Область техники
Изобретение в общем относится к химерным полипептидам OspA, нуклеиновым кислотам, кодирующим полипептиды, композициям, содержащим такие молекулы, и способам их применения.
Известный уровень техники
Лаймская болезнь представляет собой переносимую клещами болезнь, вызываемую Borrelia burgdorferi sensu lato (s.l.). Болезнь типично характеризуется появлением распространяющейся красной сыпи на месте укуса клеща, за которой могут последовать системные осложнения, включая менингит, кардит или артрит. Почти все случаи лаймской болезни вызываются одним из трех геновидов - Borrelia afzelii, Borrelia garinii и Borrelia burgdorferi sensu stricto (s.s.). В Европе обнаружены все три вида, инфицирующие людей. Однако в Северной Америке найден только один вид - Borrelia burgdorferi sensu stricto. Borrelia burgdorferi представляет собой вид грамотрицательных бактерий класса спирохет рода Borrelia. Лечение лаймской болезни антибиотиками обычно является эффективным, но у некоторых пациентов развивается хроническая инвалидизирующая форма болезни, затрагивающая суставы или нервную систему, существенно не улучшающаяся даже после терапии парентеральными антибиотиками, что указывает на потребность в вакцине для популяций с высокой степенью риска.
Внешний поверхностный белок A (OspA) представляет собой 31-кДа антиген, экспрессируемый видом Borrelia burgdorferi s.l., присутствующим в средней кишке иксодовых клещей. Была продемонстрирована эффективность OspA для профилактики лаймской болезни в Северной Америке (Steere et al., N. Engl. J. Med. 339: 209-15,1998; Sigal et al., N. Engl. J. Med. 339:216-22, 1998; erratum in: N. Engl. J. Med. 339:571, 1998). Амино-конец полностью процессированного OspA представляет собой цистеиновый остаток, который посттрансляционно модифицируется тремя жирными ацильными цепями, закрепляющими белок на внешней поверхности бактериальной мембраны (Bouchon et al., Anal. Biochem. 246: 52-61, 1997). Сообщается, что липидирование OspA стабилизирует молекулу (Luft, личное сообщение) и является существенным для защиты в отсутствие сильного адъюванта (Erdile et al., Infect. Immun. 61: 81-90, 1993). Растворимая рекомбинантная форма белка, не имеющая амино-концевого липидного мембраносвязывающего фрагмента, была со-кристаллизована с фрагментом Fab агглютинирующего мышиного моноклонального антитела для определения структуры OspA, который, как было продемонстрировано, содержит 21 антипараллельную β-нить с последующей одной α-спиралью (Li et al., Proc. Natl. Acad. Sci. U.S.A. 94:3584-9, 1997).
Моновалентная вакцина на основе OspA (LYMErix®) для профилактики лаймской болезни выпущена в продажу в США. Однако, в Европе гетерогенность последовательностей OspA у трех геновидов препятствует широкой защите вакциной на основе OspA от одного штамма (Gern et al., Vaccine 15:1551-7, 1997). В европейских изолятах были определены семь основных серотипов OspA (обозначенных как серотипы 1-7, Wilske et al., J. Clin. Microbiol. 31:340-50, 1993). Серотипы OspA коррелируют с видами; серотип 1 соответствует B. burgdorferi s.s., серотип 2 соответствует B. afzelii, и серотипы 3-7 соответствуют B. garinii.
Защитный иммунитет, приобретенный в результате иммунизации с помощью OspA, является исключительным, поскольку взаимодействие между иммунным ответом хозяина и патогеном происходит не в хозяине, а в средней кишке клеща-переносчика. В случае лаймской болезни, клещ выступает в роли переносчика или носителя при передаче лаймской болезни от животных людям. OspA-специфическое антитело, приобретенное во время кормления инфицированного клеща, препятствует переносу B. burgdorferi s.l. иммунизированному млекопитающему-хозяину (de Silva et al., J. Exp. Med. 183: 271-5, 1996). Защита является антитело-медиируемой и преимущественно обеспечивается бактерицидным антителом, хотя антитело, блокирующее прикрепление спирохеты к рецептору выстилки кишечного эпителия клеща, также может быть эффективным (Pal et al., J. Immunol. 166: 7398-403, 2001).
Рациональная разработка эффективных вакцин OspA требует идентификации защитных эпитопов, таких как определяемых защитным моноклональным антителом LA-2 (Golde et al., Infect. Immun. 65: 882-9, 1997). Анализ способами рентгеновской кристаллографии и ЯМР был использован для идентификации иммунологически важных гипервариабельных доменов OspA и локализации эпитопа LA-2 путем картирования на аминокислотах 203-257 (Ding et al., J. Mol. Biol. 302: 1153-64, 2000; Luft et al. J Infect Dis. 185 (Suppl. 1): S46-51, 2002).
В данной области техники существует потребность в разработке вакцины OspA, обеспечивающей широкую защиту против различных видов Borrelia, встречающихся в США, Европе и других регионах. Приведенное далее описание содержит характеристики такой вакцины.
Сущность изобретения
Изобретение направлено на удовлетворение одной или более потребностей в данной области техники, касающихся профилактики и лечения лаймской болезни или лайм-боррелиоза.
Изобретение включает химерный полипептид, содержащий первый полипептидный фрагмент внешнего поверхностного белка A (OspA) серотипа 3 Borrelia garinii и второй полипептидный фрагмент белка OspA серотипа 5 Borrelia garinii, обладающий способностью индуцировать иммунный ответ против белка OspA серотипа 3 и белка OspA серотипа 5. В некоторых аспектах, химерный полипептид содержит N-концевой полипептидный фрагмент белка OspA серотипа 5 и C-концевой полипептидный фрагмент белка OspA серотипа 3. В других аспектах, химерный полипептид содержит N-концевой полипептидный фрагмент белка OspA серотипа 3 и C-концевой полипептидный фрагмент белка OspA серотипа 5. В определенных аспектах, химерный полипептид дополнительно содержит N-концевой полипептидный фрагмент внешнего поверхностного белка B (OspB) Borrelia, причем полипептидный фрагмент OspB содержит лидерную последовательность OspB. В конкретных аспектах, химерный полипептид содержит аминокислотную последовательность, состоящую из по меньшей мере 200 аминокислотных остатков, с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO: 173. В различных аспектах, химерный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 173. В других аспектах, химерный полипептид состоит из аминокислотной последовательности, приведенной в SEQ ID NO: 173.
Изобретение включает композиции, содержащие химерный полипептид по изобретению и фармацевтически приемлемый носитель. В некоторых аспектах, такие композиции дополнительно содержат дополнительный полипептид внешнего поверхностного белка A (OspA) Borrelia. В некоторых аспектах, такие композиции дополнительно содержат дополнительный полипептид внешнего поверхностного белка B (OspB) Borrelia. В конкретных аспектах, дополнительный полипептид содержит N-концевой полипептидный фрагмент внешнего поверхностного белка B (OspB) Borrelia, причем полипептидный фрагмент OspB содержит лидерную последовательность OspB. В различных аспектах, Borrelia представляет собой Borrelia burgdorferi, Borrelia afzelii, Borrelia garinii, Borrelia japonica, Borrelia andersonii, Borrelia bissettii, Borrelia sinica, Borrelia turdi, Borrelia tanukii, Borrelia valaisiana, Borrelia lusitaniae, Borrelia spielmanii, Borrelia miyamotoi или Borrelia lonestar.
В некоторых аспектах, дополнительный полипептид представляет собой химерный полипептид, содержащий первый полипептидный фрагмент внешнего поверхностного белка A (OspA) серотипа 4 Borrelia garinii и второй полипептидный фрагмент белка OspA серотипа 6 Borrelia garinii, обладающий способностью индуцировать иммунный ответ против белка OspA серотипа 4 и белка OspA серотипа 6. В конкретных аспектах, дополнительный полипептид содержит N-концевой полипептидный фрагмент белка OspA серотипа 6 и C-концевой полипептидный фрагмент белка OspA серотипа 4. В других аспектах, дополнительный полипептид содержит N-концевой полипептидный фрагмент белка OspA серотипа 4 и C-концевой полипептидный фрагмент белка OspA серотипа 6. В определенных аспектах, дополнительный полипептид содержит аминокислотную последовательность, имеющую по меньшей мере 200 аминокислотных остатков, с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO: 171. В конкретных аспектах, дополнительный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 171. В дополнительных аспектах, дополнительный полипептид состоит из аминокислотной последовательности, приведенной в SEQ ID NO: 171.
В некоторых аспектах, дополнительный полипептид представляет собой химерный полипептид, содержащий первый полипептидный фрагмент внешнего поверхностного белка A (OspA) серотипа 1 Borrelia burgdorferi sensu stricto и второй полипептидный фрагмент белка OspA серотипа 2 Borrelia afzelii, обладающий способностью индуцировать иммунный ответ против белка OspA серотипа 1 и белка OspA серотипа 2. В конкретных аспектах, дополнительный полипептид содержит N-концевой полипептидный фрагмент белка OspA серотипа 1 и C-концевой полипептидный фрагмент белка OspA серотипа 2. В других аспектах, дополнительный полипептид содержит N-концевой полипептидный фрагмент белка OspA серотипа 2 и C-концевой полипептидный фрагмент белка OspA серотипа 1. В конкретных аспектах, дополнительный полипептид дополнительно содержит N-концевой полипептидный фрагмент внешнего поверхностного белка B (OspB) Borrelia, причем полипептидный фрагмент OspB содержит лидерную последовательность OspB. В определенных аспектах, дополнительный полипептид содержит аминокислотную последовательность, включающую по меньшей мере 200 аминокислотных остатков, с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO: 169. В конкретных аспектах, дополнительный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 169. В дополнительных аспектах, дополнительный полипептид состоит из аминокислотной последовательности, приведенной в SEQ ID NO: 169.
Изобретение включает композиции, содержащие по меньшей мере три химерных полипептида OspA, причем полипептиды имеют разные последовательности. В некоторых аспектах, химерные полипептиды OspA индивидуально содержат аминокислотные последовательности, приведенные в SEQ ID NO: 169, 171 и 173. В других аспектах, химерные полипептиды OspA индуцируют иммунный ответ против по меньшей мере белков OspA серотипов 1, 2, 3, 4, 5 и 6.
Изобретение включает молекулу химерной нуклеиновой кислоты, содержащую первый фрагмент нуклеотидной последовательности кодирующей области внешнего поверхностного белка A (OspA) серотипа 3 Borrelia garinii и второй фрагмент нуклеотидной последовательности кодирующей области белка OspA серотипа 5 Borrelia garinii, причем молекула нуклеиновой кислоты, кодирующая полипептид, обладает способностью индуцировать иммунный ответ против белка OspA серотипа 3 и белка OspA серотипа 5. В некоторых аспектах, молекула химерной нуклеиновой кислоты содержит 5’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 5 и 3’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 3. В других аспектах, молекула химерной нуклеиновой кислоты содержит 3’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 5 и 5’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 3. В различных аспектах, молекула химерной нуклеиновой кислоты дополнительно содержит 5’-концевой фрагмент нуклеотидной последовательности внешнего поверхностного белка B (OspB) Borrelia, причем фрагмент нуклеотидной последовательности OspB содержит лидерную последовательность OspB. В определенных аспектах, молекула химерной нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности с по меньшей мере примерно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с последовательностью нуклеиновой кислоты, приведенной в SEQ ID NO: 172; и (б) нуклеотидной последовательности, комплементарной к (а). В других аспектах, молекула химерной нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности, кодирующей полипептид с по меньшей мере примерно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO: 173; и (б) нуклеотидной последовательности, комплементарной к (а). В конкретных аспектах изобретения, молекула химерной нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 173, причем полипептид содержит замещение от одной до 25 консервативных аминокислот; (б) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 173, причем полипептид содержит инсерцию от одной до 25 консервативных аминокислот; (в) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 173, причем полипептид содержит внутреннюю делецию от одной до 25 консервативных аминокислот; (г) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 173, причем полипептид содержит C- и/или N-терминальное усечение от одной до 25 аминокислот; (д) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 173, причем полипептид содержит модификацию от одной до 25 аминокислот, выбранную из аминокислотных замещений, аминокислотных инсерций, аминокислотных делеций, C-терминального усечения, или N-терминального усечения; и (е) нуклеотидной последовательности, комплементарной к любой из (а)-(д). В различных аспектах, такие замещения, инсерции, делеции или модификации находятся в любых из положений аминокислотных остатков 1-4, 6, 8, 9, 11, 16, 18, 20-28, 47, 49, 50, 81, 82, 83, 100, 139, 155, 160, 176, 189, 190 и 250 SEQ ID NO: 173. В некоторых аспектах, молекула химерной нуклеиновой кислоты содержит нуклеотидную последовательность, приведенную в SEQ ID NO: 172. В других аспектах, молекула химерной нуклеиновой кислоты состоит из нуклеотидной последовательности, приведенной в SEQ ID NO: 172.
Изобретение включает векторы, клетки-хозяева и способы получения полипептидов путем культивации клеток-хозяев, описанных тут. В некоторых аспектах, изобретение включает вектор, содержащий любые молекулы нуклеиновой кислоты, описанные тут. В других аспектах, изобретение включает клетку-хозяина, которая содержит такие векторы. В некоторых аспектах, клетка-хозяин представляет собой эукариотическую клетку. В других аспектах, клетка-хозяин представляет собой прокариотическую клетку. В различных аспектах, способ получения полипептида включает культивацию клеток-хозяев, описанных тут, в условиях, пригодных для экспрессии полипептида и, необязательно, выделение полипептида из культуры. В различных аспектах, изобретение включает композиции, содержащие любые из таких химерных молекул нуклеиновой кислоты или любые векторы, содержащие такие молекулы нуклеиновой кислоты и фармацевтически приемлемый носитель или носители.
Как описано выше, изобретение включает композицию, содержащую молекулу химерной нуклеиновой кислоты, содержащую первый фрагмент нуклеотидной последовательности кодирующей области внешнего поверхностного белка A (OspA) серотипа 3 Borrelia garinii и второй фрагмент нуклеотидной последовательности кодирующей области белка OspA серотипа 5 Borrelia garinii, причем молекула нуклеиновой кислоты, кодирующая полипептид, обладает способностью индуцировать иммунный ответ против белка OspA серотипа 3 и белка OspA серотипа 5. В некоторых аспектах, композиция дополнительно содержит дополнительную молекулу нуклеиновой кислоты, кодирующую внешний поверхностный белок A (OspA) Borrelia. В других аспектах, композиция дополнительно содержит дополнительную молекулу нуклеиновой кислоты, кодирующую внешний поверхностный белок B (OspB) Borrelia. В конкретных аспектах, дополнительная молекула нуклеиновой кислоты дополнительно содержит 5’-концевой фрагмент нуклеотидной последовательности внешнего поверхностного белка B (OspB) Borrelia, причем фрагмент нуклеотидной последовательности OspB содержит лидерную последовательность OspB. В различных аспектах, Borrelia представляет собой Borrelia burgdorferi, Borrelia afzelii, Borrelia garinii, Borrelia japonica, Borrelia andersonii, Borrelia bissettii, Borrelia sinica, Borrelia turdi, Borrelia tanukii, Borrelia valaisiana, Borrelia lusitaniae, Borrelia spielmanii, Borrelia miyamotoi или Borrelia lonestar.
В некоторых аспектах, дополнительная молекула нуклеиновой кислоты представляет собой химерную молекулу нуклеиновой кислоты, содержащую первый фрагмент нуклеотидной последовательности кодирующей области внешнего поверхностного белка A (OspA) серотипа 6 Borrelia garinii и второй фрагмент нуклеотидной последовательности кодирующей области белка OspA серотипа 4 Borrelia garinii, причем молекула нуклеиновой кислоты, кодирующая полипептид, обладает способностью индуцировать иммунный ответ против белка OspA серотипа 6 и белка OspA серотипа 4. В других аспектах, дополнительная молекула нуклеиновой кислоты содержит 5’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 6 и 3’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 4. В различных аспектах, дополнительная молекула нуклеиновой кислоты содержит 5’-концевую нуклеотидную последовательность, кодирующую фрагмент белка OspA серотипа 4, и 3’-концевую нуклеотидную последовательность, кодирующую фрагмент белка OspA серотипа 6. В некоторых аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с нуклеотидной последовательностью, приведенной в SEQ ID NO: 170; и (б) нуклеотидной последовательности, комплементарной к (а). В других аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности, кодирующей полипептид с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полипептидом, содержащим аминокислотную последовательность, приведенную в SEQ ID NO: 171; и (б) нуклеотидной последовательности, комплементарной к (а). В конкретных аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 171, причем полипептид содержит замещение от одной до 25 консервативных аминокислот; (б) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 171, причем полипептид содержит инсерцию от одной до 25 консервативных аминокислот; (в) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 171, причем полипептид содержит внутреннюю делецию от одной до 25 консервативных аминокислот; (г) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 171, причем полипептид содержит C- и/или N-терминальное усечение от одной до 25 аминокислот; (д) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 171, причем полипептид содержит модификацию от одной до 25 аминокислот, выбранную из аминокислотных замещений, аминокислотных инсерций, аминокислотных делеций, C-терминального усечения, или N-терминального усечения; и (е) нуклеотидной последовательности, комплементарной к любой из (а)-(д). В различных аспектах, замещения, инсерции, делеции или модификации находятся в любых из положений аминокислотных остатков 1-4, 6, 8, 9, 11, 16, 18, 20-28, 47, 49, 50, 81, 82, 83, 100, 139, 155, 160, 176, 189, 190 и 250 SEQ ID NO: 171. В некоторых аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, приведенную в SEQ ID NO: 170. В других аспектах, дополнительная молекула нуклеиновой кислоты состоит из нуклеотидной последовательности, приведенной в SEQ ID NO: 170.
В других аспектах, дополнительная молекула нуклеиновой кислоты представляет собой химерную молекулу нуклеиновой кислоты, содержащую первый фрагмент нуклеотидной последовательности кодирующей области внешнего поверхностного белка A (OspA) серотипа 1 Borrelia burgdorferi sensu stricto и второй фрагмент нуклеотидной последовательности кодирующей области белка OspA серотипа 2 Borrelia afzelii, причем молекула нуклеиновой кислоты, кодирующая полипептид, обладает способностью индуцировать иммунный ответ против белка OspA серотипа 1 и белка OspA серотипа 2. В определенных аспектах, дополнительная молекула нуклеиновой кислоты содержит 5’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 1 и 3’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 2. В других аспектах, дополнительная молекула нуклеиновой кислоты содержит 5’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 2 и 3’-концевую нуклеотидную последовательность, кодирующую фрагмент кодирующей области белка OspA серотипа 1. В различных аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с нуклеотидной последовательностью, приведенной в SEQ ID NO: 168; и (б) нуклеотидной последовательности, комплементарной к (а). В дополнительных аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности, кодирующей полипептид с по меньшей мере или приблизительно 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полипептидом, содержащим аминокислотную последовательность, приведенную в SEQ ID NO: 169; и (б) нуклеотидной последовательности, комплементарной к (а). В некоторых аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы, состоящей из: (а) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 169, причем полипептид содержит замещение от одной до 25 консервативных аминокислот; (б) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 169, причем полипептид содержит инсерцию от одной до 25 консервативных аминокислот; (в) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 169, причем полипептид содержит внутреннюю делецию от одной до 25 консервативных аминокислот; (г) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 169, причем полипептид содержит C- и/или N-терминальное усечение от одной до 25 аминокислот; (д) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 169, причем полипептид содержит модификацию от одной до 25 аминокислот, выбранных из аминокислотных замещений, аминокислотных инсерций, аминокислотных делеций, C-терминального усечения, или N-терминального усечения; и (е) нуклеотидной последовательности, комплементарной к любой из (а)-(д). В различных аспектах, замещения, инсерции, делеции или модификации находятся в любых из положений аминокислотных остатков 1-4, 6, 8, 9, 11, 16, 18, 20-28, 47, 49, 50, 81, 82, 83, 100, 139, 155, 160, 176, 189, 190 и 250 SEQ ID NO: 169. В некоторых аспектах, дополнительная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, приведенную в SEQ ID NO: 168. В других аспектах, дополнительная молекула нуклеиновой кислоты состоит из нуклеотидной последовательности, приведенной в SEQ ID NO: 168.
Как описано выше, изобретение включает композицию, содержащую молекулу химерной нуклеиновой кислоты, содержащую первый фрагмент нуклеотидной последовательности кодирующей области внешнего поверхностного белка A (OspA) серотипа 3 Borrelia garinii и второй фрагмент нуклеотидной последовательности кодирующей области белка OspA серотипа 5 Borrelia garinii, причем молекула нуклеиновой кислоты, кодирующая полипептид, обладает способностью индуцировать иммунный ответ против белка OspA серотипа 3 и белка OspA серотипа 5. В некоторых аспектах, композиция дополнительно содержит по меньшей мере две дополнительные молекулы нуклеиновой кислоты, кодирующие внешний поверхностный белок A (OspA) Borrelia. В различных аспектах, такие дополнительные молекулы нуклеиновой кислоты имеют разные нуклеотидные последовательности. В определенных аспектах, композиция по изобретению содержит по меньшей мере три молекулы нуклеиновой кислоты, кодирующие внешний поверхностный белок A (OspA) Borrelia, причем молекулы нуклеиновой кислоты имеют разные нуклеотидные последовательности. В конкретных аспектах, композиция по изобретению содержит молекулы нуклеиновой кислоты, причем молекулы нуклеиновой кислоты индивидуально содержат нуклеотидные последовательности, приведенные в SEQ ID NO: 168, 170 и 172. В некоторых аспектах, композиция по изобретению содержит химерные молекулы нуклеиновой кислоты, причем молекулы нуклеиновой кислоты кодируют полипептиды, индуцирующие иммунный ответ против по меньшей мере белков OspA серотипов 1, 2, 3, 4, 5 и 6.
Изобретение также включает иммуногенные композиции. В некоторых аспектах, иммуногенная композиция по изобретению содержит любую из описанных тут композиций и фармацевтически приемлемый носитель. В различных аспектах, иммуногенная композиция обладает способностью индуцировать продуцирование антитела, которое специфически связывается с внешним поверхностным белком A (OspA). В определенных аспектах, иммуногенная композиция обладает способностью индуцировать продуцирование антитела, которое специфически связывается с Borrelia. В конкретных аспектах, иммуногенная композиция обладает способностью индуцировать продуцирование антитела, нейтрализующего Borrelia. В некоторых аспектах, антитело продуцируется животным. В дополнительных аспектах, животное представляет собой млекопитающее. В других аспектах, млекопитающее является человеком.
Изобретение дополнительно включает композиции вакцины. В некоторых аспектах, композиция вакцины по изобретению содержит любую иммуногенную композицию, описанную тут, и фармацевтически приемлемый носитель. В различных аспектах, изобретение включает комбинированную вакцину. В определенных аспектах, комбинированная вакцина по изобретению содержит любую композицию вакцины, описанную тут, в сочетании с по меньшей мере второй композицией вакцины. В некоторых аспектах, вторая композиция вакцины защищает от переносимой клещами болезни. В различных аспектах, переносимая клещами болезнь представляет собой пятнистую лихорадку Скалистых гор, бабезиоз, возвратную лихорадку, колорадскую клещевую лихорадку, моноцитарный эрлихиоз человека (HME), гранулоцитарный эрлихиоз человека (HGE), клещевую сыпную лихорадку STARI (Southern Tick-Associated Rash Illness), туляремию, клещевой паралич, энцефалит Повассан, ку-лихорадку, конго-крымскую геморрагическую лихорадку, цитауксзооноз (cytauxzoonosis), марсельскую лихорадку или клещевой энцефалит. В других аспектах, вторая композиция вакцины представляет собой вакцину, выбранную из группы, состоящей из: вакцины клещевого энцефалита, вакцины японского энцефалита и вакцины пятнистой лихорадки Скалистых гор. В различных аспектах, вторая композиция вакцины имеет сезонный график иммунизации, совместимый с иммунизацией против инфекции Borrelia или лаймской болезни.
Изобретение также включает способы индуцирования иммунологического ответа у субъекта. В различных аспектах, такие способы включают стадию введения субъекту любой из иммуногенных композиций или композиций вакцины, описанных тут, в количестве, эффективно индуцирующем иммунологический ответ. В определенных аспектах, иммунологический ответ включает продуцирование анти-OspA антитела.
Изобретение включает способы профилактики или лечения инфекции Borrelia или лаймской болезни у субъекта. В различных аспектах, такие способы включают стадию введения субъекту любой из композиций вакцины, описанных тут, или любой из комбинированных вакцин, описанных тут, в количестве, обеспечивающем эффективную профилактику или лечение инфекции Borrelia или лаймской болезни.
Изобретение включает использование композиций по изобретению для изготовления лекарственных препаратов. Настоящее изобретение также предусматривает другие родственные аспекты.
Изложенная выше сущность изобретения не предназначена для описания всех аспектов изобретения, и дополнительные аспекты описаны в других разделах, таких как приведенное далее детальное описание. Весь документ должен рассматриваться как единое раскрытие изобретения, и следует понимать, что предусматриваются все комбинации признаков, описанных тут, даже если комбинация признаков не встречается вместе в одном и том же предложении, или абзаце, или разделе данного документа. Другие признаки и преимущества изобретения будут понятны из приведенного далее детального описания. Следует понимать, однако, что детальное описание и конкретные примеры, указывающие конкретные варианты воплощения изобретения, приводятся только с целью иллюстрации, потому что различные изменения и модификации сущности и объема изобретения будут понятны квалифицированным специалистам в данной области техники из данного детального описания.
Краткое описание чертежей
Фигура 1 представляет собой общую схему получения липидированных химерных конструктов OspA.
Фигура 2 представляет собой аминокислотную последовательность lipB sOspA 1/2251 (SEQ ID NO: 2).
Фигура 3 изображает нуклеотидную (SEQ ID NO: 1) и расшифрованные аминокислотные последовательности (SEQ ID NO: 2) lipB sOspA 1/2251.
Фигура 4 представляет собой аминокислотную последовательность lipB sOspA 6/4 (SEQ ID NO: 4).
Фигура 5 изображает нуклеотидную (SEQ ID NO: 3) и расшифрованные аминокислотные последовательности (SEQ ID NO: 4) lipB sOspA 6/4.
Фигура 6 представляет собой аминокислотную последовательность lipB sOspA 5/3 (SEQ ID NO: 6).
Фигура 7 изображает нуклеотидную (SEQ ID NO: 5) и расшифрованные аминокислотные последовательности (SEQ ID NO: 6) lipB sOspA 5/3.
Фигура 8 изображает оптимизацию использования кодонов для высокого уровня экспрессии.
Фигура 9 изображает различия между последовательностями липидированных и нелипидированных конструктов.
Фигура 10 представляет собой описание системы экспрессии T7.
Фигура 11 представляет собой результаты анализа способом ДСН-ПААГ, показывающие экспрессию новых рекомбинантных белков OspA индуцированными и неиндуцированными культурами.
Фигура 12 представляет собой карту плазмиды pUC18.
Фигура 13 представляет собой карту плазмиды pET30a.
Фигура 14 изображает стратегию создания фрагмента Kpn I - Bam HI lipB sOspA 5/3.
Фигура 15 изображает выравнивание с указанием аминокислотной замены (SEQ ID NO: 39) в lipB sOspA 1/2251 и праймерные последовательности PCR (SEQ ID NO: 21 и 41), используемые для введения замены (lipB OspA 1/2 mod (SEQ ID NO: 38); консенсусная последовательность (SEQ ID NO: 40)).
Фигура 16 изображает выравнивание OspA-последовательности Blip OspA BPBP/A1 с модифицированной молекулой lipB sOspA 1/2251. Верхняя нить представляет собой исходную последовательность (SEQ ID NO: 42), и нижняя нить представляет собой оптимизированную последовательность (SEQ ID NO: 43). Примечание: три основания (CAT) в начале последовательности не показаны; они образуют часть сайта Nde I CATATG.
Фигура 17 изображает выравнивание OspA-последовательности Blip OspA KT с модифицированной молекулой lipB sOspA 6/4. Верхняя нить представляет собой исходную последовательность (SEQ ID NO: 44), и нижняя нить представляет собой оптимизированную последовательность (SEQ ID NO: 45). Примечание: Одно основание (C) в начале последовательности не показано; оно входит в состав сайта Nde I CATATG.
Фигура 18 изображает выравнивание OspA-последовательности Blip OspA 5/3 с модифицированной молекулой lipB sOspA 5/3. Верхняя нить представляет собой исходную последовательность (SEQ ID NO: 46), и нижняя нить представляет собой оптимизированную последовательность (SEQ ID NO: 47).
Фигура 19 изображает распределение функциональных анти-OspA ответов по результатам проведения анализов на поверхностное связывание антител и ингибирование роста у защищенных и инфицированных животных, иммунизированных 3 нг OspA 1/2 перед контрольным заражением штаммом B. burgdorferi s.s. B31. p-Значения Манна-Уитни продемонстрировали высокозначимые различия по содержанию функциональных антител между защищенными и инфицированными животными.
Фигура 20 изображает распределение функциональных анти-OspA ответов по результатам проведения анализов на поверхностное связывание антител и ингибирование роста у защищенных и инфицированных животных, иммунизированных 3 нг OspA 1/2 перед контрольным заражением природными клещами. p-Значения Манна-Уитни продемонстрировали высокозначимые различия по содержанию функциональных антител между защищенными и инфицированными животными.
Фигура 21 изображает поверхностное связывание (средние интенсивности флуоресценции (MFI)) и ингибирование роста (титры GI-50) в пуле мышиной сыворотки после иммунизации тремя дозами 3-компонентной вакцины химерного OspA. Эффективное поверхностное связывание и ингибирование роста наблюдалось для всех шести штаммов Borrelia, экспрессирующих типы OspA, гомологичные типам OspA в вакцине (типы 1-6).
Фигура 22 изображает титры средней интенсивности флуоресценции (MFI), которые были получены с использованием сывороток дня 42 индивидуальных мышей, иммунизированных комбинациями вакцин rOspA путем проведения анализа на поверхностное связывание (SBA). Результаты показали, что все три компонента rOspA (1/2, 6/4 и 5/3) являются необходимыми в мультивалентной вакцине для индуцирования высоких титров поверхностного связывания IgG-антител против всех 6 штаммов у C3H-мышей. Двухкомпонентные вакцины не покрывают полностью 2 отсутствующие штамма.
Фигура 23 изображает ингибирование роста Borreliae с использованием сыворотки дня 42 индивидуальных мышей (в группах по 10), иммунизированных комбинациями вакцин rOspA. Только мультивалентная вакцина (вакцина, содержащая все три штамма) давала >50% ингибирования роста у >90% животных (n=10). Черные столбики (со сплошной заливкой) указывают штаммы, гомологичные использованной вакцине.
Фигура 24 изображает покрытие Borreliae, экспрессирующих внутритиповые варианты OspA. Поверхностное связывание было классифицировано на сильное (усиление флуоресценции >10-кратного) или более слабое (2-10-кратное усиление флуоресценции).
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение предусматривает химерные молекулы OspA, пригодные для использования в качестве антигенов, которые могут быть введены как иммуногенная композиция или композиция вакцины при лаймской болезни или инфекции Borrelia. Перед детальным пояснением каких-либо вариантов воплощения изобретения укажем, что изобретение не должно рассматриваться как ограниченное в его применении деталями конструкции и конфигурацией компонентов, указанных в приведенном ниже описании, или проиллюстрированных на фигурах и в примерах. Используемые тут заголовки разделов приводятся только в организационных целях и не должны пониматься как ограничивающие описываемый предмет. Все упоминаемые в данной заявке первоисточники явным образом включены сюда в качестве ссылок.
Изобретение охватывает другие варианты воплощения и практикуется или осуществляется различными способами. Также, следует понимать, что фразеология и терминология, используемые тут, предназначены для описания и не должны рассматриваться как ограничивающие. Термины «включающий», «содержащий» или «имеющий» и их варианты следует понимать как охватывающие перечисленные далее позиции и их эквиваленты, а также дополнительные позиции.
Варианты воплощения изобретения проиллюстрированы на примерах конструирования и синтеза трех химерных кодирующих последовательностей OspA, которые кодируют три различные липидированные молекулы OspA, обладающие некоторыми общими признаками. Каждая химерная кодирующая последовательность представляет два серотипа OspA, и химерные кодирующие последовательности были сконструированы таким образом, чтобы они кодировали стабильные химерные молекулы OspA, являющиеся безопасными и сильно иммуногенными, и обеспечивающие защиту субъекта против инфекции B. burgdorferi sensu lato (s.l.).
В одном аспекте, химерные молекулы OspA содержат проксимальный участок от одного серотипа OspA, вместе с дистальным участком от другого серотипа OspA, сохраняя при этом защитные свойства обоих родительских полипептидов. Химерные молекулы нуклеиновой кислоты OspA экспрессировали в Escherichia coli (E. coli) для получения антигенов, которые могли быть введены в комбинированную вакцину для обеспечения защиты против всех шести распространенных серотипов (серотипы 1-6), ассоциированных с лаймской болезнью или инфекцией Borrelia в Европе, и против единственного серотипа OspA, ассоциированного с лаймской болезнью или инфекцией Borrelia в Северной Америке. Поскольку вакцина, содержащая серотипы 1-6, обеспечивает защиту против B. afzelii, B. garinii и B. burgdorferi, вакцина предназначена для глобального применения.
Изобретение также включает приготовление второго набора химерных кодирующих последовательностей OspA которые, в одном аспекте, являются производными от первого набора из трех генов, путем удаления последовательностей нуклеиновых кислот, кодирующих лидерную последовательность, необходимую для продуцирования липидированной молекулы OspA. Два набора конструктов (дающих липидированные и нелипидированные полипептиды) были необходимы для оценки простоты их продуцирования в ферментере (биомасса, стабильность, выход продуктов и т.д.), для определения того, насколько легко может осуществляться очистка разных типов антигена и для сравнения их биологических характеристик (профиль безопасности и эффективность защиты).
Изобретение включает иммуногенные композиции, содержащие химерные молекулы OspA по изобретению. Изобретение аналогично включает вакцины и наборы вакцин, содержащих такие молекулы OspA, способы получения иммуногенных композиций и вакцин и использование иммуногенных композиций и вакцин в медицинской терапии и профилактике заболеваний человека и животных. Изобретение дополнительно включает способы иммунизации против лаймской болезни или инфекции Borrelia с использованием композиций OspA, описанных тут, и использование композициЙ OspA в производстве лекарственных средств для профилактики лаймской болезни или инфекция Borrelia.
Определения
Если не указано иное, то все технические и научные термины, используемые тут, имеют значения, общеизвестные рядовым специалистам в области техники, к которой принадлежит данное изобретение. Следующие источники содержат общие определения многих терминов, используемых в данном изобретении: Singleton, et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY (2d ed. 1994); THE CAMBRIDGE DICTIONARY OF SCIENCE AND TECHNOLOGY (Walker ed., 1988); THE GLOSSARY OF GENETICS, 5TH ED., R. Rieger, et al. (eds.), Springer Verlag (1991); и Hale и Marham, THE HARPER COLLINS DICTIONARY OF BIOLOGY (1991).
В тексте используются следующие аббревиатуры:
Отметим, что, в используемом в данном описании и приложенной формуле изобретения, формы единственного числа (в английском тексте - с артиклями «a», «an» и «the») включают ссылки на множественное число, если из контекста явным образом не следует иное.
В используемом тут значении, следующие термины имеют указанные для них значения, если не будет указано иное.
Термин «ген» относится к ДНК-последовательности, кодирующей последовательность аминокислот, которая содержит все или часть одного или более полипептидов, белков или ферментов, и может включать или не включать интроны и регуляторные ДНК-последовательности, такие как промоторные или энхансерные последовательности, 5'-нетранслируемую область или 3'-нетранслируемую область, которые влияют, например, на условия, в которых ген экспрессируется. В данном описании, ген OspA является бактериальным и потому не содержит интронов. Термин «кодирующая последовательность» относится к ДНК-последовательности, которая кодирует последовательность аминокислот, но не содержит интронов или регуляторных последовательностей. Аналогично, в данном описании кодирующая последовательность OspA не содержит регуляторных последовательностей.
«Нуклеиновая кислота» или «последовательность нуклеиновой кислоты» или «молекула нуклеиновой кислоты» относится к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам в одно- или двухцепочечной форме. Термин охватывает нуклеиновые кислоты, которые содержат известные нуклеотидные аналоги или модифицированные остатки в основной цепи или связи, являются синтетическими, природными и неприродными, обладают способностью к связыванию, близкой к эталонной нуклеиновой кислоте, и подвергаются метаболизму аналогично эталонным нуклеотидам. Примеры таких аналогов включают, без ограничений, фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептидо-нуклеиновые кислоты (PNAs). Термины охватывают молекулы, состоящие из любых известных аналогов оснований ДНК и РНК, таких как, без ограничений, 4-ацетилцитозин, 8-гидрокси-N6-метиладенин, азиридинилцитозин, псевдоизоцитозин, 5-(карбоксигидроксилметил)урацил, 5-фторурацил, 5-бромурацил, 5-карбоксиметиламинометил-2-тиоурацил, 5-карбоксиметиламинометилурацил, дигидроурацил, инозин, N6-изопентениладенин, 1-метиладенин, 1-метилпсевдоурацил, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-метиладенин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеозин, 5'-метоксикарбонилметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, урацил-5-оксиуксусной кислоты сложный метиловый эфир, урацил-5-оксиуксусная кислота, оксибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, N-урацил-5-оксиуксусной кислоты сложный метиловый эфир, урацил-5-оксиуксусная кислота, псевдоурацил, квеозин, 2-тиоцитозин и 2,6-диаминопурин.
Если не указано иное, конкретная последовательность нуклеиновой кислоты также охватывает по умолчанию ее консервативно модифицированные варианты (например, замещения вырожденного кодона) и комплементарные последовательности, а также явным образом указанную последовательность. Конкретнее, замещения вырожденного кодона, в некоторых аспектах, получают путем генерирования последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов замещено остатками смешанных оснований и/или дезоксиинозина (Batzer et al., Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)). Термин «нуклеиновая кислота» используется взаимозаменяемо с геном, кДНК, мРНК, олигонуклеотидом и полинуклеотидом.
Термины «полипептид», «пептид» и «белок» используются в данном описании взаимозаменяемо по отношению к полимеру, состоящему из аминокислотных остатков, соединенных пептидными связями. Термины относятся к аминокислотным полимерам, в которых один или более аминокислотных остатков представляют собой искусственный химический миметик соответствующей природной аминокислоты, а также к природным аминокислотным полимерам и неприродным аминокислотным полимерам. Термин «белок» типично относится к крупным полипептидам. Термин «пептид» типично относится к коротким полипептидам. Синтетические полипептиды могут быть синтезированы, например, с помощью автоматического полипептидного синтезатора.
Термин «молекула OspA» или «химерная молекула OspA» относится, в одном аспекте, к «нуклеиновой кислоте OspA», содержащей нуклеотидную последовательность SEQ ID NO: 1 (lipB sOspA 1/2251), SEQ ID NO: 3 (lipB sOspA 6/4), SEQ ID NO: 5 (lipB sOspA 5/3), SEQ ID NO: 7 (sOspA 1/2251), SEQ ID NO: 9 (sOspA 6/4), SEQ ID NO: 11 (sOspA 5/3), SEQ ID NO: 168 (orig sOspA 1/2), SEQ ID NO: 170 (orig sOspA 6/4), или SEQ ID NO: 172 (orig sOspA 5/3), или, в другом аспекте, к «полипептиду OspA», содержащему аминокислотную последовательность SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3).
Термин «молекула lipB sOspA» относится, в одном аспекте, к «нуклеиновой кислоте OspA», содержащей нуклеотидную последовательность SEQ ID NO: 1 (lipB sOspA 1/2251), SEQ ID NO: 3 (lipB sOspA 6/4), или SEQ ID NO: 5 (lipB sOspA 5/3) или, в другом аспекте, к «полипептиду OspA», содержащему аминокислотную последовательность SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), или SEQ ID NO: 6 (lipB sOspA 5/3). Последовательности нуклеиновой кислоты SEQ ID NO: 7, 9 и 11 не содержат последовательности нуклеиновой кислоты, кодирующей лидерную последовательность lipB (MRLLIGFALALALIG (SEQ ID NO: 13). Кроме того, последовательности нуклеиновой кислоты SEQ ID NO: 7, 9 и 11 кодируют остаток метионина на амино-конце SEQ ID NO: 8, 10 и 12 вместо цистеинового остатка, присутствующего на карбокси-конце лидерной последовательности lipB в SEQ ID NO: 2, 4 и 6.
Термин «молекула orig sOspA» или «исходная молекула sOspA» относится, в одном аспекте, к «нуклеиновой кислоте OspA», содержащей нуклеотидную последовательность SEQ ID NO: 168 (orig sOspA 1/2), SEQ ID NO: 170 (orig sOspA 6/4), или SEQ ID NO: 172 (orig sOspA 5/3) или, в другом аспекте, к «полипептиду OspA», содержащему аминокислотную последовательность SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3). Такие «исходные» молекулы представляют собой химерные конструкты без мутаций и без оптимизации кодонов.
Изобретение включает химерные молекулы «липидированного OspA» и «нелипидированного OspA». В различных аспектах, липидирование придает OspA свойства адъюванта. В некоторых аспектах изобретения, липидированные молекулы OspA содержат лидерную последовательность OspB. В некоторых аспектах изобретения, лидерная последовательность OspB содержит аминокислоты MRLLIGFALALALIG (SEQ ID NO: 13). В других аспектах, лидерная последовательность OspB содержит другие аминокислоты.
Термины «идентичный» или процент «идентичности», как известно специалистам в данной области техники, относятся к соотношению между последовательностями двух или более полипептидных молекул или двух или более молекул нуклеиновой кислоты, определенному путем сравнения последовательностей. В данной области техники «идентичность» также обозначает степень родства последовательности между молекулами нуклеиновой кислоты или полипептидами, в зависимости от обстоятельств, определяемую путем попарного сравнения цепей двух или более нуклеотидов или двух или более аминокислотных последовательностей. «Идентичность» определяет процент идентичных совпадений между меньшей из двух или более последовательностей с выравниваниями пропусков (если они есть), обрабатываемыми с помощью определенной математической модели или компьютерной программы (т.е. «алгоритмов»). «Существенная идентичность» относится к последовательности с по меньшей мере примерно 70%, примерно 71%, примерно 72%, примерно 73%, примерно 74%, примерно 75%, примерно 76%, примерно 77%, примерно 78%, примерно 79%, примерно 80%, примерно 81%, примерно 82%, примерно 83%, примерно 84%, примерно 85%, примерно 86%, примерно 87%, примерно 88%, примерно 89%, примерно 90%, примерно 91%, примерно 92%, примерно 93%, примерно 94%, примерно 95%, примерно 96%, примерно 97%, примерно 98%, или примерно 99% идентичности последовательностей по длине указанной последовательности. В некоторых аспектах наблюдается идентичность на участке, состоящем из по меньшей мере примерно 50-100 аминокислот или нуклеотидов в длину. В других аспектах, наблюдается идентичность на участке, состоящем из по меньшей мере примерно 100-200 аминокислот или нуклеотидов в длину. В других аспектах, наблюдается идентичность на участке, состоящем из по меньшей мере примерно 200-500 аминокислот или нуклеотидов в длину. В определенных аспектах, процент идентичности последовательностей определяется с использованием компьютерной программы, выбранной из группы, состоящей из GAP, BLASTP, BLASTN, FASTA, BLASTA, BLASTX, BestFit и алгоритма Смита-Уотермана.
Следует также определенно понимать, что любое численное значение, указанное тут, включает все значения от наименьшего значения до наибольшего значения, т.е. все возможные комбинации численных значений между наименьшим и наибольшим указанными значениями должны рассматриваться как явным образом указанные в данной заявке. Например, если диапазон концентраций указан как составляющий от примерно 1% до 50%, то следует понимать, что такие значения, как от 2% до 40%, от 10% до 30%, или от 1% до 3% и т.д., считаются явным образом приведенными в данном описании изобретения. Перечисленные выше значения являются только примерами конкретного намерения.
Интервалы значений, в различных аспектах, выражены тут как от «примерно» или «приблизительно» одного конкретного значения и/или до «примерно» или «приблизительно» другого конкретного значения. В тех случаях, когда значения выражены приблизительно, с использованием местоимения «примерно», следует понимать, что такой интервал включает некоторую величину изменчивости.
Термин «подобие» является родственным понятием, но, в отличие от «идентичности», относится к мере подобия, которая включает как идентичные совпадения, так и совпадающие консервативные замещения. Если две полипептидные последовательности содержат, например, 10/20 идентичных аминокислот, и все остальные являются неконсервативными замещениями, то проценты идентичности и подобия оба будут составлять 50%. Если, в этом же примере, еще в пяти положениях находятся консервативные замещения, то процент идентичности остается равным 50%, но процент подобия будет равен 75% (15/20). Таким образом, в тех случаях, где присутствуют консервативные замещения, степень процента подобия между двумя полипептидами будет выше, чем процент идентичности между этими двумя полипептидами.
Термин «изолированная молекула нуклеиновой кислоты» относится к молекуле нуклеиновой кислоты по изобретению, которая (1) была отделена в любой степени от белков, липидов, углеводов или других материалов, с которыми она в естественных условиях объединена при выделении цельной ДНК из клеток-источников, (2) не связана со всем или частью полинуклеотида, с которым «изолированная молекула нуклеиновой кислоты» связана в природе, (3) является функционально связанной с полинуклеотидом, с которым она не связана в природе, или (4) не встречается в природе как часть большей по размеру полинуклеотидной последовательности. «По существу не содержащий», в используемом тут значении, указывает, что молекула нуклеиновой кислоты не содержит каких-либо других загрязняющих молекул нуклеиновой кислоты (кислот) или других загрязнений, присутствующих в его природном окружении, которые мешали бы его использованию при продуцировании полипептида или его терапевтическому, диагностическому, профилактическому или исследовательскому использованию.
Термин «изолированный полипептид» относится к полипептиду по настоящему изобретению, который (1) был отделен в любой степени от полинуклеотидов, липидов, углеводов или других материалов, с которыми он в естественных условиях объединен при выделении из клетки-источника, (2) не связан (ковалентными или нековалентными взаимодействиями) со всем или частью полипептида, с которым «изолированный полипептид» связан в природе, (3) является функционально связанным (ковалентными или нековалентными взаимодействиями) с полипептидом, с которым он не связан в природе, или (4) не встречается в природе. В одном аспекте, изолированный полипептид по существу не содержит каких-либо других загрязняющих полипептидов или других загрязнений, присутствующих в его природном окружении, которые мешали бы его терапевтическому, диагностическому, профилактическому или исследовательскому использованию.
В используемом тут значении «фрагмент» полипептида относится к любой части полипептида, меньшей полноразмерного полипептидного или белкового продукта экспрессии. Фрагменты типично являются имеющими делеции аналогами полноразмерного полипептида, в которых удалены один или более аминокислотных остатков с амино-конца и/или карбокси-конца полноразмерного полипептида. Соответственно, «фрагменты» представляют собой подмножество делеционных аналогов, описанных ниже.
В используемом тут значении, «аналог» относится к полипептиду по существу сходной структуры и обладающему такой же биологической активностью, хотя, в определенных случаях и в разной степени, к встречающейся в природных условиях молекуле. Аналоги отличаются по составу их аминокислотных последовательностей от встречающегося в природных условиях полипептида, из которого был получен аналог с помощью одной или более мутаций, включая (i) делецию одного или более аминокислотных остатков на одном или более концов полипептида (включающего описанные выше фрагменты) и/или одного или более внутренних участков встречающейся в природных условиях полипептидной последовательности, (ii) инсерцию или прибавление одной или более аминокислот на одном или более концов (типично, аналог «прибавления») полипептида и/или в одном или более внутренних участков (типично аналог «инсерции») встречающейся в природных условиях полипептидной последовательности, или (iii) замещение одной или более аминокислот на другие аминокислоты во встречающейся в природных условиях полипептидной последовательности. Замещения считаются консервативными или неконсервативными на основании физико-химического или функционального родства заменяемой аминокислоты и замещающей ее аминокислоты.
«Консервативно модифицированные аналоги» относится как к аминокислотным, так и к нуклеиново-кислотным последовательностям. Что касается конкретных последовательностей нуклеиновой кислоты, консервативно модифицированные нуклеиновые кислоты относятся к нуклеиновым кислотам, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или, в случае нуклеиновых кислот, не кодирующих аминокислотную последовательность, к по существу идентичным последовательностям. Вследствие вырожденности генетического кода, большое число функционально идентичных нуклеиновых кислот кодирует любой определенный белок. Например, кодоны GCA, GCC, GCG и GCU все кодируют аминокислоту аланин. Таким образом, в каждом положении, для которого кодон соответствует аланину, этот кодон может быть заменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие варианты нуклеиновой кислоты представляют собой «молчащие варианты», являющиеся одной из разновидностей консервативно модифицированных аналогов. Каждая последовательность нуклеиновой кислоты по настоящему изобретению, кодирующая полипептид, также описывает все возможные молчащие варианты нуклеиновой кислоты. Специалисту будет понятно, что каждый кодон нуклеиновой кислоты (кроме AUG, который обычно является единственным кодоном, соответствующим метионину, и TGG, который обычно является единственным кодоном триптофана) может быть модифицирован с образованием функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, кодирующей полипептид, неявным образом включен в каждую описанную последовательность.
Что касается аминокислотных последовательностей, специалисту будет понятно, что индивидуальные замещения, инсерции, делеции, аддиции или усечения нуклеиново-кислотной, пептидной, полипептидной или белковой последовательности, которые изменяют, прибавляют или удаляют отдельную аминокислоту или небольшой процент аминокислот кодированной последовательности, представляют собой «консервативно модифицированный аналог», в котором изменение приводит к замещению аминокислоты на химически подобную аминокислоту. Таблицы консервативных замещений, обеспечивающих введение функционально подобных аминокислот, хорошо известны специалистам. Такие консервативно модифицированные варианты рассматриваются в дополнение к и не исключают полиморфные варианты, межвидовые гомологи и аллели по изобретению.
Каждая из следующих восьми групп содержит аминокислоты, являющиеся консервативными замещениями друг друга:
1) Аланин (A), Глицин (G);
2) Аспарагиновая кислота (D), Глутаминовая кислота (E);
3) Аспарагин (N), Глутамин (Q);
4) Аргинин (R), Лизин (K);
5) Изолейцин (I), Лейцин (L), Метионин (M), Валин (V);
6) Фенилаланин (F), Тирозин (Y), Триптофан (W);
7) Серин (S), Треонин (T); и
8) Цистеин (C), Метионин (M) (см., например, Creighton, Proteins (1984)).
В используемом тут значении «вариант» относится к полипептиду, белку или его аналогу, который содержит по меньшей мере одно аминокислотное замещение, делецию, инсерцию или модификацию, при условии, что вариант сохраняет биологическую активность нативного полипептида.
В используемом тут значении, «аллельный вариант» относится к любым двум или более полиморфным формам гена, занимающим один и тот же генетический локус. Аллельные варианты возникают естественным путем вследствие мутаций и, в некоторых аспектах, приводят к фенотипическому полиморфизму в популяциях. В определенных аспектах, генные мутации являются молчащими (без изменений в кодируемом полипептиде) или, в других аспектах, кодируют полипептиды, имеющие измененные аминокислотные последовательности. «Аллельные варианты» также относится к кДНК, полученной из транскриптов мРНК генетических аллельных вариантов, а также кодируемым ими белкам.
Термин «производный» относится к полипептидам, ковалентно модифицированным путем конъюгации с терапевтическими или диагностическими агентами, мечения (например, радионуклидами или различными ферментами), ковалентного присоединения полимера, такого как пегилирование (дериватизация полиэтиленгликолем) и инсерции или замещения способами химического синтеза неприродных аминокислот. В некоторых аспектах, производные модифицируют таким образом, чтобы они содержали дополнительные химические фрагменты, нормально не входящие в состав молекулы. Такие фрагменты, в различных аспектах, модулируют растворимость молекулы, абсорбцию и/или биологический период полувыведения. Фрагменты, в различных других аспектах, альтернативно снижают токсичность молекулы и устраняют или ослабляют любые нежелательные побочные эффекты молекулы и т.д. Фрагменты, способные медиировать такие эффекты, раскрыты в Remington's Pharmaceutical Sciences (1980). Процедуры присоединения таких фрагментов к молекуле хорошо известны специалистам. Например, в некоторых аспектах, производное OspA представляет собой молекулу OspA, имеющую химическую модификацию, придающую белку более длительный период полувыведения in vivo. В одном варианте воплощения полипептиды модифицируют путем прибавления водорастворимого полимера, известного специалистам. В родственном варианте воплощения полипептиды модифицируют путем гликозилирования, пегилирования и/или полисиалилирования.
Термин «рекомбинантный», при использовании по отношению к, например, клетке, или нуклеиновой кислоте, белку или вектору, указывает, что клетка, нуклеиновая кислота, белок или вектор, были модифицированы путем введения гетерологичной нуклеиновой кислоты или белка или изменения нативной нуклеиновой кислоты или белка, или что клетка является потомком модифицированной таким образом клетки. Таким образом, например, рекомбинантные клетки экспрессируют гены, не присутствующие в нативной (не-рекомбинантной) форме клетки, или экспрессируют нативные гены, которые иначе аномально экспрессируются, экспрессируются на низком уровне или совершенно не экспрессируются.
В используемом тут значении «селектируемый маркер» относится к гену, кодирующему фермент или другой белок, который придает клетке или организму, в которых он экспрессируется, идентифицируемое фенотипическое изменение, такое как резистентность к лекарственному средству, антибиотику или другому агенту, так чтобы экспрессия или активность маркера селектировались по наличию признака (например, но без ограничений, позитивный маркер, такой как ген neo) или его отсутствию (например, и без ограничений, негативный маркер, такой как ген дифтерии). «Гетерологичный селектируемый маркер» относится к гену селектируемого маркера, который был введен в геном животного, у которого он в естественных условиях отсутствует.
Примеры селектируемых маркеров включают, без ограничений, ген резистентности к антибиотику, такому как неомицин (neo), пуромицин (Puro), дифтерийный токсин, фосфотрансферазу, гигромицин фосфотрансферазу, ксантингуанин-фосфорибозилтрансферазу, тимидинкиназу вируса простого герпеса типа 1, аденин-фосфорибозилтрансферазу и гипоксантин-фосфорибозилтрансферазу. Рядовому специалисту в данной области техники будет понятно, что любой селектируемый маркер, известный специалистам, является пригодным для использования в способах, описанных тут.
Термин «гетерологичный», при использовании по отношению к частям нуклеиновой кислоты, указывает, что нуклеиновая кислота содержит две или более субпоследовательностей, которые не встречаются в природе в такой же взаимосвязи друг с другом. Например, типично рекомбинантно продуцируют нуклеиновую кислоту, содержащую две или более последовательностей из неродственных генов, расположенных таким образом, чтобы получить новую функциональную нуклеиновую кислоту, например, промотор из одного источника и кодирующую область из другого источника. Аналогично, гетерологичный белок указывает, что белок содержит две или более субпоследовательностей, которые не встречаются в природе в такой же взаимосвязи друг с другом (например, слитый белок).
В используемом тут значении, термин «гомологичный» относится к взаимосвязи между белками, имеющими «общее эволюционное происхождение», включая белки из надсемейств (например, надсемейство иммуноглобулинов) и гомологичные белки от разных биологических видов (например, легкая цепь миозина и т.д.) (Reeck et al., Cell 50:667, 1987). Такие белки (и кодирующие их гены) обладают гомологией последовательностей, проявляющейся в подобии их последовательностей, по показателю процента подобия или в присутствии определенных остатков или мотивов в консервативных положениях.
Оптимальное выравнивание последовательностей для сравнения проводится, например, и без ограничений, по алгоритму местной гомологии Smith et al., Adv. Appl. Math. 2:482, 1981; по алгоритму выравнивания областей гомологии Needleman et al., J. Mol. Biol. 48:443, 1970; по способу поиска подобия Pearson et al., Proc. Natl. Acad. Sci. USA 85:2444, 1988; способами компьютерной реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Madison, WI), или путем визуального сравнения (см. в общем Ausubel et al., supra). Другим примером алгоритма, пригодного для определения процента идентичности последовательностей и подобия последовательностей, является алгоритм BLAST, описанный Altschul et al., J. Mol. Biol. 215:403-410, 1990. Программное обеспечение для проведения анализов BLAST является общедоступным и распространяется Национальным центром биотехнологической информации (National Center for Biotechnology Information). В дополнение к расчету процента идентичности последовательностей, алгоритм BLAST также выполняет статистический анализ подобия между двумя последовательностями (см., например, Karlin et al., Proc. Natl. Acad. Sci. USA 90:5873-5787, 1993).
Термин «вектор» используется по отношению к любой молекуле (например, нуклеиновой кислоте, плазмиде или вирусу), используемой для переноса кодирующей информации в клетку-хозяина.
«Клонирующий вектор» представляет собой маленький кусочек ДНК, в который может быть вставлен фрагмент чужеродной ДНК. Инсерция фрагмента в клонирующий вектор осуществляется путем обработки носителя и чужеродной ДНК одним и тем же рестрикционным ферментом, а затем лигированием фрагментов друг с другом. Существует большое количество типов клонирующих векторов, и все типы клонирующих векторов используются в изобретении. Генно-инженерные плазмиды и бактериофаги (такие как фаг λ), возможно, чаще всего используются для этих целей. Другие типы клонирующих векторов включают бактериальные искусственные хромосомы (BACs) и искусственные хромосомы дрожжей (YACs).
«Экспрессионный вектор» представляет собой конструкт нуклеиновой кислоты, полученный рекомбинантно или способами синтеза, с рядом определенных элементов нуклеиновой кислоты, обеспечивающих транскрипцию конкретной нуклеиновой кислоты в клетке-хозяине. Экспрессионный вектор может быть частью плазмиды, вируса или фрагмента нуклеиновой кислоты. В определенных аспектах, экспрессионный вектор включает функционально связанную с промотором нуклеиновую кислоту, которая должна транскрибироваться.
Термин «кодирующая последовательность» определяется тут как последовательность нуклеиновой кислоты, которая транскрибируется в мРНК, которая транслируется в полипептид при помещении под контроль соответствующих контрольных последовательностей. Границы кодирующей последовательности в общем определяются старт-кодоном ATG, который обычно является началом открытой рамки считывания на 5′-конце мРНК, и последовательностью терминации транскрипции, расположенной непосредственно за открытой рамкой считывания на 3′-конце мРНК. Кодирующая последовательность может включать, без ограничений, геномную ДНК, кДНК, полусинтетические, синтетические и рекомбинантные последовательности нуклеиновой кислоты. В одном аспекте, промоторная ДНК-последовательность определяется как ДНК-последовательность, расположенная перед ассоциированной с ней кодирующей последовательностью и способная контролировать экспрессию этой кодирующей последовательности.
«Промотор» определяется как совокупность контрольных последовательностей нуклеиновой кислоты, которые направляют транскрипцию нуклеиновой кислоты. В используемом тут значении, промотор включает необходимые последовательности нуклеиновой кислоты вблизи сайта начала транскрипции, такие как, в случае промотора полимеразы II типа, элемент TATA. Промотор также необязательно включает дистальные энхансерные или репрессорные элементы, которые могут быть расположены на расстоянии нескольких тысяч пар оснований от сайта начала транскрипции. «Конститутивный» промотор представляет собой промотор, активный при большинстве условий окружающей среды и развития. «Индуцируемый» промотор представляет собой промотор, активный при регуляции условий окружающей среды или эволюционной регуляции.
Термин «функционально связанный» относится к функциональной связи между контрольными последовательностями экспрессии нуклеиновой кислоты (такими как промотор, или совокупность сайтов связывания фактора транскрипции) и второй последовательностью нуклеиновой кислоты, где контрольная последовательность экспрессии направляет транскрипцию нуклеиновой кислоты, соответствующей второй последовательности.
Термин «трансдукция» используется для обозначения переноса нуклеиновых кислот от одной бактерии к другой, обычно с помощью фага. «Трансдукция» также относится к поглощению и переносу эукариотических клеточных последовательностей ретровирусами.
Термин «трансфекция» используется для обозначения поглощения чужеродной или экзогеннй ДНК клеткой, и клетка «трансфицирована», если экзогенная ДНК была введена внутрь клеточной мембраны. Ряд способов трансфекции хорошо известен специалистам в данной области техники и раскрыт в данном описании. См., например, Graham et al., Virology, 52:456 (1973); Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratories, New York, (1989); Davis et al., Basic Methods in Molecular Biology, Elsevier, (1986); и Chu et al., Gene, 13:197 (1981). Такие методики могут быть использованы для введения одного или более фрагментов экзогенной ДНК в пригодные клетки-хозяева.
Термин «трансформация», в используемом тут значении, относится к изменению генетических характеристик клетки и клетка считается трансформированной, если она была модифицирована путем введения в нее новой ДНК. Например, клетка является трансформированной, если она была генетически модифицирована по сравнению с ее нативным состоянием. После трансфекции или трансдукции, трансформирующая ДНК может рекомбинироваться с ДНК клетки путем физической интеграции в хромосому клетки. В некоторых случаях, ДНК временно поддерживается как эписомальный элемент без репликации, или независимо реплицируется в виде плазмиды. Клетка считается стабильно трансформированной, если ДНК реплицируется при делении клетки.
Термин «эндогенный» относится к полипептиду или полинуклеотиду или другому соединению, которое экспрессируется в естественных условиях в организме-хозяине, или возникает в клетке, ткани или организме. «Экзогенный» относится к полипептиду, полинуклеотиду или другому соединению, которое возникает вне клетки, ткани или организма.
Термин «агент» или «соединение» описывает любую молекулу, например, белковую или фармацевтическую, обладающую способностью влиять на биологический параметр по изобретению.
«Контроль», в используемом тут значении, может обозначать активный, позитивный, негативный контроль или контрольный эксперимент с растворителем. Квалифицированным специалистам в данной области техники будет понятно, что контроли используются для установления релевантности экспериментальных результатов и обеспечения сравнения для исследуемого состояния.
Термин «уменьшает тяжесть», по отношению к симптому лайм-боррелиоза или лаймской болезни, означает, что симптом имеет задержку проявления, пониженную тяжесть, или причиняет субъекту меньший вред. В общем, тяжесть симптома сравнивают с контролем, например, не получающим активной профилактической или терапевтической композиции. В этом случае можно сказать, что композиция снижает тяжесть симптома лайм-боррелиоза, если симптом уменьшается на 10%, 25%, 30%, 50%, 80%, или 100% (т.е. по существу исчезает), по сравнению с контрольным уровнем симптома.
Термин «антиген» относится к молекуле или части молекулы, способным связываться селективным связывающим агентом, таким как антитело, и дополнительно пригодным для использования у субъекта для продуцирования антител, способных связываться с эпитопом каждого антигена. Антиген, в различных аспектах, имеет один или более эпитопов.
Термин «антитело» относится к молекуле или молекулам, обладающим специфичностью к полипептиду OspA. В используемом тут значении, термины «специфический», «специфичность» и «специфически связывается с» относятся к способности антитела связываться с полипептидами OspA и не связываться с не-OspA полипептидами. В определенных аспектах, антитело является «нейтрализующим антителом», если антитело реагирует с инфекционным агентом и разрушает или ингибирует его инфекционность или вирулентность. Изобретение включает иммуногенные композиции, содержащие антитела, которые «нейтрализуют» Borrelia.
Термины «фармацевтически приемлемый носитель» или «физиологически приемлемый носитель», в используемом тут значении, относятся к одному или более материалов композиции, пригодных для обеспечения или увеличения доставки полипептида OspA, молекулы нуклеиновой кислоты OspA или антитела OspA в фармацевтической композиции.
Термин «стабилизатор» относится к веществу или эксципиенту вакцины, который защищает иммуногенную композицию вакцины от неблагоприятных условий, таких как условия, возникающие при нагреве или замораживании, и/или пролонгирует стабильность или срок годности при хранении иммуногенной композиции в стабильном и иммуногенном состоянии или статусе. Примеры стабилизаторов включают, без ограничений, сахара, такие как сахароза, лактоза и манноза; сахароспирты, такие как маннит; аминокислоты, такие как глицин или глутаминовая кислота; и белки, такие как человеческий сывороточный альбумин или желатин.
Термин «антимикробный консервант» относится к любому веществу, прибавляемому к иммуногенной композиции или вакцине, ингибирующей рост микроорганизмов, которая может быть введена путем многоразового отбора доз из мультидозных флаконов, в случае использования таких контейнеров. Примеры антимикробных консервантов включают, без ограничений, такие вещества как тимеросал, 2-феноксиэтанол, бензетония хлорид и фенол.
Термин «иммуногенная композиция» относится к композиции, содержащей антиген (например, химерные молекулы OspA), к которому вырабатываются антиген-специфические антитела, адъювант, стимулирующий иммунный ответ субъекта-хозяина, и пригодный иммунологически инертный, фармацевтически приемлемый носитель. Необязательно, иммуногенная композиция содержит один или более стабилизаторов. Необязательно, иммуногенная композиция содержит один или более антимикробных консервантов.
Термины «вакцина» или «композиция вакцины» относятся к биологическому препарату, который повышает иммунитет к определенной болезни (например, лаймской болезни или инфекции Borrelia). Вакцина типично содержит агент, напоминающий микроорганизм, вызывающий болезнь (например, химерные молекулы OspA (антиген) Borrelia). Агент стимулирует иммунную систему организма распознавать агент как чужеродный, уничтожать его и «запоминать» его, так, чтобы иммунная система могла легче распознавать и уничтожать любые такие микроорганизмы, с которыми она позднее столкнется. Вакцины, в различных аспектах, являются профилактическими (предотвращают или ослабляют эффекты будущей инфекции любым природным или «диким» патогеном) или терапевтическими (вакцины против присутствующей инфекции). Как было указано выше, такие композиции вакцины включают рецептуры композиций, содержащие фармацевтически приемлемые носители. Необязательно, вакцина также содержит один или более стабилизаторов и/или один или более антимикробных консервантов.
Каждый из терминов «эффективное количество» и «терапевтически эффективное количество» относится к используемому количеству молекул нуклеиновой кислоты, полипептида, композиции, или антитела для поддержания наблюдаемого уровня одной или более биологических активностей полипептидов OspA, как описано тут. Например, эффективное количество, в некоторых аспектах изобретения, будет количеством, необходимым для предотвращения, нейтрализации или ослабления инфекции Borrelia.
Термин «комбинация» относится к двум или более молекулам нуклеиновой кислоты по изобретению, или двум или более полипептидам по изобретению. В некоторых аспектах, комбинации молекул по изобретению вводятся для обеспечения иммунитета или борьбы с инфекцией по меньшей мере четырьмя из шести серотипов (1-6) Borrelia, описанных тут. В различных аспектах, используются комбинации двух или трех молекул или полипептидов по изобретению. В определенных аспектах, комбинации молекул по изобретению вводятся субъекту для обеспечения иммунитета от всех шести серотипов (1-6) Borrelia, описанных тут. Было показано, что последняя комбинация обеспечивает иммунитет к гетерологичным штаммам Borrelia, экспрессирующим типы OspA, не присутствующие в комбинации молекул нуклеиновой кислоты или полипептидов.
Термин «комбинированная вакцина» относится к рецептуре композиции вакцины, содержащей более одной композиции вакцины или более одного защитного антигена против одной или более болезней. Изобретение включает комбинированную вакцину, содержащую химерные антигены OspA против лаймской болезни или Borrelia в дополнение к антигену против одной или нескольких других болезней. В различных аспектах, одна или более других болезней представляет собой переносимую клещами болезнь. В определенных аспектах, другой переносимой клещами болезнью является пятнистая лихорадка Скалистых гор, бабезиоз, возвратная лихорадка, колорадская клещевая лихорадка, моноцитарный эрлихиоз человека (HME), гранулоцитарный эрлихиоз человека (HGE), клещевая сыпная лихорадка STARI, туляремия, клещевой паралич, энцефалит Повассан, ку-лихорадка, конго-крымская геморрагическая лихорадка, цитауксзооноз, марсельская лихорадка или клещевой энцефалит. В конкретных аспектах, изобретение включает комбинированную вакцину, которая содержит одну или более вакцин, включая вакцину клещевого энцефалита, вакцину японского энцефалита и вакцину пятнистой лихорадки Скалистых гор. В некоторых аспектах, комбинированная вакцина содержит композиции вакцин, имеющих сезонный график иммунизации, совместимый с иммунизацией инфекции Borrelia или лаймской болезни. В более конкретных аспектах, комбинированные вакцины пригодны для использования при профилактике множества болезней в географических регионах, где распространены такие болезни.
Термин «Borrelia» относится к видам грамотрицательных бактерий рода Borrelia класса спирохет. В одном аспекте, «Borrelia burgdorferi sensu lato (s.l.)» относится к Borrelia burgdorferi в широком понимании. Почти все случаи лаймской болезни или боррелиоза вызываются одним из трех геновидов - Borrelia afzelii, Borrelia garinii и Borrelia burgdorferi sensu stricto (s.s.), который относится к B. burgdorferi в более строгом понимании). Серотипы OspA Borrelia коррелируют с видами; серотип 1 соответствует B. burgdorferi s.s., серотип 2 соответствует B. afzelii и серотипы 3-7 соответствуют B. garinii. В различных аспектах, иммуногенные композиции или композиции вакцин по изобретению также обеспечивают защиту против других видов Borrelia, включая, без ограничений, Borrelia japonica, Borrelia andersonii, Borrelia bissettii, Borrelia sinica, Borrelia turdi, Borrelia tanukii, Borrelia valaisiana, Borrelia lusitaniae, Borrelia spielmanii, Borrelia miyamotoi или Borrelia lonestar.
«Субъект» рассматривается в его общепринятом значении не-растительного, не-простейшего живого существа. В большинстве аспектов, субъект является животным. В конкретных аспектах, животное является млекопитающим. В более конкретных аспектах, млекопитающее является человеком. В других аспектах, млекопитающее представляет собой комнатное животное или животное-компаньона, одомашненное сельскохозяйственное животное, или животное в зоопарке. В определенных аспектах, млекопитающее представляет собой кошку, собаку, лошадь или корову. В различных других аспектах, млекопитающее представляет собой оленя, мышь, бурундука, белку, опоссума или енота.
Лаймская болезнь (боррелиоз или лайм-боррелиоз)
В некоторых аспектах, изобретение включает химерные молекулы OspA и композиции, содержащие такие молекулы, для использования при профилактике лаймской болезни или инфекции Borrelia. Лаймская болезнь также известна специалистам как боррелиоз или лайм-боррелиоз и, поэтому все эти термины включены в изобретение. Аналогично, изобретение включает способы профилактики или лечения лаймской болезни, включающие введение химерных молекул OspA, описанных тут. Лаймская болезнь, или боррелиоз, представляет собой инфекционную болезнь, вызываемую по меньшей мере тремя видами грамнегативных спирохетозных бактерий, принадлежащих к роду Borrelia. Было открыто по меньшей мере 13 видов Borrelia, из которых три определенно связаны с лайм-боррелиозом. Виды Borrelia, вызывающие лаймскую болезнь, коллективно известны как Borrelia burgdorferi sensu lato, и демонстрируют значительное генетическое разнообразие. Группа Borrelia burgdorferi sensu lato состоит из трех близкородственных видов, которые, вероятно, отвечают за подавляющее большинство случаев заболевания. Borrelia burgdorferi sensu stricto является основной причиной лаймской болезни в США (но также присутствует в Европе), в то время как Borrelia afzelii и Borrelia garinii вызывают большинство европейских случаев болезни. Некоторые исследования также позволяют предположить, что виды Borrelia (например, Borrelia bissettii, Borellia spielmanii, Borrellia lusitaniae и Borrelia valaisiana) могут иногда инфицировать людей. Хотя эти виды, по-видимому, не являются важными факторами болезни, иммуногенная защита против этих видов также включена в изобретение.
Лаймская болезнь является наиболее распространенной переносимой клещами болезнью в Северном полушарии. Название болезни происходит от поселка Лайм (Lyme), штат Коннектикут, где в 1975 г. был идентифицирован ряд случаев заболевания. Borrelia передается людям при укусе инфицированных клещей, принадлежащих к нескольким видам рода Ixodes («твердые клещи»). Ранние симптомы, в некоторых случаях, включают лихорадку, головную боль, утомление, депрессию и характерную круговую кожную сыпь, называемую мигрирующей эритемой. Без лечения, более поздние симптомы часто могут охватывать суставы, сердце и центральную нервную систему. В большинстве случаев, инфекция и ее симптомы устраняются антибиотиками, особенно при лечении болезни на ранней стадии. Однако позднее, запаздывающее или недостаточное лечение может привести к более серьезным симптомам, которые могут приводить к потере трудоспособности и затруднению лечения. В некоторых случаях симптомы, такие как артрит, сохраняются после устранения инфекции антибиотиками.
Некоторые группы исследователей утверждали, что «хроническая» лаймская болезнь является ответственной за ряд медицински необъяснимых симптомов, помимо общепризнанных симптомов поздней стадии лаймской болезни, и что требуется дополнительное длительное лечение антибиотиками. Однако длительное лечение является спорным, и конфликт, связанный с таким лечением, привел к судебному иску по поводу рекомендаций по лечению.
Лаймская болезнь классифицируется как зооноз, поскольку она передается человеку из естественного резервуара, включающего грызунов и птиц, клещами, которые кормятся на обеих группах хозяев. Твердые клещи рода Ixodes являются основными переносчиками лаймской болезни. Большинство случаев инфекции человека вызываются клещами на нимфальной стадии, потому что нимфальные клещи имеют очень маленькие размеры и могут питаться в течение длительных периодов времени незамеченными. Укусы клещей часто остаются незамеченными из-за малых размеров клеща на нимфальной стадии, а также секрета клеща, который предотвращает у хозяина ощущения зуда или боли от укуса.
Лаймская болезнь клинически диагностируется по симптомам, объективным физикальным показателям (таким как мигрирующая эритема, поражение лицевого нерва или артрит), истории возможного контакта с инфицированными клещами, а также серологическим анализам крови. Приблизительно у половины пациентов с лаймской болезнью будет развиваться характерная концентрическая сыпь, но многие не могут вспомнить укуса клеща. Лабораторные анализы не рекомендуются для особ, не имеющих симптомов лаймской болезни.
Из-за сложности культивации бактерий Borrelia в лаборатории, диагноз лаймской болезни типично основан на результатах клинического исследования и истории посещения эндемических областей лайм-боррелиоза. Мигрирующая эритемная (EM) сыпь, которая наблюдается только в примерно 50% случаев, считается достаточной для установления диагноза лаймской болезни, даже если серологические анализы крови являются негативными. Серологические анализы могут быть использованы для подтверждения клинически подозрительных случаев, но сами по себе не являются диагностическими. Диагноз поздней стадии лаймской болезни часто трудно установить из-за многоаспектных проявлений, которые могут походить на симптомы многих других болезней. По этой причине, автор одного из обзоров назвал лайм-боррелиоз новым «великим имитатором». Лаймская болезнь, в некоторых случаях, ошибочно диагностируется как множественный склероз, ревматоидный артрит, фибромиалгия, синдром хронической усталости (CFS), волчанка, или другие аутоиммунные и нейродегенеративные болезни. Таким образом, у специалистов существует большая потребность в вакцине для предотвращения или лечения лаймской болезни.
Внешний поверхностный белок A (OspA) Borrelia
В различных аспектах, изобретение включает химерные молекулы OspA Borrelia и композиции, содержащие такие молекулы, для использования в профилактике и лечении лаймской болезни или инфекции Borrelia. За последнее десятилетие было идентифицировано несколько внешних поверхностных белков Borrelia, подверженных повышающей регуляции под действием температурных и/или специфических для млекопитающего-хозяина сигналов при переносе такой спирохеты от клещей к млекопитающим.
Основной внешний поверхностный белок, OspA, Borrelia burgdorferi является липопротеином, представляющим особый интерес вследствие его потенциальной пригодности для использования в качестве вакцины-кандидата. Серотипический и генетический анализ OspA как европейских, так и североамериканских штаммов Borrelia продемонстрировали антигенные и структурные гетерогенные признаки. OspA описан в опубликованной патентной заявке PCT WO 92/14488, в работе Jiang et al. (Clin. Diagn. Lab. Immunol. 1: 406-12, 1994) и известен специалистам. Было показано, что Osp A индуцирует защитный иммунитет в экспериментах с провокационной пробой у мышей, хомяков и собак. Клинические испытания на людях показали, что композиции OspA являются безопасными и иммуногенными для людей (Keller et al., JAMA (1994) 271:1764 1768).
Хотя OspA экспрессируется в подавляющем большинстве клинических изолятов Borrelia burgdorferi из Северной Америки, другая картина наблюдается при исследованиях клинических изолятов Borrelia в Европе. В Европе, лаймская болезнь вызывается преимущественно тремя геновидами Borrelia, а именно, B. burgdorferi, B. garinii и B. afzelii. Изобретение касается химерных молекул OspA, обеспечивающих защитный иммунитет против всех геновидов Borrelia. Изобретение описывает конструирование и синтез трех химерных генов OspA, которые кодируют три разных липидированных молекулы OspA, обладающих общими признаками. Каждый ген представляет два серотипа OspA, и гены были сконструированы так, чтобы они кодировали стабильные молекулы OspA, являющиеся безопасными и высокоиммуногенными, и обеспечивающие защиту субъекта против инфекции B. burgdorferi sensu lato (s.l.). Изобретение также описывает три исходные химерные гена OspA без мутаций и без оптимизации кодонов, которые кодируют три разных липидированных молекулы OspA, обладающих общими признаками. Каждый ген представляет два серотипа OspA и кодирует молекулы, обеспечивающие защиту субъекта против инфекции B. burgdorferi sensu lato (s.l.).
В европейских изолятах было найдено семь основных серотипов OspA (обозначенных как серотипы 1-7, Wilske et al., J. Clin. Microbiol. 31:340-50, 1993). Серотипы OspA коррелируют с видами; серотип 1 соответствует B. burgdorferi s.s., серотип 2 соответствует B. afzelii, и серотипы 3-7 соответствуют B. garinii. Эпидемиологические исследования европейских изолятов Borrelia показывают, что вакцина на основе OspA типов 1, 2, 3, 4, 5 и 6 обеспечит теоретическое покрытие в Европе 98,1% случаев лаймской болезни, и охватывают 96,7% изолятов инвазивных заболеваний. Изобретение предусматривает шесть химерных молекул нуклеиновой кислоты OspA (SEQ ID NO: 1, 3 и 5 и SEQ ID NO: 168, 170 и 172) и шесть химерных полипептидных молекул OspA (SEQ ID NO: 2, 4 и 6 и SEQ ID NO: 169, 171 и 173), которые могут обеспечить защитный иммунитет против всех шести серотипов 1-6. Были сконструированы шесть синтетических генов OspA, кодирующих молекулы OspA с защитными эпитопами OspA серотипов 1 и 2 (lipB sOspA 1/2251 (SEQ ID NO: 1 (нуклеиновая кислота) и 2 (аминокислота) и orig sOspA 1/2 (SEQ ID NO: 168 (нуклеиновая кислота) и 169 (аминокислота)); OspA серотипов 6 и 4 (lipB sOspA 6/4 (SEQ ID NO: 3 (нуклеиновая кислота) и 4 (аминокислота) и orig sOspA 6/4 (SEQ ID NO: 170 (нуклеиновая кислота) и 171 (аминокислота)); и OspA серотипов 5 и 3 (lipB sOspA 5/3 (SEQ ID NO: 5 (нуклеиновая кислота) и 6 (аминокислота) и orig sOspA 5/3 (SEQ ID NO: 172 (нуклеиновая кислота) и 173 (аминокислота)). Химерные гены OspA были получены с использованием синтетических перекрывающихся олигонуклеотидов. Такие рекомбинантные белки, в определенных аспектах, продуцируются с высоким выходом и чистотой и, в различных аспектах, подвергаются манипуляциям для максимизации желательных активностей и минимизации нежелательных.
Химерные молекулы нуклеиновой кислоты и полипептидные молекулы OspA
В различных аспектах, изобретение включает химерные молекулы нуклеиновой кислоты OspA и полипептидов Borrelia. Нуклеиновые кислоты OspA по изобретению включают молекулу нуклеиновой кислоты, содержащую, состоящую по существу из, или состоящую из нуклеотидной последовательности, приведенной в SEQ ID NO: 1 (lipB sOspA 1/2251), SEQ ID NO: 3 (lipB sOspA 6/4), SEQ ID NO: 5 (lipB sOspA 5/3), SEQ ID NO: 7 (sOspA 1/2251), SEQ ID NO: 9 (sOspA 6/4), SEQ ID NO: 11 (sOspA 5/3), SEQ ID NO: 168 (orig sOspA 1/2), SEQ ID NO: 170 (orig sOspA 6/4), или SEQ ID NO: 172 (orig sOspA 5/3), или нуклеотидной последовательности, кодирующей полипептид, приведенной в SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3).
Последовательности нуклеиновой кислоты SEQ ID NO: 7, 9 и 11 не содержат последовательности нуклеиновой кислоты, кодирующей лидерной последовательности lipB (MRLLIGFALALALIG (SEQ ID NO: 13). Кроме того, последовательности нуклеиновой кислоты SEQ ID NO: 7, 9 и 11 кодируют остаток метионина на амино-конце SEQ ID NO: 8, 10 и 12 вместо цистеинового остатка, присутствующего на карбокси-конце лидерной последовательности lipB в SEQ ID NO: 2, 4 и 6. SEQ ID NO: 1, 3 и 5 представляют собой полинуклеотиды lipB sOspA, а SEQ ID NO: 2, 4 и 6 - полипептиды lipB sOspA.
В некоторых аспектах, изобретение включает исходные молекулы («orig») химерной нуклеиновой кислоты OspA и полипептида Borrelia без мутаций и без оптимизации кодонов. Таким образом, нуклеиновые кислоты OspA по изобретению включают молекулу нуклеиновой кислоты, содержащую, состоящую по существу из, или состоящую из нуклеотидной последовательности, приведенной в SEQ ID NO: 168 (orig sOspA 1/2), SEQ ID NO: 170 (orig sOspA 6/4), или SEQ ID NO: 172 (orig sOspA 5/3), или нуклеотидной последовательности, кодирующей полипептид, приведенной в SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3).
Идентификационные номера последовательностей ДНК и аминокислотных последовательностей для химерных молекул OspA приведены в Таблице 1 ниже.
Химерные ДНК и аминокислотные последовательности OspA
SEQ ID NO:
SEQ ID NO:
SEQ ID NO:
--->
lipB sOspA 1/2251
Аминокислотная последовательность (SEQ ID NO: 2)
MRLLIGFALALALIGCAQKGAESIGSVSVDLPGEMKVLVSKEKDKNGKYDLIATVDKLELKGTSDKNNGS
GVLEGVKTNKSKVKLTISDDLGQTTLEVFKEDGKTLVSKKVTSKDKSSTEEKFNEKGEVSEKIITMADGT
RLEYTGIKSDGTGKAKYVLKNFTLEGKVANDKTTLEVKEGTVTLSMNISKSGEVSVELNDTDSSAATKKT
AAWNSKTSTLTISVNSKKTTQLVFTKQDTITVQKYDSAGTNLEGTAVEIKTLDELKNALK
ДНК-последовательность (SEQ ID NO: 1)
catatgcgtctgttgatcggctttgctctggcgctggctctgatcggctgcgcacagaaaggtgctgagt
ctattggttccgtttctgtagatctgcccggtgaaatgaaggttctggtgagcaaagaaaaagacaagaa
cggcaagtacgatctcatcgcaaccgtcgacaagctggagctgaaaggtacttctgataaaaacaacggc
tctggtgtgctggagggcgtcaaaactaacaagagcaaagtaaagcttacgatctctgacgatctcggtc
agaccacgctggaagttttcaaagaggatggcaagaccctcgtgtccaaaaaagtaacttccaaagacaa
gtcctctacggaagaaaaattcaacgaaaaaggtgaggtgtctgaaaagatcatcaccatggcagacggc
acccgtcttgaatacaccggtattaaaagcgatggtaccggtaaagcgaaatatgttctgaaaaacttca
ctctggaaggcaaagtggctaatgataaaaccaccttggaagtcaaggaaggcaccgttactctgagcat
gaatatctccaaatctggtgaagtttccgttgaactgaacgacactgacagcagcgctgcgactaaaaaa
actgcagcgtggaattccaaaacttctactttaaccattagcgttaacagcaaaaaaactacccagctgg
tgttcactaaacaagacacgatcactgtgcagaaatacgactccgcaggcaccaacttagaaggcacggc
agtcgaaattaaaacccttgatgaactgaaaaacgcgctgaaataagctgagcggatcc
Комплементарная нить (SEQ ID NO: 48)
catatgcgtctgttgatcggctttgctttggcgctggctttaatcggctgtgcacagaaaggtgctgagt
ctattggttccgtttctgtagatctgcccgggggtatgaaagttctggtaagcaaagaaaaagacaaaaa
cggtaaatacagcctgatggcaaccgtagaaaagctggagcttaaaggcacttctgataaaaacaacggt
tctggcaccctggaaggtgaaaaaactaacaaaagcaaagtaaagcttactattgctgaggatctgagca
aaaccacctttgaaatcttcaaagaagatggcaaaactctggtatctaaaaaagtaaccctgaaagacaa
gtcttctaccgaagaaaaattcaacgaaaagggtgaaatctctgaaaaaactatcgtaatggcaaatggt
acccgtctggaatacaccgacatcaaaagcgataaaaccggcaaagctaaatacgttctgaaagacttta
ctctggaaggcactctggctgctgacggcaaaaccactctgaaagttaccgaaggcactgttactctgag
catgaacatttctaaatccggcgaaatcaccgttgcactggatgacactgactctagcggcaataaaaaa
tccggcacctgggattctgatacttctactttaaccattagcaaaaacagccagaaaactaaacagctgg
tattcaccaaagaaaacactatcaccgtacagaactataaccgtgcaggcaatgcgctggaaggcagccc
ggctgaaattaaagatctggcagagctgaaagccgctttgaaataagctgagcggatcc
lipB sOspA 6/4
Аминокислотная последовательность (SEQ ID NO: 4)
MRLLIGFALALALIGCAQKGAESIGSVSVDLPGGMTVLVSKEKDKNGKYSLEATVDKLELKGTSDKNNGS
GTLEGEKTNKSKVKLTIADDLSQTKFEIFKEDAKTLVSKKVTLKDKSSTEEKFNEKGETSEKTIVMANGT
RLEYTDIKSDGSGKAKYVLKDFTLEGTLAADGKTTLKVTEGTVVLSMNILKSGEITVALDDSDTTQATKK
TGKWDSNTSTLTISVNSKKTKNIVFTKEDTITVQKYDSAGTNLEGNAVEIKTLDELKNALK
ДНК-последовательность (SEQ ID NO: 3)
catatgcgtctgttgatcggctttgctctggcgctggctctgatcggctgcgcacagaaaggtgctgagt
ctattggttccgtttctgtagatctgcccggtggcatgaccgttctggtcagcaaagaaaaagacaaaaa
cggtaaatacagcctcgaggcgaccgtcgacaagcttgagctgaaaggcacctctgataaaaacaacggt
tccggcaccctggaaggtgaaaaaactaacaaaagcaaagtgaaactgaccattgctgatgacctcagcc
agaccaaattcgaaattttcaaagaagatgccaaaaccttagtatccaaaaaagtgaccctgaaagacaa
gtcctctaccgaagaaaaattcaacgaaaagggtgaaacctctgaaaaaaccatcgtaatggcaaatggt
acccgtctggaatacaccgacatcaaaagcgatggctccggcaaagccaaatacgttctgaaagacttca
ccctggaaggcaccctcgctgccgacggcaaaaccaccttgaaagttaccgaaggcactgttgttttaag
catgaacatcttaaaatccggtgaaatcaccgttgcgctggatgactctgacaccactcaggccactaaa
aaaaccggcaaatgggattctaacacttccactctgaccatcagcgtgaattccaaaaaaactaaaaaca
tcgtgttcaccaaagaagacaccatcaccgtccagaaatacgactctgcgggcaccaacctcgaaggcaa
cgcagtcgaaatcaaaaccctggatgaactgaaaaacgctctgaaataagctgagcggatcc
Комплементарная нить (SEQ ID NO: 49)
ggatccgctcagcttatttcagcgcgtttttcagttcatcaagggttttaatttcgactgccgtgccttc
taagttggtgcctgcggagtcgtatttctgcacagtgatcgtgtcttgtttagtgaacaccagctgggta
gtttttttgctgttaacgctaatggttaaagtagaagttttggaattccacgctgcagtttttttagtcg
cagcgctgctgtcagtgtcgttcagttcaacggaaacttcaccagatttggagatattcatgctcagagt
aacggtgccttccttgacttccaaggtggttttatcattagccactttgccttccagagtgaagtttttc
agaacatatttcgctttaccggtaccatcgcttttaataccggtgtattcaagacgggtgccgtctgcca
tggtgatgatcttttcagacacctcacctttttcgttgaatttttcttccgtagaggacttgtctttgga
agttacttttttggacacgagggtcttgccatcctctttgaaaacttccagcgtggtctgaccgagatcg
tcagagatcgtaagctttactttgctcttgttagttttgacgccctccagcacaccagagccgttgtttt
tatcagaagtacctttcagctccagcttgtcgacggttgcgatgagatcgtacttgccgttcttgtcttt
ttctttgctcaccagaaccttcatttcaccgggcagatctacagaaacggaaccaatagactcagcacct
ttctgtgcgcagccgatcagagccagcgccagagcaaagccgatcaacagacgcatatg
lipB sOspA 5/3
Аминокислотная последовательность (SEQ ID NO: 6)
MRLLIGFALALALIGCAQKGAESIGSVSVDLPGGMKVLVSKEKDKNGKYSLMATVEKLELKGTSDKNNGS
GTLEGEKTNKSKVKLTIAEDLSKTTFEIFKEDGKTLVSKKVTLKDKSSTEEKFNEKGEISEKTIVMANGT
RLEYTDIKSDKTGKAKYVLKDFTLEGTLAADGKTTLKVTEGTVTLSMNISKSGEITVALDDTDSSGNKKS
GTWDSDTSTLTISKNSQKTKQLVFTKENTITVQNYNRAGNALEGSPAEIKDLAELKAALK
ДНК-последовательность (SEQ ID NO: 5)
catatgcgtctgttgatcggctttgctttggcgctggctttaatcggctgtgcacagaaaggtgctgagt
ctattggttccgtttctgtagatctgcccgggggtatgaaagttctggtaagcaaagaaaaagacaaaaa
cggtaaatacagcctgatggcaaccgtagaaaagctggagcttaaaggcacttctgataaaaacaacggt
tctggcaccctggaaggtgaaaaaactaacaaaagcaaagtaaagcttactattgctgaggatctgagca
aaaccacctttgaaatcttcaaagaagatggcaaaactctggtatctaaaaaagtaaccctgaaagacaa
gtcttctaccgaagaaaaattcaacgaaaagggtgaaatctctgaaaaaactatcgtaatggcaaatggt
acccgtctggaatacaccgacatcaaaagcgataaaaccggcaaagctaaatacgttctgaaagacttta
ctctggaaggcactctggctgctgacggcaaaaccactctgaaagttaccgaaggcactgttactctgag
catgaacatttctaaatccggcgaaatcaccgttgcactggatgacactgactctagcggcaataaaaaa
tccggcacctgggattctgatacttctactttaaccattagcaaaaacagccagaaaactaaacagctgg
tattcaccaaagaaaacactatcaccgtacagaactataaccgtgcaggcaatgcgctggaaggcagccc
ggctgaaattaaagatctggcagagctgaaagccgctttgaaataagctgagcggatcc
Комплементарная нить (SEQ ID NO: 50)
ggatccgctcagcttatttcagagcgtttttcagttcatccagggttttgatttcgactgcgttgccttc
gaggttggtgcccgcagagtcgtatttctggacggtgatggtgtcttctttggtgaacacgatgttttta
gtttttttggaattcacgctgatggtcagagtggaagtgttagaatcccatttgccggtttttttagtgg
cctgagtggtgtcagagtcatccagcgcaacggtgatttcaccggattttaagatgttcatgcttaaaac
aacagtgccttcggtaactttcaaggtggttttgccgtcggcagcgagggtgccttccagggtgaagtct
ttcagaacgtatttggctttgccggagccatcgcttttgatgtcggtgtattccagacgggtaccatttg
ccattacgatggttttttcagaggtttcacccttttcgttgaatttttcttcggtagaggacttgtcttt
cagggtcacttttttggatactaaggttttggcatcttctttgaaaatttcgaatttggtctggctgagg
tcatcagcaatggtcagtttcactttgcttttgttagttttttcaccttccagggtgccggaaccgttgt
ttttatcagaggtgcctttcagctcaagcttgtcgacggtcgcctcgaggctgtatttaccgtttttgtc
tttttctttgctgaccagaacggtcatgccaccgggcagatctacagaaacggaaccaatagactcagca
cctttctgtgcgcagccgatcagagccagcgccagagcaaagccgatcaacagacgcatatg
sOspA 1/2251
инокислотная последовательность (SEQ ID NO: 8)
MAQKGAESIGSVSVDLPGEMKVLVSKEKDKNGKYDLIATVDKLELKGTSDKNNGSGVLEGVKTNKSKVKL
TISDDLGQTTLEVFKEDGKTLVSKKVTSKDKSSTEEKFNEKGEVSEKIITMADGTRLEYTGIKSDGTGKA
KYVLKNFTLEGKVANDKTTLEVKEGTVTLSMNISKSGEVSVELNDTDSSAATKKTAAWNSKTSTLTISVN
SKKTTQLVFTKQDTITVQKYDSAGTNLEGTAVEIKTLDELKNALK
ДНК-последовательность (SEQ ID NO: 7)
catatggcacagaaaggtgctgagtctattggttccgtttctgtagatctgcccggtgaaatgaaggttc
tggtgagcaaagaaaaagacaagaacggcaagtacgatctcatcgcaaccgtcgacaagctggagctgaa
aggtacttctgataaaaacaacggctctggtgtgctggagggcgtcaaaactaacaagagcaaagtaaag
cttacgatctctgacgatctcggtcagaccacgctggaagttttcaaagaggatggcaagaccctcgtgt
ccaaaaaagtaacttccaaagacaagtcctctacggaagaaaaattcaacgaaaaaggtgaggtgtctga
aaagatcatcaccatggcagacggcacccgtcttgaatacaccggtattaaaagcgatggtaccggtaaa
gcgaaatatgttctgaaaaacttcactctggaaggcaaagtggctaatgataaaaccaccttggaagtca
aggaaggcaccgttactctgagcatgaatatctccaaatctggtgaagtttccgttgaactgaacgacac
tgacagcagcgctgcgactaaaaaaactgcagcgtggaattccaaaacttctactttaaccattagcgtt
aacagcaaaaaaactacccagctggtgttcactaaacaagacacgatcactgtgcagaaatacgactcca
acggcaccaacttagaaggcacggcagtcgaaattaaaacccttgatgaactgaaaaacgcgctgaaata
agctgagcggatcc
Комплементарная нить (SEQ ID NO: 56)
gtataccgtgtctttccacgactcagataaccaaggcaaagacatctagacgggccactttacttccaag
accactcgtttctttttctgttcttgccgttcatgctagagtagcgttggcagctgttcgacctcgactt
tccatgaagactatttttgttgccgagaccacacgacctcccgcagttttgattgttctcgtttcatttc
gaatgctagagactgctagagccagtctggtgcgaccttcaaaagtttctcctaccgttctgggagcaca
ggttttttcattgaaggtttctgttcaggagatgccttctttttaagttgctttttccactccacagact
tttctagtagtggtaccgtctgccgtgggcagaacttatgtggccataattttcgctaccatggccattt
cgctttatacaagactttttgaagtgagaccttccgtttcaccgattactattttggtggaaccttcagt
tccttccgtggcaatgagactcgtacttatagaggtttagaccacttcaaaggcaacttgacttgctgtg
actgtcgtcgcgacgctgatttttttgacgtcgcaccttaaggttttgaagatgaaattggtaatcgcaa
ttgtcgtttttttgatgggtcgaccacaagtgatttgttctgtgctagtgacacgtctttatgctgaggt
tgccgtggttgaatcttccgtgccgtcagctttaattttgggaactacttgactttttgcgcgactttat
tcgactcgcctagg
sOspA 6/4
Аминокислотная последовательность (SEQ ID NO: 10)
MAQKGAESIGSVSVDLPGGMTVLVSKEKDKNGKYSLEATVDKLELKGTSDKNNGSGTLEGEKTNKSKVKL
TIADDLSQTKFEIFKEDAKTLVSKKVTLKDKSSTEEKFNEKGETSEKTIVMANGTRLEYTDIKSDGSGKA
KYVLKDFTLEGTLAADGKTTLKVTEGTVVLSMNILKSGEITVALDDSDTTQATKKTGKWDSNTSTLTISV
NSKKTKNIVFTKEDTITVQKYDSAGTNLEGNAVEIKTLDELKNALK
ДНК-последовательность (SEQ ID NO: 9)
catatggcacagaaaggtgctgagtctattggttccgtttctgtagatctgcccggtggcatgaccgttc
tggtcagcaaagaaaaagacaaaaacggtaaatacagcctcgaggcgaccgtcgacaagcttgagctgaa
aggcacctctgataaaaacaacggttccggcaccctggaaggtgaaaaaactaacaaaagcaaagtgaaa
ctgaccattgctgatgacctcagccagaccaaattcgaaattttcaaagaagatgccaaaaccttagtat
ccaaaaaagtgaccctgaaagacaagtcctctaccgaagaaaaattcaacgaaaagggtgaaacctctga
aaaaaccatcgtaatggcaaatggtacccgtctggaatacaccgacatcaaaagcgatggctccggcaaa
gccaaatacgttctgaaagacttcaccctggaaggcaccctcgctgccgacggcaaaaccaccttgaaag
ttaccgaaggcactgttgttttaagcatgaacatcttaaaatccggtgaaatcaccgttgcgctggatga
ctctgacaccactcaggccactaaaaaaaccggcaaatgggattctaacacttccactctgaccatcagc
gtgaattccaaaaaaactaaaaacatcgtgttcaccaaagaagacaccatcaccgtccagaaatacgact
ctgcgggcaccaacctcgaaggcaacgcagtcgaaatcaaaaccctggatgaactgaaaaacgctctgaa
ataagctgagcggatcc
Комплементарная нить (SEQ ID NO: 57)
gtataccgtgtctttccacgactcagataaccaaggcaaagacatctagacgggccaccgtactggcaag
accagtcgtttctttttctgtttttgccatttatgtcggagctccgctggcagctgttcgaactcgactt
tccgtggagactatttttgttgccaaggccgtgggaccttccacttttttgattgttttcgtttcacttt
gactggtaacgactactggagtcggtctggtttaagctttaaaagtttcttctacggttttggaatcata
ggttttttcactgggactttctgttcaggagatggcttctttttaagttgcttttcccactttggagact
tttttggtagcattaccgtttaccatgggcagaccttatgtggctgtagttttcgctaccgaggccgttt
cggtttatgcaagactttctgaagtgggaccttccgtgggagcgacggctgccgttttggtggaactttc
aatggcttccgtgacaacaaaattcgtacttgtagaattttaggccactttagtggcaacgcgacctact
gagactgtggtgagtccggtgatttttttggccgtttaccctaagattgtgaaggtgagactggtagtcg
cacttaaggtttttttgatttttgtagcacaagtggtttcttctgtggtagtggcaggtctttatgctga
gacgcccgtggttggagcttccgttgcgtcagctttagttttgggacctacttgactttttgcgagactt
tattcgactcgcctagg
sOspA 5/3
Аминокислотная последовательность (SEQ ID NO: 12)
MAQKGAESIGSVSVDLPGGMKVLVSKEKDKNGKYSLMATVEKLELKGTSDKNNGSGTLEGEKTNKSKVKL
TIAEDLSKTTFEIFKEDGKTLVSKKVTLKDKSSTEEKFNEKGEISEKTIVMANGTRLEYTDIKSDKTGKA
KYVLKDFTLEGTLAADGKTTLKVTEGTVTLSMNISKSGEITVALDDTDSSGNKKSGTWDSDTSTLTISKN
SQKTKQLVFTKENTITVQNYNRAGNALEGSPAEIKDLAELKAALK
ДНК-последовательность (SEQ ID NO: 11)
catatggcacagaaaggtgctgagtctattggttccgtttctgtagatctgcccgggggtatgaaagttc
tggtaagcaaagaaaaagacaaaaacggtaaatacagcctgatggcaaccgtagaaaagctggagcttaa
aggcacttctgataaaaacaacggttctggcaccctggaaggtgaaaaaactaacaaaagcaaagtaaag
cttactattgctgaggatctgagcaaaaccacctttgaaatcttcaaagaagatggcaaaactctggtat
ctaaaaaagtaaccctgaaagacaagtcttctaccgaagaaaaattcaacgaaaagggtgaaatctctga
aaaaactatcgtaatggcaaatggtacccgtctggaatacaccgacatcaaaagcgataaaaccggcaaa
gctaaatacgttctgaaagactttactctggaaggcactctggctgctgacggcaaaaccactctgaaag
ttaccgaaggcactgttactctgagcatgaacatttctaaatccggcgaaatcaccgttgcactggatga
cactgactctagcggcaataaaaaatccggcacctgggattctgatacttctactttaaccattagcaaa
aacagccagaaaactaaacagctggtattcaccaaagaaaacactatcaccgtacagaactataaccgtg
caggcaatgcgctggaaggcagcccggctgaaattaaagatctggcagagctgaaagccgctttgaaata
agctgagcggatcc
Комплементарная нить (SEQ ID NO: 58)
gtataccgtgtctttccacgactcagataaccaaggcaaagacatctagacgggcccccatactttcaag
accattcgtttctttttctgtttttgccatttatgtcggactaccgttggcatcttttcgacctcgaatt
tccgtgaagactatttttgttgccaagaccgtgggaccttccacttttttgattgttttcgtttcatttc
gaatgataacgactcctagactcgttttggtggaaactttagaagtttcttctaccgttttgagaccata
gattttttcattgggactttctgttcagaagatggcttctttttaagttgcttttcccactttagagact
tttttgatagcattaccgtttaccatgggcagaccttatgtggctgtagttttcgctattttggccgttt
cgatttatgcaagactttctgaaatgagaccttccgtgagaccgacgactgccgttttggtgagactttc
aatggcttccgtgacaatgagactcgtacttgtaaagatttaggccgctttagtggcaacgtgacctact
gtgactgagatcgccgttattttttaggccgtggaccctaagactatgaagatgaaattggtaatcgttt
ttgtcggtcttttgatttgtcgaccataagtggtttcttttgtgatagtggcatgtcttgatattggcac
gtccgttacgcgaccttccgtcgggccgactttaatttctagaccgtctcgactttcggcgaaactttat
tcgactcgcctagg
Orig sOspA 1/2
Аминокислотная последовательность (SEQ ID NO: 169)
MKKYLLGIGLILALIACKQNVSSLDEKNSVSVDLPGEMKVLVSKEKNKDGKYDLIATVDKLEL
KGTSDKNNGSGVLEGVKADKSKVKLTISDDLGQTTLEVFKEDGKTLVSKKVTSKDKSSTEEKF
NEKGEVSEKIITRADGTRLEYTGIKSDGSGKAKEVLKNFTLEGKVANDKVTLEVKEGTVTLSK
NISKSGEVSVELNDTDSSAATKKTAAWNSKTSTLTISVNSKKTTQLVFTKQDTITVQKYDSAG
TNLEGTAVEIKTLDELKNALK
ДНК-последовательность (SEQ ID NO: 168)
atgaaaaaatatttattgggaataggtctaatattagccttaatagcatgtaagcaaaatgt
tagcagccttgacgagaaaaacagcgtttcagtagatttgcctggtgaaatgaaagttcttg
taagcaaagaaaaaaacaaagacggcaagtacgatctaattgcaacagtagacaagcttgag
cttaaaggaacttctgataaaaacaatggatctggagtacttgaaggcgtaaaagctgacaa
aagtaaagtaaaattaacaatttctgacgatctaggtcaaaccacacttgaagttttcaaag
aagatggcaaaacactagtatcaaaaaaagtaacttccaaagacaagtcatcaacagaagaa
aaattcaatgaaaaaggtgaagtatctgaaaaaataataacaagagcagacggaaccagact
tgaatacacaggaattaaaagcgatggatctggaaaagctaaagaggttttaaaaaacttta
ctcttgaaggaaaagtagctaatgataaagtaacattggaagtaaaagaaggaaccgttact
ttaagtaaaaatatttcaaaatctggggaagtttcagttgaacttaatgacactgacagtag
tgctgctactaaaaaaactgcagcttggaattcaaaaacttctactttaacaattagtgtta
acagcaaaaaaactacacaacttgtgtttactaaacaagacacaataactgtacaaaaatac
gactccgcaggtaccaatttagaaggcacagcagtcgaaattaaaacacttgatgaacttaa
aaacgctttaaaatag
Orig sOspA 6/4
Аминокислотная последовательность (SEQ ID NO: 171)
MKKYLLGIGLILALIACKQNVSTLDEKNSVSVDLPGGMTVLVSKEKDKDGKYSLEATVDKLE
LKGTSDKNNGSGTLEGEKTDKSKVKLTIADDLSQTKFEIFKEDAKTLVSKKVTLKDKSSTEE
KFNEKGETSEKTIVRANGTRLEYTDIKSDGSGKAKEVLKDFTLEGTLAADGKTTLKVTEGTV
VLSKNILKSGEITVALDDSDTTQATKKTGKWDSNTSTLTISVNSKKTKNIVFTKEDTITVQK
YDSAGTNLEGNAVEIKTLDELKNALK
ДНК-последовательность (SEQ ID NO: 170)
atgaaaaaatatttattgggaataggtctaatattagccttaatagcatgtaagcaaaatgt
tagcacgcttgatgaaaaaaatagcgtttcagtagatttacctggtggaatgacagttcttg
taagtaaagaaaaagacaaagacggtaaatacagtctagaggcaacagtagacaagcttgag
cttaaaggaacttctgataaaaacaacggttctggaacacttgaaggtgaaaaaactgacaa
aagtaaagtaaaattaacaattgctgatgacctaagtcaaactaaatttgaaattttcaaag
aagatgccaaaacattagtatcaaaaaaagtaacccttaaagacaagtcatcaacagaagaa
aaattcaacgaaaagggtgaaacatctgaaaaaacaatagtaagagcaaatggaaccagact
tgaatacacagacataaaaagcgatggatccggaaaagctaaagaagttttaaaagacttta
ctcttgaaggaactctagctgctgacggcaaaacaacattgaaagttacagaaggcactgtt
gttttaagcaagaacattttaaaatccggagaaataacagttgcacttgatgactctgacac
tactcaggctactaaaaaaactggaaaatgggattcaaatacttccactttaacaattagtg
tgaatagcaaaaaaactaaaaacattgtatttacaaaagaagacacaataacagtacaaaaa
tacgactcagcaggcaccaatctagaaggcaacgcagtcgaaattaaaacacttgatgaact
taaaaacgctttaaaataa
Orig sOspA 5/3
Аминокислотная последовательность (SEQ ID NO: 173)
MKKYLLGIGLILALIACKQNVSSLDEKNSVSVDLPGGMKVLVSKEKDKDGKYSLMATVEKLE
LKGTSDKNNGSGTLEGEKTDKSKVKLTIAEDLSKTTFEIFKEDGKTLVSKKVTLKDKSSTEE
KFNEKGEISEKTIVRANGTRLEYTDIKSDKTGKAKEVLKDFTLEGTLAADGKTTLKVTEGTV
TLSKNISKSGEITVALDDTDSSGNKKSGTWDSDTSTLTISKNSQKTKQLVFTKENTITVQNY
NRAGNALEGSPAEIKDLAELKAALK
ДНК-последовательность (SEQ ID NO: 172)
atgaaaaaatatttattgggaataggtctaatattagccttaatagcatgtaagcaaaatgt
tagcagccttgatgaaaaaaatagcgtttcagtagatttacctggtggaatgaaagttcttg
taagtaaagaaaaagacaaagatggtaaatacagtctaatggcaacagtagaaaagcttgag
cttaaaggaacttctgataaaaacaacggttctggaacacttgaaggtgaaaaaactgacaa
aagtaaagtaaaattaacaattgctgaggatctaagtaaaaccacatttgaaatcttcaaag
aagatggcaaaacattagtatcaaaaaaagtaacccttaaagacaagtcatcaacagaagaa
aaattcaacgaaaagggtgaaatatctgaaaaaacaatagtaagagcaaatggaaccagact
tgaatacacagacataaaaagcgataaaaccggaaaagctaaagaagttttaaaagacttta
ctcttgaaggaactctagctgctgacggcaaaacaacattgaaagttacagaaggcactgtt
actttaagcaagaacatttcaaaatccggagaaataacagttgcacttgatgacactgactc
tagcggcaataaaaaatccggaacatgggattcagatacttctactttaacaattagtaaaa
acagtcaaaaaactaaacaacttgtattcacaaaagaaaacacaataacagtacaaaactat
aacagagcaggcaatgcgcttgaaggcagcccagctgaaattaaagatcttgcagagcttaa
agccgctttaaaataa
<---
Полипептиды OspA по изобретению включают полипептид, содержащий, состоящий по существу из, или состоящий из аминокислотной последовательности SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3) и родственные полипептиды. Родственные полипептиды включают аналоги полипептида OspA, варианты полипептида OspA и производные полипептида OspA. В некоторых аспектах, полипептид OspA имеет амино-концевой остаток метионина, в зависимости от способа их получения. В родственных аспектах, полипептид OspA по изобретению содержит OspA активность.
В одном варианте воплощения, родственные молекулы нуклеиновой кислоты содержат или состоят из нуклеотидной последовательности, примерно на 70 процентов (70%) идентичной или подобной нуклеотидной последовательности, приведенной в SEQ ID NO: 1 (lipB sOspA 1/2251), SEQ ID NO: 3 (lipB sOspA 6/4), SEQ ID NO: 5 (lipB sOspA 5/3), SEQ ID NO: 7 (sOspA 1/2251), SEQ ID NO: 9 (sOspA 6/4), SEQ ID NO: 11 (sOspA 5/3), SEQ ID NO: 168 (orig sOspA 1/2), SEQ ID NO: 170 (orig sOspA 6/4), или SEQ ID NO: 172 (orig sOspA 5/3), в определенных аспектах, содержат, состоят по существу из, или состоят из нуклеотидной последовательности, кодирующей полипептид, примерно на 70 процентов (70%) идентичный полипептиду, приведенному в SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3). В различных вариантах воплощения нуклеотидные последовательности примерно на 70 процентов, или примерно на 71, 72, 73, 74, 75, 76, 77, 78, или 79 процентов, или примерно на 80 процентов, или примерно на 81, 82, 83, 84, 85, 86, 87, 88, или 89 процентов, или примерно на 90 процентов, или примерно на 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентов идентичны нуклеотидной последовательности, приведенной в SEQ ID NO: 1 (lipB sOspA 1/2251), SEQ ID NO: 3 (lipB sOspA 6/4), SEQ ID NO: 5 (lipB sOspA 5/3), SEQ ID NO: 7 (sOspA 1/2251), SEQ ID NO: 9 (sOspA 6/4), SEQ ID NO: 11 (sOspA 5/3), SEQ ID NO: 168 (orig sOspA 1/2), SEQ ID NO: 170 (orig sOspA 6/4), или SEQ ID NO: 172 (orig sOspA 5/3), или нуклеотидные последовательности кодируют полипептид, примерно на 70 процентов, или примерно на 71, 72, 73, 74, 75, 76, 77, 78, или 79 процентов, или примерно на 80 процентов, или примерно на 81, 82, 83, 84, 85, 86, 87, 88 или 89 процентов, или примерно на 90 процентов, или примерно на 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентов идентичный полипептидной последовательности, приведенной в SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3).
В некоторых вариантах воплощения разработаны способы определения идентичности и/или подобия последовательностей для получения наибольшего совпадения сравниваемых последовательностей. Способы определения идентичности и подобия описаны в публично доступных компьютерных программах. В некоторых аспектах, способы определения идентичности и подобия двух последовательностей с помощью компьютерной программы включают, без ограничений, программный пакет GCG, включая GAP (Devereux et al., Nucl. Acid. Res., 12:387 (1984); Genetics Computer Group, University of Wisconsin, Madison, WI, BLASTP, BLASTN и FASTA (Altschul et al., J. Mol. Biol., 215:403-410 (1990)). Программа BLASTX является общедоступной от Национального центра биотехнологической информации (National Center for Biotechnology Information, NCBI) и из других источников (BLAST Manual, Altschul et al. NCB/NLM/NIH Bethesda, MD 20894; Altschul et al., supra (1990)). Хорошо известный алгоритм Смита-Уотермана также используется для определения идентичности.
Определенные схемы выравнивания для сравнения двух аминокислотных последовательностей, в некоторых аспектах, приводят к совмещению только короткого участка двух последовательностей, и этот маленький совмещенный участок может иметь очень высокую идентичность последовательностей, даже если между двумя полноразмерными последовательностями нет значительного родства. Соответственно, в одном варианте воплощения выбранный способ выравнивания (программа GAP) будет приводить к выравниванию, охватывающему по меньшей мере 50 последовательно расположенных аминокислот целевого полипептида. Например, при использовании компьютерного алгоритма GAP (Genetics Computer Group, University of Wisconsin, Madison, WI), два полипептида, для которых нужно определить процент идентичности последовательностей, выравнивают для достижения оптимального совпадения их соответствующих аминокислот («совпадающая протяженность», как определено в алгоритме). В сочетании с алгоритмом используются штраф за открытие делеции (который рассчитывается как 3× среднюю диагональ; «средняя диагональ» представляет собой среднее значение диагонали используемой матрицы сравнения; «диагональ» является количественным показателем или числом, присваиваемым каждому полному совпадению аминокислот в конкретной матрице сравнения) и штраф за продолжение делеции (который обычно составляет 1/10 от штрафа за открытие делеции), а также матрица сравнения, такая как PAM 250 или BLOSUM 62. Алгоритмом также используется стандартная матрица сравнения (см. Dayhoff et al., Atlas of Protein Sequence and Structure, 5(3)(1978) - про матрицу сравнения PAM 250; Henikoff et al., Proc. Natl. Acad. Sci USA, 89:10915-10919 (1992) - про матрицу сравнения BLOSUM 62).
В различных аспектах, параметры сравнения полипептидных последовательностей включают:
Алгоритм: Needleman et al., J. Mol. Biol., 48:443-453 (1970);
Матрица сравнения: BLOSUM 62 по Henikoff et al., supra (1992);
Штраф за открытие делеции: 12
Штраф за продолжение делеции: 4
Порог подобия: 0
Программа GAP является пригодной для использования с приведенными выше параметрами. Вышеуказанные параметры являются параметрами по умолчанию для сравнения полипептидов (вместе с отсутствием штрафа за завершение делеции) с использованием алгоритма GAP.
В некоторых аспектах, параметры сравнения последовательностей молекул нуклеиновой кислоты включают:
Алгоритм: Needleman et al., supra (1970);
Матрица сравнения: совпадение = +10, несовпадение = 0
Штраф за открытие делеции: 50
Штраф за продолжение делеции: 3
Программа GAP также пригодна для использования с приведенными выше параметрами. Вышеуказанные параметры являются параметрами по умолчанию для сравнения молекул нуклеиновой кислоты. Другие типичные примеры алгоритмов, штрафов за открытие делеции, штрафов за продолжение делеции, матриц сравнения, порогов подобия и т.п. используются квалифицированными специалистами в данной области техники, включая те, которые приведены в Руководстве программы, Wisconsin Package, Version 9, September, 1997. Процедура выбора конкретных параметров понятна квалифицированным специалистам в данной области техники и будет зависеть от конкретного сравнения, которое необходимо провести, такого как ДНК-с-ДНК, белок-с-белком, белок-с-ДНК; и, дополнительно, от того, проводится ли сравнение между данными парами последовательностей (в этом случае в общем предпочтительными являются GAP или BestFit) или между одной последовательностью и большой базой данных последовательностей (в этом случае предпочтительны FASTA или BLASTA).
Различия в последовательностях нуклеиновой кислоты, в некоторых аспектах, приводят к консервативным и/или неконсервативным модификациям аминокислотной последовательности по сравнению с аминокислотной последовательностью SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3).
Консервативные модификации аминокислотной последовательности SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3) (и соответствующие модификации кодирующих нуклеотидов) будут давать полипептиды OspA, имеющие функциональные и химические характеристики, схожие с природным полипептидом OspA. В отличие от этого, существенные модификации функциональных и/или химических характеристик полипептидов OspA осуществляются путем выбора замещений в аминокислотной последовательности SEQ ID NO: 2 (lipB sOspA 1/2251), SEQ ID NO: 4 (lipB sOspA 6/4), SEQ ID NO: 6 (lipB sOspA 5/3), SEQ ID NO: 8 (sOspA 1/2251), SEQ ID NO: 10 (sOspA 6/4), SEQ ID NO: 12 (sOspA 5/3), SEQ ID NO: 169 (orig sOspA 1/2), SEQ ID NO: 171 (orig sOspA 6/4), или SEQ ID NO: 173 (orig sOspA 5/3), которые бы значительно отличались по своему влиянию на поддержание (а) структуры основной цепи молекулы в области замещения, например, в листовой или спиральной конформации, (б) заряда или гидрофобности молекулы в сайте-мишени, или (в) объема боковой цепи.
Например, «консервативное аминокислотное замещение», в некоторых аспектах, включает замещение нативного аминокислотного остатка не-нативным остатком так, чтобы при этом отсутствовал или возникал незначительный эффект на полярность или заряд аминокислотного остатка в данном положении. Кроме того, любой нативный остаток в полипептиде, в определенных аспектах, также замещается аланином, как было ранее описано для «сканирующего аланинового мутагенеза».
Консервативные аминокислотные замещения также охватывают неприродные аминокислотные остатки, которые типично вводятся способами химического синтеза пептидов, а не синтезом в биологических системах. Они включают пептидомиметики и другие среверсированные или инвертированные формы аминокислотных фрагментов.
Природные остатки, в различных аспектах, делятся на классы по общим свойствам боковых цепей:
гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
кислотные: Asp, Glu;
основные: His, Lys, Arg;
остатки, влияющие на ориентацию цепи: Gly, Pro; и
ароматические: Trp, Tyr, Phe.
Например, неконсервативные замещения, в некоторых аспектах, включают замену члена одного из этих классов на член из другого класса. Такие замещенные остатки, в различных аспектах, вводятся в области полипептида OspA, являющиеся гомологичными, или подобными ортологам полипептида OspA, или в негомологичные области молекулы.
При выполнении таких изменений часто принимается во внимание гидропатический индекс аминокислот. Каждой аминокислоте был присвоен гидропатический индекс на основании ее характеристик гидрофобности и заряда. Они имеют следующие значения: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серин (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9); и аргинин (-4,5).
Важность гидропатического индекса аминокислоты в придании белку функций интерактивного биологического взаимодействия понятна специалистам (Kyte et al., J. Mol. Biol., 157:105-131 (1982)). Известно, что определенные аминокислоты могут быть замещены другими аминокислотами, имеющими схожий гидропатический индекс или балльную оценку, и сохранять при этом схожую биологическую активность. При внесении изменений на основе гидропатического индекса замещение аминокислот с гидропатическими индексами в пределах ±2, в определенных аспектах, является предпочтительным, в других аспектах, с индексами в пределах ±1 - является особенно предпочтительным, и в пределах ±0,5, в различных аспектах - наиболее предпочтительным.
Специалистам в данной области техники понятно также, что замещение подобных аминокислот может быть эффективно проведено на основе гидрофильности, особенно в тех случаях, когда в результате этого должен быть получен биологически функциональный эквивалент белка или пептида, в частности, для использования в иммунологических вариантах воплощения, как в данном случае. Наибольшая локальная средняя гидрофильность белка, определяемая гидрофильностью его расположенных рядом аминокислот, коррелирует с его иммуногенностью и антигенностью, т.е. с биологическими свойствами белка.
Указанным аминокислотным остаткам были присвоены следующие значения гидрофильности: аргинин (+3,0); лизин (+3,0); аспартат (+3,0±1); глутамат (+3,0±1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5±1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5) и триптофан (-3,4). При внесении изменений на основе сходных значений гидрофильности, замещение аминокислот, значения гидрофильности которых отличаются в пределах ±2, в определенных аспектах, являются предпочтительными, отличающиеся на ±1, в других аспектах, являются особенно предпочтительными, и на ±0,5 - в различных аспектах, являются наиболее предпочтительными. Специалист также идентифицирует эпитопы по первичным аминокислотным последовательностям на основе гидрофильности. Такие участки также называются «участки ядра эпитопа».
Желательные аминокислотные замещения (консервативные или неконсервативные) могут быть определены квалифицированными специалистами в данной области техники в момент времени, когда такие замещения будут желательными. Например, аминокислотные замещения могут быть использованы для идентификации важных остатков полипептида OspA, или для увеличения или уменьшения аффинности полипептидов OspA к их субстратам, описанным тут.
В некоторых аспектах, замещения нуклеотидов в нуклеотидных последовательностях и аминокислот в аминокислотных последовательностях включены в изобретение. Замещения включают от одного до 5, от одного до 10, от одного до 15, от одного до 20, от одного до 25, от одного до 30, от одного до 35, от одного до 40, от одного до 45, от одного до 50, от одного до 55, от одного до 60, от одного до 65, от одного до 70, от одного до 75, от одного до 80, от одного до 85, от одного до 90, от одного до 95, от одного до 100, от одного до 150, и от одного до 200 нуклеотидов. Аналогично, замещения включают от одного до 5, от одного до 10, от одного до 15, от одного до 20, от одного до 25, от одного до 30, от одного до 35, от одного до 40, от одного до 45, от одного до 50, от одного до 55, от одного до 60, от одного до 65, от одного до 70, от одного до 75, от одного до 80, от одного до 85, от одного до 90, от одного до 95 и от одного до 100 аминокислот. Замещения, в различных аспектах, являются консервативными или неконсервативными.
Типичные примеры аминокислотных замещений приведены в Таблице 2.
Аминокислотные замещения
Квалифицированный специалист может определить пригодные аналоги или варианты полипептида, приведенные в SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173, используя хорошо известные методики. Для идентификации пригодных областей молекулы, которые могут быть изменены без уничтожения активности, квалифицированный специалист в данной области техники может выбрать в качестве мишени области, которые считаются неважными для проявления активности. Например, если известны схожие полипептиды со схожими активностями от одного и того же вида или от других видов, квалифицированный специалист в данной области техники может сравнивать аминокислотную последовательность полипептида OspA с такими схожими полипептидами. При таком сравнении, можно идентифицировать остатки и участки молекул, являющиеся консервативными в схожих полипептидах. Следует понимать, что изменения в областях полипептида OspA, не являющихся консервативными по сравнению с такими схожими полипептидами, будут с меньшей вероятностью оказывать нежелательное влтяние на биологическую активность и/или структуру полипептида OspA. Квалифицированный специалист в данной области техники будет также знать, что, даже в относительно консервативных областях, можно заменять химически схожие аминокислоты на природные остатки, сохраняя при этом активность (консервативные замещения аминокислотных остатков).
В некоторых вариантах воплощения полипептидные варианты OspA включают гликозилирование варианты, в которых число и/или тип сайтов гликозилирования были изменены по сравнению с аминокислотной последовательностью, приведенной в SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173. В одном варианте воплощения полипептидные варианты OspA содержат большее или меньшее число N-связанных сайтов гликозилирования, чем аминокислотная последовательность, приведенная в SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173. N-связанный сайт гликозилирования характеризуется последовательностью: Asn-X-Ser или Asn-X-Thr, где аминокислотный остаток, обозначенный как X, может быть любым аминокислотным остатком, кроме пролина. Замещение аминокислотных остатков для создания этой последовательности создает потенциальный новый сайт для аддиции N-связанной углеводной цепи. Альтернативно, замещения, уничтожающие такую последовательность, будут удалять существующую N-связанную углеводную цепь. Также предусматривается перегруппировка N-связанной углеводной цепи, когда удаляется один или более N-связанных сайтов гликозилирования (типично являющихся природными) и создается один или более новых N-связанных сайтов. Дополнительные варианты OspA включают цистеиновые варианты, в которых один или более цистеиновых остатков удалены или замещены на другую аминокислоту (например, серин) по сравнению с аминокислотной последовательностью, приведенной в SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173. Цистеиновые варианты полезны в тех случаях, когда полипептиды OspA должны быть подвергнуты переукладке в биологически активную конформацию, такой как после выделения нерастворимых телец включения. Цистеиновые варианты в общем содержат меньше цистеиновых остатков, чем нативный белок, и типично имеют четное их количество, для минимизации взаимодействий, вызываемых неспаренными цистеинами.
Изобретение далее предусматривает полипептиды, содержащие несущий эпитоп участок белка, как показано в SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173. Термин «эпитоп» относится к области белка, с которой может связываться антитело. См., например, Geysen et al., Proc. Natl. Acad. Sci. USA 81:3998-4002 (1984). Эпитопы могут быть линейными или конформационными, причем последние состоят из прерывистых участков белка, которые образуют эпитоп при укладке белка. Линейные эпитопы обычно имеют по меньшей мере 6 аминокислотных остатков в длину. Относительно короткие синтетические пептиды, имитирующие часть белковой последовательности, нормально способны вызывать образование антисыворотки, которая реагирует с частично имитируемым белком. См. Sutcliffe et al., Science 219:660-666 (1983). Антитела, распознающие короткие линейные эпитопы, являются особенно пригодными для использования в аналитических и диагностических областях применения, использующих денатурированный белок, таких как вестерн-блоттинг. См. Tobin, Proc. Natl. Acad. Sci. USA, 76:4350-4356 (1979). Антитела к коротким пептидам, в определенных случаях, также распознают белки в нативной конформации и, таким образом, будут полезны для контроля экспрессии белка и выделения белка и для детектирования OspA-белков в растворе, например, способом ELISA (иммуноферментного твердофазного анализа) или при проведении исследований способом иммунопреципитации.
Синтез химерных молекул нуклеиновой кислоты и полипептидных молекул OspA
Молекулы нуклеиновой кислоты кодируют полипептид, содержащий аминокислотную последовательность полипептида OspA, и могут быть легко получены различными способами, включая, без ограничений, способы рекомбинантных ДНК и химический синтез.
Способы рекомбинантных ДНК в общем описаны в Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989), и/или Ausubel et al., eds., Current Protocols in Molecular Biology, Green Publishers Inc. и Wiley and Sons, NY (1994). Методики рекомбинантной экспрессии, которые проводят в соответствии с приведенным ниже описанием, в различных аспектах, используют для продуцирования таких полинуклеотидов и для экспрессии кодируемых полипептидов. Например, с помощью инсерции последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность полипептида OspA в пригодный вектор, квалифицированный специалист в данной области техники может легко получить большие количества желательной нуклеотидной последовательности. Последовательности могут быть затем использованы для получения зондов для детектирования или праймеров для амплификации. Альтернативно, полинуклеотид, кодирующий аминокислотную последовательность полипептида OspA, может быть вставлен в экспрессионный вектор. В результате введения экспрессионного вектора пригодному хозяину, в некоторых аспектах, получают в больших количествах кодируемый полипептид OspA или полипептиды OspA.
Аналогично, химический синтез нуклеиновых кислот и полипептидов хорошо известен специалистам, такой как описанный Engels et al., Angew. Chem. Intl. Ed., 28:716-734 (1989). Такие способы включают, inter alia, фосфотриэфирный, фосфорамидитный и H-фосфонатный способы синтеза нуклеиновой кислоты. В одном аспекте, способ такого химического синтеза представляет собой синтез на полимерной подложке с использованием стандартной фосфорамидитной химии. Типично, ДНК, кодирующая аминокислотную последовательность полипептида OspA, будет содержать несколько сотен нуклеотидов в длину. Нуклеиновые кислоты размером более примерно 100 нуклеотидов синтезируют с использованием таких способов в виде нескольких фрагментов. Фрагменты затем лигируют вместе для получения полноразмерных нуклеотидных последовательностей по изобретению. В конкретных аспектах, фрагмент ДНК, кодирующий амино-конец полипептида, содержит ATG, кодирующий остаток метионина.
В конкретном аспекте изобретения, химерные кодирующие последовательности OspA получают с использованием синтетических перекрывающихся олигонуклеотидов. Поскольку ДНК из клеток Borrelia не используется, дополнительный полезный эффект синтетического подхода заключается в предотвращении загрязнения случайными агентами, содержащимися в материале животного происхождения (т.е. сыворотке или сывороточном альбумине), присутствующем в культуральной среде Borrelia. Эта стратегия также существенно уменьшает число манипуляций, необходимых для получения химерных генов, поскольку она позволяет осуществлять изменения последовательности за одну стадию, такие как модификации с целью оптимизации экспрессии (лидерная последовательность OspB), для введения сайтов рестрикции для облегчения клонирования, или во избежание потенциальных проблем, связанных с интеллектуальной собственностью. Она также позволяет оптимизировать использование кодонов для использования в качестве хозяина E. coli, поскольку известно, что присутствие кодонов, редко используемых в E. coli, представляет собой потенциальные препятствия для высоких уровней экспрессии чужеродных генов (Makoff et al., Nucleic Acids Res. 17:10191-202, 1989; Lakey et al., Infect. Immun. 68:233-8, 2000). Также используются другие способы, известные квалифицированному специалисту.
В определенных вариантах воплощения варианты нуклеиновой кислоты содержат кодоны, которые были изменены для оптимальной экспрессии полипептида OspA в данной клетке-хозяине. Конкретные изменения кодонов зависят от полипептида (полипептидов) OspA и клетки-хозяина (клеток-хозяев), выбранных для экспрессии. Такая «оптимизация кодонов» может быть проведена различными способами, например, путем выбора кодонов, являющихся предпочтительными для использования в генах с высокой экспрессией в данной клетке-хозяине. В некоторых случаях используются компьютерные алгоритмы, которые включают таблицы частоты встречаемости кодонов, такие как «Ecohigh.cod» для кодонов, предпочтительных для бактериальных генов с высокой экспрессией, предоставленные University of Wisconsin Package Version 9.0, Genetics Computer Group, Madison, WI. Другие пригодные таблицы частоты встречаемости кодонов включают «Celegans_high.cod», «Celegans_low.cod», «Drosophila_high.cod», «Human_high.cod», «Maize_high.cod» и «Yeast_high.cod».
Молекулу нуклеиновой кислоты, кодирующую аминокислотную последовательность полипептида OspA, в определенных аспектах, вставляют в пригодный экспрессионный вектор с помощью стандартных способов лигирования. Вектор типично выбирают таким образом, чтобы он был функциональным в конкретной используемой клетке-хозяине (т.е. вектор является совместимым с механизмами клетки-хозяина, так чтобы могла происходить амплификация гена и/или экспрессия гена). Молекула нуклеиновой кислоты, кодирующая аминокислотную последовательность полипептида OspA, в различных аспектах, амплифицируется/экспрессируется в прокариотических, дрожжевых, принадлежащих насекомым (бакуловирусные системы) и/или эукариотических клетках-хозяевах. Выбор клетки-хозяина зависит частично от того, должен ли полипептид OspA подвергаться посттрансляционному модифицированию (например, гликозилированию и/или фосфорилированию). Если должен, то предпочтительными клетками - хозяевами являются клетки дрожжей, насекомых или млекопитающего. Обзор экспрессионных векторов приведен в Meth. Enz., vol.185, D.V. Goeddel, ed., Academic Press Inc., San Diego, CA (1990).
Клонирующие векторы включают все векторы, известные специалистам. См., например, Sambrook, Fritsch & Maniatis, Molecular Cloning: A Laboratory Manual, Second Edition. Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press, 1989. В одном аспекте, в качестве клонирующего вектора для всех промежуточных стадий используется pUC18, потому что генетические манипуляции и секвенирование с этой плазмидой проводить проще, чем с вектором pET30a. Основными отличительными признаками являются генный фрагмент lacZ, кодирующий альфа-пептид LacZ пары оснований от 149 до 469 (промотор lac от пары оснований 507), ген bla, кодирующий детерминанту резистентности к ампициллину пары оснований от 1629 до 2486 (промотор bla от пары оснований 2521), точка начала репликации у пары оснований 867 и сайты множественного клонирования пары оснований от 185 до 451 (Фиг. 12).
Экспрессионные векторы включают все векторы, известные специалистам, включая, без ограничений, космиды, плазмиды (например, «голые» или содержащиеся в липосомах) и вирусы, включающие рекомбинантный полинуклеотид. Экспрессионный вектор вставляют (например, путем трансформации или трансдукции) в пригодную клетку-хозяина для экспрессии полинуклеотида и полипептида с помощью трансформации или трансфекции с использованием способов, известных специалистам. См., например, Sambrook, Fritsch & Maniatis, Molecular Cloning: A Laboratory Manual, Second Edition. Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press, 1989. В одном аспекте, pET30a (Novagen) используется в качестве экспрессионного вектора для конечной вставки полного гена OspA. В векторах pET, гены клонируются под контролем промотора T7 и экспрессия индуцируется путем обеспечения источника T7 РНК-полимеразы в клетке-хозяине (экспрессия не протекает до обеспечения источника T7 РНК-полимеразы). Основными отличительными признаками являются ген, кодирующий резистентность к канамицину (kan) от пары оснований 4048 до 4860, ген lacI пары оснований 826-1905, точка начала репликации F1 пары оснований 4956-5411 и сайты множественного клонирования пары оснований от 158 до 346 (Фиг. 13).
После того, как вектор будет сконструирован, и молекула нуклеиновой кислоты, кодирующая полипептид OspA, будет вставлена в требуемый сайт вектора, готовый вектор вводят в пригодную клетку-хозяина для амплификации и/или экспрессии полипептида. Трансформация экспрессионного вектора для полипептида OspA в выбранную клетку-хозяина, в различных аспектах, осуществляется хорошо известными способами, такими как трансфекция, инфекция, медиируемая хлоридом кальция трансформация, электропорация, микроинъекция, липофекция или способ DEAE-декстран, или другими известными способспособами. Выбранный способ будет частично определяться типом клетки-хозяина, которая должна быть использована. Такие способы и другие пригодные способы хорошо известны квалифицированному специалисту и приведены, например, в Sambrook et al., supra.
Клетки-хозяева, в некоторых аспектах, являются прокариотическими клетками-хозяевами (такими как E. coli) или эукариотическими клетками-хозяевами (такими как клетки дрожжей, насекомых или позвоночных). Клетка-хозяин, при культивации в пригодных условиях, синтезирует полипептид OspA, который затем может быть выделен из культуральной среды (если клетка-хозяин секретирует его в среду) или непосредственно из продуцирующей его клетки-хозяина (если он не секретируется). Выбор пригодной клетки-хозяина зависит от различных факторов, таких как желательные уровни экспрессии, модификации полипептида, желательные или необходимые для проявления активности (такие как гликозилирование или фосфорилирование) и легкость укладки в биологически активную молекулу. Такие клетки-хозяева включают, без ограничений, клетки-хозяева бактериального, дрожжевого, грибкового, вирусного происхождения, беспозвоночных и млекопитающих. Примеры таких клеток-хозяев приведены в Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1989). В дополнительных аспектах, клетки-хозяева, которые стали использоваться специалистами после публикации руководства Maniatis (supra), также используются в изобретении.
В одном аспекте, клетка-хозяин представляет собой клетку E. coli. Пригодные штаммы E. coli включают, без ограничений, BL21, DH5α, HMS174(DE3), DH10B или E. CLONI 10G (Lucigen, Middleton, Wis.). В некоторых вариантах воплощения клетки-хозяева подвергают генно-инженерным способам воздействия для повышения эффективности трансформации и/или поддержания вектора.
В одном аспекте, штамм DH5α E. coli [генотип: end A1 hsdR17 (rK-mK+) supE44 thi-1 recA1 gyrA (Nalr) relA1 D(lacZYA-argF)U169 deoR (F80dlacD(lacZ)M15] (Gibco BRL) используется для всех промежуточных стадий клонирования. Этот штамм был получен из штамма K12 E. coli, одного из наиболее широко используемых хозяев в генной инженерии. Штамм является amp- для обеспечения возможности селекции трансформантов векторами, содержащими ген резистентности к ампициллину (amp).
В другом аспекте, в качестве хозяина для экспрессии используется штамм E. coli HMS174(DE3). Клетки-хозяева E. coli HMS174(DE3) [генотип: F- recA1 hsdR (rk12- mk12+) RifR (DE3)] (Novagen) используются в различных описанных тут примерах для конечных стадий клонирования. Штамм является kan- для обеспечения возможности селекции трансформантов векторами, содержащими ген резистентности к канамицину (kan).
Клетки-хозяева, содержащие экспрессионный вектор полипептида OspA, культивируют с использованием стандартных сред, хорошо известных квалифицированному специалисту. Среды обычно будут содержать все питательные вещества, необходимые для роста и выживания клеток. Пригодные среды для культивации клеток E. coli включают, например, среду Луриа (LB) и/или Terrific Broth (TB). Пригодные среды для культивации эукариотических клеток включают среду Roswell Park Memorial Institute 1640 (RPMI 1640), минимальную существенную среду (MEM) и/или модифицированную по Дульбекко среду Игла (DMEM), все, в некоторых случаях, с добавкой сыворотки и/или факторов роста, требуемых для конкретной культивируемой клеточной линии. Пригодной средой для культур насекомых является среда Грейса (Grace's) с добавкой автолизата дрожжей Yeastolate, гидролизата лактальбумина и/или фетальной телячьей сыворотки, при необходимости.
Типично, антибиотик или другое соединение, пригодное для селективного выращивания трансформированных клеток, прибавляют к среде в качестве добавки. Какое соединение должно быть использовано - будет определяться селектируемым маркерным элементом, присутствующим в плазмиде, которой трансформирована клетка-хозяин. Например, в тех случаях, когда селектируемым маркерным элементом является резистентность к канамицину, прибавляемое к культуральной среде соединение будет канамицином. Другие соединения для селективного выращивания включают ампициллин, тетрациклин и неомицин.
Количество полипептида OspA, продуцируемого клеткой-хозяином, можно оценить с помощью стандартных способов, известных специалистам. Такие способы включают, без ограничений, анализ способом вестерн-блоттинга, электрофорез на полиакриламидном геле с добавкой ДСН, электрофорез в неденатурирующем геле, хроматографическое разделение, такое как высокоэффективная жидкостная хроматография (ВЭЖХ), иммунодетектирование, такое как иммунопреципитация, и/или анализы активности, такие как анализы связывания ДНК способом задержки в геле (gel shift assays).
В некоторых случаях, полипептид OspA не является биологически активным после выделения, и различные способы «переукладки» или перевода полипептида в его третичную структуру и создания дисульфидных связей используются для восстановления биологической активности. Такие способы включают воздействие на солюбилизированный полипептид значениями pH обычно выше 7 и в присутствии определенной концентрации хаотропного агента. Выбор хаотропного агента очень похож на выбор вариантов, используемых для солюбилизации телец включения, но обычно хаотропный агент используется в меньших концентрациях и не обязательно совпадает с хаотропными агентами, используемыми для солюбилизации. В некоторых случаях, раствор для переукладки/оксидации также содержит восстановитель или восстановитель плюс его окисленная форма в определенном соотношении для создания определенного значения окислительно-восстановительного потенциала, позволяющего проводить перетасовку дисульфидных связей при образовании цистеинового мостика (мостиков) белка. Некоторые из широко используемых окислительно-восстановительных пар включают цистеин/цистамин, глутатион (GSH)/дитиобис-GSH, хлорид меди, дитиотреит (DTT)/дитиан-DTT и 2-2-меркаптоэтанол (bME)/дитио-b(ME). Сорастворитель часто используется для увеличения эффективности переукладки, и наиболее часто используемые с этой целью реагенты включают глицерин, полиэтиленгликоль различного молекулярного веса, аргинин и т.п.
Если тельца включения не образуются в значительных количествах при экспрессии полипептида OspA, то после центрифугирования клеточного гомогената полипептид будет находиться преимущественно в супернатанте. Полипептид затем выделяют из супернатанта с помощью способов, описанных тут, или других способов, известных специалистам.
Очистка полипептида OspA из раствора может быть проведена с использованием различных методик, известных специалистам. Если полипептид был синтезирован так, чтобы он содержал метку, такую как гексагистидин (полипептид OspA/hexaHis) или другой малый пептид, такой как FLAG (Eastman Kodak Co., New Haven, CT) или myc (Invitrogen, Carlsbad, CA) на любом из его карбоксильного или амино-концов, полипептид часто очищают одностадийным способом, путем пропускания раствора через аффинную колонку, где матрикс колонки имеет высокую аффинность к метке. Например, полигистидин связывается с высокой аффинностью и специфичностью с никелем; таким образом, аффинная колонка с никелем (такая как никелевые колонки Qiagen®) может быть использована для очистки полипептида OspA/polyHis. См., например, Ausubel et al., eds., Current Protocols in Molecular Biology, Section 10.11.8, John Wiley & Sons, New York (1993).
Дополнительно, полипептид OspA может быть очищен путем использования моноклонального антитела, способного специфически распознавать и связываться с полипептидом OspA. Таким образом, пригодные процедуры очистки включают, без ограничений, аффинную хроматографию, иммуноаффинную хроматографию, ионообменную хроматографию, молекулярно-ситовую хроматографию, высокоэффективную жидкостную хроматографию (ВЭЖХ), электрофорез (включая нативный гелевый электрофорез) с последующим элюированием геля и препаративным изоэлектрическим фокусированием (аппарат/методика «Isoprime», Hoefer Scientific, San Francisco, CA). В некоторых случаях, комбинируют две или более методики очистки для достижения повышенной степени чистоты.
Полипептиды OspA также получают способами химического синтеза (такими как твердофазовый пептидный синтез) с использованием способов, известных специалистам, таких как описанные Merrifield et al., J. Am. Chem. Soc., 85:2149 (1963), Houghten et al., Proc. Natl. Acad. Sci. USA, 82:5132 (1985), и Stewart and Young, «Solid Phase Peptide Synthesis», Pierce Chemical Co., Rockford, IL (1984). Такие полипептиды синтезируют с метионином на амино-конце или без него. Химически синтезированные полипептиды OspA, в некоторых аспектах, окисляют с использованием способов, описанных в этих первоисточниках, с образованием дисульфидных мостиков. Ожидается, что химически синтезированные полипептиды OspA будут иметь сравнимую биологическую активность с соответствующими полипептидами OspA, полученными рекомбинантными способами или очисткой из природных источников и, таким образом, часто используются взаимозаменяемо с рекомбинантным полипептидом OspA. Подразумевается, что специалистам известен ряд дополнительных способов получения нуклеиновых кислот и полипептидов, и такие способы могут быть использованы для продуцирования полипептидов OspA.
Химические производные молекул полипептида OspA
Химически модифицированные производные полипептидов OspA могут быть получены квалифицированным специалистом в данной области техники с учетом приведенного ниже описания. Производные полипептида OspA модифицируют способами, различающимися по типу или положению молекул, в естественных условиях присоединенных к полипептиду. Производные, в некоторых аспектах, включают молекулы, образующиеся в результате удаления одной или более присоединенных в естественных условиях химических групп. Полипептид, содержащий аминокислотную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173, или вариант полипептида OspA, в одном аспекте, модифицируют путем ковалентного присоединения одного или более полимеров. Например, выбранный полимер типично является водорастворимым, чтобы белок, к которому он присоединен, не выпадал в осадок в водной среде, такой как физиологическая среда. Пригодные полимеры включают смесь полимеров. В определенных аспектах, для терапевтического применения готового препарата, полимер будет фармацевтически приемлемым.
Каждый из полимеров, в различных аспектах, может иметь любой молекулярный вес и быть разветвленным или неразветвленным. Каждый полимер типично имеет средний молекулярный вес от примерно 2 кДа до примерно 100 кДа (термин «примерно» указывает, что в препаратах водорастворимого полимера некоторые молекулы будут весить больше, а некоторые меньше, чем указанный молекулярный вес). Средний молекулярный вес каждого полимера, в различных аспектах, составляет от примерно 5 кДа до примерно 50 кДа, от примерно 12 кДа до примерно 40 кДа и от примерно 20 кДа до примерно 35кДа.
Пригодные водорастворимые полимеры или их смеси включают, без ограничений, N-связанные или O-связанные углеводы; сахара; фосфаты; полиэтиленгликоль (ПЭГ) (включая формы ПЭГ, используемые для дериватизации белков, включая моно-(C1-C10)-алкокси- или арилокси-полиэтиленгликоль); монометоксиполиэтиленгликоль; декстран (такой как низкомолекулярный декстран, например, примерно 6 кДа); целлюлозу; или другие полимеры на основе углеводов, поли-(N-винилпирролидон) полиэтиленгликоль, гомополимеры пропиленгликоля, сополимер полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин) и поливиниловый спирт. Настоящее изобретение также охватывает бифункциональные сшивающие молекулы, которые иногда используются для получения ковалентно присоединенных мультимеров полипептида, содержащего аминокислотную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173, или вариант полипептида OspA.
В некоторых аспектах, химическая дериватизация осуществляется в любых пригодных условиях, используемых для проведения реакции белка с активированной полимерной молекулой. Способы получения химических производных полипептидов в общем включают стадии (а) проведения реакции полипептида с активированной полимерной молекулой (такой как реакционноспособное сложноэфирное или альдегидной производное полимерной молекулы) в условиях, при которых полипептид, содержащий аминокислотную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 или 173, или вариант полипептида OspA, присоединяется к одной или более полимерных молекул, и (б) получения продукта (продуктов) реакции. Оптимальные реакционные условия определяют на основании известных параметров и желательного результата. Например, чем больше величина отношения полимерные молекулы: белок, тем выше процентная доля присоединенных полимерных молекул. В одном варианте воплощения производное полипептида OspA содержит один фрагмент полимерной молекулы на аминоконце (см., например, патент США № 5234784).
Пегилирование полипептида, в определенных аспектах, конкретнее осуществляют с помощью любой из реакций пегилирования, известных специалистам, как описано, например, в следующих источниках: Francis et al., Focus on Growth Factors, 3:4-10 (1992); EP 0154316; EP 0401384 и патент США № 4179337. Например, пегилирование проводят по реакции ацилирования или реакции алкилирования с реакционноспособной молекулой полиэтиленгликоля (или аналогичного реакционноспособного водорастворимого полимера), как описано тут. Для реакций ацилирования, выбранный полимер(ы) должен иметь одну реакционноспособную сложноэфирную группу. Для восстановительного алкилирования, выбранный полимер(ы) должен иметь одну реакционноспособную альдегидную группу. Реакционноспособным альдегидом является, например, полиэтиленгликоль-пропиональдегид, который является водостойким, или его моно-C1-C10-алкокси- или -арилокси-производные (см. патент США № 5252714).
В другом варианте воплощения полипептиды OspA химически присоединяют к биотину, и конъюгированным молекулам биотин/полипептид OspA затем позволяют связываться с авидином с образованием тетравалентных молекул авидин/биотин/полипептид OspA. Полипептиды OspA также ковалентно присоединяют к динитрофенолу (DNP) или тринитрофенолу (TNP) и полученные конъюгаты осаждают анти-DNP или анти-TNP-IgM с образованием декамерных конъюгатов с валентностью 10. Производные полипептида OspA, раскрытые тут, в определенных аспектах, обладают дополнительными активностями, повышенной или пониженной биологической активностью, или другими характеристиками, такими как увеличенный или уменьшенный период полувыведения, по сравнению с недериватизированными молекулами.
Иммуногенные композиции, вакцины и антитела
Некоторые аспекты изобретения включают иммуногенные композиции и вакцины. Иммуногенные химерные молекулы OspA по изобретению используются в комбинации в качестве антигена (антигенов), вызывающего анти-OspA иммунный ответ у субъекта (т.е., действуют как вакцина). Типичные примеры иммуногенных полипептидов OspA (SEQ ID NO: 2, 4, 6, 169, 171 и 173) вводятся в комбинации для того, чтобы вызвать иммунный ответ на любой один или более серотипов 1-6 Borrelia и, в более общем случае, на многие другие виды Borrelia, описанные тут. Иммунный ответ также может быть вызван путем доставки плазмидных векторов, кодирующих полипептиды OspA по изобретению (т.е., введение «голых ДНК»). В некоторых аспектах, молекулы нуклеиновой кислоты OspA (SEQ ID NO: 1, 3, 5, 168, 170 и 172) вводятся инъекцией, с помощью липосом, или другими средствами введения, описанными тут. После иммунизации, субъект вырабатывает усиленный иммунный ответ против белка OspA серотипов 1-6 Borrelia и против других видов Borrelia.
Таким образом, как описано выше, как полипептиды OspA, так и молекулы нуклеиновой кислоты OspA включаются в качестве антигенов для использования в иммуногенных композициях и/или композициях вакцин по изобретению. В определенных аспектах субъекту вводятся как нуклеиновая кислота, так и белок. В конкретных аспектах, предлагается усиливать иммунный ответ на вакцину на основе нуклеиновой кислоты путем одновременного введения когнатного белка (см. WO 99/30733). Нуклеиновая кислота и белок не должны обязательно вводиться в одной и той же композиции. Оба просто должны быть введены во время фазы индукции иммунного ответа белком, в некоторых аспектах, замаскированной или задержанной до тех пор, пока нуклеиновая кислота не сенсибилизирует иммунную систему. В конкретном аспекте, вакцины предназначены для доставки нуклеиновой кислоты и белкового антигена в антигенпрезентирующие клетки (см. WO 97/28818). В различных аспектах, нуклеиновая кислота и белок закомплексованы, например, путем ковалентной конъюгации. В дополнительных аспектах, для усиления иммуногенности антигенов вакцины также включают липосомальные композиции.
В определенных аспектах, иммуногенная композиция по изобретению включает любую одну или более молекул OspA, описанных тут, в сочетании с фармацевтическим носителем, причем композиция индуцирует продуцирование антитела, которое специфически связывается с внешним поверхностным белком A (OspA). В некоторых аспектах, иммуногенная композиция также содержит стабилизатор или антимикробный консервант. В конкретных аспектах, иммуногенная композиция индуцирует продуцирование антитела, которое специфически связывается с Borrelia. В других аспектах, композиция индуцирует продуцирование антитела, нейтрализующего Borrelia.
В некоторых аспектах, изобретение включает использование адъювантов в иммуногенных композициях, содержащих химерные молекулы OspA (антигены), описанные тут. В определенных аспектах, иммуногенность значительно усиливается, если антиген вводится совместно с адъювантом. В некоторых аспектах, адъювант используется в виде 0,001%-50% раствора в фосфатно-солевом буфере (PBS). Адъюванты усиливают иммуногенность антигена, но сами не обязательно являются иммуногенными.
Адъюванты, в различных аспектах, оказывают ряд позитивных эффектов на вакцинацию. В некоторых случаях, адъюванты ускоряют выработку сильного иммунного ответа у субъектов. Адъюванты, в других случаях, повышают уровень иммунного ответа, пролонгируют его длительность и улучшают иммунную память. Адъюванты часто используются для борьбы с ослабленным иммунитетом в определенных группах субъектов (например, у лиц пожилого возраста или пациентов с иммунной суппрессией) или для усиления иммуногенности в конкретной «группе риска» (такой как, без ограничений, очень молодые или лица пожилого возраста). Эффекты иммунного усиления адъюванта, в различных случаях, приводят к снижению требуемого количества антигена в готовой композиции для получения защитного ответа (т.е. уменьшению дозы).
В общем, адъюванты классифицируют по их преимущественному механизму действия на две основные группы: Первая группа представляет собой агонисты рецепторов или сенсоры врожденной иммунной системы, такие как агонисты Toll-подобного рецептора (TLR), агонисты лектинового рецептора C-типа, агонисты (индуцируемый ретиноевой кислотой ген-1 (RIG-1))-подобного рецептора (RLR) и агонисты (нуклеотид-связывающий домен и лейцин-богатый повтор)-содержащего рецептора (NLR). Вторая группа представляет собой вещества, действующие как системы доставки, также известные как TLR-независимые адъюванты. Примерами адъювантов TLR-агонистов являются ASO4 (Glaxo Smith Kline), TLR-4 агонист, используемый в качестве адъюванта в коммерческих вакцинах против гепатита B и вируса папилломы; Vaxinate, агонист флагеллин-слитого белка TLR-5; и многочисленные адъюванты TLR-9 агонистов, такие как использующие двухцепочечную ДНК (дцДНК) и олигонуклеотиды CpG или ODN1a. Другие TLR-агонисты, попадающие в эту категорию адъювантов, включают гликолипиды (TLR-1), липотейхоевую кислоту и липопротеин (TLR-1/TLR-2 и TLR-2/TLR-6) липополисахарид, липоолигосахариды и монофосфорил липид A (MPL) (TLR-4), двухцепочечную РНК (TLR-3); пептидогликан (TLR-6), одноцепочечную РНК (TLR-7). Примеры двух адъювантов агониста лектинового рецептора C-типа включают ß-глюканы (Dectin-1) и маннаны (Dectin-2), оба выделенные из клеточных оболочек грибов. Адъюванты агониста RLR-рецептора включают одноцепочечные вирусные РНК и двухцепочечные вирусные ДНК, в то время как адъюванты NLR-агониста включают продукты деградации пептидогликана, микробные продукты и неинфекционные кристаллические частицы. Во всех случаях, агонисты действуют путем прямой активации рецептора врожденной иммунной системы для запуска, усиливающего иммунитет воспалительного ответа. Вторая группа адъювантов, TLR-независимые адъюванты, преимущественно действуют как системы доставки и усиливают поглощение антигена и его презентацию антигенпрезентирующей клеткой. В некоторых случаях, такие адъюванты могут также действовать путем локального удерживания антигена вблизи места введения для продуцирования эффекта депо, способствующего медленному пролонгированному выделению антигена в клетки иммунной системы. Адъюванты также привлекают клетки иммунной системы к депо антигена и стимулируют такие клетки для того, чтобы вызвать иммунные ответы. Примеры TLR-независимых адъювантов включают минеральные соли, такие как гидроксид алюминия и фосфат алюминия (коллективно называемые квасцами) и фосфат кальция; эмульсию типа масло-в-воде (например, MF59, AS03 и ProVax); эмульсию типа вода-в-масле (Montanide, TiterMax); биополимеры (Advax); растительные производные, особенно фракции сапонина, тритерпеноидный экстракт коры южноамериканского мыльного дерева Quillaja saponaria Molina (SFA-1, QS21, Quil A); иммуностимулирующие комплексы (ISCOM и ISCOM matrix), состоящие из фракций сапонина, стерола и, необязательно, фосфолипидов (ISCOMATRIX и Matrix-M); липосомы, представляющие собой фосфолипидные сферы различного размера и заряда (Vaxfectin и Vaxisome); вирусоподобные частицы и виросомы, представляющие собой липосомы, содержащие вирусные поверхностные антигены, такие как гемагглютинин и нейраминидаза Influenza; наночастицы различного состава; хитозан, пептиды, такие как полиаргинин и пептид, известный как пептид KLK.
Адъюванты, перечисленные выше, используются по одному или в комбинации. Комбинации TLR-зависимых и TLK-независимых адъювантов часто являются предпочтительными для использования в качестве антигена, и считается, что TLR-зависимые адъюванты транспортируются к антигенпрезентирующим клеткам TLR-независимым адъювантом, который также будет стимулировать поглощение и стабильность, в то время как TLR-зависимый адъювант будет непосредственно усиливать иммунитет путем активации сигнализации TLR.
Примеры комбинаций TLR-зависимых и TLR-независимых адъювантов включают AS01: смесь MPL (агонист TLR-4), липосомы и QS-21 (оба - TLR-независимые адъюванты); AS04: MPL (агонист TLR-4) и гидроксид алюминия/фосфат; IC31: ODN1a (агонист TLR-9) и пептид KLK (TLR-независимый адъювант); и полный адъювант Фрейнда, экстракт мембран Mycobacterium tuberculosis (агонист TLR-4) и эмульсию типа масло-в-воде (TLR-независимый адъювант).
Комбинации, состоящие из множества TLR-зависимых адъювантов, также используются для максимизации иммуноусиливающего эффекта композиций вакцин с адъювантами. Агонисты TLR, использующие разные адапторные белки, часто комбинируют (например, комбинация агониста к связанному с плазматической мембраной рецептору TLR-3 или TLR-4, использующему TRIF (Toll/интерлейкин 1 рецепторный домен-содержащий адапторный белок, индуцирующий INF-ß) адапторный путь с агонистом TLR (TLR-7, TLR-8 и TLR-9), которые экспрессируются в эндосомальных или лизосомальных органеллах и используют путь адапторного белка MyD88 (белок первичного ответа миелоидной дифференциации).
Такие иммуностимулирующие агенты или адъюванты также усиливают иммунный ответ хозяина на вакцины. В некоторых случаях такие вещества как липополисахариды могут выступать в роли внутренних адъювантов, поскольку они нормально являются компонентами убитых или аттенуированных бактерий, используемых в качестве вакцины. Внешние адъюванты, такие как перечисленные выше, являются иммуномодуляторами, которые типично нековалентно связаны с антигенами и включаются в рецептуру композиции для усиления иммунного ответа хозяина.
Широкий спектр внешних адъювантов может вызывать сильные иммунные ответы на антигены. Они включают сапонины в виде комплексов с антигенами мембранных белков (иммуностимулирующие комплексы), полимеры Pluronic с минеральным маслом, убитые микобактерии в минеральном масле, полный адъювант Фрейнда, бактериальные продукты, такие как мурамилдипептид (MDP) и липополисахарид (LPS), а также липид A и липосомы. Для эффективного индуцирования гуморального иммунного ответа (HIR) и клеточно-медиируемого иммунитета (CMI), иммуногены, в определенных аспектах, эмульгируют в адъювантах.
Желательные характеристики идеальных адъювантов включают любые или все из: отсутствия токсичности; способности стимулировать длительный иммунный ответ; простоты производства и стабильности при длительном хранении; способности вызывать как CMI, так и HIR к антигенам, введенным различными путями; синергии с другими адъювантами; способности к селективному взаимодействию с популяциями антигенпрезентирующих клеток (APC); способности специфически вызывать соответствующие TH1 или TH2 клеточно-специфические иммунные ответы; и способности селективно повышать уровни соответствующего изотипа антитела (например, IgA) против антигенов.
Патент США № 4855283, включенный сюда в качестве ссылки, описывает для этих же целей гликолипидые аналоги, включая N-гликозиламиды, N-гликозилмочевины и N-гликозилкарбаматы, каждый из которых замещен в остатке сахара аминокислотой, в качестве иммунномодуляторов или адъювантов. Патент США № 4855283 сообщает, что N-гликолипидные аналоги, демонстрирующие структурное сходство с природными гликолипидами, такие как гликосфинголипиды и гликоглицеролипиды, способны вызывать сильные иммунные ответы в вакцинах вируса простого герпеса и вируса псевдобешенства. Некоторые гликолипиды были синтезированы из длинноцепочечных алкиламинов и жирных кислот, связанных непосредственно с сахаром через аномерный атом углерода, для имитации функций природных липидных остатков.
В некоторых аспектах, иммуногенная композиция содержит количество адъюванта, достаточное для усиления иммунного ответа на иммуноген. Пригодные адъюванты включают, без ограничений, соли алюминия (фосфат алюминия или гидроксид алюминия), смеси сквалена (SAF-1), мурамилпептид, производные сапонина, препараты клеточных мембран микобактерий, монофосфорил липид A, производные миколовой кислоты, неионные блок-coполимерные поверхностно-активные вещества, Quil A, B-субъединицу холерного токсина, полифосфазен и производные, и иммуностимулирующие комплексы (ISCOM), такие как описанные Takahashi et al. (Nature 344:873-875, 1990). В некоторых аспектах, адъювант представляет собой синтетический адъювант. В конкретном аспекте, синтетический адъювант является глюкопиранозил-липидным адъювантом (GLA).
Дополнительным аспектом изобретения является вакцина, содержащая иммуногенную композицию по изобретению и фармацевтически приемлемый носитель. Как было описано выше, вакцина, в определенных аспектах, включает один или более стабилизаторов и/или один или более консервантов.
В одном аспекте, предусматривается вакцина, содержащая по меньшей мере один рекомбинантный экспрессионный конструкт, который включает промотор, функционально связанный с последовательностью нуклеиновой кислоты, кодирующей антиген (химерный полипептид OspA, описанный тут) и адъювант. В одном варианте воплощения рекомбинантный экспрессионный конструкт (экспрессионный вектор, содержащий полинуклеотид OspA) присутствует в вирусном векторе, который в определенных дополнительных вариантах воплощения присутствует в вирусе, выбранном из аденовируса, адено-ассоциированного вируса, герпесвируса, лентивируса, поксвируса и ретровируса.
Дополнительные аспекты изобретения включают антитела к химерным молекулам OspA, описанным тут. В различных аспектах, изобретение включает химерные молекулы OspA для получения анти-OspA антител и для обеспечения иммунитета от инфекции Borrelia. В некоторых аспектах, такие анти-OspA антитела, например, мышиные, человеческие, или гуманизированные моноклональные антитела или одноцепочечные антитела, вводятся субъекту (например, пассивная иммунизация) для того, чтобы вызвать иммунный ответ против белка OspA любого одного или более серотипов 1-6 Borrelia. В используемом тут значении, термин «антитела» относится к молекуле, которая обладает специфичностью по отношению к одному или более полипептидов OspA. Пригодные антитела получают с использованием способов, известных специалистам. В определенных аспектах, антитело OspA способно связывать определенный участок полипептида OspA, тем самым ингибируя связывание полипептида с рецептором (рецепторами) полипептида OspA. Антитела и фрагменты антител, которые связывают химерные полипептиды OspA по изобретению, входят в объем настоящего изобретения.
В некоторых аспектах, антитела по изобретению включают антитело или его фрагмент, которые специфически связываются с одним или более полипептидов OspA, продуцируемых путем иммунизации животного полипептидом, содержащим аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 169, 171 и 173. В других аспектах, изобретение включает антитело или его фрагмент, которые специфически связываются с полипептидом, кодируемым последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 1, 3, 5, 7, 9, 11, 168, 170 и 172. В различных аспектах, антитело или его фрагмент являются человеческими, гуманизированными, поликлональными или моноклональными. В дополнительных аспектах, антитело представляет собой антитело Fab или Fab'. В конкретных аспектах, антитело содержит детектируемую метку. В некоторых аспектах, антитело представляет собой химически модифицированное производное антитела.
Введение химерных молекул OspA в соответствии с изобретением стимулирует иммунный или гуморальный ответ у людей или животных. В некоторых аспектах, три химерные молекулы OspA (например, липидированный OspA 1/2251, липидированный OspA 6/4 OspA и липидированный OspA 5/3; или исходный OspA 1/2, исходный OspA 6/4 и исходный OspA 5/3) вводятся вместе для того, чтобы вызвать гуморальный ответ против всех шести серотипов (1-6), описанных тут. Этот гуморальный ответ означает, что способы по изобретению, в различных аспектах, используются просто для стимулирования иммунного ответа (в отличие от обеспечения также защитного ответа), потому, что полученные антитела (без защиты), тем не менее, являются полезными. Из полученных антител, хорошо известными специалистам способами, получают моноклональные антитела; и используют такие моноклональные антитела в хорошо известных анализах связывания антител, диагностических наборах или тестах для определения присутствия или отсутствия Borrelia burgdorferi s.l., или для определения того, был ли иммунный ответ на спирохету просто стимулированным. Моноклональные антитела, в определенных аспектах, используются в иммуноадсорбционной хроматографии для извлечения или выделения антигенов Borrelia, таких как OspA.
Антитела OspA по изобретению, в различных аспектах, являются поликлональными, включая моноспецифические поликлональные, моноклональные (MAb), рекомбинантные, химерные, гуманизированные, такие как CDR-привитые, человеческие, одноцепочечные и/или биспецифические, а также их фрагментами, вариантами или производными. Фрагменты антитела включают участки антитела, которые связываются с эпитопом полипептида OspA. Примеры таких фрагментов включают фрагменты Fab и F(ab’), получаемые ферментативным расщеплением полноразмерных антител. Другие связывающие фрагменты включают получаемые способами рекомбинантных ДНК, такими как экспрессия рекомбинантных плазмид, содержащих последовательности нуклеиновой кислоты, кодирующей вариабельные области антитела.
Поликлональные антитела, направленные против полипептида OspA, в общем, продуцируются у субъекта (включая кроликов, мышей или другое животное или млекопитающее) с помощью множества подкожных, внутримышечных или интраперитонеальных инъекций полипептида OspA и адъюванта. В определенных аспектах является полезным конъюгировать полипептид OspA по изобретению с белком-носителем, иммуногенным в иммунизируемом виде, таким как гемоцианин моллюска блюдечко, сывороточный альбумин, бычий тиреоглобулин, или соевый ингибитор трипсина. Для усиления иммунного ответа также используются адъюванты, такие как квасы. После иммунизации берут образцы крови у иммунизированного субъекта и сыворотку оценивают на титр антител против полипептида OspA.
Моноклональные антитела, направленные против полипептида OspA, продуцируются с использованием любого способа, обеспечивающего продуцирование молекул антитела непрерывными клеточными линиями в культуре. Примеры пригодных способов получения моноклональных антител включают гибридомные способы Kohler et al., Nature, 256:495-497 (1975) и способ человеческой B-клеточной гибридомы, Kozbor, J. Immunol., 133:3001 (1984), и Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987). Изобретение также предусматривает гибридомные клеточные линии, продуцирующие моноклональные антитела, реакционноспособные по отношению к полипептидам OspA.
Моноклональные антитела по изобретению, в некоторых случаях, модифицируют для использования в качестве терапевтических средств. Один из вариантов воплощения представляет собой «химерное» антитело, в котором часть тяжелой и/или легкой цепи идентична с или гомологична соответствующей последовательности антител, полученных от определенного вида или принадлежащих к определенному классу или подклассу антител, в то время как остальная цепь (цепи) идентична (идентичны) с или гомологична (гомологичны) соответствующей последовательности антител, полученных от другого вида, или принадлежащей к другому классу или подклассу антител. Также включены фрагменты таких антител, при условии, что они проявляют желательную биологическую активность. См. патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1985).
В другом варианте воплощения моноклональное антитело по изобретению представляет собой «гуманизированное» антитело. Способы гуманизации не-человеческих антител хорошо известны специалистам (см. патенты США №№ 5585089 и 5693762). В общем, гуманизированное антитело содержит один или более аминокислотных остатков, введенных в него из источника, не принадлежащего человеку. Гуманизация может быть осуществлена, например, с использованием способов, описанных в литературе (Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-327 (1988); Verhoeyen et al., Science 239:1534-1536 (1988)), путем замещения по меньшей мере частью гипервариабельного участка (CDR) грызуна соответствующих областей человеческого антитела.
В альтернативном варианте воплощения человеческие антитела продуцируются из библиотеки фагового дисплея (Hoogenboom et al., J. Mol. Biol. 227:381 (1991) и Marks et al., J. Mol. Biol. 222:581 (1991)). Эти способы имитируют иммунную идентификацию путем экспонирования репертуаров антител на поверхности нитчатого бактериофага и последующей селекции фагов по их связыванию с выбранным антигеном. Одна такая методика описана в заявке PCT № PCT/US98/17364 (Adams et al.), которая описывает выделение высокоаффинных и функционально агонистических антител к MPL- и msk-рецепторам с использованием такого подхода.
Химерные, CDR-привитые и гуманизированные антитела типично продуцируют рекомбинантными способами. Нуклеиновые кислоты, кодирующие антитела, вводятся в клетки-хозяева и экспрессируются с использованием материалов и процедур, описанных тут, или известных специалистам. В одном варианте воплощения антитела продуцируются в клетках-хозяевах млекопитающего, таких как клетки CHO. Моноклональные (например, человеческие) антитела, в различных аспектах, продуцируются путем экспрессии рекомбинантной ДНК в клетках-хозяевах или путем экспрессии в гибридомных клетках, как описано тут. В некоторых аспектах, моноклональное антитело или его фрагмент является гуманизированным. В конкретном аспекте, моноклональное антитело представляет собой F237/BK2, как описано тут.
В определенных аспектах, изобретение включает способы профилактики или лечения инфекции Borrelia или лаймской болезни у субъекта, включающие стадию введения субъекту антитела или его фрагмента, как описано тут, в количестве, обеспечивающем эффективную профилактику или лечение инфекции Borrelia или лаймской болезни. В конкретных аспектах, антитело или его фрагмент представляет собой гипериммунную сыворотку, гипериммунную плазму или его очищенную иммуноглобулиновую фракцию. В других аспектах, антитело или его фрагмент представляет собой очищенный препарат иммуноглобулина или препарат иммуноглобулинового фрагмента.
Анти-антитела OspA по изобретению, в различных аспектах, используются в любом известном способе анализа, таком как конкурентные анализы связывания, прямые и непрямые сэндвич-анализы и анализы иммунопреципитации (Sola, Monoclonal Antibodies: A Manual of Techniques, pp. 147-158 (CRC Press, Inc., 1987)) для детектирования и количественного определения полипептидов OspA. Антитела будут связывать полипептиды OspA с аффинностью, соответствующей используемому способу анализа.
Для диагностического или клинического применения, в определенных вариантах воплощения анти-антитела OspA метят детектируемым фрагментом. Детектируемый фрагмент может быть любым, способным продуцировать, прямо или опосредованно, детектируемый сигнал. Например, в определенных аспектах, детектируемый фрагмент представляет собой радиоактивный изотоп, такой как 3H, 14C, 32P, 35S или 125I; флуоресцентное или хемолюминесцентное соединение, такое как флуоресцеинизотиоцианат, родамин или люциферин; или фермент, такой как щелочная фосфатаза, β-галактозидаза, или пероксидаза хрена (Bayer et al., Meth. Enzym. 184:138-163 (1990)).
Конкурентные анализы связывания основаны на способности меченого стандарта (например, полипептида OspA, или его иммунологически реакционноспособного участка) конкурировать с исследуемым образцом аналита (полипептид OspA) за связывание с ограниченным количеством анти-OspA антитела. Количество полипептида OspA в исследуемом образце обратно пропорционально количеству стандарта, связывающегося с антителами. Для облегчения определения количества связываемого стандарта, антитела типично переводят в нерастворимую форму перед или после конкурентного анализа, так чтобы стандарт и аналит, связанные с антителом, можно было удобно отделить от остающихся несвязанными стандарта и аналита.
Сэндвич-анализы типично предусматривают использование двух антител, каждое из которых способно связываться с разными иммуногенными участками, или эпитопами, детектируемого и/или количественно определяемого белка. При сэндвич-анализе, исследуемый образец аналита типично связывается первым антителом, которое иммобилизовано на твердом носителе, и после этого второе антитело связывается с аналитом, тем самым образуя нерастворимый трехкомпонентный комплекс. См., например, патент США № 4376110. Второе антитело, в некоторых случаях, является меченым детектируемым фрагментом (прямые сэндвич-анализы) или измеряется с помощью анти-иммуноглобулинового антитела, меченного детектируемым фрагментом (непрямые сэндвич-анализы). Например, одним из типов сэндвич-анализа является твердофазовый иммуноферментный анализ (ELISA), в этом случае детектируемый фрагмент представляет собой фермент.
Анти-OspA антитела также пригодны для in vivo визуализации. Антитело, меченное детектируемым фрагментом, в определенных аспектах, вводят животному в кровоток и оценивают присутствие и местонахождение меченого антитела в хозяине. Антитело, в различных аспектах, метят любым фрагментом, являющимся детектируемым у животного способами ядерного магнитного резонанса, радиологии или другими способами детектирования, известными специалистам. В некоторых аспектах изобретения, антитела OspA используются в качестве терапевтических средств.
Композиции химерного OspA и их введение
Для введения субъектам химерных полипептидов OspA, описанных тут, готовят композиции полипептидов OspA, содержащие один или более фармацевтически приемлемых носителей. Фраза «фармацевтически или фармакологически приемлемый» относится к молекулярным частицам и композициям, которые не вызывают аллергических или других нежелательных реакций при введении путями, хорошо известными специалистам, как описано ниже. «Фармацевтически приемлемые носители» включают любые из и все клинически пригодные растворители, дисперсионные среды, покрытия, антибактериальные и антигрибковые агенты, изотонические и замедляющие всасывание агенты и т.п. В некоторых аспектах, композиция образует сольваты с водой или обычными органическими растворителями. Такие сольваты также включены в настоящее изобретение.
Иммуногенная композиция или композиция вакцины по изобретению, в различных аспектах, вводится перорально, местно, трансдермально, парентерально, с помощью спрея для ингаляции, вагинально, ректально или с помощью внутричерепной инъекции. Термин «парентеральный» в используемом тут значении включает подкожные инъекции, внутривенные, внутримышечные, интрацистернальные инъекции, или инфузионные способы. Также предусматривается введение с помощью внутривенной, интрадермальной, внутримышечной, интрамаммарной, интраперитонеальной, интратекальной, ретробульбарной, внутрилегочной инъекции и или хирургической имплантации в определенное место. В общем, композиции по существу не содержат пирогенов, а также других примесей, которые могли бы быть вредными для реципиента.
Рецептура фармацевтической композиции будет меняться в зависимости от выбранного пути введения (например, раствор, эмульсия). Пригодную композицию, содержащую предназначенную для введения композицию, готовят в физиологически приемлемом растворителе или носителе. Для растворов или эмульсий, пригодные носители включают, например, водные или спиртово-водные растворы, эмульсии или суспензии, включая солевые и забуференные среды. Парентеральные носители, в некоторых аспектах, включают раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия, лактатный раствор Рингера или нелетучие масла. Внутривенные носители, в определенных аспектах, включают различные добавки, консерванты, или жидкости, питательные вещества или средства для восстановления электролитного баланса.
Фармацевтические композиции, пригодные для использования в соединениях и способах по настоящему изобретению, включающих полипептиды OspA в качестве активного ингредиента, содержат, в различных аспектах, фармацевтически приемлемые носители или добавки, определяемые в зависимости от пути введения. Примеры таких носителей или добавок включают воду, фармацевтически приемлемый органический растворитель, коллаген, поливиниловый спирт, поливинилпирролидон, карбоксивинильный полимер, карбоксиметилцеллюлозу натрия, полиакрилат натрия, альгинат натрия, водорастворимый декстран, карбоксиметилкрахмал натрия, пектин, метилцеллюлозу, этилцеллюлозу, ксантановую камедь, гуммиарабик, казеин, желатин, агар, диглицерин, глицерин, пропиленгликоль, полиэтиленгликоль, вазелин, парафин, стеариловый спирт, стеариловую кислоту, человеческий сывороточный альбумин (HSA), маннит, сорбит, лактозу, фармацевтически приемлемое поверхностно-активное вещество и т.п. Используемые добавки выбирают, без ограничений, из вышеперечисленных материалов или их комбинаций, при необходимости, в зависимости от лекарственной формы по настоящему изобретению.
Различные водные носители, например, вода, забуференная вода, 0,4% солевой раствор, 0,3% глицин, или водные суспензии содержат, в различных аспектах, активное соединение в смеси с эксципиентами, пригодными для производства водных суспензий. Такие эксципиенты представляют собой суспендирующие агенты, например, натрия карбоксиметилцеллюлоза, метилцеллюлоза, гидроксипропилметилцеллюлоза, альгинат натрия, поливинилпирролидон, трагакантовая камедь и аравийская камедь; диспергирующие или смачивающие агенты, в некоторых случаях, природный фосфатид, например, лецитин, или продукты конденсации алкиленоксида с жирными кислотами, например, полиоксиэтиленстеарат, или продукты конденсации этиленоксида с длинноцепочечными алифатическими спиртами, например, гептадекаэтиленоксицетанол, или продукты конденсации этиленоксида с неполными сложными эфирами жирных кислот и гексита, такими как полиоксиэтиленсорбитмоноолеат, или продукты конденсации этиленоксида с неполными сложными эфирами жирных кислот и ангидридов гексита, например, полиэтиленсорбитанмоноолеат. Водные суспензии, в некоторых аспектах, содержат один или более консервантов, например, этил-, или н-пропил, п-гидроксибензоат.
В некоторых аспектах, композиции OspA лиофилизируют для хранения и восстанавливают в пригодном носителе перед использованием. Эффективность этой методики была продемонстрирована на обычных иммуноглобулинах. Используются любые пригодные методики лиофилизации и восстановления, известные специалистам. Квалифицированным специалистам в данной области техники понятно, что лиофилизация и восстановление приводят к различным степеням потери активности антител, и что уровни использования часто регулируют в целях компенсации.
Диспергируемые порошки и гранулы, пригодные для получения водных суспензий путем прибавления воды, дают активное соединение в смеси с диспергирующим или смачивающим агентом, суспендирующим агентом и одним или более консервантов. Типичные примеры пригодных диспергирующих или смачивающих агентов и суспендирующих агентов были уже указаны выше.
В определенных аспектах, концентрация OspA в таких композициях меняется в широких пределах, например, от менее чем примерно 0,5%, обычно около или по меньшей мере примерно 1%, и до 15 или 20% мас., и будет выбираться преимущественно на основании объемов жидкости, вязкостей и т.д., в соответствии с выбранным конкретным способом введения. Так, например, и без ограничений, типичная фармацевтическая композиция для парентеральной инъекции готовится таким образом, чтобы она содержала 1 мл стерильной забуференной воды и 50 мг фактора свертывания крови. Типичная композиция для внутривенной инфузии может быть приготовлена так, чтобы она содержала 250 мл стерильного раствора Рингера и 150 мг фактора свертывания крови. Фактические способы получения пригодных для парентерального введения композиций известны или понятны квалифицированным специалистам в данной области техники и описаны более подробно, например, в Remington’s Pharmaceutical Science, 15th ed., Mack Publishing Company, Easton, Pa. (1980). Эффективная доза обычно находится в интервале значений от 0,01 мг до 1000 мг на кг веса тела на введение.
В различных аспектах, фармацевтические композиции имеют вид стерильных водных, масляных суспензий для инъекции, дисперсий или стерильных порошков для приготовления предназначенных для немедленного применения стерильных растворов или дисперсий для инъекций. Суспензия, в некоторых аспектах, готовится в соответствии с известным уровнем техники с использованием пригодных диспергирующих или смачивающих агентов и суспендирующих агентов, указанных выше. Стерильный препарат для инъекций, в определенных аспектах, представляет собой стерильный раствор или суспензию для инъекций в нетоксичном приемлемом для парентерального введения разбавителе или растворителе, например, в виде раствора в 1,3-бутандиоле. В некоторых вариантах воплощения носитель представляет собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.), их пригодные смеси, растительные масла, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве растворителя или суспендирующей среды обычно используются стерильные нелетучие масла. С этой целью используют любое обычное нелетучее масло, в различных аспектах, включая синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, находят применение при приготовлении композиций для инъекций.
Во всех случаях, лекарственная форма должна быть стерильной и текучей до степени, позволяющей легко вводить ее шприцом. Надлежащая текучесть поддерживается, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии, и путем использования поверхностно-активных веществ. Она должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Предотвращение воздействия микроорганизмов обеспечивается различными антибактериальными и антигрибковыми агентами, например, парабенами, хлорбутанолом, фенолом, сорбиновой кислотой, тимеросалом и т.п. Во многих случаях будет желательным включение изотонических агентов, например, сахара или хлорида натрия. В определенных аспектах, пролонгирование всасывание композиций для инъекций обеспечивается путем использования в композициях агентов, замедляющих всасывание, например, моностеарата алюминия и желатина.
Композиции, пригодные для введения, в определенных аспектах, готовят с использованием средств, усиливающих всасывание или абсорбцию для увеличения их эффективности. Такие средства-энхансеры, включают, например, салицилат, гликохолат/линолеат, глихолат, апротинин, бацитрацин, ДСН, капрат и т.п. См., например, Fix (J. Pharm. Sci., 85:1282-1285, 1996) и Oliyai et al. (Ann. Rev. Pharmacol. Toxicol., 32:521-544, 1993).
Кроме того, свойства гидрофильности и гидрофобности композиций, используемых в составах и способах по изобретению, хорошо сбалансированы, что повышает их пригодность для in vitro и особенно in vivo применения, в то время как другие композиции, не имеющие такой сбалансированности, являются значительно менее пригодными. Конкретнее, композиции по изобретению имеют соответствующую степень растворимости в водных средах, что обеспечивает абсорбцию и биодоступность в организме, и в то же время имеют также определенную степень растворимости в липидах, что позволяет соединениям проходить через клеточные мембраны к предполагаемому месту действия.
В конкретных аспектах, из полипептидов OspA, описанных тут, готовят композиции вакцины, содержащие адъювант. Любой адъювант, известный специалистам, используется в различных аспектах композиции вакцины, включая масляные адъюванты, такие как полный адъювант Фрейнда и неполный адъювант Фрейнда, адъюванты на основе миколата (например, трегалозы димиколат), бактериальный липополисахарид (LPS), пептидогликаны (т.е., муреины, мукопептиды, или гликопротеины, такие как N-Opaca, мурамилдипептид [MDP], или аналоги MDP), протеогликаны (например, экстрагированные из Klebsiella pneumoniae), стрептококковые препараты (например, OK432), Biostim™ (например, 01K2), иммуностимулирующие комплексы «Iscoms» по EP 109942, EP 180564 и EP 231039, гидроксид алюминия, сапонин, DEAE-декстран, нейтральные масла (такие как миглиол), растительные масла (такие как арахисовое масло), липосомы, полиолы Pluronic®, система адъюванта Ribi (см., например, GB-A-2189141), или интерлейкины, особенно стимулирующие клеточно-медиируемый иммунитет. Альтернативный адъювант, состоящий из экстрактов Amycolata, рода бактерий порядка Actinomycetales, был описан в патенте США № 4877612. В дополнение к этому, имеются коммерчески доступные фирменные смеси адъювантов. Используемый адъювант зависит, частично, от субъекта-реципиента. Количество адъюванта для введения зависит от типа и размеров субъекта. Оптимальные дозировки легко определяются обычными способами.
Композиция вакцины необязательно включает совместимые с вакциной фармацевтически приемлемые (т.е. стерильные и нетоксичные) жидкие, полутвердые или твердые разбавители, которые служат фармацевтическими носителями, эксципиентами или средами. Используется любой разбавитель, известный специалистам. Типичные примеры разбавителей включают, без ограничений, полиоксиэтиленсорбитан монолаурат, стеарат магния, метил- и пропилгидроксибензоат, тальк, альгинаты, крахмалы, лактозу, сахарозу, декстрозу, сорбит, маннит, аравийскую камедь, фосфат кальция, минеральное масло, масло какао и масло купуасу.
Композиция вакцины упаковывается в формы, удобные для доставки. Композиции помещают в капсулу, таблетку в виде капсулы, саше, облатку, желатиновый, бумажный или другой контейнер. Такие формы для доставки являются предпочтительными, когда они совместимы с введением иммуногенной композиции в организм реципиента и, в частности, когда иммуногенная композиция доставляется в виде дозированной лекарственной формы. Дозированные формы упаковывают, например, в виде таблеток, капсул, суппозиториев, флаконов или облаток.
Изобретение включает способы индуцирования иммунологического ответа у субъекта, в том числе антител OspA у млекопитающего-хозяина, включающие введение эффективного количества композиций OspA, описанных тут. Аналогично, изобретение включает способы профилактики или лечения инфекции Borrelia или лаймской болезни у субъекта, включающие стадию введения субъекту эффективного количества композиций вакцины, описанных тут.
Композиция вакцины вводится субъекту, иммунизация которого проводится, любым обычным способом, как детально описано выше. В определенных аспектах, композицию вводят в виде разовой дозы или множества доз на протяжении периода времени (как описано более подробно ниже).
Дозировка композиций химерного OspA/Способы индуцирования иммунологического ответа
Пригодная для введения доза иммуногенной композиции или композиции вакцины будет меняться в зависимости от различных факторов, модифицирующих действие лекарственных средств, например, возраста, состояния, веса тела, пола и режима питания субъекта, тяжести любой инфекции, времени введения, способа введения и других клинических факторов.
В некоторых аспектах, составы или композиции по изобретению вводятся с помощью начальной ударной дозы с последующей доставкой бустер-доз по истечении периода времени. В определенных аспектах, композиции по изобретению вводятся с помощью начальной ударной дозы с последующей непрерывной инфузией для поддержания терапевтических уровней готовой лекарственной формы в циркуляции. В конкретных аспектах, иммуногенные композиции или композиции вакцины по изобретению вводятся по схеме вакцинации через различные периоды времени. В некоторых аспектах, вакцинация проводится по схеме быстрой иммунизации для людей, совершающих поездки в регионы, подверженные инфекции Borrelia. В качестве другого примера, состав или композицию по изобретению вводят в виде разовой дозы. Рядовые специалисты в данной области техники могут легко оптимизировать эффективные дозы и графики введения, определяемые добросовестной медицинской практикой и клиническим состоянием индивидуума. Частота доз зависит от фармакокинетических параметров агентов и пути введения.
Рецептура фармацевтической композиции определяется квалифицированным специалистом в данной области техники в зависимости от пути введения и желательной дозы. См., например, Remington’s Pharmaceutical Sciences, 18th Ed. (1990, Mack Publishing Co., Easton, PA 18042) pages 1435-1712, описание которого включено сюда в качестве ссылки. Такие композиции, в некоторых случаях, влияют на физическое состояние, стабильность, скорость высвобождения in vivo и скорость клиренса in vivo введенной композиции. В зависимости от пути введения, пригодную дозу рассчитывают, в конкретных аспектах, с учетом веса тела, площади поверхности тела или размера органа. В некоторых аспектах, пригодные дозы устанавливают с помощью общепризнанных анализов для определения уровней доз в крови в сочетании с соответствующими данными доза-реакция. В определенных аспектах, измеряют титр антитела у индивидуума для определения оптимальной дозы и схемы введения. Конечная схема введения дозы будет определяться лечащим врачом или терапевтом, с учетом различных факторов, модифицирующих действие фармацевтических композиций, например, специфической активности композиции, восприимчивости субъекта, возраста, состояния, веса тела, пола и режима питания субъекта, тяжести какой-либо инфекции или злокачественного состояния, времени введения и других клинических факторов. При проведении исследований будет появляться дополнительная информация, касающаяся пригодных уровней доз и продолжительности лечения для профилактики и/или лечения релевантных состояний.
В определенных аспектах, иммуногенная композиция или композиция вакцины OspA содержит любую дозу молекул нуклеиновой кислоты (кислот) или полипептида (полипептидов) OspA, достаточную для того, чтобы вызвать иммунный ответ у субъекта. Эффективное количество иммуногенной композиции или композиции вакцины OspA для терапевтического использования будет зависеть, например, от терапевтического контекста и целей. Квалифицированному специалисту в данной области техники будет понятно, что, таким образом, соответствующие уровни доз для вакцинации или лечения будут различаться, в зависимости, частично, от доставляемой молекулы, показания для использования молекулы (молекул) OspA, пути введения и размеров (веса тела, поверхности тела или размера органа) и состояния (возраста и общего состояния здоровья) пациента. Соответственно, клинический врач, в некоторых случаях, титрует дозу и модифицирует путь введения для достижения оптимального терапевтического эффекта.
Типичная доза, в различных аспектах, находится в интервале значений от примерно 0,1 мкг/кг до примерно 100 мг/кг или более, в зависимости от вышеуказанных факторов. В других вариантах воплощения доза может составлять от 0,1 мкг/кг до примерно 100 мг/кг; или от 1 мкг/кг до примерно 100 мг/кг; или от 5 мкг/кг до примерно 100 мг/кг. В качестве примера, доза полипептида OspA, пригодная для использования по настоящему изобретению, составляет приблизительно 10 мкг/мл, 20 мкг/мл, 30 мкг/мл, 40 мкг/мл, 50 мкг/мл, 60 мкг/мл, 70 мкг/мл, 80 мкг/мл, 90 мкг/мл, 100 мкг/мл, 110 мкг/мл, 120 мкг/мл, 130 мкг/мл, 140 мкг/мл, 150 мкг/мл, 160 мкг/мл, 170 мкг/мл, 180 мкг/мл, 190 мкг/мл, 200 мкг/мл, 210 мкг/мл, 220 мкг/мл, 230 мкг/мл, 240 мкг/мл, 250 мкг/мл, 260 мкг/мл, 270 мкг/мл, 280 мкг/мл, 290 мкг/мл, 300 мкг/мл, 320 мкг/мл, 340 мкг/мл, 360 мкг/мл, 380 мкг/мл, 400 мкг/мл, 420 мкг/мл, 440 мкг/мл, 460 мкг/мл, 480 мкг/мл, 500 мкг/мл, 520 мкг/мл, 540 мкг/мл, 560 мкг/мл, 580 мкг/мл, 600 мкг/мл, 620 мкг/мл, 640 мкг/мл. В конкретных аспектах, типичная доза содержит от 0,1 до 5,0 мл на субъекта. В более конкретных аспектах, типичная доза содержит от 0,2 до 2,0 мл на субъекта. В определенных аспектах, доза содержит от 0,5 до 1,0 мл на субъекта.
Частота введения доз будет зависеть от фармакокинетических параметров молекулы OspA в используемой композиции. Типично, клинический врач будет вводить композицию до достижения дозы, обеспечивающей желательный эффект. Композиция, в различных аспектах, вводится поэтому в виде разовой дозы, или в виде двух или более доз (которые могут содержать одинаковое или разное количество желательных молекул) со временем, или в виде непрерывной инфузии с помощью имплантированного приспособления или катетера. Дополнительное уточнение соответствующей дозы выполняется в повседневной практике рядовыми специалистами в данной области техники и не выходит за пределы выполняемых ими повседневных работ. Требуемые дозы часто уточняют путем использования соответствующих данных доза-реакция, которые получают в обычном порядке.
Наборы
Как дополнительный аспект, изобретение включает наборы, содержащие одну или более фармацевтических композиций для введения полипептида (полипептидов) OspA субъекту, упакованные так, чтобы облегчить их использование для введения субъектам.
В конкретном варианте воплощения изобретение включает наборы для приготовления устройства для введения разовой дозы. Каждый набор, в различных аспектах, содержит первый контейнер с сухим белком и второй контейнер с водной композицией. В объем настоящего изобретения также входят наборы, содержащие одно- и многокамерные предварительно наполненные шприцы (например, шприцы для жидкостей и лиошприцы).
В другом варианте воплощения такой набор включает фармацевтическую композицию, описанную тут (например, композицию, содержащую терапевтический белок или пептид), упакованную в контейнер, такой как герметически закрытая бутылка или сосуд, с прикрепленным к контейнеру или входящим в состав упаковки ярлыком, описывающим использование соединения или композиции для практической реализации способа. В одном варианте воплощения фармацевтическая композиция упакована в контейнер так, чтобы объем свободного пространства в контейнере (например, количество воздуха между жидкой композицией и верхушкой контейнера) был очень маленьким. Предпочтительно, объем свободного пространства является пренебрежимо малым (т.е. почти отсутствует).
В одном аспекте, набор содержит первый контейнер, содержащий терапевтическую композицию белка или пептида, и второй контейнер, содержащий физиологически приемлемый раствор для восстановления композиции. В одном аспекте, фармацевтическая композиция упакована в виде дозированной лекарственной формы. Набор необязательно дополнительно включает приспособление, пригодное для введения фармацевтической композиции определенным путем введения. В некоторых аспектах, набор содержит ярлык, описывающий использование фармацевтических композиций.
Каждая публикация, патентная заявка, патент и другие первоисточники, упоминаемые в данном описании, целиком включены сюда в качестве ссылок в степени, не противоречащей целям данного описания.
Следует понимать, что примеры и варианты воплощения, описанные тут, приведены только с иллюстративными целями, и что квалифицированные специалисты в данной области техники смогут предложить на их основе различные модификации или изменения, которые должны считаться входящими в сущность и границы данной заявки и в объем приложенной формулы изобретения. Все публикации, патенты и патентные заявки, упоминаемые тут, настоящим по существу целиком включены сюда в качестве ссылок.
ПРИМЕРЫ
Дополнительные аспекты и детали изобретения будут понятны из следующих примеров, которые должны рассматриваться как иллюстративные, а не ограничительные.
Пример 1
Анализ последовательности OspA европейских штаммов Borrelia burgdorferi sensu lato (молекулярная эпидемиология) для определения рецептуры композиции вакцины OspA
Целью данных исследований было определение пригодной рецептуры композиции вакцины OspA лаймской болезни для Европы. Исследования были основаны на анализе последовательности гена OspA (молекулярная эпидемиология) из большой и разнообразной коллекции штаммов B. burgdorferi sensu lato, которая адекватно представляет обширный географический регион Европы, различных клинических синдромов, ассоциированных с болезнью, и каждого из трех патогенных геновидов (B. afzelii, B. garinii и B. burgdorferi ss), ассоциированных с лаймской болезнью. Лаймская болезнь вызывается видом Borrelia burgdorferi sensu lato, который включает в общей сложности 13 геновидов, три из которых (B. afzelii, B. garinii и B. burgdorferi ss) признаны патогенными для человека.
Вначале были проведены крупномасштабные эпидемиологические исследования (см. Таблицу 3 ниже), в которых были проанализированы штаммы Borrelia burgdoferi sensu lato от пациентов с лаймской болезнью (и от клещей) из 21 европейской страны. Было изучено в общей сложности 553 изолята европейских Borrelia, собранных в 16 европейских странах. Каждый вид определяли способом ПЦР с использованием наборов праймеров, специфических по отношению к генам 16s rРНК каждого вида.
Изоляты каждого из трех видов Borrelia, которые, как известно, вызывают у человека лаймскую болезнь, в Европе были хорошо представлены: B. afzelii (n=309, 55,9%), B. burgdorferi sensu stricto (n=67, 12,1%) и B. garinii (n=173, 31,3%). Из 359 человеческих изолятов, 56,8% принадлежали к B. аfzelii, и B. afzelii был преобладающим видом, определенным в человеческих изолятах в большинстве регионов. Аналогично, B. afzelii был выделен из 54,1% изолятов клещей. B. burgdorferi s.s. был выделен из 11,7% человеческих штаммов и 12,9% клещевых изолятов. B. burgdorferi s.s. был выделен из человеческих изолятов из юго-восточной Европы, а именно, Италии, Венгрии, Словении и Австрии. Штаммы B. garinii были выделены из 30,4% человеческих изолятов и 33% изолятов клещей. Штаммы B. garinii, выделенные у людей и клещей, были получены из большей части географических регионов по всей Европе. Данные этих исследований хорошо коррелируют с данными, содержащимися в других европейских исследованиях, и позволяют предположить, что изученная коллекция изолятов представляет точную картину лаймской болезни в Европе.
Секвенирование OspA проводилось для определения оптимальной рецептуры композиции вакцины для Европы. На основании этих данных, вакцина, включающая типы OspA 1-6, будет охватывать 98,1% штаммов и 96,7% случаев инвазивного заболевания. Результаты эпидемиологических исследований европейских изолятов Borrelia показывают, что вакцина на основе типов OspA 1, 2, 3, 4, 5 и 6 будет обеспечивать в Европе теоретический охват 98% случаев лаймской болезни и 96,7% инвазивных изолятов нейроборрелиоза.
Результаты эпидемиологических исследований
2 Прогнозируемый уровень охвата вакцины изолятов случаев нейроборрелиоза; итоговые значения являются кумулятивными.
Таким образом, вакцина, содержащая три новых рекомбинантных OspA (1/2, 6/4 и 5/3), каждый из которых представляет 2 серотипа OspA, сохраняет ключевые структурные элементы, необходимые для защиты против всех 6 распространенных серотипов OspA (1-6), ассоциированных с лайм-боррелиозом в Европе, и против единственного серотипа OspA, ассоциированного с лайм-боррелиозом в США.
Включение конструкта OspA 5/3, представляющего серотипы OspA 5 и 3 B. garinii (вместе с серотипами OspA 1/2 и 6/4), должно обеспечивать защиту против 98,1% случаев болезни и 96,7% инвазивных изолятов. Можно ожидать, что вакцина без OspA 5/3 будет защищать только от приблизительно 88,9% случаев болезни и только от приблизительно 73,4% инвазивных случаев болезни. Таким образом, вакцина, содержащая все шесть серотипов, является более эффективной для профилактики лаймской болезни, чем вакцина с лишь четырьмя серотипами.
Пример 2
Стратегия конструирования синтетических генов OspA, кодирующих липидированный OspA
Целью исследований было получение липидированных химерных конструктов OspA из нескольких штаммов Borrelia для приготовления вакцины, которая защищала бы реципиента от лаймской болезни, вызываемой любыми из таких нескольких штаммов Borrelia. Общая стратегия представлена на Фиг. 1 и описана ниже.
Для каждого нового гена OspA было синтезировано четыре набора олигонуклеотидов размером 30-60 оснований. Каждый набор олигонуклеотидов состоял из 8-12 комплементарных перекрывающихся олигонуклеотидов. Олигонуклеотиды из каждого набора отжигают вместе, в отдельных экспериментах, для получения двухцепочечных фрагментов ДНК со специфическими сайтами распознавания рестрикционных ферментов на обоих концах, т.е. фрагментов N-H (Nde I - Hind III), H-K (Hind III - Kpn I), K-E (Kpn I - EcoR I) и E-B (EcoR I - BamH I). Каждый из четырех фрагментов независимо клонируют в pUC18, разрезают соответствующими рестрикционными ферментами и трансформируют в хозяина E. coli DH5α, после чего проверяют последовательность клонированного фрагмента.
Штамм E. coli DH5α [генотип: end A1 hsdR17 (rk-mk+) supE44 thi-1 recA1 gyrA (Nalr) relA1 Δ(lacZYA-argF)U169 deoR (Φ80dlacΔ(lacZ)M15] (Gibco BRL) используют для всех промежуточных стадий клонирования. Этот штамм получен из штамма E. coli K12, одного из наиболее широко используемых хозяев в генной инженерии. Штамм является amp- для обеспечения возможности селекции трансформантов с помощью векторов, содержащих ген резистентности к ампициллину (amp). E. coli HMS174(DE3) была выбрана в качестве хозяина для экспрессии. Клетки-хозяева E. coli HMS174(DE3) [генотип : F- recA1 hsdR (rk12- mk12+) RifR (DE3)] (Novagen) использовались для конечных стадий клонирования. Штамм является kan- для обеспечения возможности селекции трансформантов с помощью векторов, содержащих ген резистентности к канамицину (kan).
pUC18 (Gibco BRL, Basel, Switzerland) используют в качестве клонирующего вектора для всех промежуточных стадий, потому что генетические манипуляции и секвенирование с этой плазмидой проще выполнять, чем с pET30a. Основными признаками являются, в частности, генный фрагмент lacZ, кодирующий альфа-пептид LacZ пары оснований от 149 до 469 (lac-промотор от пары оснований 507), ген bla, кодирующий детерминанту резистентности к ампициллину пары оснований от 1629 до 2486 (промотор bla от пары оснований 2521), точка начала репликации от пары оснований 867 и сайты множественного клонирования пары оснований от 185 до 451 (Фиг. 12).
pET30a (Novagen) используют в качестве экспрессионного вектора для окончательной вставки полного гена OspA. В векторах pET гены клонируются под контролем промотора T7, и экспрессия индуцируется путем обеспечения источника РНК-полимеразы T7 в клетке-хозяине (экспрессия не происходит до тех пор, пока не обеспечен источник РНК-полимеразы T7). Основными признаками являются ген, кодирующий резистентность к канамицину (kan) от пары оснований 4048 до 4860, ген lacI пары оснований 826-1905, точка начала репликации F1 пары оснований 4956-5411, и сайты множественного клонирования пары оснований от 158 до 346 (Фиг. 13).
Четыре фрагмента, необходимые для приготовления полноразмерного гена OspA, были выделены с помощью набора DNA miniprep. ДНК выделяют из каждого из четырех клонов с использованием тех же рестрикционных ферментов, что и используемые для начальной стадии клонирования. Фрагменты ДНК очищают и лигируют вместе с ДНК pUC18, разрезанной Nde I и BamH I, и трансформируют в DH5α-компетентные клетки E. coli. Полноразмерный ген OspA, клонированный в pUC18, секвенируют для подтверждения того, что на этой стадии не было внесено ошибок.
Ген OspA затем субклонируют в экспрессионный вектор pET-30a с помощью рестрикционных ферментов Nde I и BamH I и трансформируют в хозяина E. coli HMS 174(DE3). В векторе pET30a, ген OspA контролируется промотором T7 бактериофага.
Три синтетических гена OspA были сконструированы так, чтобы они кодировали молекулы OspA с защитными эпитопами из OspA серотипов 1 и 2 (lipB sOspA 1/2251), OspA серотипов 6 и 4 (lipB sOspA 6/4) и OspA серотипов 5 и 3 (lipB sOspA 5/3) Borrelia. Первичные аминокислотные последовательности этих молекул (SEQ ID NO: 2, 4 и 6, соответственно) приведены на Фигурах 2-8 и описаны тут, причем в их конструкцию включено полное описание основных отличительных признаков.
Олигонуклеотиды для конструкта lipB sOspA 1/2 были синтезированы собственными силами на синтезаторе ABI 394 ДНК/РНК. Олигонуклеотиды для конструктов lipB sOspA 5/3 и lipB sOspA 6/4 были приобретены у GenXpress (Wiener Neudorf, Austria) и очищены способом ВЭЖХ.
Олигонуклеотиды для генных фрагментов lipB sOspA 1/2*
L - Длина олигонуклеотида в основаниях
S - Нить: C - кодирующая, или C’ – комплементарная
Олигонуклеотиды для генных фрагментов lipB sOspA 5/3
S - Нить: C - кодирующая, или C’ - комплементарная
Олигонуклеотиды для генных фрагментов lipB sOspA 6/4
CGACTGCGTTGCCTTCGA
S - Нить: C - кодирующая, или C’ - комплементарная
Приготовление компетентных клеток E. coli. Используют одну колонию для инокуляции 5 мл модифицированного бульона LB (5,5 г NaCl, 5 г экстракта дрожжей, 10 г соевого пептона, который не был получен от животного или генетически модифицированного растительного источника - на литр воды). Культуру инкубируют, пока она не помутнеет, после чего культуру разводят до объема 25 мл предварительно подогретым модифицированным бульоном LB. Культуру инкубируют далее до достижения значения OD600нм в интервале от 0,2 до 0,6 (40-60 мин.) и разводят до объема 125 мл, переносят в колбу на 500 мл и инкубируют до достижения OD600нм, равного 0,6. Культуру быстро охлаждают осторожным встряхиванием в течение 5 мин. на ледяной бане и клетки осаждают центрифугированием (центрифуга Beckman, 4000 об/мин в течение 10 мин.), осторожно промывают буфером TfBI (Teknova Hollister, CA) (30 мМ K-ацетата, 50 мМ MnCl2, 100 мМ KCl, 10 мМ CaCl2 15% глицерина), ресуспендируют в 5 мл TfBII (10 мМ Na-MOPS, 75 мМ CaCl2, 10 мМ KCL, 15% глицерина) и выдерживают на льду в течение 15 мин. Затем пипеткой берут аликвоты клеток 100 мкл и подвергают быстрому замораживанию непосредственно в сухом льду.
Отжиг олигонуклеотидных смесей для получения фрагментов гена OspA (синтез de novo). Были сконструированы три синтетических гена OspA, кодирующие молекулы OspA с защитными эпитопами из OspA серотипов 1 и 2 (lipB sOspA 1/2), OspA серотипов 6 и 4 (lipB sOspA 6/4) и OspA серотипов 5 и 3 (lipB sOspA 5/3). Для каждого нового гена OspA (липидированного) были синтезированы четыре набора олигонуклеотидов размером 30-60 пар оснований (см. Таблицы 4-6). Фигуры 16-18 изображают оптимизированные по кодонам последовательности каждого из конструктов, выровненные с нуклеотидными последовательностями, расшифрованными из опубликованных последовательностей). Каждый набор олигонуклеотидов состоит из 8-12 комплементарных перекрывающихся олигонуклеотидов. Олигонуклеотиды из каждого набора отжигают вместе, в отдельных экспериментах, для получения двухцепочечных фрагментов ДНК со специфическими сайтами распознавания рестрикционных ферментов на обоих концах, т.е. фрагменты N-H (Nde I - Hind III), H-K (Hind III - Kpn I), K-E (Kpn I - EcoR I) и E-B (EcoR I - BamH I).
Лиофилизированные олигонуклеотиды восстанавливают дистиллированной водой, измеряют OD260 нм и концентрацию доводят до 10 мкМ. Для каждого фрагмента OspA, 2 мкл каждого из олигонуклеотидов смешивают с 1 мкл T4 полинуклеотидкиназы и буфером T4 ДНК-лигазы (10×) и смесь инкубируют при комнатной температуре в течение 30 минут для обеспечения возможности фосфорилирования олигосоединений (для конструкта lipB sOspA 6/4 эту стадию опускают, поскольку олигосоединения уже фосфорилированы). Смесь нагревают до 95°C в течение 1 минуты (стадия денатурирования) и затем олигосоединения подвергают отжигу, оставляя смесь медленно охлаждаться до комнатной температуры. Подвергнутую отжигу смесь непосредственно используют для лигирования, или хранят при -20°C до дальнейшей необходимости.
Клонирование фрагментов гена OspA. Каждый из четырех фрагментов, требуемых для конструирования индивидуального синтетического гена OspA, независимо клонируют в pUC18 и трансформируют в хозяина E. coli DH5α (см. Фигуру 1).
Для каждого нового гена OspA были синтезированы четыре набора олигонуклеотидов размером 30-60 оснований. Каждый набор олигонуклеотидов состоит из 8-12 комплементарных перекрывающихся олигонуклеотидов. Олигонуклеотиды из каждого набора отжигают вместе, в отдельных экспериментах, для получения двухцепочечных фрагментов ДНК со специфическими сайтами распознавания рестрикционных ферментов на обоих концах, т.е. фрагменты N-H (Nde I - Hind III), H-K (Hind III - Kpn I), K-E (Kpn I - EcoR I) и E-B (EcoR I - BamH I). Каждый из четырех (4) фрагментов независимо клонируют в pUC18, разрезают соответствующими рестрикционными ферментами и трансформируют в хозяина E. coli DH5α, после чего проверяют последовательность клонированного фрагмента.
Плазмиду ДНК (pUC18) очищают из ночной культуры E. coli (бульон LB) с помощью системы очистки плазмид QIAGEN в соответствии с протоколом производителя. Вектор ДНК затем гидролизуют парами рестрикционных ферментов; Nde I и Hind III, Hind III и Kpn I, Kpn I и EcoR I, EcoR I и BamH I, в соответствии с протоколами производителя. Гидролизованные образцы наносят на 0,8% агарозный гель и электрофоретически разделяют. Линеаризованный вектор ДНК вырезают и элюируют с помощью коммерческого набора для элюирования геля (QIAquick Gel Extraction Kit, Qiagen) в соответствии с протоколом производителя, и лигируют, используя T4 ДНК-лигазу, с отожженной смесью олигонуклеотидов. Продукты лигирования трансформируют в конкурентные клетки E. coli DH5α и трансформанты, содержащие плазмиду, селектируют на агаре LB, содержащем ампициллин (100 мкг/мл).
Присутствие вставки ожидаемого размера в клонирующем векторе, pUC18, подтверждают путем очистки плазмидной ДНК, гидролиза ДНК ферментами, используемыми для клонирования, и анализа фрагментов ДНК электрофорезом на агарозном геле с использованием ранее описанных процедур. Клонированный фрагмент ДНК секвенируют, используя очищенную плазмидную ДНК в качестве ДНК-матрицы и праймеры секвенирования 5’-TCGGGGCTGGCTTAACTATG-3 (SEQ ID NO: 14) и 5’-GCTTCCGGCTCGTAT (SEQ ID NO:15) (которые находятся в векторе pUC18 за пределами сайтов множественного клонирования, п.о. 130-150 и п.о. 530-515, соответственно). Последовательность реакций выполняется на автоматическом секвенаторе (ABI 310). Последовательности редактируют с помощью SequenceEditor и последовательности импортируют в VectorNTI для анализа. Только клоны с правильными последовательностями используют в качестве строительных блоков для конструирования полноразмерных генов OspA.
Для гена lipB sOspA 5/3 используют другую стратегию, поскольку во фрагменте Kpn I - BamH I не удалось найти пригодного уникального внутреннего сайта, и аминокислотная последовательность не позволяла использовать внутренний сайт EcoR I (см. Фигуру 14). Сайт Pvu II имеется во фрагменте Kpn I - BamH I, однако в векторе pUC18 присутствует два сайта Pvu II, что означает невозможность прямого клонирования фрагментов в pUC18. Поэтому, олигосоединения для конструктов были сконструированы так, чтобы сайт EcoR I был вставлен вовне и рядом с сайтом Pvu II, позволяя клонировать фрагменты Kpn I - EcoR I и EcoR I - BamH I в pUC18. Последующий гидролиз вставленных фрагментов с помощью Kpn I, EcoR I и BamH I давал фрагменты, которые затем гидролизуют Pvu II. Гидролизованные Pvu II фрагменты (Kpn I - Pvu II и Pvu II - BamH I) затем используют для тройного лигирования с ДНК вектора pUC18, разрезанной Kpn I и BamH I для получения фрагмента Kpn I - BamH I.
Конструирование полноразмерных генов OspA. На следующей стадии, каждый из четырех фрагментов, необходимых для конструирования индивидуального синтетического гена OspA, вырезают из вектора pUC18 и повторно клонируют, за одну стадию, в вектор pUC18 для получения полноразмерного гена OspA (см. Фигуру 1).
Четыре фрагмента, необходимые для приготовления полноразмерных генов, вырезают с помощью набора miniprep ДНК, выделенную с использованием одних и тех же рестрикционных ферментов, используют для начальной стадии клонирования. Гидролизованные образцы наносят на агарозный гель и электрофоретически разделяют. ДНК каждого из соответствующих 4 фрагментов вставок вырезают и элюируют с помощью коммерческого набора для элюирования геля (QiaQuick Gel Extraction Kit) в соответствии с протоколом производителя и лигируют, используя T4 ДНК-лигазу, с линеаризованной векторной ДНК, гидролизованной Nde I и BamH I и очищенной с помощью QIAquick Gel Extraction Kit. Лигированную ДНК трансформируют в конкурентные клетки E. coli DH5α и клоны, содержащие плазмиду, селектируют на LB-агаре, содержащем ампициллин (100 мкг/мл). Колонии подвергают скринингу способом ПЦР на присутствие вставок ожидаемого размера (ок. 830 п.о.).
Единичные колонии используют в качестве матричной ДНК в ПЦР-реакциях с использованием 10Х буфера (15 мМ Tris-HCl (pH 8,0), 50 мМ KCl, 1,5 мМ MgCl2), 200 мкМ dNTPs, 1,25 ед. Amplitaq ДНК-полимеразы, 400 нМ прямого праймера 5’-TCGGGGCTGGCTTAACTATG-3 (SEQ ID NO: 14) и 400 нМ обратного праймера 5’-GCTTCCGGCTCGTAT (SEQ ID NO: 15). Используют следующие условия ПЦР-реакции: 94°C в течение 5 мин., 35×(94°C в течение 30 с, 48°C в течение 30 с, 72°C в течение 1 мин 30 с) с последующим вымачиванием при 72°C в течение 5 минут и выдерживанием при 4°C. Продукты ПЦР используют непосредственно или хранят при ≤15°C до дальнейшего использования. Продукты ПЦР анализируют электрофорезом на агарозном геле на присутствие вставок правильного размера (ок. 980 п.о.). Вставки правильного размера секвенируют для подтверждения того, что не было внесено ошибок, т.е. последовательность реакций была проведена с использованием плазмидной ДНК, выделенной (QIAGEN Plasmid Purification kit) из ночных культур (бульон LB для амплификации) и с использованием праймеров секвенирования, фланкирующих сайты клонирования (5’-TCGGGGCTGGCTTAACTATG-3’(SEQ ID NO: 14) и 5’-GCTTCCGGCTCGTATGTTGT-3’ (SEQ ID NO: 16), п.о. 130-150 и 530-510, соответственно). Последовательность реакций выполняется с помощью автоматического секвенатора (ABI 310). Последовательности редактируют с помощью SequenceEditor и последовательности импортируют в VectorNTI для анализа.
Субклонирование новых генов OspA в экспрессионный вектор pET30a. После проверки полноразмерного гена OspA в pUC18, гены OspA субклонируют в экспрессионный вектор pET-30a с помощью рестрикционных ферментов NdeI и BamH I и трансформируют в хозяина E. coli HMS 174(DE3).
Miniprep-ДНК из клонов pUC18 с правильными последовательностями гидролизуют с помощью Nde I и BamH I. Аналогично, ДНК вектора pET30a гидролизуют с помощью Nde I и BamH I. Гидролизованные ДНК наносят на агарозный гель и электрофоретически разделяют. Вырезают фрагмент вставки размером приблизительно 830 п.о. и линеаризованный вектор ДНК и очищают, как описано ранее. Вектор и ДНК вставки лигируют, используя T4 ДНК-лигазу, и продукты лигирования трансформируют в конкурентные клетки E. coli HMS174(DE3) (Novagen). Трансформанты высеивают на чашки со средой LB, содержащей канамицин (30 мкг/мл). Единичные колонии подвергают скринингу способом ПЦР с использованием праймеров 5’-TTATGCTAGTTATTGCTCAGCG-3’ (SEQ ID NO:17) и 5’-TTCCCCTCTAGAAATAATTTTGT-3’ (SEQ ID NO: 18). Продукты ПЦР наносят на агарозный гель и электрофоретически разделяют. Колонии, вырабатывающие продукт правильного размера (ок. 1 т.п.н.) впоследствии используют для получения ночных культур, из которых выделяют miniprep-ДНК с помощью набора для очистки плазмид QIAGEN (QIAGEN Plasmid Purification kit) в соответствии с протоколом производителя. Последовательность снова проверяют (с использованием праймеров 5’-TTATGCTAGTTATTGCTCAGCG-3’ (SEQ ID NO: 17) и 5’-TTCCCCTCTAGAAATAATTTTGT-3’ (SEQ ID NO:18), п.о. 65-86 и 395-373, соответственно) и селектируют колонии для испытаний экспрессии.
Получение lipB sOspA 1/2251 из lipB sOspA 1/2. Изменяют единичную аминокислоту в конструкте lipB sOspA 1/2, а именно аминокислоту аланин в положении 251 заменяют на остаток аспарагина для усиления иммуногенности. Замену аминокислоты проводят способом ПЦР. Во-первых, проводят ПЦР с внешним прямым праймером и внутренним обратным праймером, получая продукт размером примерно 730 п.о. с внесенной заменой аминокислоты (см. Фигуру 15). Во-вторых, проводят ПЦР с внутренним прямым праймером и внешним обратным праймером, получая продукт размером 100 п.о., содержащий внесенную замену аминокислоты. Два продукта ПЦР, последовательности которых перекрываются, затем используют в качестве матричной ДНК для конечной ПЦР-реакции с внешним прямым и внешним обратным праймерами для получения конечного полноразмерного продукта OspA, содержащего внесенную замену аминокислоты.
Конструкт pET30a используют в качестве источника матричной ДНК. Проводят ПЦР-реакции с 10Х буфером [15 мМ Tris-HCl (pH 8,0), 50 мМ KCl, 1,5 мМ MgCl2], 200 мкМ dNTPs, 1,25 ед. Amplitaq ДНК-полимеразы и 400 нМ каждой пары праймеров (пара праймеров 5’-GGA ATT CCA TAT GCG TCT GTT GAT CGG CT (SEQ ID NO: 19) и 5’-TTG GTG CCT GCG GAG TCG (SEQ ID NO:20) и пара праймеров 5’-AAT ACG ACT CCG CAG GCA CC (SEQ ID NO: 21) и 5’-CTG-GGA TCC GCT CAG CTT ATT TCA (SEQ ID NO: 22)). ПЦР-реакции проводят в таких условиях: 94°C в течение 5 мин., 35×(94°C в течение 30 с, 48°C в течение 30 с, 72°C в течение 1 мин 30 с) с последующим вымачиванием при 72°C в течение 5 минут и выдерживанием при 4°C. Реакции дают 2 разных перекрывающихся продукта, и 2 продукта используют в качестве матричной ДНК для третьей ПЦР-реакции с использованием внешних праймеров 5’-GGA ATT CCA TAT GCG TCT GTT GAT CGG CT (SEQ ID NO:19) и 5’-CTG-GGA TCC GCT CAG CTT ATT TCA (SEQ ID NO: 22), включающих сайты рестрикции Nde I и BamH I. Используют условия реакции 94°C в течение 60 с с последующими 35 циклами (30 с при 94°C, 60 с при 49°C, 90 с при 72°C), а затем 72°C в течение 5 мин. Амплифицированный продукт очищают с помощью набора для очистки QiaQuick (Qiagen) в соответствии со спецификацией производителя и продукт гидролизуют с помощью Nde I и BamH I и лигируют в векторную ДНК pET30a, разрезанную Nde I и BamH I. Продукты лигирования трансформируют в конкурентные клетки E. coli DH5α. Трансформанты высеивают на чашки со средой LB, содержащей канамицин (30 мкг/мл). Единичные колонии подвергают скринингу способом ПЦР с использованием праймеров 5’-TTATGCTAGTTATTGCTCAGCG-3’ (SEQ ID NO:17) и 5’-TTCCCCTCTAGAAATAATTTTGT-3’ (SEQ ID NO: 18). Продукты ПЦР наносят на агарозный гель и электрофоретически разделяют. Колонии, дающие продукт правильного размера (ок. 1 т.п.н.) используют впоследствии для получения ночных культур, из которых выделяют miniprep-ДНК с помощью системы для очистки плазмид QIAGEN в соответствии с протоколом производителя. Последовательность подтверждают (с использованием праймеров 5’-TTATGCTAGTTATTGCTCAGCG-3’ (SEQ ID NO: 17) и 5-TTCCCCTCTAGAAATAATTTTGT-3’ (SEQ ID NO: 18)) и полученный конструкт трансформируют в компетентные клетки E. coli HMS174(DE3), и полученным позитивным трансформантам было присвоено наименование lipB sOspA 1/2251.
Получение конструктов без лидерной последовательности. Были получены конструкты с лидерной последовательность lipB, в которых липидный фрагмент типично присоединен к амино-концевому цистеиновому остатку. Экспериментальные исследования рекомбинантных липидированных OspA подтвердили присутствие липидного фрагмента. Однако, были также получены конструкты, не содержащие лидерной последовательности lipB. Конструкты, не содержащие лидерную последовательность lipB, были приготовлены способом ПЦР амплификации каждого из трех lipB конструктов (в pET30a) с использованием праймеров, выбранных для получения конечного продукта размером 769-771 п.о. без последовательности нуклеиновой кислоты, кодирующей лидерную последовательность и с заменой кодона цистеинового остатка на кодон остатка метионина.
ПЦР-реакции используют 10Х буфер [15 мМ Tris-HCl (pH 8,0), 50 мМ KCl, 1,5 мМ MgCl2], 200 мкМ dNTPs, 1,25 ед. Amplitaq ДНК-полимеразы, 400 нМ прямого праймера 5’-CGTGCGTACCATATGGCACAGAAAGGTGCTGAGTCT-3’ (SEQ ID NO: 23) и 400 нМ обратного праймера 5’-CTGGGATCCGCTCAGCTTATTTCA-3’ (SEQ ID NO: 22) и матричной ДНК. Используют условия ПЦР: 94°C в течение 5 мин., 35×(94°C в течение 30 с, 48°C в течение 30 с, 72°C в течение 1 мин 30 с) с последующим вымачиванием при 72°C в течение 5 мин. и выдерживанием при 4°C. ПЦР-реакции используют непосредственно или хранят при ≤15°C до дальнейшего использования.
Продукты ПЦР очищают с помощью набора для очистки ПЦР QiaQuick (Qiagen), гидролизуют с помощью Nde I и BamH I и лигируют с векторной ДНК pET30a, гидролизованной с помощью Nde I и BamH I. Смеси лигирования используют для трансформации E. coli HMS174(DE3) и колонии, содержащие рекомбинантные плазмиды, селектируют по их резистентности к канамицину и проверяют последовательность продуктов ПЦР.
Оценка экспрессии в E. coli HMS 174(DE3). Выбранные колонии испытывают на их способность к экспрессии соответствующего нового белка OspA. В каждом случае, используют единичные колонии для инокуляции бульона LB, содержащего канамицин (30 мкг/мл) и инкубируют при 37°C на протяжении 1-5 часов до достижения значения OD (600 нм) более 0,6 и менее 1. В этот момент времени, берут образец культуры (представляющий собой неиндуцированный образец) и остальную культуру индуцируют прибавлением IPTG до конечной концентрации 1 мМ. Неиндуцированный образец (1 мл) центрифугируют и осадок собирают и хранят при -20°C. Индуцированной культуре позволяют расти еще три часа, после чего берут образец 1 мл, измеряют OD (600 нм), образец центрифугируют и осадок собирают и хранят при -20°C.
Приготовление первичных клеток. Первичные клетки готовят для каждого из трех липидированных конструктов и для каждого из трех нелипидированных конструктов. Первичные клетки представляют собой клетки E. coli (HMS174(DE3)), несущие плазмиду pET30a, экспрессирующую соответствующий OspA. Для получения первичных клеток, берут единичную колонию из соответствующего экземпляра культуры с чашки, содержащей канамицин (30 мкг/мл) и рифампицин (200 мкг/мл) и используют для инокуляции 500 мкл среды SMK (SOP 8114) и инкубируют в течение ночи. Сто микролитров этой культуры затем используют для инокуляции 100 мл среды SMK (в двух экземплярах) и культуру инкубируют в течение 17-20 часов при 37°C со встряхиванием. Затем прибавляют к культуре стерильный глицерин в конечной концентрации 15% и материал отбирают пипеткой аликвотами по 500 мкл в 60 ампул, получая таким образом 60 ампул с первичными клетками, которые немедленно направляют на хранение при -80°C.
Три синтетических гена OspA были сконструированы так, чтобы они кодировали молекулы OspA с защитными эпитопами OspA серотипов 1 и 2 (lipB sOspA 1/2251), OspA серотипов 6 и 4 (lipB sOspA 6/4) и OspA серотипов 5 и 3 (lipB sOspA 5/3). Первичные аминокислотные последовательности этих молекул и описание основных отличительных признаков, включенных в их конструкцию, приведены в следующих Примерах.
Пример 3
Описание липидированного 1/2251 OspA (lipB sOspA1/2251)
Целью исследований было конструирование нового антигена OspA, липидированного 1/2251 OspA (lipB sOspA 1/2251), содержащего серотипы 1 и 2. LipB sOspA 1/2251 содержит проксимальный участок последовательности OspA серотипа 1 (штамм B31, номер доступа GenBank X14407), слитый с дистальным участком последовательности серотипа 2 (штамм Pko, номер доступа GenBank S48322). Началом последовательности, уникальным для серотипа типа 2, является лизиновый (K) остаток в положении 216. Конструкт сначала был сконструирован так, чтобы он кодировал аминокислоту аланин (A) в положении 251. Однако конструкт был впоследствии изменен способом ПЦР для кодирования аспарагинового (N) остатка (фактический остаток в опубликованной последовательности Pko) для усиления иммуногенности, вследствие чего он получил наименование lipB sOspA 1/2251.
Вторичные признаки lipB sOspA 1/2251 изображены на аннотированной аминокислотной последовательности lipB sOspA 1/2251 на Фигуре 2 и включают:
- замещение предположительного артритогенного эпитопа (Gross et al., 1998), hLFA-1 (YVLEGTLTA) (SEQ ID NO:24), в проксимальном участке молекулы (аминокислоты 161-185) на эквивалентную последовательность (выделенную курсивом и фланкирующую последовательность) из последовательности OspA серотипа 2 (штамм Pko; номер доступа GenBank S48322): последовательность, которая отличается от эпитопа hLFA-1;
- лидерную последовательность OspB (аминокислоты 1-15 Фигуры 2) и различные замещения для того, чтобы обеспечить отличия от известного уровня техники. Остатки аспарагина (N) и аспарагиновой кислоты (D) в положениях 44 и 46 были замещены на аспарагиновую кислоту (D) и аспарагин (N), соответственно, для получения последовательности KEKDKN (SEQ ID NO: 25). Остатки аланина (A) и аспарагиновой кислоты (D) в положениях 78 и 79 были замещены на треонин (T) и аспарагин (N), соответственно, для получения последовательности KTNKSK (SEQ ID NO: 26);
- стабилизацию мутаций, как описано в международной патентной публикации № WO 02/16421A2 (Luft & Dunn). Например, метионин (M) замещает аргинин (R) в положении аминокислоты 136 (R139M); тирозин (Y) замещает глутаминовую кислоту (E) в положении аминокислоты 157 (E160Y); и метионин (M) замещает лизин (K) в положении аминокислоты 186 (K189M); и
- дополнительную стабилизацию мутаций. Например, треонин (T) замещает валин (V) в положении аминокислоты 173 (аминокислота (aa) 176 описания). Удаление предположительного артритогенного эпитопа (положения 161-185) путем замещения последовательности B. burgdorferi на последовательность B. afzelii, приводит к разрыву водородной связи между аминокислотами 173 и 174 (aa 176 и 177 описания). Это приводит к ослаблению связывания с защитными моноклональными антителами (105,5 и LA-2 (Jiang et al., J. Immunol. 144: 284-9, 1990; Golde et al., Infect. Immun. 65: 882-9, 1997; и Ding et al., J. Mol. Biol. 302: 1153-64, 2000). Треонин (T) вводят в положение 173, вместо валина (V), для восстановления водородной связи и увеличения реакционной способности с защитными моноклональными антителами 105,5 и LA2.
Кроме того, аминокислоты 16-25 (начало зрелого белка) идентичны последовательности OspB (номер доступа GenBank X74810).
Нуклеотид и расшифрованные аминокислотные последовательности lipB sOspA 1/2251 приведены на Фигуре 3. Лидерная последовательность (зеленая) отщепляется во время секреции белка. Последовательность зрелого белка OspA начинается с цистеинового остатка (подчеркнут), который образует сайт присоединения липидного якоря белка.
Пример 4
Описание липидированного 6/4 OspA (lipB sOspA 6/4)
Целью исследований было конструирование нового антигена OspA, липидированного sOspA 6/4 OspA (lipB sOspA 6/4), содержащего серотипы 4 и 6. LipB sOspA 6/4 содержит проксимальный участок последовательности OspA серотипа 6 (штамм K48, номер доступа GenBank I40098), слитый с дистальным участком последовательности серотипа 4 (штамм pTroB; номер доступа GenBank I40089). Начало последовательности, уникальное для серотипа типа 4, представляет собой остаток аспарагина (N) в положении 217. Вторичные признаки показаны на аннотированной аминокислотной последовательности lipB sOspA 6/4 на Фигуре 4 и включают:
- стабилизацию мутаций, описанных в международной патентной заявке № WO 02/16421A2 (Luft и Dunn): метионин (M) вместо аргинина (R) в положении аминокислоты 136, тирозин (Y) вместо глутаминовой кислоты (E) в положении аминокислоты 157 и метионин (M) вместо лизина (K) в положении аминокислоты 187; и
- аналогично lipB sOspA 1/2251, описанному выше, используют лидерную последовательность OspB (аминокислоты 1-15 на Фигуре 4), и аминокислоты 16-25 являются идентичными последовательности OspB (номер доступа GenBank X74810).
Хотя пептидная последовательность KEKNKD (SEQ ID NO: 27) отсутствует в последовательности родительского типа OspA 6 (KEKDKD) (SEQ ID NO: 28), остаток аспарагиновой кислоты (D) в положении 46 замещают на остаток аспарагина (N) согласно эквивалентной замене, выполненной в конструкте lipB sOspA 1/2251 для получения последовательности KEKDKN (SEQ ID NO:25).
Хотя пептидная последовательность KADKSK (SEQ ID NO:29) отсутствует в последовательности родительского типа OspA 6 (KTDKSK) (SEQ ID NO: 30), остаток аспарагиновой кислоты (D) в положении 79 замещают на остаток аспарагина (N) согласно эквивалентной замене, выполненной в конструкте lipB sOspA 1/2251 для получения последовательности KTNKSK (SEQ ID NO:26).
Аминокислоту 37 меняют с глутаминовой кислоты (E), присутствующей в родительской последовательности (штамм K48; номер доступа GenBank I40098), на валин (V), потому что почти все последовательности типа 6 содержат валин в этом положении.
Нуклеотид и расшифрованные аминокислотные последовательности lipB sOspA 6/4 приведены на Фигуре 5. Лидерная последовательность (зеленая) отщепляется во время секреции белка. Последовательность зрелого белка OspA начинается с цистеинового остатка (подчеркнут, см. Фигуру 5), который образует сайт присоединения липидного якоря белка.
Пример 5
Описание липидированного 5/3 OspA (lipB sOspA 5/3)
Целью исследований было конструирование нового антигена OspA, липидированного sOspA 5/3 OspA (lipB sOspA 5/3), содержащего серотипы 3 и 5. LipB sOspA 5/3 содержит проксимальный участок последовательности OspA серотипа 5 [№ доступа в базе данных emb|X85441|BGWABOSPA, ген OspA B. garinii (субштамм WABSou)], слитый с дистальным участком последовательности серотипа 3 (штамм PBr; № доступа Genbank X80256, ген OspA B. garinii) с модификациями, указанными в SEQ ID NO: 5 и 6. Начало последовательности, уникальное для серотипа типа 3, представляет собой остаток аспарагиновой кислоты (D) в положении 216. Вторичные признаки показаны на аннотированной аминокислотной последовательности lipB sOspA 5/3 на Фигуре 6 и включают:
- стабилизацию мутаций, описанных в международной патентной заявке № WO 02/16421A2 (Luft and Dunn): метионин (M) вместо аргинина (R) в положении аминокислоты 136; тирозин (Y) вместо глутаминовой кислоты (E) в положении аминокислоты 157; и метионин (M) вместо лизина (K) в положении аминокислоты 187; и
- аналогично lipB sOspA 1/2251 и lipB sOspA 6/4, описанным выше, используют лидерную последовательность OspB (аминокислоты 1-15 на Фигуре 6), и аминокислоты 16-25 являются идентичными последовательности OspB (номер доступа GenBank X74810).
Хотя пептид последовательность KEKNKD (SEQ ID NO:27) отсутствует в последовательности родительского типа OspA 5 (KEKDKD) (SEQ ID NO: 28), остаток аспарагиновой кислоты (D) в положении 46 замещают на остаток аспарагина (N) согласно эквивалентной замене, выполненной в конструкте lipB sOspA 1/2251, с образованием последовательности KEKDKN (SEQ ID NO:25).
Хотя пептидная последовательность KADKSK (SEQ ID NO:29) отсутствует в последовательности родительского типа OspA 5 (KTDKSK) (SEQ ID NO: 30), остаток аспарагиновой кислоты (D) в положении 79 замещают на остаток аспарагина (N) согласно эквивалентной замене, выполненной в конструкте lipB sOspA 1/2251 с образованием последовательности KTNKSK (SEQ ID NO: 26).
Нуклеотид и расшифрованные аминокислотные последовательности lipB sOspA 5/3 приведены на Фигуре 7. Лидерная последовательность (зеленая) отщепляется во время секреции белка. Последовательность зрелого белка OspA начинается с кодона цистеина (подчеркнут, см. Фигуру 7), который образует сайт присоединения липидного якоря белка.
Пример 6
Оптимизация использования кодонов для высокого уровня экспрессии в E. coli
Поскольку известно, что присутствие кодонов, редко используемых в E. coli, является потенциальным препятствием для высокого уровня экспрессии чужеродных генов, малоиспользуемые кодоны были замещены на кодоны, используемые в генах E. coli с высокой экспрессией. Нуклеотидные последовательности новых генов OspA были сконструированы с использованием кодонов, наиболее часто встречающихся (предпочтительных кодонов) в гены E. coli класса II с высокой экспрессией (Guerdoux-Jamet et. al., DNA Research 4:257-65,1997). Данные по использованию кодонов в новых генах OspA и в генах класса II E. coli с высокой экспрессией приведены в Таблицах 7 и 8. Данные для реже используемых аминокислот, для которых молекулы тРНК с меньшей вероятностью являются ограничивающим скорость фактором, представлены отдельно (Таблица 7) от данных для аминокислот, встречающихся наиболее часто (Таблица 8).
Использование кодонов в новых генах OspA (реже встречающиеся аминокислоты*)
Использование кодонов в новых генах OspA (более распространенные аминокислоты)
Высокая степень согласия между использованием кодонов, выбранных для новых генов OspA (только обычные аминокислоты) и генов E. coli класса II является очевидной (т.е. график зависимости значений процентного содержания из Таблицы 8 для генов класса II по сравнению с индивидуальными новыми генами OspA; см. Фигуру 8). Для трех липидированных конструктов, исходные последовательности имеют содержание GC в интервале значений от 32,8% до 33,8%, в то время как оптимизированные по кодонам последовательности имеют содержание GC в интервале значений от 43,8% до 46,8%, что близко к 50% содержания GC для E. coli.
Пример 7
Конструирование синтетических нелипидированных генов OspA
Были также получены конструкты, не содержащие лидерной последовательности lipB. Два набора конструктов (липидированных и нелипидированных) необходимы для оценки простоты их продуцирования в ферментере (биомасса, стабильность, выход продукта и т.п.), для оценки легкости очистки разных типов антигена и для сравнения их биологических характеристик (профиль безопасности и эффективность защиты).
Конструкты (SEQ ID NO: 7, 9 и 11) были получены способом ПЦР-амплификации из каждого из трех конструктов lipB OspA (SEQ ID NO: 1, 3 и 5) с использованием ПЦР-праймеров с включенными сайтами рестрикции. Продукты ПЦР очищают, гидролизуют Nde I и BamH I и лигируют с гидролизованной векторной ДНК pET30a. Смеси лигирования используют для трансформации E. coli DH5α и проверяют последовательности OspA. Готовят Miniprep-ДНК, выделяют и используют для трансформации клеток-хозяев HMS 174(DE3). Последовательности нелипидированных производных идентичны липидированным вариантам, за исключением того, что в них отсутствуют первые 45 пар оснований, кодирующих лидерную последовательность, и содержат сайт Nde I, который содержит кодон метионина замещающий цистеиновый кодон в липидированных вариантах (см. Фигуру 9).
Пример 8
Экспрессия новых рекомбинантных антигенов OspA
Для экспрессии/продуцирования новых рекомбинантных генов OspA для использования в качестве антигенов, используют систему экспрессии E. coli, контролируемую РНК-полимеразой бактериофага T7 (Studier et al., J. Mol. Biol. 189:113-30, 1986). В этой системе экспрессии, новые гены OspA клонируют в сайт множественного клонирования одной из плазмид серии pET (например, pET30a). Поскольку экспрессия чужеродного гена находится под контролем промотора бактериофага T7, который не распознается РНК-полимеразой E. coli, экспрессия является зависимой от источника РНК-полимеразы T7. Этот фермент обеспечивается, когда рекомбинантные плазмиды перемещают в пригодного хозяина экспрессии, такого как E. coli HMS174(DE3), который содержит хромосомную копию гена T7 РНК-полимеразы. Экспрессия хромосомно интегрированного гена РНК-полимеразы T7 находится под контролем промотора lacUV5, который может быть включен (т.е. индуцирован) путем прибавления изопропил β-D-1-тиогалактопиранозида (IPTG) или лактозы (см. Фигуру 10). Следовательно, экспрессия чужеродного гена также регулируется путем добавления молекулы индуктора.
Клетки индуцируют на поздней лог-фазе и собирают через 3-4 часа после индукции. В индуцированных клетках, химерный антиген OspA был наиболее высокоэкспрессируемым белком, по результатам определения способом ДСН-ПААГ клеточных лизатов. Большая часть химер OspA была обнаружена в супернатанте. Загрязняющие белки E. coli удаляют анионообменной хроматографией и химерные белки OspA, элюируемые в объем пустот, концентрируют ультрафильтрацией.
Экспрессию новых рекомбинантных белков OspA из каждого из конструктов испытывают и образцы из индуцированных и неиндуцированных культур разделяют на полиакриламидном геле с добавкой ДСН (Фигура 11). Для липидированных (SEQ ID NO: 2, 4 и 6) и нелипидированных (SEQ ID NO: 8, 10 и 12) антигенов, в каждом случае наблюдалась полоса размером приблизительно 31 кДа (см. Фигуру 11). Белки охарактеризовывают и определенные молекулярные веса коррелируют (+/-0,5 дальтон) с теоретическими молекулярными весами, предполагая отщепление терминального метионина. Фигура 11 показывает, что экспрессируемые рекомбинантные липидированные белки OspA составляют по меньшей мере 10% от общего выхода белка, подтверждая пригодность конструктов для их предполагаемого назначения.
Пример 9
Отдельно взятый рекомбинантный антиген OspA (rOspA 1/2) защищает против инфекции B. burgdorferi s.s. и B. afzelii
Целью данных исследований было определение того, будет ли способен отдельно взятый рекомбинантный антиген (rOspA 1/2; полипептид, содержащий SEQ ID NO: 2 (lipB sOspA 1/2251)), сконструированный для сохранения защитных свойств серотипов OspA 1 и 2, индуцировать гуморальные реакции, защищающие мышей от инфекции B.burgdorferi s.s. (серотип OspA 1) или B. afzelii (серотип OspA 2). Приводятся данные, демонстрирующие, что включение дополнительных антигенов rOspA не оказывает антагонистического эффекта на защитный иммунитет, создаваемый антигеном rOspA 1/2.
Проектирование и конструкция rOspA 1/2. Для устранения риска внесения случайных агентов используют комплементарные перекрывающиеся синтетические олигонуклеотиды для получения фрагментов ДНК, которые лигируют вместе и клонируют в вектор pET30a, и последовательность верифицируют. Этот подход также позволяет оптимизировать использование кодонов для хозяина E. coli HMS174 (DE3), используемого для экспрессии гена OspA. Новый ген основан на проксимальном участке последовательности OspA серотипа 1 (аминокислоты 29-218, штамм B31; номер доступа GenBank X14407), слитым с дистальным участком последовательности серотипа 2 (аминокислоты 219-273, штамм PKo; номер доступа S48322). Фрагмент размером 25 аминокислот из штамма B31 B. burgdorferi (аминокислоты 164-188) замещают на последовательность из штамма PKo B. afzelii (аминокислоты 164-188), потому что этот участок B31 OspA (аминокислоты 165-173) имеет высокую степень родства с областью, окружающей эпитоп hLFA-1 (аминокислоты 332-340). N-Концевая последовательность, включая лидерную последовательность и первые 11 аминокислот, была получена из OspB (штамм B31; номер доступа GenBank X74810) для оптимизации экспрессии липидированного белка. Другие специфические изменения аминокислот были проведены для повышения иммуногенности и конформационной стабильности молекулы rOspA 1/2, и последовательность rOspA 1/2 (lipB sOspA 1/2251) приведена в SEQ ID NO: 2.
Испытания на животных. Способность отдельно взятого рекомбинантного антигена OspA (rOspA 1/2) предотвращать инфекцию двумя видами Borrelia, которые экспрессируют разные антигены OspA, оценивают на мышах C3H/HeJ, иммунизированных подкожно (дни 0 и 28) очищенным антигеном OspA (дозы 0,1 мкг или 0,03 мкг), приготовленным с 0,2% (мас/об) гидроксида алюминия в качестве адъюванта. Проводят контрольное заражение мышей через 2 недели после бустер-иммунизации, путем интрадермальной инъекции (заражение иглой; 7×104 клеток) или природным путем передачи инфекции (заражение от клещей). Для последних экспериментов, сажают 8 нимфальных клещей на мышь и позволяют им питаться в течение до 5 дней. Нимф собирают в окрестностях Ческе-Будеевице (Budweis) (Республика Чехия), регионе эндемической лаймской болезни. Большинство этих клещей были инфицированы B. afzelii по результатам определения способом испытаний некормленых клещей способом ПЦР. Инфекционный статус мышей определяют через четыре недели. В экспериментах с заражением от клещей, присутствие Borrelia подтверждают культивацией (мочевой пузырь) и путем детектирования Borrelia ДНК способом ПЦР в реальном масштабе времени (сердце). Эксперименты на животных проводят в соответствии с законодательством Австрии о проведении экспериментов на животных и международными рекомендациями (AAALAC и OLAW) и контролируются Институциональным комитетом по уходу за животными и их использованию (Institutional Animal Care and Use Committee) и они получали одобрение регуляторных органов Австрии. Иммуногенность. Гуморальный ответ (мкг IgG/мл) на антиген rOspA 1/2 определяют способом ELISA с использованием rOspA 1/2 в качестве покрывающего антигена и специфических моноклональных антител OspA (собственного производства) с определенным содержанием IgG в качестве стандарта.
Диагностические процедуры. Для экспериментов с заражением иглой, присутствие антител к консервативному эпитопу в поверхностно-экспонированном белке липопротеина VlsE (C6 ELISA; планшеты с покрытием фирмы Immunetics® C6 Lyme ELISA™) или к антигенам Borrelia, отличным от иммуногена OspA (вестерн-блоттинг) используют для диагностирования инфекции. Для вестерн-блоттинга используют клеточный лизат, приготовленный из штамма B. burgdorferi s.s. ZS7, поскольку он использовался в качестве организма контрольного заражения. Животные считаются инфицированными, если они были позитивными по результатам обоих анализов.
Для экспериментов с заражением от клещей проводились также анализы способами C6 ELISA и вестерн-блоттинга. Однако для вестерн-блоттинга были использованы лизаты B. burgdorferi s.s. ZS7, B. afzelii ACA1 и B. garinii KL11, потому что идентичность инфицирующего организма была неизвестной. Считалось, что животные подверглись сероконверсии, только если результаты обоих анализов были позитивными. Кроме того, инфекцию Borrelia оценивают по культуре из мочевого пузыря и путем детектирования нуклеиновых кислот B.burgdorferi s.l. в геномной ДНК, экстрагированной из ткани сердца, способом анализа ПЦР в реальном масштабе времени, нацеленного на 5’-участок OspA, и генетического анализа рРНК 16S. Животных определяют как ПЦР-позитивных, только если продукт ПЦР был детектирован обоими способами анализа. В общем, для определения животного как инфицированного, мыши должны были быть позитивными по результатам определения в культуре, способом ПЦР или серологии.
Характеризация инфицирующих Borrelia. В тех случаях, когда это возможно, инфицирующий организм культивируют и последовательность OspA и расшифрованную аминокислотную последовательность определяют для остатков 38-262 OspA (B. afzelii VS461, номер доступа GenBank Z29087). Эту информацию сравнивают с эталонными последовательностями OspA, чтобы можно было определить тип OspA и вид Borrelia. Для видов, экспрессирующих единственный серотип OspA, последовательность OspA для типичного штамма данного вида выбирают в качестве эталона, например, B. afzelii VS461 или B. valaisiana VS116 (номер доступа GenBank Z29087; AF095940). Поскольку B. garinii имеет множество типов OspA, используют последовательности OspA для генотипов OspA 3-7 (т.е. штаммы PBr, PTrob, WABSou, TlsI и T25; номера доступа GenBank X80256, X80186, X85441, X85440 и X80254, соответственно). Для типирования способом ПЦР в реальном масштабе времени, последовательность выравнивания гена OspA для 124 разновидностей B. burgdorferi s.l., депонированных в GenBank, проверяют на серотип-специфические последовательности и пригодные комбинации праймер-зонда конструируют с помощью Primer Express 3.0 (Applied Biosystems). Все анализы проводят на приборе для детектирования последовательностей ABI Prism® 7900HT с использованием универсальных условий циклирования.
Профилактика инфекции B.burgdorferi s.s (серотип OspA-1) путем иммунизации rOspA 1/2. Все мыши, иммунизированные низкими дозами двух разных партий антигена rOspA 1/2, вырабатывают IgG-антитела, специфические по отношению к иммуногену, по результатам определения способом ELISA. Никаких антител не обнаружено у контрольных мышей, получавших буфер композиции вакцины, содержащий гидроксид алюминия. Для оценки способности этого иммунного ответа предотвращать инфекцию B. burgdorferi s.s., вида, кодирующего OspA серотипа 1, мышам вводили интрадермальными инъекциями 7×104 клеток штамма ZS7 B. burgdorferi s.s. Все контрольные мыши, получавшие буфер, содержащий адъювант, продемонстрировали серологическое подтверждение инфекции, определенное способами C6 ELISA и вестерн-блоттинга. Ни одна из мышей, иммунизированных антигеном rOspA 1/2, не была инфицирована, и сыворотки этих мышей были негативными по результатам обоих анализов. Всего 0,03 мкг антигена rOspA 1/2, в виде композиции с гидроксидом алюминия в качестве адъюванта, при введении по схеме иммунизации двумя дозами, обеспечивает 100% защиты (P<0,0001, двусторонний точный критерий Фишера) от заражения иглой вирулентным штаммом ZS7 B. burgdorferi s.s.
Профилактика инфекции B. afzelii (серотип OspA-2) путем иммунизации rOspA 1/2. Для оценки способности иммунизации антигеном rOspA 1/2 для предотвращения инфекции B. afzelii, видом, кодирующим OspA серотипа 2, мышей иммунизируют, в двух отдельных экспериментах, с использованием тех же партий антигена и схемы исследований, что и в эксперименте с заражением иглой, описанном выше. Однако, в этом случае, проводят контрольное заражение иммунизированных мышей природными клещами (нимфы), определенными как инфицированные преимущественно B. afzelii. Способность этих природных клещей переносить мышам B. burgdorferi s.l. подтверждают путем контрольного заражения неиммунизированных контрольных животных.
Большинство контрольных мышей (в общей сложности 11/14, 79%) были инфицированы. Все инфицированные контрольные животные были позитивными по ДНК Borrelia по результатам двух независимых анализов способом ПЦР в реальном масштабе времени (16S-рРНК и гены OspA). В 10/11 случаев, удалось выделить Borrelia способом культуры мочевого пузыря. Остальные мыши были позитивными по результатам серологии и ПЦР. Для 9 из 10 культур изолятов, были выделены последовательности OspA, которые все были типированы как B. afzelii (>99% идентичности последовательностей OspA). Кроме того, все инфицирующие организмы были типированы как B. afzelii способом ПЦР-анализа ДНК, экстрагированной из сердца, способом ПЦР в реальном масштабе времени, специфически направленного на гены OspA серотипа 2. Эти данные подтверждают, что B. afzelii был основным видом Borrelia, переносимым инфицированными природными клещами мышам-хозяевам.
Несколько мышей, иммунизированных rOspA 1/2 (в общей сложности 3/32, 9%) были инфицированы. Из этих трех мышей, одна была инфицирована по результатам определения всех трех диагностических критериев (серология, ПЦР и культура), и анализ последовательности показал, что инфицирующим организмом был B. garinii серотип 6 (>99% идентичности последовательности OspA). Остальные два животных, считавшиеся инфицированными, были позитивными только по двум из трех критериев. Одна мышь была позитивной по серологии и ПЦР. Однако, инфицирующий организм не удалось выделить в культуре. Тем не менее, этот организм удалось типировать как B. garinii серотип 7 способом ПЦР-анализа ДНК, экстрагированной из сердца с использованием ПЦР, специфической по отношению к гену OspA серотипа 7. Третья мышь была позитивной по ПЦР и культуре, но серологически негативной. Культура изолята этой мыши принадлежала B. valaisiana по результатам определения способом секвенирования (идентичность последовательности OspA со штаммом VS116 B. valaisiana). Важно, что ни одна из иммунизированных мышей (0/32) не была инфицирована B. afzelii. Всего лишь 0,03 мкг антигена rOspA 1/2, в виде композиции с гидроксидом алюминия в качестве адъюванта, и при введении по схеме иммунизации двумя дозами, обеспечивают полную защиту против B. afzelii, переносимых природными клещами.
Заключение. Антиген отдельно взятого рекомбинантного внешнего поверхностного белка A (OspA), сконструированого таким образом, чтобы он содержал защитные элементы от двух разных серотипов OspA (1 и 2), был способен индуцировать гуморальные ответы, защищающие мышей от инфекции B. burgdorferi sensu stricto (OspA серотипа 1) или B. afzelii (OspA серотипа 2). Защита против инфекции B. burgdorferi s.s., штамм ZS7, была продемонстрирована в модели заражения иглой. Защита против вида B. afzelii была продемонстрирована на модели заражения от клещей с использованием природных клещей. В обеих моделях, всего лишь 0,03 мкг антигена, при введении по графику иммунизации двумя дозами с гидроксидом алюминия в качестве адъюванта, было достаточно для обеспечения полной защиты против видов-мишеней. Как и предполагалось, защита, создаваемая этим новым антигеном, не распространяется на другие виды Borrelia, что было продемонстрировано неспособностью антигена обеспечивать защиту против инфекции штаммами B. garinii и B. valaisiana. Такое подтверждение основного положения исследований показывает, что знание защитных эпитопов может быть использовано для рационального конструирования эффективных генетически-модифицированных вакцин, требующих использования меньшего количества антигенов OspA, и позволяет предположить, что этот подход может облегчить разработку вакцины OspA, пригодной для использования в глобальном масштабе.
Пример 10
Эффективность связывания мышиных анти-OspA антител с поверхностью живых Borrelia или ингибирования их роста коррелирует с защитой против заражения иглой штаммом B. burgdorferi s.s. типа 1
Целью данных исследований было установление соотношений корреляции при защите мышей, иммунизированных антигеном rOspA 1/2 в модели заражения иглой с использованием штамма Borrelia burgdorferi sensu stricto типа OspA 1. Анализируемыми параметрами была способность анти-OspA антител связываться с поверхностью живых Borreliae или ингибировать рост Borreliae.
98 мышей были преднамеренно иммунизированы субоптимальной дозой 3 нг антигена rOspA 1/2 с адъювантом 0,2% AI(OH3), которая была в 10 раз меньше низшей дозы, использованной в Примере 9, по схеме первичной бустерной дозы, так, чтобы при контрольном заражении можно было наблюдать как защищенных, так и инфицированных животных. Вакцинация проводилась подкожно с использованием объема дозы 100 мкл в дни 0, 14 и 28. В день 38, у 96 мышей брали образцы сыворотки перед контрольным заражением, и через 10 дней проводили контрольное заражение животных с использованием 19,4×ID50, выращенного в культуре B.burgdorferi s.s. ZS7, и инфекционный статус определяли через четыре недели. 71 из 96 мышей (72%) были определены как защищенные после иммунизации такой низкой дозой антигена.
Через четыре недели после контрольного заражения берут кровь для идентификации инфицированных мышей способом вестерн-блоттинга их сыворотки против мембранной фракции штамма ZS7 B.burgdorferi s.s. При использованных для контрольного заражения дозах, только инфицированные мыши имели гуморальный ответ на мембранные антигены штамма ZS7, отличные от OspA (ответ на OspA, индуцированные вакциной, не оценивался).
Количественное определение связывания антитела OspA с поверхностью живых Borreliae. В этом анализе, штамм B31 B. burgdorferi s.s., экспрессирующий OspA типа 1, инкубируют при постоянном разбавлении (1:100) со взятой перед контрольным заражением мышиной сывороткой при комнатной температуре в присутствии ЭДТА для предотвращения активации комплемента. После промывки для удаления несвязанного антитела, специфически связанные с поверхностью клеток антитела метят путем инкубации обработанных клеток с r-фикоэритрин-конъюгированным анти-мышиным Ig поликлональным антителом. После этого используют способ ДНК-окрашивания (LDS-751), дающий красную флуоресценцию, что облегчает детектирование, и бактерии затем анализируют способом проточной цитометрии (FACSCalibur, Beckton-Dickinson). Интенсивность флуоресценции, которая коррелирует с числом молекул антитела, связанных с поверхностью клеток, регистрируют для по меньшей мере 2000 индивидуальных Borreliae и рассчитывают среднюю интенсивность флуоресценции (MFI). Нормальная мышиная сыворотка служит негативным контролем для оценки величины неспецифического поверхностного связывания антител, в то время OspA серотип 1-специфическое mAb служит позитивным контролем для подтверждения идентичности типа OspA и для верификации уровня экспрессии OspA клетками в бактериальной культуре.
Анализ ингибирования роста бактерий. Для измерения способности взятой перед контрольным заражением сыворотки ингибировать рост Borreliae, штамм B31 B. burgdorferi s.s., экспрессирующий OspA типа 1, культивируют при 33°C в присутствии серийных разбавлений термоинактивированной взятой перед контрольным заражением или неиммунной мышиной сыворотки (негативный контроль) в присутствии комплемента (нормальная сыворотка морской свинки). Когда бактерии в контрольных культурах, инкубируемых с неиммунными сыворотками, в достаточной степени вырастут, при определении микроскопическим способом, проводят точный подсчет клеток с помощью анализа способом проточной цитометрии. Клеточные культуры смешивают с раствором, содержащим определенное число флуоресцентно-меченых бусин, и ДНК-краситель прибавляют для флуоресцентного мечения клеток Borrelia. Образцы обрабатывают с помощью проточного цитометра FACSCalibur до подсчета 100 бусин и рассчитывают абсолютные концентрации клеток (клеток/мл) путем сравнения числа событий в гейте бусин и в гейте Borreliae. Разбавление сыворотки, при котором рост бактерий ингибируется на 50%, рассчитывают по сравнению с NMS-контролем и приводят как титр GI-50. Стандартный препарат сыворотки используют для нормирования титров для разных анализов. Распределение измеренных параметров сыворотки сравнивают для инфицированных и защищенных животных с помощью непараметрического U-критерия Манна-Уитни (Graphpad Prism Vers. 5.0).
Результаты этих исследований (см. Фигуру 19) четко демонстрируют существование высокозначимой корреляции между содержанием функционального антитела в иммунной сыворотке во время контрольного заражения и защитой против инфекции высокой дозой (19,4×ID50) B. burgdorferi s.s. (ZS7) при заражении иглой. Измерения интенсивности флуоресценции способом FACS (возбужденная флуоресценция сортированных клеток) живых Borreliae, экспрессирующих OspA типа 1, которая отражает число молекул анти-OspA антител, связанных с поверхностью клеток, проводимые после инкубации бактерий с взятой перед контрольным заражением сывороткой при постоянном разбавлении, наилучшим образом коррелируют с защитой (p<0,0001, U-критерий Манна-Уитни). Однако титры ингибирования роста также высокозначимо коррелируют с защитой (p=0,0002, U-критерий Манна-Уитни, Фигура 19).
Пример 11
Эффективность связывания мышиных анти-OspA антител с поверхностью живых borrelia или ингибирования роста коррелирует с защитой против заражения от клещей с помощью штамма B. afzelii типа 2
Целью данных исследований было установление корреляций при защите мышей, иммунизированных химерным антигеном OspA 1/2 в модели заражения от клещей, использующей для инфицирования мышей природный путь передачи инфекции с помощью природных клещей, собранных в районе г. Ческе-Будеевице (Budweis) в Республике Чехии. Поскольку нимфальные клещи из данного эндемического региона инфицированы преимущественно B. afzelii, считается, что они обеспечивают контрольное заражение штаммом B. afzelii, OspA типа 2. Как указано в Примере 10, анализируемыми параметрами была способность анти-OspA антител связываться с поверхностью живых Borreliae или ингибировать рост Borreliae, обе из которых, как было показано, демонстрируют хорошую корреляцию с противодействием заражению иглой Borrelia bugdorferi s.s. Таким образом, данные исследования направлены на расширение применимости этих двух параметров как коррелирующих с защитой против природной инфекции B. afzelii, наиболее важным геновидом, ассоциированным с болезнями человека в Европе.
Сорок мышей иммунизируют субоптимальной 3 нг дозой антигена rOspA 1/2 с адъювантом 0,2% Al(OH)3), что в 10 раз меньше низшей дозы, использованной в Примере 9, по схеме первичной бустерной дозы. Как и в Примере 10, эту субоптимальню дозу выбирают для того, чтобы обеспечить наблюдение после контрольного заражения как защищенных, так и инфицированных животных. Вакцинация проводилась подкожно с использованием объема инъекции 100 мкл в дни 0, 14 и 28. В день 40, индивидуальные образцы крови брали у мышей для получения сыворотки перед контрольным заражением. Поскольку ограниченное число доступных клещей не позволяло провести контрольное заражение всех 40 мышей, выбрали 20 мышей на основании данных по поверхностному связыванию и концентраций анти-тип 2 IgG, чтобы охватить широкий спектр защитных реакций. На каждую мышь посадили восемь клещей и позволили им кормиться на этих мышах в течение 5 дней. Через четыре недели после контрольного заражения, мышей умертвили и инфекционный статус иммунизированных и контрольных мышей определили способом вестерн-блоттинга сыворотки против мембранных антигенов B. burgdorferi s.s., B. afzelii и B. garinii; культуры организмов Borrelia из мочевого пузыря; и путем детектирования Borrelia способом ПЦР в реальном масштабе времени в ДНК, экстрагированной из мочевого пузыря.
Количественное определение связывания антитела OspA с поверхностью живых Borreliae. В этом анализе, штамм Arcon B. afzelii, экспрессирующий OspA типа 2, инкубируют при постоянном разбавлении (1:100) с взятой перед контрольным заражением мышиной сывороткой при комнатной температуре в присутствии ЭДТА для предотвращения активации комплемента. После промывки для удаления несвязанного антитела, специфически связанные с поверхностью клеток антитела метят путем инкубации обработанных клеток с r-фикоэритрин-конъюгированным анти-мышиным Ig поликлональным антителом. Все последующие стадии анализа аналогичны описанным в Примере 10. Нормальная мышиная сыворотка служит негативным контролем для неспецифического связывания антитела. Высокий титр мышиной сыворотки против композиции трехкомпонентной рецептуры rOspА вакцины, вместе с OspA серотип 2-специфическими mAbs, служат позитивными контролями для подтверждения серотип-специфичности OspA и уровня экспрессии OspA клетками в бактериальной культуре.
Анализ ингибирования роста бактерий. Для измерения способности взятой перед контрольным заражением сыворотки ингибировать рост Borreliae, штамм Arcon B. afzelii, экспрессирующий OspA типа 2, культивируют при 33°C в присутствии серийных разбавлений термоинактивированной взятой перед контрольным заражением или неиммунной мышиной сыворотки (негативный контроль) без комплемента. В тех случаях, когда бактерии в контрольных культурах, инкубируемых с неиммунными сыворотками, в достаточной степени вырастут, при определении микроскопическим способом, проводят точный подсчет клеток путем анализа способом проточной цитометрии. Процедура, используемая для подсчета бактерий, аналогична описанной ранее для анализа ингибирования роста в Примере 10. Разбавление сыворотки, ингибирующее бактериальный рост на 50%, рассчитывают по сравнению с NMS-контролем и обозначают как титр GI-50. Стандартный препарат сыворотки используют для нормирования титров для разных анализов.
Статистический анализ. Сравнивают распределение измеренных параметров сыворотки у инфицированных и защищенных животных с помощью непараметрического U-критерия Манна-Уитни (Graphpad Prism Version 5.0).
Результаты. Из 20 животных, иммунизированных три раза по 0,003 пг rOspA 1/2 и контрольно зараженных 8 природными клещами, 7/20 (35%) были инфицированы. Из-за ограниченного количества клещей было невозможно определить точные показатели инфицирования при контрольном заражении контрольной группы неиммунизированных мышей. Однако, такое контрольное заражение не было необходимым для данных исследований, и типично в экспериментах с контрольным заражением природными клещами из Ческе-Будеевице (Budweis) достигался показатель инфицирования 70-80%.
Были обнаружены существенные различия между защищенными и инфицированными группами по результатам анализов поверхностного связывания (p=0,007) и ингибирования роста (p=0,03) (Фигура 20).
Заключение. В данных исследованиях было показано, что существует статистически значимая корреляция между содержанием функционального антитела в мышиной сыворотке во время контрольного заражения и защитой против природной инфекции при заражении 8 клещами на мышь. Измерения интенсивности флуоресценции способом FACS (возбужденная флуоресценция сортированных клеток) живых Borreliae, экспрессирующих OspA типа 2, которая отражает число молекул анти-OspA антител, связанных с поверхностью клеток, проведенные после инкубации бактерий с взятой перед контрольным заражением сывороткой при постоянном разбавлении, наилучшим образом коррелируют с защитой. Титры ингибирования роста также хорошо коррелируют с защитой. В отличие от штаммов Borrelia burgdorferi s.s., для эффективного уничтожения которых необходим комплемент, антиген rOspA1/2 индуцирует антитела, эффективно ингибирующие рост Borrelia даже в отсутствие комплемента.
Результаты исследований, представленных в Примерах 10 и 11, при их совместном рассмотрении, устанавливают in vitro параметры средней интенсивности флуоресценции (MFI) антител, поверхностно-связанных живыми Borreliae и титр GI-50 иммунных мышиных сывороток как «коррелирующие с защитой» в обоих примерах, для которых в настоящее время доступны модели активной защиты мышей (например, конкретнее, модель заражения иглой для штаммов B. burgdorferi s.s. OspA типа 1 и модель заражения от клещей для штаммов B. afzelii OspA типа 2). Более того, в отсутствие надежных моделей активной защиты для оценки защиты против гомологичных штаммов B. garinii, экспрессирующих OspA типов 3-6, путем умозаключений, вышеуказанные модели могут быть использованы как in vitro «суррогатные маркеры защиты» для оценки защитного потенциала и перекрестного покрытия штаммов различными композициями вакцин к штаммам, экспрессирующим все гомологичные типы OspA вакцины, и даже к экспрессирующим гетерологичные типы OspA. Действительно, при проведении исследований с использованием этих анализов функционального иммунного ответа на иммунных сыворотках мышей, иммунизированных 3-компонентной химерной композицией вакцины rOspA, были получены сравнимые величины MFI и титров GI-50 для B. garinii (OspA типов 3, 4, 5, 6) (см. Пример 13), что указывает, при использовании этих суррогатных маркеров защиты, что защитные ответы были также получены против штаммов, для которых в настоящее время не существует модели активной защиты для мышей. Кроме того, путем сравнения иммунных ответов у мышей, иммунизированных (а) индивидуальными химерными антигенами rOspA; или (б) любой одной из возможных комбинаций 2-компонентных композиций вакцин химерных антигенов rOspA; или (в) 3-компонентной композицией химерных антигенов rOspA, удалось показать, что последняя 3-компонентная вакцина была необходима для оптимального охвата штаммов, экспрессирующих OspA типов 1-6 (Пример 14). Более того, путем использования этих in vitro анализов суррогатных маркеров, удалось показать, что иммунные ответы, возникающие после иммунизации мышей композициями 3-компонентных вакцин химерных rOspA (rOspA 1/2, rOspA 6/4 и rOspA 5/3), действительно индуцируют функциональные иммунные ответы на все внутритиповые варианты (или субтипы) типов 1, 2, 3, 5 и 6, испытанных до настоящего времени (см. Пример 15) и даже на гетерологичные типы OspA,, отличные от гомологичных OspA типов 1-6, присутствующих в вакцине (см. Пример 16).
Пример 12
Мультивалентная рекомбинантная композиция OspA, содержащая 3 антигена (1/2, 6/4 и 5/3), проявляет высокую иммуногенность на мышах
Оценивают мультивалентную вакцину OspA (rOspA 1/2, rOspA 5/3 и rOspA 6/4) в модели заражения от клещей. Вакцина содержит комбинацию трех рекомбинантных антигенов OspA, содержащих защитные эпитопы из OspA серотипов 1 и 2 (SEQ ID NO: 2), OspA серотипов 6 и 4 (SEQ ID NO: 4) и OspA серотипов 5 и 3 (SEQ ID NO: 6).
Группы по десять самок мышей C3H/HeJ (возраст на момент иммунизации: 11 недель) иммунизируют подкожно в дни 0 и 28 фиксированной дозой 0,3 мкг мультивалентной вакцины (0,1 мкг каждого из rOspA 1/2, rOspA 5/3 и rOspA 6/4). Заражение от клещей выполняют, как описано выше, с использованием клещей из Ческе-Будеевице (Республика Чехия). Способность природных клещей переносить B.burgdorferi s.l. мышам подтверждают путем контрольного заражения неиммунизированных контрольных животных. Инфекционный статус контрольно-зараженных мышей определяют способами вестерн-блоттинга, ПЦР в реальном масштабе времени и культивацией.
Промежуточные образцы крови берут в день 41 путем орбитальной пункции. Конечные образцы крови (день 70/71) берут пункцией сердца. Индивидуальные сыворотки получают из цельной крови центрифугированием (10 минут; 1000-2000×G; комнатная температура (RT)). Сыворотки хранят при ≤ -20°C до использования.
В этом эксперименте некормленых клещей, взятых из той же партии, которая используется для контрольного заражения мышей, охарактеризовывают для определения общего показателя инфекционности и для подтверждения видов инфицирующих организмов. В случае тестирования 80 нимфальных клещей на присутствие ДНК B.burgdorferi s.l. способом ПЦР в реальном масштабе времени 16S-рРНК, 32,5% (26/80) были инфицированными. Серотип OspA мог быть определен способом ПЦР-ELISA для 22 из 26 инфицированных нимф; 86% (19/22) были типированы как B. afzelii и 14% (3/22) - как B.burgdorferi s.s.
Все неиммунизированные контрольные мыши (100%; 10/10) были инфицированы, в то время как только одна из мышей, иммунизированных мультивалентной вакциной rOspA, была инфицирована (10%; 1/10). Наблюдалось 100% согласие для разных способов, используемых для идентификации инфицированных мышей. Мультивалентная вакцина rOspA обеспечивает статистически высокозначимую защиту (p=0,00012; двусторонний точный критерий Фишера) по сравнению с контрольной группой.
Эти данные показывают, что иммунизация мультивалентной вакциной rOspA, содержащей антиген rOspA 1/2, способна предотвращать инфекцию B. afzelii, видом Borrelia, экспрессирующим OspA серотипа 2. Кроме этого, нет данных, указывающих на то, что включение дополнительных антигенов rOspA имеет антагонистический эффект на защитный иммунитет, создаваемый антигеном rOspA 1/2.
Эта вакцина обеспечивает защиту против переносимой клещами инфекции B. afzelii, эквивалентную наблюдающейся для антиген OspA 1/2; 0,3 мкг вакцины (0,1 мкг каждого антигена) в композиции с 0,2% Al(OH)3) при введении по графику двух доз обеспечивают 90% защиты по результатам определения способами вестерн-блоттинга, культивации Borrelia и детектирования ДНК Borrelia способом ПЦР.
Пример 13
Вакцина, представляющая собой трехкомпонентную вакцину (OspA 1/2, OspA 6/4 и OspA 5/3), индуцирует высокие уровни функциональных анти-OspA антител, которые связываются с и ингибируют рост штаммов Borrelia, экспрессирующих OspA типов 1-6
Поскольку было показано, что как поверхностное связывание (MFI), так и ингибирование роста (титры GI-50) хорошо коррелируют с защитой в модели заражения иглой (B. burgdorferi s.s.) (Пример 10) и в модели заражения мыши от клещей (B. afzelii) (Пример 11), данные исследования были предприняты для определения того, будут ли эквивалентные функциональные иммунные ответы индуцироваться 3-компонентной композицией вакцины химерных антигенов rOspA против B. garinii OspA серотипов 3-6, для которых отсутствует in vivo модель защиты для исследования эффективности вакцины.
Иммунизация мышей. Группы по 10 самок мышей C3H/HeJ иммунизируют подкожно три раза (день 0, день 14, день 28) 1:1:1 смесью rOspA-1/2, rOspA-6/4 и rOspA-5/3) в трех разных дозах (1, 0,1, 0,03 мкг белка на дозу) в сочетании с 0,2% Al(OH) 3) в качестве адъюванта. Получают сыворотку из образцов крови, взятых в день 40.
Количественное определение связывания антитела OspA с поверхностью живых Borreliae. В этом анализе, выращенные in vitro культуры шести репрезентативных штаммов Borrelia, экспрессирующих OspA типов 1-6 (B. burgdorferi sensu stricto B31/OspA-1; B. afzelii Arcon/OspA-2; B. garinii PBr/OspA-3; B. garinii DK6/OspA-4; B. garinii W/OspA-5; и B. garinii KL11/0spA-6) инкубируют при постоянном разбавлении (1:100) пулами мышиной сыворотки с пиковыми значениями титров при комнатной температуре в присутствии ЭДТА для предотвращения активации комплемента. Последующие процедуры промывки, мечения, детектирования и анализа были аналогичны описанным в Примере 10. Нормальная мышиная сыворотка служит негативным контролем для неспецифического связывания антител.
Анализ ингибирования роста бактерий. Для измерения способности взятой перед контрольным заражением сыворотки ингибировать рост Borreliae, шесть репрезентативных штаммов, экспрессирующих OspA типов 1-6 (B31, Arcon, PBr, DK6, W и KL11) культивируют при 33°C в присутствии серийных разбавлений пулов термоинактивированной сыворотки с пиковыми титрами или неиммунной мышиной сыворотки (негативный контроль). B31 культивируют в присутствии комплемента (сыворотка морской свинки), тогда как остальные пять штаммов тестируют в отсутствие комплемента. Затем снова анализы ингибирования роста проводят, как описано в Примере 10. Стандартный препарат сыворотки используют для нормирования титров для разных анализов.
Поверхностное связывание и эффективность ингибирования роста анти-OspA гуморального иммунного ответа. Интенсивное флуоресцентное окрашивание со значениями MFI в интервале от 50 до 200 наблюдалось для всех шести штаммов Borrelia при тестировании с тремя пулами сыворотки, полученной от групп с разными дозами иммунизации (1,0, 0,1 и 0,03 мкг белка на дозу) 3-компонентной вакциной при разбавлении 1:100 (Фигура 21). При тестировании пулов сыворотки от групп с 3 разными дозами на их способность ингибировать бактериальный рост, было обнаружено, что 3-компонентная вакцина также индуцирует высокие титры GI-50 ко всем шести типам штаммов OspA, в интервале значений от 1000 (штамм типа 4, доза 0,03 мкг) до 20000 (штамм типа 6).
Заключение. Взятые вместе, эти результаты демонстрируют, что антигены rOspA являются высокоиммуногенными и индуцируют большие количества функциональных антител, которые могут связываться с поверхностью живых Borreliae и ингибировать рост Borreliae. Степень покрытия для шести испытанных штаммов была полной, поскольку наблюдались высокие интенсивности флуоресценции и высокие титры ингибирования роста, сравнимые с уровнями, наблюдаемыми для OspA типов 1 и 2. В итоге, результаты, представленные в данных исследованиях, показывают, что гуморальные ответы, индуцированные трехкомпонентной вакциной rOspA (1/2+5/3+6/4), приготовленной в виде композиции с Al(OH)3 предотвращают инфекцию штаммами, экспрессирующими OspA типов 1-6, которые, как показали эпидемиологические исследования, теоретически охватывают более 99% изолятов, вызывающих болезни человека в Европе и Северной Америке и, таким образом, являются высокоэффективными для профилактики лайм-боррелиоза.
Пример 14
Вакцина, представляющая собой трехкомпонентную вакцину (OspA 1/2, OspA 6/4 и OspA 5/3), необходима для оптимального охвата Borrelia, экспрессирующих OspA типов 1-6
Целью данных исследований было изучение и сравнение иммуногенности и перекрестного покрытия штаммов по показателям функционального поверхностного связывания и/или ингибирования роста антителами, индуцированными одно- и мультикомпонентными композициями вакцин rOspA лайм-боррелиоза, также с использованием эффективности связывания анти-OspA антител с поверхностью живых Borreliae и ингибирования роста Borreliae in vitro в качестве параметров, коррелирующих с защитой.
Иммунизация мышей. Иммунизируют по десять самок мышей (C3H) на группу 0,1 мкг однокомпонентной вакцины, содержащей rOspA 1/2 антиген, rOspA 6/4 антиген, или rOspA 5/3 антиген; двухкомпонентной вакцины, содержащей 0,1 мкг обоих антигенов 1/2+5/3, антигенов 1/2+6/4, или антигенов 5/3+6/4; или трехкомпонентной вакцины, содержащей комбинацию 0,1 мкг всех трех 1/2+5/3+6/4 антигенов с адъювантом 0,2% AI(OH)3, по схеме первичной бустерной дозы. Вакцинация проводилась подкожно с использованием объема дозы 200 мкл в дни 0, 14 и 28. В день 42, индивидуальные образцы крови были взяты у мышей для получения сыворотки.
Анализы поверхностного связывания антител и ингибирования роста. Используют слегка модифицированный вариант анализа поверхностного связывания для определения эффективности связывания анти-OspA IgG с поверхностью живых Borreliae. Серийные разбавления пула сыворотки с определенными титрами MFI включают в анализы для построения калибровочной кривой, по которой определяют относительные титры тестируемых сывороток после интерполяции кривой нелинейной регрессии. Титр MFI стандартной сыворотки индивидуальных штаммов, экспрессирующих OspA типов 1-6, определяют как наибольшее разбавление, при котором определяемая интенсивность флуоресценции Borreliae по меньшей мере в 3 раза превышает интенсивность флуоресценции, наблюдаемую для нормальной мышиной сыворотки. Все определения проводят с двумя параллельными измерениями.
Диаграммы рассеяния, представленные на Фигуре 22, сравнивают титры MFI для шести штаммов, экспрессирующих гомологичные OspA типов, наблюдаемых для иммунных сывороток индивидуальных C3H мышей после иммунизации отдельно взятыми антигенами rOspA или комбинациями антигенов rOspA. Результаты показали, что композиция, содержащая все три антигена rOspA (1/2, 5/3 и 6/4) была необходима для индуцирования высоких титров MFI против всех шести штаммов Borrelia, экспрессирующих OspA типов 1-6, и что композиции, состоящие из двух антигенов rOspA (т.е. охватывающие четыре штамма) не обеспечивали полное покрытие для штаммов, экспрессирующих два типа OspA, не присутствующие в композиции.
Для определения эффективности различных комбинаций вакцин по индуцированию антитела ингибирующего рост, шесть репрезентативных штаммов Borreliae (B31, Arcon, PBr, DK6, W, KL11), экспрессирующих OspA типов 1-6, соответственно, культивируют при 33°C в присутствии пулов термоинактивированной иммунной или неиммунной мышиной сыворотки. Все сыворотки испытывают при единственном разбавлении. Использовали следующие разбавления: B31, PBr и KL11 1:200, Arcon, DK6 и W 1:100. PBr культивируют в отсутствие 20% комплемента, в то время как другие 5 штаммов испытывают в присутствии комплемента. Комплемент детенышей кролика используют для DK6, W и KL11, и сыворотку морской свинки используют для B31 и Arcon. Когда бактерии в контрольных культурах, инкубируемые с неиммунными сыворотками, вырастут в достаточной степени, при определении микроскопическим способом, проводят точный подсчет клеток, как описано ранее (см. Пример 10). Процент ингибирования роста бактерий рассчитывают по результатам подсчета клеток, полученным для тестируемой сыворотки, по сравнению с контролем нормальной мышиной сыворотки. Общее ингибирование роста, наблюдаемое для разных испытываемых композиций, затем представляют (Фигура 23) как число животных в разных группах по десять мышей C3H, которые продемонстрировали более чем 50% ингибирования роста. Результаты продемонстрировали, что 3-компонентная композиция была единственной композицией, способной индуцировать высокие титры ингибирующих рост антител против всех шести репрезентативных штаммов, экспрессирующих OspA типов 1-6 (Фигура 23). Во всех случаях, 3-компонентная композиция вакцины обеспечивала >50% ингибирования роста у >90% иммунизированных животных. 2-компонентные композиции вакцин не обеспечивали полного покрытия для двух штаммов, экспрессирующих OspA типов, не присутствующих в вакцине. Композиция, содержащая rOspA 1/2+6/4, не покрывает штамм типа 3; композиция, содержащая комбинацию rOspA 1/2+5/3, не покрывает типы 4 или 6; и композиция, содержащая rOspA 5/3+6/4, не покрывает тип 1.
Пример 15
Мультивалентные композиции вакцины OspA охватывают Borrelia, экспрессирующие внутритиповые варианты или субтипы OspA типов 1-6
Хотя в качестве основы для разработки и конструирования мультивалентной вакцины rOspA были выбраны OspA Borrelia типов 1-6, были также выделены Borreliae, экспрессирующие варианты белка OspA типов 1, 2, 3, 5 и 6. Такие варианты, хотя и классифицированные как принадлежащие к одному и тому же типу, имеют слегка измененные генные последовательности нуклеотидов и аминокислотные последовательности белков. Таким образом, существуют внутритиповые варианты или субтипы OspA типов 1, 2, 3, 5 и 6 (см. Фигуру 24). До настоящего времени не было обнаружено внутритиповых вариантов или субтипов для OspA типа 4.
Целью данных исследований было подтверждение того, что иммунная сыворотка, полученная при иммунизации мышей 3-компонентной мультивалентной вакциной rOspA, содержит функциональные антитела, которые могут связываться с поверхностью живых Borreliae, экспрессирующих такие внутритиповые варианты или субтипы.
Для данных исследований, получают пул мышиной иммунной сыворотки путем иммунизации 70 самок C3H мышей три раза по 0,3 мкг 3-компонентной мультивалентной вакцины rOspA в дни 0, 14 и 28. В день 42, мышей обескровливают и сыворотку получают и объединяют в пул. Пул иммунной сыворотки затем используют для тестирования связывания антител с поверхностью живых Borreliae. Культуры Borrelia инкубируют с пулом иммунной сыворотки или контрольной нормальной мышиной сывороткой в разбавлении 1:100 с двумя параллельными измерениями, и интенсивности флуоресценции измеренного для Borreliae связывания анти-OspA антител с бактериями контролируют с помощью анализов способом FACS, как описано выше.
Высокие уровни поверхностного связывания антитела (определяемые как интенсивность флуоресценции, более чем в 10 раз превышающая наблюдаемую для контрольной сыворотки неиммунизированных мышей) при разбавлении сыворотки 1:100, наблюдались для большинства штаммов, экспрессирующих OspA субтипов 1-6. В частности, высокие уровни связывания антител наблюдались для штаммов Borreliae, экспрессирующих субтипы OspA 1a, 1b, 1c, 1d, 1f, 1h, 1J, 1k и 1l; 2a, 2b, 2e, 2g, 2k, 2l и 2n; 3a, 3c, 3d и 3e; 5a и 5c; и 6a, 6e, 6f, 6g и 6k (Фигура 24). Более слабое связывание (определяемое как интенсивность флуоресценции, в 2-10 раз превышающая наблюдаемую для контрольной сыворотки неиммунизированных мышей) наблюдалось для штаммов Borreliae, экспрессирующих субтипы OspA 1g, 2j, 2m, 3b, 5d и 6l (Фигура 24), но такое слабое связывание было вызвано преимущественно низкой экспрессией белка OspA в используемых условиях роста.
Заключение. 3-компонентная химерная вакцина rOspA индуцирует функциональные связывающиеся с поверхностью антитела против всех внутритиповых вариантов или субтипов OspA типов 1, 2, 3, 5 и 6 у C3H мышей.
Пример 16
Мультивалентная композиция вакцины OspA обеспечивает защиту против других типов Borrelia, в дополнение к экспрессирующим OspA типов 1-6
Целью данных исследований было определение того, будет ли 3-компонентная композиция вакцины химерных антигенов rOspA (содержащая все 3 химерных антигена - 1/2, 6/4 и 3/5) также обеспечивать защиту против Borrelia, экспрессирующих OspA типов, отличных от гомологичных OspA типов 1-6. 40 C3H мышей иммунизируют три раза по 0,3 мкг 3-компонентной вакцины в дни 0, 14 и 28. В день 42, мышей обескровливают и готовят пул сыворотки, который используют для оценки эффективности поверхностного связывания и ингибирования роста против штаммов, экспрессирующих гетерологичные типы OspA.
Результаты этих исследований показали, что 3-компонентная химерная вакцина rOspA действительно индуцирует антитела, которые связываются с поверхностью Borreliae и ингибируют рост других типов Borreliae, включая штаммы B. spielmanii, B. valaisiania, B. lusitaniae и B. japonica (см. Таблицу 9). В случае B. garinii, экспрессирующего OspA типа 7 наблюдалось только слабое поверхностное связывание и незначительное ингибирование роста или его отсутствие; однако, это слабое связывание и маленькая величина ингибирования роста могут быть вызваны низкими уровнями экспрессии OspA в используемых условиях культуры in vitro, а не отсутствием связывания антител иммунной сыворотки.
Поверхностное связывание и ингибирование роста
других типов Borreliae
-: отсутствие значительное связывания/ингибирования роста
(+-): низкая интенсивность поверхностного связывания
Пример 17
Мультивалентные композиции вакцины OspA индуцируют антитела к общему эпитопу на N-конце молекулы OspA, которые могут способствовать защите против любого штамма Borrelia, экспрессирующего OspA
В ходе исследований защитной эффективности композиций мультивалентных химерных rOspA, было получено моноклональное антитело (F237/BK2) против 2-компонентной вакцины rOspA, содержащей rOspA-1/2 и rOspA-6/4. Способом анти-OspA ELISA было показано, что F237/BK2 связывается со всеми исследованными на сегодня типами OspA (OspA типов 1-7), а также с 3 химерными антигенами rOspA (rOspA-1/2, rOspA-5/3 и rOspA-6/4). Такие результаты показывают, что F237/BK2 распознает общий эпитоп, присутствующий во всех молекулах OspA. Более того, предварительные исследования по картированию эпитопа показывают, что этот общий эпитоп расположен в менее вариабельной N-концевой половине молекулы (т.е. в сторону N-конца от аминокислоты 130), где обычно наблюдается гомологичность последовательностей OspA.
Интересно, что было также продемонстрировано связывание F237/BK2 с поверхностью Borreliae, экспрессирующих гомологичные OspA типов 1-6 и гетерологичные типы OspA, включая экспрессируемые B. spielmanii, B. valaisiania и B. japonica, хотя и менее эффективное, чем для моноклональных антител, направленных против специфических эпитопов C-концевого типа. Было также обнаружено, с использованием способов, аналогичных описанным в предыдущих примерах, что F237/BK2 ингибирует рост репрезентативных штаммов, экспрессирующих OspA типов 1, 2, 4, 5 и 6.
При испытаниях F237/BK2 в in vivo модели пассивной защиты на мышах наблюдали, что F237/BK2 создает защиту против заражения природными клещами, соответствующего контрольному заражению B. afzelii типа 2. Собирают в Вундшу (Wundschuh) (Штирия, Австрия) клещей, о которых известно, что они инфицированы преимущественно B. afzelii. Десяти самкам C3H мышей делают интраперитонеально инъекции 500 мкг аффинно-очищенного mAb F237/BK2. Через два часа сажают 10 пассивно иммунизированным мышам по 8 клещей на животное, а также 10 псевдо-иммунизированным животным. Через четыре дня накормленных клещей удаляют. В день 90 мышей умерщвляют и анализируют на инфекцию с помощью серологического тестирования, ПЦР-анализа и культуры Borrelia, как описано выше. В группе, получавшей F237/BK2, ни одно животное не было инфицировано, в то время как в контрольной группе 5 животных (50%) были инфицированы B. afzelii. Таким образом, моноклональное антитело F237/BK2 обеспечивает статистически значимую (p = 0,0325) пассивную защиту против заражения от клещей по сравнению с псевдо-иммунизированными контрольными мышами. Это первое сообщение о том, что моноклональное антитело, которое связывается с общим эпитопом в N-концевой половине молекулы, принимает участие в защите. Более того, если вакцина может индуцировать антитела, распознающие такой общий эпитоп, то такое антитело будет, несомненно, усиливать перекрестную эффективность защиты вакцины.
Для проверки того, действительно ли такие антитела индуцируются 3-компонентными композициями вакцин химерных rOspA, проводили анализ ингибирования моноклональным антителом способом ELISA с использованием пероксидаза-меченого F237/BK2. В этих экспериментах, белок GST-OspA типа 3 используют в качестве покрывающего антигена, и нормальную мышиную сыворотку или пул сыворотки C3H мышей, иммунизированных три раза 3-компонентной химерной вакциной rOspA, добавляют в лунки при разбавлении 1:100. Через шестьдесят минут прибавляют пероксидаза-меченный F237/BK2 в предварительно оптимизированной концентрации для получения в конечном счете величины оптической плотности (OD), равной приблизительно 1 для контрольной неингибирующей нормальной мышиной сыворотки, и инкубацию продолжают в течение еще 60 мин. Наконец, планшеты для ELISA промывают и проявляют субстратом TMB.
С помощью такого анализа ингибирования моноклонального антитела способом ELISA можно продемонстрировать, что 3-компонентная композиция химерного rOspA действительно индуцирует антитела, которые связываются с эпитопом, идентичным с или очень близким к эпитопу, распознаваемому mAb F237/BK2. Значения OD были значительно сниженными (например, типично на 20-30%) анти-OspA иммунными сыворотками по сравнению с контрольной неингибирующей нормальной мышиной сывороткой.
Заключение. Данные исследования показывают, что 3-компонентная химерная вакцина rOspA способна индуцировать как типоспецифический, так и широкий перекрестный защитный иммунный ответ.
Пример 18
Дополнительные синтетические молекулы нуклеиновой кислоты и полипептида OspA
Целью исследований было конструирование дополнительных новых антигенов OspA, включающих серотипы 1 и 2, 6 и 4 и 5 и 3, соответственно. Были сконструированы три синтетических гена OspA (SEQ ID NO: 168 (orig sOspA 1/2), 170 (orig sOspA 6/4) и 172 (orig sOspA 5/3)), кодирующие полипептидные молекулы OspA с защитными эпитопами OspA серотипов 1 и 2 (orig sOspA 1/2), OspA серотипов 6 и 4 (orig sOspA 6/4) и OspA серотипов 5 и 3 (orig sOspA 5/3) Borrelia. Первичные аминокислотные последовательности этих молекул (SEQ ID NO: 169, 171 и 173, соответственно) приведены в Таблице 1. Эти последовательности представляют собой исходные химерные конструкты, т.е. без мутаций и без оптимизации кодонов.
Пример 19
Мультивалентная композиция рекомбинантных OspA, содержащая 3 антигена (1/2, 6/4 и 5/3), является иммуногенной на мышах
Мультивалентную вакцину OspA, содержащую композиции исходных конструктов без оптимизации кодонов и без мутаций (orig OspA 1/2, orig OspA 5/3 и orig OspA 6/4), оценивают в модели заражения от клещей. Вакцина содержит комбинацию трех рекомбинантных антигенов OspA, содержащих защитные эпитопы OspA серотипов 1 и 2 (SEQ ID NO: 169), OspA серотипов 6 и 4 (SEQ ID NO: 171) и OspA серотипов 5 и 3 (SEQ ID NO: 173).
Группы по десять самок мышей C3H/HeJ (возраст на момент иммунизации: 11 недель) иммунизируют подкожно в дни 0 и 28 фиксированной дозой 0,3 мкг мультивалентной вакцины (по 0,1 мкг каждого из orig OspA 1/2, orig OspA 5/3 и orig OspA 6/4). Заражение от клещей проводят, как описано выше, с использованием клещей из Ческе-Будеевице (Республика Чехия). Способность природных клещей переносить B.burgdorferi s.l. мышам подтверждают путем контрольного заражения неиммунизированных контрольных животных. Инфекционный статус контрольно-зараженных мышей определяется способами вестерн-блоттинга, ПЦР в реальном масштабе времени и культивации.
Промежуточные образцы крови берут в день 41 путем орбитальной пункции. Конечные образцы крови (день 70/71) собирают пункцией сердца. Индивидуальные сыворотки получают из цельной крови центрифугированием (10 минут; 1000-2000×G; комнатная температура). Сыворотки хранят при ≤ -20°C до использования.
В этом эксперименте некормленых клещей, взятых из той же партии, что и используемые для контрольного заражения мышей, охарактеризовывают для определения общего показателя инфекционности и для подтверждения вида инфицирующих организмов.
Пример 20
Вакцина, содержащая трехкомпонентную вакцину (orig OspA 1/2, orig OspA 6/4 и orig OspA 5/3), индуцирует высокие уровни функциональных анти-OspA антител, которые связываются с и ингибируют рост штаммов Borrelia, экспрессирующих OspA типов 1-6
Результаты, представленные в Примере 13 показывают, что гуморальные ответы, индуцированные трехкомпонентной вакциной rOspA (lipB sOspA1/2+lipB sOspA 5/3+lipB sOspA 6/4), приготовленной в виде композиции с Al(OH)3, предотвращают инфекцию штаммами, экспрессирующими OspA типов 1-6 и, таким образом, являются эффективными при профилактике лайм-боррелиоза. Таким образом, данные исследования проводятся для определения того, будут ли индуцироваться эквивалентные функциональные иммунные ответы трехкомпонентной вакциной OspA, содержащей химерные исходные (orig) антигены OspA (orig sOspA1/2+orig sOspA 5/3+orig sOspA 6/4).
Иммунизация мышей. Группы по 10 самок мышей C3H/HeJ иммунизируют подкожно три раза (день 0, день 14, день 28) 1:1:1 смесью orig sOspA1/2+orig sOspA 5/3+orig sOspA 6/4) в трех разных дозах (1, 0,1, 0,03 мкг белка на дозу), в сочетании с 0,2% Al(OH)3 в качестве адъюванта. Сыворотку получают из образцов крови, взятых в день 40.
Количественное определение связывания антитела OspA с поверхностью живых Borreliae. В этом анализе, выращенные in vitro культуры шести репрезентативных штаммов Borrelia, экспрессирующих OspA типов 1-6 (B. burgdorferi sensu stricto B31/OspA-1; B. afzelii Arcon/OspA-2; B. garinii PBr/OspA-3; B. garinii DK6/OspA-4; B. garinii W/OspA-5; и B. garinii KL11/0spA-6) инкубируют при постоянном разбавлении (1:100) пулами мышиной сыворотки с пиковыми значениями титров при комнатной температуре в присутствии ЭДТА для предотвращения активации комплемента. Последующие процедуры промывки, мечения, детектирования и анализа аналогичны описанным в Примерах 10 и 13. Нормальная мышиная сыворотка служит негативным контролем для неспецифического связывания антител.
Анализ ингибирования роста бактерий. Для измерения способности взятой перед контрольным заражением сыворотки ингибировать рост Borreliae, шесть репрезентативных штаммов, экспрессирующих OspA типов 1-6 (B31, Arcon, PBr, DK6, W и KL11) культивируют при 33°C в присутствии серийных разбавлений пулов термоинактивированной сыворотки с пиковыми титрами или неиммунной мышиной сыворотки (негативный контроль). B31 культивируют в присутствии комплемента (сыворотка морской свинки), тогда как остальные пять штаммов тестируют в отсутствие комплемента. Анализы ингибирования роста проводят, как описано в Примерах 10 и 13. Стандартный препарат сыворотки используется для нормирования титров для разных анализов.
Поверхностное связывание и эффективность ингибирования роста анти-OspA гуморального иммунного ответа. Флуоресцентное окрашивание измеряют для всех шести штаммов Borrelia при тестировании с тремя пулами сыворотки, полученной от групп с разными дозами иммунизации (1,0, 0,1 и 0,03 мкг белка на дозу) 3-компонентной вакцины при разбавлении 1:100.
Пример 21
Вакцина, содержащая трехкомпонентную вакцину (OspA 1/2, OspA 6/4 и OspA 5/3), необходима для оптимального покрытия Borrelia, экспрессирующих OspA типов 1-6
Целью данных исследований является исследование и сравнение иммуногенности и перекрестного покрытия штаммов для способных к функциональному поверхностному связыванию и/или ингибированию роста антител, индуцированных одно- и мультикомпонентными композициями вакцины лайм-боррелиоза orig sOspA, с использованием эффективности связывания анти-OspA антител с поверхностью живых Borreliae и ингибирования роста Borreliae in vitro в качестве параметров, коррелирующих с защитой.
Иммунизация мышей. Десять самок мышей (C3H) на группу иммунизируют 0,1 мкг однокомпонентной вакцины, содержащей антиген orig sOspA1/2, антиген orig sOspA 5/3 или антиген orig sOspA 6/4; двухкомпонентной вакцины, содержащей 0,1 мкг обоих из антигенов 1/2+5/3, антигенов 1/2+6/4, или антигенов 5/3+6/4; или трехкомпонентной вакцины, содержащей комбинацию 0,1 мкг всех трех антигенов 1/2+5/3+6/4 с адъювантом 0,2% AI(OH)3 по схеме первичной бустерной дозы. Вакцинацию проводят подкожно с использованием объема дозы 200 мкл в дни 0, 14 и 28. В день 42 у мышей берут индивидуальные образцы крови для получения сыворотки.
Поверхностное связывание антител и анализы ингибирования роста. Слегка модифицированный вариант анализа поверхностного связывания, описанного выше, используется для определения эффективности связывания анти-OspA IgG с поверхностью живых Borreliae. Серийные разбавления пула сыворотки с определенными титрами MFI включают в анализы для построения калибровочной кривой, по которой определяют относительные титры тестируемых сывороток после интерполяции кривой нелинейной регрессии. Титр MFI стандартной сыворотки для индивидуальных штаммов, экспрессирующих OspA типов 1-6, определяется как наибольшее разбавление, при котором определяемая интенсивность флуоресценции Borreliae по меньшей мере в 3 раза превышает интенсивность флуоресценции, наблюдаемую для нормальной мышиной сыворотки. Все определения проводятся с двумя параллельными измерениями.
Для определения активности различных комбинаций вакцин при индуцировании ингибирующих рост антител, шесть репрезентативных штаммов Borreliae (B31, Arcon, PBr, DK6, W, KL11), экспрессирующих OspA типов 1-6, соответственно, культивируют при 33°C в присутствии пулов термоинактивированной иммунной или неиммунной мышиной сыворотки. Все сыворотки анализируют при единственном разбавлении. Используются следующие разбавления: B31, PBr и KL11-1:200, Arcon, DK6 и W-1:100. PBr культивируют в отсутствие 20% комплемента, в то время как другие 5 штаммов тестируют в присутствии комплемента. Для DK6, W и KL11 используют комплемент детенышей кролика, а для B31 и Arcon используют сыворотку морской свинки. Когда бактерии в контрольных культурах, инкубируемые с неиммунными сыворотками, вырастут в достаточной степени, при определении микроскопическим способом, проводят точные подсчеты клеток, как описано ранее (см. Пример 10). Процент ингибирования роста бактерий рассчитывают по результатам подсчета клеток, полученным для исследуемой сыворотки, по сравнению с контрольной нормальной мышиной сывороткой. Общее ингибирование роста, наблюдаемое для разных испытываемых композиций, представляют затем как число животных в разных группах по десять C3H-мышей, продемонстрировавших более 50% ингибирования роста.
Пример 22
Мультивалентная композиция вакцины OspA обеспечивает покрытие Borrelia, экспрессирующих внутритиповые варианты или субтипы OspA типов 1-6
Целью данных исследований было подтверждение того, что иммунная сыворотка, полученная в результате иммунизации мышей 3-компонентной мультивалентной вакциной orig OspA (orig sOspA 1/2, orig sOspA 6/4 и orig sOspA 5/3), содержит функциональные антитела, которые могут связываться с поверхностью живых Borreliae, экспрессирующих такие внутритиповые варианты или субтипы.
Для данных исследований, готовят пул мышиной иммунной сыворотки путем иммунизации 70 самок C3H мышей три раза по 0,3 мкг 3-компонентной мультивалентной вакцины orig OspA в дни 0, 14 и 28. В день 42 мышей обескровливают, получают сыворотку и объединяют. Пул иммунной сыворотки затм используют для тестирования связывания антител с поверхностью живых Borreliae. Культуры Borrelia инкубируют с пулом иммунной сыворотки или контрольной нормальной мышиной сывороткой в разбавлении 1:100 с двумя параллельными измерениями и интенсивности флуоресценции Borreliae при измерениях связывания анти-OspA антител с бактериями контролируют с помощью анализов способом FACS, как описано выше.
Изобретение было описано на примере конкретных вариантов воплощения, подтверждено или предположительно включающих конкретные способы реализации данного изобретения. Различные модификации и варианты описанного изобретения будут очевидны квалифицированным специалистам в данной области техники без выхода за пределы объема и сущности изобретения. Хотя изобретение было описано в связи с конкретными вариантами воплощения, следует понимать, что заявляемое изобретение не должно ненадлежащим образом ограничиваться такими конкретными вариантами воплощения. Вместо этого, различные модификации описанных способов воплощения изобретения, очевидные для квалифицированных специалистов в соответствующих областях техники, должны рассматриваться как входящие в объем приложенной формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CROWE, et al.
<120> ХИМЕРНЫЕ ГЕНЫ OSPA, БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> 31315/44557A1
<150> 61/334,901
<151> 2010-05-14
<160> 173
<170> Патентная версия 3.5
<210> 1
<211> 829
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 1
catatgcgtc tgttgatcgg ctttgctctg gcgctggctc tgatcggctg cgcacagaaa 60
ggtgctgagt ctattggttc cgtttctgta gatctgcccg gtgaaatgaa ggttctggtg 120
agcaaagaaa aagacaagaa cggcaagtac gatctcatcg caaccgtcga caagctggag 180
ctgaaaggta cttctgataa aaacaacggc tctggtgtgc tggagggcgt caaaactaac 240
aagagcaaag taaagcttac gatctctgac gatctcggtc agaccacgct ggaagttttc 300
aaagaggatg gcaagaccct cgtgtccaaa aaagtaactt ccaaagacaa gtcctctacg 360
gaagaaaaat tcaacgaaaa aggtgaggtg tctgaaaaga tcatcaccat ggcagacggc 420
acccgtcttg aatacaccgg tattaaaagc gatggtaccg gtaaagcgaa atatgttctg 480
aaaaacttca ctctggaagg caaagtggct aatgataaaa ccaccttgga agtcaaggaa 540
ggcaccgtta ctctgagcat gaatatctcc aaatctggtg aagtttccgt tgaactgaac 600
gacactgaca gcagcgctgc gactaaaaaa actgcagcgt ggaattccaa aacttctact 660
ttaaccatta gcgttaacag caaaaaaact acccagctgg tgttcactaa acaagacacg 720
atcactgtgc agaaatacga ctccgcaggc accaacttag aaggcacggc agtcgaaatt 780
aaaacccttg atgaactgaa aaacgcgctg aaataagctg agcggatcc 829
<210> 2
<211> 270
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 2
Met Arg Leu Leu Ile Gly Phe Ala Leu Ala Leu Ala Leu Ile Gly Cys
1 5 10 15
Ala Gln Lys Gly Ala Glu Ser Ile Gly Ser Val Ser Val Asp Leu Pro
20 25 30
Gly Glu Met Lys Val Leu Val Ser Lys Glu Lys Asp Lys Asn Gly Lys
35 40 45
Tyr Asp Leu Ile Ala Thr Val Asp Lys Leu Glu Leu Lys Gly Thr Ser
50 55 60
Asp Lys Asn Asn Gly Ser Gly Val Leu Glu Gly Val Lys Thr Asn Lys
65 70 75 80
Ser Lys Val Lys Leu Thr Ile Ser Asp Asp Leu Gly Gln Thr Thr Leu
85 90 95
Glu Val Phe Lys Glu Asp Gly Lys Thr Leu Val Ser Lys Lys Val Thr
100 105 110
Ser Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu Lys Gly Glu
115 120 125
Val Ser Glu Lys Ile Ile Thr Met Ala Asp Gly Thr Arg Leu Glu Tyr
130 135 140
Thr Gly Ile Lys Ser Asp Gly Thr Gly Lys Ala Lys Tyr Val Leu Lys
145 150 155 160
Asn Phe Thr Leu Glu Gly Lys Val Ala Asn Asp Lys Thr Thr Leu Glu
165 170 175
Val Lys Glu Gly Thr Val Thr Leu Ser Met Asn Ile Ser Lys Ser Gly
180 185 190
Glu Val Ser Val Glu Leu Asn Asp Thr Asp Ser Ser Ala Ala Thr Lys
195 200 205
Lys Thr Ala Ala Trp Asn Ser Lys Thr Ser Thr Leu Thr Ile Ser Val
210 215 220
Asn Ser Lys Lys Thr Thr Gln Leu Val Phe Thr Lys Gln Asp Thr Ile
225 230 235 240
Thr Val Gln Lys Tyr Asp Ser Ala Gly Thr Asn Leu Glu Gly Thr Ala
245 250 255
Val Glu Ile Lys Thr Leu Asp Glu Leu Lys Asn Ala Leu Lys
260 265 270
<210> 3
<211> 832
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 3
catatgcgtc tgttgatcgg ctttgctctg gcgctggctc tgatcggctg cgcacagaaa 60
ggtgctgagt ctattggttc cgtttctgta gatctgcccg gtggcatgac cgttctggtc 120
agcaaagaaa aagacaaaaa cggtaaatac agcctcgagg cgaccgtcga caagcttgag 180
ctgaaaggca cctctgataa aaacaacggt tccggcaccc tggaaggtga aaaaactaac 240
aaaagcaaag tgaaactgac cattgctgat gacctcagcc agaccaaatt cgaaattttc 300
aaagaagatg ccaaaacctt agtatccaaa aaagtgaccc tgaaagacaa gtcctctacc 360
gaagaaaaat tcaacgaaaa gggtgaaacc tctgaaaaaa ccatcgtaat ggcaaatggt 420
acccgtctgg aatacaccga catcaaaagc gatggctccg gcaaagccaa atacgttctg 480
aaagacttca ccctggaagg caccctcgct gccgacggca aaaccacctt gaaagttacc 540
gaaggcactg ttgttttaag catgaacatc ttaaaatccg gtgaaatcac cgttgcgctg 600
gatgactctg acaccactca ggccactaaa aaaaccggca aatgggattc taacacttcc 660
actctgacca tcagcgtgaa ttccaaaaaa actaaaaaca tcgtgttcac caaagaagac 720
accatcaccg tccagaaata cgactctgcg ggcaccaacc tcgaaggcaa cgcagtcgaa 780
atcaaaaccc tggatgaact gaaaaacgct ctgaaataag ctgagcggat cc 832
<210> 4
<211> 271
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 4
Met Arg Leu Leu Ile Gly Phe Ala Leu Ala Leu Ala Leu Ile Gly Cys
1 5 10 15
Ala Gln Lys Gly Ala Glu Ser Ile Gly Ser Val Ser Val Asp Leu Pro
20 25 30
Gly Gly Met Thr Val Leu Val Ser Lys Glu Lys Asp Lys Asn Gly Lys
35 40 45
Tyr Ser Leu Glu Ala Thr Val Asp Lys Leu Glu Leu Lys Gly Thr Ser
50 55 60
Asp Lys Asn Asn Gly Ser Gly Thr Leu Glu Gly Glu Lys Thr Asn Lys
65 70 75 80
Ser Lys Val Lys Leu Thr Ile Ala Asp Asp Leu Ser Gln Thr Lys Phe
85 90 95
Glu Ile Phe Lys Glu Asp Ala Lys Thr Leu Val Ser Lys Lys Val Thr
100 105 110
Leu Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu Lys Gly Glu
115 120 125
Thr Ser Glu Lys Thr Ile Val Met Ala Asn Gly Thr Arg Leu Glu Tyr
130 135 140
Thr Asp Ile Lys Ser Asp Gly Ser Gly Lys Ala Lys Tyr Val Leu Lys
145 150 155 160
Asp Phe Thr Leu Glu Gly Thr Leu Ala Ala Asp Gly Lys Thr Thr Leu
165 170 175
Lys Val Thr Glu Gly Thr Val Val Leu Ser Met Asn Ile Leu Lys Ser
180 185 190
Gly Glu Ile Thr Val Ala Leu Asp Asp Ser Asp Thr Thr Gln Ala Thr
195 200 205
Lys Lys Thr Gly Lys Trp Asp Ser Asn Thr Ser Thr Leu Thr Ile Ser
210 215 220
Val Asn Ser Lys Lys Thr Lys Asn Ile Val Phe Thr Lys Glu Asp Thr
225 230 235 240
Ile Thr Val Gln Lys Tyr Asp Ser Ala Gly Thr Asn Leu Glu Gly Asn
245 250 255
Ala Val Glu Ile Lys Thr Leu Asp Glu Leu Lys Asn Ala Leu Lys
260 265 270
<210> 5
<211> 829
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 5
catatgcgtc tgttgatcgg ctttgctttg gcgctggctt taatcggctg tgcacagaaa 60
ggtgctgagt ctattggttc cgtttctgta gatctgcccg ggggtatgaa agttctggta 120
agcaaagaaa aagacaaaaa cggtaaatac agcctgatgg caaccgtaga aaagctggag 180
cttaaaggca cttctgataa aaacaacggt tctggcaccc tggaaggtga aaaaactaac 240
aaaagcaaag taaagcttac tattgctgag gatctgagca aaaccacctt tgaaatcttc 300
aaagaagatg gcaaaactct ggtatctaaa aaagtaaccc tgaaagacaa gtcttctacc 360
gaagaaaaat tcaacgaaaa gggtgaaatc tctgaaaaaa ctatcgtaat ggcaaatggt 420
acccgtctgg aatacaccga catcaaaagc gataaaaccg gcaaagctaa atacgttctg 480
aaagacttta ctctggaagg cactctggct gctgacggca aaaccactct gaaagttacc 540
gaaggcactg ttactctgag catgaacatt tctaaatccg gcgaaatcac cgttgcactg 600
gatgacactg actctagcgg caataaaaaa tccggcacct gggattctga tacttctact 660
ttaaccatta gcaaaaacag ccagaaaact aaacagctgg tattcaccaa agaaaacact 720
atcaccgtac agaactataa ccgtgcaggc aatgcgctgg aaggcagccc ggctgaaatt 780
aaagatctgg cagagctgaa agccgctttg aaataagctg agcggatcc 829
<210> 6
<211> 270
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 6
Met Arg Leu Leu Ile Gly Phe Ala Leu Ala Leu Ala Leu Ile Gly Cys
1 5 10 15
Ala Gln Lys Gly Ala Glu Ser Ile Gly Ser Val Ser Val Asp Leu Pro
20 25 30
Gly Gly Met Lys Val Leu Val Ser Lys Glu Lys Asp Lys Asn Gly Lys
35 40 45
Tyr Ser Leu Met Ala Thr Val Glu Lys Leu Glu Leu Lys Gly Thr Ser
50 55 60
Asp Lys Asn Asn Gly Ser Gly Thr Leu Glu Gly Glu Lys Thr Asn Lys
65 70 75 80
Ser Lys Val Lys Leu Thr Ile Ala Glu Asp Leu Ser Lys Thr Thr Phe
85 90 95
Glu Ile Phe Lys Glu Asp Gly Lys Thr Leu Val Ser Lys Lys Val Thr
100 105 110
Leu Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu Lys Gly Glu
115 120 125
Ile Ser Glu Lys Thr Ile Val Met Ala Asn Gly Thr Arg Leu Glu Tyr
130 135 140
Thr Asp Ile Lys Ser Asp Lys Thr Gly Lys Ala Lys Tyr Val Leu Lys
145 150 155 160
Asp Phe Thr Leu Glu Gly Thr Leu Ala Ala Asp Gly Lys Thr Thr Leu
165 170 175
Lys Val Thr Glu Gly Thr Val Thr Leu Ser Met Asn Ile Ser Lys Ser
180 185 190
Gly Glu Ile Thr Val Ala Leu Asp Asp Thr Asp Ser Ser Gly Asn Lys
195 200 205
Lys Ser Gly Thr Trp Asp Ser Asp Thr Ser Thr Leu Thr Ile Ser Lys
210 215 220
Asn Ser Gln Lys Thr Lys Gln Leu Val Phe Thr Lys Glu Asn Thr Ile
225 230 235 240
Thr Val Gln Asn Tyr Asn Arg Ala Gly Asn Ala Leu Glu Gly Ser Pro
245 250 255
Ala Glu Ile Lys Asp Leu Ala Glu Leu Lys Ala Ala Leu Lys
260 265 270
<210> 7
<211> 784
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 7
catatggcac agaaaggtgc tgagtctatt ggttccgttt ctgtagatct gcccggtgaa 60
atgaaggttc tggtgagcaa agaaaaagac aagaacggca agtacgatct catcgcaacc 120
gtcgacaagc tggagctgaa aggtacttct gataaaaaca acggctctgg tgtgctggag 180
ggcgtcaaaa ctaacaagag caaagtaaag cttacgatct ctgacgatct cggtcagacc 240
acgctggaag ttttcaaaga ggatggcaag accctcgtgt ccaaaaaagt aacttccaaa 300
gacaagtcct ctacggaaga aaaattcaac gaaaaaggtg aggtgtctga aaagatcatc 360
accatggcag acggcacccg tcttgaatac accggtatta aaagcgatgg taccggtaaa 420
gcgaaatatg ttctgaaaaa cttcactctg gaaggcaaag tggctaatga taaaaccacc 480
ttggaagtca aggaaggcac cgttactctg agcatgaata tctccaaatc tggtgaagtt 540
tccgttgaac tgaacgacac tgacagcagc gctgcgacta aaaaaactgc agcgtggaat 600
tccaaaactt ctactttaac cattagcgtt aacagcaaaa aaactaccca gctggtgttc 660
actaaacaag acacgatcac tgtgcagaaa tacgactcca acggcaccaa cttagaaggc 720
acggcagtcg aaattaaaac ccttgatgaa ctgaaaaacg cgctgaaata agctgagcgg 780
atcc 784
<210> 8
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 8
Met Ala Gln Lys Gly Ala Glu Ser Ile Gly Ser Val Ser Val Asp Leu
1 5 10 15
Pro Gly Glu Met Lys Val Leu Val Ser Lys Glu Lys Asp Lys Asn Gly
20 25 30
Lys Tyr Asp Leu Ile Ala Thr Val Asp Lys Leu Glu Leu Lys Gly Thr
35 40 45
Ser Asp Lys Asn Asn Gly Ser Gly Val Leu Glu Gly Val Lys Thr Asn
50 55 60
Lys Ser Lys Val Lys Leu Thr Ile Ser Asp Asp Leu Gly Gln Thr Thr
65 70 75 80
Leu Glu Val Phe Lys Glu Asp Gly Lys Thr Leu Val Ser Lys Lys Val
85 90 95
Thr Ser Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu Lys Gly
100 105 110
Glu Val Ser Glu Lys Ile Ile Thr Met Ala Asp Gly Thr Arg Leu Glu
115 120 125
Tyr Thr Gly Ile Lys Ser Asp Gly Thr Gly Lys Ala Lys Tyr Val Leu
130 135 140
Lys Asn Phe Thr Leu Glu Gly Lys Val Ala Asn Asp Lys Thr Thr Leu
145 150 155 160
Glu Val Lys Glu Gly Thr Val Thr Leu Ser Met Asn Ile Ser Lys Ser
165 170 175
Gly Glu Val Ser Val Glu Leu Asn Asp Thr Asp Ser Ser Ala Ala Thr
180 185 190
Lys Lys Thr Ala Ala Trp Asn Ser Lys Thr Ser Thr Leu Thr Ile Ser
195 200 205
Val Asn Ser Lys Lys Thr Thr Gln Leu Val Phe Thr Lys Gln Asp Thr
210 215 220
Ile Thr Val Gln Lys Tyr Asp Ser Ala Gly Thr Asn Leu Glu Gly Thr
225 230 235 240
Ala Val Glu Ile Lys Thr Leu Asp Glu Leu Lys Asn Ala Leu Lys
245 250 255
<210> 9
<211> 787
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 9
catatggcac agaaaggtgc tgagtctatt ggttccgttt ctgtagatct gcccggtggc 60
atgaccgttc tggtcagcaa agaaaaagac aaaaacggta aatacagcct cgaggcgacc 120
gtcgacaagc ttgagctgaa aggcacctct gataaaaaca acggttccgg caccctggaa 180
ggtgaaaaaa ctaacaaaag caaagtgaaa ctgaccattg ctgatgacct cagccagacc 240
aaattcgaaa ttttcaaaga agatgccaaa accttagtat ccaaaaaagt gaccctgaaa 300
gacaagtcct ctaccgaaga aaaattcaac gaaaagggtg aaacctctga aaaaaccatc 360
gtaatggcaa atggtacccg tctggaatac accgacatca aaagcgatgg ctccggcaaa 420
gccaaatacg ttctgaaaga cttcaccctg gaaggcaccc tcgctgccga cggcaaaacc 480
accttgaaag ttaccgaagg cactgttgtt ttaagcatga acatcttaaa atccggtgaa 540
atcaccgttg cgctggatga ctctgacacc actcaggcca ctaaaaaaac cggcaaatgg 600
gattctaaca cttccactct gaccatcagc gtgaattcca aaaaaactaa aaacatcgtg 660
ttcaccaaag aagacaccat caccgtccag aaatacgact ctgcgggcac caacctcgaa 720
ggcaacgcag tcgaaatcaa aaccctggat gaactgaaaa acgctctgaa ataagctgag 780
cggatcc 787
<210> 10
<211> 256
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 10
Met Ala Gln Lys Gly Ala Glu Ser Ile Gly Ser Val Ser Val Asp Leu
1 5 10 15
Pro Gly Gly Met Thr Val Leu Val Ser Lys Glu Lys Asp Lys Asn Gly
20 25 30
Lys Tyr Ser Leu Glu Ala Thr Val Asp Lys Leu Glu Leu Lys Gly Thr
35 40 45
Ser Asp Lys Asn Asn Gly Ser Gly Thr Leu Glu Gly Glu Lys Thr Asn
50 55 60
Lys Ser Lys Val Lys Leu Thr Ile Ala Asp Asp Leu Ser Gln Thr Lys
65 70 75 80
Phe Glu Ile Phe Lys Glu Asp Ala Lys Thr Leu Val Ser Lys Lys Val
85 90 95
Thr Leu Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu Lys Gly
100 105 110
Glu Thr Ser Glu Lys Thr Ile Val Met Ala Asn Gly Thr Arg Leu Glu
115 120 125
Tyr Thr Asp Ile Lys Ser Asp Gly Ser Gly Lys Ala Lys Tyr Val Leu
130 135 140
Lys Asp Phe Thr Leu Glu Gly Thr Leu Ala Ala Asp Gly Lys Thr Thr
145 150 155 160
Leu Lys Val Thr Glu Gly Thr Val Val Leu Ser Met Asn Ile Leu Lys
165 170 175
Ser Gly Glu Ile Thr Val Ala Leu Asp Asp Ser Asp Thr Thr Gln Ala
180 185 190
Thr Lys Lys Thr Gly Lys Trp Asp Ser Asn Thr Ser Thr Leu Thr Ile
195 200 205
Ser Val Asn Ser Lys Lys Thr Lys Asn Ile Val Phe Thr Lys Glu Asp
210 215 220
Thr Ile Thr Val Gln Lys Tyr Asp Ser Ala Gly Thr Asn Leu Glu Gly
225 230 235 240
Asn Ala Val Glu Ile Lys Thr Leu Asp Glu Leu Lys Asn Ala Leu Lys
245 250 255
<210> 11
<211> 784
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 11
catatggcac agaaaggtgc tgagtctatt ggttccgttt ctgtagatct gcccgggggt 60
atgaaagttc tggtaagcaa agaaaaagac aaaaacggta aatacagcct gatggcaacc 120
gtagaaaagc tggagcttaa aggcacttct gataaaaaca acggttctgg caccctggaa 180
ggtgaaaaaa ctaacaaaag caaagtaaag cttactattg ctgaggatct gagcaaaacc 240
acctttgaaa tcttcaaaga agatggcaaa actctggtat ctaaaaaagt aaccctgaaa 300
gacaagtctt ctaccgaaga aaaattcaac gaaaagggtg aaatctctga aaaaactatc 360
gtaatggcaa atggtacccg tctggaatac accgacatca aaagcgataa aaccggcaaa 420
gctaaatacg ttctgaaaga ctttactctg gaaggcactc tggctgctga cggcaaaacc 480
actctgaaag ttaccgaagg cactgttact ctgagcatga acatttctaa atccggcgaa 540
atcaccgttg cactggatga cactgactct agcggcaata aaaaatccgg cacctgggat 600
tctgatactt ctactttaac cattagcaaa aacagccaga aaactaaaca gctggtattc 660
accaaagaaa acactatcac cgtacagaac tataaccgtg caggcaatgc gctggaaggc 720
agcccggctg aaattaaaga tctggcagag ctgaaagccg ctttgaaata agctgagcgg 780
atcc 784
<210> 12
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 12
Met Ala Gln Lys Gly Ala Glu Ser Ile Gly Ser Val Ser Val Asp Leu
1 5 10 15
Pro Gly Gly Met Lys Val Leu Val Ser Lys Glu Lys Asp Lys Asn Gly
20 25 30
Lys Tyr Ser Leu Met Ala Thr Val Glu Lys Leu Glu Leu Lys Gly Thr
35 40 45
Ser Asp Lys Asn Asn Gly Ser Gly Thr Leu Glu Gly Glu Lys Thr Asn
50 55 60
Lys Ser Lys Val Lys Leu Thr Ile Ala Glu Asp Leu Ser Lys Thr Thr
65 70 75 80
Phe Glu Ile Phe Lys Glu Asp Gly Lys Thr Leu Val Ser Lys Lys Val
85 90 95
Thr Leu Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu Lys Gly
100 105 110
Glu Ile Ser Glu Lys Thr Ile Val Met Ala Asn Gly Thr Arg Leu Glu
115 120 125
Tyr Thr Asp Ile Lys Ser Asp Lys Thr Gly Lys Ala Lys Tyr Val Leu
130 135 140
Lys Asp Phe Thr Leu Glu Gly Thr Leu Ala Ala Asp Gly Lys Thr Thr
145 150 155 160
Leu Lys Val Thr Glu Gly Thr Val Thr Leu Ser Met Asn Ile Ser Lys
165 170 175
Ser Gly Glu Ile Thr Val Ala Leu Asp Asp Thr Asp Ser Ser Gly Asn
180 185 190
Lys Lys Ser Gly Thr Trp Asp Ser Asp Thr Ser Thr Leu Thr Ile Ser
195 200 205
Lys Asn Ser Gln Lys Thr Lys Gln Leu Val Phe Thr Lys Glu Asn Thr
210 215 220
Ile Thr Val Gln Asn Tyr Asn Arg Ala Gly Asn Ala Leu Glu Gly Ser
225 230 235 240
Pro Ala Glu Ile Lys Asp Leu Ala Glu Leu Lys Ala Ala Leu Lys
245 250 255
<210> 13
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 13
Met Arg Leu Leu Ile Gly Phe Ala Leu Ala Leu Ala Leu Ile Gly
1 5 10 15
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 14
tcggggctgg cttaactatg 20
<210> 15
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 15
gcttccggct cgtat 15
<210> 16
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 16
gcttccggct cgtatgttgt 20
<210> 17
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 17
ttatgctagt tattgctcag cg 22
<210> 18
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 18
ttcccctcta gaaataattt tgt 23
<210> 19
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 19
ggaattccat atgcgtctgt tgatcggct 29
<210> 20
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 20
ttggtgcctg cggagtcg 18
<210> 21
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 21
aatacgactc cgcaggcacc 20
<210> 22
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 22
ctgggatccg ctcagcttat ttca 24
<210> 23
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 23
cgtgcgtacc atatggcaca gaaaggtgct gagtct 36
<210> 24
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 24
Tyr Val Leu Glu Gly Thr Leu Thr Ala
1 5
<210> 25
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 25
Lys Glu Lys Asp Lys Asn
1 5
<210> 26
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 26
Lys Thr Asn Lys Ser Lys
1 5
<210> 27
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 27
Lys Glu Lys Asn Lys Asp
1 5
<210> 28
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 28
Lys Glu Lys Asp Lys Asp
1 5
<210> 29
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 29
Lys Ala Asp Lys Ser Lys
1 5
<210> 30
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 30
Lys Thr Asp Lys Ser Lys
1 5
<210> 31
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 31
catatgcgtc tgttgatcgg ctttgctctg gcgctggctc tgatcggctg cgcacagaaa 60
<210> 32
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 32
catatggcac agaaa 15
<210> 33
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 33
Met Arg Leu Leu Ile Gly Phe Ala Leu Ala Leu Ala Leu Ile Gly Cys
1 5 10 15
Ala Gln Lys
<210> 34
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 34
Met Ala Gln Lys
1
<210> 35
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 35
gcaaaaacag ccagaaaact aaacagctgg tattcaccaa agaaaacact 50
<210> 36
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 36
gcaaaaacag ccagaaaact aaacagctgg g 31
<210> 37
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 37
aattcaaaca gctggtattc accaaagaaa acact 35
<210> 38
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 38
gatcactgtg cagaaatacg actccaacgg caccaactta gaaggcacgg cagtc 55
<210> 39
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 39
gatcactgtg cagaaatacg actccgcagg caccaactta gaaggcacgg cagtc 55
<210> 40
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 40
gatcactgtg cagaaatacg actccggcac caacttagaa ggcacggcag tc 52
<210> 41
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 41
cgactccgca ggcaccaa 18
<210> 42
<211> 813
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 42
atgagattat taataggatt tgctttagcg ttagctttaa taggatgtgc acaaaaaggt 60
gctgagtcaa ttggatccgt ttcagtagat ttgcctggtg aaatgaaagt tcttgtaagc 120
aaagaaaaag acaaaaacgg caagtacgat ctaattgcaa cagtagacaa gcttgagctt 180
aaaggaactt ctgataaaaa caatggatct ggagtacttg aaggcgtaaa aactaacaaa 240
agtaaagtaa aattaacaat ttctgacgat ctaggtcaaa ccacacttga agttttcaaa 300
gaagatggca aaacactagt atcaaaaaaa gtaacttcca aagacaagtc atcaacagaa 360
gaaaaattca atgaaaaagg tgaagtatct gaaaaaataa taacaatggc agacggaacc 420
agacttgaat acacaggaat taaaagcgat ggaactggaa aagctaaata tgttttaaaa 480
aattttactc ttgaaggaaa agtagctaat gataaaacaa cattggaagt aaaagaagga 540
accgttactt taagtatgaa tatttcaaaa tctggggaag tttcagttga acttaatgac 600
actgacagta gtgctgctac taaaaaaact gcagcttgga attcaaaaac ttctacttta 660
acaattagtg ttaacagcaa aaaaactaca caacttgtgt ttactaaaca agacacaata 720
actgtacaaa aatacgactc caacggtacc aatttagaag gcacagcagt cgaaattaaa 780
acacttgatg aacttaaaaa cgctttaaaa taa 813
<210> 43
<211> 826
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 43
atgcgtctgt tgatcggctt tgctctggcg ctggctctga tcggctgcgc acagaaaggt 60
gctgagtcta ttggttccgt ttctgtagat ctgcccggtg aaatgaaggt tctggtgagc 120
aaagaaaaag acaagaacgg caagtacgat ctcatcgcaa ccgtcgacaa gctggagctg 180
aaaggtactt ctgataaaaa caacggctct ggtgtgctgg agggcgtcaa aactaacaag 240
agcaaagtaa agcttacgat ctctgacgat ctcggtcaga ccacgctgga agttttcaaa 300
gaggatggca agaccctcgt gtccaaaaaa gtaacttcca aagacaagtc ctctacggaa 360
gaaaaattca acgaaaaagg tgaggtgtct gaaaagatca tcaccatggc agacggcacc 420
cgtcttgaat acaccggtat taaaagcgat ggtaccggta aagcgaaata tgttctgaaa 480
aacttcactc tggaaggcaa agtggctaat gataaaacca ccttggaagt caaggaaggc 540
accgttactc tgagcatgaa tatctccaaa tctggtgaag tttccgttga actgaacgac 600
actgacagca gcgctgcgac taaaaaaact gcagcgtgga attccaaaac ttctacttta 660
accattagcg ttaacagcaa aaaaactacc cagctggtgt tcactaaaca agacacgatc 720
actgtgcaga aatacgactc cgcaggcacc aacttagaag gcacggcagt cgaaattaaa 780
acccttgatg aactgaaaaa cgcgctgaaa taagctgagc ggatcc 826
<210> 44
<211> 818
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 44
atatgagatt attaatagga tttgctttag cgttagcttt aataggatgt gcacaaaaag 60
gtgctgagtc aattggatcc gtttcagtag atttacctgg tggaatgaca gttcttgtaa 120
gtaaagaaaa agacaaagac ggtaaataca gtctagaggc aacagtagac aagcttgagc 180
ttaaaggaac ttctgataaa aacaacggtt ctggaacact tgaaggtgaa aaaactgaca 240
aaagtaaagt aaaattaaca attgctgatg acctaagtca aactaaattt gaaattttca 300
aagaagatgc caaaacatta gtatcaaaaa aagtaaccct taaagacaag tcatcaacag 360
aagaaaaatt caacgaaaag ggtgaaacat ctgaaaaaac aatagtaaga gcaaatggaa 420
ccagacttga atacacagac ataaaaagcg atggatccgg aaaagctaaa gaagttttaa 480
aagactttac tcttgaagga actctagctg ctgacggcaa aacaacattg aaagttacag 540
aaggcactgt tgttttaagc aagaacattt taaaatccgg agaaataaca gttgcacttg 600
atgactctga cactactcag gctactaaaa aaactggaaa atgggattca aatacttcca 660
ctttaacaat tagtgtgaat agcaaaaaaa ctaaaaacat tgtatttaca aaagaagaca 720
caataacagt acaaaaatac gactcagcag gcaccaatct agaaggcaac gcagtcgaaa 780
ttaaaacact tgatgaactt aaaaacgctt taaaataa 818
<210> 45
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 45
atatgcgtct gttgatcggc tttgctctgg cgctggctct gatcggctgc gcacagaaag 60
gtgctgagtc tattggttcc gtttctgtag atctgcccgg tggcatgacc gttctggtca 120
gcaaagaaaa agacaaaaac ggtaaataca gcctcgaggc gaccgtcgac aagcttgagc 180
tgaaaggcac ctctgataaa aacaacggtt ccggcaccct ggaaggtgaa aaaactaaca 240
aaagcaaagt gaaactgacc attgctgatg acctcagcca gaccaaattc gaaattttca 300
aagaagatgc caaaacctta gtatccaaaa aagtgaccct gaaagacaag tcctctaccg 360
aagaaaaatt caacgaaaag ggtgaaacct ctgaaaaaac catcgtaatg gcaaatggta 420
cccgtctgga atacaccgac atcaaaagcg atggctccgg caaagccaaa tacgttctga 480
aagacttcac cctggaaggc accctcgctg ccgacggcaa aaccaccttg aaagttaccg 540
aaggcactgt tgttttaagc atgaacatct taaaatccgg tgaaatcacc gttgcgctgg 600
atgactctga caccactcag gccactaaaa aaaccggcaa atgggattct aacacttcca 660
ctctgaccat cagcgtgaat tccaaaaaaa ctaaaaacat cgtgttcacc aaagaagaca 720
ccatcaccgt ccagaaatac gactctgcgg gcaccaacct cgaaggcaac gcagtcgaaa 780
tcaaaaccct ggatgaactg aaaaacgctc tgaaataagc tgagcggatc c 831
<210> 46
<211> 816
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 46
catatgagat tattaatagg atttgcttta gcgttagctt taataggatg tgcacaaaaa 60
ggtgctgagt caattggatc cgtttcagta gatttacctg gtggaatgaa agttcttgta 120
agtaaagaaa aagacaaaga tggtaaatac agtctaatgg caacagtaga aaagcttgag 180
cttaaaggaa cttctgataa aaacaacggt tctggaacac ttgaaggtga aaaaactgac 240
aaaagtaaag taaaattaac aattgctgag gatctaagta aaaccacatt tgaaatcttc 300
aaagaagatg gcaaaacatt agtatcaaaa aaagtaaccc ttaaagacaa gtcatcaaca 360
gaagaaaaat tcaacgaaaa gggtgaaata tctgaaaaaa caatagtaag agcaaatgga 420
accagacttg aatacacaga cataaaaagc gataaaaccg gaaaagctaa agaagtttta 480
aaagacttta ctcttgaagg aactctagct gctgacggca aaacaacatt gaaagttaca 540
gaaggcactg ttactttaag caagaacatt tcaaaatccg gagaaataac agttgcactt 600
gatgacactg actctagcgg caataaaaaa tccggaacat gggattcaga tacttctact 660
ttaacaatta gtaaaaacag tcaaaaaact aaacaacttg tattcacaaa agaaaacaca 720
ataacagtac aaaactataa cagagcaggc aatgcgcttg aaggcagccc agctgaaatt 780
aaagatcttg cagagcttaa agccgcttta aaataa 816
<210> 47
<211> 829
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 47
catatgcgtc tgttgatcgg ctttgctttg gcgctggctt taatcggctg tgcacagaaa 60
ggtgctgagt ctattggttc cgtttctgta gatctgcccg ggggtatgaa agttctggta 120
agcaaagaaa aagacaaaaa cggtaaatac agcctgatgg caaccgtaga aaagctggag 180
cttaaaggca cttctgataa aaacaacggt tctggcaccc tggaaggtga aaaaactaac 240
aaaagcaaag taaagcttac tattgctgag gatctgagca aaaccacctt tgaaatcttc 300
aaagaagatg gcaaaactct ggtatctaaa aaagtaaccc tgaaagacaa gtcttctacc 360
gaagaaaaat tcaacgaaaa gggtgaaatc tctgaaaaaa ctatcgtaat ggcaaatggt 420
acccgtctgg aatacaccga catcaaaagc gataaaaccg gcaaagctaa atacgttctg 480
aaagacttta ctctggaagg cactctggct gctgacggca aaaccactct gaaagttacc 540
gaaggcactg ttactctgag catgaacatt tctaaatccg gcgaaatcac cgttgcactg 600
gatgacactg actctagcgg caataaaaaa tccggcacct gggattctga tacttctact 660
ttaaccatta gcaaaaacag ccagaaaact aaacagctgg tattcaccaa agaaaacact 720
atcaccgtac agaactataa ccgtgcaggc aatgcgctgg aaggcagccc ggctgaaatt 780
aaagatctgg cagagctgaa agccgctttg aaataagctg agcggatcc 829
<210> 48
<211> 829
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 48
ggatccgctc agcttatttc agcgcgtttt tcagttcatc aagggtttta atttcgactg 60
ccgtgccttc taagttggtg cctgcggagt cgtatttctg cacagtgatc gtgtcttgtt 120
tagtgaacac cagctgggta gtttttttgc tgttaacgct aatggttaaa gtagaagttt 180
tggaattcca cgctgcagtt tttttagtcg cagcgctgct gtcagtgtcg ttcagttcaa 240
cggaaacttc accagatttg gagatattca tgctcagagt aacggtgcct tccttgactt 300
ccaaggtggt tttatcatta gccactttgc cttccagagt gaagtttttc agaacatatt 360
tcgctttacc ggtaccatcg cttttaatac cggtgtattc aagacgggtg ccgtctgcca 420
tggtgatgat cttttcagac acctcacctt tttcgttgaa tttttcttcc gtagaggact 480
tgtctttgga agttactttt ttggacacga gggtcttgcc atcctctttg aaaacttcca 540
gcgtggtctg accgagatcg tcagagatcg taagctttac tttgctcttg ttagttttga 600
cgccctccag cacaccagag ccgttgtttt tatcagaagt acctttcagc tccagcttgt 660
cgacggttgc gatgagatcg tacttgccgt tcttgtcttt ttctttgctc accagaacct 720
tcatttcacc gggcagatct acagaaacgg aaccaataga ctcagcacct ttctgtgcgc 780
agccgatcag agccagcgcc agagcaaagc cgatcaacag acgcatatg 829
<210> 49
<211> 832
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 49
ggatccgctc agcttatttc agagcgtttt tcagttcatc cagggttttg atttcgactg 60
cgttgccttc gaggttggtg cccgcagagt cgtatttctg gacggtgatg gtgtcttctt 120
tggtgaacac gatgttttta gtttttttgg aattcacgct gatggtcaga gtggaagtgt 180
tagaatccca tttgccggtt tttttagtgg cctgagtggt gtcagagtca tccagcgcaa 240
cggtgatttc accggatttt aagatgttca tgcttaaaac aacagtgcct tcggtaactt 300
tcaaggtggt tttgccgtcg gcagcgaggg tgccttccag ggtgaagtct ttcagaacgt 360
atttggcttt gccggagcca tcgcttttga tgtcggtgta ttccagacgg gtaccatttg 420
ccattacgat ggttttttca gaggtttcac ccttttcgtt gaatttttct tcggtagagg 480
acttgtcttt cagggtcact tttttggata ctaaggtttt ggcatcttct ttgaaaattt 540
cgaatttggt ctggctgagg tcatcagcaa tggtcagttt cactttgctt ttgttagttt 600
tttcaccttc cagggtgccg gaaccgttgt ttttatcaga ggtgcctttc agctcaagct 660
tgtcgacggt cgcctcgagg ctgtatttac cgtttttgtc tttttctttg ctgaccagaa 720
cggtcatgcc accgggcaga tctacagaaa cggaaccaat agactcagca cctttctgtg 780
cgcagccgat cagagccagc gccagagcaa agccgatcaa cagacgcata tg 832
<210> 50
<211> 829
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 50
ggatccgctc agcttatttc aaagcggctt tcagctctgc cagatcttta atttcagccg 60
ggctgccttc cagcgcattg cctgcacggt tatagttctg tacggtgata gtgttttctt 120
tggtgaatac cagctgttta gttttctggc tgtttttgct aatggttaaa gtagaagtat 180
cagaatccca ggtgccggat tttttattgc cgctagagtc agtgtcatcc agtgcaacgg 240
tgatttcgcc ggatttagaa atgttcatgc tcagagtaac agtgccttcg gtaactttca 300
gagtggtttt gccgtcagca gccagagtgc cttccagagt aaagtctttc agaacgtatt 360
tagctttgcc ggttttatcg cttttgatgt cggtgtattc cagacgggta ccatttgcca 420
ttacgatagt tttttcagag atttcaccct tttcgttgaa tttttcttcg gtagaagact 480
tgtctttcag ggttactttt ttagatacca gagttttgcc atcttctttg aagatttcaa 540
aggtggtttt gctcagatcc tcagcaatag taagctttac tttgcttttg ttagtttttt 600
caccttccag ggtgccagaa ccgttgtttt tatcagaagt gcctttaagc tccagctttt 660
ctacggttgc catcaggctg tatttaccgt ttttgtcttt ttctttgctt accagaactt 720
tcataccccc gggcagatct acagaaacgg aaccaataga ctcagcacct ttctgtgcac 780
agccgattaa agccagcgcc aaagcaaagc cgatcaacag acgcatatg 829
<210> 51
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 51
tttctgtgcg cagccgatca gagccagcgc cagagcaaag ccgatcaaca gacgcatatg 60
<210> 52
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 52
tttctgtgcc atatg 15
<210> 53
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 53
agtgttttct ttggtgaata ccagctgttt agttttctgg ctgtttttgc 50
<210> 54
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 54
aattcccagc tgtttagttt tctggctgtt tttgc 35
<210> 55
<211> 784
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 55
gtataccgtg tctttccacg actcagataa ccaaggcaaa gacatctaga cgggccactt 60
tacttccaag accactcgtt tctttttctg ttcttgccgt tcatgctaga gtagcgttgg 120
cagctgttcg acctcgactt tccatgaaga ctatttttgt tgccgagacc acacgacctc 180
ccgcagtttt gattgttctc gtttcatttc gaatgctaga gactgctaga gccagtctgg 240
tgcgaccttc aaaagtttct cctaccgttc tgggagcaca ggttttttca ttgaaggttt 300
ctgttcagga gatgccttct ttttaagttg ctttttccac tccacagact tttctagtag 360
tggtaccgtc tgccgtgggc agaacttatg tggccataat tttcgctacc atggccattt 420
cgctttatac aagacttttt gaagtgagac cttccgtttc accgattact attttggtgg 480
aaccttcagt tccttccgtg gcaatgagac tcgtacttat agaggtttag accacttcaa 540
aggcaacttg acttgctgtg actgtcgtcg cgacgctgat ttttttgacg tcgcacctta 600
aggttttgaa gatgaaattg gtaatcgcaa ttgtcgtttt tttgatgggt cgaccacaag 660
tgatttgttc tgtgctagtg acacgtcttt atgctgaggt tgccgtggtt gaatcttccg 720
tgccgtcagc tttaattttg ggaactactt gactttttgc gcgactttat tcgactcgcc 780
tagg 784
<210> 56
<211> 787
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 56
gtataccgtg tctttccacg actcagataa ccaaggcaaa gacatctaga cgggccaccg 60
tactggcaag accagtcgtt tctttttctg tttttgccat ttatgtcgga gctccgctgg 120
cagctgttcg aactcgactt tccgtggaga ctatttttgt tgccaaggcc gtgggacctt 180
ccactttttt gattgttttc gtttcacttt gactggtaac gactactgga gtcggtctgg 240
tttaagcttt aaaagtttct tctacggttt tggaatcata ggttttttca ctgggacttt 300
ctgttcagga gatggcttct ttttaagttg cttttcccac tttggagact tttttggtag 360
cattaccgtt taccatgggc agaccttatg tggctgtagt tttcgctacc gaggccgttt 420
cggtttatgc aagactttct gaagtgggac cttccgtggg agcgacggct gccgttttgg 480
tggaactttc aatggcttcc gtgacaacaa aattcgtact tgtagaattt taggccactt 540
tagtggcaac gcgacctact gagactgtgg tgagtccggt gatttttttg gccgtttacc 600
ctaagattgt gaaggtgaga ctggtagtcg cacttaaggt ttttttgatt tttgtagcac 660
aagtggtttc ttctgtggta gtggcaggtc tttatgctga gacgcccgtg gttggagctt 720
ccgttgcgtc agctttagtt ttgggaccta cttgactttt tgcgagactt tattcgactc 780
gcctagg 787
<210> 57
<211> 784
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 57
gtataccgtg tctttccacg actcagataa ccaaggcaaa gacatctaga cgggccccca 60
tactttcaag accattcgtt tctttttctg tttttgccat ttatgtcgga ctaccgttgg 120
catcttttcg acctcgaatt tccgtgaaga ctatttttgt tgccaagacc gtgggacctt 180
ccactttttt gattgttttc gtttcatttc gaatgataac gactcctaga ctcgttttgg 240
tggaaacttt agaagtttct tctaccgttt tgagaccata gattttttca ttgggacttt 300
ctgttcagaa gatggcttct ttttaagttg cttttcccac tttagagact tttttgatag 360
cattaccgtt taccatgggc agaccttatg tggctgtagt tttcgctatt ttggccgttt 420
cgatttatgc aagactttct gaaatgagac cttccgtgag accgacgact gccgttttgg 480
tgagactttc aatggcttcc gtgacaatga gactcgtact tgtaaagatt taggccgctt 540
tagtggcaac gtgacctact gtgactgaga tcgccgttat tttttaggcc gtggacccta 600
agactatgaa gatgaaattg gtaatcgttt ttgtcggtct tttgatttgt cgaccataag 660
tggtttcttt tgtgatagtg gcatgtcttg atattggcac gtccgttacg cgaccttccg 720
tcgggccgac tttaatttct agaccgtctc gactttcggc gaaactttat tcgactcgcc 780
tagg 784
<210> 58
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 58
agtgttttct ttggtgaata ccagctgttt g 31
<210> 59
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 59
tatgcgtctg ttgatcggct ttgctctggc gctggctctg atcgg 45
<210> 60
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 60
ctgcgcacag aaaggtgctg agtctattgg ttccgtttct gtagatctgc 50
<210> 61
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 61
ccggtgaaat gaaggttctg gtgagcaaag aaaaagacaa gaacggcaag 50
<210> 62
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 62
tacgatctca tcgcaaccgt cgacaagctg gagctgaaag gtacttctga 50
<210> 63
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 63
taaaaacaac ggctctggtg tgctggaggg cgtcaaaact aacaagagca aagtaa 56
<210> 64
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 64
agctttactt tgctcttgtt agttttgacg ccctccagca 40
<210> 65
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 65
caccagagcc gttgttttta tcagaagtac ctttcagctc cagcttgtcg 50
<210> 66
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 66
acggttgcga tgagatcgta cttgccgttc ttgtcttttt ctttgctcac 50
<210> 67
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 67
cagaaccttc atttcaccgg gcagatctac agaaacggaa ccaatagact 50
<210> 68
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 68
cagcaccttt ctgtgcgcag ccgatcagag ccagcgccag agcaaagccg atcaacagac 60
gca 63
<210> 69
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 69
agcttacgat ctctgacgat ctcggtcaga ccac 34
<210> 70
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 70
gctggaagtt ttcaaagagg atggcaagac cctcgtgtcc aaaaaagtaa 50
<210> 71
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 71
cttccaaaga caagtcctct acggaagaaa aattcaacga aaaaggtgag 50
<210> 72
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 72
gtgtctgaaa agatcatcac catggcagac ggcacccgtc 40
<210> 73
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 73
ttgaatacac cggtattaaa agcgatggta c 31
<210> 74
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 74
catcgctttt aataccggtg tattcaagac gggtgccgtc tgccatg 47
<210> 75
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 75
gtgatgatct tttcagacac ctcacctttt tcgttgaatt tttcttccgt 50
<210> 76
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 76
agaggacttg tctttggaag ttactttttt ggacacgagg gtcttgccat 50
<210> 77
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 77
cctctttgaa aacttccagc gtggtctgac cgagatcgtc agagatcgta 50
<210> 78
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 78
cggtaaagcg aaatatgttc tgaaaaactt cactctgga 39
<210> 79
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 79
aggcaaagtg gctaatgata aaaccacctt ggaagtcaag gaaggcaccg 50
<210> 80
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 80
ttactctgag catgaatatc tccaaatctg gtgaagtttc cgttgaactg 50
<210> 81
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 81
aacgacactg acagcagcgc tgcgactaaa aaaactgcag cgtgg 45
<210> 82
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 82
aattccacgc tgcagttttt ttagtcgca 29
<210> 83
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 83
gcgctgctgt cagtgtcgtt cagttcaacg gaaacttcac cagatttgga 50
<210> 84
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 84
gatattcatg ctcagagtaa cggtgccttc cttgacttcc aaggtggttt 50
<210> 85
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 85
tatcattagc cactttgcct tccagagtga agtttttcag aacatatttc gctttaccgg 60
tac 63
<210> 86
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 86
aattccaaaa cttctacttt aaccattagc gttaacagca aaaaa 45
<210> 87
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 87
actacccagc tggtgttcac taaacaagac acgatcactg tgcagaaata 50
<210> 88
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 88
cgactccaac ggcaccaact tagaaggcac ggcagtcgaa attaaaaccc 50
<210> 89
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 89
ttgatgaact gaaaaacgcg ctgaaataag ctgagcg 37
<210> 90
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 90
gatccgctca gcttatttca gcgcgttttt cagttcatca agggttttaa tttcgactgc 60
c 61
<210> 91
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 91
gtgccttcta agttggtgcc gttggagtcg tatttctgca cagtgatcgt 50
<210> 92
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 92
gtcttgttta gtgaacacca gctgggtagt ttttttgctg ttaacgctaa 50
<210> 93
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 93
tggttaaagt agaagttttg g 21
<210> 94
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 94
tatgcgtctg ttgatcggct ttgctttggc gctggcttta atcggctg 48
<210> 95
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 95
tgcacagaaa ggtgctgagt ctattggttc cgtttctgta gatctgcccg 50
<210> 96
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 96
ggggtatgaa agttctggta agcaaagaaa aagacaaaaa cggtaaatac 50
<210> 97
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 97
agcctgatgg caaccgtaga aaagctggag cttaaaggca cttctgataa 50
<210> 98
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 98
aaacaacggt tctggcaccc tggaaggtga aaaaactaac aaaagcaaag taa 53
<210> 99
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 99
agctttactt tgcttttgtt agttttttca ccttcca 37
<210> 100
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 100
gggtgccaga accgttgttt ttatcagaag tgcctttaag ctccagcttt 50
<210> 101
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 101
tctacggttg ccatcaggct gtatttaccg tttttgtctt tttctttgct 50
<210> 102
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 102
taccagaact ttcatacccc cgggcagatc tacagaaacg gaaccaatag 50
<210> 103
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 103
actcagcacc tttctgtgca cagccgatta 30
<210> 104
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 104
aagccagcgc caaagcaaag ccgatcaaca gacgca 36
<210> 105
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 105
agcttactat tgctgaggat ctgagcaaaa ccacctttga aatcttc 47
<210> 106
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 106
aaagaagatg gcaaaactct ggtatctaaa aaagtaaccc tgaaagacaa 50
<210> 107
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 107
gtcttctacc gaagaaaaat tcaacgaaaa gggtgaaatc 40
<210> 108
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 108
tctgaaaaaa ctatcgtaat ggcaaatggt ac 32
<210> 109
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 109
aaggtggttt tgctcagatc ctcagcaata gta 33
<210> 110
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 110
agagttttgc catcttcttt gaagatttca 30
<210> 111
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 111
atttttcttc ggtagaagac ttgtctttca gggttacttt tttagatacc 50
<210> 112
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 112
catttgccat tacgatagtt ttttcagaga tttcaccctt ttcgttga 48
<210> 113
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 113
ccgtctggaa tacaccgaca tcaaaagcga taaaaccggc aaagctaa 48
<210> 114
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 114
atacgttctg aaagacttta ctctggaagg cactctggct gctgacggca 50
<210> 115
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 115
aaaccactct gaaagttacc gaaggcactg ttactctgag catgaacatt 50
<210> 116
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 116
tctaaatccg gcgaaatcac cgttgcactg gatgacactg actctagcgg 50
<210> 117
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 117
caataaaaaa tccggcacct gggattctga tacttctact ttaaccatta 50
<210> 118
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 118
gcaaaaacag ccagaaaact aaacagctgg g 31
<210> 119
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 119
gcttttgatg tcggtgtatt ccagacgggt ac 32
<210> 120
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 120
ccttccagag taaagtcttt cagaacgtat ttagctttgc cggttttatc 50
<210> 121
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 121
cagtgccttc ggtaactttc agagtggttt tgccgtcagc agccagagtg 50
<210> 122
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 122
cagtgcaacg gtgatttcgc cggatttaga aatgttcatg ctcagagtaa 50
<210> 123
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 123
tcagaatccc aggtgccgga ttttttattg ccgctagagt cagtgtcatc 50
<210> 124
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 124
aattcccagc tgtttagttt tctggctgtt tttgctaatg gttaaagtag aagta 55
<210> 125
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 125
aattcaaaca gctggtattc accaaagaaa acactatcac cgtac 45
<210> 126
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 126
agaactataa ccgtgcaggc aatgcgctgg aaggcagccc 40
<210> 127
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 127
ggctgaaatt aaagatctgg cagagctgaa agccgctttg aaataagctg agcg 54
<210> 128
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 128
gatccgctca gcttatttca aagcggct 28
<210> 129
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 129
ttcagctctg ccagatcttt aatttcagcc gggctgcctt ccagcgcatt 50
<210> 130
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 130
gcctgcacgg ttatagttct gtacggtgat agtgttttct ttggtgaata ccagctgttt 60
g 61
<210> 131
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 131
tatgcgtctg ttgatcggct ttgctctggc gctggctctg atcggctg 48
<210> 132
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 132
cgcacagaaa ggtgctgagt ctattggttc cgtttctgta gatctgcccg 50
<210> 133
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 133
gtggcatgac cgttctggtc agcaaagaaa aagacaaaaa cg 42
<210> 134
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 134
gtaaatacag cctcgaggcg accgtcgaca 30
<210> 135
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 135
agcttgtcga cggtcgcctc gaggctgtat ttaccgtttt tgtctttttc tttgct 56
<210> 136
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 136
gaccagaacg gtcatgccac cgggcagatc tacagaaacg 40
<210> 137
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 137
gaaccaatag actcagcacc tttctgtgcg cagccgatca gagccagcgc 50
<210> 138
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 138
cagagcaaag ccgatcaaca gacgca 26
<210> 139
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 139
agcttgagct gaaaggcacc tctgataaaa acaacggttc cggcaccctg 50
<210> 140
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 140
gaaggtgaaa aaactaacaa aagcaaagtg aaactgacca ttgctgat 48
<210> 141
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 141
gacctcagcc agaccaaatt cgaaattttc aaagaagatg ccaaaacctt 50
<210> 142
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 142
agtatccaaa aaagtgaccc tgaaagacaa gtcctctacc gaagaaaaat 50
<210> 143
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 143
tcaacgaaaa gggtgaaacc tctgaaaaaa ccatcgtaat ggcaaatggt ac 52
<210> 144
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 144
catttgccat tacgatggtt ttttcaga 28
<210> 145
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 145
ggtttcaccc ttttcgttga atttttcttc ggtagaggac 40
<210> 146
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 146
ttgtctttca gggtcacttt tttggatact aaggttttgg catcttcttt 50
<210> 147
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 147
gaaaatttcg aatttggtct ggctgaggtc atcagcaatg gtcagtttca 50
<210> 148
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 148
ctttgctttt gttagttttt tcaccttcca gggtgccgga 40
<210> 149
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 149
accgttgttt ttatcagagg tgcctttcag ctca 34
<210> 150
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 150
ccgtctggaa tacaccgaca tcaaaagcga tggctccggc aaagccaa 48
<210> 151
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 151
atacgttctg aaagacttca ccctggaagg caccctcgct gccgacgg 48
<210> 152
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 152
caaaaccacc ttgaaagtta ccgaaggcac tgttgtttta ag 42
<210> 153
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 153
catgaacatc ttaaaatccg gtgaaatcac cgttgcgctg 40
<210> 154
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 154
gatgactctg acaccactca ggccactaaa aaaaccggca aatgggattc 50
<210> 155
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 155
taacacttcc actctgacca tcagcgtg 28
<210> 156
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 156
aattcacgct gatggtcaga gtggaagtgt tagaatccca tttgccg 47
<210> 157
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 157
gtttttttag tggcctgagt ggtgtcagag tcatccagcg caacggtgat ttcac 55
<210> 158
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 158
cggattttaa gatgttcatg cttaaaacaa cagtgccttc ggtaactttc 50
<210> 159
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 159
aaggtggttt tgccgtcggc agcgagggtg ccttccaggg 40
<210> 160
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 160
tgaagtcttt cagaacgtat ttggctttgc cggagccatc 40
<210> 161
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 161
gcttttgatg tcggtgtatt ccagacgggt ac 32
<210> 162
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 162
aattccaaaa aaactaaaaa catcgtgttc accaaagaag acaccatcac cg 52
<210> 163
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 163
tccagaaata cgactctgcg ggcaccaacc tcgaaggcaa cgcagtcgaa 50
<210> 164
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 164
atcaaaaccc tggatgaact gaaaaacgct ctgaaataag ctgagcg 47
<210> 165
<211> 71
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 165
gatccgctca gcttatttca gagcgttttt cagttcatcc agggttttga tttcgactgc 60
gttgccttcg a 71
<210> 166
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 166
ggttggtgcc cgcagagtcg tatttctgga cggtgatggt gtcttctttg 50
<210> 167
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 167
gtgaacacga tgtttttagt ttttttgg 28
<210> 168
<211> 822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 168
atgaaaaaat atttattggg aataggtcta atattagcct taatagcatg taagcaaaat 60
gttagcagcc ttgacgagaa aaacagcgtt tcagtagatt tgcctggtga aatgaaagtt 120
cttgtaagca aagaaaaaaa caaagacggc aagtacgatc taattgcaac agtagacaag 180
cttgagctta aaggaacttc tgataaaaac aatggatctg gagtacttga aggcgtaaaa 240
gctgacaaaa gtaaagtaaa attaacaatt tctgacgatc taggtcaaac cacacttgaa 300
gttttcaaag aagatggcaa aacactagta tcaaaaaaag taacttccaa agacaagtca 360
tcaacagaag aaaaattcaa tgaaaaaggt gaagtatctg aaaaaataat aacaagagca 420
gacggaacca gacttgaata cacaggaatt aaaagcgatg gatctggaaa agctaaagag 480
gttttaaaaa actttactct tgaaggaaaa gtagctaatg ataaagtaac attggaagta 540
aaagaaggaa ccgttacttt aagtaaaaat atttcaaaat ctggggaagt ttcagttgaa 600
cttaatgaca ctgacagtag tgctgctact aaaaaaactg cagcttggaa ttcaaaaact 660
tctactttaa caattagtgt taacagcaaa aaaactacac aacttgtgtt tactaaacaa 720
gacacaataa ctgtacaaaa atacgactcc gcaggtacca atttagaagg cacagcagtc 780
gaaattaaaa cacttgatga acttaaaaac gctttaaaat ag 822
<210> 169
<211> 273
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 169
Met Lys Lys Tyr Leu Leu Gly Ile Gly Leu Ile Leu Ala Leu Ile Ala
1 5 10 15
Cys Lys Gln Asn Val Ser Ser Leu Asp Glu Lys Asn Ser Val Ser Val
20 25 30
Asp Leu Pro Gly Glu Met Lys Val Leu Val Ser Lys Glu Lys Asn Lys
35 40 45
Asp Gly Lys Tyr Asp Leu Ile Ala Thr Val Asp Lys Leu Glu Leu Lys
50 55 60
Gly Thr Ser Asp Lys Asn Asn Gly Ser Gly Val Leu Glu Gly Val Lys
65 70 75 80
Ala Asp Lys Ser Lys Val Lys Leu Thr Ile Ser Asp Asp Leu Gly Gln
85 90 95
Thr Thr Leu Glu Val Phe Lys Glu Asp Gly Lys Thr Leu Val Ser Lys
100 105 110
Lys Val Thr Ser Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu
115 120 125
Lys Gly Glu Val Ser Glu Lys Ile Ile Thr Arg Ala Asp Gly Thr Arg
130 135 140
Leu Glu Tyr Thr Gly Ile Lys Ser Asp Gly Ser Gly Lys Ala Lys Glu
145 150 155 160
Val Leu Lys Asn Phe Thr Leu Glu Gly Lys Val Ala Asn Asp Lys Val
165 170 175
Thr Leu Glu Val Lys Glu Gly Thr Val Thr Leu Ser Lys Asn Ile Ser
180 185 190
Lys Ser Gly Glu Val Ser Val Glu Leu Asn Asp Thr Asp Ser Ser Ala
195 200 205
Ala Thr Lys Lys Thr Ala Ala Trp Asn Ser Lys Thr Ser Thr Leu Thr
210 215 220
Ile Ser Val Asn Ser Lys Lys Thr Thr Gln Leu Val Phe Thr Lys Gln
225 230 235 240
Asp Thr Ile Thr Val Gln Lys Tyr Asp Ser Ala Gly Thr Asn Leu Glu
245 250 255
Gly Thr Ala Val Glu Ile Lys Thr Leu Asp Glu Leu Lys Asn Ala Leu
260 265 270
Lys
<210> 170
<211> 825
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 170
atgaaaaaat atttattggg aataggtcta atattagcct taatagcatg taagcaaaat 60
gttagcacgc ttgatgaaaa aaatagcgtt tcagtagatt tacctggtgg aatgacagtt 120
cttgtaagta aagaaaaaga caaagacggt aaatacagtc tagaggcaac agtagacaag 180
cttgagctta aaggaacttc tgataaaaac aacggttctg gaacacttga aggtgaaaaa 240
actgacaaaa gtaaagtaaa attaacaatt gctgatgacc taagtcaaac taaatttgaa 300
attttcaaag aagatgccaa aacattagta tcaaaaaaag taacccttaa agacaagtca 360
tcaacagaag aaaaattcaa cgaaaagggt gaaacatctg aaaaaacaat agtaagagca 420
aatggaacca gacttgaata cacagacata aaaagcgatg gatccggaaa agctaaagaa 480
gttttaaaag actttactct tgaaggaact ctagctgctg acggcaaaac aacattgaaa 540
gttacagaag gcactgttgt tttaagcaag aacattttaa aatccggaga aataacagtt 600
gcacttgatg actctgacac tactcaggct actaaaaaaa ctggaaaatg ggattcaaat 660
acttccactt taacaattag tgtgaatagc aaaaaaacta aaaacattgt atttacaaaa 720
gaagacacaa taacagtaca aaaatacgac tcagcaggca ccaatctaga aggcaacgca 780
gtcgaaatta aaacacttga tgaacttaaa aacgctttaa aataa 825
<210> 171
<211> 274
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 171
Met Lys Lys Tyr Leu Leu Gly Ile Gly Leu Ile Leu Ala Leu Ile Ala
1 5 10 15
Cys Lys Gln Asn Val Ser Thr Leu Asp Glu Lys Asn Ser Val Ser Val
20 25 30
Asp Leu Pro Gly Gly Met Thr Val Leu Val Ser Lys Glu Lys Asp Lys
35 40 45
Asp Gly Lys Tyr Ser Leu Glu Ala Thr Val Asp Lys Leu Glu Leu Lys
50 55 60
Gly Thr Ser Asp Lys Asn Asn Gly Ser Gly Thr Leu Glu Gly Glu Lys
65 70 75 80
Thr Asp Lys Ser Lys Val Lys Leu Thr Ile Ala Asp Asp Leu Ser Gln
85 90 95
Thr Lys Phe Glu Ile Phe Lys Glu Asp Ala Lys Thr Leu Val Ser Lys
100 105 110
Lys Val Thr Leu Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu
115 120 125
Lys Gly Glu Thr Ser Glu Lys Thr Ile Val Arg Ala Asn Gly Thr Arg
130 135 140
Leu Glu Tyr Thr Asp Ile Lys Ser Asp Gly Ser Gly Lys Ala Lys Glu
145 150 155 160
Val Leu Lys Asp Phe Thr Leu Glu Gly Thr Leu Ala Ala Asp Gly Lys
165 170 175
Thr Thr Leu Lys Val Thr Glu Gly Thr Val Val Leu Ser Lys Asn Ile
180 185 190
Leu Lys Ser Gly Glu Ile Thr Val Ala Leu Asp Asp Ser Asp Thr Thr
195 200 205
Gln Ala Thr Lys Lys Thr Gly Lys Trp Asp Ser Asn Thr Ser Thr Leu
210 215 220
Thr Ile Ser Val Asn Ser Lys Lys Thr Lys Asn Ile Val Phe Thr Lys
225 230 235 240
Glu Asp Thr Ile Thr Val Gln Lys Tyr Asp Ser Ala Gly Thr Asn Leu
245 250 255
Glu Gly Asn Ala Val Glu Ile Lys Thr Leu Asp Glu Leu Lys Asn Ala
260 265 270
Leu Lys
<210> 172
<211> 822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический нуклеотид
<400> 172
atgaaaaaat atttattggg aataggtcta atattagcct taatagcatg taagcaaaat 60
gttagcagcc ttgatgaaaa aaatagcgtt tcagtagatt tacctggtgg aatgaaagtt 120
cttgtaagta aagaaaaaga caaagatggt aaatacagtc taatggcaac agtagaaaag 180
cttgagctta aaggaacttc tgataaaaac aacggttctg gaacacttga aggtgaaaaa 240
actgacaaaa gtaaagtaaa attaacaatt gctgaggatc taagtaaaac cacatttgaa 300
atcttcaaag aagatggcaa aacattagta tcaaaaaaag taacccttaa agacaagtca 360
tcaacagaag aaaaattcaa cgaaaagggt gaaatatctg aaaaaacaat agtaagagca 420
aatggaacca gacttgaata cacagacata aaaagcgata aaaccggaaa agctaaagaa 480
gttttaaaag actttactct tgaaggaact ctagctgctg acggcaaaac aacattgaaa 540
gttacagaag gcactgttac tttaagcaag aacatttcaa aatccggaga aataacagtt 600
gcacttgatg acactgactc tagcggcaat aaaaaatccg gaacatggga ttcagatact 660
tctactttaa caattagtaa aaacagtcaa aaaactaaac aacttgtatt cacaaaagaa 720
aacacaataa cagtacaaaa ctataacaga gcaggcaatg cgcttgaagg cagcccagct 780
gaaattaaag atcttgcaga gcttaaagcc gctttaaaat aa 822
<210> 173
<211> 273
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 173
Met Lys Lys Tyr Leu Leu Gly Ile Gly Leu Ile Leu Ala Leu Ile Ala
1 5 10 15
Cys Lys Gln Asn Val Ser Ser Leu Asp Glu Lys Asn Ser Val Ser Val
20 25 30
Asp Leu Pro Gly Gly Met Lys Val Leu Val Ser Lys Glu Lys Asp Lys
35 40 45
Asp Gly Lys Tyr Ser Leu Met Ala Thr Val Glu Lys Leu Glu Leu Lys
50 55 60
Gly Thr Ser Asp Lys Asn Asn Gly Ser Gly Thr Leu Glu Gly Glu Lys
65 70 75 80
Thr Asp Lys Ser Lys Val Lys Leu Thr Ile Ala Glu Asp Leu Ser Lys
85 90 95
Thr Thr Phe Glu Ile Phe Lys Glu Asp Gly Lys Thr Leu Val Ser Lys
100 105 110
Lys Val Thr Leu Lys Asp Lys Ser Ser Thr Glu Glu Lys Phe Asn Glu
115 120 125
Lys Gly Glu Ile Ser Glu Lys Thr Ile Val Arg Ala Asn Gly Thr Arg
130 135 140
Leu Glu Tyr Thr Asp Ile Lys Ser Asp Lys Thr Gly Lys Ala Lys Glu
145 150 155 160
Val Leu Lys Asp Phe Thr Leu Glu Gly Thr Leu Ala Ala Asp Gly Lys
165 170 175
Thr Thr Leu Lys Val Thr Glu Gly Thr Val Thr Leu Ser Lys Asn Ile
180 185 190
Ser Lys Ser Gly Glu Ile Thr Val Ala Leu Asp Asp Thr Asp Ser Ser
195 200 205
Gly Asn Lys Lys Ser Gly Thr Trp Asp Ser Asp Thr Ser Thr Leu Thr
210 215 220
Ile Ser Lys Asn Ser Gln Lys Thr Lys Gln Leu Val Phe Thr Lys Glu
225 230 235 240
Asn Thr Ile Thr Val Gln Asn Tyr Asn Arg Ala Gly Asn Ala Leu Glu
245 250 255
Gly Ser Pro Ala Glu Ile Lys Asp Leu Ala Glu Leu Lys Ala Ala Leu
260 265 270
Lys
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕННЫЕ ПОЛИПЕПТИДЫ НА ОСНОВЕ ПОСЛЕДОВАТЕЛЬНОСТИ OspA | 2019 |
|
RU2816208C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| ХИМЕРНЫЕ ГЕНЫ OSPA, БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2636455C2 |
| ХИМЕРНЫЕ ГЕНЫ OSPA, БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2583289C2 |
| НОВЫЙ КОРОНАВИРУС РЫБ | 2020 |
|
RU2813731C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| ВАКЦИНА ПРОТИВ АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ | 2020 |
|
RU2819672C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ТРИ СЛИТЫХ БЕЛКА OSPA, ДЛЯ ПРИМЕНЕНИЯ В МЕДИЦИНЕ | 2021 |
|
RU2837669C1 |
| НОВЫЙ TOTIVIRUS РЫБ | 2020 |
|
RU2798051C2 |
| КОМПОЗИЦИЯ МИКОБАКТЕРИАЛЬНОГО АНТИГЕНА | 2015 |
|
RU2702194C2 |
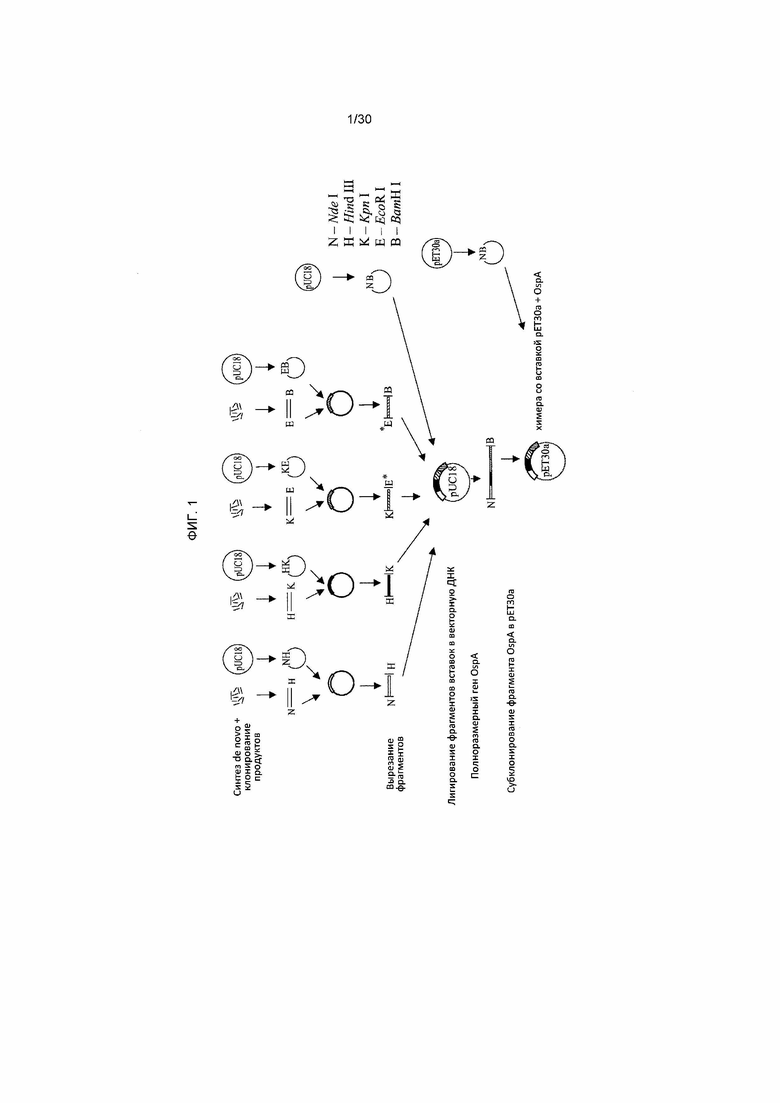











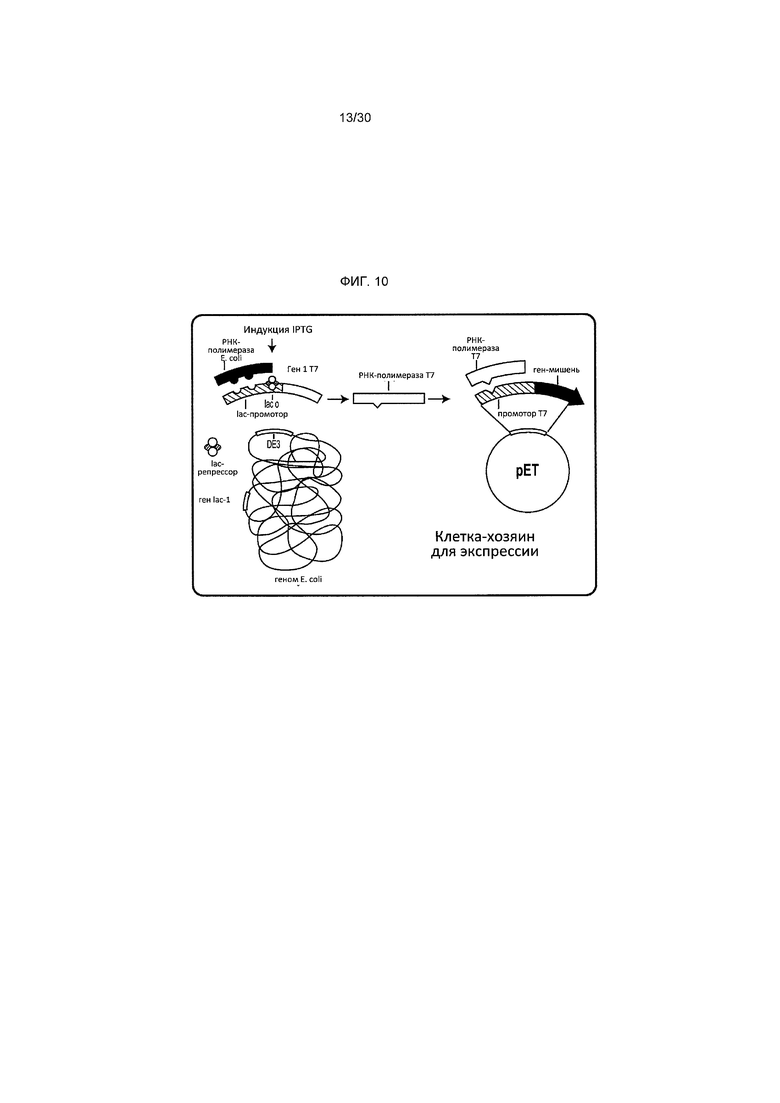
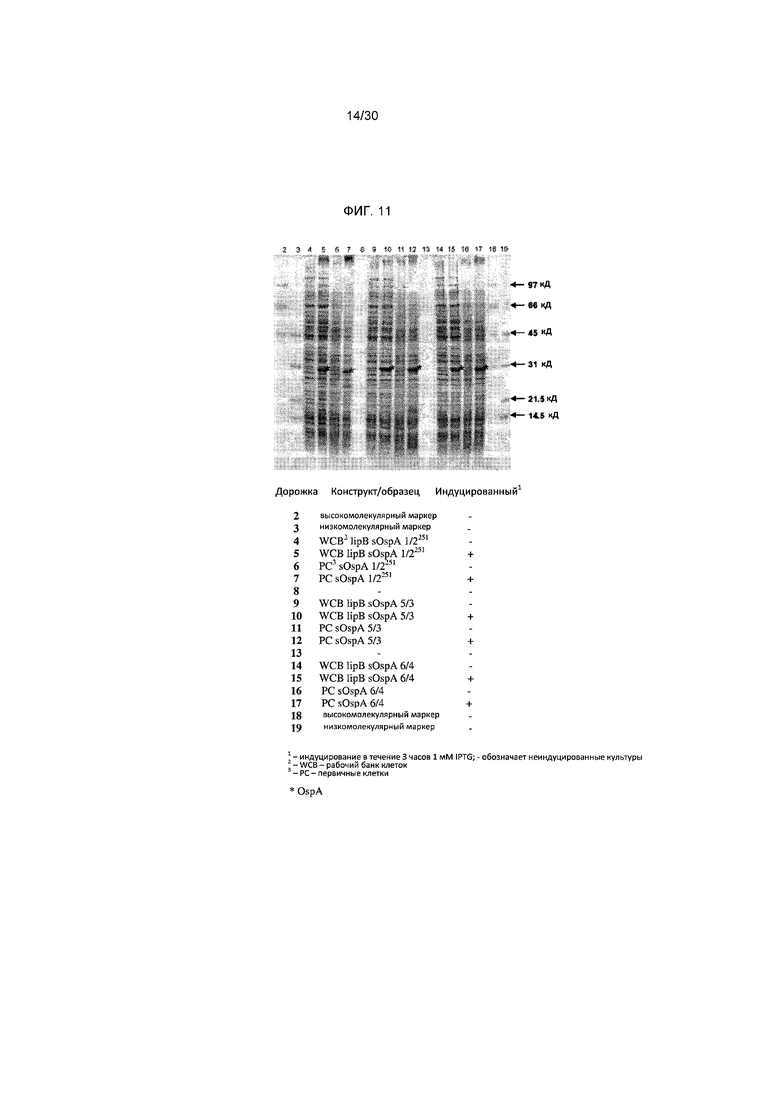

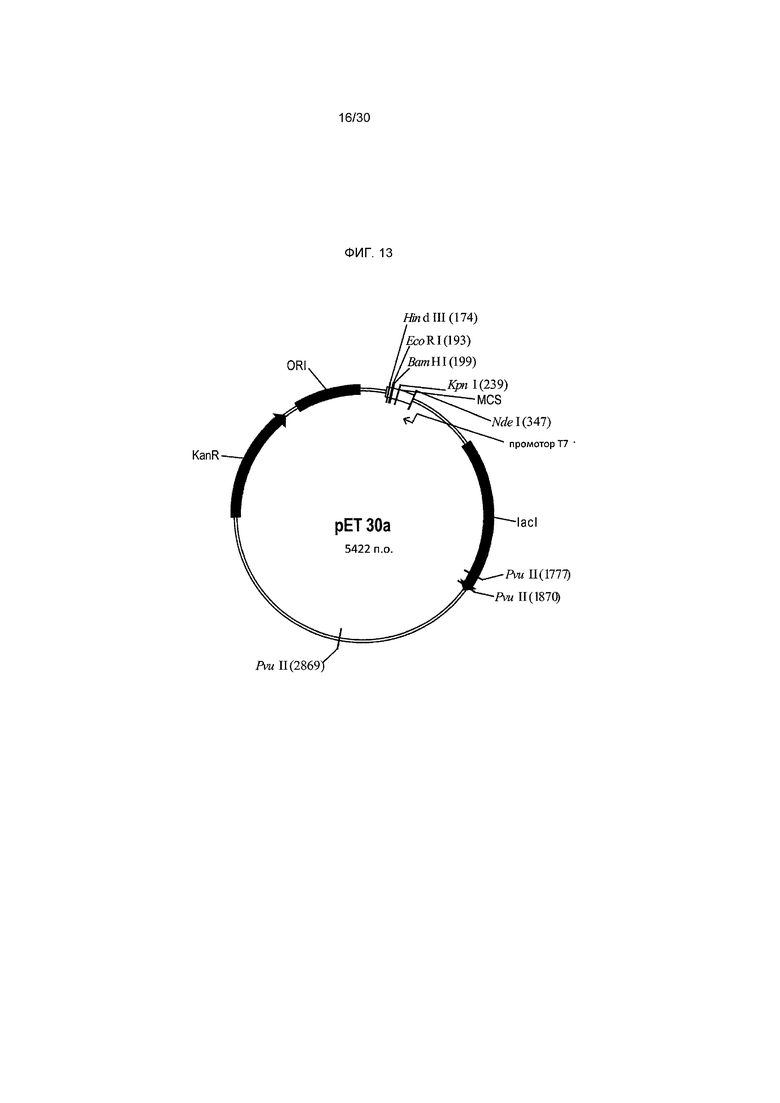








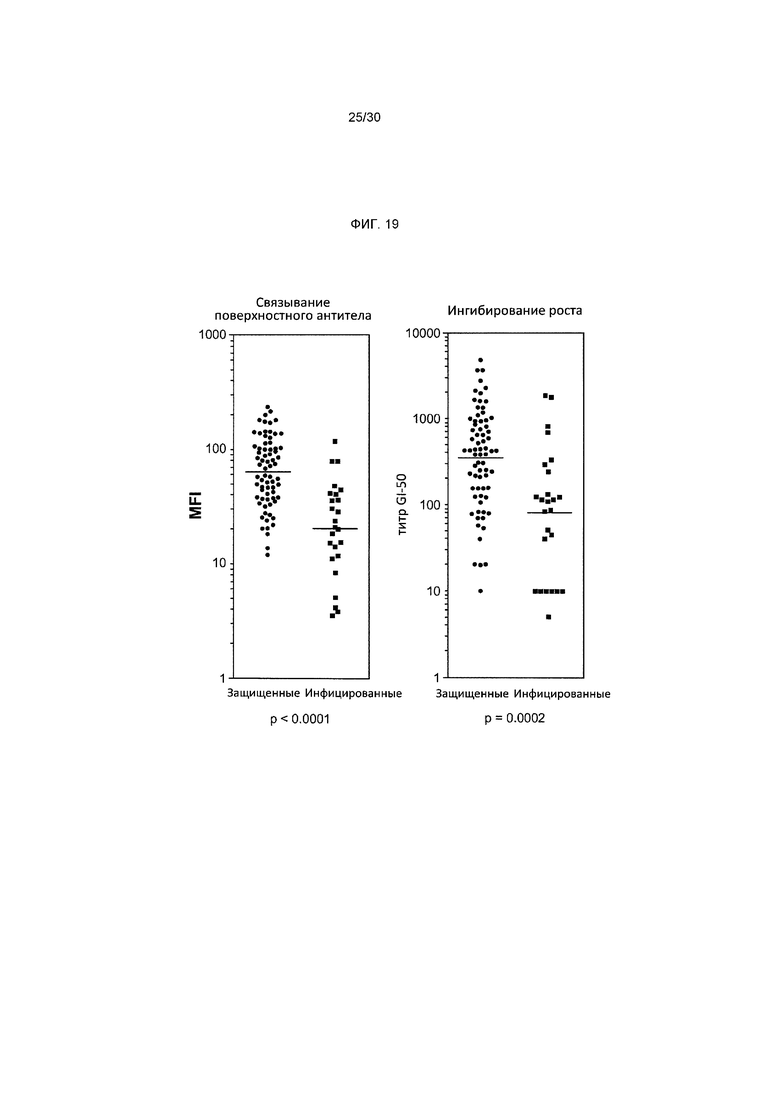


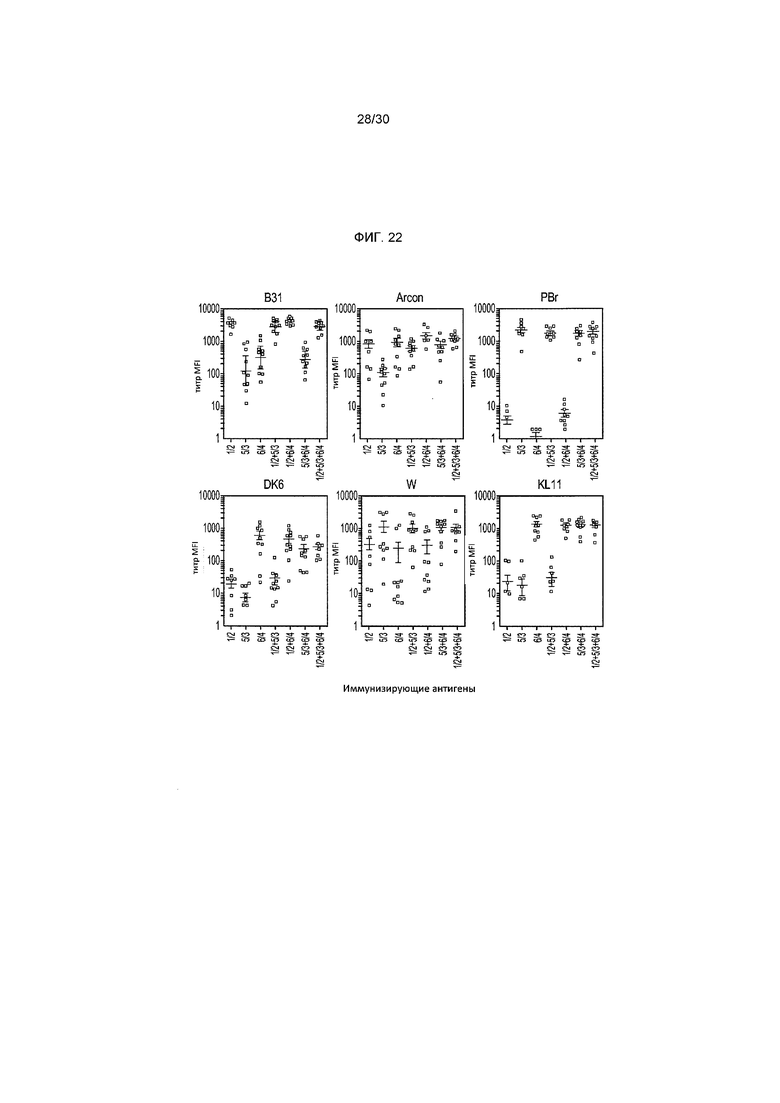
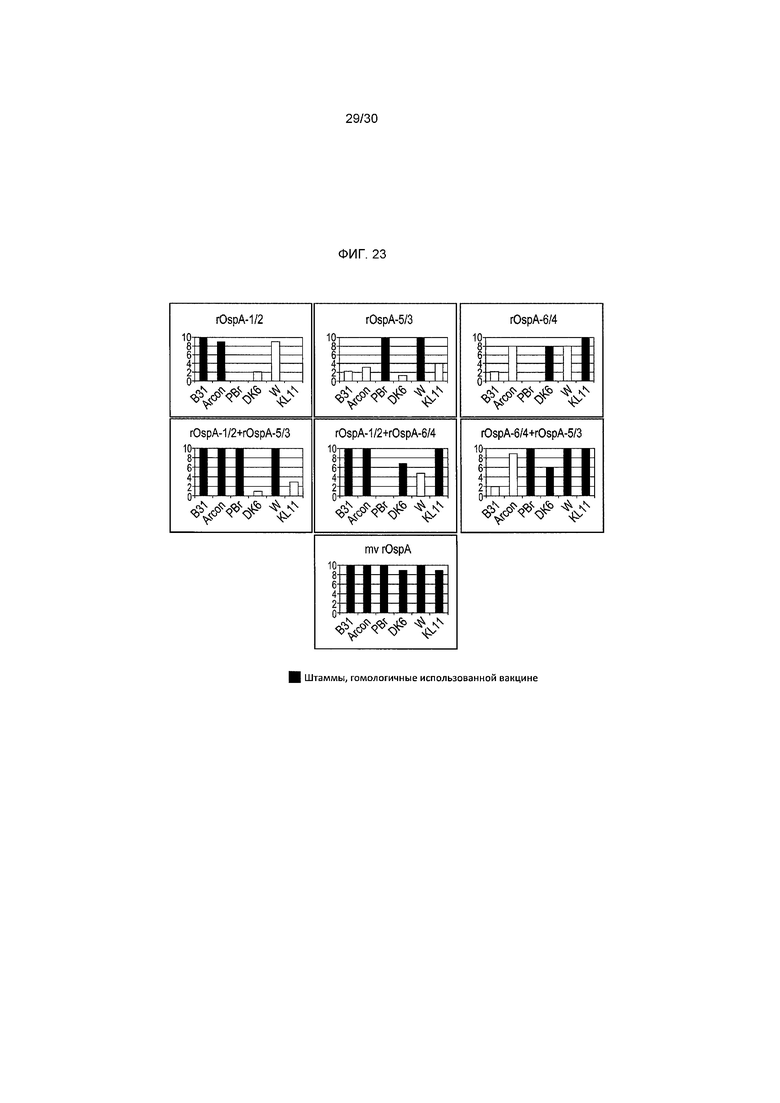
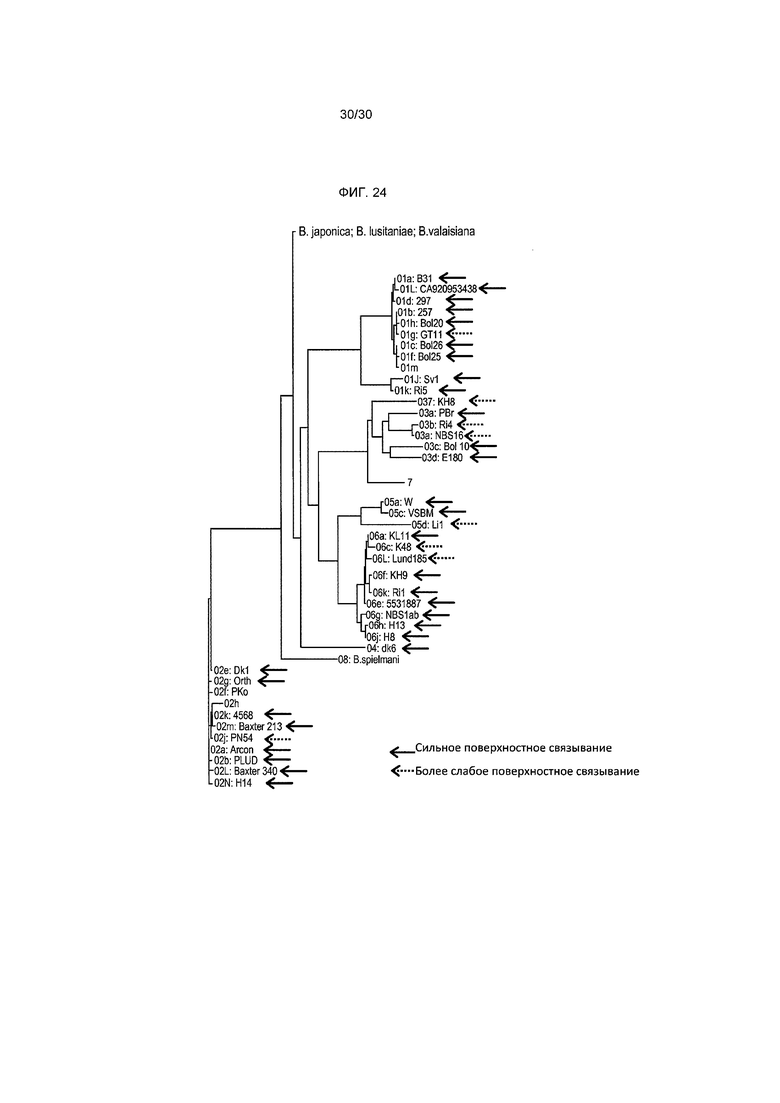
Изобретение относится к области биотехнологии, в частности к иммуногенным композициям, способным вызывать специфический иммунный ответ против бактерий Borrelia, и может быть использовано в медицине. Более конкретно изобретение относится к новым химерным молекулам OspA, содержащим проксимальный участок одного серотипа OspA вместе с дистальным участком другого серотипа OspA, сохраняя при этом антигенные свойства обоих родительских полипептидов, а также кодирующим их нуклеиновым кислотам. Фармацевтические композиции, содержащие указанные нуклеиновые кислоты в качестве активного ингредиента, могут быть применимы в качестве лекарственного средства при генной терапии боррелиоза, болезни Лайма и иных переносимых клещами болезней. 5 н. и 7 з.п. ф-лы, 22 пр., 9 табл., 24 ил.
1. Иммуногенная композиция, способная вызывать специфический иммунный ответ против бактерий рода Borrelia, содержащая эффективное количество:
(i) молекулы нуклеиновой кислоты, выбранной из группы, состоящей из:
(a) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью с по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полноразмерной нуклеотидной последовательностью, приведенной в SEQ ID NO: 1;
(b) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, выбранной из группы, состоящей из последовательностей, приведенных в SEQ ID NO: 1 или SEQ ID NO: 7;
(c) молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, выбранной из группы, состоящей из последовательностей, приведенных в SEQ ID NO: 1 или SEQ ID NO: 7;
(d) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид с по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полноразмерным полипептидом, содержащим аминокислотную последовательность, приведенную в SEQ ID NO: 2;
(e) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 2, где полипептид имеет замену от одной до 10 аминокислот;
(f) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 2, где полипептид имеет инсерцию от одной до 10 аминокислот;
(g) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 2, где полипептид имеет внутреннюю делецию от одного до 10 аминокислот;
(h) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 2, где полипептид имеет C- и/или N-терминальное усечение от одной до 10 аминокислот; и
(i) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 2, где полипептид имеет модификацию от одной до 10 аминокислот, выбранных из аминокислотных замен, аминокислотных инсерций, аминокислотных делеций, C-терминального усечения или N-терминального усечения; или
(j) молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 2;
где молекула нуклеиновой кислоты кодирует иммуногенный полипептид, способный вызывать специфический иммунный ответ против бактерий рода Borrelia,
(ii) молекулы нуклеиновой кислоты, выбранной из группы, состоящей из:
(a) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью с по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полноразмерной нуклеотидной последовательностью, приведенной в SEQ ID NO: 3;
(b) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, выбранной из группы, состоящей из последовательностей, приведенных в SEQ ID NO: 3 или SEQ ID NO: 9;
(c) молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, выбранной из группы, состоящей из последовательностей, приведенных в SEQ ID NO: 3 или SEQ ID NO: 9;
(d) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид с по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности с полноразмерным полипептидом, содержащим аминокислотную последовательность, приведенную в SEQ ID NO: 4;
(e) молекулы нуклеиновой кислоты, характеризующейся последовательностью нуклеотидной кислоты, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 4, где полипептид имеет замену от одной до 10 аминокислот;
(f) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 4, где полипептид имеет инсерцию от одной до 10 аминокислот;
(g) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 4, где полипептид имеет внутреннюю делецию от одной до 10 аминокислот;
(h) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 4, где полипептид имеет С- и/или N-терминальное усечение от одной до 10 аминокислот; и
(i) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 4, где полипептид имеет модификацию от одной до 10 аминокислот, выбранных из аминокислотных замен, аминокислотных инсерций, аминокислотных делеций, С- и/или N-терминального усечения; и
(j) молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO:4;
где молекула нуклеиновой кислоты кодирует иммуногенный полипептид, способный вызывать специфический иммунный ответ против бактерий рода Borrelia,
(iii) молекулы нуклеиновой кислоты, выбранной из группы, состоящей из:
(а) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью с по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полноразмерной нуклеотидной последовательностью, приведенной в SEQ ID NO: 5;
(b) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, выбранной из группы, состоящей из последовательностей, приведенных в SEQ ID NO: 5 или SEQ ID NO: 11;
(с) молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, выбранной из группы, состоящей из последовательностей, приведенных в SEQ ID NO: 5 или SEQ ID NO: 11;
(d) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид с по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности последовательности с полноразмерным полипептидом, содержащим аминокислотную последовательность, приведенную в SEQ ID NO: 6;
(e) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 6, где полипептид имеет замену от одной до 10 аминокислот;
(f) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 6, где полипептид имеет инсерцию от одной до 10 аминокислот;
(g) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 6, где полипептид имеет внутреннюю делецию от одной до 10 аминокислот;
(h) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 6, где полипептид имеет С- и/или N-терминальное усечение от одной до 10 аминокислот; и
(i) молекулы нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, кодирующей полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 6, где полипептид имеет модификацию от одной до 10 аминокислот, выбранных из аминокислотных замен, аминокислотных инсерций, аминокислотных делеций, С- или N-терминального усечения; и
(j) молекулы нуклеиновой кислоты, состоящей из нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, приведенный в SEQ ID NO: 6;
где молекула нуклеиновой кислоты кодирует иммуногенный полипептид, способный вызывать специфический иммунный ответ против бактерий рода Borrelia;
(iv) адъюванта; и
(v) фармацевтически приемлемого носителя.
2. Иммуногенная композиция, способная вызывать специфический иммунный ответ против бактерий рода Borrelia, содержащая эффективное количество:
(i) полипептида, выбранного из группы, состоящей из:
(а) полипептида, характеризующегося аминокислотной последовательностью, имеющей по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентов идентичности последовательности с полноразмерной аминокислотной последовательностью, приведенной в SEQ ID NO: 2;
(b) полипептида, характеризующегося аминокислотной последовательностью, приведенной в SEQ ID NO: 2 или SEQ ID NO: 8; и
(с) полипептида, состоящего из аминокислотной последовательности, приведенной в SEQ ID NO: 2 и SEQ ID NO:8;
где полипептид способен вызывать специфический иммунный ответ против бактерий рода Borrelia;
(ii) полипептида, выбранного из группы, состоящей из:
(a) полипептида, характеризующегося аминокислотной последовательностью, имеющей по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентов идентичности последовательности с полноразмерной аминокислотной последовательностью, приведенной в SEQ ID NO: 4;
(b) полипептида, характеризующегося аминокислотной последовательностью, приведенной в SEQ ID NO: 4 или SEQ ID NO: 10; и
(с) полипептида, состоящего из аминокислотной последовательности множества указанных в SEQ ID NO: 4 или SEQ ID NO: 10,
где полипептид способен вызывать специфический иммунный ответ против бактерий рода Borrelia;
(iii) полипептида, выбранного из группы, состоящей из:
(а) полипептида, характеризующегося аминокислотной последовательностью, имеющей по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентов идентичности последовательности с полноразмерной аминокислотной последовательностью, приведенной в SEQ ID NO: 6;
(b) полипептида, характеризующегося аминокислотной последовательностью, указанной в SEQ ID NO: 6 или SEQ ID NO: 12; и
(с) полипептида, состоящего из аминокислотной последовательности, приведенной в SEQ ID NO: 6 или SEQ ID NO: 12,
где полипептид способен вызывать специфический иммунный ответ против бактерий рода Borrelia;
(iv) адъюванта; и
(v) фармацевтически приемлемого носителя.
3. Иммуногенная композиция по п. 1 или 2, где композицию выбирают из группы, состоящей из:
(а) композиции, обладающей способностью индуцировать продуцирование антитела, которое специфически связывается с внешним поверхностным белком A (OspA);
(b) композиции, обладающей способностью индуцировать продуцирование антитела, которое специфически связывается с Borrelia;
(с) композиции, обладающей способностью индуцировать продуцирование антитела, нейтрализующего Borrelia;
(d) композиции, где Borrelia представляет собой Borrelia burgdorferi sensu lato;
(e) композиции, где Borrelia представляет собой Borrelia afzelii, Borrelia garinii или Borrelia burgdorferi sensu stricto; и
(f) композиции, где Borrelia представляет собой Borrelia japonica, Borrelia andersonii, Borrelia bissettii, Borrelia sinica, Borrelia turdi, Borrelia tanukii, Borrelia valaisiana, Borrelia lusitaniae, Borrelia spielmanii, Borrelia miyamotoi или Borrelia lonestar.
4. Композиция вакцины, способная вызывать специфический иммунный ответ против бактерий рода Borrelia, содержащая эффективное количество иммуногенной композиции по любому из пп. 1-3 и фармацевтически приемлемый носитель.
5. Комбинированная вакцина, способная вызывать специфический иммунный ответ против бактерий рода Borrelia и второй переносимой клещами болезни, содержащая эффективное количество композиции вакцины по п. 4 в сочетании с по меньшей мере второй композицией вакцины.
6. Комбинированная вакцина по п. 5, где переносимая клещами болезнь представляет собой пятнистую лихорадку Скалистых гор, бабезиоз, возвратную лихорадку, колорадскую клещевую лихорадку, моноцитарный эрлихиоз человека (HME), гранулоцитарный эрлихиоз человека (HGE), клещевую сыпную лихорадку STARI, туляремию, клещевой паралич, энцефалит Повассан, ку-лихорадку, конго-крымскую геморрагическую лихорадку, цитауксзооноз, марсельскую лихорадку или клещевой энцефалит.
7. Комбинированная вакцина по п. 5, где вторая композиция вакцины представляет собой вакцину, выбранную из группы, состоящей из: вакцины клещевого энцефалита, вакцины японского энцефалита и вакцины пятнистой лихорадки Скалистых гор.
8. Комбинированная вакцина по п. 5, отличающаяся тем, что вторая композиция вакцины имеет сезонный график иммунизации, совместимый с иммунизацией против инфекции Borrelia или лаймской болезни.
9. Иммуногенная композиция по любому из пп. 1-3 для индуцирования иммунологического ответа.
10. Композиция по п. 9, где иммунологический ответ включает продуцирование анти-OspA антитела.
11. Композиция вакцины по п. 4 или комбинированная вакцина по любому из пп. 5-8 для профилактики или лечения инфекции Borrelia или лаймской болезни.
12. Применение композиции по любому из пп. 1-4 или комбинированной вакцины по любому из пп. 5-8 в качестве лекарственного средства для предотвращения или лечения инфекции Borrelia или лаймской болезни.
| УСТРОЙСТВО ДЛЯ ХОЛОДНОЙ СВАРКИ | 1966 |
|
SU216421A1 |
| EP 1939294 A1, 02.07.2008 | |||
| US 6143872 A1, 07.11.2000 | |||
| GERN L | |||
| et al | |||
| Immunization with a polyvalent OspA vaccine protects mice against Ixodes ricinus tick bites infected by Borrelia burgdorferi ss, Borrelia garinii and Borrelia afzelii, VACCINE, 1997, v.15, n.14, p | |||
| Электролизер с биполярными электродами | 1923 |
|
SU1551A1 |
| ХОЛЕСТЕРИНСОДЕРЖАЩИЕ СОЕДИНЕНИЯ И ИХ ПРИМЕНЕНИЕ В КАЧЕСТВЕ ИММУНОГЕНОВ ПРОТИВ BORRELIA BURGDORFERI | 2004 |
|
RU2385323C2 |

