Настоящее изобретение относится к области онкологии и молекулярной биологии и может быть использовано в качестве дополнительного метода обследования пациентов с немелкоклеточным раком легких для определения чувствительности опухолевых клеток к действию препаратов, реактивирующих белок р53.
Перспективным направлением лечения онкологических заболеваний считается таргетная терапия, подразумевающая направленное фармакологическое воздействие на ключевой для клеток опухоли регуляторный белок (так называемую мишень для лекарственного препарата). Одним из таких белков-мишеней является белок р53. Белок р53 представляет собой многофункциональный опухолевый супрессор. Его активация в ответ на стрессовые воздействия или нарушение целостности геномной ДНК приводит к остановке клеточного цикла и апоптотической гибели клетки. Потеря клеткой функции белка р53 является центральным событием в формировании злокачественного фенотипа клетки, а также служит одной из причин лекарственной устойчивости опухолевых клеток к действию генотоксических химиопрепаратов. Таким образом, восстановление нормальной активности р53 в клетке считается перспективным подходом к лечению опухолевых заболеваний.
Ключевую роль в негативной регуляции р53 в клетках опухоли играет белок Mdm2, представляющий собой Е3 убиквитин лигазу. В клетке Mdm2 связывается с N-концевым доменом белка р53, при этом происходит инактивация последнего и последующая протеолитическая убиквитинзависимая деградация [1]. Структурный гомолог Mdm2 - Mdm4 (или Mdmx) также участвует в негативной регуляции р53, формируя гетерокомплексы с Mdm2 за счет взаимодействия RING доменов [2]. В клетках опухоли, как правило, имеет место увеличение экспрессии гена MDM2 и (или) MDM4. В результате этого в опухолевых клетках происходит быстрая деградация белка р53, что позволяет им избегать р53-зависимого апоптоза. Разрушение комплекса Mdm2-p53 способствует стабилизации р53 и восстановлению его активности, что в свою очередь приводит к остановке пролиферации и (или) гибели опухолевых клеток, а также к увеличению чувствительности опухоли к лучевой и генотоксической химиотерапии. Эта стратегия может быть перспективна при лечении злокачественных заболеваний, при которых сохраняется экспрессия гена ТР53 дикого типа.
В качестве препаратов, реактивирующих р53, известны соединения, вытесняющие белок р53 из комплекса Mdm2-p53. К таким соединениям относятся (но ими не ограничиваются) Nutlins (нутлины) [3], BDAs (бензодиазепиндионы) [4] и серия Mdm2 ингибиторов (Ml) производных спирооксиндолов, включающих MI-63, MI-219 и MI-43 [5, 6]. Все три серии соединений имитируют аминокислотные остатки F19, W23 и L26 белка р53 и с высокой афинностью связываются с Mdm2 в области р53-специфичного кармана и, таким образом, вытесняют белок р53 из Mdm2-р53 комплекса. В таблице 1 приведен список химических соединений - антагонистов Mdm2, которые на сегодняшний день исследуются в качестве терапевтических препаратов. Информация о текущих клинических испытаниях получена с сайта .
Из всех соединений, ингибирующих взаимодействие белков Mdm2 и р53, наиболее хорошо изучены Nutlins (нутлины). Данные доклинического испытания соединения Nutlin-3а для лечения острого миелоидного лейкоза [7, 8] подтверждают способность Nutlin-3а индуцировать апоптоз бластных клеток (но не нормальных гематопоэтических клеток крови). Результаты доклинического испытания Nutlin-3а при лечения нейробластомы [9] также подтверждают способность Nutlin-3а индуцировать апоптоз в клетках нейробластомы, резистентных к химиотерапии. При этом в обоих случаях цитотоксический эффект Nutlin-3 наблюдался только в случае опухоли с немутантным геном ТР53.
Для назначения таргетной терапии, основанной на использовании лекарственных соединений, реактивирующих белок р53, необходимо определить критерии, характеризующие категорию пациентов, с высокой вероятностью реагирующих на лечение препаратами рассматриваемой группы. Следовательно, актуальной задачей является разработка способов определения чувствительности опухолевых клеток, в частности, клеток немелкоклеточного рака легких (НМРЛ), к лечению препаратами, реактивирующими белок р53.
Наиболее ближайшим к заявляемому способу - прототипом является способ определения чувствительности клеток лимфобластной лейкемии к действию препарата (Nutlin-3), реактивирующего белок р53 [10], включающий следующие стадии: выделение РНК из клеток крови при помощи RNeasy Mini Kit (Qiagen, Germany); синтез кДНК в реакции обратной транскрипции с использованием случайных гексануклеотидных и oligo(dT) праймеров; амплификация фрагмента кДНК гена MDM2 при помощи ПНР с детекцией результатов в режиме реального времени, определение количества кДНК гена MDM2 и определение уровня эксперссии гена MDM2 путем нормирования полученных результатов относительно количества кДНК нормировочного гена. В качестве нормировочного гена авторы используют ген домашнего хозяйства GAPDH.
Недостатком известного способа является значительная вариабельность результатов анализа при неверном выборе одного нормировочного гена. При выборе нормировочного гена предполагается, что уровень его экспрессии в клетках не изменяется в зависимости от типа образца. Однако не существует идеального нормировочного гена, для которого уровень экспрессии будет оставаться постоянным в независимости от ткани и состояния клеток в анализируемом образце.
Задачей предлагаемого изобретения является разработка более достоверного и быстрого способа определения чувствительности клеток немелкоклеточного рака легких к действию препаратов, реактивирующих белок р53.
Технический результат, достигаемый при использовании предложенного способа, заключается в повышении достоверности способа за счет уменьшения ошибки вычисления, вносимой при использовании одного нормировочного гена; в уменьшении числа амплификации за счет исключения необходимости анализировать уровень экспрессии нескольких нормировочных генов.
Поставленная задача достигается предлагаемым способом, заключающимся в том, что в качестве критерия чувствительности клеток НМРЛ рассматривают соотношение количества кДНК гена E2F1 и количества кДНК гена CDKN1A или гена BTG2, при этом относительное количество кДНК генов CDKN1A, BTG2 и E2F1 определяют методом обратной транскрипции и полимеразной цепной реакции с детекцией результатов в режиме реального времени.
Метод обратной транскрипции и полимеразной цепной реакции (ОТ-ПЦР) с регистрацией результатов в режиме реального времени позволяет определить относительное или абсолютное количество молекул мРНК с заданной нуклеотидной последовательностью в образце. Одно из применений метода - это определение уровня экспрессии генов, кодирующих диагностически значимые белковые факторы в клетках различных тканей. Метод ОТ-ПЦР включает в себя три этапа: синтез молекул кДНК (ДНК комплиментарных клеточной РНК) при помощи реакции обратной транскрипции (ОТ); амплификацию последовательности кДНК, соответствующей фрагменту анализируемого гена; анализ полученных результатов и определение относительного количества и уровня экспрессии кДНК анализируемого гена в образце. Количество специфической кДНК в образце с одной стороны зависит от уровня экспрессии мРНК соответствующего гена в клетках образца, с другой стороны, оно определяется объемом клеточного материала, степенью деградации анализируемой РНК и наличием в образце примесей ингибирующих реакцию ОТ. Для того чтобы учесть влияние дополнительных факторов уровень экспрессии кДНК анализируемого гена рассчитывается относительно экспрессии кДНК нормировочного гена. В качестве таких нормировочных генов используют, как правило, гены «домашнего хозяйства». Однако не существует идеального нормировочного гена, для которого уровень экспрессии будет оставаться постоянным в независимости от ткани и состояния клеток в анализируемом образце, поэтому на практике нормирование осуществляют с использованием одновременно нескольких нормировочных генов. При этом нормировочный коэффициент вычисляется как среднее арифметическое от значений относительного количества кДНК всех нормировочных генов [11].
Более конкретно, предложенный способ заключается в определении относительного количества кДНК генов CDKNJA, BTG2 и E2F1 методом ОТ-ПЦР в режиме реального времени с последующим расчетом соотношения количества кДНК гена E2F1 к количеству кДНК гена CDKN1A или гена BTG2 и при величине соотношения E2F1/CDKN1A>3 или E2F1/BTG2>1,5 считают клетки НМРЛ чувствительными к препаратам, реактивирующим белок р53.
Относительное количество кДНК рассчитывают при помощи калибровочной кривой, построенной при амплификации стандартных образцов ДНК, взятых в 5-ти последовательно убывающих известных концентрациях. Стандарт представляет собой смесь фрагментов ДНК, несущих амплифицируемую последовательность к ДНК генов CDKN1A, BTG2 и E2F1 длиной по 500 пар нуклеотидов. Относительное количество кДНК каждого гена определяют исходя из уравнения калибровочной кривой, при этом учитывают эффективность амплификации каждого образца. Далее вычисляют маркеры, учитывающие динамику клеточного цикла (баланс факторов, регулирующих G1/S арест клеточного цикла и стадию репликации).
Маркеры вычисляют по формуле:
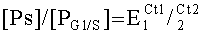
где PS - количество кДНК гена фактора, контролирующего репликацию;
PG1/S- количество к ДНК гена фактора, контролирующего G1/S арест клеточного цикла;
E1 и Е2 - эффективность амплификации генов Ps и PG]/S соответственно.
Ct1 и Ct2 - значения порогового цикла ПЦР, автоматически определяемые прибором при амплификации фрагментов генов PS и PG1/S соответственно.
Предлагаемый способ включает следующие стандартные стадии.
Выделение РНК из образцов.
В качестве образцов для проведения анализа могут быть взяты биоптаты и(или) пунктаты ткани опухоли, культуры.
Способ выделения суммарной РНК из образцов ткани млекопитающих включает следующие стадии: измельчение образцов в гомогенизаторе или в жидком азоте (относится к плотным, сложно гомогенизируемым фрагментам ткани), лизис клеток, выделение РНК и ее очистку, проверку качества РНК электрофорезом в 1-1,5%-ном агарозном геле в присутствии красителя бромида этидия, а также спектрофотометрическое определение количества РНК. Гомогенизацию кусочков ткани проводят вручную, растирая пестиком в керамической ступке, или с помощью механических гомогенизаторов, например TissueLyser (Qiagen). Для лизиса клеток и выделения РНК используют сильные хаотропные агенты в сочетании с детергентами, такими как гуанидинизотиоцианат и саркозил, далее последовательно проводят экстракцию фенолом при рН5 5,0 и хлороформом для денатурации и удаления белков и геномной ДНК [Chomczynsci P., Sacci N. Single-step method of RNA isolation by Acid guanidinium thiocyanate-phenol-chloroform extraction. Analytical Biochemistry. 1987/ Vol 162. P 156-159]. Далее РНК осаждают в присутствии изопропанола или этанола или сорбируют на твердом носителе, например, Silica S-5631(Sigma) или спин-колонки (Qiagen; Promega). Для предотвращения разрушения РНК РНКазами могут быть использованы ингибиторы рибонуклеаз, такие, например, как рекомбинантный RNAsin (Promega). Для элиминации примесей геномной ДНК можно обработать полученный препарат РНК очищенной от рибонуклеаз ДНАказой. Выделение РНК осуществляют с использованием реагента Trizol®Reagent [Invitrogen] или при помощи коммерчески доступных наборов, таких как RNeasy kits (Qiagen), SV Total RNA Isolation System (Promega) и т.д.
Синтез кДНК при помощи реакция обратной транскрипции.
В результате реакции обратной транскрипции на РНК-матрице синтезируется одноцепочечная цепь ДНК. Образующиеся при этом молекулы кДНК более стабильны по сравнению с нестабильными молекулами РНК и могут быть амплифицированы с помощью полимеразной цепной реакции до количеств, необходимых для детекции.
Реакцию обратной транскрипции проводят при помощи различных коммерческих препаратов обратных транскриптаз, например, обратная транскриптаза вируса лейкоза мышей Молони (M-MLV), вируса миелобластоза птиц (AMV), PowerScript (точечная мутация M-MLV-RT), С.Therm Polymerase и др. Может быть использована термостабильная ДНК-полимераза Thermus thermophilus (Tth), обладающая обратной транскриптазной активностью в присутствии ионов Мn2+.
Реакция обратной транскрипции начинается с формирования двухцепочечной затравки, для этого используют различные типы праймеров. Например, oligo(dT)n праймеры (число n обычно равно 12-18), связывающиеся с эндогенным полиА-хвостом на 3'-конце мРНК. Эти праймеры используют для получения полноразмерных кДНК. К 3'-концу oligo(dT) последовательности могут быть добавлены нуклеотиды А, С или G, чтобы «заякорить» праймер на границе транскрипта и поли-А тракта.
Также могут быть использованы случайные гексануклеотидные праймеры (статистические праймеры), гибридизующиеся в нескольких участках на протяжении РНК. В реакции обратной транскрипции с использованием статистических праймеров синтезируются более короткие (в среднем около 500 п.н.) фрагменты кДНК. Реакция обратной транскрипции с использованием статистических праймеров, как правило, проходит более эффективно, в особенности при обратной транскрипции GC-богатых районов и 5'-областей мРНК.
Кроме гексамерных праймеров могут быть использованы и более длинные 9-12-мерные праймеры. Также можно использовать статистические праймеры в комбинации с oligo(dT) праймерами или специфическими праймерами, комплиментарными последовательности анализируемого гена.
Амплификация кДНК генов CDKN1A, BTG2 и E2F1 при помощи ПЦР с детекцией в режиме реального времени.
Амплификацию фрагментов транскриптов генов CDKN1A, BTG2 и E2F1 выполняют при помощи пар праймеров, комплиментарных транскриптам. При выборе праймеров для амплификации кДНК учитывают экзон-интронную структуру гена и размещают праймеры таким образом, чтобы их сайты гибридизации располагались в различных экзонах. Таким образом, достигается супрессирование возможных примесей геномной ДНК в образце.
Реакцию амплификации выполняют с использованием коммерчески доступного препарата ДНК полимеразы Thermus aquaticus, например HS Taq ДНК полимераза, (Евроген) GoTaq© Hot Start Polymerase (Promega) и др. В качестве матрицы используют кДНК, синтезированную на матрице суммарной клеточной РНК.
Накопление продуктов амплификации кДНК CDKN1A, BTG2 и E2F1 детектируют в режиме реального времени при помощи одного из способов:
- или при помощи интекалирующего с двухцепочечной ДНК флуоресцентного красителя с высоким индексом дискриминации флуоресценции у связанной и несвязанной форм, например, SYBR GreenI (Invitrogen);
- или при помощи олигонуклеотидной пробы типа TaqMan комплиментарной центральной части амплифицируемого фрагмента транскрипта. Проба длиной 20-30 нуклеотидов несет на 5'-конце флуорофор (FAM, R6G, ROX) и на 3'-конце не флуоресцентный тушитель (BHQ, FTQ и др.). На каждом цикле амплификации гибридизованная проба гидролизуется со стороны 5'-конца, при этом флуорофор пространственно разобщается с тушителем и, как следствие, увеличивается флуоресценция реакционной смеси.
Для выбора последовательности праймеров и проб типа TaqMan используют доступное программное обеспечение, например OligoAnalyser v. 1.0.3, BeaconDisigner v. 7.0 и другие. Структуры предпочтительных пар праймеров и гибридизационных проб приведены в таблице 2.
Определение количества кДНК генов CDKN1A, BTG2 и E2F1 выполняют при помощи уравнения калибровочной кривой с учетом эффективности каждой реакции амплификации. Для построения калибровочной кривой одновременно с анализируемыми образцами амплифицируют стандартные образцы ДНК с известной последовательностью, уменьшающейся в три раза концентрацией ДНК. Стандартный образец представляет собой смесь двуцепочечных фрагментов ДНК длиной 500 п.н., несущих последовательности амплифицируемых фрагментов транскриптов CDKN1A, BTG2 и E2F1. Калибровочную кривую строят в зависимости от значений Ct и Log количества ДНК в стандартном образце. В соответствии с полученным графиком калибровочной кривой при помощи регрессионного анализа определяют коэффициенты линейного уравнения калибровочной кривой. Полученное уравнение используют для определения количества кДНК CDKN1 A, BTG2 и E2F1 в анализируемых образцах.
Ниже изобретение подробно проиллюстрировано конкретным примером.
Пример.
Материалом исследования являлись клеточные линии НМРЛ, несущие ген ТР53 дикого типа. Всего было проанализировано 8 клеточных линий: А549, NCI-H292, А427, COR-L23, DV-90, NCI-H1395, NCI-H1944, NCI-H2228.
Культивирование клеточных линий НМРЛ.
Чувствительность клеток к соединению Nutlin3(±) (Cayman Chemical) оценивали по значению параметра IC50 (inhibitory concentration; концентрация препарата, при которой в культуре наблюдается деление 50% клеток). Для этого клетки А427, находящиеся в экспоненциальной фазе роста, высевали в лунки 24-луночного планшета по 60 тыс.клеток на лунку. Клетки культивировали в среде DMEM/F12, содержащей 10% FBS, 0.03% глутамина и канамицин в концентрации 100 мкг/мл, при 37°С и 5%-ном содержании СO2 в атмосфере. Через 24 часа культуральную среду заменяли на свежую, содержащую Nutlin3(±) в концентрации от 50 мкМ до 1 мкМ (каждая точка была представлена в тройном повторе). Через 48 часов после добавления химиопрепарата определяли долю жизнеспособных клеток при помощи теста на жизнеспособность с резазурином. Резазурин (резазурина натриевая соль, Диаэм) добавляли к клеткам в концентрации 50 мкг/мл. Объем культуральной среды составлял 500 мкл. В качестве контроля (фон) использовали 500 мкл культуральной среды, не содержащей клеток НМРЛ. Продолжительность времени инкубации клеток с резазурином составляло 1 час. Инкубирование осуществлялось в стерильных условиях, при 99% влажности, температуре 37°С и концентрации СO2 - 5%. Накопление резоруфина (восстановленной формы резазурина) оценивали при помощи детекции флуоресценции в диапазоне 587/607 нм. Для оценки результатов теста строилась калибровочная кривая изменения флуоресценции в зависимости от числа клеток. Значения IC50 для каждой клеточной линии приведены в таблице 3.
Выделение РНК и синтез кДНК в реакции обратной транскрипции.
Клетки, не подвергавшиеся воздействию Nutlin3(±) и находящиеся в экспоненциальной фазе роста, высевали в лунки 6-луночного планшета и культивировали до 70%-ной плотности монослоя. После этого культуральную среду удаляли, а клетки лизировали реагентом Trisol («Invitrogen»). Полученные препараты хранили при -70°С. Суммарную клеточную РНК выделяли согласно рекомендациям Invitrogen. Качество препаратов РНК оценивали по итогам электрофоретического разделения в 1,3% агарозном геле. Реакцию обратной транскрипции проводили при 42°С в течение 45 мин в 20 мкл реакционной смеси, содержащей 10мМ Tris-HCl (рН 8,3), 5 мМ MgC12, 10 мМ DTT, 50 мМ KСl, 0,2 мМ dNTP, Stat-9 праймер (10нг/мкл), 100 ед.акт. ДНК зависимой РНК полимеразы MoMLV (Biosan, ИХБФМ СОР АН) и 500 нг суммарной клеточной РНК.
Амплификация кДНК генов CDKN1A, BTG2 и E2F1 с детекцией в режиме реального времени.
Реакция амплификации выполнялась при помощи термоциклера с оптическим блоком для детекции флуоресценции CFX96 (Bio-Rad). Возможность регистрации флуоресценции в режиме реального времени достигалась при помощи гибридизационных проб типа TaqMan.
ПНР проводилась в объеме 25 мкл, реакционная смесь содержала 65 мМ Tris-HCl (рН 8.9); 24 мМ (NH4)2SO4; 0,05% Tween 20; 2,5 мМ MgCl2, 0,2mM dNTP, 300 nM праймеры (прямой и обратный), 100 nM Taq-Man проба, 0.5 ед.акт.Taq-ДНК полимеразы (Biosan, ИХБФМ СОРАН) и 0.1-5 нг кДНК.
Протокол амплификации:
- первичная денатурация - 3 минуты при 96°С;
- амплификационный цикл (×40);
- денатурация - 10 секунд при 96°С;
- отжиг праймеров, элонгация и съем флуоресцентного сигнала - 60 секунд при 60°С.
Специфичность амплификации подтверждалась электрофоретическим анализом в полиакриламидном геле.
Для амплификации использовались соответствующие гибридизационные пробы и прямой и обратный праймеры. Структуры олигонуклеотидных праймеров и гибридизационных проб представлены в таблице 2.
Анализ результатов ОТ-ПЦР.
Всего было проанализировано 8 образцов. Для каждого образца при помощи программного обеспечения прибора определяли номер порогового цикла (Ct), при котором величина флуоресценции реакционной смеси достигал некоторого «порогового» значения (Т) одинакового для всех сравниваемых образцов. Определение относительного количества кДНК CDKN1A, BTG2 и E2F1 (Х0) в анализируемых образцах рассчитывали по формуле:
Log(X0)=а+b·Ct,
где а=log(T); b=log(1+E); Е - эффективность ПЦР, одинаковая для всех анализируемых образцов.
Эффективность амплификации определялась как угол наклона калибровочной кривой. Для построения калибровочной кривой одновременно с анализируемыми образцами амплифицировались 5 стандартных образцов с последовательно уменьшающейся в три раза концентрацией ДНК (1 у.е., 3 у.е., 9 у.е., 27 у.е. и 81 у.е.). Линейное уравнение калибровочной кривой определяло взаимосвязь значений Ct, полученных при амплификации пяти стандартных образцов и значений lg(Xo) этих образцов (в нашем случае это lg1, lg3, lg9, lg27, lg81).
Для всех пар праймеров эффективность амплификации составила не меньше 90%.
Для оценки воспроизводимости значений порогового цикла (Ct) все анализируемые образцы с каждой парой праймеров амплифицировались в тройном повторе. Разница значений Ct между повторами была не более 0,3 цикла.
Сравнение значений IC50 соединения Nutlin3(±) и уровня экспрессии генов CDKN1A, BTG2 и E2F1 в клетках НМРЛ.
Среди восьми клеточных линий НМРЛ были выделены три линии (NCI-H2228, NCI-H1944, А549), резистентных к действию Nutlin3(±) для которых среднее значение IC50 составило 24 мкМ±4,5 мкМ и пять линий (NCI-H292, А427, COR-L23, DV-90, NCI-H1395), чувствительных к действию Nutlin3(±) для которых среднее значение IC50 составило 7,9 мкМ±2,9 мкМ. Значения IC50 соединения Nutlin3(±) для восьми клеточных линий НМРЛ приведены в таблице 3.
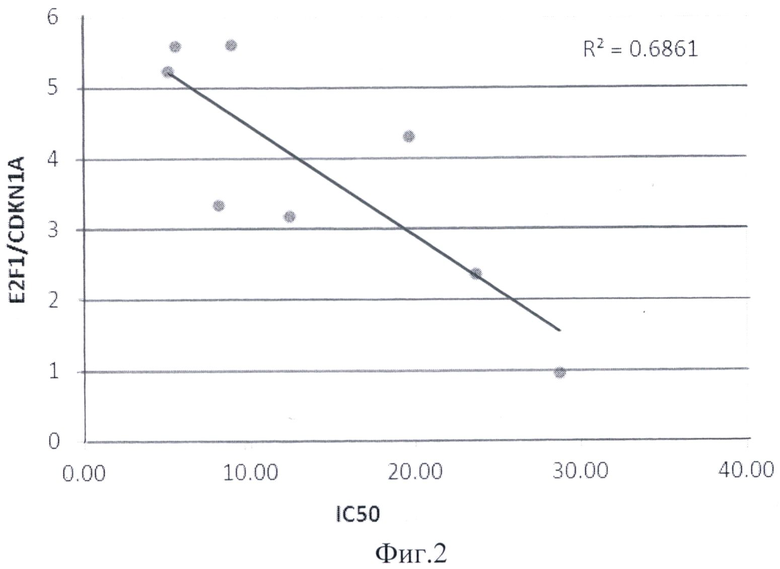
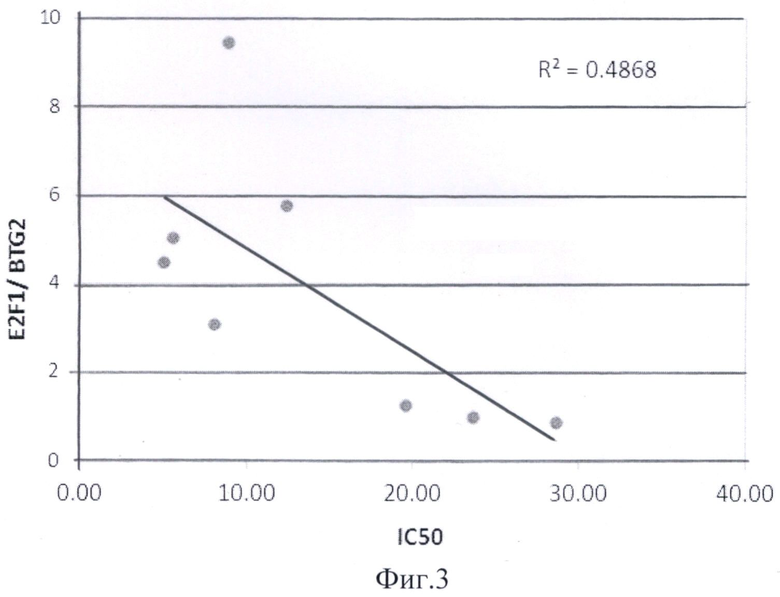
Соотношение уровня экспрессии E2F1/CDKN1A и E2F1/ BTG2 было достоверно выше в группе чувствительных к действию Nutlin3(±) линий НМРЛ по сравнению с группой резистентных к действию Nutlin3(±) линий НМРЛ (р=0,045 и 0,007 соответственно). Значения соотношений E2F1/CDKN1A и E2F1/ BTG2 для каждой клеточной линии приведены на фиг.1.
Зависимость соотношения E2F1/CDKN1 и соотношения E2F1/BTG2 от значения IC50 соединения Nutlin3(±) для восьми клеточных линий НМРЛ представлены на фиг.2, 3. Полученные значения соотношений уровней экспрессии E2F1/CDKN1A и E2F1/ BTG2 достоверно коррелировали со значениями IC50 с коэффициентом корреляции R=0,64 и R2=0,48. Использование предлагаемого способа позволит более точно прогнозировать чувствительность клеток немелкоклеточного рака легких к действию препаратов, реактивирующих белок р53.
Источники информации
1. Haupt, Y., Maya, R., Kazaz, A. & Oren, M. Mdm2 promotes the rapid degradation of p53. Nature (1997) 387, 296-299.
2. Wade, M., Wang, Y. V. & Wahl, G. M. The p53 orchestra: Mdm2 and Mdmx set the tone. Trends Cell Biol. (2010) 20, 299-309.
3. Vassilev LT, Vu ВТ, Graves B, Carvajal D, Podlaski F, Filipovic Z, Kong N, Kammlott U, Lukacs C, Klein C, et al. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science. (2004); 303(5659), 844-848.
4. Grasberger BL, Lu T, Schubert C, Parks DJ, Carver ТЕ, Koblish HK, Cummings MD, LaFrance LV, Milkiewicz KL, Calvo RR, et al. Discovery and cocrystal structure of benzodiazepinedione HDM2 antagonists that activate p53 in cells. J Med Chem (2005) 48: 909-912.
5. Shangary S, Qin D, McEachern D, Liu M, Miller RS, Qiu S, Nikolovska-Coleska Z, Ding K, Wang G, Chen J, et al. Temporal activation of p53 by a specific MDM2 inhibitor is selectively toxic to tumors and leads to complete tumor growth inhibition. Proc Natl Acad Sci USA. 2008; 105(10): 3933-3938.
6. Ding K, Lu Y, Nikolovska-Coleska Z, Wang G, Qiu S, Shangary S, Gao W, Qin D, Stuckey J, Krajewski K, et al. 2006. Structure-based design of spiro-oxindoles as potent, specific small-molecule inhibitors of the MDM2-p53 interaction. J Med Chem 49: 3432-3435.
7. Kojima, K. et al. MDM2 antagonists induce p53-dependent apoptosis in AML: implications for leukemia therapy. Blood (2005) 106, 3150-3159.
8. Carter, B. Z. et al. Simultaneous activation of p53 and inhibition of XIAP enhance the activation of apoptosis signaling pathways in AML. Blood (2010) 115,306-314. 9. Van Maerken, Т. et al. Antitumor activity of the selective MDM2 antagonist nutlin-3 against chemoresistant neuroblastoma with wild-type p53. J. Natl Cancer Inst. (2009) 101, 1562-1574.
10. L Gu, N Zhu, H W Findley and M Zhou. MDM2 antagonist nutlin-3 is a potent inducer of apoptosis in pediatric acute lymphoblastic leukemia cells with wild-type p53 and overexpression of MDM2Nutlin-3 induces apoptosis in ALL cells. Leukemia (2008) 22, 730-739.
11. Tricarico, C, Pinzani, P., Bianchi, S., Paglierani, M., Distante, V., Pazzagli, M., Bustin, S.A., Orlando, C. Quantitative real-time reverse transcription polymerase chain reaction: normalization to rRNA or single housekeeping genes is inappropriate for human tissue biopsies. // Anal Biochem. (2002) - N309. - P.293-300.
Способ определения чувствительности клеток немелкоклеточного рака легких к действию препаратов, реактивирующих белок р53
Стадия-I NCT00623870 и NCT00559533
(гематологические неоплазии)
5' -CGTTTGG AGTGGTAG A AATCTGT-3'
5'-FAM- CTTGTGCCTCGCTCAGGGGAGC-BHQ- 3'
5 '-GCTGCGTAGTACAGATATTCATCA-3'
5'-FAM- CCACATCCAGTGGCTGGGCAGC-BHQ-3'
5 '-TTCAGCC AAGGAATACATGCAA-3'
5'- FAM-CTGCTCTCCTTGGGATGATGGC-BHQ-3'
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ УРОВНЯ ТРАНСКРИПЦИИ ГЕНА, КОДИРУЮЩЕГО ХЕМОКИН CCL2 (МСР-1) ЧЕЛОВЕКА И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2522801C2 |
| ПЕПТИДЫ, СПОСОБНЫЕ РЕАКТИВИРОВАТЬ МУТАНТЫ Р53 | 2014 |
|
RU2693487C2 |
| ЛЕЧЕНИЕ ОПУХОЛЕЙ КОМБИНАЦИЕЙ ОНКОЛИТИЧЕСКОГО АДЕНОВИРУСА, ИНГИБИТОРА CDK4/6 И ДОПОЛНИТЕЛЬНОГО ТЕРАПЕВТИЧЕСКОГО АГЕНТА | 2020 |
|
RU2835886C1 |
| ЛЕЧЕНИЕ ОПУХОЛЕЙ КОМБИНАЦИЕЙ ОНКОЛИТИЧЕСКОГО АДЕНОВИРУСА И ИНГИБИТОРА CDK4/6 | 2019 |
|
RU2811278C2 |
| ИММОРТАЛИЗОВАННЫЕ ЛИНИИ КЛЕТОК ПТИЦ ДЛЯ ПОЛУЧЕНИЯ ВИРУСОВ | 2004 |
|
RU2359999C2 |
| ПЕПТИДЫ, СПОСОБНЫЕ РЕАКТИВИРОВАТЬ МУТАНТЫ Р53 | 2014 |
|
RU2721423C2 |
| СПОСОБ ДИАГНОСТИКИ НЕМЕЛКОКЛЕТОЧНОГО РАКА ЛЕГКИХ И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2010 |
|
RU2445627C1 |
| СПОСОБ ДИАГНОСТИКИ ПЛОСКОКЛЕТОЧНОГО РАКА ЛЕГКИХ И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2006 |
|
RU2327162C1 |
| СПОСОБ ДИАГНОСТИКИ НЕМЕЛКОКЛЕТОЧНОГО РАКА ЛЕГКИХ И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2006 |
|
RU2330285C1 |
| СПОСОБ ДИАГНОСТИКИ ПЛОСКОКЛЕТОЧНОГО РАКА ЛЕГКОГО И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2014 |
|
RU2586779C2 |
Изобретение относится к области онкологии и молекулярной биологии. Предложен способ определения чувствительности клеток немелкоклеточного рака легких к действию препаратов, реактивирующих р53 белок, включающий выделение РНК из образцов, синтез кДНК генов CDKN1A, BTG2 и E2F1 методом обратной транскрипции и полимеразной цепной реакции с детекцией результатов в режиме реального времени с последующим определением соотношения количества кДНК гена E2F1 к количеству кДНК гена CDKN1A или гена BTG2, где при величине соотношения E2F1/CDKN1A>3 или E2Fl/BTG2>1,5 считают клетки немелкоклеточного рака легких чувствительными к препаратам, реактивирующим белок р53. Изобретение может быть использовано при лечении онкологических заболеваний. 3 ил., 3 табл., 1 пр. 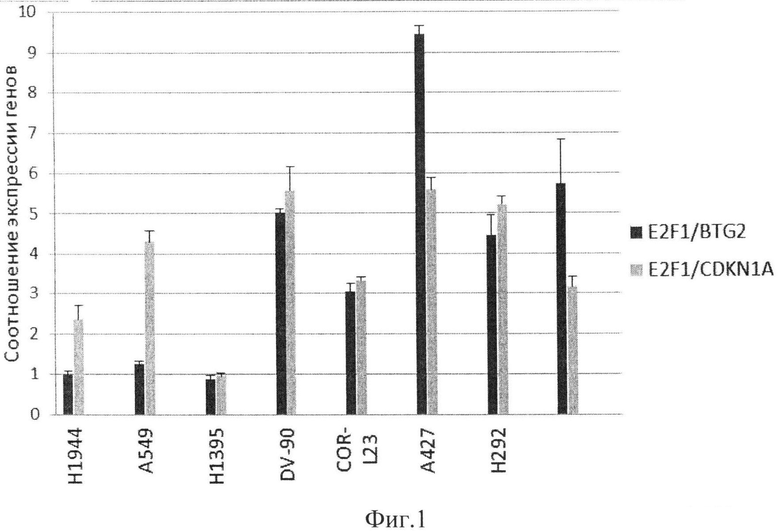
Способ определения чувствительности клеток немелкоклеточного рака легких (НМРЛ) к действию препаратов, реактивирующих белок р53, включающий выделение РНК из образцов, синтез кДНК при помощи реакции обратной транскрипции, амплификацию кДНК соответствующих генов при помощи ПЦР с детекцией в режиме реального времени с последующим анализом уровня экспрессии кДНК выбранных генов, отличающийся тем, что определяют относительное количество кДНК генов CDKN1A, BTG2 и E2F1 методом ОТ-ПЦР в режиме реального времени, далее рассчитывают соотношение количества кДНК гена E2F1 к количеству кДНК гена CDKN1A или гена BTG2 и при величине соотношения E2F1/CDKN1A>3 или E2F1/BTG2>1,5 считают клетки НМРЛ чувствительными к препаратам, реактивирующим белок р53.
| US 20120136583 A1, 31.05.2012 | |||
| Прибор для посадки уплотнительных колец на цилиндрические золотники или поршни | 1930 |
|
SU19877A1 |
| БИОМАРКЕРЫ | 2005 |
|
RU2429297C2 |
| US 20110097717 A1, 28.04.2011 | |||
| US 20110269145 A1, 03.11.2011 | |||
| СОРТОВОЙ ПРОКАТ КРУГЛЫЙ ИЗ БОРСОДЕРЖАЩЕЙ СТАЛИ ДЛЯ ХОЛОДНОЙ ОБЪЕМНОЙ ШТАМПОВКИ | 2006 |
|
RU2336316C2 |
| ЛОГАРИФМИЧЕСКИЙ АРИФМОМЕТР | 1928 |
|
SU12452A1 |

