Настоящее изобретение относится к способам получения продуктов питания или напитков на основе солода, подходящих для потребления лицом, страдающим целиакией. В частности настоящее изобретение относится к способам получения продуктов питания или напитков на основе солода с низким содержанием гордеинов. Также предлагаются растения ячменя, дающие зерно, которое может применяться в способах изобретения.
Целиакия (глютеновая энтеропатия) (CD, называемая также целиакия-спру) является опосредованным T-клетками аутоиммунным заболеванием тонкой кишки, которая вызывается у восприимчивых людей при потреблении в пищу определенных запасных белков, известных в целом как проламины, из пшеницы (глютена, состоящего из глютенинов и глиадинов), ячменя (гордеинов) или ржи (секалинов). Проламины овса (авенины), по-видимому, переносятся большинством больных целиакией (Hogberg et al., 2004; Peraaho et al, 2004a), однако у меньшего числа больных целиакией они могут вызвать положительные реакции (Lundin et al. 2003; Peraaho et al. 2004b). CD встречается у приблизительно 0,25-1% населения, по меньшей мере, в Австралии, Северной и Южной Америке, Европе, Африке и Индии (Hovell et al. 2001; Fasano et al. 2003; Treem 2004), однако данное заболевание, вероятно, диагностируется на недостаточном уровне. Увеличение информированности по поводу симптомов и последствий неподвергавшихся лечению случаев CD в Австралии приводит к повышению количества диагностируемых случаев на 15% ежегодно. Приблизительно 1-4 европейцев и жителей западной Азии имеют аллели HLA-DQ8 или -DQ2, которые являются необходимой, но не достаточной детерминантой CD (Treem 2004). При этом лишь у приблизительно 1 - 20 человек с данными аллелями развивается CD. В настоящее время единственным лечением является полный отказ от пшеницы, ячменя и ржи, поскольку рецидивы могут быть вызваны потреблением всего лишь 10 миллиграммов глютена в день (Biagi et al., 2004).
В случае невыявления заболевания или отсутствия лечения, CD приводит к серьезным последствиям для здоровья, которые могут угрожать жизни, особенно у детей первого года жизни. CD вызывает деформацию абсорбирующих ворсинок тонкой кишки и может приводить к разрушению ворсинок. В результате питательные вещества плохо всасываются, причем это может быть связано с потерей веса, усталостью, минеральной недостаточностью, дерматитом и потерей ночного зрения, а также интенсивного кишечного расстройства, которое обычно включает вздутие, диарею и спазмы. Лица с нелеченной CD подвергаются повышенному риску развития рака, например, 10-кратному повышению риска возникновения карциномы тонкой кишки, 3-6-кратному повышению риска возникновения неходжкинской лимфомы и 28-кратному повышению риска возникновения Т-клеточной лимфомы кишечника. CD также представляет 3-кратное повышение риска развития диабета I типа (Peters et al., 2003; Peters et al., 2003; Verkarre et al., 2004). Также сообщали о пятикратном повышении частоты психической депрессии у пациентов с целиакией (Pynnonen et al., 2004).
Молекулярная основа целиакии в настоящее время довольно хорошо изучена (Sollid 2002; Hadjivassiliou et al. 2004) и представляет собой реакцию на определенную последовательность аминокислот в проламинах. Плохо переваренные пептиды проламинов, богатые пролином и глутамином, соответствуют субстратному мотиву, с которым в слизистой оболочке кишечника взаимодействует тканевая трансглутаминаза (tTG) человека, которая дезамидирует ключевые остатки глутамина. Образующаяся в результате отрицательно заряженная глутаминовая кислота позволяет дезамидированному проламину связываться со специфическим классом молекул HLA (DQ2 или DQ8) (Kim et al. 2004). Стимулируется пролиферация специфичных клонов T-клеток, так называемых DQ2 (8)/CD4+ несущих T-клеток, направленных на эндотелий кишечника, которые секретируют лимфокины, вызывающие атрофию кишечных ворсинок или выработку антител (Hadjivassiliou et al. 2004). Указанные клоны T-клеток достигают максимальной концентрации в периферической крови больных целиакией приблизительно через шесть дней после пищевой провокации (Anderson et al. 2000). Токсичность очищенных белков при целиакии, таким образом, может быть точно и специфично определена путем измерения их способности стимулировать T-клетки к выработке IFN-γ - цитокина, являющегося основным в патогенезе энтеропатии, наблюдаемой при целиакии. Поэтому вероятно, что заболевание вызывается иммунной системой хозяина, которая реагирует на проламины, формируя мощный иммунный ответ, как если бы они являлись скорее патогеном, чем аллергеном.
Глютен пшеницы состоит из многих сотен различных, но родственных белков, включая мономерные глиадины и полимерные глютенины. Глиадины составляют приблизительно половину глютеновой фракции, при этом α-глиадины составляют более 50% глиадинов (Wieser et al., 1994; Gellrich et al., 2003). До настоящего времени большинство данных по токсичности при целиакии было сосредоточено на α-глиадине - первом клонированном и полностью отсеквенированном проламине (Kasarda et al., 1984). Токсичность α-глиадина пшеницы при целиакии в значительной степени определяется одним остатком глутамина в пределах ключевого эпитопа длиной 17 аминокислот (Arentz-Hansen et al., 2000; Anderson et al., 2000; Shan et al., 2002). Были идентифицированы природные и синтетические пептиды, несущие точковые мутации в данной области, которые не являлись токсичными (Vader et al., 2003). Таким образом, кажется возможным, что могут быть идентифицированы другие нетоксичные, но функциональные проламиновые молекулы. В настоящий момент практическое предсказание токсичности при целиакии ограничивается небольшой фракцией проламинов, которые были описаны по аминокислотной последовательности или нуклеотидной последовательности генов, кодирующих их.
Ячмень представляет собой диплоидную злаковую культуру, которую широко выращивают в зонах с более прохладным климатом для производства напитков и продуктов питания. Белки семени ячменя подразделяются на альбумин, глобулин, проламин (гордеин) и глютелин в зависимости от их растворимости в воде, растворе соли, водном растворе спирта, а также в щелочных или кислотных растворах соответственно. Приблизительно половина запасных белков семени в ячмене присутствует в проламиновой фракции. Данные проламины, прежде всего, являются запасными белками, которые служат в качестве источников углерода, азота или серы для роста и развития после прорастания. Гордеин составляет приблизительно 40% белка семени, хотя это зависит от обеспечения растения азотом в процессе роста. Локусы, кодирующие проламины ячменя, были характеризованы главным образом из-за их вклада в качество ячменного солода, а также в пенообразование и мутность в производстве пива. В ячмене присутствует четыре класса проламинов - B-, C-, D- и γ-гордеины, кодируемые локусами Hor2, Horl, Hor3 и Hor5 соответственно, на хромосоме 1H (Shewry et al. 1999). Указанные локусы кодируют белки, которые варьируют от одного проламина (например, D-гордеин) до семейств белков, содержащих 20-30 членов (например, B- и C-гордеины). B- и C-гордеины встречаются относительно более часто, включая приблизительно 70% и 24% от общего количества гордеинов соответственно. D- и γ-гордеины представляют минорные компоненты, приблизительно по 2-4% каждый. Молекулярная масса гордеинов изменяется приблизительно от 35 кДа до 100 кДа. Каких-либо проламинов ячменя, которые имеют близкую гомологию с α-глиадинами пшеницы, нет, однако общеизвестно, что гордеины токсичны для больных целиакией (Williamson & Marsh 2000). Относительно степени, в которой отдельные гордеины ячменя стимулируют CD, не сообщали.
Пиво является широко потребляемым продуктом, производимым из соложеного ячменя, поэтому пиво, как широко предполагается, не подходит для больных целиакией и, как правило, исключено из их диеты. Kanerva et al. (2005) смогли идентифицировать проламины в низких уровнях во всех сортах из множества сортов пива, кроме одного. Врачи и диетологи обычно убеждают своих пациентов с CD стараться избегать потребления любых продуктов, содержащих пшеницу, ячмень или рожь, включая пиво. В США определение FDA "не содержит глютена" требует, чтобы продукт был произведен только из сырья, не содержащего глютен, то есть вообще не содержащего пшеницу, ячмень или рожь. Кодекс алиментариус допускает пометку "не содержит глютена" на пищевых продуктах, содержащих не более 200 м.д. глютена (0,2 г на килограмм или литр), это также соответствует европейскому стандарту для продуктов, которые "не содержат глютена". Большинство больных целиакией может потреблять приблизительно до 10 мг глютена в день без развития основного эффекта (Thompson, 2001).
Проламины, токсичные для пациентов с целиакией, можно специфично обнаружить с помощью иммуноанализов, таких как ELISA (Ellis et al., 1990; Sorell et al., 1998). Данные анализы основаны на специфичном взаимодействии между белком и антителом. Также использовали электрофорез в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ) и ВЭЖХ (Kanerva et al., 2005; Marchylo et al., 1986; Sheehan and Skerritt, 1997).
Таким образом, существует потребность в ячмене со значительно более низким содержанием CD-индуцирующих гордеинов, который может применяться в продуктах питания и напитках, предназначенных для лиц, подверженных риску развития CD.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В ячмене присутствует четыре класса проламинов - B-, C-, D-и γ-гордеины, кодируемые локусами Hor2, Horl, Hor3 и Hor5 соответственно. Авторы настоящего изобретения обнаружили, что, по меньшей мере, B, C и D классы вызывают нежелательные воспалительные ответы у лиц с целиакией.
Тогда как ранее были идентифицированы различные мутанты ячменя с продукцией определенных классов гордеинов в уменьшенных уровнях, также наблюдали, что это, по меньшей мере, компенсировалось увеличенной продукцией гордеинов других классов. Это позволяет предположить, что семя ячменя имеет компенсационные механизмы, которые обеспечивают определенные уровни гордеинов, необходимые для того, чтобы семя являлось жизнеспособным. Неожиданно авторы настоящего изобретения определили, что большая часть, если не все количество, гордеинов, вырабатываемых в ячмене, можно удалить и получить при этом жизнеспособные семена, которые способны прорастать и давать растения ячменя в полевых условиях, несмотря на потерю основной запасной формы азота в семени. Указанные семена в частности полезны для производства продуктов питания и напитков, пригодных для потребления лицами, страдающими целиакией.
Таким образом, в одном аспекте настоящее изобретение обеспечивает способ получения продукта питания или напитка на основе солода, включающий смешивание зерна ячменя или солода, муки или непросеянной муки, полученной из указанного зерна с по меньшей мере другим компонентом продукта питания или напитка, причем зерно, солод, мука или непросеянная мука содержит около 25% или менее от содержания гордеинов в зерне, от соответствующего растения ячменя дикого типа, или солоде, муке или непросеянной муке, полученных аналогичным путем из зерна, получаемого из соответствующего растения ячменя дикого типа, с получением в результате продукта питания или напитка на основе солода.
Предпочтительно, зерно, солод, мука или непросеянная мука содержит около 15% или менее, около 10% или менее, около 7,5% или менее, около 5% или менее или, более предпочтительно около 2,5% или менее от содержания гордеинов в зерне соответствующего растения ячменя дикого типа, или солоде, муке или непросеянной муке, полученных аналогичным путем из зерна дикого типа.
Примеры растения ячменя дикого типа включают, помимо прочих, Bomi, Sloop, Carlsberg II, K8 или L1.
В другом варианте осуществления зерно содержит около 25% или менее, около 20% или менее, около 15% или менее, около 10% или менее, около 7,5% или менее, около 5% или менее или более предпочтительно около 2,5% или менее от содержания B-, C- и/или D-гордеинов или любых комбинаций перечисленного в зерне соответствующего растения ячменя дикого типа. Солод, мука или непросеянная мука могут включать аналогичную степень снижения уровня гордеинов B, C и/или D, или любых комбинаций перечисленного.
В другом варианте осуществления мука содержит менее чем около 0,4%, менее чем около 0,3%, менее чем около 0,2% и более предпочтительно менее чем около 0,1% гордеинов. Уровни гордеина в муке, полученной из указанного зерна, могут быть определены с помощью любого метода, известного в уровне техники, такого как спиртовое фракционирование.
В одном из вариантов осуществления зерно имеет средний вес (ср. вес 100 зерен), по меньшей мере, около 2,4 г. Предпочтительно, зерно имеет средний вес от около 2,4 до около 6 г, более предпочтительно средний вес от около 3,5 до около 6 г.
В другом варианте осуществления, содержание крахмала в зерне составляет, по меньшей мере, приблизительно 50% (по весу). Более предпочтительно, содержание крахмала в зерне составляет от приблизительно 50% до приблизительно 70% (по весу). Содержание крахмала может быть определено с использованием любой методики, известной в уровне техники. Например, может применяться способ, приведенный в Примере 4.
В следующем варианте осуществления токсичность при целиакии муки, полученной из зерна, составляет менее чем приблизительно 50%, менее чем приблизительно 25%, более предпочтительно приблизительно 10% или менее, от соответствующего показателя муки, полученной из зерна соответствующего растения ячменя дикого типа. Токсичность при целиакии может быть определена, используя любую технику, известную в уровне техники. Например, может применяться способ, приведенный в Примере 1.
В еще одном варианте осуществления солод, полученный из зерна, содержит менее чем приблизительно 200 м.д. гордеинов, менее чем приблизительно 125 м.д. гордеинов, более предпочтительно менее чем приблизительно 75 м.д. гордеинов. Содержание гордеина также может быть определено с помощью любого метода, известного в уровне техники. Например, может применяться способ, приведенный в Примере 7.
В другом варианте осуществления, по меньшей мере, приблизительно 50% генома зерна ячменя идентичны геному ячменя сорта Sloop.
Предпочтительно, зерно получено из растения, которое является гомозиготным, по меньшей мере, по одному, по меньшей мере, по двум, по меньшей мере, по трем или более локусам для генетической вариации (вариаций), которая приводит к пониженному содержанию гордеина, по меньшей мере, одного, по меньшей мере, двух или всех трех классов B, C и D, по сравнению с соответствующим растением ячменя дикого типа. Более предпочтительно, указанные генетические вариации представляют собой аллели, в которых удалена большая часть или все гены, кодирующие B-гордеин, в локусе Hor2, и/или мутантный аллель в локусе Lys3 ячменя.
В одном варианте осуществления зерно получено из нетрансгенного растения. Например, зерно может быть получено в результате скрещивания между Riso 56 и Riso 1508 или их потомства, включающих мутации hor2 и Lys3 соответственно, присутствующие в указанных родительских линиях. Предпочтительно, такие растения, полученные в результате скрещивания, включают генетический фон, по существу отличный от Riso 56 или Riso 1508, например, содержащий менее чем приблизительно 25% генетического фона указанных родительских линий.
В другом варианте осуществления зерно получено из трансгенного растения. Одним из вариантов осуществления трансгенного растения является растение, которое включает трансген, кодирующий полинуклеотид, который подавляет продукцию, по меньшей мере, одного гордеина в зерне. Предпочтительно, полинуклеотид данного варианта осуществления является антисмысловым полинуклеотидом, смысловым полинуклеотидом, каталитическим полинуклеотидом, искусственной микроРНК или РНК дуплексом, которые подавляют экспрессию одного или предпочтительно нескольких генов, кодирующих гордеины.
Другим вариантом осуществления трансгенного растения является растение, которое включает трансген, кодирующий проламин, который менее токсичен, или предпочтительно нетоксичен, для лица, страдающего целиакией. Пример проламина, который является нетоксичным для лица, страдающего целиакией, включает, помимо прочего, авенины овса и зеины кукурузы.
В одном из вариантов осуществления способ включает получение из зерна муки или непросеянной муки.
В особенно предпочтительном варианте осуществления способ дополнительно включает солод, полученный из зерна. В варианте осуществления способ дополнительно включает фракционирование высушенного, пророщенного зерна на две или более из фракции эндосперма, фракции эндотелиального слоя, фракции шелухи, фракции зародышей и фракции солодовых ростков; а также комбинирование и смешивание заданного количества двух или более фракций.
Что касается производства солода и пива, важным компонентом семени ячменя является крахмал. При этом уровни крахмала в мутантах ячменя с уменьшенным содержанием гордеинов, как было показано ранее, снижены, что может привести к тому, что семя будет неподходящим для производства солода и пива. Авторы изобретения в частности неожиданно обнаружили, что семена ячменя, в которых большая часть, если не все количество, продуцируемого гордеина была удалена, могут использоваться для производства солода и пива с подходящими свойствами для коммерческого производства. Таким образом, в наиболее предпочтительном варианте осуществления напитком на основе солода является пиво или виски, причем способ включает проращивание зерна.
В одном из вариантов осуществления напитком на основе солода является пиво, которое содержит, по меньшей мере, около 2%, более предпочтительно, по меньшей мере, около 4% спирта. Предпочтительно, спирт является этанолом.
В еще одном варианте осуществления напитком на основе солода является пиво, которое содержит менее чем около 1 м.д. гордеинов.
В следующем варианте осуществления, по меньшей мере, приблизительно 50% зерна прорастает в течение 3 дней после набухания в стандартных условиях, используемых при соложении.
Примеры продуктов питания, которые могут быть произведены с применением способов изобретения, включают, помимо прочих, муку, крахмал, дрожжевой или бездрожжевой хлеб, пасту, лапшу, корм для животных, готовые завтраки (хлопья), закусочные пищевые продукты, пироги, солод, мучные кондитерские изделия или продукты питания, содержащие соусы на основе муки.
Предпочтительно, продукт питания или напиток на основе солода предназначен для потребления человеком. В другом предпочтительном варианте осуществления после употребления продукта питания или напитка, по меньшей мере, один симптом целиакии не развивается у лица с указанным заболеванием.
В другом аспекте настоящее изобретение обеспечивает способ получения продукта питания или напитка на основе солода, включающий смешивание солода, включающего один или более белков зерна ячменя и менее чем приблизительно 200 м.д. гордеинов, и/или муки, включающей один или более белков зерна ячменя и менее чем приблизительно 0,4% гордеинов, по меньшей мере, с одним другим компонентом продукта питания или напитка, с получением в результате продукта питания или напитка на основе солода.
В одном из вариантов осуществления способ включает получение солода и/или муки.
В еще одном аспекте настоящее изобретение обеспечивает способ получения продукта питания или напитка на основе солода, включающий смешивание зерна ячменя или солода, муки или непросеянной муки, полученной из указанного зерна, по меньшей мере, с одним другим компонентом продукта питания или напитка, с получением в результате продукта питания или напитка на основе солода, где мука, полученная из зерна, содержит менее чем приблизительно 0,4% гордеинов, и/или солод, полученный из зерна, содержит менее чем приблизительно 200 м.д. гордеинов.
В одном из вариантов осуществления способ включает получение солода и/или муки.
В другом аспекте настоящее изобретение обеспечивает растение ячменя, из которого получают зерно, включающее приблизительно 25% или меньшее количество гордеинов, от уровня в зерне, полученном из соответствующего растения ячменя дикого типа.
Предпочтительно, зерно содержит около 15% или менее, около 10% или менее, около 7,5% или менее, около 5% или менее, или более предпочтительно около 2,5% или менее гордеинов, от содержания в зерне от соответствующего растения ячменя дикого типа. Примеры растения ячменя дикого типа включают, помимо прочих, Bomi, Sloop, Carlsberg II, K8 или L1.
В другом варианте осуществления зерно содержит около 25% или менее, около 20% или менее, около 15% или менее, около 10% или менее, около 7,5% или менее, около 5% или менее или более предпочтительно около 2,5% или менее от содержания B-, C- и/или D-гордеинов, или любых комбинаций перечисленного, в зерне от соответствующего растения ячменя дикого типа.
В другом варианте осуществления мука, полученная из зерна, содержит менее чем около 0,4%, менее чем около 0,3%, менее чем около 0,2% и более предпочтительно менее чем около 0,1% гордеинов.
В варианте осуществления зерно имеет средний вес (ср. вес 100 зерен), по меньшей мере, около 2,4 г. Предпочтительно, зерно имеет средний вес от около 2,4 г до около 6 г, более предпочтительно средний вес от около 3,5 г до около 6 г.
В другом варианте осуществления содержание крахмала в зерне составляет, по меньшей мере, около 50% (по весу). Более предпочтительно, содержание крахмала в зерне составляет от около 50% до около 70 % (по весу).
В следующем варианте осуществления токсичность при целиакии муки, полученной из зерна, составляет менее чем приблизительно 50%, менее чем приблизительно 25%, более предпочтительно приблизительно 10% или меньше, от уровня токсичности муки, полученной из зерна соответствующего растения ячменя дикого типа.
В еще одном варианте осуществления солод, полученный из зерна, содержит менее чем около 200 м.д. гордеинов, менее чем около 125 м.д. гордеинов, более предпочтительно менее чем около 75 м.д. гордеинов.
В другом варианте осуществления, по меньшей мере, около 50% генома зерна ячменя идентичны геному сорта ячменя Sloop.
Предпочтительно, зерно получено из растения, которое является гомозиготным, по меньшей мере, по одному, по меньшей мере, по двум, по меньшей мере, по трем или более локусов для генетической вариации (вариаций), которая приводит к снижению уровня гордеина, по меньшей мере, одного, по меньшей мере, двух или всех трех классов B, C и D, по сравнению с уровнем в соответствующем растении ячменя дикого типа.
В одном варианте осуществления зерно получено из нетрансгенного растения. Например, зерно может быть получено в результате скрещивания Riso 56 и Riso 1508 или их потомства, включающих мутации hor2 и Lys3 соответственно, присутствующие в указанных родительских линиях. Предпочтительно, такое зерно включает генетический фон, по существу отличный от Riso 56 или Riso 1508, например, содержащий менее чем приблизительно 25% генетического фона указанных родительских линий.
В другом варианте осуществления зерно получено из трансгенного растения.
Один из вариантов осуществления трансгенного растения является растением, которое включает трансген, кодирующий полинуклеотид, который подавляет продукцию в зерне, по меньшей мере, одного гордеина. Предпочтительно, полинуклеотид данного варианта осуществления является антисмысловым полинуклеотидом, смысловым полинуклеотидом, каталитическим полинуклеотидом, искусственной микроРНК или РНК дуплексом, которые подавляют экспрессию одного или предпочтительно нескольких генов, кодирующих гордеины.
Другим вариантом осуществления трансгенного растения является растение, которое включает трансген, кодирующий проламин, который менее токсичен, предпочтительно нетоксичен для лица с целиакией. Пример проламина, который является нетоксичным для лица с целиакией, включает, помимо прочих, авенин овса.
В следующем варианте осуществления, по меньшей мере, приблизительно 50% зерна прорастает в течение 3 дней после набухания в стандартных условиях, используемых при соложении.
В другом аспекте настоящее изобретение обеспечивает растение ячменя, из которого получают зерно, где мука, полученная из зерна, включает менее чем приблизительно 0,4% гордеины, и/или солод, полученный из зерна, включает менее чем приблизительно 200 м.д. гордеинов.
В другом аспекте настоящее изобретение обеспечивает зерно растения ячменя согласно изобретению.
В следующем аспекте настоящее изобретение обеспечивает способ получения зерна ячменя, включающий:
a) выращивание растения ячменя изобретения,
b) сбор урожая зерна, и
c) возможно, обработка зерна.
Предпочтительно, растения выращивают в коммерческом масштабе в полевых условиях. Например, в одном варианте осуществления способ включает выращивание, по меньшей мере, 1000, более предпочтительно, по меньшей мере, 5000 растений в поле на площади, по меньшей мере, один гектар.
Также предлагается способ получения муки, непросеянной муки, крахмала или другого продукта, полученного из зерна, включающий:
a) получение зерна изобретения, и
b) обработку зерна для получения муки, непросеянной муки, крахмала или другого продукта.
В другом аспекте настоящее изобретение обеспечивает продукт, полученный из растения ячменя изобретения или зерна изобретения.
В одном из вариантов осуществления продукт является продуктом питания или напитком на основе солода.
Предпочтительно напиток на основе солода является пивом или виски.
В другом варианте осуществления продукт является непищевым продуктом, предпочтительно включающим крахмал или состоящим, по меньшей мере, из приблизительно 50% крахмала. Примеры включают, помимо прочего, пленки, покрытия, клеящие материалы, бумагу, строительные материалы и упаковочные материалы, или продукты, не содержащие крахмал, такие как этанол.
В еще одном аспекте настоящее изобретение обеспечивает продукт питания или напиток на основе солода, произведенный с применением способа изобретения.
В одном из вариантов осуществления напиток на основе солода является пивом, которое включает, по меньшей мере, приблизительно 2%, более предпочтительно, по меньшей мере, приблизительно 4% спирта. Предпочтительно, спирт является этанолом.
В еще одном варианте осуществления напиток на основе солода является пивом, которое включает менее чем приблизительно 1 м.д. гордеинов.
В другом аспекте настоящее изобретение обеспечивает пиво, включающее один или более белков зерна ячменя и менее чем приблизительно 1 м.д. гордеинов. В одном из вариантов осуществления пиво содержит менее чем приблизительно 0,5 м.д. гордеинов.
Предпочтительно пиво включает, по меньшей мере, приблизительно 2%, более предпочтительно, по меньшей мере, приблизительно 4% спирта. Предпочтительно спирт является этанолом. Примеры белков зерна ячменя включают, помимо прочих, 9 кДа белок переноса липидов 1 ячменя (LTP1) и белок Z.
В другом аспекте настоящее изобретение обеспечивает муку, включающую один или более белков зерна ячменя и менее чем приблизительно 0,4% гордеинов.
В одном из вариантов осуществления мука включает менее чем приблизительно 0,3%, менее чем приблизительно 0,2% и более предпочтительно менее чем приблизительно 0,1% гордеинов.
Предпочтительно мука включает менее чем приблизительно 7 мг, более предпочтительно менее чем приблизительно 5 мг спирторастворимого белка/г сухого веса муки.
В еще одном аспекте настоящее изобретение обеспечивает солод, включающий один или более белков зерна ячменя и менее чем приблизительно 200 м.д. гордеинов.
В одном из вариантов осуществления солод включает менее чем приблизительно 125 м.д. гордеинов, более предпочтительно менее чем приблизительно 75 м.д. гордеинов.
В другом аспекте настоящее изобретение обеспечивает способ идентификации зерна ячменя, которое может использоваться для получения продукта питания и/или напитка на основе солода, пригодного для употребления лицом с целиакией, включающий:
a) получение одного или нескольких следующих материалов:
i) образца из растения, способного давать указанное зерно,
ii) зерна,
iii) солода, полученного из зерна, и/или
iv) экстракта указанного зерна,
b) анализ материала стадии a) на наличие, по меньшей мере, одного гордеина и/или, по меньшей мере, одного гена, кодирующего гордеин,
где чем больше количество гордеинов, продуцированных в зерне, тем зерно меньше подходит для производства продукта питания и/или напитка на основе солода, пригодного для употребления лицом с целиакией.
В варианте осуществления образец является зерном, а стадия b) включает анализ материала на наличие гордеинов B и/или C. Это может быть выполнено с использованием любого метода, известного в уровне техники, например, с использованием иммунологического метода, такого как анализ ELISA. Может применяться способ, описанный в Примере 1. В одном из вариантов осуществления стадия b) включает пероральное введение материала из стадии a) лицу с целиакией и определение иммунореактивности T-клеток, полученных у лица, в отношении к одному или нескольким гордеинам ячменя.
В другом варианте осуществления материал образца стадии a) включает геномную ДНК, а стадия b) включает обнаружение отсутствия одного или нескольких функциональных генов гордеинов. Опять же, это может быть выполнено с использованием любого метода, известного в уровне техники. Например, выполняя стадию амплификации гена, как описано в Примере 9.
В одном из вариантов осуществления способ включает стадию селекции растения ячменя, зерна или солода согласно изобретению из множества образцов растений, зерна или солода для культивирования или применения. Такая селекция основана, прямо или косвенно, на уменьшенной токсичности материала при целиакии.
В другом аспекте настоящее изобретение обеспечивает способ профилактики или снижения частоты или тяжести целиакии у лица, включающий пероральное введение лицу продукта питания или напитка на основе солода изобретения, или зерна изобретения. Частота или тяжесть заболевания в данном контексте, как понимают, снижены относительно введения равного количества продукта питания или напитка, приготовленного из ячменя дикого типа. Продукт питания или напиток могут применяться для обеспечения питательными веществами, или повышенным количеством питательных веществ, лица, страдающего целиакией, уменьшая при этом риск провоцирования симптомов заболевания.
В другом аспекте настоящее изобретение предусматривает применение продукта питания или напитка на основе солода изобретения, или зерна изобретения, в целях производства лекарственного средства, предназначенного для перорального введения лицу питательных веществ, предотвращая или уменьшая в то же время частоту или тяжесть целиакии.
Как очевидно, предпочтительные признаки и особенности одного из аспектов изобретения применимы к любому другому аспекту изобретения.
По всему тексту настоящего описания слово "включает", или такие вариации, как "включающий", следует понимать, как включение указанного элемента, целого числа или стадии, или группы элементов, целых чисел или стадий, а не исключение какого-либо другого элемента, целого числа или стадии, или группы элементов, целых чисел или стадий.
Далее изобретение описано посредством неограничивающих Примеров со ссылкой на сопровождающие иллюстрации.
КРАТКОЕ ОПИСАНИЕ СОПРОВОЖДАЮЩИХ ИЛЛЮСТРАЦИЙ
Фиг.1: Обращено-фазовая FPLC суммарного экстракта проламинов, на которой показаны хроматограммы A280 нм пшеницы (a), ячменя (b), овса (c); кукурузы (d) или чистый градиент (e). Наносили проламины, эквивалентные 0,2 г муки. Показан окисленный ДТТ (DTTox); хроматограммы были сведены для ясности.
Фиг.2: Обращено-фазовая FPLC гордеинов. На репрезентативной хроматограмме показаны A280 нм (сплошная линия) и смесь растворителей (пунктирная линия) в ходе выделения гордеиновой фракции 1 (#1), 2 (#2), 3 (#3), 4 (#4), 5 (#5), или 6 (#6) из экстракта ячменя. Указанные фракции были объединены, как показано (жирная линия), из последовательных введений проб.
Фиг.3: Анализ 20 мкг гордеиновых фракций #1-6 с помощью электрофореза в ДСН-ПААГ, гель окрашен 0,06% Coomassie Blue G250. На левой дорожке указано положение стандартов молекулярной массы (в кДа, BenchMark, Invitrogen).
Фиг.4: Стимуляция выработки IFN-γ в T-клетках, выделенных у больных целиакией через шесть дней после диетической провокации, суммарными препаратами проламинов, приготовленными из ячменя, пшеницы, овса или кукурузы в присутствии (•, n=21)) или отсутствии (o, n=13) предварительной обработки tTG. IFN-γ положительные колонии были подсчитаны и представлены, как среднее SFU ± SE. Планки погрешностей не показывали, когда SE было меньше, чем символы.
Фиг.5: Стимуляция выработки IFN-γ в T-клетках, выделенных у больных целиакией через шесть дней после диетической провокации гордеиновыми фракциями #1, 2, 3, 4, 5 и 6, в присутствие (•, n=21)), или отсутствие (o, n=13) предварительной обработки tTG. IFN-γ положительные колонии были подсчитаны и представлены, как среднее SFU ± SE. Планки погрешностей не показывали, когда SE было меньше, чем символы.
Фиг.6: Хроматограммы аналитической обращено-фазовой ВЭЖХ выделенных гордеиновых фракций. На репрезентативных хроматограммах показаны A280 нм в ходе ВЭЖХ гордеиновых фракций #1, 2, 3, 4, 5, 6, выделенных из ячменя. Для сравнения показаны хроматограммы для ячменя дикого типа (Himalaya), на которых показана элюция (сплошная линия) гордеины D, C, и B, а также мутант R56, в котором накапливаются главным образом C гордеины.
Фиг.7: Характеристика проламинов в Riso56 и Riso1508 с помощью электрофореза в ДСН-ПААГ и Вестерн-блоттинга. Двадцать мкг проламина, очищенного, как в Примере 1, из указанной линии ячменя, инкубировали в течение 30 минут при комнатной температуре в буфере, содержащем 6,6 М мочевины, 2% (вес./об.) ДСН, 1% (вес./об.) ДТТ, 62,5 мМ Трис-HCl (pH 6,8) и 0,01% (вес./об.) бромфенолового синего, а затем наносили на две дорожки в 12% акриламидный гель и проводили электрофорез при 200 V в течение 40 мин. Гель промывали в буфере для переноса, содержащем 192 мМ глицина, 25 мМ Трис-основания и 20% (об./об.) метанола, в течение 10 минут, после чего выполняли перенос на нитроцеллюлозу (Amersham Hybond C+) при 100 V в течение 1 часа. Мембрану слева окрашивали в 0,2% Ponceau S (вес./об.) в 3% (вес./об.) трихлоруксусной кислоте, 3% 5-сульфосалициловой кислоте и быстро удаляли излишки красителя в воде; мембрану справа блокировали в 5% обезжиренном молоке в PBST в течение 1 часа, затем инкубировали с мышиным моноклональным антителом 12224 (Skerrit, 1988) в PBST, 3x промывали в PBST в течение 10 минут, инкубировали с антителом овцы против мышиного антитела, конъюгированным с HRP (Selenius), в PBST, 3x промывали в PBST 5 минут, инкубировали в ECL-реагенте Amersham согласно инструкции изготовителя, и выдерживали на Amersham Hyperfilm. MAb 12224 индуцировали против суммарного глютенинового экстракта, поэтому детектировали все гордеины и проламины (Skerrit, 1988).
Фиг.8: Обращено-фазовая FPLC гордеиновых экстрактов в Riso56 и Riso1508 в сравнении с диким типом Bomi и Carlsberg II. Гордеины выделяли из указанных линий, как в Примере 1, при этом на FPLC колонку наносили количество, эквивалентное 0,2 г муки, используя первый метод FPLC в Примере 1. Указано время элюирования C-гордеина (C-Hor) и B-гордеина (B-Hor).
Фиг.9: Репрезентативный электрофорез в ДСН-ПААГ спирторастворимых белков, нанесенных в расчете на семя. Проламиновые экстракты (10 мкл) отдельных семян ячменя F2 из скрещивания Riso1508 и Riso56 получали, как описано выше. Положения белковых стандартов 30, 50, 70 и 100 кДа указаны на дорожке слева. Также показаны белковые профили родительских линий Riso1508 и Riso56, и дикого типа (Bomi). Шесть дорожек из предполагаемых двойных нулевых проб содержат очень мало белка (ноль), шесть других дорожек содержат низкие уровни белка (сниженный).
Фиг.10: Репрезентативный электрофорез в ДСН-ПААГ спирторастворимых белков, нанесенных исходя из равного количества белка. Пробы, содержащие 20 мкг спирторастворимого белка, экстрагированного из отдельных семян ячменя F2, разделяли с помощью электрофореза, а гель окрашивали Coomassie blue. Наносили пробы из родительских линий (Riso 1508 и Riso 56), а также дикого типа (Bomi). Внешние и центральные дорожки (10 кДа) содержали белковые стандарты с известным молекулярным весом, указаны положения полос 30, 50, 70 и 100 кДа.
Фиг.11: Хроматограммы ОФ FPLC спирторастворимых экстрактов из семян ячменя F3. Спирторастворимые белки экстрагировали из отдельных семян F3, как описано; супернатанты двух семян из каждой линии объединяли, 50 мкл вводили в колонку ОФ FPLC и элюировали, как описано в Примере 1.
Фиг.12: Содержание водорастворимого (A), солерастворимого (B), спирторастворимого (C) и растворимого в растворе мочевины (D) белка в продублированных образцах муки из ячменя дикого типа (Sloop, Carlsberg II, Bomi), одно-нулевых родителей (Riso 56, Riso 1508) и семенах F4 растений линий J4, J1, BB5, G1, 5RB определяли, как в Примере 4. Определяли содержание суммарного экстрагируемого белка (E), суммируя содержание в отдельных фракциях. Содержание суммарного белка также оценивали с помощью элементного анализа согласно методу Дюма (F). Содержание белка приведено, как среднее ± SE.
Фиг.13: Токсичность при целиакии гордеинов, выделенных из различных образцов муки, определяли с использованием T-клеток, выделенных у больных целиакией, через 6 дней после провокации, как в Примере 5, и привели среднее число пятнообразующих единиц (SFU) + SE в зависимости от сырого веса муки. Для ясности, среднее SFU показано только для гордеинов, выделенных из ячменя дикого типа (Sloop) или двойной нулевой линии (G1) в присутствии (+tTG) или отсутствии (-tTG) фермента, тканевой трансглутаминазы (A). Во всех случаях обработка tTG повышала токсичность гордеинов, как ожидается при целиакии. SFU также приведено для обработанных tTG гордеинов (B), выделенных из образцов муки ячменя дикого типа (Sloop), одно-нулевых родителей (Р56, R1508) и семян F4 (4BH).
Фиг.14: Последовательности генов, специфичные либо к контрольному гену (gamma3-Hor), либо к гену B-гордеина (B-Hor), амплифицировали из ДНК экстрактов отдельных всходов F4 или линий 9RE, 4BH, или родительской линии Р56, как в Примере 9.
Общие методики и определения
Если конкретно не определено иное, все технические и научные термины, используемые в настоящей заявке, имеют стандартное значение, под которым они обычно понимаются средним специалистом в данной области техники (например, в растениеводстве, технологии пищевых продуктов, культивировании клеток, молекулярной генетике, иммунологии, химии белков и биохимии).
Если не указано иное, методы генной инженерии, культивирования клеток, а также иммунологические методы, используемые в настоящем изобретении, представляют собой стандартные методики, известные специалистам, квалифицированным в данной области техники. Такие методы широко описаны и объяснены в литературе, например, в J. Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984), J. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989), T. A. Brown (редактор), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991), D.M. Glover and B.D. Hames (редакторы), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996), а также F.M. Ausubel et al., (редакторы), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, включая все редакции на данный момент), Ed Harlow and David Lane (редакторы) Antibodies: A Laboratory Manual, Cold Spring Harbour Laboratory, (1988), and J.E. Coligan et al., (редакторы) Current Protocols in Immunology, John Wiley & Sons (включая все редакции на данный момент).
Используемый в настоящей заявке термин "ячмень" относится к любым видам рода Hordeum, включая предшественников данного рода, а также потомство, полученное в результате скрещивания с другими видами. Предпочтительной формой ячменя являются виды Hordeum vulgare.
Целиакия или глютеновая энтеропатия является аутоиммунным заболеванием тонкой кишки, которое развивается у генетически предрасположенных людей во всех возрастных группах после первого года жизни. Она затрагивает приблизительно 1% индоевропейских популяций, хотя она диагностируется в значительной степени недостаточно. Целиакия вызвана реакцией на глиадин, глютеновый белок, присутствующий в пшенице (а также подобными белками пшеницевых (Triticeae), которые включает другие культуры, такие как ячмень и рожь). При контакте с глиадином фермент тканевая трансглутаминаза модифицирует белок, а иммунная система перекрестно реагирует с тканью кишечника, вызывая воспалительную реакцию. Это приводит к выравниванию оболочки тонкой кишки и, как следствие, нарушению всасывания питательных веществ. Единственным эффективным методом лечения является пожизненная аглютеновая диета. Данное заболевание имеет несколько других названий, включающих: целиакию (cœliac disease, с лигатурой), целиакию- (c(o)eliac) спру, нетропическую спру, эндемическую спру, глютеновую энтеропатию или глютензависимую энтеропатию, а также непереносимость глютена. Симптомы целиакии значительно различаются от человека к человеку. Симптомы целиакии могут включить одно или более из следующего: газообразование, периодическое абдоминальное вздутие и боли, хроническая диарея, запор, бледность, зловонный или жирный стул, потеря веса/увеличение веса, утомляемость, необъяснимая анемия (низкое количество эритроцитов, вызывающее утомляемость), боли в костях или суставах, остеопороз, остеопения, поведенческие изменения, покалывание с онемением (от повреждения нервов), мышечные спазмы, судороги, нарушение менструального цикла (часто из-за чрезмерной потери веса), бесплодие, привычное невынашивание, задержка роста, задержка в развитии у детей, сероватые язвы во рту, называемые афтозными язвами, обесцвечивание зубов или потеря эмали, а также зудящая кожная сыпь, называемая герпетиформным дерматитом. Некоторые из наиболее часто встречаемых симптомов включают: усталость, перемежающуюся диарею, абдоминальные боли или спазмы, расстройство пищеварения, метеоризм, вздутие, а также потерю веса. Целиакия может быть диагностирована, например, как описано в WO 01/025793.
Используемый в настоящей заявке термин "нетоксичный для лица с целиакией" относится к употреблению продукта питания или напитка, не приводящего к развитию симптома целиакии у лица, страдающего от указанного заболевания. Как описано в настоящей заявке, продукт питания или напиток, приготовленный из соответствующего растения ячменя дикого типа, приводит к развитию симптомов заболевания.
Термины "семя" и "зерно" используются в настоящей заявке попеременно. "Зерно" обычно относится к зрелому, собранному зерну, но может также относиться к зерну после обработки, например, после размалывания или полировки, при которой состав большей части зерна остается в неизменном виде, или после набухания или прорастания, согласно контексту. Зрелое зерно обычно имеет влагосодержание менее чем приблизительно 18-20%. Зерно ячменя дикого типа (цельное зерно) обычно содержит 9-12% белка, из которого приблизительно 30-50% составляет проламин, обычно 35%, таким образом, зерно ячменя дикого типа содержит приблизительно 3-4% проламина по весу. Проламины присутствуют практически только в эндосперме, который составляет приблизительно 70% веса цельного зерна.
Используемый в настоящей заявке термин "соответствующий дикий тип" растения ячменя относится к растению, которое включает, по меньшей мере, 50%, более предпочтительно, по меньшей мере, 75%, более предпочтительно, по меньшей мере, 95%, более предпочтительно, по меньшей мере, 97%, более предпочтительно, по меньшей мере, 99%, и наиболее предпочтительно, 99,5% генотипа растения изобретения, но дает зерно с неизмененным содержанием гордеинов. В одном варианте осуществления "соответствующий дикий тип" растения ячменя представляет собой сорт растения, используемый в экспериментах по селекции растений с целью введения генетических вариантов, которые приводят к уменьшению продукции гордеинов в зерне. В другом варианте осуществления, "соответствующий дикий тип" растения ячменя является родительским сортом, в который был введен трансген, который уменьшает продукцию гордеинов в зерне. В следующем варианте осуществления, "соответствующий дикий тип" растения ячменя является сортом, который применяется на дату регистрации коммерческого производства зерна ячменя, такого как, помимо прочих, Bomi, Sloop, Carlsberg II, K8, L1, Vlamingh, Stirling, Hamelin, Schooner, Baudin, Gairdner, Buloke, WI3586-1747, WI3416, Flagship, Cowabbie, Franklin, SloopSA, Sloop Vic, Quasar, VB9104, Grimmett, Cameo*Arupo 31-04, Prior, Schooner, Unicom, Harrington, Torrens, Galleon, Morex, Dhow, Capstan, Fleet, Keel, Maritime, Yarra, Dash, Doolup, Fitzgerald, Molloy, Mundah, Onslow, Skiff, Unicorn, Yagan, Chebec, Hindmarsh, Chariot, Diamant, Koral, Rubin, Bonus, Zenit, Akcent, Forum, Amulet, Tolar, Heris, Maresi, Landora, Caruso, Miralix, Wikingett Brise, Caruso, Potter, Pasadena, Annabell, Maud, Extract, Saloon, Prestige, Astoria, EIo, Cork, Extract, Laura. В варианте осуществления "соответствующий дикий тип" растения ячменя дает зерно с неизмененным содержанием гордеинов, поскольку он включает полный набор функциональных гордеиновых генов, кодирующих функциональные белки гордеинов, включая B, C, D и γ-гордеины, кодируемые локусами Hor2, Hor1, Hor3 и Hor5.
Используемый в настоящей заявке термин "один или более белков зерна ячменя" относится к природным белкам, продуцируемым зерном ячменя. Примеры таких белков известны специалистам, квалифицированным в данной области техники. Конкретные примеры включают, помимо прочих, альбумины ячменя, например, 9 кДа белок переноса липидов 1 (LTP1) (см. обзор Douliez et al. (2000) и Swiss-prot P07597, в качестве примера), а также белок Z (см. Brandt et al. (1990) и Genbank P06293), включая соответствующие процессированные (зрелые) формы, а также их денатурированные формы и/или фрагменты, полученные в результате производства солода, муки, непросеянной муки, продукта питания или напитка на основе солода согласно изобретению.
Используемый в настоящей заявке термин "солод" относится к ячменному солоду, "мука" относится к ячменной муке, "мука из цельного зерна" относится к ячменной муке из цельного зерна, и "пиво" относится к ячменному пиву. Более конкретно, источник солода, муки, пива, муки из цельного зерна, продукта питания и т.д. согласно изобретению получают в результате обработки (например, помола и/или ферментации) зерна ячменя. Указанные термины включают солод, муку, пиво, муку из цельного зерна, продукт питания и т.д., полученные из смеси зерен. В предпочтительном варианте осуществления, по меньшей мере, 50% зерна, используемого для производства солода, муки, пива, муки из цельного зерна, продукта питания и т.д., является зерном ячменя.
Термин "растение", также используемый в настоящей заявке, относится к целому растению, такому как, например, растение, растущее в поле с целью коммерческого производства ячменя. "Часть растения" относится к вегетативным структурам растения (например, листьям, стеблям), корням, цветковым органам/структурам, семени (включая зародыш, эндосперм и оболочку семени), ткани растения (например, сосудистой ткани, покровной ткани и т.п.), клеткам, крахмальным зернам или потомству вышеперечисленного.
"Трансгенное растение", "генетически модифицированное растение" или варианты перечисленного, относятся к растению, которое содержит генную конструкцию ("трансген"), не присутствующую в растении дикого типа того же вида, расы или сорта. "Трансген", как указано в настоящей заявке, имеет нормальное значение из области биотехнологии и включает генетическую последовательность, которая была получена или изменена с помощью генноинженерных методов ДНК или РНК, и которая была введена в клетку растения. Трансген может включать генетические последовательности, полученные из клетки растения. Как правило, трансген введен в растение искусственно, например, с помощью трансформации, однако при этом может использоваться любой метод, известный специалисту в данной области техники.
"Молекула нуклеиновой кислоты" относится к полинуклеотиду, такому как, например, ДНК, РНК или олигонуклеотиды. Она может представлять собой ДНК или РНК геномного или синтетического происхождения, двунитевую или однонитевую, а также связанную с углеводом, липидами, белком или другими материалами для выполнения специфической функции, определенной в настоящей заявке.
"Функционально связанный", используемый в настоящей заявке, относится к функциональному отношению между двумя или более сегментами нуклеиновых кислот (например, ДНК). Как правило, это относится к функциональному отношению элемента регуляции транскрипции (промотора) с транскрибируемой последовательностью. Например, промотор функционально связан с кодирующей последовательностью, такой как полинуклеотид, определенный в настоящей заявке, если он стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке. В большинстве случаев промоторные элементы регуляции транскрипции, которые функционально связаны с транскрибируемой последовательностью, физически прилегают к транскрибируемой последовательности, то есть, они действуют в cis-положении. Однако, некоторые элементы регуляции транскрии, такие как энхансеры, не обязательно должны физически прилегать или располагаться в непосредственной близости к кодирующим последовательностям, транскрипцию которых они усиливают.
Используемый в настоящей заявке термин "ген" следует понимать в его наиболее широком смысле, при этом он включает дезоксирибонуклеотидные последовательности, включающие область структурного гена, кодирующую белок, и включает последовательности, прилегающие к кодирующей области на 5' и на 3' концах на протяжении, по меньшей мере, приблизительно 2 тпн в любом конце, и которые участвуют в экспрессии гена. Последовательности, которые расположены на 5'-конце кодирующей области и которые присутствуют на мРНК, называются 5'-нетранслируемыми последовательностями. Последовательности, которые расположены на 3'-конце или справа от кодирующей области и которые присутствуют на мРНК, называются 3'-нетранслируемыми последовательностями. Термин "ген" охватывает и кДНК, и геномные формы гена. Геномная форма или клон гена содержат кодирующую область, которая может быть прервана некодирующими последовательностями, которые называют "интронами" или "промежуточными областями" или "промежуточными последовательностями". Интроны представляют собой сегменты гена, которые транскрибируются в ядерную РНК (гяРНК), интроны могут содержать регуляторные элементы, такие как энхансеры. Интроны удаляются, или "вырезаются" из ядерного или первичного транскрипта, поэтому интроны отсутствуют в транскрипте информационной РНК (мРНК). Функционирует мРНК в ходе трансляции, определяя последовательность или порядок аминокислот в синтезируемом полипептиде. Термин "ген" включает синтетическую или слитую молекулу, кодирующую все или часть белков изобретения, описанных в настоящей заявке, а также нуклеотидную последовательность, комплементарную любой из вышеуказанных. Используемый в настоящей заявке термин "другой компонент продукта питания или напитка" относится к любому веществу, пригодному для употребления животным, предпочтительно к любому веществу, пригодному для употребления человеком. Примеры включают, помимо прочего, воду, зерно растений других видов, сахар и т.д.
Используемый в настоящей заявке термин "генетическая вариация, которая приводит к уменьшению уровня, по меньшей мере, одного гордеина", относится к любому полиморфизму растения ячменя, который приводит к уменьшению продукции гордеина. Генетическая вариация может являться, например, делецией гордеинового гена (генов) или его части, или мутацией, которая приводит к уменьшению транскрипции гена в ячмене. Примеры таких генетических вариаций присутствуют в Riso 56, Riso 527 и Riso 1508. Следовательно, такие растения могут применяться в способах изобретения. Кроме того, растение изобретения может являться гибридом любого из указанных мутантов ячменя. В предпочтительном варианте осуществления растение изобретения является гибридом Riso 56 и Riso 1508 или их потомства, включающего мутации hor2 и Lys3, присутствующие в данных линиях. В варианте осуществления растение не является гибридом Riso 527 и Riso 1508.
Используемая в настоящей заявке фраза "приблизительно", если не указано обратное, относится к любому допустимому диапазону с учетом рассматриваемого значения. В предпочтительном варианте осуществления термин "приблизительно" соответствует +/- 10%, более предпочтительно +/- 5% от указанного значения.
Проламины и гордеины
Проламины злаковых культур (известные как глиадины в пшенице, гордеины в ячмене, секалины во ржи, авенины в овсе и зеины в кукурузе) являются главными запасными белками эндосперма во всех зернах злаковых культур, за исключением овса и риса (Shewry and Halford, 2002). Гордеины составляют 35-50% суммарного белка в семенах ячменя (Jaradat, 1991). Они подразделяются на четыре группы (также известные как γ-гордеин), B, C, и D, в порядке уменьшения подвижности (Field et al., 1982). B-гордеины являются главной фракцией белка и отличаются от C-гордеинов по содержанию серы (Kreis and Shewry, 1989). B-гордеины составляют 70-80% от общего количества, а C-гордеины - 10-20% (Davies et al., 1993). A-гордеины в целом не рассматривают как фракцию запаса, тогда как D-гордеины гомологичны глютенинам с высокой молекулярной массой. Гордеины, наряду с остальной частью родственных проламинов злаковых культур, не экспрессируются в зиготном эмбрионе непосредственно, в отличие от других запасных белков, таких как напины, при этом они, как полагают, экспрессируются исключительно в крахмалистом эндосперме в течение средних/поздних стадий развития семени.
Примеры аминокислотных последовательностей гордеинов ячменя (приведенные в форме номера NCBI с описанием, идентификатором gi), включают, помимо прочих:







Примеры генов и/или кДНК, кодирующих гордеины ячменя (приведенные в форме номера NCBI с описанием, идентификатором gi), включают, помимо прочих:







Один из вариантов осуществления настоящего изобретения относится к трансгенным растениям ячменя, включающим проламин, который не токсичен для лица с целиакией. Как показано в настоящей заявке, примерами такого проламина являются авенин овса и зеин кукурузы.
Примеры аминокислотных последовательностей авенинов овса (приведенные в форме номера NCBI с описанием, идентификатором gi), включают, помимо прочих:



Соложение
Напиток на основе солода, обеспечиваемый настоящим изобретением, включает алкогольные напитки (включая напитки, полученные перегонкой) и безалкогольные напитки, которые производят с использованием солода в качестве части или всего исходного материала. Примеры включают пиво, хаппошу (напиток наподобие пива, с низким содержанием солода), виски, слабоалкогольные напитки на основе солода (например, напитки на основе солода, содержащие менее 1% спиртов), а также безалкогольные напитки.
Соложение представляет собой процесс контролируемого замачивания и проращивания с последующей сушкой зерна ячменя. Данная последовательность событий важна для синтеза многочисленных ферментов, которые вызывают модификацию зерна, процесс, в котором преимущественно деполимеризуются стенки мертвых клеток эндосперма и мобилизуются питательные вещества зерна. В последующем процессе сушки в результате химических реакций потемнения развивается аромат и цвет. Хотя основным применением солода является производство напитков, он может также применяться в других производственных процессах, например как источник ферментов в хлебопекарной промышленности или в качестве вкусовой добавки и красителя в пищевой промышленности, например, в виде солода или солодовой муки, или опосредованно, в виде солодового сиропа, и т.д.
В одном из вариантов осуществления настоящее изобретение относится к способу получения солодовой композиции. Способ предпочтительно включает следующие стадии:
(i) получение зерен растения ячменя изобретения,
(ii) замачивание указанного зерна,
(iii) проращивание замоченного зерна в заданных условиях, и
(iv) сушка указанного, пророщенного зерна.
Например, солод может быть получен любым из способов, описанных в Hoseney (Principles of Cereal Science and Technology, Second Edition, 1994: American Association of Cereal Chemists, St. Paul, Minn.). Впрочем, в настоящем изобретении может также использоваться любой другой подходящий способ получения солода, такой как способы получения специальных сортов солода, включающие, помимо прочих, способы обжарки солода. Один из неограничивающих примеров описан в Примере 6.
Солод может быть приготовлен с использованием зерна, полученного только из растений ячменя изобретения, или смесей, включающих другие зерна.
Солод главным образом используют в пивоварении, однако он также применяется и в производстве крепких спиртных напитков. Варка пива включает производство затора, главную и вспомогательную ферментации, а также постобработку. Сначала солод промалывают, размешивают в воде и нагревают. В ходе данного "затирания" ферменты, активированные при соложении, расщепляют крахмал зерна с образованием сбраживаемых сахаров. Приготовленный затор осветляют, добавляют дрожжи, смесь сбраживают и проводят постобработку.
В другом варианте осуществления из солода могут быть приготовлены заторные композиции. Указанный затор может являться первым и/или вторым, и/или следующим затором. Обычно заторная композиция имеет высокое содержание амидного азота и сбраживаемых углеводов, главным образом мальтозы. Как правило, затор приготавливают, выдерживая солод с водой, то есть, при затирании. В процессе затирания в композицию из солода/воды могут быть добавлены дополнительные богатые углеводами композиции, например ячменные, кукурузные или рисовые добавки. Неосоложенные зерновые добавки обычно не содержат каких-либо активных ферментов и поэтому, чтобы обеспечить ферменты, необходимые для гидролиза полисахаридов, необходим солод или вводимые извне ферменты.
Обычно первой стадией в процессе производства затора является размалывание солода, необходимое для того, чтобы вода могла получить доступ к частицам зерна в фазе затирания, которое по сути является продолжением процесса соложения с ферментативной деполимеризацией субстратов. В ходе затирания размолотый солод выдерживают с жидкой фракцией, такой как вода. Температуру либо поддерживают на постоянном уровне (изотермическое затирание), либо постепенно повышают. В любом случае, растворимые вещества, образующиеся при соложении и затирании, переходят в указанную жидкую фракцию перед ее разделением с помощью фильтрации на затор и остаточные твердые частицы, называемые дробиной. Указанный затор может также называться первым затором. После фильтрации получают второй затор. Следующие заторы могут быть получены при повторе данной процедуры. Неограничивающие примеры подходящих методик приготовления затора описаны в Hoseney (выше).
Заторная композиция может быть также приготовлена путем выдерживания растений ячменя изобретения или их частей с одним или несколькими подходящими ферментами, такими как ферментные композиции или смешанные ферментные композиции, например Ultraflo или Cereflo (Novozymes). Заторная композиция может быть также приготовлена с использованием смеси солода и неосоложенных растений ячменя или их частей, необязательно с добавкой одного или нескольких подходящих ферментов в ходе указанного приготовления. Кроме того, ферменты пролил-эндопептидазы, которые специфично разрушают токсичные аминосвязи, вовлеченные в целиакию, могут быть добавлены в процессе сбраживания затора с целью уменьшения токсичности остаточных гордеинов (De Angelis et al., 2007; Marti et al., 2005; Stepniak et al, 2006).
Обработка зерна
Зерно ячменя изобретения может быть обработано с целью получения продукта питания или непищевого продукта, с использованием любой методики, известной в данной области техники.
В одном из вариантов осуществления продуктом является мука из цельного зерна (мука из цельного зерна ультратонкого помола, например, мука из цельного зерна ультратонкого помола; мука из цельного зерна, или мука, полученная из приблизительно 100% зерна). Мука из цельного зерна включает очищенную составляющую муки (очищенную муку), а также грубую фракцию (грубую фракцию ультратонкого помола).
Очищенная мука может являться мукой, которую получают, например, при размалывании и просеивании очищенного ячменя. Управление по контролю качества продуктов питания и лекарственных средств (FDA) требует, чтобы мука удовлетворяла определенным стандартам крупности частиц, чтобы она могла быть включена в категорию очищенной ячменной муки. Крупность частиц очищенной муки описывается как мука, в которой не менее 98% частиц проходит через сито с отверстиями, размер которых не превышает размер отверстий тканой проволочной сетки, определяемых как "212 микрометров (U.S. Wire 70)".
Грубая фракция включает, по меньшей мере, одно из: отрубей и проростков. Например, проросток представляет собой зародыш растения, находящийся в зерне ячменя. Проростки включают липиды, волокно, витамины, белок, минеральные вещества и питательные вещества растительного происхождения, такие как флавоноиды. Отруби включают несколько клеточных слоев и содержат существенное количество липидов, волокна, витаминов, белка, минеральных веществ и питательных веществ растительного происхождения, таких как флавоноиды. Кроме того, грубая фракция может включать алейроновый слой, который также включает липиды, волокно, витамины, белок, минеральные вещества и питательные вещества растительного происхождения, такие как флавоноиды. Алейроновый слой, который фактически считается частью эндосперма, проявляет многие из тех же особенностей, что и отруби, и поэтому обычно удаляется вместе с отрубями и проростками в ходе процесса помола. Алейроновый слой содержит белки, витамины и питательные вещества растительного происхождения, такие как феруловую кислоту.
Далее грубая фракция может быть смешана с очищенной составляющей муки. Предпочтительно, грубую фракцию гомогенно смешивают с очищенным элементом муки. Гомогенное смешивание грубой фракции и очищенной составляющей муки может помочь уменьшить расслоение частиц по размеру в процессе отгрузки. Грубая фракция может быть смешана с очищенной составляющей муки, с формированием муки из цельного зерна, обеспечивая, таким образом, муку из цельного зерна с повышенной пищевой ценностью, содержанием волокон и антиоксидантной способностью по сравнению с очищенной мукой. Например, грубая фракция или мука из цельного зерна могут применяться в различных количествах для замены очищенной муки или муки из цельного зерна в хлебобулочных изделиях, закусочных продуктах и продуктах питания. Мука из цельного зерна настоящего изобретения (напр. мука из цельного зерна ультратонкого помола) может также поставляться непосредственно потребителям для применения при изготовлении их домашней выпечки. В примере осуществления крупность частиц муки из цельного зерна является такой, что 98% по весу частиц муки из цельного зерна меньше, чем 212 микрометров.
В других вариантах осуществления ферменты, присутствующие в отрубях и проростках муки из цельного зерна и/или грубой фракции инактивированны для стабилизации муки из цельного зерна и/или грубой фракции. В настоящем изобретении также рассматривается, что инактивированный может означать ингибированный, денатурированный или т.п. Стабилизация является процессом, в котором используется пар, нагревание, облучение или другие способы обработки, предназначенные для инактивации ферментов, присутствующих в слое отрубей и проростков. Природные ферменты в отрубях и проростках катализируют изменения соединений в муке, отрицательно влияя на кулинарные свойства муки и срок годности. Инактивированные ферменты не катализируют изменения соединений, присутствующих в муке, поэтому мука, которая была стабилизирована, сохраняет свои кулинарные свойства и имеет более длительный срок годности. Например, настоящее изобретение может обеспечивать технологию помола с двумя потоками для размола грубой фракции. После отделения и стабилизации грубой фракции, грубая фракция проходит через мельницу, предпочтительно вальцовую мельницу, с формированием грубой фракции, имеющей распределение размера частиц, меньшее или равное приблизительно 500 микрометрам. В примере осуществления вальцовая мельница обычно работает со скоростью вальца 115 - 144 м/с, высокая скорость вальца приводит к нагреву. Тепло, выделяемое в ходе процесса, и поток воздуха, приводят к уменьшению содержания микроорганизмов в грубой фракции. В других вариантах осуществления перед размолом на вальцовой мельнице грубая фракция может иметь среднее содержание аэробных микроорганизмов 95000 колониеобразующих единиц/грамм (КОЕ/г) и среднее содержание бактерий группы кишечной палочки 1200 КОЕ/г. После прохождения через вальцовую мельницу грубая фракция может иметь среднее содержание аэробных микроорганизмов 10000 КОЕ/г и среднее содержание бактерий группы кишечной палочки 900 КОЕ/г. Таким образом, содержание микроорганизмов в грубой фракции настоящего изобретения может быть существенно снижено. После просеивания измельченная грубая фракция с размером частиц более 500 микрометров может быть возвращена в процесс для дополнительного помола.
В дополнительных вариантах осуществления мука из цельного зерна или грубая фракция могут являться компонентом продукта питания. Например, продукт питания может представлять собой рогалик, бисквит, хлеб, булочку, круассан, клецки, английскую сдобную булочку, сдобную булочку, питу, бездрожжевой хлеб, охлажденные/замороженные изделия из теста, тесто, готовые бобы, буррито, чили, тако, тамал, тортилью, пирог, готовый завтрак, готовый обед, начинку, обед для приготовления в микроволновой печи, брауни, пирог, чизкейк, булочку к кофе, печенье, десерт, кондитерские изделия, сладкий рулет, сладкий батончик, хрустящую основу для пирога, начинку для пирога, продукт для детского питания, смесь для выпечки, кляр, панировку, смесь для соуса, наполнитель для мяса, заменитель мяса, смесь приправ, суповую смесь, соус, заправку для соуса, заправку к салату, суп, сметану, лапшу, пасту, макароны, лапшу чоу-мейн, лапшу ло-мейн, наполнитель для мороженого, брикет мороженого, мороженое-рожок, мороженое в вафлях, крекер, гренки, пончик, яичный рулетик, экструдированные снеки, прессованные мюсли с фруктами, закусочные продукты для приготовления в микроволновой печи, питательный батончик, блин, хлебобулочное изделие-полуфабрикат, претцель, пудинг, продукт на основе гранолы, чипсы, снеки, смесь снеков, вафлю, основу для пиццы, корм для животных или корм для домашних животных.
В альтернативных вариантах осуществления мука из цельного зерна или грубая фракция могут являться компонентом пищевой добавки. Например, пищевая добавка может являться продуктом, который добавляют к пище, и который содержит один или более компонентов, обычно включающих: витамины, минеральные вещества, травы, аминокислоты, ферменты, антиоксиданты, травы, специи, пробиотики, экстракты, пребиотики и волокна. Мука из цельного зерна или грубая фракция настоящего изобретения включают витамины, минеральные вещества, аминокислоты, ферменты и волокна. Например, грубая фракция содержит концентрированное количество пищевого волокна, а также другие необходимые питательные вещества, например, витамины группы B, селен, хром, марганец, магний и антиоксиданты, которые необходимы для здорового питания. Например, 22 грамма грубой фракции настоящего изобретения обеспечивают 33% рекомендуемого ежедневного потребления волокна для человека. Далее, для обеспечения человека 20% ежедневного рекомендуемого потребления волокна необходимо 14 граммов. Таким образом, грубая фракция является превосходным дополнительным источником для удовлетворения потребности человека в волокне. Таким образом, в настоящем варианте осуществления мука из цельного зерна или грубая фракция могут являться компонентом пищевой добавки. Пищевая добавка может включать любые известные пищевые компоненты, которые способствуют улучшению общего самочувствия человека, примеры включают, помимо прочего, витамины, минеральные вещества, другие волокнистые компоненты, жирные кислоты, антиоксиданты, аминокислоты, пептиды, белки, лютеин, рибозу, омега-3 жирные кислоты и/или другие пищевые компоненты.
В дополнительных вариантах осуществления мука из цельного зерна или грубая фракция могут являться волокнистой добавкой или ее компонентом. Многие существующие волокнистые добавки, такие как шелуха семян подорожника, производные целлюлозы и гидролизованная гуаровая камедь кроме содержания волокна не обладают существенной пищевой ценностью. Кроме того, многие волокнистые добавки имеют нежелательную структуру и плохой вкус. Волокнистые добавки, приготовленные из муки из цельного зерна или грубой фракции, могут способствовать обеспечению волокном, а также белком и антиоксидантами. Волокнистые добавки могут быть изготовлены, помимо прочего, в следующих формах: быстрорастворимые смеси для приготовления напитков, готовые к употреблению напитки, пищевые брикеты, вафли, печенье, крекеры, гель, капсулы, жевачки, жевательные таблетки и пилюли. Один из вариантов осуществления обеспечивает волокнистые добавки в форме ароматизированного коктейля или солодового напитка, данный вариант осуществления может быть особенно привлекательным в качестве волокнистой добавки для детей.
В дополнительном варианте осуществления мукомольный процесс может использоваться для получения муки из нескольких видов зерна, муки из нескольких видов ячменя или грубой фракции из нескольких видов зерна. Например, отруби и проростки из одного типа ячменя могут быть измельчены и смешаны с измельченным эндоспермом или мукой из цельного зерна ячменя другого типа. В альтернативе отруби и проростки одного типа зерна могут быть измельчены и смешаны с измельченным эндоспермом или мукой из цельного зерна другого типа. В дополнительном варианте осуществления отруби и проростки из первого типа ячменя или зерна могут быть смешаны с отрубями и проростками из второго типа ячменя или зерна, с получением грубой фракции из нескольких видов зерна. Рассматривается, что настоящее изобретение охватывает смешивание любой комбинации одного или более из отрубей, проростков, эндосперма и муки из цельного зерна, полученных из зерна одного или нескольких видов. Данный многозерновой, многоячменный подход может использоваться для получения муки специальных сортов и выгодного использования свойств и пищевой ценности зерна или ячменя нескольких видов, при изготовлении муки одного сорта.
Мука из цельного зерна настоящего изобретения может быть получена с помощью различных мукомольных процессов. Пример осуществления включает помол зерна в одном потоке, без отделения эндосперма, отрубей и проростков зерна в отдельные потоки. Чистое и кондиционированное зерно поступает в первую мельницу, такую как молотковая дробилка, вальцовая дробилка, стержневая мельница, ударная дробилка, дисковая мельница, роторно-струйная мельница, вальцовая мельница и т.п. В одном варианте осуществления мельница может являться вальцовой мельницей. После помола зерно выгружают и подают на сито. Может использоваться любое сито, известное из уровня техники и применяемое для просеивания измельченных частиц. Материал, проходящий через сетку сита, является мукой из цельного зерна настоящего изобретения и не требует никакой дополнительной обработки. Материал, который остается на сетке, называется второй фракцией. Вторая фракция требует дополнительного измельчения частиц. Таким образом, указанная вторая фракция может подаваться во вторую дробилку. После помола вторая фракция может подаваться во второе сито. Материал, проходящий через сетку второго сита, является мукой из цельного зерна настоящего изобретения. Материал, который остается на сетке, называется четвертой фракцией и требует дополнительной обработки для уменьшения размера частиц. Четвертая фракция на сетке второго сита поступает обратно, либо в первую дробилку, либо во вторую дробилку для дополнительной обработки через контур обратной связи. В альтернативном варианте осуществления изобретения процесс может включать ряд первых дробилок для обеспечения более высокой емкости системы.
Предусмотрено, что мука из цельного зерна, грубая фракция и/или зерновые продукты настоящего изобретения могут быть изготовлены с помощью любого мукомольного процесса, известного в уровне техники. Кроме того, предусмотрено, что мука из цельного зерна, грубая фракция и/или зерновые продукты настоящего изобретения могут быть модифицированы или улучшены посредством ряда других процессов, таких как: брожение, инстантизация, экструзия, инкапсулирование, обжарка, запекание и т.п.
Полинуклеотиды, которые подавляют продукцию гордеина
В одном варианте осуществления зерно, изобретения и/или применяемое в способах изобретения, получено из трансгенного растения ячменя, которое включает трансген, кодирующий полинуклеотид, который подавляет продукцию, по меньшей мере, одного гордеина в зерне. Примеры таких полинуклеотидов включают, помимо прочих, антисмысловой полинуклеотид, смысловой полинуклеотид, каталитический полинуклеотид, искусственную микроРНК или РНК дуплекс. В случае присутствия в зерне каждого из перечисленных полинуклеотидов приводит к снижению количества мРНК гордеина, доступной для трансляции.
Антисмысловые полинуклеотиды
Термин "антисмысловой полинуклеотид" следует понимать, как означающий молекулу ДНК или РНК, или их комбинацию, которая является комплементарной, по меньшей мере, к части специфичной молекулы мРНК, кодирующей гордеин и способной препятствовать ходу посттранскрипционного события, такого как трансляция мРНК. Применение методов с использованием антисмысловых полинуклеотидов известно в уровне техники (см. например, Hartmann and S. Endres, Manual of Antisense Methodology, Kluwer (1999)). Применение методов с использованием антисмысловых полинуклеотидов в растениях было рассмотрено в обзорах Bourque (1995) и Senior (1998). Senior (1998) констатировал, что в настоящее время методы с использованием антисмысловых полинуклеотидов являются хорошо отработанной методикой для управления экспрессией генов.
Антисмысловой полинуклеотид в растении ячменя изобретения гибридизуется с целевым полинуклеотидом при физиологических условиях. Используемый в настоящей заявке термин "антисмысловой полинуклеотид, который гибридизуется при физиологических условиях" означает, что полинуклеотид (который является полностью или частично однонитевым) способен, по меньшей мере, к формированию двунитевого полинуклеотида с мРНК, кодирующей белок, такой как гордеин ячменя, при нормальных условиях в клетке ячменя.
Антисмысловые молекулы могут включать последовательности, которые соответствуют структурным генам или последовательностям, которые регулируют экспрессию гена или сплайсинг. Например, антисмысловая последовательность может соответствовать целевой кодирующей области генов изобретения или 5'-нетранслируемой области (UTR), или 3'-UTR, или их комбинации. Она может являться частично комплементарной последовательностям интронов, которые могут вырезаться во время или после транскрипции, предпочтительно только последовательностям экзонов целевого гена. Принимая во внимание в целом более высокую дивергенцию UTR, нацеливание на данные области обеспечивает более высокую специфичность ингибирования гена.
Длина антисмысловой последовательности должна составлять, по меньшей мере, 19 непрерывных нуклеотидов, предпочтительно, по меньшей мере, 50 нуклеотидов и, более предпочтительно, по меньшей мере, 100, 200, 500 или 1000 нуклеотидов. Может использоваться полноразмерная последовательность, комплементарная полному генному транскрипту. Длина наиболее предпочтительно составляет 100-2000 нуклеотидов. Степень идентичности антисмысловой последовательности целевому транскрипту должна составлять, по меньшей мере, 90%, а более предпочтительно 95-100%. Антисмысловая молекула РНК, безусловно, может включать чужеродные последовательности, которые могут стабилизировать молекулу.
Каталитические полинуклеотиды
Термин каталитический полинуклеотид/нуклеиновая кислота относятся к молекуле ДНК или молекуле, содержащей ДНК (также известной в уровне техники как "дезоксирибозим"), или молекуле РНК или молекуле, содержащей РНК (также известный как "рибозим"), которая специфично узнает определенный субстрат и катализирует химическую модификацию данного субстрата. Основаниями нуклеиновой кислоты в каталитической нуклеиновой кислоте могут являться основания A, C, G, T (а также U для РНК).
Как правило, каталитическая нуклеиновая кислота содержит антисмысловую последовательность для специфичного узнавания целевой нуклеиновой кислоты, а также нуклеиновую кислоту, проявляющую гидролизирующую ферментативную активность (также упоминаемую в настоящей заявке как "каталитический домен"). Типы рибозимов, которые являются наиболее предпочтительными в настоящем изобретении, включают рибозим типа hammerhead (Haseloff and Gerlach, 1988, Perriman et al., 1992) и шпилечный рибозим (Shippy et al., 1999).
Рибозимы в растениях ячменя изобретения и ДНК, кодирующая рибозимы, могут быть химически синтезированы с использованием способов, известных в уровне техники. Рибозимы могут быть также получены из молекулы ДНК (которая после транскрипции дает молекулу РНК), функционально связанной с промотором РНК-полимеразы, например, промотором РНК-полимеразы T7 или РНК-полимеразы SP6. В случае, когда вектор также содержит промотор РНК-полимеразы, функционально связанный с молекулой ДНК, рибозим может быть получен in vitro при инкубировании с РНК-полимеразой и нуклеотидами. В отдельном варианте осуществления ДНК может быть встроена в кассету экспрессии или кассету транскрипции. После синтеза молекула РНК может быть модифицирована посредством лигирования с молекулой ДНК, обладающей способностью стабилизировать рибозим и делающей его устойчивым к гидролизу РНКазой.
Как и в случае с антисмысловыми полинуклеотидами, описанными в настоящей заявке, каталитические полинуклеотиды также должны быть способны к гибридизации с целевой молекулой нуклеиновой кислоты (например, мРНК, кодирующей гордеин ячменя) при "физиологических условиях", а именно, при условиях внутри клетки ячменя.
РНК-интерференция
РНК-интерференция (РНКи) особенно полезна в целях специфичного ингибирования продукции специфического белка. Не желая ограничиваться в соответствии с какой-либо теорией, Waterhouse и сотр. (1998) предложили модель механизма, по которому днРНК (двунитевая РНК) может использоваться для уменьшения продукции белка. Данная технология основана на присутствии молекул днРНК, которые содержат последовательность, которая по существу идентична последовательности мРНК целевого гена или его части, в данном случае мРНК, кодирующей полипептид согласно изобретению. Удобно то, что днРНК может синтезироваться с одного промотора в рекомбинантном векторе или хозяйской клетке, в которых смысловая и антисмысловая последовательности фланкированы чужеродной последовательностью, которая обеспечивает гибридизацию смысловой и антисмысловой последовательности с формированием молекулы днРНК, в которой чужеродная последовательность формирует петлевую структуру. Создание и получение подходящих молекул днРНК для настоящего изобретения лежит в пределах компетенции специалиста, квалифицированного в данной области техники, в частности см. Waterhouse et al. (1998), Smith et al. (2000), WO 99/32619, WO 99/53050, WO 99/49029, а также WO 01/34815.
В одном примере вводят ДНК, которая направляет синтез, по меньшей мере, частично двунитевого (дуплексного) РНК-продукта (продуктов), обладающего гомологией к целевому гену, который надлежит инактивировать. Таким образом, ДНК включает и смысловую, и антисмысловую последовательности, которые при транскрипции в РНК могут гибридизоваться с формированием области двунитевой РНК. В предпочтительном варианте осуществления смысловая и антисмысловая последовательности разделены спейсерной областью, которая включает интрон, подвергаемый сплайсингу при транскрипции в РНК. Данная схема, как было показано, приводит к более высокой эффективности сайленсинга гена. Двунитевая область может включать одну или две молекулы РНК, транскрибируемые либо с одной области ДНК, либо с двух. Присутствие двунитевой молекулы, как полагают, индуцирует ответ эндогенной системы растения, которая разрушает как двунитевую РНК, так и гомологичный РНК-транскрипт целевого гена растения, эффективно уменьшая или устраняя активность целевого гена.
Длина смысловой и антисмысловой последовательностей, которые гибридизуются, должна, для каждой, составлять, по меньшей мере, 19 непрерывных нуклеотидов, предпочтительно, по меньшей мере, 30 или 50 нуклеотидов и более предпочтительно, по меньшей мере, 100, 200, 500 или 1000 нуклеотидов. Может использоваться полноразмерная последовательность, соответствующая полному транскрипту гена. Длины, наиболее предпочтительно составляют 100-2000 нуклеотидов. Степень идентичности смысловой и антисмысловой последовательностей целевому транскрипту должна составлять, по меньшей мере, 85%, предпочтительно, по меньшей мере, 90%, а более предпочтительно 95-100%. Молекула РНК, безусловно, может включать чужеродные последовательности, которые могут стабилизировать молекулу. Молекула РНК может экспрессирваться под контролем промоторов РНК-полимеразы II или РНК-полимеразы III. Примеры последнего включают промоторы тРНК или мяРНК.
Предпочтительная молекула малой интерферирующей РНК ("миРНК") включает нуклеотидную последовательность, которая идентична последовательности из приблизительно 19-21 нуклеотида целевой мРНК. Предпочтительно, целевая последовательность мРНК начинается с динуклеотида AA, имеет содержание GC приблизительно 30-70% (предпочтительно 30-60%, более предпочтительно 40-60% и, наиболее предпочтительно приблизительно 45-55%), и не имеет высокой идентичности к любой нуклеотидной последовательности, за исключением целевой последовательности в геноме растения ячменя, в которое она должна быть введена, например, при определении с помощью стандартного поиска BLAST.
МикроРНК
МикроРНК регуляция представляет собой специализированное ответвление пути РНК сайленсинга, которая развилась в способ регуляции генов, отличный от обычного РНКи/ПТСГ. МикроРНК является особым классом малых РНК, которые кодируются геноподобными элементами, организованными в характерные инвертированные повторы. При транскрипции с генов микроРНК синтезируются РНК-предшественники со структурой стебель-петля, из которых впоследствии процессируются микроРНК. МикроРНК обычно имеют длину приблизительно 21 нуклеотид. Образовавшиеся микроРНК включаются в RISC-подобные комплексы, содержащие специфическое семейство белков Argonaute, которые вызывают сиквенс-специфичную репрессию гена (Millar and Waterhouse, 2005; Pasquinelli et al, 2005; Almeida and Allshire, 2005).
Косупрессия
Другим молекулярным биологическим подходом, который может использоваться, является косупрессия. Механизм косупрессии полностью не изучен, однако как предполагают, она включает посттранскрипционный сайленсинг генов (ПТСГ), и в данном отношении может быть весьма схож со многими примерами антисмысловой супрессии. Она включает введение дополнительной копии гена или его фрагмента в растение, в смысловой ориентации относительно промотора для его экспрессии. Размер смыслового фрагмента, его соответствие областям целевого гена, а также степень идентичности его последовательности целевому гену - такие же, как и в случае антисмысловых последовательностей, описанных выше. В некоторых случаях дополнительная копия генной последовательности нарушает экспрессию целевого гена растения. Для получения информации относительно методов осуществления косупрессии см. WO 97/20936 и EP 0465572.
Конструкции нуклеиновых кислот
Конструкции нуклеиновых кислот, пригодные для получения трансгенных растений, могут быть легко созданы с использованием стандартных методов.
При введении области, кодирующей мРНК, конструкция может включать последовательности интронов. Указанные последовательности интронов могут способствовать экспрессии трансгена в растении. Термин "интрон" используется в стандартном значении и означает генетический сегмент, который транскрибируется, но при этом не кодирует белок и вырезается из РНК перед трансляцией. Интроны могут быть введены в 5'-UTR или кодирующую область, если трансген кодирует транслируемый продукт, или в любой другой участок транскрибируемой области, если трансген не кодирует транслируемый продукт. Впрочем, в предпочтительном варианте осуществления, любая область, кодирующая полипептид, представлена в виде единой открытой рамки считывания. Как осведомлен квалифицированный специалист, такие открытые рамки считывания могут быть получены путем обратной транскрипции мРНК, кодирующей полипептид.
Чтобы обеспечить надлежащую экспрессию гена, кодирующего целевую мРНК, конструкция нуклеиновой кислоты обычно включает один или более регуляторных элементов, таких как промоторы, энхансеры, а также последовательности терминации транскрипции или полиаденилирования. Такие элементы известны в уровне техники.
Область инициации транскрипции, включающая регуляторный элемент(ы), может присутствовать для обеспечения регулируемой или конститутивной экспрессии в растении. Предпочтительно, экспрессия, по меньшей мере, происходит в клетках семени.
Был описан ряд конститутивных промоторов, которые активны в клетках растения. Подходящие промоторы для конститутивной экспрессии в растениях включают, помимо прочих, 35S промотор вируса мозаики цветной капусты (CaMV), 35S промотор вируса мозаики норичника (FMV), промотор палочковидного вируса сахарного тростника, промотор вируса желтой мозаики коммелины, легкоиндуцируемый промотор маленькой субъединицы рибулозо-1,5-бисфосфаткарбоксилазы, промотор цитозольной триозофосфат-изомеразы риса, промотор аденинфосфорибозилтрансферазы Arabidopsis, промотор гена актина 1 риса, промоторы маннопинсинтазы и октопинсинтазы, промотор Adh, промотор сахарозосинтазы, промотор комплекса R генов, а также промотор гена хлорофилл α/β связывающего белка. Перечисленные промоторы использовались для создания ДНК векторов, которые экспрессировались в растениях; см., например, WO 84/02913. Все указанные промоторы использовались для создания различных типов экспрессируемых в растениях рекомбинантных ДНК векторов.
Промотор может регулироваться такими факторами, как температура, свет или стресс. Обычно регуляторные элементы расположены на 5'-конце экспрессируемой генетической последовательности. Конструкция может также содержать другие элементы, которые усиливают транскрипцию, такие как nos-3' или ocs-3' области полиаденилирования или терминаторы транскрипции.
5'-нетранслируемые лидерные последовательности могут быть получены из промотора, выбранного для экспрессии последовательности гетерологичного гена, и могут быть специфично модифицированы, если необходимо увеличить трансляцию мРНК. Для получения информации относительно оптимизации экспрессии трансгенов, см. обзор Koziel et al., (1996). 5'-нетранслируемые области могут быть также получены из РНК вирусов растений (вируса табачной мозаики, вируса гравировки табака, вируса карликовой мозаики кукурузы, вируса мозаики люцерны и т.д.), из подходящих эукариотических генов, генов растений (пшеницы и сигнальной последовательности гена хлорофил a/b связывающего белка кукурузы), или из последовательности синтетического гена. Настоящее изобретение не ограничивается применением конструкций, в которых нетранслируемая область получена из 5'-нетранслируемой последовательности, которая сопровождает последовательность промотора. Лидерная последовательность может быть также получена от чужеродного промотора или кодирующей последовательности. Лидерные последовательности, которые могут применяться в рамках настоящего изобретения, включают лидерную последовательность белка Hsp70 кукурузы (US 5362865 и US 5859347), а также омега-элемент ВТМ.
Терминация транскрипции обеспечивается посредством 3'-нетранслируемой ДНК последовательности, функционально связанной в химерном векторе с целевым полинуклеотидом. 3'-нетранслируемая область рекомбинантной молекулы ДНК содержит сигнал полиаденилирования, который функционирует в растениях, вызывая присоединение аденилат нуклеотидов к 3'-концу РНК. 3'-нетранслируемая область может быть получена из различных генов, которые экспрессируются в клетках растений. Обычно используют 3'-нетранслируемую область нопалинсинтазы, 3'-нетранслируемую область гена малой субъединицы Rubisco, 3'-нетранслируемую область гена запасного белка 7S сои. Также подходящими являются транскрибируемые 3'-нетранслируемые области, содержащие сигнал полиаденилирования генов Ti-плазмиды (от англ. tumor-inducing -опухолегенный) Agrobacterium tumefaciens.
Как правило, конструкция нуклеиновых кислот включает селективный маркер. Селективные маркеры помогают в идентификации и скрининге растений или клеток, которые были трансформированы экзогенной молекулой нуклеиновой кислоты. Селективный маркерный ген может придавать клеткам ячменя устойчивость к антибиотику или гербициду, или позволять утилизацию таких субстратов, как манноза. Селективный маркер предпочтительно придает клеткам ячменя устойчивость к гигромицину.
Предпочтительно, конструкция нуклеиновых кислот стабильно интегрирована в геном растения. Соответственно, нуклеиновая кислота включает необходимые элементы, которые позволяют молекуле интегрироваться в геном, или конструкцию помещают в подходящий вектор, который может интегрироваться в хромосому растительной клетки.
Один из вариантов осуществления настоящего изобретения включает применение рекомбинантного вектора, который включает, по меньшей мере, трансген, описанный в настоящей заявке, встроенный в любой вектор, способный доставлять молекулу нуклеиновой кислоты в хозяйскую клетку. Такой вектор содержит гетерологичные последовательности нуклеиновых кислот, которые являются последовательностями нуклеиновых кислот, которые не присутствуют в природе, слитые с молекулами нуклеиновых кислот настоящего изобретения, и которые предпочтительно происходят из вида, отличного от видов, из которых происходит молекула (молекулы) нуклеиновой кислоты. Вектор может являться РНК или ДНК, прокариотическим или эукариотическим, и обычно является вирусом или плазмидой.
Множество векторов, подходящих для стабильной трансфекции клеток растений или создания стабильных трансгенных растений были описаны, например, в Pouwels et al., Cloning Vectors: A Laboratory Manual, 1985, supp. 1987; Weissbach and Weissbach, Methods for Plant Molecular Biology, Academic Press, 1989; а также Gelvin et al., Plant Molecular Biology Manual, Kluwer Academic Publishers, 1990. Как правило, растительные векторы экспрессии включают, например, один или несколько растительных генов, клонированных под транскрипционным контролем 5'- и 3'-регуляторных последовательностей, а также доминантный селективный маркер. Такие растительные векторы экспрессии могут также содержать промоторную регуляторную область (например, регуляторную область, контролирующую индуцируемую или конститутивную, регулируемую внешними условиями или развитием, или клеточно- или тканеспецифичную экспрессию), сайт инициации транскрипции, сайт связывания рибосомы, сигнал процессинга РНК, сайт терминации транскрипции и/или сигнал полиаденилирования.
Трансгенные растения
Трансгенные растения ячменя, как определено в рамках настоящего изобретения, включают растения (а также части и клетки указанных растений) и их потомство, которые были генетически изменены с использованием методов генной инженерии, с целью вызвать продукцию, по меньшей мере, одного полинуклеотида и/или полипептида в желаемом растении или органе растения. Трансгенные растения могут быть получены с использованием методов, известных из уровня техники, таких как описанные в общих чертах в A. Slater et al., Plant Biotechnology - The Genetic Manipulation of Plants, Oxford University Press (2003), а также в P. Christou and H. Klee, Handbook of Plant Biotechnology, John Wiley and Sons (2004).
В предпочтительном варианте осуществления трансгенные растения являются гомозиготными по каждому гену, который был введен (трансгену), в результате чего их потомство не расщепляется по желаемому фенотипу. Трансгенные растения могут также являться гетерозиготными по введенному трансгену (трансгенам), например, в потомстве F1, которое было выращено из гибридного семени. Такие растения могут обеспечить преимущества, такие как гибридная сила, известные из уровня техники.
Были описаны четыре общих метода прямой доставки гена в клетки: (1) химические методы (Graham et al., 1973); (2) физические методы, такие как микроинъекции (Capecchi, 1980); электропорация (см., например, WO 87/06614, US 5472869, 5384253, WO 92/09696 и WO 93/21335), а также генная пушка (см., например, US 4,945,050 и US 5,141,131); (3) вирусные векторы (Clapp, 1993; Lu et al., 1993; Eglitis et al., 1988); и (4) рецепторно-опосредованные механизмы (Curiel et al., 1992; Wagner et al., 1992).
Методы ускорения, которые могут использоваться, включают, например, бомбардировку микрочастицами и т.п. Одним из примеров метода доставки трансформирующей молекулы нуклеиновой кислоты в клетки растений является бомбардировка микрочастицами. Данный метод был описан в обзоре Yang et al., Particle Bombardment Technology for Gene Transfer, Oxford Press, Oxford, England (1994). Небиологические частицы (микрочастицы), которые могут быть покрыты нуклеиновыми кислотами и введены в клетки посредством движущей силы. Примеры частиц включают частицы из вольфрама, золота, платины и т.п. Особым преимуществом бомбардировки микрочастицами, в дополнение к тому, что данный метод является эффективным способом воспроизводимой трансформации однодольных, состоит в том, что не требуется ни выделение протопластов, ни восприимчивость к инфекции Agrobacterium. Иллюстративным вариантом осуществления способа доставки ДНК в клетки кукурузы (Zea mays) с помощью ускорения является биолистическая система доставки α-частиц, которая может использоваться, для пропускания частиц, покрытых ДНК, через экран, например, из нержавеющей стали или экран Nytex, на поверхность фильтра, покрытую суспензией клеток кукурузы. Система доставки частиц, подходящая для применения в настоящем изобретении, является пушкой PDS-1000/He с ускорением гелием, поставляемой Bio-Rad Laboratories.
Для бомбардировки, клетки в суспензии могут быть сконцентрированы на фильтрах. Фильтры, содержащие бомбардируемые клетки, помещают на соответствующем расстоянии под пластиной, останавливающей микрочастицы. Если необходимо, между пушкой и бомбардируемыми клетками также помещают один или более экранов.
В альтернативе незрелые зародыши или другие целевые клетки могут быть нанесены на твердую культуральную среду. Бомбардируемые клетки помещают на соответствующем расстоянии под пластиной, останавливающей микрочастицы. Если необходимо, между ускорителем и бомбардируемыми клетками также помещают один или более экранов. С помощью способов, изложенных в настоящей заявке, можно получить до 1000 или более очагов клеток, транзиентно экспрессирующих маркерный ген. Число клеток в очаге, которые экспрессируют продукт экзогенного гена через 48 часов после бомбардировки обычно варьирует от одной до десяти и в среднем составляет одну - три клетки.
В трансформации с помощью бомбардировки можно оптимизировать условия культивирования перед бомбардировкой и параметры бомбардировки, чтобы получить максимальное количество стабильных трансформантов. В данном методе важны как физические, так и биологические параметры бомбардировки. Физические факторы включают манипуляции с осаждением ДНК/микрочастиц или факторы, которые влияют на полет и скорость макро- или микрочастиц. Биологические факторы включают все стадии, включаемые в манипуляции с клетками до и непосредственно после бомбардировки, осмотическое регулирование целевых клеток, направленное на смягчение травмы, связанной с бомбардировкой, а также свойства трансформирующей ДНК, такие как линеаризованная ДНК или интактные суперскрученные плазмиды. Предполагают, что манипуляции, проводимые перед бомбардировкой, особенно важны для успешной трансформации незрелых зародышей.
В другом альтернативном варианте осуществления могут быть стабильно трансформированы пластиды. Описан способ трансформации пластид высших растений, включающий доставку с помощью пушки на частицах ДНК, содержащей селективный маркер, и направление ДНК в геном пластиды посредством гомологичной рекомбинации (US 5451513, US 5545818, US 5877402, US 5932479 и WO 99/05265).
Таким образом, рассматривается, что может потребоваться регулировать различные аспекты параметров бомбардировки в мелкомасштабных исследованиях, чтобы полностью оптимизировать условия. Может в частности потребоваться регулировать физические параметры, такие как расстояние промежутка, расстояние полета, расстояние ткани, а также давление гелия. Можно также минимизировать травматичные факторы, изменяя условия, которые влияют на физиологическое состояние реципиентных клеток, и поэтому это может влиять на эффективность трансформации и интеграции. Например, осмотическое состояние, гидратация ткани и фаза субкультуры или клеточный цикл реципиентных клеток могут регулироваться для оптимальной трансформации. Выполнение других обычных регуляций известно специалистам в данной области техники с учетом настоящего описания.
Перенос с помощью Agrobacterium является широко применяемой системой для введения генов в клетки растения, поскольку ДНК может быть введена в целые ткани растения, обходя, таким образом, потребность в регенерации интактного растения из протопласта. Применение агробактериальных растительных, интегративных векторов для введения ДНК в клетки растения известно в уровне техники (см., например, US 5177010, US 5104310, US 5004863, US 5159135). Кроме того, интеграция T-ДНК является относительно точным процессом, который приводит к малому числу перестроек. Область ДНК, которая подвергается переносу, обозначена бордерными последовательностями, при этом находящаяся между ними ДНК обычно встраивается в геном растения.
Современные векторы для агробактериальной трансформации способны к репликации в E. Coli, как и в Agrobacterium, обеспечивая удобство при манипуляциях, как описано в Klee et al., In: Plant DNA Infectious Agents, Hohn and Schell, eds., Springer-Verlag, New York, pp. 179-203 (1985). Кроме того, технологические достижения в области векторов для переноса генов с помощью Agrobacterium привели к улучшениям в расположении генов и сайтов рестрикции в векторах, что облегчает конструирование векторов, способных к экспрессии генов, кодирующих различные полипептиды. Описанные векторы несут удобные мультилинкерные области между промотором и сайтом полиаденилирования для прямой экспрессии встраиваемых генов, кодирующих полипептид, и являются подходящими в рамках настоящего изобретения. Кроме того, для трансформации может использоваться Agrobacterium, содержащая активные и неактивные Ti-гены. В тех разновидностях растений, в которых агробактериальная трансформация эффективна, данный метод является предпочтительным благодаря простоте и изученности переноса генов.
Трансгенное растение, полученное с применением методов агробактериальной трансформации, обычно содержит один генетический локус на одной хромосоме. Такие трансгенные растения могут быть упомянуты как гемизиготные по введенному гену. Более предпочтительным является трансгенное растение, которое является гомозиготным по введенному структурному гену, то есть, трансгенное растение, которое содержит два добавленных гена, один ген в одном и том же локусе на каждой хромосоме в паре хромосом. Гомозиготное трансгенное растение может быть получено при половом размножении (самоопылении) поколения независимого трансгенного растения после расщепления, которое содержит один добавленный ген, проращивании части полученных семян и анализе полученных растений на присутствие целевого гена.
Следует также понимать, что два различных трансгенных растения могут быть скрещены с получением потомства, которое содержит два независимо расщепленных экзогенных гена. Самоопыление соответствующего потомства может дать растения, которые являются гомозиготными по обоим экзогенным генам. Обратное скрещивание с родительским растением и ауткросс с нетрансгенным растением также рассматривается, поскольку является вегетативным размножением. Описания других способов размножения, которые обычно используются для различных признаков и культур, можно найти в Fehr, In: Breeding Methods for Cultivar Development, Wilcox J. ed., American Society of Agronomy, Madison Wis (1987).
Трансформация протопластов растения может быть проведена с использованием методов, основанных на преципитации с фосфатом кальция, обработке полиэтиленгликолем, электропорации и комбинациях перечисленных методов. Применение указанных систем к различным видам растений зависит от способности восстанавливать специфическую линию растения из протопластов. Примеры способов регенерации злаков из протопластов описаны в Fujimura et al., 1985; Toriyama et al., 1986; Abdullah et al., 1986.
Также могут использоваться другие методы трансформации клетки, которые включают, помимо прочего, введение ДНК в растения посредством прямого переноса ДНК в пыльцу, прямую инъекцию ДНК в репродуктивные органы растения или прямую инъекцию ДНК в клетки незрелых зародышей, с последующей регидратацией высушенных зародышей.
Регенерация, развитие и культивирование растений из одного трансформированного протопласта растения или из различных трансформированных эксплантов известны в уровне техники (Weissbach et al., In: Methods for Plant Molecular Biology, Academic Press, San Diego, Calif., (1988)). Указанный процесс регенерации и роста обычно включает стадии селекции трансформированных клеток, культивирования полученных отдельных клеток в обычных стадиях развития эмбриона до стадии давшего корни ростка. Трансгенные зародыши и семена регенерируют аналогичным путем. Затем полученные трансгенные побеги с корнями высаживают в подходящую среду для роста растения, такую как почва.
Развитие или регенерация растений, содержащих чужеродный, экзогенный ген, известны из уровня техники. Предпочтительно, регенерированные растения являются самоопыляемыми, чтобы обеспечивать получение гомозиготных трансгенных растений. В других случаях, пыльца, получаемая от регенерированных растений, опыляет выращенные из семян растения агротехнически важных линий. В свою очередь, пыльца из растений указанных важных линий используется для опыления регенерированных растений. Трансгенное растение настоящего изобретения, содержащее целевую экзогенную нуклеиновую кислоту, выращивают с использованием методов, известных специалисту, квалифицированному в данной области техники.
Способы трансформации двудольных растений, прежде всего при помощи Agrobacterium tumefaciens, и получения трансгенных растений были описаны для хлопка (US 5004863, US 5159135, US 5518908), сои (US 5569834, US 5416011), капусты (US 5463174), арахиса (Cheng et al., 1996) и гороха (Grant et al., 1995).
Способы трансформации злаковых растений, таких как ячмень, введения генетической модификации в растение путем введения экзогенной нуклеиновой кислоты и регенерация растений из протопластов или незрелых зародышей растения известны в уровне техники, см. например, CA 2092588, AU 61781/94, AU 667939, US 6100447, PCT/US97/10621, US 5589617, US 6541257 и WO 99/14314. Предпочтительно, трансгенные растения ячменя получают с помощью методов трансформации, опосредованной Agrobacterium tumefaciens. Векторы, несущие целевую конструкцию нуклеиновой кислоты, могут быть введены в регенерируемые клетки ячменя культивируемых тканей растений или эксплантов, или подходящие растительные системы, такие как протопласты.
Регенерируемые клетки ячменя предпочтительно взяты из щитка незрелых зародышей, зрелых зародышей, каллуса, полученного из них, или меристематической ткани. В целях подтверждения присутствия трансгенов в трансгенных клетках и растениях могут быть выполнены амплификация с помощью полимеразной цепной реакции (ПЦР) или Саузерн блот-анализ, с использованием методов, известных специалистам, квалифицированным в данной области техники. Продукты экспрессии трансгенов, в зависимости от природы продукта, могут быть обнаружены различными способами, которые включают Вестерн-блоттинг и ферментный анализ. Одним из наиболее удобных способов количественного анализа экспрессии белка и обнаружения репликации в различных тканях растения состоит использовании репортерного гена, такого как GUS. После того, как трансгенные растения были получены, они могут быть выращены с получением растительной ткани или частей растения, имеющих нужный фенотип. Могут быть собраны ткань растения или части растения, и/или получены семена. Семя может служить источником для выращивания дополнительных растения, ткани или части которых имеют нужные характеристики.
TILLING-введение индуцированных локальных повреждений в геномах
Растения изобретения могут быть получены с применением способа, известного как TILLING (от англ. Targeting Induced Local Lesions in Genomes - введение индуцированных локальных повреждений в геномах). На первом этапе в популяции растений индуцируют мутации, например, новые замены одной пары оснований, с помощью обработки семян (или пыльцы) химическим мутагеном, а затем отбирают растения из тех поколений, в которых мутации стабильно наследованы. ДНК экстрагируют, а семена, полученные от всех представителей популяции, сохраняют, чтобы создать ресурс, к которому можно периодически обращаться с течением времени.
Для анализа TILLING создают ПЦР праймеры, чтобы специфично амплифицировать единственный целевой ген-мишень. Специфичность особенно важна, если мишенью является член семейства генов или часть полиплоидного генома. Затем для амплификации ПЦР-продуктов из объединенной ДНК некоторого количества отдельных растений могут применяться меченые красителем праймеры. Указанные продукты ПЦР денатурируют и повторно отжигают с формированием несовпадающих пар оснований. Мизматчи, или гетеродуплексные пары, представляют собой как природные однонуклеотидные полиморфизмы (SNP или снипы) (то есть, несколько растений из популяции могут нести один и тот же полиморфизм), так и индуцированные SNP (то есть, только редкие отдельные растения могут проявлять мутацию). После формирования гетеродуплекса используют эндонуклеазу, такую как Cel I, которая узнает и расщепляет неспаренную ДНК, что служит ключом к обнаружению новых SNPs в пределах TILLING популяции.
С помощью данного подхода можно провести скрининг многих тысяч растений, чтобы идентифицировать растение с заменой одного основания, а также с короткими вставками или делециями (1-30 п.н.), в любом гене или определенной области генома. Анализируемые геномные фрагменты могут отличаться по размеру приблизительно от 0,3 до 1,6 тпн. При 8-кратном объединении, фрагменты 1,4 тпн (без учета концов фрагментов, на которых обнаружение SNP проблематично из-за шума) и 96 дорожек в анализе, данная комбинация позволяет провести скрининг до миллиона пар оснований геномной ДНК за один анализ, что делает TILLING высокопроизводительным методом. TILLING также описан в Slade and Knauf (2005), а также в Henikoff et al., (2004).
В дополнение к возможности эффективного обнаружения мутаций, высокая производительность метода TILLING идеальна для обнаружения природных полиморфизмов. Таким образом, детальное сравнение неизвестной гомологичной ДНК посредством образования гетеродуплексов с известной последовательностью показывает число и положение полиморфных участков. Идентифицируют и нуклеотидные замены, и короткие вставки, и делеции, которые включают, по меньшей мере, некоторое количество повторных полиморфизмов. Данный метод назвали Ecotilling (Comai et al., 2004).
Каждый SNP регистрируют по его приблизительному положению в пределах нескольких нуклеотидов. Таким образом, каждый гаплотип может быть внесен в базу на основе его изменчивости. Данные последовательности могут быть получены с относительно малым возрастающим усилием, используя аликвоты той же амплифицированной ДНК, которая используется в анализе с расщеплением мизматчей. Левый или правый секвенирующие прамеры для одной реакции выбрают по близости его положения к полиморфизму. Программное обеспечение секвенатора выполняет многократное выравнивание и обнаруживает изменение оснований, которое в каждом случае подтверждает полосу на геле.
“Ecotilling” может быть выполнен с меньшими затратами по сравнению с полным секвенированием, методом, который в настоящее время используется для обнаружения большинства SNP. Планшеты, содержащие матрицы экотипической ДНК, могут быть подвергнуты скринингу, без потребности в объединении ДНК, полученной из растений, подвергнутых мутагенезу. Поскольку детектирование выполняют на гелях с разрешением почти в пару оснований, а фоновые образцы однородны вдоль дорожек, полосы, которые имеют идентичный размер, могут быть сравнены, что позволяет, таким образом, обнаружить и провести генотипирование SNP в одной стадии. Таким образом, полное секвенирование SNP является простым и эффективным, что более всего обусловлено тем фактом, что аликвоты одних и тех же ПЦР-продуктов, используемых в скрининге, могут быть подвергнуты секвенированию ДНК.
ПРИМЕРЫ
Пример 1. Материалы и Методы
Выделение и очистка проламинов
Для выделения проламинов из злаков муку из цельного зерна (10 г) перемешивали в течение 30 минут при 25°C в 200 мл буфера, содержащего 20 мМ триэтаноламина-HCl (TEA), 1% (вес./об.) аскорбата натрия, 1% (вес./об.) полиэтиленгликоля (мол.вес 6000; PEG6000) и 200 мкл смеси ингибиторов протеаз растений (Sigma #P9599), все с pH 8. Суспензию центрифугировали при 7000 g в течение 15 минут, осадок промывали не менее двух раз, чтобы удалить белки, растворимые в водном буфере. Проламины в промытом осадке растворяли в 40 мл 50% (об./об.) пропан-2-ола, содержащего 1% (вес./об.) дитиотреитола (ДТТ), 1% (вес./об.) PEG6000, 1% (вес./об.) аскорбата натрия, перемешивали в течение 30 мин при 25°C. Суспензию центрифугировали, а проламины осаждали из супернатанта 2 объемами пропан-2-ола и хранили при -20°C. В случае необходимости аликвоту, эквивалентную 10 г муки, осаждали центрифугированием при 160 g при 4°C в течение 10 мин, осадок повторно растворяли в 10 мл буфера (буфера A), который содержал 25 мМ TEA, 8 M свежеприготовленной деионизированной мочевины и 1% ДТТ, все с pH 6, или в других буферах, как описано.
Суммарную проламиновую фракцию выделяли из каждого образца зерна с помощью обращено-фазовой быстрой жидкостной хроматографии белка (ОФ FPLC) следующим образом: Проламины (200 мкл) вводили в 1 мл колонку RPC Resource (Pharmacia), соединенную с аналогичной 3 мл колонкой. Колонку промывали 2 мл 95% растворителя A/5% растворителя B и элюировали проламины в 30 мл, в линейном градиенте с 95% растворителя A/5% растворителя B до 100% растворителя B со скоростью 2 мл/мин. Растворителем A являлась 0,1% (об./об.) трифторуксусная кислота (ТФУ) в воде, растворителем B являлась 0,1% (об./об.) ТФУ в 60% (об./об.) водном ацетонитриле. Фракции элюата, соответствующие пикам белка, объединяли. Контрольные фракции растворителя аналогично объединяли из прогонов без ввода белка.
Далее гордеины ячменя фракционировали с помощью ОФ FPLC следующим образом: Использовали методики, как и приведенные выше, за исключением того, что градиент элюции отличался тем, чтобы концентрация растворителя B составляла 50% при 4 мл, 52% при 17 мл, 56% при 34 мл, 58% при 37 мл, 60% при 41 мл, 62% при 44 мл, 64% при 47 мл, 66% при 50 мл, 100% при 53 мл, 100% при 57 мл. Отбирали фракции объемом 1 мл, а фракции 11-14 (#1), 19-23 (#2), 31-34 (#3), 43-51 (#4), 53-58 (#5) и 63-64 (#6), соответствующие пикам A280, объединяли.
Аналитические методы
Проламиновые фракции растворяли в 6 M мочевине, 2% (вес./об.) ДСН, 1% (вес./об.) ДТТ, 0,01% (вес./об.) бромфенолового синего, 0,0625 М Трис-HCl (pH 6,8) при 25°C, и анализировали с помощью электрофореза в ДСН-ПААГ следующим образом. Аликвоту 5 мкл раствора проламин-ДСН наносили на гель ДСН-ПААГ, используя готовые гели 245x110x0,5 мм с градиентом полиакриламида 8-18% (ExcelGel Pharmacia), и проводили форез при 600В в течение 90 мин при 15°C. Гели промывали в 40% MeOH/10% уксусной кислоте в течение 30 мин, а затем в воде в течение 10 мин. Проламины окрашивали путем погружения геля в 0,06% (вес./об.) коллоидный Coomassie G250 в 8,5% фосфорной кислоте в течение 30 мин, после чего гель отмывали в воде в течение ночи. Каждый гель калибровали стандартным 10 кДа белковым маркером (BenchMark, Invitrogen).
Гордеиновые фракции также растворяли в 50% (об./об.) водном изопропиловом спирте, 1% (вес./об.) ДТТ, обрабатывали в избытке винилпиридина с целью восстановления дисульфидных связей и анализировали с помощью обращено-фазовой ВЭЖХ (ОФ-ВЭЖХ, Larroque et al., 2000), калибровали с проламинами, выделенными из линий ячменя Riso56 или Riso1508, в которых, в результате мутации, не накапливаются гордеины групп B или C соответственно (Doll, 1983). Уровни белка в экстрактах или фракциях определяли методом Брэдфорд (1976). Как правило, содержание белка измеряли в формате 96-луночных планшетов, добавляя 10 мкл каждого ДТТ/пропан-2-ол супернатанта к 200 мкл разведения 1 к 5 концентрата для анализа белка Coomassie protein assay concentrate (BioRAD) в воде, калибровали против гамма-глобулина, и измеряли поглощение при 595 нм.
Ex vivo анализы T-клеточной токсичности
Проламины (50 мг/мл в 2M мочевине) растворяли в PBS, содержащем 1 мМ CaCl2 с получением концентрации проламина 62,5, 250, 625, 2500 или 6250 мкг/мл, дезамидировали, добавляя 25 мкл каждого раствора к 100 мкл tTG из печени морской свинки (трансглутаминаза (Sigma, T5398), 25мкг/мл tTG в PBS, содержащем 1 мМ CaCl2), и инкубировали в течение 6 ч при 37°C. Недезамидированные растворы приготовляли аналогично, путем инкубирования в отсутствие tTG. Растворители в качестве контроля добавляли, как при максимальных концентрациях проламина. Другие контроли содержали либо известный токсичный пептид ω-глиадин, обозначенный как 626fEE, в концентрации 50 мкг/мл, пептид 626fEE в чистом виде или с анатоксином столбняка (50 светообразующих частиц/мл). Пептид ω-глиадин 626fEE, также известный как DQ2-ω-1, имел аминокислотную последовательность QPEQPFPQPEQPFPWQP (SEQ ID NO:1) и был синтезирован фирмой Mimotopes, Melbourne, Australia. Его идентичность и чистоту (91%) подтверждали с помощью масс-спектрометрии и ВЭЖХ. Анатоксин столбняка получили от Commonwealth Serum Laboratories, Melbourne. Затем все растворы замораживали при -20°C.
Двадцать один пациент с HLA-DQ2+ целиакией, подтвержденной биопсией, придерживавшиеся строгой аглютеновой диеты в течение, по меньшей мере, трех месяцев, ежедневно в течение 3 дней получали 150 г вареного ячменя, употребляемого в качестве части их диеты, которая в остальном оставалась аглютеновой. Гепаринизированную венозную кровь забирали либо непосредственно до, либо через шесть дней после начала диетической провокации, и из каждого образца крови с помощью центрифугирования в градиенте плотности Фиколл-Гипак (Anderson et al., 2000) выделяли мононуклеары периферической крови (PBMC). Клетки PBMC ресуспендировали в полной среде HT-RPMI (Invitrogen), содержащей 10% термоинактивированной, смешанной человеческой AB сыворотки. Дезамидированные или недезамидированные проламины и контрольные растворы размораживали и 25 мкл вносили в лунки, содержащие 100 мкл PBMC (3-8×105 PBMC в лунке). Их культивировали при 37°C в течение ночи в 96-луночных планшетах (MAIP-S-45; Millipore, Bedford, MA). Контрольные культуры получали, добавляя 25 мкл PBS, содержащего 1 мМ CaCl2 (контроль, содержащий только буфер). Конечные концентрации проламина составляли 2,5, 10, 25, 100 или 250 мкг/мл, а конечная концентрация мочевины составляла 50 мМ. Уровень IFN-γ, выработанного в каждой культуре, служивший показателем токсичности каждого проламина, оценивали визуально по формированию пятна, используя вторичные антитела согласно инструкциям производителей (Mabtech, Stockholm, Sweden), и подсчитывали количество пятнообразующих единиц (SFU) с использованием автоматизированного ELISPOT планшет-ридера (AID Autoimmun Diagnostika GmbH, Germany). Результаты были представлены как среднее количество пятнообразующих единиц (SFU) ±SE. Как правило, процентный коэффициент вариации SFU/106 PBMC в анализе составлял 14% на основе шести дублированных анализов положительного контроля, инкубируемого с 0,5x106 клеток у шести пациентов с CD (у всех >20 SFU/лунка).
Статистический анализ
Дисперсионный анализ (ANOVA) или t-критерии с использованием GenStat применяли для определения величины расхождения, наблюдаемого для среднего SFU, показанного T-клетками, выделенными у пациентов с целиакией до (n=10) или после (n=21) диетической провокации, и инкубируемыми с гордеинами, проламинами или контролем.
Кривые отклика для 21 человека после провокации сильно отличались, и существенная доля дисперсии была обусловлена данными различиями. Чтобы принять во внимание различные ответы пациентов, была применена модель случайных коэффициентов. Это - смешанный модельный анализ, который выполняли, используя метод остаточного наибольшего правдоподобия (REML), который позволяет вводить поправку случайных показателей, касающихся субъекта (пациента) и провокации (концентрации белка) в организме пациента. В целях стабилизации существенной неоднородности вариации, перед указанным анализом данные преобразовали логарифмически. Чтобы решить проблему нулевых значений, ко всем данным до логарифмического преобразования прибавляли единицу. Постоянными значениями в модели являлись присутствие или отсутствие tTG, а также используемая гордеиновая фракция, вместе с их взаимодействием.
Гиперболическую модель также применили к непреобразованному среднему SFU для T-клеток от 21 пациента после провокации, взаимодействовавших с шестью tTG гордеиновыми фракциями или четырьмя препаратами злакового проламина, обработанного tTG.
Трансформация ячменя
Трансформированные растения ячменя могут быть получены методом Tingay et al, (1997). Генные конструкции в бинарных векторах могут быть введены в высоко вирулентный штамм Agrobacterium (AGL1) посредством трехродительского конъюгирования, которые затем используют для введения T-ДНК, содержащей трансген и селективный маркерный ген (кодирующий устойчивость к гигромицину, экспрессируемый с промотора CaMV35S), в регенерируемые клетки щитка незрелых зародышей ячменя, следующим образом.
Развивающиеся семена ячменя сорта Golden Promise, через 12-15 дней после периода цветения, удаляли из растущего колоса выращиваемых в теплице растений и стерилизовали в течение десяти минут в 20% (об./об.) растворе гипохлорита натрия с последующей однократной промывкой 95% этанолом и семь раз стерильной водой. Затем зародыши (размером приблизительно 1,5 - 2,5 мм) отделяли от семян в стерильных условиях, и из каждого зародыша вырезали рубчик. Зародыши помещали вырезом вниз на чашку Петри, содержащую среду индукции каллусообразования. Трансконъюгаты Agrobacterium выращивали в питательной среде MG/L (содержащей 5 г маннита, 1 г L-глутаминовой кислоты, 0,2 г KH2PO4, 0,1 г NaCl, 0,1 г MgSO4·7H2O, 5 г триптона, 2,5 г дрожжевого экстракта и 1 мкг биотина в литре, pH 7,0), содержащей спектиномицин (50 мг/л) и рифампицин (20 мг/л) с аэрацией при 28°C до концентрации приблизительно 2-3×108 клеток/мл. Затем приблизительно 300 мкл суспензии клеток добавляли к зародышам в чашке Петри. Через 2 минуты сливали лишнюю жидкость с чашки, а зародышей переворачивали так, чтобы сторона с вырезом (пазушная сторона щитка зародыша) оказывалась сверху. Затем зародышей переносили в новую чашку со средой индукции калусообразования и помещали в темноту на 2-3 дня при 24°C. Зародышей переносили в среду индукции калусообразования с селекцией (50 мкг/мл гигромицина и 15 мкг/мл тиментина).
Зародышей оставляли в данной среде на 2 недели в темноте при 24°C. Затем здоровый каллус разделяли, и помещали в свежую селективную среду, после чего инкубировали еще в течение двух недель при 24°C в темноте. После этого зародышей инкубировали при 24°C на свету в течение 2 недель на среде регенерации, содержащей цитокинин, а затем переносили в среду для корнеобразования, содержащую цитокинин и ауксин в течение трех 2-недельных периодов. Затем молодые растения пересаживали в почвенную смесь и держали на полке с мелкокапельным орошением в течение двух недель, после чего, наконец, переносили в теплицу.
Методы мутагенеза, включающие гамма-облучение
Мутацию генов в ячмене, приводящую к снижению экспрессии D-, C-, B- или γ-гордеинов, можно ввести либо с помощью гамма-облучения, либо посредством химического мутагенеза, например, с использованием этилметансульфоната (EMS). В случае мутации, вызываемой гамма-излучением, семена могут быть облучены при дозе 20-50 кР источником 60Co (Zikiryaeva and Kasimov, 1972). EMS мутагенез может быть выполнен путем обработки семян EMS (0,03%, об./об.) согласно Mullins et al., (1999). В двойном нулевом фоне B+C мутантные зерна могут быть идентифицированы на основе уменьшенного содержания белка или гордеина, или измененной морфологии зерна, и подтверждены методами, описанными выше. Мутанты по одному гену гордеина могут быть скрещены со вторым мутантом для объединения мутаций и получения нетрансгенного сорта ячменя, по существу не содержащего гордеинов в эндосперме.
Пример 2. Токсичность гордеинов ячменя для больных целиакией
Состав проламинов ячменя и других злаков
Проламины выделяли в форме белков, растворимых в водноспиртовой смеси, из злаков, токсичных для больных целиакией, ячменя и пшеницы, менее токсиченого овса и нетоксичной кукурузы, и очищали в одну стадию с помощью ОФ FPLC, как описано в Примере 1. Профили элюции белка проламинов, определенные при A280нм в ОФ FPLC (Фиг.1), показали ряд частично разрешенных пиков отдельных белков, элюированных при резком повышении градиента растворителя. Фракции, содержащие белок после 10 очисток для каждого злака, объединяли и лиофилизировали. Обычный выход проламина из различных злаков (2 г) составил: кукуруза - 10 мг, овес - 23 мг, ячмень - 73 мг, и пшеница - 114 мг. Суммарные проламины из каждого злака лиофилизировали и хранили для тестирования в ex vivo анализе на T-клетках (ниже). Контроли растворителей также получили с помощью ОФ FPLC.
Проламины ячменя (гордеины) также фракционировали с помощью ОФ FPLC, как описано в Примере 1. Профиль элюции, полученный в процессе фракционирования в первоначальном эксперименте, показан на Фиг.2. Получили шесть пиков и выделили из каждого белок. Соответствующие объединенные фракции из двадцати последовательных вводов проб объединяли и лиофилизировали. Обычные выходы из 4 г муки из цельного зерна составляли: фракция 1-19 мг, фракция 2-26 мг, фракция 3-14 мг, фракция 4-104 мг, фракция 5-24 мг, и фракция 6-11 мг.
Идентичность гордеинов в каждой фракции определяли с помощью электрофореза в ДСН-ПААГ, как описано в Примере 1, и подтверждали с помощью аналитической ОФ ВЭЖХ. Результаты показаны на Фиг.3 и 6. ВЭЖХ показала, что фракция #1 содержала приблизительно 39% D-гордеина, масса которого достигала 90 кДа на электрофорезе в ДСН-ПААГ, и приблизительно 61% C-гордеина, масса которого достигала 47 и 48 кДа на электрофорезе в ДСН-ПААГ (Фиг.3, #1). Фракция #2 содержала C-гордеины, как показал электрофорез в ДСН-ПААГ и ВЭЖХ. Фракция #3 содержала широкую полосу белка, которая шла приблизительно на уровне 45 кДа на электрофорезе в ДСН-ПААГ, но которая разделялась на 6 пиков в ВЭЖХ, что соответствует элюции и C-, и B-гордеинов. Состав оценивали с помощью ВЭЖХ, определив содержание приблизительно 43% и 57% гордеинов C и B соответственно. Фракции #4, 5 и 6, содержали B-гордеины; данные фракции могут также содержать небольшое количество гамма-гордеина. Двумерный электрофорез и масс-спектрометрия триптических пептидов (фингерпринт) указанных гордеиновых фракций не дали достаточно уникальных пептидных фрагментов, чтобы однозначно идентифицировать отдельные гордеины. Это может происходить из-за небольших вариаций между последовательностями выделенных гордеинов и последовательностями, доступными в базах данных. Поэтому фракционирование в данном эксперименте привело к обогащению специфических гордеинов из ячменя, но не к полной очистке. Дальнейшая очистка может быть достигнута с помощью дополнительных стадий ОФ FPLC или ОФ FLPC, комбинированной с ионообменными методами.
Образцы каждой гордеиновой фракции обрабатывали или не обрабатывали tTG, которая превращает остатки глутамина в белках в глутамат, а затем лиофилизировали для применения в анализе на T-клетках.
Анализы токсичности
Анализы на T-клетках с использованием PBMC, выделенных у пациентов с подтвержденными случаями целиакии, проводили, как описано в Примере 1, с целью определения токсичности суммарных препаратов проламинов и гордеиновых фракций. PBMC выделяли до и после диетической провокации ячменем, а образцы проламинов обрабатывали tTG, или не обрабатывали. T-клетки, выделенные в группе из 10 больных целиакией до диетической провокации, были невосприимчивы к проламинам. Статистический анализ с использованием ANOVA показал, что отсутствовало какое-либо существенное различие (P=0,77) между средним числом IFN-γ положительных пятен для максимально высоких концентраций обработанных tTG фракций проламинов, пептидов или гордеинов (среднее значение в группе SFU±SE - 1,52±0,18), и контрольной культурой (среднее SFU±SE - 1,40±0,45). Напротив, анализ показал, что T-клетки, не подвергавшиеся провокации, активно реагировали (P<0,001) с анатоксином столбняка, служившим в качестве положительного контроля (среднее SFU±SE - 22,3±4,72), по сравнению с проламинами. Это указывает на то, что выделенные T-клетки являлись функциональными и были способны к реакции с известным токсином, и подтверждает, что в популяциях, выделенных перед диетической провокацией, присутствовало малое количество проламин-реактивных T-клеток.
В отличие от слабого ответа на проламины перед диетической провокацией, T-клетки, выделенные после диетической провокации, являлись очень активными. T-клетки, выделенные у 21 больных целиакией, через 6 дней после провокации, давали сильный ответ на проламины, обработанные tTG, в отличие от T-клеток из подгруппы (n=13) данной группы, реагировавших с недезамидированными проламинами. На Фиг.4 показаны результаты злаков, суммарные проламины ячменя индуцировали максимальное значение SFU, за которым следуют в порядке убывания проламины из пшеницы, овса и затем кукурузы (на Фиг.4 блоки A, B, C, D соответственно). Хотя проламин кукурузы вызывал слабый дозозависимый T-клеточный ответ в данном анализе, обычно он не вызывает ответ при диетической провокации и считается безопасным при целиакии злаком. Переваривание в кишечнике может разрушать эпитопы, присутствующие в цельных проламинах кукурузы, которые оставались интактными в настоящем анализе и стимулировали T-клетки in vitro.
Из гордеиновых фракций фракции #1, #2 и #3 давали более высокие значения SFU по сравнению с гордеиновыми фракциями #4, #5 и #6 (Фиг.5).
Поскольку концентрация проламинов в анализе была увеличена, значение IFN-γ пятен росло в гиперболической манере, наподобие ферментативной кинетики Михаэлиса-Ментен, часто наблюдаемой между ферментом и его субстратом (Фиг.4 и 5), хотя не было ясно, почему это происходило в данных клеточных анализах.
Каждый 96-луночный планшет содержал некоторое количество внутренних положительных и отрицательных контролей. Наблюдали малое, но существенное различие (P<0,001) между средним SFU, при сравнении контрольных культур и контрольных растворителей (контрольные культуры SFU 2,75±0,67 и 1,49±0,24; контрольные растворители SFU 2,64±0,23 и 2,75±0,23, в отсутствие и в присутствии tTG соответственно). Хотя данные значения являлись статистически значимыми, они были очень малыми по сравнению с SFU после провокации в положительных контролях или анализах, включавших проламин. Это подтверждало, что примеси в растворителях не давали ложноположительные сигналы. Контрольный пептид 626fEE давал соответственно высокий ответ (среднее SFU±SE 29,55±4,38 и 33,60±2,97 в отсутствие и в присутствии tTG соответственно). Недостаточное усиление ответа 626fEE при обработке tTG ожидали, поскольку указанный пептид был синтезирован с глутаматом в 10-ом положении, и не требовал обработки tTG для повышения токсичности. Добавление контрольного растворителя существенно не ингибировало положительный ответ пептида 626fEE (P=0,13), подтверждая, что примеси в растворителях не давали ложноотрицательный сигнал. Воспроизводимость контрольных показателей анатоксина столбняка (P=0,193) от планшета к планшету подтверждала, что различия в ответе T-клеток на проламины не были обусловлены расхождениями от планшета к планшету, но отражала различную чувствительность популяций T-клеток, полученных от различных пациентов.
Разброс чувствительности различных пациентов к одной и той же концентрации проламинов составлял до 200 раз. Поэтому к нормализованным данным SFU применили модель случайных коэффициентов REML, и установили, что модель, учитывающая поправку на отклонение в ответах пациентов из-за различных концентраций белка, показала значительно лучшее соответствие данным (P<0,001), чем модель, в которой применяли ответ одного пациента независимо от концентрации, с отклонением в пределах от 1982,28 (1616 df) до 1640,91 (1613 df). Главные эффекты, вызываемые tTG (P<0,001) и проламиновой фракцией (P<0,001), являлись значимыми, причем между ними отсутствовало какое-либо взаимодействие. Это подтверждало, что чувствительные к проламину T-клетки были индуцированы у пациентов с целиакией через шесть дней после диетической провокации ячменем. Обработанные средние значения, на логарифмической шкале, для нормализованных данных SFU составляли 1,613 (без tTG) и 2,026 (с tTG) со среднеквадратической ошибкой (SED) 0,0527, подтверждая, что предварительная обработка tTG оказывала существенное влияние на ответы. Обработанные средние значения для гордеиновых фракций #1 - #6 составляли 1,903, 1,909, 1,956, 1,693, 1,724 и 1,733 соответственно с SED 0,0826. Приведенные результаты указывают, что гордеиновые фракции подразделяются на две группы, различных по токсичности, при этом гордеиновые фракции #1, #2 и #3 формируют более токсичную группу, чем гордеиновые фракции #4, #5 и #6.
Интересно отметить, что наиболее токсичные гордеиновые фракции, были элюированы в обращено-фазовой FPLC и ВЭЖХ вначале, и поэтому являлись более полярными, чем элюированные позже, менее токсичные фракции.
Выводы
T-клетки, выделенные у 21 больного целиакией, через 6 дней после провокации, сильно взаимодействовали с проламинами, обработанными tTG, по сравнению с недезамидированными проламинами, как и следует ожидать для целиакии (Hadjivassiliou et al, 2004; Kim et al., 2004). Это можно объяснить взаимодействием между дезамидированными проламинами и связывающим сайтом в ключевом белке, таком как молекула HLA-DQ2, которая презентирует стимулирующие белки рецепторам на CD4+ T-клетках, вовлекаемых в воспалительный ответ.
Хотя наблюдали измеримые различия в токсичности гордеиновых фракций, все гордеины были значительно более токсичны по сравнению с проламинами кукурузы и овса, которые считаются безопасными для большинства больных целиакией. Статистические анализы показали, что проламины ячменя и гордеиновые фракции #1, #2 и #3 (содержащие D и C гордеины) формировали наиболее токсичную группу. Гордеиновые фракции #4, #5 и #6, содержащие главным образом B-гордеины, а также проламины пшеницы, формировали вторую, менее токсичную группу. Проламины овса и кукурузы формировали наименее токсичную группу. Это указывает, что T-клетки, индуцируемые у больных целиакией в результате провокации ячменем, являлись менее чувствительными к пшенице и овсу. Это может объясняться тем, что доминирующие эпитопы в проламинах ячменя значительно отличаются от эпитопов в проламинах пшеницы и овса. Хотя обработанные данные показывали, что проламины овса являлись значительно менее токсичными по сравнению с проламинами ячменя, между различными пациентами наблюдали пятидесятикратное расхождение результатов при одной и той же концентрации проламина овса, при этом T-клетки пяти из 21 пациента показывали >20 SFU при наибольшей концентрации проламина. Это согласуется с другими сообщениями о существенных ответах на овес при целиакии (Arentz-Hansen et al., 2004; Lundin et al., 2003).
На основе приведенных данных сочли возможным, что при диетической провокации все гордеинсодержащие фракции вызывают существенную кишечную реакцию у больных целиакией. Это позволяет предположить, что все гордеинсодержащие фракции необходимо удалить или модифицировать, чтобы получить ячмень, который является абсолютно нетоксичным для больных целиакией. Также предложили, что в первую очередь необходимо удалить или модифицировать основные компоненты - гордеины B и C.
Пример 3. Получение зерна ячменя с пониженным содержанием гордеинов B и C
Ряд мутантов ячменя, с изменениями в синтезе или накоплении гордеинов, были идентифицированы ранее. Указанные мутанты ячменя были выделены не с целью снижения содержания гордеинов в зерне, а были выделены и отобраны по показателю повышенного содержания лизина в зерне, а впоследствии установили, что они имели пониженное содержание гордеинов.
Мутант Riso 7, впервые описанный Doll et al. (1976), был идентифицирован после обработки быстрыми нейтронами исходного сорта Bomi. Он содержал рецессивную мутацию в гене, которая приводила к 29%-ому снижению уровня проламинов и 10%-ому повышению содержания лизина в белках, по сравнению с Bomi. Снижение содержания бедных лизином проламинов компенсировалось повышением содержания других, относительно богатых лизином заасных белков, что приводило к повышению содержания лизина. Урожай зерна и содержание крахмала снизились на 6% и 7%, соответственно, по сравнению с исходным сортом (Talberg, 1982; Doll, 1983).
Riso 56, впервые описанный Doll et al. (1973), был создан путем вызванной гамма-излучением мутации родительского сорта Carlsberg II. Размер зерна, урожай зерна и содержание проламинов снизилось на 30%, 47% и 25% соответственно, по сравнению с исходным сортом, тогда как содержание лизина в белках мутантного зерна выросло на 13% по сравнению с родительским сортом. Уменьшенное содержание гордеинов было связано с повышением содержания небелкового азота и воды, а также солерастворимых белков (Shewry PR et al., 1980). Высокое содержание лизина в белках Riso 56 было обусловлено рецессивной мутацией на хромосоме 5 (Ullrich and Eslick, 1978) в генетическом локусе, обозначенном Hor2ca (Doll, 1980). Мутация включала делецию 80-90 тпн ДНК из локуса Hor2, который кодировал B-гордеины в ячмене. Экспрессия B-гордеинов в мутанте снизилась на 75%, тогда как экспрессия C-гордеинов выросла в 2 раза (Kreis et al., 1983). Делеция не была связана с транслокацией между хромосомами 2 и 5, которая также присутствовала в Riso 56 (Olsen, 1977).
Riso 527, впервые описанный Doll et al. (1973), также был получен с помощью вызванной гамма-излучением мутацией, но из родительского сорта Bomi. Размер зерна, урожай зерна и содержание проламинов в зерне были снижены на 13%, 25% и 20% соответственно, по сравнению с исходным сортом, тогда как содержание лизина в белках мутанта выросло на 12%. Мутация являлась рецессивной и была локализована в гене на хромосоме 6, обозначенной lys6i (Jensen, 1979). Данный мутант имел пониженное содержание D-гордеинов и повышенное содержание B1-гордеинов (Klemsdal et al., 1987).
Riso 1508 был идентифицирован после EMS-индуцированной мутации родительского сорта Bomi (Doll et al., 1973; Ingerversen et al., 1973; Doll, 1973). Размер зерна, урожай зерна и содержание проламинов в зерне снизились на 8%, 12% и 70% соответственно, по сравнению с исходным сортом, тогда как содержание лизина в белках мутанта выросло на 42%. Высокое содержание лизина было обусловлено рецессивной мутацией в гене, расположенном вблизи центромерной области хромосомы 7 ячменя (Karlsson, 1977). Указанный ген вначале обозначили как sexSc ксении с морщинистым эндоспермом (Ullrich and Eslick, 1977), однако теперь он известен как lysSa (Tallberg, 1977). Сравнительные уровни типов белков в мутантном зерне изменились, при этом выросло содержание водо-растворимого белка (альбумин/глобулины) - с 27% до 46% от общего азота белка семени, и снизилось количество проламинов - на 70% по сравнению с родительским сортом, с 29% до 9% от общего азота белка семени (Ingerversen et al., 1973; Doll, 1973). В Riso 1508 наблюдали четырехкратное увеличение содержания свободных аминокислот и небелкового N, по сравнению с родительским сортом, при выращивании растений в условиях высокого содержания азотных удобрений (Koeie and Kreis, 1978). Shewry с сотр. (1978) подтвердили, что уровень солерастворимого небелкового азота удвоился. Процентное содержание азота семян в форме гордеинов снизилось на 70%, а содержание солерастворимых белков в Riso 1508 увеличилось по сравнению с Bomi на 70%. Детальный молекулярный анализ показал, что уровни B- и C-гордеинов снизились на 80% и 93% соответственно, тогда как содержание D-гордеинов выросло в четыре раза. Указанное влияние на накопление белка было обусловлено изменениями в копийности или стабильности мРНК (Kreis et al., 1984). Это могло быть вызвано повышением метилированием промоторов генов, кодирующих B- и C-гордеины в мутанте Riso 1508 (Sorensen et al., 1996). Меньший размер семени Riso 1508 главным образом был вызван уменьшением синтеза крахмала (Koeie and Breis, 1978; Kreis and Doll, 1980; Doll, 1983). Содержание сахаров выросло в два раза, тогда как синтез крахмала в Riso 1508 снизился примерно на 20-30% по сравнению с исходным сортом. Kreis (1979) сообщил, что в Riso 1508 уровни β-амилазы снизились, тогда как Hejgaard и Boisen (1980) сообщали об аналогичных уровнях β-амилазы.
Hiproly являлся спонтанным мутантом, идентифицированным из зародышевой плазмы эфиопского ячменя CI 3947 (Munck et al., 1970), и имел повышенное на 20-30% содержание суммарного белка и лизина в белке по сравнению с ячменем дикого типа (Doll, 1983). При скрещивании с ячменем дикого типа высокое содержание белка было утеряно, в то время как повышенное содержание лизина в белке сохранилось, показав, что данные признаки наследовались независимо. Повышенное содержание лизина было обусловлено одной рецессивной мутацией в гене lys на хромосоме 7. Мутация вызвала повышение содержания воды и солерастворимых белков и, следовательно, содержание лизина. В отличие от Riso с высоким содержанием лизина, уровни гордеинов и вес семян в мутантах Hiproly в потомстве от беккросса не снизились. Небелковый азот также не был повышен. Содержание β-амилазы выросло в 4 раза (Hejgaard and Boisen, 1980).
Характеристика родительских линий Riso 56 и Riso 1508
Свойства проламинов, накапливаемых родительскими линиями Riso 56 и Riso 1508, подтверждали с помощью электрофореза в ДСН-ПААГ и обращенно-фазовой ВЭЖХ. Солерастворимые белки, экстрагированные из зерна, разделяли с помощью гель-электрофореза и переносили на мембраны (Вестерн-блоттинг). Белки на мембранах окрашивали для определения суммарного белка (Фиг.7, левая сторона) или обрабатывали проламин-специфичным моноклональным антителом (мышиным моноклональным антителом MAb 12224, индуцированным против суммарного глютенинового экстракта, которое детектировало все гордеины и проламины (Skerritt, 1988) (правая сторона), что показало то, что уровни B-гордеинов в Riso 56 были очень низкими, тогда как содержание C-гордеинов, по сравнению с уровнями в Riso 527, выросло. Детектирование антителом подтверждало, что уровень B-гордеинов в экстракте Riso 56 являлся чрезвычайно низким (пунктирный квадрат). Указанные три белка, наблюдаемые в Riso 56, которые перемещались вместе с B-гордеинами, наиболее вероятно, являлись γ-гордеинами. В Riso 1508 было снижено накопление B-гордеинов, в то время как C-гордеины различались с трудом (пунктирный квадрат). Это согласуется с опубликованной литературой. Уровни D-гордеина, который являлся относительно минорным проламиновым компонентом, не казались повышенными при нанесении белка в количествах, используемых в данном геле.
На Фиг.8 показаны относительные уровни различных гордеинов в очищенных экстрактах после анализа обращено-фазовой FPLC. Экстракты гордеинов, эквивалентные 0,2 г муки, анализировали с помощью FPLC, как описано в Примере 1. Таким образом, площадь под хроматограммами A280 являлась пропорциональной относительному содержанию белка в каждом образце. В Riso 56 уровни C-гордеинов выросли на 400%, а B-гордеинов - снизились на 86% по сравнению с родительским сортом Carlsberg II. В Riso 1508 C- и B-гордеины снизились (на 91% и 86% соответственно) по сравнению с родительским сортом Bomi. Полученные результаты были аналогичны опубликованным данным.
Идентификация семян с двумя гордеиновыми мутациями
Растения линий Riso 56 и Riso 1508 скрещивали путем удаления пестиков Riso 1508, а через два дня опыляли их свежей пыльцой Riso 56. Десять семян F1 проращивали и выращивали растения F1, которым позволяли самооплодотворяться. Семена F2 собирали в зрелом виде.
С целью идентификации двойных мутантов в популяции, половину каждого из 288 семян F2 отдельно дробили и истирали в порошок в пластиковой микропробирке, используя шарик из нержавеющей стали, при встряхивании с частотой 30/сек в течение 3 × 1,5 мин в 96-луночной вибромельнице Vibration Mill (Retsch Gmbh, Rheinische). В каждую пробирку добавляли аликвоту (400 мкл) водного буфера для экстракции водорастворимых белков. Буфер содержал 20 мМ триэтиламина-HCl (TEA), 1% (вес./об.) аскорбата натрия, 1% (вес./об.) PEG6000 и ингибитор протеаз растений в разведении 1/1000 (Sigma P9599), pH 8 при комнатной температуре (КТ). Содержимое каждой пробирки снова встряхивали, а затем центрифугировали при 160 g в течение 10 мин при КТ. Нерастворимый в воде осадок муки промывали не менее двух раз аналогичным путем, после чего соответствующие супернатанты объединяли, получив водорастворимые фракции. Затем из осадка экстрагировали спирторастворимые проламины, добавляя 400 мкл 50% (об./об.) водного пропан-2-ола, содержащего 1% (вес./об.) ДТТ, и встряхивали пробирку, как указано выше, с последующим инкубированием в течение 30 минут при КТ и второй стадией встряхивания и центрифугирования, как указано выше. Соответствующие супернатанты, содержащие экстрагированные проламины, объединяли и переносили в новые пробирки. Содержание белка в ДТТ/пропан-2-ол супернатантах измеряли реактивом Coomassie (BioRAD), после чего проламины в аликвотах по 200 мкл осаждали 400 мкл пропан-2-ола и хранили в течение ночи при -20°C.
Аликвоту каждого проламинового экстракта из половины семени анализировали на предмет снижения B- и C-гордеинов с помощью электрофореза в ДСН-ПААГ, как описано в Примере 1 (Фиг.9). На анализируемые гели наносили пробы, соответствующие отдельным семенам, причем на каждую дорожку наносили количество белка, эквивалентное 1/20 семени. В частности, экстракты анализировали на предмет отсутствия или снижения характеристических белковых полос гордеинов 40 кДа (специфичной B-гордеину) и 70 кДа (специфичной C-гордеину). Семена родительских линий Riso 56 и Riso 1508 имели пониженное содержание B- и C-гордеинов соответственно, но все еще содержали низкие уровни D-гордеинов, соответствующих полосе 100 кДа (Фиг.9). Большинство экстрактов семян F2 содержало профиль дикого типа, включавший наличие гордеинов D, C и B (Фиг.9), подтверждая то, что было выполнено эффективное скрещивание двух родительских линий. Шестнадцать семян, как оказалось, не содержали ни B-, ни C-гордеинов, и поэтому их оценивали как гомозиготные по обоим генетическим нарушениям, присутствующим в родительских линиях. Их идентифицировали из 288 полусемян (частота 0,055). Это соответствовало частоте 1 к 16 (0,0625), ожидаемой для комбинации двух простых, рецессивных мутаций.
Содержание суммарного белка в спирторастворимых экстрактах половин семян F2 сравнивали с аналогичными показателями семян дикого типа и родительских семян. Данные приведены в Таблице 1. Уровни белка в экстрактах семян F2 были снижены менее чем до 20%, в некоторых случаях менее чем до 15% от уровня дикого типа. Указанные значения, возможно, были завышены небелковыми азотсодержащими соединениями, такими как свободные аминокислоты, которые присутствуют в экстрактах.
(мкг/семя ± SE)
Наблюдаемые различия в уровнях проламинов между линиями F2, возможно, обусловлены расщеплением других генов или мутаций из родителей.
Дополнительные белковые гели прогоняли, взяв объем супернатанта в ДТТ/пропан-2-оле, содержащий 20 мкг белка, высушивали каждый в вакууме в SpeediVac, растворяли белок в 20 мкл буфера, содержащего 62,5 мМ Трис-HCl (pH 6,8), 12,5% (вес./об.) глицерина, 2% (вес./об.) ДСН, 1% (вес./об.) ДТТ и 0,112% (вес./об.) бромфенолового синего, а затем нагревали в бане с кипящей водой в течение 90 сек. Каждый раствор наносили на заранее залитый ДСН-полиакриламидный гель, проводили электрофорез, после чего окрашивали и анализировали гель, как описано выше. Стандартный гель показан на Фиг.10. Большинство отобранных семян F2, как оказалось, не содержало ни B-, ни C-гордеинов и, как предполагали, являлось "двойными нулями". Даже при том, что на каждую дорожку наносили одно и то же количество белка, измеренное в анализе связывания белка с красителем, большинство экстрактов из двойных нулей, как оказалось, содержали значительно меньше белка, чем контроль, в особенности они содержали малое количество белкового материала с массой более чем 20 кДа. Это можно объяснить присутствием небелковых азотсодержащих соединений, таких как свободные аминокислоты, содержащиеся в экстрактах, которые, возможно, вызывали завышение истинных уровней белка при оценке с помощью анализа связывания белка с красителем. Данный эффект также наблюдали для экстрактов Riso 1508, в которых общее количество окрашиваемого материала, идущего в виде полос белка, было уменьшено по сравнению с Riso 56 или Bomi. Riso 1508, как было показано, накапливал большее количество небелкового N в форме свободных аминокислот (Koie and Kreis, 1978).
Также исследовали срез семян F2. По сравнению с диким типом, в некоторых случаях эндосперм предполагаемых двойных нулевых семян оказывался умеренно морщинистым, в других - более морщинистым.
Вторую половину каждого семени F2 проращивали на сырой фильтровальной бумаге, проростки F2 переносили в почву в теплице и выращивали до зрелого состояния, с получением семян F3. Измеряли различные параметры роста и урожая растений (Таблица 2).
колос
В Red
Измеряли высоту растения, вес колоса и стебля, число побегов, число семян в колосе и средний вес 100 семян. Индекс урожайности вычисляли из отношения веса колоса/(вес стебля + вес колоса). Затем семена F3 выращивали в поле с получением семян F4 каждой линии.
Семена F3 показали значительное изменение по всем измеренным параметрам, по сравнению с родительскими линиями и контрольной линией, Sloop. Многие из предполагаемых двойных нулевых линий, таких как J4 и 6RF, имели сниженный средний вес 100 семян, приблизительно до 40%, или уменьшенное количество семян в колосе по сравнению с сибсом дикого типа К8. Это позволяет предположить, что другие гены, расщепляющиеся в популяции, как и мутации гордеинов B или C, оказывали влияние на урожай. Однако несколько линий F3 имели вес семян выше или равный весу семян в родительских линиях, и по-видимому другие гены могли расщепляться от мутаций B-гордеина и lys3a.
Внешний вид семян F3 на срезе варьировал от морщинистого (подобно Riso 1508) до слегка морщинистого (подобно Riso 56), по сравнению с сибсами дикого типа или контрольным Sloop.
Суммарные водорастворимые и спирторастворимые белки из восьми семян F3 из нескольких линий экстрагировали, как описано выше. Содержание белка в спирторастворимых и водорастворимых фракциях измеряли, как описано в Примере 1, используя известные количества гамма-глобулина в качестве белкового стандарта. Однако уровни суммарного спирторастворимого белка в некоторых образцах семян F3 по существу являлись такими же, как у Riso 1508. Впоследствии определили, что данные образцы семян расщеплялись по аллелям дикого типа гена Lys3a и не являлись однородно "двойными нулями".
Количественное определение уровней гордеина в семенах F3 посредством ОФ FPLC
Спирторастворимые экстракты из двух семян каждой линии объединяли и 50 мкл анализировали с помощью ОФ FPLC, как описано выше. Хроматограммы показаны на Фиг.11. Полную площадь под хроматограммами, соответствующую гордеинам, вычисляли и выражали относительно уровнем в линии дикого типа. Данные (Таблица 3) показали, что в зерне F3 уровни гордеина были менее 30% от уровня дикого типа, в некоторых случаях менее 20%, даже равные 5,3%. Отсутствие существенных полос белка на ДСН-ПААГ поддерживает утверждение, что уровни суммарного спиртового белка были завышены вследствие повышенных уровней небелкового азота в семенах F3.
Пример 4. Свойства выращенного в поле зерна ячменя F4
Сравнивали свойства выращенных в теплице и выращенных в поле семян F4 отобранных линий (9RE, J1, G1, 4BH), однонулевых родительских линий (Riso 56 и Riso 1508) и ячменя дикого типа (Sloop, Bomi и K8, воссозданного сибса дикого типа из того же скрещивания, что и двойные нулевые линии).
Вес семян
Средний вес 100 семян F4, выращенных в теплице, варьировал в пределах 60-76% от веса Sloop (5,47 + 0,16 г на 100 семян), тогда как вес 100 семян F4, выращенных в поле, был ниже, и варьировал в пределах 58-65% от веса Sloop (4,75 + 0,04 г).
Проращивание зерна
Проращивание семян двух отобранных линий ячменя F4 сравнивали с сортом дикого типа Sloop, выдерживая каждый образец из 100 семян на влажной бумаге в течение шести дней. Прорастание наблюдали, как появление кончика корня из оболочки семени. Зерна F4, как оказалось, прорастали с такой же скоростью, как и зерно дикого типа, с приблизительно 60-70%-ным прорастанием через 3 дня. Хранение зерна при 37°C в течение 4 недель перед набуханием несколько повышало % прорастания обоих линий F4. Обработка при 4°C в течение 3 дней также обеспечивала аналогичное повышение в случае свежесобранного материала.
Это демонстрирует, что зерно линий F4 не показывало какого-либо серьезного замедления прорастания, и поэтому было сочтено агронотехнически пригодным.
Уровни белка в зерне F4
Уровни водо-, соле-, спирторастворимых, а также растворимых в мочевине белков в зерне линий F4 измеряли, используя дублированные образцы по 20 мг муки из цельного зерна выращенных в теплице семян F4 отобранных линий (9RE, J1, G1, 4BH), однонулевых родительских линий (Riso 56 и Riso 1508), и ячменя дикого типа (Sloop, Bomi и K8).
Водорастворимые белки экстрагировали из каждого образца муки с использованием 0,5 мл воды, перемешивали в течение 30 минут, центрифугировали смесь при 13000 об/мин в течение 5 минут, удаляли супернатант, после чего экстрагирование дважды повторяли на осадке. Супернатанты объединяли (водорастворимый экстракт), а осадок последовательно экстрагировали три раза аналогичным путем, используя 0,5 мл 0,5 M NaCl (солерастворимый экстракт), а затем 0,5 мл 50% (об./об.) пропан-1-ола, содержащего 1% (вес./об.) ДТТ (спирторастворимый экстракт (гордеины)), с последующей экстракцией 8 M мочевиной, содержащей 1% (вес./об.) ДТТ (растворимый в мочевине экстракт). Содержание белка в каждой фракции измеряли с использованием анализа связывания красителя (BioRad) согласно инструкциям изготовителя, с калибровкой по гамма-глобулину в качестве белкового стандарта. Данные показаны на Фиг.12. Содержание суммарного экстрагируемого белка (Фиг.12E) вычисляли как суммарное количество белка, содержащееся во всех растворимых фракциях.
Кроме того, общее количество азота (Общий N; Фиг.12F) измеряли, используя дублированные образцы по 2,5 мг той же муки, с помощью элементного анализа, после сгорания при 1800°C и восстановления до N2 при 600°C, и количественного определения посредством масс-спектрометрии (метод Дюма). Полное содержание белка вычисляли, используя следующую формулу: содержание белка = 6,63 × количество общего N. Данные, полученные для уровней суммарного белка с помощью МС, являлись обоснованно аналогичными предполагаемому содержанию суммарного экстрагируемого белка, демонстрируя, что экстрагирование белка являлось эффективным.
Содержание гордеинов (измеренное как уровень спирторастворимого белка) в зерне F4 было снижено до 17-39% от уровня родительских линий (R1508 и R56) и до 7-16% от уровня сорта Sloop дикого типа. Это соответствует приблизительно 10-кратному снижению общего количества гордеинов, которые, как показано выше, являются токсичными для больных целиакией, в данных образцах зерна, по сравнению с ячменем дикого типа, Sloop.
Другие типы белков, в особенности водо- и солерастворимые белки, как считают, оказывают благоприятные воздействия на пивоваренные свойства зерна ячменя. Поскольку уровни водо- и солерастворимых белков зерна F4 были аналогичны соответствующим показателям дикого типа Sloop, предположили, что зерна F4 содержат достаточное в пивоваренных целях количество указанных белков.
Содержание и состав жирных кислот
Так как главный запас азота, необходимого в процессе роста и развития семян, был удален вследствие снижения уровня гордеинов, мутантное зерно анализировали с целью определения, скомпенсировано ли развитие семени повышением накопления других компонентов, некоторые из которых могут быть нежелательны при применении зерна. Жирные кислоты в дублированном образце 50 мг муки из цельного зерна F4 экстрагировали, метилировали и анализировали с помощью количественной газовой хроматографии (ГХ), используя метод Folich et al. (1957).
Суммарная концентрация жирных кислот в зерне F4 линий G1, BB5, J1 и J4 варьировала в пределах приблизительно от 2,5% до 3% (по весу), и была аналогична уровню в зерне ячменя однонулевой линии и дикого типа. Установили, что двойные нулевые зерна не содержали повышенных уровней жирных кислот.
Жирные кислоты в липидах зерна включали главным образом линолевую (C18:2), олеиновую (C18:1) и пальмитиновую кислоты (C16:0), а также другие жирные кислоты с более низкими уровнями. В концентрациях отдельных жирных кислот, которые накапливались в отобранном зерне F4, по сравнению с однонулевыми родительскими линиями или ячменем дикого типа, не наблюдали каких-либо существенных различий. В частности концентрация эруковой кислоты (C22:1n-9), токсичной для людей в высокой концентрации, в зерне F4 не была повышена. Мутантное зерно, таким образом, имело нормальное содержание и состав жирных кислот.
Уровни крахмала
Крахмал является главным компонентом зерна злаков и обычно составляет приблизительно 55-65% сухого веса. Уровни крахмала особенно важны в ячмене, который используется для соложения. Содержание крахмала, которое является слишком низким, может приводить к получению солода с содержанием сахаров, недостаточным для поддержания эффективного брожения в пивоваренном процессе, и поэтому измеряли содержание крахмала в зерне ячменя.
Крахмал мутантного зерна выделяли и анализировали по существу как описано в Megazyme Method (AACC76.13), используя образцы по 20 мг муки из цельного зерна. Общее содержание крахмала в зерне F4 находилось в пределах от 57% до 66% (по весу), и было аналогично содержанию крахмала в однонулевых родительских линиях и ячмене дикого типа, которые находилось в пределах 51-64% (по весу).
Установили, что зерна ячменя F4 имели достаточное содержание крахмала, чтобы обеспечивать получение солода из зерна.
Уровни β-глюканов
Содержание β-глюканов в мутантном зерне оценивали, как описано в Megazyme Method (AACC32.23), используя образцы по 20 мг муки из цельного зерна. Уровни β-глюканов в зерне линий G1, BB5, J1, J4 находились в пределах от 1,2 до 2,6% (по весу), и были аналогичны содержанию β-глюканов в однонулевом родительском зерне и зерне ячменя дикого типа, которое находилось в пределах от 2,4-3,3% (по весу).
Высокие уровни β-глюканов способствуют формированию помутнения в пиве в процессе хранения. Установили, что содержание β-глюканов в зерне F4 не было повышено по сравнению с зернами дикого типа, при этом маловероятно, что подобные уровни будут неблагоприятно влиять на пивоваренные характеристики данного зерна.
Уровни свободных аминокислот
Повышенное накопление свободных аминокислот, возможно, может являться неблагоприятным при применении зерна. Например, свободный аспарагин в достаточных количествах может формировать токсичное соединение акриламид при нагревании до высоких температур в присутствии крахмала.
Содержание и состав свободных аминокислот в зерне оценивали, используя дублированные образцы по 20 мг муки из цельного зерна, выращенных в теплице семян F4. Образцы растворяли в 0,1 Н HCl, отобрали аликвоту и высушивали, после чего анализировали аминокислоты с использованием набора Waters AccQTag chemistry, на базе Австралийского Центра Протеомного Анализа (Australian Proteome Analysis Facility, Сидней).
Наиболее преобладающими аминокислотами в ячменной муке являлись пролин, аспарагин, глутаминовая кислота и аспарагиновая кислота в порядке убывания, в пределах от приблизительно 1,5 мг/г муки до 0,5 мг/г муки. Содержание свободного пролина в отобранном зерне F4 находилось в пределах 0,6-1,5 мг/г, и было аналогично содержанию свободного пролина в однонулевых родительских линиях и ячмене дикого типа, которое находилось в диапазоне 0,2-1,2 мг/г. Уровни остальных свободных аминокислот являлись соответственно аналогичными уровням в контрольных зернах и F4. В частности содержание свободного аспарагина в зерне F4 составляло приблизительно 0,5 мг/г для линий G1, BB5 и J1, и приблизительно 1,0 мг/г в линии J4. В однонулевых родительских зернах уровень свободного аспарагина составлял 0,3 или 0,9 мг/г, а в зернах ячменя дикого типа свободный аспарагин присутствовал в диапазоне от 0,3-0,6 мг/г. Поскольку содержание свободного аспарагина в зерне F4 было аналогично уровням в соответствующем зерне дикого типа, предположили, что образование акриламида из свободного аспарагина в процессе соложения или другого применения зерна не будет отличаться по сравнению с зерном дикого типа.
Свободный лизин, как известно, является критической аминокислотой в корме животных, и поэтому уровни указанной аминокислоты представляют интерес для потенциального применения зерна в качестве корма. Содержание свободного лизина в зерне F4 линий G1, BB5 и J1 составляло приблизительно 0,5 мг/г, и 1,0 мг/г в зерне линии J4. Это соответствовало 181% - 1,020%-ому увеличению по сравнению с уровнем в зерне дикого типа сорта Sloop. Таким образом, линии F4 являлись более ценным пищевым источником свободного лизина по сравнению со Sloop.
Пример 5. Анализ зерна F4 - тест токсичности T-клеток
Для анализа токсичности зерна F4 при целиакии, гордеины выделяли и очищали из 10 г образцов муки из цельного зерна, полученной из выращенных в поле семян отобранных линий 9JAE, J1, G1 и 4BH, однонулевых родительских линий (Riso 56 и Riso 1508), и ячменя дикого типа (Sloop, Bomi и K8), как описано ниже. Очищенные гордеины добавляли к T-клеткам, выделенным в популяции больных целиакией, с целью проверки на токсичность при целиакии. Тест включал измерение количества T-клеток, которые вырабатывали гамма-интерферон после ночного инкубирования с очищенным белком, используя иммуноанализ уровня гамма-интерферона. Таким образом, уровень гамма-интерферона служил показателем степени токсичности белков в зерне. Затем данное измерение токсичности муки при целиакии наносили на график как функцию от сырого веса муки, полученной из зерна.
Очистка проламинов (гордеинов)
Муку из цельного зерна (10 г) перемешивали в течение 30 минут при 25°C в 200 мл буфера, содержащего 20 мМ триэтаноламина-HCl (TEA), 1% (вес./об.) аскорбата натрия, 1% (вес./об.) полиэтиленгликоля (мол.вес 6000; PEG 6000), и 1 мкг/мл ингибиторов протеазы E64 и AEBSF (Sigma); буфер доводили до pH 8. Суспензию центрифугировали при 5000 g в течение 5 минут, супернатант отбрасывали, а осадок промывали не менее двух раз. Белки в промытом осадке растворяли в 80 мл 50% (об./об.) пропан-2-ола, содержащего 1% (вес./об.) ДТТ, перемешивали в течение 30 минут при 60°C. Суспензию охлаждали при 4°C в течение 10 минут и центрифугировали при 10000 g в течение 10 минут при 4°C. Белки, включающие гордеины, в супернатанте осаждали 2 объемами пропан-2-ола в течение ночи при -20°C, после чего центрифугировали при 10000 g в течение 10 минут при 4°C, а осадок растворяли в 10 мл буфера, который содержал 8 M свежеприготовленного деионизированного раствора мочевины, 1% ДТТ, 20 мМ TEA, pH 6.
Гордеины очищали с помощью FPLC следующим образом. Раствор гордеинов (1 мл) вводили в 8 мл колонку для обращенно-фазовой хроматографии Source 15 (ОФХ, Pharmacia). Колонку промывали 4 мл 5%-ного растворителя B, а гордеины элюировали 2,5 мл в линейном градиенте с 5% растворителя B до 35% растворителю B со скоростью 4 мл/мин, а затем в линейном градиенте с 35% растворителя B до 83% растворителя B, в объеме более 36 мл. Растворитель A представлял собой 0,1% (об./об.) трифторуксусную кислоту (ТФУ) в воде, растворитель B представлял собой 0,1% (об./об.) ТФУ в 60% (об./об.) водном ацетонитриле. Фракции, элюируемые между 25 и 43 мл, объединяли. Контрольный растворитель аналогичным путем объединяли из прогонов без ввода пробы. Соответствующие пулы 10 последовательных инъекций объединяли и лиофилизировали.
Ex vivo анализ на T-клетках
Гордеины, очищенные с помощью FPLC (50 мг/мл в 2 M мочевине), растворяли в PBS, содержащем 1 мМ CaCl2, получив растворы с концентрацией 25, 62,5, 125, 250, 625, 3750 или 6250 мкг гордеина/мл, после чего гордеины дезамидировали, добавляя по 25 мкл каждого раствора к 100 мкл tTG из печени морской свинки (Sigma; 25 мкг/мл tTG в PBS, содержащем 1 мМ CaCl2), с последующим инкубированием в течение 6 часов при 37°C. Недезамидированные растворы получали аналогично, путем инкубирования в отсутствие tTG. Контрольные растворители добавляли, как при максимальных концентрациях гордеинов. Другие контрольные образцы содержали либо контрольный растворитель, контрольный растворитель и известный токсин, анатоксин столбняка (50 светообразующих частиц/мл, полученный из Commonwealth Serum Laboratories, Melbourne), либо анатоксин столбняка (50 светообразующих частиц/мл) в чистом виде. Затем все растворы замораживали при -20°C.
T-клетки получали следующим образом. Шесть пациентов с HLA-DQ2+ целиакией, подтвержденной посредством биопсии, которые придерживались строгой аглютеновой диеты в течение, по меньшей мере, трех месяцев, употребляли 150 г вареного ячменя ежедневно в течение 3 дней. PBMC выделяли с помощью центрифугирования в градиенте плотности Фиколл-Гипак из гепаринизированной венозной крови, забранной либо непосредственно до, либо через шесть дней после начала диетической провокации, и ресуспендировали в полной среде HT-RPMI, содержащей 10% термоинактивированную, смешанную человеческую сыворотку AB. Дезамидированные или недезамидированные гордеины и контрольные растворы размораживали, и добавляли по 25 мкл в лунки, содержащие 100 мкл PBMC (3-8×105 PBMC в лунке), культивировали при 37°C в течение ночи в 96-луночных планшетах (MAIP-S-45; Millipore, Bedford, MA) и сравнивали с контрольными культурами (без добавки), к которым добавляли 25 мкл PBS, содержащего только 1 мМ CaCl2. Конечные концентрации гордеинов составляли 0, 1, 2,5, 5, 10, 25, 150 или 250 мкг/мл. Максимальная конечная концентрация мочевины составляла 10 мМ. IFN-γ визуализировали, используя вторичные антитела согласно инструкциям изготовителей (Mabtech, Stockholm, Sweden), как ранее описано Anderson et al. (2005), и измеряли пятнообразующие единицы (SFU), используя автоматизированный ELISPOT планшет-ридер (AID Autoimmun Diagnostika GmbH; Germany). Результаты представлены как среднее значение пятнообразующих единиц (SFU) ± SE в зависимости от эквивалентной массы муки, которая содержит расчетное количество гордеина. Содержание гордеинов в каждом образце муки вычисляли в Примере 5, принимая в расчет вычисление веса муки.
Данные анализировали с использованием GraphPAD Prism, рассчитывали кривые наибольшего соответствия и показывали со средними значениями + SE. Значения r2 для данных превышали 0,83, что указывает на хорошее соответствие между экспериментальными данными и кривой наибольшего соответствия (Фиг.13).
Результаты
T-клетки, выделенные у одного пациента с целиакией перед диетической провокацией, были менее чувствительны к проламинам, добавляемым в концентрации 25 мкг/мл, по сравнению с T-клетками, выделенными у того же человека после диетической провокации ячменем. Среднее SFU±SE 29,5+3,0 и 104+15,9 регистрировали для T-клеток, выделенных до и после диетической провокации. Это указывает, что специфичные при целиакии T-клетки были индуцированы диетической провокацией.
Используемые T-клетки, выделенные через шесть дней после указанной диетической провокации, положительный контроль, анатоксин столбняка, давали сопоставимый ответ в отсутствие и в присутствии tTG (среднее SFU±SE 28,1±5,9 и 20,2±7,4 соответственно). Добавление контрольного растворителя существенно не ингибировало ответ положительного контроля, анатоксина столбняка (среднее SFU±SE 20,5+4,1 и 17,6±6,0, в отсутствие и в присутствии tTG соответственно), подтверждая, что примеси в растворителе не давали ложноотрицательные и не ингибировали положительные ответы.
T-клетки, выделенные у больных целиакией через 6 дней после провокации, более активно взаимодействовали со всеми гордеиновыми фракциями, обработанными tTG, по сравнению с T-клетками, взаимодействовавшими с недезамидированными гордеинами, как и ожидается при целиакии (Hadjivassiliou et al., 2004, Kim et al., 2004) (Фиг.13 A; для ясности показан ответ только к двум образцам гордеинов, Sloop и G1). Это подтверждало то, что измеряемый ответ T-клеток был связан с токсичностью при целиакии.
Поскольку концентрация гордеина была увеличена, число SFU также возросло в гиперболической манере, ожидаемой для стандартной ферментативной кинетики Михаэлиса-Ментен между ферментом и его субстратом. Для описания подобных кривых обычно используют два параметра: Bmax, максимальное число SFU, ожидаемое при наиболее высокой концентрации; и Kd, концентрацию белка, требуемую для индуцирования половины от максимального числа SFU. Чем более токсичен образец муки, тем ниже Kd.
Коэффициенты Kd и Bmax вычисляли по кривым наибольшего соответствия. Значения Bmax значительно не отличались между диким типом и мутантами, как и ожидали. В противоположность, значения Kd для линий F4 были выше в 10 раз по сравнению с линиями дикого типа (Таблица 4). Таким образом, приблизительно в 10 раз большее количество муки, полученной из мутантных линий, должно вызывать половинный максимальный ответ токсичности, по сравнению с мукой дикого типа (Таблица 4). Таким образом, установили, что токсичность зерна F4 при целиакии была снижена примерно в 10 раз по сравнению с линиями дикого типа. Данный уровень снижения хорошо согласуется с уменьшенным уровнем гордеина, обнаруженным при определении белка в зерне F4.
Токсичность зерна F4 была ниже, чем токсичность Riso 56, как и ожидали. Однако токсичность зерна F4 была подобна токсичности другой родительской линии - Riso 1508. Впоследствии, при дальнейшем генетическом исследовании зерна F4 обнаружили, что это было обусловлено гетерозиготностью мутации гена, кодирующего белок B-гордеин в отобранных линиях F4, что приводило к повышению содержания гордеинов выше ожидаемого.
Пример 6. Соложение зерна F4
Чтобы определить пригодность зерна ячменя для соложения, проводили исследования, включающие тесты с мелкомасштабным соложением (микросоложением).
Одним из факторов, который влияет на способность к соложению, является размер семени. Образцы зерна F4 анализировали на распределение размера семян с помощью подсчета процентной доли семян из 1000 семян, которые задерживались ситом с ячейками 2,8, 2,5 или 2,2 мм. Зерно F4 в среднем имело меньшую крупность по сравнению с диким типом, и аналогичную родительским зернам Riso 1508 и Riso 56, при этом менее 5% семян задерживались ситом с ячейками 2,5 мм (Таблица 5). Это контрастировало с контрольными линиями Galleon и Sloop, в которых крупность 90% семян превышала 2,5 мм. Было отмечено, что зерна линии K8, которая является линией дикого типа, полученной из того же кросса Riso 1508XRiso56, также имели уменьшенный размер, поэтому, по меньшей мере, частично, уменьшение размера семян было связано с генетическим фоном и, опосредованно, снижение уровня гордеинов. Кроме того, меньший размер семян можно скомпенсировать модификациями в способе вымачивания зерна.
Уровни влагосодержания семян могут повлиять на эффективность соложения. Перед микросоложением измеряли % влаги и % азота с помощью анализа в ближней ИК-области (NIR). Уровень влажности семян всех образцов зерна F4 находился в диапазоне 11 -11,4% и был аналогичен контрольным линиям, за исключением зерна сорта Galleon (GA1, 8,9%). Содержание азота в семени двойных нулевых линий варьировало от 2,3% до 2,5%, что было выше по сравнению с контрольной линией соложения сорта Galleon (1,6%). Для соложения уровни азота в семени оптимально составляют 1,5 - 2,0%.
Образцы ячменя (170 г) выращенного в поле зерна F4 отобранных линий 5RB, G1, J1, 9RE, 4BH, однонулевых родительских линий (Riso 56 и Riso 1508) и ячменя дикого типа K8, сортов Bomi, Carlsberg II, Sloop и Galleon вымачивали при 16°C в течение 6 ч, с последующим отстоем в течение 7 ч на воздухе, после чего вымачивали в течение 6 ч, а затем проращивали при 15°C в течение 4 дней в системе микросоложения JWM. Проросшее зерно высушивали в течение 21 ч при минимальной температуре 50°C и максимальной температуре 80°C, после чего полученный солод очищали от корней путем перетирания и просеивания.
Солод анализировали по показателям влажности (%), общего содержания азота (% сухого веса) с помощью NIR цельного зерна, а также выхода (выраженного как вес очищенного солода в процентах от исходного веса ячменя).
Кроме того, образцы солода дробили в молотковой дробилке и образцы весом 50 г растворяли в воде, нагретой до 45°C - 70°C, получив раствор с конечным весом 450 г, который анализировали на экстрактивность (солюбилизируемый % от веса зерна), цвет, растворимый азот (N), число Кольбаха (KI: % растворимого белка/общее содержание белка), β-глюкан, вязкость, AAL (предел видимого сбраживания или ферментируемость, % падения плотности в процессе сбраживания пивными дрожжами), все согласно стандартным технологиям Европейской Пивоваренной Конвенции (European Brewery Convention, http://www.ebc-nl.com/) (Таблица 6).
Содержание белка в солоде в целом превышало требуемую норму. Это было связано с общим N солода и растворимым N, однако процентное содержание растворимого белка по сравнению с общим количеством (KI) было близко к норме. Цвет и вязкость сусла F4 были близки к норме, а уровни β-глюканов в сусле были низкими. Данные показатели приемлемы для соложения.
Процесс соложения включал три стадии: соложение, приготовление сусла и ферментацию. Полную эффективность вычисляли из трех значений эффективности каждой стадии: выход, экстрактивность и AAL соответственно. Они указывают, что в каждой стадии зерно F4 являлось приблизительно на 10% менее эффективным, чем эталонное зерно сорта Galleon. В итоге приблизительно в 1,3 раза больше зерна линий F4 потребуется для производства пива с крепостью, эквивалентной коммерческому стандарту, по сравнению с Galleon.
Все указанные признаки показали, что из зерна F4 может быть приготовлен солод.
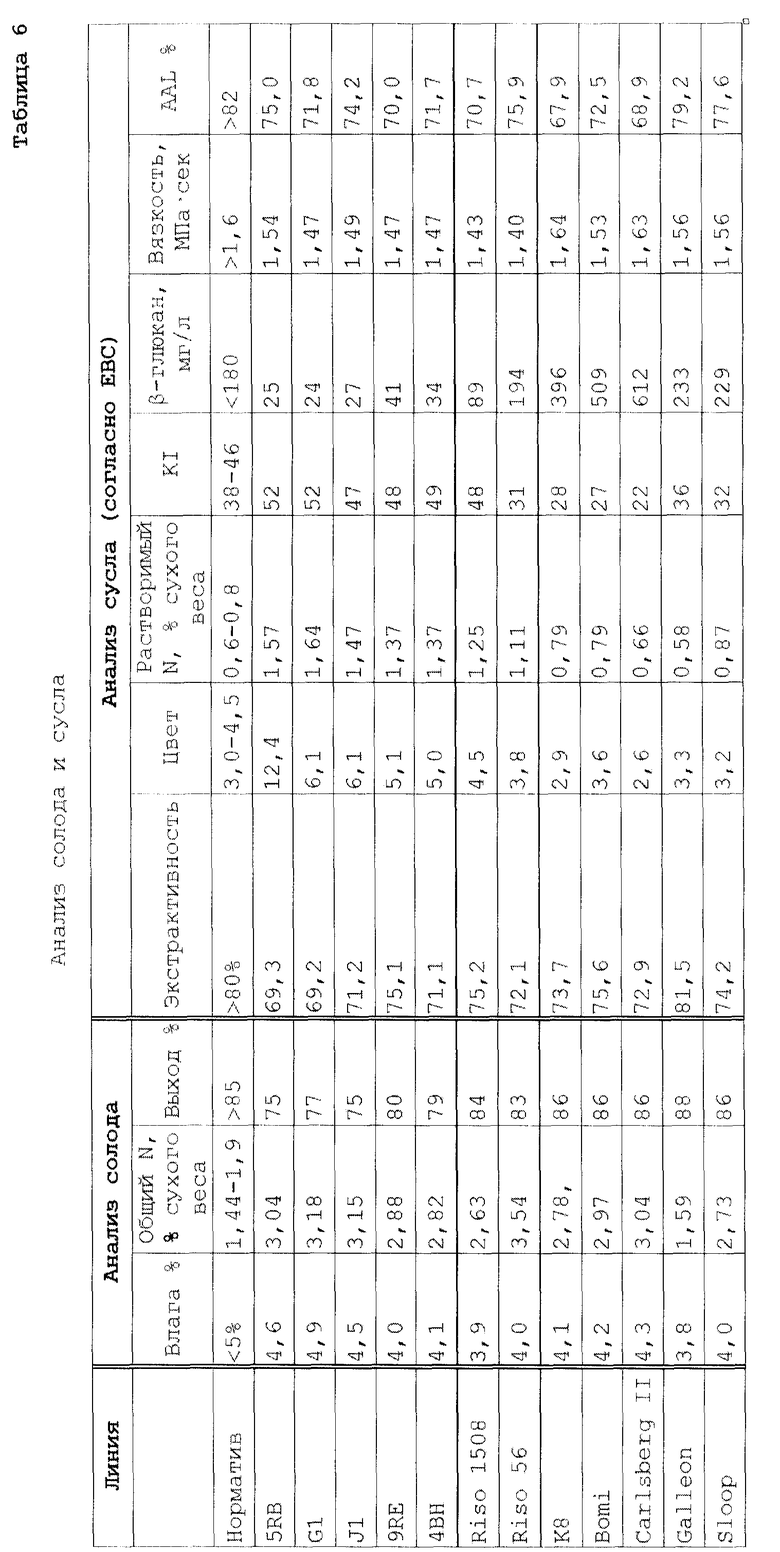
Пример 7. Анализ ELISA неочищенных образцов солода
Образцы объемом приблизительно 40 мл сусла из Примера 6 замораживали, лиофилизировали и растворяли в 20 мл 6M мочевины, 1% (вес./об.) ДТТ, 20 мМ TEA (pH 6) при комнатной температуре. Содержание белка в каждом образце определяли, используя метод Брэдфорд. Последовательные разведения, содержащие 20 мкг белка солода в 100 мкл 6М мочевины, 1% ДТТ и 20 мМ TEA (pH 6) наносили на нитроцеллюлозную мембрану (Amersham Hybond C+), которую предварительно уравновешивали буфером PBS, в приборе для дот-блоттинга (BioRad), и калибровали очищенным стандартом (C-гордеин, 2 мкг). Раствор пропускали через мембрану при пониженном давлении, затем мембрану промывали буфером PBS, содержащим 0,1% Tween 20 (PBST), прибор разбирали, а мембрану блокировали путем инкубирования в 5% (вес./об.) сухом обезжиренном молоке в буфере PBS, содержащем 0,1% Tween 20, в течение 1 часа при комнатной температуре. Гордеины детектировали первичным антителом (антителом кролика против глиадинов пшеницы, конъюгированным с пероксидазой хрена, Sigma), разбавив 1 к 2000 в буфере PBST, в течение 30 минут при комнатной температуре. Мембрану промывали в трех сменах буфера PBST и окрашивали, инкубируя в 10 мл 1:1 (об./об.) смеси Amersham ECL реагентов A и B для Вестерн-блоттинга (GE HealthCare), после чего детектировали сигнал, выдерживая с Amersham Hyperfilm в течение 30 сек. После окрашивания пленки проводили количественный анализ, используя программное обеспечение Total Lab TL100 (Non-linear dynamics, 2006).
Неочищенные солодовые растворы, полученные из отобранного зерна F4, имели среднее содержание гордеина 58+12,7 м.д.
Указанный уровень был существенно ниже предельного уровня 200 м.д., установленного FSANZ для низкоглютеновых продуктов питания в Австралии, и значительно ниже, чем среднее содержание 687±158 м.д. для солода из сортов дикого типа, Galleon, Sloop, K8, Bomi и Carlsberg II. Он также был значительно ниже, чем содержание гордеинов в солоде, полученном из родительских линий Riso 56 и Riso 1508.
Обычно содержание глютена (гордеинов) в смесях резко падает в процессах соложения, приготовления сусла и брожения, в результате чего готовое стабилизированное пиво может содержать 1/1000 от уровня в сыром солоде (Dostalek et al., 2006).
Поэтому предположили, что уровень гордеинов в готовом пиве, приготовленном из солода F4, снизится приблизительно до 0,05 м.д., что значительно ниже диапазона 3-40 м.д., присутствующего в пиве, приготовленном из зерна ячменя дикого типа (Dostalek et al., 2006).
В литературе существует несколько новых рекомендаций относительно предельного содержания глютена в диете больных целиакией. Наиболее надежные из них основаны на многоцентровом, с контролем плацебо, двойном слепом испытании, и показывают, что употребление менее чем 10 мг/день безопасно для больных целиакией, а также рекомендуют, чтобы потребление сохранялось на уровне менее 50 мг/день (Catassi et al., 2007). Другое недавнее исследование подтверждает вышеуказанные данные и (Collin et al., 2004) рекомендует, что употребление пищи с содержанием глютена 100 м.д. приведет к потреблению приблизительно 30 мг/день и вызовет небольшое повреждение у больных целиакией. FSANZ устанавливает пищевые стандарты для Новой Зеландии и Австралии. Комиссия "Кодекс алиментариус" (от лат. "Продовольственный кодекс") была создана в 1963 ФАО и ВОЗ с целью разработки пищевых стандартов, руководств и подобных документов, таких как свод правил под контролем Программы стандартов на пищевые продукты ФАО/ВОЗ, и является общепринятым нормативным актом для Европы и Северной Америки. Кодекс алиментариус в настоящее время устанавливает аглютеновый предел менее 0,05 г N (в виде глютена) на 100 граммов пищевого продукта. Имеется предложение к пересмотру стандарта Кодекса, предлагающее предел 20 м.д. для пищевого продукта, изготовленного из злаков, не содержащих глютен, и 200 м.д. для пищевого продукта, изготовленного из злаков, содержащих глютен (p32, PROPOSAL P264, REVIEW OF GLUTEN CLAIMS WITH SPECIFIC REFERENCE TO OATS AND MALT, FSANZ web site: http;//www,foodstandards.gov.au/_srcfiles/P264 Gluten Claims FAR-pάfffsearcbr^/o 22gluten%20free%22s).
Из вышеописанного анализа установили, что употребление пива, произведенного из линий ячменя F4, будет значительно ниже предела безопасности, установленного для аглютеновых пищевых продуктов, предназначенных для больных целиакией, в вышеупомянутых исследованиях, включая инструкции, установленные FSANZ и Кодекс алиментариус.
Пример 8. Дополнительное исследование линий F4
Спирторастворимые белки очищали из массы семян F4, собранных для каждой из обозначенных линий, как описано выше. Очищенные образцы белка (20 мкг) из зерна F4 линий G1, J1, 4BH, 5RB и 9RE растворяли в 6M мочевине с 2% (вес./об.) ДСН, 1% (вес./об.) ДТТ, 0,01% (вес./об.) бромфеноловым синим, 0,0625 М Трис-HCl (pH 6,8) при 25°C, анализировали с помощью электрофореза в ДСН-ПААГ, окрашивали 0,006% коллоидным Commassie Blue, сравнивали с гордеинами, выделенными из Riso 56, Riso 1508, а также линии дикого типа (K8). Движение белков соотносили со стандартами молекулярного веса для определения молекулярной массы (Таблица 7).
Последовательности белков получали с помощью масс-спектрометрии трипсиновых переваров, полученных из пятен белка, вырезанных из гелей, и обрабатывали с целью секвенирования белка посредством фрагментации при тандемной масс-спектрометрии, как описано ранее (Campbell et al., 2001), с поиском в нерезервированной базе данных NCBI.
(% белка)
балл
MS-MS
A: Все перевары, как ожидалось, также содержат пептиды из свиного трипсина.
B: Также содержится низкий уровень D-гордеина
C: Также содержится низкий уровень B-гордеина.
D: Также содержится низкий уровень предшественника гамма-гордеина-1.
E: Суммарный балл поиска MS-MS указывает достоверность отнесения идентичности. Из прошлых опытов, для надежной идентификации необходим балл выше 15, а балл выше 50 указывает на практически безошибочную идентификацию.
Пептиды из каждого образца связывали на колонке Agilent Zorbax SB-C18 5 мкм, 150x0,5 мм с расходом 0,1% (об./об.) муравьиной кислоты/5% (об./об.) ацетонитрила 20 мкл/мин в течение одной минуты, а затем элюировали в градиентах с повышением концентрации ацетонитрила 0,1% (об./об.) муравьиной кислоты/20% (об./об.) ацетонитрила за одну мин при 5 мкл/мин, затем 0,1% (об./об.) муравьиной кислоты/50% (об./об.) ацетонитрила за 28 мин, затем 0,1% (об./об.) муравьиной кислоты/95% (об./об.) ацетонитрила за одну мин. Колонку промывали в градиенте от 0,1% (об./об.) муравьиной кислоты/95% (об./об.) ацетонитрила до 0,1% (об./об.) муравьиной кислоты/100% (об./об.) ацетонитрила в течение 5 минут при 20 мкл/мин и повторно уравновешивали 0,1% (об./об.) муравьиной кислотой/5% (об./об.) ацетонитрилом в течение 7 минут перед нанесением пептидов из образца.
Элюат из колонки вводили в масс-спектрометрическую ионную ловушку Agilent XCT через микронебулайзер источника ионов электрораспыления. По мере элюции пептидов с колонки ионная ловушка регистрировала широкодиапазонные сканы положительных ионов (100-2200 m/z) с последующими четырьмя MS/MS сканами ионов, наблюдаемых в полном спектре согласно настройка прибора 'SmartFrag' и 'Peptide Scan'. Как только два спектра фрагментации получали для каких-либо определенных значений m/z, их исключали из выбора для анализа в течение последующих 30 секунд, чтобы избежать сбора избыточных данных.
Наборы масс-спектрометрических данных сравнивали с базами последовательностей, используя программное обеспечение Agilent's Spectrum Mill (Rev A.03.02.060). Ложноположительных совпадений избегали при использовании программной настройки по умолчанию 'автовалидация'. Это включает требование, чтобы совпадения пептидов являлись значительно лучше, чем наилучшее совпадение по обратным базам данных и различным оценка, способствующим более вероятным профилям ионизации и фрагментации ('оценка мобильности протонов'). Окисленный метионин допускали в качестве вариабельной модификации.
Результаты секвенирования белков показали, что семя F4 из отобранных линий неожиданно содержало полосу B3-гордеина, в дополнение к ожидаемым гамма-1-гордеину и D-гордеину. Идентичность полос гамма-1 и 3 гордеинов установили при секвенировании белков из мутанта Riso 56, в котором указанные белки при движении не маскировались наложением полосы B-гордеина. Это указывало, что отобранные линии F4 были не полностью лишены B3-гордеина.
Пример 9. Идентификация зерна ячменя, не содержащего B и C гордеины
Отдельные половины семян из одного колоса выращенных в поле растений F4 линии G1 оставили на ночь для набухания в воде, содержащей ингибиторы протеаз E64 и AEBSF (1 мкг/мл), по отдельности дробили и истирали в пластмассовой микропробирке с помощью шарика из нержавеющей стали, встряхивали с частотой 30/сек в течение 3×1,5 мин в 96-луночной вибромельнице Vibration Mill (Retsch Gmbh, Rheinische), а затем центрифугировали при 3000 g в течение 5 минут при КТ и отбрасывали супернатант. Нерастворимый в воде осадок муки промывали не менее двух раз аналогичным путем и отбрасывали супернатанты. Затем из осадка экстрагировали спирторастворимые гордеины, добавляя по 400 мкл 50% (об./об.) водного пропан-2-ола, содержащего 1% (вес./об.) ДТТ, с последующим встряхиванием и центрифугированием, как описано выше. Супернатанты, содержащие экстрагированные гордеины, переносили в новые пробирки, после чего в ДТТ/пропан-2-ол супернатантах измеряли содержание белка, используя Coomassie реагент (BioRAD).
Аликвоту каждого гордеинового экстракта, соответствующего 20 мкг гордеина, лиофилизовали в вакууме в течение ночи, растворяли в 15 мкл кипящего ДСН-буфера, нагревали в течение 3 минут при 90°C, наносили на заранее залитый гель 12-18% Excell gradient gel (Pharmacia) и анализировали с помощью электрофореза в ДСН-ПААГ, как описано в Примере 1. Заметная полоса, соответствующая приблизительно 43 кДа, как наблюдали, выделялась в отдельных семенах и отсутствовала в экстрактах 5 из 16 семян. Положение указанной полосы соответствовало полосе B3-гордеина, идентифицированной ранее.
Данные анализа белка подтверждали, что семя F4 из линии G1 являлось гетерозиготным и расщеплялось по одному или нескольким B-гордеиновым белкам. Данная ситуация также подтверждалась для других линий F4.
Генетическое исследование
Генетические анализы выполняли с целью подтверждения данных белкового анализа. Отдельные половины семян выращенных в поле, отобранных линий F4 проращивали во влажной почве, а затем выращивали в течение 2 недель в теплице при 25°C днем и 20°C ночью. ДНК выделяли из 0,5 см пластинки листа, используя набор REDExtract-N-Amp Plant PCR Kit (Sigma) согласно инструкциям. Генные последовательности, специфичные B1-гордеинам и гамма-гордеинам, амплифицировали в отдельных реакциях ПЦР, добавляя 10 мкл RMix, по 1 мкл каждого праймера для B1-гордеина (5'B1hor и 3'B1hor) или по 0,5 мкл каждого праймера для гамма-3-гордеина (5'gamma hor3 и 3'gamma 3-full), 4 мкл растительной ДНК и воду MilliQ до 20 мкл, при комнатной температуре, а затем подвергали следующей температурной программе в термоциклере Eppendorf: 95°C в течение 10 мин, затем 35 циклов: 95°C в течение 30 сек, 56°C в течение 30 сек и 72°C в течение 1 мин. После этого выполняли нагрев при 72°C в течение 10 минут и охлаждали до 10°C.
Последовательности ПЦР праймеров являлись следующими:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2)
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3)
5'gamma hor3: 5'-CGAGAAGGTACCATTACTCCAG-3' (SEQ ID NO:4)
3'gamma 3-full: 5'-AGTAACAATGAAGGTCCATCG-3' (SEQ ID NO:5).
По 20 мкл каждой ПЦР-смеси наносили на 1 см, 1% (вес./об.) агарозный гель, содержащий EtBr, проводили электрофорез при 100В в течение 1 часа в TBE буфере, и получали изображение флюоресценции ДНК-продуктов, используя систему GelDoc image system (uvitec) (Фиг.14).
Полоса амплифицированной ДНК контрольного гена гамма-3-гордеина присутствовала во всех дорожках, как ожидали (Фиг.14, нижний блок, gamma3-Hor). Амплифицированная ДНК B-гордеина отсутствовала во всех дорожках ПЦР из Riso 56, как ожидали, поскольку ген в Riso 56 был удален (Фиг.14, верхний блок, R56). Полосы амплифицированной ДНК генов B-гордеина выделялись в экстрактах из семян, взятых из одного колоса, F4 линий 9RE и 4BH (Фиг.14, верхний блок 9RE, 4BH). Это указывало, что один или несколько генов B-гордеина присутствовали в части семян F4, и что семя F3 не являлось гомозиготным по делеции локуса B-гордеина в Riso 56. Это также показали для других линий F4. Данный способ являлся удобным методом на основе ДНК, позволяющим идентифицировать и отбирать семена, несодержащие B1-гордеин.
Результаты генетического исследования использовали для отбора растений, которые не содержали B-гордеиновые гены. Двенадцать индивидуальных растений F5, нулевых согласно ПЦР по B-гордеиновым генам, отбирали и выращивали с целью получения популяции семян F5, известной как G1*. Индивидуальные G1*, F5 половины семян, взятые из одного колоса, проращивали во влажной почве, и выращивали в течение 2 недель в теплице при 25°C днем и 20°C ночью перед выделением ДНК/ПЦР-анализом, описанным выше. Соответствующую половину семени использовали для выделения и анализа гордеинов, для чего брали аликвоту, соответствующую 40 мкг гордеина, лиофилизовали в вакууме в течение ночи, растворяли в 15 мкл кипящего ДСН-буфера, нагревали в течение 3 минут при 90°C, наносили на предварительно залитый 1 мм 12% гель Longlife (Longlife Gels) и проводили электрофорез при 150В в течение 40 минут, а затем окрашивали, как в Примере 1.
ПЦР-анализ показал, что ДНК, выделенная из линий положительного контроля, Sloopa и Riso 1508, дала полосу B-гордеина, как и ожидали. Размер полосы из Sloop был больше, чем ПЦР-продукт из Riso 1508, так как гены B1-гордеина несколько отличались. Контрольный ген, гамма-3-гордеин, амплифицировали во всех растениях. Полоса ПЦР не была амплифицирована из экстрактов шести индивидуальных образцов Gl*, подтверждая отсутствие гена в данных растениях. Профиль гордеинов в соответствующих половинах семян подтверждал это - в Gl* какие-либо полосы B-гордеинов не наблюдали. Поэтому сделали вывод, что Gl* не содержал детектируемых B-гордеинов и являлся гомозиготным нулем по локусу, кодирующему B-гордеин.
Остальные 250 F5 G1* семян проращивали, анализировали всходы и подтверждали как нули по гену B-гордеина. Последующие поколения использовали для получения семян данной линии.
Анализ содержания гордеинов
Сорта ячменя Sloop, R56, R1508 и G1* выращивали на смежных участка в поле, зрелое зерно собирали и обрабатывали с получением муки. Уровни гордеинов в образцах муки анализировали, как описано выше. Фракции белков, растворимых в воде, растворе соли, спирте/ДТТ и растворе мочевины, получали, как описано в Примере 4, после чего в каждой фракции измеряли содержание белка. Содержание белка приведено в Таблице 8 и выражено как мг белка/грамм сухого веса муки. Каждое содержание суммарного белка определяли, суммируя содержание белка во фракциях соответствующего образца. Гордеины содержались в спирторастворимой фракции наряду с другими спирторастворимыми белками, такими как серпины, ингибиторы протеазы, LTP1 и Z белок.
Данные показали, что содержание спирторастворимого белка в зерне G1* и, следовательно, муке было снижено менее чем до 22% по сравнению с сортом дикого типа Sloop.
Фракции спирторастворимого белка, полученные выше, обогащали по гордеинам с помощью FPLC, как в Примере 5. Белки в каждом элюате FPLC лиофилизировали и определяли выход FPLC-очищенного белка с 10 г муки. Это показало, что содержание гордеина в G1* было уменьшено менее чем до 8 мг/10 г муки по сравнению со 105 мг/10 г муки для Sloop, 38 для R56 и 24 для R1508. Это соответствовало снижению содержания гордеина в зерне и муке G1*, по меньшей мере, на 92% по сравнению со Sloop.
Пример 10. Более масштабное соложение и производство пива с применением зерна F4
Эксперименты по соложению проводили в более крупном масштабе, чтобы получить достаточное количество солода для пробного производства пива с применением зерна F4. В указанных тестах использовали измененные схемы вымачивания, с учетом, среди прочих факторов, меньшего размера зерна, следующим образом. Образцы зерна по 800 г помещали в емкости для соложения. Применяли схему вымачивания 17°C в течение 5 часов, температура проращивания составляла 15°C в течение 94 часов. Программа сушки являлась следующей: 50-78°C в течение 17 часов, 50-74°C в течение 17 часов. При производстве солода не использовали гиббереллиновую кислоту, это не являлось необходимым.
Рецепт приготовления затора: 4,65 кг низкоглютенового солода, 10 литров воды, 10 г хлорида кальция, 2 г сульфата кальция, 64-65°C в течение 2 часов.
Котел: добавляли 17 г хмелевых гранул Target (содержание альфа-кислот (AA) 10,0%) в течение 60 минут, 21 г хмелевых гранул Hallertau (AA - 4,5%) в течение 10 минут.
Брожение проводили в 19-литровой емкости периодического цикла, при температуре ферментации 12°C, с использованием 12 г сухих дрожжей Fermentis W34/70, с первичным брожением в течение 8 дней, затем при охлаждении до 0°C в течение 9 дней. Затем пиво фильтровали через фильтр с порами 1 микрон, под давлением углекислого газа заливали в кег, и разливали в бутылки с помощью машины для розлива под противодавлением. Исходная удельная плотность составляла 1,044, конечная плотность ферментированного продукта составила 1,013 с приблизительной горечью 30 Единиц (IBU), и приблизительное содержание спирта - 4,0 об%. Другие параметры, измеренные в ходе процесса производства, являлись следующими: влажность солода: 4,2%, экстрактивность 71,5; цвет 3,9; WC 1,0; TN 2,63% сухого веса; SN 1,11; KI 51; Вязкость 1,52; AAL 71,8%; бета- глюкозидаза 130 мг/л; DP 24.
Все указанные параметры показали, что пиво может быть приготовлено из солода, полученного из зерна F4.
Более масштабные тесты соложения и производства пива также выполняли с применением зерна ячменя G1*. Восемьсот граммов зерна осолаживали в автоматизированном солодорастильном аппарате Joe White Maltings, согласно указанному протоколу. Предпочтительные условия соложения для зерна G1* определяли как следующие: вымачивание 3 часа при 17°C, проращивание 4 дня при 15°C, с последующей сушкой при 50-80°C в сушильной печи. Оптимальный промежуток времени для вымачивания зерна G1* несколько отличался по сравнению с другими зернами: Sloop: программа 8ч-9ч-5ч вымачивание/выстаивание/вымачивание при 17°C; R1508: программа 7ч-8ч-3ч вымачивание/выстаивание/вымачивание шага в 17°C; R56: программа 8ч-10ч-5ч вымачивание/выстаивание/ вымачивание при 17°C. Применяли методики анализа, установленные в соответствии с Европейской Конвенцией Пивоваренных производств (EBC) или Институтом Пивоварения (IOB). Влагосодержание зерна определяли с помощью спектроскопии в ближней ИК-области (NIR). Полное содержание азота определяли методом Дюма. Данные для солода приведены в Таблице 9. Одно существенное различие между зерном G1* и другими протестированными сортами заключалось в том, что показатели диастатической активности, определенные для G1* и R1508, были намного ниже, чем для зерна Sloop или R56. Это связали, таким образом, с lys3 мутацией в G1* и R1508.
Соложения повторяли и объединяли для каждого сорта. Приблизительно из 4 кг солода каждой линии G1*, R56, R1508 и Sloop (дикий тип) приготовили пиво и разлили в бутылки следующим образом. В образцы солода добавляли горечь, хмель Tettnang, в течение 60 мин при температуре кипения, достигнув 21-22 единиц горечи (IBU). Брожение проводили с использование дрожжей US-05 (Fermentis) при 18-20°C. Ферментированный продукт разливали в кеги без фильтрации и газировали под давлением перед розливом в бутылки. Все образцы пива еще являлись мутными при розливе, но через 2-4 недели хранения стали менее мутными. Пиво обладало заметным ароматом и привкусом "молочной карамели" из-за наличия диацетила при розливе в кеги и бутылки, но он также исчез при хранении.
Данные по приготовленным пивным продуктам приведены в Таблице 10. Содержание спирта в пиве, приготовленном из зерна G1*, составило 4,2 об%. Пиво G1* имело несколько пониженный, но удовлетворительный уровень пены после разлива.

(об%)
Данные эксперименты показали, что зерно G1* может успешно применяться в производстве солода и пива.
Уровни гордеинов в пиве, приготовленном из зерна G1*, измеренные с помощью иммуноанализа, как ожидается, ниже 1 м.д., а в некоторых случаях, ниже 0,5 м.д. Это соответствует диапазону содержания гордеинов в пшеничном пиве 10-41 м.д., в стауте 9-15 м.д., в лагерном пиве 3-9 м.д.
Специалистам в данной области будет понятно, что возможны многочисленные изменения и/или модификации, как показано в некоторых вариантах осуществления, без отступления от сущности или объема изобретения, как в целом описано. Настоящие варианты осуществления, поэтому, следует расценивать во всех отношениях как иллюстративные и неограничивающие.
Настоящая заявка имеет приоритет заявки США Серийный № 60/964672, все содержание которой включено в настоящее описание путем ссылки.
Все публикации, описанные и/или процитированные в настоящей заявке, полностью включены в настоящее описание путем отсылки.
Любое рассмотрение документов, действий, материалов, устройств, изделий и т.п., которые были включены в настоящее описание, предназначено исключительно в целях обеспечения контекста для настоящего изобретения. Не следует рассматривать как признание, что какие-либо из указанных объектов являются частью предшествующего уровня техники или являлись общеизвестным, доступным знанием в области, к которой относится настоящее изобретение, поскольку они существовали до даты приоритета каждого пункта формулы настоящей заявки.


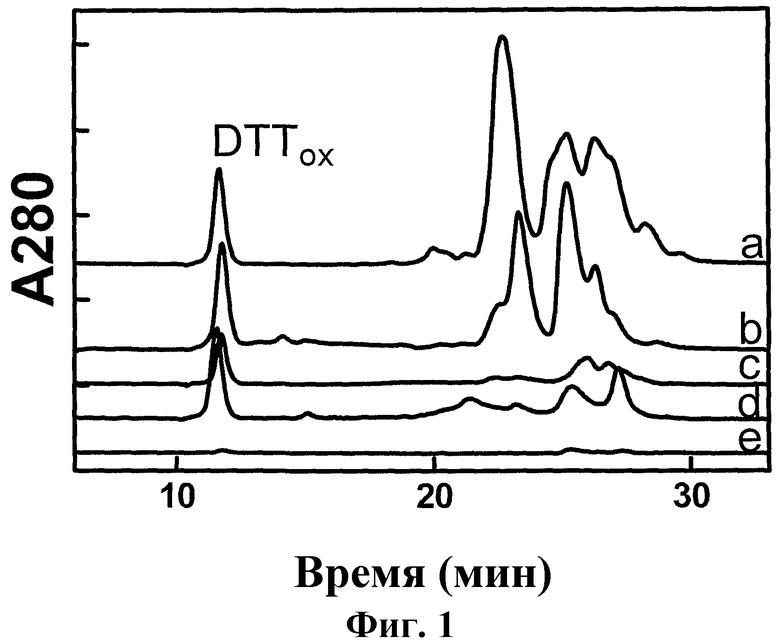
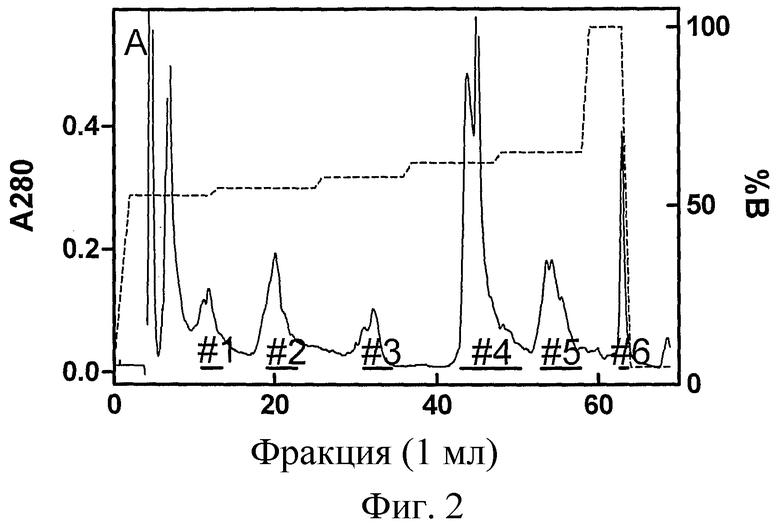
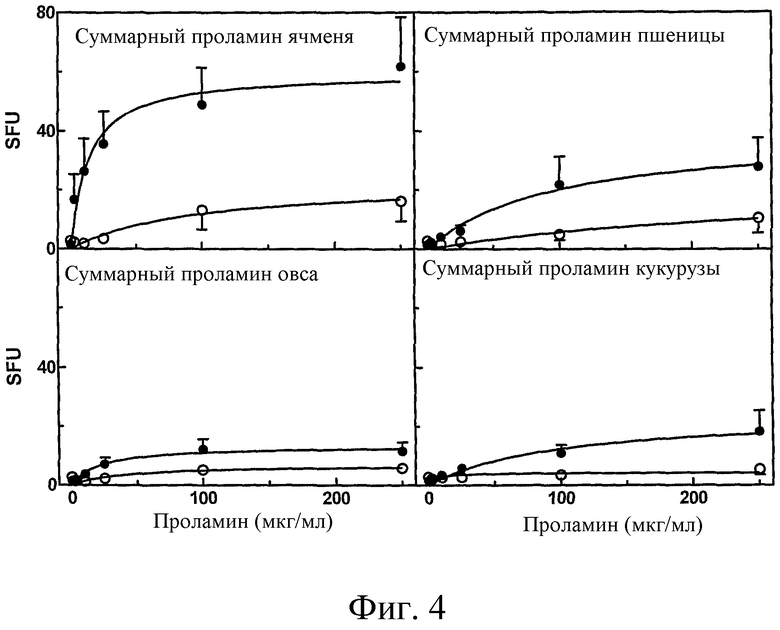
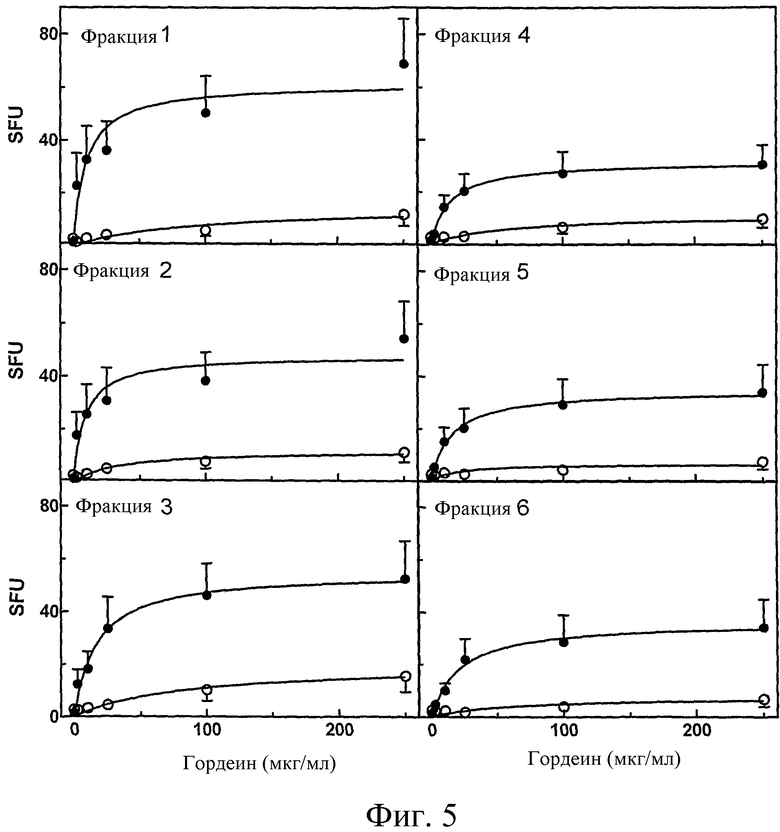
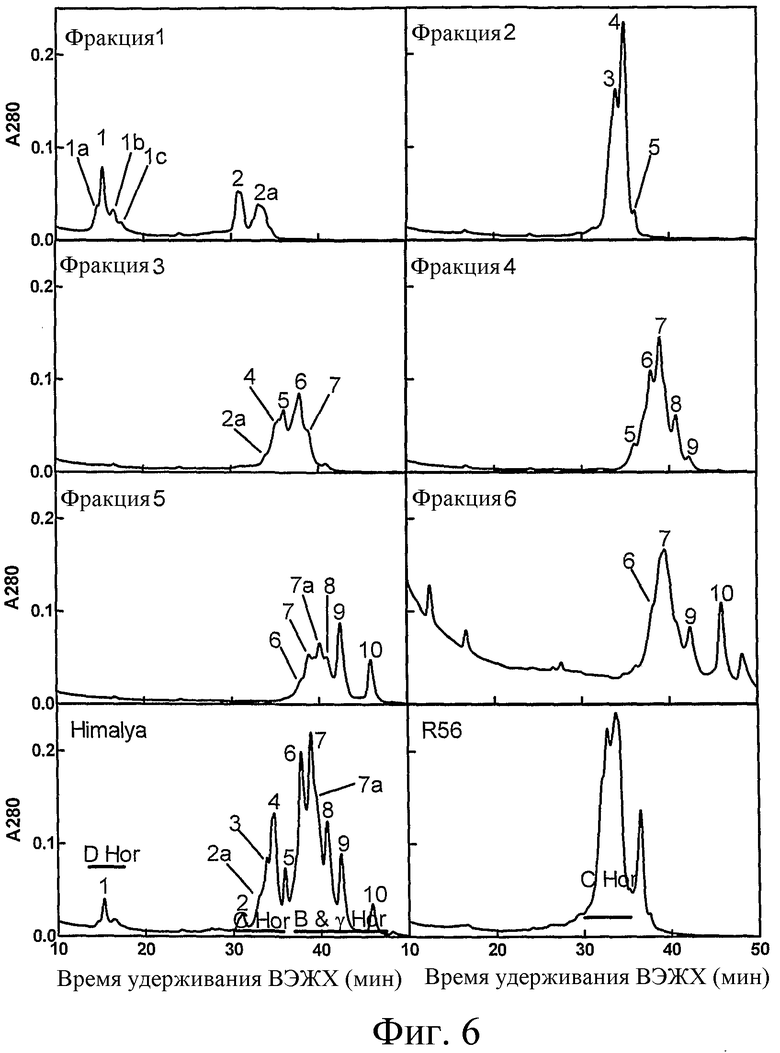
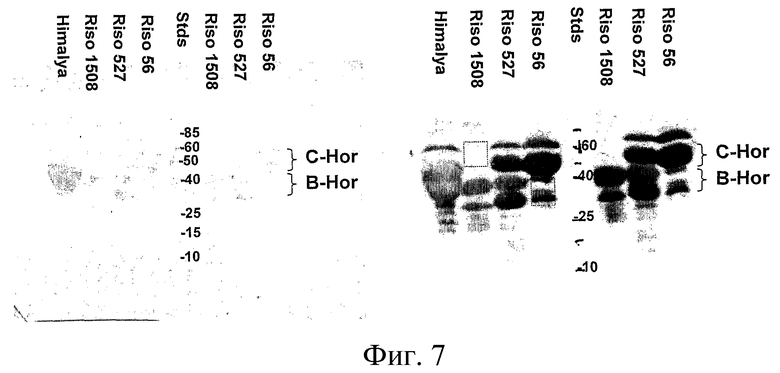
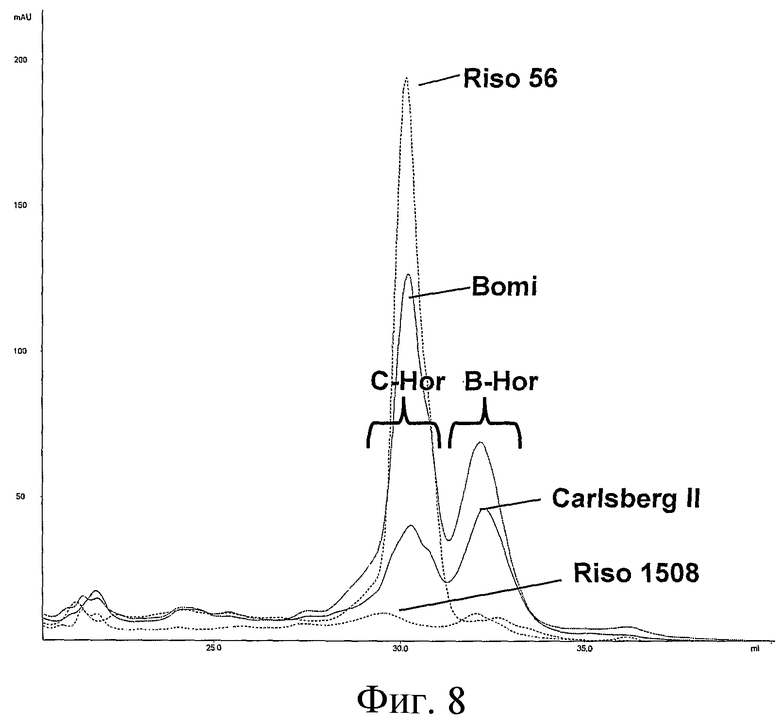
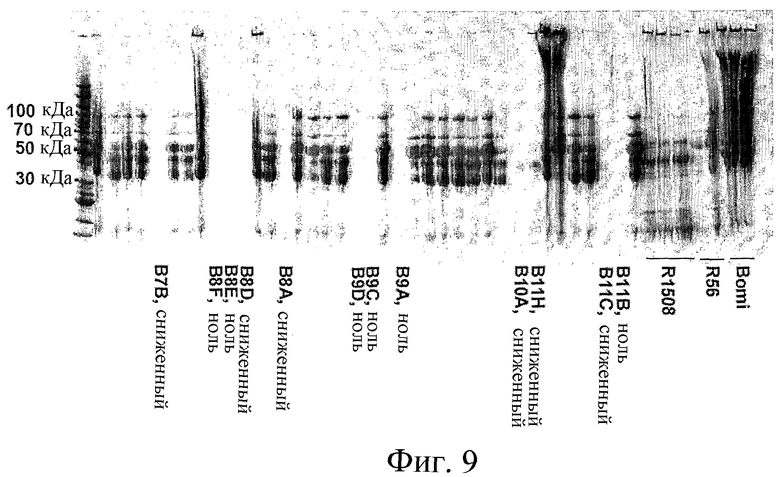
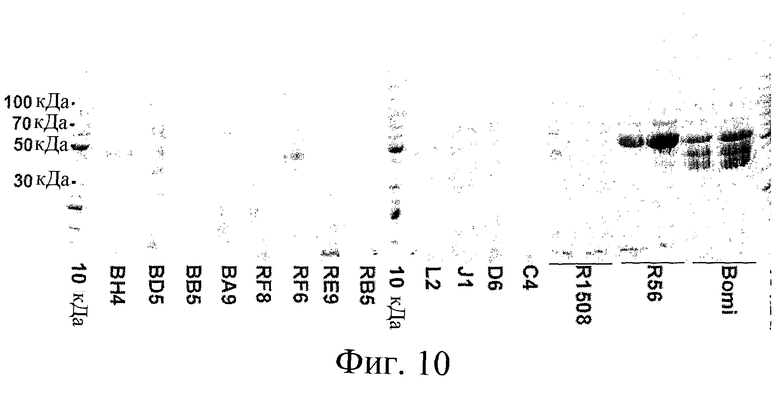
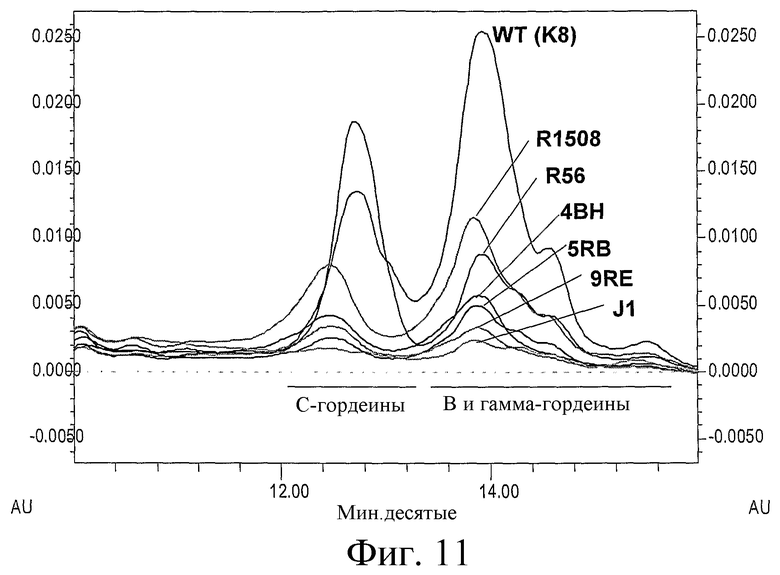
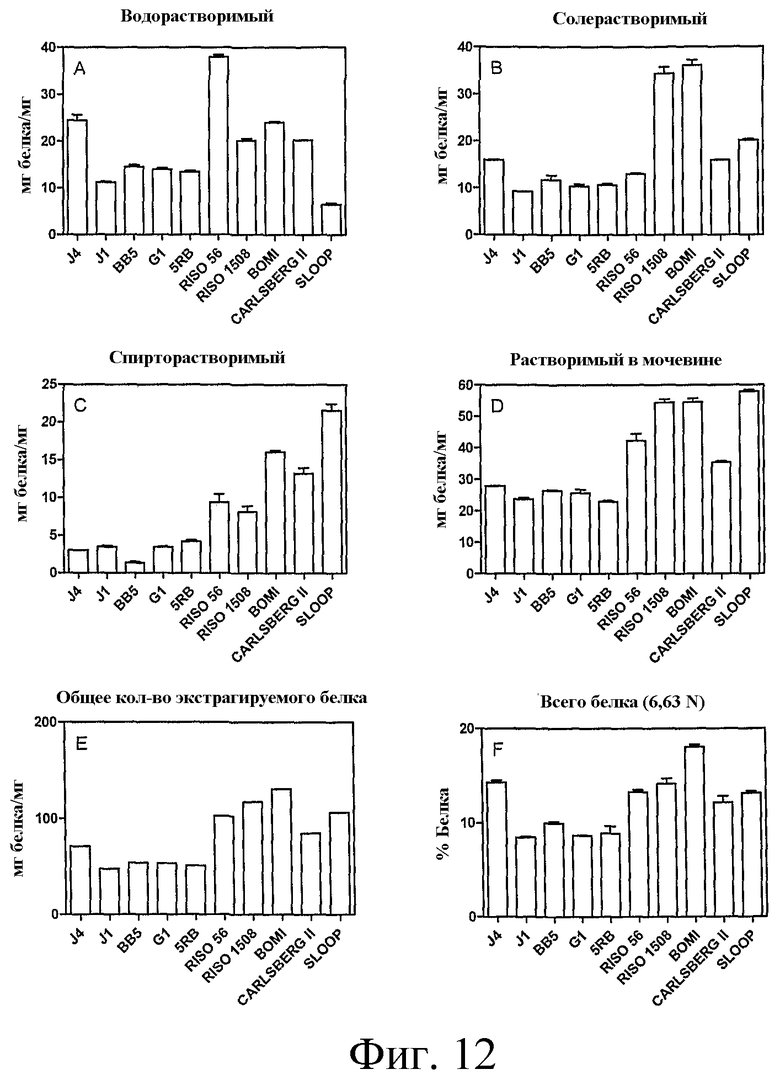
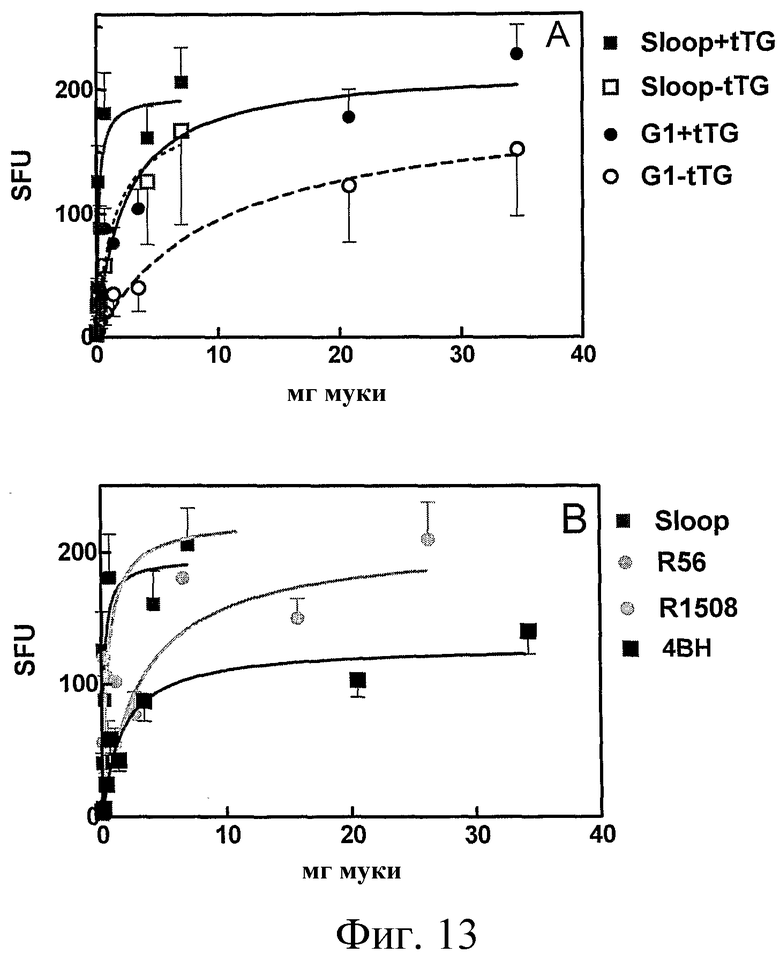
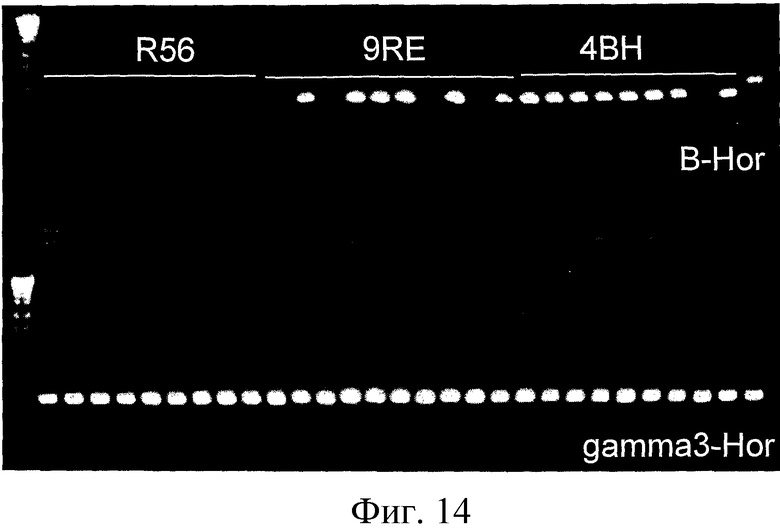
Изобретение относится к области биотехнологии и пищевой промышленности. Представлено растение ячменя, которое дает зерно и является гомозиготным, по меньшей мере, в двух локусах для введенных генетических вариаций, которые представляют собой: a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и b) мутантную аллель в локусе Lys3 ячменя, так что зерно не содержит ни В-, ни С- гордеинов, и указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно, при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров: 5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3', 3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3', а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ. Также представлены: зерно ячменя, полученное от указанного растения; пищевые продукты, не содержащие В- и С-гордеинов, полученные из указанного зерна, такие как мука, солод, пиво. Кроме того, описаны способы получения пищевых продуктов (муки, цельнозерновой муки, крахмала, солода) и напитков с использованием зерна, полученного от растения ячменя, имеющего указанные выше признаки. Предложен способ идентификации ячменного зерна, пригодного для производства пищевого продукта и/или напитка на основе солода, пригодного для употребления лицом с целиакией, включающий: a) получение одного или нескольких материалов: i) образца растения, способного давать указанное зерно, ii) зерна, iii) солода, полученного из зерна, и/или iv) экстракта указанного зерна; b) анализ материала со стадии а) на наличие, по меньшей мере, одного гордеина и/или, по меньшей мере, одного гена, кодирующего гордеин, с выбором зерна, имеющего генный набор указанного выше растения. Изобретение позволяет получать пищевые продукты или напитки на основе солода, которые не содержат В- и С-гордеинов. 14 н. и 13 з.п. ф-лы, 14 ил., 10 табл., 10 пр.
1. Способ производства пищевого продукта для потребления человеком, включающий смешивание ячменного зерна или солода, муки или цельнозерновой муки, полученных из указанного зерна, по меньшей мере, с одним другим компонентом пищевого продукта с получением пищевого продукта, причем зерно получено из растения, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
2. Способ по п.1, в котором указанный пищевой продукт является дрожжевым или бездрожжевым хлебом, макаронами, вермишелью, зерновым завтраком, закусочным продуктом, пирогом, кондитерским изделием или пищевым продуктом, содержащим соус на основе муки.
3. Способ производства напитка на основе солода для потребления человеком, включающий получение солода из ячменного зерна и смешивание этого солода с, по меньшей мере, одним другим компонентом напитка для получения напитка, причем указанное ячменное зерно получено из растения, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
4. Способ по п.3, в котором получение солода из ячменного зерна включает замачивание зерна, проращивание вымоченного зерна в заданных условиях и сушку пророщенного зерна.
5. Способ по п.3, в котором указанным напитком на основе солода является пиво или виски.
6. Способ по любому из пп.1-5, в котором электрофорез на полиакриламидном геле не обнаруживает В- и С-гордеинов.
7. Способ по любому из пп.1-5, в котором зерно имеет один или более из следующих признаков:
i) имеет средний вес 100 зерен по меньшей мере около 2,4 г,
ii) имеет средний вес 100 зерен от около 2,4 г до около 6 г,
iii) содержит по меньшей мере около 50 вес.% крахмала или
iv) содержит от около 50 вес.% до около 70 вес.% крахмала.
8. Способ по любому из пп.1-5, в котором токсичность при целиакии муки, полученной из указанного зерна, соответствует:
i) менее чем около 50% токсичности муки, полученной из зерна родительского растения ячменя,
ii) менее чем около 25% токсичности муки, полученной из зерна родительского растения ячменя,
iii) менее чем около 10% токсичности муки, полученной из зерна родительского растения ячменя.
9. Способ по любому из пп.1-5, в котором по меньшей мере около 50% зерна прорастает в течение 3 дней после замачивания.
10. Растение ячменя, которое дает зерно и является гомозиготным, по меньшей мере, в двух локусах для введенных генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
11. Растение ячменя по п.10, в котором содержание В- и С-гордеинов является необнаружимым посредством электрофореза на полиакриламидном геле.
12. Зерно растения ячменя по п.10 или 11, используемое для производства пищевого продукта или напитка на основе солода для потребления человеком.
13. Зерно по п.12, имеющее один или более из следующих признаков:
i) имеет средний вес 100 зерен по меньшей мере около 2,4 г,
ii) имеет средний вес 100 зерен от около 2,4 г до около 6 г,
iii) содержит по меньшей мере около 50 вес.% крахмала или
iv) содержит от около 50 вес.% до около 70 вес.% крахмала.
14. Способ получения ячменного зерна, включающий
выращивание растения ячменя по п.10 или 11, и
сбор урожая зерна.
15. Способ получения муки, включающий
получение зерна по п.12 или 13, и
обработку зерна для получения муки.
16. Способ получения цельнозерновой муки, включающий
получение зерна по п. 12 или 13, и
обработку зерна для получения цельнозерновой муки.
17. Способ получения крахмала, включающий
получение зерна по п. 12 или 13, и
обработку зерна для получения крахмала.
18. Способ получения солода, включающий
получение зерна по п.12 или 13, и
обработку зерна для получения солода.
19. Пиво для потребления человеком, полученное способом по любому из пп.3-9, в котором зерно получено от растения, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
20. Пиво по п.19, в котором содержание В- и С-гордеинов является необнаружимым посредством электрофореза на полиакриламидном геле.
21. Мука для потребления человеком, полученная способом по любому из пп.1,6-9, в котором зерно получено от растения, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В- ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
22. Мука по п.21, в которой содержание В- и С-гордеинов является необнаружимым посредством электрофореза на полиакриламидном геле.
23. Солод для потребления человеком, полученный способом по любому из пп.1,6-9, в котором зерно получено от растения, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
24. Солод по п.23, в котором содержание В- и С-гордеинов является необнаружимым посредством электрофореза на полиакриламидном геле.
25. Пищевой продукт для потребления человеком, полученный способом по любому из пп.1,6-9, в котором зерно получено от растения, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
26. Пищевой продукт по п.25, в котором содержание В- и С-гордеинов является необнаружимым посредством электрофореза на полиакриламидном геле.
27. Способ идентификации ячменного зерна, пригодного для производства пищевого продукта и/или напитка на основе солода, пригодного для употребления лицом с целиакией, включающий:
a) получение одного или нескольких следующих материалов:
i) образец растения, способного давать указанное зерно,
ii) зерно,
iii) солод, полученный из зерна, и/или
iv) экстракт указанного зерна,
b) анализ материала со стадии а) на наличие, по меньшей мере, одного гордеина и/или, по меньшей мере, одного гена, кодирующего гордеин,
причем выбирают ячменное зерно, которое является гомозиготным, по меньшей мере, в двух локусах для генетических вариаций, которые представляют собой:
a) аллель, в которой удалена большая часть или все гены, кодирующие В-гордеин в локусе Hor2, и
b) мутантную аллель в локусе Lys3 ячменя,
так что зерно не содержит, ни В-, ни С-гордеинов, и
указанные генетические вариации присутствуют в ячмене линий Riso 56 и Riso 1508 соответственно,
при этом отсутствие В-гордеинов является обнаружимым по отсутствию амплифицированной ДНК с использованием праймеров:
5'B1hor: 5'-CAACAATGAAGACCTTCCTC-3' (SEQ ID NO:2),
3'B1hor: 5'-TCGCAGGATCCTGTACAACG-3' (SEQ ID NO:3),
а отсутствие С-гордеинов является обнаружимым по отсутствию 70 кДа полосы при исследовании спирторастворимого экстракта зерна посредством ДСН-ПААГ.
| СПОСОБ ПОЛУЧЕНИЯ СЫРЬЯ ДЛЯ ПИВОВАРЕНИЯ, СЫРЬЕ ДЛЯ ПИВОВАРЕНИЯ, ПОЛУЧЕННОЕ СОГЛАСНО СПОСОБУ, И СПОСОБ ПИВОВАРЕНИЯ | 1993 |
|
RU2109040C1 |
| СПОСОБ ПОВЫШЕНИЯ КАЧЕСТВА ЗЕРНА КУКУРУЗЫ | 1991 |
|
RU2083088C1 |
| Привод | 1983 |
|
SU1210869A1 |
| US 2007116846 A1, 24.05.2007 | |||
| Генератор импульсов | 1988 |
|
SU1657300A1 |
| US 2008152780 A1, 26.06.2008 | |||

