Изобретение относится к области биотехнологии и медицины и касается способа микробиологического синтеза зрелого (правильно свернутого, имеющего корректно замкнутые дисульфидные связи и не содержащего N-концевого метионина) интерферона альфа-2 человека.
К интерферону альфа-2 человека (IFN-α2) будем относить два аллельных варианта интерферон альфа-2b и интерферон альфа-2а, отличающиеся в кодирующей области одиночной нуклеотидной заменой в позиции 137 (2а:А, 2b:G). [Kaluz et al., 1993 и 1994; Lee et al., 1995], которая приводит к аминокислотной замене (2а:Lys, 2b:Arg).
Интерфероны альфа-2 человека относятся к классу цитокинов, обладающих антивирусной активностью, и являются представителями семейства альфа-интерферонов, секретируемых практически всеми тирами вирус-инфицированных клеток человека [Pfeffer et al., 1998].
Рекомбинантные IFN-α2 применяют для терапии вирусных и опухолевых заболеваний [Samuel, 2001; Schadendorf et al., 2009; Pfeffer et al., 1998], в том числе для лечения твердых опухолей, таких как рак мочевого пузыря, рак почки, ВИЧ-индуцированная саркома Капоши и др. [Torti et al., 1988; Vugrin, D. et al., 1985; Rios el al., 1985]. IFN-α2 являются основными терапевтическими средствами, используемыми для лечения хронических форм гепатитов В и С [Clark & Nelson, 2009].
Основным способом получения рекомбинантных IFN-α2 в настоящее время является микробиологический синтез в клетках бактерий Escherichia coli [Eur. Pharm., 6th ed.: 2007]. Однако в то время как природным N-концевым аминокислотным остатком зрелого IFN-α2 является цистеин, у белков бактериального происхождения на N-конце может находиться дополнительный остаток метионина (формилметионина) [Voss et al., 1994], наличие которого способно влиять на стабильность и иммуногенность белка [Ben-Bassat & Bauer, 1987], а также в ряде случаев приводить к потере белком биологической активности [Rabbani et al., 1988]. С начала 2005 года Европейская Фармакопея ограничила использование на территории европейских стран препаратов, содержащих модифицированный интерферон [Eur. Pharm., 5th ed.: 2004]. В этой связи проблема получения зрелого IFN-α2 приобрела еще большее значение.
В клетках E.coli удаление (процессинг) из состава синтезируемых белков N-концевого метионина детерминируется радиусом бокового радикала следующего за метионином аминокислотного остатка [Hirel et al., 1989; Dalboge et al., 1990]. Однако процессинг часто оказывается неполным в случае гиперэкспрессии белков в виде телец включения [Yasueda et al., 1991; Hwang et al., 1999], как это характерно для способа получения IFN-α2 [RU 2319502].
Из числа ферментативных способов удаления N-концевого метионина следует отметить: 1) способы, основанные на использовании метионинаминопептидазы [Shapiro et al., 1988; Solbiati et al., 1999; Liao et al., 2004]; 2) способы, основанные на биосинтезе удлиненных белков, содержащих в N-концевой области сайт узнавания какой-либо протеиназы. Протеиназная обработка такого белка приводит к удалению введенной последовательности вместе с N-концевым метионином [US 5665566; Belagaje et al., 1997; Hosfield & Lu, 1999; Jenny et al., 2003]. Однако применение этих способов значительно усложняет технологию получения препарата зрелого IFN-α2.
Значительно более эффективным способом получения зрелого белка IFN-α2 является биологический способ, основанный на процессинге белка в ходе секреции. Так секреция интерферона, сопряженная с получением зрелого белка, реализована в клетках E.coli [Voss et al., 1994]. Для этой технологии характерен высокий уровень 250-300 мг/л продукции IFN-α2, однако белок накапливается в периплазме, что затрудняет его очистку, содержание белка в клетках невелико - 1.5 мг/г биомассы, доля корректно процессированного белка не превышает 60%, и общий выход очищенного IFN-α2 составляет всего около 14%.
Другой подход основан на секреции зрелого IFN-a2 культурой клеток животных [Rossmann et al., 1996; Ceaglio et al., 2008]. Недостатком метода является его относительная дороговизна.
Секреция IFN-α2 в клетках дрожжей является альтернативой обоим перечисленным способам, поскольку соединяет в себе высокий уровень продукции зрелого IFN-α2 и относительно недорогой способ производства.
В недавних работах [Shi et al., 2007; Ghosalkar et al., 2008; Salunkhe et al., 2010] показано, что пре-про область α-фактора дрожжей эффективно направляет секрецию зрелого интерферона альфа-2b в клетках дрожжей Pichia pastoris. Уровень накопления биологически активного интерферона в среде культивирования составляет до 300 мг/л. В условиях высокоплотного культивирования Р.pastoris получают до 600 мг/л зрелого IFN-α2 [Ayed et al., 2008].
Известно, что дрожжи Saccharomyces cerevisiae также способны секретировать в среду культивирования зрелый IFN-α2. Однако при использовании для секреции лидерной области α-фактора дрожжей более 50% от общего количества секретированного белка накапливается в среде в форме незрелого предшественника. Выход зрелого IFN-α2 в этом случае невелик, в 100 раз меньше, чем в Р.pastoris, он лимитирован недостаточной активностью протеиназы KEX2, отвечающей за отщепление лидерной области от зрелого белка [Singh et al., 1984; Zsebo et al., 1986; Hitzeman et al., 1990].
Протеиназа KEX2 участвует в процессинге секретируемых белков, содержащих в своем составе лидерную область α-фактора дрожжей. Известны две формы протеиназы KEX2: нативная (далее KEX2A), являющаяся трансмембранным белком, локализованным в комплексе Гольджи [Fuller et al., 1989], и секретируемая в среду культивирования дрожжей (далее KEX2B) [Brenner & Fuller, 1992]. В некоторых случаях увеличение экспрессии протеиназы KEX2A в клетках S. cerevisiae приводит к увеличению секреции белков, например гирудина пиявки [FR 2645174]. Влияние повышенной экспрессии протеиназы KEX2 (как KEX2A, так и KEX2B) на увеличение секреции IFN-α2 не показано.
Известен способ избежать влияния протеиназы KEX2 на уровень секреции белков путем замены лидерной пре-про области α-фактора на сигнальный пептид. Однако такой подход приводит к значительному снижению секреции IFN-α2 [Hitzman et al., 1983; Degryse et al., 1992].
В противовес низкому уровню продукции секретируемого IFN-α2 в клетках S.cerevisiae, характерному для ранних исследований, в работе Chu et al. [2003] показана возможность использования дрожжей S.cerevisiae для получения до 280 мг/л биологически активного интерферона альфа-2а.
Известно, что ряд гетерологичных белков, секретируемых клетками дрожжей в среду культивирования, подвергаются деградации при участии дрожжевых протеиназ. В некоторых случаях протеиназами, отвечающими за деградацию, оказываются япсины - аспартильные протеиназы, локализованные на поверхности клеточной стенки дрожжей [Krysan et al., 2005]. К ним относится Протеиназа YPS1, вызывающая Arg- и Lys- специфическую деградацию таких секретируемых белков, как глюкагон, предшественник бета-амилоидного белка человека, просоматостатин, специфический ингибитор эластазы - пре-элафин [Egel-Mitani et al., 2000; Bourbonnais et al., 1994 и 2000; Komano el al., 1998]. Участие протеиназы YPS1 в деградации IFN-α2, секретируемого клетками дрожжей, показано не было.
Присутствие деградированных форм белков в составе препаратов фармацевтического назначения жестко ограничивается требованиями Европейской Фармакопеи [Eur. Pharm., 6th ed.: 2007].
В качестве ближайших аналогов заявляемого способа и заявляемых штаммов рассмотрим способ биосинтеза биологически активного интерферона альфа-2а человека и штамм-продуцент S.cerevisiae DCO4 (Cir0, MATa, adel, leu2-04)/pHC11-IFNαA1, сведения о которых приведены в работе Chu et al. [2003], в которой показана возможность получения до 280 мг/л зрелого биологически активного интерферона альфа-2а человека.
Задача заявляемой группы изобретений - повысить выход зрелого интерферона альфа-2 человека, секретируемого дрожжами Saccharomyces cerevisiae в культуральную среду.
Задача решена путем
- разработки способа микробиологического синтеза зрелого интерферона альфа-2 человека, в котором в качестве продуцента используют секретирующие интерферон альфа-2 человека дрожжи Saccharomyces cerevisiae, содержащие инактивирующую мутацию в структурном гене протеиназы YPS1 и/или дополнительные гены протеиназы KEX2 (KEX2A или KEX2B);
- конструирования штаммов Saccharomyces cerevisiae ВКПМ Y-3549, ВКПМ Y-3548, ВКПМ Y-3493 и ВКПМ Y-3550 - продуцентов зрелого интерферона альфа-2b человека;
- конструирования штаммов Saccharomyces cerevisiae ВКПМ Y-3561, ВКПМ Y-3562, ВКПМ Y-3563 и ВКПМ Y-3564 - продуцентов зрелого интерферона альфа-2а человека.
В основу изобретения положены установленные нами факты, влияющие на выход зрелого интерферона: 1) деградация IFN-α2 снижается в результате инактивации протеиназы YPS1; 2) повышение уровня экспрессии протеиназы KEX2A или экспрессия протеиназы KEX2B приводят к увеличению эффективности процессинга и выхода секретируемого IFN-α2. Эти факты использованы при конструировании штаммов дрожжей S.cerevisiae - продуцентов зрелого секретируемого IFN-α2, в клетках которых одновременно с экспрессией интерферона осуществляют экспрессию одной из двух форм протеиназы KEX2 - внутриклеточной KEX2A или секретируемой KEX2B, а также в ряде случаев инактивируют ген протеиназы YPS1, отвечающей за деградацию интерферона в среде культивирования дрожжей.
Этапы конструирования заявляемых штаммов
Этап 1. Конструируют экспрессионные бирепликонные векторы pPDX2-IFN2b и pPDX2-IFN2a. Для этого последовательности генов интерферона IFN-α2 (альфа-2b или альфа-2а) человека амплифицируют в реакции ПЦР и клонируют в плазмиде. При этом в каждом случае осуществляют прецизионное слияние последовательности гена интерферона и лидерной пре-про области α-фактора дрожжей, слитой с промотором GAL1 дрожжей. В ходе секвенирования подтверждают корректность полученных конструкций. Затем полученные конструкции в виде фрагментов ДНК, кодирующих промотор GAL1 и гены интерферона альфа-2b или альфа-2а человека, слитые с лидерной пре-про областью α-фактора дрожжей, клонируют в лабораторном векторе pPDX2.
Этап 2. Конструируют реципиентные штаммы дрожжей. Для этого используют лабораторные штаммы А2 (МАТа ura3-52 leu2-3,112 trp1-289 lys7 pgkl::URA3 gal80::LEU2) и В2 (МАТα ura3-52 leu2-3,112 trp1-289 his3-Δ1 his4-519 pgk1::URA3 gal80::LEU2). Для удобства манипуляции с клетками этих штаммов на разных средах в них, используя метод трансформации, временно вводят лабораторный вектор pPDX2, не содержащий гена интерферона. В результате получают штаммы A2/pPDX2 и B2/pPDX2.
Этап 3. Конструируют фрагмент ДНК yps1::TRP1 для сайт-направленного разрушения гена протеиназы YPS1. Для этого ДНК-последовательность гена YPS1 амплифицируют в реакции ПЦР и клонируют в подходящий вектор. В полученной плазмиде замещают структурную часть гена YPS1 на фрагмент ДНК, кодирующий ген TRP1 дрожжей S.cerevisiae. При этом сохраняют фланкирующие последовательности гена YPS1, которые служат для направления интеграции этого фрагмента в хромосомный локус YPS1.
Этап 4. В клетках реципиентных штаммов дрожжей инактивируют ген протеиназы YPS1. Для этого фрагментом ДНК yps1::TRP1 трансформируют клетки штаммов A2/pPDX2 и B2/pPDX2. Используя метод ПЦР, доказывают, что в клетках полученных трансформантов инактивирован ген YPS1. Полученные штаммы называют A2yps1/pPDX2 и B2yps1/pPDX2.
Этап 5. Получают диплоидные штаммы дрожжей. Для этого, используя стандартные методы, скрещивают между собой гаплоидные штаммы A2/pPDX2 и B2/pPDX2, а также A2yps1/pPDX2 и B2yps1/pPDX2. В результате получают диплоидные штаммы D721/pPDX2 и Y21/pPDX2.
Этап 6. Получают бесплазмидные (свободные от вектора) диплоидные реципиентные штаммы. Для освобождения от вектора pPDX2 штаммы D721/pPDX2 и Y21/pPDX2 культивируют на средах, не содержащих сбраживаемые источники углерода, например, на среде с этанолом и глицерином. В этих условиях функциональный ген PGK1, локализованный в векторе pPDX2, не является жизненно необходимым для клеток дрожжей и может быть утрачен вместе с вектором в процессе клеточного деления. Бесплазмидные штаммы D721 и Y21 селектируют, используя стандартные микробиологические методы.
Этап 7. Получают штаммы-продуценты интерферона альфа-2b человека. Для этого клетки бесплазмидных штаммов D721 и Y21 трансформируют вектором pPDX2-IFN2b. Полученные трансформанты называют D721/pPDX2-IFN2b и Y21/pPDX2-IFN2b.
Этап 8. Конструируют интегративные вектора, несущие гены KEX2. Для этого, используя ПЦР, амплифицируют два варианта гена KEX2: полноразмерный структурный ген KEX2A и ген секретируемой формы протеиназы KEX2B, кодирующий N-концевую часть белка, содержащую каталитически активный домен. Амплифицированные гены KEX2A и KEX2B клонируют в интегративные векторы под контролем сильных дрожжевых промоторов.
Этап 9. Получают штаммы дрожжей - продуценты интерферона альфа-2b, секретирующие его в культуральную среду и несущие дополнительные копии генов KEX2A и KEX2B. Для этого клетки штаммов D721/pPDX2-IFN2b и Y21/pPDX2-IFN2b трансформируют интегративными векторами, несущими гены KEX2A и KEX2B. В результате трансформации получают 4 набора прототрофных штаммов, продуцирующих зрелый секретируемый интерферон альфа-2b человека и различающихся наличием мутантного аллеля yps1. Из каждой серии трансформантов отбирают наиболее продуктивные варианты, которые депонируют во Всероссийской коллекции промышленных микроорганизмов (ВКПМ).
В результате получают заявляемые штаммы:
- D721/pPDX2-IFN2b/pTRP1-GAL1-KEX2A (лаб. название: SCR-2b-1, Saccharomyces cerevisiae ВКПМ Y-3549)
- D721/pPDX2-IFN2b/pTRP1-GAL1-KEX2B (лаб. название: SCR-2b-1G3, Saccharomyces cerevisiae ВКПМ Y-3548)
- Y21/pPDX2-IFN2b/pTRP1-TDH3-KEX2A (лаб. название: SCR-2b-8, Saccharomyces cerevisiae ВКПМ Y-3493)
- Y21/pPDX2-IFN2b/pTRP1-GAL1-KEX2B (лаб. название: SCR-2b-8G1, Saccharomyces cerevisiae ВКПМ Y-3550)
Этап 10. Получают бесплазмидные диплоидные реципиентные штаммы, несущие дополнительные копии генов KEX2A и KEX2B. Для этого клетки штаммов ВКПМ Y-3549, ВКПМ Y-3548, ВКПМ и ВКПМ Y-3550 освобождают от вектора pPDX2-IFN2b, как на этапе 6.
Этап 11. Получают штаммы-продуценты интерферона альфа-2а человека. Для этого клетки бесплазмидных диплоидных реципиентных штаммов, полученных на этапе 10, трансформируют вектором pPDX2-IFN2a. Подтверждают способность полученных трансформантов секретировать интерферон и депонируют их во Всероссийской коллекции промышленных микроорганизмов (ВКПМ).
В результате получают заявляемые штаммы:
- D721/pPDX2-IFN2a7pTRP1-GAL1-KEX2A (лаб. название: SCR-2a-1, Saccharomyces cerevisiae ВКПМ Y-3561)
- D721/pPDX2-IFN2a/pTRP1-GAL1-KEX2B (лаб. название: SCR-2a-1G3, Saccharomyces cerevisiae ВКПМ Y-3562)
- Y21/pPDX2-IFN2a/pTRP1-TDH3-KEX2A (лаб. название: SCR-2a-8, Saccharomyces cerevisiae ВКПМ Y-3563)
- Y21/pPDX2-IFN2a/pTRP1-GAL1-KEX2B (лаб. название: SCR-2a-8G1, Saccharomyces cerevisiae ВКПМ Y-3564)
Заявляемые штаммы несут экспрессионные плазмиды, имеющие следующие свойства:
- содержат область инициации репликации эндогенной 2-мкм плазмиды дрожжей, т.е. способны поддерживаться в клетках дрожжей в эписомном многокопийном состоянии, ген дрожжей PGK1, используемый в качестве селективного маркера, последовательность бактериальной плазмиды pUC18, а также экспрессионную кассету с геном IFN-α2;
- в состав экспрессионных кассет входит промоторная область гена дрожжей GAL1, область структурного гена MFα1, кодирующая пре-про лидерный пептид предшественника полового феромона дрожжей, область структурного гена зрелого IFN-α2 и область терминации транскрипции гена CYC1 дрожжей. Наличие данной кассеты в составе вектора pPDX2 приводит к тому, что в клетках заявляемых штаммов транскрипция гена IFN-α2 направляется высокоэффективным регулируемым промотором GAL1 и терминируется в области терминации транскрипции гена CYC1 дрожжей; секреция зрелого IFN-α2 направляется лидерным пре-про пептидом. На стыке между лидерной областью и последовательностью зрелого IFN-α2 в составе экспрессируемого белка находится сайт узнавания дрожжевой протеиназы KEX2, непосредственно за которым следует цистеин - первый аминокислотный остаток зрелого IFN-α2. Вследствие этого в процессе секреции экспрессируемый белок подвергается процессингу под действием протеиназы KEX2, в результате чего из его состава выщепляется зрелый IFN-α2. Сочетание сильного промотора и высокой дозы гена IFN-α2 в совокупности обеспечивает высокий уровень секреции IFN-α2 в клетках дрожжей S.cerevisiae.
Заявляемые штаммы дрожжей S.cerevisiae являются диплоидными прототрофными штаммами. Диплоидный статус клеток обеспечивает им повышенную стабильность. В составе хромосом клеток этих штаммов присутствует гомозиготная мутация в гене фосфоглицераткиназы - генотип pgk1/pgk1. Такой генотип гарантирует невозможность роста потерявших плазмиду клеток этого штамма на любых средах, содержащих любой единственный источник углерода, усваиваемый дрожжами S.cerevisiae. Наличие гена PGK1 в составе вектора pPDX2 обеспечивает рост плазмидосодержащих клеток заявляемых штаммов и селективное поддержание этого вектора в клетках.
Заявляемые штаммы несут гомозиготную мутацию в гене GAL80 - генотип gal80::LEU2/gal80::LEU2. Мутация gal80 отвечает за инактивацию белка, репрессирующего промотор GAL1. Такой генотип клеток заявляемых штаммов приводит к изменению регуляции и позволяет осуществлять галактозо-независимую активацию промотора GAL1, контролирующего экспрессию IFN-α2, а также в ряде штаммов - экспрессию протеиназы KEX2.
Заявляемые штаммы несут дополнительные копии генов одной из двух форм протеиназы KEX2, внутриклеточной KEX2A (ВКПМ Y-3549, ВКПМ Y-3493, ВКПМ Y-3561 и ВКПМ Y-3563) или секретируемой KEX2 В (ВКПМ Y-3548, ВКПМ Y-3550, ВКПМ Y-3562 и ВКПМ Y-3564), что обеспечивает увеличение эффективности процессинга IFN-α2.
Заявляемые штаммы ВКПМ Y-3493, ВКПМ Y-3550, ВКПМ Y-3563 и ВКПМ Y-3564 несут мутацию yps1, приводящую к инактивации протеиназы YPS1, участвующей в деградации IFN-α2, благодаря чему понижается количество деградированных форм IFN-α2.
Характеристика заявляемых штаммов
Генотип заявляемых штаммов
SCR-2b-1 (a/α leu2/leu2 ura3/ura3 trp1/trp1::(pTRP1-GAL1-KEX2A) pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 STA2/STA2 suc0/SUC2)/pPDX2-IFN2b
SCR-2a-1 (a/α leu2/leu2 ura3/ura3 trp1/trp1::(pTRP1-GAL1-KEX2A) pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 STA2/STA2 suc0/SUC2)/pPDX2-IFN2a
SCR-2b-1G3 (a/α leu2/leu2 ura3/ura3 trp1/trp1::(pTRP1-GAL1-KEX2B) pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 STA2/STA2 suc0/SUC2)/pPDX2-IFN2b
SCR-2a-1G3 (a/α leu2/leu2 ura3/ura3 trp1/trp1::(pTRP1-GAL1-KEX2B) pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 STA2/STA2 suc0/SUC2)/pPDX2-IFN2a
SCR-2b-8 (a/α leu2/leu2 ura3/ura3 trp1/trp1 pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys71LYS7 his3/HIS3 his4/HIS4 yps1::TRP1/yps1::TRP1 ATH1/ath1::(ADH1-G418, TDH3-KEX2A) STA2/STA2 suc0/SUC2)/pPDX2-IFN2b
SCR-2a-8 (a/α leu2/leu2 ura3/ura3 trp1/trp1 pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 yps1::TRP1/yps1::TRP1 ATH1/ath1::(ADH1-G418, TDH3-KEX2A) STA2/STA2 suc0/SUC2)/pPDX2-IFN2a
SCR-2b-8G1 (a/α leu2/leu2 ura3/ura3 trp1/trp1 pgk1::URA3/pgk1::URA3 gal80::LEU21gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 yps1:TRP1/yps1:TRP1 ATH1/ath1::(ADH1-G418, GAL1-KEX2B) STA2/STA2 suc0/SUC2)/pPDX2-IFN2b
SCR-2a-8G1 (a/α leu2/leu2 ura3/ura3 trp1/trp1 pgk1::URA3/pgk1::URA3 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 yps1::TRP1/yps1::TRP1 ATH1/ath1::(ADH1-G418, GAL1-KEX2B) STA2/STA2 suc0/SUC2)/pPDX2-IFN2a
Все заявляемые штаммы характеризуются следующими культурально морфологическими и физиолого-биохимическими признаками:
Морфологические признаки:
При культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (мас.%): пептон 2, дрожжевой экстракт 1, глюкоза 2, агар 2, вода остальное, клетки заявляемых штаммов Saccharomyces cerevisiae имеют овальную форму, 3-7 мкм в диаметре. Клетки почкуются. Почкование истинное, многостороннее. Истинного мицелия не образуют.
Колонии имеют следующий вид:
1) на агаризованной среде YPD колонии белого цвета с ровным краем, матовой поверхностью, линзовидным профилем и сметанообразной консистенцией;
2) на агаризованной среде с крахмалом (состав, мас.%: пептон 2, дрожжевой экстракт 1, крахмал 1, агар 2, вода остальное) колонии белого цвета с узорчатым краем, матовой поверхностью, линзовидным профилем и крупчатой консистенцией.
Рост в жидкой среде с крахмалом: при 28°С в течение первых 24 ч культивирования -жидкость мутная, осадок белый, не комкуется, пристеночных пленок не образует. Физико-химические признаки: Все заявляемые штаммы - факультативные анаэробы. Температура роста 20-33°С (оптимум 28°С). рН культивирования 3,8-7,4 (оптимум 5,0).
Ассимиляция источников углерода: Все заявляемые штаммы сбраживают глюкозу, фруктозу, мальтозу, сахарозу, декстрины, крахмал. Не сбраживают лактозу, галактозу, инулин, ксилозу, арабинозу.
Ассимиляция источников азота: Все заявляемые штаммы усваивают аминокислоты, сернокислый аммоний, азотнокислый аммоний.
Патогенностъ: Заявляемые штаммы Saccharomyces cerevisiae непатогененны.
Хранение: Штаммы хранят при температуре -70°С в 20% водном растворе глицерина. Возможно хранение на агаризованной богатой среде с глюкозой в течение 3 месяцев при +4°С.
Стабильность: Стабильность заявляемых штаммов сохраняется при 20 последовательных пересевах на агаризованной среде YPD при температуре 28°С.
Продукция IFN-α2: При выращивании в ферментере клетки заявляемых штаммов Saccharomyces cerevisiae продуцируют зрелый интерферон IFN-α2.
Способ в общем виде
Для засева ферментера получают посевную культуру путем культивирования штаммов-продуцентов IFN-α2 в среде следующего состава, мас.%: пептон 1-3; дрожжевой экстракт 0,5-3; глюкоза 1-3, вода остальное, рН среды - естественный в течение 16-40 часов при температуре 22-32°С на орбитальной качалке 100-350 об/мин.
Посевной культурой засевают ферментер (0,5-1000 л), содержащий ферментационную среду, следующего состава, мас.%: пептон 1-3; дрожжевой экстракт 0,5-3, глюкоза или сахароза 1-4, вода остальное, рН среды - естественный. Количество посевной культуры, вносимой в ферментер составляет 3-15% от объема ферментационной среды.
Ферментацию проводят при 22-32°С, аэрации 0,5-1000 л/мин и скорости перемешивания культуры 100-1500 оборотов/мин. Через 16-30 ч после засева ферментера начинают подпитку 30-60% раствором глюкозы или сахарозы со скоростью 1-10 г./л./ч. и устанавливают рН-статирование культуры в диапазоне рН 5,0-рН 7,2. Общая продолжительность ферментации составляет 68-96 часов.
Выход секретируемого IFN-α2 в таких условиях составляет не менее 0,3 г/л.
Настоящее изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование кассеты для экспрессии гена секретируемого интерферона альфа-2b человека
Ген интерферона альфа-2b человека амплифицируют в реакции ПЦР с использованием праймеров N466 (ataccatggaaaagagatgtgatctgcctcaaacccacagcctaggtagccgt) и N467 (atetcgagtcattctttacttcttaaggattcttgcaagttt). Матрицей служит ДНК плазмиды pSX50 [RU 2319502]. Полученный в результате амплификации фрагмент ДНК размером 510 п.о. элюируют из агарозного геля с использованием кита Qiagen (Qiagen, cat. JMo 28706), обрабатывают рестриктазами Ncol и Xhol и клонируют в лабораторной плазмиде pUC18x-GAL1ppI. В результате осуществляют прецизионное слияние последовательности гена интерферона альфа-2b человека и последовательности, кодирующей лидерную пре-про область α-фактора дрожжей.
В результате получают плазмиду pUC18x-GAL1ppI-IFN2b, в составе которой с помощью секвенирования подтверждают корректность первичной последовательности клонированного гена интерферона альфа-2b человека.
Полученная плазмида pUC18x-GAL1ppI-IFN2b содержит фрагмент ДНК (SEQ ID: NO 1), в составе которого ген интерферона альфа-2b человека слит с последовательностью, кодирующей пре-про область α-фактора дрожжей, которая, в свою очередь, слита с промоторной областью гена GAL1 дрожжей.
Пример 2. Конструирование рекомбинантной плазмиды pPDX2-IFN2b
Рекомбинантная плазмида pPDX2-IFN2b представляет собой совокупность HindIII/XhoI фрагмента ДНК лабораторной векторной плазмиды pPDX2 и HindIII/XhoI фрагмента ДНК (SEQ ID: NO 1) плазмиды pUC18x-GAL1ppI-IFN2b, заключающего ген пре-про предшественника интерферона альфа-2b под контролем промотора GAL1.
Для получения плазмиды pPDX2-IFN2b плазмиду pUC18x-GAL1ppI-IFN2b расщепляют с помощью рестриктаз HindIII и XhoI, образовавшийся фрагмент ДНК (SEQ ID: NO 1) элюируют из геля и лигируют с ДНК лабораторного вектора pPDX2, расщепленного с помощью тех же рестриктаз. Лигирование проводят с помощью ДНК-лигазы фага Т4. В результате получают плазмиду pPDX2-IFN2b.
Сконструированную плазмиду pPDX2-IFN2b используют для биосинтеза в клетках дрожжей зрелого секретируемого интерферона альфа-2b человека.
Пример 3. Конструирование кассеты для экспрессии гена секретируемого интерферона альфа-2а человека
Ген интерферона альфа-2а человека амплифицируют в реакции ПЦР с использованием праймеров N617 (agcctaggtagccgtcgtaccttgatgctcctggcacagatgcgtaagatctctctttt) и N467 (atctcgagtcattctttacttcttaaggattcttgcaagttt). Матрицей служит ДНК плазмиды pSX50 [RU 2319502]. Использование таких праймеров для амплификации фрагмента ДНК обеспечивает одновременное изменение последовательности гена интерферона альфа-2b человека на последовательность гена интерферона альфа-2а человека. Амплифицированный фрагмент ДНК размером 480 п.о. элюируют из агарозного геля, как в примере 1, обрабатывают рестриктазами XmaJI и XhoI и клонируют в плазмиде pUC18x-GAL1ppI-IFN2b, расщепленной по тем же сайтам.
В результате получают плазмиду pUC18x-GAL1ppI-IFN2a, в составе которой с помощью секвенирования подтверждают корректность первичной последовательности клонированного гена интерферона.
Полученная плазмида pUC18x-GAL1ppI-IFN2a содержит ген интерферона альфа-2а человека, слитый с последовательностью, кодирующей пре-про область α-фактора дрожжей, слитой с промоторной областью гена GAL1 дрожжей.
Пример 4. Конструирование рекомбинантной плазмиды pPDX2-IFN2a
Рекомбинантную плазмиду pPDX2-IFN2a конструируют, как в примере 2, за исключением того, что донором фрагмента ДНК, содержащего ген интерферона альфа-2а человека, является плазмида pUC18x-GAL1ppI-IFN2a (пример 3).
Пример 5. Клонирование структурного гена протеиназы KEX2A
Структурный ген нативной протеиназы KEX2A амплифицируют в ходе ПЦР с использованием в качестве матрицы хромосомной ДНК лабораторного штамма S.cerevisiae Y618 [Kartasheva et al., 1996]. Праймерами для амплификации служат N500 (5'-ataggatccaccatgaaagtgaggaaatatatta) и N117 (5'-tttgctcgagtcgacatatcgaatcacgatcgtcc). Амплифицированный фрагмент ДНК размером 2476 пар оснований (п.о.) элюируют из агарозного геля, как в примере 1, обрабатывают рестриктазами BamHI и XhoI и клонируют в расщепленный по этим же сайтам лабораторный вектор pUC19x, содержащий в составе модифицированного полилинкера сайт XhoI вместо сайта HindIII.
В результате получают плазмиду pUC19x-KEX2A, в составе которой секвенированием подтверждают наличие последовательности гена KEX2A.
Плазмида pUC19x-KEX2A содержит уникальный BamHI/XhoI фрагмент ДНК, кодирующий ген нативной протеиназы KEX2A.
Пример 6. Конструирование гена секретируемой протеиназы KEX2B
Для получения гена секретируемой протеиназы KEX2B из состава клонированного гена KEX2A (пример 5) делегируют фрагмент ДНК, кодирующий трансмембранный и цитоплазматический домены нативной протеиназы KEX2A.
Для этого последовательность гена KEX2A в составе плазмиды pUC19x-KEX2A расщепляют с помощью рестриктазы Kpn2I, образовавшиеся липкие концы достраивают с помощью фрагмента Klenow ДНК полимеразы-1 E.coli, после чего полимеразу инактивируют и проводят дополнительное расщепление ДНК с помощью рестриктазы HindIII. Больший из образовавшихся фрагментов ДНК элюируют из геля, как в примере 1. Из этой же плазмиды pUC19x-KEX2A выщепляют и элюируют HindIII/PvuII фрагмент ДНК размером 1486 п.о., являющийся фрагментом гена протеиназы KEX2A. Оба элюированных фрагмента ДНК лигируют между собой, в результате чего получают плазмиду pUC19x-KEX2B.
Плазмида pUC19x-KEX2B содержит BamHI/XhoI фрагмент ДНК, кодирующий ген секретируемой формы протеиназы KEX2, называемой KEX2B.
Пример 7. Конструирование интегративного вектора athAG-TDH3-KEX2A
Для введения и экспрессии генов KEX2A и KEX2B в клетках дрожжей конструируют интегративные экспрессионные вектора. Для этого используют лабораторный вектор pTRP1, несущий селективный маркер - ген TRP1 дрожжей S.cerevisiae, и athAG, несущий ген АРН под контролем промотора ADH1 дрожжей S.cerevisiae. Ген АРН кодирует аминогликозид-3'-O-фосфотрансферазу E.coli и при введении в клетки дрожжей обеспечивает им устойчивость к антибиотику генетицину (G418).
Для конструирования плазмиды athAG-TDH3-KEX2A элюируют (1) HindIII/BamHI фрагмент ДНК лабораторной плазмиды pUC18x-TDH3 размером 1061 п.о., заключающий промоторную область гена TDH3, не содержащий в своем составе стартового ATG-кодона; (2) BamHI/XhoI фрагмент ДНК плазмиды pUC19x-KEX2A (пример 5) размером 2467 п.о., заключающий структурный ген протеиназы KEX2A, и (3) HindIII/XhoI фрагмент лабораторной плазмиды athAG, заключающий ее векторную часть. В результате одновременного лигирования трех элюированных фрагментов ДНК получают плазмиду athAG-TDH3-KEX2A.
Вектор athAG-TDH3-KEX2A при введении в клетки дрожжей обеспечивает экспрессию гена KEX2A под контролем промотора TDH3.
Пример 8. Конструирование интегративного вектора athAG-GAL1-KEX2B
Для конструирования плазмиды athAG-GAL1-KEX2B элюируют (1) HindIII/BamHI фрагмент ДНК лабораторной плазмиды pUC18x-GAL1 размером 531 п.о., заключающий промоторную область гена GAL1, не содержащий в своем составе стартового ATG-кодона; (2) BamHI/XhoI фрагмент ДНК плазмиды pUC19x-KEX2B (пример 6) размером 1879 п.о., заключающий структурный ген секретируемой протеиназы KEX2B, и (3) HindIII/XhoI фрагмент лабораторной плазмиды athAG, заключающий ее векторную часть. В результате одновременного лигирования трех элюированных фрагментов ДНК получают плазмиду athAG-GAL1-KEX2B.
Плазмида athAG-GAL1-KEX2B при введении в клетки дрожжей обеспечивает экспрессию гена KEX2B под контролем промотора GAL1.
Пример 9. Конструирование интегративного вектора pTRP1-GAL1-KEX2A
Для конструирования плазмиды pTRP1-GAL1-KEX2A элюируют (1) AatI/BamHI фрагмент ДНК плазмиды pUC18x-GAL1 размером 995 п.о., заключающий промоторную область гена GAL1, не содержащий в своем составе стартового ATG-кодона; (2) BamHI/XhoI фрагмент ДНК плазмиды pUC19x-KEX2A (пример 5) размером 2467 п.о., заключающий структурный ген протеиназы KEX2A, и (3) AatII/XhoI фрагмент лабораторной плазмиды pTRP1, заключающий ее векторную часть. В результате одновременного лигирования трех элюированных фрагментов ДНК получают плазмиду pTRP1-GAL1-KEX2A.
Плазмида pTRP1-GAL1-KEX2A при введении в клетки дрожжей обеспечивает экспрессию гена KEX2A под контролем промотора GAL1.
Пример 10. Конструирование интегративного вектора pTRP1-GAL1-KEX2B
Для конструирования плазмиды pTRPI-GAL1-KEX2 В элюируют (1) AatI/BamHI фрагмент ДНК плазмиды pUC18x-GAL1 размером 995 п.о., заключающий промоторную область гена GAL1, не содержащий в своем составе стартового ATG-кодона; (2) BamHI/XhoI фрагмент ДНК плазмиды pUC19x-KEX2B (пример 6) размером 1879 п.о., заключающий структурный ген протеиназы KEX2B, и (3) AatII/XhoI фрагмент лабораторной плазмиды pTRP1, заключающий ее векторную часть. В результате одновременного лигирования трех элюированных фрагментов ДНК получают плазмиду pTRP1-GAL1-KEX2B.
Плазмида pTRPI-GAL1-KEX2 В при введении в клетки дрожжей обеспечивает экспрессию гена KEX2B под контролем промотора GAL1.
Пример 11. Конструирование фрагмента ДНК для направленной инактивации гена YPS1
В ходе собственных исследований мы установили, что протеиназа YPS1 дрожжей участвует в деградации интерферона альфа-2b, накапливаемого в среде культивирования дрожжей.
Для направленной инктивации гена YPS1 конструируют фрагмент ДНК yps1::TRP1. Для этого ген YPS1 амплифицируют с помощью ПЦР. Для ПЦР используют в качестве матрицы хромосомную ДНК дрожжей штамма YBS618, как в примере 1. Праймерами служат N126 (5'-tctgacgcgtcaattagatcgtca) и N127 (5'-ccacgtacgcgttagtcaaaaatg). Амплифицированный фрагмент ДНК размером ок. 1800 п.о. элюируют из геля, как в примере 1, и после обработки концов фрагмента рестриктазой MluI клонируют в составе лабораторного вектора pUC19m. Вектор pUC19m является производным стандартного вектора pUC19, в котором участок полилинкера был заменен на сайт MluI. В результате клонирования получают плазмиду pUC19m-YPS1.
В плазмиде pUC19m-YPS1 производят следующие модификации: 1) в сайт MunI в промоторной области гена YPS1 клонируют последовательность линкера SmaI (gcccgggc); 2) HindIII/HindIII фрагмент ДНК структурного гена YPS1 замещают на последовательность линкера PstI (gctgcagc). Результирующую плазмиду называют pUC19m-yps1.
Лабораторную плазмиду pUC19-TRP1, содержащую геномный PstI/EcoRI фрагмент ДНК, кодирующей ген TRP1, модифицируют путем клонирования последовательности линкера SmaI (gcccgggc) в сайт EcoRI. Результирующую плазмиду называют pUC19-TRP1-Sma.
Элюируют SmaI/PstI фрагмент ДНК плазмиды pUC19-TRP1-Sma, заключающий ген TRP1, и SmaI/PstI векторную часть плазмиды pUC19m-yps1. В результате лигирования этих фрагментов ДНК получают плазмиду pUC19m-yps1::TRP1.
Плазмида pUC19m-yps1::TRP1 содержит MluI/MluI фрагмент ДНК yps1::TRP1, используемый для направленного разрушения гена YPS1 в геноме дрожжей.
Пример 12. Инактивация гена YPS1 в клетках штамма дрожжей S.cerevisiae A2
Для инактивации гена YPS1 используют лабораторный штамм A2 (МАТа ura3-52 leu2-3,112 trp1-289 lys7 pgk1::URA3 gal80::LEU2). Для удобства манипуляции с клетками этого штамма на разных средах в них, используя метод трансформации, вводят вектор pPDX2, не несущий целевого гена. Для этого клетки штамма A2 культивируют в течение 18-24 часов при температуре 28°С на агаризованной среде YPGE, следующего состава, мас.%: бактопептон 2, дрожжевой экстракт 1, бактоагар 2, этанол 2, глицерин 3, вода остальное. Трансформацию проводят по методу Ito et al. [1983]. Трансформанты отбирают по способности расти на среде YPD. Один из полученных трансформантов называют A2/pPDX2.
Для инактивации гена YPS1 клетки штамма A2/pPDX2 трансформируют MluI/MluI фрагментом ДНК плазмиды pUC19m-yps1::TRP1, заключающим кассету yps1::TRP1 (пример 11). Для этого клетки штамма A2/pPDX2 выращивают на агаризованной среде YPD при температуре 28°С в течение 16-20 часов и трансформируют, как описано выше. Для отбора трансформантов используют синтетическую среду с добавлением L-аминокислот лейцина, лизина и гистидина до конечной концентрации 30 мкг/л. Состав синтетической среды (мас.%): KH2PO4 0,1, MgSO4 0,05, NaCl 0,01, CaCl2 0,01, (NH4)2SO4 0,35, глюкоза 2, тиамин (витамина B1) 0,02, рибофлавин (витамина В2) 0,02, никотиновая кислота (витамина РР) 0,02, п-аминобензойная кислота 0,02, пантотенат кальция 0,02, биотин 0,0002, пиридоксин (витамина В6) 0,02, инозит 1, фолиевая кислота 0,02, вода остальное.
Разрушение гена YPS1 доказывают методом ПЦР с использованием праймеров N126 и N127 (пример 11). Один из отобранных штаммов называют A2yps1/pPDX2.
Клетки полученного штамма A2yps1/pPDX2 несут мутацию в гене протеиназы YPS1
Пример 13. Инактивация гена YPS1 в клетках штамма дрожжей S.cerevisiae B2
Инактивацию гена YPS1 в клетках лабораторного штамма B2 (МАТα ura3-52 leu2-3,112 trp1-289 his3-Δ1 his4-519 pgk1::URA3 gal80::LEU2) проводят, как в примере 12, но вместо клеток штамма А2 используют клетки штамма B2.
В результате получают штамм B2yps1/pPDX2, клетки которого несут мутацию в гене протеиназы YPS1.
Пример 14. Конструирование штамма D721/pPDX2-IFN2b - продуцента интерферона альфа-2b человека
Получают диплоидный штамм D721/pPDX2. Для этого скрещивают между собой клетки штаммов A2/pPDX2 (пример 12) и B2/pPDX2 (пример 13). Диплоидный штамм отбирают на синтетической среде, в которую дополнительно вносят триптофан конечной концентрации 30 мкг/л. Один из отобранных штаммов называют D721/pPDX2.
Клетки штамма D721/pPDX2 освобождают от эписомной плазмиды pPDX2. Для этого их культивируют на среде YPGE (пример 12). На этой среде одинаково эффективно растут клетки, несущие плазмиду pPDX2, в составе которой находится ген PGK1, и клетки, их утратившие. Бесплазмидные клоны селектируют по признаку отсутствия роста на среде YPD (пример 12) с глюкозой. Отобранный штамм называют D721.
Клетки штамма D721 трансформируют вектором pPDX2-IFN2b (пример 2), как в примере 12. В результате получают штамм D721/pPDX2-IFN2b.
Пример 15. Конструирование штамма Y21/pPDX2-IFN2b - продуцента интерферона альфа-2b человека
Конструирование штамма Y21/pPDX2-IFN2b проводят, как в примере 14, за исключением того, что для скрещивания берут штаммы A2/pPDX2 (пример 12) и B2/pPDX2 (пример 13).
Пример 16. Определение характеристик интерферона, продуцируемого штаммами дрожжей D721/pPDX2-IFN2b или Y21/pPDX2-IFN2b
Клетки штаммов D721/pPDX2-IFN2b или Y21/pPDX2-IFN2b культивируют в колбах при 30°С на качалке (250 об/мин) на жидкой среде YPD, засевая в титре 5×105-5×106 мл-1. Образцы для анализа отбирают через 46 ч выращивания культуры. Конечная оптическая плотность культуры составляет ОД600=40-45. Культуральную жидкость освобождают от клеток центрифугированием при 10000 об/мин в течение 1 мин и используют для последующего анализа.
Накопление интерферона в среде культивирования штаммов D721/pPDX2-IFN2b или Y21/pPDX2-IFN2b подтверждают методом электрофореза в денатурирующих восстанавливающих условиях в 18% полиакриламидном геле [Laemmli., 1970], а также методом иммуноблоттинга [Harlow & Lane, 2006] с использованием моноклональных мышиных антител H11F6, специфичных к интерферону альфа-2 человека (ГосНИИгенетика, Москва, Россия), и козьих антимышиных поливалентных антител (SIGMA, А-0412), конъюгированных с пероксидазой хрена. При необходимости белки культуральной жидкости концентрируют [Kozlov & Yagudin, 2007]. В качестве стандарта используют препарат интерферона альфа-2b бактериального происхождения (ООО «Фармапарк», Россия), имеющего известную концентрацию.
Количественное содержание интерферона в образцах культуральной жидкости определяют с помощью аналитической обращеннофазовой высокоэффективной жидкостной хроматографии (офВЭЖХ) на колонке Vydac Protein&Peptides C18 218TP5410, размер частиц 5 мкм, размер пор 300 Å, 4,6 мм внутр. диам., длина 250 мм (GRACE, США). В качестве контрольного образца используют препарат интерферона альфа-2b человека (ООО «Фармапарк», Россия).
Продукция интерферона составляет 50 мг/л для обоих штаммов.
У белка, синтезированного клетками штамма Y21/pPDX2-IFN2b, определяют последовательность пяти N-концевых аминокислот. Анализ проводят методом Эдмана [Edman, 1950] на секвенаторе Precise Sequencing System, model 492 (Applied Biosystems). Последовательность, определенная в результате анализа, в точности совпадает с последовательностью пяти первых аминокислотных остатков зрелого природного интерферона альфа-2 человека.
На основании вышеприведенных данных делают вывод об уровне секреции зрелого интерферона альфа-2b человека штаммами дрожжей D721/pPDX2-IFN2b и Y21/pPDX2-IFN2b.
Пример 17. Конструирование штамма ВКПМ Y-3549
Помимо зрелого интерферона альфа-2b штаммы D721/pPDX2-IFN2b и Y21/pPDX2-IFN2b секретируют значительное количество побочного продукта, являющегося высокомолекулярным предшественником интерферона, сохранившим лидерную про-область, наличие которого регистрируют методом иммуноблоттинга. Для превращения предшественника в целевой продукт - зрелый интерферон альфа-2b в клетки штаммов D721/pPDX2-IFN2b и Y21/pPDX2-IFN2b вводят дополнительные копии гена протеиназы KEX2.
С целью увеличения экспрессии нативной протеиназы KEX2A в клетки штамма D721/pPDX2-IFN2b вводят интегративную конструкцию pTRP1-GAL1-KEX2A (пример 9). Для этого вектор pTRP1-GAL1-KEX2A линеаризуют, используя уникальный сайт рестриктазы KpnI, расположенный в области гена TRP1, что позволяет направить интеграцию вектора в локус TRP1 генома дрожжей. Используемый способ трансформации позволяет получать многокопийные интегранты.
Трансформацию клеток штамма D721/pPDX2-IFN2b проводят, как в примере 7. Трансформанты отбирают на синтетической среде (пример 7), не содержащей триптофана.
В результате трансформации получают набор трансформантов D721(KEX2A)/pPDX2-IFN2b. Полученные трансформанты сравнивают с реципиентным штаммом по продукции секретируемого интерферона альфа-2b. Секрецию интерферона анализируют, как в примере 16.
Один из трансформантов, демонстрирующий наибольшее увеличение секреции зрелого интерферона альфа-2b и уменьшение секреции высокомолекулярного предшественника, называют SCR-2b-1. Данные офВЭЖХ (пример 16) свидетельствуют, что секреция зрелого интерферона клетками штамма SCR-2b-1 увеличена по сравнению с реципиентным штаммом в 1.6 раза.
Штамм SCR-2b-1 депонируют во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3549.
Пример 18. Конструирование штамма ВКПМ Y-3548
Конструирование и селекцию штамма ВКПМ Y-3548 проводят так же, как штамма ВКПМ Y-3549 (пример 17), за исключением того, что в клетки реципиентного штамма дрожжей вводят ген секретируемой формы протеиназы KEX2B, для чего для трансформации реципиентного штамма используют интегративную конструкцию pTRP1-GAL1-KEX2B (пример 10).
Отобранный штамм называют SCR-2b-1G3. Данные офВЭЖХ (пример 16) свидетельствуют, что секреция зрелого интерферона клетками штамма SCR-2b-1G3 увеличена по сравнению с реципиентным штаммом в 2.1 раза.
Штамм SCR-2b-1G3 депонируют во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3548.
Пример 19. Конструирование штамма ВКПМ Y-3493
Для увеличения экспрессии нативной протеиназы KEX2A в клетки штамма Y21/pPDX2-IFN2b (пример 15) вводят интегративную конструкцию, которую получают, обрабатывая вектор athAG-TDH3-KEX2A (пример 7) рестриктазой MluI, выщепляя из его состава бактериальную часть. Интегративная конструкция содержит области, позволяющие направлять ее интеграцию в локус АТН1 генома дрожжей и получать многокопийные интегранты.
Трансформацию клеток штамма Y21/pPDX2-IFN2b проводят, как в примере 7. Трансформанты отбирают на среде YPD, содержащей антибиотик генетицин (G418) в концентрации 200 мкг/мл.
В результате трансформации получают набор трансформантов Y21(KEX2)/pPDX2-IFN2b. Полученные трансформанты сравнивают с реципиентным штаммом по продукции секретируемого интерферона альфа-2b. Секрецию интерферона анализируют, как в примере 16.
Один из трансформантов, демонстрирующий наибольшее увеличение секреции зрелого интерферона альфа-2b и уменьшение секреции высокомолекулярного предшественника, называют SCR-2b-8. Данные офВЭЖХ (пример 16) свидетельствуют, что секреция зрелого интерферона клетками штамма SCR-2b-8 увеличена по сравнению с реципиентным штаммом в 1.6 раза.
Штамм SCR-2b-8 депонируют во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3493.
Пример 20. Конструирование штамма ВКПМ Y-3550
Конструирование и селекцию штамма ВКПМ Y-3550 проводят так же, как штамма ВКПМ Y-3493 (пример 19), за исключением того, что в клетки реципиентного штамма дрожжей вводят ген секретируемой формы протеиназы KEX2B, для трансформации реципиентного штамма используют интегративную конструкцию athAG-GAL1-KEX2B (пример 8).
Отобранный штамм называют SCR-2b-8G1. Данные офВЭЖХ (пример 16) свидетельствуют, что секреция зрелого интерферона клетками штамма SCR-2b-8G1 увеличена по сравнению с реципиентным в 2.1 раза.
Штамм SCR-2b-8G1 депонируют во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3550.
Пример 21. Конструирование штамма ВКПМ Y-3561
Штамм дрожжей S.cerevisiae ВКПМ Y-3561 - продуцент зрелого секретируемого интерферона альфа-2а человека - получают с использованием штамма ВКПМ Y-3549. Для этого клетки штамма ВКПМ Y-3549 освобождают от эписомной плазмиды pPDX2-IFN2b, как в примере 14, а полученный бесплазмидный штамм трансформируют вектором pPDX2-IFN2a (пример 4), как в примере 12.
После подтверждения секреции интерферона, как в примере 16, один из отобранных трансформантов называют SCR-2a-1 и депонируют его во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3561.
Пример 22. Конструирование штамма ВКПМ Y-3562
Штамм дрожжей S.cerevisiae ВКПМ Y-3562 - продуцент зрелого секретируемого интерферона альфа-2а человека - получают с использованием штамма ВКПМ Y-3548. Для этого клетки штамма ВКПМ Y-3548 освобождают от эписомной плазмиды pPDX2-IFN2b, как в примере 14, а полученный бесплазмидный штамм трансформируют вектором pPDX2-IFN2a (пример 4), как описано в примере 12.
После подтверждения секреции интерферона, как в примере 16, один из отобранных трансформантов называют SCR-2a-1G3 и депонируют его во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3562.
Пример 23. Конструирование штамма ВКПМ Y-3563
Штамм дрожжей S.cerevisiae ВКПМ Y-3563 - продуцент зрелого секретируемого интерферона альфа-2а человека - получают с использованием штамма ВКПМ Y-3493. Для этого клетки штамма ВКПМ Y-3493 освобождают от эписомной плазмиды pPDX2-IFN2b, как в примере 14, а полученный бесплазмидный штамм трансформируют вектором pPDX2-IFN2a (пример 4), как описано в примере 12.
После подтверждения секреции интерферона, как в примере 16, один из отобранных трансформантов называют SCR-2a-8 и депонируют его во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3563.
Пример 24. Конструирование штамма ВКПМ Y-3564
Штамм дрожжей S.cerevisiae ВКПМ Y-3564 - продуцент зрелого секретируемого интерферона альфа-2а человека - получают с использованием штамма ВКПМ Y-3550. Для этого клетки штамма ВКПМ Y-3550 освобождают от эписомной плазмиды pPDX2-IFN2b, как в примере 14, а полученный бесплазмидный штамм трансформируют вектором pPDX2-IFN2a (пример 4), как описано в примере 12.
После подтверждения секреции интерферона, как в примере 16, один из отобранных трансформантов называют SCR-2a-8G1 и депонируют его во Всероссийской Коллекции Промышленных Микроорганизмов как штамм Saccharomyces cerevisiae ВКПМ Y-3564.
Пример 25. Микробиологический синтез интерферона альфа-2b человека с использованием штамма ВКПМ Y-3549
Для получения посевного материала штамм ВКПМ Y-3549 выращивают в среде YPD на качалке (250 об/мин) при температуре 28°С в течение 20-24 ч.
50 мл посевного материала используют для засева 3 л ферментера Anglicon, содержащего 950 мл среды YPD. Ферментацию проводят при температуре 28°С, аэрации 1 л/мин и скорости перемешивания 1000 об/мин. Через 24 часа после засева ферментера начинают подпитку 50% раствором глюкозы со скоростью 2 мл/ч и устанавливают рН-статирование культуры на уровне рН 6.8, используя для подтитровки растворы 10% серной кислоты и 10% NaOH. Общее время ферментации составляет 72 часа.
Продукция зрелого секретируемого интерферона альфа-2b человека штаммом ВКПМ Y-3549 составляет не менее 350 мг/л.
Пример 26. Микробиологический синтез интерферона альфа-2b человека с использованием штамма ВКПМ Y-3548
Микробиологический синтез интерферона альфа-2b человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3548.
Продукция зрелого секретируемого интерферона альфа-2b человека штаммом ВКПМ Y-3548 составляет не менее 450 мг/л.
Пример 27. Микробиологический синтез интерферона альфа-2b человека с использованием штамма ВКПМ Y-3493
Микробиологический синтез интерферона альфа-2b человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3493.
Продукция зрелого секретируемого интерферона альфа-2b человека штаммом ВКПМ Y-3493 составляет не менее 350 мг/л.
Пример 28. Микробиологический синтез интерферона альфа-2b человека с использованием штамма ВКПМ Y-3550
Микробиологический синтез интерферона альфа-2b человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3550.
Продукция зрелого секретируемого интерферона альфа-2b человека штаммом ВКПМ Y-3550 составляет не менее 450 мг/л.
Пример 29. Микробиологический синтез интерферона альфа-2а человека с использованием штамма ВКПМ Y-3561
Микробиологический синтез интерферона альфа-2а человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3561.
Продукция зрелого секретируемого интерферона альфа-2а человека штаммом ВКПМ Y-3561 составляет не менее 350 мг/л.
Пример 30. Микробиологический синтез интерферона альфа-2а человека с использованием штамма ВКПМ Y-3562
Микробиологический синтез интерферона альфа-2а человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3562.
Продукция зрелого секретируемого интерферона альфа-2а человека штаммом ВКПМ Y-3562 составляет не менее 450 мг/л.
Пример 31. Микробиологический синтез интерферона альфа-2а человека с использованием штамма ВКПМ Y-3563
Микробиологический синтез интерферона альфа-2а человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3563.
Продукция зрелого секретируемого интерферона альфа-2а человека штаммом ВКПМ Y-3563 составляет не менее 350 мг/л.
Пример 32. Микробиологический синтез интерферона альфа-2а человека с использованием штамма ВКПМ Y-3564
Микробиологический синтез интерферона альфа-2а человека проводят, как описано в примере 25, но с использованием штамма ВКПМ Y-3564.
Продукция зрелого секретируемого интерферона альфа-2а человека штаммом ВКПМ Y-3564 составляет не менее 450 мг/л.
Пример 33. Очистка синтезированного интерферона альфа-2b и определение его специфической активности
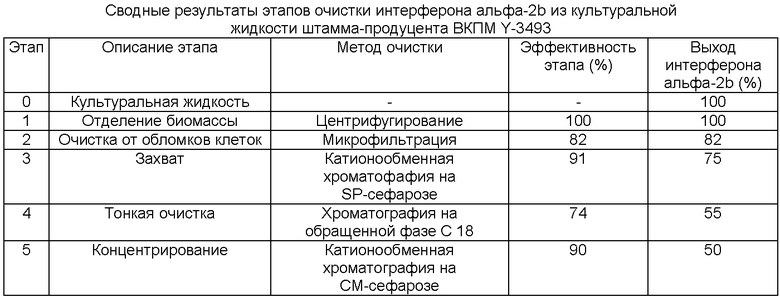
Выделение и очистку интерферона альфа-2b из культуральной жидкости, полученной в результате микробиологического синтеза интерферона с использованием штамма ВКПМ Y-3493 (пример 27), проводят при 4-8°С, используя следующие этапы:
1. Культуральную жидкость, содержащую интерферон альфа-2b, отделяют от биомассы дрожжей с помощью центрифугирования при 8500 g в течение 10 минут;
2. Полученный центрифугат культуральной жидкости очищают от клеточных обломков с помощью микрофильтрации на мембранном кассетном модуле из полипропилена с номинальным отсечением 0,2 мкм и номинальной площадью фильтрации 0,1 кв.м (ЗАО «Владисарт», Россия);
3. Захват интерферона альфа-2b из полученного микрофильтрата осуществляют, используя катионообменную хроматографию на носителе SP-Sepharose Fast Flow (GE Healthcare). Для этого полученный фильтрат подкисляют уксусной кислотой до значения рН 4,4-4,5. Подкисленный фильтрат пропускают через колонку с SP-Сефарозой, уравновешенной 40 мМ буферным раствором ацетата аммония, рН 4,5, содержащим 0,05% Tween 80 (буфер А). После прохождения фильтрата через колонку и последующей промывки колонки раствором 0,12 М NaCl в буфере А интерферон альфа-2b элюируют раствором 0,4 М NaCl в буфере А.
4. Проводят тонкую очистку интерферона альфа-2b из полученного элюата с использованием препаративной офВЭЖХ и последующее концентрирование очищенного белка с использованием катионнообменной хроматографии [RU 2319502].
Характеристики эффективности этапов выделения и очистки интерферона альфа-2b из культуральной жидкости приведены в таблице.
Специфическую биологическую активность выделенного и очищенного интерферона альфа-2b определяют методом La Bonnardiere & Laude [1981] в культуре перевиваемых линий клеток MDBK (АТСС № CCL-22), чувствительных к интерферону альфа-типа в сравнении с международным стандартным образцом (МСО) (WHO International Standard INTERFERON ALPHA 2b, (Human rDNA derived) NIBSC code 95/566) против индикаторного вируса. В качестве индикаторного вируса используют вирус везикулярного стоматита (ВВС) штамм «Индиана» ГКВ ГУ НИИ Вирусологии им. Д.И.Ивановского РАМН (депозит №600) с инфекционным титром не менее 10-5 ТЦД50/мл или вирус энцефаломиокардита мышей (ЕМС) ГКВ ГУ НИИ Вирусологии им. Д.И.Ивановского РАМН (депозит №787) с инфекционным титром не менее 10-5 ТЦД50 в 1 мл.
Специфическая активность полученного интерферона альфа-2b составляет 2,0×108 ME/мг.
Таким образом, разработан способ микробиологического синтеза зрелого интерферона альфа-2 человека, в котором увеличение продукции зрелого интерферона альфа-2 достигнуто за счет снижения уровня деградации секретируемого интерферона путем инактивации гена протеиназы YPS1 дрожжей, а также за счет улучшения эффективности процессинга предшественника секретируемого интерферона путем увеличения экспрессии дрожжевой протеиназы KEX2 в нативной либо секретируемой форме.
Общий выход высокоочищенного интерферона альфа-2b составляет 50% от его содержания в культуральной жидкости. Выделенный интерферон альфа-2b содержит на N-конце остаток цистеина и обладает специфической активностью 2,0×108 МЕ/мг, при этом по данным офВЭЖХ содержание нативной формы интерферона альфа-2b в конечном продукте составляет не менее 98%.
Источники информации
Ayed A., Rabin I., Dellagi K., Kallel H. (2008) High level production and purification of human interferon a2b in high cell density culture of Pichia pastoris, Enzyme Microb. Technol. 42: 173-180.
Ben-Bassat A., Bauer K. (1987) Amino-terminal processing of proteins. Nature 326: 315.
Belagaje R.M., Reams S.G., Ly S.C., Prouty W.F. (1997) Increased production of low molecular weight recombinant proteins in Escherichia coli. Protein Sci, 6: 1953-1962.
Brenner С., Fuller R.S. (1992) Structural and enzymatic characterization of a purified prohormone-processing enzyme: Secreted, soluble Kex2 protease. Proc. Natl. Acad. Sci. USA, 89: 922-926.
Bourbonnais, Y., Germain, D., Ash, J. & Thomas, D.Y. (1994) Cleavage of prosomatostatins by the yeast Yap3 and Kex2 endoprotease. Biochimie 76, 226-233.
Bourbonnais, Y., Larouche, C. & Tremblay, G.M. (2000) Production of full-length human pre-elafin, an elastase specific inhibitor, from yeast requires the absence of a functional yapsin 1 (Yps1p) endoprotease. Protein Expr. Purif 20, 485-491.
Ceaglio N., Etcheverrigaray M., Kratje R., Oggero M. (2008) Novel long-lasting interferon alpha derivatives designed by glycoengineering. Biochimie 90: 437-449.
Chu J, Zhang S, Zhuang Y. (2003). Fermentation process optimization of recombinant Saccharomyces cerevisiae for the production of human interferon-alpha2a. Appl Biochem Biotechnol., 111: 129-38.
Clark V., Nelson D.R., (2009) Novel interferons for treatment of hepatitis с virus. Clin Liver Dis, 13: 351-363.
Dalboge H., Bayne S., Pedersen J. (1990) In vivo processing of N-terminal methionine in E.coli. FEBS Lett. 266: 1-3.
Degryse E., Dietrich M., Nguyen M., Achstetter Т., Charlier M., Charpigny G., Gaye P., Martal J. (1992) Addition of a dipeptide spacer significantly improves secretion of ovine trophoblast interferon in yeast. Gene, 118: 17-53.
Edman P. (1950) Method for the determination of the amino acid sequence in peptides. Acta Chem. Scand., 4: 283.
Egel-Mitani, M., Andersen, A.S., Diers, I.I., Hach, M., Thim, L., Hastrup, S., Vad, K. (2000) Yield improvement of heterologous peptides expressed in yps1-disrupted Saccharomyces cerevisiae strains. Enzyme Microb. Technol. 26: 671-677.
Fuller R.S., Brakes A., Thorner J. (1989) Yeast prohormone processing enzyme (KEX2 gene product) is a Ca2+-dependent serine protease. Proc. Nati. Acad. Sci. USA, 86: 1434-1438.
Ghosalkar A., Sahai V., Srivastava A. (2008). Secretory expression of interferon-alpha2b in recombinant Pichia pastoris using three different secretion signals. Protein Expression and Purification, 60: 103-109.
Harlow E., Lane D. (2006) Immunoblotting: Antigen Detection Using Chemiluminescence. Cold Spring Harb Protoc; 2006; doi:10.1101/pdb.prot4271.
Hirel P.H., Schmitter M.J., Dessen P., Fayat G., Blanquet S. (1989) Extent of N-terminal methionine excision from Escherichia coli proteins is governed by the side-chain length of the penultimate amino acid. Proc Natl Acad Sci USA, 86: 8247-51.
Hitzman R.A., Leung D.W., Perry L.J., Kohr W.J., Levine H.L., Goeddel D. (1983) Secretion of human interferons by yeast, Science 219: 620-625.
Hitzeman R.A., Chen C.Y., Dowbenko D.J., Renz M.E., Liu C., Pai R., Simpson N.J., Kohr W.J., Singh A., Chisholm V., Hamilton. R., Chang C.N. (1990) Use of heterologous and homologous signal sequences for secretion of heterologous proteins from yeast. Methods Enzymol. 185: 421-440.
Hosfield Т., Lu Q. (1999) Influence of the Amino Acid Residue Downstream of (Asp)4Lys on Enterokinase Cleavage of a Fusion Protein. Anal Biochem, 269: 10-16.
Hwang D.D.W., Liu L.F., Kuan I.C., Lin L.Y., Tam T.C.S., Tam M.F. (1999) Co-expression of glutathione S-transferase with methionine aminopeptidase: a system of producing enriched N-terminal processed proteins in Escherichia coli. Biochem J, 338: 335-342.
Ito H., Fukuda Y, Murata K, Kimura A. (1983) Transformation of intact yeast cells treated with alkali cations. J. Bacterial. 153: 163-168.
Jenny R.J., Mann K.G., Lundblad R.L. (2003) A critical review of the methods for cleavage of fusion proteins with thrombin and factor Xa. Protein Express Purif, 31: 1-11.
Kartasheva N.N., Kuchin S.V., Benevolensky S.V. (1996) Genetic aspects of carbon catabolite repression of the STA2 glucoamylase gene in Saccharomyces cerevisiae. Yeast 12: 1297-13.
Kaluz S, Gibadulinova A., Kontsek P. (1993) Interferon alpha 2b but not interferon alpha 2a detected in human genomic DNA. Acta Virol. 37: 97-100.
Kaluz S., Kabat P., Gibadulinova A., Vojtassak J., Fuchsberger N., Kontsek P. (1994) Interferon alpha 2b is the predominant subvariant detected in human genomic DNAs. Acta Virol. 38: 101-4.
Komano, H., Seeger, M., Gandy, S., Wang, G.T., Krafft, G.A. & Fuller, R.S. (1998) Involvement of cell surface glycosylphosphatidylinositol - linked aspartyl proteases in alpha-secretasetype cleavage and ectodomain solubilization of human Alzheimer beta-amyloid precursor protein in yeast. J. Biol. Chem. 273, 31648-31651.
Kozlov D.G., Yagudin T.A. (2008) Antibody fragments may be incorrectly processed in the yeast Pichia pastoris. Biotechnology Letters (2008) 30: 1661-1663.
Krysan D.J., Ting E.L., Abeijon C., Kroos L., Fuller R.S. (2005) Yapsins Are a Family of Aspartyl Proteases Required for Cell Wall Integrity in Saccharomyces cerevisiae. Eukaryotic Cell 4: 1364-1374.
La Bonnardiere С., Laude H. (1981). High interferon titer in newborn pig intestine during experimentally induced viral enteritis. Infection and Immunity 32, 28-31.
Laemmli U.K. (1970) Cleavage of Structural Proteins During the Assembly of the Head of Bacteriophage T4. Nature, 227: 680-685.
Lee N., Ni D., Brissette R., Chou M., Hussain M., Gill D.S., Liao M.J., Testa D. (1995) Interferon-alpha 2 variants in the human genome. J Interferon Cytokine Res. 15: 341-9.
Liao Y.D., Jeng J.C., Wang C.F., Wang S.C., Chang S.T. (2004) Removal of N-terminal methionine from recombinant proteins by engineered E.coli methionine aminopeptidase. Protein Science, 13: 1802-1810.
Pfeffer L.M., Dinarello C.A., Herberman R.B., Williams B.R.G., Borden E.C., Bordens R., Walter M.R., Nagabhushan T.L., Trotta P.P., Pestka S. (1998) Biological properties of recombinant α-interferons: 40th anniversary of the discovery of interferons. Cancer Research 58: 2489-2499.
Rabbani S.F. et al., (1988) Recombinant human parathyroid hormone synthesized in Escherichia coli. J Biol Chem, 263: 1307-1313.
Rios A, Mansell P, Newell GR. Reuben JM, Hersh EM, Gutterman JU. (1985) Treatment of acquired immunodeficiency syndrome-related Kaposi's sarcoma with lymphoblastoid interferon. J. Clin. Oncol., 3: 506-512.
Rossmann C., Sharp N., Alien G., Gewert D. (1996) Expression and purification of recombinant, glycosylated human interferon alpha2b in murine myeloma NSo cells. Protein Expr Purif. 7: 335-42.
Salunkhe S, Soorapaneni S, Prasad KS, Raiker VA, Padmanabhan S. (2010) Strategies to maximize expression of rightly processed human interferon alpha2b in Pichia pastoris. Protein Expr Purif. 71 (2): 139-46.
Samuel C.E. (2001) Antiviral actions of interferons. Clinical Microbiology Reviews, 14: 778-809.
Schadendorf D., Algarra S.M., Bastholt L., Cinat G., Dreno В., Eggermont A.M.M., Espinosa E., Guo J., Hauschild A., Petrella Т., Schachter J., Hersey P. (2009) Immunotherapy of distant metastatic disease. Annals of Oncology, 20 (Supplement 6): vi41-vi50.
Shapiro R., Harper J.W., Fox E.A., Jansen H.W., Hein F., Uhlmann E. (1988) Expression of Met-(-1) angiogenin in Escherichia coli: Conversion to the authentic <Glu-1 protein. Analytical Biochemistry 175: 450-461.
Shi L., Wang D., Chan W., Cheng L. (2007). Efficient expression and purification of human interferon alpha2b in the methylotrophic yeast, Pichia pastoris. Protein Expr Purif., 54: 220-6.
Singh A., Lugovoy J.M., Kohr W.J., Perry L.J. (1984) Synthesis, secretion and processing of alpha-factor-interferon fusion proteins in yeast, Nucleic Acids Res. 12: 8927-8938.
Solbiati J., Chapman-Smith A., Miller J.L., Miller C.G., Cronan J.E. (1999) Processing of the N termini of nascent polypeptide chains requires deformylation prior to methionine removal. J Mol Biol 290: 607-614.
Torti F.M., Shortliffe L.D., Williams R.D., Pitts W.C., Kempson R.L., Ross J.C., Palmer J., Meyers F., Ferrari M., Hannigan J. (1988) Alpha-interferon in superficial bladder cancer: a Northern California Oncology Group Study. J. Clin. Oncol., 6: 476-483.
Voss Т., Falkner E., Ahorn H., Krystek E., Maurer-Fogy I., Bodo G., Hauptmann R. (1994). Periplasmic expression of human interferon-α2c in Escherichia coli results in a correctly folded molecule. Biochem. J., 298: 719-725.
Vugrin D, Hood L, Taylor W, Laszlo J. (1985) Phase II study of human lymphoblastoid interferon in patients with advanced renal carcinoma. Cancer Treat. Rep., 69: 817-820.
Yasueda H., Kikuchi Y., Kojima H., Nagase K. (1991) In-vivo processing of the initiator methionine from recombinant methionyl human interleukin-6 synthesized in Escherichia coli overproducing aminopeptidase-P. Appl Microbiol Biotechnol, 36: 211-215.
Zsebo K.M., Lu H.S., Fieschko J.C., Goldstein L., Davis J., Duker K., Suggs S.V., Lai P.H., Bitter G.A. et al. (1986). Protein secretion from Saccharomyces cerevisiae directed by the prepro-α-factor leader region. J. Biol. Chem. 261: 5858-5865.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE И ШТАММ-ПРОДУЦЕНТ ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2446172C1 |
| ГИБРИДНЫЙ БЕЛОК, ОБЛАДАЮЩИЙ ПРОЛОНГИРОВАННЫМ ДЕЙСТВИЕМ, НА ОСНОВЕ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ), СПОСОБ ЕГО ПОЛУЧЕНИЯ И ШТАММ Saccharomyces cerevisiae ДЛЯ ОСУЩЕСТВЛЕНИЯ ЭТОГО СПОСОБА (ВАРИАНТЫ) | 2013 |
|
RU2515913C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА | 2011 |
|
RU2460795C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ ШТАММОВ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫХ ПРОДУЦЕНТОВ СОМАТОТРОПИНА ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫЙ ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2009 |
|
RU2420567C1 |
| ГИБРИДНЫЙ БЕЛОК (ВАРИАНТЫ), ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА | 2011 |
|
RU2453604C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА E7-HSP70 И ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2489481C1 |
| ГИБРИДНЫЙ БЕЛОК, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА ИЗ ЭТОГО ГИБРИДНОГО БЕЛКА | 2010 |
|
RU2441072C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ, СЛИТЫЙ БЕЛОК, РЕКОМБИНАНТНАЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И ШТАММЫ-ПРОДУЦЕНТЫ | 2010 |
|
RU2451023C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЦЕЛЕВОГО СЕКРЕТИРУЕМОГО БЕЛКА В ДРОЖЖАХ Saccharomyces cerevisiae | 2012 |
|
RU2502805C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА КЛЕТКАМИ ДРОЖЖЕЙ, СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE-ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1998 |
|
RU2180003C2 |
Изобретение относится к области генной инженерии, конкретно к рекомбинантной продукции интерферона человека, и может быть использовано для получения рекомбинантного интерферона альфа-2 человека. Способ микробиологического синтеза зрелого интерферона альфа-2 человека осуществляют путем культивирования дрожжей Saccharomyces cerevisiae, содержащих инактивирующую мутацию в структурном гене протеиназы YPS1 и/или дополнительные гены протеиназы КЕХ2. Рекомбинантным путем получают штаммы дрожжей Saccharomyces cerevisiae, способные секретировать зрелый интерферон альфа-2 человека в культуральную среду. Изобретение позволяет увеличить продукцию зрелого интерферона альфа-2 за счет снижения деградации секретируемого интерферона путем инактивации гена протеиназы YPS1 дрожжей, а также за счет улучшения эффективности процессинга предшественника секретируемого интерферона путем увеличения экспрессии протеиназы KEX2 в нативной либо секретируемой форме. 9 н.п. ф-лы, 1 табл.
1. Способ микробиологического синтеза зрелого интерферона альфа-2 человека путем культивирования дрожжей Saccharomyces cerevisiae, секретирующих зрелый интерферон альфа-2 человека в культуральную среду, отличающийся тем, что в качестве продуцента используют дрожжи Saccharomyces cerevisiae, содержащие инактивирующую мутацию в структурном гене протеиназы YPS1 и/или дополнительные гены протеиназы KEX2.
2. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3549 - продуцент зрелого интерферона альфа-2b человека для осуществления способа по п.1.
3. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3548 - продуцент зрелого интерферона альфа-2b человека для осуществления способа по п.1.
4. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3493 - продуцент зрелого интерферона альфа-2b человека для осуществления способа по п.1.
5. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3550 - продуцент зрелого интерферона альфа-2b человека для осуществления способа по п.1.
6. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3561 - продуцент зрелого интерферона альфа-2а человека для осуществления способа по п.1.
7. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3562 - продуцент зрелого интерферона альфа-2а человека для осуществления способа по п.1.
8. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3563 - продуцент зрелого интерферона альфа-2а человека для осуществления способа по п.1.
9. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3564 - продуцент зрелого интерферона альфа-2а человека для осуществления способа по п.1.
| CHU J | |||
| et al., Fermentation process optimization of recombinant Saccharomyces cerevisiae for the production of human interferon-alpha2a, Appl | |||
| Biochem | |||
| Biotechnol., 2003, v.111, n.3, p.129-138. | |||
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА КЛЕТКАМИ ДРОЖЖЕЙ, СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE-ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1998 |
|
RU2180003C2 |
| КАРАБЕЛЬСКИЙ A.B | |||
| и др | |||
| Изучение протеолитического расщепления химерных рекомбинантных белков, секретируемых метилотрофными | |||

