Изобретение относится к средству для нейтрализации вируса натуральной оспы на основе одноцепочечного антитела человека, сконструированного in vitro на основе генов, кодирующих вариабельные домены иммуноглобулинов людей, вакцинированных вирусом осповакцины, рекомбинантную фагмидную ДНК pHEN-1A, содержащую полученный генно-инженерными методами искусственный ген одноцепочечного антитела человека под контролем промотора лактозного оперона, обеспечивающий в клетках Escherichia coli синтез одноцепочечного антитела человека в составе химерного белка с оболочечным белком p3 бактериофага M13, способного нейтрализовать ортопоксвирусы, а также одноцепочечное антитело человека 1A, экспонированное на поверхности нитчатого бактериофага M13 в составе оболочечного белка p3, способное нейтрализовать вирус натуральной оспы, вирус осповакцины и вирус оспы коров, и может быть использовано в медицине, биотехнологии, белковой и генной инженерии.
Род Orthopoxvirus, входящий в семейство Poxviridae, включает в себя сложные ДНК-содержащие вирусы, реплицирующиеся в цитоплазме клеток позвоночных и беспозвоночных. К этому роду относятся патогенные для человека вирусы - вирус натуральной оспы и вирус оспы обезьян, вирусы, в норме вызывающие только местные инфекции - вирус осповакцины и вирус оспы коров, и непатогенные для человека вирусы - вирус эктромелии, вирус оспы верблюдов и др. [1]. Следует отметить, что у людей с ослабленным иммунным статусом вирусы осповакцины и оспы коров могут стать причиной тяжелых заболеваний или даже генерализованной инфекции.
В связи с прекращением естественной трансмиссии вируса натуральной оспы была прекращена массовая вакцинация вирусом осповакцины. Однако, несмотря на ликвидацию натуральной оспы, существуют причины, по которым ортопоксвирусы продолжают оставаться источником биологической опасности для людей, причем стихийному или преднамеренному распространению натуральной оспы может способствовать то, что в настоящее время большинство населения не имеет иммунитета к этим вирусам. Кроме того, возобновление массовых вакцинаций вирусом осповакцины в США подтвердило неизбежность возникновения поствакцинальных осложнений в ряде случаев, тем более что большая часть вакцинированных взрослых не были привиты осповакциной в детстве [2].
Вакцинация вирусом осповакцины, давая надежную защиту [3], может сопровождаться рядом осложнений, особенно у людей с врожденным или приобретенным иммунодефицитом, поэтому существует необходимость в разработке специфических средств профилактики поствакцинальных осложнений. В настоящее время таким средством является сывороточный вакцинный иммуноглобулин (VIG) [1], однако использование препаратов, полученных из человеческой крови, всегда сопровождается определенным биологическим риском, к тому же этот препарат дорог и малодоступен. Альтернативу вакцинному иммуноглобулину могли бы составить рекомбинантные антитела человека, специфичные к ортопоксвирусам.
В настоящее время наиболее предпочтительными являются полностью человеческие рекомбинантные антитела (fully human antibodies). Для их создания используют объединение вариабельных доменов антител человека, обладающих целевой активностью, с константными доменами иммуноглобулинов человека нужного изотипа. Одним из способов получения вариабельных доменов является их отбор из комбинаторных фаговых библиотек мини-антител человека. Важно, чтобы отобранные вариабельные домены обладали вируснейтрализующими свойствами в отношении целевых вирусов. К настоящему времени таким способом сконструированы полноразмерные антитела человека против ряда вирусных агентов [4, 5, 6, 7]. Известны сконструированные таким способом полноразмерные антитела человека против ортопоксвирусов [8, 9, 10]. Эти антитела были способны связывать вирусы осповакцины оспы коров [8, 9, 10] и нейтрализовать вирус осповакцины [10]. Антитела b9 и с4 также способны подавлять инфекционность вируса оспы обезьян [10]. Полноразмерные антитела человека, обладающие вируснейтрализующими и протективными свойствами в отношении вируса осповакцины, были получены с использованием трансгенных мышей [11].
Наиболее близким аналогом (прототипом) является антитело 2А8 [12], отобранное по способности связывать вирус оспы коров и подавлять инфекционность вирусов осповакцины и оспы коров из иммунной комбинаторной фагмидной библиотеки одноцепочечных антител человека, сконструированной in vitro на основе генов, кодирующих вариабельные домены иммуноглобулинов людей, вакцинированных вирусом осповакцины [13].
Однако в вышеприведенных технических решениях [8-12] свойства подавлять инфекционность вируса натуральной оспы для антител 2А8, b9 и с4 показано не было.
Техническим результатом заявляемого изобретения является создание с помощью фагового дисплея плазмидной ДНК pHEN-1A, обеспечивающей синтез в клетках Escherichia coli одноцепочечного антитела человека 1А, экспонированного на поверхности бактериофага М13К07 в составе оболочечного белка р3 и способного осуществлять нейтрализацию инфекционности вируса натуральной оспы, вируса осповакцины и вируса оспы коров.
Указанный технический результат достигается отбором из фагмидной коллекции рекомбинантной фагмидной ДНК pHEN-1A, содержащей уникальный ген одноцепочечного антитела человека 1А, способного нейтрализовать вирус натуральной оспы, вирус осповакцины и вирус оспы коров; получением одноцепочечного антитела человека 1A, экспонированного на поверхности бактериофага М13К07 в составе оболочечного белка р3 и способного осуществлять нейтрализацию инфекционности вируса натуральной оспы, вируса осповакцины и вируса оспы коров.
Сущность изобретения заключается в следующем:
- из иммунной фаговой библиотеки отобрана фагмидная ДНК pHEN-1A, содержащая уникальный ген одноцепочечного антитела человека 1А, способного нейтрализовать вирус натуральной оспы, вирус осповакцины и вирус оспы коров;
- выделено фаговое одноцепочечное антитело 1А, экспонированное на поверхности нитчатого бактериофага М13К07 в составе оболочечного белка р3, способное нейтрализовать вирусы натуральной оспы, осповакцины и оспы коров.
Методами фагового дисплея отбирают одноцепочечное антитело человека 1А, экспонированное на поверхности нитчатого бактериофага М13К07 в составе оболочечного белка р3, кодируемое фагмидной ДНК pHEN-1A. Полученное антитело способно нейтрализовать вирусы натуральной оспы, осповакцины и оспы коров.
Для получения антител против ортопоксвирусов, например вируса эктромелии, проводят аффинное обогащение комбинаторной иммунной библиотеки специфическими клонами. Комбинаторная библиотека антител представляет собой коллекцию бактерофагов, каждый из которых экспонирует на своей поверхности уникальное одноцепочечное антитело в составе оболочечного фагового белка [14]. Обогащение библиотеки проводят с использованием вируса эктромелии в качестве антигена с последующим отбором индивидуальных клонов, способных продуцировать бактериофаги, экспонирующие на своей поверхности одноцепочечные антитела человека против вируса эктромелии. Для этого аликвоту трансформированных клеток Escherichia coli размораживают, растят до среднелогарифмической фазы и инфицируют фагом-помощником М13К07, при этом получают популяцию нитчатых бактериофагов, каждый из которых экспонирует на своей поверхности уникальное одноцепочечное антитело человека в виде химерного белка с оболочечным белком p3 фага М13К07.
Аффинное обогащение библиотеки проводят в ходе двух последовательных раундов. Для проведения первого раунда обогащения в лунках полистиролового планшета сорбируют вирус эктромелии (штамм К-1) в концентрации 100 мкг/мл. По окончании сорбции места неспецифического связывания насыщают 5%-ным раствором сухого обезжиренного молока в фосфатно-солевом буфере (ФСБР), после чего в каждую лунку добавляют аликвоту (1011 БОЕ) библиотеки в виде коллекции нитчатых бактериофагов. Неспецифически связавшиеся фаговые частицы удаляют при промывке ФСБР, а фаги, несущие на своей поверхности одноцепочечные антитела, способные связываться с вирусом эктромелии, элюируют раствором триэтиламина. Полученным элюатом инфицируют культуру клеток Е.coli TG1 в стадии экспоненциального роста для амплификации элюированных фаговых антител.
Второй раунд аффинного обогащения проводят аналогично. При этом концентрации вируса эктромелии уменьшают до 50 мкг/мл соответственно. Степень обогащения библиотеки антителами, специфическими к вирусу эктромелии, проверяют с использованием непрямого иммуноферментного анализа (ИФА) по способности популяций фаговых антител, полученных в результате каждого раунда аффинного обогащения, связывать вирус эктромелии (фиг.1). При этом фаговые антитела выделяют, как описано в Примере 1. В качестве отрицательного контроля используют связывание популяций фаговых антител с сорбированным 5% обезжиренным молоком в ФСБР.
Далее, из популяции фаговых антител, обогащенной против вируса эктромелии, проводят отбор моноклональных фаговых антител, специфически связывающих вирус эктромелии, с помощью ИФА. Вирус эктромелии сорбируют на 96-луночные иммунологические планшеты («Медполимер», Россия) в концентрации 3 мкг/мл. Места неспецифического связывания блокируют раствором 3%-ного бычьего сывороточного альбумина (БСА, «Sigma») в фосфатном буфере, рН 7.2. Затем в лунки вносят выделенные, как описано в Примере 1, фаговые антитела, разведенные ФСБР, содержащим 0,1% Твин, и инкубируют 1 час при 37°С. После промывки в лунки вносят поликлональные анти-М13 антитела кролика в разведении 1:16000, а затем антивидовой конъюгат щелочной фосфатазы («Sigma») в разведении 1:4000. В качестве хромогена используют паранитрофенилфосфат. Контролем неспецифического связывания служит связывание вируса с фагом-помощником М13К07, не несущим фрагментов антител на своей поверхности. В качестве отрицательного контроля используют связывание фаговых антител с БСА. Клоны, продемонстрировавшие сигнал, значение которого в 3 и более раз превышает контроль неспецифического связывания и отрицательный контроль, отбирают как положительные (фиг.2).
Отобранные фаговые антитела выделяют и исследуют с помощью ИФА при последовательном разведении антитела. При этом в качестве антигенов для более полной характеристики исследуемых антител можно использовать помимо вируса эктромелии другие ортопоксвирусы, например вирусы осповакцины и оспы коров (фиг.3).
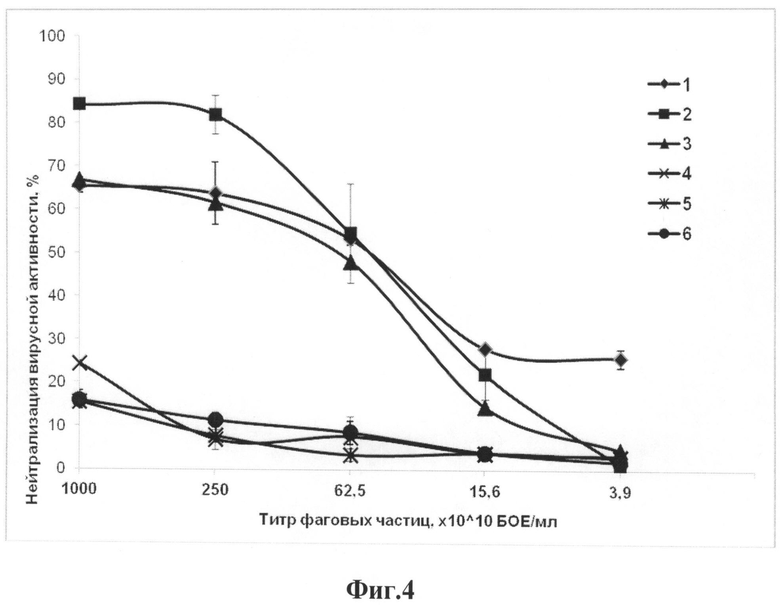
Стандартный анализ вируснейтрализующей активности проводят для всех фаговых антител, специфически связывающих вирус эктромелии. В ходе этого анализа проверяют способность ингибировать образование бляшек вирусами натуральной оспы, осповакцины и оспы коров на монослое клеток Vero E6. В качестве отрицательного контроля используют бактериофаг-помощник М13К07, не несущий на своей поверхности одноцепочечного антитела. Способность нейтрализовать вирусную инфекционность оценивают по титру нейтрализации, который определяется как разведение фаговых антител, демонстрирующее уменьшение количества вирусных бляшек не менее чем на 50% (фиг.4).
Далее проводят определение нуклеотидных последовательностей генов, кодирующих одноцепочечное антитело, нейтрализующее вирусную инфекционность. Для амплификации гена, кодирующего одноцепочечное антитело 1А, в качестве матрицы используют фагмидные ДНК, выделенные методом щелочного лизиса [15], и олигонуклеотидные праймеры: LMB 5'-CAGGAAACAGTCATGAC, pHEN-SEQ 5'-CTATGGGGCCCCATTCA. Амплификацию осуществляют методом ПЦР с использованием TaqSE ДНК-полимеразы при температуре отжига праймеров LMB и pHEN-SEQ 52°С. Определение нуклеотидных последовательностей очищенных Vh и Vl фрагментов проводят в обоих направлениях с использованием автоматического секвенатора ABI 3730XL Genetic Analyser (Applied Biosystems) и набора BigDye® Terminator v3.1 Cycling Sequencing Kit (Applied Biosystems, США. Для анализа последовательностей используют программы SeqMan Lasergene, Vector NTI Suite 8 и MEGA 4.1. Полученные последовательности сравнивают с последовательностями, размещенными в базе данных NCBI BLAST. На фиг.5 представлена нуклеотидная последовательность и кодируемая ею аминокислотная последовательность уникального гена одноцепочечного антитела человека, обладающего вируснейтрализующей активностью в отношении вирусов натуральной оспы, осповакцины и оспы коров, а на фиг.6 представлена аминокислотная последовательность этого рекомбинантного антитела 1А.
Полученная из описанной фагмидной коллекции рекомбинантная фагмидная ДНК pHEN-1A, содержащая уникальный ген одноцепочечного антитела человека 1А, способного нейтрализовать вирусы натуральной оспы, осповакцины и оспы коров, характеризуется следующими признаками (фиг.7):
- имеет молекулярную массу 3,47 МДа и размер 5255 п.о.;
- кодирует одноцепочечное антитело человека 1А в составе химерного белка с оболочечным белком р3 фага М13К07;
- состоит из следующих элементов:
а) фрагмента ДНК, размером 840 п.о., включающего искусственный ген одноцепочечного антитела человека 1А, способного нейтрализовать ортопоксвирусы, в котором вариабельный домен тяжелой цепи соединен с вариабельным доменом легкой цепи иммуноглобулина человека с помощью ДНК последовательности, кодирующей гибкий пептидный линкер Ser(Gly4 Ser)2AlaArgGlySerGly4 Ser, последовательность, кодирующую пептид HHHHHH, и имеющего нуклеотидную последовательность SEQ ID NO:1, представленную на фиг.4;
б) фагмидного вектора pHEN2 (MRC, Великобритания), обеспечивающего эффективную транскрипцию полученного гена, кодирующего одноцепочечное антитело человека 1А, и его экспрессию;
содержит:
а) сайт инициации репликации ColE1 из плазмиды pBR322;
б) промотор Lac-оперона Е.coli;
в) генетический маркер: AMPr - ген β-лактамазы (bla), определяющий устойчивость трансформированных фагмидой pHEN-1A клеток Escherichia coli к ампициллину;
г) искусственный ген одноцепочечного антитела человека 1А, включающий последовательность, кодирующую пептид HHHHHH, способного нейтрализовать ортопоксвирусы, в котором вариабельный домен тяжелой цепи соединен с вариабельным доменом легкой цепи иммуноглобулина человека с помощью ДНК последовательности, кодирующей гибкий пептидный линкер Ser(Gly4 Ser)2AlaArgGlySerGly4 Ser,
д) уникальные сайты узнавания эндонуклеазами рестрикции, имеющие следующие координаты: HindIII(235), SfiI(328), NotI(1088), BamHI(1749), ClaI(2057), AvaI(2951), EcoRI(2379).
Таким образом, впервые получена плазмидная ДНК pHEN-1A, обеспечивающая синтез в клетках Escherichia coli одноцепочечного антитела человека 1А, экспонированного на поверхности бактериофага М13К07 в составе оболочечного белка р3 и способного осуществлять нейтрализацию инфекционности вируса натуральной оспы, вируса осповакцины и вируса оспы коров.
Изобретение иллюстрируется следующими графическими материалами, представленными на фиг.1-7:
Фиг.1. Связывание исходной библиотеки и популяций фаговых антител, полученных после 1-го и 2-го раундов аффинного обогащения, с вирусом эктромелии (темные столбики) и 5% раствором обезжиренного молока в ФСБР (светлые столбики) в ИФА.
Фиг.2. Связывание фаговых антител с вирусом эктромелии (темные столбики) и 3% раствором БСА в ФСБР (светлые столбики) в ИФА.
Фиг.3. Связывание фагового одноцепочечного антитела 1А с различными ортопоксвирусами при последовательных разведениях антитела. Обозначения: 1 - кривая связывания антитела 1А с вирусом эктромелии, 2 - связывания антитела 1А с вирусом осповакцины, 3 - связывания антитела 1А с вирусом оспы коров, 4 - связывания контрольного фага-помощника М13К07 с вирусом эктромелии, 5 - с вирусом осповакцины, 6 - с вирусом оспы коров.
Фиг.4. Нейтрализация инфекционности вирусов натуральной оспы, осповакцины и оспы коров фаговым одноцепочечным антителом 1А и фагом-помощником М13К07. Обозначения: 1 - нейтрализация инфекционности вируса осповакцины одноцепочечным антителом 1А; 2 - нейтрализация вируса оспы коров одноцепочечным антителом 1А; 3 - нейтрализация вируса натуральной оспы одноцепочечным антителом 1А; 4 - нейтрализация инфекционности вируса осповакцины фагом-помощником М13К07; 5 - нейтрализация вируса оспы коров фагом-помощником М13К07; 6 - нейтрализация вируса натуральной оспы фагом-помощником М13К07.
Фиг.5. Нуклеотидная и кодируемая ею аминокислотная последовательность фрагмента плазмиды pHEN-1A, кодирующая одноцепочечное антитело человека 1А в составе химерного белка с оболочечным белком р3 фага М13К07.
Фиг.6. Аминокислотная последовательность одноцепочечного антитела человека 1А.
Фиг.7. Общая схема структурной организации фагмиды pHEN-1A (физическая карта). 1А - ген, кодирующий одноцепочечное антитело человека 1А, p3 - ген, кодирующий оболочечным белок р3 фага М13К07; lac Z - промотор лактозного оперона Р, AMPr - ген устойчивости к ампициллину; amber - супрессируемый стоп кодон amber TAG; 6 His-tag - последовательность, кодирующая пептид HHHHHH, указаны некоторые сайты рестрикции.
Для лучшего понимания сущности предлагаемого изобретения оно иллюстрируется следующими примерами его осуществления.
Пример 1. Аффинное обогащение полученной библиотеки фаговыми антителами, специфичными к вирусу эктромелии.
Аликвоту клеточного варианта библиотеки одноцепочечных антител человека, содержащую около 1010 клонов (25 мкл), инокулируют в 50 мл среды 2×YT, содержащей 100 мкг/мл ампициллина и 1% глюкозу, и растят при постоянном перемешивании при 37°С до OD 600=0.5. После этого полученную культуру инфицируют фагом-помощником М13К07 (New England Biolabs, Великобритания), добавляя его в соотношении 1:20 (количество бактериальных клеток: количество фаговых частиц), и инкубируют 30 мин при 37°С. Затем 20 мл культуры осаждают центрифугированием в течение 10 мин при 3300 g. Осадок ресуспендируют в 10 мл 2×YT, содержащей 100 мкг/мл ампициллина и 25 мкг/мл канамицина, и переносят суспензию в 200 мл 2×YT, содержащей 100 мкг/мл ампициллина и 25 мкг/мл канамицина. Культуру инкубируют ночь при постоянном перемешивании при 30°С. На следующий день клетки Е.coli осаждают центрифугированием 10800 g, 10 мин в центрифуге "Beckman J2-21", к супернатанту добавляют 1/5 объема раствора PEG/NaCl (20% ПЭГ6000 и 2,5 М NaCl), хорошо перемешивают и в течение 1 ч инкубируют на льду. После этого суспензию центрифугируют при 10800 g 30 мин. Полученный осадок растворяют в 40 мл ФСБР рН 7,2 и добавляют 8 мл раствора PEG/NaCl, инкубируют во льду не менее 20 мин и проводят осаждение бактериофагов в центрифуге "Beckman J2-21" при 3300 g 30 минут. Супернатант вместе с остатками PEG/NaCl тщательно удаляют, а осадок ресуспендируют в 2 мл стерильного ФСБР. Для удаления остатков бактериальных клеток фаговую суспензию центрифугируют при 3300 g 10 мин. Выход фаговых частиц составляет приблизительно 1012-1013 бляшкообразующих единиц (БОЕ).
Аффинное обогащение библиотеки проводят в ходе двух последовательных раундов. Для проведения первого раунда селекции в лунках 96-луночного планшета («Медполимер», Россия) сорбируют вирус эктромели в концентрации 100 мкг/мл в ФСБР и инкубируют в течение ночи при комнатной температуре. На следующий день после удаления антигена лунки трижды промывают ФСБР и места неспецифического связывания насыщают 5% раствором сухого обезжиренного молока в буфере ФСБР в течение 2 ч при 37°С. Кроме того, 5% раствор обезжиренного молока сорбируют в дополнительные лунки в качестве отрицательного контроля. После этого лунки трижды промывают ФСБР и в каждую лунку добавляют 1011 БОЕ фаговой библиотеки в 100 мкл раствора ФСБР с 0,1% Твин-20. Планшеты инкубируют 1,5 часа при постоянном перемешивании при 37°С, после чего лунки планшета промывают 20 раз раствором ФСБР с 0,1% Твин-20, а затем 20 раз ФСБР.
Элюцию связавшихся бактериофагов проводят с помощью триэтиламина. Для этого в лунки добавляют по 100 мкл 100 мМ триэтиламина, инкубируют 2-3 минуты, непрерывно перемешивая, затем элюированные фаги переносят в 350 мкл 1М трис-HCl, рН 7,4.
Для получения фагового репертуара культуру клеток Е.coli TG1 в стадии экспоненциального роста (по 10 мл для каждого элюата) инфицируют полученным фаговым элюатом (по 400 мкл), инкубируют 30 мин при 37°С, затем центрифугируют 10 мин при 3000 g. Осадки ресуспендируют в 1 мл 2×YT и высевают на чашки с агаризованной средой 2×YT, содержащей 100 мкг/мл ампициллина и 1% глюкозу. Чашки инкубируют ночь при 30°С. Выросшие клетки собирают и хранят в среде 2×YT, содержащей 30% глицерин, при -70°С.
Амплификацию отобранных после первого раунда бактериофагов проводят как описано ранее.
Второй раунд аффинного обогащения проводят аналогично. При этом концентрацию вируса эктромелии уменьшают до 50 мкг/мл.
Степень обогащения библиотеки антителами, специфичеными к вирусу эктромелии, проверяют с использованием непрямого ИФА по способности популяций фаговых антител, полученных в результате каждого раунда аффинного обогащения, связывать вирус эктромелии (фиг.1). Для этого вирус эктромелии сорбируют на 96-луночные иммунологические планшеты («Медполимер», Россия) в количестве 300 нг на лунку. Места неспецифического связывания блокируют раствором 3%-ного БСА («Sigma») в фосфатном буфере, рН 7.2. Затем в лунки вносят фаговые антитела, выделенные, как описано ранее, разведенные ФСБР буфером, содержащим 0,1% Твин, и инкубируют 1 час при 37°С. После промывки в лунки вносят анти-М13 сыворотку мыши в разведении 1:16000, а затем антивидовой конъюгат щелочной фосфатазы («Sigma») в разведении 1:6000. В качестве хромогена используют паранитрофенилфосфат. В качестве отрицательного контроля используют связывание фаговых антител с обезжиренным молоком. Результаты приведены на фиг.1.
Пример 2. Отбор клонов, способных продуцировать антитела, связывающие вирус эктромелии.
Для получения моноклональных фаговых антител отдельные колонии E.coli TG1 из обогащенной против вируса эктромелии библиотеки пересевают с агаризованной среды в жидкую среду 2×YT, содержащую 100 мкг/мл ампициллина и 1%-ную глюкозу, и растят до плотности, равной 0,5 при OD 600. Затем клетки инфицируют фагом-помощником М13К07, осаждают центрифугированием 3300 g, 10 мин и ресуспендируют клеточный осадок в среде, содержащей 100 мкг/мл ампициллина и 50 мкг/мл канамицина. Клетки культивируют при 30°С при перемешивании в течение ночи и затем осаждают центрифугированием 10800 g, 10 мин.
Супернатант, содержащий фаговые антитела, исследуют на способность связываться с вирусом эктромелии методом ТИФА. Вирус осповакцины сорбируют в лунки 96-луночных иммунологических планшетов («Медполимер», Россия) в концентрации 3 мкг/мл. Места неспецифического связывания блокируют раствором 3%-ного БСА («Sigma») в фосфатном буфере, рН 7.2. Затем в лунки вносят супернатанты, разведенные ФСБР буфером, содержащим 0,1% Твин, в объемном соотношении 1:1 и инкубируют 1 час при 37°С. После промывки в лунки вносят анти-М13К07 сыворотку кролика в разведении 1:16000, а затем антивидовой конъюгат щелочной фосфатазы («Sigma») в разведении 1:6000. В качестве хромогена используют паранитрофенилфосфат. Контролем неспецифического связывания служит связывание вируса с фагом-помощником М13К07, не несущим фрагментов антител на своей поверхности. В качестве отрицательного контроля используют связывание фаговых антител с БСА. Результаты приведены на фиг.2.
В дальнейшем из супернатантов, продемонстрировавших сигнал, значение которого в 3 и более раз превышает контроль неспецифического связывания и отрицательный контроль, выделяют фаговые антитела с помощью преципитации раствором PEG/NaCl и последующего центрифугирования, как описано в Примере 1.
Пример 3. Иммуноферментный анализ связывания фагового антитела 1А с вирусами осповакцины, оспы коров и эктромелии.
Последовательные разведения вирусов осповакцины, оспы коров и эктромелии, начиная с 600 нг на лунку, с шагом 1:4 сорбируют в лунки 96-луночных иммунологических планшетов («Медполимер», Россия). Места неспецифического связывания блокируют раствором 3%-ного БСА («Sigma») в фосфатном буфере, рН 7.2. Затем в лунки вносят фаговые частицы, несущие на своей поверхности антитело 1А, в количестве 1011 БОЕ на лунку в ФСБР буфере, содержащем 0,1% Твин, и инкубируют 1 час при 37°С. После промывки в лунки вносят поликлональные анти-М13К07 антитела кролика в разведении 1:16000, а затем антивидовой конъюгат щелочной фосфатазы («Sigma») в разведении 1:6000. В качестве хромогена используют паранитрофенилфосфат. Результаты экспериментов приведены на фиг.3.
Пример 4. Исследование вируснейтрализующей активности фагового антитела 1А.
Антитела последовательно разводят в ФСБР буфере с шагом 1:4, начиная с концентрации 1013 фаговых частиц, и смешивают с равными объемами суспензии вирусов натуральной оспы, осповакцины и оспы коров, разведенных до примерно 300 БОЕ/мл. Смеси инкубируют при 37°С в течение 1 часа. Аликвоты из каждой смеси наносят на монослой клеток линии Vero E6 в 24-луночные планшеты при 37°С на 1 час, после чего смывают. Затем лунки покрывают средой MEM, содержащей 2% FBS и 0.24% агарозу. Жизнеспособные клетки окрашивают через 3-4 дня после инфицирования, визуализируя бляшки. В качестве отрицательного контроля используется бактериофаг-помощник М13К07, не несущий на своей поверхности одноцепочечного антитела. Титр нейтрализации определяют как разведение фаговых антител, при котором происходит 50%-ное уменьшение количества бляшек. Результаты приведены на фиг.4.
Из изложенного выше видно, что получены фагмидная ДНК pHEN-1A, содержащая ген одноцепочечного антитела человека 1А, обеспечивающий под контролем промотора лактозного оперона в клетках E.coli синтез одноцепочечного антитела человека 1А, экспонированного на поверхности нитчатого бактериофага М13, способного нейтрализовать вирусы натуральной оспы, вируса осповакцины и вируса оспы коров, включающий аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NO: 1. (фиг.5).
Источники информации
1. Fenner F., Wittek R., Dumbel K. The Orthopoxviruses. San Diego, New York, Berkeley, Boston, London, Sydney, Tokyo, Toronto: Academic Press, Inc. 1989.
2. Маренникова С.С., Щелкунов С.Н. Патогенные для человека ортопоксвирусы. - М.: КМК Scientific Press Ltd., 1998. - 386 с.
3. Галактионов В.Г. Иммунология. - М.: Изд-во "РИЦ МКД", 2000. - 488 с.
4. Lim A.P.C., Chan C.E.Z., Wong S.K.K., Chan A.H.Y., Ooi E.E. and Hanson B.J. Neutralizing human monoclonal antibody against H5N1 influenza HA selected from a Fab-phage display library // Virol J. - 2008. - V.5. - Р.130.
5. Clementi N., Mancini N., Solforosi L., Castelli M., Clementi M., Burioni R. Phage Display-based Strategies for Cloning and Optimization of Monoclonal Antibodies Directed against Human Pathogens // Int J Mol Sci. - 2012. - V. 13(7). - P. 8273-92.
6. Nimmagadda S.V., Aavula S.M., Biradhar N., Sula S., Lingala R., Chandran D., Villuppanoor S.A. Development of recombinant single-chain variable fragment against hepatitis A virus and its use in quantification of hepatitis A antigen // Biologicals. - 2012. - V. 40(4). - P.299-308.
7. Тикунова Н.В., Колокольцов А.А., Чепурнов А.А. Рекомбинантные моноклональные антитела человека против вируса Эбола. //ДАН, 2001, т. 378, № 4, с. 551-554.
8. Юн Т.Э., Тикунова Н.В., Шингарова Л.Н., Алиев Т.К., Болдырева Е.Ф., Морозова В.В., Швалов А.А., Некрасова О.В., Полыхалова И.В., Панина А.А., Ильичев А.А., Кирпичников М.П., Сандахчиев Л.С. Полноразмерное рекомбинантное антитело человека против вируса осповакцины. //Доклады Российской Академии Наук (ДАН), 2006, т. 407: №5, с. 702-705.
9. Тикунова Н.В., Шингарова Л.Н., Юн Т.Э., Морозова В.В., Алиев Т.К., Болдырева Е.Ф., Некрасова О.В., Полыхалова И.В., Панина А.А., Кирпичников М.П., Ильичев А.А. и Сандахчиев Л.С. Рекомбинантные плазмидные ДНК pCL37 и pCH37, кодирующие полипептиды со свойствами легкой и тяжелой цепей антитела человека против вируса осповакцины, обеспечивающие совместно биосинтез полноразмерных антител человека против вируса осповакцины в клетках млекопитающих, и рекомбинантное полноразмерное антитело человека класса IgG1, взаимодействующее с вирусом осповакцины. Патент РФ № 2317330. 2008.
10. Tikunova N., Dubrovskaya V., Morozova V., Yun T., Khlusevich Y., Bormotov N., Laman A., Brovko F., Shvalov A., Belanov E. The neutralizing human ecombinant antibodies to pathogenic Orthopoxviruses derived from a phage display immune library // Virus Research. - 2012. - V. 163. - P. 141-150.
11. McCausland M.M., Benhnia M.R., Crickard L., Laudenslager J., Granger S.W., Tahara T., Kubo R., Koriazova L., Kato S., Crotty S. Combination therapy of vaccinia virus infection with human anti-H3 and anti-B5 monoclonal antibodies in a small animal model // Antivir. Ther. - 2010. - V. 15(4). - P. 661-675.
12. Дубровская В.В., Тикунова Н.В., Морозова В.В., Бормотов Н.И., Ламан А.Г., Улитин А.Б., Бровко Ф.А., Беланов Е.Ф., Ильичев А.А. Комбинаторная фагмидная библиотека одноцепочечных антител человека, обогащенная антителами против вируса осповакцины, рекомбинантная фагмидная ДНК pHEN-2A8, содержащая уникальный ген одноцепочечного антитела человека, способного нейтрализовать вирус осповакцины и вирус оспы коров, и искусственное одноцепочечное антитело человека 2A8, способное нейтрализовать вирус осповакцины и вирус оспы коров. Патент РФ №2311927. 2005 г. (прототип).
13. Дубровская В.В., Улитин А.Б., Ламан А.Г., Гилева И.П., Бормотов Н.И., Ильичев А.А., Бровко Ф.А., Щелкунов С.Н., Беланов Е.Ф., Тикунова Н.В. Конструирование комбинаторной иммунной клонотеки одноцепочечных антител человека против ортопоксвирусов и селекция из нее антител к рекомбинантному белку prF30L вируса натуральной оспы // Молекулярная биология. - 2007. - Т.41, N1. - С. 173-185.
14. Hoogenboom H.R. and Winter G. By-passing immunization: Human antibodies from synthetic repertoires of germline VH gene segments rearranged in vitro// J. Mol. Biol. - 1992. - Vol.227. - P.381-388.
15. Birnboim H.C. and Dolly J. A rapid aikalin extraction procedure for screening recombinant plasmid DNA // Nucl. Acids Res. 1979. v.7. p.1513-1523.






Изобретение относится к иммунологии и медицине и представляет собой средство для нейтрализации вируса натуральной оспы, представляющее собой искусственное одноцепочечное антитело человека 1A, имеющее аминокислотную последовательность, приведенную в материалах заявки, экспонированное на поверхности нитчатого бактериофага М13. Изобретение позволяет нейтрализовать вирус натуральной оспы. 7 ил., 4 пр.
Средство для нейтрализации вируса натуральной оспы, представляющее собой искусственное одноцепочечное антитело человека 1A, имеющее аминокислотную последовательность, приведенную на фиг.6, экспонированное на поверхности нитчатого бактериофага М13.
| Катодный умножитель частоты | 1933 |
|
SU36489A1 |
| NINA TIKUNOVA, et al., "The neutralizing human recombinant antibodies to pathogenic Orthopoxviruses derived from a phage display immune library", Virus Research 163 (2012) , pp | |||
| Топливник с глухим подом | 1918 |
|
SU141A1 |
| КОМБИНАТОРНАЯ ФАГМИДНАЯ БИБЛИОТЕКА ОДНОЦЕПОЧЕЧНЫХ АНТИТЕЛ ЧЕЛОВЕКА, ОБОГАЩЕННАЯ АНТИТЕЛАМИ ПРОТИВ ВИРУСА ОСПОВАКЦИНЫ, РЕКОМБИНАНТНАЯ ФАГМИДНАЯ ДНК pHEN-2A8, СОДЕРЖАЩАЯ УНИКАЛЬНЫЙ ГЕН ОДНОЦЕПОЧЕЧНОГО АНТИТЕЛА ЧЕЛОВЕКА, СПОСОБНОГО НЕЙТРАЛИЗОВАТЬ ВИРУС ОСПОВАКЦИНЫ И ВИРУС ОСПЫ КОРОВ, И ИСКУССТВЕННОЕ ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО ЧЕЛОВЕКА 2A8, СПОСОБНОЕ НЕЙТРАЛИЗОВАТЬ ВИРУС ОСПОВАКЦИНЫ И ВИРУС ОСПЫ КОРОВ | 2005 |
|
RU2311927C2 |
| Edited by JOHN N | |||
| ABELSON "Combinatorial Chemistry" (Methods in Enzymology), volume 267, 493 | |||

