Группа изобретений относится к области биотехнологии и касается белка, обладающего пролонгированным действием (более длительным временем жизни в организме животных по сравнению с природным белком), на основе рекомбинантного интерферона альфа-2 человека и способа его получения путем культивирования дрожжей Saccharomyces cerevisiae.
Интерфероны альфа-2 человека (ИФН) относятся к классу цитокинов, к группе альфа-интерферонов, секретируемых практически всеми типами вирус-инфицированных клеток. Лекарственные формы ИФН являются препаратами широкого спектра действия, обладающими иммуномодулирующей, противовирусной и противоопухолевой активностью. В настоящее время лекарственные средства на основе ИФН являются наиболее востребованными противовирусными препаратами, входящими в «золотые лекарственные стандарты» для лечения различных вирусных заболеваний, в том числе вирусного гепатита В и вирусного гепатита С (ВГС), требующих пожизненной терапии.
Фармакокинетические характеристики рекомбинантного немодифицированного (стандартного) ИФН не позволяют поддерживать постоянным его уровень в плазме, что частично объясняет недостаточно высокий терапевтический эффект [Reddy, 2004].
Известны способы модификации белков, в том числе ИФН, увеличивающие их время жизни в плазме крови животных. Большинство известных модификаций направлено на увеличение физических размеров (гидродинамического объема) или заряда (отрицательного) молекул лекарственных белков, которое осуществляют с целью замедления процесса их фильтрации из плазмы крови сквозь поры почечных канальцев [Therapeutic Proteins, ed. R.Kontermann, 2012].
Химическая модификация ИФН с помощью полиэтиленгликолей (ПЭГ) позволила получить молекулы ПЭГ-ИФН, для которых характерны увеличенное время жизни в плазме крови и улучшенный фармакокинетический профиль [Zeuzem et al., 2000; Heathcote et al., 2000; Lindsay et al., 2001]. Масштабные клинические испытания продемонстрировали, что эффективность комбинированных препаратов на основе ПЭГ-ИФН в 1,5-2 раза превышает эффективность стандартного интерферона. При этом уровень излечивания ВГС достигает значений 54-56% (табл.1), и одновременно улучшается качество жизни пациентов [Fried et al., 2002; Manns et al., 2001; Hadziyannis, 2004; Clark & Nelson, 2009].
Однако высокая стоимость ПЭГ-ИФН в значительной степени ограничивает их массовое применение. Другими неблагоприятными свойствами являются неоднородность химически модифицированных молекул ПЭГ-ИФН, необходимость значительно (до 10 раз) увеличивать дозировку препаратов на их основе [табл.1 и Pasut, 2009], а также накопление в организме производных ПЭГ, не подверженных биодеградации. Кроме того, около половины больных ВГС остаются невосприимчивы к курсу лечения ПЭГ-ИФН [Pasut, 2009].
Для получения пролонгированных производных ИФН также использованы следующие способы его биомодификации:
• генетическая гибридизация ИФН с долгоживущими белками крови человека, например сывороточным альбумином [Bain et al., 2006; Subramanian et al., 2007; Nelson et al., 2009a, 2009b; Zeuzem 2009, 2009], или фрагментом иммуноглобулина [Jones et al., 2004], или с фрагментом антитела, обеспечивающего нековалентное связывание с сывороточным альбумином человека непосредственно в кровотоке [Walker et al., 2010]. Однако препарат Albinterferon-alfa-2b (Human Genome Sciences Inc и Novartis AG), предложенный для лечения хронического гепатита С, не получил одобрения контролирующих организаций в США (FDA) и ЕС (ЕМА) [http://seekingalpha.com/article/258673-human-genome-sciences-evenly-poised-after-fda-approval];
• генетическая гибридизация ИФН с искусственными полипептидами, в которых содержатся сайты N-гликозилирования и преобладают повторы из остатков глицина, аспарагиновой кислоты и глутаминовой кислоты [US 20090298762];
• генетическая гибридизация ИФН с полианионным полипептидом из остатков глутаминовой кислоты (84 и 173 остатка) в клетках Е.coli [US 20020169125, AU 2002252429, WO 02077036]. Белок не секретируется и требует продолжительной и трудоемкой очистки как от других внутриклеточных белков, так и от клеточных эндотоксинов;
• генетическая гибридизация ИФН с С-концевым пептидом (СТР) β-субъединицы хорионического гонадотропина человека, содержащего 28 аминокислотных остатков, формирующих четыре сайта О-гликозилирования [US 20120015437]. Одна или несколько копий СТР, присоединяемых к С-концу молекул, существенно удлиняют время жизни терапевтических белков в плазме крови [US 2012035101, US 2012015437, US 2012004286, US 2011286967, US 2010317585, US 2010081614, US 5759818]. Использование СТР-технологии ограничено необходимостью получать модифицированные белки исключительно в клетках животных, обеспечивающих корректное О-гликозилирование;
• генетическое изменение природной последовательности ИФН с целью конструирования сайтов N-гликозилирования. Модифицированный таким образом ИФН, продуцируемый в культуре клеток СНО, содержит четыре или пять N-углеводных цепочек и обладает в 25 раз увеличенным временем жизни в кровотоке [US 2012134960]. Для него характерно увеличение противоопухолевой активности, но снижение специфической противовирусной активности in vitro. Такой способ получении пролонгирования, основанный на изменении природной последовательности ИФН, чреват нарушением иммунного статуса пациента, а его получение основано на использовании дорогостоящих клеточных культур.
Практическое воплощение вышеуказанных способов в большинстве случаев не позволяет оптимизировать процесс получения пролонгированного ИФН путем использования более продуктивных и/или более дешевых систем экспрессии, поскольку ограничено выбором клеток-хозяев, подходящих для биосинтеза модифицированного белка.
Таких ограничений лишен способ пролонгирования с использованием полипептидной технологии, которая предполагает генетическую гибридизацию (коньюгацию) белков с высокорастворимыми, химически и иммунологически инертными неструктурированными полипептидами (НП), способными в физиологических условиях имитировать поведение молекул ПЭГ. Такие НП в водных растворах при физиологических солевых, температурных и буферных условиях устойчивы к фолдингу (формированию третичной структуры), агрегации, неспецифической адсорбции и поддерживают стабильное неупорядоченное состояние типа “клубок”, даже будучи генетически коньюгированными с обычными структурированными белками. Постоянное изменение конформации НП, вызываемое термическим движением, приводит к более или менее постоянной усредненной ватообразной структуре, молекулярный размер которой зависит только от числа аминокислотных остатков в составе НП, их собственного размера (диаметра) и конформационной гибкости. В этой связи НП способны опосредовать значительное увеличение гидродинамического объема коньюгированных белков, вызывающее ослабление почечной фильтрации и удлиняющее время жизни белков в плазме крови [Binder & Skerra, 2012].
Использование НП для пролонгирования белков имеет преимущества перед химической модификацией с помощью ПЭГ, в частности:
• белки, модифицированные по этой технологии, могут быть экспрессированы и внутриклеточно, и в секретированом виде в различных типах клеток, включая бактерии и дрожжи, что позволяет значительно удешевить и ускорить их получение и очистку;
• модифицирующие НП обладают крайне низкой иммуногенностью и подобно ПЭГ способны экранировать чужеродные белки от распознавания иммунной системой;
• модифицирующие НП являются биоразлагаемыми, что значительно снижает риск появления токсичности или накопления в каких-либо органах (например, в почках или печени) или клетках (например, в макрофагах);
• генетически кодируемая последовательность НП гарантирует неизменность и точность модификации белка;
• поскольку внутренние последовательности НП, как правило, не взаимодействуют ни между собой, ни с последовательностями белка, с которым коньюгированы НП, модифицированные белки сохраняют специфическую активность на уровне немодифицированных аналогов;
• подбор размера пролонгирующего НП позволяет конструировать белки, обладающие медицински обоснованным уровнем пролонгирования.
В основных чертах эти характерные особенности свойственны трем известным типам НП, способным пролонгировать действие белков до 100 раз:
1) XTEN-полипептидам, содержащим случайное чередование 6 аминокислотных остатков аланина, глутаминовой кислоты, глицина, пролина, серина и треонина [ЕР 2402754, US 20110312881, WO 2011123830, WO 2011123813, US 20110151433, US 20100323956, WO 2010144502, WO 2010091122, СА 2748314, WO 2007103515, US 7846445, US 7855279; Geething et al., 2010; Schellenberger et al; 2009];
2) PAS-полипептидам, содержащим остатки пролина, аланина и серина [WO 2011144756, NZ 580670, US 20100292130, ЕР 2173890, WO 2008155134, Skerra, 2010];
3) НАР (или ОЯ8)-полипептидам, являющимися глицин-богатыми последовательностями [Schlapschy et al., 2007; WO 2007103515], содержащими остатки глицина (G) и серина (S).
Показано пролонгирование действия ИФН за счет использования НП PAS, наименьший размер которых составлял около 200 аминокислотных остатков [Skerra, 2010; ЕА 201000040].
У НАР-полипептидов наличие пролонгирующей активности в отношении ИФН не установлено, более того известен пример, доказывающий отсутствие пролонгирующей активности у НП НАР размером менее 200 аминокислотных остатков в отношении фрагмента одного из антител [Schlapschy et al., 2007].
В качестве ближайшего аналога заявляемого белка рассмотрим гибридный белок, обладающий пролонгированным действием в организме животных и представляющий собой гибридный белок PAS-ИФН, включающий в свой состав, начиная с N-конца (1) аффинную метку “Strep-tag II”, облегчающую очистку гибридного белка с использованием аффинной хроматографии со стрептавидином, (2) НП PAS, состоящий из 200 остатков пролина, аланина и серина, и (3) последовательность интерферона альфа-2b, имеющего массу 20,9 кДа, которая отличается от расчетной массы нативного интерферона альфа-2b человека - 19,3 кДа [ЕА 201000040].
Способ получения белка-ближайшего аналога осуществляют в клетках бактерий E.coli путем микробиологического синтеза с секрецией в периплазму и последующим осторожным отделением фракции периплазматических белков от белков цитоплазмы клеток, аффинной очистки целевого белка с использованием стрептавидина и доочистку его от клеточных эндотоксинов. При этом выход белка-аналога после аффинной очистки не превышал 0,1 мг/л/ОД.
Полученный белок-ближайший аналог демонстрировал специфическую активность на уровне немодифицированного ИФН и увеличенное время жизни в организмах крыс, подобно вариантам ПЭГ-ИФН.
Способ получения белка-ближайшего аналога путем культивирования бактерий E.coli в отличие от заявляемого не позволяет получать секретированный во внеклеточную среду целевой белок, а уровень продукции белка-аналога почти в 30 раз ниже уровня продукции заявляемого белка по заявляемому способу. При этом фактически выход белка-ближайшего аналога даже завышен, поскольку примененный для его очистки метод аффинной хроматографии со стрептавидином требует доочистки целевого белка от бактериальных эндотоксинов и, следовательно, приводит к дополнительным потерям.
Известно, что одной из главных оценок эффективности препаратов ИФН служит уровень излечивания (SVR) больных ВГС, который коррелирует с величиной разовой дозы, обусловленной специфической активностью ИФН, и «временем полувыведения» препаратов ИФН из организма [Pasut, 2009]. Анализ данных [Pasut, 2009], характеризующих препараты ПЭГ-ИФН в сравнении со стандартным ИФН (табл.1), позволяет полагать, что значительным терапевтическим потенциалом будет обладать белковый препарат, для которого характерно физиологически умеренное 3-4-кратное пролонгирование и высокая на уровне стандартного ИФН специфическая активность. Преимуществами такого препарата также могут быть облегченный по сравнению со стандартным ИФН режим приема, например 2 раза вместо 3 раз в неделю, при сохранении себестоимости на уровне стандартного ИФН.
Задача заявляемой группы изобретений - расширить арсенал белков на основе рекомбинантного интерферона альфа-2 человека, обладающих пролонгированным действием, и разработать способ их получения.
Задача решена путем разработки:
- гибридного белка GFN80, обладающего пролонгированным действием и представляющего собой рекомбинантный интерферон альфа-2 человека, слитый с N-конца с аминокислотной последовательностью полипептида S(G4S)16, где S - остаток серина, a G - остаток глицина;
- гибридного белка GFN100, обладающего пролонгированным действием и представляющего собой рекомбинантный интерферон альфа-2 человека, слитый с N-конца с аминокислотной последовательностью полипептида S(G4S)20, где S - остаток серина, а G - остаток глицина;
- способа получения гибридного белка GFN80 или GFN100 путем культивирования в подходящих условиях дрожжей Saccharomyces cerevisiae, трансформированных вектором экспрессии, который содержит область инициации репликации эндогенной 2-мкм плазмиды дрожжей Saccharomyces cerevisiae, а также промотор GAL1 дрожжей, контролирующий экспрессию гена, включающего последовательность ДНК SEQ ID NO:1 или SEQ ID NO:2, соответственно, с последующим выделением гибридного белка из культуральной жидкости, а также конструирования:
- штамма Saccharomyces cerevisiae ВКПМ Y-3927 - продуцента гибридного белка GFN80, полученного путем введения нуклеотидной последовательности SEQ ID NO:1 в составе плазмиды р71-81 в клетки реципиента, являющегося бесплазмидным производным штамма Saccharomyces cerevisiae ВКПМ Y-3550 [RU 2427645];
- штамма Saccharomyces cerevisiae ВКПМ Y-3928 - продуцента гибридного белка GFN100, полученного путем введения нуклеотидной последовательности SEQ ID NO:2 в составе плазмиды р71-82 в клетки реципиента, являющегося бесплазмидным производным штамма Saccharomyces cerevisiae ВКПМ Y-3550.
Этапы конструирования штаммов-продуцентов заявляемых гибридных белков
Этап 1. Синтезируют фрагмент ДНК, имеющий открытые «липкие» концы, идентичные «липким» концам, создаваемым рестриктазами BglII и BamHI, и кодирующий полипептид S(G4S)4, где S - остаток серина, a G - остаток глицина. Направленно клонируют этот синтетический фрагмент ДНК в подходящем лабораторном векторе, расщепленном по уникальным сайтам узнавания рестриктаз BglII и BamHI.
Этап 2. Используя рутинные методы генетической инженерии, осуществляют мультимеризацию оригинального синтетического фрагмента ДНК, клонированного на этапе 1, и получают плазмиды, содержащие фрагменты ДНК, кодирующие полипептиды S(G4S)16 и S(G4S)20, где S - остаток серина, а G - остаток глицина.
Этап 3. Конструируют вспомогательные плазмиды и целевые экспрессионные плазмиды р71-81 и р71-82, в составе которых осуществляют корректное слияние последовательностей ДНК, кодирующих промоторную область гена GAL1 дрожжей, лидерную пре-про область α-фактора дрожжей и аминокислотную последовательность S(G4S)16 или S(G4S)20, соответственно, и структурный ген зрелого интерферона альфа-2b человека.
Этап 4. Получают штамм-реципиент S. cerevisiae SCR8G1. С этой целью клетки музейного штамма S. cerevisiae ВКПМ Y-3550, содержащего рекомбинантную плазмиду, освобождают от содержащейся в них рекомбинантной плазмиды. Клоны штамма S. cerevisiae SCR8G1 селектируют по признаку отсутствия роста на агаризованной среде с глюкозой 20 мг/мл, детерминированного отсутствием в клетках штамма S. cerevisiae SCR8G1 рекомбинантной плазмиды, несущей функциональный аллель гена PGK1.
Этап 5. Получают заявляемые штаммы S. cerevisiae ВКПМ Y-3927 и S. cerevisiae ВКПМ Y-3928. С этой целью клетки штамма-реципиента S. cerevisiae SCR8G1 трансформируют соответственно рекомбинантными плазмидами р71-81 или р71-82, содержащими функциональный аллель гена PGK1. Трансформанты отбирают по способности расти при температуре 30°C на агаризованной среде с глюкозой 20 мг/мл.
Подтверждают способность полученных трансформантов секретировать целевые гибридные белки и депонируют их во Всероссийской коллекции промышленных микроорганизмов (ВКПМ).
В результате получают заявляемые штаммы:
- SCR-8G1/p71-81 (лаб.название GFN80-Sc:, Saccharomyces cerevisiae ВКПМ Y-3927)
- SCR-8G1/p71-82 (лаб.название GFN100-Sc:, Saccharomyces cerevisiae ВКПМ Y-3928)
Характеристика заявляемых штаммов-продуцентов гибридных белков
Описание штаммов-продуцентов
Заявляемые штаммы ВКПМ Y-3927 и ВКПМ Y-3928 содержат гены целевых гибридных белков GFN80 или GFN100 в составе рекомбинантных эписомных плазмид р71-81 или р71-82, соответственно, каждая из которых обеспечивает экспрессию гена целевого гибридного белка под контролем промотора дрожжей GAL1 и имеет следующие свойства:
- плазмида содержит: ориджин репликации эндогенной 2-мкм плазмиды дрожжей, т.е. способна поддерживаться в клетках дрожжей в эписомном многокопийном состоянии; гены оротидин 5-фосфат декарбоксилазы (URA3) и 3-фосфоглицерат киназы (PGK1) дрожжей, способные служить селективными маркерами, последовательность бактериальной плазмиды pUC19, а также экспрессионную кассету с геном гибридного белка GFN80 или GFN100;
- в состав экспрессионной кассеты входит последовательность ДНК промоторной области гена дрожжей GAL1, последовательность ДНК структурного гена MFα1, кодирующая пре-про лидерный пептид предшественника полового феромона (α-фактора) дрожжей, последовательность ДНК структурного гена целевого гибридного белка GFN80 или GFN100 и последовательность ДНК области терминации транскрипции гена CYC1 дрожжей. Секрецию целевого гибридного белка в клетках штамма-продуцента направляет N-концевая лидерная последовательность α-фактора дрожжей, удаляемая из состава целевого гибридного белка в процессе внутриклеточного транспорта. Сочетание сильного промотора и высокой дозы гена в составе многокопийной плазмиды в совокупности обеспечивают высокий уровень секреции целевых гибридных белков GFN80 или GFN100 в клетках заявляемых штаммов.
Заявляемые штаммы-продуценты S. cerevisiae ВКПМ Y-3927 и ВКПМ Y-3928 являются диплоидными прототрофными штаммами. Диплоидный статус клеток этих штаммов обеспечивает повышенную стабильность их характеристик. В геноме заявляемых штаммов присутствует гомозиготная мутация в гене 3-фосфоглицерат киназы, обеспечивающая генотип pgk1/pgk1. Эта мутация ограничивает способность бесплазмидных клеток заявляемых штаммов к росту на любых средах, содержащих любой единственный источник углерода, усваиваемый дрожжами S.cerevisiae. Присутствие функционального аллеля гена 3-фосфоглицерат киназы (PGK1) в составе эписомной экспрессионной плазмиды р71-81 или р71-82 обеспечивает селективный рост содержащих плазмиду клеток заявляемых штаммов в этих условиях и селективное поддержание плазмид в растущих клетках.
Клетки заявляемых штаммов содержат гомозиготную мутацию в гене GAL80 - генотип gal80::LEU2/gal80::LEU2. Мутация gal80 отвечает за инактивацию белка, репрессирующего промотор GAL1, что приводит к изменению регуляции и позволяет осуществлять галактозо-независимую активацию промотора GAL1 контролирующего экспрессию целевых гибридных белков.
Клетки заявляемых штаммов содержат в геноме ген протеиназы Кех2 дрожжей S.cerevisiae, секретируемой во внеклеточную среду, под контролем промотора GAL1 дрожжей, что способствует увеличению продукции целевых гибридных белков.
Клетки заявляемых штаммов содержат в геноме гомозиготную мутацию в гене YPS1 - генотип yps1::TRP1/ypsfile://l:/TRPl, приводящую к инактивации протеиназы Yps1, отвечающей за деградацию интерферона и, предположительно, интерферонсодержащих гибридных целевых белков [RU 2427645].
В связи с тем, что в клетках заявляемых штаммов синтез целевых гибридных белков осуществляется под контролем промотора GAL1, который репрессируется глюкозой, индукция секреции целевых гибридных белков в культуральную среду происходит при снижении концентрации сахара в среде до уровня менее 2 мг/мл.
Генотипические признаки Saccharomyces cerevisiae ВКПМ Y-3927:
а/α leu2/leu2 ura3/ura3 trp1/trp1 pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 yps1::TRP1/yps1v::TRP1 ATH1/ath1::(ADH1-G418, TDH3-KEX2A) STA2/STA2 suc0/SUC2)/р71-81
Клетки штамма ВКПМ Y-3927 содержат эписомную плазмиду р71-81. Saccharomyces cerevisiae ВКПМ Y-3928:
а/α leu2/leu2 ura3/ura3 trp1/trp1 pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 yps1::TRP1/yps1v::TRP1 ATH1/ath1::(ADH1-G418, TDH3-KEX2A) STA2/STA2 suc0/SUC2)/р71-82
Клетки штамма ВКПМ Y-3928 содержат эписомную плазмиду р71-82.
Культурально-морфологические признаки
При культивировании при температуре 28°C в течение 48-96 часов на агаризованных питательных средах заявляемые штаммы образуют колонии. При этом клетки заявляемых штаммов имеют округлую или овальную форму, 3-7 мкм в диаметре, почкуются. Почкование истинное, многостороннее, истинного мицелия не образуют.
Колонии имеют следующий вид:
1) на агаризованной среде YPD (состав в мас.%: бактопептон - 2, дрожжевой экстракт - 1, бактоагар - 2, глюкоза - 2, вода - остальное) - колонии белого цвета, непрозрачные, с ровным краем, матовой поверхностью, линзовидным профилем и сметанообразной консистенцией;
2) на агаризованной среде с крахмалом (состав в мас.%: пептон - 2, дрожжевой экстракт - 1, крахмал - 1, агар - 2, вода - остальное) - колонии белого цвета с узорчатым краем, матовой поверхностью, линзовидным профилем и крупитчатой консистенцией.
При росте в жидкой среде при температуре 28°C в течение первых 24 ч культивирования - жидкость мутная, осадок белый, не комкуется, пристеночных пленок не образует.
Физико-биохимические признаки
Заявляемые штаммы прототрофные, факультативные анаэробы, мезофилы. Рост возможен в интервале температур 10-37°С, оптимальная для роста температура 29±1°C, однако устойчивы к повышенной температуре, способны к росту при 37°C. Способны к росту при значениях рН 3,8-7,4 (оптимальный диапазон рН 4,7-5,5).
Ассимиляция источников углерода
Заявляемые штаммы сбраживают глюкозу, фруктозу, мальтозу, сахарозу, декстрины, крахмал. Не сбраживают лактозу, галактозу, инулин, ксилозу, арабинозу. Штаммы способны к росту в среде с крахмалом.
Ассимиляция источников азота
В качестве источников азота используют пептон, дрожжевой экстракт, аминокислоты, сернокислый аммоний, азотнокислый аммоний.
Синтез целевого белка
Заявляемые штаммы синтезируют гибридные целевые белки, в состав которых входит аминокислотная последовательность нативного интерферона альфа-2b человека. Полная активация синтеза целевых гибридных белков происходит при снижении концентрации сбраживаемого источника углерода в среде культивирования до неферментируемого уровня или при росте штаммов в средах, содержащих исключительно неферментируемые источники углерода. В этих условиях клетки штамма секретируют целевые гибридные белки в среду культивирования.
Условия и состав сред для длительного хранения штаммов
Суспензия клеток заявляемых штаммов хранится в 15%-ном растворе глицерина при минус 70°C.
Способ получения гибридных белков в общем виде
Для засева ферментера получают посевную культуру путем культивирования штамма-продуцента целевого гибридного белка в среде следующего состава, в мас.%: пептон 1-3; дрожжевой экстракт 0,5-3; глюкоза 1-3, вода - остальное, рН среды - естественный в течение 16-40 часов при температуре 22-32°C на орбитальной качалке 100-350 об/мин.
Посевной культурой засевают ферментер (0,5-1000 л), содержащий ферментационную среду следующего состава, в мас.%: пептон 1-3; дрожжевой экстракт 0,5-3, глюкоза или сахароза 1-4, вода - остальное, рН среды - естественный. Количество посевной культуры, вносимой в ферментер составляет 3-15% от объема ферментационной среды.
Ферментацию проводят при 22-32°C, аэрации 0,5-1000 л/мин и скорости перемешивания культуры 100-1500 об/мин. Через 16-30 ч после засева ферментера процесс проводят без подпитки или начинают подпитку 30-60% раствором глюкозы или сахарозы со скоростью 2-10 г/л/ч и устанавливают pH-статирование культуры в диапазоне рН 5,0-7,2. Общая продолжительность ферментации составляет 38-68 ч.
Продукция секретируемого целевого гибридного белка в таких условиях составляет не менее 0,12 г/л.
Очистку гибридного белка из среды культивирования дрожжей производят как описано [RU 2427645].
В результате получают не менее 0,05 г целевого гибридного белка из 1 л культуральной жидкости.
Пример 1. Конструирование рекомбинантной плазмиды pGS-4.
Синтезируют фрагмент ДНК, имеющий открытые «липкие» концы, идентичные «липким» концам, создаваемым рестриктазами BglII и BamHI, и кодирующий полипептид S(G4S)4, где S - остаток серина, a G - остаток глицина.
SGGGGSGGGGSGGGGSGGGGS
5'-GATCTGGTGGTGGTGGTTCTGGTGGTGGTGGTTCCGGAGGTGGTGGTTCTGGTGGTGGTG
ACCACCACCACCAAGACCACCACCACCAAGGCCTCCACCACCAAGACCACCACCACCTAG-5'
Рекомбинантную плазмиду pGS-4 конструируют на основе любого подходящего лабораторного вектора, например на основе модифицированного вектора pUC19Bgl, в котором сайт узнавания рестриктазы EcoRI замещен на сайт узнавания рестриктазы BglI. С этой целью синтетический фрагмент ДНК клонируют в выбранном векторе, расщепленном по сайтам BglII и BamHI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены.
В результате получают рекомбинантную плазмиду pGS-4, содержащую клонированный синтетический фрагмент ДНК.
Пример 2. Конструирование рекомбинантной плазмиды pGS-16.
Стадия 1. Рекомбинантную плазмиду pGS-4 расщепляют с использованием рестриктаз AatII и BamHI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-1.
Стадия 2. Рекомбинантную плазмиду pGS-4 расщепляют с использованием рестриктаз AatII и BglII и образовавшиеся фрагменты рестрикции разделяют в геле. Больший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-2.
Фрагменты 1 и 2, полученные на стадиях 1 и 2, соответственно, лигируют между собой и полученную плазмиду называют pGS-8.
Повторяют стадии 1 и 2 за исключением того, что на каждой стадии используют плазмиду pGS-8. В результате получают плазмиду pGS-16. Плазмида pGS-16 содержит BglII/BamHI фрагмент ДНК, кодирующий полипептид S(G4S)16, где S - остаток серина, а G - остаток глицина.
Пример 3. Конструирование рекомбинантной плазмиды pGS-20.
AatII/BamHI фрагмент-1 ДНК рекомбинантной плазмиды pGS-4, полученный как описано на стадии 1 примера 2, лигируют с AatII/BglII фрагментом-2 ДНК плазмиды pGS-16, полученным как описано на стадии 2 примера 2.
В результате получают плазмиду pGS-20. Плазмида pGS-20 содержит BglII/BamHI фрагмент ДНК, кодирующий полипептид S(G4S)20, где S - остаток серина, a G - остаток глицина.
Пример 4. Конструирование рекомбинантной плазмиды pUC18x-(Ser)IFN.
Для конструирования рекомбинантной плазмиды pUC18x-(Ser)IFN осуществляют лигирование двух фрагментов ДНК: 1) BamHI/XhoI фрагмента, заключающего векторную часть лабораторной плазмиды pUC18x, содержащей в составе модифицированного полилинкера сайт XhoI вместо сайта EcoRI; 2) BamHI/XhoI фрагмента ДНК, который получают в результате ПЦР-амплификации фрагмента ДНК плазмиды pUC18x-GAL1ppI-IFN2b [RU 2427645] с использованием праймеров N499 (5'-ataggatcctgtgatctgcctcaaacccac) и N169 (5-gagcggataacaatttcacacagg) и последующего открытия его липких концов с использованием рестриктаз BamHI и XhoI.
Плазмида pUC18x-(Ser)IFN заключает нуклеотидную последовательность структурного гена интерферона альфа-2b человека в составе BamHI/XhoI фрагмента ДНК.
Пример 5. Конструирование рекомбинантной плазмиды pUC18x-GAL1ppI-(Ser).
Плазмиду pUC18x-GAL1ppI-(Ser) конструируют путем замещения NcoI/XhoI фрагмента ДНК плазмиды pUC18x-GAL1ppI-IFN2b [RU 2427645] на двуцепочечный синтетический фрагмент ДНК, полученный в результате отжига соответствующих олигонуклеотидов:
5′-catggaaaaaagatctc
cttttttctagagagct-5′
В результате клонирования получают плазмиду pUC18x-GAL1ppI-(Ser), в составе которой в последовательности ДНК, слитой с промоторной областью гена GAL1 дрожжей и кодирующей лидерную пре-про область α-фактора дрожжей, сконструирован сайт узнавания рестриктазы BglII, формируемый кодоном аргинина, входящим в сайт узнавания протеиназы Кех2, и кодоном серина, следующим непосредственно за кодоном аргинина.
Пример 6. Конструирование рекомбинантной плазмиды р71-81.
Используют плазмиду pPDX3-IFN2b, отличающуюся от плазмиды pPDX2-IFN2b [RU 2427645] одиночной нуклеотидной заменой G на А в положении +211 относительно стартового ко дона ATG в последовательности ДНК структурного гена URA3 дрожжей [в кодоне 71 (GAG→AAG)], приводящей к инактивации сайта узнавания рестриктазы NcoI.
Конструирование производят путем одновременного направленного лигирования 4-х фрагментов ДНК, которыми являются:
HindIII/BglII фрагмент ДНК плазмиды pUC18x-GAL1ppI-(Ser), заключающий промоторную область гена GAL1 дрожжей, слитый с последовательностью ДНК, кодирующей лидерную пре-про область α-фактора дрожжей;
BglII/BamHI фрагмент ДНК плазмиды pGS-16, кодирующий полипептид S(G4S)16, где S - остаток серина, a G - остаток глицина;
BamHI/XhoI фрагмента ДНК плазмиды pUC18x-(Ser)IFN, заключающий нуклеотидную последовательность структурного гена интерферона альфа-2b человека;
XhoI/HindIII фрагмент ДНК, заключающий векторную часть плазмиды pPDX3-IFN2b.
В результате получают рекомбинантную плазмиду р71-81 размером 9352 п.о., в составе которой корректно слиты генетические элементы, кодирующие лидерную пре-про область α-фактора дрожжей, аминокислотную последовательность S(G4S)16 и последовательность нативного интерферона альфа-2b человека.
Пример 7. Конструирование рекомбинантной плазмиды р71-82.
Рекомбинантную плазмиду р71-82 конструируют как описано в примере 6, но вместо фрагмента ДНК плазмиды pGS-16 для лигирования используют BglII/BamHI фрагмент ДНК плазмиды pGS-20, кодирующий полипептид S(G4S)20, где S - остаток серина, a G - остаток глицина.
В результате получают рекомбинантную плазмиду р71 - 82 размером 9412 п.о., в составе которой корректно слиты генетические элементы, кодирующие лидерную пре-про область α-фактора дрожжей, аминокислотную последовательность S(G4S)20 и последовательность нативного интерферона альфа-2b человека.
Пример 8. Получение штамма-реципиента Saccharomyces cerevisiae SCR8G1.
Штамм-реципиент является бесплазмидным производным содержащего плазмиду штамма Saccharomyces cerevisiae ВКПМ Y-3550.
Для получения штамма-реципиента клетки штамма S. cerevisiae ВКПМ Y-3550 освобождают от содержащейся в них плазмиды путем культивирования в течение 4 суток при температуре 30°C на агаризованной среде YPGE следующего состава, в мас.%: бактопептон - 2, дрожжевой экстракт - 1, бактоагар - 2, этанол - 2, глицерин - 3, вода - остальное. На этой среде способностью к росту обладают и клетки штамма ВКПМ Y-3550, несущие плазмиду, в составе которой находится ген PGK1, и клетки штамма-реципиента SCR8G1, утратившие эту плазмиду. Затем смесь выросших клеток клонируют.
Утратившие плазмиду клоны селектируют по признаку отсутствия роста на агаризованной среде YPD следующего состава, в мас.%: бактопептон - 2, дрожжевой экстракт - 1, бактоагар - 2, глюкоза - 2, вода - остальное. В результате получают штамм-реципиент S. cerevisiae SCR8G1, который отличается от штамма S. cerevisiae ВКПМ Y-3550 отсутствием способности к росту на среде с глюкозой 20 мг/мл, детерминированной отсутствием в клетках штамма SCR8G1 плазмиды, несущей функциональный аллель гена PGK1.
Пример 9. Получение штамма ВКПМ Y-3927.
Штамм S. cerevisiae ВКПМ Y-3927 - продуцент белка GFN80 получают трансформацией клеток штамма-реципиента S. cerevisiae SCR8G1 рекомбинантной плазмидой р71-81.
Для подготовки к трансформации клетки исходного штамма S. cerevisiae SCR8G1 культивируют в течение 18-24 часов при температуре 28°C на агаризованной среде YPGE. Трансформацию выросших клеток плазмидой р71-82 проводят по методу Ito [Ito et al., 1983] за исключением того, что перед высевом на селективную среду трансформированные клетки инкубируют в течение 3 часов при температуре 30°C на шейкере с орбитальной скоростью 250 об/мин в жидкой среде YPGE. Трансформанты отбирают по способности расти при температуре 30°C на агаризованной среде YPD.
Оценивают уровень экспрессии белка GFN80 клетками полученных трансформантов. Для этого 3 независимо полученных клона трансформантов выращивают в течение 46 часов при температуре 30°C на шейкере с орбитальной скоростью 250 об/мин в жидкой среде YPD до стационарной фазы роста (до оптической плотности 45±5 опт. ед.). Уровень накопления заявляемого белка определяют в освобожденной от клеток среде культивирования. Отбирают клон, продуцирующий белок, имеющий подвижность в условиях электрофореза на уровне 25 килодальтон.
В результате получают штамм S. cerevisiae ВКПМ Y-3927 - продуцент белка GFN80.
Штамм-продуцент S.cerevisiae ВКПМ Y-3927 отличается от штамма-реципиента S. cerevisiae SCR8G1 способностью к росту на среде с глюкозой 20 мг/мл, а от штамма S. cerevisiae ВКПМ Y-3550 способностью к продукции белка GFN80, детерминированной введенной в состав штамма плазмидой р71-81.
Пример 10. Получение штамма ВКПМ Y-3928
Штамм S. cerevisiae ВКПМ Y-3928 - продуцент белка GFN100 получают, как описано в примере 9, за исключением того, что для трансформации используют рекомбинантную плазмиду р71-82.
Отбирают клон, продуцирующий белок GFN100, имеющий подвижность в условиях электрофореза на уровне 31 килодальтон.
В результате получают штамм S. cerevisiae ВКПМ Y-3928 - продуцент белка GFN100.
Штамм-продуцент S.cerevisiae ВКПМ Y-3928 отличается от штамма-реципиента S. cerevisiae SCR8G1 способностью к росту на среде с глюкозой 20 мг/мл, а от музейного штамма ВКПМ Y-3550 - способностью к продукции белка GFN100, детерминированной, введенной в состав штамма плазмидой р71-82.
Пример 11. Микробиологический синтез белка GFN8Q с использованием штамма ВКПМ Y-3927.
Для получения посевного материала штамм S. cerevisiae ВКПМ Y-3927 выращивают в среде YPD на качалке (250 об/мин) при температуре 28°C в течение 20-24 ч.
50 мл посевного материала используют для засева 3 л ферментера Anglicon, содержащего 950 мл среды 223 следующего состава, в мас.%: бактопептон - 2, дрожжевой экстракт - 2, сахароза - 3, вода - остальное. Ферментацию проводят при температуре 28°С, аэрации 1 л/мин и скорости перемешивания 1000 об/мин. Через 24 часа после засева ферментера осуществляют pH-статирование культуры на уровне pH 5,9, используя для подтитровки растворы 10% серной кислоты и 10% NaOH. Общее время ферментации составляет 46 ч.
Продукция зрелого секретируемого белка GFN80 штаммом S. cerevisiae ВКПМ Y-3927 в этих условиях составляет более 150 мг/л.
Пример 12. Микробиологический синтез белка GFN100 с использованием штамма ВКПМ Y-3928.
Микробиологический синтез белка GFN100 проводят, как описано в примере 11, но с использованием штамма S. cerevisiae ВКПМ Y-3928.
Продукция зрелого секретируемого белка GFN100 штаммом S. cerevisiae ВКПМ Y-3928 в этих условиях составляет более 120 мг/л.
Пример 13. Очистка заявляемого белка GFN80 и определение его специфической активности.
Выделение и очистку белка GFN80 из культуральной жидкости, полученной в результате микробиологического синтеза интерферона с использованием штамма ВКПМ Y-3927 (пример 11), проводят при 4-8°C, используя следующие этапы:
1. Культуральную жидкость, содержащую белок GFN80, отделяют от биомассы дрожжей с помощью центрифугирования при 8500g в течение 20 минут.
2. Дальнейшую очистку белка GFN80 из полученного центрифугата осуществляют, используя катионообменную хроматографию на носителе SP-Sepharose Fast Flow (GE Healthcare). Для этого полученный после отделения клеток раствор подкисляют уксусной кислотой до значения pH 4,3±0,1. Подкисленный раствор пропускают через колонку с SP-сефарозой, уравновешенной 50 мМ раствором натрий ацетатного буфера рН 4,3, содержащего 1 мМ ЭДТА (буфер А). После прохождения фильтрата через колонку и последующей промывки колонки буфером А белок GFN80 элюируют раствором 0,5 М NaCl в буфере А.
3. Тонкую очистку белка GFN80 из полученного элюата проводят с использованием метода препаративной обратно-фазовой Высокоэффективной жидкостной хроматографии (офВЭЖХ). Для этого раствор белка наносят на колонку с сорбентом Vydac C18, уравновешенную 0,2%-ным раствором трифторуксусной кислоты (буфер В). Белок GFN80 элюируют градиентом ацетонитрила в буфере В (по программе: от 0 мин до 15 мин - 0%В; от 15 мин до 40 мин - от 0% до 45%В; от 40 мин до 60 мин - 45%В). Элюат сразу же разводят в 5 раз 5 мМ натрий ацетатным буфером, pH 4,0 (буфер C).
4. Полученный разведенный раствор белка подвергают дополнительной очистке методом ионообменной хроматографии на колонке с сорбентом SP-Сефароза. Колонку уравновешивают буфером C, наносят раствор белка и промывают колонку этим же буфером. Белок GFN80 элюируют 0,5 М хлоридом натрия в буфере C.
Характеристика эффективности этапов выделения и очистки белка GFN80 из культуральной жидкости приведены в табл.2.
Специфическую биологическую активность выделенного и очищенного белка GFN80 определяют методом La Bonnardiere & Laude [1981] в культуре перевиваемых линий клеток MDBK (АТСС №CCL-22), чувствительных к интерферону альфа-типа, в сравнении с международным стандартным образцом (МСО) (WHO International Standard INTERFERON ALPHA 2b, (Human rDNA derived) NIBSC code 95/566) против индикаторного вируса. В качестве индикаторного вируса используют вирус везикулярного стоматита (ВВС) штамм «Индиана» ГКВ ГУ НИИ Вирусологии им. Д.И. Ивановского РАМН (депозит №600) с инфекционным титром не менее 10-5 ТЦД50/мл или вирус энцефаломиокардита мышей (ЕМС) ГКВ ГУ НИИ Вирусологии им. Д.И. Ивановского РАМН (депозит №787) с инфекционным титром не менее 10-5 ТЦД50 в 1 мл.
Специфическая активность полученного белка GFN80 составляет 1,3×10 МЕ/мг.
Пример 14. Очистка синтезированного белка GFN100 и определение его специфической активности.
Выделение из культуральной жидкости, очистку и определение специфической активности белка GFN100 проводят, как в примере 13, за исключением того, что используют штамм ВКПМ Y-3928.
Специфическая активность полученного белка GFN100 составляет 1,1×10 МЕ/мг.
Пример 15. Определение фармакокинетических параметров заявляемых белков относительно стандартного интерферона.
Используют самцов мышей Balb/C×DBA весом 25-27 г, которые получают стандартный гранулированный корм и воду ad libitum. Накануне вечером перед введением препаратов корм убирают, непосредственно перед экспериментом убирают также питьевую воду, возвращая доступ к ней для животных после забора крови через 2 часа после введения исследуемого препарата. Животных кормят в день эксперимента после забора крови через 6 часов после введения исследуемых препаратов.
Растворы исследуемых препаратов для введения животным готовят непосредственно перед использованием. Препараты разводят стерильным 10 мМ фосфатным буфером pH 7,4, содержащим NaCl в концентрации 150 мМ, до концентрации 20 мкг/мл для в/в (внутривенного) или 40 мкг/мл для в/м (внутримышечного) введения. Для всех способов введения доза на животное составляет 2 мкг. Внутривенно растворы препаратов вводили в правый ретроорбитальный синус по 100 мкл на мышь. При внутримышечном введении растворы препаратов вводили в мышцы бедра по 50 мкл на мышь. В эксперименте используют 10 животных.
После введения у животных в установленные временные точки берут кровь из левого ретроорбитального синуса по 50 мкл. В каждом эксперименте используют 8 временных точек для отбора от момента введения: через 1, 5, 15, 30, 120, 300-360 и 1440 минут после введения. Образцы крови от каждого животного берут не более 5 раз по схеме (табл.3):
Образцы крови отбирают в пробирки эппендорф, инкубируют при комнатной температуре до образования сгустка, после чего сыворотку отделяют центрифугированием и замораживают при -20°C до определения уровня целевого белка.
Анализ содержания целевого белка в полученных образцах сыворотки крови проводят с помощью иммуноферментной тест-системы, специфичной к интерферону альфа-2 (набор «альфа-ИНТЕРФЕРОН-ИФА-БЕСТ», А-8758, производитель ЗАО «Вектор-Бест»).
В качестве стандарта для иммуноферментного анализа (ИФА) используют тот препарат белка, который вводили животным в опыте. Перед анализом образцы сыворотки разводят до рабочей концентрации буфером для стандарта, прилагаемым к тест-системе.
В качестве стандартного интерферона, служащего образцом сравнения, используют препарат «Интерфераль» производства ГосНИИ ОЧБ (Санкт-Петербург, Россия).
Полученные данные подвергают математической обработке. Анализ фармакокинетических параметров выполняют на основе экспериментально полученных значений концентрации исследуемых препаратов в сыворотке крови, измеренных в разное время после введения препарата. Для описания фармакокинетики используют и определяют следующие стандартные показатели [Руководство под ред. Хабриева, 2005].
D - введенная доза (нг/особь)
C(ti) - концентрация в момент ti (нг/мл)
Cmax - максимальная концентрация (нг/мл)
Tmax - время достижения максимальная концентрация (мин)
Thalf - время полувывода препарата(мин)
AUC∝ - полная площадь под кривой концентраций (мин·нг/мл)
AUCM∝ - полная площадь под 1-моментной кривой концентраций (мин2·нг/мл)
MRT=AUCM∝/AUCM∝ - среднее время удерживания (мин)
Cl=D/AUC∝ - общий клиренс (мл/нг/мин)
F%=Cliv/Cl=AUC*Div/AUCiv*D - биодоступность относительно внутривенного (iv) введения.
Показатели оценивают модельно-независимым способом, не опирающимся на согласование экспериментальных данных с какой-либо фармакологической моделью (1-, 2-, .. частной), т.е. не предполагающим конкретный тип функции от времени (сумма 1, 2,.. экспонент), значениями которой должны быть экспериментально измеренные концентрации.
При вычислении основного показателя AUS∝ (а также AUCM∝), зная который легко определить остальные показатели, за кривую концентраций берут кусочно-линейную кривую, соединяющую экспериментальные точки C(ti,) (t·C(ti) для AUCM∝), с продолжением до бесконечности экспонентой B·exp(-βt), проведенной через две последние экспериментальные точки и, в случае внутривенного введения, дополнением до нуля экспонентой A·exp(-αt), проведенной через две первые экспериментальные точки. Площадь под такой кривой есть сумма площадей трапеций плюс площади под одним-двумя одноэкспоненциальными участками.
Расчет параметров проводят по средним значениям для каждой временной точки («функция от среднего»).
Усредненные результаты измерений (табл.4) показывают, что после в/в введения мышам 2 мкг интерферона-альфа наблюдается быстрое экспоненциальное снижение его концентрации; через 2 часа уровень в сыворотке составляет около 2 нг/мл, а через 6 достигает почти неопределяемых значений. После в/м введения 2 мкг интерферона-альфа максимальная концентрация составляет около 30 нг/мл и наблюдается через 15-30 минут после введения. Через сутки после в/м введения данного препарата в сыворотке мышей не определяется его следов.
Заявляемые белки демонстрируют очевидные отличия от интерферона-альфа в профиле зависимости концентрации препарата от времени.
После в/в введения препаратов белка GFN80 наблюдается существенно замедленное по сравнению с интерфероном-альфа снижение концентрации белка от времени, причем некоторое количество белка определяется в крови животных даже через 24 часа после введения.
После в/м введения препаратов белка GFN80 или GFN100 максимальный уровень наблюдается примерно через 2 часа после введения, а через сутки определяются следовые количества.
Расчет фармакокинетических параметров (табл.5) показывает, что биодоступность интерферона-альфа (F%) при в/м введении составляет около 72%. Следует отметить, что ввиду сравнительно быстрого выведения данного препарата полная площадь под кривой концентраций (AUG∝) при в/в введении 2 мкг интерферона-альфа составляет около 10 мкг/мл в минуту. Время полувыведения (Thaif) интерферона-альфа после в/в введения составляет около 8 минут, после в/м введения - около 2 часов.
Расчет фармакокинетических показателей для в/в введения препарата белка GFN80 подтверждает значительно замедленное выведение из организма мышей по сравнению с интерфероном-альфа (табл.6). Клиренс (Cl) GFNa80 - показатель, определяющий скорость выведения белка из сыворотки крови, по данным расчетов составляет около 0,05 мл/нг/мин при том, что клиренс интерферона-альфа после в/в введения мышам составляет 0,23 мл/нг/мин. Полная площадь под кривой концентраций для белка GFN80 - составляет около 40 мкг/мл в минуту, что превышает уровень соответствующего показателя для интерферона-альфа в 4 раза. Время полувыведения белка GFN80 составляет около 5 часов, тогда как для интерферона-альфа этот показатель не превышает 8 минут.
После в/м введения (табл.7) препарата белка GFN80 максимальная концентрация препарата в сыворотке составляет около 40 нг/мл, что сопоставимо с соответствующим показателем для интерферона-альфа, однако из-за значительно более медленного выведения (время полувыведения около 4 часов) полная площадь под кривой концентраций составляет около 14 мкг/мл в минуту, что более чем в 2 раза превышает данный показатель, определенный после в/в введения интерферона-альфа.
Сравнение фармакокинетических показателей препаратов белка GFN80 или GFN100 при в/м введении (табл.7) демонстрирует очевидные признаки более длительной циркуляции белка GFN100. Не смотря на сравнительно небольшие различия по времени полувыведения, за счет более медленного выведения (клиренс (Cl) белка GFN100 - 0,08 мл/нг/мин, GFNa80 - 0,14 мл/нг/мин) полная площадь под кривой концентраций белка GFN100 почти в 2 раза превышает данный показатель для белка GFN80.
Таким образом,
- на основе интерферона альфа-2 человека получены варианты гибридного белка, обладающие удельной противовирусной активностью, превышающей 75% активности стандартного немодифицированного интерферона и при этом в 2-4 раза превосходящие его по времени жизни в организмах животных, в то время как применяющиеся в настоящее время пролонгированные варианты ПЭГ-ИФН обладают существенно сниженной удельной активностью;
- впервые для получения на основе интерферона альфа-2 человека гибридного белка обладающего пролонгированным действием, использованы полипептиды, состоящие из 81 или 101 аминокислотных остатков глицина и серина, в то время как для пролонгирования интерферона альфа-2 человека в ближайшем аналоге использован полипептид, включающий более 200 аминокислотных остатков, представленных пролином, аланином и серином;
- заявляемый белок имеет потенциальную терапевтическую ценность, так как позволяет предложить облегченный по сравнению со стандартным интерфероном режим приема, например 2 раза вместо 3 раз в неделю, при этом себестоимость заявляемого белка предположительно останется на уровне себестоимости стандартного немодифицированного интерферона;
- штаммы-продуценты вариантов заявляемого белка сконструированы на основе дрожжей Saccharomyces cerevisiae, секретирующих этот белок в культуральную жидкость в отличие от бактериального продуцента белка - ближайшего аналога, синтезирующего этот белок внутриклеточно;
- разработанный способ получения заявляемого белка, включающий его микробиологический синтез и последующее выделение, проще и экономичнее способа получения ближайшего аналога, так как не требует стадии разрушения клеток продуцента, и позволяет упростить процедуру выделения структурно полноценного заявляемого белка благодаря использованию в качестве продуцента эукариотического организма, не образующего бактериальных токсинов. Уровень продукции заявляемого белка по заявляемому способу почти в 30 раз выше уровня продукции способа получения белка-ближайшего аналога;
- экспрессия заявляемого белка осуществлена с использованием экспрессионной системы дрожжей Saccharomyces cerevisiae, входящих в список безопасных для человека организмов, что открывает возможности получения на основе этих дрожжей и других белков медицинского назначения, обладающих пролонгированным действием.
Источники информации
Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ (под ред. Хабриева Р.У.), - М.: «Медицина». - 2005. - 832 с.]
Therapeutic Proteins: Strategies to Modulate Their Plasma Half-Lives, First Edition. Edited by Roland Kontermann. © 2012 Wiley-VCH Verlag GmbH & Co. KGaA
Bain VG, Kaita KD, Yoshida EM, et al. (2006) A phase 2 study to evaluate the antiviral activity, safety, and pharmacokinetics of recombinant human albumin-interferon alfa fusion protein in genotype 1 chronic hepatitis С patients. J Hepatol; 44:671-678
Binder U., Skerra A. (2012) Half - Life Extension of Therapeutic Proteins via Genetic Fusion to Recombinant PEG Mimetics. Therapeutic Proteins: Strategies to Modulate Their Plasma Half-Lives, First Edition. Edited by Roland Kontermann. © 2012 Wiley-VCH Verlag GmbH & Co. KGaA
Clark V., Nelson D.R. (2009) Novel Interferons forTreatment of Hepatitis С Virus. Clin Liver Dis 13:351-363
Fried MW, Shiffman ML, Reddy KR, et al. (2002) Peginterferon alfa-2a plus ribavirin for chronic hepatitis С virus infection. N Engl JMed;347:975-82
Geething N.C., To W., Spink B.J., Scholle M.D., Wang C.W., Yin Y., Yao Y., Schellenberger V., Cleland J.L.. Stemmer W.P.C., Silverman J. (2010) Gcg-XTEN: an improved glucagon capable of preventing hypoglycemia without increasing baseline blood glucose. PLoS ONE, 5:el0175
Hadziyannis S.J. (2004) Peginterferon-alpha2a and ribavirin combination therapy in chronic hepatitis C: a randomized study of treatment duration and ribavirin dose. Ann Intern Med;140:346-355
Ito H., Fukuda Y, Murata K, Kimura A.(1983) Transformation of intact yeast cells treated with alkali cations. J.Bacteriol 153:163-168
Jones TD, Hanlon M, Smith BJ, Heise CT, Nayee PD, Sanders DA, Hamilton A, Sweet C, Unitt E, Alexander G, Lo KM, Gillies SD, Carr FJ, Baker MP. (2004) The development of a modified human IFN-alpha2b linked to the Fc portion of human IgGl as a novel potential therapeutic for the treatment of hepatitis С virus infection. J Interferon Cytokine Res. 24(9):560-72
Manns MP, McHutchison JG, Gordon SC, et al. (2001) Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis С: a randomised trial. Lancet;358:958-65
Nelson DR, Rustgi V, Balan V, et al. (2009a) Safety and antiviral activity of albinterferon alfa-2b in prior interferon nonresponders with chronic hepatitis C. Clin Gastroenterol Hepatol;7:212-218
Nelson D, Benhamou Y, Chuang WL, et al. (2009b) Efficacy and safety results of albinterferon alfa-2b in combination with ribavirin in treatment-nai..ve patients with chronic hepatitis С genotype 2 or 3 [oral presentation]. 44th Annual Meeting of the European Association for the Study of the Liver
Pasut G. (2009) PEGylated interferons: two different strategies to achieve increased efficacy. In PEGylated Protein Drugs: Basic Science and Clinical Applications. Edited by F.M. Veronese. Birkhauser Verlag/Switzerland
Subramanian GM, Fiscella M, Lamouse-Smith A, et al. (2007) Albinterferon [alpha]-2b: a genetic fusion protein for the treatment of chronic hepatitis C. Nat Biotechnol; 25:1411-9
Zeuzem S, Yoshida EM, Benhamou Y, et al. (2008) Albinterferon alfa-2b dosed every two or four weeks in interferon-nai..ve patients with genotype 1 chronic hepatitis C. Hepatology 2008;48:407-417
Zeuzem S, Sulkowski M, Lawitz E, et al. (2009) Efficacy and safety of albinterferon alfa-2b in combination with ribavirin in treatment-naf..ve patients with chronic hepatitis С genotype 1 [oral presentation]. 44th Annual Meeting of the European Association for the Study of the Liver
Walker A., Dunlevy G., Rycroft D., Topley P., Holt L.J., Herbert Т., Davies M., Cook F., Holmes S., Jespers L., Herring C. (2010). Anti-serum albumin domain antibodies in the development of highly potent, efficacious and long-acting interferon. Protein Engineering. Design & Selection, 23(4):271-278
Reddy KR. (2004) Development and pharmacokinetics and pharmacodynamics of pegylated interferon alfa-2a (40 kD). Semin Liver Dis; 24(Suppl 2):33-38
Schellenberger V., Wang C.W., Geething N.C., Spink B.J., Campbell A., To W., Scholle M.D., Yin Y., Yao Y., Bogin O. et al. (2009) A recombinant polypeptide extends the in vivo half-life of peptides and proteins in a tunable manner. Nat Biotechnol, 27:1186-1190
Skerra A. (2010) ′PASylation′: a superior technology to extend the plasma half-life of biologicals http://www.ideas-exchange.org/Upload/Content/Documents/Skerra_ASylation_13-10-10.pdf
Schlapschy M., Theobald I., Mack H., Schottelius M., Wester H.J., Skerra A. (2007) Fusion of a recombinant antibody fragment with a homo-amino-acid polymer: effects on biophysical properties and prolonged plasma half-life. Protein Engineering, Design & Selection, 20:273-284
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE И ШТАММ-ПРОДУЦЕНТ ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2446172C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА, ШТАММ Saccharomyces cerevisiae - ПРОДУЦЕНТ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2010 |
|
RU2427645C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ ШТАММОВ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫХ ПРОДУЦЕНТОВ СОМАТОТРОПИНА ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫЙ ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2009 |
|
RU2420567C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЦЕЛЕВОГО СЕКРЕТИРУЕМОГО БЕЛКА В ДРОЖЖАХ Saccharomyces cerevisiae | 2012 |
|
RU2502805C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА | 2011 |
|
RU2460795C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИБРИДНОГО БЕЛКА Е7-HSP70 (ВАРИАНТЫ) | 2013 |
|
RU2546917C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА E7-HSP70 И ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2489481C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ, СЛИТЫЙ БЕЛОК, РЕКОМБИНАНТНАЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И ШТАММЫ-ПРОДУЦЕНТЫ | 2010 |
|
RU2451023C1 |
| ГИБРИДНЫЙ БЕЛОК (ВАРИАНТЫ), ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА | 2011 |
|
RU2453604C1 |
| ГИБРИДНЫЙ БЕЛОК, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА ИЗ ЭТОГО ГИБРИДНОГО БЕЛКА | 2010 |
|
RU2441072C1 |
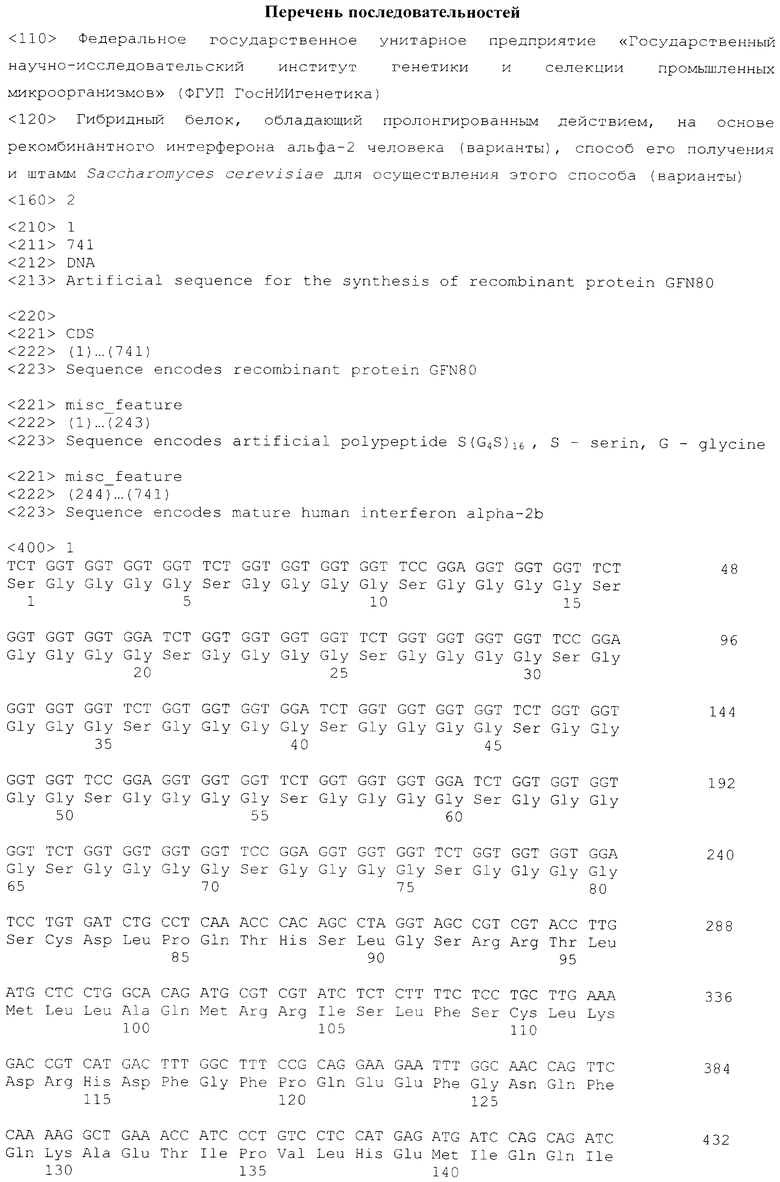


Изобретение относится к области биотехнологии и может быть использовано для получения рекомбинантного интерферона альфа-2 человека. Гибридные белки GFN80 и GFN100 конструируют на основе рекомбинантныго интерферона альфа-2 человека, слитого с N-конца с аминокислотной последовательностью полипептида S(G4S)16 или S(G4S)20 соответственно. Рекомбинантным путем получают штаммы продуценты Saccharomyces cerevisiae ВКПМ Y-3927 и Saccharomyces cerevisiae ВКПМ Y-3928. Штаммы используют в способе получения гибридного белка GFN80 и GFN100, который предусматривает культивирование в подходящих условиях дрожжевых клеток, трансформированных вектором экспрессии, который содержит область инициации репликации эндогенной 2-мкм плазмиды дрожжей Saccharomyces cerevisiae, а также промотор GAL1 дрожжей, контролирующий экспрессию гена, включающего последовательность ДНК SEQ ID NO:1 или SEQ ID NO:2, соответственно, с последующим выделением гибридного белка из культуральной жидкости. Изобретение позволяет получить гибридный рекомбинантный интерферон альфа-2 человека с пролонгированным действием в организме животных. 5 н.п. ф-лы, 7 табл., 15 пр.
1. Гибридный белок GFN80, обладающий пролонгированным действием и представляющий собой рекомбинантный интерферон альфа-2 человека, слитый с N-конца с аминокислотной последовательностью полипептида S(G4S)16.
2. Гибридный белок GFN100, обладающий пролонгированным действием и представляющий собой рекомбинантный интерферон альфа-2 человека, слитый с N-конца с аминокислотной последовательностью полипептида S(G4S)20.
3. Способ получения гибридного белка по п.1 или 2 путем культивирования в подходящих условиях дрожжей Saccharomyces cerevisiae, трансформированных вектором экспрессии, который содержит область инициации репликации эндогенной 2-мкм плазмиды дрожжей Saccharomyces cerevisiae, а также промотор GAL1 дрожжей, контролирующий экспрессию гена, включающего последовательность ДНК SEQ ID NO:1 или SEQ ID NO:2, соответственно, с последующим выделением гибридного белка из культуральной жидкости.
4. Штамм Saccharomyces cerevisiae ВКПМ Y-3927, предназначенный для осуществления способа по п.3 и полученный путем введения нуклеотидной последовательности SEQ ID NO:1 в составе плазмиды р71-81 в клетки штамма-реципиента, являющегося бесплазмидным производным штамма Saccharomyces cerevisiae ВКПМ Y-3550.
5. Штамм Saccharomyces cerevisiae ВКПМ Y-3928, предназначенный для осуществления способа по п.3 и полученный путем введения нуклеотидной последовательности SEQ ID NO:2 в составе плазмиды р71-82 в клетки штамма-реципиента, являющегося бесплазмидным производным штамма Saccharomyces cerevisiae ВКПМ Y-3550.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| КОНЪЮГАТЫ ИНТЕРФЕРОНОВ И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2011 |
|
RU2466138C1 |
| НОВЫЙ ФУНКЦИОНАЛЬНО АКТИВНЫЙ ВЫСОКООЧИЩЕННЫЙ СТАБИЛЬНЫЙ КОНЪЮГАТ ИНТЕРФЕРОНА α С ПОЛИЭТИЛЕНГЛИКОЛЕМ, ПРЕДСТАВЛЕННЫЙ ОДНИМ ПОЗИЦИОННЫМ ИЗОМЕРОМ ПЭГ-NH-ИФН, С УМЕНЬШЕННОЙ ИММУНОГЕННОСТЬЮ, С ПРОЛОНГИРОВАННЫМ БИОЛОГИЧЕСКИМ ДЕЙСТВИЕМ, ПРИГОДНЫЙ ДЛЯ МЕДИЦИНСКОГО ПРИМЕНЕНИЯ, И ИММУНОБИОЛОГИЧЕСКОЕ СРЕДСТВО НА ЕГО ОСНОВЕ | 2010 |
|
RU2447083C1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |

