Изобретение относится к медицине, микробиологической промышленности и может быть использовано для получения методами генной инженерии рекомбинантных химерных полипептидов, несущих эпитопы различных иммунодоминантных белков спирохет комплекса Borrelia burgdorferi sensu lato. Указанные полипетиды могут быть использованы для диагностики иксодового клещевого боррелиоза (Лайм-боррелиоз, болезнь Лайма) методами иммуноферментного анализа (ИФА), иммунохроматографического анализа и вестерн-блот анализа, а также для получения сывороток животных, содержащих поликлональные антитела к соответствующим рекомбинантным химерным полипептидам, и создания молекулярных вакцин против возбудителя иксодового клещевого боррелиоза.
Иксодовый клещевой боррелиоз (ИКБ) - это инфекционное трансмиссивное природноочаговое заболевание, вызываемое спирохетами комплекса Borrelia burgdorferi sensu lato (B.burgdorferi s.l.), передающееся через иксодовых клещей. В настоящее время охарактеризовано 18 видов спирохет, входящих в состав комплекса В.burgdorferi s.l., из которых только четыре: В.burgdorferi sensu stricto, B.garinii, B.afzelii и B.spielmanii, являются патогенными для человека (1). По широте распространения и уровню заболеваемости ИКБ занимает одно из ведущих мест среди природно-очаговых заболеваний. Географическое распространение ИКБ обширно, оно встречается на всех континентах (кроме Антарктиды). В нашей стране природные очаги этой инфекции широко распространены в лесной зоне, от границ с Прибалтикой на западе до Южного Сахалина - на востоке (2). В России обнаружены четыре из шести видов боррелий, встречающихся в мире. Виды B.afzelii и B.garinii распространены по всей территории страны, от европейской части до Дальневосточного региона (3). B.burgdorferi sensu stricto обнаружена только на европейской части территории России - в Московской и Ленинградской областях (4, 5, 6). За последнее десятилетие проведены работы по изучению зараженности иксодовых клещей, распространенных на территории Западной Сибири, спирохетами комплекса Borrelia burgdorferi sensu lato и определению видового состава боррелий (B.afzelii, B.garinii и, предположительно B.japonica, патогенность последнего не выявлена) (7, 8, 9).
Иксодовый клещевой боррелиоз представляет серьезную опасность для здоровья людей, поскольку может приводить к длительной нетрудоспособности и инвалидности. Клинически заболевание протекает с преимущественным поражением кожи, нервной системы, опорно-двигательного аппарата, сердца и характеризуется склонностью к хроническому, а также латентному течению (10).
Диагностика ИКБ является одной из актуальных проблем здравоохранения, и это обусловлено многими факторами: разнообразием клинических проявлений, наличием сопутствующих инфекций, возбудители которых переносятся иксодовыми клещами (анаплазмоз, бабезиоз, вирусный клещевой энцефалит и др.) (11), зависимостью проявлений инфекции как от видовой принадлежности возбудителя, так и от индивидуальных особенностей организма и др. На ранней стадии ИКБ хорошо поддается лечению антибиотикотерапией. На поздних стадиях лечение является более длительным и не всегда успешным. В раннем периоде развития ИКБ выделяют раннюю стадию инфекции (стадию локальной инфекции) и следующую за ней стадию дисссеминированной инфекции, соответствующую диссеминации боррелий в различные органы. Заболевание может иметь стертую клиническую картину течения или латентную форму или маскироваться под другие заболевания, что затрудняет его диагностику.
Основными методами, позволяющими быстро диагностировать ИКБ, являются методы, основанные на выявлении антител к возбудителю, при этом в качестве антигенов используют лизат боррелий или белки, входящие в их состав.
Известен лизат боррелий B.burgdorferi s.1., содержащий белки боррелий, которые используют в качестве антигенов для диагностики ИКБ (12). Недостатками использования антигенов в составе лизата боррелий является то, что лизат включает также белки теплового шока р75 и р60, имеющие гомологию аминокислотных последовательностей с белками, выполняющими сходные функции у ряда грам-отрицательных, грам-положительных микроорганизмов и микобактерий (13). Кроме того, флагеллин боррелий имеет общие эпитопы с флагеллинами других спирохет (T.pallidum, T.phagedenis B.persica, B.hermsii, B.duttoni и др.). Поэтому применение антигенов лизата боррелий в лабораторной практике приводит к получению ложноположительных результатов при тестировании сывороток больных сифилисом, пародонтозом, лептоспирозом, туберкулезом и многими другими заболеваниями (14). Специфичность анализа при использовании антигенов лизата боррелий невелика и составляет 60-70%.
Известны антигены внешних поверхностных белков и декорин-связывающих белков нативных штаммов боррелий (спирохет B.burgdorferi s.1.), которые используют для диагностики ИКБ (15). Однако при многократном пассировании культуры in vitro, боррелии теряют свои внехромосомные элементы (плазмиды), кодирующие иммунодоминантные белки, в том числе внешние поверхностные и декорин-связывающие белки. Это ограничивает количество получаемых антигенов ввиду небольшого числа циклов размножения возбудителя в условиях in vitro и делает их использование непрактичным.
В последние 15 лет для диагностики ИКБ в качестве антигенов используют иммунодоминантные белки возбудителей ИКБ, синтезируемые рекомбинантными штаммами Escherichia coli.
Известен лизат клеток Е.coli, продуцирующих поверхностный иммунодоминантный белок боррелий BmpA, используемый для диагностики ИКБ (16). Недостатком заявленного диагностического средства является то, что использование неочищенного из лизата белка BmpA существенно снижает чувствительность диагностики. Кроме того, он не может быть использован для диагностики ранней стадии ИКБ.
Известен рекомбинантный флагеллярный белок спирохет B.burgdorferi s.1. FlaA массой 38 кДа, используемый в качестве антигена для диагностики ИКБ на ранней стадии заболевания (17). Однако он не может быть использован для серодиагностики стадии диссеминированной инфекции ИКБ.
Известен рекомбинантный флагеллярный белок спирохет B.burgdorferi s.1. FlaB массой 41 кДа, используемый в качестве антигена для диагностики ИКБ, который вызывает иммунный ответ уже на ранних стадиях развития ИКБ, а иммуноглобулины к FlaB сохраняются в высоком титре на протяжении всей инфекции (18). Недостатком данного антигена является то, что он не позволяет дифференцировать раннюю стадию и стадию диссеминированной инфекции ИКБ.
Известен рекомбинантный белок OspC из семейства внешних поверхностных липопротеинов боррелий, используемый в качестве антигена для диагностики ИКБ (15), который является сильным иммунодоминантным белком B.burgdorferi s.1., индуцирующим гуморальный иммунный ответ на ранней стадии инфекции. Недостатком OspC является его непригодность для диагностики стадии диссеминированной инфекции ИКБ.
Известен рекомбинантный белок OspA из семейства внешних поверхностных белков боррелий, используемый в качестве антигена для диагностики ИКБ (19). Данный маркер позволяет диагностировать ИКБ с преимущественным поражением суставов на стадии диссеминированной инфекции, он не пригоден для диагностики ранней стадии ИКБ
Известна комбинация рекомбинантных белков OspA и VIsE (variable major protein-like sequence, expressed), совместно используемых в качестве антигенов для диагностики ИКБ на стадии диссеминированной инфекции. Данная комбинация антигенов недостаточна для исключения ложноположительной реакции у здоровых пациентов (20). Кроме того, она не позволяет диагностировать раннюю стадию ИКБ.
Известен рекомбинантный декоринсвязывающий белок (DbpA) боррелий, используемый в качестве антигена для диагностики ИКБ на стадии диссеминированной инфекции, что позволяет выявлять 100% больных с такой формой ИКБ, как нейроборрелиоз и 62% при ИКБ с преимущественным поражением суставов (21). Использование DbpA без сочетания с другими маркерами ИКБ не позволяет диагностировать раннюю стадию заболевания.
Диагностическую значимость для выявления ИКБ на стадии диссеминированной инфекции имеет также другой декоринсвязывающий белок боррелий - DbpB (22). Однако установлено, что использование декоринсвязывающих белков в качестве антигенов для диагностики ИКБ не позволяет диагностировать раннюю стадию заболевания (22).
Известен рекомбинантный белок боррелий ВтрА (р39) из семейства основных мембранных белков, используемый в качестве антигена для диагностики ИКБ на стадии диссеминированной инфекции. BmpA вызывает сильный иммунный ответ в организме. Зрелый белок BmpA имеет молекулярную массу около 39 кДа и является наряду с флагеллярными белками достаточно консервативным иммуногенным белком спирохет комплекса B.burgdorferi s. 1.: гомология аминокислотных последовательностей BmpA различных видов спирохет В.burgdorferi s.1., патогенных для человека и распространенных в Европе, составляет от 86% до 91% (23). Чувствительность диагностических тест-систем, основанных на использовании BmpA, полученного на основе одного вида боррелий зависит от распространенности данного вида в том регионе, в котором инфицирован пациент. Использование BmpA для диагностики ИКБ не позволяет выявлять раннюю стадию заболевания.
Известен рекомбинантный полипептид, включающий фрагмент аминокислотной последовательности (позиции 139-273) OspC боррелий штамма Borrelia burgdorferi sensu stricto, рекомбинантный полипептид, включающий фрагмент 160-170 указанной аминокислотной последовательности OspC, а также рекомбинантный химерный полипептид, включающий комбинацию фрагментов аминокислотных последовательностей OspC (1-164, 165-179, 180-216, 217-273) от четырех разных штаммов боррелий, которые могут быть использованы для получения молекулярных вакцин против ИКБ (24). Недостатком применения названных рекомбинантных полипептидов и химерного полипептида для диагностики ИКБ в Сибири является то, что источником генов для получения данных полипептидов являются типичные европейские изоляты боррелий, существенно отличающиеся по антигенным свойствам от спирохет, распространенных в Сибири. Кроме того, использование OspC или его фрагментов, в том числе в составе рекомбинантного химерного полипептида, для диагностики ИКБ не позволяет диагностировать стадию диссеминированной инфекции.
Известны рекомбинантные белки OspA, OspB, их фрагменты, рекомбинантный химерный полипептид OspA-OspB, полипептид FlaB, которые могут быть использованы для диагностики ИКБ (25). Недостатком применения названных рекомбинантных полипептидов для диагностики ИКБ в России является то, что они получены на основе генов североамериканских изолятов Borrelia burgdorferi sensu stricto (штаммы N40, В31 и ZS7), которые в России встречаются только в Московской и Ленинградской областях и отсутствуют на других территориях, в том числе в Сибири. Кроме того, выбранная структура гена flaB кодирует перекрестные эпитопы с флагеллярными белками других видов патогенов, главным образом, возбудителя сифилиса - Treponema palladium (29), что обусловливает получение ложноположительных результатов при использовании для диагностики рекомбинантного полипептида FlaB, кодируемого данным геном. Рекомбинантные белки, получаемые согласно данному аналогу, синтезируются слитыми с глутатионтрансферазой, что может приводить к искажениям результатов при диагностике ИКБ.
Известны рекомбинантные белки OspA и FlaB, кодируемые полноразмерными генами ospA и flab, полученными амплификацией методом ПЦР на ДНК североамериканского изолята Borrelia burgdorferi, штамм ZS7 (26). Недостатком применения их для диагностики ИКБ является то, что они обладают отличающимися антигенными свойствами, по сравнению с аналогичными полипептидами, кодируемыми генами изолятов боррелий, встречающихся на территории России, в частности в Сибири, что снижает эффективность использования основанных на их применении тест-систем диагностики ИКБ на территории России. Кроме того, как отмечено выше, OspA пригоден для диагностики ИКБ главным образом на стадии диссеминированной инфекции, и только к FlaB имеются антитела, как на ранней стадии заболевания, так и на стадии диссеминированной инфекции. Таким образом, совместное использование этих антигенов для серодиагностики ИКБ не обеспечивает надежную постановку диагноза на ранней стадии инфекции.
Известен рекомбинантный белок OspC, кодируемый геном, относящимся к инвазивным группам A, B, I и K Borrelia burgdorferi s.1. или группам A, B Borrelia afzelii (27). Недостатком применения данного полипептида для диагностики ИКБ в России является то, что он получен на основе гена боррелий, не встречающихся в популяциях иксодовых клещей на территории России. Кроме того, OspC не пригоден для диагностики ИКБ на стадии диссеминированной инфекции.
Известен рекомбинантный белок OspC, и рекомбинантный полипетид, соответствующий фрагменту белка FlaB западносибирского изолята B.garinii подгруппы NT29, совместно используемые в качестве антигенов для диагностики ИКБ. Недостатком фрагмента белка FlaB является то, что он содержит восемь дополнительных аминокислотных остатков зрелого α-токсина Staphylococcus aureus, что дает при диагностике ИКБ ложноположительные результаты, т.е. не может обеспечить высокую специфичность диагностики ИКБ (28). Кроме того, сочетание двух указанных рекомбинантных полипептидов не обеспечивает надежной диагностики ИКБ на стадии диссеминированной инфекции, т.к. антитела к OspC нарабатываются у больных только на ранней стадии, и только антитела к FlaB могут присутствовать как на ранней стадии, так и на стадии диссеминированной инфекции.
Известен рекомбинантный белок FlaA вида B.burgdorferi s.s. (штамм В31), распространенного на североамериканском континенте и в странах Западной Европы (29). Недостатком применения этого белка для диагностики ИКБ в России является то, что он отличается по антигенным свойствам от белков FlaA, свойственных боррелиям, циркулирующим в популяциях иксодовых клещей на территории Сибири и Дальнего Востока. Кроме того, FlaA не пригоден для диагностики ИКБ на стадии диссеминированной инфекции.
Наиболее близкими к заявленным являются химерные рекомбинантные полипептиды, несущие эпитопы различного размера из белков OspA, свойственных видам североамериканских и европейских изолятов боррелий и одного или двух других рекомбинантных полипептидов указанных боррелий в следующих комбинациях: OspA и OspA, OspA-OspB, OspA-OspB-OspC, OspA-p83, а также химерные рекомбинантные полипептиды OspB-FlaB и OspB-FlaB-OspC (30). Применение названных химерных антигенов для диагностики ИКБ в Сибири не приемлемо ввиду следующих причин:
- в их состав входят преимущественно эпитопы высоковариабельных антигенов североамериканских и европейских изолятов боррелий, таких как: OspA, OspB и OspC, существенно отличающихся от аналогичных аминокислотных последовательностей сибирских изолятов боррелий, что обусловливает их различную аффинность к антителам сывороток больных и снижает чувствительность анализа;
- количество консервативных антигенов, по которым различия между видами боррелий различных регионов менее выражены, ограничивается белками р83 и FlaB, входящими в состав химерных антигенов OspA-p83, OspB-FlaB-OspC и OspB-FlaB, однако сочетание в составе одного рекомбинантного химерного полипептида маркеров ранней стадии и стадии диссеминированной инфекции затрудняет дифференциальную диагностику этих стадий заболевания.
Задачей настоящего изобретения является расширение арсенала рекомбинантных полипептидов, пригодных для диагностики ИКБ, повышение специфичности и чувствительности диагностики ИКБ в регионах распространения западносибирских изолятов Borrelia burgdorferi s.1., в том числе дифференциальной диагностики ранней стадии и стадии диссеминированной инфекции.
Решение поставленной задачи достигается тем, что предложены следующие новые рекомбинантные химерные полипептиды, полученные на основе экспрессии генов, синтезированных методом ПЦР на ДНК, выделенных из западносибирских изолятов B.garinii и B.afzelii:
1. Рекомбинантный химерный полипептид - rmBmpA-frp83 (SEQ ID NO 4) размером 591 а.о. с молекулярной массой 65,89 кДа, содержащий с N-конца молекулы аминокислотную последовательность зрелого белка BmpA западносибирского изолята Borrelia garinii (rmBmpA) размером 324 а.о. с молекулярной массой 35,3 кДа, а с С-конца - аминокислотную последовательность C-концевого фрагмента белка р83 (frp83) западносибирского изолята Borrelia garinii размером 248 а.о. с молекулярной массой 28,17 кДа.
2. Рекомбинантный химерный полипептид - mOspA-frp83 (SEQ ID NO 6) размером 521 а.о. с молекулярной массой 57,99 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка OspA (rmOspA) западносибирского изолята Borrelia garinii размером 257 а.о. с молекулярной массой 27,79 кДа, а с С-конца - аминокислотную последовательность C-концевого фрагмента белка р83 (frp83) западносибирского изолята Borrelia garinii размером 248 а.о. с молекулярной массой 28,17 кДа.
3. Рекомбинантный химерный полипептид - rmDbpB-rmOspA (SEQ ID NO 8) размером 432 а.о. с молекулярной массой 47,13 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка DbpB (rmDbpB) западносибирского изолята Borrelia garinii размером 156 а.о. с молекулярной массой 17,02 кДа, а с С-конца - рекомбинантного зрелого белка OspA (rmOspA) западносибирского изолята Borrelia garinii размером 257 а.о. с молекулярной массой 27,79 кДа.
4. Рекомбинантный химерный полипептид - rmFlaA-frFlaB (SEQ ID NO 10) размером 587 а.о. с молекулярной массой 64,76 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка FlaA (rmFlaA) западносибирского изолята Borrelia garinii размером 318 а.о. с молекулярной массой 35,82 кДа, а с С-конца - аминокислотную последовательность фрагмента флагеллярного белка FlaB (frFlaB) специфичного для спирохет B.burgdorferi s.1. размером 250 а.о. с молекулярной массой 26,62 кДа.
5. Рекомбинантный химерный полипептид - rmOspCBg-rmOspCBa (SEQ ID NO 12) размером 390 а.о. с молекулярной массой 40,83 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка OspC западносибирского изолята Borrelia garinii (rmOspCBg) размером 188 а.о. с молекулярной массой 19,70 кДа, а с С-конца - аминокислотную последовательность рекомбинантного зрелого белка OspC западносибирского изолята Borrelia afzelii (rmOspCBa) размером 186 а.о. с молекулярной массой 19,19 кДа.
В кратких обозначениях белков нижние индексы означают: «m» - зрелый (mature), «fr» - фрагмент «r» - рекомбинантный, Bg и Ba - принадлежность белка к боррелиям видов Borrelia garinii и Borrelia afzelii соответственно.
Известны способы серодиагностики иксодового клещевого боррелиоза методами иммуноферментного анализа с применением коммерческих тест-систем.
Известен способ серодиагностики ИКБ методом иммуноферментного анализа, включающий применение нативного антигена в виде лизата спирохет, полученного из штамма П1 В.afzelii, выделенного на территории Московской области (производитель:
Государственный научный центр прикладной микробиологии, г.Оболенск, Россия) (31). Недостатком известного способа является то, что он не обеспечивает удовлетворительной специфичности по отношению к ИКБ в регионах распространения западносибирских изолятов боррелий, т.к. они существенно отличаются по структуре антигенов от изолятов боррелий, распространенных в Московской области. Кроме того, способ не позволяет дифференцировать раннюю стадию и стадию диссеминированной инфекции. Использование лизатов спирохет обеспечивает высокую чувствительность способа, но он обладает очень низкой специфичностью ввиду присутствия в лизатах антигенов, имеющих перекрестную антигенную реактивность с антигенами других микроорганизмов, включая симбионтов человека. Кроме того, получение лизатов связано с дорогостоящим культивированием спирохет, причем в процессе нескольких пассажей спирохеты могут терять антигены, кодируемые плазмидными ДНК этих микроорганизмов.
Известен способ серодиагностики ИКБ методом иммуноферментного анализа, включающий применение природного антигена, выделенного из штамма РКо В.afzelii (производитель тест-системы: «Dade Behring Marburg GmbH», Германия) (32). Известный способ обладает теми же недостатками.
Известен способ серодиагностики ИКБ методом иммуноферментного анализа, включающий применение в качестве антигенов смеси следующих рекомбинантных полипептидов трех видов боррелий: р21 и OspC из В.burgdorferi sensu stricto и В.garinii, p41i - внутренняя часть флагеллина, из В.garinii, p18 и р100 из В.afzelii (производитель: «Biomedica», Австрия) (33). Недостатком заявленного способа является то, что известные антигены получены на основе боррелий, распространенных в Западной Европе, что ограничивает его применение на территориях, где распространены западносибирские изоляты боррелий. Кроме того, известный способ не позволяет дифференцировать раннюю стадию и стадию диссеминированной инфекции. Другим недостатком способа является то, что использование трех-четырех антигенов недостаточно для достижения высокой чувствительности и специфичности способа.
Известен способ серодиагностики ИКБ методом иммуноферментного анализа, включающий применение в качестве антигенов рекомбинантного полипептида VlsE и лизата клеток В.afzelii, В.arinii для диагностики стадии диссеминированной инфекции ИКБ (выявления антител к боррелиям класса IgG), рекомбинантного полипептида FlaB и природного очищенного OspC для диагностики ранней стадии ИКБ (для выявления антител к боррелиям класса IgM) (производитель тест-ситемы: компания «NovaTec GmbH», Германия) (34). Недостатком заявленного способа является то, что известные антигены получены на основе боррелий, распространенных в Западной Европе, что ограничивает его применение на территориях, где распространены западносибирские изоляты боррелий. Использование лизатов клеток обеспечивает высокую чувствительность способа, но он обладает очень низкой специфичностью ввиду присутствия в лизатах антигенов, имеющих перекрестную антигенную реактивность с антигенами других микроорганизмов.
Известен способ серодиагностики ИКБ инфекции методом иммуноферментного анализа, включающий применение в качестве антигенов смеси рекомбинантных полипетидов трех видов боррелий: VlsE из В.burgdorferi sensu stricto, FlaB (p41) и BmpA из В.garinii, OspC из В.afzelii (производитель: ЗАО «Вектор-Бест», г.Новосибирск, Россия). Известный способ является недостаточно чувствительным и специфичным, в том числе на территории Сибири. Это обусловлено небольшим количеством используемых антигенов, а также использованием в качестве антигенов рекомбинантных белков боррелий, которые не встречаются на территории Сибири (B.burgdorferi sensu stricto) либо существенно отличаются по аминокислотной последовательности от белков западносибирских изолятов боррелий. Кроме того, в способе используют рекомбинантный белок OspC только вида В.afzelii, который в Сибири менее распространен, чем В.garinii.
Наиболее близким к заявляемому является способ серодиагностики ИКБ методом иммуноферентного анализа, включающий применение в качестве антигенов смеси рекомбинантных антигенов DbpA из В.afzelii и В.garinii, FlaB (p41) из В.garinii, OspC из В.afzelii и В.garinii, p35 из В.afzelii (производитель тест-ситемы: ООО «Омникс», г.Санкт-Петербург, Россия) (31). Поскольку он основан на антигенах, свойственных европейским изолятам боррелий, то обладает сниженной чувствительностью и специфичностью при диагностике ИКБ, вызываемого боррелиями, циркулирующими на территории Сибири. Кроме того, он недостаточно надежно выявляет ИКБ на ранней стадии, т.к. для этого используется только один маркер - OspC. Наличие же в сыворотке антител к FlaB встречается у пациентов как на ранней стадии, так и на стадии диссеминированной инфекции.
Задачей, на решение которой направлено изобретение, является повышение специфичности и чувствительности способа диагностики ИКБ, дифференциальной диагностики ранней стадии и стадии диссеминированной инфекции ИКБ на территории распространения западносибирских изолятов Borrelia burgdorferi s.1.
Решение поставленной задачи достигается тем, что в качестве антигенов для иммуноферментного анализа используют рекомбинантные химерные полипептиды rmBmpA-frp83, rmOspA-frp83, rmDbpB-rmOspA, rmFlaA-frFlaB, rmOspCBg-rmOspCBa, заявленные в настоящем изобретении, полученные на основе ДНК западносибирских изолятов Borrelia garinii, Borrelia afzelii; для диагностики ИКБ на ранней стадии используют рекомбинантные химерные полипептиды rmFlaA-frFlaB и rmOspCBg-rmOspCBa;
для диагностики ИКБ на стадии диссеминированной инфекции - рекомбинантные химерные полипептиды rmBmpA-frp83, rmOspA-frp83 и rmDbpB-rmOspA.
Описание сущности изобретения
Согласно заявленному изобретению предложены следующие новые рекомбинантные химерные полипептиды, пригодные для диагностики иксодового клещевого боррелиоза на территории распространения западносибирских изолятов боррелий:
1. Рекомбинантный химерный полипептид - rmBmpA-frp83 (SEQ ID NO 4) размером 591 а.о. с молекулярной массой 65,89 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка BmpA (rmBmpA) западносибирского изолята Borrelia garinii размером 324 а.о. с молекулярной массой 35,3 кДа, а с С-конца - аминокислотную последовательность С-концевого фрагмента белка р83 (frp83) западносибирского изолята Borrelia garinii размером 248 а.о. с молекулярной массой 28,17 кДа.
2. Рекомбинантный химерный полипептид - rmOspA-frp83 (SEQ ID NO 6) размером 521 а.о. с молекулярной массой 57,99 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка OspA (rmOspA) западносибирского изолята Borrelia garmii размером 257 а.о. с молекулярной массой 27,79 кДа, а с С-конца - аминокислотную последовательность С-концевого фрагмента белка р83 (frp83) западносибирского изолята Borrelia garinii размером 248 а.о. с молекулярной массой 28,17 кДа.
3. Рекомбинантный химерный полипептид - rmDbpB-rmOspA (SEQ ID NO 8) размером 432 а.о. с молекулярной массой 47,13 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка DbpB (rmDbpB) западносибирского изолята Borrelia garmii размером 156 а.о. с молекулярной массой 17,02 кДа, а с С-конца - аминокислотную последовательность рекомбинантного зрелого белка OspA (rmOspA) западносибирского изолята Borrelia garmii размером 257 а.о. с молекулярной массой 27,79 кДа.
4. Рекомбинантный химерный полипептид - rmFlaA-frFlaB (SEQ ID NO 10) размером 587 а.о. с молекулярной массой 64,76 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка FlaA (rmFlaA) западносибирского изолята Borrelia garmii размером 318 а.о. с молекулярной массой 35,82 кДа, а с С-конца - аминокислотную последовательность фрагмента флагеллярного белка FlaB (frFlaB) специфичного для спирохет B.burgdorferi s.1. размером 250 а.о. с молекулярной массой 26,62 кДа.
5. Рекомбинантный химерный полипептид - rmOspCBg-rmOspCBa (SEQ ID NO 12) размером 390 а.о. с молекулярной массой 40,83 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка OspC западносибирского изолята Borrelia garmii (rmOspCBg) размером 188 а.о. с молекулярной массой 19,70 кДа, а с С-конца - аминокислотную последовательность рекомбинантного зрелого белка OspC западносибирского изолята Borrelia afzelii (rmOspCBa) размером 186 а.о. с молекулярной массой 19,19 кДа.
Предложенные рекомбинантные химерные полипептиды позволяют существенно расширить арсенал средств, которые могут быть использованы для диагностики ИКБ. Они пригодны в качестве антигенов для диагностики ИКБ на территориях распространения западносибирских изолятов боррелий, т.к. они созданы на основе ДНК боррелий этих регионов. Одновременное использование при диагностике ИКБ заявленных рекомбинантных химерных полипепидов, включающих в составе химер различные комбинации из 8 белков западносибирских изолятов боррелий, позволяет обеспечить высокую специфичность и чувствительность диагностики. Повышение специфичности и чувствительности дифференциальной диагностики ранней стадии и стадии диссеминированной инфекции ИКБ обеспечивается тем, что в состав рекомбинантных химерных полипептидов согласно пп.1-3 входят 4 маркера стадии диссеминированной инфекции, а рекомбинантны химерные полипептиды по пп.4, 5 включают 4 маркера ранней стадии ИКБ.
Заявленные рекомбинантные химерные полипептиды получены следующим образом.
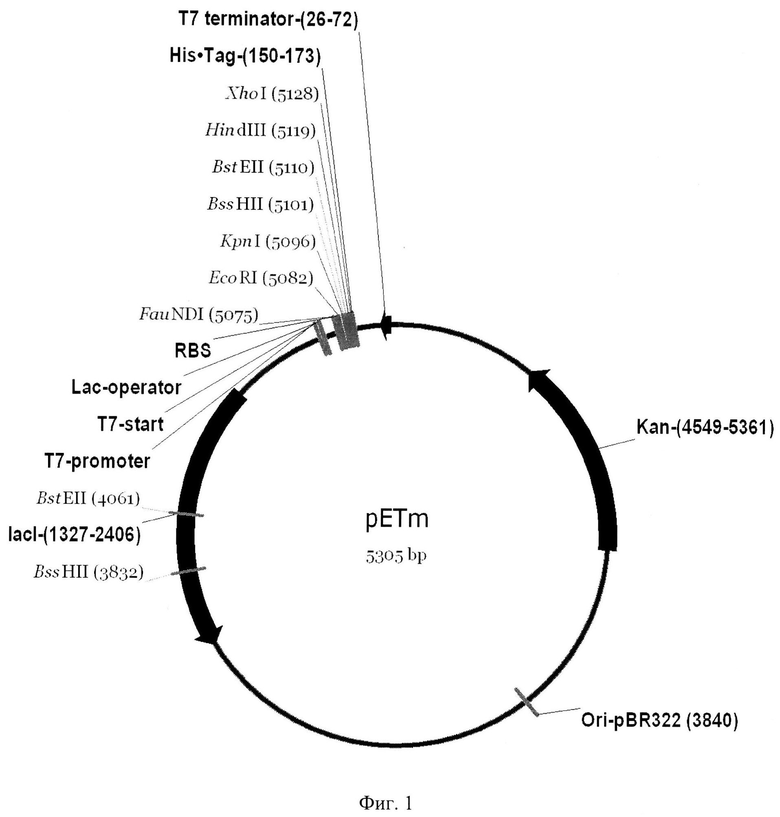
Экспрессирующий вектор pET36b(+) («Novagen», США) был модифицирован путем встраивания под регуляторную область промотора бактериофага Т7 полилинкера, представляющего из себя фрагмент ДНК, содержащий сайты узнавания эндонуклеазами рестрикции EcoRI, KpnI, BssHII, BstEII, HindIII и XhoI. Такая структура полилинкера была получена для удобства клонирования выбранных генов боррелий. За полилинкером следует нуклеотидная последовательность, кодирующая 8-ми гистидиновый аминокислотный фрагмент в C-концевой области рекомбинантных белков. Это позволяет очищать рекомбинантные полипептиды с помощью аффинной хроматографии на металло-хелатных сорбентах. Модифицированный вектор был назван pETm, структура встроенного полилинкера приведена в перечне последовательностей под номером SEQ ID NO 1. Целевые рекомбинантные полипептиды, получаемые с помощью модифицированного экспрессирующего вектора pETm, не содержат протяженных чужеродных аминокислотных последовательностей, вследствие чего не дают неспецифических сигналов в иммуноферментных анализах. Это, в свою очередь, обеспечивает повышение специфичности диагностики.
Рекомбинантная плазмидная ДНК pETm размером 5305 п.н. содержит:
- фрагмент ДНК размером 48 п.н. - полилинкер, содержащий сайты рестрикции EcoRI, KpnI, BssHII, BstEII, HindIII;
- уникальные сайты рестрикции XbaI, FauNDI, EcoRI, BssHII, BstEII, KpnI, HindIII, XhoI, DraIII, XmaI, SapI;
- XhoI-FauNDI фрагмент плазмидной ДНК pET36b(+) размером 5257 п.н., включающий:
1) фрагмент ДНК размером 47 п.н., содержащий последовательность терминатора транскрипции гена 10 бактериофага Т7;
2) фрагмент ДНК размером 468 п.н., содержащий ориджин репликации однонитчатого бактериофага f1;
3) фрагмент ДНК, размером 813 п.н., содержащий ген неомициптрансферазы, обусловливающей устойчивость клеток E.coli к капамицину;
4) фрагмент ДНК размером 468 п.н., содержащий ориджин репликации плазмиды рМВ1;
5) фрагмент ДНК размером 1080 п.н., кодирующий область гена-регулятора lacI оперона lac E.coli;
6) фрагмент ДНК размером 468 п.н., содержащий промотор гена 10 бактериофага Т7.
Структура вектора pETm приведена в перечне последовательностей под номером SEQ ID NO 2.
Вектор pETm был использован для клонирования и экспрессии индивидуальных генов (или фрагментов генов) белков rmOspCBg, rmDbpB, rmOspA, rmFlaA, frFlaB, rmBmpA и frp83, свойственных западносибирским изолятам Borrelia garinii, белка rmOspCBa западносибирского изолята Borrelia afzelii, а также химерных генов данных белков в заявленных сочетаниях.
В настоящем изобретении были получены и клонированы в составе вектора pETm рекомбинантные химерные ДНК, кодирующие рекомбинантные химерные полипептиды следующего состава: rmBmpA-frp83, rmOspA-frp83, rmDbpB-rmOspA, rmFlaA-frFlaB и rmOspCBg-rmOspCBa. В кратких обозначениях белков нижние индексы означают: «m» - зрелый (mature), «fr» - фрагмент, Bg и Ba - принадлежность белка к боррелиям видов Borrelia garinii и Borrelia afzelii соответственно.
Аналогичным образом были названы рекомбинантные химерные ДНК, кодирующие вышеперечисленные полипептиды. Сконструированные плазмиды, содержащие рекомбинантные химерные ДНК, были названы соответственно: pETm-rmbmpA-frp83, pETm-rmospA-frp83, pETm-rmdbpB-rmospA, pETm-rmflaA-frflaB и pETm-rmospCBg-rmospCBa.
Рекомбинантная химерная ДНК rmbmpA-frp83 в составе вектора pETm обеспечивает синтез рекомбинантного химерного полипептида, содержащего эпитопы иммунодоминантных белков rmBmpA и frp83, специфичных для сибирского изолята В.garinii подгруппы 20047T. Рекомбинантная химерная ДНК rmbmpA-frp83 имеет размер 1773 п.н. и включает следующие элементы (все позиции нуклеотидных остатков указаны относительно плазмиды pETm-rmbmpA-frp83):
- фрагмент гена bmpA B.garinii размером 972 п.н., кодирующий рекомбинантный зрелый белок BmpA (rmBmpA) западносибирского изолята Borrelia garimi, позиция фрагмента 5097-6068 п.н.;
- сайты эндонуклеазы рестрикции XhoI размером 6 п.н., позиции сайтов 6069-6074 п.н. и 6819-6824;
- фрагмент гена р83 B.garinii размером 744 п.н., кодирующий C-концевой фрагмент белка р83 западносибирского изолята Borrelia garimi (frp83), позиции фрагмента 6075-6818 п.п.;
- уникальные сайты рестрикции: KpnI (GGTACC), позиция сайта 5091-5096; BglII (AGATCT) позиция сайта 5454-5459 п.н.; HindIII (AAGCTT) позиции 6522-6527 п.н.
Структура рекомбинантной химерной ДНК mbmpA-frp83 представлена в перечне последовательностей SEQ ID NO 3, кодируемая ею аминокислотная последовательность рекомбинантного химерного полипептида mBmpA-frp83 отдельно представлена под номером SEQ ID NO 4.
Рекомбинантная химерная ДНК rmospA-frpSS в составе вектора pETm обеспечивает синтез рекомбинантного химерного полипептида, содержащего эпитопы иммунодоминантных белков rmOspA и frp83, специфичных для сибирского изолята В.garinii подгруппы 20047T. Рекомбинантная химерная ДНК rmospA-frp83 имеет размер 1563 п.н. и включает следующие элементы (все позиции нуклеотидных остатков указаны относительно плазмиды pETm-rmospA-frp83):
- фрагмент гена ospA B.garinii размером 771 п.н., кодирующий рекомбинантный зрелый белок OspA западносибирского изолята Borrelia garimi (rmOspA), позиция фрагмента 5088-5858 п.н.;
- сайты эндонуклеазы рестрикции XhoI размером 6 п.н., позиции сайтов 5859-5864 п.н. и 6609-6614;
- фрагмент гена р83 B.garinii размером 744 п.н., кодирующий C-концевой фрагмент белка р83 западносибирского изолята Borrelia garinii (frp83), позиции фрагмента 5865-6608 п.п.;
- уникальные сайты рестрикции: EcoRI (GAATTC), позиция сайта 5081-5086; BamHI (GGATTC), позиция сайта 5496-5501 п.н.; BglII (AGATCT), позиция сайта 5828-5833 п.н.; HindIII (AAGCTT), позиция сайта 6312-6317 п.н.
Структура рекомбинантной химерной ДНК rmospA-frp83 представлена в перечне последовательностей SEQ ID NO 5, кодируемая ею аминокислотная последовательность рекомбинантного химерного полипептида rmOspA-frp83 отдельно представлена под номером SEQ ID NO 6.
Рекомбинантная химерная ДНК rmdbpB-rmospA в составе вектора pETm обеспечивает синтез рекомбинантного химерного полипептида, содержащего эпитопы иммунодоминантных белков rmDbpB и rmOspA, специфичных для сибирского изолята B.garinii подгруппы 20047T. Рекомбинантная химерная ДНК rmbpB-rmOspA имеет размер 1296 п.н. и включает следующие элементы (все позиции нуклеотидных остатков указаны относительно плазмиды pETm-rmdbpB-rmospA):
- фрагмент гена rmdbpB B.garinii размером 468 п.н., кодирующий зрелый белок rmDbpB, позиция фрагмента 5097-5564 п.н.;
- сайты эндонуклеазы рестрикции XhoI размером 6 п.н., позиции сайтов 5565-5570 п.н. и 6342-6347;
- фрагмент гена rmospA B.garinii размером 771 п.н., кодирующий зрелый белок rmOspA, позиции фрагмента 5571-6341 п.н.;
- уникальные сайты рестрикции: EcoRI (GAATTC), позиция сайта 5081-5086; KpnI (GGTACC), позиция сайта 5091-5096; BamHI (GGATTC), позиция сайта 5979-5984 п.н.; BglII (AGATCT) позиция сайта 6311-6316 п.н.
Структура рекомбинантной химерной ДНК rmdbpB-rmospA представлена в перечне последовательностей SEQ ID NO 7, кодируемая ею аминокислотная последовательность рекомбинантного химерного полипептида rmDbpB-rmOspA отдельно представлена под номером SEQ ID NO 8.
Рекомбинантная химерная ДНК rmflaA-frflaB в составе вектора pETm обеспечивает синтез рекомбинантного полипептида, содержащего эпитопы иммунодоминантных белков frFlaB и rmFlaA, специфичные для сибирского изолята B.garinii подгруппы 20047T. Рекомбинантная химерная ДНК rmflaA-frflaB имеет размер 1761 п.н. и включает следующие элементы (все позиции нуклеотидных остатков указаны относительно плазмиды рЕТ- rmflaA-frflaB):
- фрагмент гена flaA B.garinii размером 954 п.н., кодирующий зрелый белок rmFlaA позиция фрагмента 5097-6050 п.п.;
- сайты эндонуклеазы рестрикции XhoI размером 6 п.н., позиции сайтов 6051-6056 п.н. и 6807-6812 п.н.;
- фрагмент кодирующей области гена flaB B.garinii размером 756 п.н., позиции фрагмента 6057-6812 п.н.;
- уникальные сайты рестрикции - EcoRI (GAATTC) позиция сайта 5081-5086 п.н. и KpnI (GGTACC), позиции сайта 5091-5096 п.н.
Структура рекомбинантной химерной ДНК rmflaA-frflaB представлена в перечне последовательностей SEQ ID NO 9, кодируемая ею аминокислотная последовательность рекомбинантного химерного полипептида rmFlsA-frFlaB отдельно представлена под номером SEQ ID NO 10.
Рекомбинантная химерная ДНК rmospCBg-rmospCBa в составе вектора pETm обеспечивает синтез рекомбинантного химерного полипептида, содержащего эпитопы иммунодоминантных белков rmOspCBg и rmOspCBa, специфичных для сибирского изолята B.garinii подгруппы 20047T и B.afzelii соответственно. Рекомбинантная химерная ДНК rmospCBg-rmospCBa имеет размер 1170 п.н. и включает следующие элементы (все позиции нуклеотидных остатков указаны относительно плазмиды pETm- rmospCBg-rmospCBa):
- фрагмент гена rmospCBg B.garinii размером 564 п.н., кодирующий зрелый белок rmOspCBg, позиция фрагмента 5088-5651 п.н.;
- фрагмент гена rmospCBa B.afzelii размером 558 п.н., кодирующий зрелый белок rmOspCBa, позиции фрагмента 5658-6215 п.н.;
- уникальные сайты рестрикции - EcoRI (GAATTC), позиция сайта 5081-5086; KpnI (GGTACC), позиция сайта 5652-5657 п.н.; XhoI (CTCGAG), позиция сайта 6216-6221 п.н.
Структура рекомбинантной химерной ДНК rmospCBg-rmospCBa представлена в перечне последовательностей SEQ ID NO 11, кодируемая ею аминокислотная последовательность рекомбинантного химерного полипептида rmOspCBg-rmOspCBa отдельно представлена под номером SEQ ID NO 12.
Заявленный способ серодиагностики иксодового клещевого боррелиоза методом иммуноферментного анализа включает использование в качестве антигенов комбинации заявленных согласно изобретению рекомбинантных химерных полипептидов rmBmpA-frp83, rmOspA-frp83, rmDbpB-rmOspA, rmFlaA-frFlaB, rmOspCBg-rmOspCBa, полученных на основе экспрессии генов, синтезированных методом ПЦР на ДНК западносибирского изолята Borrelia garinii или в случае белка rmOspCBa на ДНК изолята Borrelia afzelii. Для серодиагностики ранней стадии ИКБ используют рекомбинантные химерные полипептиды rmFlsA-frFlaB, rmOspCBg-rmOspCBa; для серодиагностики стадии диссеминированной инфекции ИКБ - рекомбинантные химерные полипептиды rmBmpA-frp83, rmOspA-frp83, rmDbpB-rmOspA. Сравнение аминокислотных последовательностей антигенов западносибирских изолятов боррелий, используемых в заявленном способе, с первичными структурами аналогичных антигенов, свойственных трем патогенным видам боррелий, распространенных в Западной Европе, Северной Америке и Азии, выявило в разной степени выраженные различия между ними: для антигена rmOspA различия в аминокислотной последовательности составили от 26,9% до 64%, для rmDbpB - от 17% до 46,5%, для rmFlaA - от 1,5% до 4%, для frFlaB - от 2% до 5%, для rmBmpA - от 5,8% до 13,8%, для frp83 - от 3,3% до 22,6%, для rmOspCBa - от 56% до 72,3%, для rmOspCBg - от 70% до 84,3%.
Способ осуществляют на основе стандартного метода твердофазного иммуноферментного анализа (Пример 5 конкретного выполнения). Заявленный способ серодиагностики иксодового клещевого боррелиоза демонстрирует высокую диагностическую чувствительность и специфичность при выявлении ИКБ в Западной Сибири, которые обусловлены следующими факторами:
а) прежде всего за счет использования химерных белков, включающих антигены, свойственные боррелиям, распространенным непосредственно на территории Западной Сибири, поскольку известно, что отличия в аминокислотных последовательностях аналогичных антигенов, свойственных изолятам боррелий разных географических зон, обуславливают необходимость использования в диагностических тестах антигенов, свойственных боррелиям, распространенным в соответствующих географических зонах;
б) используемые в анализе заявляемые химерные полипептиды состоят из полноразмерных природных аминокислотных последовательностей зрелых иммунодоминантных антигенов, содержащих полный набор соответствующих им В-эпитопов (исключение составляет иммунодоминантный С-концевой фрагмент белка р83);
в) использование в анализе одновременно 8 антигенов не снижает чувствительность анализа, поскольку в сенсибилизации лунок планшет участвуют, по существу, пять химерных антигенов, а не 8 входящих в их состав индивидуальных антигенов;
г) химерный полипептид mOspCBg-mOspCBa содержит аминокислотные последовательности двух иммунодоминантных высоко вариабельных антигенов, свойственных спирохетам B.garinii и B.afzelii, циркулирующим на территории Западной Сибири.
Изобретение иллюстрируется следующими графическими материалами.
Перечень фигур и иных материалов
Фиг.1. Физико-генетическая карта рекомбинантной плазмидной ДНК pETm. lacI - кодирующая область гена-регулятора lacI оперона lac E.coli; T7-promotor - промотор гена 10 бактериофага Т7; TV-start - точка инициации транскрипции в промоторсодержащей области бактериофага Т7; lac-operator - последовательность, связывающая репрессор транскрипции гена lacZ; RBS - рибосомосвязывающий сайт (последовательность Шайна-Дальгарно); His-Tag - последовательность, кодирующая восемь гистидиновых а.о.; Т7-terminator - последовательность терминатора транскрипции гена 10 бактериофага Т7; Ori-pBR322 - ориджин репликации плазмиды pBR322; Kan - ген неомицин-фосфотрансферазы II, обусловливающей устойчивость клеток E.coli к канамицину.
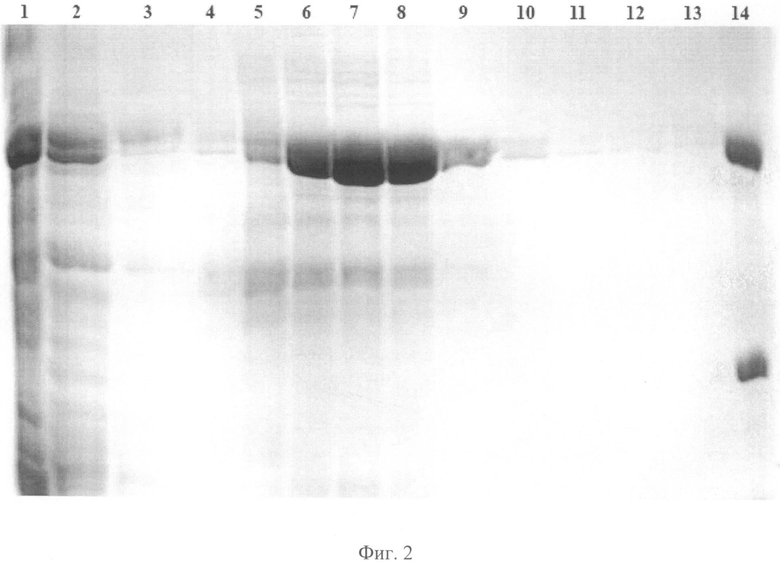
Фиг.2. Электрофореграмма фракций, полученных в процессе аффинной хроматографии на Ni-хелатном сорбенте в денатурирующих условиях лизата клеток E.coli, продуцирующих химерный полипептид rmBmpA-frp83. Дорожки: (1) - лизат клеток продуцента после индукции; (2) - растворимая фракция лизата клеточной биомассы; (3) - фракция белков, несвязавшихся со смолой; (4-5) - элюаты, содержащие имидазол с конечной концентрацией 20 мМ и 50 мМ, соответственно; (6-8) - элюаты, содержащие имидазол с конечной концентрацией 250 мМ; (9-13) - элюаты, содержащие имидазол с конечной концентрацией 500 мМ, (14) - маркерные белки (66 и 28 кДа).
Таблица 1. Нуклеотидные последовательности праймеров, используемых для амплификации генов B.burgdorferi s.1. (сайт рестрикции подчеркнут).
Таблица 2. Условия амплификации фрагментов генов B.burgdorferi s.1.
Таблица 3. Выявление методом ИФА в сыворотках больных ИКБ и здоровых доноров антител классов IgM и IgG к рекомбинантным химерным антигенам боррелий.
Примеры конкретного выполнения
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Конструирование плазмидного вектора pETm.
Для конструирования экспрессирующего вектора pETm используют вектор pET36b(+) и синтетический дуплекс ДНК - полилинкер. Синтетический дуплекс ДНК в своей структуре содержит набор уникальных сайтов рестрикции: EcoRI, KpnI, BssHII, BstEII, HindIII. За синтетическим дуплексом следует участок, кодирующий восьмигистидиновую аминокислотную последовательность и стоп-кодон терминации трансляции мРНК. При этом продукты экспрессии встраиваемых в сконструированный вектор генов содержат дополнительно 8-ми гистидиновую аминокислотную последовательность в C-концевой области полипептидной цепи, что позволяет осуществлять аффинную очистку полипептидов на металлохелатных сорбентах. Синтетический дуплекс ДНК получают гибридизацией пары комплементарных друг другу олигонуклеотидов размером 53 и 55 п.п., структуры олигонуклеотидов приведены в перечне последовательностей под номерами SEQ ID NO 13 и 14 соответственно. Олигонуклеотиды синтезированы таким образом, что при гибридизации образуется двунитевой фрагмент ДНК, содержащий на 5' и 3' концах «липкие концы» для эндонуклеаз рестрикции FauNDI nXhoI соответственно (сайты подчеркнуты):
FauNDI XhoI TATGCAGAATTCTATGGGTACCGTTGCGCGCAGCGGTCACCCGAAGCTTCTGCACGTCTTAAGATACCCATGGCAACGCGCGTCGCCAGTGGGCTTCGAAGACGAGCT
Гибридизацию олигонуклеотидов проводят следующим образом. В эквимолярном соотношении смешивают олигонуклеотиды в одной пробирке, нагревают пробирку на водяной бане и постепенно остужают раствор при комнатной температуре. Полученный раствор, содержащий синтетический дуплекс, используют для лигировапия с XhoI-FauNDI фрагментом плазмидной ДНК pET36b(+). Все процедуры, связанные со встройкой и клонированием дуплекса, а в последующем и ампликонов генов проводят с использованием стандартных генно-инженерных методик, описанных в руководстве (36).
Для получения фрагмента, плазмидную ДНК pET36b(+) гидролизуют эндонуклеазами рестрикции FauNDI и XhoI. Линейную форму плазмидной ДНК размером 5252 п.н. - фрагмент XhoI-FauNDI, отделяют от продуктов гидролиза в 0,8% агарозном геле. Необходимый фрагмент плазмидной ДНК вырезают из агарозного геля и извлекают ДНК с помощью набора фирмы «Биосилика» (Новосибирск, Россия) согласно прилагаемому протоколу. Очищенный фрагмент ДНК XhoI-FanNDI плазмиды pET36b(+) лигируют с синтетическим дуплеком ДНК. Лигирование проводят в молярном соотношении 20:1 (вставка / фрагмент XhoI-FauNDI) с помощью Т4 ДНК-лигазы фирмы «Сибэнзим» (Россия) согласно инструкции производителя фермента. Лигазной смесью методом электропорации трансформировали клетки E.coli штамма BL21 (DE3). Трансформацию клеток E.coli лигазной смесью проводят в соответствии с руководством фирмы-производителя электропоратора - «PeqLab, Biotechnologie GmbH» (Германия). Трансформированные клетки рассевают на агаризованую среду Луриа-Бертани (LB), содержащую канамицин 30 мкг/мл и выращивают в течение ночи при 37°С. Полученные клоны клеток анализируют на наличие вставки синтетического дуплекса с помощью полимеразной цепной реакции (ПЦР) с использованием внешних праймеров PR268 и PR269, т.е. праймеров, лежащих «левее» и «правее» области встроенного синтетического фрагмента ДНК. Структуры праймеров PR268 и PR269 указаны в перечне последовательностей под номерами SEQ ID NO 15 и 16 соответственно. Для ПЦР использовали Taq ДНК- полимеразу «Сибэнзим» (Россия).
Условия амплификации:
1 цикл (96°С - 2 мин, 64°С - 0,5 мин, 72°С - 0,5 мин);
25 циклов (96°С - 0,5 мин, 64°С - 0,5 мин, 72°С - 0,5 мин);
1 цикл (96°С - 0,5 мин, 64°С - 5 мин, 72°С - 10 мин).
Нужные клоны идентифицируют по размеру ампликона - продукта ПЦР, который должен составлять 296 п.н. Продукты ПЦР анализируют в 2% агарозном геле. Из нескольких положительных клонов выделяют плазмиды щелочным методом и используют для рестрикционного анализа ферментами, сайты для которых присутствуют только во встроенном полилинкере. Плазмидную ДНК одного из клонов, удовлетворяющую всем условиям, секвенируют для подтверждения нуклеотидной последовательности клонированного синтетического фрагмента ДНК. Новая конструкция плазмидной ДНК названа pETm, ее используют в дальнейшем для клонирования кодирующих областей генов спирохет комплекса B.burgdorferi sensu lato. Физико-генетическая карта плазмидного вектора pETm представлена на Фиг.1. Нуклеотидная последовательность вектора - в перечне последовательностей SEQ ID NO 2.
Пример 2. Конструирование рекомбинантных химерных ДНК, содержащих в составе плазмидного вектора pETm нуклеотидные последовательности, кодирующие рекомбинантные химерные иммунодоминантные белки спирохет комплекса Borrelia burgdorferi sensu lato.
Кодирующие области генов заявленных рекомбинантных химерных полипептидов получают в препаративных количествах с помощью амплификации соответствующих фрагментов геномной ДНК спирохет B.garinii и B.afzelii методом ПЦР. В качестве матрицы используют геномные ДНК, полученные ранее из культур B.garinii и B.afzelii (37).
Праймеры для амплификации кодирующих областей генов подбирают исходя из сравнения известных нуклеотидных последовательностей, опубликованных в базах данных Интернета (GenBank). На основании сравнения выбирают наиболее консервативные участки, чтобы вероятность того, что праймер окажется комплементарен участку гена западносибирских изолятов боррелий, была высокой. Структуры подобранных праймеров представлены в табл.1. Каждый праймер на 5'-конце содержит последовательность, кодирующую сайт узнавания какой-либо эндонуклеазой рестрикции.
Амплификацию генов проводят с Pfu-ДНК полимеразой, фирмы «Сибэнзим» (Россия) согласно прилагаемой инструкции. Условия амплификации генов представлены в таблице 2. Начальная температура денатурации составляет 96°С в течение 3 мин, последующие циклы по 20 с; общее количество циклов реакции составляет 30. Концентрация каждого праймера в реакционной смеси составляет 0,2 нМ (наномоль).
Продукты амплификации анализируют в 1% агарозном геле, фрагменты генов идентифицируют по размеру. Каждый фрагмент гена нарабатывают в количестве 1-2 мкг. Ампликоны очищают стандартной процедурой фенол-хлороформной экстракции, переосаждают этанолом, растворяют в 100 мкл бидистиллированной воды и используют для гидролиза рестриктазами. Гидролиз ампликонов проводят из расчета 2-3 кратного избытка фермента в течение 16-20 часов, при температуре 37°С, в случае использования эндонуклеазы рестрикции XhoI - при 50°С. По окончании реакции ампликоны снова очищают фенол-хлороформной экстракцией, переосаждают и растворяют в 10-20 мкл бидистиллированной воды, 1/5 часть используют для электрофоретического анализа. Основную часть препаратов ДНК хранят при -20°С до лигирования, но не более месяца.
Ампликоны соответствующих генов боррелий клонируют в составе вектора pETm. На первом этапе клонируют первый из пары генов в составе вектора pETm, а именно гены rmospCBg, rmflaA, rmdbpB, rmospA и rmbmpA. Для этого вектор гидролизуют эндонуклеазами рестрикции, сайты которых имеются на 3'- и 5'-концах клонируемого гена, отделяют от продуктов гидролиза в агарозном геле и выделяют вектор из геля с помощью специального набора (процедуры аналогичны описанным в примере 1). Очищенные гены и очищенный вектор лигируют в эквимолярном соотношении 5:1 с помощью Т4 ДНК лигазы фирмы «Сибэнзим» (Россия) согласно инструкции производителя фермента. Лигазной смесью методом электропорации трансформируют клетки E.coli штамма BL21 (DE3). Трансформацию клеток E.coli лигазной смесью проводят в соответствии с руководством фирмы-производителя электропоратора - «PeqLab, Biotechnologie GmbH» (Германия). Трансформированные клетки рассевают на агаризованную среду LB, содержащую канамицин 30 мкг/мл, и выращивают в течение ночи при 37°С. Полученные клоны клеток анализируют на наличие встроенного гена боррелий с помощью ПЦР с использованием внешних праймеров PR268 и PR269 аналогично описанному методу в Примере 1. Продукты ПЦР анализируют в 2% агарозном геле, нужные клоны идентифицируют по размеру ампликонов. Из нескольких положительных клонов выделяют плазмиды стандартным щелочным методом и секвенируют для подтверждения нуклеотидной последовательности клонированного гена боррелий. В случае удовлетворительного результата приступают ко второму этапу сборки рекомбинантной ДНК, кодирующей полипептиды. В результате выполнения первого этапа получают плазмиды pETm-rmospCBg, pETm-rmflaA, pETm-rmdbpB, pETm-rmospA и pETm-rmbmpA, содержащие соответствующие гены боррелий.
На втором этапе в состав полученных рекомбинантных плазмид встраивают второй из пары генов, а именно гены rmospCBa, frflaB, rmospA, ДНК-фрагмент, кодирующий C-концевой фрагмент белка р83 западносибирского изолята Borrelia garinii (frp83). Для этого плазмиды pETm-rmospCBg, pETm-rmflaA, pETm-rmdbpB, pETm-rmospA, pETm-rmBmpA гидролизуют эндонуклеазами рестрикции, сайты которых имеются на 3'- и 5'-концах соответствующего клонируемого гена, очищают от продуктов гидролиза в агарозном геле и выделяют из геля с помощью набора (процедуры описаны в Примере 1). Очищенный ген (второй) и очищенную соответствующую плазмиду лигируют в эквимолярном соотношении 5:1, лигазной смесью трансформируют клетки E.coli и рассевают на агаризованную среду аналогично вышеописанной процедуре (первый этап). Полученные клоны клеток анализируют на наличие встроенного гена боррелий с помощью ПЦР с использованием внешних праймеров PR268 и PR269 аналогично описанному методу в Примере 1. Продукты ПЦР анализируют в 2% агарозном геле, нужные клоны идентифицируют по размеру ампликонов. Из нескольких положительных клонов выделяют плазмиды стандартным щелочным методом и секвенируют для подтверждения нуклеотидной последовательности клонированного гена боррелий.
Плазмиды с подтвержденным содержанием встроенных в них пар генов названы соответственно pET-rmbmpA-frp83, pETm-rmospA-frp83, pET-rmdbpB-rmospA, pET-rmflaA-frflaB и pET-rmospCBg-rmoSpCBa.
Пример 3. Экспрессия в клетках E.coli рекомбинантных химерных ДНК, кодирующих рекомбинантные химерные полипептиды, несущие эпитопы различных иммунодоминантных белков спирохет комплекса Borrelia burgdorferi sensu lato.
Экспрессию вышеназванных химерных генов, клонированных в составе вектора рЕТ, осуществляют в клетках E.coli штаммов BL21 (DE3) и Rosetta 2. В настоящем изобретении в качестве основного продуцента использован штамм BL21 (DE3).
Музейную культуру рекомбинантного клона либо колонию клеток с чашки подращивают в течение ночи в 5 мл среды LB при 37°С. Затем ночную культуру переносят в 500 мл свежей среды LB, содержащей канамицин 30 мкг/мл, и выращивают при 37°С до плотности клеток 0,8-1,0 о. е. длине волны 600 нм, затем добавляют изопропил-β-D-1-тиогалактопироназид (ИПТГ) до конечной концентрации 0,5 мМ. Культуру клеток инкубируют при 30°С и интенсивной аэрацией в течение 4 часов. По окончании клетки осаждают центрифугированием 10 мин при 3500 об/мин и хранят при -20°С до выделения рекомбинантных полипептидов.
Пример 4. Выделение и очистка рекомбинантных химерных полипептидов.
Выделение и очистку рекомбинантных химерных полипептидов из полученной биомассы (Пример 3) осуществляют с помощью одностадийной аффинной хроматографии лизата клеток на колонке с Ni-NTA-сефарозой CL-6B как в нативных, так и в денатурирующих условиях согласно протоколу фирмы-производителя "Quiagen" (США).
Для выделения рекомбинантных химерных полипептидов используют денатурирующие условия с применением гуанидина и мочевины. Клетки лизируют в 6М растворе гуанидинхлорида. Затем дебрис осаждают центрифугированием при 16000 g в течение 30 мин. Лизат инкубируют со смолой в течение часа при комнатной температуре, после чего суспензию загружают в колонку. Колонку промывают буфером, содержащим 8 М мочевину. Элюцию рекомбинантных химерных полипептидов осуществляют буфером, содержащим ступенчатый градиент имидазола: 20, 40, 60, 80, 250 и 500 мМ. Рекомбинантные химерные полипептиды, содержащие С-концевой 8-ми гистидиновый фрагмент, остаются связанными с сорбентом, в то время как бактериальные белки свободно проходят через колонку. Основной пул целевого рекомбинантного химерного полипептида смывают с колонки раствором 250 мМ имидазола. В процессе хроматографии собирают фракции, затем анализируют их электрофорезом в пластинах SDS-ПААГ по Леммли. Гели окрашивают раствором кумаси R250. Пример электрофоретического анализа фракций, полученных в процессе хроматографической очистки химерного белка rmBmpA-frp83, представлен на фиг.2.
Наиболее чистые фракции объединяют, замеряют концентрацию белка по Лоури и хранят при -20°С до использования в ИФА. Выход белков из одного литра культуры клеток Е.coli штамма BL21 (DE3) в среднем составляет 40-50 мг/л. Чистота рекомбинантных химерных белков достигает не менее 95%.
Пример 5. Использование очищенных рекомбинантных химерных полипептидов для иммуноферметного анализа.
Исследование иммунохимических свойств рекомбинантных химерных полипептидов проводят с помощью стандартного твердофазного иммуноферментного анализа (ИФА). Твердой фазой во всех вариантах ИФА служат 96-луночные плоскодонные планшеты из полистирола. Для исследования используют образцы сывороток крови различных групп доноров:
- группа больных с достоверным диагнозом ИКБ (сыворотка взята в период 2-8 недель с начала заболевания);
- группа здоровых доноров (сыворотки тестировались на наличие антител классов IgG и IgM к спирохетам комплекса B.burgdorferi s.1. и Treponema pallidum с помощью доступных коммерческих диагностических наборов).
Сыворотки были получены из 1-й городской инфекционной больницы г.Новосибирска и областного центра «СПИД» (г.Новосибирск).
Для приготовления рабочего раствора антигена и его последующей сорбции на планшетах используют 0,02 М карбонатно-бикарбонатный буферный раствор с рН-9,6. Для промывки планшетов, разведения сывороток и приготовления рабочего раствора конъюгата используют фосфатно-солевой буфер, содержащий 0,15М хлористого натрия и 0,1% твин-20 (ФСБ). Субстратный раствор включает 0,05М цитратный буфер (рН-5,0), перекись водорода (0,035%) и ортофенилендиамин (0,05%). Для выявления специфического комплекса антиген-антитело используют пероксидазные конъюгаты на основе моноклональных антител (МА) к IgG и IgM человека. Конъюгаты на основе МА были получены из лаборатории гибридомных технологий ЦНИИ рентгенорадиологии МЗ РФ.
Антиген разводят в карбонатно-бикарбонатном буфере (рН 9.6) до рабочей концентрации, вносят в лунки планшета по 100 мкл и оставляют для сорбции при +4°С на 15-18 часов. Рабочую концентрацию определяют при помощи тестирования сывороток здоровых доноров и сывороток пациентов с достоверным диагнозом клещевого боррелиоза на разных стадиях заболевания. После завершения сорбции планшеты с иммобилизованным антигеном промывают дважды карбонатно-бикарбонатным буферным раствором с 0.05% Твином-20, при этом в каждую лунку планшета вносят объем раствора не менее 250 мкл. По окончании промывки остатки жидкости удаляют, во все лунки планшета вносят по 100 мкл блокирующего раствора для сывороток, а затем по 10 мкл исследуемых сывороток, в разведении 1:10, планшет закрывают крышкой и инкубируют 30 мин при температуре 37°С. В качестве раствора для разведения сывороток используют глициновый буфер (ГСБ) с добавлением 2,0 мкг/мл лизата клеток E.coli BL21 (DE3) для устранения неспецифического связывания.
После окончания инкубации планшеты промывают моющим раствором (не менее 6 раз) и после удаления влаги в каждую лунку вносят 100 мкл конъюгата в рабочем разведении. Планшет инкубируют 30 мин при 37°С и затем промывают моющим раствором не менее 6 раз. Остатки влаги удаляют из планшета и в каждую лунку вносят по 100 мкл субстрата для пероксидазы. Субстрат готовят непосредственно перед использованием. Таблетку о-фенилендиамина растворяют в цитратно-фосфатном буфере до конечной концентрации 0,5 мкг/мл. После добавления субстрата планшет помещают в защищенное от света место и выдерживают 20 мин при комнатной температуре. Реакцию останавливают внесением в каждую лунку по 50 мкл 2 М раствора серной кислоты.
Результаты ИФА регистрируют на ридере Multiscan (Финляндия). Оптическую плотность (ОП) измеряют при длине волны 450 нм. Результат учитывают только в том случае, если среднее значение ОП в лунках с контролем конъюгата (ОПКк) не превышает 0.2, в лунках с К(-) среднее значение ОП (ОПК.) не более 0.2, а в лунках с К(+) среднее значение ОП (ОПк+) не менее 0.7. Результаты ИФА определяют по формуле:
ОП крит=ср.знач. ОП К(-)+0,2;
где 0,2 - коэффициент, установленный методом статистической обработки результатов.
Пробы, значения ОП которых ниже значения критической ОП, относят к серонегативным. Пробы, значения ОП которых равны или превышают значение критической ОП, оценивают как серопозитивные. Результаты исследования реактивности рекомбинатных химерных полипептидов с сыворотками представлены в табл.3.
Все сыворотки из группы здоровых доноров (n=10) показали отрицательный результат.
Данные ИФА, представленные в табл.3, убедительно доказывают специфичность взаимодействия рекомбинантных химерных полипептидов с IgG и IgM антителами больных иксодовым клещевым боррелиозом. Каждый заявленный рекомбинантный химерный полипептид может быть пригоден для диагностики иксодового клещевого боррелиоза, в том числе и для дифференцированного выявления ранней стадии и стадии диссеминированной инфекции.
СПИСОК ЛИТЕРАТУРЫ
1. Stanek G, Reiter M. The expanding Lyme Borrelia complex - clinical significance of genomic species? // Clin Microbiol Infect. 2011. 17. (4). 487-493. doi: 10.1111/j.1469-0691.2011.03492.x.
2. Postic D., Korenberg E., Gorelova N., Kovalevski Y.V., Bellenger E., Baranton G. Borrelia burgdorferi sensu lato in Russia and neighbouring countries: high incedence of mixed isolates // Res. Microbiol. 1997. 148. 691-702.
3. Коренберг Э.И., Нефедова В.В., Фадеева И.А., Горелова Н.Б. Основные итоги генотипирования боррелий в России // Бюл. сибирской медицины. 2006. Приложение 1. 87-92.
4. Масузава Т., Наумов Р.Л., Кудекен M., Харитоненков И.Г. Обнаружение Borrelia burgdorferi sensu stricto в Московской области, Россия // Медицинская паразитология и паразитарные болезни. 2001. (2). 52.
5. Gorelova N.B., Korenberg E.I., Postik D., lunicheva Iu.V., Riabova Т.Е. The first isolation of Borrelia burgdorferi sensu stricto in Russia // Zh. Mikrobiol. Epidemiol. Immunobiol.2001.4. 10-12.
6. Рудакова С.А., Коломеец А.Н., Самойленко И.Е., Кузьминов A.M., Рудаков Н.В. Экспресс-диагностика трансмиссивных патогенов как основа дифференцированного подхода к профилактике инфекций, передающихся иксодовыми клещами // Бюл. СО РАМН. 2007. №4. С.116-119.
7. Beklemishev А.В., А.К.Dobrotvorsky, A.V.Piterina, I.D.Ivanov, N.Yu.Nomokonova, N.N.Livanova. Detection and typing of Borrelia burgdorferi sensu lato genospecies in Ixodes persulcatus ticks in West Siberia, Russia. // FEMS Microbiol Lett. 2003 V.227, N.2, - P.157-161.
8. Рябченко А.В., Беклемишев А.Б. Мониторинг зараженности клещей рекреационной зоны г.Новосибирска возбудителями Лайм-боррелиоза. Вестник НГУ, 2007, Т.5, вып.1, С.106-110.
9. Мамаев А.Л., Рябченко А.В. и Беклемишев А.Б. Геновидовой состав спирохет Borrelia burgdorferi sensu lato, циркулировавших в иксодовых клещах, отловленных весной 2008 г. в рекреационной зоне г.Новосибирска. // Бюллетень СО РАМН. 2010, Т.30, №2. С.13-16.
10. Wang, G., van Dam A.P., Schwartz // J.Clin. Microbiol. Rev. - Molecular typing of Borrelia burgdorferi sensu lato: taxonomic, epidemiological, and clinical implications. - 1999. - Vol.12. - P.633-653.
11. Parola, P., and D.Raoult. Tick-borne bacterial diseases emerging in Europe // Clin. Microbiol. Infect. - 2001. - V.7. - P.80-83.
12. Bruckbauer H.R., Preac-Mursic V., Fuchs R., Wilske B. Cross-reactive proteins of Borrelia burgdorferi // Eur J Clin Microbiol Infect Dis. - 1992. - V.11. - N.3 - P.224-232.
13. Carreiro MM, Laux DC, Nelson DR. Characterization of the heat shock response and identification of heat shock protein antigens of Borrelia burgdorferi // Infect Immun. - 1990. - V.58. - N.7. - P.2186-2191.
14. Magnarelli LA, Miller JN, Anderson JF, Riviere GR. Cross-reactivity of nonspecific treponemal antibody in serologic tests for Lyme disease // J Clin Microbiol. - 1990. - V.28. - N.6. - P.1276-1279.
15. Wilske В, Fingerle V, Schulte-Spechtel U. Microbiological and serological diagnosis of Lyme borreliosis // FEMS Immunol Med Microbiol. - 2007. - V.49. - N.1. - P.13-21.
16. Ge Y., li С., Corum L., et al. Structure and expression of the FlaА periplasmic flagellar protein of Borrelia burgdorferi. // Journal of Bacteriology. - 1998. - V.180. - P.2418-2425.
17. Gilmore R.D. Jr., Murphree R.L., James A.M., Sullivan S.A., Johnson B.J. The Borrelia burgdorferi 37-kilodalton immunoblot band (P37) used in serodiagnosis of early lyme disease is the flaA gene product // J. Clin. Microbiol. - 1999. - V.37. - P.548-552).
18. Berland R., Fikrig E., Rahn D., Hardin J. and FlaVell R.. Molecular characterization of the humoral response to the 41-kilodalton flagellar antigen of Borrelia burgdorferi, the Lyme disease agent. // Infection and Immunity. - 1991. - V.59. -. P.3531-35).
19. Steere A.C., Klitz W., Drouin E.E., Falk B.A., Kwok W.W., Nepom G.T., Baxter-Lowe L.A. Antibiotic-refractory Lyme arthritis is associated with HLA-DR molecules that bind a Borrelia burgdorferi peptide // J. Exp. Med. - 2006. - V.203. - P.961-971.
20. Aberer E. Lyme borreliosis - an update // JDDG. - 2007. - №5. - P.406-413.
21. Heikkila Т., Seppala I., Saxen H., Panelius J., Yrjanainen H., Lahdenne P. Species-specific serodiagnosis of Lyme arthritis and neuroborreliosis due to Borrelia burgdorferi sensu stricto, B.afzelii, and B.garinii by using decorin binding protein A // J. Clin. Microbiol. - 2002. - V.40. - P.453-460.
22. Heikkila Т., Seppala I., Saxen H., Panelius J., Peltomaa M., Huppertz H.I., Lahdenne P. Cloning of the gene encoding the decorin-binding protein В (DbpB) in Borrelia burgdorferi sensu lato and characterisation of the antibody responses to DbpB in Lyme borreliosis // J. Med. Microbiol. - 2002. - V.51. - P.641-648.
23. Roessler D., Hauser U. and Wilske В. Heterogeneity of BmpA (P39) among European isolates of Borrelia burgdorferi sensu lato and influence of interspecies variability on serodiagnosis. // Journal of Clinical Microbiology. - 1997. - V.35. - P.2752-2758).
24. EP 1865062 «Altered OspA of Borrelia burgdorferi)), A16K 39/02, C07K 14/20, опубл. 12.12.2007 г.
25. US 5747294 «Compositions and methods for the prevention and diagnosis of Lyme disease», C07K 14/20, A61K 39/00, опубл. 05.05.1998 г.
26. US 5686267 «Nucleic acid molecule encoding antigen associated with lyme disease», C07K 14/20, опубл. 11.11.1997 г.
27. WO 0078966 «Groups of Borrelia burgdorferi and Borrelia afzelii that cause Lyme disease in humans», A61K 39/02, опубл. 28.12.2000 г.
28. RU №2260047 «Рекомбинантная плазмидная ДНК, обеспечивающая синтез иммунодоминантного белка Borrelia garinii NT29, используемого для диагностики Лайм-боррелиоза (варианты)», C12N 15/09 опубл. 10.09.2005 БИ №25.
29. US №20040142399 « Recombinant FlaA as a diagnostic reagent», C07K 14/20, опубл. 22.07.2004 г.
30. US №6248562 «Chimeric proteins comprising borrelia polypeptides and uses therefor», C07K 14/20, опубл. 19.06.2001 г.
31. Манзенюк И.Н., Манзенюк 0.10. Клещевые боррелиозы (болезнь Лайма). Кольцове. 2005 г.// http://entomologs.ru/laima_prodrobno_7.php
32. Marangoni A., Sparacino M., Cavrini F., Stomi E., Mondardini V., Sambri V. and Cevenini R. Comparative evaluation of three different ELISA methods for the diagnosis of early culture-confirmed Lyme disease in Italy.// J Med Microbiol. - 2005. - V.54. - N.4. - P.361-367.
33. Лайм-боррелиоз (Borrelios) // http://laboratory.rusmedserv.com/diaginfect/sintez/laimborrelioz/
34. Компания NovaTec представляет новые наборы для диагностики боррелиоза // Сайт «Медико-диагностическая лаборатория», рубрика «Инновации в медицине». http://www.meddialab.ru/text/v_poslednee_.htm?PHPSESSID=7ff9945d79e7e1bf13410b48a611ca04
35. National Center for Biotechnology Information, USA. http://www.ncbi.nlm.nih.gov
36. Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии. Молекулярное клонирование // М., 1984. С.157-185.
37. Рябченко А.В., И.В. Ивлева, А.Б. Беклемишев. Комплексная оценка зараженности клещей Ixodes persulcatus, распространенных в рекреационной зоне Новосибирского научного центра, спирохетами Borrelia burgdorferi s.1. // Ж. инфекционной патологии, 2004, Т.11, №3-5, - С.107-110.











Изобретение относится к области медицины и касается рекомбинантных химерных полипептидов, несущих эпитопы различных иммунодоминантных белков спирохет комплекса Borellia Burgdorferisensu lato, и способа серодиагностики иксодового клещевого боррелиоза. Предложены рекомбинантные химерные полипептиды rmBmpA-frp83, rmOspA-frp83, rmDbpB-rmOspA, rmFlaA-frFlaB и rmOspCBg-rmOspCBa, полученные на основе экспрессии генов, амплифицированных методом ПЦР на ДНК западносибирского изолята Borrelia garinii 20047T или, в случае белка rmOspCBa, на ДНК изолята Borrelia afzelii. Представленные изобретения расширяют арсенал рекомбинантных полипептидов, пригодных для серодиагностики иксодового клещевого боррелиоза, повышения специфичности и чувствительности диагностики ИКБ, в том числе дифференциальной диагностики ранней стадии и стадии диссеминированной инфекции в регионах распространения западносибирских изолятов Borrelia burgdorferi s.1., и могут быть использованы в качестве антигенов для иммуноферментного анализа. 6 н. и 2 з.п. ф-лы, 2 ил., 3 табл., 5 пр.
1. Рекомбинантный химерный полипептид для серодиагностики иксодового клещевого боррелиоза - rmBmpA-frp83 (SEQ ID NO 4) размером 591 а.о. с молекулярной массой 65,89 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка BmpA западносибирского изолята Borrelia garinii (rmBmpA) размером 324 а.о. с молекулярной массой 35,3 кДа, а с С-конца - аминокислотную последовательность C-концевого фрагмента белка р83 западносибирского изолята Borrelia garinii (frp83) западносибирского изолята Borrelia garinii размером 248 а.о. с молекулярной массой 28,17 кДа.
2. Рекомбинантный химерный полипептид для серодиагностики иксодового клещевого боррелиоза - rmOspA-frp83 (SEQ ID NO 6) размером 521 а.о. с молекулярной массой 57,99 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка OspA западносибирского изолята Borrelia garinii (rmOspA) размером 257 а.о. с молекулярной массой 27,79 кДа, а с C-конца - аминокислотную последовательность С-концевого фрагмента белка р83 западносибирского изолята Borrelia garinii (frp83) размером 248 а.о. с молекулярной массой 28,17 кДа.
3. Рекомбинантный химерный полипептид для серодиагностики иксодового клещевого боррелиоза - rmDbpB-rmOspA (SEQ ID NO 8) размером 432 а.о. с молекулярной массой 47,13 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка DbpB западносибирского изолята Borrelia garinii (rmDbpB) размером 156 а.о. с молекулярной массой 17,02 кДа, а с С-конца - аминокислотную последовательность рекомбинантного зрелого белка OspA западносибирского изолята Borrelia garinii (rmOspA) размером 257 а.о. с молекулярной массой 27,79 кДа.
4. Рекомбинантный химерный полипептид для серодиагностики иксодового клещевого боррелиоза - rmFlaA-frFlaB (SEQ ID NO 10) размером 587 а.о. с молекулярной массой 64,76 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка FlaA западносибирского изолята Borrelia garinii (rmFlaA) размером 318 а.о. с молекулярной массой 35,82 кДа, а с С-конца - аминокислотную последовательность фрагмента флагеллярного белка FlaB, специфичного для спирохет B.burgdorferi s.1. (frFlaB) размером 250 а.о. с молекулярной массой 26,62 кДа.
5. Рекомбинантный химерный полипептид для серодиагностики иксодового клещевого боррелиоза - rmOspCBg-rmOspCBa (SEQ ID NO 12) размером 390 а.о. с молекулярной массой 40,83 кДа, содержащий с N-конца молекулы аминокислотную последовательность рекомбинантного зрелого белка OspC западносибирского изолята Borrelia garinii (rmOspCBg) размером 188 а.о. с молекулярной массой 19,70 кДа, а с С-конца - аминокислотную последовательность рекомбинантного зрелого белка OspC западносибирского изолята Borrelia afzelii (rmOspCBa) размером 186 а.о. с молекулярной массой 19,19 кДа.
6. Способ серодиагностики иксодового клещевого боррелиоза методом иммуноферментного анализа, отличающийся тем, что в качестве антигенов используют комбинации рекомбинантных химерных полипептидов по пп.1-5.
7. Способ по п.6, отличающийся тем, что для диагностики ранней стадии в качестве антигенов используют комбинацию рекомбинантных химерных полипептидов по пп.4, 5.
8. Способ по п.6, отличающийся тем, что для диагностики стадии диссеминированной инфекции в качестве антигенов используют комбинацию рекомбинантных химерных полипептидов по пп.1-3.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ИММУНОДОМИНАНТНОГО БЕЛКА BORRELIA GARINII, ИСПОЛЬЗУЕМОГО ДЛЯ ДИАГНОСТИКИ ЛАЙМ-БОРРЕЛИОЗА (ВАРИАНТЫ) | 2003 |
|
RU2260047C2 |
| СПОСОБ ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКИ КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ИКСОДОВОГО КЛЕЩЕВОГО БОРРЕЛИОЗА И ИХ МИКСТИНФЕКЦИЙ У ДЕТЕЙ | 2010 |
|
RU2433404C1 |
| US 20040142399 A1, 22.07.2004 | |||
| US 20090060932 A2, 05.03.2009 | |||
| US 0005965702 A1, 12.10.1999 | |||

