Изобретение относится к области медицинской биотехнологии и касается способа получения белка медицинского назначения E7-HSP70 путем микробиологического синтеза с использованием в качестве продуцента дрожжей Saccharomyces cerevisiae.
Белок E7-HSP70 является гибридным, он включает в свой состав последовательности онкобелка Е7 вируса папилломы человека (ВПЧ) 11 типа и белка теплового шока HSP70 бактерий Mycobacterium tuberculosis, 16 С-концевых аминокислотных остатков которого замещены на искусственную последовательность, включающую 6 остатков гистидина для облегчения очистки гибридного белка с использованием металло-хелатной аффинной хроматографии на Ni-NTA сорбенте [Porath et al., 1975].
Вирусы папилломы человека способны поражать все слои дифференцированного кожного или слизистого эпителия различных анатомических областей тела человека. Многочисленные штаммы ВПЧ классифицируют на штаммы высокого или низкого риска. Штаммы ВПЧ высокого риска вызывают внутриэпителиальные поражения, которые с течением времени способны прогрессировать в инвазивные карциномы, наиболее значимой из которых является рак шейки матки [McLaughlin-Drubin & Münger, 2009b]. Co штаммами ВПЧ низкого риска, такими как ВПЧ типа 6 или типа 11, ассоциируются большинство доброкачественных образований, известных как бородавки в случае, если они обнаруживаются на коже, или как кондиломы при поражении генитальных областей тела [Howley, 1996; Laimins, 1993; zur Hausen, 2000].
Входящий в состав гибридного белка E7-HSP70 белок Е7 играет центральную роль в жизненном цикле вирусов ВПЧ, который тесно ассоциирован с процессом дифференцировки инфицированных клеток. Для собственного воспроизводства ВПЧ используют терминально дифференцированные клетки эпителия [обзор Lee et al., 2007], в которых в норме блокирован синтез ДНК, что делает их неспособными поддерживать репликацию ВПЧ. Чтобы обеспечить собственное развитие, ВПЧ вынуждены разобщать между собой процессы клеточной дифференцировки и пролиферации [McLaughlin-Drubin & Munger, 2009а]. Решающую роль в этом процессе играет вирусный белок Е7 [Cheng et al., 1995]. Доказано, что несмотря на возможные штамм-специфические различия в экспрессии, белок Е7 необходим для эписомной репликации и продолжения продуктивной фазы развития вируса [Oh et al., 2004].
По причине постоянного присутствия в инфицированных клетках, а также поскольку Е7 является белком исключительно вирусного происхождения, отсутствующим в неинфицированных клетках, он является идеальной мишенью для разработки высокоспецифичных способов иммунотерапии папилломатозов, направленных на их предотвращение и лечение. При этом разрабатываются и терапевтические вакцины, нацеленные на активацию Т-клеточного иммунного ответа против антигенов ВПЧ, экспрессируемых в инфицированных клетках, и профилактические вакцины, индуцирующие выработку нейтрализующих антител [Kanodia et al., 2008].
В процессе разработки вакцин против ВПЧ показано, что иммуногенность белка Е7 в составе терапевтических препаратов может быть увеличена за счет его соединения с белком-носителем, стимулирующим Т-клеточный ответ. Таким белком служит, в частности, белок теплового шока HSP70 Mycobacterium tuberculosis, в составе которого иммунные системы животных способны распознавать многочисленные В и Т клеточные эпитопы, что позволяет ему выполнять функцию мощного неспецифического стимулятора иммунной системы [Qian et al., 2006]. Показано, что для того чтобы стимулировать полноценный иммунный ответ, Е7 должен располагаться на N-конце относительно белка HSP70 [Li et al, 2006].
Известен способ получения вакцины HspE7, которая была использована компанией Stressgen для лечения рецидивирующего респираторного папилломатоза (РРП), вызываемого вирусами низкого риска, в основном 6 и 11 типа [US 6524825]. Несмотря на то, что основным компонентом вакцины является белок Е7 ВПЧ типа 16, а не ВПЧ типа 11, являющийся наиболее частым индуктором РРП, клинические испытания вакцины дали положительный результат. [Derkay et al., 2005].
В качестве ближайшего аналога заявляемого способа рассмотрим разработанный отечественной компанией ООО «Аванген» способ получения препарата на основе гибридного белка E7-HSP70, состоящего из онкобелка Е7 ВПЧ 11 типа и белка HSP70 Mycobacterium tuberculosis [RU 2290204]. Выбор в качестве мишени белка Е7 ВПЧ 11, а в качестве иммуномодулятора - белка HSP70 - определил улучшение иммунологических характеристик разработанного препарата по отношению к препарату HspE7 компании Stressgen. Дозировка белка E7-HSP70 в препарате «Аванген» предпочтительно составляет от 100 до 2500 мкг, при этом препарат обладает высокой степенью стандартности и слабой реактогенностью.
Согласно ближайшему аналогу биосинтез целевого белка осуществляют в клетках E.coli путем введения плазмиды pQE30-E711-dnaK. Белок получают в виде телец включения. После разрушения клеток бактерий тельца включения отмывают, для извлечения из них целевого белка и восстановления его структуры и растворимости используют процедуру денатурации-ренатурации. Исходя из описания RU 2290204 можно предположить, что отсутствие в процессе получения белка E7-HSP70 специфических стадий очистки препарата от комплекса липополисахаридов (ЛПС) будет приводить к тому, что препараты белка E7-HSP70, получаемые согласно описанной процедуре, будут содержать значительное и варьирующее от образца к образцу количество бактериального ЛПС, обладающего токсичным и пирогенным действием. Тем самым применение в лечебных целях белка E7-HSP70, синтезированного в клетках E.coli, будет затруднено. Введение стадии очистки препаратов от ЛПС может существенно удорожить производство. В этой связи разработка альтернативного способа получения белка E7-HSP70, обеспечивающего постоянство его конформации и состава и исключающего наличие в препарате токсичного комплекса ЛПС, является актуальной задачей.
Обеспечить выполнение этих требований позволяет использование дрожжей Saccharomyces cerevisiae в качестве альтернативных продуцентов белка E7-HSP70. Эти дрожжи, входящие в список микроорганизмов, безопасных для человека, не содержат токсичный и пирогенный бактериальный ЛПС [Gordonova et al., 2002]. В настоящее время их используют для производства 28 разрешенных к применению в медицине белков, включая вакцины [Ferrer-Miralles et al., 2009]. Однако до появления настоящего изобретения никому не удавалось осуществить сверхэкспрессию белков семейства HSP70 в клетках дрожжей S. cerevisiae [Wegele et al., 2003].
Сведений о штаммах S.cerevisiae - продуцентах белка E7-HSP70 в источниках информации не обнаружено.
Задача заявляемой группы изобретений:
- разработка способа получения белка медицинского назначения E7-HSP70, включающего в свой состав последовательности онкобелка Е7 ВПЧ 11 типа и белка теплового шока HSP70 М.tuberculosis, 16 С-концевых аминокислотных остатков которого замещены на искусственную последовательность, включающую 6 остатков гистидина, с использованием рекомбинантных дрожжей S.cerevisiae.
Задача решена путем:
- разработки способа получения белка медицинского назначения E7-HSP70 путем культивирования продуцента этого белка - дрожжей Saccharomyces cerevisiae, включающих последовательность ДНК, кодирующую слитый белок SEQ ID NO:1, составными частями которого являются аминокислотные последовательности белка E7-HSP70 и белка убиквитина дрожжей S.cerevisiae, занимающего в составе слитого белка N-концевое положение и содержащего сайт процесинга, распознаваемый природными убиквитин-специфичными протеиназами дрожжей, с последующим выделением и очисткой целевого белка (при этом в ходе экспрессии в дрожжах S.cerevisiae слитый белок подвергается процессингу, что обеспечивает биосинтез зрелого водорастворимого белка E7-HSP70, не содержащего убиквитиновый компонент);
- конструирования штамма дрожжей Saccharomyces cerevisiae ВКПМ Y-3853 - продуцента белка E7-HSP70.
Этапы конструирования заявляемого штамма
Этап 1
Конструируют дрожжевой вектор экспрессии, несущий в своем составе рекомбинантную ДНК, кодирующую слитый белок, включающий в свой состав последовательность убиквитина дрожжей S.cerevisiae, располагающуюся на N-конце слитого белка, и последовательность рекомбинантного белка E7-HSP70, содержащего последовательности онкобелка Е7 ВПЧ 11 типа и белка теплового шока Hsp70 M.tuberculosis, содержащего на С-конце последовательность, включающую 6 остатков гистидина.
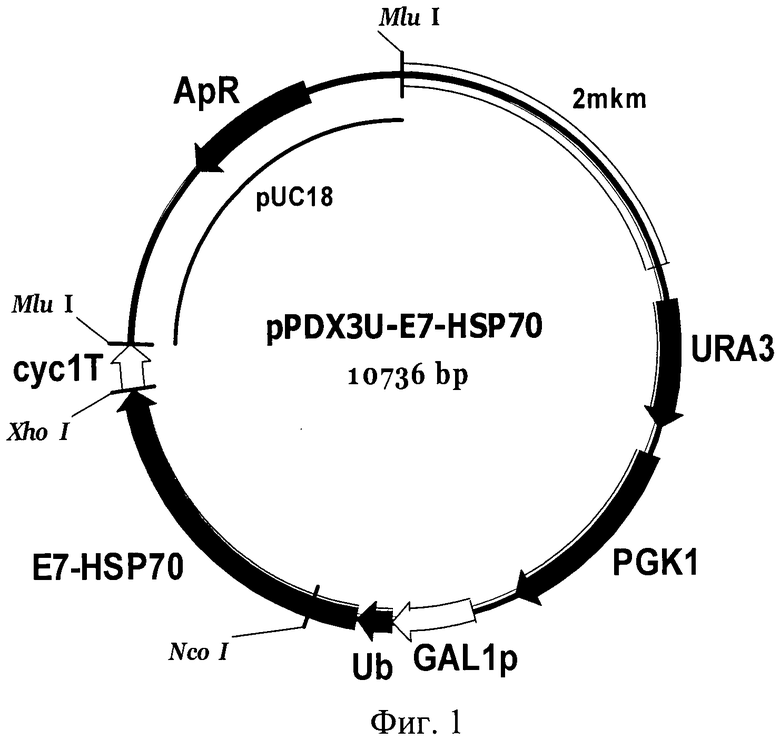
Сконструированный вектор экспрессии pPDX3U-E7-HSP70 (Фиг.1), содержит область инициации репликации эндогенной 2-мкм плазмиды дрожжей S.cerevisiae, что обеспечивает ему способность поддерживаться в клетках дрожжей S. cerevisiae в эписомном многокопийном состоянии.
Вектор pPDX3U-E7-HSP70 получают путем клонирования последовательности структурного гена белка E7-HSP70 в плазмиде pPDX3-GAL1U, несущей в своем составе под контролем промоторной области GAL1 дрожжей ген убиквитина дрожжей S.cerevisiae, причем в результате клонирования осуществляют слияние в одной рамке считывания последовательности структурного гена белка E7-HSP70 и гена убиквитина.
Этап 2
Конструируют заявляемый штамм S. cerevisiae - продуцент гибридного белка Е7-HSP70.
В качестве реципиентного используют лабораторный штамм S.cerevisiae D702 (а/α leu2/leu2 ura3/ura3 trp1/trp1 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 pgk1:URA3/pgk1:URA3 GAL4::(STA2p-GAL4, TRP1)/GAL4::(STA2p-GAL4, TRP1) STA2/STA2 suc0/SUC2), который является диплоидным, что обеспечивает повышенную стабильность его экспрессионных характеристик. Штамм S.cerevisiae D702 содержит гомозиготные мутации в хромосомных аллелях структурного гена PGK1, кодирующего фосфоглицерат киназу, что обеспечивает стабильное поддержание вектора, несущего структурный ген PGK1, на средах, содержащих любой единственный источник углерода, усваиваемый дрожжами S.cerevisiae, и гена GAL80, кодирующего белок - репрессор промотора GAL1, а также гомозиготную мутацию, приводящую к изменению регуляции гена GAL4, кодирующего белок - активатор промотора GAL1, вследствие чего осуществляется глюкозо-регулируемая экспрессия генов, находящихся под контролем промотора GAL1 [Kozlov et al., 1996].
Заявляемый штамм конструируют путем трансформации реципиентного штамма S.cerevisiae D702 вектором экспрессии pPDX3U-E7-HSP70. В результате трансформации получают штамм, продуцирующий рекомбинантный белок E7-HSP70, который депонируют во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) как штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3853.
Характеристика заявляемого штамма
Генотип заявляемого штамма:
ВКПМ Y-3853 (а/α leu2/leu2 ura3/ura3 trp1/trp1 gal80::LEU2/gal80::LEU2 lys7/LYS7 his3/HIS3 his4/HIS4 pgk1:URA3/pgk1:URA3 GAL4::(STA2p-GAL4,TRP1)/GAL4::(STA2p-GAL4, TRP1) STA2/STA2 suc0/SUC2)/pPDX3U-E7-HSP70
Заявляемый штамм характеризуется следующими культурально-морфологическими и физиолого-биохимическими признаками:
Морфологические признаки:
При культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (в мас.%): пептон 2, дрожжевой экстракт 1, глюкоза 2, агар 2, вода остальное, клетки имеют овальную форму, 3-7 мкм в диаметре. Клетки почкуются. Почкование истинное, многостороннее. Истинного мицелия не образует.
Колонии имеют следующий вид:
1) на агаризованной среде YPD колонии белого цвета с ровным краем, матовой поверхностью, линзовидным профилем и сметанообразной консистенцией;
2) на агаризованной среде с крахмалом (состав в мас.%: пептон 2, дрожжевой экстракт 1, крахмал 1, агар 2, вода остальное) колонии белого цвета с узорчатым краем, матовой поверхностью, линзовидным профилем и крупчатой консистенцией.
Рост в жидкой среде с крахмалом: при 28°С в течение первых 24 ч культивирования - жидкость мутная, осадок белый, не комкуется, пристеночных пленок не образует.
Физико-химические признаки: факультативный анаэроб, температура роста 18-35°С (оптимум 28°С), рН культивирования 3,8-7,4 (оптимум 5,0).
Ассимиляция источников углерода: сбраживает глюкозу, фруктозу, мальтозу, сахарозу, декстрины, крахмал. Не сбраживает лактозу, галактозу, инулин, ксилозу, арабинозу.
Ассимиляция источников азота: усваивает аминокислоты, сернокислый аммоний, азотнокислый аммоний.
Патогенность: штамм Saccharomyces cerevisiae ВКПМ Y-3853 непатогенен.
Хранение: Штамм хранят при температуре -70°С в 20% водном растворе глицерина. Возможно хранение на агаризованной богатой среде с глюкозой в течение 3 месяцев при +4°С.
Стабильность: штамм сохраняет характеристики при 20 последовательных пересевах на агаризованной среде YPD при температуре 28°С.
Продукция белка E7-HSP70: при культивировании в комплексной среде YPD, содержащей менее 0,2% глюкозы, продуцирует гибридный белок E7-HSP70.
Способ в общем виде
Для засева ферментера получают посевную культуру штамма S. cerevisiae -продуцента белка E7-HSP70.
Эту культуру получают путем культивирования штамма-продуцента белка E7-HSP70 в течение 16-40 часов при температуре 22-32°С на орбитальной качалке 100-350 об/мин в среде следующего состава, в мас.%: пептон 1-3; дрожжевой экстракт 0,5-3; глюкоза 1-3, вода остальное, рН среды - естественный.
Посевной культурой засевают ферментер (0,5-1000 л), содержащий ферментационную среду следующего состава, в мас.%: пептон 1-3; дрожжевой экстракт 0,5-3, глюкоза или сахароза 1-4, вода остальное, рН среды - естественный. Количество посевной культуры, вносимой в ферментер составляет 3-15% от объема ферментационной среды.
Ферментацию проводят при 22-32°С, аэрации 0,5-1000 л/мин и скорости перемешивания культуры 100-1500 оборотов/мин. Общая продолжительность ферментации составляет 40-66 часов.
Уровень биосинтеза гибридного белка E7-HSP70 в таких условиях составляет не менее 100 мг/л.
Выделение и очистку целевого продукта осуществляют путем последовательного выполнения следующих процедур:
- разрушения клеток дрожжей в «буфере для разрушения», не содержащем ЭДТА, с использованием подходящего гомогенизатора, например, FastPrep-24 (MP Biomedicals LLC, USA);
- отделения фракции растворимых внутриклеточных белков путем отделения клеточных обломков центрифугированием, например, в течение 10 минут при ускорении 10000 g;
- экстрагирования белка E7-HSP70 из фракции растворимых внутриклеточных белков путем специфического связывания на колонке с Ni-NTA агарозой и последующей элюцией раствором имидазола (например, с использованием набора Qiagen cat. #31314);
- переводом экстрагированного белка в подходящий «буфер для хранения» (например, с использованием колонки GE Healthcare PD10).
Выход белка, получаемого заявляемым способом, составляет не менее 10 мг/10 г влажной клеточной биомассы. Чистота получаемого препарата составляет не менее 90%.
Заявляемое изобретение проиллюстрировано следующими фигурами:
Фиг.1. Карта вектора pPDX3U-E7-HSP70.
Обозначения: E7-HSP70 - структурный ген рекомбинантного белка E7-HSP70; HUB - ген убиквитина дрожжей S.cerevisiae; GAL1 - промоторная область гена GAL1 дрожжей S.cerevisiae; URA3 и PGK1 - структурные гены URA3 и PGK1 дрожжей S.cerevisiae, соответственно; cyc1T - последовательность терминатора транскрипции гена CYC1 дрожжей S.cerevisiae; 2 mkm - фрагмент эндогенной 2-микронной плазмиды дрожжей S.cerevisiae, содержащий область начала репликации; pUC18 - фрагмент плазмиды pUC18, содержащий ген бета-лактамазы (ApR) и область начала репликации, обеспечивающий селективную амплификацию вектора в клетках E.coli.
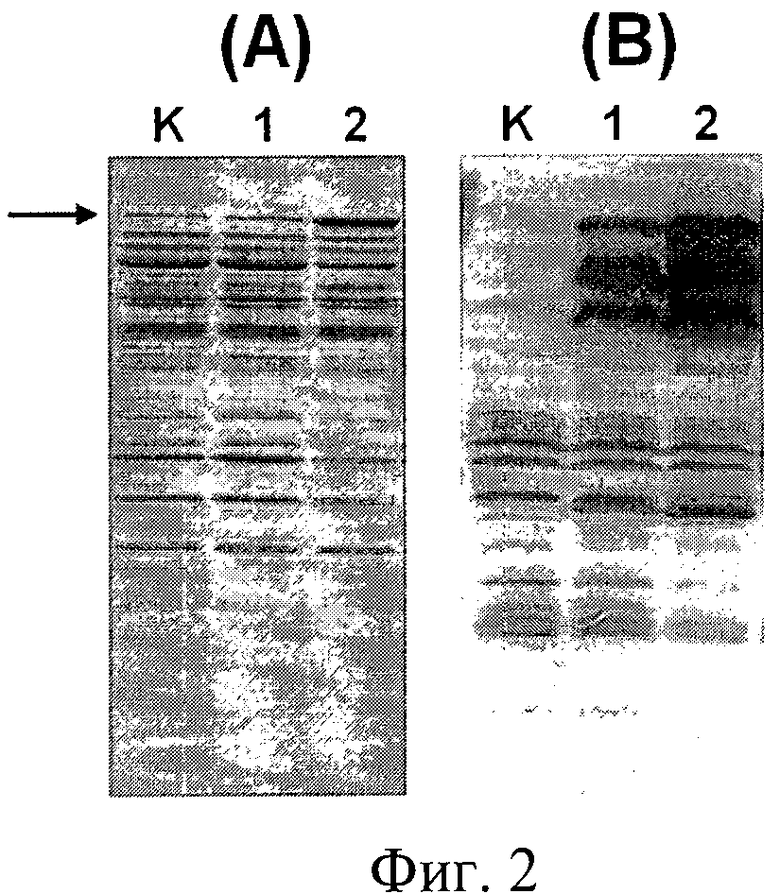
Фиг.2. Электрофореграмма (А) и Western-анализ (В) внутриклеточных белков штаммов дрожжей. На дорожки нанесены внутриклеточные белки штаммов: K - D702/pPDX3, отрицательный контроль; 1 - D702/pPDX3-E7-HSP70; 2 - ВКПМ Y-3853. Для Western-анализа использовали антитела MAb TS31, специфичные HSP70 (пример 10). Стрелкой показано положение полноразмерного белка E7-HSP70.
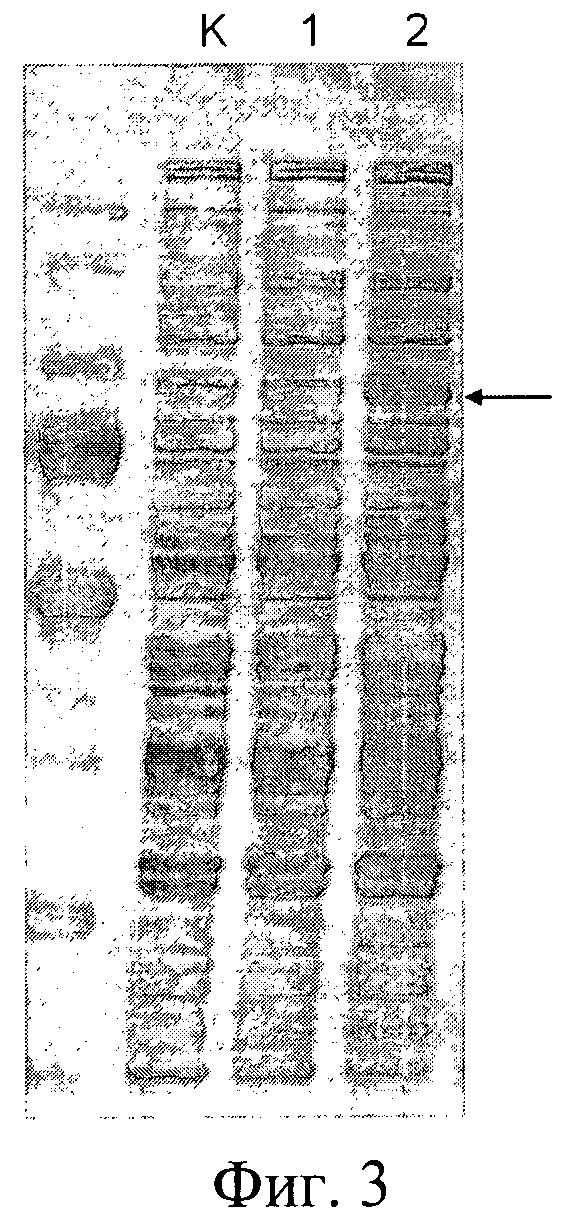
Фиг.3. Электрофореграмма внутриклеточных белков штаммов дрожжей. На дорожки нанесены внутриклеточные белки штаммов: К - YBS618/pPDX3, отрицательный контроль; 1 - YBS618/pPDX3-E7-HSP70; 2 - YBS618/pPDX3U-E7-HSP70. Стрелкой показано положение полноразмерного белка E7-HSP70.
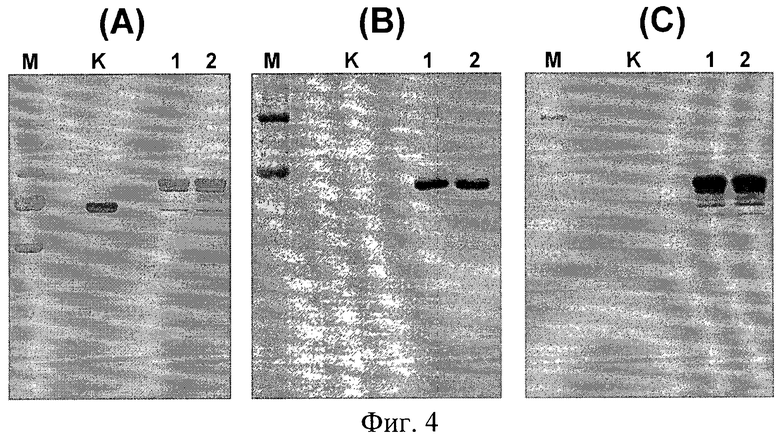
Фиг.4. Электрофореграмма (А) и Western-анализ (В и С) белков E7-HSP70, очищенных из клеток штамма дрожжей ВКПМ Y-3853 с помощью металл-хелатной хроматографии (пример 8). На дорожки нанесены: М - маркеры молекулярного веса (SM0671, Fermentas), K - бычий сывороточный альбумин, 0,5 мкг, отрицательный контроль; 1 - аналитический образец белка E7-HSP70 (0,4 мкг), очищенный методом очистки "batch" (пример 10); 2 - образец белка E7-HSP70 (0,4 мкг), полученный с использованием препаративных методов выделения и очистки (пример 12). Для Western-анализа использовали антитела (В) MAb 711-13, специфичные к белку Е7, и (С) MAb TS31, специфичные к белку HSP70 (пример 10).
Пример 1. Конструирование последовательности ДНК, кодирующей белок E7-HSP70
Конструирование проводят с использованием плазмиды pQE30-E711-dnaK, несущей структурные гены белка Е7 ВПЧ-11 и белка теплового шока Hsp70 из Mycobacterium tuberculosis [RU2290204], а также лабораторной плазмиды pUC19MX-His6, содержащей EcoRI/XhoI фрагмент ДНК, кодирующий аминокислотную последовательность из 6 остатков гистидина. Для этого:

2) ДНК плазмиды pQE30-E711-dnaK расщепляют по сайтам BamHI и KpnI. Из полученной смеси фрагментов рестрикции изолируют фрагмент ДНК размером 1614 пар оснований, кодирующий протяженную 5'-концевую часть структурного гена белка теплового шока Hsp70 из М.tuberculosis, и называют его фрагментом-2;

4) ДНК плазмиды лабораторной плазмиды pUC19MX-His6 расщепляют по сайтам BglII и EcoRI. Из полученной смеси фрагментов рестрикции изолируют фрагмент ДНК, представляющий собой векторную часть pUC19MX-His6, содержащей EcoRI/XhoI фрагмент ДНК, кодирующий последовательность из 6 остатков гистидина, и называют фрагментом-4.
Затем в результате совместного лигирования четырех полученных фрагментов ДНК получают плазмиду pUC19MX-E7-HSP70.
Полученная плазмида pUC19MX-E7-HSP70 содержит ген рекомбинантного белка E7-HSP70, состоящего из онкобелка Е7 ВПЧ 11 типа и белка теплового шока Hsp70 из M.tuberculosis и содержащего на С-конце последовательность, включающую 6 остатков гистидина.
Пример 2. Конструирование экспрессионного вектора рРРХ3-GAL1
Конструирование проводят с использованием лабораторного вектора pPDX3. Вектор pPDX3 отличается от вектора pPDX2 [RU2010123696] тем, что включает вариант структурного гена URA3, содержащий мутацию в последовательности сайта NcoI (ccatgg изменен на ccatga), не приводящую к инактивации гена URA3. Конструирование проводят следующим образом. HindIII/XhoI фрагмент ДНК вектора pPDX3 лигируют с HindIII/XhoI фрагментом ДНК лабораторной плазмиды pUC18x-GAL1, заключающим нуклеотидную последовательность промотора гена GAL1 дрожжей S. cerevisiae, слитую с нуклеотидной последовательностью открытой рамки считывания, содержащей сайты BamHI и XhoI как показано ниже (сайты подчеркнуты).


В результате клонирования получают экспрессионный вектор pPDX3-GAL1U, ДНК которого содержит нуклеотидную последовательность, кодирующую убиквитин дрожжей S.cerevisiae, слитую с нуклеотидной последовательностью промоторной области гена GAL1 дрожжей S.cerevisiae.
Пример 4. Конструирование экспрессионного вектора pPDX3-E7-HSP70
BglII/XhoI фрагмент ДНК плазмиды pUC19MX-E7-HSP70, заключающий структурный ген белка E7-HSP70, включающего аминокислотные последовательности онкобелка Е7 ВПЧ 11 типа и белка теплового шока Hsp70 M tuberculosis, содержащего на С-конце последовательность, включающую 6 остатков гистидина, лигируют с ДНК экспрессионного вектора pPDX3-GAL1, расщепленного по сайтам BamHI и XhoI.
В результате лигирования получают экспрессионный вектор pPDX3-E7-HSP70, включающий в свой состав последовательность структурного гена белка E7-HSP70 под контролем промотора гена GAL1 дрожжей S.cerevisiae.
Пример 5. Конструирование экспрессионного вектора pPDX3U-E7-HSP70
Конструирование проводят, как описано в примере 4, за исключением того, что ген белка E7-HSP70 клонируют в экспрессионный вектор pPDX3-GAL1U.
В результате клонирования получают экспрессионный вектор pPDX3U-E7-HSP70 (Фиг.1), включающий последовательность структурного гена рекомбинантного белка E7-HSP70, слитую с последовательностью гена убиквитина дрожжей S.cerevisiae под контролем промотора гена GAL1 дрожжей S.cerevisiae.
Пример 6. Конструирование заявляемого штамма
Штамм ВКПМ Y-3853 получают в результате трансформации лабораторного штамма D702 экспрессионным вектором pPDX3U-E7-HSP70. Для трансформации клетки штамма D702 подращивают в течение 24 часов при температуре 28°С на агаризованной среде YPGE, следующего состава в мас.%: бактопептон 2, дрожжевой экстракт 1, бактоагар 2, этанол 2, глицерин 3, вода остальное. Трансформацию выращенных клеток штамма D702 проводят по методу Ito [Ito et al., 1983]. Трансформанты отбирают по способности расти на среде YPD следующего состава, в мас.%: бактопептон 2, дрожжевой экстракт 1, глюкоза 2, бактоагар 2, вода остальное. Один из полученных трансформантов называют штаммом S.cerevisiae ВКПМ Y-3853.
Пример 7. Конструирование штамма S.cerevisiae SCR-702-E7-HSP70
Штамм SCR-702-E7-HSP70 получают в результате трансформации лабораторного штамма D702 экспрессионным вектором pPDX3-E7-HSP70. Трансформацию осуществляют как в примере 6. Один из полученных трансформантов называют штаммом S.cerevisiae SCR-702-E7-HSP70.
Пример 8. Конструирование штамма S. cerevisiae YBS618U-E7-HSP70
Штамм YBS618U-E7-HSP70 получают в результате трансформации лабораторного штамма S.cerevisiae Y618 [Kartasheva et al., 1996] экспрессионным вектором pPDX3U-E7-HSP70. Трансформацию осуществляют как в примере 6, за исключением того, что трансформанты отбирают по способности расти на синтетической среде с добавлением аминокислот гистидина, лейцина и лизина до конечной концентрации 30 мкг/л. Состав синтетической среды, в мас.%: KH2PO4 0,1, MgSO4 0,05, NaCl 0,01, CaCl2 0,01, (NH4)2SO4 0,35, глюкоза 2, тиамин (витамина В1) 0,02, рибофлавин (витамина В2) 0,02, никотиновая кислота (витамина РР) 0,02, п-аминобензойная кислота 0,02, пантотенат кальция 0,02, биотин 0,0002, пиридоксин (витамина В6) 0,02, инозит 1, фолиевая кислота 0,02, вода остальное.
Один из полученных трансформантов называют штаммом S.cerevisiae YBS618U-E7-HSP70.
Пример 9. Конструирование штамма S.cerevisiae YBS618-E7-HSP70
Штамм YBS618-E7-HSP70 получают в результате трансформации лабораторного штамма S. cerevisiae Y618 [Kartasheva et al., 1996] экспрессионным вектором pPDX3-E7-HSP70. Трансформацию осуществляют как в примере 8. Один из полученных трансформантов называют штаммом S.cerevisiae YBS618-E7-HSP70.
Пример 10. Оценка уровня экспрессии белка E7-HSP70 в клетках сконструированных штаммов
Клетки дрожжевых штаммов, содержащих ген белка E7-HSP70, включающий убиквитиновую компоненту: заявляемого ВКПМ Y-3853 и S. cerevisiae YBS618U-E7-HSP70, а также не содержащих убиквитиновую компоненту штаммов: SCR-702-E7-HSP70 (контроль 1) и YBS618-E7-HSP70 (контроль 2), культивируют в колбах при 30°С на ротационной качалке со скоростью 250 об/мин на жидкой среде YPD следующего состава, мас.%: бактопептон 2, дрожжевой экстракт 1, глюкоза 2, вода остальное, засевая в титре 5×105-5×106 мл-1. Образцы для анализа отбирают через 46 часов роста культуры. Конечная оптическая плотность культур составляет ОД600=40-45. Клетки дрожжей отделяют от среды культивирования центрифугированием при 12000 g в течение 2 мин в пробирке на 1,5 мл. Осадок клеток, собранный из 1.5 мл стационарной культуры, суспендируют в "буфере для разрушения" (50 мМ фосфата натрия, 100 мМ NaCl, 10 мМ имидазол, 1 мМ ЭДТА, 2 мМ PMSF, 0.1% Tween-20, pH8.0) из расчета 500 мкл буфера на ~100 мкл влажного осадка клеток. Разрушение клеток осуществляют с помощью стеклянных шариков (d=0,45-0,65 мм) на встряхивателе для пробирок типа «Вортекс». Для этого ~300 мг шариков добавляют к суспензии клеток, смесь встряхивают 10 раз по 1 мин, с охлаждением во льду, к содержимому пробирок добавляют 500 мкл «буфера для разрушения», и встряхивание повторяют еще 60 сек. Содержимое пробирок центрифугируют в течение 10 мин при 16000 g. Надосадочную жидкость, содержащую водорастворимые клеточные белки дрожжей, используют для анализа уровня продукции белка E7-HSP70.
Электрофоретический анализ образцов водорастворимых клеточных белков дрожжей проводят в 10% ПААГ-ДДС-Na в восстанавливающих условиях по стандартной процедуре Лэммли [Laemmli, 1970] с использованием системы Mini-PROTEAN Tetra Cell (#165-8000). Приготовление буферов, подготовку образцов, нанесение и проведение электрофореза выполняют согласно инструкции "BIO-RAD", прилагаемой к системе. В качестве маркеров используют смесь предокрашенных белков с известной молекулярной массой в диапазоне от 10 до 170 kDa (реагент фирмы "Fermentas", PageRuler Prestained Protein Ladder, #SM0671). Окрашивание геля нитратом серебра проводят при помощи кита фирмы "Fermentas" (PageSilver Silver Staining Kit, #K0681) согласно прилагаемой инструкции.
Вестерн-блот анализ образцов водорастворимых клеточных белков дрожжей проводят следующим образом. Сначала, после проведения электрофореза (как описано выше), белки переносят на нитроцеллюлозную мембрану ("Sigma", #Z361380) при помощи электропереноса, проводимого согласно методике фирмы "BIO-RAD" с использованием системы Mini Trans-Blot Electrophoretic Transfer Cell, #170-3930. Качество переноса контролируют полным переносом преокрашенных белковых маркеров из геля на мембрану. На втором этапе мембрану инкубируют (все инкубации и отмывки мембраны проводят при комнатной температуре на качалке) 1 час в блокирующем растворе (1% BSA в TBS-T буфере: 20 mM Tris-HCl, 150 mM NaCl, 0.1% Tween-20, рН 7.5), чтобы исключить неспецифическое связывыние антител с пустыми местами мембраны. Затем мембрану в течение 1 ч инкубируют с моноклональными мышиными антителами (разведение 1:2000 в TBS-T буфере), специфичными либо к белку Е7 (MAb 711-13, распознающие эпитоп 10-18 ак), либо к белку HSP70 (MAb TS31 или MAb TS29, распознающие N-конец или середину белка соответственно). Для этого используют моноклональные антитела фирмы HyTest Ltd, Finland. После отмывки мембраны от несвязавшихся антител в буфере TBS-T 6×5 мин, к мембране добавляют козьи антимышиные поливалентные антитела ("Sigma", A-0412), коньюгированные с пероксидазой хрена (разведение 1:5000 в TBS-T буфере), и инкубируют 1 ч. После тщательной отмывки мембраны от вторичных антител (2×10 мин, 6×5 мин), ее проявляют с использованием кита Pierce ECL Western Blotting Substrate ("Pierce", #32106) согласно прилагаемой инструкции. Фотографии блота получают на рентгеновской пленке.
Аналитическую очистку рекомбинантного белка E7-HSP70 из фракции водорастворимых белков сконструированных штаммов дрожжей проводят с использованием металло-хелатной аффинной хроматографии [Porath et al.., 1975, Nature, 258: 598-599]. Используют реагент Ni-NTA Agarose (QIAGEN, #30210) и методику очистки «batch» согласно прилагаемому протоколу.
Анализ АТФ-связывающей активности белка HSP70 в составе рекомбинантного белка E7-HSP70 проводят с использованием реагента ATP-Agarose (Sigma, #A2767) согласно инструкции, прилагаемой к реагенту.
Сравнительный электрофоретический анализ (Фиг.2 и Фиг.3) и Вестерн-анализ экспрессии целевого белка E7-HSP70 (Фиг.2) в клетках сконструированных штаммов дрожжей S.cerevisiae свидетельствует о том, что:
- Целевой белок накапливается в клетках штаммов дрожжей во фракции водорастворимых белков;
- Целевой белок, экспрессируемый в клетках всех анализируемых штаммов, обладает одинаковой электрофоретической подвижностью, которая соответствует молекулярному весу белка E7-HSP70 (78 кДа), не содержащего убиквитиновую компоненту;
- Уровень накопления целевого белка в клетках заявляемого штамма ВКПМ Y-3853 и штамма YBS618U-E7-HSP70, сконструированных для осуществления заявляемого способа, в 7-10 раз превышает уровень накопления целевого белка контрольными штаммами SCR-702-E7-HSP70 и YBS618-E7-HSP70, не содержащими убиквитиновую компоненту.
Результаты Вестерн-анализа препаратов очищенного целевого белка из клеток заявляемого штамма (Фиг.4) свидетельствуют о специфическом взаимодействии очищенного целевого белка с антителами против белка Е7 и с антителами против белка HSP70, что доказывает наличие этих белков в его составе.
Способность целевого белка к хроматографической очистке на Ni-NTA сорбенте подтверждает наличие в его составе последовательности, включающей 6 остатков гистидина, а взаимодействие с АТР-агарозой подтверждает нативную конформацию белка HSP70.
Пример 11. Микробиологический синтез белка E7-HSP70 в клетках заявляемого штамма
Для получения посевного материала клетки штамма ВКПМ Y-3853 выращивают в среде YPD на ротационной качалке со скоростью 250 об/мин при температуре 28°С в течение 24 часов. 50 мл посевного материала используют для засева 3-литрового ферментера Anglicon, содержащего 950 мл среды YPD. Ферментацию проводят при температуре 28°С, аэрации 1 л/мин и скорости перемешивания 1000 об/мин. Через 24 часа после засева ферментера начинают подпитку среды культивирования 50%-м раствором глюкозы со скоростью 2 мл/ч и устанавливают рН-статирование культуры на уровне рН 6.8±0.1, используя для подтитровки растворы 10% серной кислоты и 10% NaOH. Общее время ферментирования составляет 72 часа. По окончании культивирования клеточную биомассу, содержащую рекомбинантный белок E7-HSP70, отделяют от культуральной жидкости с помощью центрифугирования 9000 g в течение 15 минут при температуре 8°С.
По данным электрофоретического анализа уровень продукции рекомбинантного белка E7-HSP70 составляет 100 мг/л среды.
Пример 12. Выделение и очистка целевого белка E7-HSP70, синтезированного в клетках заявлемого штамма
10 г биомассы дрожжей, полученных как описано в примере 10, смешивают с 20 мл «буфера для разрушения», не содержащего ЭДТА, и 10 мл стеклянных шариков (пример 10). Разрушение клеток дрожжей проводят в течение 1 минуты при максимальной мощности на дезинтеграторе FastPrep-24 (MP Biomedicals LLC, USA). Лизат клеток дрожжей центрифугируют при 4°С сначала 10 минут 9000 g, затем 30 минут 30000 g. Осадок отбрасывают.
Надосадочную жидкость наносят на колонку, содержащую 1.5 мл Ni-NTA агарозы (Qiagen), уравновешенную «буфером для разрушения» без ЭДТА, со скоростью 0.25 мл/мин на холоду. По окончании нанесения колонку промывают «буфером для нанесения» без ЭДТА до тех пор, пока оптическая плотность промывочного буфера не достигнет фонового значения.
Элюцию белка E7-HSP70 с колонки проводят буфером следующего состава, мас.%: NaH2PO4 - 0.78, NaCl - 1.75, имидазол - 1.7, Твин-20 - 0,05, вода - остальное, рН 8.5. По окончании элюции в раствор белка E7-HSP70 вносят ЭДТА до концентрации 1 мМ.
Элюированный белок E7-HSP70 переводят в буфер для хранения (50 мМ фосфата натрия, 100 мМ NaCl, 1 мМ ЭДТА, рН 8.5) для чего используют колонку PD10 (GE Healthcare). Концентрация белка в буфере для хранения составляет 1.2±0.3 мг/мл. Выход очищенного белка составляет 12 мг/10 г влажной клеточной биомассы, а чистота - 92%.
Таким образом, заявляемый способ обеспечивает:
- осуществление биосинтеза белка E7-HSP70 в клетках дрожжей Saccharomyces cerevisiae, входящих в список микроорганизмов, безопасных для человека (GRAS, generally regarded as safe);
- высокий уровень биосинтеза и выход очищенного белка E7-HSP70 на уровне не менее 10 мг/10 г влажной клеточной биомассы, с чистотой - не менее 90%;
- получение водорастворимого корректно процессированного белка E7-HSP70, не требующего применения корректирующих процедур растворения и денатурации-ренатурации, приводящих к формированию различных укладок белка, в том числе и отличных от природных;
- принципиальное отсутствие в препаратах белка E7-HSP70 токсичного и пирогенного ЛПС бактерий.
Источники информации


| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИБРИДНОГО БЕЛКА Е7-HSP70 (ВАРИАНТЫ) | 2013 |
|
RU2546917C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ, СЛИТЫЙ БЕЛОК, РЕКОМБИНАНТНАЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И ШТАММЫ-ПРОДУЦЕНТЫ | 2010 |
|
RU2451023C1 |
| СПОСОБ ПОЛУЧЕНИЯ СУСПЕНЗИЙ ГИДРОГЕЛЕВЫХ МИКРОЧАСТИЦ С ЗАДАННЫМИ РАЗМЕРАМИ НА ОСНОВЕ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2478706C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА | 2011 |
|
RU2460795C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE И ШТАММ-ПРОДУЦЕНТ ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2446172C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА, ШТАММ Saccharomyces cerevisiae - ПРОДУЦЕНТ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2010 |
|
RU2427645C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ ШТАММОВ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫХ ПРОДУЦЕНТОВ СОМАТОТРОПИНА ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫЙ ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2009 |
|
RU2420567C1 |
| ГИБРИДНЫЙ БЕЛОК, ОБЛАДАЮЩИЙ ПРОЛОНГИРОВАННЫМ ДЕЙСТВИЕМ, НА ОСНОВЕ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ), СПОСОБ ЕГО ПОЛУЧЕНИЯ И ШТАММ Saccharomyces cerevisiae ДЛЯ ОСУЩЕСТВЛЕНИЯ ЭТОГО СПОСОБА (ВАРИАНТЫ) | 2013 |
|
RU2515913C1 |
| КАССЕТА И РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЗКСПРЕССИИ И СЕКРЕЦИИ МЕХАНОЗАВИСИМОГО ФАКТОРА РОСТА ЧЕЛОВЕКА (MGF), ШТАММ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ MGF И СПОСОБ ПОЛУЧЕНИЯ MGF | 2006 |
|
RU2344173C2 |
| Фосфолипаза А2 для экспрессии в дрожжах (варианты) | 2018 |
|
RU2716087C1 |
Группа изобретений относится к области биотехнологии. Культивируют микроорганизм-продуцент в подходящей питательной среде с последующим выделением и очисткой целевого белка. При этом в качестве продуцента используют дрожжи Saccharomyces cerevisiae, трансформированные вектором экспрессии, который содержит область инициации репликации эндогенной 2-мкм плазмиды дрожжей Saccharomyces cerevisiae, а также промотор GAL1 дрожжей, контролирующий экспрессию гена, включающего последовательность ДНК SEQ ID NO:1, кодирующую слитый белок, составными частями которого являются аминокислотные последовательности белка E7-PISP70 и белка убиквитина дрожжей Saccharomyces cerevisiae, занимающего в составе слитого белка N-концевое положение, заканчивающегося сайтом процессинга, отделяющим его от последовательности белка E7-HSP70 и распознаваемым природными убиквитин-специфичными протеиназами дрожжей. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3853 - продуцент белка E7-HSP70 - получают путем трансформации вектором экспрессии pPDX3U-E7-HSP70 штамма дрожжей Saccharomyces cerevisiae D702. Группа изобретений обеспечивает высокий уровень биосинтеза и выход очищенного белка E7-HSP70, получение водорастворимого корректно процессированного белка E7-HSP70, принципиальное отсутствие в препаратах белка E7-HSP70 токсичного и пирогенного ЛПС бактерий. 2 н.п. ф-лы, 4 ил., 12 пр.
1. Способ получения белка E7-HSP70 путем культивирования микроорганизма-продуцента в подходящей питательной среде с последующим выделением и очисткой целевого белка, отличающийся тем, что в качестве продуцента используют дрожжи Saccharomyces cerevisiae, трансформированные вектором экспрессии, который содержит область инициации репликации эндогенной 2-мкм плазмиды дрожжей Saccharomyces cerevisiae, а также промотор GAL1 дрожжей, контролирующий экспрессию гена, включающего последовательность ДНК SEQ ID NO:1, кодирующую слитый белок, составными частями которого являются аминокислотные последовательности белка E7-HSP70 и белка убиквитина дрожжей Saccharomyces cerevisiae, занимающего в составе слитого белка N-концевое положение, заканчивающегося сайтом процессинга, отделяющим его от последовательности белка E7-HSP70 и распознаваемым природными убиквитин-специфичными протеиназами дрожжей.
2. Штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-3853 - продуцент белка E7-HSP70 для осуществления способа по п.1, полученный путем трансформации вектором экспрессии pPDX3U-E7-HSP70 штамма дрожжей Saccharomyces cerevisiae D702.
| РЕКОМБИНАНТНЫЙ ГИБРИДНЫЙ БЕЛОК, ПРЕПАРАТ ДЛЯ ИММУНОТЕРАПИИ НА ЕГО ОСНОВЕ И СПОСОБ ИММУНОТЕРАПИИ РЕЦИДИВИРУЮЩЕГО ПАПИЛЛОМАТОЗА ГОРТАНИ | 2005 |
|
RU2290204C1 |
| МУКОЗАЛЬНАЯ ВАКЦИНА ДЛЯ ИММУНОТЕРАПИИ ЗАБОЛЕВАНИЙ, ОБУСЛОВЛЕННЫХ ВИРУСАМИ ПАПИЛЛОМЫ ЧЕЛОВЕКА, И СПОСОБ ЛЕЧЕНИЯ С ЕЕ ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2008 |
|
RU2377305C1 |
| LI H et al | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Gynecol | |||
| Oncol | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| CHEN CH et al | |||
| Enhancement of DNA vaccine potency by | |||

