Настоящее изобретение относится к области медицины. Точнее настоящее изобретение относится к иммуногенным белкам Streptococcus, а также к их иммуногенным частям, производным и аналогам.
Род Streptococcus включает широкий круг патогенных и комменсальных грамположительных бактерий, которые существуют в различных хозяевах, в том числе в организмах людей, лошадей, свиней и коров. В организме хозяина стрептококки обычно поражают слизистые поверхности верхних дыхательных путей. Однако в некоторых обстоятельствах стрептококки также могут вызывать заболевания, тяжесть которых варьирует от подострой до острой и даже хронической.
По настоящее время многие коммерческие вакцины против Streptococcus основаны на бактеринах целых клеток. Обычно такие бактерины действительно оказывают существенную защиту против заражения гомологичными серотипами, но не защищают против заражения гетерологичными серотипами. Вакцинация целыми клетками Streptococcus часто приводит к иммунному ответу, который направлен против того же штамма Streptococcus, но который не направлен (существенным образом) против других штаммов Streptococcus, не говоря уже о других серотипах Streptococcus. В результате многие вакцины представляют недостаточную защиту против гетерологичных штаммов и/или серотипов, поскольку вакцинация против одного штамма Streptococcus обычно недостаточна для противодействия инфицированию другим штаммом Streptococcus. Кроме того, вакцинация против одного серотипа Streptococcus обычно недостаточна для противодействия инфицированию другим серотипом Streptococcus. Таким образом, желательно, чтобы иммуногенные композиции были способны вызывать иммунный ответ против по меньшей мере двух штаммов Streptococcus, предпочтительно против двух серотипов Streptococcus.
Одна из задач настоящего изобретения заключается в получении белков Streptococcus, а также их иммуногенных частей, производных и/или аналогов, и молекул кодирующих их нуклеиновых кислот таким образом, что они способны индуцировать иммунный ответ против по меньшей мере двух штаммов Streptococcus.
Настоящее изобретение предусматривает способ идентификации белка Streptococcus, который способен вызвать иммунный ответ против по меньшей мере двух штаммов Streptococcus, и включает:
а) идентификацию по меньшей мере части секретированного белка, поверхностно-ассоциированного белка и/или белка, который идентичен по меньшей мере на 50% последовательности бактериального фактора вирулентности,
б) выбор по меньшей мере одного белка, идентифицированного на стадии а), который сохраняется по меньшей мере в двух штаммах Streptococcus, и
в) определение способности по меньшей мере одного белка, выбранного на стадии б), или его иммуногенной части, производного и/или аналога, специфически связывать антитело и/или иммунные клетки животного, инфицированного первым штаммом Streptococcus, и вторым антителом и/или иммунными клетками животного, инфицированного вторым штаммом Streptococcus. Указанный первый штамм Streptococcus и указанный второй штамм Streptococcus предпочтительно относятся к одному виду Streptococcus. Предпочтительно указанный белок, последовательность которого по меньшей мере на 50% идентична последовательности бактериального фактора вирулентности, обладает по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, наиболее предпочтительно по меньшей мере на 80%, идентичностью с последовательностью бактериального фактора вирулентности.
По настоящему изобретению идентифицируют по меньшей мере один белок Streptococcus, который способен вызвать иммунный ответ против по меньшей мере двух штаммов Streptococcus. Указанный белок может применяться для иммунизации конкретного животного и/или человека, поскольку может вызывать широкий иммунный ответ. Таким образом, настоящее изобретение не требует получения вакцины для каждого штамма и/или серотипа Streptococcus. Применение иммуногенного белка Streptococcus по настоящему изобретению, таким образом, сохраняет время и деньги. Важнее, что иммуногенный белок Streptococcus по настоящему изобретению в принципе способен вызывать иммунный ответ против штамма Streptococcus, который пока еще неизвестен, или против штамма, в отношении которого еще нет специфической вакцины (например, штамма, который недавно возник в природных условиях). Предпочтительно белок Streptococcus по настоящему изобретению способен индуцировать иммунный ответ против по меньшей мере двух серотипов Streptococcus. Предпочтительный вариант осуществления настоящего изобретения, таким образом, предусматривает способ идентификации белка Streptococcus, который способен индуцировать иммунный ответ против по меньшей мере двух серотипов Streptococcus, включающий:
а) идентификацию по меньшей мере части секретированного белка, поверхностно-ассоциированного белка и/или белка, который идентичен по меньшей мере на 50% последовательности бактериального фактора вирулентности,
б) выбор по меньшей мере одного белка, выявленного на стадии а), который сохраняется по меньшей мере в двух серотипах Streptococcus, и
в) определение способности по меньшей мере одного белка, выбранного на стадии б), или его иммуногенной части, производного и/или аналога, специфически связывать антитело и/или иммунные клетки животного, инфицированного первым серотипом Streptococcus, и антителом и/или иммунными клетками животного, инфицированного вторым серотипом Streptococcus.
Иммунный ответ против по меньшей мере двух штаммов Streptococcus и/или двух серотипов Streptococcus в контексте настоящего изобретения выражают в качестве гуморального и/или клеточного иммунного ответа, направленного против Streptococcus по меньшей мере двух разных штаммов и/или серотипов. Указанный иммунный ответ индуцируется, например, в животном, но не в организме человека. Также возможно индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus в организме конкретного человека, для того, чтобы предупредить и/или воспрепятствовать заболеванию, связанному со Streptococcus. Гуморальный иммунный ответ приводит к выработке антител, несмотря на то, что клеточный иммунный ответ преимущественно повышает формирование реактивных иммунных клеток, например, Т-клеток-киллеров. Обычно обе части иммунного ответа индуцируются введением иммуногенного белка или его иммуногенной части. Иммунный ответ против по меньшей мере двух штаммов/серотипов Streptococcus предпочтительно включает выработку антитела. Указанный иммунный ответ предпочтительно способен по меньшей мере частично понизить число микроорганизмов Streptococcus в организме конкретного человека и/или животного. Указанный иммунный ответ также предпочтительно способен по меньшей мере частично, препятствовать расстройству, вызываемому Streptococcus.
Штамм Streptococcus идентифицируют по морфологическим, биохимическим и серологическим признакам, известным в данной области. Серотип Streptococcus является группой Streptococcus, классификация которой основывается на наличии специфических антигенных полисахаридов. Классификация серотипов Streptococcus также известна в данной области.
Способ по настоящему изобретению включает идентификацию по меньшей мере части секретированного белка, поверхностно-ассоциированного белка и/или белка, последовательность которого идентична по меньшей мере на 50% последовательности бактериального фактора вирулентности. Указанный белок идентифицируют разными способами. В одном из вариантов осуществления настоящего изобретения применяют геномный подход. Ген, кодирующий секретируемый белок и/или поверхностно-ассоциированный белок, идентифицируют, например, по поиску мотива указанного секретированного белка и/или поверхностно-ассоциированного белка. Указанный мотив предпочтительно включает сайт присоединения липида, сайт расщепления сигнальной пептидазы и/или сайт присоединения сортазы. Безусловно возможно обнаружить другие мотивы, известные в данной области. В одном из вариантов осуществления настоящего изобретения, таким образом, предусматривают способ по настоящему изобретению, в котором указанный секретированный белок и/или поверхностно-ассоциированный белок выявляют путем идентификации по меньшей мере части последовательности генома гена Streptococcus, включающего мотив секретированного и/или поверхностно-ассоциированного белка. Дополнительно или в другом варианте ген, кодирующий секретируемый белок и/или поверхностно-ассоциированный белок, идентифицируют одним или несколькими методами, известными в данной области. Например, если ген бактерий вида рода Streptococcus, кодирующий секретируемый белок и/или поверхностно-ассоциированный белок, известен, можно обнаружить другую геномную последовательность Streptococcus по наличию гена с высоким процентом идентичности последовательности.
В одном из вариантов осуществления настоящего изобретения указанный метод скрининга включает способ, в котором указанная другая геномная последовательность Streptococcus подвергается скринингу для выявления способности гибридизироваться с нуклеотидной последовательностью, кодирующей секретированный и/или поверхностно-ассоциированный белок Streptococcus. Настоящее изобретение, таким образом, предусматривает способ по настоящему изобретению, в котором указанный белок, имеющий по меньшей мере 50% идентичность с бактериальным фактором вирулентности, идентифицируют по меньшей мере у части геномной последовательности гена Streptococcus, который способен к гибридизации с какой-либо из нуклеотидных последовательностей, перечисленных на фиг.4, при 65°С в буфере, содержащем 0,5 М натрий фосфат, 1 мМ EDTA и 7% натрий додецилсульфат при pH 7,2, причем молекулы нуклеиновой кислоты остаются гибридизированными после двухкратной промывки буфером, содержащим 40 мМ натрий фосфат (pH 7,2), 1 мМ EDTA и 5% натрий додецилсульфат в течение 30 мин при 65°С, и промывают дважды буфером, содержащим 40 мМ натрий фосфат (pH 7,2), 1 мМ EDTA и 1% натрий додецилсульфат в течение 30 мин при 65°С. Предпочтительно указанный белок, обладающий по меньшей мере 50% идентичностью к последовательности бактериального фактора вирулентности, идентичен по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере, на 75%, наиболее предпочтительно по меньшей мере на 80% последовательности бактериального фактора вирулентности.
В данной области также предусматривают различные способы определения, имеет ли последовательность белка Streptococcus идентичность по меньшей мере равную 50%, по отношению к последовательности бактериального фактора вирулентности. Например, аминокислотную последовательность белка Streptococcus сравнивают с аминокислотной последовательностью бактериального фактора вирулентности. Также возможно применение геномного подхода. Ген, кодирующий белок Streptococcus, последовательность которого по меньшей мере на 50% идентична последовательности бактериального фактора вирулентности, выявляют, например, скринингом геномной последовательности Streptococcus по нуклеотидной последовательности, которая обладает по меньшей мере 50% идентичностью последовательности бактериального гена, кодирующего фактор вирулентности. В одном из вариантов осуществления настоящего изобретения, таким образом, предусматривают способ по настоящему изобретению, в котором белок, который обладает по меньшей мере 50% идентичностью последовательности бактериального фактора вирулентности, идентифицируют путем идентификации по меньшей мере части геномной последовательности гена Streptococcus, которая обладает по меньшей мере 50% идентичностью последовательности гена бактериального фактора вирулентности. Однако многие другие способы определения, обладает ли белок Streptococcus по меньшей мере 50% идентичностью последовательности к последовательности бактериального фактора вирулентности, известны в данной области.
Если идентифицирован по меньшей мере один ген Streptococcus, кодирующий секретируемый белок, поверхностно-ассоциированный белок и/или белок, который идентичен по меньшей мере на 50% последовательности бактериального фактора вирулентности, предпочтительно определить, может ли по меньшей мере один из указанных генов сохраняться по меньшей мере в двух штаммах Streptococcus. Ген первого штамма Streptococcus сохраняется по меньшей мере в двух штаммах Streptococcus, если геном второго штамма Streptococcus включает а последовательность нуклеиновой кислоты, которая по меньшей мере примерно на 60% идентична указанному гену указанного первого штамма first Streptococcus. Предпочтительно, указанная последовательность нуклеиновой кислоты по меньшей мере на 70%, более предпочтительно по меньшей мере, на 75%, более предпочтительно по меньшей мере, на 80% более предпочтительно по меньшей мере, на 90%, наиболее предпочтительно по меньшей мере на 95% идентична последовательности указанного гена. Понятие «идентичность последовательностей» относится к процентной идентичности между двумя последовательностями нуклеиновой кислоты или последовательностями аминокислот. Две последовательности нуклеиновой кислоты обладают идентичностью по меньшей мере на 60% по отношению друг к другу после выравнивания и внедрения гэпов, при необходимости, для достижения максимального процента идентичности последовательностей. Способы и компьютерные программы для выравнивания известны в данной области. Одной из компьютерных программ, которая может быть применена или адаптирована для целей определения, попадает ли последовательность-кандидат под данное определение, является программа «Align 2», разработанная фирмой Genentech, Inc., которая прилагается к документации для пользователя Ведомства по охране авторских прав США, Вашингтон, округ Колумбия 20559, 10 декабря 1991 года.
В одном из вариантов осуществления настоящего изобретения, если ген по настоящему изобретению сохраняется по меньшей мере в двух штаммах Streptococcus, белок, кодируемый указанным геном, является хорошим кандидатом для оценки, является ли указанный белок или его иммуногенная часть, производное и/или аналог, способным вызвать иммунный ответ против более чем одного штамма Streptococcus. Указанный первый штамм Streptococcus и указанный второй штамм Streptococcus предпочтительно являются одним и тем же видом Streptococcus.
Предпочтительно определяют, является ли указанный ген сохраняется по меньшей мере в двух серотипах Streptococcus, для того, чтобы идентифицировать хороший белок-кандидат (кодируемый указанным геном), который тестируют по способности вызывать иммунный ответ против более чем одного серотипа Streptococcus. Способ по настоящему изобретению, который также включает выбор гена, который сохраняется по меньшей мере в двух штаммах и/или серотипах Streptococcus, таким образом, является предпочтительным.
Если идентифицируют ген, который сохраняется по меньшей мере в двух штаммах/серотипах Streptococcus, предпочтительно получают белок, кодируемый указанным геном. Дополнительно, или в другом варианте, получают иммуногенную часть, производное и/или аналог указанного белка. В данной области известны разные методы получения белка, кодируемого геном, или его иммуногенной части, производного и/или аналога. Указанный ген, например, экспрессируют в соответствующей системе экспрессии. Примерами, которые не ограничивают настоящее изобретение, являются эукариотические клетки-хозяева, например, дрожжи и прокариотические клетки-хозяева, например, Escherichia coli. Предпочтительно ген по настоящему изобретению, кодирующий секретированный белок, ассоциированный с поверхностью белок и/или белок, который по меньшей мере на 50% идентичен последовательности бактериального фактора вирулентности, и который сохраняется по меньшей мере в двух штаммах Streptococcus, белок, кодируемый указанным геном, экспрессируется в прокариотической системе экспрессии. Прокариотичекая система экспрессии является предпочтительной, поскольку белок (прокариотический) бактерии Streptococcus в принципе лучше экспрессируется в прокариотической системе экспрессии. Кроме того, прокариотическую систему экспрессии обычно легче создать и использовать.
Способ по настоящему изобретению включает выяснение, является ли по меньшей мере один белок по настоящему изобретению или его иммуногенная часть, производное и/или аналог, способными специфически связывать антитело и/или иммунные клетки животного, инфицированного первым штаммом Streptococcus, и антителом и/или иммунными клетками животного, инфицированного вторым штаммом Streptococcus. Предпочтительно определяют, может ли по меньшей мере один белок по настоящему изобретению или его иммуногенная часть, производное и/или аналог, обладать способностью специфически связывать антитело и/или иммунные клетки животного, инфицированного первым серотипом Streptococcus, и антитело и/или иммунные клетки животного, инфицированного вторым серотипом Streptococcus. Многие способы известны в данной области для выполнения указанного теста. Предпочтительно используют сыворотку по меньшей мере двух животных, инфицированных по меньшей мере двумя разными штаммами Streptococcus. В другом варианте используют сыворотку только одного животного, инфицированного по меньшей мере двумя разными штаммами Streptococcus. В одном из вариантов осуществления настоящего изобретения одно животное, но не человека, инфицируют по меньшей мере первым штаммом и/или серотипом Streptococcus, и второе животное, но не человека, инфицируют по меньшей мере вторым штаммом и/или серотипом Streptococcus. Указанные штаммы и/или серотипы Streptococcus, например, вводят внутривенно указанному животному. Затем по одному из вариантов осуществления настоящего изобретения отбирают сыворотку от указанных животных, имеющих Streptococcus-специфические антитела и/или иммунные клетки. Указанную сыворотку необязательно перерабатывают перед применением. Например, антитела и/или иммунные клетки, являются по меньшей мере частично сконцентрированными и/или выделенными. Белок по настоящему изобретению и/или его иммуногенная часть, производное и/или аналог, предпочтительно выделяют и/или получают рекомбинантно и затем инкубируют с указанной сывороткой - или с (частично) выделенным антителом и/или иммунными клетками - производными от указанных животных. Возможно применение сыворотки, антитела и/или иммунных клеток, полученных от первого животного, вместе с сывороткой, антителом и/или иммунными клетками, полученными от второго животного. В другом варианте, сыворотка, антитело и/или иммунные клетки, полученные от первого животного, вводят сначала, затем дополнительно вводят сыворотку, антитела и/или иммунные клетки от второго животного. В еще одном из вариантов осуществления настоящего изобретения сыворотку, антитела и/или иммунные клетки от первого животного вводят в одной отдельной группе, включающей по меньшей мере один белок и/или иммуногенную часть, производное и/или аналог по настоящему изобретению, а также сыворотку, антитела и/или иммунные клетки второго животного вводят в другой группе, включающей по меньшей мере один белок и/или иммуногенную часть, производное и/или аналог по настоящему изобретению. После инкубирования указанную сыворотку, антитела и/или иммунные клетки промывают и связанные антитела и/или иммунные клетки визуализируют, используя какой-либо известный в данной области способ. Связанные антитела, например, инкубированные со вторым антителом, способны специфически связывать указанные связанные антитела, причем второе антитело конъюгировано с пероксидазой хрена. После отмывания второго несвязанного антитела вводят перекись водорода. Разрушение перекиси водорода пероксидазой хрена связано с окислением хромогенного соединения таким образом, что реакция становится видимой.
Если белок по настоящему изобретению и/или его иммуногенная часть, производное и/или аналог оказались специфически связанными антителом и/или иммунными клетками, индуцированными первым штаммом Streptococcus, и антителом и/или иммунными клетками, индуцированными вторым штаммом Streptococcus, означает, что указанный белок, иммуногенная часть, производное и/или аналог способны индуцировать иммунный ответ против по меньшей мере двух штаммов Streptococcus.
В предпочтительном варианте осуществления настоящего изобретения используют антитело и/или иммунные клетки, производные сыворотки в период выздоровления животного, которое было инфицировано Streptococcus. Сыворотку в период выздоровления получают от животного, которое эффективно выздоравливает, избавляясь от инфекции. Таким образом, сыворотка в период выздоровления животного, которое было инфицировано бактерией Streptococcus, включает антитела и/или иммунные клетки, которые способны защитить указанное животное от заражения тем же штаммом Streptococcus. Таким образом, инкубирование со средой выздоровления предпочтительно для определения, способен ли белок и/или его иммуногенная часть, производное и/или аналог по настоящему изобретению вызвать защитный иммунный ответ.
В одном из вариантов осуществления настоящего изобретения предусматривается способ идентификации белка Streptococcus, который способен вызывать иммунный ответ против по меньшей мере двух штаммов Streptococcus, включающий:
- получение выделенных и/или рекомбинантных белков Streptococcus,
- инкубирование указанных белков с антителом и/или иммунными клетками животного, инфицированного первым штаммом и/или серотипом Streptococcus, и с антителом и/или иммунными клетками животного, инфицированного вторым штаммом и/или серотипом Streptococcus, и
- определение, способен ли белок связывать антитело и/или иммунные клетки животного, инфицированного первым штаммом и/или серотипом Streptococcus, и антитело и/или иммунные клетки животного, инфицированного вторым штаммом и/или серотипом Streptococcus.
Белки Streptococcus получают разными способами. Предпочтительно секретированные белки, поверхностно-ассоциированные белки и/или белки, которые идентичны по меньшей мере на 50% последовательности бактериального фактора вирулентности, выделены из культуры Streptococcus. В одном из вариантов осуществления настоящего изобретения поверхностно-ассоциированные белки выделяют из Streptococcus, используя, например, лизоцим.
В одном из вариантов осуществления настоящего изобретения белки Streptococcus получают рекомбинантно, используя по меньшей мере одну последовательность нуклеиновой кислоты, кодирующей по меньшей мере один из указанных белков. Выше было указано, что предпочтительно используют секретированные белки, поверхностно-ассоциированные белки и/или белки, которые идентичны по меньшей мере на 50%, последовательности бактериального фактора вирулентности. Более предпочтительно, указанный ген сохраняют по меньшей мере в двух штаммах и/или серотипах Streptococcus. В другом варианте или дополнительно белок Streptococcus или его иммуногенная часть, производное и/или аналог, получают, используя другой метод, известный в данной области. Например, иммуногенный белок или пептид Streptococcus получают, используя обычный метод синтеза, например, твердофазный синтез. В другом примере белок Streptococcus выделен из Streptococcus или получен рекомбинантно, после чего его модифицируют для того, чтобы получить иммуногенную часть, производное и/или аналог.
В одном из предпочтительных вариантов осуществления настоящего изобретения белки Streptococcus разделяют на полиакриламидном геле и последовательно инкубируют с антителом и/или иммунными клетками животного, инфицированного первым штаммом и/или серотипом Streptococcus, и с антителом и/или иммунными клетками животного, инфицированного вторым штаммом и/или серотипом Streptococcus. Предпочтительно применяют двухмерный полиакриламидный гель.
В предпочтительном варианте осуществления настоящего изобретения идентифицируют белок Streptococcus, который способен вызвать антитела, индуцирующие опсонофагоцитоз. Опсонофагоцитоз является природным процессом, в котором микроорганизм опсонизируется опсонинами, после чего указанный микроорганизм фагоцитируется фагоцитными клетками и уничтожается. Многие микроорганизмы нуждаются в опсонизации опсонинами для повышения их фагоцитоза. Опсонизация - это процесс, в результате которого микроорганизм становится более чувствительным для поглощения в результате фагоцитоза. В указанном процессе опсонизированные антитела и/или белки связаны с указанным микроорганизмом, тем самым, облегчая поглощение указанного микроорганизма в результате указанного фагоцитоза. Таким образом, белок Streptococcus по настоящему изобретению или его иммуногенная часть, производное и/или аналог, индуцирующие антитела, способные вызывать опсонофагоцитоз, является предпочтительным, поскольку введение такого белка, и/или его иммуногенной части, производного и/или аналога животному приводит к наличию опсонофагоцитоз-индуцирующих антител в указанном животном, способном фагоцитировать Streptococcus.
Белок Streptococcus по настоящему изобретению, или его иммуногенная часть, производное и/или аналог способны вызвать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus. Для того чтобы вызвать еще более широкий иммунный ответ, предпочтительно выявить по меньшей мере два разных белка Streptococcus, и/или иммуногенную часть, производное и/или аналог по меньшей мере одного из указанных белков. Более предпочтительно по меньшей мере выявление трех разных белков Streptococcus, и/или иммуногенной части, производного и/или аналога по меньшей мере одного из указанных белков, и т.д. Повышенное число выявленных белков Streptococcus, и/или его иммуногенных частей, производных и/или аналогов по настоящему изобретению индуцирует более широкий иммунный ответ.
В другом предпочтительном варианте осуществления настоящего изобретения выявляют по меньшей мере один белок Streptococcus и/или иммуногенную часть, производное и/или аналог по настоящему изобретению, которые способны вызывать иммунный ответ против по меньшей мере трех штаммов Streptococcus. Указанный белок и/или иммуногенная часть, производное и/или аналог особо применимы для индукции широкого иммунного ответа у конкретного человека и/или животного. Более предпочтительно выявляют по меньшей мере один идентифицированный белок, и/или иммуногенная часть, производное, и/или аналог по настоящему изобретению из Streptococcus индуцировать иммунный ответ против по меньшей мере трех серотипов Streptococcus.
Иммуногенную часть белка определяют в качестве той части белка, которая способна вызвать иммунный ответ у конкретного человека и/или животного. Предпочтительно указанная иммуногенная часть способна вызвать тот же тип иммунного ответа, хотя необязательно в том же количестве, что и указанный белок. Иммуногенная часть белка предпочтительно включает один или несколько эпитопов указанного белка. Эпитоп белка определяют в качестве части указанного белка, состоящего по меньшей мере примерно из 5 аминокислот в длину, способной индуцировать специфическое антитело и/или иммунные клетки, способные специфически связывать указанный эпитоп. Существует два разных типа эпитопов: линейные эпитопы и конформационные эпитопы. Линейный эпитоп представляет последовательность расположенных подряд аминокислот. Конформационный эпитоп сформирован несколькими последовательностями расположенных подряд аминокислот, которые складываются определенным образом и вместе формируют эпитоп правильно сложенного белка. Иммуногенная часть по настоящему изобретению способна включать эпитопы или одного, или обоих указанных типов.
Иммуногенная часть белка представляет по меньшей мере 5 аминокислотных остатков. Предпочтительно указанная иммуногенная часть представляет по меньшей мере 10, более предпочтительно по меньшей мере 15, более предпочтительно по меньшей мере 25, и наиболее предпочтительно по меньшей мере 30 последовательно расположенных аминокислот. Указанная иммуногенная часть предпочтительно представляет самое большее примерно 500 аминокислотных остатков, более предпочтительно самое большее 250 аминокислотных остатков, в зависимости от типа белка, из которого указанная иммуногенная часть происходит.
Производное белка определяют в качестве молекулы, которая обладает теми же иммуногенными свойствами, необязательно выраженными количественно. Специалист в данной области способен изменить белок таким образом, что иммуногенные свойства указанной молекулы в существенной степени того же типа, но необязательно выраженные количественно, по сравнению с указанным белком. Производное белка, например, получают за счет мутации по меньшей мере одного аминокислотного остатка указанного белка и/или замещения одного аминокислотного остатка на другой аминокислотный остаток. Предпочтительно производят консервативные аминокислотные замещения, например, замещение аминокислоты, включающей кислую боковую цепочку, другой аминокислотой, включающей кислую боковую цепочку, замещение объемной аминокислоты другой объемной аминокислотой, замещение аминокислоты, включающей щелочную боковую цепочку, другой аминокислотой, включающей щелочную боковую цепочку, и т.д.
Специалист в данной области может получать аналоги белка. Так, например, это возможно путем скрининга библиотеки пептидов или изменением пептидных программ. Аналог по настоящему изобретению имеет в значительно степени те же иммуногенные свойства указанного белка, но не обязательно выраженные количественно. Аналог белка по настоящему изобретению, например, представляет гибридный белок и/или химерный белок.
Для способности индукцировать иммунный ответ, иммуногенной части, производному и/или аналогу по настоящему изобретению предпочтительно предаются свойства для обеспечения выработки антитела и/или иммунных клеток. Указанные свойства, известные в данной области, например, включают соответствующие фланкирующие последовательности и/или сайты протеолитического расщепления. В другом варианте, или дополнительно, белок, иммуногенная часть, производное и/или аналог по настоящему изобретению, предпочтительно обеспечиваются иммуногенным носителем.
Если белок или иммуногенная часть, производное и/или аналог по настоящему изобретению вводят человеку или животному, оно обычно подвергается риску разрушения, вызванному рядом разных факторов, например, протеолизом, нарушением складчатости, экстремальными величинами pH, детергентами и высокими концентрациями солей. Для продления периода целостности белка, или его иммуногенной части, производного и/или аналога, его устойчивость к разрушению предпочтительно повышают, например, синтезом пептида с С-концевым карбоксамидом, и/или ацетилированием N-конца пептида для поддержания исходных свойств заряда молекулы. В одном из вариантов осуществления настоящего изобретения устойчивость к разрушению дополнительно повышают путем мутирования белка или иммуногенной части, производного и/или аналога по настоящему изобретению таким образом, что локальный процесс нарушения складывания указанного белка или его иммуногенной части, производного и/или аналога, чувствительных к автолизу по меньшей мере частично подавлен. Стратегии стабилизирующих мутаций известны и описаны, например, Matthews (1991), Alber (1991), Vriend, Eijsink (1993) и Fersht, Serrano (1993).
Секретированный белок выражают в качестве белка, который естественным образом вырабатывается в клетке и/или организме и по меньшей мере частично секретируются из указанных клеток и/или организма во внешнюю среду. Таким образом, при культивировании Streptococcus секретированный белок по меньшей мере частично, содержится по меньшей мере в части культуральной среды по меньшей мере в некоторых временных точках. Секретируемый белок необязательно должен вырабатываться и/или секретироваться постоянно. Секретированный белок может, например, вырабатываться и/или секретироваться во время определенной фазы жизненного цикла бактерии. Кроме того, выработка и секреция секретируемого белка не обязательно происходит в одно и то же время. Например, некоторые секретированные белки сначала аккумулируются внутри клетки и сектерируются позднее.
Ассоциированный с поверхностью белок определяют в качестве белка, который в норме формирует часть поверхности клетки, или который присоединен к поверхности клетки. Если указанный белок, ассоциированный с поверхностью клетки, присоединен к поверхности клетки, присоединение является или непосредственным, или непрямым. Непрямое присоединение, например, включает наличие по меньшей мере одного линкера.
Понятие «выделенный белок» относится к белку, который по меньшей мере частично выделен из природной среды, и/или к белку, который не содержит по меньшей мере части последовательности, имеющейся у природной формы.
Понятие «рекомбинантный белок» относится к белку, который вырабатывается выделенной и/или искусственной системой экспрессии, предпочтительно используя последовательность нуклеиновой кислоты, кодирующей указанный белок. Указанная последовательность нуклеиновой кислоты предпочтительно оперативно связано по меньшей мере с одной регуляторной последовательностью, например, промотором, энхансером и/или теминатором. Предпочтительно указанная регуляторная последовательность является индуцируемой, поэтому возможен контроль длительности экспрессии указанного белка. В одном из вариантов осуществления настоящего изобретения указанная последовательность нуклеиновой кислоты включает экзогенную последовательность нуклеиновой кислоты. Экзогенная последовательность нуклеиновой кислоты является последовательностью нуклеиновой кислоты, которая содержится в сайте генома организма, в котором указанная последовательность нуклеиновой кислоты в природном варианте отсутствует.
После идентификации способом по настоящему изобретению способности белка Streptococcus индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus, предпочтительно осуществляют его выработку. Выработанный белок, например, пригоден для выработки иммуногенной композиции и/или индукции иммунного ответа против по меньшей мере двух штаммов и/или серотипов Streptococcus в организме животного. Согласно указанному выше, различные способы выработки белка известны в данной области, например, выработка с применением рекомбинации. Настоящее изобретение, таким образом, предусматривает способ получения по меньшей мере одного белка, выявленного по способу настоящего изобретения. Белок Streptococcus, который способен индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus, получаемый по способу по настоящему изобретению, также описан в настоящем изобретении.
Белок Streptococcus и/или иммуногенная часть, производное и/или аналог по настоящему изобретению особенно применимы для приготовления иммуногенной композиции. Указанная иммуногенная композиция способна индуцировать широкий гуморальный и/или клеточный иммунный ответ против по меньшей мере, двух штаммов Streptococcus. Предпочтительно белок Streptococcus и/или его иммуногенная часть, производное и/или аналог, способные вызывать иммунный ответ против по меньшей мере двух серотипов Streptococcus, используют для приготовления иммуногенной композиции таким образом, что достигается широкий иммунный ответ против по меньшей мере, двух серотипов Streptococcus. Применение белка, который может быть получен по способу настоящего изобретения, или его иммуногенной части, производного и/или аналога для приготовления иммуногенной композиции, способной индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus, таким образом, также предусматривают наряду с иммуногенной композицией, способной индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus, включая по меньшей мере один выделенный и/или рекомбинантный белок, получаемый способом по настоящему изобретению, или его иммуногенную часть, производное и/или аналог. Для того, чтобы обеспечить более широкую защиту по меньшей мере два или несколько белков и/или иммуногенных частей, производных и/или аналогов по настоящему изобретению предпочтительно используют для приготовления иммуногенной композиции. В одном из вариантов осуществления настоящего изобретения комбинация по меньшей мере одного белка и по меньшей мере одной иммуногенной части, производного и/или аналога по настоящему изобретению, используют для приготовления иммуногенной композиции.
Помимо более широкой защиты, применение по меньшей мере двух белков и/или иммуногенных частей, производных и/или аналогов по настоящему изобретению снижает изменение «ускользнувших» выделенных мутантов бактерий Streptococcus. «Ускользнувшие» мутанты бактерий обычно возникают при стрессовых условиях внешней среды, например, в присутствии антибиотика и/или в присутствии антитела против эпитопа указанного организма. Благодаря природной вариабельности в популяции организма некоторые организмы избегают ингибирующего эффекта указанного стресса внешней среды, например, антибиотика и/или антитела, и способны к размножению. Изменение развития «ускользнувшего» мутанта для нескольких разных эпитопов одновременно меньше, чем изменение развития «ускользнувшего» мутанта только для одного эпитопа.
Таким образом, иммуногенная композиция по настоящему изобретению предпочтительно включает по меньшей мере два выделенных и/или рекомбинантных белка, и/или по меньшей мере одну их иммуногенную часть, производное и/или аналог, получаемые по способу настоящего изобретения. Для того, чтобы еще лучше избежать формирования «ускользнувших» мутантов, белок по настоящему изобретению предпочтительно представляет существенно важный белок. Это белок, который важен - предпочтительно существенно важен - для метаболизма, выживания и/или размножения бактерий Streptococcus. Таким образом, возможный «ускользнувший» мутант с измененным существенно важным белком менее жизнеспособен или нежизнеспособен.
Таблицы 5 и 6 включают перечень предпочтительных белков Streptococcus uberis, которые идентифицированы по способу настоящего изобретения. Эти белки, или по меньшей мере одна их иммуногенная часть, производное и/или аналог, применимы для приготовления иммуногенной композиции по настоящему изобретению. Применение по настоящему изобретению, в котором указанная белок выбран из табл.5 и/или табл.6, таким образом, также предусматривается, наряду с иммуногенной композицией по настоящему изобретению, включающей по меньшей мере один выделенный и/или рекомбинантный белок, представленный в табл.5 и/или табл.6, или его иммуногенную часть, производное и/или аналог. Чтобы обеспечить еще более широкую защиту, указанная иммуногенная композиция предпочтительно включает по меньшей мере два белка, представленных в табл.5 и/или табл.6, и/или их иммуногенные части, производные и/или аналоги. Наиболее предпочтительно указанная иммуногенная композиция включает по меньшей мере три белка, представленных в табл.5 и/или табл.6, и/или их иммуногенные части, производные и/или аналоги.
В предпочтительном варианте осуществления настоящего изобретения по меньшей мере один, по меньшей мере два или по меньшей мере три белка, представленные в табл.5 и/или табл.6, взяты из группы, включающей Р15, Р16, Р17, Р19, Р20, Р22, Р27, Р54, Р28, Р63, Р64, Р68, Р75, Р81, Р93, Р100, Р105, белок поверхностного исключения, триггерный фактор (ropA) и нуклеозиддифосфаткиназу. Эти белки либо распознаются антителом, имеющимся в сыворотке животных, инфицированных S. uberis, указывая на то, что эти белки экспрессируются in vivo и являются иммуногенными в организмах коров, либо обладают перекрестной реактивной способностью между по меньшей мере двумя штаммами S. uberis, представленными в табл.5. Нумерация описанных выше белков, например, в табл.5, относится к белкам, представленным, например, в табл.1, 2 и 3, которые показывают, но ими не ограничиваются, примеры обычных поверхностных белков S. uberis. Кроме того, фиг.4 показывает, но ими не ограничиваются, примеры последовательностей нуклеиновых кислот и аминокислот этих выбранных предполагаемых поверхностных белков/факторов вирулентности бактерий вида S. uberis.
Белки, которые высоко консервативны, экспрессируются in vivo и высоко иммуногенны, например белки, которые распознаются сыворотками в период выздоровления от коров, инфицированных разными штаммами согласно примеру 11, являются особенно применимыми в иммуногенных композициях по настоящему изобретению. В еще более предпочтительном варианте осуществления настоящего изобретения, таким образом, выбор белков из табл.5 и/или табл.6 включает белок, выбранный из группы, включающей Р15, Р16, Р20, Р27, Р54, Р28, Р63, Р68, Р93 и Р105. Наиболее предпочтительно выбор белков из табл.5 и/или табл.6 представляет белок, выбранный из группы, состоящей из Р15, Р16, Р54, Р28, Р63 и Р105. В примере 11 показано, что последующий отбор распознается всеми применяемыми конвалесцентными сыворотками, свидетельствуя, что эти антигены экспрессируются всеми штаммами S. uberis, которые вызывают соответствующую инфекцию, что эти антигены экспрессируются во время инфицирования организма хозяина и что эти антигены в высокой степени иммуногенны.
В еще одном из вариантов осуществления настоящего изобретения предусматривают иммуногенную композицию, способную индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus, включающих по меньшей мере одну молекулу нуклеиновой кислоты, кодирующую по меньшей мере один белок, получаемый по способу настоящего изобретения, или иммуногенную часть, производное и/или аналог указанного белка. При введении указанной иммуногенной композиции животному указанная молекула нуклеиновой кислоты экспрессируется в организме животного, приводя к экспрессии по меньшей мере одного белка и/или иммуногенной части, производного и/или аналога по настоящему изобретению. Выработка и, необязательно, внеклеточная экскреция указанного белка и/или иммуногенной части, производного и/или аналога приводит к иммунному ответу.
В одном из вариантов осуществления настоящего изобретения белок по настоящему изобретению и/или его иммуногенная часть, производное и/или аналог получают путем рекомбинации. Настоящее изобретение предусматривает способ выработки иммуногенной композиции, способной индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus, причем указанный способ включает введение клеток или другой системы экспрессии по меньшей мере с одним рекомбинантным вектором, причем указанный по меньшей мере один вектор, включает последовательность нуклеиновой кислоты, кодирующую по меньшей мере один белок, получаемый способом по настоящему изобретению, и/или по меньшей мере один белок, выбранный из табл.5 и/или табл.6, и/или иммуногенную часть, производное и/или аналог указанного белка. Соответствующие векторы экспрессии известны в данной области. В одном из вариантов осуществления настоящего изобретения экспрессируется по меньшей мере одна последовательность нуклеиновой кислоты, кодирующая один белок по настоящему изобретению или его иммуногенную часть. В другом варианте осуществления настоящего изобретения используют по меньшей мере одну молекулу нуклеиновой кислоты, кодирующую по меньшей мере два белка и/или иммуногенные части. Также возможно применение по меньшей мере двух молекул нуклеиновой кислоты, каждая из которых кодирует один или несколько белков и/или иммуногенных частей по настоящему изобретению, и др. Например, можно использовать одну молекулу, кодирующую (по меньшей мере) один белок, и одну молекулу нуклеиновой кислоты, кодирующую (по меньшей мере) одну иммуногенную часть. Таким образом, возможны вариации числа молекул нуклеиновой кислоты и числа белков и/или иммуногенных частей, кодируемых указанными молекулами нуклеиновой кислоты.
Последовательность нуклеиновой кислоты по настоящему изобретению инсертирована, например, в геном клетки гомологичной рекомбинацией. Также возможно инсертировать последовательность нуклеиновой кислоты случайным образом, например, электропорацией. В другом варианте или дополнительно, указанная последовательность нуклеиновой кислоты помещается в вектор, например, плазмидный вектор или фаговый вектор, который стабилен в выбранной системе экспрессии, представляющей микроорганизм и/или клетку. Указанная последовательность нуклеиновой кислоты по настоящему изобретению предпочтительно транскрибируется и транслируется под контролем регуляторной последовательности, например промотора, энхансера и/или терминатора. Предпочтительно указанный промотор, энхансер и/или терминатор применим для использования в выбранной системе экспрессии. Более предпочтительно указанная регуляторная последовательность индуцируема для того, чтобы допустить контролируемую экспрессию. Промоторы и терминаторы, применимые для различных микроорганизмов, описаны в кн.: Biseibutsugaku Kisokoza (Basic Microbiology), 1990, т.8, Genetic Technology, Kyoritsu Shuppan. Например, допустимыми плазмидными векторами для Escherichia, точнее, для Escherichia coli, являются плазмиды серий pBR и pUC, а также допустимы промоторы, например, промотор lac (β-галактозидазы), оперон trp (триптофановый оперон) и промотор tac (гибридный промотор lac-trp) и промоторы, производные от λ-faag PL или PR. К предпочтительным терминаторам относятся trpA- или производный от фага rrnB рибосомальный терминатор. Плазмидные векторы, применимые для рекомбинантной выработки у Streptococcus, включают, например, pHV1301 (FEMS Microbiol. Lett. 26, 1985, с.239), pGK1 (Appl. Environ. Microbiol. 50, 1985, с.94).
Настоящее изобретение также предусматривает молекулу рекомбинантной нуклеиновой кислоты, включающую последовательность нуклеиновой кислоты, кодирующей по меньшей мере два белка Streptococcus, которые могут быть получены способом по настоящему изобретению, и/или выбраны из табл.5 и/или табл.6, и/или иммуногенную часть по меньшей мере одного из указанных белков, под контролем функционально связанной регуляторной последовательностью, например, промотора. Выделенные клетки-хозяева, включающие последовательность нуклеиновой кислоты, которые кодируют по меньшей мере два белка, получаемые способом по настоящему изобретению и/или выбранные из табл.5 и/или табл.6, и/или их иммуногенную часть, также предусмотрены в настоящем изобретении. Указанные клетки-хозяева предпочтительно представляют прокариотические клетки-хозяева.
В предпочтительном варианте осуществления настоящего изобретения молекулу нуклеиновой кислоты по настоящему изобретению используют для индукции иммунного ответа против Streptococcus. Индукцию предпочтительно осуществляют с рекомбиантным носителем, включающим нуклеиновую кислоту, кодирующую по меньшей мере один белок, получаемый способом по настоящему изобретению, и/или выбранный из табл.5 и/или табл.6, и/или иммуногенную часть указанного по меньшей мере одного белка, или рекомбинантную молекулу нуклеиновой кислоты по настоящему изобретению. Указанный рекомбинантный носитель, таким образом, также предусмотрен в настоящем изобретении. Наиболее предпочтительно также предусмотрен рекомбинантный носитель, включающий нуклеиновую кислоту, кодирующую по меньшей мере один белок, выбранный из табл.5 и/или табл.6. В одном особенно предпочтительном варианте осуществления настоящего изобретения указанный рекомбинантный носитель включает нуклеиновую кислоту, кодирующую по меньшей мере два белка, выбранных из табл.5 и/или табл.6. В одном из вариантов осуществления настоящего изобретения допускается выработка указанного рекомбинантного носителя по меньшей мере один белок по настоящему изобретению, после чего по меньшей мере один рекомбинантный белок и сам носитель используют для индукции иммунного ответа против по меньшей мере двух штаммов и/или серотипов Streptococcus. В другом варианте осуществления настоящего изобретения предусмотрен неживой рекомбинантный носитель по настоящему изобретению. В еще одном из вариантов осуществления настоящего изобретения также предусматривается живой рекомбинантный носитель по настоящему изобретению. В другом варианте осуществления настоящего изобретения указанный живой носитель является аттенуированным носителем. Живой носитель по настоящему изобретению предпочтительно способен инфицировать человека и/или животное, после чего индуцируется иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus.
Рекомбинантный носитель по настоящему изобретению предпочтительно представляет виды Streptococcus. При этом иммунный ответ, направленный против Streptococcus, индуцируется и белком (белками), и/или иммуногенной частью (частями), производным (производными), и/или аналогом (аналогами), кодируемыми указанным носителем, и самим указанным рекомбинантным носителем. Продукты экспрессии капсулярного гена Streptococcus часто являются высоко иммуногенными и серотип-специфичными. Таким образом, наличие продуктов экспрессии капсулярного гена препятствует индукции иммунного ответа, направленного против различных штаммов и/или серотипов Streptococcus. В одном из вариантов осуществления настоящего изобретения, таким образом, если рекомбинантный носитель по настоящему изобретению включает Streptococcus, указанный Streptococcus утрачивает по меньшей мере часть продукта экспрессии капсулярного гена. В одном из вариантов осуществления настоящего изобретения указанная бактерия Streptococcus относится к некапсулярным стрептококкам.
Согласно описанному выше, иммунизация по меньшей мере двумя белками и/или иммуногенными частями, производными и/или аналогами, производными по меньшей мере от двух разных штаммов и/или серотипов Streptococcus, предусматривает широкую защиту и минимизацию изменения формирования «ускользнувших» мутантов. Предпочтительный вариант осуществления настоящего изобретения, таким образом, предусматривает рекомбинантный носитель по настоящему изобретению, включающий последовательность нуклеиновой кислоты, кодирующую по меньшей мере один белок и/или его иммуногенная часть, производную от первого штамма и/или серотипа Streptococcus, и последовательность нуклеиновой кислоты, кодирующую по меньшей мере один белок и/или его иммуногенную часть, производную от второго штамма и/или серотипа Streptococcus. Указанный рекомбинантный носитель предпочтительно включает живой рекомбинантный носитель.
Рекомбинантный носитель, например, вырабатывается в соответствующих клетках-хозяевах. Таким образом, клетки-хозяева, включающие рекомбинантный носитель по настоящему изобретению, также предусмотрены.
Рекомбинантный носитель по настоящему изобретению применим для выработки иммуногенной композиции, способной индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus. Таким образом, в настоящем изобретении также предусмотрена иммуногенная композиция, способная индуцировать иммунный ответ против Streptococcus, причем также предусмотрена указанная композиция, включающая рекомбинантный носитель по настоящему изобретению.
После введения иммуногенной композиции по настоящему изобретению человеку и/или животному, индуцируется иммунный ответ против Streptococcus. Указанный иммунный ответ предпочтительно способен по меньшей мере частично, противодействовать заболеванию, связанному с бактериями рода Streptococcus. Таким образом, в настоящем изобретении также предусмотрена иммуногенная композиция по настоящему изобретению для применения в качестве лекарственного средства, а также применение иммуногенной композиции по настоящему изобретению для приготовления лекарственного средства против заболевания, связанного с бактериями рода Streptococcus.
Иммуногенная композиция по настоящему изобретению также применима для выработки вакцины. Указанная вакцина предпочтительно способна по меньшей мере частично, обеспечивает защиту от заболевания, связанного с бактерией рода Streptococcus. Предпочтительно, указанная вакцина способна обеспечить защиту от инфекции Streptococcus. Настоящее изобретение, таким образом, предусматривает применение иммуногенной композиции по настоящему изобретению для приготовления вакцины.
Белок, иммуногенная часть, производное, аналог и/или рекомбинантный носитель по настоящему изобретению предпочтительно вводят конкретному человеку и/или животному вместе с соответствующим носителем. Указанный носитель предпочтительно облегчает восприятие указанным конкретным человеком и/или животным указанного белка, иммунногенной части, производного, аналога и/или рекомбинантного носителя по настоящему изобретению и предпочтительно повышает иммуногенный эффект. Соответствующий носитель по настоящему изобретению, например, включает соответствующий адъювант, способный повышать иммунизирующий эффект иммуногенной композиции по настоящему изобретению. Многие соответствующие адъюванты на основе масла и воды известны специалистам в данной области. В одном из вариантов осуществления настоящего изобретения указанным адъювантом является Diluvac Forte и/или Specol. В другом варианте осуществления настоящего изобретения указанный соответствующий носитель представляет раствор, например, физиологический раствор, например, для разведения белков или его иммуногенных частей, производных и/или аналогов. Таким образом, настоящее изобретение также описывает иммуногенную композицию по настоящему изобретению, включающую по меньшей мере один белок, иммуногенную часть, производное, аналог и/или рекомбинантный носитель по настоящему изобретению и соответствующий носитель.
Иммуногенная композиция по настоящему изобретению способна вызвать иммунный ответ против Streptococcus у конкретного человека и/или животного и, тем самым, она снижает и/или уничтожает ряд бактерий Streptococcus у указанного конкретного человека и/или животного. Настоящее изобретение, таким образом, предусматривает способ снижения и/или уничтожения ряда бактерий Streptococcus у человека и/или животного, включающий получение указанным индивидуумом и/или животным иммуногенной композиции по настоящему изобретению.
Иммуногенная композиция по настоящему изобретению предпочтительно способна по меньшей мере частично, препятствовать развитию заболевания и/или предупреждать заболевание, связанное с бактериями рода Streptococcus. Если заболевание, связанное с бактериями рода Streptococcus, уже возникло, иммуногенная композиция по настоящему изобретению предпочтительно способна по меньшей мере частично, противодействовать указанному заболеванию. Фармацевтическая композиция, представляющая иммуногенную композицию по настоящему изобретению и, предпочтительно, соответствующий носитель, например, адъюванты Diluvac Forte и/или Specol, таким образом, также предусмотрена в настоящем изобретении.
В другом варианте осуществления настоящего изобретения предусматривают способ измерения иммунитета конкретного человека и/или животного против Streptococcus, который включает определение по меньшей мере в одном образце указанного индивидуума и/или животного, наличия антитела и/или иммунных клеток, направленных против белка, получаемого способом по настоящему изобретению, и/или выбранного из табл.5 и/или табл.6, или его иммуногенной части. Диагностический набор, включающий по меньшей мере один белок, получаемый способом по настоящему изобретению, и/или выделенный из табл.5 и/или табл.6, или его иммуногенную часть, и средства выявления связывания антитела и/или связывания иммунных клеток с указанным белком или его иммуногенной частью, также представлены в настоящем изобретении. В предпочтительном варианте осуществления настоящего изобретения указанный диагностический набор включает по меньшей мере два белка, выбранных из табл.5 и/или табл.6.
Подробное описание изобретения
Способ по настоящему изобретению является предпочтительным вариантом осуществления настоящего изобретения, применимым для идентификации белка Streptococcus uberis, способного вызвать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis. Указанный белок Streptococcus uberis предпочтительно используют для приготовления иммуногенной композиции, способной вызвать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis. Иммуногенная композиция по настоящему изобретению, способная вызвать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, включающая по меньшей мере один, предпочтительно по меньшей мере два, выделенных и/или рекомбинантных белка, получаемых способом по настоящему изобретению, или по меньшей мере одну его иммуногенную часть, производное и/или аналог, таким образом, также предусмотрены в настоящем изобретении, а также предусмотрены его применения для приготовления лекарственного средства против мастита, вызываемого Streptococcus uberis. Настоящее изобретение также предусматривает выделенную или рекомбинантную молекулу нуклеиновой кислоты, включающую последовательность нуклеиновой кислоты, кодирующей по меньшей мере два белка Streptococcus uberis, получаемых способом по настоящему изобретению, и/или выбранных из табл.5 и/или табл.6. Также предусмотрены рекомбинантные носители, клетки-хозяева и иммуногенные композиции, включающие указанную нуклеиновую кислоту, а также их применение.
Бактерия Streptococcus uberis ассоциирована с маститом у коров. Мастит у коров представляет инфекцию молочной железы, обычно вызванную бактериями. Воспалительный ответ после инфицирования приводит к снижению продуктивности и качества молока и вызывает значительные ежегодные экономические потери при производстве молока. Экономическая потеря в Нидерландах составляет примерно 100 евро на корову в год.
Среди видов бактерий большинство видов, обычно ассоциированных с маститом, являются разными видами рода Streptococcus, включая Streptococcus uberis (нетипируемый вид). Streptococcus agalactiae (группа В по классификации Лэнсфилда), Streptococcus dysgalactiae (группа С по классификации Лэнсфилда), Streptococcus zooepidemicus и стрептококки групп D, G, L и N по классификации Лэнсфилда. Некоторые из этих видов являются заразными (например, S. agalactiae), хотя другие представляют патогены, распространенные в окружающей среде (например, S. dysgalactiae и S. uberis).
Мастит, возникающий в результате заражения S. uberis, обычно протекает бессимптомно, отличаясь внешне нормальным молоком с повышенным количеством соматических клеток из-за притока лейкоцитов.
Мастит может быть разным по тяжести, исходя из клинических проявлений, вызванных инфекцией. Умеренная форма мастита может несколько повысить температуру тела и/или повысить температуру вымени. В более тяжелых случаях мастит, вызываемый S. uberis, может также протекать в острой клинической форме с очевидными признаками заболевания, например, сгустками в молоке или с изменением цвета молока, набухания или уплотнения молочной железы. Некоторые случаи клинического заболевания могут быть тяжелыми, и может наблюдаться гипертермия. Обзор клинических проявлений мастита, вызванного S. uberis, см. в работах Bramley (1991) и Schalm и др. (1971). Традиционные способы уничтожения бактериальных инфекций, например, пальпирование соска и лечение антибиотиками, эффективны для уничтожения многих типов контагиозных маститов, но бактерии, в норме присутствующие в окружающей среде и обычно связанные со всеми молочными производствами, часто устойчивы к таким способам. Следовательно, указанные способы не эффективны в случае мастита, вызванного патогенами внешней среды, например, Streptococcus uberis и Escherichia coli, которые в настоящее время ответственны за более чем 95% случаев мастита. Из этих двух видов S. uberis является наиболее важным патогеном, поступающим из окружающей среды, что следует из исследований, проведенных в Великобритании (Hillerton и др., 1993), в Новой Зеландии (McDougall, 1998), в США (Hogan и др., 1989) и в Нидерландах (Animal Health Service, 2000). Также установлено, что S. uberis, если инфекция происходит из внешней среды, может непосредственно распространиться от инфицированной коровы другому чувствительному животному (Neave и др., 1969, Oliver и др., 1999, Zadoks и др., 2001). Известно несколько штаммов S. uberis, которые отличаются по вирулентности и антигенности.
Несостоятельность современных способов, касающихся уничтожения возбудителя мастита S. uberis, привела к поиску иных средств борьбы с инфекцией, например, более эффективных вакцин. До настоящего времени было получено и исследовано на коровах несколько типов вакцин.
Повторная иммунизация молочного скота убитыми целыми бактериями приводит к снижению числа бактерий, содержащихся в молоке после экспериментального заражения тем же штаммом (Leigh, 1999; Leigh, 2000). Однако убитая вакцина не предупреждает ни инфицирование, ни воспалительного ответа в молочной железе, и не влияет на заболеваемость маститом, вызываемым S. uberis, в поле (Leigh, 1999). Таким образом, был сделан вывод, что иммунизация убитыми бактериями не решает проблемы мастита, вызванного S. uberis. Иммунизация живыми бактериями S. uberis вызывает частичную защиту против экспериментального заражения тем же (или гомологичным) штаммом (Finch и др., 1997). Защита достигалась в отсутствии опсонизирующей активности и без большого сосредоточения нейтрофилов. Однако представляется, что вакцина не защищает от других штаммов S. uberis. Относительно небольшая польза от применения вакцин из таких целых клеток показывает, что трудно защитить животных против S. uberis, используя обычные вакцины из целых бактерий.
Позднее была получена субъединичная вакцина, основанная на одном белке S. uberis (Fontaine и др. 2002). Публикация указанной субъединичной вакцины по настоящее время не привела к последующему развитию, в результате было сделано заключение, что изменения в обнаружении единственного белка, который может защитить животное от нескольких типов S. uberis, незначительны, и субъединичные вакцины этого типа обычно не соответствуют решению проблемы борьбы с маститом, вызываемым S. uberis.
Таким образом, мастит, вызванный S. uberis, эффективно не предупреждается или не вылечивается вакцинацией целыми живыми или убитыми бактериями или субъюнитной вакциной, включающей один белок.
Несмотря на описанные выше неудачи вакцинации против мастита, вызываемого S. uberis, в настоящем изобретении описывают успешное предупреждение и/или ослабление проявления заболеваний мастита, вызванных различными штаммами S. uberis, за счет применения антигенной композиции, способной индуцировать иммунный ответ против S. uberis по настоящему изобретению.
Настоящее изобретение предусматривает способ идентификации белка Streptococcus uberis, способного индуцировать иммунный ответ против по меньшей мере двух штаммов и/или типов Streptococcus uberis, включающий:
а) идентификацию по меньшей мере, части секретированного белка, ассоциированного с поверхностью белка и/или белка, последовательность которого по меньшей мере на 50% идентична последовательности бактериального фактора вирулентности;
б) выбор по меньшей мере одного белка, выявленного на стадии а), который сохраняется по меньшей мере в двух штаммах и/или типах Streptococcus uberis; и
в) определение, может ли по меньшей мере один белок, выбранный на стадии б), или его иммуногенная часть, производное и/или аналог, специфически связываться с антигеном, и/или иммунными клетками животного, инфицированного первым штаммом и/или типом Streptococcus uberis, и антителом и/или иммунными клетками животного, инфицированного вторым штаммом и/или типом Streptococcus uberis. Предпочтительно, указанный белок, последовательность которого по меньшей мере на 50%, по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, наиболее предпочтительно по меньшей мере на 80% идентична последовательности бактериального фактора вирулентности.
Настоящее изобретение, кроме того, представляет комбинацию по меньшей мере двух выделенных или рекомбинантных поверхностных белков S. uberis или их иммуногенных частей в антигенной композиции, которая существенно повышает иммунный ответ против штаммов S. uberis. Хотя вакцины из целых бактериальных клеток, включающих много бактериальных иммуногенных белков, не вызывают широкой защиты против разных штаммов S. uberis, два или несколько белков или их иммуногенных частей в иммуногенной композиции по настоящему изобретению обладают требуемым эффектом повышения иммунного ответа против S. uberis. В настоящем изобретении установлено, что выбор по меньшей мере двух иммуногенных белков или их иммуногенной части бактерии S. uberis, и предпочтительно по меньшей мере двух штаммов или типов бактерий S. uberis, и комбинирование указанных по меньшей мере двух иммуногенных белков или их иммуногенной части в иммуногенной композиции повышает иммунитет против разных штаммов S. uberis, поскольку иммунный ответ желателен против широкого круга бактерий S. uberis.
Для индукции иммунного ответа у человека или животного предпочтительно иммуногенная часть белка поступает указанному человеку или животному. В настоящем изобретении понятие «иммуногенный сайт» используют взаимозаменяемо с понятием «иммуногенная часть». Понятие «иммуногенный сайт (иммуногенная часть)» означает часть белка, способную индуцировать иммунный ответ у субъекта. Предпочтительно указанная иммуногенная часть белка включает один или несколько эпитопов и соответственно индуцирует иммунный ответ. Иммуногенная часть включает по меньшей мере 5 аминокислот, предпочтительно по меньшей мере 10-15, и наиболее предпочтительно 25 или нескольких последовательно расположенных аминокислот. Таким образом, настоящее изобретение в другом варианте своего осуществления предусматривает белок или его иммуногенную часть, включающую по меньшей мере отрезок из 30 последовательно расположенных аминокислот белковой молекулы, кодируемой нуклеиновой кислотой по настоящему изобретению. Конформационный эпитоп обычно формируется несколькими отрезками последовательно расположенных аминокислот, которые складываются и вместе формируют эпитоп, когда белок принимает свойственную ему трехмерную структуру. Настоящее изобретение также описывает применение конформационных эпитопов в качестве иммуногенных частей.
Производное белка означает белок, обладающий тем же типом иммуногенных свойств, но не обязательно в том же количестве. Специалист в данной области способен изменить белок таким образом, что иммуногенные свойства указанной молекулы в существенной степени того же типа, но не обязательно в том же количестве. Производное белка может быть получено многими способами, например, путем консервативного замещения аминокислот, например, замещением одной аминокислоты в белке на другую аминокислоту. При традиционном заместительном картировании предпочтительно производят, например, замещение аминокислот, включающих кислую боковую цепочку, на другую аминокислоту, включающую кислую боковую цепочку, объемную аминокислоту на другую объемную аминокислоту, аминокислоту, включающую основную боковую цепочку, на аминокислоты, включающие основную боковую цепочку, аминокислоты, включающие незаряженную полярную боковую цепочку на аминокислоты, включающие незаряженные полярные боковые цепочки, и аминокислоты, включающие неполярную боковую цепочку на аминокислоты, включающие неполярную боковую цепочку. Специалист в данной области способен вырабатывать аналогичные белковые соединения. Например, путем скрининга пептидной библиотеки или по программе изменения пептидов. При использовании в качестве иммуногена пептид синтезируют с улучшенными свойствами, чтобы повысить высокую вероятность успешной выработки антитела. К ним относится С-концевая свободная карбоксильная группа, если пептид представляет С-концевую последовательность нативного белка, и свободная N-концевая аминогруппа, если пептид представляет реальную N-концевую последовательность нативного белка. Такой аналог обладает в существенной степени теми же иммуногенными свойствами указанного белка, но не обязательно в том же количестве.
Белок или пептид является объектом для разрушения рядом разнообразных средств, например, протеолизом, нарушением складчатости, предельными величинами pH, детергентами и высокими концентрациями солей. Для продления срока сохранности рекомбинантного белка или пептида, указанный белок или пептид делают более стабильным для противостояния разрушению, например, путем синтеза указанного пептида с С-концевым карбоксамидом и/или ацетилирования N-конца для поддержания свойств нативного заряда. Этого также можно достичь за счет мутаций, используя стратегию стабилизирующих мутаций для подавления процессов местного развертывания, которые обычно делают белок чувствительным к автолизу. Указанная стратегия стабилизирующей мутации основана на обычно принятых принципах структуры белка и стабильности, согласно описанию, например, Matthews (1991), Alber (1989), Vriend и Eijsink (1993), Fersht и Serrano (1993).
В одном из вариантов осуществления настоящего изобретения иммуногенная композиция по настоящему изобретению включает композицию, содержащую по меньшей мере два рекомбинантных или выделенных поверхностных белка, или их производные, или аналоги, и/или иммуногенные части, причем введение композиции человеку или животному, предпочтительно корове, приводит к развитию гуморального и/или клеточного иммунного ответа в отношении указанных поверхностных белков или их иммуногенных частей.
Иммуногенный ответ включает формирование гуморального и/или клеточного иммунного ответа, направленного против указанного белка или его иммуногенной части у человека или животного, предпочтительно коровы. Гуморальный иммунный ответ приводит к выработке антитела у человека или животного, поскольку клеточный иммунный ответ преимущественно повышает формирование реактивных иммунных клеток. В целом обе составляющие иммунного ответа индуцируются введением иммуногенного белка или его части. Предпочтительный иммунный ответ против S. uberis заключается в выработке антител. Предпочтительно указанный иммунный ответ предупреждает и/или снижает проявление мастита, и/или снижает число бактерий S. uberis в вымени. Настоящее изобретение описывает способы отбора и выработки белков и эпитопов для индукции указанного ответа в виде антител. Другим предпочтительным иммунным ответом против S. uberis является клеточный иммунный ответ. Настоящее изобретение также описывает способы отбора эпитопов Т-клеток поверхностных белков и выработки эпитопов Т-клеток, вызывающих повышенную Т-клеточную реактивность, например, путем соединения составных предварительно выбранных эпитопов Т-клеток по типу цепочки бус (string-of bead fashion), например, согласно описанию Van der Burg и др. (WO 97/41440).
В одном из вариантов осуществления настоящего изобретения указанная иммуногенная композиция способна снижать длительность и/или тяжесть инфекции и/или повышать устойчивость животных к инфицированию бактерией S. uberis.
Настоящее изобретение описывает иммунный ответ, направленный против внешней поверхности S. uberis, являющийся предпочтительным. Таким образом, настоящее изобретение описывает иммуногенную композицию или ее иммуногенную часть, которая способна индуцировать иммунный ответ на антигены, предпочтительно расположенные на поверхности клеток или около поверхности клеток S. uberis. К поверхностным белкам по настоящему изобретению относятся белки, которые в естественном состоянии предпочтительно находятся около или на поверхности бактерий S. uberis, и/или белков, которые в естественном состоянии предпочтительно вырабатываются и/или экстрагируются бактериями S. uberis внеклеточно. Указанные поверхностные белки предпочтительно имеют гомологичные белки в других штаммах S. uberis. Таким образом, иммунный ответ индуцируется иммуногенными белками или их частями, производными от одного штамма S. uberis, и он также эффективен против других штаммов S. uberis. Таким образом, настоящее изобретение описывает иммуногенную композицию, способную индуцировать иммунный ответ против S. uberis, причем указанная композиция включает по меньшей мере два рекомбинантный и/или выделенных поверхностных белка, производных от Streptococcus uberis, и/или иммуногенные части одного или обоих указанных белков.
Понятие «рекомбинантный белок» относится к белку, получаемому методами рекомбинации ДНК, т.е. вырабатываемому клетками, трансформированными конструкцией нуклеиновой кислоты, кодирующей требуемый белок. Указанная конструкция нуклеиновой кислоты является примером конструкции рекомбинантной ДНК с регуляторной последовательностью, например, промоторной последовательностью, и/или терминирующей последовательностью, и/или энхансерной последовательностью, которая контролирует экспрессирующую последовательность.
Понятие «выделенный белок» относится к белку, выделенному и очищенному из целого организма, в составе которого молекула обнаружена в природе; и/или белку, не содержащему, полностью или частично, веществ, которые в норме сопутствуют белку в природе. Указанная иммуногенная композиция включает или по меньшей мере два белка, или их иммуногенные части, производные от той же бактерии S. uberis, или она включает по меньшей мере один белок или его иммуногенную часть от одного типа бактерий S. uberis и по меньшей мере один белок или его иммуногенную часть от бактерии S. uberis другого типа. Настоящее изобретение также описывает комбинацию по меньшей мере 3, или 4, или большего количества белков или их иммуногенных частей, из которых один, или два или более производны от других типов S. uberis.
Предпочтительно иммуногенная композиция или ее иммуногенная часть по настоящему изобретению включает белки по меньшей мере двух разных организмов S. uberis, поскольку получаемый широкий иммунный ответ обеспечивает перекрестную защиту, т.е. направленный против разных типов S. uberis. Кроме того, применение иммуногенных белков или их иммуногенных частей по меньшей мере двух типов штаммов S. uberis, снижает изменения в развитии «ускользнувших» мутантов бактерий S. uberis. Выщепление мутантных бактерий обычно происходит в стрессовых условиях внешней среды, например, в присутствии антибиотика, или в присутствии антитела против эпитопа указанного организма. За счет природных вариаций, например, вызванных низкой частотой мутирования в популяции бактерий, у некоторых бактерий указанной популяции репликация подавляется больше указанными антителами, чем у других, которые «ускользнули» от ингибирующего действия присутствующего указанного антитела и сохраняют возможность размножаться, тем самым, занимая преобладающее положение в новой популяции. Изменение развития «ускользнувшего» мутанта по нескольким разным эпитопам одновременно меньше, чем изменение развития «ускользнувшего» мутанта только для одного эпитопа. Иммуногенная композиция и/или ее иммуногенная часть предпочтительно индуцирует иммунный ответ против по меньшей мере двух белков, предпочтительно вызывая широкую защиту против инфекции и снижения клинических признаков мастита. Таким образом, настоящее применение предусматривает иммуногенную композицию, способную индуцировать иммунный ответ против Streptococcus uberis, включающую по меньшей мере два рекомбинантных и/или выделенных поверхностных белка, производных по меньшей мере от одного штамма Streptococcus uberis, и/или иммуногенную часть, или аналог, или производное одного или обоих указанных белков.
Белки, важные для метаболизма, или выживания, или размножения бактерии, обычно известны в качестве значимых для бактерии белков. Последовательность и функция указанных значимых белков обычно предпочтительно сохраняется у разных типов S. uberis. В предпочтительном варианте осуществления настоящего изобретения указанные иммуногенные белки являются значимыми белками S. uberis. Таким образом, иммунный ответ направлен против существенно важного белка или его иммуногенной части, тем самым, формируя защиту против гомологичной бактерии S. uberis, но, также обеспечивая перекрестную защиту против разных типов S. uberis, поскольку указанный сохраненный белок или значимый белок также присутствует на поверхности других типов S. uberis. Таким образом, применение значимых поверхностных белков бактерии S. uberis в качестве иммуногенного белка по настоящему изобретению повышает эффективность защиты иммунного ответа против инфекции разными типами бактерий S. uberis и снижает способность указанных организмов избежать иммунного ответа.
Капсулярные антигены S. uberis обычно являются хорошими иммуногенными эпитопами, поскольку капсулярные антигены легко выявляются сывороткой выздоравливающих коров (которые перенесли мастит, вызванный S. uberis). Указанные иммуногенные свойства способны повышать иммунный ответ против родственных иммуногенных эпитопов S. uberis. Таким образом, в другом варианте осуществления настоящего изобретения иммуногенная композиция включает по меньшей мере один капсулярный антиген дополнительно к иммуногенным белкам, поскольку указанный капсулярный антиген повышает иммунный ответ против указанной иммуногенной композиции.
В настоящей заявке на патент в табл.5 и табл.6 представлены предпочтительные рекомбинантные и или выделенные поверхностные белки, производные от S. uberis и выбранные по присущей им способности индуцировать иммунный ответ против разных штаммов S. uberis. Таким образом, применение настоящего изобретения предусматривает иммуногенную композицию по настоящему изобретению, и/или an иммуногенную часть, или аналог, или производное одного или обоих указанных белков, в которой по меньшей мере два белка, выбранные из табл.5 и/или табл.6.
В предпочтительном варианте осуществления настоящего изобретения выбор производят из табл.5 и/или табл.6, и комбинацию получают из двух или нескольких белков, например, белка №63 и/или его иммуногенной части из штамма S. uberis 0140J, вместе с белком №15 или 22, и/или обоими, и/или их иммуногенной частью из штамма S. uberis 41-241. Такой выбор предусматривает белки или их иммуногенные части из двух разных штаммов S. uberis, тем самым, обеспечивая широкую защиту для нескольких штаммов S. uberis.
В предпочтительном варианте осуществления настоящего изобретения выбор белков из табл.5 и/или табл.6 включает белок, выбранный из группы, включающей Р15, Р16, Р17, Р19, Р20, Р22, Р27, Р54, Р28, Р63, Р64, Р68, Р75, Р81, Р93, Р100 и Р105. Согласно указанному ранее, эти белки или распознаются антителами, имеющимися в сыворотке животных, инфицированных S. uberis, показывая, что эти белки экспрессируются in vivo и иммуногены в организмах коров, или перекрестие взаимодействуют по меньшей мере между двумя штаммами S. uberis, представленными в табл.5.
Белки, выявленные в примере пример 11, особенно применимы для индукции иммунного ответа. В еще более предпочтительном варианте осуществления настоящего изобретения, таким образом, выбор белков из табл.5 и/или табл.6 включает белок, выбранный из группы, состоящей из Р15, Р16, Р20, Р27, Р54, Р28, Р63, Р68, Р93, и Р105. Наиболее предпочтительно выбор белков из табл.5 и/или табл.6 включает белок, выбранный из группы, состоящей из Р15, Р16, Р54, Р28, Р63 и Р105. Согласно указанному выше, последующая селекция белков представляется всеми штаммами S. uberis, которые вызывают соответствующую инфекцию в примере 11, экспрессированными во время инфицирования организма хозяина, и в высокой степени иммуногенными. Приведенная выше нумерация белков, представленных, например в табл.5, относится к белкам, представленным, например, в табл.1, 2 и 3, которые показывают неограничительные примеры распространенных поверхностных белков S. uberis. Кроме того, фиг.4 показывает неограничительные примеры последовательностей нуклеиновых кислот и аминокислот таких выбранных предполагаемых поверхностных белков/факторов вирулентности S. uberis.
Небольшое количество бактерий в молоке или на/в вымени коров часто обнаруживают в полевых условиях и они не вредны для животного. Мастит может развиться, если число бактерий, например, бактерий S. uberis, возрастает в молоке или в вымени. Иммунный ответ, индуцированный белками или их иммуногенными частями по настоящему изобретению, предпочтительно эффективен в подавлении по меньшей мере частичном, бактериального роста бактерий Streptococcus uberis в вымени. Снижение количества бактерий S. uberis в среде, непосредственно окружающей животное, также способствует предупреждению мастита. Настоящее изобретение описывает предупреждение и/или понижение проявления мастита, вызываемого S. uberis, путем иммунизации коров, тем самым, сохраняя число бактерий S. uberis низким. Указанный низкий уровень бактерий S. uberis далее сохраняется низким при применении гигиенического режима при дойке, согласно которому следует, например, мыть вымя, соски и всю аппаратуру, которая приходит в контакт с выменем и/или сосками.
Рекомбинантные и/или выделенные поверхностные белки, производные бактерии S. uberis, получаемые по настоящему изобретению, в одном из вариантов его осуществления получают с помощью системы выработки, используя прокариотические клетки или эукариотические клетки. Примерами клеток с хорошо сформированными системами хозяина/вектора для выработки рекомбинантного белка являются, например, бактерии: Escherichia, Bacillus, Pseudomonas, Serratia, Brevibacterium, Corynebacterium, Streptococcus и Lactobacillus, дрожжи: Saccharomyces, Kluyveromyces, Schizosaccharomyces, Zygosaccharomyces, Yarrowia, Trichosporon, Rhodosporidium, Hansenula, Pichia и Candida, и грибы: Neurospora, Aspergillus, Cephalosporium и Trichoderma.
Для получения рекомбинантного исследуемого белка ген, кодирующий указанный белок или его часть, либо интегрирован в геном, например, путем гомологичной рекомбинации или случайно, либо указанный ген помещен в плазмидный вектор или в фаговый вектор, который стабильно поддерживается и экспрессируется в выбранных микроорганизмах или клетках. Для экспрессии выбранной конструкции ДНК в микроорганизме или клетках ген транскрибируют и транслируют под контролем промотора и терминатора. Предпочтительно указанный промотор и терминатор применимы для выбранных микроорганизмов. Промоторы и терминаторы, применимые для разных микроорганизмов, описаны в «Biseibutsugaku Kisokoza (Basic Microbiology)», 1990, т.8, Genetic Technology, Kyoritsu Shuppan (1990)», а предпочтительные для дрожжей описаны в Adv. Biochem. Eng. 43, 1990, cc.75-102 и в Yeast 8, 1992, cc.423-488. Например, применимыми плазмидными векторами для Escherichia, более конкретно для Escherichia со/г, являются плазмиды серий pBR и pUC, и применимыми промоторами являются промотор lac (β-галактозидаза), оперон trp (триптофановый оперон) и промотор tac (гибридный промотор lac-trp) и промоторы, производные от λ-faag PL или PR. Предпочтительные терминаторы представляют trpA- или производный от фага rrnB рибосомальный терминатор. К плазмидным векторам, применимым для рекомбинантной выработки в бактериях Streptococcus, относятся, например, pHV1301 (FEMS Microbiol. Lett. 26, 1985, с.239) и pGK1 (Appl. Environ. Microbiol. 50, 1985. с.94). Плазмидные векторы, применимые для рекомбинантной выработки в Lactobacillus, включают, например, векторы, описанные для Streptococcus, например, рАМβ1 (J. Bacteriol. 137, 1979, с.614). Плазмидные векторы, применимые для рекомбинантной выработки в Saccharomyces, предпочтительно Saccharomyces cerevisiae, включают, например, векторы, серий YRp, YEp, YCp и YIp. Вектор интеграции (ЕР 5327456), сконструированный с помощью применения гомологической рекомбинации рибосомальной ДНК с копией в хромосоме, применим для инсерции копии и для стабильного генного контроля. Плазмидные векторы, применимые для рекомбинантной выработки у Kluyveromyces, предпочтительно Kluyveromyces lactis, включают, например, серии плазмид размером 2 мкм, производные от Saccharomyces cerevisiae, серии плазмид pKD1 (J. Bacteriol. 145, 1981, cc.382-390) и pGK11-производную плазмиду, участвующую в действии по уничтожению, плазмиду серий KARS с геном аутоиммунной репликации Kluyveromyces и интегрирующим вектором (ЕР 537456). Плазмидные векторы, применимые для рекомбинантной выработки у Pichia, включают, например, систему вектора хозяина, разработанную в Pichia pastoris, используя ген, который участвует в автономной репликации у Pichia (Mol. Cell. Biol. 5, 1985, с.3376). Плазмидные векторы, применимые для рекомбинантной выработки у Candida включают, например, систему вектора хозяина сформированного у Candida maltosa, Candida albicans и Candida tropicalis. (Agri. Biol. Chem. 51, 1987, с.1587. Плазмидные векторы, применимые для получения путем рекомбинации у Aspergillus, включают, например, вектор, сконструированный путем интеграции гена в плазмиду или хромосому и промотора для внеклеточной протеазы или амилазы (Trends in Biotechnology 7, 1989, cc.283-287). Плазмидные векторы, применимые для рекомбинантной выработки у Trichoderma, включают, например, систему вектора хозяина, сформированного у Trichoderma reesei, и промотор внеклеточной целлюлазы, который применима для конструирования этого вектора (Biotechnology 7, 1989, cc.596-603).
Предпочтительно указанная система выработки предусматривается с конструкцией нуклеиновой кислоты, предпочтительно с конструкцией ДНК, кодирующей белок, представленный в табл.5 и/или табл.6, или его иммуногенную часть.
В другом варианте осуществления настоящего изобретения указанная система выработки предусматривает конструкцию ДНК, кодирующую два, или три, или четыре, или даже большее количество белков, представленных в табл.5 и/или табл.6, или их иммуногенных частей.
В еще одном из вариантов осуществления настоящего изобретения предусмотрена указанная система выработки по меньшей мере с двумя конструкциями ДНК, каждая из которых кодирует по меньшей мере один белок, представленный в табл.5 и/или табл.6, или его иммуногенную часть.
В предпочтительном варианте осуществления настоящего изобретения указанный белок, представленный в табл.5 и/или табл.6, выбран из группы, состоящей из Р15, Р16, Р17, Р19, Р20, Р22, Р27, Р54, Р28, Р63, Р64, Р68, Р75, Р81, Р93, Р100 и Р105. В еще более предпочтительном варианте осуществления настоящего изобретения указанный белок, представленный в табл.5 и/или табл.6, выбран из группы, состоящей из Р15, Р16, Р20, Р27, Р54, Р28, Р63, Р68, Р93 и P105. Наиболее предпочтительно указанный белок, представленный в табл.5 и/или табл.6, выбран из группы, включающей Р15, Р16, Р54, Р28, Р63 и Р105.
В более предпочтительном варианте осуществления настоящего изобретения указанная конструкция нуклеиновой кислоты кодирует гибридный белок, включающий иммуногенные эпитопы, производные более чем от одного белка S. uberis. Более предпочтительно, указанный гибридный белок включает эпитопы, производные от белков, производных от более чем одного штамма S. uberis. В другом варианте осуществления настоящего изобретения указанная конструкция нуклеиновой кислоты кодирует эпитопы, которые модифицируют для повышения гуморального и/или клеточного иммунного ответа. Таким образом, настоящая заявка предусматривает способ получения иммуногенной композиции, включающей по меньшей мере два белка S. uberis, которые способны вызвать иммунный ответ против Streptococcus uberis, указанная способ, включающий получение клеток с рекомбинантным вектором, причем указанный вектор, включающий нуклеиновую кислоту, колирующую по меньшей мере два белка, представленных в табл.5 и/или табл.6, и/или иммуногенную часть, или аналог, или производное одного или обоих из указанных белков. Для рекомбинантного получения белка, кодируемого нуклеиновой кислотой, указанную нуклеиновую кислоту предпочтительно помещают под контроль индуцируемой регуляторной последовательности, способной повысить экспрессию указанного белка, предпочтительно приводя к повышенному уровню белка. Предпочтительно накопление указанного рекомбинантного белка или его иммуногенной части происходит либо в цитоплазме, например, в случаях, в которых вырабатываемый рекомбинантный белок собирают из клеток, либо белок экскретируется, например, если собирают из культуральной жидкости. Таким образом, рекомбинантную конструкцию, кодирующую указанный иммуногенный белок, получают с точными регуляторными последовательностями и/или функционально связанным промотором для внутриклекточного или внеклеточного накопления.
Таким образом, настоящее изобретение предусматривает рекомбинантную молекулу, включающую последовательность нуклеиновой кислоты, кодирующую по меньшей мере два белка, представленных в табл.5 и/или табл.6, и/или иммуногенную часть, или аналог, или производное одного или обоих указанных белков под контролем функционально связанного промотора. В предпочтительном варианте осуществления настоящего изобретения указанный белок, из числа представленных в табл.5 и/или табл.6, выбран из группы, состоящей из Р15, Р16, Р17, Р19, Р20, Р22, Р27, Р54, Р28, Р63, Р64, Р68, Р75, Р81, Р93, Р100 и Р105. Еще более предпочтителен белок, из числа представленных в табл.5 и/или табл.6 и выбранный из группы, состоящей из Р15, Р16, Р20, Р27, Р54, Р28, Р63, Р68, Р93 и Р105. Наиболее предпочтителен белок, из числа представленных в табл.5 и/или табл.6, выбранный из группы, состоящей из Р15, Р16, Р54, Р28, Р63 и Р105.
В настоящем изобретении также предусматривается живой рекомбинантный носитель, включающий последовательность нуклеиновой кислоты по настоящему изобретению или рекомбинантную молекулу ДНК по настоящему изобретению. В предпочтительном варианте осуществления настоящего изобретения носитель является первым штаммом S. uberis и рекомбинантная нуклеиновая кислота кодирует иммуногенные эпитопы другого штамма S. uberis. Таким образом, настоящее изобретение предусматривает живой рекомбинантный носитель, включающий последовательность нуклеиновой кислоты, кодирующей один или несколько белков, представленных в табл.5 и/или табл.6, и/или иммуногенную часть, или аналог, или производное одного или обоих указанных белков под контролем функционально связанного промотора. Безусловно, указанный живой рекомбинантный носитель в неживой форме также является иммуногенным. Таким образом, настоящее изобретение также описывает неживой рекомбинантный носитель.
В настоящем изобретении также предусматриваются выделенные клетки-хозяева, включающие последовательность нуклеиновой кислоты, кодирующую один или несколько белков, из представленных в табл.5 и/или табл.6, и/или иммуногенную часть, или аналог, или производное одного или обоих указанных белков под контролем функционально связанного промотора. Выделенные клетки-хозяева, например, включают бактериальные клетки, например: Escherichia, Bacillus, Pseudomonas, Serratia, Brevibacterium, Corynebacterium, Streptococcus uberis, Streptococcus suis, Lactobacillus, или клетки дрожжей, например: Saccharomyces, Kluyveromyces, Schizosaccharomyces, Zygosaccharomyces, Yarrowia, Trichosporon, Rhodosporidium, Hansenula, Pichia, Candida, или клетки грибов, например: Neurospora, Aspergillus, Cephalosporium и/или Trichoderma.
С помощью указанных выше выделенных клеток-хозяев, включающих рекомбинантную молекулу, содержащую последовательность нуклеиновой кислоты, кодирующую один или несколько белков, представленных в табл.5 и/или табл.6, и/или иммуногенную часть, или аналог, или производное одного или обоих указанных белков под контролем функционально связанного промотора, настоящее изобретение описывает для специалистов способ получения рекомбинантной белковой молекулы. Из-за различий между разными штаммами S. uberis белки и пептиды от разных штаммов могут показывать небольшую вариабельность аминокислотной последовательности и по-прежнему имеют ту же функцию. Таким образом, белковая молекула, полученная от одного штамма S. uberis, по последовательности идентична функционально идентичной белковой молекуле другого штамма S. uberis. Понятие «идентичность последовательности» относится к процентной идентичности двух аминокислотных последовательностей или нуклеотидных последовательностей после выравнивания двух последовательностей и внедрения гэпов, при необходимости, для достижения максимальной процентной идентичности. Способы и компьютерные программы для выравнивания известны специалистам в данной области. Одной из компьютерных программ, которая может использоваться или может быть адаптирована для целей определения, попадает ли последовательность-кандидат в данное определение, является программа «Align 2», разработанная фирмой Genentech, Inc., которая в Агентстве охраны авторских прав США (United States Copyright Office, Washington, D.C. 20559) получила дату приоритета 10 декабря 1991 г. Две аминокислотные последовательности имеют высокую степень «идентичности последовательностей» друг к другу, если последовательности проявляют по меньшей мере примерно на 80%, предпочтительно по меньшей мере примерно на 90%, и наиболее предпочтительно по меньшей мере примерно на 95% идентичность молекул после выравнивания. Таким образом, настоящее изобретение описывает выделенную и/или рекомбинантную белковую молекулу, которая имеет по меньшей мере на 80% идентичную последовательность к белку, кодируемому нуклеиновой кислотой по настоящему изобретению. В предпочтительном варианте осуществления настоящего изобретения описывают выделенную и/или рекомбинантную белковую молекулу, которая имеет по меньшей мере на 95% идентичную последовательность к белковой молекуле, кодируемой нуклеиновой кислотой по настоящему изобретению.
Для индукции иммунного ответа у животного против белка или иммуногенной части по настоящему изобретению, указанное животное обеспечивают белком и/или иммуногенной частью, которая включает по меньшей мере один иммуногенный сайт. Иммуногенная часть или сайт белка сформированы одним или несколькими эпитопами и, таким образом, способны индуцировать иммунный ответ. Иммуногенный сайт включает предпочтительно по меньшей мере 5 аминокислот, более предпочтительно по меньшей мере 10-15, и наиболее предпочтительно 25 или более последовательно расположенных аминокислот. Настоящее изобретение в другом предпочтительном варианте его осуществления предусматривает белок или его иммуногенную часть, которые включают по меньшей мере отрезок из 25 последовательно расположенных аминокислот белковой молекулы, кодируемой нуклеиновой кислотой по настоящему изобретению. Предпочтительно указанный отрезок, состоящий по меньшей мере из 25 последовательно расположенных аминокислот, включает иммуногенный сайт. Рекомбинантная молекула нуклеиновой кислоты в предпочтительном варианте осуществления настоящего изобретения кодирует по меньшей мере один белок или гибридный белок, кодирующий по меньшей мере два белка или их иммуногенные части. В предпочтительном варианте осуществления настоящего изобретения указанный гибридный белок включает иммуногенные части белков по меньшей мере двух штаммов S. uberis. Таким образом, настоящее изобретение описывает нуклеиновую кислоту, кодирующую белковую молекулу по настоящему изобретению.
Экспрессия указанной нуклеиновой кислоты в клетках-хозяевах предусматривает иммуногенный белок по настоящему изобретению. Указанный иммуногенный белок предпочтительно включен в иммуногенную композицию по настоящему изобретению. Таким образом, настоящее изобретение предусматривает иммуногенную композицию, способную индуцировать иммунный ответ против Streptococcus uberis, причем указанная композиция включает выделенную и/или рекомбинантную белковую молекулу по настоящему изобретению.
Указанный рекомбинантный белок по настоящему изобретению вырабатывается клетками-хозяевами. Для выработки используют клетки-хозяева бактериальных видов, например, Е. coli, и/или дрожжей, или грибов, или эукариотических клеток.
Иммуногенная композиция, включающая выделенный или рекомбинантный белок, и/или клетки, экспонирующие указанный белок, способна индуцировать иммунный ответ против Streptococcus uberis после введения животному, предпочтительно корове. Предпочтительно указанные клетки, включающие нуклеиновую кислоту по настоящему изобретению под контролем соответствующей регуляторной последовательности, экспрессируют указанный белок на поверхности. Такие клетки называются клетками-носителями. Указанные клетки-носители предпочтительно включены в иммуногенную композицию по настоящему изобретению. Предпочтительно, указанные клетки, несущие указанный иммуногенный белок, являются живым рекомбинантным носителем, но, в другом варианте осуществления настоящего изобретения, указанные клетки-носители способны индуцировать иммунный ответ, после того, как клетки убивают, например, обработкой формалином. Таким образом, настоящее изобретение предусматривает иммуногенную композицию, способную индуцировать иммунный ответ против Streptococcus uberis, причем указанная композиция включает живой или убитый рекомбинантный носитель по настоящему изобретению. Поскольку причиной мастита у коров является бактерия Streptococcus uberis, в предпочтительном варианте осуществления настоящего изобретения указанным живым или убитым рекомбинантным носителем является вид Streptococcus. В одном из вариантов осуществления настоящего изобретения указанные клетки-хозяева относятся к клеткам бактерий видов рода стрептококков. Белковая молекула затем предпочтительно содержится в составе других стрептококковых белков, которые повышают индуцируемый иммунный ответ. Кроме того, иммуногенность может быть повышена за счет сверхэкспрессии рекомбинантных белков на поверхности клеток-хозяев, предпочтительно клеток стрептококков. Кроме того, применение разных штаммов стрептококковых клеток-хозяев повышает иммуногенность иммуногенных белков или их частей. Таким образом, настоящее изобретение также предусматривает иммуногенную композицию по настоящему изобретению, в которой указанные клетки-хозяева относятся к бактериям видов стрептококков. Несколько видов стрептококков также могут применяться в качестве клеток-хозяев, например, S. suis, или S. agalactiae, или S. dysgalactiae.
Из-за различий между разными видами стрептококков, а, также учитывая, что некоторые белки, естественным образом присутствующие на бактериях Streptococcus uberis, могут способствовать индукции иммунного ответа против Streptococcus uberis, в предпочтительном варианте осуществления настоящего изобретения, аттеннуированную бактерию Streptococcus uberis используют в качестве живого рекомбинантного носителя в иммуногенной композиции по настоящему изобретению. Предпочтительно указанные бактерии S. uberis экспрессируют белки другого штамма S. uberis, тем самым, обнаруживая различные вакцины, поскольку сыворотка животного, вакцинированного указанной вакциной, отлична от сыворотки животного, инфицированного полевой инфекцией, поскольку выявляет антитела против белков обоих штаммов. В другом варианте осуществления настоящего изобретения указанная бактерия S. uberis в качестве клеток-хозяев или в качестве живого или убитого носителя замещена на бактерию S. suis, или виды Staphylococcus, или Е. coli, либо живые, либо убитые.
Введение иммуногенного белка по настоящему изобретению или его части, или введение нуклеиновой кислоты, кодирующей указанный белок по настоящему изобретению, индуцирует иммунный ответ. Указанная нуклеиновая кислота при введении корове экспрессируется в клетках указанной коровы и распознается иммунной системой указанной коровы. Нуклеиновая кислота, тем самым, участвует в качестве иммуногенной композиции, например, ДНК вакцины против мастита. Таким образом, настоящее изобретение предусматривает иммуногенную композицию, способную индуцировать иммунный ответ против Streptococcus uberis, причем указанная композиция включает нуклеиновую кислоту по настоящему изобретению.
Поскольку иммуногенная композиция по настоящему изобретению индуцирует иммунный ответ у животного, предпочтительно у коровы, белок по настоящему изобретению снижает проявление заболевания, связанного с маститом, и улучшает состояние здоровья указанных коров, тем самым, делая коров более устойчивыми к другим (вторичным) инфекциям. Таким образом, настоящее изобретение предусматривает иммуногенную композицию по настоящему изобретению для применения в качестве лекарственного средства.
Предпочтительно, иммуногенную композицию по настоящему изобретению используют для получения лекарственного средства против мастита, вызываемого Streptococcus uberis, которое снижает специфическое проявление болезни, связанной с маститом, вызванным Streptococcus uberis. Таким образом, настоящее изобретение предусматривает применение иммуногенной композиции по настоящему изобретению для приготовления лекарственного средства против мастита, вызываемого Streptococcus uberis.
Иммуногенную композицию по настоящему изобретению также используют для получения или переработки вакцины против мастита. Вакцина обычно предохраняет животных или людей от заражения. Предпочтительно иммуногенная композиция по настоящему изобретению способна предупредить мастит. Таким образом, настоящее изобретение описывает в предпочтительном варианте осуществления применение иммуногенной композиции по настоящему изобретению для приготовления вакцины.
В другом варианте осуществления настоящего изобретения иммуногенную композицию по настоящему изобретению предпочтительно используют для снижения количества и/или уничтожения бактерий S. uberis в молоке и/или в вымени коровы. Процесс дойки на молочных фермах представляет серьезную угрозу переноса бактерий S. uberis от больной коровы к здоровой. Снижение количества бактерий S. uberis, таким образом, наиболее важно для подавления распространения инфекции из одного соска вымени к другому соску и/или от животного к животному.
Для введения субъекту иммуногенной композиции по настоящему изобретению смешивание белков или его иммуногенных частей, или клеток-хозяев с соответствующим носителем облегчает получение субъектом иммуногенной композиции и повышает иммуногенный эффект композиции. К соответствующим носителям по настоящему изобретению, например, относится соответствующий адъювант для повышения иммунизирующего эффекта указанной иммуногенной композиции. Многие соответствующие адъюванты, основанные и на масле, и на воде, известны специалистам в данной области, например, адъювант Diluvacforte® или адъювант Specol®. В одном из вариантов осуществления настоящего изобретения соответствующий носитель включает, например, раствор, например, физиологический раствор для разведения бактерий, или белков, или их иммуногенных частей. Таким образом, настоящее изобретение описывает фармацевтическую композицию, включающую иммуногенную композицию по настоящему изобретению и соответствующий носитель.
В организме животного, предпочтительно коровы, иммунизированного иммуногенной композицией по настоящему изобретению, вырабатываются антитела, направленные против S. uberis. Наличие и уровень указанных антител являются показателями иммунитета после иммунизации иммуногенной композицией или вакциной по настоящему изобретению. Указанные антитела предпочтительно не направлены против эпитопов, которые имеются у полевых штаммов S. uberis дикого типа. Чтобы отличить вакцинированное животное от животного, которое инфицировано штаммом полевого типа, иммунитет указанного вакцинированного животного в одном из вариантов осуществления настоящего изобретения предпочтительно определяют по измерению антител, направленных против указанной иммуногенной композиции по настоящему изобретению. Антисыворотку указанной вакцинированной коровы также исследуют на наличие антител против антигенов S. uberis, которые отсутствуют в иммуногенной композиции. Выявление антител против антигена S. uberis, отсутствующего в иммуногенной композиции или вакцине по настоящему изобретению, является показателем того, что инфекция относится к дикому типу. Таким образом, настоящее изобретение описывает способ измерения иммунитета животного против S. uberis, причем указанный способ, включающий определение по меньшей мере в одном образце от указанного животного наличия антитела, направленного против белка, выбранного из представленных в табл.5 и/или табл.6 белков или их иммуногенных частей. В предпочтительном варианте осуществления настоящего изобретения указанный белок выбран из таблицы 5 и/или таблицы 6 из группы, состоящей из Р15, Р16, Р17, Р19, Р20, Р22, Р27, Р54, Р28, Р63, Р64, Р68, Р75, Р81, Р93, Р100 и Р105. Еще более предпочтительно, указанный белок выбран из таблицы 5 и/или таблицы 6, из группы, состоящей из Р15, Р16, Р20, Р27, Р54, Р28, Р63, Р68, Р93 и Р105. Наиболее предпочтительно, указанный белок выбран из белков, представленных в табл.5 и/или табл.6, из группы, состоящей из Р15, Р16, Р54, Р28, Р63 и Р105.
Для выявления антитела, которое связывается с иммуногенной композицией по настоящему изобретению, применим диагностический набор, который включает по меньшей мере один из белков, представленных в табл.5 и/или табл.6, или его иммуногенную часть. Связывание антитела с указанным белком или его иммуногенной частью выявляют с помощью, например, выявления иммуннофлуоресцентного антитела, или выявления фермент-связанного антитела, или других средств выявления антитела, связанного с указанным белком или его иммуногенной частью.
В предпочтительном варианте осуществления настоящего изобретения указанный по меньшей мере один белок из представленных в табл.5 и/или табл.6, выбран из группы, включающей Р15, Р16, Р17, Р19, Р20, Р22, Р27, Р54, Р28, Р63, Р64, Р68, Р75, Р81, Р93, Р100 и Р105. Еще более предпочтителен указанный по меньшей мере один из белков, представленных в табл.5 и/или табл.6, выбранный из группы, включающей Р15, Р16, Р20, Р27, Р54, Р28, Р63, Р68, Р93 и Р105. Наиболее предпочтительно указанный по меньшей мере один из белков, представленных в табл.5 и/или табл.6, выбран из группы, включающей Р15, Р16, Р54, Р28, Р63 и Р105.
Таким образом, настоящее изобретение описывает диагностический набор, включающий по меньшей мере один белок, выбранный из представленных в табл.5 и/или табл.6, или его иммуногенную часть и средства выявления антитела, связывающегося с указанным белком или его иммуногенной частью. Таким диагностическим набором, например, является тест ELISA, или какой-либо другой тест, применимый для скрининга сывороток. Предпочтительно указанная тестовый набор применим для скрининга большого количества сывороток.
В другом варианте осуществления настоящего изобретения нуклеиновую кислоту по настоящему изобретению используют для выявления животных, зараженных штаммами дикого типа S. uberis в популяции животных, вакцинированных с иммуногенной композицией или вакциной по настоящему изобретению. Этого выявления, например, можно достичь методом ПЦР.
Настоящее изобретение описывает, что полезные иммуногенные белки по настоящему изобретению являются белковыми молекулами и/или белками, доступными для антитела на поверхности бактерии и общими для ряда штаммов S. uberis. В качестве примера поверхностные белки идентифицируют из последовательностей генома штаммов 41-241 и O140J путем выбора генов, содержащих одну или несколько последовательностей, обычно обнаруживаемых у поверхностных белков грамположительных бактерий, например, мотив LPXTG сортазы, требуемый для заякоривания белка клеточной стенкой, или мотив присоединения липида, необходимого для липопротеинов, или сигнальная последовательность, или трансмембранная область, упреждающая поверхностную локализацию кодируемого белка.
Настоящее изобретение описывает наличие выбранных белков у штаммов S. uberis, что подтверждается зондированием хромосомальной ДНК ряда штаммов S. uberis с продуктами ПЦР, полученными из генов, например, отобранных выше.
Настоящее изобретение также поясняется приводимыми ниже примерами. Эти примеры не ограничивают рамок охвата настоящего изобретения, а только поясняют настоящее изобретение. Многие другие варианты его осуществления могут быть выполнены в рамках охвата настоящего изобретения.
Подписи к фигурам
Фиг.1. Анализ FACS, выполненный на интактных штаммах S. uberis. Штаммы S. uberis 41-241 (А) и O140J (Б) инкубируют с сывороткой иммунизированных мышей (темные столбцы) или с соответствующей сывороткой до иммунизации (светлые столбцы). Связанные антитела выявляют с помощью FITC-конъюгированных вторичных антител. Данные выражают в виде средней флуоресценции, связанной с бактериальными клетками. Флуоресценцию ≥10 (в 2 раза выше исходной) оценивают в качестве положительной. Номера сыворотки соответствуют номерам генов/белков, указанных в табл.1-4.
Фиг.2. Окрашенные кумасси бриллиантовым синим 2D протеомные образцы. Лизаты штаммов 41-241 (А) и O140J (Б) S. uberis на экспоненциальной стадии роста были зондированы сывороткой, полученной от коров после экспериментального заражения штаммом O140J. Обведенные кружком белки идентифицируют в качестве иммуногенных белков. Свойства идентифицированных белков по анализу в геле при расщеплении трипсином и по данным масс-спектрометрии MALDI-TOF показаны в табл.6.
Фиг.3. Заражение коров штаммом 0140J или штаммом 41-421 S. uberis.
3А. Коров 6716 и 6717 заражают через млечный проток штаммом 0140J S. uberis.
Коров 6720 и 6721 заражают через млечный проток штаммом S. uberis 41-421. Следует отметить, что корова 6720 заражена 5000 KOE S. uberis и корова 6721 заражена 500 KOE S. uberis.
SSC означает подсчет соматических клеток в молоке. ВО означает исследование бактерий и означает число бактерий в качестве колониеобразующих единиц (KOE), выделенных из молока.
RV означает правую переднюю четверть, LA означает левую заднюю четверть.
3Б. Клинические признаки, бактериальные и цитологические последствия заражения коров 6718 и 6719 штаммом 0140J S. uberis.
3В. Клинические признаки, бактериальные и цитологические последствия заражения коров 6722 и 6723 штаммом 41-241 S. uberis.
Фиг.4. Последовательности нуклеиновых кислот и аминокислот белков S. uberis, представленных в табл.5.
Примеры
Пример 1. Отбор общих поверхностных антигенов
Анализ последовательности ДНК. Последовательность ДНК штамма 41-241 S. uberis определяют с 2 × охватом. Данные по секвенированию собирают для получения 572 соприкасающихся последовательностей, содержащих 1815 ORF. В Центре Sanger штамм O140J S. uberis (Hill, 1988) был секвенирован. Данные о последовательности, доступные на сайте Sanger, были собраны в апреле 2002 г. для получения 61 контига, содержащего 1938 ORF.
Выбор общих поверхностных антигенов. Успешными вакцинными антигенами являются белки, доступные для антитела на бактериальной поверхности и общие для ряда штаммов S. uberis. Поверхностные белки идентифицируют из геномных последовательностей штаммов 41-241 и O140J путем выбора генов, содержащих одну или несколько последовательностей, которые формируют мотив подписи (см. М&М), обычно обнаруживаемый в поверхностных белках грамположительных бактерий. Среди всех исследованных ORF, 17 ORF содержат LPXTG мотив сортазы (табл.1), необходимый для заякоривания белка на клеточной стенке. Четыре белка из семнадцати указанных белков (Р12, Р23, Р24 и Р25) были обнаружены исключительно у штамма 41-241.
Тридцать одна открытая рамка считывания (ORF) содержит мотив присоединения липида, необходимый для липопротеинов (табл.2).
Все эти белки обнаружены у штамма O140J, а также у штамма 41-241. Кроме того, выбирают 87 ORF, которые содержат сигнальную последовательность или трансмембранную область, прогнозируя поверхностную локализацию кодируемого белка (табл.3).
Эти белки обнаруживают у штамма O140J, а также у штамма 41-241.
Пример 2. Распространение выбранных генов среди различных клинических и субклинических изолятов S. uberis
Для исследования наличия выбранных генов среди различных штаммов S. uberis, проводят эксперименты по точечной гибридизации, в которых хромосомальную ДНК значительного числа клинических штаммов S. uberis зондируют ПЦР-продуктами, полученными от 99 выбранных генов. Полученные данные (табл.4) показывают, что большинство выбранных генов гибридизируется с большинством штаммов S. uberis, следовательно, большинство выбранных генов обычно имеется у различных штаммов S. uberis. Напротив, 4 из 99 исследованных генов гибридизуется только с ограниченным числом штаммов. Все эти гены имеются в штамме 41-241 и кодируют белки, имеющие LPXTG мотив сортазы, требуемый для заякоривания белка на клеточной стенке.
Пример 3. Иммуногенность выбранных поверхностных белков
Для оценки роли белков в качестве белков-кандидатов для вакцины, белки, кодируемые 115 выбранными генами, клонируют и экспрессируют в Е. coli с полигистидиновыми метками. Продукты 106 из этих генов успешно клонируют и экспрессируют в Е. coli. Затем сыворотку, полученную от зараженных S. uberis коров и от кроликов, иммунизированных клетками S. uberis, убитыми формалином или ультразвуком, исследуют на наличие антител, направленных против экспрессированных белков, методом вестерн-блоттинга. Результаты (табл.5) показывают, что 19 из экспрессированных белков выявлены антителами, имеющимися в сыворотке животных, зараженных S. uberis, следовательно, эти белки экспрессируются in vivo и являются иммуногенными для коров. Кроме того, 30 экспрессированных белков были распознаны антителами, содержащимися в сыворотке от кроликов, иммунизированных клетками S. uberis, убитых формалином или ультразвуком. Двенадцать из числа экспрессированных белков распознаются и сывороткой, полученной от коров, зараженных S. uberis, и сывороткой от кроликов, иммунизированных клетками S. uberis, убитыми формалином или ультразвуком. Эти данные показывают, что большинство белков являются антигенными.
Таблица 5 также показывает, что некоторые белки распознаются антисыворотками, индуцированными после экспериментального заражения штаммами 41-241 и O140J. Однако, другие белки реагируют положительно исключительно с сывороткой, полученной после заражения штаммов O140J, или с сывороткой, полученной после заражения штаммом 41-241. Это предположительно указывает на различия между двумя штаммами, или в экспрессии белка in vivo, или в доступности этих белков для иммунной системы.
Пример 4. Очистка выбранных белков
Для дальнейшей оценки роли белков в качестве вакцин-кандидатов очищают все 36 белков, распознанных или сывороткой от зараженных коров, и/или сывороткой от иммунизированных кроликов. Кроме того, очищают 4 белка, которые содержат мотив сортазы, но которые не реагируют с обеими сыворотками. Тридцать один из 40 выбранных рекомбинантных белков успешно сверхэкспрессируют в растворимой форме и они могут быть очищены в нативных условиях, хотя 5 белков экспрессируются в виде нерастворимых телец включений. Эти белки очищают, используя условия для денатурирования, и после очистки заново свертывают. Для 4 белков не удалось получить достаточного для иммунизации количества. Все 36 очищенных белков затем используют для иммунизации мышей. Из результатов вестерн-блоттинга следует, что ни один из белков не взаимодействует с сывороткой, полученной от мышей до иммунизации. Напротив, белки, прочно взаимодействующие с иммунной сывороткой, полученной от мышей (табл.5), показывают, что белки в высокой степени иммуногенны у мышей. Специфичность индуцированного антитела подтверждают иммуноблоттингом против лизатов (или супернатантов протопластов) клеток S. uberis. Результаты показывают, что большинство индуцированных антител специфически реагируют с белком S. uberis ожидаемой молекулярной массы, четко указывая, что эти белки представляют (поверхностные) антигены, которые способны индуцировать иммунный ответ у мышей.
Пример 5. Функциональные свойства индуцированных антител
Используют сыворотку мышей в анализе методом FACS для изучения связывания антитела с целыми инкапсулированными клетками S. uberis (выращенными в среде Todd-Hewitt). На фиг.1 показано, что ни одна из сывороток, полученных от мышей до иммунизации, не способна связываться с целыми клетками S. uberis. Напротив, только восемь из иммунных сывороток (сывороток, индуцированных против белков Р11, Р15, Р17, Р20, Р25, Р26, Р27, Р63) сильно связываются с целыми бактериальными клетками, хотя две сыворотки (нацеленные против белков Р17, P18) показывают слабое связывание с целыми бактериальными клетками. Из указанных результатов с очевидностью следует, что белки, распознанные этими сыворотками, экспрессируются в условиях, используемых для роста бактериальных клеток S. uberis, и применимы для связывания с антителами. Кроме того, фиг.1 четко показывает, что экспрессия и/или поверхностная доступность белков различны у двух разных штаммов. Экспрессия и/или поверхностная доступность сохраняются в трех белках (Р17, Р19 и Р20). Напротив, Р11 и Р63 обнаруживают исключительно при использовании штамма O140J, хотя Р15, Р18, Р25 и Р26 обнаруживают исключительно при использовании штамма 41-241. Белок Р27 доступен на поверхности и штамма 41-241, и O140J, но лишь слабо распознается антисывороткой штамма 41-241.
В совокупности приведенные данные с очевидностью свидетельствуют о том, что 10 из выбранных антигенов экспрессируются на поверхности бактерий, растущих in vitro, и применимы для связывания антитела на интактных инкапсулированных клетках. Трое из этих белков сохраняют в двух исследуемых штаммах.
Пример 6. Серологический протеомный подход
В качестве другого подхода к идентификации вакцин-кандидатов S. uberis используют серологический протеомный анализ.
Белки штаммов S. uberis, выращенных в бульоне ТН, разделяют 2D гель-электрофорезом и зондируют антителами, содержащимися в сыворотках животных, зараженных S. uberis. Ряд высоко иммунногенных белков S. uberis был идентифицирован (данные не представлены). Три пятна успешно спаривают с белками, присутствующими в геле 2D, окрашенном красителем бриллиантовым синим кумаси (фиг.2). Продукты расщепления этих белков трипсином анализируют методом Q-TOF и получаемые фингерпринты пептидной массы сравнивают с in silico выработанными фингерпринтами пептидной массы всех белков, рассчитанных из анализа геномной последовательности штаммов 41-241 и O140J. Кроме того, два больших триптических пептида, выбранных из каждого фингерпринта, используют для тандемной MS. Получаемую аминокислотную последовательность пептидов затем сравнивают с последовательностями, рассчитанными по геномным последовательностям S. uberis. Все три белка могут быть успешно спарены с помощью этой процедуры. Свойства идентифицированных белков перечислены в табл.6. Одну из этих вакцин-кандидатов также идентифицируют, используя геномный подход (Р63). Это подчеркивает важность этого белка в качестве вакцины-кандидата.
Пример 7. Анализ методом ELISA культуральных супернатантов с мышиной иммунной сывороткой
Четко устанавливают Р93 и Р105 в супернатантах культур обоих штаммов, свидетельствуя о том, что эти белки секретируются бактериями.
Пример 8. Анализ FACS с применением клеток S. uberis, выращенных на молочной сыворотке, для имитации условий in vivo
Анализ, представленный в примере 5, проводят для исследования антитела к клеткам S. uberis, выращенным на среде, имитирующей молоко.
Пример 9. Консервация антигенов среди различных штаммов S. uberis
Исследуют анализом FACS сохранение выбранных белков и доступности к антителам среди разных изолятов S. uberis. Эти исследования позволяют выбрать вакцины-кандидаты, направленные на различные штаммы S. uberis.
Пример 10. Вакцинация и заражение скота
Экспериментальное инфицирование невакцинированных животных
Вирулентность штаммов S. uberis O140J и 41-241 определяют после экспериментального инфицирования. Вымя разделяют на четыре части, которые обычно называют квадратами. Каждый квадрат включает клетки, секретирующие молоко, млечные протоки, полости для молока и сосок. В этом эксперименте каждый квадрат отдельно инокулируют в полость для молока через млечный проток и сосок. Шесть из восьми квадратов, инокулированных штаммом O140J, становятся успешно инфицированными (фиг.3, табл.7). Чистые культуры S. uberis O140J выделяют из молока, полученного из таких квадратов, и определяют повышенные уровни количества соматических клеток (КСК). В двух четвертях после заражения не могут обнаружить инфекцию (двух разных коров, коровы №№6717 и 6719, фиг.3А и 3Б). Обе оставшиеся четверти остаются отрицательными по бактериологическим параметрам на протяжении эксперимента. В одной из двух четвертей наблюдают небольшое повышение КСК. Напротив, во всех восьми четвертях, зараженных штаммом 41-241, устанавливают инфекцию (фиг.3А и 3В; табл.7). Четыре из этих восьми четвертей (коровы 6721 и 6723) заражают дозой 5х10 КОЕ штамма 41-241, а другие четыре четверти заражают дозой 5х10 КОЕ (коровы 6720 и 6722). Более тяжелые эффекты наблюдают после инокуляции повышенной дозой: температура тела коров повышена более существенно и наблюдают более тяжелые клинические симптомы заболевания (сгустки в молоке и уплотнения вымени) (табл.7, фиг.3А и 3В). Однако клинические симптомы мастита также возникают при использовании штамма 41-241 при дозе заражения 5×102 КОЕ.
Сходные данные были ранее получены для штамма O140J (Hill, 1988).
По сравнению со штаммом 41-241 клинические признаки мастита, вызванного штаммом O140J (доза заражения 5×102 КОЕ, исследуют в течение 16 суток), представляются более тяжелыми (фиг.3Б и 3В). Три из четырех четвертей становятся эффективно инфицированными штаммом O140J и все три показывают клинические признаки мастита в течение по меньшей мере 16 суток после заражения. Две из этих четвертей остаются бактериологически положительными в течение 16 суток после заражения (фиг.3Б), и в одной четверти выявляют повышенный уровень КСК в течение 35 суток (данные не представлены). Все четыре четверти, зараженные штаммом 41-241, показывают клинические признаки мастита в течение 10-13 суток после заражения, но становятся отрицательными по клиническим признакам после 13 суток (фиг.3В). Две из этих четвертей (корова 6723) остаются бактериологически позитивными во время эксперимента (16 суток), свидетельствую о наличии S. uberis в молочной железе (фиг.3В).
Гистологическое исследование материала вымени, собранного через 3-5 суток после заражения обычно соответствует клиническим наблюдениям. Обе коровы, инфицированные штаммом 41-241, и одна корова, инфицированная штаммом O140J, болеют маститом в форме от умеренной до тяжелой по всей железе с много очаговым внутри альвеолярным накоплением полиморфно-ядерных гранулоцитов, фокальным разрушением эпителиального слоя в альвеолах и умеренной интерстициальной инфильтрацией мононуклеарных клеток (данные не представлены). Вторая корова, инфицированная штаммом O140J, больна умеренным много очаговым катаральным маститом.
В совокупности эти данные показывают, что оба штамма, 0140J и 41-241, патогенны для коров.
Иммунизация молочных коров
Повышается содержание поликлональных антител против белков S. uberis у коров. Коров иммунизируют по разным схемам иммунизации, используя подкожное заражение, и/или внутримышечное, и/или в молочную железу.
Иммуногенную композицию перерабатывают с растворителем, например, фосфатно-солевым буфером и адъювантом, например, адъювантом вода-в-масле или адъювантом без масла.
После иммунизации образец крови отбирают и сыворотку исследует на антитела против S. uberis.
Экспериментальное инфицирование вакцинированных животных
Вакцинированных и невакцинированных коров заражают 500 КОЕ штамма S. uberis O140J. Каждую четверть вымени отдельно заражают через млечный проток каждого соска.
После заражения коровы, вакцинированные иммуногенной композицией по настоящему изобретению, показывают слабые клинические признаки мастита, меньшее изменение молока, пониженные уровни КСК, сокращенный период клинической стадии мастита, меньшую лихорадку. Клинические оценки и гистологическое подтверждение при анализе вымени четко показывают, что иммунизация по настоящему изобретению эффективна против мастита, вызванного S. uberis.
Пример 11. Реактивность антигенов с сывороткой реконвалесцентов
Способность выбранных белков (Р15, Р16, Р20, Р22, Р27, Р54, Р28, Р63, Р68, Р75, Р93 и Р105) индуцировать антитела выздоравливающих исследуют методом вестерн-блоттинга, используя полевую сыворотку, полученную от 14 коров, которые выздоровели от имевшейся ранее инфекции S. uberis. Шесть из 12 выбранных антигенов (Р15, Р16, Р54, Р28, Р63, Р105) распознают все 14 используемых сывороток реконвалесцентов. Эти данные показывают, что указанные антигены экспрессируются всеми штаммами S. uberis, которые вызывают соответствующие инфекции, что эти антигены экспрессируются во время инфицирования в организме хозяина, и что эти антигены в высокой степени иммуногенны. Пять антигенов (Р68, Р27, Р20, Р93 и Р22) распознают сыворотки реконвалесцентов 8, 9, 10, 11 или 12, соответственно. С одним из антигенов нельзя обнаружить реакции с какой-либо сывороткой реконвалесцентов (Р75).
Пример 12. Сохранение антигенов в разных штаммах S. uberis
Чтобы показать применимость антигенов для выработки защиты против различных штаммов S. uberis, определяют сохранение и экспрессию 12 выбранных антигенов (Р15, Р16, Р20, Р22, Р27, Р54, Р28, Р63, Р68, Р75, Р93 и Р105) среди коллекции ранее выделенных полевых штаммов (35 штаммов). Экспрессию антигенов показывают скринингом методом вестерн-блоттинга с мышиной иммунной сывороткой против очищенных антигенов. Пять из 12 антигенов (Р15, Р16, Р28, Р75 и Р105) экспрессируются у более чем 97% исследованных штаммов, два из антигенов (Р27, Р63) экспрессируются у 94% штаммов и четыре антигена (Р20, Р22, Р54, Р93) экспрессируются у 81-92% штаммов. Эти данные с очевидностью показывают, что экспрессия большинства антигенов высоко консервативна среди различных штаммов S. uberis.
Материалы и методы
Бактериальные штаммы и условия роста
Один штамм S. uberis 41-241 был выделен в 1998 г.на коммерческой молочной ферме, на которой была вспышка мастита, вызванного S. uberis (Hill, 1988). Штамм 41-241 показывает, что методом RAPD (random amplified polymorphic DNA - случайно амплифицированной полиморфной ДНК) преимущественно выявляют фингерпринтинг типа В в определенном стаде во время вспышки инфекции (Zadoks и др., 2003). Штамм выделяют у коров, инфицированных S. uberis в течение по меньшей мере двух месяцев. Начало инфицирования протекает бессимптомно и затем сопровождается резким проявлением различных клинических симптомов.
Штаммы S. uberis O140J и EF20 были любезно предоставлены доктором J. Leigh, Институт ветеринарии, Комптон, Англия. Другие штаммы S. uberis и S. para uberis, выделенные при клиническом проявлении симптомов мастита на разных молочных фермах, были любезно предоставлены доктором D.Mevius, фирма CIDC, Leiystad, Нидерланды, докторами O.Sampimon, фирма Animal Health Service, Deventer, Нидерланды, или Dierenartsen Praktijk, Diessen, Нидерланды. Все другие виды стрептококков получены из лабораторной коллекции ASG, Leiystad, Нидерланды. Штаммы стрептококков выращивают в бульоне Todd-Hewitt (код СМ 189, фирма Oxoid) и помещают на кровяной агар Columbia (код СМ331, фирма Oxoid), содержащий 6 об.% крови лошади и 0,1 мас.% аэскулина если не указано иначе. Штаммы Е. coli выращивают в бульоне Luria (18) и помещают на агар Luria, содержащий 1,5 мас.% агара. При необходимости вносят 50 мкг/мл канамицина.
Приготовление сыворотки. Молоко получают с молочных ферм, о которых известно, что у них ранее было заражение S. uberis (Waiboerhoeve, Leiystad, Нидерланды). Молоко центрифугируют в течение 30 мин при 12800 g и удаляют масло. Затем добавляют 40 мкл/мл реннина (Lactoferm, Brouwiand, Belgium) и молоко инкубируют в течение 2 ч при 37°С с регулярным перемешиванием. Коагулированное молоко удаляют фильтрацией и оставшийся супернатант центрифугируют в течение 30 мин при 12800 g. Осветленный супернатант стерилизуют фильтрацией через фильтр Sartobran P с размером пор 0,2 мкм (фирма Sartonus, Goettingen, Германия).
Образцы молока и сыворотка. Образцы молока и сыворотку получают от животных, больных маститом, вызванным S. uberis, на разных молочных фермах Голландии (от Drs. O.Sampimon, Ветеринарная служба, Дэвентер, Нидерланды). Ни одно из животных не лечили антибиотиками до взятия образцов.
Кроме того, молоко и сыворотку получают у коров в разное время после экспериментального заражения штаммами S. uberis O140J и 41-241.
Антисыворотка кролика. Поликлональные антитела, направленные против убитых формалином целых клеток S. uberis, а также против обработанных ультразвуком клеток S. uberis, повышаются у кроликов. Кроликов иммунизируют подкожно, используя 2-4×109 убитых клеток в адъюванте вода-в-масле. Инокулирования повторяют спустя две, три и четыре недели. Через 6 недель кроликов умерщвляют и собирают сыворотку.
Для приготовления антигенов штаммы S. uberis выращивают в течение 16 ч в бульоне Todd-Hewitt. Культуры разводят в 10 раз в 1 л предварительно подогретого бульона Todd-Hewitt и клетки выращивают до оптической плотности 0,5 (600 нм). Культуры центрифугируют в течение 15 мин при 10000 g и осадки растворяют в 100 мл ФСБ (136,89 мМ NaCl, 2,68 мМ KCl, 8,1 мМ Na2HPO4, 2,79 мМ KH2PO4 pH 7,2). Затем оптическую плотность (600 нм) доводят до 1,0 с помощью ФСБ. Для приготовления фиксированных формалином клеток порции этих клеток объемом 10 мл центрифугируют в течение 20 мин при 10000 g и осадки ресуспендируют в 2,5 мл ФСБ. К этой суспензии добавляют 250 мкл 3% формалина и поддерживают в течение 16 ч при комнатной температуре. Суспензию проверяют на отсутствие живых бактерий путем высева на агаровую среду Columbia. Для удаления формалина клетки дважды промывают ФСБ. Для приготовления обработанных ультразвуком клеток порции по 10 мл клеток центрифугируют в течение 20 мин при 10000 g, осадок ресуспендируют в 250 мкл ФСБ. Клетки обрабатывают ультразвуком в течение 15 мин, используя ультразвуковой генератор с наконечником при 100% выходе, 50% нагрузке. После обработки ультразвуком клетки разводят в 10 раз в ФСБ. Оба антигена смешивают 1:1 со Specol для получения эмульсий вода-в масле.
Геномные последовательности. Геномную ДНК выделяют из штамма S. uberis 41-241 по описанию Sambrook и др. (1989). ДНК нарезают и используют для создания плазмидной библиотеки. Случайные клоны секвенируют, используя метод химии красителей-терминаторов (dye-terminator chemistry), и анализируют на анализаторе ABI PRISM 3700 ДНК (фирма Applied Biosystems, Warrington, Великобритания). Данные по секвенированию собирают для получения 572 непрерывных последовательностей. Первоначальный набор открытых рамок считывания (open reading frames - ORF) идентифицируют с помощью программного обеспечения GLIMMER и GENEMARK.
Трансмембранные спирали и субклеточные локализации в генах прогнозируют с помощью компьютерной программы ТМНММ, которую можно получить на сайте www.cbs.dtu.dk/services/TMHMM. Для поиска в каждой ORF сигнальных пептидов используют программу SignalP (Nielsen и др., 1999). Другие прогнозы по сигнальным пептидам получают, используя программу PSORT, доступную на сайте http://psort.nibb.ac.jp, и программу GCG-SPS, доступную на сайте http://www.biology.wustl.edu /gcg/spscan.html. Липопротеины обнаруживают, используя программу GCG-Fmdpatterns со следующими выражениями: PS00013: ~(D,E,R,K)6(L,I,V,M,F,W,S,T,A,G)2(L,I,V,M,F,Y,S,T,A,G,C,Q)(A,G,S)C; g-lpp:<(M,V)X{0,13}(R,K)~(D,E,R,K,Q){6,20}(L,I,V,M,F,E,S,T,A,G)(L,V,I,A,M)(I,V, M,S,T,A,F,G)(A,G)C и g-lpp_rvh: (M,V,L)X{0,13}(R,K)~(D,E,R,K,Q){6,20}(L,I,V,M,F,E,S,T,A,G)(L,V,I,A,M)(I,V,M,S,T,A,F,G)(A,G)C. Белки с якорными доменами в клеточной стенке идентифицируют, используя добавление к базе InterPro IPR001899. Программу BLAST используют для последовательностей белков с последовательностью, идентичной с вычисленными аминокислотными последовательностями.
Спот-блоттинг, блоттинг по Саузерну и гибридизация. Хромосомальную ДНК выделяют согласно описанию Sambrook и др. (1989). Для спот-блоттинга один мкг хромосомальной ДНК наносят пятном на мембраны Genescreen Plus. Мембраны инкубируют в 0,4 М NaOH-1 M NaCl при комнатной температуре в течение 10 мин для денатурации ДНК в течение 10 мин в 0,6 М NaCl, 0,06 М цитрате натрия (pH 7,0) для нейтрализации. Для блоттинга по Саузерну фрагменты ДНК разделяют в 0,8% агарозных гелях и переносят на мембраны Gene-Screen Plus (NEN) согласно описаниям Sambrook и др. (1989). Зонды ДНК метят с помощью [(α-32P]dCTP (3000 Кюри × ммоль-1; фирма Amersham), используя набор для случайного нанесения метки (фирма Boehringer). ДНК в блотах гибридизируют при 65°С в буфере, содержащем 0,5 М фосфат натрия, 1 мМ EDTA и 7% натрия додецилсульфата при pH 7,2, с соответствующими зондами ДНК согласно рекомендациям поставщика мембран Gene Screen Plus. После гибридизации мембраны промывают дважды раствором 40 мМ фосфата натрия, pH 7,2, 1 мМ EDTA, 5% SDS в течение 30 мин при 65°С и дважды раствором 40 мМ фосфата натрия, pH 7,2, 1 мМ EDTA, 1% SDS в течение 30 мин при 65°С. Сигналы выявляют на приборе Phosphor-imager (Storm; фирма Molecular Dynamics).
Клонирование и экспрессия выбранных белков. Выбранные ORF амплифицируют методом ПЦР со специфическими олигонуклеотидными праймерами для клонирования в pET200/D-TOPO (фирма Invitrogen). Белки клонируют без предполагаемых сигнальных последовательностей или расчетных трансмембранных областей. Конструкции трансформируют в Escherichia coli BL21 Star (DE3) (фирма Invitrogen) для экспрессии рекомбинантных белков.
Для ПЦР-реакции (25 мкл) Platinum Pfx ДНКполимеразу (фирма Invitrogen) используют согласно рекомендациям производителя. Амплификацию ДНК проводят в термическом циклере Perkin Elmer 9700 по программе, включающей инкубирование в течение 5 мин при 94°С, 35 циклов по 15 сек при 94°С, 30 сек при 57°С, 2 мин при 68°С и 5 мин при 68°С.
Иммунодетекция экспрессированных антигенов. Белки разделяют SDS-полиакриламидным гель-электрофорезом, используя систему мини-клеток XCell SureLock (фирма Invitrogen). Белки в геле визуализируют, используя краситель SYPRO-orange (фирма Molecular Probes, Sunnyvale, Калифорния) и окрашивая по рекомендациям производителя. Сигналы выявляют на приборе Phosphor-imager (Storm; фирма Molecular Dynamics).
Белки переносят на мембрану из нитроцеллюлозы стандартными методами (19). Мембраны блокируют в Blotto: Tris-забуференный физиологический раствор (TBS) (50 мМ Tris-HCl [pH 7,5], 150 мМ NaCl), содержащий 4% снятого молока, 5% фетальной сыворотки теленка и 0,05% Tween 20, при комнатной температуре в течение 16 ч. Для выявления рекомбинантных антигенов мембраны инкубируют с моноклональным антителом против метки 6 × HIS (фирма Clontech, Palo Alto, Калифорния). Связанные антитела выявляют и визуализируют, используя конъюгат анти-мышиного антитела и щелочной фосфатазы и нитро-синий-тетразолий/5-бром-4-хлор-3-индолил-фосфат по описанию Sambrook и др. (1989).
Иммуногенность экспрессированных антигенов исследуют, используя образцы сыворотки, полученные от коров, которые клинически или субклинически заражают S. uberis, или используя кроличью анти-S. uberis антисыворотку. Связанные антитела выявляют с помощью крольчьих-анти-коровьих или козьих-анти-кроличьих иммуноглобулинов, конъюгированных со щелочной фосфатазой (фирма Jackson Immunoresearch), и визуализируют, используя нитро-синий-тетразолий/5-бром-4-хлор-3-индолилфосфат по описанию Sambrook и др. (1989).
Очистка белка. Белки очищают аффинно от клеточного осадка, используя Ni-нитрилтриуксусную кислоту (Ni2+-NTA) колоночную хроматографию по описанию производителя (фирма Qiagen). Вкратце, клетки выращивают экспоненциально, затем добавляют 1 мМ IPTG и клетки оставляют расти еще 4 ч при 37°С. Затем клетки собирают и лизируют. Осветленные супернатанты загружают в Ni2+-NTA агарозные колонки. Колонки промывают и белок элюируют. Разные буферы используют для нативной и для денатурирующей очистки. Белки, очищенные в условиях денатурирования, ренатурируют диализом, используя линейный 6 М - 0 М градиент мочевины в 286,89 мМ NaCl, 2,68 мМ KCl, 8,1 мМ Na2HPO4, 2,79 мМ KH2PO4 pH 7,2. Очищенные белки дополнительно концентрируют, используя фильтры Amicon Ultra-4 5000 MWCO (фирма Millipore).
Концентрация белка. Концентрацию белка в образцах определяют после гель-электрофореза в SDS полиакриламидном геле. Белки в геле визуализируют, используя краситель SYPRO-orange (фирма Molecular Probes, Sunnyvale, Калифорния), которым окрашивают согласно рекомендациям производителя. Сигналы выявляют на визуализирующем устройстве Phosphor-imager (Storm; фирмы Molecular Dynamics). Известный диапазон концентраций сывороточного альбумина быка используют в качестве стандарта для подсчета количеств белка, содержащихся в геле. Для подсчетов используют программу Molecular Dynamics.
Иммуногенность очищенных белков. Мышей линии OF1 иммунизируют подкожно, используя 20 мкг очищенных белков в полном адъюванте Фрейнда. Инокуляцию повторяют через три недели, используя 20 мкг очищенных белков в неполном адъюванте Фрейнда. Через три недели после второй инокуляции мышей умерщвляют и получают сыворотку.
Анализ FACS. Клетки S. uberis выращивают в бульоне Todd-Hewitt до оптической плотности OD600=0,5. Клетки собирают центрифугированием, один раз промывают в буфере FACS (ФСБ-13, pH 7,2 [137 мМ NaCl, 2,68 мМ KCl, 8,1 мМ Na2HPO4, 2,8 мМ KH2PO4] - 0,5% БСА) и корректируют плотность клеток до величины примерно OD600=1,0 в буфере FACS. Клетки (250 мкл) собирают центрифугированием и ресуспендируют в 50 мкл буфера FACS, содержащего мышиную антисыворотку (в разведении 1:50). Образцы инкубируют в течение 45 мин на льду. Для удаления несвязанных антител клетки промывают дважды в 250 мкл буфера FACS. Затем клетки инкубируют с 50 мкл буфера FACS, содержащего меченое флюоресцеинизотиоцианатом (fluorescein isothiocyanate - FITC) кроличье антимышиное вторичное антитело (разведение 1:100; фирма DAKO A/S, Glostrup, Дания), в течение 30 мин на льду. Клетки промывают дважды в 250 мкл буфера FACS, ресуспендируют в 100 мкл буфера FACS и связанное антитело обнаруживают сортировщиком флуоресцентно активированных клеток (прибор FACS Calibur, фирма Benton Dickinson, Franklin Lakes, США).
ELISA целых клеток. Клетки S. uberis на стадии экспоненциального роста собирают центрифугированием, ресуспендируют в покровном буфере с величиной pH 9,6 (0,05М NaHCO3, 0,05М Na2CO3) и оптическую плотность клеток корректируют до величины OD600=1,0. Высокого связывания 96-луночные планшеты покрывают такой суспензией в количестве 100 мкл/лунку и выдерживают 16 ч при 4°С. Лунки промывают 4 раза буфером ELISA (5% Tween-80, 0,02% азида Na), сыворотку (разведенную 1:20 в буфере ФСБ-13, содержащем 0,05% Tween-80, 2% NaCl и 5% фетальной сыворотки теленка) добавляют и планшеты инкубируют в течение 1 ч при 37°С. Для удаления несвязанного антитела лунки промывают четыре раза буфером ELISA. Затем добавляют вторичное антитело (100 мкл пероксидазы хрена, конъюгированной с кроличьим-анти-мышиным антителом (DAKO), разведенным 1:250 в буфере ФСБ-13, содержащем 0,05% Tween-80, 2% NaCl и 5% фетальной сыворотки теленка) и планшеты инкубируют в течение 1 ч при 37°С. Лунки опять промывают четыре раза буфером ELISA, и связанные антитела обнаруживают при комнатной температуре, используя 100 мкл тетраметилбензидина (tetramethylbenzidine - ТМВ) (фирма CeDi-Diagnostics, Leiystad, Нидерланды). Реакции останавливают через 15 мин, добавляя 100 мкл 0,5М H2SO4 в лунку. Поглощение считывают, используя ридер ELISA (фирма Thermo Labsystems, Франклин, США), при 450 нм.
Подготовка образцов для двухмерного гель-электрофореза. Штаммы S. uberis выращивают в течение 16 ч в 100 мл бульона Todd-Hewitt. Культуры разводят в 20 раз в 1 л предварительно подогретого бульона Todd-Hewitt и клетки выращивают до оптической плотности 0,5 (при 600 нм). Культуры центрифугируют в течение 20 мин при 10000×g, и осадки однократно промывают равным объемом 250 мМ сахарозы/25 мМ Tris, pH 8,0 и один раз равным объемом superQ. Получаемые осадки растворяют в 5 мл superQ. Эти суспензии по частям, объемом 1,5 мл, облучают в течение 15 мин, используя ультразвуковой дезинтегратор с наконечником (прибор Branson sonifier 250, 50% интервал, амплитуда 3). Затем суспензии обрабатывают ДНКазой I и MgCl2 (конечные концентрации 6,5 мкг/мл и 10 мМ, соответственно) в течение 10 мин при 37°С. Ингибиторы протеазы пепстатин А, лейпептин, пефаблок и апротинин вносят до конечной концентрации 2,5 мкг/мл, 5 мкг/мл, 25 мкг/мл и 1 мкг/мл, соответственно. Мочевину, дитиотрейтол и Triton-X100 вносят до конечной концентрации 9М, 70 мМ и 2%, соответственно. Образцы центрифугируют в течение 30 мин при 10000 g, супернатанты собирают и центрифугируют в течение дополнительных 30 мин при 100000 g. Супернатанты собирают и концентрации белка в образцах определяют, используя метод анализа белка RC DC (фирма BioRad) согласно инструкциям производителя.
Двухмерный гель-электрофорез. Образцы, содержащие 50-100 мкг белка, растворяют в 450 мкл образца буфера (8М мочевины, 2% CHAPS, 0,5% IPG-буфера 3-10, 70 мМ дитиотрейтола и следы красителя бромфенолового синего). Белки разделяют в первом направлении путем изоэлектрического фокусирования, используя иммобилиновые длиной 18 см полоски Dry Strips (3-10 NL фирмы Amershan Pharmacia Biotech) в камере IPGphor (фирма Amershan Pharmacia Biotech) после повторного увлажнения полосок согласно инструкциям производителя. Сразу же после фокусирования полоски впоследствии уравновешивают в течение 15 мин в выравнивающем буфере (6М мочевина, 30% глицерин, 2% SDS, 50 мМ Tris-HCL-pH 8,8, следы бром-фенолового синего), содержащем 10 мг/мл дитиотрейтола и в течение 15 мин в выравнивающем буфере, содержащем 25 мг/мл иодацетамида. Белки разделяют во втором направлении методом SDS-полиакриламидного гель-электрофореза в 12,5% гелях заводского изготовления Ettan DALT (фирма Amershan Pharmacia Biotech) в системе с 12 ячейками Ettan DALT (фирма Amersham Pharmacia Biotech) no инструкциям производителя.
Окрашивание. Белки в гелях окрашивают серебром, используя набор для окрашивания PlusОдин™ (фирма Amershan Pharmacia Biotech) по инструкциям производителя, или бриллиантовым синим кумаси по описанию Sambrook и др. (1989) с пролонгированным временем инкубирования из-за пластической основы гелей.
Расщепление белков из гелей 2D. Белковые пятна, идентифицированные в геле по окраске красителем кумаси, вырезают вручную. Кусочки геля замораживают в 0,1% уксусной кислоте при -80°С и хранят до использования. Белки в гелях расщепляю трипсином по описанию Li и др. (2003).
Масс-спектрометрия белковых пятен из гелей 2D, расщепленных трипсином. Micromass Q-TOF масс-спектрометр используют для анализа масс белковых пятен, расщепленных трипсином, по описанию Li и др. (2004).
Эксперименты по искусственному заражению
Животные. Клинически здоровых коров породы Голыптейн Фрезиан на 2-4 неделю после первой лактации, используют для инфицирования. Коров доят дважды в сутки в 7 ч утра и в 4 ч дня. У всех коров количество соматических клеток (КСК) ниже 2,0×105 клеток/мл, они отрицательны по возбудителям мастита, судя по повторной микробиологической оценке молока во время последних 14 суток до инфицирования, и ранее не болели маститом.
Приготовление инокулюма и заражение. Штаммы S. uberis O140J и 41-241 используют в качестве посевного материала (инокулюма). Отдельные колонии, выращенные в чашках на агаровой среде Columbia, содержащей 6 об.% крови лошади и 0,1 мас.% аэскулина, переносят в 90 мл бульона Todd-Hewitt (фирмы Oxoid) и культивируют в течение ночи при 37°С. Ночные культуры разводят 1:10 в той же среде и выращивают бактерии до концентрации примерно 3×108 КОЕ/мл (логарифмическая фаза роста). Затем клетки собирают центрифугированием, заново суспензируют и разводят в ФСБ.
Две четверти железы у каждой коровы заражают внутрь полостей количеством бактерий или 5×102 (штамм O140J или штамм 41-241), или 5×103 (штамм 41-241) КОЕ в 5 мл ФСБ. В контрольные четверти вводят 5 мл ФСБ.
Инъекции осуществляют сразу после полуденной дойки, используя имеющиеся сосочковые канюли. Перед инокулированием сосочки промывают спиртом. Инокулюм массируют по направлению к полостям железы.
Одну группу из четырех коров заражают 5×102 КОЕ штамма O140J. Другую группу из четырех коров заражают штаммом 41-241. Двух из этих коров заражают из расчета 5×102 КОЕ, а двух коров - 5×103 КОЕ. Коров используют в двух последовательных экспериментах, из которых один заканчивают через 45-80 ч, а второй через 16 суток.
Отбор образцов и клинические оценки. Образцы молока собирают асептически из всех четвертей вымени при каждой дойке. Образцы исследуют бактериологически в чашках с кровяным агаром, содержащим 6 об.% крови лошади и 0,1 мас.% аэскулина, и количество соматических клеток (КСК) определяют, используя стандартные процедуры (Международной федерации молочного производства, 1981). Кроме того, образцы молока хранят при -20°С до проведения анализа на ответы антител. В каждую дойку определяют температуру тела коров и выработку молока, а у коров проводят мониторинг клинических признаков мастита (последовательно по вымени и по комкам в молоке). Раз в неделю берут образцы крови на анализ ответов антител в сыворотке.
Патология. Для гистологического исследования ткань молочной железы из каждой четверти вымени отбирают из разных мест трех разных горизонтальных поперечных срезов, т.е. в основании железы, по середине от основания железы до полости железы и в полости железы. Образцы ткани фиксируют в 4% буферном формалине и погружают в парафин. Для гистологических исследований срезы тканей нарезают и окрашивают гематоксилином/эозином.
СПИСОК ЛИТЕРАТУРЫ
1. Alber T. Mutational effects on protein stability. Annu. Rev. Biochem. 58, 1989, cc.765-798.
2. Adv. Biochem. Eng. 43, 1990, cc.75-102.
3. Agri. Biol. Chem. 51, 1987, с.1587.
4. Appl. Environ. Microbiol. 50, 1985, с.94.
5. Biotechnology 7, 1989, cc. 596-603.
6. Bramley, Mastitis: physiology or pathology, cc.3-9. В кн.: (ed.), «New insights into the pathogenesis of mastitis», 1991, под ред. Burvenich С., G.Vandeputte-van Messom, A.W.Hill, изд-во Rijksuniversiteit Gent, Бельгия.
7. Biseibutsugaku Kisokoza (Basic Microbiology), 8, 1990, Genetic Technology, Kyoritsu Shuppan.
8. De Greeff и др., Infect Immun. 70, 2002, cc.1319-1325.
9. FEMS Microbiol. Lett. 26, 1985, с.239.
10. Fersht A.R., L.Serrano, Principles of protein stability derived from protein engineering experiments. Curr. Opinion Struct. Biol., 3, 1993, cc.75-83.
11. Finch J.M., A.Winter, A.W.Walton, J.A.Leigh. Further studies on the efficacy of a live vaccine versus mastitis caused by Streptococcus uberis. Vaccine 15, 1997, cc.1138-1143.
12. Fontaine M.C., J.Perez-Casal, X.M.Song, J.Shelford, P.J.Willson, A.A.Potter, Immunisation of dairy cattle with рекомбинантный Streptococcus uberis GapC or a chimeric CAMP antigen confers protection versus heterologous bacterial challenge. Vaccine 20, 2002, cc.2278-2286.
13. Grandi G. Antimicrobial vaccine design using genomics and proteomics. Trends in Microbiol. 19, 2001, cc.181-188.
14. Hill A.W. Pathogenicity of two strain Streptococcus uberis infused into lactating and non-lactating bovine mammary glands. Res Vet Sci. 45, 1988, cc.400-404.
15. Hillerton J.E., M.F.Shearn, R.M.Teverson, S.Langridge, J.M., Booth Effect of pre-milking teat dipping on clinical mastitis on dairy farms in England. J. of Dairy Research 60, 1993, cc.31-41.
16. Hogan, J.S., K.L.Smith, K.H.Hoblet, D.A.Todhunter, P.S.Schoenberger, W.D.Hueston, D.E.Pritchard, G.L.Bowman, L.E.Heider, B.L.Brockett и др. Bacterial counts in bedding materials used on nine commercial dairies. J. Dairy Sci 72, 1989, сс.250-258.
17. J. Bacteriol. 137, 1979, с.614.
18. J. Bacteriol. 145, 1981, сс.382-390.
19. Leigh J.A. Streptococcus uberis: a permanent barrier to the control of bovine mastitis?. Vet. J. 157, 1999, сс.225-238.
20. Leigh J.A. Vaccines versus bovine mastitis due to Streptococcus uberis current status and future prospects. Adv. Exp. Med. Biol. 480, 2000, сс.307-311.
21. Li K.W., M.P.Hornshaw, R.C. van der Schors, R.Watson, S.Tate, B.Casetta, C.R.Jimenez, Y.Gouwenberg, E.D.Gundelfinger, K-H.Smalla, A.B.Smit, Proteomics Analysis of Rat Brain Postsynaptic Density. J. Biol. Chem. 279, 2004, сс.987-1002.
22. Matthews B.W. Mutational analysis of protein stability. Curr. Opinion Struct. Biol., 1, 1991, сс.17-21.
23. McDougall, Efficacy of two antibiotic treatments in curing clinical and sub-clinical mastitis in lactating dairy cows. New Zealand Vet. J. 46, 1998, сс.226-232.
24. Mol. Cell. Biol. 5, 1985, с.3376.
25. Neave F.K., F.H.Dodd, R.G.Kingwell, D.R.Westgarth. Control of mastitis in the dairy herd by hygiene and management. J Dairy Sci.52, 1969, сс.696-707.
26. Nielsen H., S.Brunak, G. von Heijne. Machine learning approaches to the prediction of signal peptides and other protein sorting signals. Protein Engineering 12, 1999, сс.3-9.
27. Oliver S.P., M.J.Lewis, B.E.Gillespie, S.J.Ivey, L.H.Coleman, R.A.Almeida, W.Fang, and K.Lamar. Evaluation of a postmilking teat disinfectant containing a phenolic combination for the prevention of mastitis in lactating dairy cows. J. of Food Protection 62, 1999, сс.1354-1357.
28. Paton J.C., P.Giammarinaro. Genome-based analysis of pneumococcal virulence factors: the quest for novel vaccine antigens and drug targets. Trends in Microbiol. 9, 2001, сс.515-518.
29. Pizza M, V.Scarlato, V.Masignani, M.M.Giuliani, B.Arico, M.Comanducci, G.T.Jennings, L.Baldi, E.Bartolim, B.Capecchi, C.L.Galeotti, E.Luzzi, R.Manetti, E.Marchetti, M.Mora, S.Nuti, G.Ratti, L.Santini, S.Savino, M.Scarselli, E.Storm, P.Zuo, M.Broeker, E.Hundt, B.Knapp, E.Blair, T.Mason, H.Tettelin, D.W.Hood, A.C.Jeffries, N.J.Saunders, D.M.Granoff, J.C.Venter, E.R.Moxon, G.Grandi, R.Rappuoli. Identification of vaccine candidates versus serotype В Meningococcus by whole-genome sequencing. Science 287, 2000, сс.1816-1820.
30. Rappuoli R. Reverse vaccinology. Current Opinion in Microbiol. 3, 2000, сс.445-450.
31. Wizemann T.M., J.H.Heinrichs, J.E.Adamou, A.L.Erwin, C.Kunsch, G.H.Choi, S.C.Barash, C.A.Rosen, H.R.Masure, E.Tuomanen, A.Gayle, Y.A.Brewah, W.Walsh, P.Barren, R.Lathigra, M.Hanson, S.Langermann, S.Johnson, S.Koenig, Use of whole genome approach to identify vaccine molecules affording protection against Streptococcus pneumoniae infection. Infect. Immun. 69, 2001, сс.1593-1598.
32. Sambrook J., E.F.Fritsch, T.Maniatis. Molecular cloning: a laboratory manual, 1989, 2-е изд., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк.
33. Schalm и др. The mastitis complex-A briea summary, в кн.: «Bovine Mastitis», 1971, изд-во Lea & Febiger, Филадельфия, сс.1-3.
34. Trends in Biotechnology 7, 1989, сс.283-287.
35. Van der Burg, S.H., W.M.Kast, R.E.M.Toes, R.Offringa, C.J.M.Melief, Спосоюз for selection and producing T cell peptide epitops and vaccines incorporating said selected epitops. International Patent Application WO 97/41440.
36. Vriend G., V.G.H.Eijsink, Prediction and analysis of structure, stability and unfolding of bacillus neutral proteases. J. Computer-Aided Mol. Design 7, 1993, cc.367-396.
37. Wren B.W. Microbial genome analysis: insights into virulence, host adaptation and evolution. Natural Genetics 1, 2000, cc.30-39.
38. Yeast 8, 1992, cc.423-488.
39. Zadoks R.N., H.G.Allore, H.W.Barkema, O.C.Sampimon, Y.T.Grohn, Y.H.Schukken. Analysis of an Outbreak of Streptococcus uberis mastitis. J. Dairy Sci. 84, 2001, cc.590-599.
40. Zadoks, R.N., B.E.Gillerpie, S.P.Oliver, H.W.Barkema, O.C.Sampimon, Y.H.Schukken. Clinical, epidemiological and molecular characteristics of Streptococcus uberis infections in dairy herds. Epidemiol Infect. 130, 2003, cc.335-349.
41. Zadoks R.N., H.G.Allore, H.W.Barkema, O.C.Sampimon, G.J.Wellenberg, Y.T.Grohn, Y.H.Schukken. Cow- and quarter-level risk factors for Streptococcus uberis and Staphylococcus aureus mastitis. J. Dairy Sci. 84, 2001, cc.2649-2663.


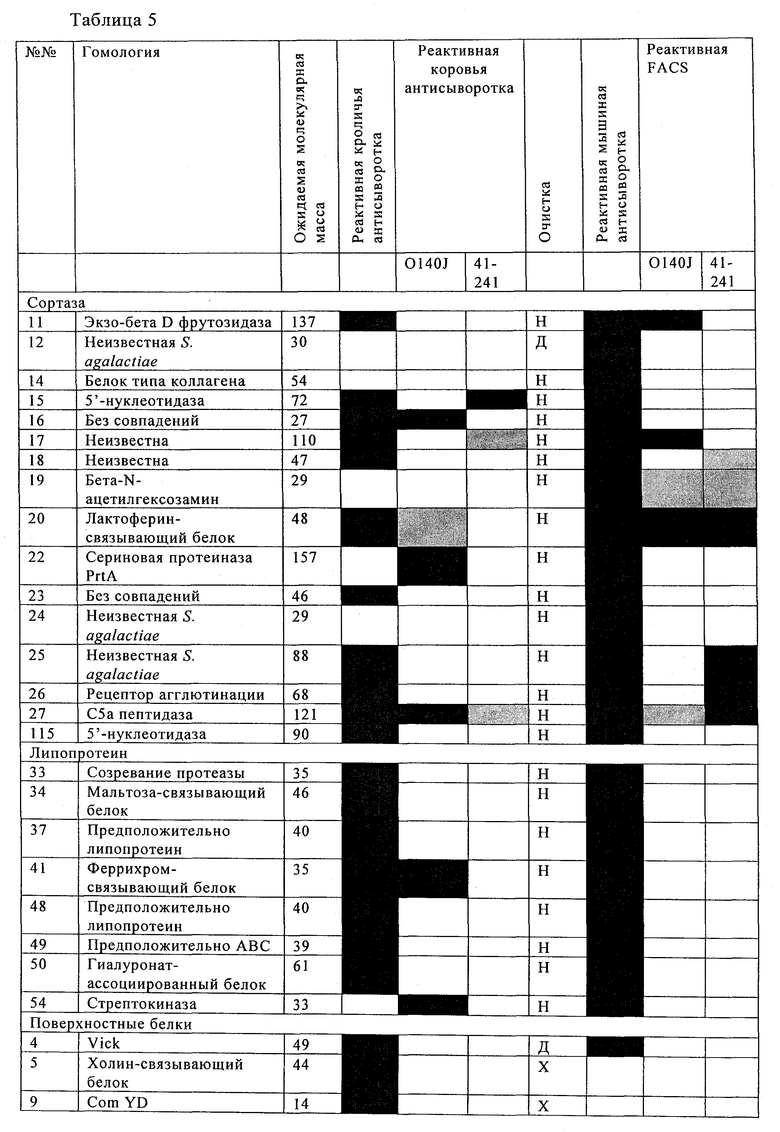
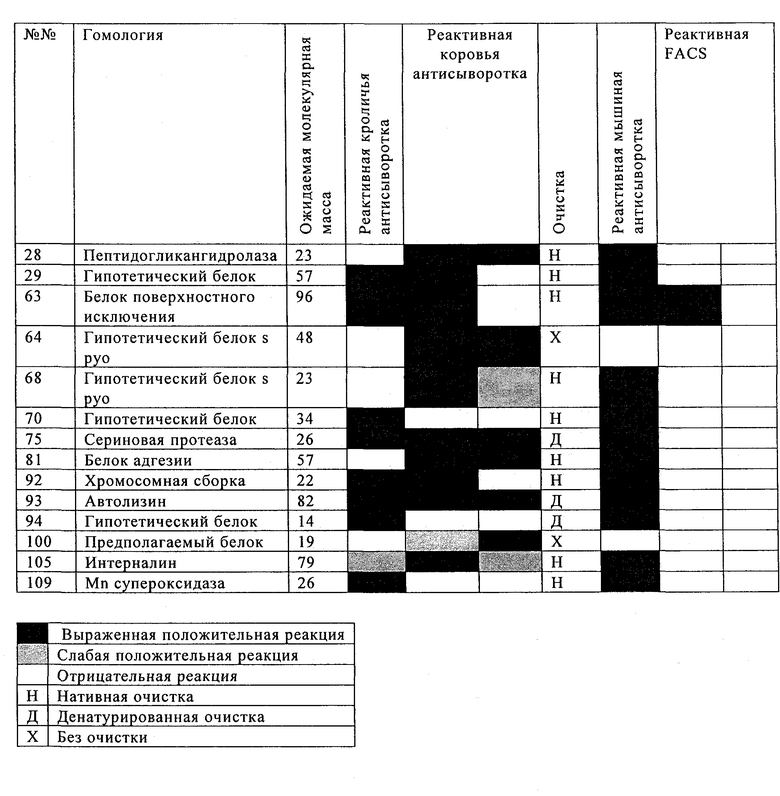
| название | год | авторы | номер документа |
|---|---|---|---|
| МНОГОКОМПОНЕНТНАЯ ИММУНОГЕННАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ ЗАБОЛЕВАНИЯ, ВЫЗВАННОГО β-ГЕМОЛИТИЧЕСКИМИ СТРЕПТОКОККАМИ (БГС) | 2009 |
|
RU2478396C2 |
| МУТАНТНЫЕ АНТИГЕНЫ GAS57 И АНТИТЕЛА ПРОТИВ GAS57 | 2008 |
|
RU2471497C2 |
| МУТАНТНЫЕ ФОРМЫ СТРЕПТОЛИЗИНА О | 2008 |
|
RU2498994C2 |
| СРЕДСТВА И СПОСОБЫ ЛЕЧЕНИЯ ВИРУСА ГЕПАТИТА B | 2017 |
|
RU2740802C2 |
| ИММУНОГЕННЫЙ ПЕПТИД ПРОТИВ СТРЕПТОКОККОВ ГРУППЫ А | 2018 |
|
RU2775621C2 |
| НОВАЯ ВАКЦИНА ПРОТИВ HAEMOPHILUS PARASUIS | 2020 |
|
RU2822516C1 |
| СЛИТЫЕ ПОЛИПЕПТИДЫ И ВАКЦИНЫ | 2014 |
|
RU2699006C2 |
| НОВАЯ ВАКЦИНА ПРОТИВ HEAMOPHILUS PARASUIS | 2020 |
|
RU2833948C1 |
| Иммунногенный слитый белок | 2016 |
|
RU2757426C2 |
| ИММУНОБИОЛОГИЧЕСКОЕ СРЕДСТВО НА ОСНОВЕ РЕКОМБИНАНТНОГО БЕЛКА ДЛЯ ПРОФИЛАКТИКИ ПНЕВМОКОККОВОЙ ИНФЕКЦИИ | 2024 |
|
RU2823752C1 |

































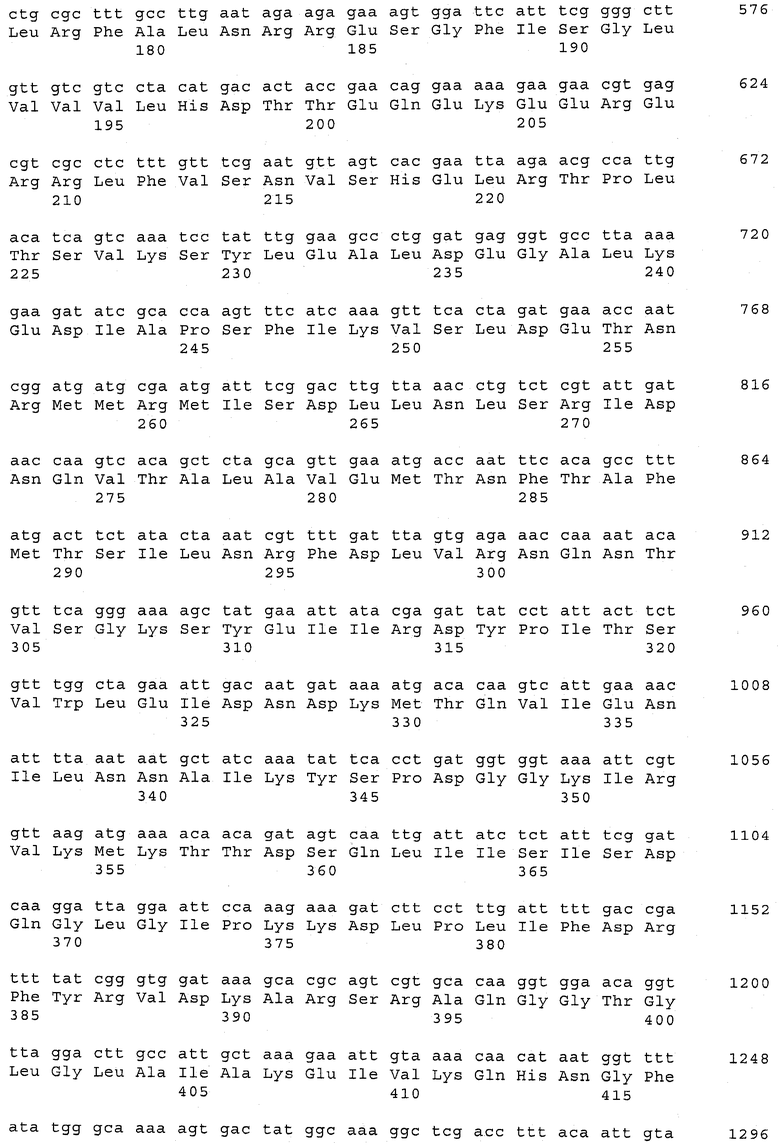



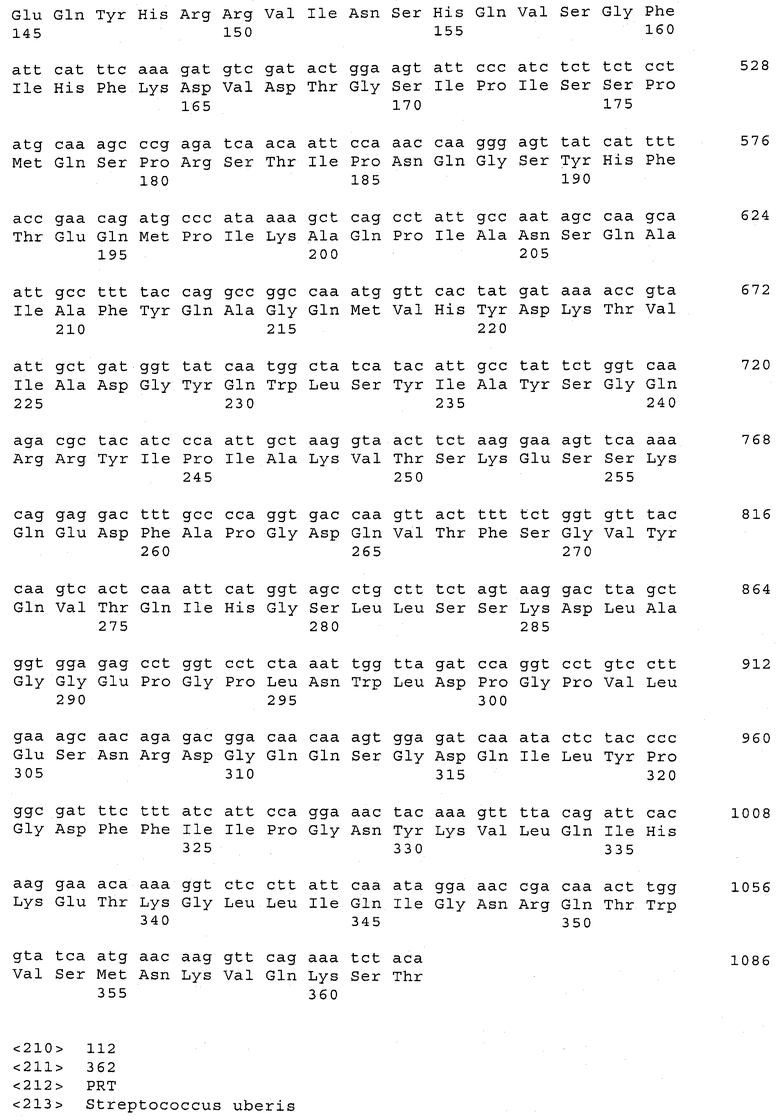




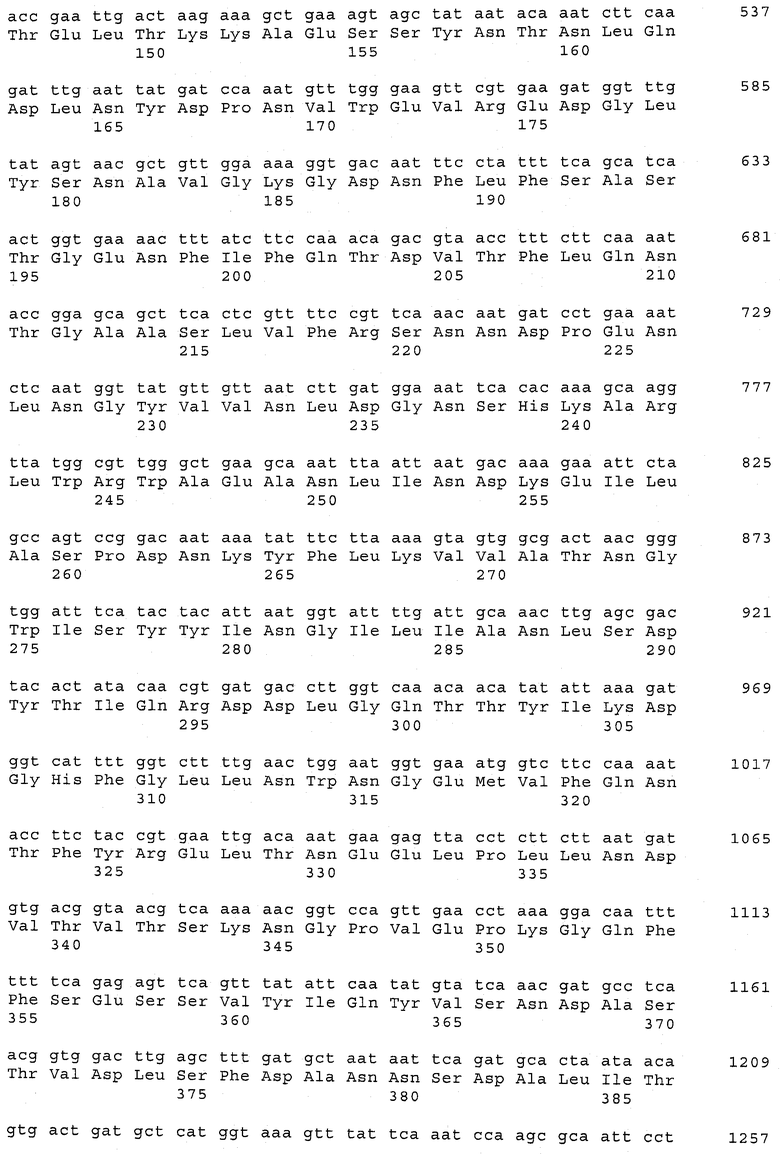
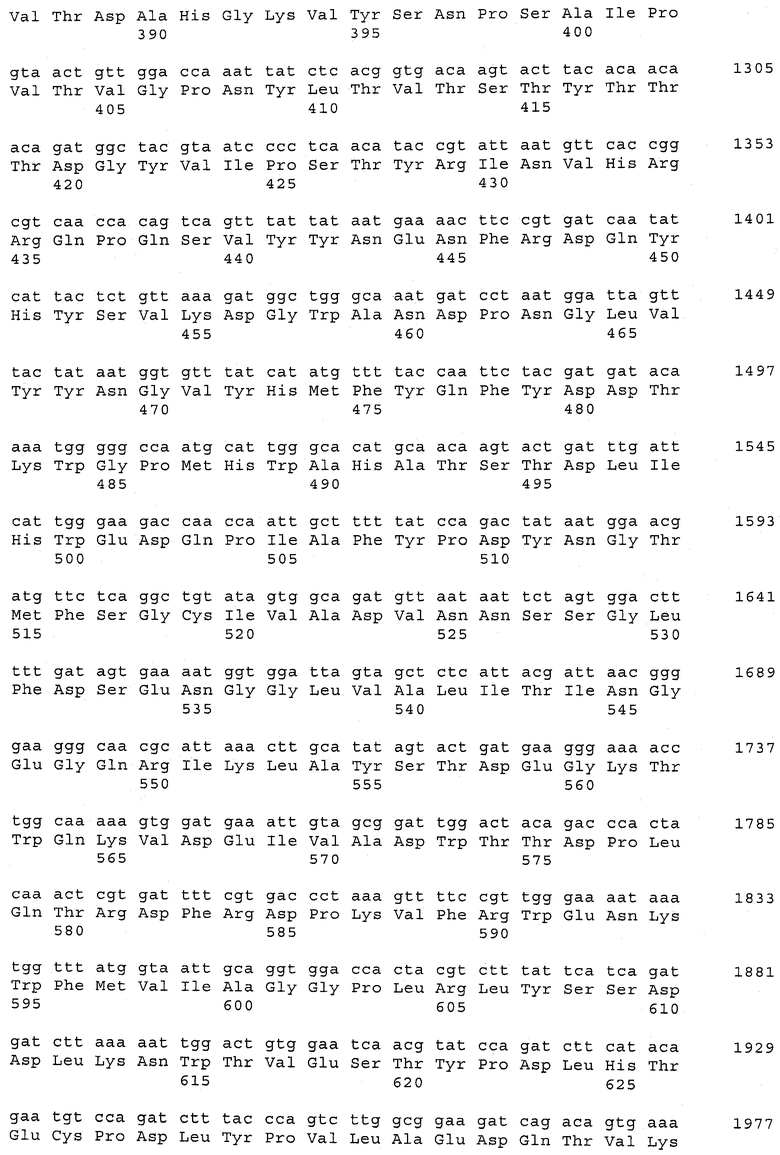

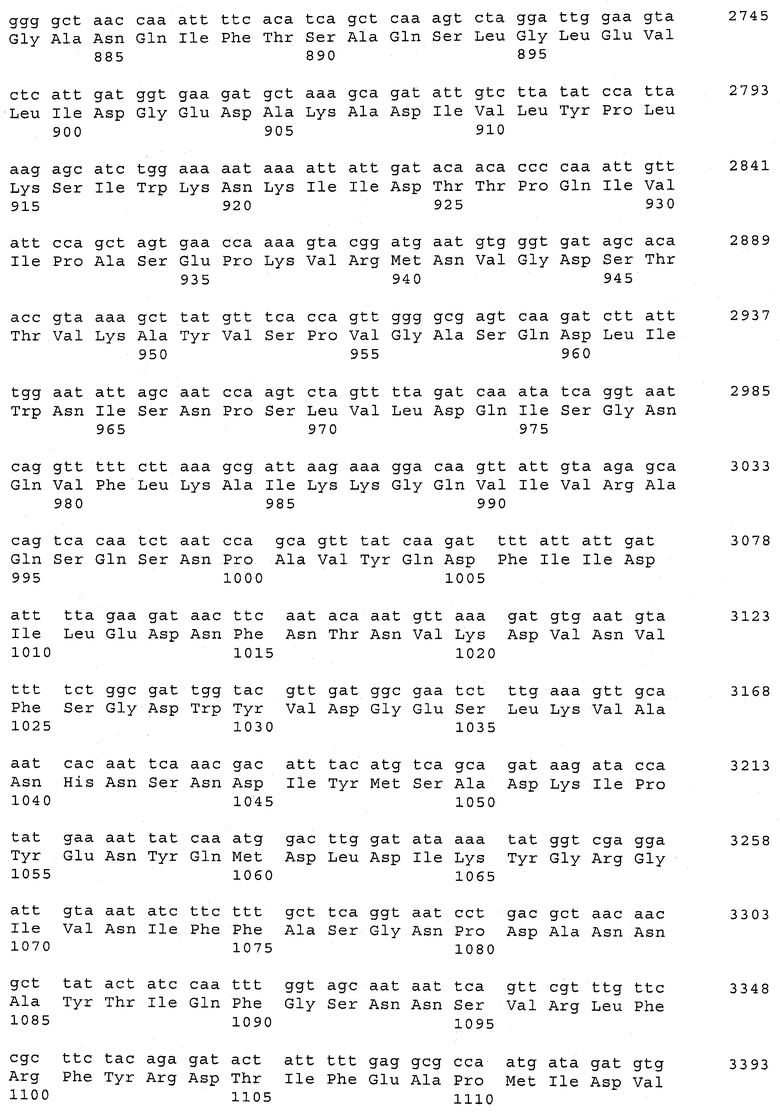





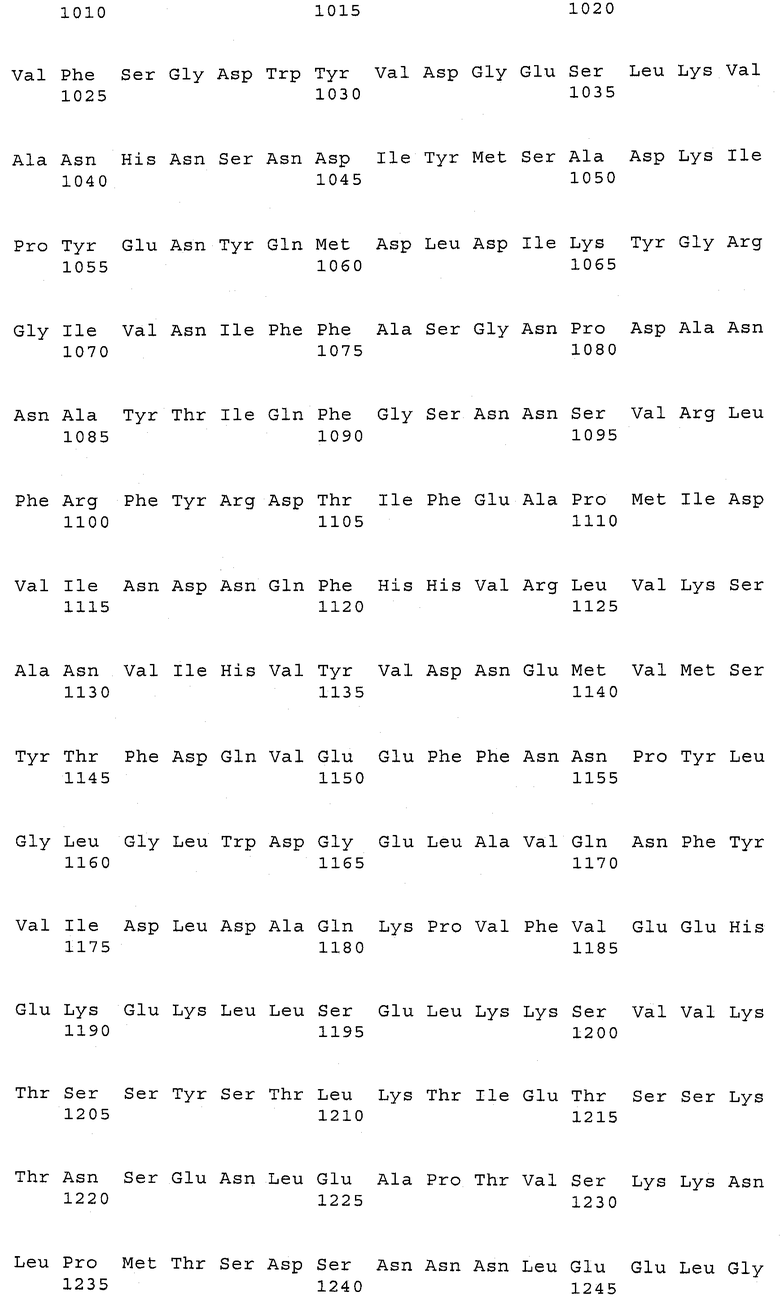




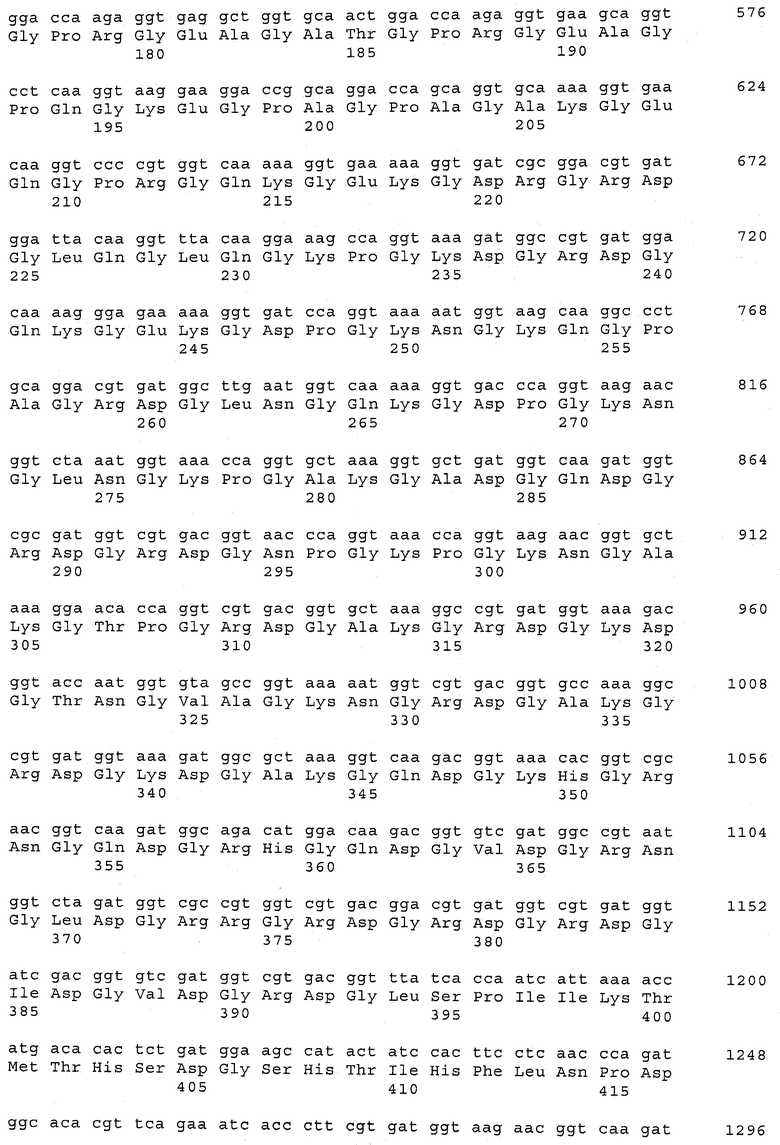




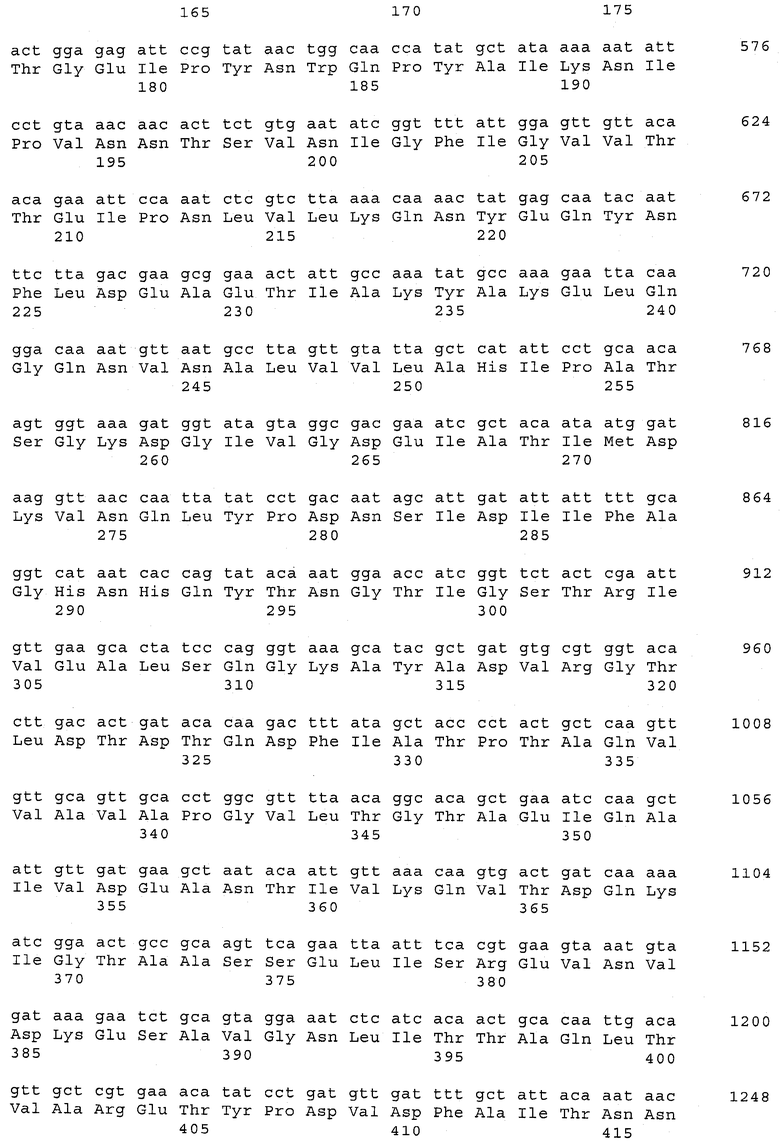
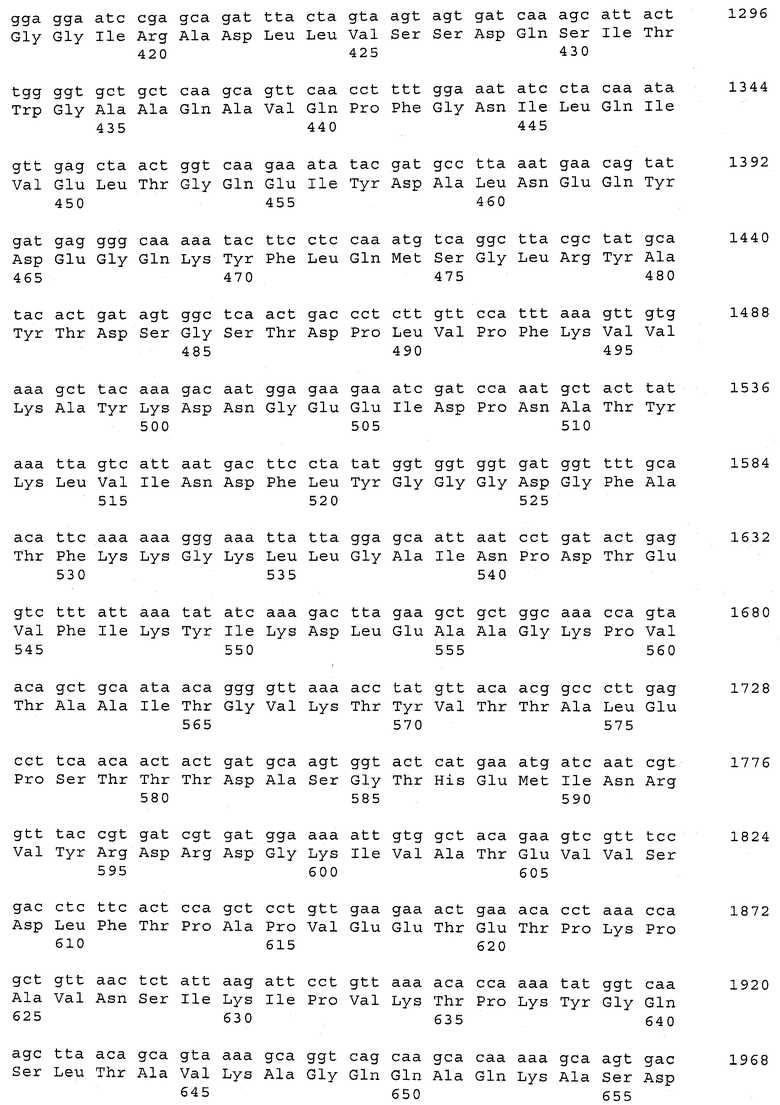




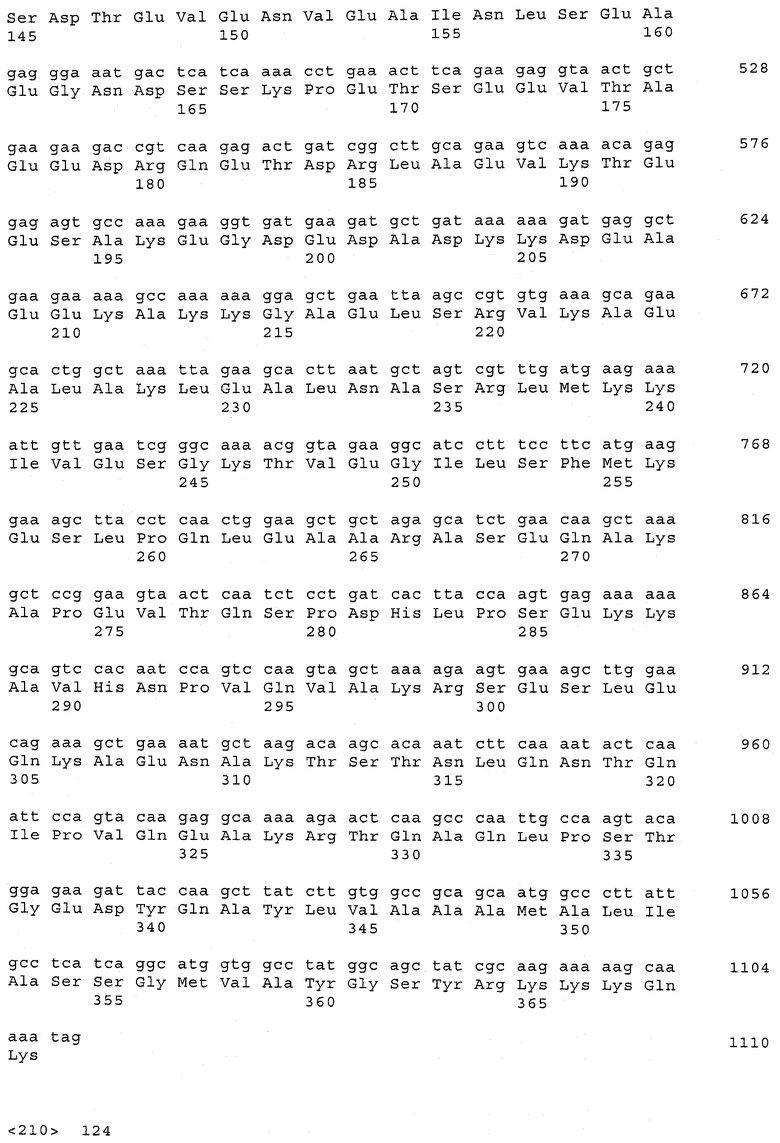



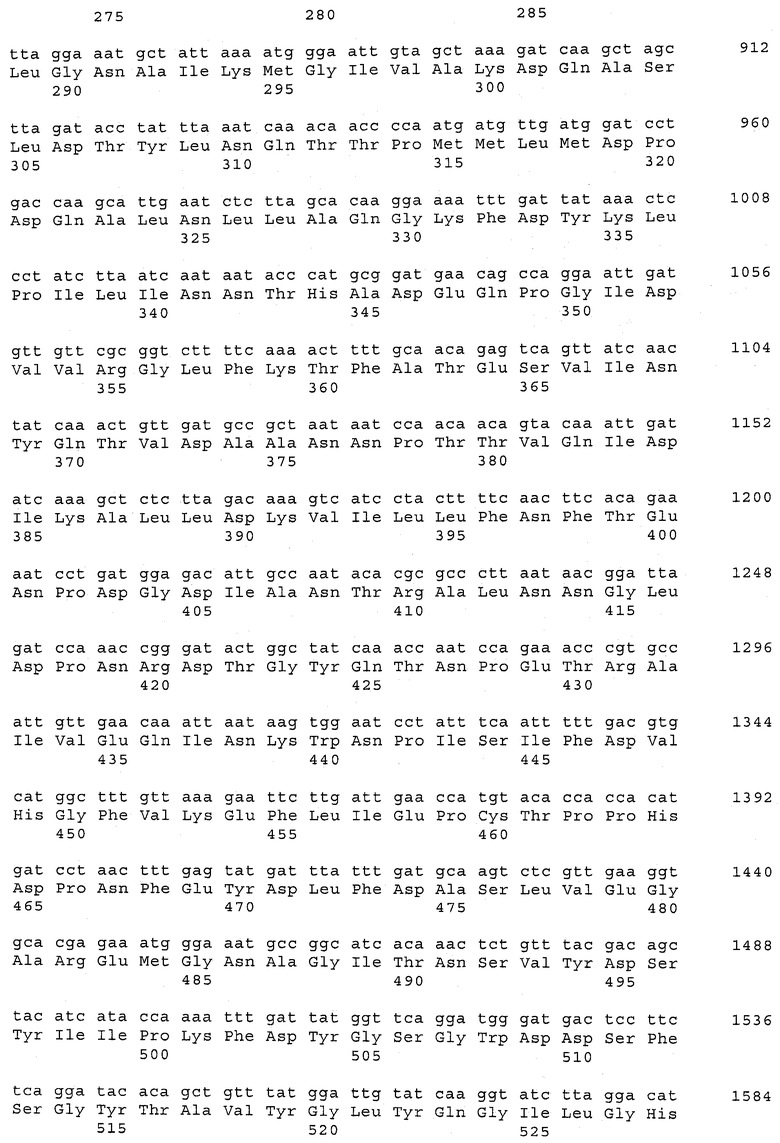
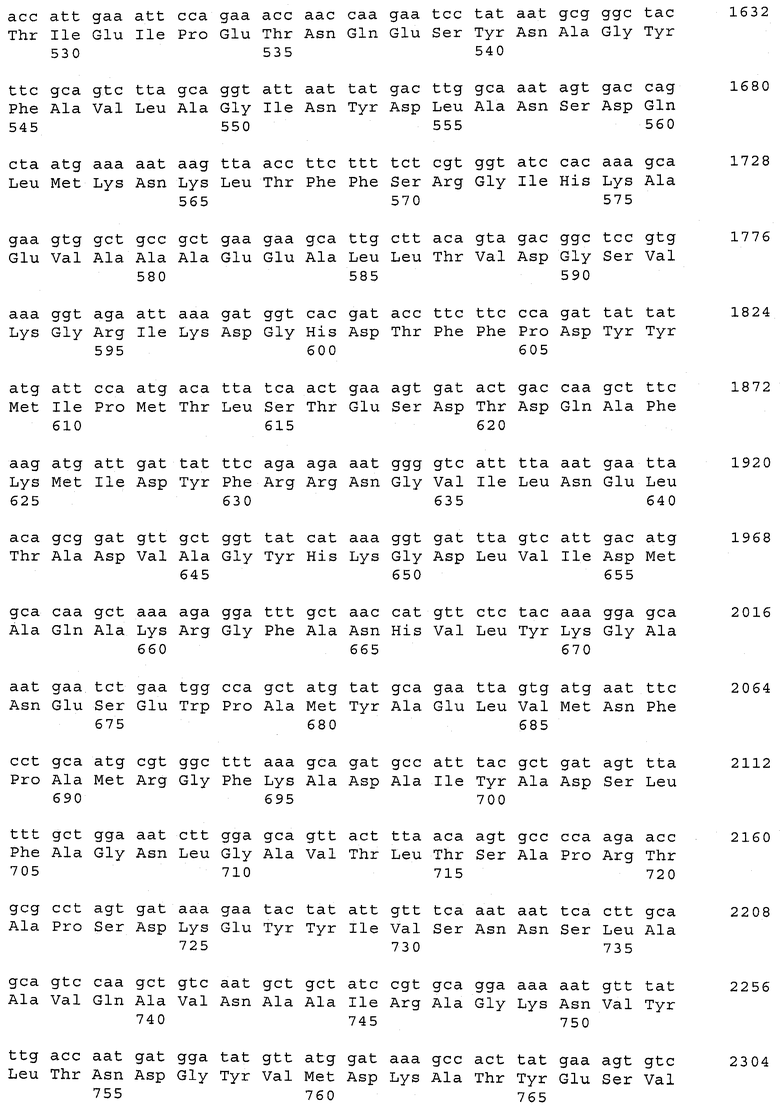
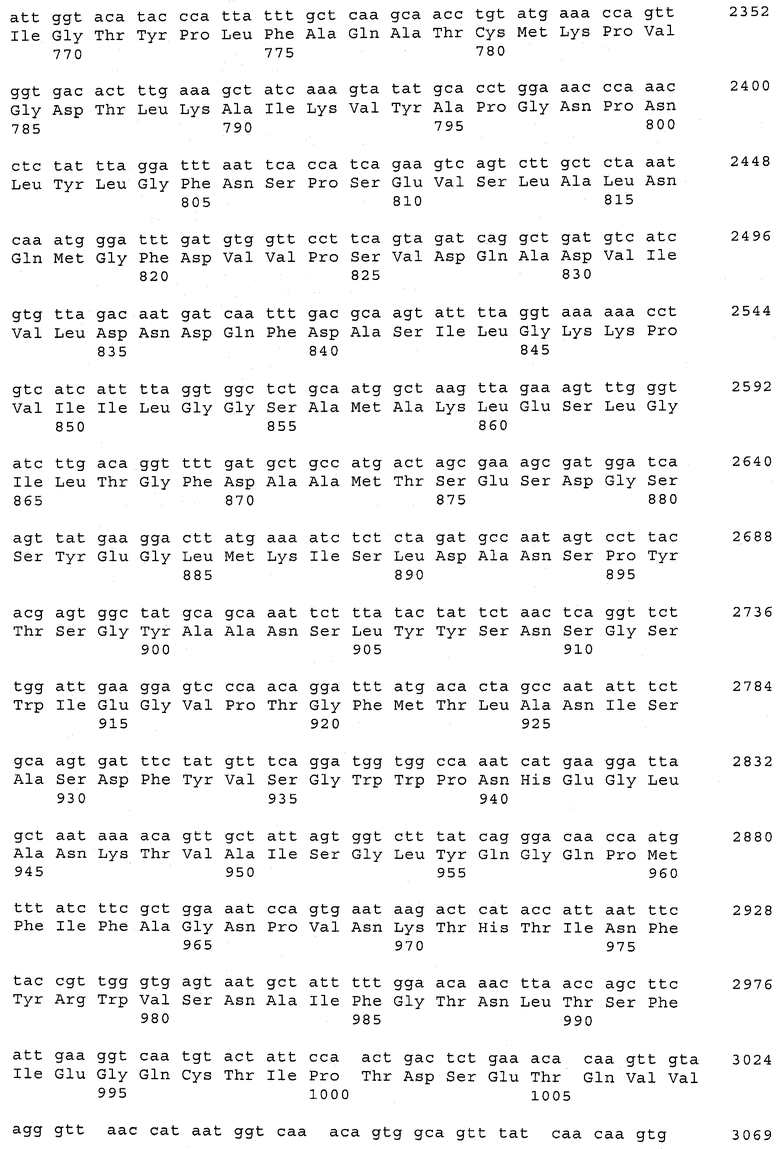





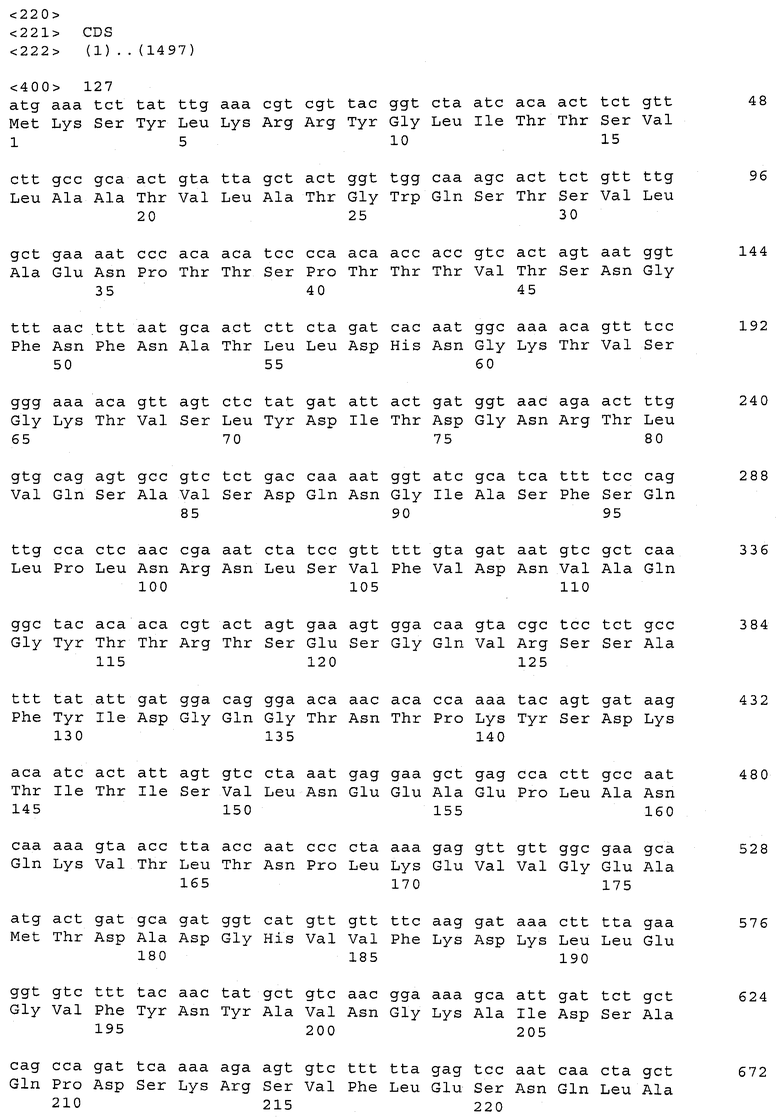
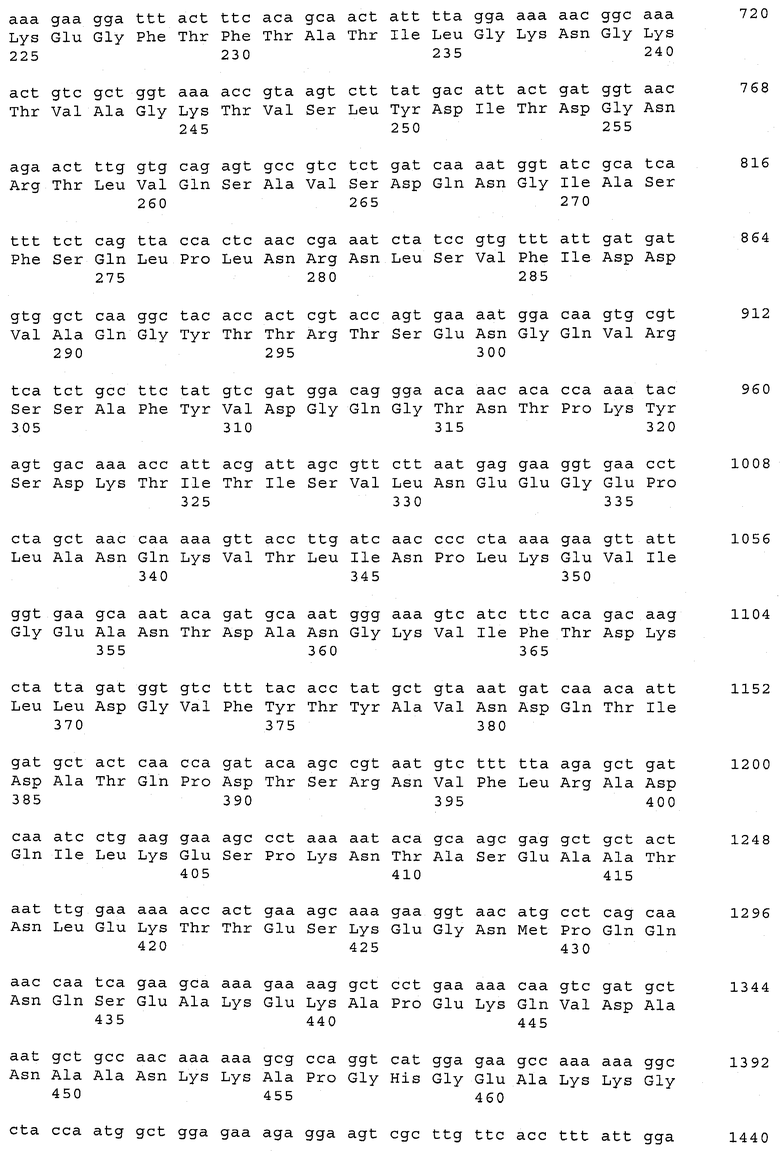






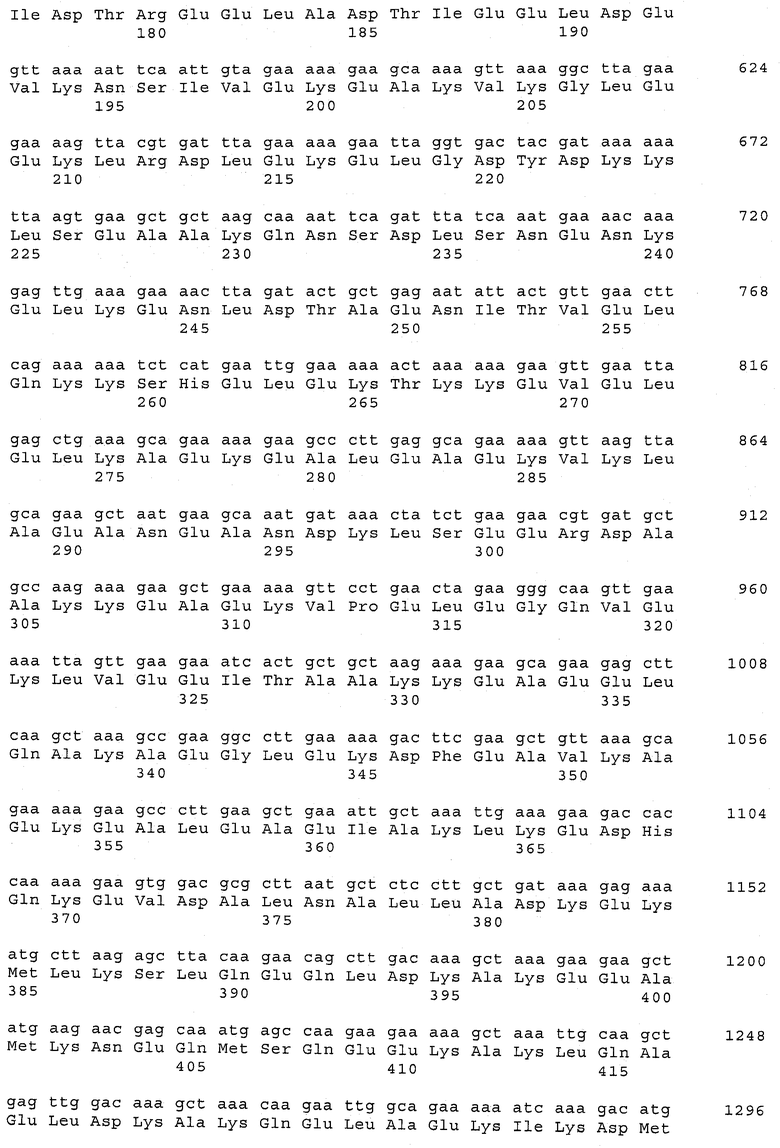










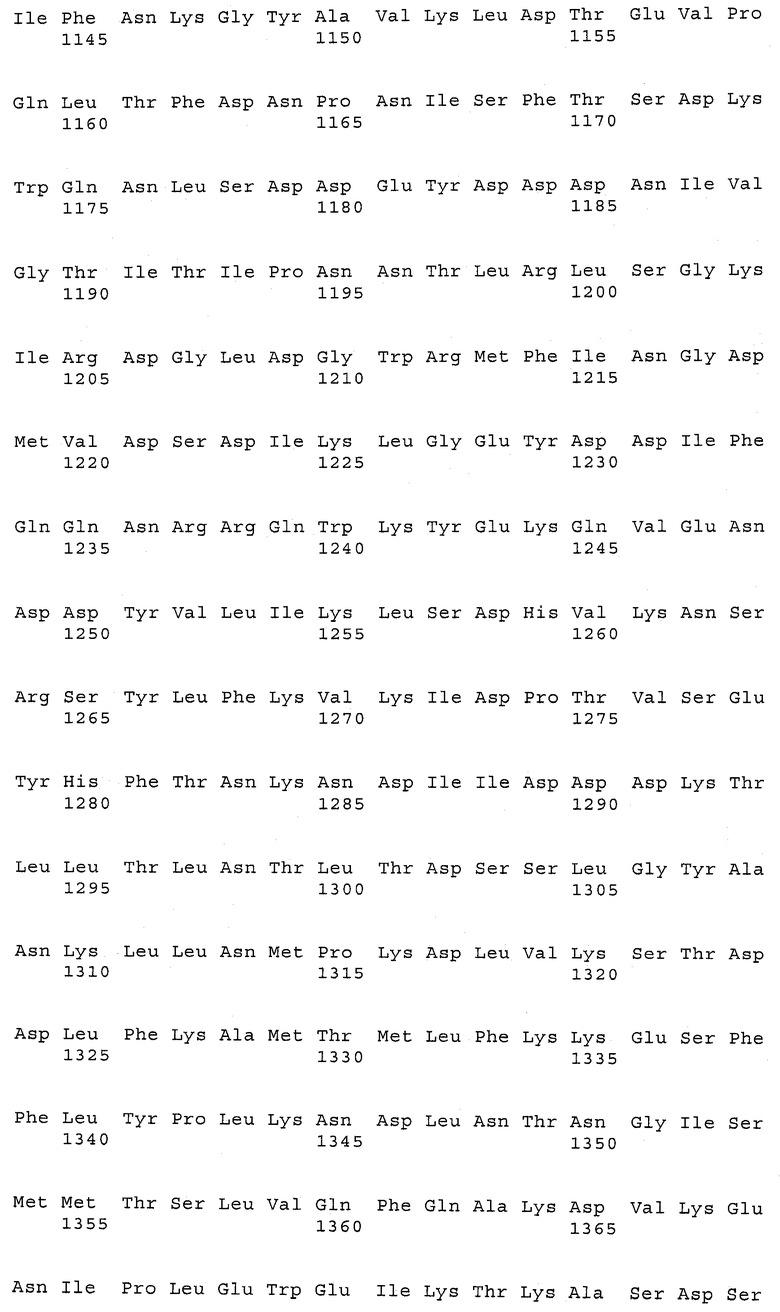








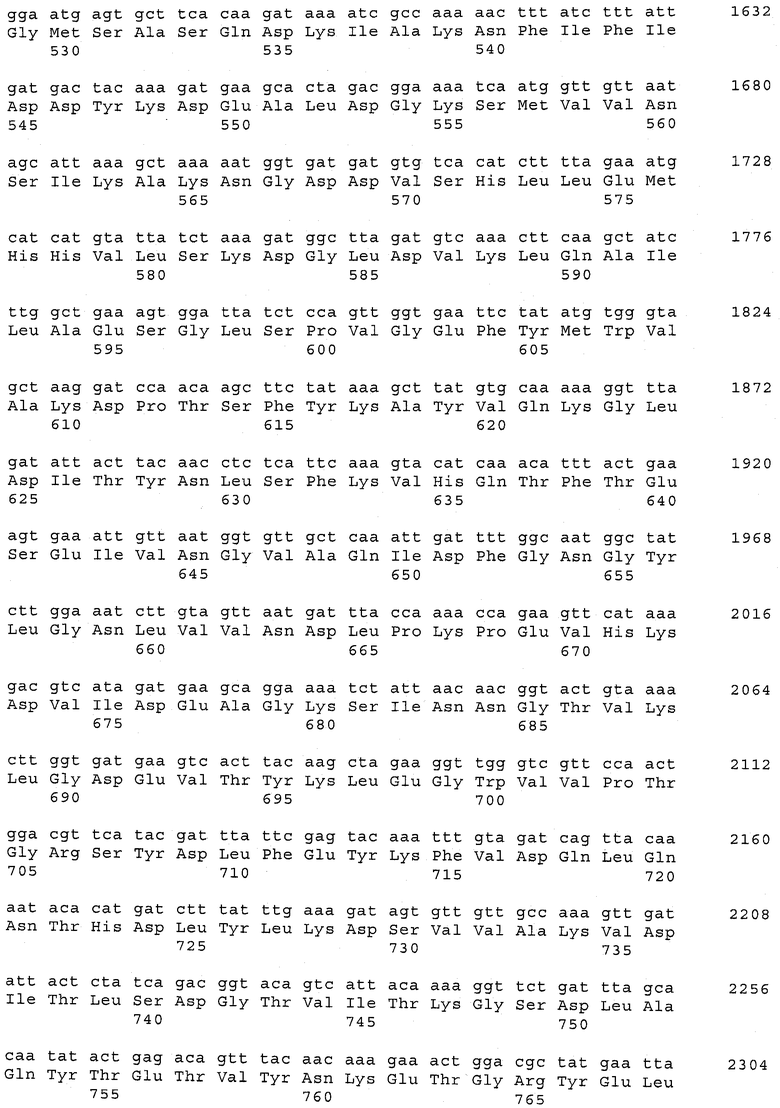





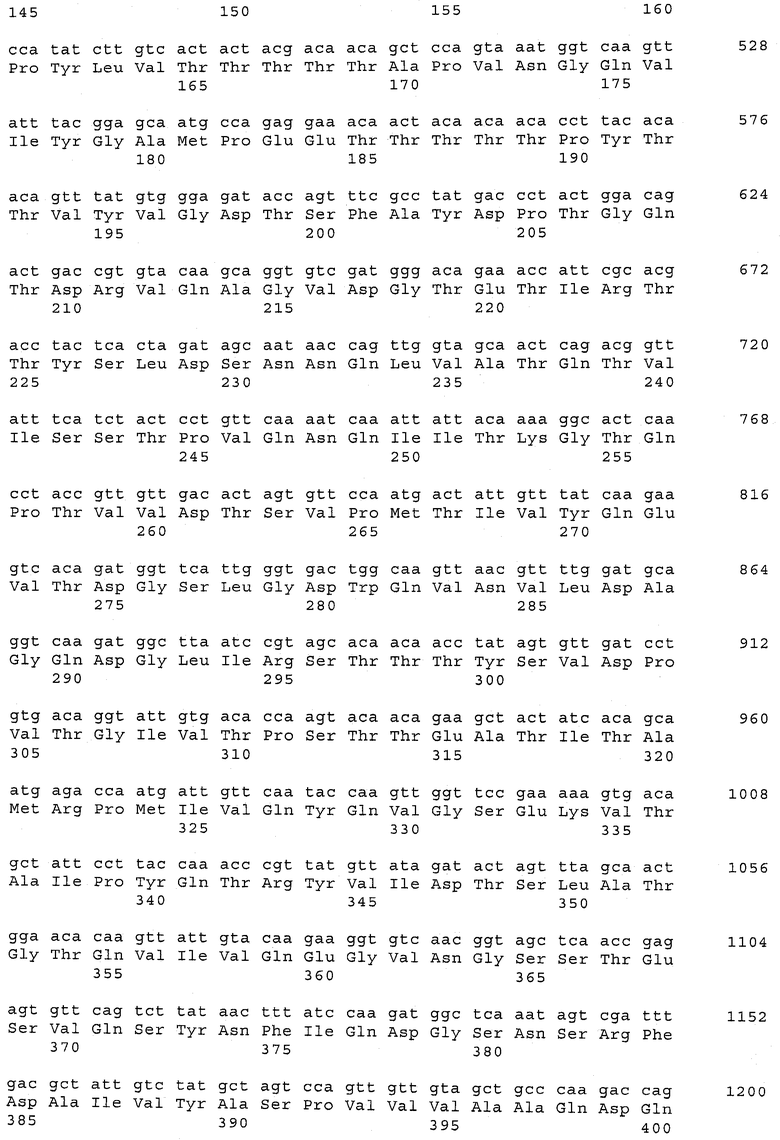
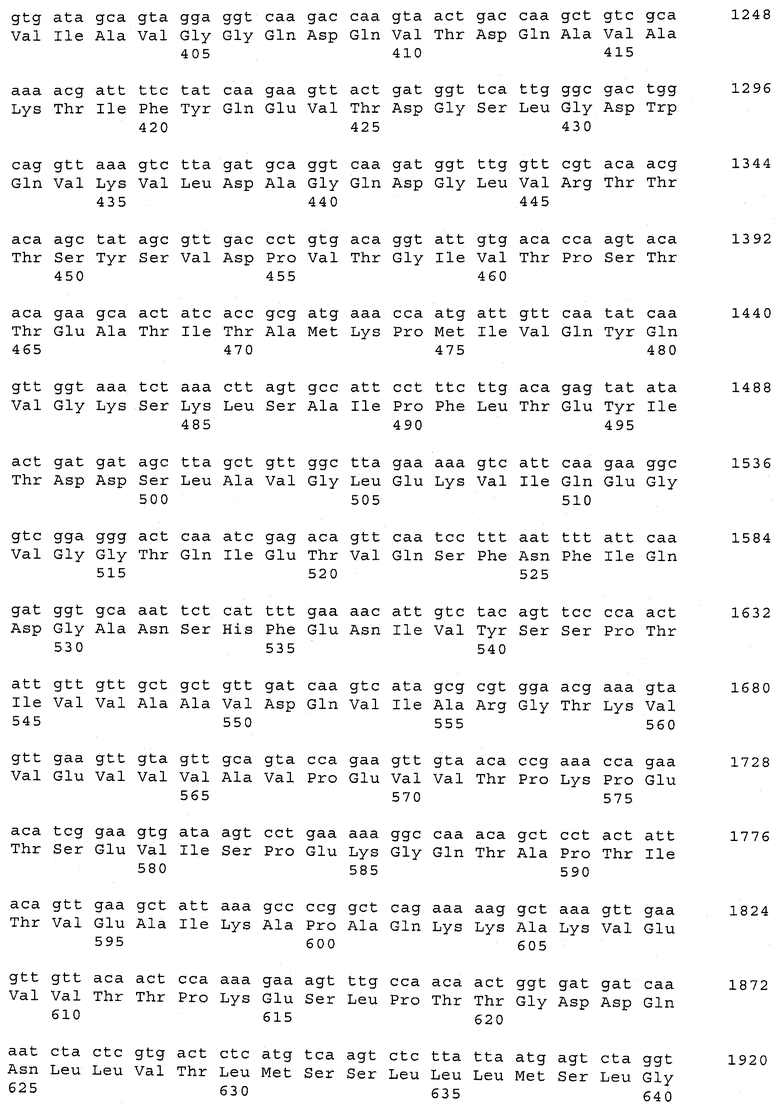




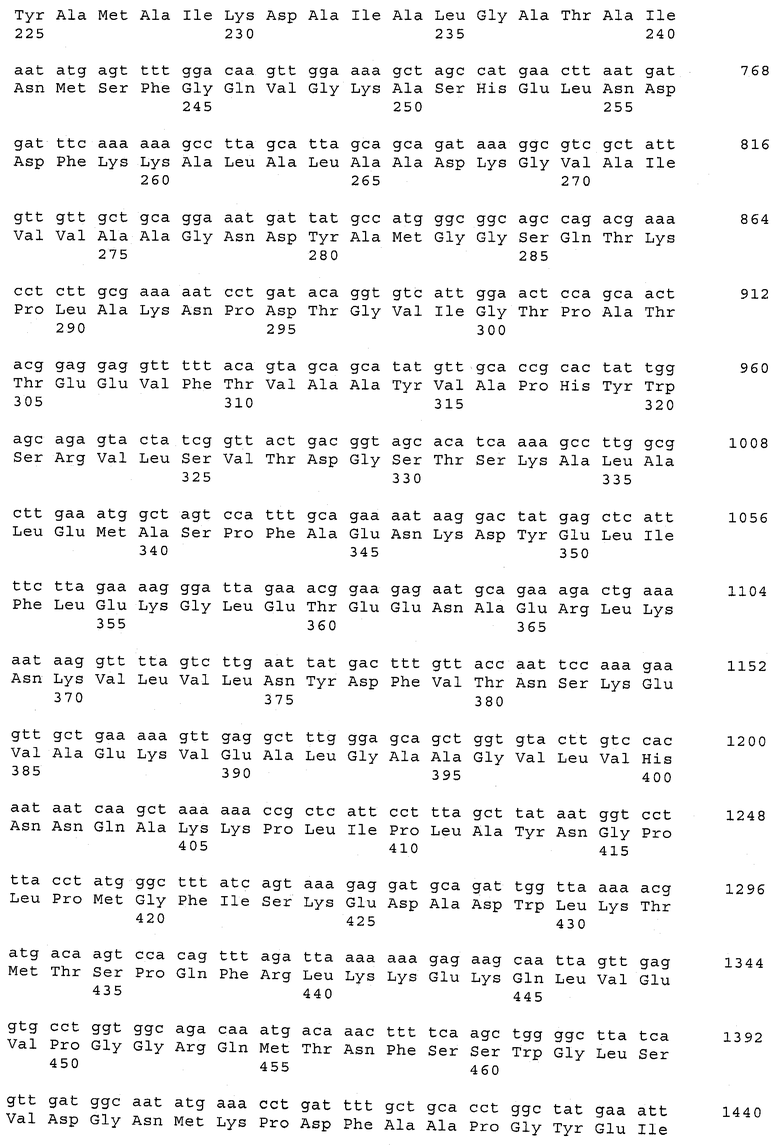

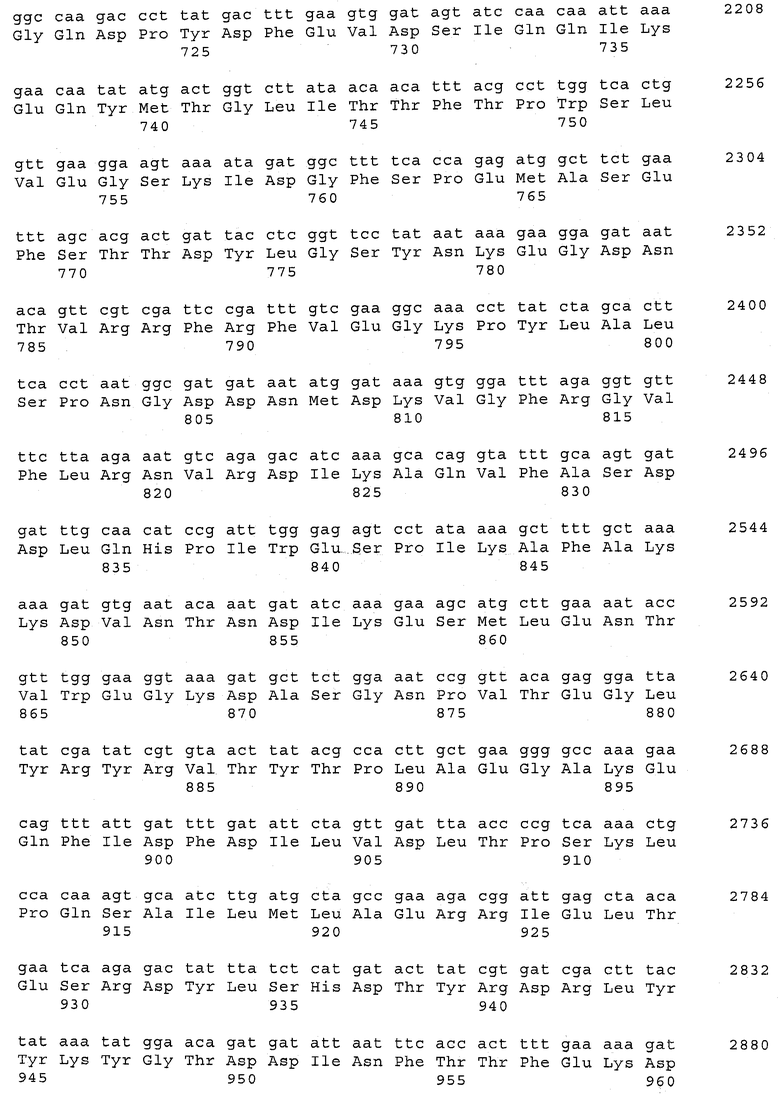






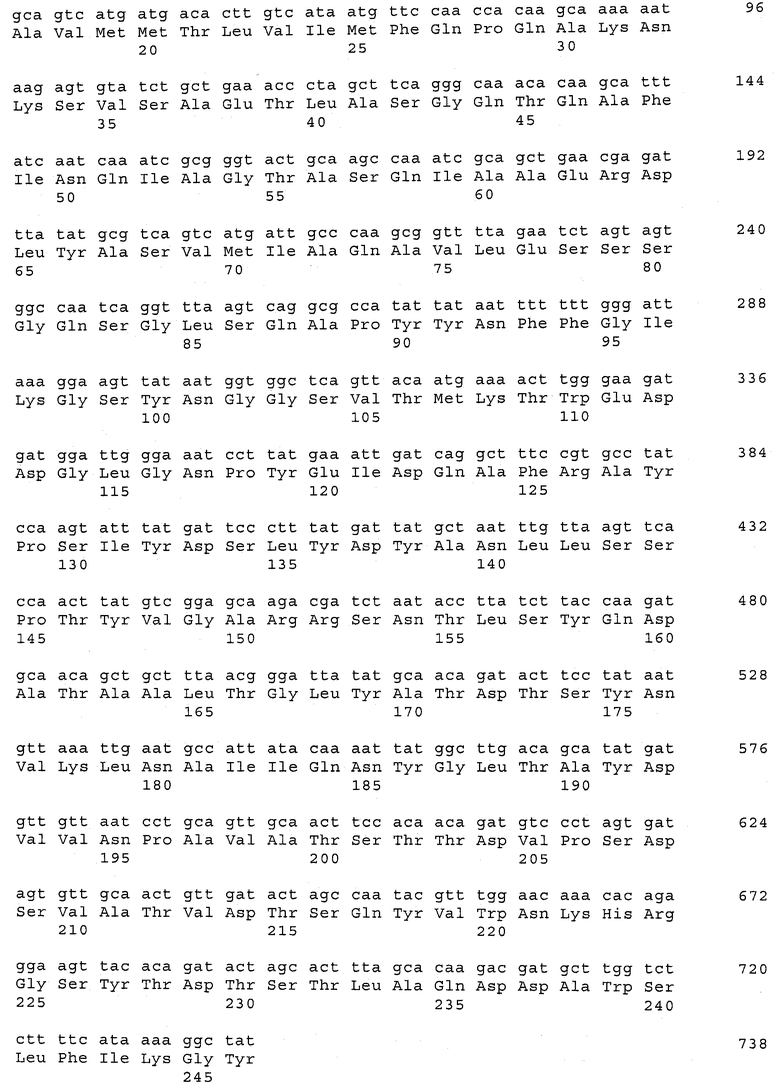


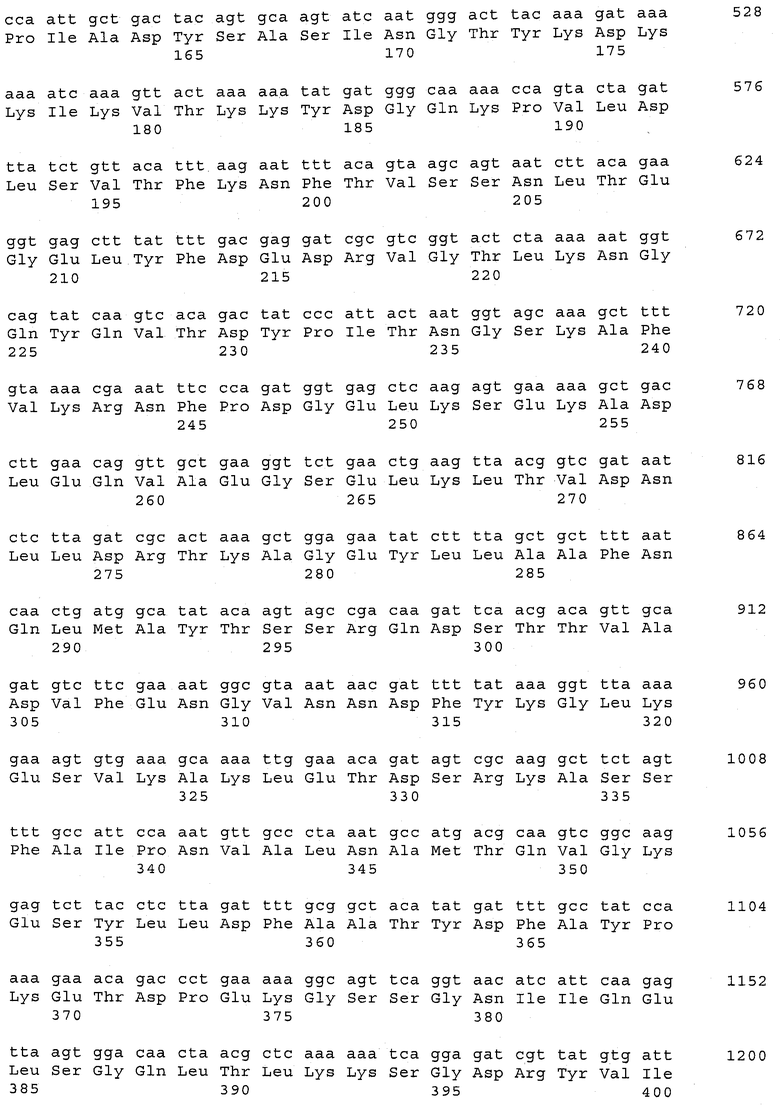






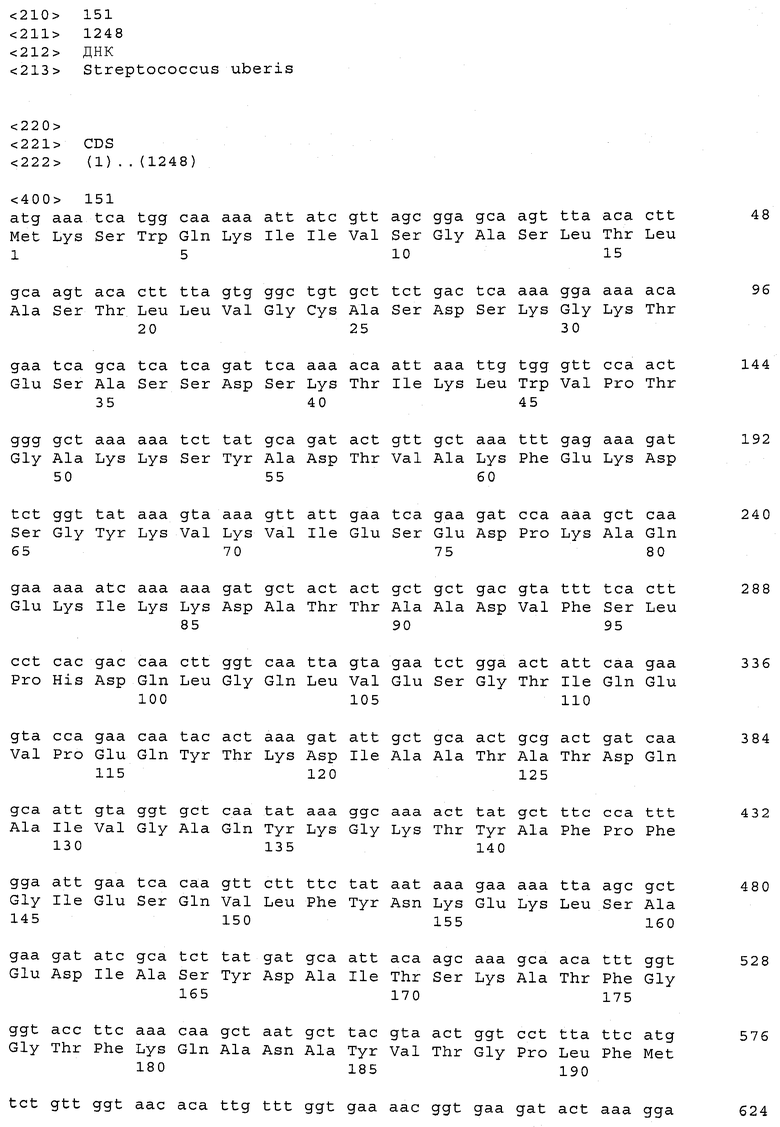





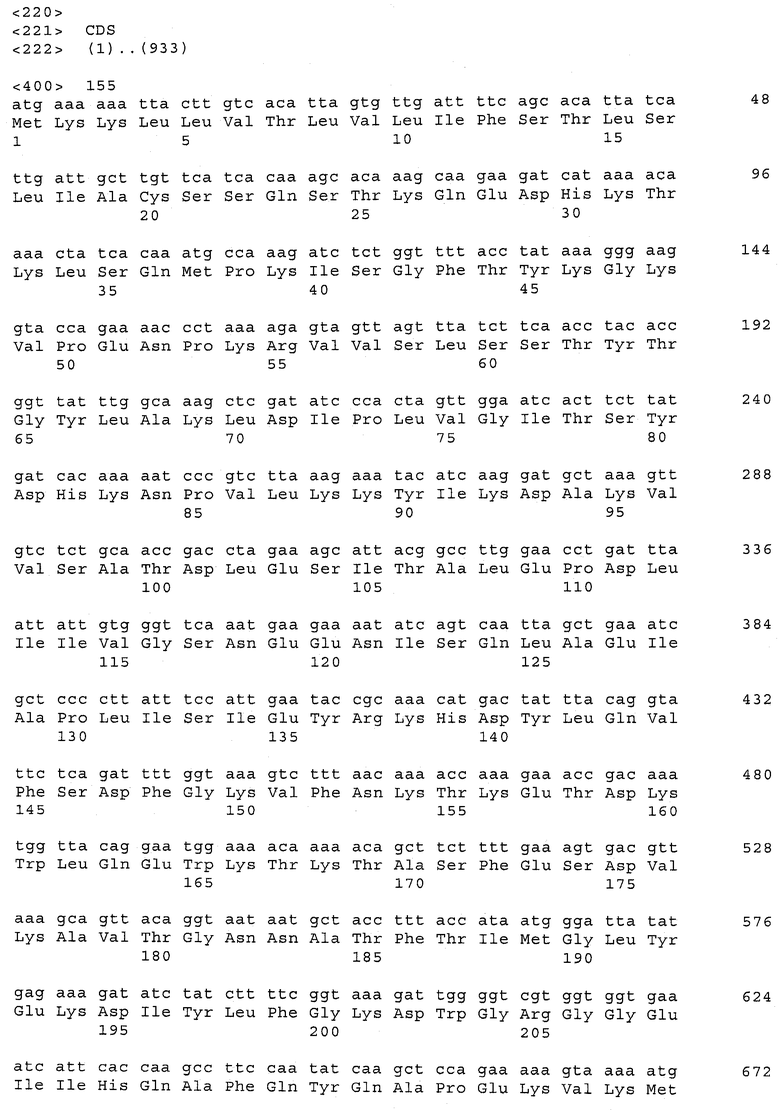


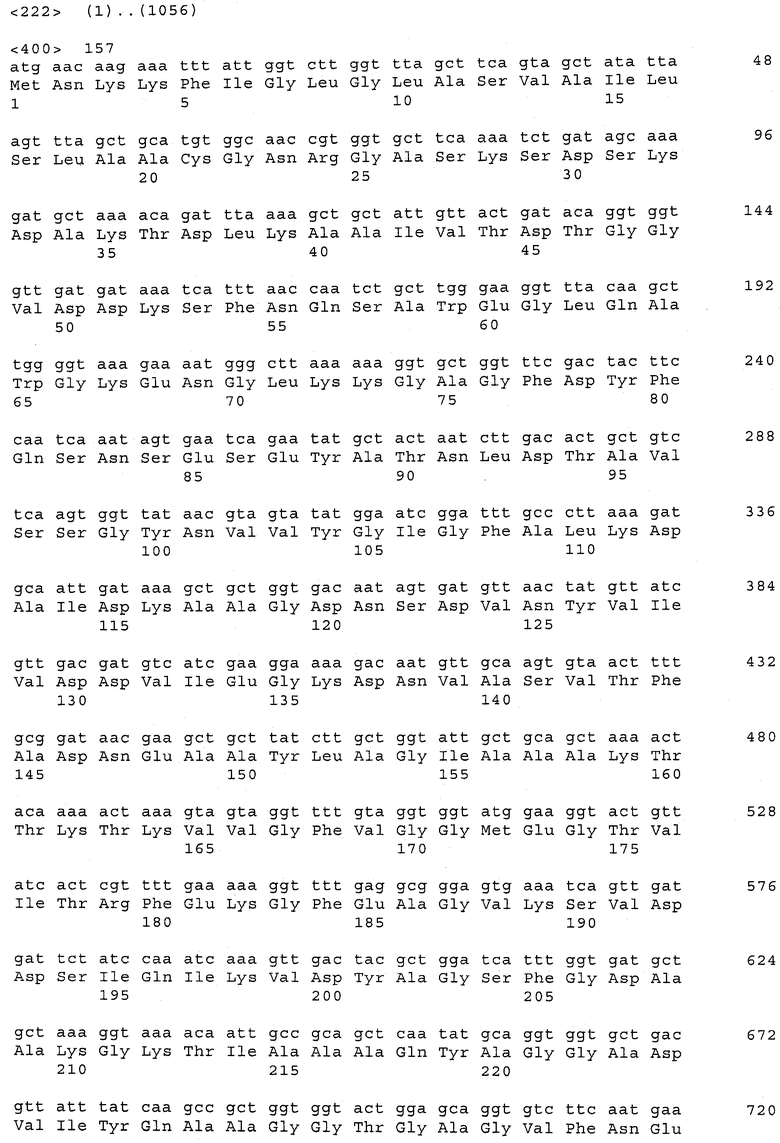






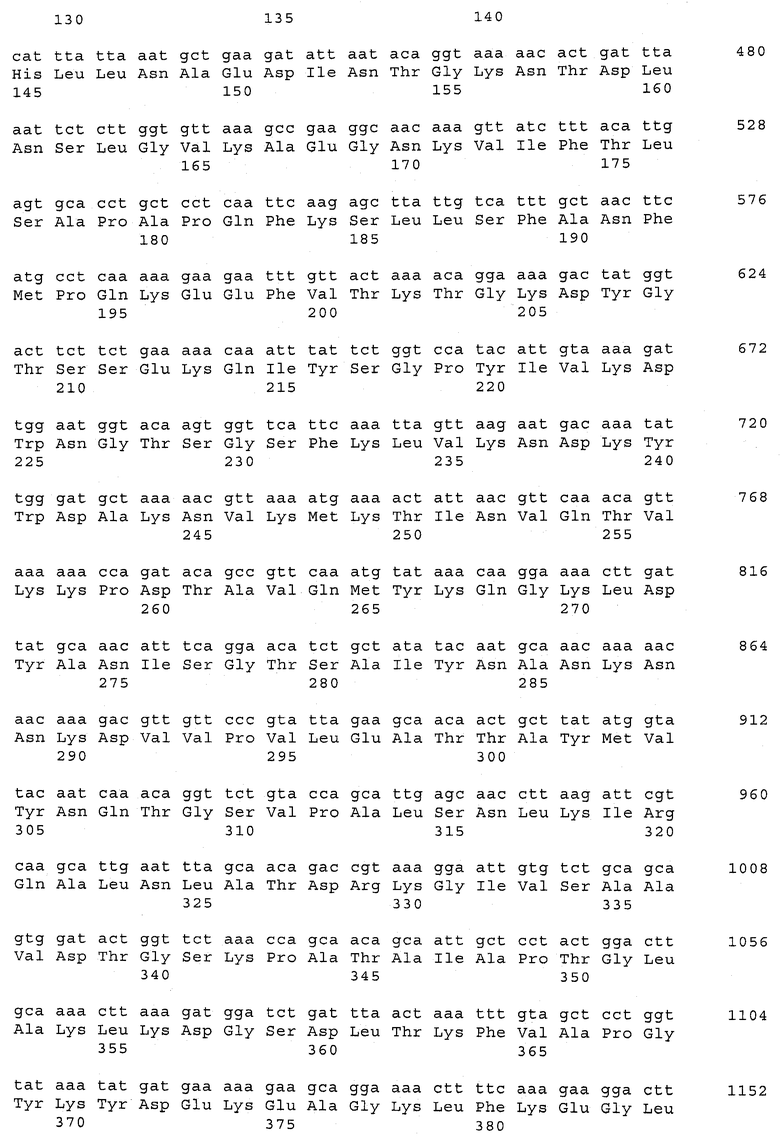






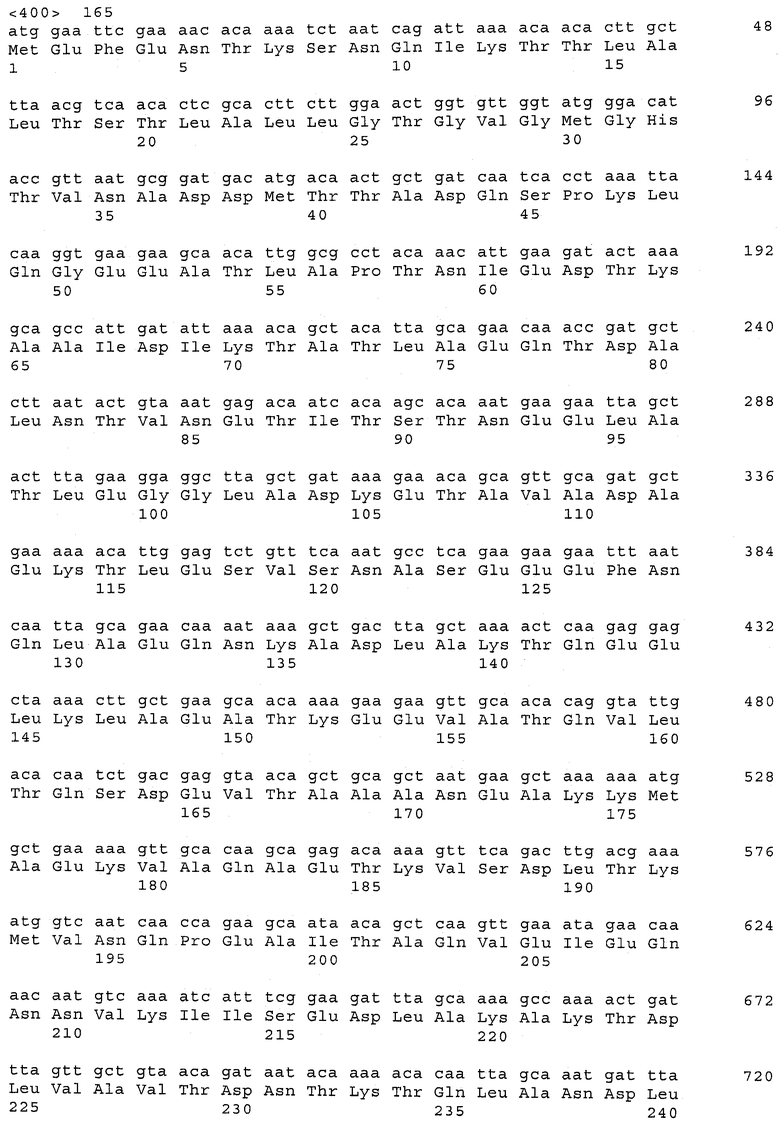
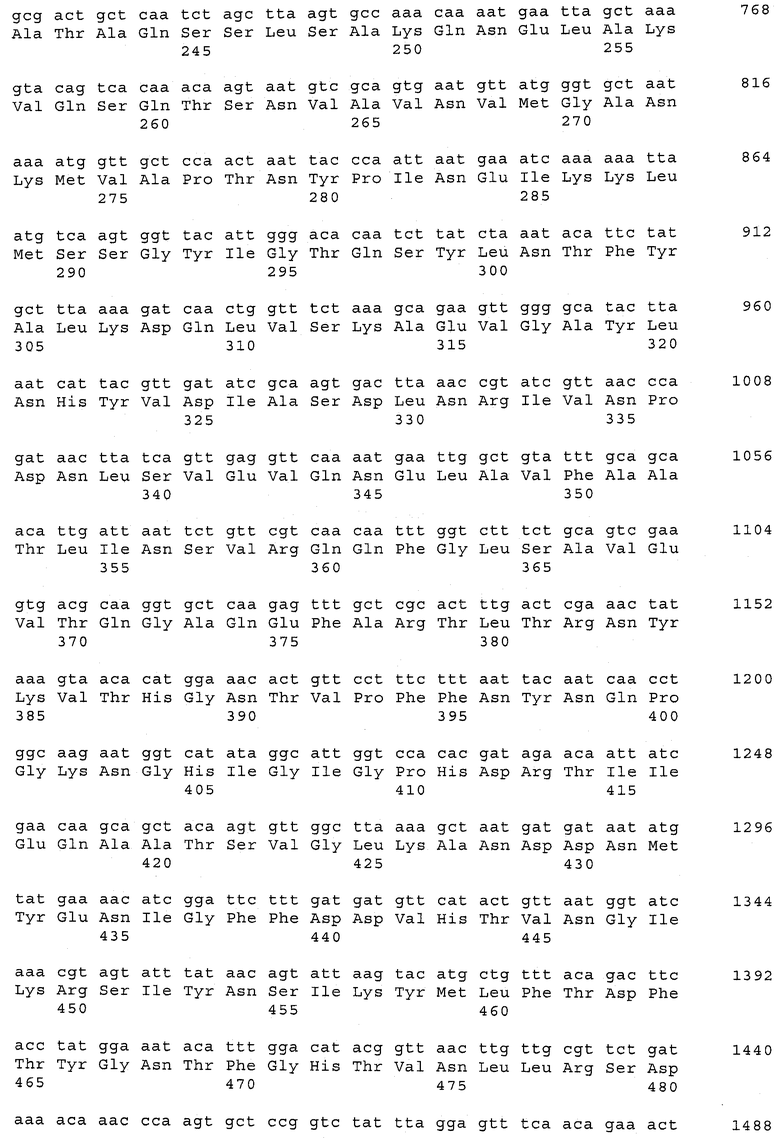






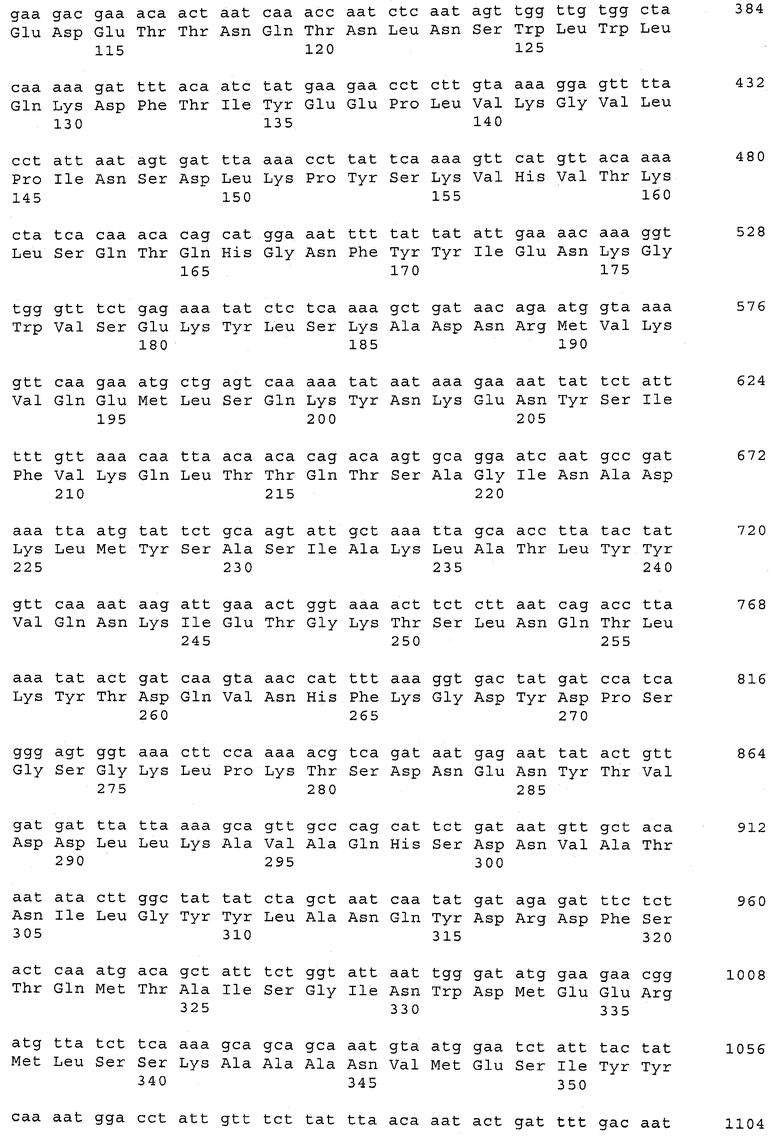










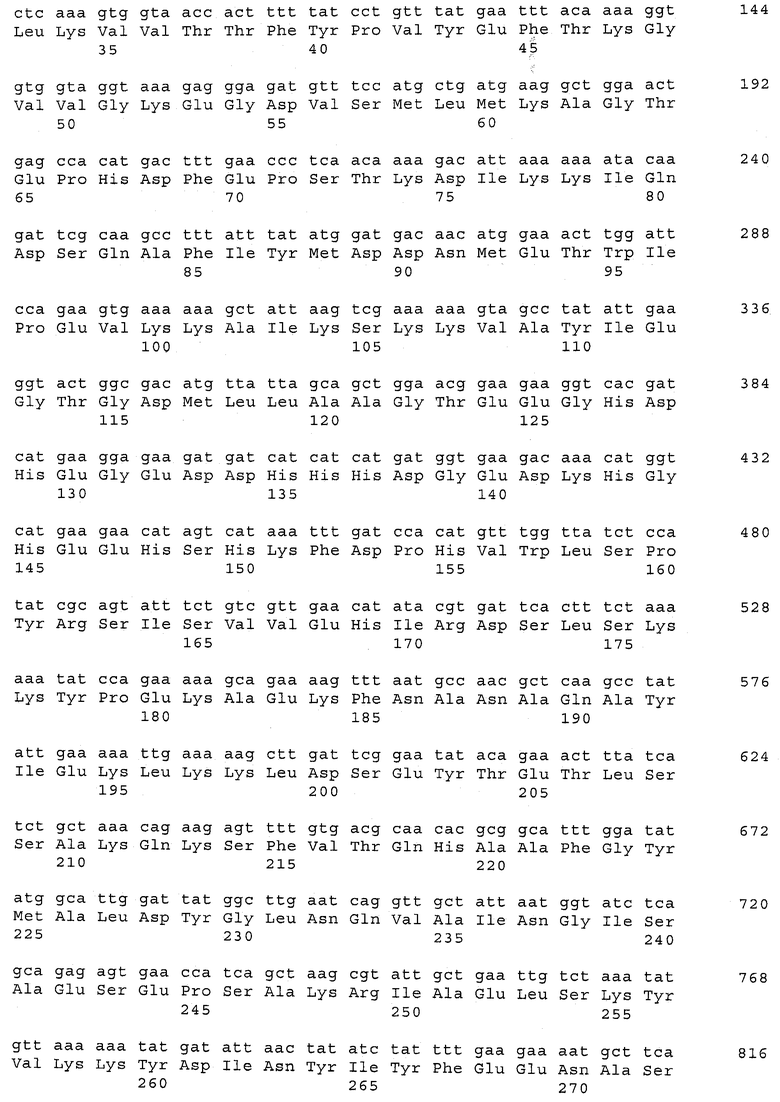
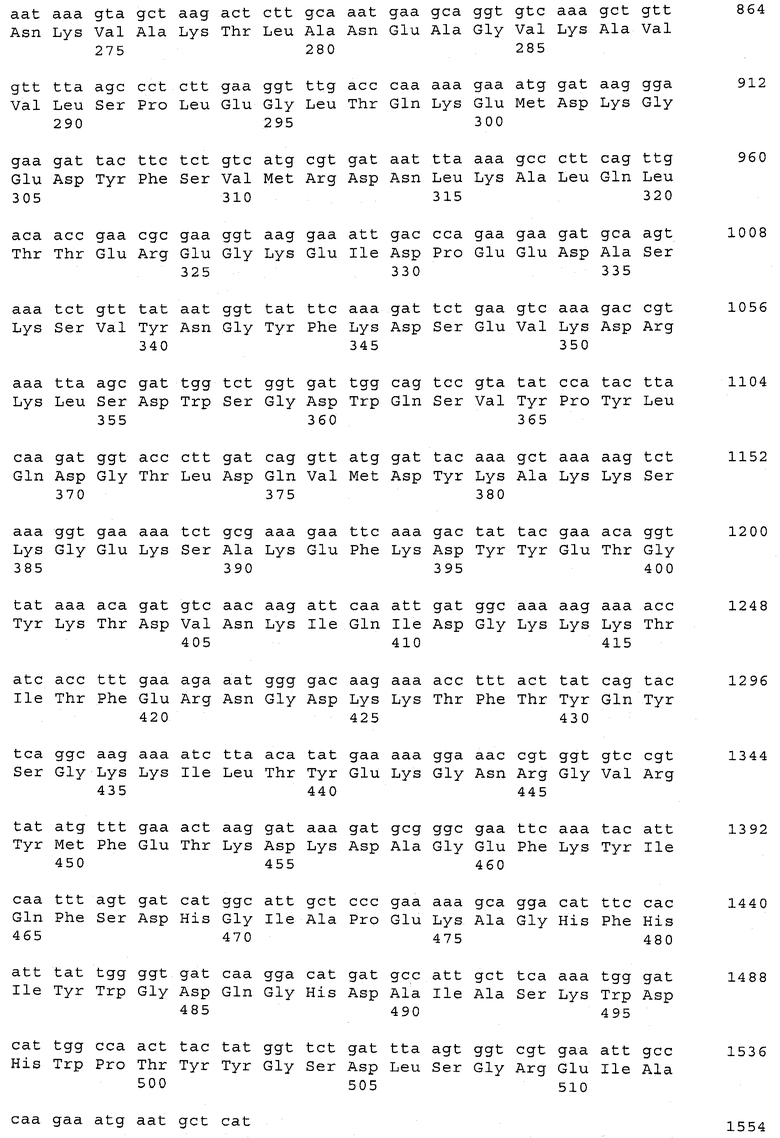













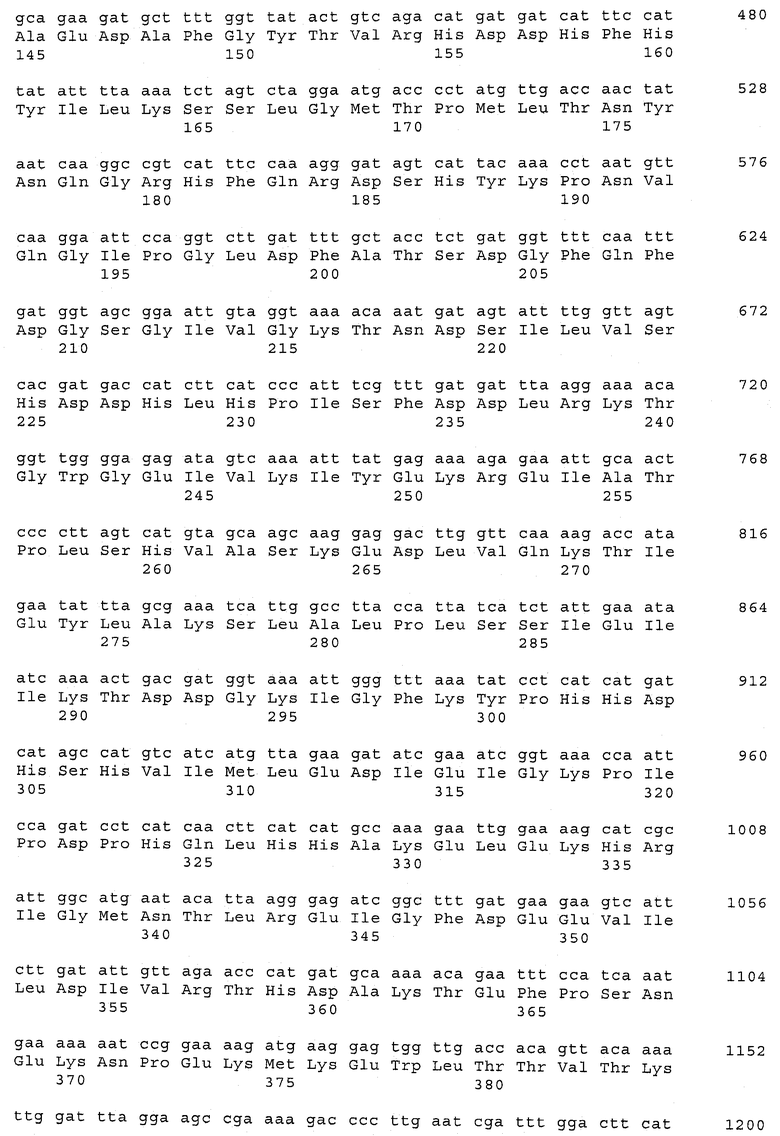
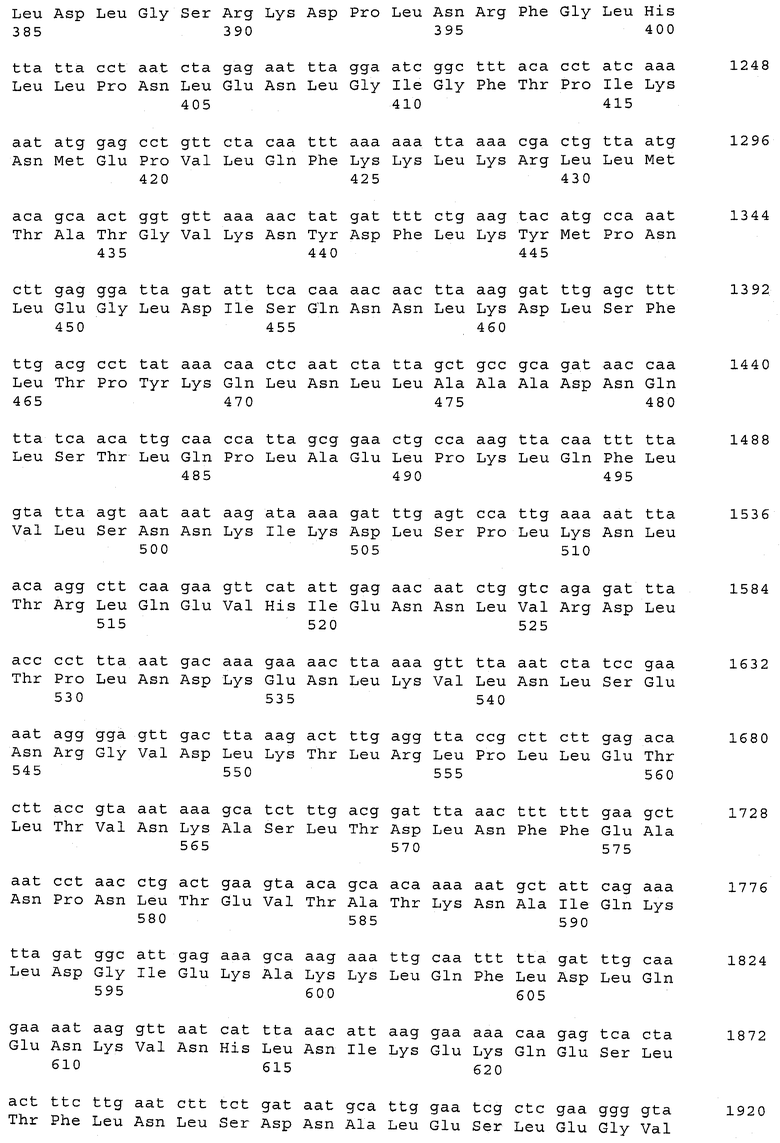






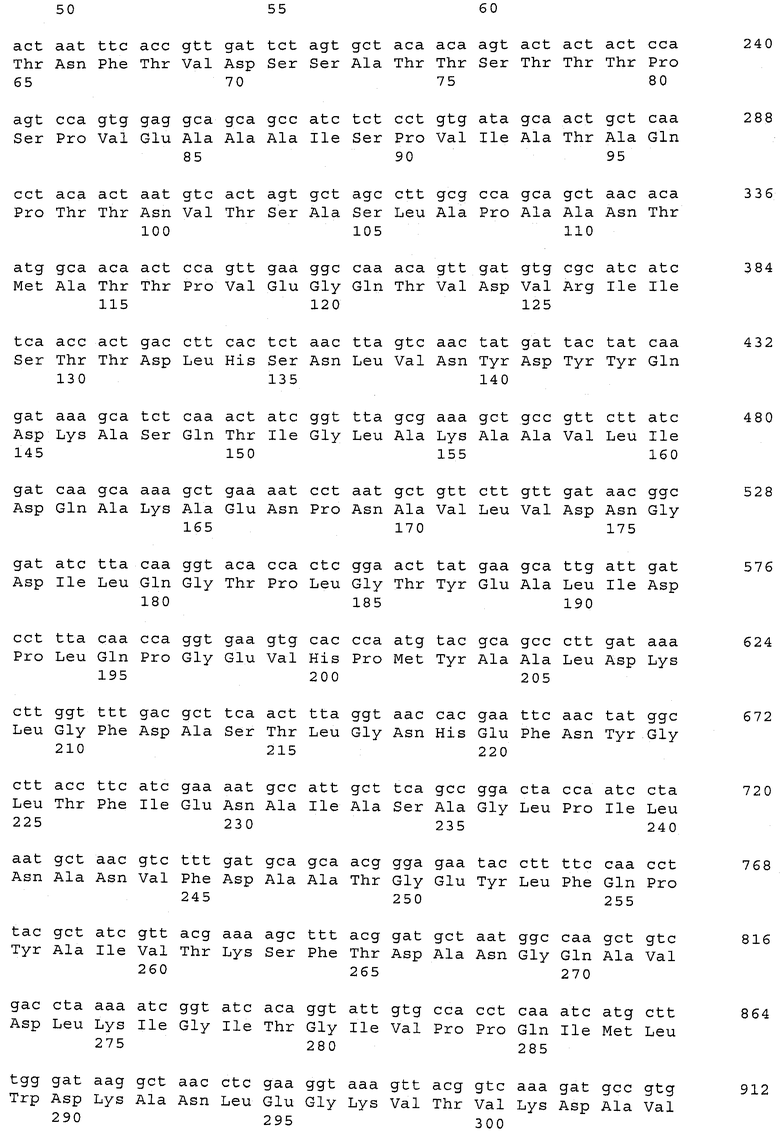
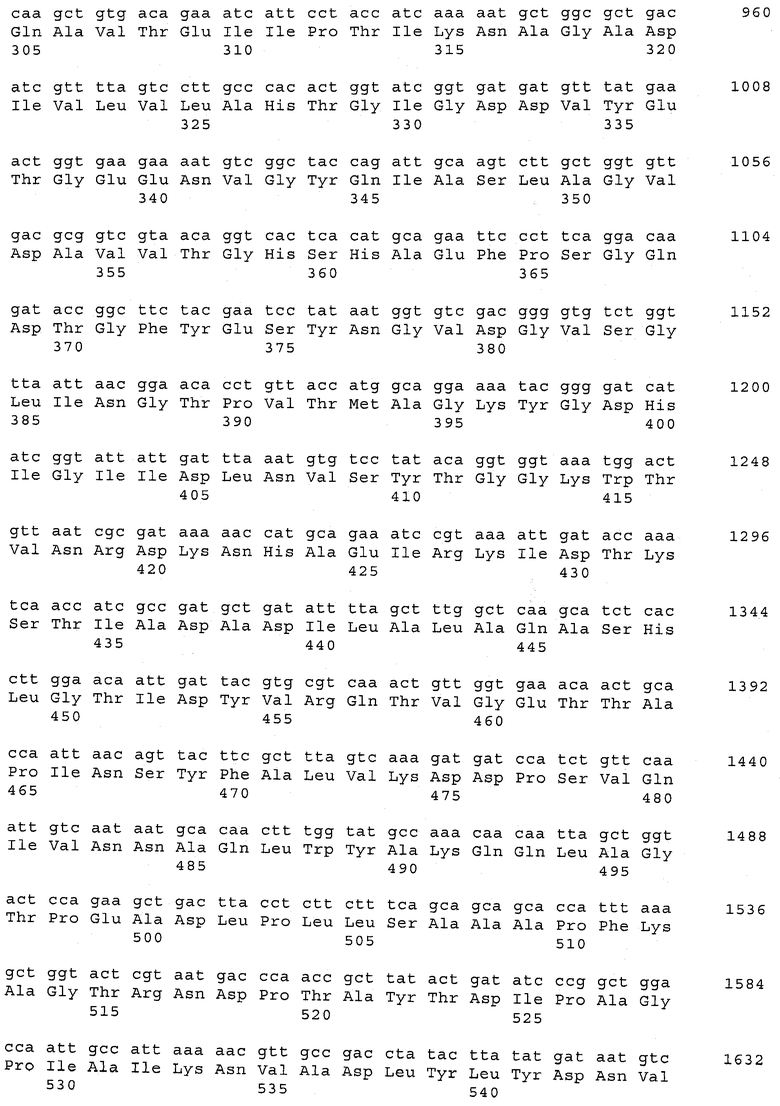
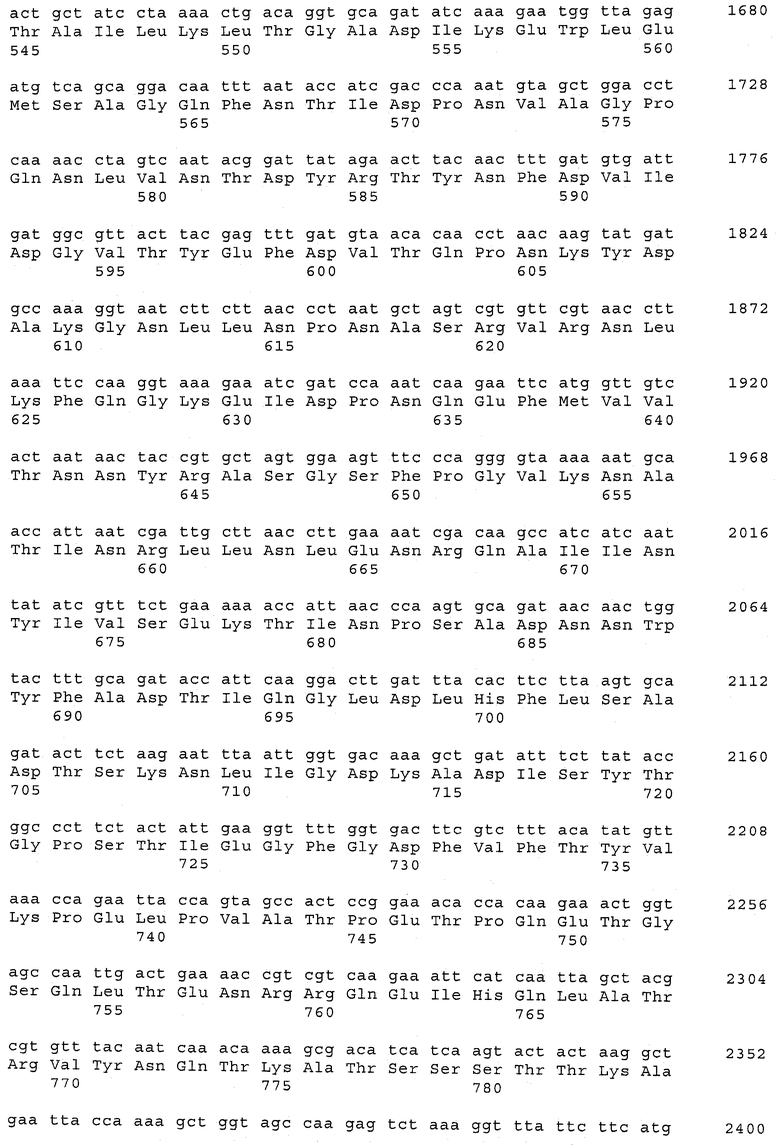




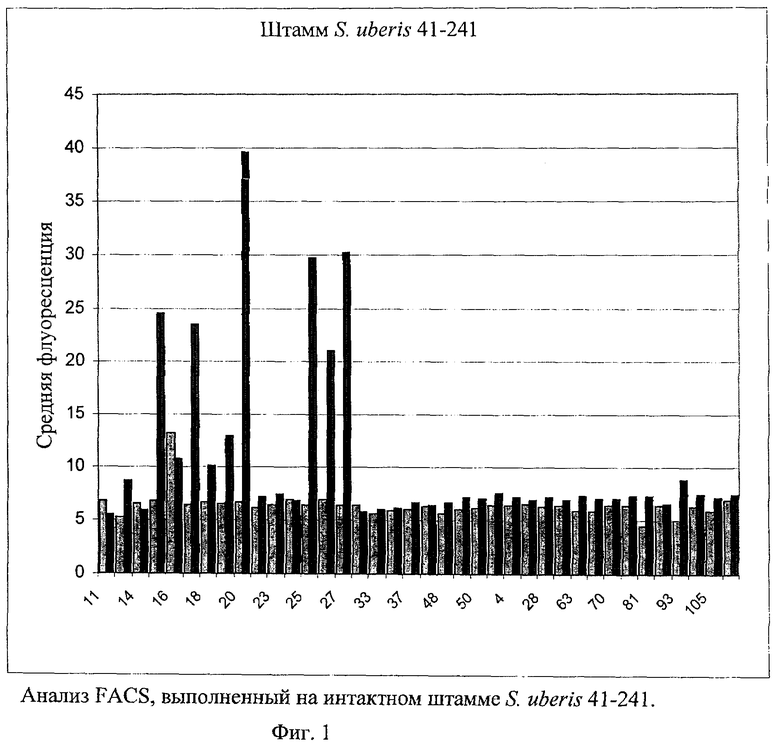
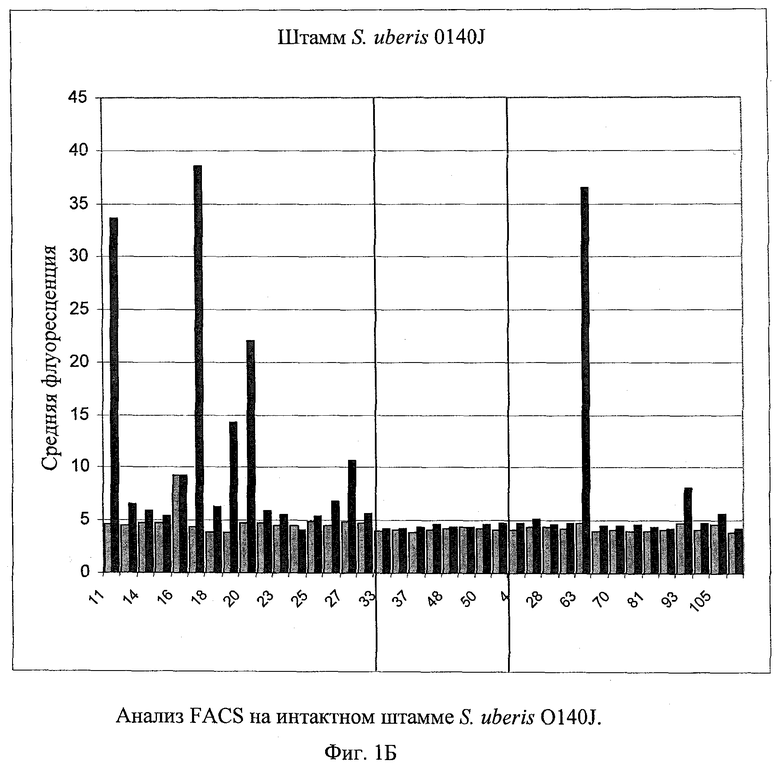
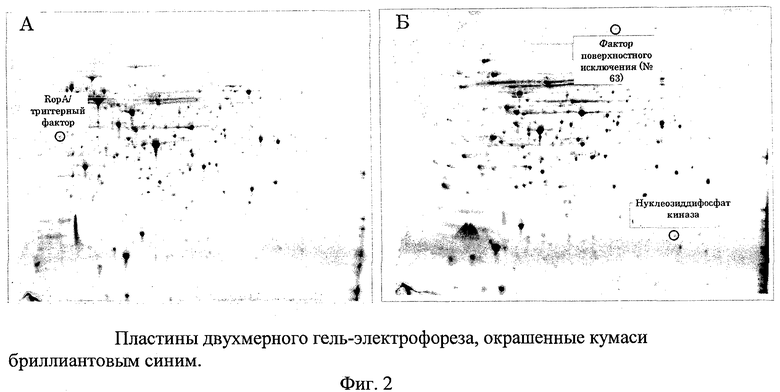
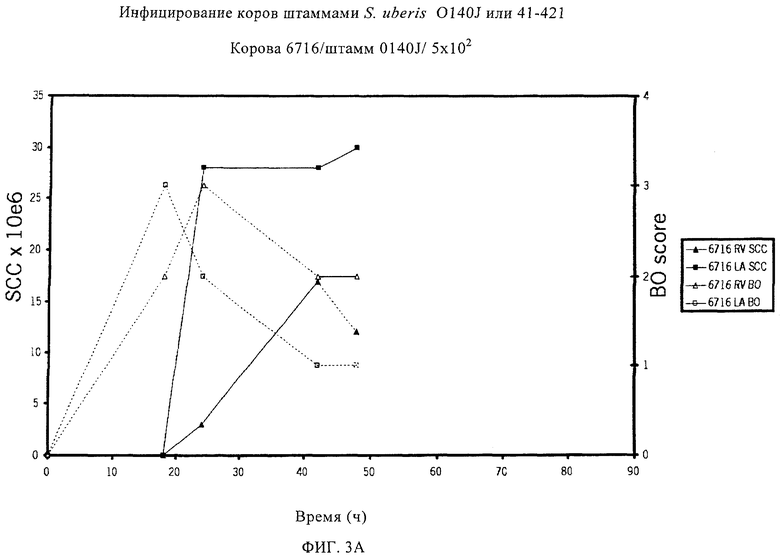
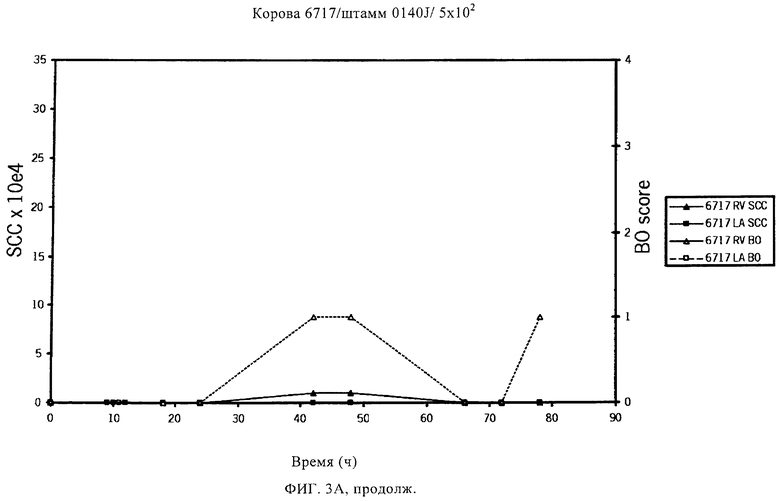
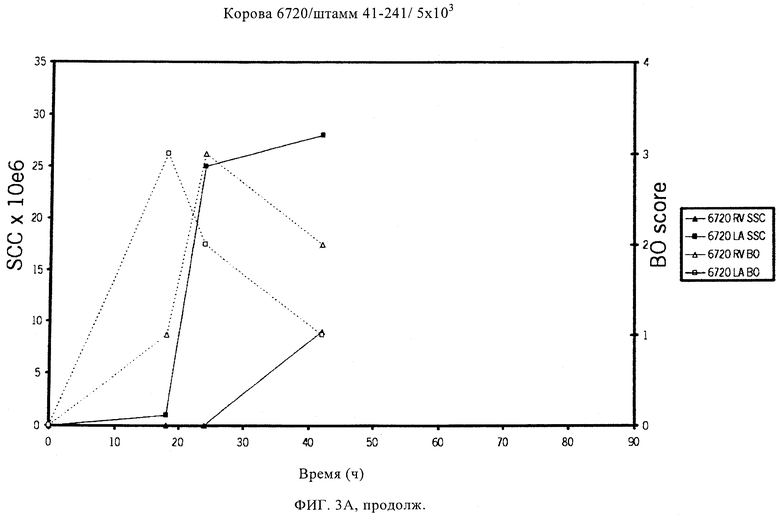
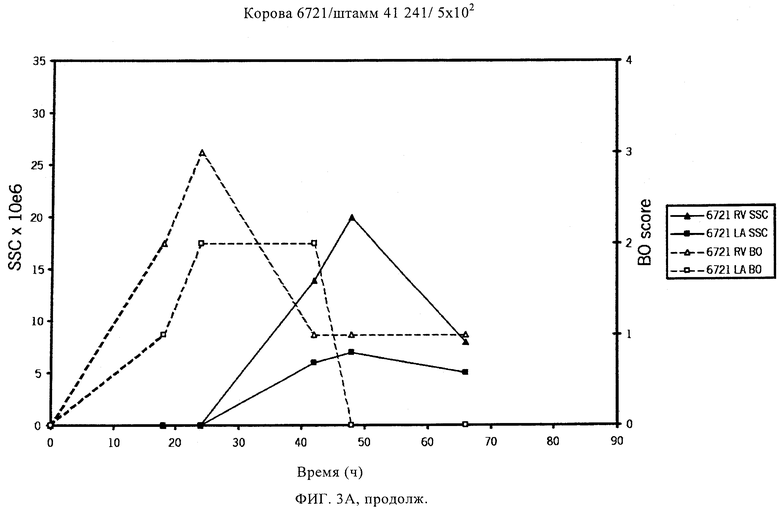
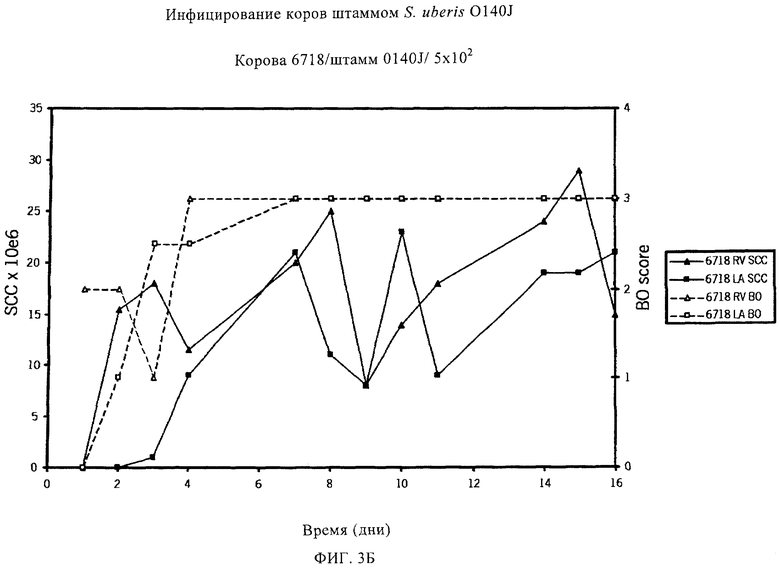
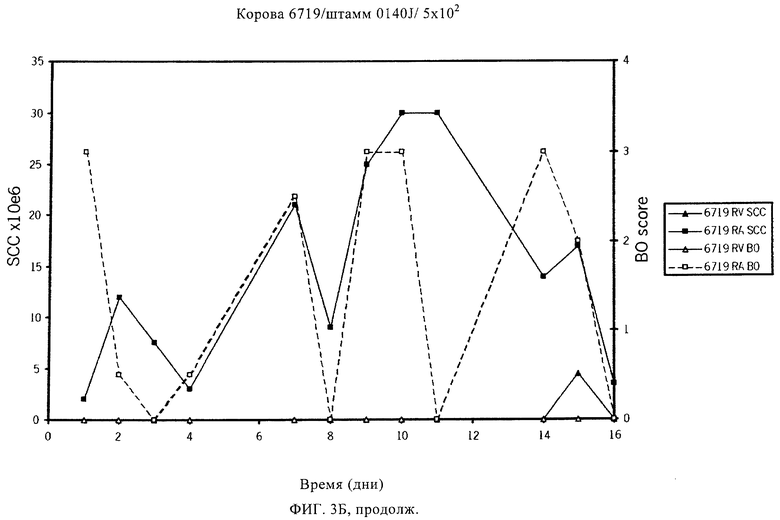
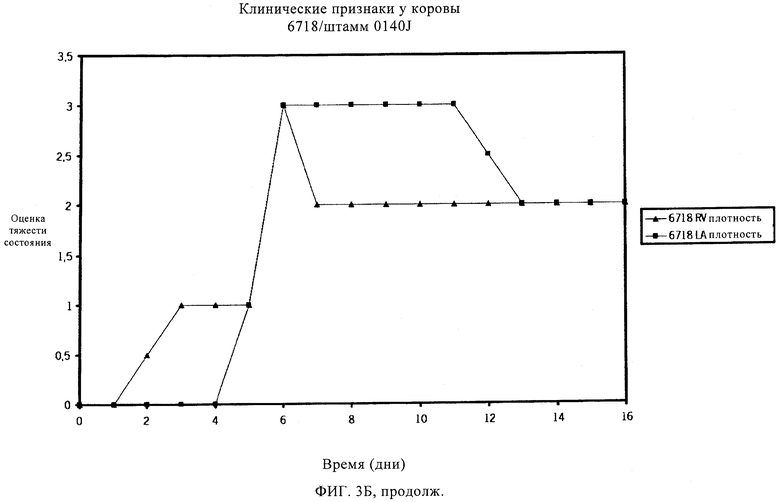
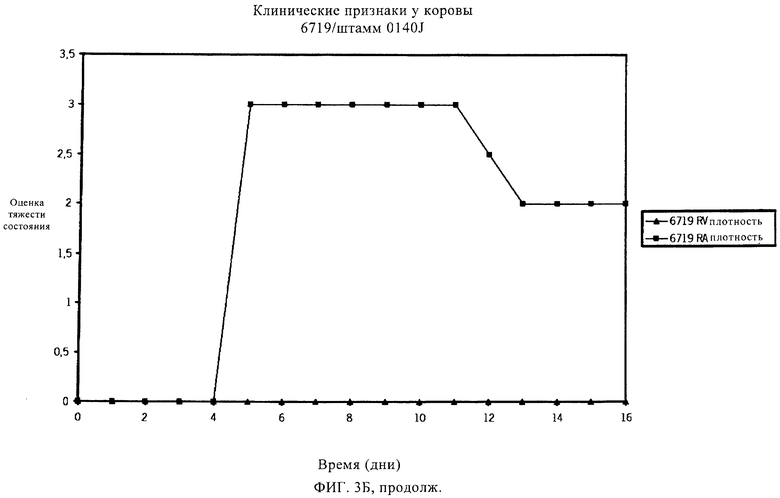
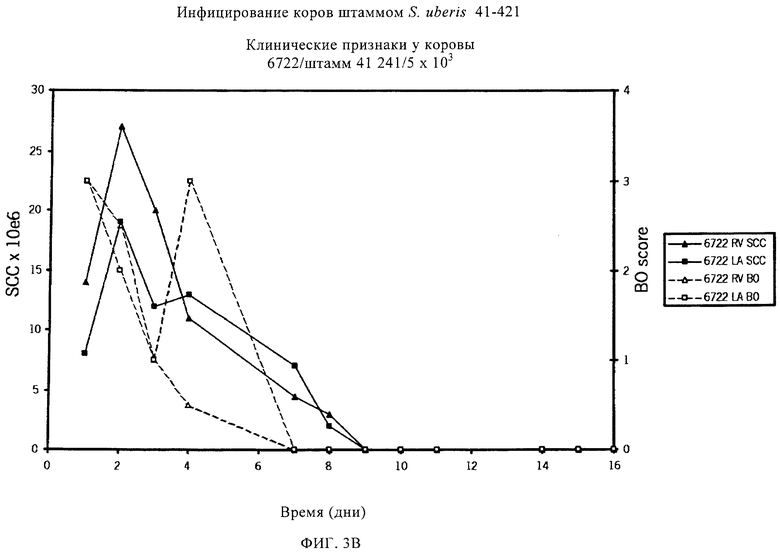
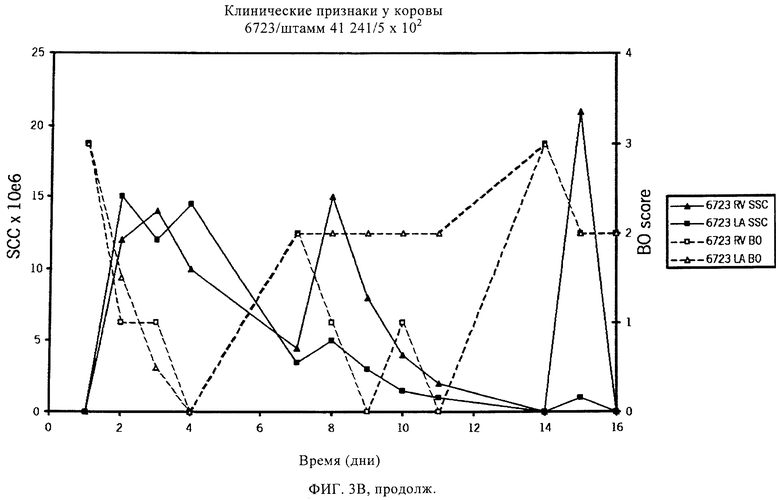
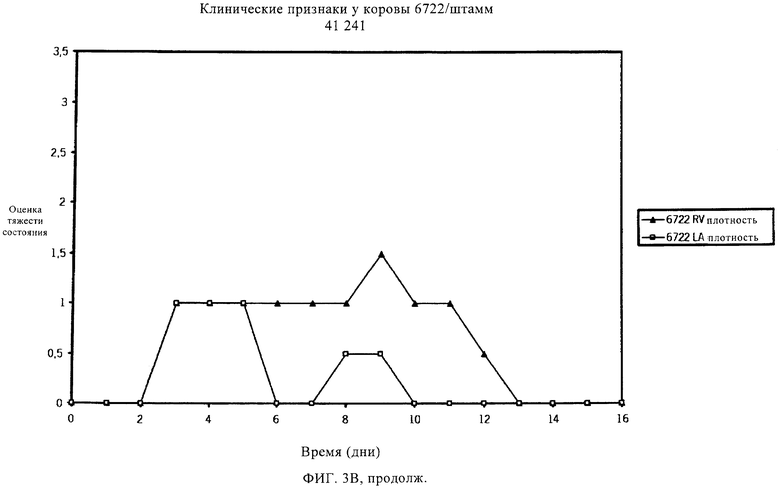
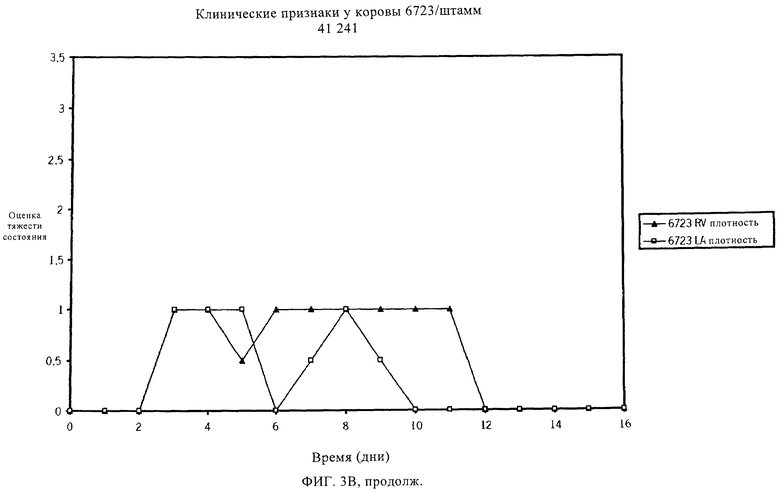



















Изобретение относится к области фармацевтики. Создана иммуногенная композиция, способная индуцировать иммунный ответ в отношении по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, включающая по меньшей мере один выделенный и/или рекомбинантный белок, имеющий определенную аминокислотную последовательность, полученный способом, включающим трансформацию прокариотической, эукариотической клеток или микроорганизма, такого как бактерия, дрожжи или грибы, конструкцией нуклеиновой кислоты, кодирующей указанный протеин, рекомбинантную молекулу нуклеиновой кислоты, рекомбинантный носитель и/или их иммуногенную часть, производное и/или аналог. Способ получения иммуногенной композиции. Способ идентификации белка бактерии Streptococcus uberis, который способен вызывать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, включающий стадии: а) идентификации по меньшей мере части секретируемого белка, поверхностно-ассоциируемого белка и/или белка, который по меньшей мере на 80% идентичен последовательности фактора бактериальной вирулентности, б) выбор по меньшей мере одного белка, идентифицированного на стадии а), который сохраняется по меньшей мере в двух штаммах и/или серотипах Streptococcus uberis, в) определение способности по меньшей мере одного белка, выбранного на стадии б), или его иммуногенной части, производного или аналога, специфически связывать антитело и/или иммунные клетки животного, инфицированного первым штаммом и/или серотипом Streptococcus uberis, и антителом и/или иммунными клетками животного, инфицированного вторым штаммом и/или серотипом Streptococcus uberis. Способ индукции иммунного ответа против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis. А также набор для диагностики мастита у крупного рогатого скота, включающий по меньшей мере один белок, имеющий определенную аминокислотной последовательность, рекомбинантную молекулу нуклеиновой кислоты, рекомбинантный носитель и средства выявления антитела. Использование изобретения позволяет индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis.. 10 н. и 11 з.п.ф-лы, 4 ил., 7 табл., 12 пр.
1. Способ идентификации белка Streptococcus uberis, который способен вызвать иммунный ответ в отношении по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, включающий:
а) идентификацию по меньшей мере части секретируемого белка, поверхностно-ассоциированного белка и/или белка, который по меньшей мере на 80% идентичен последовательности фактора бактериальной вирулентности;
б) выбор по меньшей мере одного белка, идентифицированного на стадии а), который сохраняется по меньшей мере в двух штаммах и/или серотипах Streptococcus uberis, и
в) определение способности по меньшей мере одного белка, выбранного на стадии б), или его иммуногенной части, производного и/или аналога, специфически связывать антитело и/или иммунные клетки животного, инфицированного первым штаммом и/или серотипом Streptococcus uberis, и антителом и/или иммунными клетками животного, инфицированного вторым штаммом и/или серотипом Streptococcus uberis.
2. Способ по п.1, в котором указанный секретированный белок и/или поверхностно-ассоциированный белок идентифицируют путем выявления по меньшей мере части геномной последовательности гена Streptococcus uberis, включающего мотив секретированного и/или поверхностно-ассоциированного белка.
3. Способ по п.1, в котором указанный белок, обладающий по меньшей мере 80% идентичностью по отношению к последовательности бактериального фактора вирулентности, идентифицируют путем выявления по меньшей мере в части геномной последовательности гена Streptococcus uberis, который обладает по меньшей мере 80% идентичностью по отношению к последовательности гена фактора бактериальной вирулентности.
4. Способ по п.1, в котором указанный белок, последовательность которого идентична по меньшей мере на 80% последовательности бактериального фактора вирулентности, идентифицируют путем выявления по меньшей мере части геномной последовательности гена Streptococcus uberis, которая гибридизуется с нуклеотидной последовательностью полной длины какой-либо из последовательностей нуклеиновой кислоты, выбранных из группы, которая состоит из SEQ ID NO:109, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:115, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:121, SEQ ID NO:123, SEQ ID NO:125, SEQ ID NO:127, SEQ ID NO:129, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137, SEQ ID NO:139, SEQ ID NO:141, SEQ ID NO:143, SEQ ID NO:145, SEQ ID NO:147, SEQ ID NO:149, SEQ ID NO:151, SEQ ID NO:153, SEQ ID NO:155, SEQ ID NO:157, SEQ ID NO:159, SEQ ID NO:161, SEQ ID NO:163, SEQ ID NO:165, SEQ ID NO:167, SEQ ID NO:169, SEQ ID NO:171, SEQ ID NO:173, SEQ ID NO:175, SEQ ID NO:177, SEQ ID NO:179, SEQ ID NO:181, SEQ ID NO:183, SEQ ID NO:185, SEQ ID NO:187 и SEQ ID NO:189 при 65°C в буфере, содержащем 0,5 М натрий фосфат, 1 мМ EDTA и 7% натрий додецилсульфат при pH 7,2, причем молекулы нуклеиновой кислоты остаются гибридизированными после двукратной промывки буфером, содержащим 40 мМ натрий фосфат (pH 7,2), 1 мМ EDTA и 5% натрий додецилсульфат в течение 30 мин при 65°C, и промывки дважды буфером, содержащим 40 мМ натрий фосфат (pH 7,2), 1 мМ EDTA и 1% натрий додецилсульфат в течение 30 мин при 65°C.
5. Способ по п.2, дополнительно включающий выбор гена, который сохраняется по меньшей мере в двух штаммах и/или серотипах Streptococcus uberis.
6. Способ по п.5, дополнительно включающий получение белка, кодируемого указанным геном, или иммуногенной части, производного и/или аналога указанного белка.
7. Способ по п.2, в котором указанный ген экспрессируется в прокариотической системе экспрессии.
8. Способ по п.1, включающий:
- получение выделенных и/или рекомбинантных белков Streptococcus uberis,
- инкубирование указанных белков с антителом и/или иммунными клетками животного, инфицированного первым штаммом и/или серотипом Streptococcus uberis, и антителом и/или иммунными клетками животного, инфицированного вторым штаммом и/или серотипом Streptococcus uberis, и
- определение способности белка связывать антитело и/или иммунные клетки животного, инфицированного первым штаммом и/или серотипом Streptococcus uberis, и антитело и/или иммунные клетки животного, инфицированного вторым штаммом и/или серотипом Streptococcus uberis.
9. Способ по п.8, дополнительно включающий экспрессию указанного белка, используя последовательность нуклеиновой кислоты, которая кодирует указанный белок.
10. Способ по п.1, в котором указанное антитело и/или иммунные клетки получают из сыворотки в период выздоровления.
11. Способ по п.1, в котором указанный белок Streptococcus uberis способен индуцировать опсонофагоцитоз-индуцирующие антитела.
12. Способ по п.1, в котором идентифицируют по меньшей мере два белка Streptococcus ubers, способных индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis.
13. Способ получения по меньшей мере одного белка, идентифицированного способом по одному из пп.1-12, который включает трансформацию прокариотической клетки, эукариотической клетки или микроорганизма такого, как бактерия, дрожжи или грибы, конструкцией нуклеиновой кислоты, кодирующей указанный протеин, и сбор продуцируемого рекомбинантного белка из клетки или культуральной жидкости.
14. Белок Streptococcus uberis, способный индуцировать иммунный ответ в отношении по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, который получен способом по одному из пп.1-13.
15. Рекомбинантная молекула нуклеиновой кислоты, включающая последовательность нуклеиновой кислоты, кодирующую по меньшей мере два белка, идентифицированных способом по п.1, и/или по меньшей мере два белка, имеющих аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:116, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:122, SEQ ID NO:124, SEQ ID NO:126, SEQ ID NO:128, SEQ ID NO:130, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138, SEQ ID NO:140, SEQ ID NO:142, SEQ ID NO:144, SEQ ID NO:146, SEQ ID NO:148, SEQ ID NO:150, SEQ ID NO:152, SEQ ID NO:154, SEQ ID NO:156, SEQ ID NO:158, SEQ ID NO:160, SEQ ID NO:162, SEQ ID NO:164, SEQ ID NO:166, SEQ ID NO:168, SEQ ID NO:170, SEQ ID NO:172, SEQ ID NO:174, SEQ ID NO:176, SEQ ID NO:178, SEQ ID NO:180, SEQ ID NO:182, SEQ ID NO:184, SEQ ID NO:186, SEQ ID NO:188 и SEQ ID NO:190 и/или иммуногенную часть по меньшей мере одного из указанных белков, под контролем функционально связанного промотора.
16. Рекомбинантный носитель, включающий нуклеиновую кислоту, кодирующую по меньшей мере два белка, идентифицированных способом по п.1, и/или имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:116, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:122, SEQ ID NO:124, SEQ ID NO:126, SEQ ID NO:128, SEQ ID NO:130, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138, SEQ ID NO:140, SEQ ID NO:142, SEQ ID NO:144, SEQ ID NO:146, SEQ ID NO:148, SEQ ID NO:150, SEQ ID NO:152, SEQ ID NO:154, SEQ ID NO:156, SEQ ID NO:158, SEQ ID NO:160, SEQ ID NO:162, SEQ ID NO:164, SEQ ID NO:166, SEQ ID NO:168, SEQ ID NO:170, SEQ ID NO:172, SEQ ID NO:174, SEQ ID NO:176, SEQ ID NO:178, SEQ ID NO:180, SEQ ID NO:182, SEQ ID NO:184, SEQ ID NO:186, SEQ ID NO:188 и SEQ ID NO:190 и/или иммуногенную часть по меньшей мере одного из указанных белков.
17. Иммуногенная композиция, способная индуцировать иммунный ответ в отношении по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, включающая по меньшей мере один выделенный и/или рекомбинантный белок по п.14, рекомбинантную молекулу нуклеиновой кислоты по п.15, рекомбинантный носитель по п.16, и/или их иммуногенную часть, производное и/или аналог.
18. Способ получения иммуногенной композиции, способной индуцировать иммунный ответ против по меньшей мере двух штаммов и/или серотипов Streptococcus uberis, который включает получение клеток по меньшей мере с одним рекомбинантным вектором, причем по меньшей мере один указанный вектор включает последовательность нуклеиновой кислоты, которая кодирует по меньшей мере один белок, идентифицированный способом по одному из пп.1-13 или полученный с помощью способа по п.13 и/или по меньшей мере один белок, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:116, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:122, SEQ ID NO:124, SEQ ID NO:126, SEQ ID NO:128, SEQ ID NO:130, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138, SEQ ID NO:140, SEQ ID NO:142, SEQ ID NO:144, SEQ ID NO:146, SEQ ID NO:148, SEQ ID NO:150, SEQ ID NO:152, SEQ ID NO:154, SEQ ID NO:156, SEQ ID NO:158, SEQ ID NO:160, SEQ ID NO:162, SEQ ID NO:164, SEQ ID NO:166, SEQ ID NO:168, SEQ ID NO:170, SEQ ID NO:172, SEQ ID NO:174, SEQ ID NO:176, SEQ ID NO:178, SEQ ID NO:180, SEQ ID NO:182, SEQ ID NO:184, SEQ ID NO:186, SEQ ID NO:188 и SEQ ID NO:190 и/или их иммуногенную часть, производное и/или аналог.
19. Способ измерения иммунитета животного в отношении S. uberis, включающий определение по меньшей мере в одном образце от указанного животного наличия антител, направленных против белка, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:116, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:122, SEQ ID NO:124, SEQ ID NO:126, SEQ ID NO:128, SEQ ID NO:130, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138, SEQ ID NO:140, SEQ ID NO:142, SEQ ID NO:144, SEQ ID NO:146, SEQ ID NO:148, SEQ ID NO:150, SEQ ID NO:152, SEQ ID NO:154, SEQ ID NO:156, SEQ ID NO:158, SEQ ID NO:160, SEQ ID NO:162, SEQ ID NO:164, SEQ ID NO:166, SEQ ID NO:168, SEQ ID NO:170, SEQ ID NO:172, SEQ ID NO:174, SEQ ID NO:176, SEQ ID NO:178, SEQ ID NO:180, SEQ ID NO:182, SEQ ID NO:184, SEQ ID NO:186, SEQ ID NO:188 и SEQ ID NO:190, или белка по п.14 и/или его иммуногенной части.
20. Способ индукции иммунного ответа против по меньшей мере двух штаммов и/или серотипов Streptococcus, uberis, включающий введение хозяину по меньшей мере одного выделенного и/или рекомбинантного белка по п.14, рекомбинантной молекулы нуклеиновой кислоты по п.15, рекомбинантного носителя по п.16 и/или их иммуногенной части, производного и/или аналога.
21. Диагностический набор для диагностики мастита крупного рогатого скота, включающий по меньшей мере один белок, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:116, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:122, SEQ ID NO:124, SEQ ID NO:126, SEQ ID NO:128, SEQ ID NO:130, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138, SEQ ID NO:140, SEQ ID NO:142, SEQ ID NO:144, SEQ ID NO:146, SEQ ID NO:148, SEQ ID NO:150, SEQ ID NO:152, SEQ ID NO:154, SEQ ID NO:156, SEQ ID NO:158, SEQ ID NO:160, SEQ ID NO:162, SEQ ID NO:164, SEQ ID NO:166, SEQ ID NO:168, SEQ ID NO:170, SEQ ID NO:172, SEQ ID NO:174, SEQ ID NO:176, SEQ ID NO:178, SEQ ID NO:180, SEQ ID NO:182, SEQ ID NO:184, SEQ ID NO:186, SEQ ID NO:188 и SEQ ID NO:190, или белок по п.14, или его иммуногенную часть, и средства выявления антитела, связывающего указанный белок или его иммуногенную часть.
| LI Y., MARTINEZ G | |||
| et al Identification of a surface protein of Streptococcus suis and evaluation of its immunogenic and protective capacity in pigs | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| ALMEIDA R | |||
| A | |||
| et al Identification, isolation, and partial characterization of a novel Streptococcus uberis adhesion molecule (SUAM) | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |

