид-N-Гликозидаза F (PNGаза F), являющаяся амидазой, расщепляет связь между самым глубоко расположенным остатком GlcNAc и остатком аспарагина олигосахаридов с высоким содержание маннозы, и гибридных и сложных олигосахаридов N-связанных гликопротеинов;  : N GlcNAc;
: N GlcNAc;  : манноза.
: манноза.
На Фиг. 2 изображены последовательности нуклеотидов и аминокислот Endo Н, которые были экспрессированы в N. benthamiana как описано в настоящем описании. На Фиг. 2А изображена последовательность нуклеотидов бактериального гена Endo Н, кодоны которой были оптимизированы с использованием кодонов N. benthamiana. На Фиг. 2Б изображена последовательность аминокислот Endo Н, экспрессированной в растении N. benthamiana. Сигнальная пептидная последовательность PR-1a (MGFVLFSQLPSFLLVSTLLLFLVISHSCRA) (SEQ ID NO: 3), добавленная на N-конец подчеркнута. Эпитоп FLAG (маркер для аффинной очистки), за которым следует последовательность KDEL (SEQ ID NO: 5) (сигнал удерживания ER) (выделена жирным), были добавлены на С-конец.
На Фиг. 3 изображены результаты Вестерн-блот анализа Endo Н в растениях N. benthamiana. Растения N. benthamiana, в которые были введены конструкции pGR-M-Endo Н или pGR-M-PNGase F, продуцировали Endo Н и PNGазу F. Образцы листьев собирали через 5 дней после инокуляции гомогенизировали в трех объемах буфера для экстракции. После центрифугирования при 13,000 g образцы наносили на ДСН-ПААГ с последующим Вестерн-блоттингом. Endo Н и PNGase F детектировали с использованием поликлонального антитела кроликов против указанного маркера (Cell signaling, кат. №2368). Антитело против IgG кролика, связанное с пероксидазой хрена, (Cell signaling, кат. №7074) использовали в качестве вторичного антитела. 1 - растение N. benthamiana, экспрессирующее PNGазу F (35 кДа); 2 - растение N. benthamiana, экспрессирующее Endo Н (30 кДа).
На Фиг. 4 показано, что листья растения N. benthamiana, в которые были введены pGR-M-PNGase F/pGR-M-PA83 (1,2) или pGR-M-Endo H/pGR-M-PA83 (3-5), как указано, анализировали с помощью Вестерн-блота с использованием поликлонального антитела кролика против указанного маркера (Cell signaling, кат. №2368) для детектирования PNGазы F или Endo Н. А: 6 - растение N. benthamiana, в которое ничего не вводили. На Фиг. 4Б изображены результаты анализа Вестерн-блот растений N. benthamiana, в которые были введены pGR-M-PNGase F/pGR-M-PA83 (1, 2) или pGR-M-Endo H/pGR-M-PA83 (3-5). Для детектирования белка РА83 использовали антитело против РА (защитное антигенное антитело ВАР0101 против Bacillus anthracis, кат.№abl988, Abeam).
На Фиг. 5 изображены результаты анализа Вестерн-блот одновременной экспрессии Bacillus anthracis РА с бактериальными PNGазой F и Endo Н, как указано в настоящем описании, в растениях N. benthamiana. В растения N. benthamiana вводили конструкции pGR-M-PNGase/ pGR-M-PA83 или pGR-M-Endo Н/ pGR-M-PA83 для продукции дегликозилированного РА83. Образцы листьев собирали через 5 дней после инокуляции и гомогенизировали в трех объемах буфера для экстракции. После центрифугирования при 13000 g образцы разбавляли в 10 раз в ДСН-буфере для образцов. 10 микролитров образцов наносили на ДСН-ПААГ с последующим Вестерн-блоттингом. Полосы РА83 получали с использованием антитела против РА (защитное антигенное антитело против Bacillus anthracis, ВАР0101, кат. № abl988, Abeam) или антитела против гистидинового маркера (Penta His Antibody, без БСА, кат. №34660), QIAGEN), как указано. РА83 экспрессировали один (-) или с PNGазой F (+) или Endo Н (+).
На Фиг. 6 изображены результаты анализа Вестерн-блот одновременной экспрессии белка Pfs48/45 из Plasmodium falciparum с бактериальным Endo Н в растениях N. benthamiana. В растения N. benthamiana вводили конструкции pGR-M-Endo Н или pGR-M-Endo H/pGR-М-РА83, как указано, для продукции дегликозилированного РА83. Образцы листьев собирали через 7 дней после инокуляции и гомогенизировали в трех объемах буфера для экстракции. После центрифугирования при 13000 g образцы разбавляли в 10 раз в ДСН-буфере для образцов. 10 микролитров образцов наносили на ДСН-ПААГ с последующим Вестерн-блоттингом. Полосы Pfs48/45 и Endo Н получали с использованием поликлонального антитела против FLAG (Cell signaling, кат. №2368).
На Фиг. 7 изображены результаты анализов ДСН-ПААГ (А, Б) и Вестерн-блот (В) дегликозилированных форм РА83, очищенных с использованием смолы HisPur™ Ni-NTA. На Фиг. 7А изображены результаты анализа ДСН-ПААГ дегликозилированных белков РА83, полученных посредством совместной экспрессии Endo Н и PNGазы F, соответственно. На Фиг. 7Б изображены результаты окрашивания белков в ДСН-ПААГ с использованием Coomassie Blue, инкубированного при 4°С в течение 48 ч. На Фиг. 7 В изображены результаты анализа Вестерн-блот дегликозилированных белков РА83, инкубированного при 37°С в течение 1 ч и при 4°С в течение 48 ч, как указано.
ПРИМЕРЫ
Пример 1 - Материалы и методы
Клонирование и экспрессия Endo H в N. benthamiana. Ген Endo Н (номер доступа в GenBank - ААА26738.1) оптимизировали для экспрессии в растениях N. benthamiana и синтезировали с помощью GENEART AG (Thermo Fisher Scientific). Для осуществления транзиентной экспрессии Endo Н в растениях N. benthamiana из последовательности Endo Н удаляли сигнальный пептид (аминокислоты 1 - 42) и на N-конец добавляли сигнальный пептид PR-1a Nicotiana tabacum (MGFVLFSQLPSFLLVSTLLLFLVISHSCRA) (SEQ ID NO: 3). В дополнение, на С-конец добавляли последовательность KDEL (SEQ ID NO: 5) (сигнал удерживания ER) и эпитоп FLAG (маркер для аффинной очистки). Полученную последовательность вводили в модифицированный бинарный вектор экспрессии pGreen II (pGR-M) или pBI121 (Chen и др., 2003) с получением pGR-M-Endo Н и pBI-Endo Н, соответственно.
Клонирование и экспрессия PNGазы F, РА83 и Pfs48/45 в N. benthamiana. Последовательности PNGазы F (Mamedov и др., 2012) и РА из В. anthracis (аминокислоты 30 - 764, номер доступа в GenBank - ААА22637, Mamedov и др., 2012) оптимизировали для экспрессии в растениях N. benthamiana и ген PNGазы F длиной 942 п. о. синтезировали из олигонуклеотидов, а ген РА длиной 2205 п. о. синтезировали с использованием компьютерной программы Gene2Oligo (Rouillard и др., 2004; Mamedov и др., 2007). Последовательность Pfs48/45 (аминокислоты 28 - 401, номер доступа в GenBank -EU366251) оптимизировали для экспрессии в растениях N. benthamiana и синтезировали с использованием GENEART AG (Thermo Fisher Scientific). Ha N-конец всех генов добавляли сигнальную последовательность PR-1a (MGFVLFSQLPSFLLVSTLLLFLVISHSCRA) (SEQ ID NO: 3). В дополнение, на С-конец добавляли последовательность KDEL (SEQ ID NO: 5) (сигнал удерживания ER) и эпитоп FLAG (для Endo Н и PNGазы F, маркер для аффинной очистки) или гистидиновый маркер (для РА83). Полученные последовательности вводили в бинарный вектор экспрессии pGR-M или pBI121 (Chen и др., 2003). Плазмиды pGreen II, pSoup получали из John Innes Centre (Норвич). Затем все плазмиды вместе с pSoup и р19 вводили в штамм Agrobacterium tumefaciens AGL1 или GV3101. Полученные штаммы бактерий выращивали в среде BBL (10 г/л гидролизата сои, 5 г/л экстракта дрожжей, 5 г/л NaCl и 50 мг/л канамицина) в течение ночи при 28°С.Бактерии вводили вручную в растения N. benthamiana возраста 6 недель, растущие в почве. Через четыре, пять, шесть и семь дней после введения собирали листья и гомогенизировали с использованием ступки и пестика. Штаммы Agrobacterium AGL1 или GV3101 и плазмида р19 были любезно предоставлены профессором Sophien Kamoun (The Sainsbury Laboratory, Норвич, Великобритания). Растения дикого типа Nicotiana benthamiana выращивали на почве в теплице Университета Акдениз.
Анализ ДСН-ПААГ и Вестерн-блот. Анализ образцов растений, продуцирующих РА83, осуществляли с помощью ДСН-ПААГ в 10% акриламидных гелях с окрашиванием Coomassie (Gel Code Blue, Пирс Рокфорд, Иллинойс). Анализ Вестерн-блот проводили после электрофореза и переноса белков на поливинилиденфторидные мембраны. После переноса, мембраны Вестерн-блот блокировали с использованием I-Block (Applied Biosystems, Карлсбад, Калифорния) и рекомбинантные белки детектировали с помощью антител против 4xHis (Qiagen, Валенсия, Калифорния) или защитного антигенного антитела против Bacillus anthracis [ВАР0101] (abl988). Затем мембраны промывали 1 раз с помощью фосфатно-солевого буфера, содержащего 0,1% Твин 20 (PBS-T), для удаления избытка первичного антитела и проявляли с использованием вторичного антитела против мыши, конъюгированного с пероксидазой хрена (HRP), (ab98790) или вторичного антитела против кролика, конъюгированного с пероксидазой хрена (HRP), (ab97051). Генерирование сигнала достигали с использованием хемилюминесцентного субстрата (SuperSignal West Pico, Thermo Fisher Scientific, Гранд-Айленд, Нью-Йорк); мембрану инкубировали в 5 мл хемилюминесцентного субстрата SuperSignal West Pico в течение 5 минут, завернутую в пластиковую пленку, а затем экспонировали рентгеновскую пленку Kodak в течение различных промежутков времени. Пленку проявляли и закрепляли в темной комнате с использованием растворов Kodak® для проявления и быстрого фиксирования, соответственно.
Пример 2 - Клонирование и экспрессия бактериального гена Endo Н в N. benthamiana
Последовательность бактериального гена Endo Н (GenBank: ААА26738.1), кодирующую 313 аминокислот (полноразмерный каталитически активный белок без сигнальной последовательности, аминокислоты 1 - 42) оптимизировали для экспрессии в растениях N. benthamiana (Фиг. 2), клонировали в вектор pGR-M и экспрессировали в растениях N. benthamiana, как описано в Материалах и методах. В дополнение, для сравнения дегликозилирования in vivo посредством Endo Н и PNGазы F, последовательность бактериального гена PNGазы F, кодирующую 314 аминокислот, оптимизировали для экспрессии в растениях N. benthamiana, и экспрессировали как описано ранее (Mamedov и др., 2012). Экспрессию Endo Н и PNGазы F в растениях N. benthamiana подтверждали с помощью анализа Вестерн-блот с использованием моноклонального антитела против FLAG (мАт) (Фиг. 3). Как показано на Фиг. 3, PNGаза F мигрирует как белок размером ~35 кДа в ДСН-ПААГ (с последующим Вестерн-блотом), когда Endo Н мигрирует как белок размером ~25 кДа, быстрее, чем белок PNGаза F. Следует отметить, что растения N. benthamiana, экспрессирующие Endo Н и PNGазу F, остаются здоровыми на 7, 8 и 9 дни после инокуляции (Фиг. 4) без видимых симптомов изменения развития или роста когда одновременная экспрессия целевых белков достигает наивысшего уровня.
Пример 3 - Способность Endo Н к дегликозилированию; дегликозилирование in vivo рекомбинантного белка РА из В. anthracis в растениях N. benthamiana при совместной экспрессии с Endo Н
Для оценки расщепления in vivo N-связанных олигосахаридов, украшающих РА83, белки Endo Н и РА83 одновременно транзиентно экспрессировали в растениях N. benthamiana посредством совместной агроинфильтрации. В качестве контроля, бактериальную PNGазу F также транзиентно совместно экспрессировали в растениях N. benthamiana посредством совместной агроинфильтрации. Следует отметить, что РА из В. anthracis не является гликопротеином, но он получается гликозилированным при экспрессии в растениях N. benthamiana. Анализ Вестерн-блот, проведенный через 5 дней после инокуляции, продемонстрировал сдвиг подвижности РА83 при совместной экспрессии с Endo Н (Фиг. 5), указывая на дегликозилирование белка. В дополнение, как показано с помощью анализов ДСН-ПААГ Вестерн-блот, совместная экспрессия с Endo Н приводит к накоплению РА83, имеющего размер, одинаковый с размером молекулы, дегликозилированной in vivo с помощью бактериальной PNGазы F (Mamedov и др., 2012), что позволяет предположить, что РА83 был ферментативно дегликозилированным (Фиг. 5).
Пример 4 - Способность Endo Н к дегликозилированию; дегликозилирование in vivo белка Pfs48/45 в растениях N. benthamiana при совместной экспрессии с Endo Н
Pfs48/45 является одним из ведущих кандидатов для разработки вакцины для блокирования передачи (ТВ). Для оценки расщепления in vivo N-связанных олигосахаридов, украшающих Pfs48/45, бактериальную Endo Н и белок Pfs48/45 из Plasmodium falciparum совместно транзиентно экспрессировали в растениях N. benthamiana посредством совместной агроинфильтрации с помощью конструкций pGR-M-Pfs48/45 и pGR-M-Endo Н. Как показывают результаты ДСН-ПААГ и Вестерн-блота, совместная экспрессия с Endo Н приводит к накоплению Pfs48/45 молекулярной массы ~60 кДа, что схоже с массой молекулы, дегликозилированной in vitro посредством PNGазы F (см. Фиг. 4, Mamedov и др., 2012), что указывает на то, что Pfs48/45 был ферментативно дегликозилирован (Фиг. 5). Эти результаты демонстрируют, что Endo Н успешно отщепляет N-связанные гликаны от белков РА83 и Pfs48/45, а произведенная в растениях Endo Н является активной in vivo, что подтверждает, что указанная стратегия совместной экспрессии Endo Н может быть использована для продукции терапевтических белков в негликозилированной форме в транзиентной системе экспрессии на основе N. benthamiana.
Пример 5 - Очистка дегликозилированных белков РА83 с использованием смолы HisPur™ Ni-NTA и предварительный анализ стабильности дегликозилированного белка РА.
Совместно экспрессировали Endo Н и PNGase F и дегликозилированные формы РА83 очищали с использованием смолы HisPur™ Ni-NTA (ThermoFisher scientific, кат. №8822). Как видно из результатов ДСН-ПААГ и окрашивания Coomassie (Фиг. 5А), очищенные белки РА83 были в высокой степени гомогенными. В дополнение, как показывают результаты анализов ДСН-ПААГ (Фиг. 5А) Вестерн-блот, совместная экспрессия с Endo Н приводит к накоплению РА83, который имеет размер сходный с размером молекулы, дегликозилированной in vivo посредством бактериальной PNGазы F. Для того, чтобы оценить эффект дегликозилирования посредством Endo Н и PNGазы F на свойства белков, дегликозилированные формы белка РА83 подвергали предварительному тесту на стабильность различных форм РА83 при температурах 37°С и 4°С и обнаружили, что РА83, дегликозилированный in vivo посредством Endo Н, оказался немного более стабильным по сравнению с формой, дегликозилированный посредством PNGазы F, в особенности при повышенной температуре (37°С).
Пример 6 - Связывание моноклонального антитела против РА с гликозилированным вариантом РА83 и с вариантами РА83, дегликозилированными in vivo посредством Endo Н или PNGазы F
Связывание моноклонального антитела против РА с гликозилированным вариантом РА83 и с вариантами РА83, дегликозилированными in vivo посредством Endo Н или PNGазы F осуществляли с использованием антигенов РА83, очищенных ГМАС, и защитного антигенного антитела против Bacillus anthracis (MA 1-21675). Антитело против РА (защитное антигенное антитело против Bacillus anthracis, ВАР0101, кат. № abl988, Abсam) показало одинаковое связывание с полученным в растениях гликозилированным белком РА83 и белками РА83, дегликозилированными in vivo Endo Н или PNGase F (данные не показаны).
Ссылки
Дегликозилирование рекомбинантных белков in vivo посредством совместной экспрессии с РNGазой F. 12, 2012, WO/2012/170678.
Mamedov Т, Ghosh A, Jones RM, Mett V, Farrance СЕ, Musiychuk K, et al. Production of non-glycosylated recombinant proteins in Nicotiana benthamiana plants by co-expressing bacterial PNGase F. Plant biotechnology journal. 2012; 10: 773-782.doi: 10.1111/j.1467-7652.2012.00694.x PMID: 22520228.
Mamedov T, Yusibov V. In vivo deglycosylation of recombinant proteins in plants by со-expression with bacterial PNGase F. Bioengineered. 2013; 4: 338-342.
Mamedov, Т.G., Padhye, N.V., Viljoen, H., Subramanian, A. 2007. "Rational de novo gene synthesis by rapid polymerase chain assembly (PCA) and expression of endothelial protein-C and thrombin receptor", Journal of Biotechnolgy, 131(4), 379-87.
Tarentino AL, Maley F. Purification and properties of an endo-beta-N-acetylglucosaminidase from hen oviduct. Journal of Biological Chemistry. 1976; 251(21): 6537-6543. PMID: 977586.
Trimble RB, Tarentino AL, Plummer TH, Maley F. Asparaginylglycopeptides with a low mannose content are hydrolyzed by endo-beta-N-acetylglucosaminidase H. Journal of Biological Chemistry. 1978; 253(13): 4508-4511. PMID: 659430.
Muramatsu T. Demonstration of an endo-glycosidase acting on a glycoprotein. Journal of Biological Chemistry. 1971; 246(17): 5535-5537. PMID: 4108054.
Rouillard, J.M., Lee, W., Truan, G., Gao, X., Zhou, X., Gulari, E., 2004.Gene2Oligo: oligonucleotide design for in vitro gene synthesis. Nucleic Acids Res. 32, W176-W180.
Wang F, Wang X, Yu X, Fu L, Liu Y, Ma L, Zhai C. High-level expression of endo-β-N-acetylglucosaminidase H from Streptomyces plicatus in Pichiapastoris and its application for the deglycosylation of glycoproteins. PLoS One. 2015 Mar 17; 10(3):e0120458. doi: 10.1371/journal.pone.0120458. eCollection 2015.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MAMMEDOV, Tarlan
<120> PRODUCTION OF IN VIVO N-DEGLYCOSYLATED RECOMBINANT PROTEINS BY
CO-EXPRESSION WITH ENDO H
<130> 10108739-50427845-0000
<150> PCT/IB2015/058781
<151> 2015-11-13
<160> 5
<170> PatentIn version 3.5
<210> 1
<211> 942
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic: recombinant DNA
<400> 1
atgggtttcg tgctgttcag ccagctgcct tctttccttc ttgtgtctac ccttctgctg 60
ttcctggtga tctctcattc ttgtagggct ccagctcctg ctcctgttaa gcaaggtcct 120
acttctgtgg cttacgttga ggtgaacaac aacagcatgc tgaacgtggg aaagtacacc 180
cttgctgatg gtggtggtaa cgctttcgat gtggctgtga ttttcgctgc taacatcaac 240
tacgataccg gtactaagac cgcttacctg cacttcaatg agaacgtgca gagggtgttg 300
gataacgctg tgactcagat taggcctctt cagcagcagg gtattaaggt gctgctttct 360
gtgcttggta accaccaggg tgctggtttc gctaattttc ctagtcagca ggctgcttcc 420
gctttcgcta agcaactttc tgatgctgtg gctaagtacg gtctggatgg tgtggatttc 480
gatgatgagt acgctgagta cggtaacaac ggtactgctc agcctaacga tagctctttc 540
gtgcatcttg tgaccgctct gagggctaac atgcctgata agatcatcag cctttacaac 600
atcggtcctg ctgcttccag gctttcttac ggtggtgttg atgtgagcga taagttcgat 660
tacgcttgga acccttacta cggaacctgg caagttcctg gtattgcttt gcctaaggct 720
cagctttctc cagctgctgt tgagattggt aggacctcta ggtctaccgt ggctgatctt 780
gctagaagga ctgtggatga gggttacggt gtgtacctta cctacaacct ggatggtggt 840
gataggaccg ctgatgtgtc tgctttcacc agagagcttt acggttctga ggctgtgagg 900
acccctgatt acaaggacga tgatgataag gatgagctgt ag 942
<210> 2
<211> 313
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic: recombinant protein
<400> 2
Met Gly Phe Val Leu Phe Ser Gln Leu Pro Ser Phe Leu Leu Val Ser
1 5 10 15
Thr Leu Leu Leu Phe Leu Val Ile Ser His Ser Cys Arg Ala Pro Ala
20 25 30
Pro Ala Pro Val Lys Gln Gly Pro Thr Ser Val Ala Tyr Val Glu Val
35 40 45
Asn Asn Asn Ser Met Leu Asn Val Gly Lys Tyr Thr Leu Ala Asp Gly
50 55 60
Gly Gly Asn Ala Phe Asp Val Ala Val Ile Phe Ala Ala Asn Ile Asn
65 70 75 80
Tyr Asp Thr Gly Thr Lys Thr Ala Tyr Leu His Phe Asn Glu Asn Val
85 90 95
Gln Arg Val Leu Asp Asn Ala Val Thr Gln Ile Arg Pro Leu Gln Gln
100 105 110
Gln Gly Ile Lys Val Leu Leu Ser Val Leu Gly Asn His Gln Gly Ala
115 120 125
Gly Phe Ala Asn Phe Pro Ser Gln Gln Ala Ala Ser Ala Phe Ala Lys
130 135 140
Gln Leu Ser Asp Ala Val Ala Lys Tyr Gly Leu Asp Gly Val Asp Phe
145 150 155 160
Asp Asp Glu Tyr Ala Glu Tyr Gly Asn Asn Gly Thr Ala Gln Pro Asn
165 170 175
Asp Ser Ser Phe Val His Leu Val Thr Ala Leu Arg Ala Asn Met Pro
180 185 190
Asp Lys Ile Ile Ser Leu Tyr Asn Ile Gly Pro Ala Ala Ser Arg Leu
195 200 205
Ser Tyr Gly Gly Val Asp Val Ser Asp Lys Phe Asp Tyr Ala Trp Asn
210 215 220
Pro Tyr Tyr Gly Thr Trp Gln Val Pro Gly Ile Ala Leu Pro Lys Ala
225 230 235 240
Gln Leu Ser Pro Ala Ala Val Glu Ile Gly Arg Thr Ser Arg Ser Thr
245 250 255
Val Ala Asp Leu Ala Arg Arg Thr Val Asp Glu Gly Tyr Gly Val Tyr
260 265 270
Leu Thr Tyr Asn Leu Asp Gly Gly Asp Arg Thr Ala Asp Val Ser Ala
275 280 285
Phe Thr Arg Glu Leu Tyr Gly Ser Glu Ala Val Arg Thr Pro Asp Tyr
290 295 300
Lys Asp Asp Asp Asp Lys Asp Glu Leu
305 310
<210> 3
<211> 30
<212> PRT
<213> Nicotiana benthamiana
<220>
<221> misc_feature
<223> recombinant protein
<400> 3
Met Gly Phe Val Leu Phe Ser Gln Leu Pro Ser Phe Leu Leu Val Ser
1 5 10 15
Thr Leu Leu Leu Phe Leu Val Ile Ser His Ser Cys Arg Ala
20 25 30
<210> 4
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic: FLAG sequence
<400> 4
Asp Tyr Lys Asp Asp Asp Asp Lys
1 5
<210> 5
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 5
Lys Asp Glu Leu
1
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Лиганды к рецептору фолликулостимулирующего гормона (FSH) в диагностике и лечении опухолей | 2017 |
|
RU2755000C2 |
| АКТИВАТОР СИГНАЛЬНОГО ПУТИ ЖАСМОНОВОЙ КИСЛОТЫ | 2016 |
|
RU2728472C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ С MUC1, И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2746413C1 |
| ХИМЕРНЫЕ ВАКЦИНЫ ПРОТИВ ГРИППА | 2021 |
|
RU2834969C1 |
| АНТИТЕЛА К MUC16 И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2758113C2 |
| УСОВЕРШЕНСТВОВАННАЯ КОРОНАВИРУСНАЯ ВАКЦИНА | 2022 |
|
RU2816182C2 |
| НОВЫЙ ВАРИАНТ АЛЬФА-1-АНТИТРИПСИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2567007C1 |
| МУТАНТЫ ГЕМАГГЛЮТИНИНА ВИРУСА ГРИППА | 2019 |
|
RU2801708C2 |
| МУТАНТЫ ГЕМАГГЛЮТИНИНА ВИРУСА ГРИППА | 2019 |
|
RU2809237C2 |


Изобретение относится к области биохимии, в частности к способу получения дегликозилированного полипептида в клетке растения. Изобретение позволяет эффективно получать дегликозилированный полипептид. 6 з.п. ф-лы, 6 пр., 7 ил.
1. Способ получения дегликозилированного полипептида в клетке растения,
включающий:
введение в контакт цитозоля указанной клетки растения с первой нуклеиновой кислотой, включающей первую последовательность для контроля экспрессии и первую экспрессируемую последовательность нуклеиновой кислоты, функционально связанную с указанной первой последовательностью для контроля экспрессии, при этом указанная первая экспрессируемая нуклеиновая кислота содержит последовательность нуклеиновой кислоты, приведённую в SEQ ID NO:1;
введение в контакт указанного цитозоля указанной клетки растения со второй нуклеиновой кислотой, включающей вторую последовательность для контроля экспрессии и вторую экспрессируемую последовательность нуклеиновой кислоты, функционально связанную с указанной первой последовательностью для контроля экспрессии, при этом указанная вторая экспрессируемая последовательность нуклеиновой кислоты кодирует иммуноактивный полипептид;
совместную экспрессию указанной первой нуклеиновой кислоты и указанной второй нуклеиновой кислоты в клетке растения для получения указанного дегликозилированного полипептида,
при этом указанной клеткой растения является клетка Nicotiana benthamiana.
2. Способ по п. 1, отличающийся тем, что указанный полипептид является иммуноактивным в людях.
3. Способ по п. 1, отличающийся тем, что указанная вторая последовательность нуклеиновой кислоты кодирует последовательность пептида, который при получении в его нативных видах не является гликозилированным.
4. Способ по п. 1, отличающийся тем, что указанный полипептид выбран из Pfs48/45, PA83 и фактора XIII человека.
5. Способ по п. 4, отличающийся тем, что указанным полипептидом является PA83, при этом указанный полипептид имеет повышенную стабильность по сравнению с гликозилированным пептидом PA83.
6. Способ по п. 4, отличающийся тем, что указанным полипептидом является PA83, при этом указанный полипептид имеет повышенную стабильность по сравнению с пептидом PA83, дегликозилированным посредством PNGазы.
7. Способ по п. 1, отличающийся тем, что введение в контакт цитозоля указанной клетки растения дополнительно включает трансформирование указанной клетки растения с использованием Agrobacterium.
| WO 2015080603 A1, 04.06.2015 | |||
| US 20150203814 A1, 23.07.2015 | |||
| WO 2013119470 A1, 15.08.2013 | |||
| US 8545909 B2, 01.10.2013 | |||
| ВЫДЕЛЕННЫЙ ОБОЛОЧЕЧНЫЙ БЕЛОК HCV, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЛЕКАРСТВО, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) ЕГО СОДЕРЖАЩИЕ | 2002 |
|
RU2274643C2 |

