Настоящая заявка притязает на приоритет заявки на выдачу патента США № 12/546292, поданной 24 августа 2009. Полное содержание вышеупомянутой заявки включено в настоящее описание в виде ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящая заявка, в общем, относится к медицинской диагностике и, в частности, к способам диагностики злокачественных состояний в различных тканях.
УРОВЕНЬ ТЕХНИКИ
Рак молочной железы является наиболее распространенной причиной злокачественных опухолей у женщин и второй из наиболее распространенных причин смертности у женщин от злокачественной опухоли в США. Хотя большинство новых случаев рака молочной железы диагностируют на основе отклонений, наблюдаемых на маммограмме, уплотнение или изменение консистенции ткани молочной железы также может быть настораживающим признаком заболевания. Повышенное внимание к определению риска рака молочной железы в последние десятилетия привело к увеличению количества женщин, подвергнутых маммографии для скрининга, что привело к выявлению рака на более ранних стадиях и в результате к улучшенным показателям выживаемости. Хотя рак молочной железы все еще является наиболее распространенной причиной смертности у женщин в возрасте от 45 до 55 лет.
Рак молочной железы можно классифицировать по нескольким стадиям. Стадия 0 означает карциному (включая дольковую карциному и протоковую карциному) in situ. Стадия I является ранней стадией инвазивного рака молочной железы. Опухоль диаметром не более 2 сантиметров. Злокачественные клетки не распространяются за пределы молочной железы. Опухоли стадии II включают опухоли, которые имеют диаметр не более 2 сантиметров, но которые распространяются в подмышечные лимфатические узлы, опухоли, имеющие диаметр от 2 до 5 сантиметров, которые могут распространяться в подмышечные лимфатические узлы, и опухоли, которые больше 5 сантиметров (2 дюймов), но которые не распространились в подмышечные лимфатические узлы. Стадия III соответствует местно распространенной злокачественной опухоли. Указанную стадию дополнительно делят на стадии IIIA, IIIB и IIIC. Стадия IV соответствует злокачественной опухоли с отдаленными метастазами. Злокачественная опухоль может распространяться в другие части тела. Способы лечения ранних стадий отличаются от способов лечения поздних стадий.
Известно, что многие типы злокачественной опухоли вызваны генетическими нарушениями, т.е. мутациями. Накопление мутаций и утрата регуляторных клеточных функций вызывают прогрессирующие фенотипические изменения от нормальной гистологии к раннему предраковому состоянию, такому как внутриэпителиальная неоплазия (IEN), затем к IEN повышенной степени тяжести, затем к поверхностному раку и, наконец, к инвазивному заболеванию. Хотя такой процесс в некоторых случаях может быть относительно агрессивным, обычно он происходит относительно медленно в течение нескольких лет и даже десятилетий. Онкогенная аддикция означает физиологическую зависимость злокачественных клеток от непрерывной активации или сверхэкспрессии отдельных онкогенов для поддержания злокачественного фенотипа. Такая зависимость возникает в условиях других изменений, которые являются маркерами прогрессирования неоплазии.
Длительный период прогрессирования инвазивного рака обеспечивает возможность клинического вмешательства. Поэтому важно идентифицировать биомаркеры, которые являются показателями предраковых состояний, так чтобы можно было предпринять меры лечения для предотвращения или замедления развития инвазивного рака.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Один аспект настоящего изобретения относится к способу мониторинга состояний молочной железы у субъекта. Способ включает определение отношения экспрессии гена парного бокса 2 (Paired Box 2) к экспрессии гена бета-дефензина 1 (PAX2-к-DEFB1) в клетках, полученных из молочной железы субъекта, при этом отношение экспрессии PAX2 к экспрессии DEFB1 коррелирует с состояниями молочной железы и может служить в качестве прогностического показателя, используемого для определения хода лечения.
В одном варианте отношение экспрессии PAX2 к экспрессии DEFB 1, составляющее 100:1 или выше, является показателем наличия рака молочной железы у субъекта, и отношение экспрессии PAX2 к экспрессии DEFB1 менее 100:1 является показателем наличия незлокачественного или предзлокачественного состояния молочной железы у субъекта.
В другом варианте определение стадии заключается в определении уровня экспрессии гена PAX2 по сравнению с уровнем экспрессии контрольного гена, определении уровня экспрессии гена DEFB1 по сравнению с уровнем экспрессии того же самого контрольного гена; и определении отношения экспрессии PAX2 к экспрессии DEFB1 на основании уровней экспрессии PAX2 и DEFB1.
В одном варианте способ дополнительно включает определение статуса рецептора эстрогена/рецептора прогестерона/рецептора эпидермального фактора роста 2 (ER/PR/HER2) в клетках, полученных из ткани молочной железы в случае данного состояния молочной железы.
Другой аспект настоящего изобретения относится к набору для мониторинга состояний молочной железы. В одном варианте набор для мониторинга состояний молочной железы содержит: одну или несколько пар олигонуклеотидных праймеров для выявления экспрессии PAX2 в образце ткани, одну или несколько пар олигонуклеотидных праймеров для выявления экспрессии DEFB1 в образце ткани и инструкции о том, как определить отношение экспрессии PAX2 к экспрессии DEFB1 в образце ткани с использованием праймеров. В другом варианте одна или несколько пар олигонуклеотидных праймеров для выявления экспрессии PAX2 содержат пару олигонуклеотидных праймеров, выбранную из группы, состоящей из последовательностей SEQ ID NO: 43 и 47, SEQ ID NO: 44 и 48, и SEQ ID NO: 45 и 49. В другом варианте одна или несколько пар олигонуклеотидных праймеров для выявления экспрессии DEFB1 содержат последовательности SEQ ID NO: 35 и 37.
В другом варианте набор дополнительно содержит одну или несколько пар контрольных олигонуклеотидных праймеров. В одном варианте одна или несколько пар контрольных олигонуклеотидных праймеров содержит олигонуклеотидные праймеры для выявления экспрессии β-актина. В предпочтительном варианте олигонуклеотидные праймеры для выявления экспрессии β-актина содержат последовательности SEQ ID NO: 34 и 36.
В другом варианте одна или несколько пар контрольных олигонуклеотидных праймеров содержат олигонуклеотидные праймеры для выявления экспрессии GAPDH. В предпочтительном варианте олигонуклеотидные праймеры для выявления экспрессии GAPDH содержат последовательности SEQ ID NO: 42 и 46.
В другом родственном варианте набор дополнительно содержит один или несколько реагентов для ПЦР-реакции.
В еще одном родственном варианте набор дополнительно содержит один или несколько реагентов для экстракции РНК.
В другом варианте набор для мониторинга состояний молочной железы содержит микроматрицу олигонуклеотидов, содержащую олигонуклеотидные зонды для выявления экспрессии PAX2 и DEFB1, и инструкции о том, как определить отношение экспрессии PAX2 к экспрессии DEFB1 в образце ткани с использованием микроматрицы олигонуклеотидов.
В родственном варианте набор дополнительно содержит реагенты для экстракции РНК из образца ткани.
Другой аспект изобретения относится к способу определения схемы лечения субъекта с данным состоянием молочной железы. Способ включает стадию определения уровня экспрессии гена PAX2 по сравнению с уровнем экспрессии контрольного гена в клетках, полученных из ткани молочной железы указанного субъекта, и определения схемы лечения указанного субъекта на основе относительного уровня экспрессии гена PAX2.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Сопровождающие фигуры, которые включены и которые составляют часть настоящего описания, иллюстрируют некоторые варианты раскрытого способа и композиций и вместе с описанием служат для объяснения принципов предлагаемого способа и композиций.
На фигуре 1 показан количественный анализ ОТ-ПЦР (КОТ-ПЦР) экспрессии бета-дефензина-1 (DEFB1). Чтобы подтвердить индукцию экспрессии DEFB1, осуществляли КОТ-ПЦР. На фигуре 1A показаны относительные уровни экспрессии DEFB1, сравниваемые в клинических образцах от 6 пациентов, которые были подвергнуты радикальной простатэктомии. На фигуре 1B показаны относительные уровни экспрессии DEFB1, сравниваемые в доброкачественных и злокачественных клинических образцах простаты, клетках hPrEC и в линиях злокачественных клеток простаты до и после индукции DEFB1. На фигуре 1C показаны относительные уровни экспрессии DEFB1, анализируемые в доброкачественной ткани, злокачественной ткани и внутриэпителиальной неоплазии простаты (PIN) в одном срезе ткани. На фигуре 1D показана экспрессия DEFB1 в доброкачественной ткани, злокачественной ткани и PIN у одного пациента по сравнению со средним уровнем экспрессии DEFB1, обнаруженном в доброкачественной ткани.
На фигуре 2 показан микроскопический анализ индуцированных DEFB1 изменений целостности мембраны и морфологии клеток. Морфологию клеток DU145, PC3 и LNCaP анализировали, используя фазово-контрастную микроскопию, через 48 часов после индукции DEFB1. Перемещение волны возбуждения в мембране указано черными стрелками и апоптозные тельца указаны белыми стрелками.
На фигуре 3 показан анализ цитотоксичности DEFB1 в злокачественных клетках простаты. Линии клеток простаты DU145, PC3 и LNCaP обрабатывали PonA, чтобы индуцировать экспрессию DEFB1, в течение 1-3 дней, затем осуществляли анализ MTT, чтобы определить жизнеспособность клеток. Результаты представлены в виде среднего ± s.d. (стандартное отклонение), n=9.
На фигуре 4 показана индукция клеточной гибели клеток DU145 и PC3 под действием DEFB1. Экспрессию DEFB1 индуцировали в линиях злокачественных клеток простаты DU145 (A) и PC3 (B) и затем подвергали окрашиванию, используя аннексин V/ФИТЦ/йодид пропидия, и проточно-цитометрическому анализу. Клетки, позитивно окрашенные йодидом пропидия и аннексином V, считали апоптозными. Время индукции показано под каждой панелью. Цифры рядом с рамками для каждой временной точки представляют процентное содержание клеток йодид пропидия (PI)-, аннексин V+ (нижний правый квадрант) и клеток PI+, аннексин V+ (верхний правый квадрант). Данные получены в одном типичном эксперименте из трех отдельных экспериментов.
На фигуре 5 показан анализ пан-каспазы после индукции DEFB1. Клетки DU145 и PC3 красили FAM-VAD-FMK-меченым фторметилкетоном, чтобы выявить активность каспазы. Клетки были видимы при дифференциально-интерференционном контрасте (ДИК) для каждого состояния. Анализ с использованием конфокальной микроскопии не выявил окрашивания каспазы в контрольных клетках DU145 (B), клетках PC3 (F) и LNCaP (J). В клетках, обработанных PonA в течение 24 часов для индукции DEFB1, выявлена каспазная активность в DU145 (D) и PC3 (H). Не выявлено каспазной активности в LNCaP (L).
На фигуре 6 показан сайленсинг экспрессии белка гомеотического гена парного бокса 2 (PAX2) после обработки ми-РНК PAX2. На фигуре 6A показан Вестер-блот-анализ клеток PC3 и DU145, трансфицированных дуплексом ми-РНК PAX2, в нулевой день (дорожка 1), второй день (дорожка 2) и четвертый день (дорожка 3). На фигуре 6B показан Вестерн-блот-анализ клеток PC3 и DU145, трансфицированных дуплексом ми-РНК PAX2 в нулевой день (дорожка 1), второй день (дорожка 2), четвертый день (дорожка 3) и 6 день (дорожка 4). Белок PAX2 не выявлялся уже после четырех дней обработки (дорожка 3) в клетках DU145 и после шести дней обработки в PC3. С блотов удаляли антитела и подвергали повторному исследованию с использованием зондов на β-актин в качестве внутреннего контроля.
На фигуре 7 показан анализ роста злокачественных клеток простаты после обработки ми-РНК PAX2. Фазово-контрастный микроскопический анализ DU145, PC3 и LNCaP на 6 день в присутствии нормальной ростовой среды. Обработка используемой в качестве негативного контроля ми-РНК не оказывала влияния на клетки. Однако имело место значимое уменьшение количества клеток во всех трех линиях после обработки ми-РНК PAX2.
На фигуре 8 показан анализ гибели клеток после вызванного ми-РНК сайленсинга PAX2. Линии злокачественных клеток простаты PC3, DU145 и LNCaP обрабатывали 0,5 мкг пула из четырех ми-РНК PAX2 или четырех неспецифичных контрольных ми-РНК в течение 2, 4 или 6 дней с последующим осуществлением MTT-анализа, чтобы определить жизнеспособность клеток. Результаты представлены в виде среднего ± s.d., n=9.
На фигуре 9 показан анализ каспазной активности. Клетки DU145, PC3 и LNCaP красили меченным карбоксифлуоресцеином фторметилкетоном, чтобы выявить каспазную активность после обработки ми-РНК PAX2. Конфокальный микроскопический анализ необработанных и обработанных клеток показывает, что клетки видимы при использовании ДИК. Анализ флуоресценции не выявил окрашивания каспазы в контрольных клетках DU145 (B), клетках PC3 (F) и клетках LNCaP (J). Однако обработка клеток ми-РНК PAX2 индуцировала каспазную активность в DU145 (D), PC3 (H) и LNCaP (L).
На фигуре 10 показан анализ апоптозных факторов после обработки ми-РНК PAX2. Изменения в экспрессии проапоптозных факторов сравнивали в необработанных контрольных клетках и в клетках, обработанных в течение шести дней ми-РНК PAX2. На фигуре 10A показаны уровни экспрессии Bcl-2-ассоциированного X-белка (BAX), повышенные в DU145, PC3 и LNCaP. На фигуре 10B показана экспрессия агониста взаимодействующего домена смерти BH3 (BID), повышенная в DU145 и LNCaP, но измененная в PC3. На фигуре 10C показаны уровни экспрессии Bcl-2-ассоциированного промотора смерти (BAD), повышенные в трех линиях клеток.
На фигуре 11 показана модель связывания PAX2 с последовательностью распознавания ДНК. Репрессор транскрипции PAX2 связывается с сайтом распознавания CCTTG (SEQ ID NO: 1) непосредственно вблизи с TATA-боксом DEFB1, предотвращая транскрипцию и экспрессию белка DEFB1. Ингибирование экспрессии белка PAX2 обеспечивает нормальную экспрессию DEFB1.
Фигура 12 иллюстрирует конструкцию репортера DEFB1. Промотор DEFB1, состоящий из первых 160 оснований выше стартового сайта мРНК, амплифицировали в ПЦР из клеток DU145 и лигировали в репортерную плазмиду с геном люциферазы pGL3.
Фигура 13 показывает, что ингибирование PAX2 приводит к экспрессии DEFB1. DU145, PC3, LNCaP и HPrEC обрабатывали в течение 48 часов ми-РНК PAX2. КОТ-ПЦР-анализ перед обработкой показал отсутствие экспрессии DEFB1 в DU145, PC3 и LNCaP. Однако экспрессия DEFB1 восстанавливалась после обработки всех линий. Изменения экспрессии DEFB1 не происходило после обработки ми-РНК PAX2-нулевых клеток HprEC.
На фигуре 14 показано, что ингибирование PAX2 приводит к повышенной активности промотора DEFB1. Создавали конструкции промотор PC3/pGL3 и промотор DU145/pGL3, и полученными конструкциями трансфицировали клетки PC3 и DU145, соответственно. Активность промотора сравнивали до и после ингибирования PAX2 посредством обработки ми-РНК. Активность промотора DEFB1 возраста после обработки в 2,65 раз в DU145 и в 3,78 раза в PC3.
На фигуре 15 показан ChIP-анализ связывания PAX2 с промотором DEFB1. ChIP-анализ осуществляли на клетках DUI45 и PC3. После иммунопреципитации с использованием анти-PAX2-антитела осуществляли ПЦР, чтобы выявить область промотора DEFB1, содержащую сайт распознавания PAX2 GTTCC (SEQ ID NO: 2). Анализ показывает, что репрессор транскрипции PAX2 связан с промотором DEFB1 в линиях злокачественных клеток простаты.
На фигуре 16 показана рассчитанная структура PrdPD и PrdHD с ДНК. Координаты структур PrdPD, связанного с ДНК (Xu et al., 1995), и PrdHD, связанного с ДНК (Wilson et al., 1995), использовали для конструирования модели двух доменов в тот момент, когда они связаны с сайтом PHO. Отдельные сайты связывания граничат друг с другом в конкретной ориентации, которая указана. Домен RED ориентировали на основании кристаллической структуры PrdPD.
На фигуре 17 показано сравнение консенсусных последовательностей разных парных доменов. Сверху на фигуре изображено схематичное представление контактов белок ± ДНК, описанных при кристаллографическом анализе комплекса Prd-парный домен ± ДНК. Незаштрихованными прямоугольниками показаны α-спирали, заштрихованными прямоугольниками показаны β-слои и жирной линией указан β-поворот. Контактирующие аминокислоты показаны с использованием однобуквенного кода. Показаны только прямые контакты аминокислота ± основание. Незаштрихованными кружками показаны контакты в большой бороздке, тогда как красными стрелками показаны контакты в малой бороздке. Представленная схема выровнена со всеми известными консенсусными последовательностями белков парных доменов (показаны только нити, совпадающие лучше всего). Вертикальными линиями между консенсусными последовательностями показаны консервативные пары оснований. Нумерация положений показана внизу фигуры.
На фигуре 18 показано направленное воздействие на PAX2 в качестве методики химиопрофилактики. Нарушенная экспрессия PAX2 является ранним событием в случае инициации и прогрессирования злокачественной опухоли. Ингибирование PAX2 во время дисплазии или другой предраковой стадии можно использовать для профилактики злокачественной опухоли.
На фигуре 19 показано влияние ангиотензина II (Ang II) на экспрессию PAX2 в клетках DU145. Чтобы определить влияние AngII на экспрессию PAX2, следили за уровнями белка DEFB1 после обработки. В данном случае уровни экспрессии PAX2 возрастали уже через 4 часа и продолжали оставаться повышенными вплоть до 48 часов.
На фигуре 20A показано влияние лозартана (Los) на экспрессию PAX2 в DU145. Клетки DU145 обрабатывали блокатором рецептора ангиотензина II типа 1 (ATR1) лозартаном. КОТ-ПЦР-анализ показал, что уровни мРНК PAX2 снижались после обработки, по меньшей мере, на половину. На фигуре 20B показано влияние блокатора рецептора ангиотензина II типа 2 (ATR2) на экспрессию PAX2 в DU145. Чтобы определить влияние рецептора ATR2 на экспрессию PAX2, клетки DU145 обрабатывали блокатором рецептора ATR2 PD123319. В данном случае экспрессия PAX2 возрастала в 7-8 раз.
На фигуре 21 показано, что лозартан (Los) блокирует влияние AngII на экспрессию PAX2 в DU145. Обработка клеток DU145 5 мкМ AngII в течение 72 часов приводила к 2-кратному увеличению экспрессии PAX2. Кроме того, обработка 10 мкМ в течение 72 часов приводила к более чем 3-кратному увеличению экспрессии. Обработка клеток 5 мкМ лозартаном подавляла пролиферацию на 50%. Кроме того, обработка лозартаном в течение 30 минут перед обработкой AngII блокировала влияние AngII на пролиферацию.
На фигуре 22 показано, что AngII усиливает пролиферацию клеток DU145. Обработка клеток DU145 5 мкМ AngII в течение 72 часов приводила к 2-кратному увеличению пролиферации. Кроме того, обработка 10 мкМ в течение 72 часов приводила к более чем 3-кратному увеличению пролиферации.
На фигуре 23 показано влияние Los и ингибиторов MAP-киназ на экспрессию PAX2 в клетках DU145. На фигуре 23A показано, что обработка клеток DU145 лозартаном подавляет экспрессию фосфор-ERK 1/2 и PAX2; на фигуре 23B показано, что ингибиторы MEK-киназы и AICAR подавляют экспрессию белка PAX2; на фигуре 23C показано, что ингибиторы MEK-киназы и лозартан подавляют экспрессию белка фосфо-STAT3.
На фигуре 24 показано влияние Los и ингибиторов MEK-киназы на активацию PAX2 в клетках DU145. На фигуре 24A показано, что обработка клеток DU145 ингибиторами передачи сигнала AT1R приводила к снижению уровней белка фосфор-PAX2, который является активной формой PAX2. Кроме того, обработка индуктором AMP-киназы AICAR приводила к подавлению экспрессии PAX2. На фигуре 24B показано, что ингибирование передачи сигнала AT1R лозартаном снижало уровни фосфо-JNK. Однако AngII повышал уровни белка фосфор-JNK.
На фигуре 25 показано, что AngII повышал экспрессию PAX2 и снижал экспрессию DEFB1 в клетках hPrEC. Чтобы определить влияние AngII на уровни PAX2 в hPrEC, клетки обрабатывали в течение 72 и 96 часов и исследовали экспрессию PAX2 и DEFB1, используя КОТ-ПЦР. В данном случае обработка AngII приводила к резкому увеличению PAX2 до уровней, сходных с уровнями в злокачественных клетках простаты PC3. Наоборот, экспрессия DEFB1 была значительно снижена после обработки AngII.
На фигуре 26 показана схема передачи сигнала AngII и PAX2 при раке простаты. Экспрессия PAX2 в злокачественных клетках простаты регулируется путем передачи сигнала AT1R. В частности, каскад передачи сигнала MEK-киназы приводит к повышенной экспрессии PAX2. Кроме того, AT1R и AngII осуществляют повышающую регуляцию активации PAX2 через JNK.
На фигуре 27 показана схема блокирования экспрессии PAX2 в ходе терапии рака простаты. На фигуре 27A показано, что экспрессия PAX2 регулируется путем передачи сигнала AT1R. Ингибирование экспрессии PAX2 приводит к возобновлению экспрессии DEFB1 и гибели злокачественных клеток. На фигуре 27B показано, что соединения, которые блокируют AT1R, расположенные ниже киназы, или непосредственно подавляют PAX2, обеспечивают новый подход к лечению рака простаты.
На фигуре 28 показано сравнение экспрессии DEFB1 и PAX2 с оценкой по шкале Глисона. Относительные уровни экспрессии DEFB1 сравнивали в доброкачественных клинических образцах от 6 пациентов, которые были подвергнуты радикальной простатэктомии. В данном случае шкала Глисона была обратно пропорциональная уровням экспрессии DEFB1 в близлежащей доброкачественной ткани простаты. Пациенты с относительными уровнями экспрессии DEFB1 выше 0,005 имели оценку в баллах по шкале Глисона 6. Однако пациенты с уровнями экспрессии ниже 0,005 имели оценку в баллах по шкале Глисона 7.
На фигуре 29 показано отношение PAX2-DEFB1 в качестве прогностического фактора развития рака простаты. КОТ-ПЦР осуществляли на срезах тканей простаты, полученных с использованием лазерной захватывающей микродиссекции (LCM), чтобы определить относительные уровни экспрессии DEFB1 и PAX2. Уровни экспрессии DEFB1 снижались в ряду нормальная ткань-PIN-злокачественная опухоль. Однако экспрессия PAX2 увеличивалась в ряду нормальная ткань-PIN-злокачественная опухоль. Кроме того, пациент №1457 со злокачественной опухолью, имеющей оценку по шкале Глисона 6, имел больше DEFB1 в нормальной ткани и PIN по сравнению с пациентом №1569 со злокачественной опухолью, оцениваемой по шкале Глисона в 7 баллов. Наоборот, пациент №1569 имел более высокие уровни PAX2 в злокачественных областях по сравнению с пациентом №1457.
На фигуре 30 показано, что прогностический фактор Дональда (DPF) основан на соотношении относительной экспрессии PAX2-DEFB1. Повышение DPF для ткани простаты увеличивает шансы развития рака простаты. Ткань с соотношением PAX2-DEFB1 от 0 до 39 на основе DPF была нормальной (доброкачественной). Ткань с соотношением PAX2-DEFB1 от 40 до 99 представляла собой PIN (предзлокачественную) по шкале DPF. Наконец, ткань с соотношением PAX2-DEFB1 от 100 до 500 была злокачественной (от низкой до высокой степени злокачественности).
На фигуре 31 показан анализ экспрессии hBD-1 в ткани простаты человека. Относительные уровни экспрессии hBD-1 сравнивали с нормальными клиническими образцами от пациентов, которые были подвергнуты радикальной простатэктомии. Пунктирная линия служит в качестве точки отсчета для сравнения значений для макропрепаратов и препаратов, полученных с использованием LCM, и соответствующие баллы по шкале Глисона указаны над каждым столбиком. На фигуре 31A показаны уровни экспрессии hBD-1, сравниваемые в тканях на макроскопических срезах. На фигуре 31B показаны уровни экспрессии hBD-1, сравниваемые в тканях, полученных с использованием лазерной захватывающей микродиссекции.
На фигуре 32 показан анализ экспрессии hBD-1 в линиях клеток простаты. На фигуре 32A показаны уровни экспрессии hBD-1 в линиях злокачественных клеток простаты по сравнению с клетками hPrEC до и после индукции hBD-1. Звездочка указывает статистически более высокие уровни экспрессии по сравнению с hPrEC. Двойные звездочки означают статистически значимые уровни экспрессии по сравнению с линий клеток до индукции hBD-1 (t-критерий Стьюдента, p<0,05). На фигуре 32B показана эктопическая экспрессия hBD-1, подтвержденная в линии злокачественных клеток простаты DU145 с использованием иммуноцитохимии. Клетки hPrEC красили на hBD-1 в качестве дополнительного контроля (a: ДИК и b: флуоресценция). Клетки DU145 трансфицировали hBD-1 и индуцировали в течение 18 часов (c: ДИК и d: флуоресценция). Размерная линия = 20 мкм.
На фигуре 33 показан анализ цитотоксичности hBD-1 в злокачественных клетках простаты. Линии клеток простаты DU145, PC3, PC3/AR+ и LNCaP обрабатывали Pon A, чтобы индуцировать экспрессию hBD-1, в течение 1-3 дней, затем проводили MTT-анализ, чтобы определить жизнеспособность клеток. Каждый столбик представляет среднее ± S.E.M. (стандартная ошибка средней) из трех независимых экспериментов, проводимых в трех повторах.
На фигуре 34 показан КОТ-ПЦР-анализ экспрессии hBD-1 и cMYC на полученных с использованием LCM срезах тканей простаты человека: нормальной, PIN и опухоли. Экспрессия каждого гена представлена в виде отношений экспрессии по сравнению с β-актином. На фигуре 34A показано сравнение уровней экспрессии hBD-1 на срезах нормальных тканей, PIN и опухолей. На фигуре 34B показано сравнение уровня экспрессии cMYC на срезах нормальных тканей, PIN и опухолей.
На фигуре 35 показан КОТ-ПЦР-анализ экспрессии hBD1 после нокдауна PAX2 с использованием ми-РНК. Уровни экспрессии hBD-1 представлены в виде отношений экспрессии по сравнению с β-актином. Звездочкой указаны статистически более высокие уровни экспрессии по сравнению с линией клеток перед обработкой ми-РНК PAX2 (t-критерий Стьюдента, p<0,05).
На фигуре 36 показан сайленсинг экспрессии белка PAX2 после обработки ми-РНК PAX2. На фигуре 36A показана экспрессия PAX2, исследованная с использованием Вестерн-блот-анализа в первичных клетках простаты HPrEC (дорожка 1) и в злокачественных клетках простаты DU145 (дорожка 2), PC3 (дорожка 3) и LNCaP (дорожка 4). С блотов удаляли антитела и подвергали повторному исследованию с использованием зондов на β-актин в качестве внутреннего контроля, чтобы удостовериться в равной загрузке. На фигуре 36B показан Вестерн-блот-анализ DU145, PC3 и LNCaP, в которых был подтвержден нокдаун экспрессии PAX2 после трансфекции дуплексом ми-РНК PAX2. С блотов также удаляли антитела и подвергали повторному исследованию с использованием зондов на β-актин в качестве внутреннего контроля.
На фигуре 37 показан анализ роста злокачественных клеток простаты после обработки ми-РНК PAX2. Фазово-контрастный микроскопический анализ HPrEC (A), LNCaP (C), DU145 (E) и PC3 (G) через 6 день в присутствии неспецифичной ми-РНК, используемой в качестве негативного контроля. Имело место значимое уменьшение количества клеток в DU145 (D), PC3 (F) и LNCaP (H) после обработки ми-РНК PAX2. Однако, не выявлено влияния в HPrEC (B). Линия = 20 мкм.
На фигуре 38 показан анализ гибели клеток после сайленсинга PAX2 с использованием ми-РНК. Линии злокачественных клеток простаты PC3, DU145 и LNCaP обрабатывали ми-РНК PAX2 или неспецифичными негативными контрольными ми-РНК в течение 2, 4 или 6 дней, затем осуществляли MTT-анализ. Нокдаун PAX2 приводил к снижению относительной жизнеспособности клеток во всех трех линиях. Результаты представляют среднее ± SD, n=9.
На фигуре 39 показан анализ каспазной активности. Клетки DU145, PC3 и LNCaP красили меченным карбоксифлуоресцеином фторметилкетоном, чтобы выявить каспазную активность после обработки ми-РНК PAX2. Флуоресцентный анализ не выявил окрашивания каспазы в контрольных клетках DU145 (A), клетках PC3 (C) и клетках LNCaP (E). Однако в клетках, обработанных ми-РНК PAX2, была индуцирована каспазная активность в DU145 (B), PC3 (D) и LNCaP (F). Линия = 20 мкм.
На фигуре 40 показан анализ апоптозных факторов после обработки ми-РНК PAX2. Изменения в экспрессии проапоптозных факторов сравнивали в необработанных контрольных клетках и в клетках, обработанных в течение 6 дней ми-РНК PAX2. На фигуре 40A показана экспрессия BAD, повышенная в DU145, PC3 и LNCaP после нокдауна PAX2. На фигуре 40B показаны уровни экспрессии BID, повышенные в LNCaP и DU145, но не в клетках PC3. На фигуре 40C показана экспрессия AKT, сниженная в LNCaP и DU145. Однако не выявлено изменения экспрессии AKT в клетках PC3 после нокдауна PAX2. Результаты представляют среднее ± SD, n=9. Звездочками показаны статистические различия (p<0,05).
ПОДРОБНОЕ ОПИСАНИЕ
Если не определено иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, которое обычно подразумевает специалист в области, к которой относятся раскрытый способ и композиции. Необходимо отметить, что в смысле, используемом в настоящем описании и в прилагаемой формуле изобретения, формы единственного числа включают и формы множественного числа, если контекст ясно не диктует иное. Таким образом, например, указание «пептид» включает множество таких пептидов, ссылка на «пептид» означает ссылку на один или несколько пептидов и их эквивалентов, известных специалистам в данной области, и так далее.
Один аспект настоящего изобретения относится к способу мониторинга развития злокачественной опухоли. В некоторых вариантах способ относится к мониторингу предраковых состояний, таких как внутриэпителиальная неоплазия, и злокачественных состояния простаты или молочной железы у субъекта. Способ включает определение отношения экспрессии гена парного бокса 2 к экспрессии гена бета-дефензина-1 (PAX2-к-DEFB1) в клетках, полученных из простаты или молочной железы субъекта, при этом отношение экспрессии PAX2 к экспрессии DEFB1 коррелирует с состояниями простаты или молочной железы. Соотношение экспрессии генов может быть определено на уровне мРНК (например, с использованием ОТ-ПЦР или олигонуклеотидной матрицы) или на уровне белка (например, с использованием Вестерн-блота или матрицы антител).
В некоторых вариантах отношение экспрессии PAX2-к-DEFB определяют на уровне мРНК и называют в настоящем описании «прогностическим фактором Дональда» или «DPF».
В некоторых вариантах отношение экспрессии PAX2-к-DEFB1 (определяемое на уровне РНК) в простате используют для того, чтобы отличать нормальное, предраковое и злокачественное состояния простаты у субъекта. В одном варианте отношение PAX2-к-DEFB1 менее чем 40:1 является показателем нормального состояния простаты, отношение PAX2-к-DEFB1, по меньшей мере, от 40:1 до менее чем 100:1 является показателем внутриэпителиальной неоплазии простаты (PIN), и отношение PAX2-к-DEFB1, по меньшей мере, 100:1 является показателем рака простаты.
Также предлагается способ диагностики рака простаты у субъекта. Способ включает определение в клетках из простаты субъекта уровней PAX2 и бета-дефензина-1 (DEFB1), при этом отношение PAX2 к DEFB1 составляет, по меньшей мере, 100:1.
Также предлагается способ диагностики внутриэпителиальной неоплазии простаты (PIN) у субъекта. Способ включает определение в клетках из простаты субъекта уровней PAX2 и бета-DEFB1, при этом отношение PAX2 к DEFB1 составляет, по меньшей мере, 40:1 и менее чем 100:1.
В некоторых других вариантах отношение экспрессии PAX2-к-DEFB1 (на уровне РНК) в молочной железе используют для того, чтобы отличить незлокачественные (доброкачественные и/или предраковые) и злокачественные состояния молочной железы у субъекта. В одном варианте отношение PAX2-к-DEFB1 менее 100:1 является показателем незлокачественного (нормального) и/или предракового (внутриэпителиальная неоплазия молочной железы (MIN)) состояния, и отношение PAX2-к-DEFB1, по меньшей мере, 100:1 является показателем рака молочной железы.
Также предлагается способ диагностики рака молочной железы у субъекта. Способ включает определение в клетках из молочной железы субъекта уровней PAX2 и DEFB1, при этом отношение PAX2 к DEFB1, по меньшей мере, 100:1 является показателем рака молочной железы у субъекта. Также предлагается способ диагностики незлокачественных состояний молочной железы (нормального и/или MIN) у субъекта. Способ включает определение в клетках из молочной железы субъекта уровней PAX2 и DEFB1, при этом отношение PAX2 к DEFB1 менее чем 100:1 является показателем незлокачественных состояний молочной железы у субъекта.
В одном варианте способ дополнительно включает определение статуса рецептора эстрогена/рецептора прогестерона (ER/PR) в клетках, полученных из ткани молочной железы с определенным состоянием молочной железы. Статус ER/PR ткани молочной железы можно использовать в сочетании с отношением PAX2-к-DEFB1 в той же ткани для определения состояний молочной железы у субъекта.
В используемом далее в настоящем описании смысле термин «внутриэпителиальная неоплазия молочной железы» включает дольковую внутриэпителиальную неоплазию и протоковую внутриэпителиальную неоплазию.
Способы мониторинга и диагностики согласно настоящему изобретению дают клиницистам средство прогнозирования наличия инициированной или предзлокачественной ткани. Кандидатами для такого тестирования являются пациенты с высоким риском (на основании возраста, расы) развития злокачественной опухоли. Что касается диагностики, то положительный или отрицательный тест на PAX2 затем может сопровождаться дополнительным скринингом в отношении биомаркера для определения места злокачественной опухоли. Кроме того, такие пациенты могут быть кандидатами на лечение модуляторами PAX2/DEFB1. Альтернативно, такой тест можно использовать на пациентах (например, пациентах с трижды негативным раком молочной железы) в качестве меры эффективности терапии злокачественной опухоли, чтобы определить ход лечения или контролировать рецидивы злокачественной опухоли.
В качестве другого примера пациенты, у которых имеются возможные показатели злокачественной опухоли, такие как определение узелков в простате в ходе ректального пальпирования врачом, или пациенты, у которых внезапно произошло повышение PSA, часто находятся в состоянии «ожидания под наблюдением». Часто бывает трудно установить, имеют ли такие пациенты злокачественную опухоль или у них будет развиваться злокачественная опухоль. Определение отношения PAX2-к-DEFB1 в образцах, таких как плазма/сыворотка, полученных от таких пациентов, можно применять в качестве помощи при решении вопроса о получении биопсии у людей с предполагаемым раком простаты, что может привести к уменьшению количества ненужных операций взятия биопсии и более раннему вмешательству в их заболевание.
Рак простаты
Скрининг рака простаты в настоящее время состоит из ректального исследования и измерения уровней специфичного для простаты антигена (PSA). Такие способы не обладают специфичностью, так как пальцевое ректальное исследование имеет значительную вариабельность в зависимости от исследователя, и уровни PSA могут быть повышены при доброкачественной гиперплазии простаты (BPH), воспалении простаты и других состояниях.
Рак простаты можно оценивать, используя систему Глисона (Gleason, et al. 1966). В системе использована структура ткани, а не цитологические признаки. Используют баллы от 1 до 5 (от хорошо до плохо дифференцированной) и общие оценки областей наиболее частых и более тяжелых поражений суммируют. Оценки в баллах по шкале Глисона дают прогностическую информацию, которая может быть полезной в дополнение к оценке стадии опухоли (стадирование). Оценки в баллах по шкале Глисона от 2 до 4 и от 8 до 10 имеют высокую прогностическую ценность, но примерно три четверти опухолей имеют промежуточные значения.
Для определения стадий рака простаты используют две главных системы: систему TNM и систему Джеветта (Benson and Olsson, et al. 1989). При стадировании учитывают любое метастатическое распространение опухоли, что оказывается сложным, так как трудно оценить либо вовлечение регионального лимфатического узла, либо локальную инвазию. Размер опухоли также трудно измерить, поскольку опухолевую ткань невозможно макроскопически отличить от нормальной ткани простаты и поскольку предстательная железа не имеет отчетливой капсулы и окружена слоем фиброзной жировой ткани.
Четыре категории описывают стадии опухолей простаты (T) в диапазоне от T1 до T4. В случае T1 злокачественная опухоль является микроскопической, односторонней и непальпируемой. Врач не может ни прощупывать опухоль, ни видеть ее при визуализации, такой как трансректальное ультразвуковое исследование. Заболевание может быть обнаружено при лечении BPH, или оно может быть подтверждено с использованием пункционной биопсии, которую делают при повышенном PSA. В случае T2 врач может прощупать злокачественную опухоль при DRE. По-видимому, заболевание затрагивает предстательную железу с одной или обеих сторон железы. В случае T3 злокачественная опухоль распространяется на близлежащую ткань за пределами железы. В случае T4 злокачественная опухоль распространяется на другие части тела.
Следовательно, имеющиеся в настоящее время способы скрининга неудовлетворительны; не существует надежного способа диагностики рака простаты, или прогнозирования, или предотвращения его возможного метастатического распространения, которое является основной причиной смерти большинства пациентов.
Рак молочной железы
Широко используемые способы скрининга рака молочной железы включают самостоятельный и клинический осмотры молочной железы, рентгеновскую маммографию и магнитно-резонансную томографию молочной железы (MRI). Наиболее современным способом скрининга рака молочной железы является ультразвуковая компьютерная томография, в которой используют звуковые волны для создания трехмерного изображения и выявления рака молочной железы без применения опасного излучения, используемого в рентгеновской маммографии. Также можно использовать генетическое тестирование. Генетическое тестирование в случае рака молочной железы обычно заключается в исследовании мутаций в генах BRCA. Такой способ не является широко рекомендованным, за исключением пациентов с повышенным риском рака молочной железы.
Частота заболеваемости раком молочной железы, лидирующей причины смерти у женщин, постепенно возрастала в Соединенных Штатах на протяжении последних тридцати лет. Хотя патогенез рака молочной железы неясен, трансформация нормального эпителия молочной железы к злокачественному фенотипу может быть результатом генетических факторов, особенно у женщин в возрасте до 30 лет (Miki et al., 1994, Science, 266: 66-71). Открытие и характеристика BRCA1 и BRCA2 недавно расширили наши знания о генетических факторах, которые могут вносить вклад в наследственный рак молочной железы. Мутации зародышевой линии в указанных локусах ассоциированы с 50-85% риском рака молочной железы и/или яичника на протяжении всей жизни (Casey, 1997, Curr. Opin. Oncol. 9:88-93; Marcus et al., 1996, Cancer 77: 697-709). Однако и другие негенетические факторы, вероятно, также оказывают значимое влияние на этиологию заболевания. Заболеваемость и смертность от рака молочных железы, независимо от происхождения, в значительной степени повышаются, если рак не выявлен на ранней стадии его прогрессирования. Соответственно, значительные усилия были сфокусированы на раннем выявлении клеточной трансформации и образования опухоли в ткани молочной железы.
В настоящее время основным способом идентификации рака молочной железы является способ на основе выявления присутствия плотной опухолевой ткани. Такое выявление можно осуществить с различной степенью эффективности прямым наружным осмотром молочной железы или посредством маммографии или другими способами визуализации с использованием рентгеновского излучения (Jatoi, 1999, Am. J. Surg. 177: 518-524). Однако последний способ требует значительных затрат. Каждый раз при маммографии пациент подвергается небольшому риску появления опухоли молочной железы, индуцированной ионизирующими свойствами излучения, используемого во время тестирования. Кроме того, способ является дорогостоящим, и субъективные интерпретации специалистов могут приводить к расхождению результатов, например, в одном исследовании обнаружены большие клинические расхождения примерно для одной трети набора маммограмм, которые были интерпретированы индивидуально группой рентгенологов, проводящих исследование. Кроме того, многие женщины признают, что процедура маммографии является мучительным испытанием. Соответственно, Национальный институт рака не рекомендовал маммограммы для женщин в возрасте до пятидесяти лет, так как для такой группы не столь вероятно развитие рака молочной железы, как для женщин более старшего возраста. Однако следует отметить, что хотя только около 22% случаев рака молочной железы возникает у женщин в возрасте до пятидесяти лет, данные свидетельствуют, что рак молочной железы является более агрессивным у женщин в пременопаузе.
PAX2
Гены PAX представляют семейство из девяти генов, регулирующих развитие, кодирующих ядерные факторы транскрипции. Они играют важную роль в эмбриогенезе и экспрессируются очень упорядоченно в пространственном и временном отношении. Все они содержат «парный бокс» из 384 пар оснований, кодирующий ДНК-связывающий домен, который высоко консервативен в эволюции (Stuart, ET, et al., 1994). Влияние генов Pax на процессы развития продемонстрировано многочисленными природными синдромами у мышей и человека, которые можно объяснить непосредственно гетерозиготной недостаточностью гена Pax. Последовательность PAX2 приведена в публикации Dressier с соавторами, 1990. Аминокислотные последовательности белка PAX2 человека и его вариантов, а также последовательности ДНК, кодирующие белки, указаны в SEQ ID NO: 58-69 (SEQ ID NO: 58, аминокислотная последовательность, кодируемая экзоном 1 гена PAX2 человека; SEQ ID NO: 59, промотор гена PAX2 человека и экзон 1; SEQ ID NO:60, аминокислотная последовательность PAX2 человека; SEQ ID NO: 61, гена PAX2 человека; SEQ ID NO: 62, аминокислотная последовательность варианта b гена PAX2 человека; SEQ ID NO: 63, вариант b гена PAX2 человека; SEQ ID NO: 64, аминокислотная последовательность варианта c гена PAX2 человека; SEQ ID NO: 65, вариант c гена PAX2 человека; SEQ ID NO: 66, аминокислотная последовательность варианта d гена PAX2 человека; SEQ ID NO: 67, вариант d гена PAX2 человека; SEQ ID NO: 68, аминокислотная последовательность варианта e гена PAX2 человека; SEQ ID NO: 69, вариант e гена PAX2 человека).
Примеры злокачественных опухолей, в которых была выявлена экспрессия PAX2, перечислены в таблице 1.
Злокачественные опухоли, экспрессирующие PAX2
DEFB1
Бета-дефензины являются катионными пептидами с широким спектром противомикробной активности, которые представляют собой продукты эпителия и лейкоцитов. Указанные продукты отдельных генов, содержащих два экзона, экспрессируются на поверхности эпителия и секретируются в определенных местах, включая кожу, роговицу, язык, десну, слюнные железы, пищевод, кишечник, почку, мочеполовые пути и респираторный эпителий. До настоящего времени у человека идентифицировано и охарактеризовано пять генов бета-дефензина эпителиального происхождения: DEFB1 (Bensch с соавторами, 1995), DEFB 2 (Harder с соавторами, 1997), DEFB3 (Harder с соавторами, 2001; Jia с соавторами, 2001), DEFB4 и HE2/EP2.
Первичная структура каждого продукта гена бета-дефензина характеризуется небольшим размером, наличием мотива из шести цистеинов, высоким катионным зарядом и тонкими различиями помимо указанных признаков. Наиболее характерным признаком белков дефензина является их мотив, состоящий из шести цистеинов, который образует сеть из трех дисульфидных связей. Три дисульфидные связи в белках бета-дефензина расположены между C1-C5, C2-C4 и C3-C6. Наиболее распространенный интервал между близлежащими остатками цистеина составляет 6, 4, 9, 6, 0. Интервал между цистеинами в белках бета-дефензина может изменяться на одну или две аминокислоты, за исключением C5 и C6, расположенных ближе всего к карбоксильному концу. Во всех известных генах бета-дефензина позвоночных такие два остатка цистеина расположены рядом друг с другом.
Вторым характерным признаком белков бета-дефензина является их небольшой размер. Каждый ген бета-дефензина кодирует препробелок, который имеет размер в диапазоне от 59 до 80 аминокислот со средним размером 65 аминокислот. Затем такой генный продукт расщепляется посредством неизвестного механизма с образованием зрелого пептида, который имеет размер в диапазоне от 36 до 47 аминокислот со средним размером 45 аминокислот. Исключениями из таких диапазонов являются продукты гена EP2/HE2, которые содержат мотив бета-дефензина и экспрессируются в эпидидимисе.
Третьим характерным признаком белков бета-дефензина является высокая концентрация катионных остатков. Количество положительно заряженных остатков (аргинин, лизин, гистидин) в зрелом пептиде находится в диапазоне от 6 до 14 со средним значением 9.
Наконец, следующим характерным признаком продуктов генов бета-дефензина является их разнообразная первичная структура, но при этом очевидный консерватизм третичной структуры. Помимо шести цистеинов нет отдельной аминокислоты в заданном положении, консервативной во всех известных представителях данного семейства белков. Однако имеются положения, которые являются консервативными, которые, по-видимому, важны для вторичной и третичной структуры и функции.
Несмотря на большое разнообразие первичной аминокислотной последовательности белков бета-дефензина, некоторые данные свидетельствуют, что третичная структура такого семейства белков является консервативной. Ядром структуры является трехнитевой, антипараллельный бета-слой, как, например, в случае белков, кодируемых BNBD-12 и DEFB2. Три бета-нити связаны бета-поворотом и петлей в виде альфа-шпильки, и вторая бета-нить также содержит бета-петлю. Когда такие структуры уложены в правильную третичную структуру, случайные последовательности катионных и гидрофобных остатков, по-видимому, концентрируются, образуя две поверхности глобулярного белка. Одна поверхность является гидрофильной и содержит много положительно заряженных боковых цепей, а другая является гидрофобной. В растворе белок HBD-2, кодируемый геном DEFB2, имел альфа-спиральный участок вблизи N-конца, что ранее не было описано для структур альфа-дефензинов или бета-дефензина BNBD-12 в растворе. Аминокислоты, боковые цепи которых направлены к поверхности белка, являются менее консервативными в белках бета-дефензина, тогда как аминокислотные остатки в трех бета-нитях центрального бета-слоя являются более высоко консервативными.
Пептиды бета-дефензина продуцируются в виде пре-про-пептидов и затем расщепляются, высвобождая C-концевой активный фрагмент пептида; однако пути внутриклеточного процессинга, хранения и высвобождения пептидов бета-дефензина человека в эпителии дыхательных путей неизвестны.
Определение отношения экспрессии PAX2-к-DEFB1
Уровни экспрессии PAX2 и DEFB1 в ткани могут быть измерены любым способом, известным в данной области. В некоторых вариантах уровни экспрессии PAX2 и DEFB1 в ткани-мишени определяют благодаря определению уровней PAX2 и DEFB1 в клетке или клетках, полученных непосредственно из ткани-мишени, такой как образец биопсии. В других вариантах уровни экспрессии PAX2 и DEFB1 в ткани-мишени определяют благодаря непосредственному определению уровней PAX2 и DEFB1 в некоторых тканях организма, таких как кровь или плазма.
В некоторых вариантах уровни экспрессии PAX2 и DEFB1 в простате или молочной железе субъекта могут быть измерены с использованием образца клеток простаты или молочной железы. Образцы клеток включают образцы биопсии простаты или молочной железы и образец крови. Биопсия представляет собой способ, при котором небольшие образцы ткани извлекают из органа-мишени для дальнейшего анализа. Биопсию простаты обычно осуществляют в том случае, когда оценка в баллах в тесте крови PSA повышается до уровня, который ассоциирован с возможным наличием рака простаты. Подобным образом, биопсию молочной железы обычно осуществляют у пациентов с уплотнениями молочной железы или подозрительными маммограммами.
Уровни экспрессии генов можно оценить на уровне РНК или белка. РНК можно измерить, например, с использованием ДНК-матриц, ОТ-ПЦР и Нозерн-блоттинга. Уровни белка могут быть измерены с использованием иммуноанализов и анализов ферментов. В некоторых вариантах отношение экспрессии PAX2-к-DEFB1 определяют посредством определения уровня экспрессии гена PAX2 по сравнению с уровнем экспрессии контрольного гена, определения уровня экспрессии гена DEFB1 по сравнению с уровнем экспрессии того же контрольного гена, и вычисления отношения экспрессии PAX2-к-DEFB1 на основании уровней экспрессии PAX2 и DEFB1. В одном варианте контрольным геном является ген глицеральдегид-3-фосфатдегидрогеназы.
Олигонуклеотидная микроматрица
Олигонуклеотидная микроматрица состоит из расположенной в определенном порядке серии множества микроскопических пятен олигонуклеотидов, называемых признаками, каждое из которых содержит небольшое количество (обычно в пикомолярном диапазоне) конкретной олигонуклеотидной последовательности. Конкретная олигонуклеотидная последовательность может представлять собой короткий участок гена или другой олигонуклеотидный элемент, который используют в качестве зондов для гибридизации с образцом кДНК или кРНК в условиях высокой жесткости. Гибридизацию зонд-мишень обычно выявляют и количественно оценивают с использованием основанной на флуоресценции регистрации меченных флуорофором мишеней, определяя относительное содержание последовательностей нуклеиновых кислот в мишени.
Зонды обычно связывают с твердой поверхностью ковалентной связью с химической матрицей (через эроксисилан, аминосилан, лизин, полиакриламид или другие). Твердой поверхностью может быть стекло, или кремниевый чип, или микроскопические шарики. Олигонуклеотидные матрицы отличаются от других типов микроматрицы только тем, что они позволяют либо измерять нуклеотиды, либо использовать олигонуклеотид в качестве части такой системы регистрации.
Для выявления экспрессии гена в ткани-мишени или клетках-мишенях с использованием олигонуклеотидной матрицы, представляющую интерес нуклеиновую кислоту очищают из ткани или клеток мишени. Нуклеотид может быть полностью РНК для определения профиля экспрессии, ДНК для сравнительной гибридизации, или ДНК/РНК, связанной с конкретным белком, который подвергают иммунопреципитации (ChIP на чипе) для эпигенетических исследований или исследований регуляции.
В одном варианте суммарную РНК выделяют (суммарную, так как она является и ядерной, и цитоплазматической), используя экстракцию смесью тиоцианат гуанидиния-фенол-хлороформ (например, тризолом). Очищенную РНК можно анализировать в отношении качества (например, капиллярным электрофорезом) и количества (например, используя нанокапельный спектрометр). Суммарную РНК обратно транскрибируют в ДНК, используя либо поли-T-праймеры, либо случайные праймеры. ДНК-продукты необязательно могут быть амплифицированы в ПЦР. Метку добавляют к продукту амплификации либо на стадии ОТ, либо на дополнительной стадии после амплификации, если таковая имеется. Метка может быть флуоресцирующей меткой или радиоактивной меткой. Затем меченые продукты ДНК гибридизуют с микроматрицей. Затем микроматрицу промывают и сканируют. Уровень экспрессии представляющего интерес гена определяют на основе результатов гибридизации, используя способ, хорошо известный в данной области.
Иммуноанализы
Иммуноанализы в наиболее простом и прямом смысле представляют собой анализы связывания, которые заключаются в связывании между антителами и антигеном. Известно много типов и форм иммуноанализов, и все они подходят для выявления предложенных биомаркеров. Примерами иммуноанализов являются твердофазные иммуноферментные анализы (ELISA), радиоиммуноанализы (РИА), радиоактивные анализы иммунопреципитации (RIPA), иммунологические анализы на основе улавливания на шариках, Вестерн-блоттинг, дот-блоттинг, анализы сдвига в геле, проточная цитометрия, белковые матрицы, мультиплексный анализ на шариках, магнитное улавливание, визуализация in vivo, резонансный перенос энергии флуоресценции (FRET) и восстановление/локализации флуоресценции после фотоотбеливания (FRAP/FLAP).
В общем, иммуноанализы заключаются в осуществлении контакта образца, который предположительно содержит представляющую интерес молекулу (такую как описанные биомаркеры), с антителом к представляющей интерес молекуле или в осуществлении контакта антитела к представляющей интерес молекуле (такой как антитела к описанным биомаркерам) с молекулой, которая может быть связана антителом, в зависимости от обстоятельств, в условиях, которые обеспечивают эффективное образование иммунокомплексов. Во многих формах иммуноанализа композицию образец-антитело, например срез ткани, планшет для ELISA, дот-блот или Вестерн-блот, затем можно промыть, чтобы удалить любые неспецифично связанные виды антител, обеспечивая специфичное связывание только таких антител в выявляемых первичных иммунных комплексах.
Радиоактивный анализ иммуннопреципитации (RIPA) является чувствительным анализом, в котором используют радиоактивно меченые антигены для выявления специфичных антител в сыворотке. Антигенам дают возможность взаимодействовать с сывороткой и затем преципитируют, используя специальный реагент, такой как, например, шарики на основе белок A-сефарозы. Затем связанный радиоактивно меченный иммунопреципитат обычно анализируют, используя гель-электрофорез. Радиоактивный анализ иммуннопреципитации (RIPA) часто применяют в качестве подтверждающего теста в случае диагностики наличия ВИЧ-антител. RIPA также называют в данной области анализом по Фарру, анализом преципитина, радиоактивным анализом иммунопреципитина; радиоиммунным анализом преципитации; и анализом радиоиммунопреципитации.
Также предполагаются иммуноанализы, в которых белок или антитело, специфичное для белка, связывают с твердой подложкой (например, пробиркой, лункой, шариком или ячейкой), чтобы уловить из образца представляющее интерес антитело или белок, соответственно, в сочетании со способом выявления белка или антитела, специфичного для белка, на подложке. Примерами таких иммуноанализов являются радиоиммуноанализ (РИА), твердофазный иммуноферментный анализ (ELISA), проточная цитометрия, белковая матрица, мультиплексный анализ на шариках и магнитное улавливание.
Белковые матрицы представляют собой системы анализа связывания лиганда с твердой фазой, в которых используют иммобилизованные белки на поверхностях, включая стекло, мембраны, лунки планшетов для микротитрования, планшеты масс-спектрометра и шарики или другие частицы. Анализы являются высокопараллельными (мультиплексными) и часто в минимальном масштабе (микроматрицы, белковые чипы). Преимуществами таких анализов являются быстрота и возможность автоматизации, высокая чувствительность, экономия реагентов и получение достаточно большого количества данных в одном эксперименте. Важным является биоинформационное сопровождение; обработка данных требует сложной компьютерной программы и сравнительного анализа данных. Однако можно адаптировать компьютерную программу, используемую для матриц ДНК, что возможно в случае применения многих систем оборудования и регистрации.
Матрицы для улавливания составляют основу диагностических чипов и матриц для определения профиля экспрессии. В данном случае используют реагенты для высокоаффинного улавливания, такие как обычные антитела, отдельные домены, сконструированные каркасы, пептиды или аптамеры нуклеиновых кислот, для связывания и выявления конкретных лигандов-мишеней с высокой пропускной способностью. Матрицы антител коммерчески доступны. Кроме обычных антител, для матриц также можно использовать Fab- и scFv-фрагменты, отдельные V-домены верблюдовых или сконструированные эквиваленты человека (Domantis, Waltham, MA).
Небелковые молекулы для улавливания, в частности, аптамеры однонитевой нуклеиновой кислоты, которые связывают белковые лиганды с высокой специфичностью и аффинностью, также используют в матрицах (SomaLogic, Boulder, CO). Аптамеры выбирают из библиотек олигонуклеотидов способом SelexTM, и их взаимодействие с белком может быть усилено ковалентным связыванием благодаря включению бромированного дезоксиуридина и УФ-активированного поперечного сшивания (фотоаптамеры). Поперечное фотосшивание с лигандом снижает перекрестную реактивность аптамеров вследствие специфичных стерических требований. Аптамеры имеют преимущества, обусловленные простым получением в результате автоматизированного синтеза олигонуклеотида и стабильностью и устойчивостью ДНК; на матрицах фотоаптамеров можно использовать универсальные флуоресцирующие красители белков для выявления связывания.
Альтернативой матрице улавливающих молекул является матрица, получаемая с использованием методики «молекулярного импринтинга», в ходе которого пептиды (например, из C-концевых областей белков) используют в качестве матриц для создания структурно комплементарных специфичных для последовательности полостей в полимеризуемом матриксе; полости затем могут специфично улавливать (денатурированные) белки, которые имеют соответствующую первичную аминокислотную последовательность (ProteinPrintTM, Aspira Biosystems, Burlingame, CA).
Другой методикой, которую можно использовать диагностически и при определении профиля экспрессии, является матрица ProteinChip® (Ciphergen, Fremont, CA), в случае которой твердофазные хроматографические поверхности связывают белки со сходными характеристиками заряда или гидрофобности из смеси, такой как плазма или экстракты опухолей, и для выявления удерживаемых белков используют SELDI-TOF-масс-спектрометрию.
Другая применимая методика заключается в использовании крупномасштабных функциональных чипов, сконструированных посредством иммобилизации большого количества очищенных белков на чипе и мультиплексный анализ на шариках.
Антитела
Термин «антитела» используют в настоящем описании в широком смысле, и термин включает как поликлональные, так и моноклональные антитела. Кроме интактных молекул иммуноглобулинов в термин «антитела» также включены фрагменты или полимеры молекул иммуноглобулинов и человеческие или гуманизированные варианты молекул иммуноглобулинов или их фрагменты, при условии, что их выбирают на основе их способности взаимодействовать, например, с PAX2 или DEFB1, так что происходит ингибирование взаимодействия PAX2 с DEFB1. Также описаны антитела, которые связывают описанные области PAX2 или DEFB1, вовлеченные во взаимодействие между PAX2 и DEFB1. Антитела можно тестировать в отношении требуемой активности, используя анализы in vitro, описанные в настоящей публикации, или аналогичные способы, и затем тестируют их терапевтические и/или профилактические активности in vivo согласно известным способам клинического исследования.
Моноклональные антитела согласно настоящему изобретению, в частности, включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, полученных из конкретного вида или относящихся к конкретному классу или подклассу антител, тогда как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям в антителах, полученных от другого вида, или относящихся к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую антагонистическую активность (см., например, патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)).
В используемом в настоящем описании смысле термин «антитело» или «антитела» также может относиться к человеческому антителу и/или гуманизированному антителу. Многие антитела животных, отличных от человека (например, антитела, полученные от мышей, крыс или кроликов), являются в природе антигенными для человека, и, следовательно, могут вызывать нежелательные иммунные ответы при введении человеку. Следовательно, использование человеческих или гуманизированных антител в способах служит для уменьшения вероятности того, что антитело, введенное человеку, будет вызывать нежелательный иммунный ответ. Способы гуманизации антител животных, отличных от человека, хорошо известны в данной области.
Фармакогеномика
В другом варианте профили экспрессии PAX2 и/или DEFB1 используют для определения фармакогеномики рака молочной железы. Фармакогеномика относится к взаимосвязи между генотипом индивидуума и ответом такого индивидуума на чужеродное соединение или лекарственное средство. Различия в метаболизме терапевтических средств может приводить к тяжелой токсичности или неблагоприятному исходу терапии из-за изменения зависимости между дозой и концентрацией в крови фармакологически активного лекарственного средства. Таким образом, лечащий врач или клиницист может использовать сведения, полученные в соответствующих фармакогеномных исследованиях, при определении того, вводить ли противоопухолевое средство, а также при подборе дозы и/или терапевтической схемы лечения противоопухолевым лекарственным средством.
Фармакогеномика изучает клинически значимые наследственные изменения ответа на лекарственные средства вследствие измененного распределения лекарственного средства и аномального действия у пораженных пациентов. В общем, можно выделить два типа фармакогенетических состояний. Генетические состояния, передаваемые в виде одного фактора, изменяющего путь, которым лекарственные средства действуют на организм (измененное действие лекарственного средства), или генетические состояния, передаваемые в виде отдельных факторов, изменяющих путь, которым организм действует на лекарственные средства (измененный метаболизм лекарственного средства). Такие фармакогенетические состояния могут возникать либо в виде редких генетических дефектов, либо в виде встречающегося в природе полиморфизма. Например, недостаточность глюкозо-6-фосфатдегидрогеназы (G6PD) является распространенной наследственной энзимопатией, при которой основным клиническим осложнением является гемолиз после приема внутрь окислительных лекарственных средств (противомалярийных средств, сульфонамидов, анальгетиков, нитрофуранов) и употребления конских бобов.
Один из используемых в фармакогеномике способов идентификации генов, который позволяет предсказать ответ на лекарственное средство, известный как «поиск ассоциации по всему геному», главным образом, основан на использовании карты высокого разрешения генома человека, состоящей из уже известных связанных с генами сайтов (например, карта «биаллельных» генных маркеров, которая состоит из 60000-100000 полиморфных или вариабельных сайтов в геноме человека, каждый из которых имеет два варианта). Такую генетическую карту высокого разрешения можно сравнить с картой генома каждого из статистически значимого количества субъектов, принимающих участие в испытании лекарственного средства в фазе II/III, чтобы идентифицировать гены, ассоциированные с конкретным наблюдаемым ответом на лекарственное средство или побочным эффектом. Альтернативно, такая карта высокого разрешения может быть создана на основе сочетания нескольких десятков миллионов известных однонуклеотидных полиморфизмов (SNP) в геноме человека. В используемом в настоящем описании смысле термин «SNP» означает обычное изменение, которое происходит в одном нуклеотидном основании на участке ДНК. Например, SNP может возникать с частотой один на каждую 1000 оснований ДНК. SNP может быть вовлечен в патологический процесс. Однако подавляющее большинство SNP может быть не ассоциировано с заболеваниями. С учетом генетической карты, основанной на возникновении таких SNP, индивидуумов можно отнести к определенным генетическим категориям в зависимости от конкретной картины SNP в их индивидуальных геномах. Таким образом, схема лечения может быть адаптирована для групп генетически сходных индивидуумов с учетом признаков, которые могут быть общими у таких генетически сходных индивидуумов. Таким образом, картирование PAX2 и/или DEFB1 на картах SNP для пациентов с раком молочной железы может обеспечить более простую идентификацию таких генов согласно генетическим способам, описанным в настоящей публикации.
Альтернативно, можно применять способ, называемый «способом на основе генов-кандидатов», чтобы идентифицировать гены, которые позволяют предсказать ответ на лекарственное средство. Согласно такому способу, если ген, который кодирует мишень лекарственного средства, известен, то все обычные варианты такого гена можно довольно легко идентифицировать в популяции и можно определить, ассоциировано ли наличие одного варианта гена по сравнению с другим с конкретным ответом на лекарственное средство.
В качестве иллюстративного варианта активность ферментов, метаболизирующих лекарственное средство, является основным фактором, определяющим как интенсивность, так и продолжительность действия лекарственного средства. Открытие генетического полиморфизма ферментов, метаболизирующих лекарственные средства (например, N-ацетилтрансферазы 2 (NAT 2) и ферментов цитохромов P450, CYP2D6 и CYPZC19) позволило объяснить, почему некоторые субъекты не получают ожидаемых эффектов от применения лекарственных средств или почему у них наблюдается усиленный ответ на лекарственное средство и серьезная токсичность после приема стандартной и безопасной дозы лекарственного средства. Такие полиморфизмы экспрессируются, давая два фенотипа в популяции, быстрых метаболизаторов и медленных метаболизаторов. Распространенность фенотипа медленных метаболизаторов различны в разных популяциях. Например, ген, кодирующий CYP2D6, является высоко полиморфным, и было идентифицировано несколько мутаций у медленных метаболизаторов, которые приводили к отсутствию функционального CYP2D6. Медленные метаболизаторы по CYP2D6 и CYP2C19 довольно часто проявляют усиленный ответ на лекарственное средство и побочные эффекты при получении ими стандартных доз. Если метаболит является активным терапевтическим компонентом, то у медленных метаболизаторов не наблюдают терапевтического ответа, как показано в случае анальгезирующего действия кодеина, опосредованного его образуемым CYP2D6 метаболитом морфином. Другим крайним случаем являются так называемые сверхбыстрые метаболизаторы, которые не отвечают на стандартные дозы. Недавно была выявлено, что молекулярной основой сверхбыстрого метаболизма является амплификация гена CYP2D6.
Альтернативно, можно применять способ, называемый «определением профиля экспрессии генов», чтобы идентифицировать гены, которые позволяют прогнозировать ответ лекарственного средства. Например, экспрессия гена животного, которому вводили дозу лекарственного средства, может быть показателем того, были ли запущены генные пути, связанные с токсичностью.
Информацию, полученную в результате применения более чем одного из описанных выше способов фармакогеномики, можно использовать для определения подходящей дозы и схем лечения в случае профилактического или терапевтического лечения индивидуума. Такие сведения в применении к дозированию или выбору лекарственного средства, могут позволить избежать неблагоприятных реакций или неблагоприятных исходов терапии и, следовательно, повысить терапевтическую или профилактическую эффективность при лечении субъекта с определенным состоянием молочной железы.
В одном варианте профили экспрессии PAX2 и/или DEFB1, а также статус ER/PR у субъекта используют для определения подходящих схем лечения пациента с определенным состоянием молочной железы.
В другом варианте уровень экспрессии PAX2 (обычно определяют по отношению к контрольному гену, такому как ген актина или ген GAPDH), используют у пациентов с трижды негативным раком молочной железы (т.е., негативным по рецептору эстрогена (ER), негативным по рецептору прогестерона (PR), негативным по рецептору 2 эпидермального фактора роста человека (HER2)), чтобы измерить эффективность противораковой терапии, определить ход лечения или контролировать рецидивы злокачественной опухоли.
Диагностические наборы
Другой аспект настоящего изобретения относится к набору для мониторинга состояний молочной железы. В одном варианте набор для мониторинга состояний молочной железы содержит: одну или несколько пар олигонуклеотидных праймеров для выявления экспрессии PAX2 в образце ткани, одну или несколько пар олигонуклеотидных праймеров для выявления экспрессии DEFB1 в образце ткани и инструкции о том, как определить отношение экспрессии PAX2-к-DEFB1 в образце ткани с использованием праймеров. В другом варианте одна или несколько пар олигонуклеотидных праймеров для выявления экспрессии PAX2 содержат пару олигонуклеотидных праймеров, выбранную из группы, состоящей из последовательностей SEQ ID NO: 43 и 47, SEQ ID NO: 44 и 48, и SEQ ID NO: 45 и 49. В другом варианте одна или несколько пар олигонуклеотидных праймеров для выявления экспрессии DEFB1 содержат последовательности SEQ ID NO: 35 и 37.
В другом варианте набор дополнительно содержит одну или несколько пар контрольных олигонуклеотидных праймеров. В одном варианте одна или несколько пар контрольных олигонуклеотидных праймеров содержат олигонуклеотидные праймеры для выявления экспрессии β-актина. В предпочтительном варианте олигонуклеотидные праймеры для выявления экспрессии β-актина содержат последовательности SEQ ID NO: 34 и 36.
В другом варианте одна или несколько пар контрольных олигонуклеотидных праймеров содержат олигонуклеотидные праймеры для выявления экспрессии GAPDH. В предпочтительном варианте олигонуклеотидные праймеры для выявления экспрессии GAPDH содержат последовательности SEQ ID NO: 42 и 46.
В другом родственном варианте набор дополнительно содержит один или несколько реагентов для ПЦР-реакции.
В еще одном родственном варианте набор дополнительно содержит один или несколько реагентов для экстракции РНК.
В другом варианте набор для мониторинга состояний молочной железы содержит микроматрицу олигонуклеотидов, имеющую олигонуклеотидные зонды для выявления экспрессии PAX2 и DEFB1, и инструкции о том, как определить отношение экспрессии PAX2-к-DEFB1 в образце ткани с использованием микроматрицы олигонуклеотидов.
В родственном варианте набор дополнительно содержит реагенты для экстракции РНК из образца ткани.
Настоящее изобретение дополнительно проиллюстрировано следующими примерами, которые не следует рассматривать как ограничивающие. Содержание всех публикаций, патентов и опубликованных заявок на выдачу патента, цитированных на протяжении настоящего описания, а также фигуры и таблицы включены в настоящее описание в виде ссылки.
ПРИМЕР 1: БЕТА-ДЕФЕНЗИН-1 ЧЕЛОВЕКА ЯВЛЯЕТСЯ ЦИТОТОКСИЧНЫМ ПО ОТНОШЕНИЮ К ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ ПРОСТАТЫ НА ПОЗДНЕЙ СТАДИИ И ИГРАЕТ РОЛЬ В ПРОТИВООПУХОЛЕВОМ ИММУНИТЕТЕ ПРИ РАКЕ ПРОСТАТЫ
В данном примере DEFB1 клонировали в системе индуцируемой экспрессии, чтобы исследовать, какое влияние он оказывает на нормальные эпителиальные клетки простаты, а также на линии злокачественных клеток простаты, позитивные по рецептору андрогена (AR+) и негативные по рецептору андрогена (AR-). Индукция экспрессии DEFB1 приводила к снижению клеточного роста (AR-)-клеток DU145 и PC3, но не оказывала влияния на рост злокачественных (AR+)-клеток простаты LNCaP. DEFB1 также вызывал быструю индукцию опосредованного каспазой апоптоза. Данные, представленные в данном примере, являются первым доказательством его роли в естественном противоопухолевом иммунитете и свидетельствуют о том, что его утрата вносит вклад в прогрессирование опухоли при раке простаты.
Материалы и способы
Линии клеток: Линию клеток DU145 культивировали в среде DMEM, PC3 выращивали в среде F12 и LNCaP выращивали в среде RPMI (Life Technologies, Inc., Grand Island, NY). В ростовые среды для всех трех линий добавляли 10% (об./об.) фетальной сыворотки теленка (Life Technologies). Клетки hPrEC культивировали в основной для культивирования эпителия простаты (Cambrex Bio Science, Inc., Walkersville, MD). Все линии клеток поддерживали при 37°C и 5% CO2.
Образцы ткани и лазерная захватывающая микродиссекция: Ткани простаты, полученные от давших согласие пациентов, которые были подвергнуты радикальной простатэктомии, приобретали из банка опухолей онкологического центра Hollings в соответствии с одобренным экспертным советом организации протоколом. Протокол включал руководство по обработке, изготовлению срезов, гистологической характеристике, очистке РНК и ПЦР-амплификации образцов. После медико-патологического исследования замороженных срезов ткани осуществляли лазерную захватывающую микродиссекцию (LCM), чтобы убедиться, что анализируемые образцы ткани состояли из чистых популяций доброкачественных клеток простаты. Для каждого анализируемого среза ткани LCM осуществляли в трех разных областях, содержащих доброкачественную ткань, и собранные клетки затем объединяли.
Клонирование гена DEFB1: кДНК DEFB1 получали на основе РНК, используя обратную транскрипцию-ПЦР. ПЦР-праймеры конструировали так, чтобы они содержали сайты рестрикции ClaI и KpnI. ПЦР-продукты DEFB1 расщепляли ферментами рестрикции ClaI и KpnI и лигировали в клонирующий вектор TA. Вектором TA/DEFB1 затем трансфицировали E. coli, используя тепловой шок, и отдельные клоны собирали и размножали. Плазмиды выделяли, используя набор для выделения ДНК из культуры клеток Midiprep (Qiagen, Valencia, CA), и целостность последовательности подтверждали автоматизированным секвенированием. Затем фрагмент гена DEFB1 лигировали в pTRE2, расщепленную ClaI и KpnI, которая служила в качестве промежуточного вектора в целях ориентирования. Затем конструкцию pTRE2/DEFB1 расщепляли ApaI и KpnI, чтобы вырезать вставку DEFB1, которую лигировали в вектор pIND системы индуцируемой экспрессии экдизона (Invitrogen, Carlsbad, CA), также дважды расщепленный ApaI и KpnI. Конструкцией снова трансфицировали E. coli и отбирали и размножали отдельные клоны. Плазмиды выделяли и целостность последовательности pIND/DEFB1 снова подтверждали автоматизированным секвенированием.
Трансфекция: Клетки (1×106) высевали в 100-мм чашки Петри и выращивали в течение ночи. Затем клетки котрансфицировали, используя липофектамин 2000 (Invitrogen, Carlsbad, CA), 1 мкг плазмиды pVgRXR, которая экспрессирует гетеродимерный рецептор экдизона, и 1 мкг векторной конструкции pIND/DEFB1 или пустого контрольного вектора pIND в среде Opti-MEM (Life Technologies, Inc., Grand Island, NY).
Выделение РНК и количественная ОТ-ПЦР: Чтобы подтвердить экспрессию белка DEFB1 в клетках, трансфицированных конструкцией DEFB1, РНК собирали после 24-часового периода индукции понастероном A (Pon A). Коротко, суммарную РНК выделяли, используя систему для выделения суммарной РНК SV (Promega, Madison, WI), примерно из 1×106 клеток, собранных трипсинизацией. В данном случае клетки лизировали и суммарную РНК выделяли центрифугированием в центрифужных колонках. В случае клеток, собранных с использованием LCM, суммарную РНК выделяли, используя набор для выделения РНК PicoPure (Arcturus Biosciences, Mt. View, CA), следуя протоколу производителя. Суммарную РНК (0,5 мкг на реакцию) из обоих источников обратно транскрибировали в кДНК, используя случайные праймеры (Promega). Фермент обратную транскриптазу II AMV (500 единиц на реакцию; Promega) использовали для синтеза первой нити, а ДНК-полимеразу Tfl использовали для синтеза второй нити (500 единиц на реакцию; Promega) согласно протоколу производителя. В каждом случае 50 пг кДНК использовали для осуществления ПЦР-реакции. Двухстадийную КОТ-ПЦР осуществляли на кДНК, созданной с использованием обратной транскриптазы MultiScribe из системы обратной транскрипции TaqMan и смеси SYBR® Green PCR Master Mix (Applied Biosystems).
Пару праймеров для DEFB1 (таблица 2) создавали на основе опубликованной последовательности DEFB1 (GenBank, номер доступа U50930). Осуществляли сорок циклов ПЦР в стандартных условиях, используя температуру отжига 56°C. Кроме того, ген β-актина (таблица 3) амплифицировали в качестве гена домашнего хозяйства, чтобы нормализовать начальное содержание суммарной кДНК. Экспрессию DEFB1 вычисляли в виде соотношения экспрессии между DEFB1 и β-актином и сравнивали в линиях клеток, в которых экспрессия DEFB1 была индуцирована или не была индуцирована, а также в полученной с использованием LCM доброкачественной ткани простаты. В качестве негативного контроля также осуществляли реакции КОТ-ПЦР без матрицы кДНК. Все реакции проводили три раза в трех повторах.
Последовательности праймеров для КОТ-ПЦР


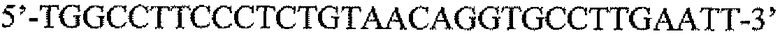
MTT-анализ жизнеспособности клеток: Чтобы исследовать влияние DEFB1 на рост клеток, осуществляли метаболические анализы с использованием бромида 3-[4,5-диметилтиазол-2-ил]-2,5-дифенилтетразолия (MTT). Клетки PC3, DU145 и LNCaP, котрансфицированные плазмидой pVgRXR и конструкцией pIND/DEFB1 или пустым вектором pIND, высевали в 96-луночный планшет по 1-5×103 клеток на лунку. Через двадцать четыре часа после посева ежедневно добавляли свежую ростовую среду, содержащую 10 мкМ понастерона A, чтобы индуцировать экспрессию DEFB1 в течение 24, 48 и 72 часов, затем осуществляли MTT-анализ согласно инструкциям производителя (Promega). Реакции осуществляли три раза в трех повторах.
Проточная цитометрия: Клетки PC3 и DU145, котрансфицированные системой экспрессии DEFB1, выращивали в 60-мм чашках и индуцировали в течение 12, 24 и 48 часов 10 мкМ понастероном A. После каждого периода инкубации среду из чашек собирали (сохраняя открепившиеся клетки) и объединяли с PBS, используемым для промывки чашек. Оставшиеся прикрепленные клетки собирали трипсинизацией и объединяли с открепленными клетками и PBS. Затем клетки осаждали при 4°C (500×g) в течение 5 минут, дважды промывали в PBS и ресуспендировали в 100 мкл 1-кратного буфера для связывания аннексина (0,1 М Hepes/NaOH при pH 7,4, 1,4 М NaCl, 25 мМ CaCl2), содержащего 5 мкл аннексина V-ФИТЦ и 5 мкл PI. Клетки инкубировали при комнатной температуре в течение 15 минут в темноте, затем разбавляли 400 мкл 1-кратного буфера для связывания аннексина и анализировали, используя FACscan (Becton Dickinson, San Jose, CA). Все реакции проводили три раза.
Микроскопический анализ: Морфологию клеток анализировали с использованием фазово-контрастной микроскопии. Клетки DU145, PC3 и LNCaP, не содержащие вектора, содержащие пустую плазмиду или плазмиду DEFB1, высевали в 6-луночные чашки для культивирования (BD Falcon, USA). На следующий день клетки, содержащие плазмиды, индуцировали в течение периода времени, составляющего 48 часов, средой, содержащей 10 мкМ понастерона A, тогда как в контрольные клетки вносили свежую среду. Затем клетки наблюдали под инвертированным микроскопом Zeiss IM 35 (Carl Zeiss, Germany). Фазово-контрастные изображения поля клеток получали, используя камеру SPOT Insight Mosaic 4.2 (Diagnostic Instruments, USA). Клетки исследовали посредством фазово-контрастной микроскопии при 32-кратном увеличении и цифровые изображения хранили в виде несжатых файлов TIFF и экспортировали в компьютерную программу Photoshop CS (Adobe Systems, San Jose, CA) для обработки изображения и представления печатных копий.
Выявление каспазы: Выявление каспазной активности в линиях злокачественных клеток простаты осуществляли, используя набор для выявления каспазы с использованием карбоксифлуоресценина APO LOGIXTM (Cell Technology, Mountain View, CA). Активные каспазы выявляли, используя ингибитор FAM-VAD-FMK, который необратимо связывается с активными каспазами. Коротко, клетки DU145 и PC3 (1,5-3×105), содержащие систему экспрессии DEFB1, высевали в 35-мм чашки с микролунками со стеклянным дном (Matek, Ashland, MA) и обрабатывали в течение 24 часов только средой или средой, содержащей PonA, как описано ранее. Затем 10 мкл 30-кратного рабочего разведения меченого карбоксифлуоресцеином пептид-фторметилкетона (FAM-VAD-FMK) добавляли к 300 мкл среды и добавляли в каждую 35-мм чашку. Затем клетки инкубировали в течение 1 часа при 37°C в 5% CO2. Затем среду аспирировали и клетки дважды промывали 2 мл 1-кратного рабочего разведения буфера для промывки. Клетки наблюдали при дифференциально-интерференционном контрасте (ДИК) или при возбуждении лазером при 488 нм. Сигнал флуоресценции анализировали, используя конфокальный микроскоп (Zeiss LSM 5 Pascal) и 63X иммерсионный ДИК-объектив с лазерным сканирующим модулем Vario 2 RGB.
Статистический анализ: Статистические различия оценивали, используя t-критерий Стьюдента для непарных выборок. Значения P определяли двухсторонними вычислениями, и P-значение меньше 0,05 считали статистически значимым.
Экспрессия DEFB1 в ткани простаты и линиях клеток: Уровни экспрессии DEFB1 измеряли посредством КОТ-ПЦР в доброкачественной и злокачественной ткани простаты, эпителиальных клетках простаты hPrEC и злокачественных клетках простаты DU145, PC3 и LNCaP. Экспрессию DEFB1 выявляли во всех доброкачественных клинических образцах. Среднее количество относительной экспрессии DEFB1 составляло 0,0073. Кроме того, относительная экспрессия DEFB1 в клетках hPrEC составляла 0,0089. Не было статистического различия в экспрессии DEFB1, выявленной в образцах доброкачественной ткани простаты и в hPrEC (фигура 1A). Анализ относительных уровней экспрессии DEFB1 в линиях злокачественных клеток простаты выявил значимо более низкие уровни в DU145, PC3 и LNCaP. В качестве следующей контрольной точки измеряли относительную экспрессию DEFB1 на срезе близлежащей к злокачественной опухоли ткани простаты от пациента №1215. Не было значимых различий в уровне экспрессии DEFB1, наблюдаемой в трех линиях рака простаты, по сравнению со злокачественной тканью простаты от пациента №1215 (фигура 1B). Кроме того, уровни экспрессии во всех четырех образцах были близки к негативным контролям без матрицы, что подтвердило, что эндогенная экспрессия DEFB1 является низкой или отсутствует (данные не показаны). Также осуществляли КОТ-ПЦР на линиях злокачественных клеток простаты, трансформированных системой экспрессии DEFB1. После 24-часового периода индукции относительные уровни экспрессии составляли 0,01360 в DU145, 0,01503 в PC3 и 0,138 в LNCaP. Продукты амплификации подтверждали, используя гель-электрофорез.
КОТ-ПЦР осуществляли в полученных с использованием LCM областях тканей, содержащих доброкачественную ткань, PIN и злокачественную опухоль. Относительная экспрессия DEFB1 составляла 0,0146 в доброкачественной области по сравнению с 0,0009 в злокачественной области (фигура 1C). Полученный результат означает 94% снижение, что снова демонстрирует значительную понижающую регуляцию экспрессии. Кроме того, анализ PIN выявил, что уровень экспрессии DEFB1 составлял 0,044, что означает снижение на 70%. Сравнение экспрессии у пациента №1457 со средним уровнем экспрессии, обнаруженным в доброкачественных областях у шести других пациентов (фигура 1A), выявило отношение 1,997, означающее почти в два раза более высокую экспрессию (фигура 1D). Однако отношение экспрессии составляло 0,0595 в PIN и 0,125 в злокачественной ткани по сравнению со средними уровнями экспрессии в доброкачественной ткани.
DEFB1 вызывает проницаемость и перемещение волн возбуждения в клеточной мембране: Индукция DEFB1 в линиях злокачественных клеток простаты приводила к значимому снижению количества клеток в DU145 и PC3, но не оказывала влияния на пролиферацию клеток LNCaP (фигура 2). В качестве негативного контроля наблюдали пролиферацию клеток во всех трех линиях, содержащих пустую плазмиду. Не было наблюдаемых изменений клеточной морфологии в клетках DU145, PC3 или LNCaP после добавления PonA. Кроме того, индукция DEFB1 приводила к морфологическим изменениям в DU145 и PC3. В данном случае клетки выглядели более округлыми, и наблюдалось перемещение волн возбуждения в мембране, которое является показателем гибели клеток. Апоптозные тельца также присутствовали в обеих линиях.
Экспрессия DEFB1 приводит к пониженной жизнеспособности клеток: MTT-анализ показал снижение жизнеспособности клеток под влиянием DEFB1 в клетках PC3 и DU145, но отсутствие значимого влияния на клетки LNCaP (фигура 3). Через 24 часа относительная жизнеспособность клеток составляла 72% в DU145 и 56% в PC3. Анализ через 48 часов после индукции выявил 49% жизнеспособность клеток в DU145 и 37% жизнеспособность клеток в PC3. Через 72 часа экспрессия DEFB1 приводила к 44% и 29% относительной жизнеспособности клеток DU145 и PC3, соответственно.
DEFB1 вызывает быстрый опосредованный каспазами апоптоз в клетках рака простаты на поздней стадии: Чтобы определить, является ли влияние DEFB1 на клетки PC3 и DU145 цитостатическим или цитотоксическим, осуществляли анализ FACS. В нормальных условиях роста более чем 90% культур PC3 и DU145 были жизнеспособными и неапоптозными (нижний левый квадрант) и не были окрашены аннексином V или PI (фигура 4). После индукции экспрессии DEFB1 в клетках PC3 количество апоптозных клеток (нижний и верхний правые квадранты) составляло 10% во временной точке 12 часов, 20% в 24 часа и 44% в 48 часов. В случае клеток DU145 количество апоптозных клеток составляло 12% после 12 часовой, 34% после 24 часовой и 59% после 48 часовой индукции. Не наблюдали увеличения апоптоза в клетках, содержащих пустую плазмиду, после индукции с использованием PonA (данные не показаны).
Каспазную активность определяли в анализе с использованием конфокальной лазерной микроскопии (фигура 5). Клетки DU145 и PC3 индуцировали в отношении экспрессии DEFB1 и активность контролировали на основе связывания зеленого флуоресцирующего FAM-VAD-FMK с каспазами в клетках, активно подвергающихся апоптозу. Анализ клеток в условиях ДИК показал присутствие жизнеспособных контрольных клеток DU145 (A), PC3 (E) и LNCaP (I) во временной точке 0 часов. Возбуждение конфокальным лазером при 488 нм не давало регистрируемого зеленого окрашивания, что свидетельствует об отсутствии каспазной активности в DU145 (B), PC3 (F) или LNCaP (J). После индукции в течение 24 часов клетки DU145 (C), PC3 (G) и LNCaP (K) были снова видимы в условиях ДИК. Конфокальный флуоресцентный анализ выявил зеленое окрашивание в клетках DU145 (D) и PC3 (H), что свидетельствует о каспазной активности. Однако зеленого окрашивания не наблюдали в LNCaP (L), что свидетельствует об отсутствии индукции апоптоза под влиянием DEFB1.
В заключение, настоящее исследование показывает функциональную роль DEFB1 при раке простаты. Кроме того, полученные данные показывают, что DEFB1 является частью естественной иммунной системы, вовлеченной в противоопухолевый иммунитет. Данные, представленные в настоящем описании, демонстрируют, что DEFB1, экспрессированный на физиологических уровнях, является цитотоксическим для рефрактерных по отношению к гормону злокачественных клеток простаты AR-, но не для чувствительных к гормону злокачественных клеток простаты AR+ и не для нормальных эпителиальных клеток простаты. Учитывая что DEFB1 конститутивно экспрессируется в нормальных клетках простаты без цитотоксичности, вероятно, что клетки рака простаты на поздней стадии AR- обладают отличающимися фенотипическими признаками, которые делают их чувствительными к цитотоксичности DEFB1. Таким образом, DEFB1 является терапевтическим средством, пригодным для лечения рака простаты на поздней стадии, и возможно также для лечения других злокачественных опухолей.
ПРИМЕР 2: ОПОСРЕДОВАННЫЙ МИ-РНК НОКДАУН ЭКСПРЕССИИ PAX2 ПРИВОДИТ К ГИБЕЛИ ЗЛОКАЧЕСТВЕННЫХ КЛЕТОК ПРОСТАТЫ, НЕЗАВИСИМО ОТ СТАТУСА P53
В данном примере исследовали влияние ингибирования экспрессии PAX2 в результате РНК-интерференции в злокачественных клетках простаты, которые отличаются по статусу гена p53. Результаты показывают, что ингибирование PAX2 приводит к гибели клеток, независимо от статуса p53, что свидетельствует о том, что существуют дополнительные гены супрессоры опухолей или пути гибели клеток, ингибированные под влиянием PAX2 в злокачественной опухоли простаты.
Материалы и способы
Сайленсинг PAX2 под влиянием ми-РНК: Чтобы добиться эффективного сайленсинга гена, синтезировали пул из четырех комплементарных малых интерферирующих рибонуклеотидов (ми-РНК), мишенью которых является мРНК PAX2 человека (номер доступа NM_003989.1) (Dharmacon Research, Lafayette, CO, USA). Второй пул из четырех ми-РНК использовали в качестве внутреннего контроля, чтобы тестировать специфичность ми-РНК PAX2. Мишенью двух синтезированных последовательностей являлась мРНК люциферазы GL2 (номер доступа X65324), и две были неспецифичными по последовательности (таблица 3). Для отжига ми-РНК 35 М отдельных нитей инкубировали в буфере для отжига (100 мМ ацетат калия, 30 мМ HEPES-KOH, pH 7,4, 2 мМ ацетат магния) в течение 1 минуты при 90°C с последующей 1-часовой инкубацией при 37°C.
Последовательности ми-РНК PAX2
Пул из четырех ми-РНК использовали для ингибирования экспрессии белка PAX2.
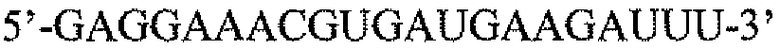


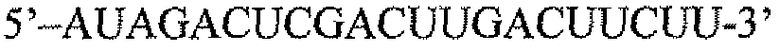
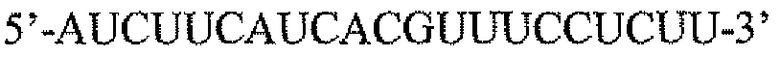
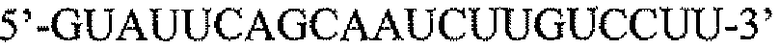

Вестерн-анализ: Коротко, клетки собирали трипсинизацией и дважды промывали в PBS. Лизирующий буфер готовили согласно инструкциям производителя (Sigma) и затем добавляли к клеткам. После 15-минутного периода инкубации при 4°C в орбитальном встряхивателе клеточный лизат собирали и центрифугировали в течение 10 минут при 12000×g, чтобы осадить обломки клеток. Затем надосадок, содержащий белок, собирали и количественно оценивали. Затем 25 мкг белкового экстракта наносили на 8-16% градиентный SDS-ПААГ (Novex). После электрофореза белки переносили на PVDF-мембраны и затем блокировали 5% обезжиренным сухим молоком в TTBS (0,05% твин 20 и 100 мМ трис-Cl) в течение 1 часа. Затем блоты исследовали, используя первое кроличье анти-PAX2-антитело (Zymed, San Francisco, CA) в разведении 1:2000. После промывки мембраны инкубировали с антителом против Ig кролика, конъюгированным с пероксидазой хрена (HRP) (разведение 1:5000; Sigma), и регистрацию сигнала визуализировали, используя хемилюминесцентные реагенты (Pierce), на устройстве Alpha Innotech Fluorchem 8900. В качестве контроля с блотов удаляли антитела и повторно исследовали, используя первое мышиное антитело против β-актина (1:5000; Sigma-Aldrich) и конъюгированное с HRP второе антитело против Ig мыши (1:5000; Sigma-Aldrich), и регистрацию сигнала снова визуализировали.
Фазово-контрастная микроскопия: Влияние нокдауна PAX2 на рост клеток анализировали с использованием фазово-контрастной микроскопии. В данном случае 1-2×104 клеток высевали в 6-луночные чашки для культивирования (BD Falcon, USA). На следующий день клетки обрабатывали только средой, негативной контрольной неспецифичной ми-РНК или ми-РНК PAX2 и инкубировали в течение шести дней. Затем клетки наблюдали под инвертированным микроскопом Zeiss IM 35 (Carl Zeiss, Germany) при 32-кратном увеличении. Фазово-контрастные изображения поля клеток получали, используя камеру SPOT Insight Mosaic 4.2 (Diagnostic Instruments, USA).
MTT-анализ цитотоксичности: клетки DU145, PC3 и LNCaP (1×105) трансфицировали 0,5 мкг пула ми-РНК PAX2 или пула контрольной ми-РНК, используя реагент для трансфекции Codebreaker согласно протоколу производителя (Promega). Затем суспензии клеток разбавляли и высевали в 96-луночный планшет по 1-5×103 клеток на лунку и давали возможность расти в течение 2, 4 или 6 дней. После культивирования определяли жизнеспособность клеток посредством измерения превращения бромида 3-[4,5-диметилтиазол-2-ил]-2,5-дифенилтетразолия, MTT (Promega), в окрашенный продукт формазан. Оптическую плотность регистрировали при 540 нм на сканирующем многолуночном спектрофотометре.
Определение пан-каспазы в линиях злокачественных клеток простаты и количественную ПЦР в реальном времени осуществляли, как описано в примере 1. Пары праймеров для BAX, BID и BAD создавали на основе опубликованных последовательностей (таблица 4). Реакции осуществляли в 96-луночном оптическом планшете для реакции MicroAmp (PE Biosystems). Проводили сорок циклов ПЦР в стандартных условиях, используя температуру отжига 60°C. Количественную оценку осуществляли на основании номера цикла, в котором начиналась экспоненциальная амплификация (пороговое значение), и усредняли, используя значения, полученные в трех повторах. Не было обратной зависимости между уровнем мессенджера и пороговым значением. Кроме того, GAPDH использовали в качестве гена домашнего хозяйства, чтобы нормализовать начальное содержание суммарной кДНК. Экспрессию гена рассчитывали в виде соотношения экспрессии между проапоптозными генами и GAPDH. Все реакции осуществляли в трех повторах.
Праймеры для количественной ОТ-ПЦР. Нуклеотидные последовательности праймеров, используемых для амплификации PAX2 и GAPDH
Ингибирование белка PAX2 с использованием ми-РНК: Чтобы подтвердить, что ми-РНК эффективно целенаправленно воздействует на мРНК PAX2, осуществляли Вестерн-анализ, контролируя уровни экспрессии белка PAX2 на протяжении шестидневного периода обработки. Клетки подвергали одному раунду трансфекции пулом ми-РНК PAX2. Результаты подтвердили специфичное целенаправленное воздействие на мРНК PAX2, так как показали нокдаун белка PAX2 на четвертый день в DU145 (фигура 6a) и на шестой день в PC3 (фигура 6b).
Нокдаун PAX2 ингибирует рост злокачественных клеток простаты: Клетки анализировали после шестидневного периода обработки только средой, негативной контрольной неспецифичной ми-РНК или ми-РНК PAX2 (фигура 7). Все клетки DU145 (a), PC3 (d) и LNCaP (g) достигали, по меньшей мере, 90% слияния в чашках для культивирования, содержащих только среду. Обработка DU145 (b), PC3 (e) и LNCaP (h) негативной контрольной неспецифичной ми-РНК не оказывало влияния на рост клеток, и клетки снова достигали слияния через шесть дней. Однако обработка ми-РНК PAX2 приводила к значимому уменьшению количества клеток. Клетки DU145 достигали примерно 15% слияния (c), а клетки PC3 достигали только 10% слияния (f). Клетки LNCaP достигали 5% слияния после обработки ми-РНК.
Анализы цитотоксичности: Жизнеспособность клеток измеряли после двух-, четырех- и шестидневного периода воздействия и выражали в виде отношения оптической плотности при 570-630 нм для обработанных клеток к оптической плотности необработанных контрольных клеток (фигура 8). Относительная жизнеспособность клеток после 2 дней обработки составляла 77% в LNCaP, 82% в DU145 и 78% в PC3. После четырех дней относительная жизнеспособность клеток составляла 46% в LNCaP, 53% в DU145 и 63% в PC3. После шестидневной обработки относительная жизнеспособность клеток снижалась до 31% в LNCaP, 37% в PCS и составляла 53% в DU145. В качестве негативных контролей жизнеспособность клеток измеряли после шестидневного периода обработки негативной контрольной неспецифичной ми-РНК или только реагентов для трансфекции. В обоих случаях не происходило статистически значимого изменения жизнеспособности клеток по сравнению с нормальной ростовой средой.
Выявление пан-каспазы: Каспазную активность регистрировали в анализе с использованием конфокальной лазерной микроскопии. Клетки DU145, PC3 и LNCaP обрабатывали ми-РНК PAX2 и наблюдение активности осуществляли на основе связывания FAM-меченого пептида с каспазами в клетках, активно подвергающихся апоптозу, которые будут давать зеленую флуоресценцию. Анализ клеток в присутствии только среды при использовании ДИК показывает присутствие жизнеспособных клеток DU145 (A), PC3 (E) и LNCaP (I) в 0 часов (фигура 9). Возбуждение конфокальным лазером при 488 нм не давало регистрируемого зеленого окрашивания, что свидетельствует об отсутствии каспазной активности в необработанных DU145 (B), PC3 (F) или LNCaP (J). После четырех дней обработки ми-РНК PAX2 клетки DU145 (C), PC3 (G) и LNCaP (K) были снова видны при ДИК. В условиях флуоресценции обработанные клетки DU145 (D), PC3 (H) и LNCaP (L) имели зеленую окраску, свидетельствующую о каспазной активности.
Влияние ингибирования PAX2 на проапоптозные факторы: Клетки DU145, PC3 и LNCaP обрабатывали ми-РНК против PAX2 в течение шести суток и измеряли экспрессию проапоптозных генов, зависимых и независимых от регуляции транскрипции p53, чтобы исследовать пути гибели клеток. В случае BAX имело место 1,81-кратное увеличение в LNCaP, 2,73-кратное увеличение в DU145 и 1,87-кратное увеличение в PC3 (фигура 10a). Уровни экспрессии BID возрастали; в 1,38 раза в LNCaP и в 1,77 раза в DU145 (фигура 10b). Однако уровни экспрессии BID снижались в 1,44 раза в PC3 после обработки (фигура 10c). Анализ BAD показал 2,0-кратное увеличение экспрессии в LNCaP, 1,38-кратное увеличение в DU145 и 1,58-кратное увеличение в PC3.
Полученные результаты свидетельствуют о зависимости жизнеспособности злокачественных клеток простаты от экспрессии PAX2. После активации p53 в результате нокдауна PAX2 в p53-экспрессирующей линии клеток LNCaP, мутантной по p53 линии DU145 и p53-нулевой линии PC3, каспазную активность выявляли во всех трех линиях, что свидетельствует об инициации запрограммированной гибели клеток. Экспрессия BAX подвергалась повышающей регуляции во всех трех линиях клеток, независимо от статуса p53. Экспрессия проапоптозного фактора BAD также увеличивалась во всех трех линиях после ингибирования PAX2. После обработки ми-РНК PAX2 экспрессия BID экспрессия возрастала в LNCaP и DU145, но даже снижалась в PC3. Полученные результаты показывают, что гибель клеток, наблюдаемая в злокачественной опухоли простаты, подвержена влиянию, но не зависит от экспрессии p53. Инициация апоптоза в злокачественных клетках простаты разными путями, приводящими к гибели клеток, независимо от статуса p53, свидетельствует, что PAX2 ингибирует другие супрессоры опухолей.
Кроме последовательностей SEQ ID NO: 3-6, другие примеры анти-PAX2-ми-РНК включают без ограничения ми-РНК, имеющие последовательности (в 5'-3'-направлении):
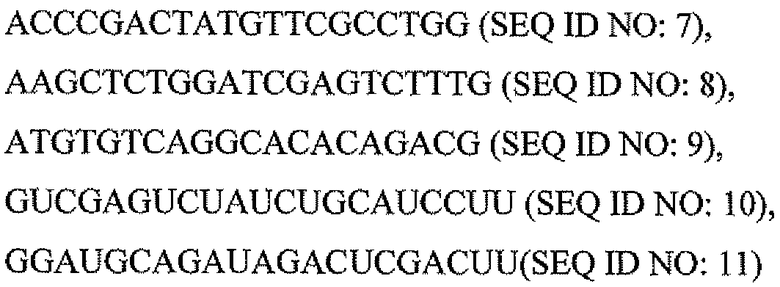 .
.
Белки PAX представляют семейство факторов транскрипции, которые являются консервативными в эволюции и способны связывать специфичные последовательности ДНК на протяжении доменов, называемых «парным доменом (PD)» и «гомеодоменом (HD)». PD представляет собой консенсусную последовательность, общую для некоторых белков PAX (например, PAX2 и PAX6). PD управляет связыванием ДНК аминокислотами, расположенными в α3-спирали, с образованием ДНК-белкового комплекса. В случае PAX2 аминокислоты в HD узнают и специфично взаимодействуют с центральной последовательностью ДНК CCTTG (SEQ ID NO: 1). Предполагается, что олигонуклеотиды, содержащие такую последовательность или ее комплемент, являются ингибиторами белков PAX2.
В одном варианте олигонуклеотид имеет последовательность V-CCTTG-W (SEQ ID NO: 12), где V означает от 1 до 35 следующих друг за другом фланкирующих нуклеотидов DEFB1, и W означает от 1 до 35 нуклеотидов. Нуклеотиды могут представлять собой непрерывно следующие друг за другом нуклеотиды, которые в норме фланкируют сайт связывания PAX2 с ДНК DEFB1. Альтернативно, они могут быть неродственными DEFB1 и выбранными обычным образом так, чтобы избежать взаимных помех с последовательностью распознавания.
Другие примеры олигонуклеотидов, которые ингибируют связывание PAX2 с промотором DEFB1, включают без ограничения олигонуклеотид, имеющий последовательности (в 5'-3'-направлении):

ПРИМЕР 3: ИНГИБИРОВАНИЕ ОНКОГЕНА PAX2 ПРИВОДИТ К DEFB1-ОПОСРЕДОВАННОЙ ГИБЕЛИ ЗЛОКАЧЕСТВЕННЫХ КЛЕТОК ПРОСТАТЫ
Идентификацию специфичных для опухоли молекул, которые служат в качестве мишеней для разработки новых противораковых лекарственных средств, считают главной целью исследований злокачественных опухолей. Пример 1 продемонстрировал высокую частоту утраты экспрессии DEFB1 в злокачественной опухоли простаты, и что индукция экспрессии DEFB1 приводит к быстрому апоптозу на негативной по рецептору андрогена стадии злокачественной опухоли простаты. Полученные данные показывают, что DEFB1 играет роль в подавлении опухоли простаты. Кроме того, учитывая, что DEFB1 является встречающимся в природе компонентом иммунной системы нормального эпителия простаты, полагают, что он является ценным терапевтическим средством с небольшими или без побочных эффектов. Пример 2 показал, что ингибирование экспрессии PAX2 приводит к гибели раковых клеток простаты, независимо от p53. Полученные данные свидетельствуют, что существует дополнительный проапоптозный фактор или супрессор опухоли, который ингибируется PAX2. Кроме того, данные показывают, что онкогенный фактор PAX2, который сверхэкспрессируется в злокачественной опухоли простаты, является репрессором транскрипции DEFB1. Целью настоящего исследования является определение того, является ли утрата экспрессии DEFB1 следствием аномальной экспрессии онкогена PAX2, и приводит ли ингибирование PAX2 к опосредованной DEFB1 гибели клеток.
Материалы и способы
Выделение РНК и количественную ОТ-ПЦР осуществляли, как описано в примере 1. Пару праймеров для DEFB1 создавали на основе опубликованной последовательности DEFB1 (номер доступа U50930). Осуществляли сорок циклов ПЦР в стандартных условиях, используя температуру отжига 56°C. Кроме того, ген GAPDH амплифицировали в качестве гена домашнего хозяйства, чтобы нормализовать начальное содержание суммарной кДНК. Экспрессию DEFB1 вычисляли в виде соотношения экспрессии между DEFB1 и GAPDH и сравнивали в линиях клеток до и после нокдауна экспрессии PAX2 с использованием ми-РНК. Все реакции проводили три раза в трех повторах.
Создание репортерной конструкции DEFB1: Репортерную плазмиду с геном люциферазы pGL3 использовали для мониторинга репортерной активности DEFB1. В данном случае использовали область длиной 160 оснований выше сайта инициации транскрипции DEFB1, которая включала TATA-бокс DEFB1. Область также содержала последовательность GTTCC (SEQ ID NO: 2), которая необходимая для связывания PAX2. Праймеры для ПЦР конструировали так, чтобы они содержали сайты рестрикции KpnI и NheI. ПЦР-продукты с промотора DEFB1 расщепляли ферментами рестрикции KpnI и NheI и лигировали в сходным образом расщепленную ферментами рестрикции плазмиду pGL3 (фигура 2). Конструкциями трансфицировали E. coli и отдельные клоны отбирали и размножали. Выделяли плазмиды и целостность последовательности конструкции DEFB1/pGL3 подтверждали автоматизированным секвенированием.
Анализ репортера люциферазы: В данном случае 1 мкг репортерной конструкции DEFB1 или контрольной плазмиды pGL3 использовали для трансфекции 1×106 клеток DU145. Затем 0,5×103 клеток высевали в каждую лунку 96-луночного планшета и давали возможность расти в течение ночи. Затем добавляли свободную среду, содержащую ми-РНК PAX2, или только среду и клетки инкубировали в течение 48 часов. Люциферазу регистрировали, используя набор BrightGlo согласно протоколу производителя (Promega), и планшеты регистрировали на автоматизированном 96-луночном люминометре Veritas. Активность промотора выражали в виде относительной люминесценции.
Анализ проницаемости мембран: Осуществляли двойное окрашивание акридиновым оранжевым (AO)/бромидом этидия (EtBr), чтобы идентифицировать изменения в целостности клеточной мембраны, а также апоптозные клетки по окрашиванию конденсированного хроматина. AO окрашивает жизнеспособные клетки, а также ранние апоптозные клетки, тогда как EtBr окрашивает апоптозные клетки на поздней стадии, которые утратили целостность мембраны. Коротко, клетки высевали на 2-камерные культуральные стекла (BD Falcon, USA). Клетки, трансфицированные пустой плазмидой pIND/pvgRXR или pIND DEFB1/pvgRXR, индуцировали в течение 24 или 48 часов средой, содержащей 10 мкМ понастерона A. Контрольные клетки получали свежую среду во временных точках 24 и 48 часов. Чтобы определить влияние ингибирования PAX2 на целостность мембран, отдельные культуральные стекла, содержащие DU145, PC3 и LNCaP, обрабатывали ми-РНК PAX2 и инкубировали в течение 4 суток. Затем клетки промывали один раз PBS и красили, используя 2 мл смеси (1:1) AO (Sigma, USA) и раствора EtBr (Promega, USA) (5 мкг/мл), в течение 5 минут. После окрашивания клетки снова промывали PBS. Флуоресценцию наблюдали в лазерном сканирующем конфокальном микроскопе Zeiss LSM 5 Pascal Vario 2 (Carl Zeiss Jena, Germany). Диск для выбора цвета возбуждения содержит блоки фильтров BS505-530 (зеленый) и LP560 (красный), которые обеспечивают разделение испускаемого зеленого света от AO в зеленый канал и красного света от EtBr в красный канал. Установки мощности лазерного излучения и регулировки уровня в каждом отдельном эксперименте были идентичны в случае контрольных клеток и клеток с индуцированным DEFB1. Возбуждение получали от лазера на смешанном газе Kr/Ar с длинами волн 543 нм для AO и 488 нм для EtBr. Стекла анализировали при 40-кратном увеличении и цифровые изображения хранили в виде несжатых файлов TIFF и экспортировали в компьютерную программу Photoshop CS (Adobe Systems, San Jose, CA) для обработки изображения и представления печатных копий.
ChIP-анализ PAX2: Иммунопреципитация хроматина (ChIP) обеспечивает возможность идентификации сайтов связывания для ДНК-связывающих белков на основе занятости промотора in vivo фактором транскрипции и обогащения связанного с фактором транскрипции хроматина иммунопреципитацией. Использовали модификацию протокола, описанного в публикации лаборатории Farnham; также в режиме онлайн на сайте http://mcardle.oncology. wisc.edu/farnham/). Линии клеток DU145 и PC3 сверхэкспрессируют белок PAX2, но не экспрессируют DEFB1. Клетки инкубировали в PBS, содержащем 1,0% формальдегида, в течение 10 минут, чтобы поперечно связать белки с ДНК. Затем образцы обрабатывали ультразвуком, получая ДНК со средней длиной 600 п.н. Обработанный ультразвуком хроматин, предварительно очищенный с использованием шариков Dynabeads с белком A, инкубировали с PAX2-специфичным антителом или в контроле «без антитела» [с совпадающими по изотипу контрольными антителами]. Затем промытые иммунопреципитаты собирали. После удаления поперечных сшивок ДНК анализировали в ПЦР, используя специфичные для промотора праймеры, чтобы определить, представлен ли DEFB1 в PAX2-иммунопреципитированных образцах. Праймеры конструировали так, чтобы амплифицировать область размером 160 п.н. непосредственно выше сайта старта мРНК DEFB1, которая содержала TATA-бокс DEFB1 и функциональный сайт распознавания PAX2 GTTCC (SEQ ID NO: 2). Для таких исследований позитивные контроли включали ПЦР аликвоты исходного хроматина (перед иммунопреципитацией, но поперечные сшивки удалены). Все стадии осуществляли в присутствии ингибиторов протеаз.
Ингибирование PAX2 с использованием ми-РНК увеличивает экспрессию DEFB1: КОТ-ПЦР-анализ экспрессии DEFB1 перед обработкой ми-РНК выявил относительные уровни экспрессии, составляющие 0,00097 в DU145, 0,00001 в PC3 и 0,00004 в LNCaP (фигура 13). После вызванного ми-РНК нокдауна PAX2 относительная экспрессия составляла 0,03294 (338-кратное увеличение) в DU145, 0,00020 (22,2-кратное увеличение) в PC3 и 0,00019 (4,92-кратное увеличение) в LNCaP. В используемой в качестве негативного контроля линии эпителиальных клеток простаты человека (hPrEC), которая является PAX2-нулевой, выявлены уровни экспрессии 0,00687 перед обработкой и 0,00661 после обработки ми-РНК, подтверждая отсутствие статистически значимого изменения экспрессии DEFB1.
DEFB1 вызывает проницаемость клеточной мембраны: Целостность мембран контролировали в конфокальном анализе (фигура 14). В данном случае интактные клетки окрашены в зеленый цвет благодаря AO, который проникает через мембрану. Кроме того, клетки с нарушенными плазматическими мембранами могут окрашиваться красным благодаря EtBr, который не проникает через мембрану. В данном случае неиндуцированные клетки DU145 (A) и PC3 (D) положительно окрашивались AO и испускали зеленый свет, но не окрашивались EtBr. Однако индукция DEFB1 в DU145 (B) и PC3 (E) приводила к накоплению EtBr в цитоплазме через 24 часа, о чем свидетельствует красное окрашивание. Через 48 часов клетки DU145 (C) и PC3 (F) имели конденсированные ядра и выглядели желтыми, что является следствием присутствия и зеленого и красного окрашивания в результате накопления AO и EtBr, соответственно.
Ингибирование PAX2 приводит к проницаемости мембраны: Клетки обрабатывали ми-РНК PAX2 в течение 4 суток и снова контролировали целостность мембран в конфокальном анализе. В данном случае и DU145, и PC3 имели конденсированные ядра и выглядели желтыми. Однако цитоплазма и ядра клеток LNCaP оставались зелеными после обработки ми-РНК. Красное окрашивание на периферии клеток также свидетельствует о сохранении целостности клеточной мембраны. Полученные данные свидетельствуют, что ингибирование PAX2 приводит к специфично опосредованной DEFB1 гибели клеток в DU145 и PC3, но не в клетках LNCaP. Гибель, наблюдаемая в LNCaP, является следствием трансактивации существующего p53 дикого типа в LNCaP после ингибирования PAX2.
Ингибирование PAX2 при использовании ми-РНК усиливает активность промотора DEFB1: Анализ активности промотора DEFB1 в клетках DU145, содержащих конструкцию DEFB1/pGL3, выявил 2,65-кратное увеличение в относительных световых единицах после 48-часовой обработки по сравнению с необработанными клетками. В клетках PC3 происходило 3,78-кратное увеличение в относительных световых единицах по сравнению с необработанными клетками.
PAX2 связывается с промотором DEFB1: Осуществляли ChEP-анализ на клетках DU145 и PC3, чтобы определить, связывается ли репрессор транскрипции PAX2 с промотором DEFB1 (фигура 15). Дорожка 1 содержит маркер молекулярной массы 100 п.н. Дорожка 2 содержит позитивный контроль, представляющий собой область длиной 160 п.н. промотора DEFB1, амплифицированную из DU145 перед поперечным сшиванием и иммунопреципитацией. Дорожка 3 является негативным контролем, представляющим ПЦР, выполненную без ДНК. Дорожки 4 и 5 являются негативными контролями, представляющими ПЦР на основе продуктов иммунопреципитации, осуществляемой с использованием IgG, после поперечного сшивания из DU145 и PC3, соответственно. ПЦР-амплификация 25 пг ДНК (дорожка 6 и 8) и 50 пг ДНК (дорожка 7 и 9), иммунопреципитированной анти-PAX2-антителом после поперечного сшивания, выявила фрагмент промотора длиной 160 п.н. в DU145 и PC3, соответственно.
Полученные результаты показывают, что онкогенный фактор PAX2 подавляет экспрессию DEFB1. Подавление происходит на уровне транскрипции. Кроме того, компьютерный анализ промотора DEFB1 выявил присутствие сайта связывания ДНК GTTCC (SEQ ID NO: 2) для репрессора транскрипции PAX2 рядом с TATA-боксом DEFB1 (фигура 1). Одним из признаков цитотоксичности дефензина является нарушение целостности мембраны. Полученные результаты показывают, что эктопическая экспрессия DEFB1 в злокачественных клетках простаты приводит к утрате мембранного потенциала вследствие нарушенных клеточных мембран. Такое же явление наблюдается после ингибирования экспрессии белка PAX2. Следовательно, подавление экспрессии или функции PAX2 приводит к восстановлению экспрессии DEFB1 и, как следствие, к опосредованной DEFB1 гибели клеток. Также полученные результаты доказывают применимость DEFB1 для направленной терапии при лечении рака простаты и возможно при лечении других злокачественных опухолей посредством естественного иммунитета.
ПРИМЕР 4: ВЛИЯНИЕ ЭКСПРЕССИИ DEFB1 НА ИМПЛАНТИРОВАННЫЕ ОПУХОЛЕВЫЕ КЛЕТКИ
Противоопухолевую способность DEFB1 оценивали посредством инъекции опухолевых клеток, которые сверхэкспрессируют DEFB1, мышам nude. DEFB1 клонировали в векторе pBI-EGFP, который имеет двунаправленный отвечающий на тетрациклин промотор. Линии клеток Tet-Off создавали посредством трансфекции pTet-Off в клетки DU145, PC3 и LNCaP и селекции с использованием G418. Плазмиду pBI-EGFP-DEFB1 котрансфицировали с pTK-Hyg в линии клеток Tet-off и подвергали селекции с использованием гигромицина. Использовали только суспензии одиночных клеток с жизнеспособностью >90%. Каждое животное получало примерно 500000 клеток, вводимых подкожно в правый бок самкам мышей nude. Использовали две группы, контрольную группу, которой инъецировали клоны только с вектором, и группу, которой инъецировали клоны, сверхэкспрессирующие DEFB1. Каждая группа содержала по 35 мышей, как было определено статистиком. Животных взвешивали дважды в неделю, за ростом опухолей наблюдали, используя штангенциркули, и объемы опухолей определяли, используя следующую формулу: объем=0,5×(ширина)2×длина. Всех животных умерщвляли, используя избыточную дозу CO2, когда размер опухоли достигал 2 мм3 или через 6 месяцев после имплантации; опухоли вырезали, взвешивали и хранили в нейтральном забуференном формалине для патологического исследования. Различия в росте опухолей между группами характеризовали, используя сводные статистические данные и графические представления. Статистическую значимость оценивали, используя либо t-критерий, либо непараметрический аналог.
ПРИМЕР 5: ВЛИЯНИЕ МИ-РНК PAX2 НА ИМПЛАНТИРОВАННЫЕ ОПУХОЛЕВЫЕ КЛЕТКИ
Матричные олигонуклеотиды ми-РНК PAX2, имеющих форму шпильки, которые использовали в исследованиях in vitro, применяли для исследования влияния повышающей регуляции экспрессии DEFB1 in vivo. Смысловую и антисмысловую нити (см. таблицу 4) отжигали и клонировали в векторе pSilencer 2.1 U6 hygro, экспрессирующем ми-РНК (Ambion) под контролем промотора РНК-полимеразы III U6 человека. Клонированную плазмиду секвенировали, проверяли и трансфицировали в линии клеток PC3, Du145 и LNCap. Рандомизированную ми-РНК клонировали и использовали в качестве негативного контроля в данном исследовании. Отбирали резистентные к гигромицину колонии, клетки подкожно вводили мышам и рост опухолей наблюдали, как описано выше.
ПРИМЕР 6: ВЛИЯНИЕ НИЗКОМОЛЕКУЛЯРНЫХ ИНГИБИТОРОВ СВЯЗЫВАНИЯ PAX2 НА ИМПЛАНТИРОВАННЫЕ ОПУХОЛЕВЫЕ КЛЕТКИ
Распознаваемая последовательность ДНК для связывания PAX2 находится в промоторе DEFB1 между нуклеотидами -75 и -71 [+1 относится к сайту старта транскрипции]. Предлагаются короткие олигонуклеотиды, комплементарные ДНК-связывающему домену PAX2. Примеры таких олигонуклеотидов включают 20-мерные и 40-мерные олигонуклеотиды, содержащие последовательность распознавания GTTCC (SEQ ID NO: 2), представленные ниже. Указанные длины были выбраны случайно, и предполагается, что олигонуклеотиды другой длины являются эффективными в качестве блокаторов связывания. В качестве негативного контроля конструировали олигонуклеотиды с рандомизированной последовательностью (CTCTG)-(SEQ ID NO: 17), чтобы подтвердить специфичность. Олигонуклеотиды трансфицируют в злокачественные клетки простаты и клетки HPrEC, используя реагент липофектамин или реагент для трансфекции Codebreaker (Promega, Inc). Чтобы подтвердить взаимодействия ДНК-белок двунитевые олигонуклеотиды могут быть помечены [32P]-dCTP и подвергнуты анализам сдвига электрофоретической подвижности. Кроме того, проводят мониторинг экспрессии DEFB1 с использованием КОТ-ПЦР и Вестерн-анализа после обработки олигонуклеотидами. Наконец гибель клеток выявляют, используя MTT-анализ и проточную цитометрию, как описано ранее.
Последовательность распознавания №1: 
последовательность распознавания №2: 
рандомизированная последовательность №1: 
последовательность распознавания №3: 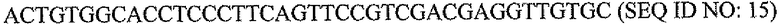
последовательность распознавания №4: 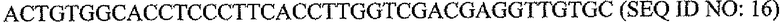
рандомизированная последовательность №2: 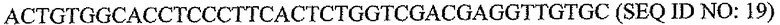
Дополнительные примеры олигонуклеотидов согласно изобретению включают:
последовательность распознавания №1: 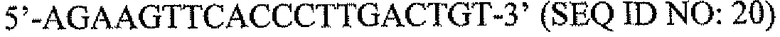
последовательность распознавания №2: 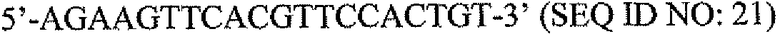
рандомизированная последовательность №1: 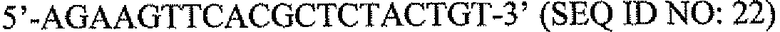
последовательность распознавания №3: 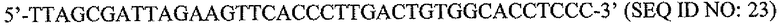
последовательность распознавания №4: 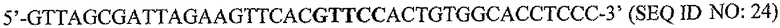
рандомизированная последовательность №2: 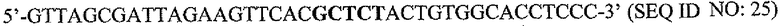
Указанная группа альтернативных ингибирующих олигонуклеотидов представляют собой последовательность распознавания (наряду с центральной последовательностью CCTTG (SEQ ID NO: 1)) для связывающего домена PAX2 и гомеобокса. Олигонуклеотиды содержат реальные последовательности из промотора DEFB1.
Ген PAX2 необходим для роста и жизнеспособности различных злокачественных клеток, включая клетки простаты. Кроме того, ингибирование экспрессии PAX2 приводит к гибели клеток, опосредованной компонентом естественного иммунитета DEFB1. Подавление экспрессии и активности DEFB1 сопровождается связыванием белка PAX2 с сайтом распознавания GTTCC (SEQ ID NO: 2) в промоторе DEFB1. Поэтому указанный путь обеспечивает ценную терапевтическую мишень при лечении рака простаты. В указанном способе последовательности связываются с ДНК-связывающим участком PAX2 и блокируют связывание PAX2 с промотором DEFB1, таким образом обеспечивая экспрессию и активность DEFB1. Олигонуклеотидные последовательности и эксперимент, описанные выше, являются примерами и демонстрируют модель конструирования дополнительных ингибирующих PAX2 лекарственных средств.
В связи с тем, что последовательность GTTCC (SEQ ID NO: 2) существует в генах интерлейкина-3, интерлейкина-4, рецептора инсулина и других, PAX2 также регулирует их экспрессию и активность. Поэтому ингибиторы PAX2, предлагаемые в настоящем изобретении, применимы в случае ряда других заболеваний, включая заболевания, связанные с воспалением, включая простатит и доброкачественную гипертрофию простаты (BPH).
ПРИМЕР 7: УТРАТА ЭКСПРЕССИИ DEFB1 ПРИВОДИТ К УСИЛЕННОМУ ОНКОГЕНЕЗУ
Создание мышей с утраченной функцией: Систему Cre/loxP использовали для выяснения молекулярных механизмов, лежащих в основе канцерогенеза простаты. В данном случае обусловленный Cre нокаут DEFB1 используют для индуцируемого нарушения в простате. Обусловленный Cre нокаут DEFB1 заключается в создании направляющего к мишени вектора, содержащего сайты loxP, фланкирующие экзоны, кодирующие DEFB1, целенаправленное воздействие на ES-клетки таким вектором и создание химерных по зародышевой линии мышей из таких подвергнутых целенаправленному воздействию ES-клеток. Гетерозигот скрещивают с трансгенными мышами по специфичному для простаты Cre и интеркросс гетерозигот используют для создания мышей со специфичным для простаты нокаутом DEFB1. Обнаружено, что четыре генотоксических химических соединения индуцируют карциномы простаты у грызунов: N-метил-N-нитрозомочевина (MNU), N-нитрозобис-2-оксопропиламин (BOP), 3,2'-диметил-4-аминобифенил (MAB) и 2-амино-1-метил-6-фенилимидазо[4,5-b]пиридин (PhIP). Трансгенных по DEFB1 мышей обрабатывают указанными канцерогенными соединениями путем внутрижелудочного введения или внутримышечной инъекции для исследований аденомы простаты и аденокарциномы. Образцы простаты исследуют в отношении различий в росте опухолей и изменений экспрессии генов, используя гистологические, иммуногистологические анализы, анализы мРНК и белка.
Создание мышей GOF: В случае мышей с индуцируемым PAX2 GOF однопометным животным PAX2 GOF (битрансгенные) и дикого типа (монотрансгенные) вводят доксициклин (Dox), начиная с 5-недельного возраста, чтобы индуцировать специфичную для простаты экспрессию PAX2. Коротко, монотрансгенных мышей PROBASIN-rtTA (специфичная для клеток простаты экспрессия tet-зависимого индуктора rtTA) скрещивают с полученными авторами отвечающими трансгенными по PAX2 линиями. Для индукции битрансгенных мышей кормят Dox в питьевой воде (500 мг/л, свежеприготовленный, дважды в неделю). Начальные эксперименты подтверждают низкие исходные уровни, хорошую способность к индукции и специфичную для типа клеток экспрессию PAX2 и репортера EGFP с использованием трансгенной линии-основателя битрансгенных мышей. Что касается размеров экспериментальных групп, то 5-7 совпадающих по возрасту и полу особей в каждой группе (дикого типа и GOF) обеспечивают статистическую значимость. В случае всех животных в данном исследовании ткани простаты сначала собирают с недельными интервалами для анализа и сравнения, чтобы определить временные параметры канцерогенеза.
ПЦР-генотипирование, ОТ-ПЦР и к-ПЦР: Трансгенных мышей PROBASIN-rtTA генотипируют, используя следующие ПЦР-праймеры и условия:
PROBASIN5 (прямой) 5'-ACTGCCCATTGCCCAAACAC-3' (SEQ ID NO: 26);
RTTA3 (обратный) 5'-AAAATCTTGCCAGCTTTCCCC-3' (SEQ ID NO: 27);
денатурация при 95°C в течение 5 минут с последующими 30 циклами при 95°C в течение 30 секунд, 57°C в течение 30 секунд, 72°C в течение 30 секунд, затем удлинение в течение 5 минут при 72°C с получением продукта длиной 600 п.н.
Трансгенных мышей с индуцируемым PAX2 генотипируют, используя следующие ПЦР-праймеры и условия: PAX2 прямой 5'-GTCGGTTACGGAGCGGACCGGAG-3' (SEQ ID NO: 28);
обратный 5'IRES 5'-TAACATATAGACAAACGCACACCG-3' (SEQ ID NO: 29);
денатурация при 95°C в течение 5 минут с последующими 34 циклами при 95°C в течение 30 секунд, 63°C в течение 30 секунд, 72°C в течение 30 секунд, затем удлинение в течение 5 минут при 72°C с получением продукта длиной 460 п.н.
Гемизигот мышей Immortomouse генотипируют, используя следующие ПЦР-праймеры и условия: Immol1, 5'-GCGCTTGTGTCGCCATTGTATTC-3' (SEQ ID NO: 30); Immol2, 5'-GTCACACCACAGAAGTAAGGTTCC-3' (SEQ ID NO: 31);
94°C 30 секунд, 58°C 1 минута, 72°C 1 минута 30 секунд, 30 циклов с получением полосы трансгена ~1 т.н.
Для генотипирования мышей, нокаутированных по PAX2, используют следующие ПЦР-праймеры и условия:
PAX2 прямой 5'-GTCGGTTACGGAGCGGACCGGAG-3' (SEQ ID NO: 32);
PAX2 обратный 5'-CACAGAGCATTGGCGATCTCGATGC-3' (SEQ ID NO: 33);
94°C 1 минута, 65°C 1 минута, 72°C 30 секунд, 36 циклов с получением полосы 280 п.н.
Исследование пептида DEFB1 на животных: Самцам бестимусных мышей (nude) шестинедельного возраста, приобретенным из Charles River Laboratories, подкожно выше лопатки инъецируют 106 жизнеспособных клеток PC3. Через неделю после инъекции животных случайным образом помещают в одну из трех групп - группу I: контрольную; группу II: внутрибрюшинные инъекции DEFB1, 100 мкг/сутки 5 дней в неделю, на протяжении недель 2-14; группу III: внутрибрюшинные инъекции DEFB1, 100 мг/сутки, 5 дней в неделю на протяжении недель 8-14. Животных содержат в стерильных условиях, по четыре животных в клетке, и наблюдение ведут ежедневно. С 10-дневными интервалами измеряют опухоли, используя штангенциркули, и объем опухолей вычисляют, используя уравнение V=(L×W2)/2.
ПРИМЕР 8: ЦЕЛЕНАПРАВЛЕННОЕ ВОЗДЕЙСТВИЕ НА ЭКСПРЕССИЮ PAX2 ДЛЯ ХИМИОПРОФИЛАКТИКИ ВНУТРИЭПИТЕЛИАЛЬНОЙ НЕОПЛАЗИИ И ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ
Химиопрофилактику злокачественной опухоли определяют как профилактику злокачественной опухоли или лечение предракового состояния или даже раньше. Длительный период прогрессирования до инвазивного рака является основной возможностью с научной точки зрения, но и помехой с экономической точки зрения, чтобы показать клиническую пользу выбранных химиопрофилактических лекарственных средств. Поэтому важным компонентом исследований по разработке химиопрофилактических средств в последние годы была идентификация более ранних (чем злокачественная опухоль) конечных точек или биомаркеров, которые позволяют точно прогнозировать клинический результат или эффект, снижающий частоту возникновения злокачественных опухолей. В случае многих злокачественных опухолей IEN является ранним эффектом, например, при раке простаты. В связи с тем, что регуляция пути PAX2/DEFB1 нарушается во время IEN, а возможно даже на более ранней гистопатологической стадии, делает его эффективным прогностическим биомаркером и прекрасной мишенью для химиопрофилактики злокачественной опухоли. Показан ряд соединений, которые подавляют PAX2 и увеличивают экспрессию DEFB1, которые могут быть применимы в качестве химиопрофилактических средств в случае рака простаты.
Как показано в таблице 1, ген PAX2 экспрессируется в нескольких злокачественных опухолях. Кроме того, показано, что несколько злокачественных опухолей имеют аномальную экспрессию PAX2 (фигура 18). Химиопрофилактика посредством целенаправленного воздействия на экспрессию PAX2 может в значительной степени повлиять на связанные со злокачественными опухолями смертельные случаи. Ангиотензин II (AngII) является основным регулятором кровяного давления и гомеостаза сердечнососудистой системы и известен как сильнодействующий митоген. AngII опосредует свои биологические эффекты связыванием с двумя подтипами рецепторов, рецептором ангиотензина типа I (AT1R) и рецептором ангиотензина типа II (AT2R), которые относятся к надсемейству сопряженных с G-белком рецепторов, но имеют разное распределение в тканях и пути внутриклеточной передачи сигналов. Показано, что кроме влияния на кровяное давление AngII играет роль в различных патологических ситуациях, включая ремоделирование ткани, такое как заживление ран, гипертрофию сердца и развитие. Действительно, недавние исследовании выявили локальную экспрессию некоторых компонентов системы ренин-ангиотензин (RAS) в различных злокачественных клетках и тканях, включая простату. Повышающая регуляция AT1R дает значительное преимущество злокачественным клеткам, которые приобретают способность ускользать от апоптоза и регулирующих рост элементов.
Материалы и способы
Реагенты и обработки: Клетки обрабатывали 5 или 10 мкМ AngII, 5 мкМ антагониста ATR1 Los, 5 мкМ антагониста ATR2 PD123319, 25 мкМ ингибитора MEK U0126, 20 мкМ ингибитора MEK/ERK PD98059 или 250 мкМ индуктора AMP-киназы AICAR.
Вестерн-анализ осуществляли, как описано в примере 2. Затем блоты исследовали, используя первое антитело (анти-PAX2, -фосфо-PAX2, -JNK, -фосфо-JNK, -ERK1/2 или -фосфо-ERK1/2) (Zymed, San Francisco, CA) в разведениях 1:1000-2000. После промывки мембраны инкубировали с антителом против Ig кролика, конъюгированным с пероксидазой хрена (HRP) (разведение 1:5000; Sigma), и регистрацию сигнала визуализировали, используя хемилюминесцентные реагенты (Pierce), на Alpha Innotech Fiuorchem 8900. В качестве контроля с блотов удаляли антитела и повторно исследовали, используя в качестве первого мышиное антитело против β-актина (1:5000; Sigma-Aidrich) и конъюгированное с HRP второе антитело против Ig мыши (1:5000; Sigma-Aldrich), и регистрацию сигнала снова визуализировали.
КОТ-ПЦР-анализ осуществляли, как описано в примере 1. Реакции осуществляли в 96-луночном оптическом планшете для реакции MicroAmp (PE Biosystems). Проводили сорок циклов ПЦР в стандартных условиях, используя температуру отжига 60°C. Количественную оценку осуществляли на основании номера цикла, в котором начиналась экспоненциальная амплификация (пороговое значение) и усредняли, используя значения, полученные в трех повторах. Имела место обратная зависимость между уровнем мессенджера и пороговым значением. Кроме того, GAPDH использовали в качестве гена домашнего хозяйства, чтобы нормализовать начальное содержание суммарной кДНК. Относительную экспрессию вычисляли в виде соотношения экспрессии между каждым геном и GAPDH. Все реакции осуществляли в трех повторах.
Включение тимидина: Пролиферацию клеток определяли по включению [3H]-тимидин-риботида ([3H]TdR) в ДНК. По 0,5×106 клеток/лунку суспензии клеток DU145 высевали в соответствующую среду. Клетки инкубировали в течение 72 часов в присутствии или в отсутствие AngII в указанных концентрациях. Клетки подвергали воздействию 37 кБк/мл [метил-3H]тимидина в той же среде в течение 6 часов. Прикрепленные клетки фиксировали 5% трихлоруксусной кислотой и лизировали в лизирующем буфере SDS/NaOH в течение ночи. Радиоактивность измеряли в жидкостно-сцинтилляционном счетчике Beckman LS3801 (Canada). Суспензионную культуру клеток собирали, используя сборщик клеток (Packard instrument Co., Meriden, CT), и радиоактивность измеряли в жидкостно-сцинтилляционном счетчике 1450 microbeta (PerkinElmer Life Sciences).
Чтобы исследовать влияние AngII на экспрессию PAX2 в злокачественных клетках простаты DU145, экспрессию PAX2 исследовали после обработки AngII в течение периода времени от 30 минут до 48 часов. Как показано на фигуре 19, экспрессия PAX2 постепенно возрастала в течение времени после обработки AngII. Блокирование передачи сигнала RAS при обработке DU145 с использованием Los значимо снижало экспрессию PAX2 (фигура 20A). В данном случае экспрессии PAX2 составляла 37% после 48 часов и 50% после 72 часов обработки Los по сравнению с необработанными контрольными клетками DU145 (фигура 21). Известно, что рецептор AT2R противодействует AT1R. Поэтому исследовали влияние блокирования рецептора AT2R на экспрессию PAX2. Обработка DU145 блокатором AT2R PD123319 приводила к 7-кратному увеличению экспрессии PAX2 после 48 часов и к 8-кратному увеличению после 96 часов обработки (фигура 20B). Полученные данные в совокупности демонстрируют, что экспрессия PAX2 регулируется рецептором ATR1.
Известно, что AngII непосредственно влияет на пролиферацию злокачественных клеток простаты в результате AT1R-опосредованной активации фосфорилирования MAPK и STAT3. Обработка DU145 с использованием AngII приводила к двух-трехкратному увеличению скорости пролиферации (фигура 21). Однако обработка Los снижала скорости пролиферации на 50%. Кроме того, блокирование рецептора AT1R предварительной обработкой Los в течение 30 минут подавляла влияние AngII на пролиферацию.
Чтобы дополнительно исследовать роль передачи сигнала AT1R в регуляции экспрессии и активации PAX2, исследовали влияние блокирования различных компонентов пути передачи сигнала MAP-киназ на экспрессию PAX2. В данном случае обработка клеток DU145 ингибитором MEK U0126 приводила к значимому снижению экспрессии PAX2 (фигура 22). Кроме того, обработка ингибитором MEK/ERK PD98059 также приводила к снижению уровня PAX2. Обработка клеток DU145 с использованием Los не оказывала влияния на уровни белка ERK, но снижала количество фосфо-ERK (фигура 23A). Однако обработка DU145 с использованием Los приводила к значимому снижению экспрессии PAX2. Сходные результаты наблюдали в случае U0126 и PD98059. Также известно, что экспрессия PAX2 регулируется STAT3, который является расположенной ниже мишенью ERK. Обработка DU145 с использованием Los, U0126 и PD98059 снижала уровни белка фосфо-STAT3 (фигура 23C). Полученные результаты демонстрируют, что PAX2 регулируется посредством AT1R в злокачественных клетках простаты.
Кроме того, исследовали влияние передачи сигнала AT1R на активацию PAX2 под действием INK. Обработка DU145 с использованием Los, U0126 и PD98059 приводила к значимому снижению или подавлению уровней белка фосфо-PAX2 (фигура 24A). Однако Los и U0126 не снижали уровни белка фосфо-JNK (фигура 24B). Следовательно, снижение уровня фосфо-PAX2, по-видимому, является следствием пониженных уровней PAX2, а не сниженного фосфорилирования.
5-Аминоимидазол-4-карбоксамид-1-β-4-рибофуранозид (AICAR) широко используют в качестве активатора AMP-киназы, который регулирует гомеостаз энергии и ответ на метаболический стресс. Недавние сообщения показали антипролиферативное и проапоптозное действие активированной AMPK при использовании фармакологических средств или сверхэкспрессии AMPK. Показано, что активация AMPK индуцирует апоптоз в злокачественных клетках желудка человека, клетках рака легкого, в злокачественной опухоли простаты, клетках поджелудочной железы и клетках карциномы печени и усиливает индуцированный окислительным стрессом апоптоз в клетках нейробластомы мыши посредством различных механизмов, которые включают ингибирование пути синтазы жирных кислот и индукцию стресс-киназ и каспазы 3. Кроме того, обработка злокачественных клеток простаты PC3 повышала экспрессию белков p21, p27 и p53 и ингибирование пути PI3K-Akt. Все указанные пути прямо или опосредованно регулируются PAX2. Обработка злокачественных клеток простаты с использованием AICAR приводила к подавлению экспрессии PAX2 (фигура 23B), а также его активированной формы фосфор-PAX2 (фигура 24A). Кроме того, фосфо-STAT3, который регулировал экспрессию PAX2, также был подавлен (фигура 23C).
Наконец, была выдвинута гипотеза о том, что аномальная передача сигнала RAS, которая приводит к повышающей регуляции и сверхэкспрессии PAX2, подавляет экспрессию гена супрессора опухолей DEFB1. Чтобы проверить указанную гипотезу первичную культуру нормальных эпителиальных клеток простаты hPrEC обрабатывали AngII и исследовали уровни экспрессии PAX2 и DEFB1. Обнаружена обратная зависимость между экспрессией DEFB1 и PAX2 в нормальных клетках простаты по сравнению со злокачественными клетками простаты. В необработанных hPrEC обнаружена 10% относительная экспрессия PAX2 по сравнению с экспрессией в злокачественных клетках простаты PC3. Наоборот, в необработанных PC3 наблюдали только 2% относительной экспрессии DEFB1 по сравнению с экспрессией в hPrEC. После 72 часов обработки 10 мкМ AngII происходило снижение на 35% экспрессии DEFB1 по сравнению с необработанными hPrEC, и к 96 часам происходило снижение на 50% экспрессии DEFB1 по сравнению с необработанными клетками hPrEC. Однако происходило увеличение на 66% экспрессии PAX2 через 72 часа и к 96 часам происходило увеличение на 79% экспрессии PAX2 по сравнению с необработанными клетками hPrEC. Кроме того, увеличение экспрессии PAX2 в hPrEC после 72 часов достигало 77% от уровня PAX2, наблюдаемого в злокачественных клетках простаты PC3. После 96-часовой обработки AngII экспрессия PAX2 достигала 89% экспрессии PAX2 в PC3. Полученные результаты свидетельствуют, что нарушенная регуляция передачи сигнала RAS подавляет экспрессию DEFB1 в результате повышающей регуляции экспрессии PAX2 в клетках простаты.
Ингибирование апоптоза является важным патофизиологическим фактором, который вносит вклад в развитие злокачественной опухоли. Несмотря на значительный прогресс в разработке противоопухолевых терапевтических средств, мало успехов достигнуто в лечении заболевания на поздних стадиях. В связи с тем, что канцерогенез является многолетним, многостадийным, прогрессирующим различными путями заболеванием, химиопрофилактика с применением лекарственного средства или других средств для ингибирования, замедления или обращения вспять данного процесса была признана очень многообещающей областью исследования злокачественных опухолей. Успешное лечение лекарственными средствами в случае химиопрофилактики рака простаты требует применения терапевтических средств, специфично воздействующих на клетки-мишени, при поддержании минимального клинического влияния на хозяина с общей целью подавления развития злокачественной опухоли. Таким образом, понимание механизмов ранних стадий канцерогенеза важно для определения эффективности конкретного лечения. Значимость аномальной экспрессии PAX2 и его способность отменять апоптоз с последующим участием в образовании опухоли свидетельствует, что он может быть подходящей мишенью при лечении рака простаты. PAX2 регулируется под действием AT1R в злокачественной опухоли простаты (фигура 26). В данном случае нарушенная регуляция передачи сигнала RAS приводила к повышенной экспрессии онкогена PAX2 и снижению экспрессии супрессора опухолей DEFB1. Следовательно, применение антагонистов AT1R снижает экспрессию PAX2 и приводит к повышенной гибели злокачественных клеток простаты посредством восстановления экспрессии DEFB1 (фигура 27). Полученные результаты дают новые данные о том, что целенаправленное воздействие на экспрессию PAX2 через путь передачи сигнала ренин-ангиотензин, путь AMP-киназ или другими способами, которые заключаются в инактивации белка PAX2 (например, вакцинацией анти-PAX2-антителом), может быть ценной мишенью для профилактики злокачественной опухоли (таблица 5).
Соединения, применимые для ингибирования экспрессии PAX2 в случае химиопрофилактики
Настоящее исследование показывает, что повышающая регуляция онкогена PAX2 в злокачественной опухоли простаты является следствием нарушенной регуляции передачи сигнала RAS. Экспрессия PAX2 регулируется путем передачи сигнала ERK 1/2, который опосредован рецептором ангиотензина типа I. Кроме того, блокирование AT1R лозартаном (Los) подавляет экспрессию PAX2. Кроме того, также показано, что AICAR, который является активатором AMPK, является многообещающим потенциальным ингибитором PAX2. Описанные исследования в сочетании четко свидетельствуют, что такие классы лекарственных средств являются потенциальными супрессорами экспрессии PAX2 и в конечном счете служат в качестве новых химиопрофилактических средств.
ПРИМЕР 9: УРОВЕНЬ ЭКСПРЕССИИ PAX2-DEFB1 В КАЧЕСТВЕ СРЕДСТВА КЛАССИФИКАЦИИ ТКАНИ ПРОСТАТЫ И ПРОГНОСТИЧЕСКОГО ФАКТОРА РАЗВИТИЯ РАКА ПРОСТАТЫ
Материалы и способы
КОТ-ПЦР-анализ: Срезы простаты получали от пациентов, которые были подвергнуты радикальной простатэктомии. После медико-патологического исследования осуществляли лазерную захватывающую микродиссекцию, чтобы выделить области нормальной ткани, пролиферативной внутриэпителиальной неоплазии (PIN) и злокачественной ткани. КОТ-ПЦР осуществляли, как описано ранее, чтобы оценить экспрессию. Экспрессию DEFB1 и PAX2 в каждой области и GAPDH использовали в качестве внутреннего контроля.
Сбор крови и выделение РНК: Для КОТ-ПЦР кровь (2,5 мл) от каждого пациента собирали в пробирки для получения РНК из крови PAXgeneTM (QIAGEN), следуя протоколу производителя. Цельную кровь тщательно смешивали с реагентом для стабилизации гена PAX и хранили при комнатной температуре в течение 6 часов перед экстракцией РНК. Затем суммарную РНК экстрагировали, используя набор для выделения РНК из крови PAXgeneTM согласно инструкциям производителя (QIAGEN). Чтобы удалить примеси геномной ДНК, образцы суммарной РНК, абсорбированные на центрифужной колонке системы выделения РНК из крови PAXgeneTM, инкубировали с ДНКазой I (QIAGEN) при 25°C в течение 20 минут, чтобы удалить геномную ДНК. Суммарную РНК элюировали, количественно оценивали и осуществляли КОТ-ПЦР, как указано ранее, чтобы сравнить соотношения экспрессии PAX2 и DEFB1.
КОТ-ПЦР-анализ полученной с использованием LCM нормальной ткани показал, что пациенты с относительными уровнями экспрессии DEFB1 выше 0,005 имеют более низкую оценку в баллах по шкале Глисона по сравнению с пациентами с уровнями экспрессии ниже, чем 0,005 (фигура 28A). Таким образом, существует обратная зависимость между экспрессией DEFB1 и оценкой по шкале Глисона. Наоборот, имеется положительная корреляция между экспрессией PAX2 и оценкой по шкале Глисона в злокачественной ткани простаты и PIN (фигура 28B).
Вычисляли и сравнивали уровни экспрессии PAX2 и DEFB1 в нормальных, PIN и злокачественных тканях от отдельных пациентов (фигура 29). В общем, уровни экспрессии PAX2 относительно внутреннего контроля GAPDH были в диапазоне от 0 до 0,2 в нормальной (доброкачественной) ткани, от 0,2 до 0,3 в PIN и от 0,3 до 0,5 в раковой (злокачественной) ткани (фигура 30). В случае DEFB1 имела место обратная зависимость при сравнении с PAX2. В данном случае уровни экспрессии DEFB1 по сравнению с внутренним контролем GAPDH были в диапазоне от 0,06 до 0,005 в нормальной (доброкачественной) ткани, от 0,005 до 0,003 в PIN и от 0,003 до 0,001 в раковой (злокачественной) ткани. Таким образом, описана прогностическая шкала (называемая «прогностическим фактором Дональда» или «DPF»), в которой использовано соотношение экспрессий PAX2-DEFB1 в качестве прогностического фактора доброкачественной, предзлокачественной (PIN) и злокачественной ткани простаты. Ткани с соотношением PAX2-DEFB1 от 0 до 39 на основе DPF будут нормальными (патологически доброкачественными). Ткани с соотношением PAX2-DEFB1 от 40 до 99 будут PIN (предзлокачественными) по шкале DPF. Наконец, ткани с соотношением PAX2-DEFB1 от 100 до 500 будут злокачественными (от низкой до высокой степени злокачественности).
В настоящее время существует острая потребность в прогностических биомаркерах развития рака простаты. Известно, что возникновение рака простаты происходит задолго до того, когда заболевание регистрируется современными способами скрининга, такими как тест PSA или ректальное пальпирование. Есть основания полагать, что надежный тест, который позволит проводить мониторинг прогрессирования и раннего проявления рака простаты, может значительно снизить частоту смертности благодаря более эффективному лечению заболевания. В настоящей публикации описан прогностический показатель, позволяющий лечащим врачам заблаговременно распознавать патологическое состояние простаты. DPF измеряет снижение соотношения экспрессии PAX2-DEFB1, ассоциированное с прогрессированием заболевания простаты. Такой эффективный критерий может не только предсказать вероятность развития рака простаты у пациента, то также может точно определить раннее появление предзлокачесвтенной стадии рака. И наконец, такое средство может позволить лечащим врачам отличить, какие пациенты имеют более агрессивное заболевание, а какие его не имеют.
Идентификация специфичных для злокачественной опухоли маркеров применялась для того, чтобы помочь идентифицировать циркулирующие опухолевые клетки (CTC). Также существуют новые доказательства, которые демонстрируют, что выявление опухолевых клеток, диссеминированных в периферической крови, может обеспечивать клинически важными данными для стадирования опухолей, прогнозирования и идентификации суррогатных маркеров для ранней оценки эффективности вспомогательной терапии. Кроме того, при сравнении профилей экспрессии генов во всех циркулирующих клетках можно исследовать экспрессию генов DEFB1 и PAX2, которые играют роль в «иммунологическом надзоре» и «жизнеспособности злокачественной опухоли», соответственно, в качестве прогностического показателя для раннего выявления рака простаты.
ПРИМЕР 10: ФУНКЦИОНАЛЬНЫЙ АНАЛИЗ ЗАЩИТНОГО ПЕПТИДА ХОЗЯИНА БЕТА-ДЕФЕНЗИНА-1 ЧЕЛОВЕКА: НОВЫЙ ВЗГЛЯД НА ЕГО ПОТЕНЦИАЛЬНУЮ РОЛЬ В ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ
Материалы и способы
Образцы ткани и лазерная захватывающая микродиссекция: Ткани простаты получали от пациентов, которые дали информированное согласие перед тем, как были подвергнуты радикальной простатэктомии. Образцы приобретали из банка опухолей онкологического центра Hollings в соответствии с протоколом, одобренным экспертным советом организации. Протокол включал руководство по обработке, изготовлению срезов, гистологической характеристике, очистке РНК и ПЦР-амплификации образцов. Образцы простаты, полученные от хирургов и патологов, сразу замораживали в соединении OCT. Каждый блок OCT разрезали, получая серийные срезы, которые красили и исследовали. Области, содержащие доброкачественные клетки, внутриэпителиальную неоплазию простаты (PIN) и злокачественные клетки, идентифицировали и использовали для ориентира при отборе областей на неокрашенных стеклах с использованием системы Arcturus PixCell II (Sunnyvale, CA). Наконечники, содержащие захваченный материал, подвергали воздействию 20 мкл лизата из набора для выделения чистой РНК Arcturus Pico и сразу обрабатывали. Количество и качество РНК оценивали, используя наборы праймеров, которые продуцируют 5'-ампликоны. Наборы включали праймеры для рибосомного белка L32 (3'-ампликон и 5'-ампликон - 298 оснований отдельно), для глюкозофосфатизомеразы (391 основание отдельно) и для глюкозофосфатизомеразы (842 основания отдельно). Обычно получали соотношения от 0,95 до 0,80 для таких наборов праймеров при использовании образцов из множества приготовленных тканей. Дополнительные опухолевые и нормальные образцы были подвергнуты макродиссекции патологами, быстро заморожены в жидком азоте и подвергнуты оценке в отношении экспрессии hBD-1 и cMYC.
Клонирование гена hBD-1: кДНК hBD-1 получали на основе РНК в результате обратной транскрипции-ПЦР с использованием праймеров, созданных на основе опубликованной последовательности hBD-1 (номер доступа U50930) (Ganz, 2004). ПЦР-праймеры конструировали так, чтобы они содержали сайты рестрикции ClaI и KpnI. ПЦР-продукты hBD-1 расщепляли ферментами рестрикции ClaI и KpnI и лигировали в клонирующий вектор TA. Вектором TA/hBD-1 затем трансфицировали E. coli, штамм XL-1 Blue, используя тепловой шок, и отдельные клоны собирали и размножали. Плазмиды выделяли, используя набор для выделения ДНК из культуры клеток Midiprep (Qiagen, Valencia, CA), и целостность последовательности подтверждали автоматизированным секвенированием. Затем фрагмент гена hBD-1 лигировали в pTRE2, расщепленную ClaI и KpnI, которая служила в качестве промежуточного вектора в целях ориентирования. Затем конструкцию pTRE2/hBD-1 расщепляли ApaI и KpnI, чтобы вырезать вставку hBD-1. Вставку лигировали в вектор pIND системы индуцируемой экспрессии экдизона (Invitrogen, Carlsbad, CA), также дважды расщепленный ApaI и KpnI. Конструкцией снова трансфицировали E. coli и отбирали и размножали отдельные клоны. Плазмиды выделяли и целостность последовательности pIND/hBD-1 снова подтверждали автоматизированным секвенированием.
Трансфекция: Клетки (1×106) высевали в 100-мм чашки Петри и выращивали в течение ночи. Затем клетки котрансфицировали, используя липофектамин 2000 (Invitrogen, Carlsbad, CA), 1 мкг плазмиды pVgRXR, которая экспрессирует гетеродимерный рецептор экдизона, и 1 мкг векторной конструкции pIND/hBD-1 или контрольного вектора pIND/β-галактозидаза (β-gal) в среде Opti-MEM (Life Technologies, Inc.). Эффективность трансфекции определяли, индуцируя экспрессию β-gal понастероном A (PonA) и окрашивая клетки с использованием набора для выявления β-галактозидазы (Invitrogen). Оценка эффективности трансфекции с помощью подсчета позитивно окрашенных колоний (синих) показала, что 60-85% клеток в клеточных линиях экспрессировали β-галактозидазу.
Иммуноцитохимия: Чтобы подтвердить экспрессию белка hBD-1, клетки DU145 и hPrEC высевали на 2-камерные культуральные стекла (BD Falcon, USA) при плотности 1,5-2×104 клеток на камеру. Клетки DU145, трансфицированные только pvgRXR отдельно (контроль) или плазмидой hBD-1, индуцировали в течение 18 часов средой, содержащей 10 мкМ Pon A, тогда как нетрансфицированные клетки получали свежую ростовую среду. После индукции клетки промывали 1×PBS и фиксировали в течение 1 часа при комнатной температуре 4% параформальдегидом. Затем клетки промывали шесть раз 1×PBS и блокировали в 1×PBS с добавлением 2% БСА, 0,8% нормальной сыворотки козы (Vector Laboratories, Inc., Burlingame, CA) и 0,4% тритона-X 100 в течение 1 часа при комнатной температуре. Затем клетки инкубировали в течение ночи с первым кроличьим поликлональным антителом против BD-1 человека (PeproTech Inc, Rocky Hill, NJ), разбавленным 1:1000 в блокирующем растворе. Затем клетки промывали шесть раз блокирующим раствором и инкубировали в течение 1 часа при комнатной температуре со вторым антителом козы против IgG (H+L) кролика Alexa Fluor 488 в разведении 1:1000 в блокирующем растворе. После промывки клеток блокирующим раствором шесть раз, помещали покровные стекла, используя Gel Mount (Biomeda, Foster City, CA). Наконец клетки рассматривали при дифференциально-интерференционном контрасте (ДИК) и возбуждении лазером при 488 нм. Сигнал флуоресценции анализировали, используя конфокальный микроскоп (Zeiss LSM 5 Pascal) и 63× иммерсионный ДИК-объектив с лазерным сканирующим модулем Vario 2 RGB. Цифровые изображения экспортировали в компьютерную программу Photoshop CS (Adobe Systems) для обработки изображения и представления печатных копий.
Выделение РНК и количественную ОТ-ПЦР осуществляли, как описано в примере 1. Пары праймеров для hBD-1 и c-MYC создавали на основе опубликованных последовательностей (таблица 6). Осуществляли сорок циклов ПЦР в стандартных условиях, используя температуру отжига 56,4°C для hBD-1 и c-MYC и 55°C для PAX2. Кроме того, ген β-актина (таблица 6) амплифицировали в качестве гена домашнего хозяйства, чтобы нормализовать начальное содержание суммарной кДНК. Экспрессию генов в образцах доброкачественной ткани простаты вычисляли в виде соотношения экспрессии по сравнению с β-актином. Уровни экспрессии hBD-1 в злокачественной ткани простаты, в первичной культуре клеток простаты hPREC и линиях злокачественных клеток простаты до и после индукции вычисляли относительно среднего уровня экспрессии hBD-1 в клетках hPrEC. В качестве негативного контроля также осуществляли реакции КОТ-ПЦР без матрицы кДНК. Все реакции проводили минимум три раза.
MTT-анализ жизнеспособности клеток осуществляли, как описано в примере 1.
Анализ целостности мембран осуществляли, как описано в примере 3. Клетки, трансфицированные пустой плазмидой или hBD-1-плазмидой, индуцировали в течение 24 или 48 часов средой, содержащей 10 мкМ Pon A, тогда как контрольные клетки получали свежую ростовую среду в каждой временной точке.
Последовательности праймеров для КОТ-ПЦР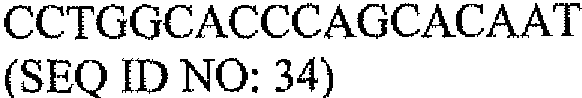
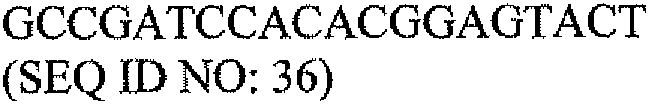
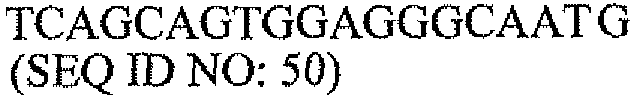
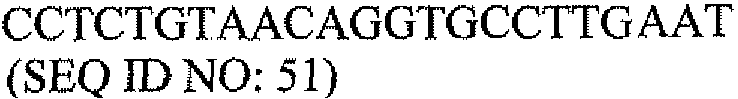
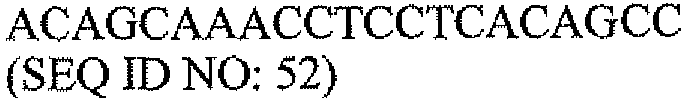
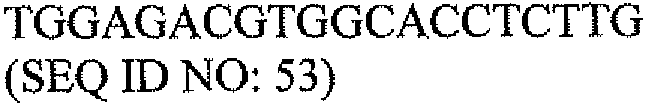
Проточную цитометрию и выявление каспаз осуществляли, как описано в примере 1.
Сайленсинг PAX2 с использованием ми-РНК осуществляли, как описано в примере 2. Молекула ми-РНК покрывали реагентом для трансфекции CodeBreaker (Promega, Inc.) согласно инструкциям производителя перед обработкой.
Статистический анализ: Статистический анализ осуществляли, используя t-критерий Стьюдента для непарных выборок. Значения P определяли двухсторонними вычислениями, и P-значение меньше 0,05 считали статистически значимым. Статистические различия указаны звездочками.
Экспрессия hBD-1 в ткани простаты: в 82% проанализированных срезов замороженных тканей злокачественных опухолей простаты не обнаружено или обнаружена небольшая экспрессия hBD-1 (Donald с соавторами, 2003). Чтобы сравнить уровни экспрессии hBD-1, осуществляли анализ КОТ-ПЦР на нормальной ткани простаты, полученной с использованием макродиссекции или LCM нормальной ткани простаты вблизи злокачественных областей, которые были выбраны случайно. В данном случае hBD-1 выявляли во всех полученных макродиссекцией нормальных клинических образцах с диапазоном экспрессии, которые охватывает примерно 6,6-кратное различие в уровнях экспрессии (фиг.31A). Полученные с использованием LCM нормальные образцы ткани экспрессировали hBD-1 на уровнях в диапазоне, которые охватывает 32-кратное различие в экспрессии (фиг.31B). Совмещение номеров образцов с соответствующими профилями для пациентов показало, что в большинстве случаев уровень экспрессии hBD-1 был выше в образцах от пациентов с оценкой по шкале Глисона 6, чем в образцах от пациентов с оценкой по шкале Глисона 7. Кроме того, сравнение уровней экспрессии hBD-1 в ткани, полученной макродиссекцией и LCM от одного и того же пациента, №1343, показало 854-кратное различие между двумя способами выделения. Таким образом, полученные результаты свидетельствуют, что LCM является более чувствительной методикой для оценки экспрессии hBD-1 в ткани простаты.
Экспрессия hBD-1 в линии клеток простаты: Чтобы подтвердить повышающую регуляцию hBD-1 в линиях злокачественных клеток простаты после трансфекции системой экспрессии hBD-1, осуществляли КОТ-ПЦР. Кроме того, также получали негативные контроли без матрицы и продукты амплификации подтверждали гель-электрофорезом. В данном случае, экспрессия hBD-1 была значительно снижена в линиях злокачественных клеток простаты по сравнению с клетками hPrEC. После 24-часового периода индукции относительные уровни экспрессии hBD-1 значимо возрастали в DU145, PC3 и LNCaP по сравнению с линиями клеток до индукции hBD-1 (фиг.32A).
Затем экспрессию белка hBD-1 подтверждали в клетках DU145, трансфицированных системой экспрессии hBD-1, после индукции Pon A с использованием иммуноцитохимиии. В качестве позитивного контроля также исследовали клетки эпителия простаты hPrEC, экспрессирующие hBD-1. Клетки красили первым антителом против hBD-1 и наблюдали за экспрессией белка на основе зеленой флуоресценции второго антитела (фиг.32B). Анализ клеток при ДИК подтвердил наличие клеток hPrEC и клеток DU145, в которых индуцирована экспрессия hBD-1, во временной точке 18 часов. Возбуждение конфокальным лазером при 488 нм вызывало регистрируемую зеленую флуоресценцию, что свидетельствовало о присутствии белка hBD-1 в hPrEC в качестве позитивного контроля. Однако не было выявлено зеленой флуоресценции в контрольных клетках DU145, и в индуцированных пустой плазмидой клетках DU145, что свидетельствует об отсутствии экспрессии hBD-1. Конфокальный анализ клеток DU145, индуцированных в отношении экспрессии hBD-1, выявил зеленую флуоресценцию, что свидетельствует о присутствии белка hBD-1 после индукции Pon A.
Экспрессия hBD-1 приводит к пониженной жизнеспособности клеток: Осуществляли MTT-анализ, чтобы оценить влияние экспрессии hBD-1 на относительную жизнеспособность клеток в линиях злокачественных клеток простаты DU145, PC3, PC3/AR+ и LNCaP. MTT-анализ в случае использования пустого вектора не выявил статистически значимого изменения жизнеспособности клеток. Через двадцать четыре часа после индукции hBD-1 относительная жизнеспособность клеток составляла 72% в DU145 и 56% в клетках PC3, и через 48 часов жизнеспособность клеток снижалась до 49% в клетках DU145 и до 37% в клетках PC3 (фиг.33A). После 72-часовой индукции hBD-1 относительная жизнеспособность клеток снижалась до 44% в клетках DU145 и до 29% в клетках PC3. Напротив, не было значимого влияния на жизнеспособность клеток LNCaP. Чтобы оценить, являлась ли резистентность к цитотоксичности hBD-1, наблюдаемая в LNCaP, следствием присутствия рецептора андрогена (AR), исследовали цитотоксичность hBD-1 в клетках PC3 с эктопической экспрессией AR (PC3/AR+). В данном случае не было различий между клетками PC3/AR+ и PC3. Следовательно, данные показывают, что hBD-1 является цитотоксическим специфично для клеток злокачественной опухоли простаты на поздней стадии.
Чтобы определить, были ли эффекты hBD-1 на PC3 и DU145 цитостатическими или цитотоксическими, осуществляли FACS-анализ, чтобы измерить гибель клеток. В нормальных условиях роста более чем 90% культур PC3 и DU145 были жизнеспособными и неапоптозными (нижний левый квадрант) и не были окрашены аннексином V или PI (фигура 4). После индукции экспрессии hBD-1 в клетках PC3 количество клеток, подвергающихся раннему апоптозу и позднему апоптозу/некрозу (нижний и верхний правые квадранты, соответственно), составляло 10% во временной точке 12 часов, 20% в 24 часа и 44% в 48 часов. В случае клеток DU145 количество клеток, подвергающихся раннему апоптозу и позднему апоптозу/некрозу, составляло 12% после 12 часовой, 34% после 24 часовой и 59% после 48 часовой индукции. Не наблюдали увеличения апоптоза в клетках, содержащих пустую плазмиду, после индукции с использованием PonA. Исследования поглощения аннексина V и йодида пропидия показали, что hBD-1 обладает цитотоксической активностью против злокачественных клеток простаты DU145 и PC3, и результаты свидетельствуют, что механизмом гибели клеток является апоптоз.
hBD-1 вызывает изменения целостности мембран и активацию каспаз: Исследовали, является ли гибель клеток, наблюдаемая в злокачественных клетках простаты после индукции hBD-1, опосредованным каспазами апоптозом. Чтобы лучше понять клеточные механизмы, вовлеченные в экспрессию hBD-1, осуществляли анализ с использованием конфокальной лазерной микроскопии (фиг.5) на клетках DU145 и LNCaP, индуцированных в отношении экспрессии hBD-1. Активацию пан-каспазы исследовали на основе связывания и отщепления зеленого флуоресцирующего FAM-VAD-FMK от каспаз в клетках, активно подвергающихся апоптозу. Анализ клеток в условиях ДИК показал присутствие жизнеспособных контрольных клеток DU145 (фиг.5A) и LNCaP (фиг.5E) во временной точке 0 часов. Возбуждение конфокальным лазером при 488 нм не давало регистрируемого зеленого окрашивания, что свидетельствует об отсутствии каспазной активности в DU145 (фиг.5B) или LNCaP (фиг.5F). После индукции в течение 24 часов клетки DU145 (фиг.5C) и LNCaP (фиг.5G) были снова видимы в условиях ДИК. Конфокальный флуоресцентный анализ выявил зеленое окрашивание в клетках DU145 (фиг.5D), что свидетельствует о пан-каспазной активности после индукции экспрессии hBD-1. Однако зеленого окрашивания не наблюдали в клетках LNCaP (фиг.5H), индуцированных в отношении экспрессии hBD-1. Следовательно, гибель клеток, наблюдаемая после индукции hBD-1, является опосредованным каспазами апоптозом.
Предполагаемым механизмом противомикробной активности дефензиновых пептидов является нарушение мембраны микроорганизмов вследствие образования пор (Papo and Shai, 2005). Чтобы определить, изменяла ли экспрессия hBD-1 целостность мембран, исследовали поглощение EtBr в конфокальном анализе. Интактные клетки были окрашены зеленым благодаря AO, который проникает через мембрану, тогда как только клетки с нарушенными плазматическими мембранами были окрашены в красный цвет благодаря включению не проникающего через мембраны EtBr. Контрольные клетки DU145 и PC3 были позитивно окрашены AO и испускали зеленый свет, но не были окрашены EtBr. Однако индукция hBD-1 и в клетках DU145, и в клетках PC3 приводила к накоплению EtBr в цитоплазме по временной точке 24 часа, что было показано по красному окрашиванию. К 48 часам клетки DU145 и PC3 имели конденсированные ядра и выглядели желтыми, что является следствием совместного присутствия и зеленого, и красного окрашивания в результате накопления AO и EtBr, соответственно. Наоборот, не было наблюдаемых изменений в целостности мембран в клетках LNCaP после 48 часовой индукции, о чем свидетельствует позитивная зеленая флуоресценция AO, но отсутствие красной флуоресценции EtBr. Полученные данные показывают, что изменения целостности мембран и проницаемость в ответ на экспрессию hBD-1 отличается в злокачественных клетках простаты на ранней и поздней стадиях.
Сравнение уровней экспрессии hBD-1 и cMYC: Осуществляли КОТ-ПЦР-анализ на полученных с использованием LCM срезах ткани простаты от трех пациентов (фиг.34). У пациента №1457 наблюдали, что экспрессия hBD-1 снижалась в 2,7 раза при переходе от нормальной ткани к PIN, снижалась в 3,5 раза при переходе от PIN к опухоли и снижалась в 9,3 раза при переходе от нормальной ткани к опухоли (фиг.34A). Подобным образом экспрессия cMYC имела сходную картину у пациента №1457, при этом экспрессия снижалась в 1,7 раза при переходе от нормальной ткани к PIN, в 1,7 раза при переходе от PIN к опухоли и 2,8 раза при переходе от нормальной ткани к опухоли (фиг.34B). Кроме того, имело место статистически значимое снижение экспрессии cMYC у двух других пациентов. Пациент №1569 имел 2,3-кратное снижение при переходе от нормальной ткани к PIN, тогда как у пациента №1586 происходило 1,8-кратное снижение при переходе от нормальной ткани к PIN, 4,3-кратное снижение при переходе от PIN к опухоли и 7,9-кратное снижение при переходе от нормальной ткани к опухоли.
Индукция экспрессии hBD-1 после ингибирования PAX2: Чтобы дополнительно исследовать роль PAX2 в регуляции экспрессии hBD-1, использовали ми-РНК для нокдауна экспрессии PAX2 и осуществляли КОТ-ПЦР, чтобы наблюдать экспрессию hBD-1. Обработка клеток hPrEC ми-РНК PAX2 не оказывала влияния на экспрессию hBD-1 (фиг.35). Однако нокдаун PAX2 приводил к 42-кратному увеличению в LNCaP, 37-кратному увеличению в PC3 и 1026-кратному увеличению в DU145 экспрессии hBD-1 по сравнению с необработанными клетками. В качестве негативного контроля клетки обрабатывали неспецифичной ми-РНК, которая не оказывала значимого влияния на экспрессию hBD-1.
ПРИМЕР 11: ИНГИБИРОВАНИЕ ЭКСПРЕССИИ PAX2 ПРИВОДИТ К АЛЬТЕРНАТИВНЫМ ПУТЯМ КЛЕТОЧНОЙ ГИБЕЛИ В ЗЛОКАЧЕСТВЕННЫХ КЛЕТКАХ ПРОСТАТЫ, ОТЛИЧАЮЩИМСЯ ПО СТАТУСУ P53
Материалы и способы
Линии клеток: Линии злокачественных клеток PC3, DU145 и LNCaP, которые отличаются по мутационному статусу p53 (таблица 7), культивировали, как описано в примере 1.
Сайленсинг PAX2 под влиянием ми-РНК и Вестерн-анализ осуществляли, как описано в примере 2. Затем блоты исследовали с использованием первого кроличьего анти-PAX2-антитела (Zymed, San Francisco, CA) в разведении 1:1000. После промывки мембраны инкубировали с антикроличьим антителом, конъюгированным с пероксидазой хрена (HRP) (разведение 1:5000; Sigma), и при регистрации сигнал визуализировали, используя хемилюминесцентные реагенты (Pierce), на Alpha Innotech Fluorchem 8900. В качестве контроля с блотов удаляли антитела и повторно исследовали, используя первое мышиное антитело против β-актина (1:5000; Sigma-Aldrich) и конъюгированное с HRP второе антитело против Ig мыши (1:5000; Sigma-Aldrich), и регистрацию сигнала снова визуализировали.
Мутация гена p53 в линиях злокачественных клеток простаты
Фазово-контрастная микроскопия и MTT-анализ цитотоксичности осуществляли, как описано в примере 2.
Определение пан-каспазы и количественную ОТ-ПЦР в реальном времени осуществляли, как описано в примере 1. Пары праймеров для BAX, BID, BCL-2, AKT и BAD создавали на основе опубликованных последовательностей (таблица 8). Реакции осуществляли в 96-луночном оптическом планшете для реакции MicroAmp (PE Biosystems). Проводили сорок циклов ПЦР в стандартных условиях, используя температуру отжига 60°C. Количественную оценку осуществляли на основании номера цикла, в котором начиналась экспоненциальная амплификация (пороговое значение), и усредняли, используя значения, полученные в трех повторах. Имела место обратная зависимость между уровнем мессенджера и пороговым значением. Кроме того, GAPDH использовали в качестве гена домашнего хозяйства, чтобы нормализовать начальное содержание суммарной кДНК. Относительную экспрессию рассчитывали в виде соотношения экспрессии между каждым геном и GAPDH. Все реакции осуществляли в трех повторах.
Праймеры для количественной ОТ-ПЦР 
Анализ проницаемости мембран осуществляли, как описано в примере 3. Клетки PC3 и LNCaP трансфицировали ми-РНК PAX2, неспецифичными ми-РНК или только средой.
Анализ экспрессии белка PAX2 в клетках простаты: Экспрессию белка PAX2 исследовали, используя Вестерн-анализ, в первичной культуре клеток простаты HPrEC и в линиях злокачественных клеток простаты LNCaP, DU145 и PC3. В данном случае белок PAX2 был выявлен во всех линиях злокачественных клетках простаты (фиг.36A). Однако белок PAX2 не выявляли в HPrEC. С блотов удаляли антитела и повторно исследовали в отношении β-актина в качестве внутреннего контроля, чтобы гарантировать равную нагрузку. Экспрессию белка PAX2 также исследовали после избирательного целенаправленного воздействия и ингибирования PAX2-специфичной ми-РНК в линиях злокачественных клеток простаты DU145, PC3 и LNCaP. Клетки подвергали одному раунду трансфекции пулом ми-РНК PAX2 в течение 6-дневного периода обработки. Белок PAX2 экспрессировался в контрольных клетках, обработанных только средой. Специфичное целенаправленное воздействие на мРНК PAX2 подтверждали, наблюдая нокдаун белка PAX2 во всех трех линиях клеток (фиг.36B).
Влияние нокдауна PAX2 на рост злокачественных клеток простаты: Влияние ми-РНК PAX2 на количество клеток и жизнеспособность клеток анализировали, используя световую микроскопию и MTT-анализ. Чтобы исследовать влияние ми-РНК PAX2 на количество клеток, линии клеток PC3, DU145 и LNCaP трансфицировали только средой, неспецифичной ми-РНК или ми-РНК PAX2 в течение 6 дней. Каждая линия клеток достигала слияния на 80-90% в 60-мм культуральных чашках, содержащих только среду. Обработка клеток HPrEC, DU145, PC3 и LNCaP неспецифичной ми-РНК очевидно оказывала мало влияния или не оказывала влияния на рост клеток по сравнению с клетками, обработанными только средой (фиг.37A, 37C и 37E соответственно). Обработка PAX2-нулевой линии клеток HPrEC ми-РНК PAX2 очевидно не оказывала значимого влияния на рост клеток (фиг.37B). Однако обработка линий злокачественных клеток простаты DU145, PC3 и LNCaP ми-РНК PAX2 приводила к значимому снижению количества клеток (фиг.37D, 37F и 37H соответственно).
Влияние нокдауна PAX2 на жизнеспособность злокачественных клеток простаты: Жизнеспособность клеток измеряли после 2-, 4- и 6-дневных периодов воздействия. Жизнеспособность в процентах вычисляли в виде отношения оптической плотности при 570-630 нм клеток, обработанных ми-РНК PAX2 к оптической плотности необработанных контрольных клеток. В качестве негативных контролей жизнеспособность клеток измеряли после каждого периода обработки используемой в качестве негативного контроля неспецифичной ми-РНК или трансфекции с использованием только реагента. Относительную жизнеспособность клеток вычисляли делением жизнеспособности в процентах после обработки ми-РНК PAX2 на жизнеспособность в процентах после обработки неспецифичной ми-РНК (фиг.38). После 2-дневной обработки относительная жизнеспособность составляла 116% в DU145, 81% в PC3 и 98% в LNCaP. После 4-дневной обработки относительная жизнеспособность клеток снижалась до 69% в DU145, 79% в PC3 и 80% в LNCaP. Наконец, к 6 дню относительная жизнеспособность составляла 63% в DU145, 43% в PC3 и 44% в LNCaP. Кроме того, жизнеспособность клеток также измеряли после обработки только реагентом для трансфекции. В данном случае, в каждой линии клеток не наблюдали значимого снижения жизнеспособности клеток.
Выявление пан-каспазной активности: Каспазную активность регистрировали в анализе с использованием конфокальной лазерной микроскопии. Клетки LNCaP, DU145 и PC3 обрабатывали ми-РНК PAX2 и наблюдение активности осуществляли на основе связывания FAM-меченого пептида с каспазами в клетках, активно подвергающихся апоптозу, которые будут давать зеленую флуоресценцию. Анализ клеток с пользованием только среды показывает присутствие жизнеспособных клеток LNCaP, DU145 и PC3, соответственно. Возбуждение конфокальным лазером при 488 нм не давало регистрируемого зеленого окрашивания, что свидетельствует об отсутствии каспазной активности в необработанных клетках (фиг.39A, 39C и 39E соответственно). После 4 дней обработки ми-РНК PAX2 клетки LNCaP, DU145 и PC3 в условиях флуоресценции имели зеленую окраску, свидетельствующую о каспазной активности (фиг.39B, 39D и 39F соответственно).
Влияние ингибирования PAX2 на апоптозные факторы: Клетки LNCaP, DU145 и PC3 обрабатывали ми-РНК против PAX2 в течение 4 дней и экспрессию проапоптозных и противоапоптозных факторов измеряли, используя КОТ-ПЦР. После нокдауна PAX2 анализ BAD выявил увеличение в 2 раза в LNCaP, в 1,58 раза в DU145 и в 1,375 раза в PC3 (фиг.40A). Уровни экспрессии BID возрастали в 1,38 раз в LNCaP и в 1,78 раза в DU145, но не было статистически значимого различия в уровне BID, наблюдаемом в PC3 после подавления экспрессии PAX2 (фиг.40B). Анализ противоапоптозного фактора AKT выявил 1,25-кратное снижение экспрессии в LNCaP и 1,28-кратное снижение в DU145 после обработки, но не наблюдали изменения в PC3 (фиг.40C).
Анализ целостности мембраны и некроза: Целостность мембран контролировали, используя конфокальный анализ, в клетках LNCaP, DU145 и PC3. В данном случае интактные клетки были окрашены в зеленый цвет благодаря AO, который проникает через мембрану, тогда как клетки с нарушенными плазматическими мембранами могут окрашиваться красным благодаря включению непроникающего через мембрану EtBr в цитоплазму и желтым благодаря совместной локализации AO и EtBr в ядрах. Необработанные клетки LNCaP, DU145 и PC3 позитивно окрашивались AO и испускали зеленый свет, но не были окрашены EtBr. После нокдауна PAX2 не было наблюдаемых изменений целостности мембран в клетках LNCaP, что показано на основании позитивной зеленой флуоресценции AO и отсутствия красной флуоресценции EtBr. Полученные данные дополнительно свидетельствуют, что клетки LNCaP могут подвергаться апоптозной, но не некротической гибели клеток после нокдауна PAX2. Наоборот, нокдаун PAX2 в DU145 и PC3 приводил к накоплению EtBr в цитоплазме, что показано на основании красного окрашивания. Кроме того, клетки DU145 и PC3 имели конденсированные ядра, которые выглядели желтыми вследствие совместной локализации зеленого и красного окрашивания за счет AO и EtBr, соответственно. Полученные результаты свидетельствуют, что DU145 и PC3 подвергаются альтернативному пути клеточной гибели, включающему некротическую гибель клеток по сравнению с LNCaP.
ПРИМЕР 12: ЭКСПРЕССИЯ PAX2 и DEFB-1 В ЛИНИЯХ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ И ТКАНЯХ МОЛОЧНОЙ ЖЕЛЕЗЫ С ПРОТОКОВОЙ ИЛИ ДОЛЬКОВОЙ ВНУТРИЭПИТЕЛИАЛЬНОЙ НЕОПЛАЗИЕЙ
Экспрессия PAX2 и DEFB-1 будет определена в образцах биопсии молочной железы с протоковой или дольковой внутриэпителиальной неоплазией и в следующих линиях злокачественных клеток молочной железы:
BT-20: выделена из первичной инвазивной протоковой карциномы; клетки экспрессируют E-кадгерин, ER, EGFR и uPA.
BT-474: выделена из первичной инвазивной протоковой карциномы; клетки экспрессируют E-кадгерин, ER, PR и имеют амплифицированный HER2/neu.
Hs578T: выделена из первичной инвазивной протоковой карциномы; также создана линия клеток из нормальной близлежащей ткани, названная Hs578Bst.
MCF-7: создана из плеврального выпота. Клетки экспрессируют ER и являются наиболее распространенным примером отвечающих на эстроген злокачественных клеток молочной железы.
MDA-MB-231: создана из плеврального выпота. Клетки являются ER-негативными, E-кадгерин-негативными и высоко инвазивными в анализах in vitro.
MDA-MB-361: создана из метастаза головного мозга. Клетки экспрессируют ER, PR, EGFR и HER2/neu.
MDA-MB-435: создана из плеврального выпота. Клетки являются ER-негативными, E-кадгерин-негативными и являются высоко инвазивными и метастатическими у иммунодефицитных мышей.
MDA-MB-468: создана из плеврального выпота. Клетки имеют амплифицированный EGFR и являются ER-негативными.
SK-BR-3: создана из плеврального выпота. Клетки имеют амплифицированный HER/2neu, экспрессируют EGFR и являются ER-негативными.
T-47D: создана из плеврального выпота. В клетках сохраняется экспрессия E-кадгерина, ER и PR.
ZR-75-1: создана из асцитной жидкости. Клетки экспрессируют ER, E-кадгерин, HER2/neu и VEGF.
Отношение экспрессии PAX2-к-DEFB-1 будет определено с применением способов, описанных в примере 9.
ПРИМЕР 13: ЭКСПРЕССИЯ DEFB1 В КЛЕТКАХ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ
DEFB1 будет экспрессирован в злокачественных клетках молочной железы с применением способов, описанных в примере 1. Жизнеспособность клеток и каспазная активность будут определены, как описано в примере 1.
ПРИМЕР 14: ИНГИБИРОВАНИЕ ЭКСПРЕССИИ PAX2 В КЛЕТКАХ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ
Экспрессия PAX2 в клетках рака молочной железы будет ингибирована с использованием ми-РНК, описанной в примере 2. Уровни экспрессии проапоптозных генов, таких как BAX, BID и BAD, жизнеспособность клеток и каспазная активность будут определены, как описано в примере 2.
ПРИМЕР 15: ВЛИЯНИЕ ЭКСПРЕССИИ DEFB1 НА РОСТ ОПУХОЛЕЙ IN VIVO
Противоопухолевая способность DEFB1 будет оценена посредством инъекции клеток рака молочной железы, которые сверхэкспрессируют DEFB1, мышам nude. Клетки рака молочной железы будут трансфицированы экспрессирующим вектором, несущим ген DEFB1. Будут отобраны и клонированы клетки, экспрессирующие экзогенный ген DEFB1. Используют только суспензии одиночных клеток с жизнеспособностью >90%. Каждое животное получает примерно 500000 клеток, вводимых подкожно в правый бок самкам мышей nude. Имеется две группы, контрольная группа, которой инъецируют клоны только с вектором, и группа, которой инъецируют клоны, сверхэкспрессирующие DEFB1. Каждая группа содержит 35 мышей, что определено статистически. Животных взвешивают два раза в неделю, рост опухолей контролируют, используя штангенциркули, и объем опухолей определяют, используя следующую формулу: объем=0,5×(ширина)2×длина. Всех животных умерщвляют, используя избыточную дозу CO2, когда размер опухоли достигает 2 мм3 или через 6 месяцев после имплантации; опухоли вырезают, взвешивают и хранят в нейтральном забуференном формалине для патологического исследования. Различия в росте опухолей между группами характеризуют, используя сводные статистические данные и графические представления. Статистическую значимость оценивают, используя либо t-критерий, либо непараметрический аналог.
ПРИМЕР 16: ВЛИЯНИЕ МИ-РНК PAX2 НА РОСТ ОПУХОЛЕЙ IN VIVO
Матричные олигонуклеотиды ми-РНК PAX2, имеющих форму шпильки, используемые в исследованиях in vitro, применяют для исследования влияния повышающей регуляции экспрессии DEFB1 in vivo. Смысловую и антисмысловую нити (см. таблицу 3) отжигают и клонируют в векторе pSilencer 2.1 U6 hygro, экспрессирующем ми-РНК (Ambion) под контролем промотора РНК-полимеразы III U6 человека. Клонированную плазмиду секвенируют, проверяют и трансфицируют в линии клеток рака молочной железы. Рандомизированную ми-РНК клонируют и используют в качестве негативного контроля в данном исследовании. Отбирают резистентные к гигромицину колонии, клетки подкожно вводят мышам и рост опухолей наблюдают, как описано выше.
ПРИМЕР 17: ВЛИЯНИЕ НИЗКОМОЛЕКУЛЯРНЫХ ИНГИБИТОРОВ СВЯЗЫВАНИЯ PAX2 НА КЛЕТКИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ
Альтернативные ингибирующие олигонуклеотиды, описанные в примере 6, будут трансфицированы в клетки рака молочной железы с использованием реагента липофектамина или реагента для трансфекции Codebreaker (Promega, Inc). Чтобы подтвердить взаимодействия ДНК-белок двунитевые олигонуклеотиды будут помечены [32P]-dCTP и подвергнуты анализам сдвига электрофоретической подвижности. Мониторинг экспрессии DEFB1 будет проведен с использованием КОТ-ПЦР и Вестерн-анализа после обработки олигонуклеотидами. Наконец, гибель клеток будет выявлена с использованием MTT-анализа и проточной цитометрии, как описано ранее.
Приведенное выше описание служит в качестве руководства для специалиста в данной области в отношении того, как осуществить настоящее изобретение на практике, и не предназначено для детализации всех его очевидных модификаций и изменений, которые будут понятны специалисту при чтении описания. Однако подразумевается, что все такие очевидные модификации и изменения включены в объем настоящего изобретения, который определен следующей далее формулой изобретения. Подразумевается, что формула изобретения охватывает заявленные компоненты и стадии в любой последовательности, которая эффективно удовлетворяет предполагаемым целям, если контекст специально не указывает обратное.
SEQ ID NO: 1-69

















| название | год | авторы | номер документа |
|---|---|---|---|
| ФОСФОДИЭСТЕРАЗА 9А В КАЧЕСТВЕ МАРКЕРА ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2010 |
|
RU2592668C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2016 |
|
RU2760577C2 |
| СПОСОБ СКРИНИНГА И МОНИТОРИНГА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ (ВАРИАНТЫ) | 2012 |
|
RU2537263C2 |
| БИОМАРКЕРЫ РАКА | 2007 |
|
RU2460075C2 |
| ИДЕНТИФИКАЦИЯ ОПУХОЛЕАССОЦИИРОВАННЫХ АНТИГЕНОВ ДЛЯ ДИАГНОСТИКИ И ТЕРАПИИ | 2013 |
|
RU2644686C2 |
| АГЕНТ, ИНДУЦИРУЮЩИЙ ИММУНИТЕТ | 2012 |
|
RU2634862C2 |
| СПОСОБЫ И НАБОРЫ ДЛЯ МОЛЕКУЛЯРНОГО СУБТИПИРОВАНИЯ ОПУХОЛЕЙ | 2014 |
|
RU2690241C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И ОДНОЦЕПОЧЕЧНЫЕ ФРАГМЕНТЫ АНТИТЕЛ ПРОТИВ КЛЕТОЧНО-ПОВЕРХНОСТНОГО СПЕЦИФИЧЕСКОГО ДЛЯ ПРОСТАТЫ МЕМБРАННОГО АНТИГЕНА | 2006 |
|
RU2458073C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2603742C2 |
| СРЕДСТВО, ИНДУЦИРУЮЩЕЕ ИММУНИТЕТ | 2012 |
|
RU2614386C2 |































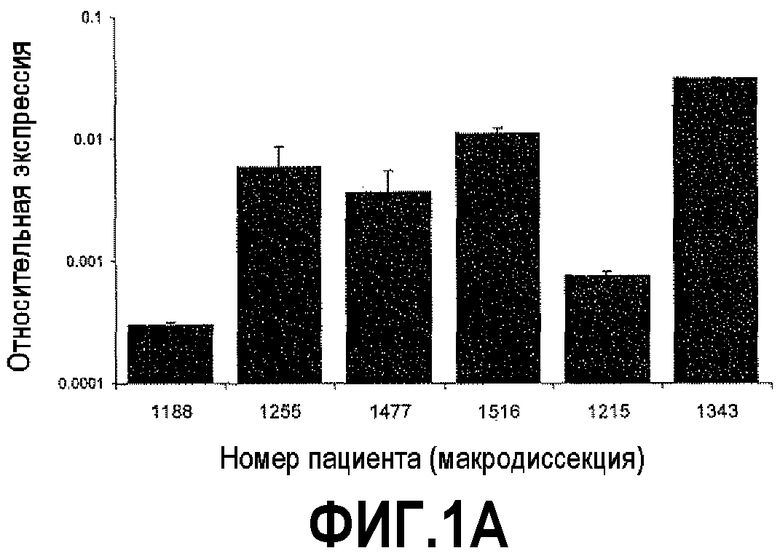
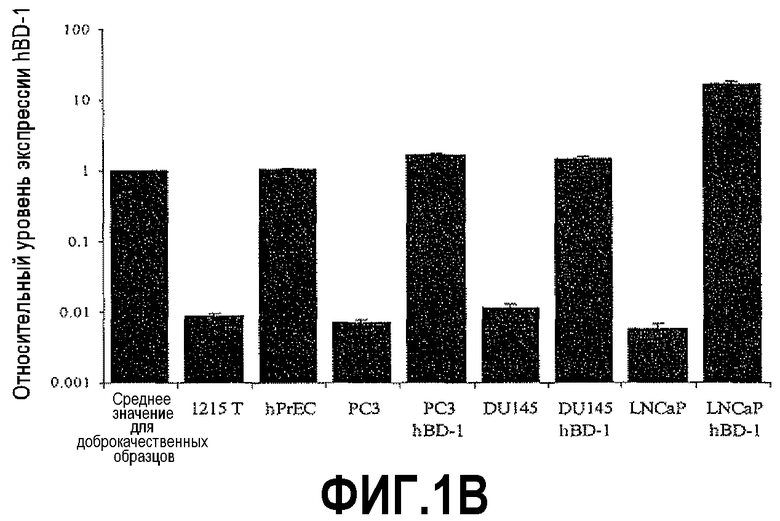
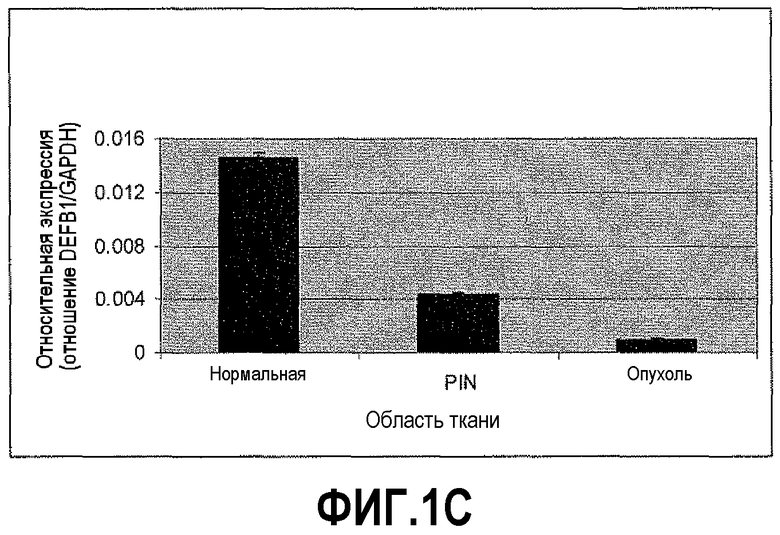
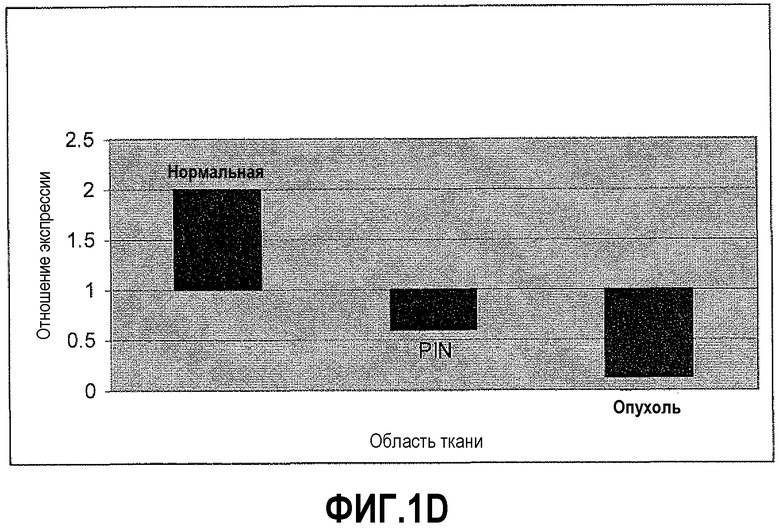
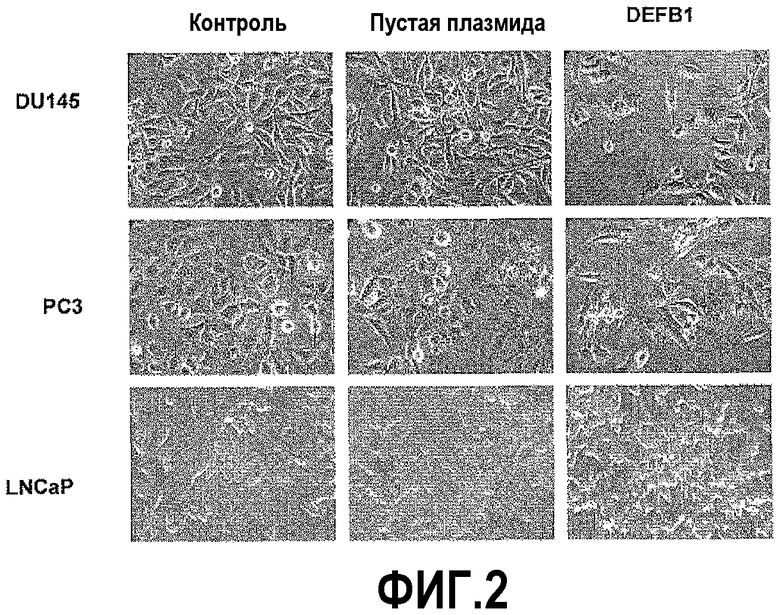
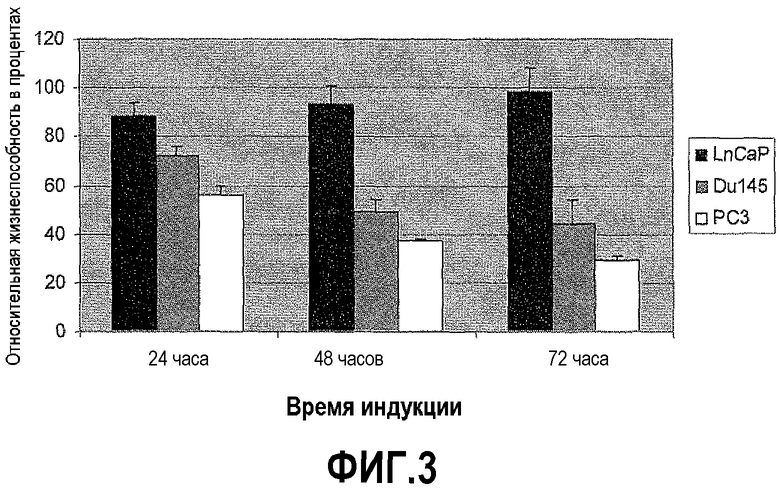
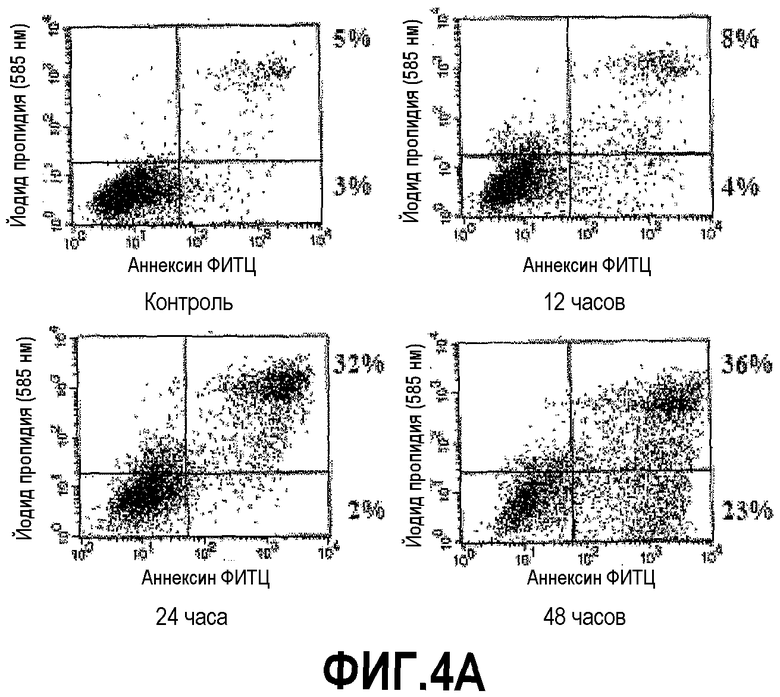
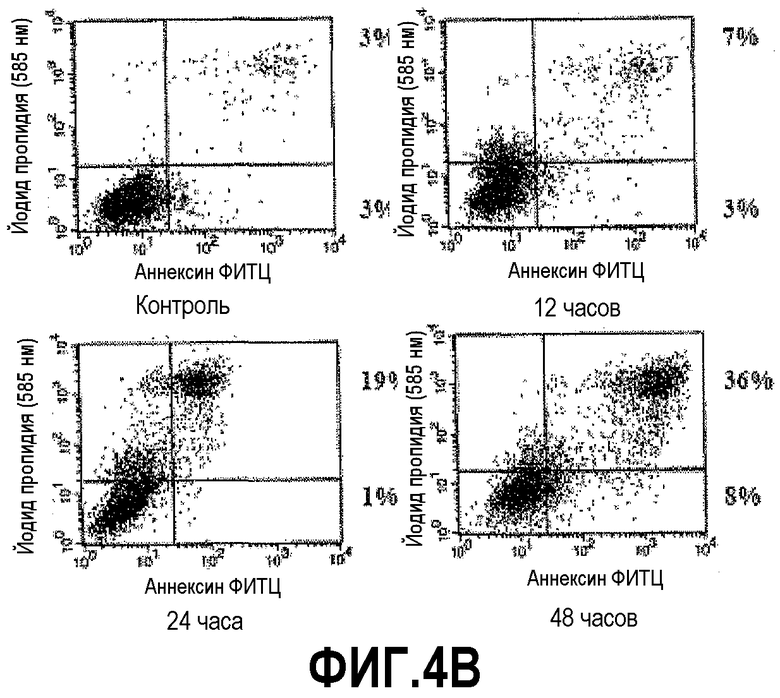
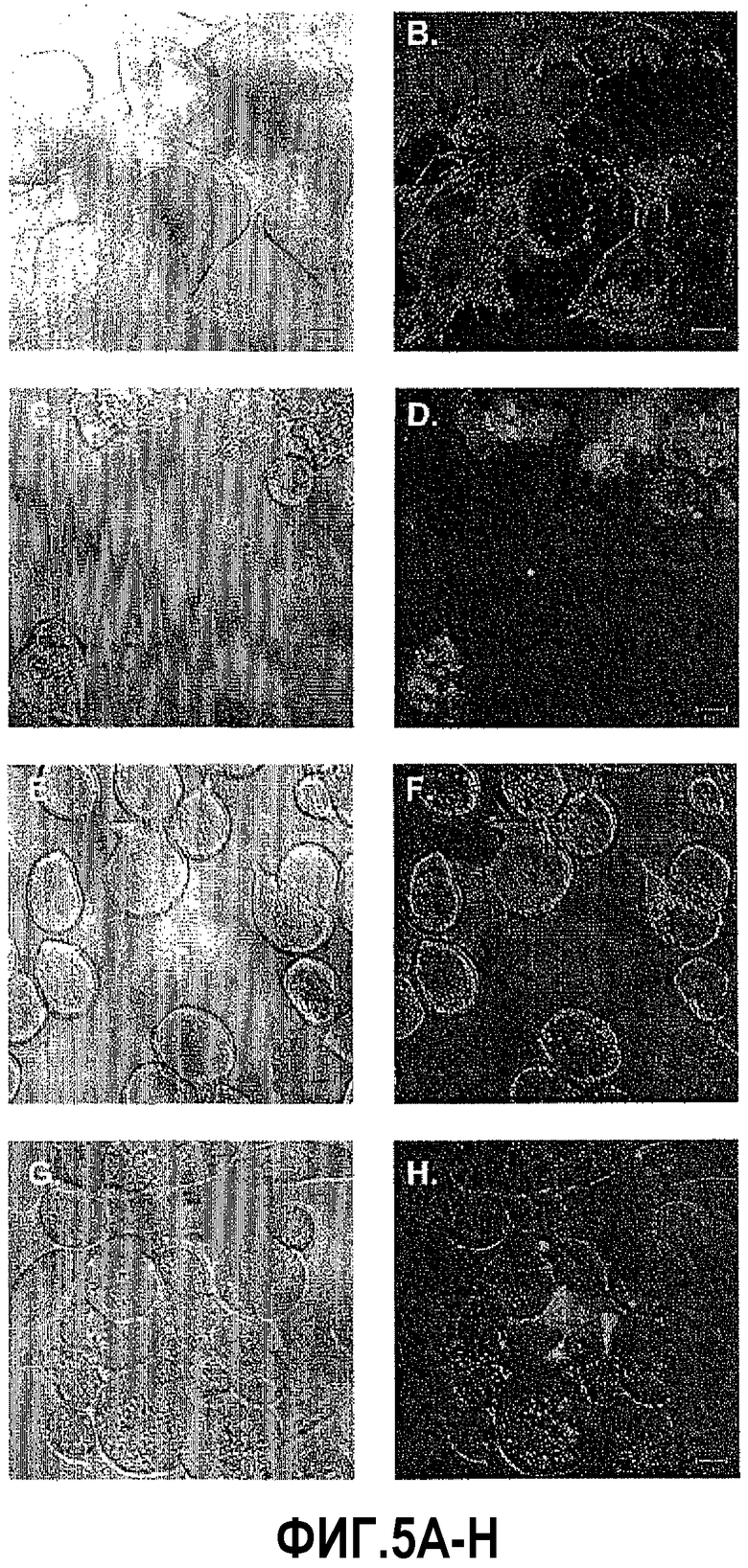
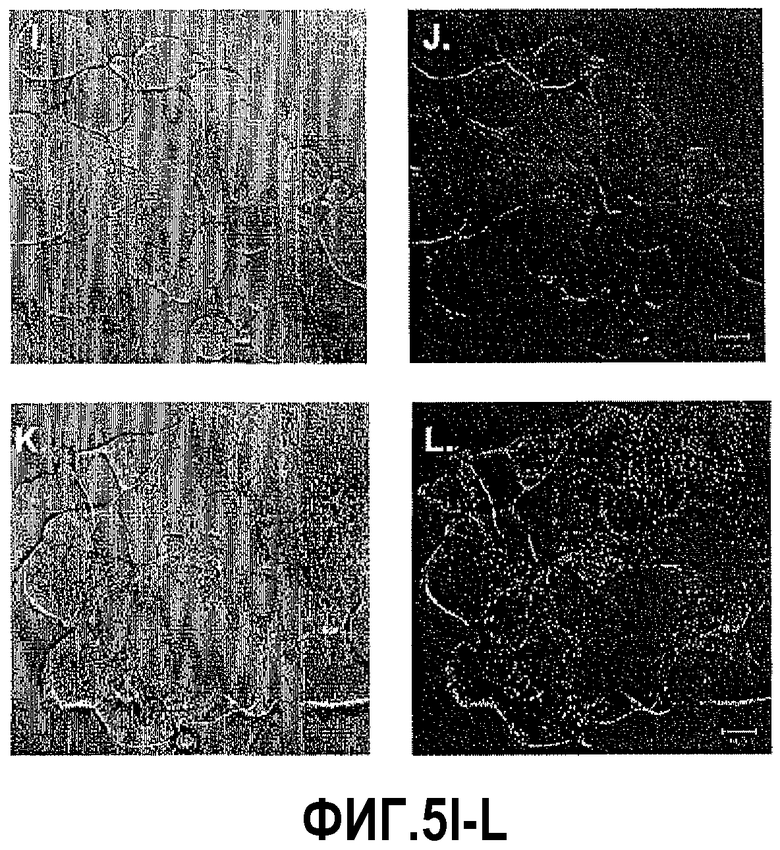
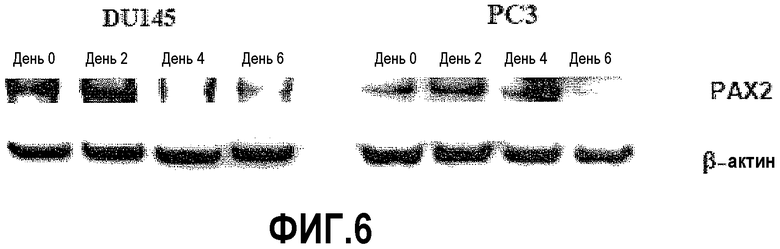
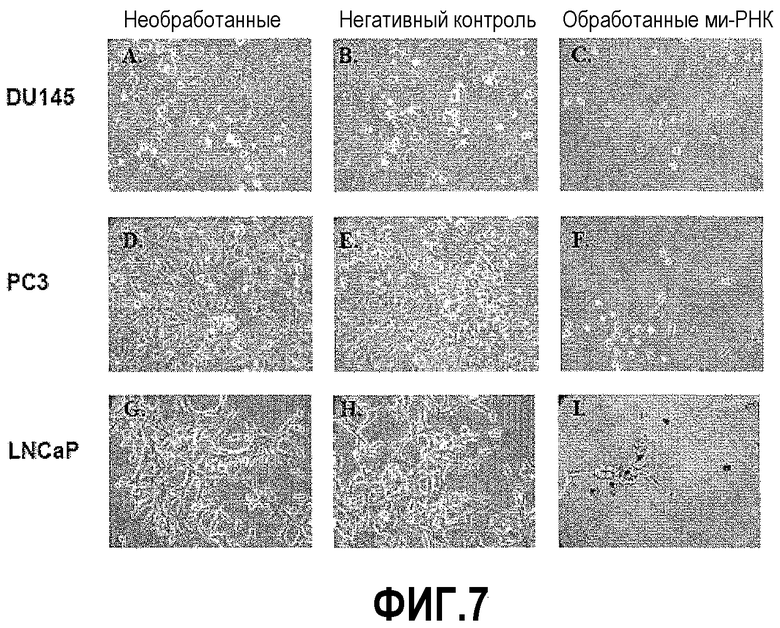
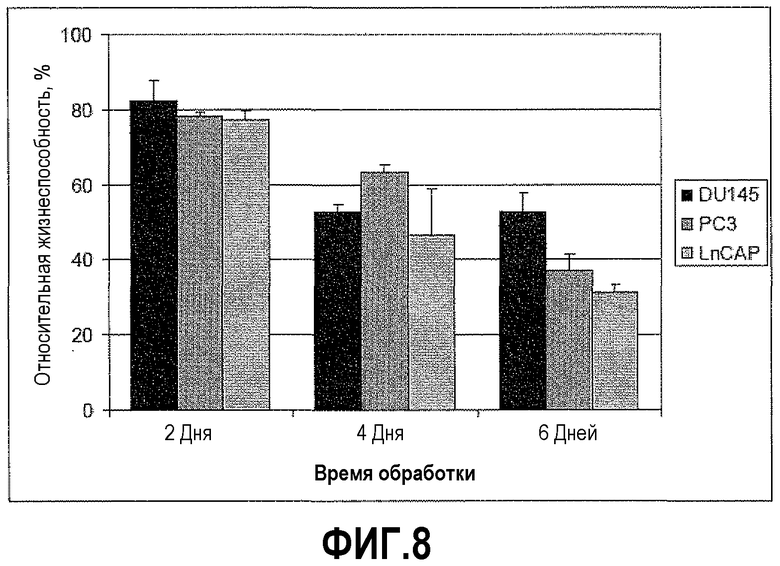
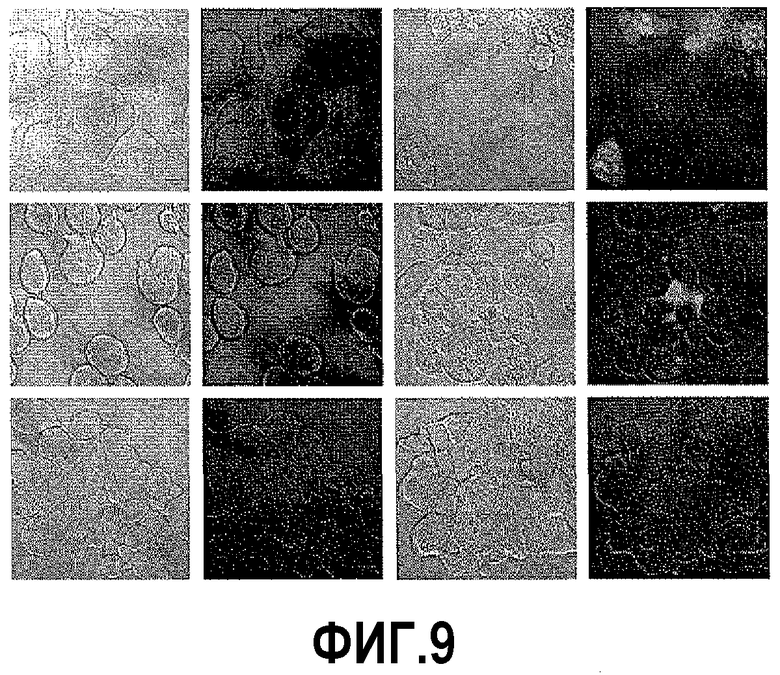
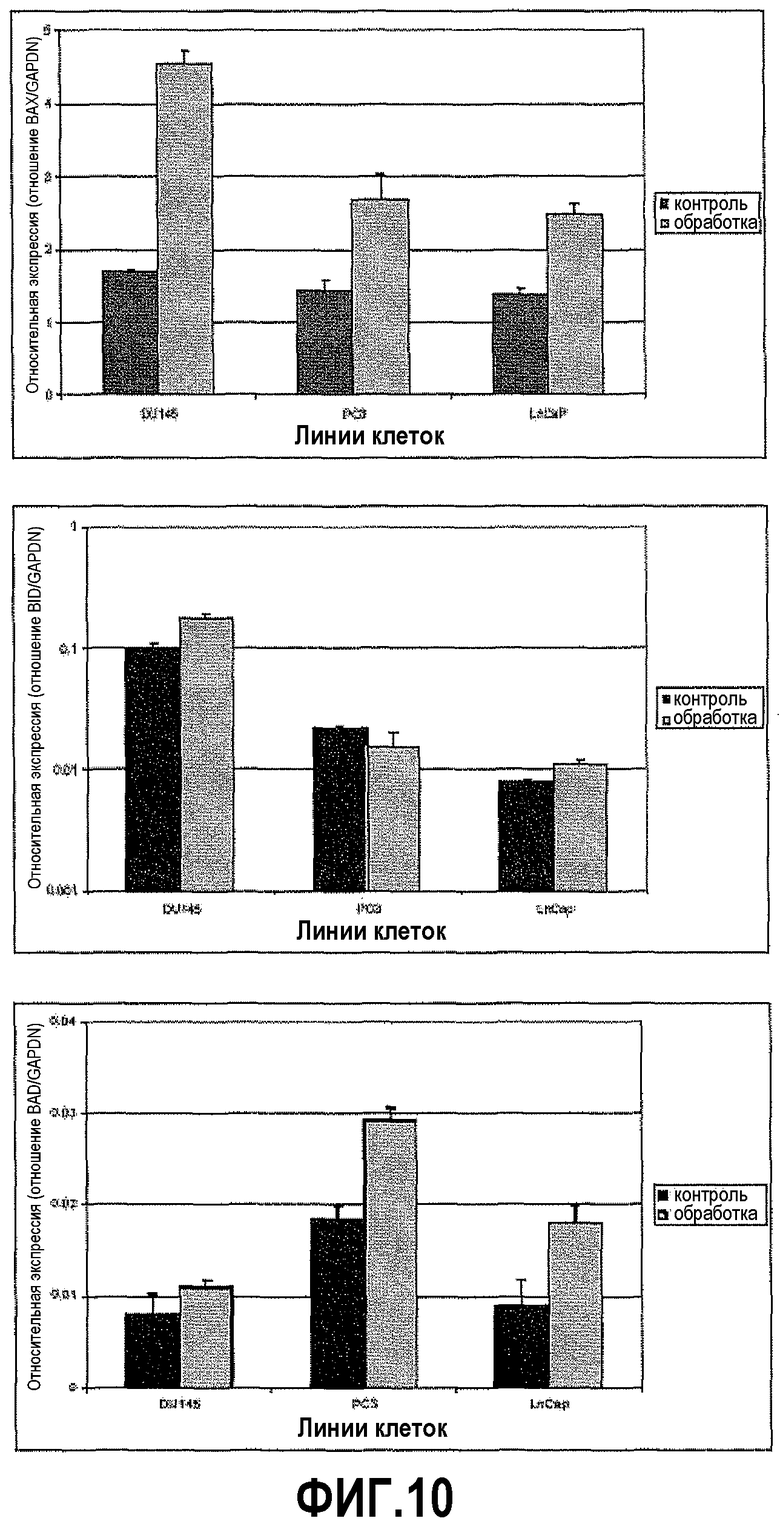
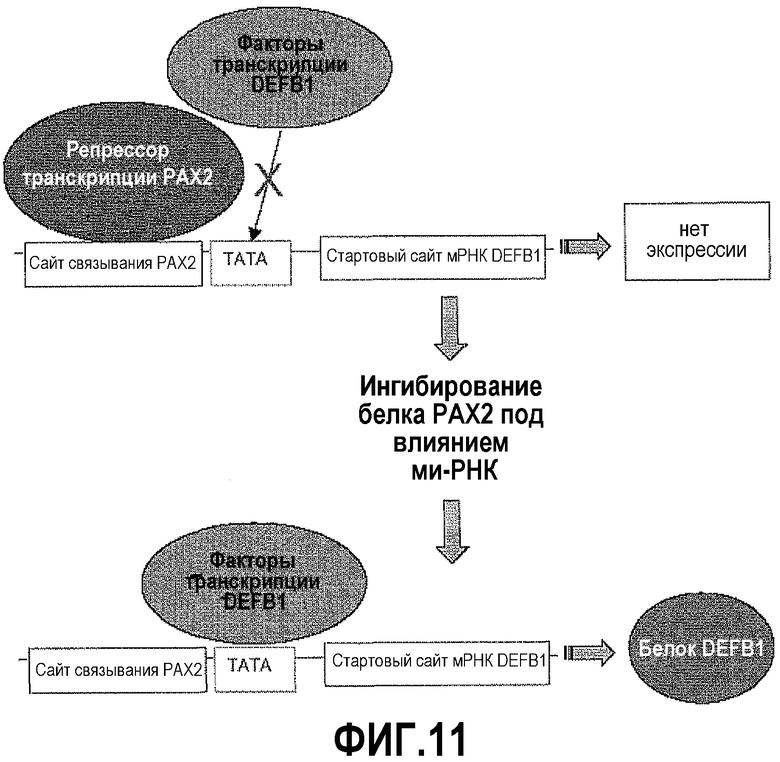
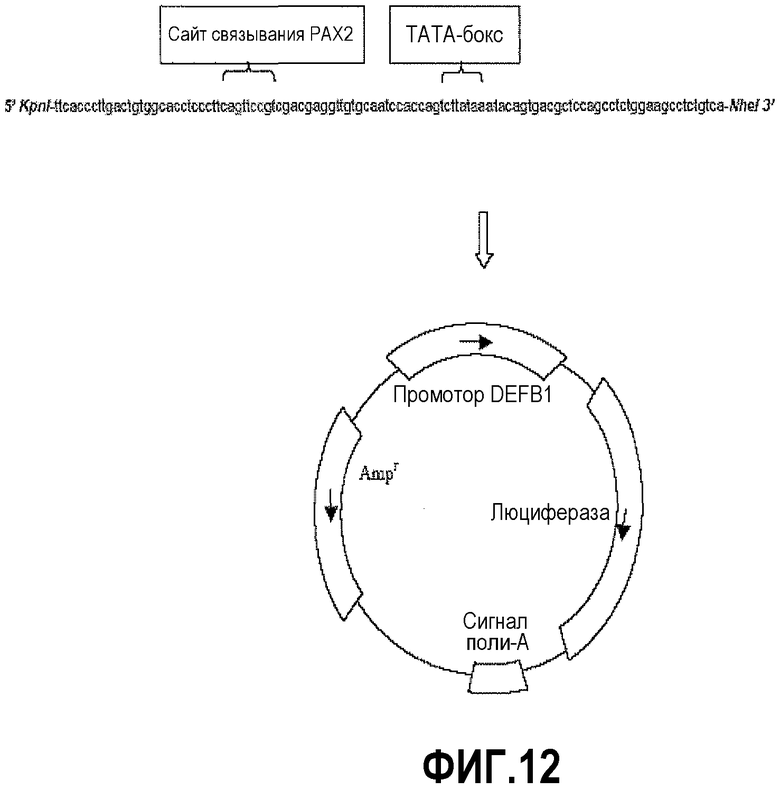
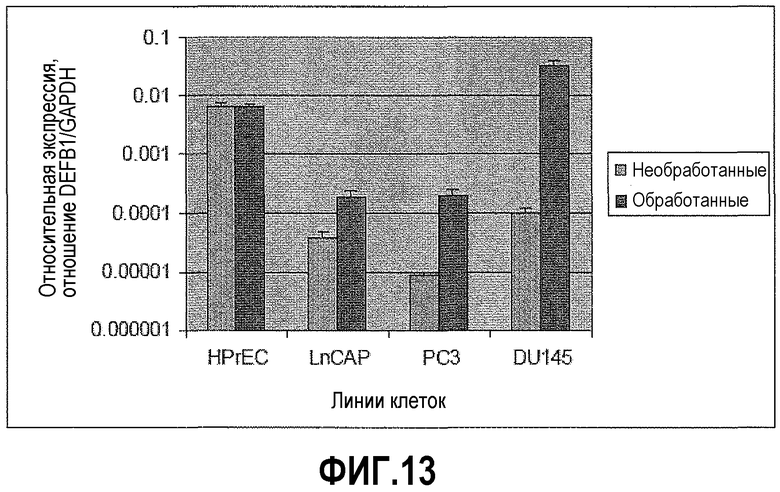
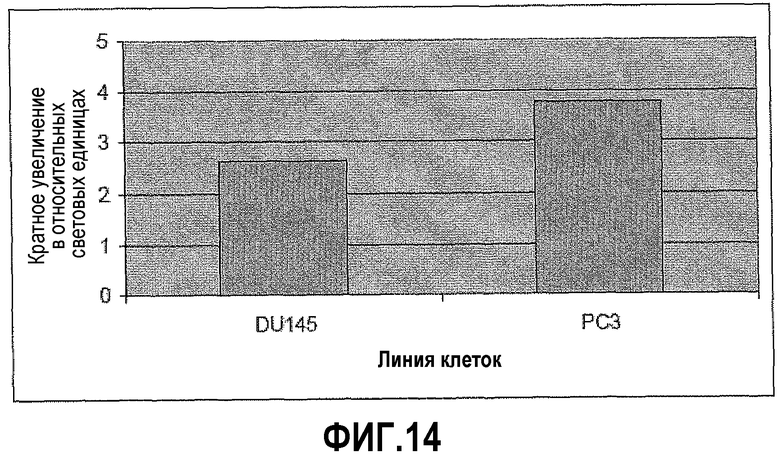
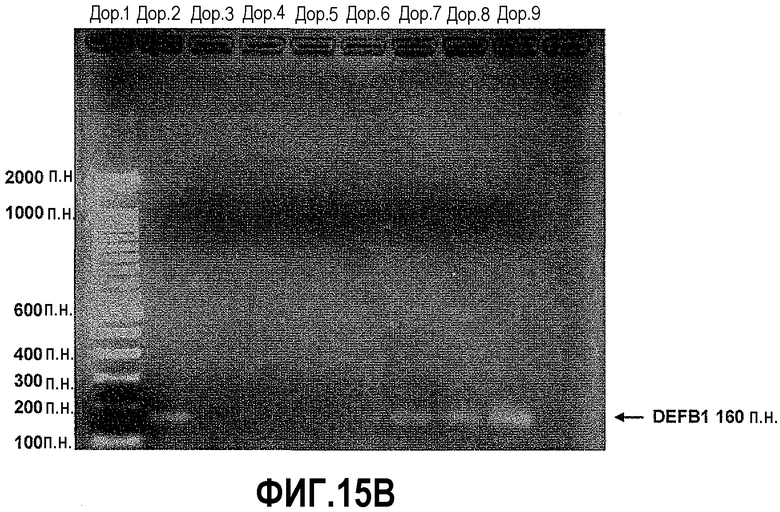

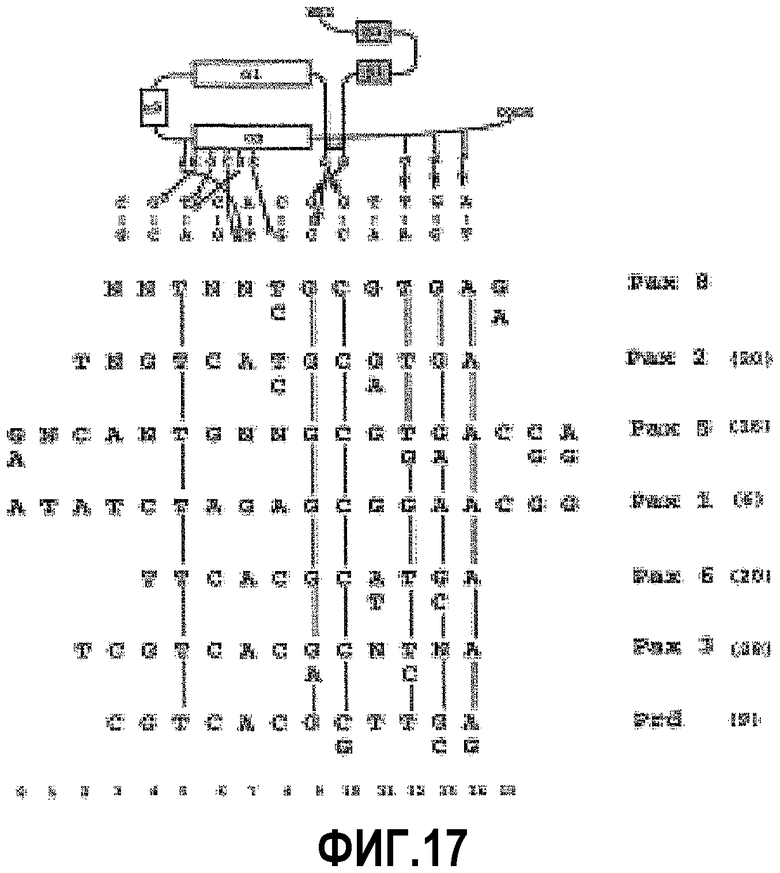
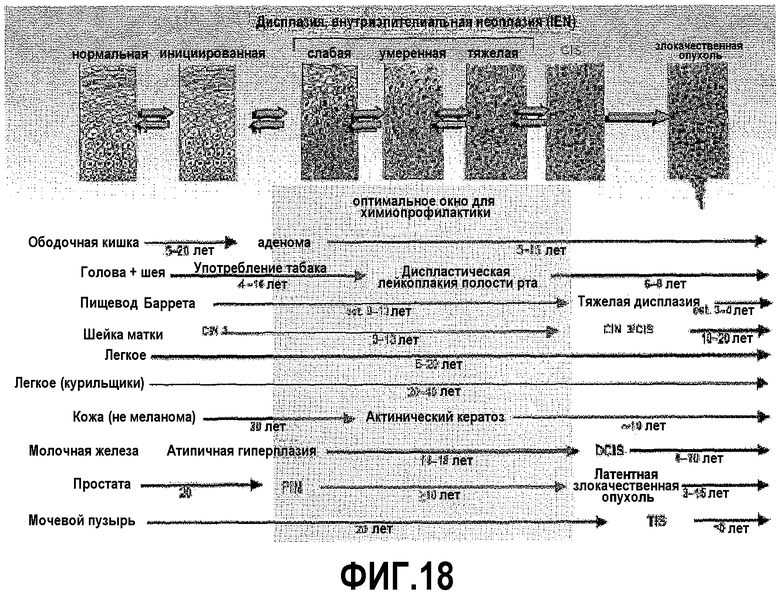
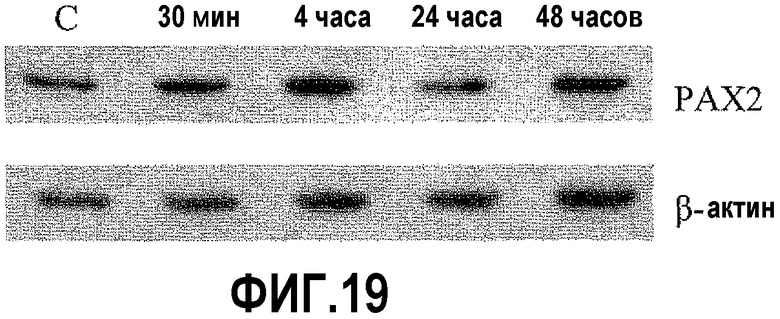
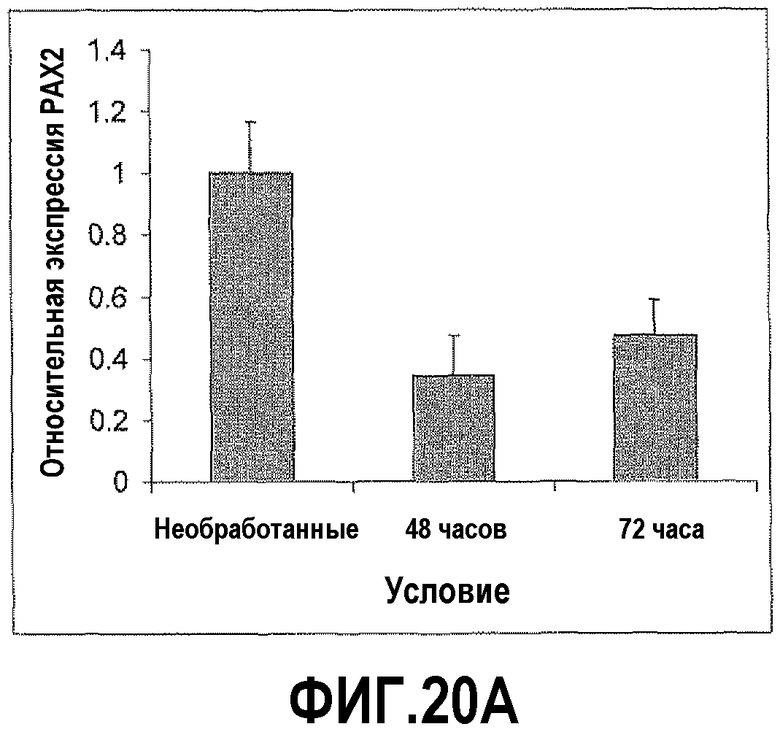
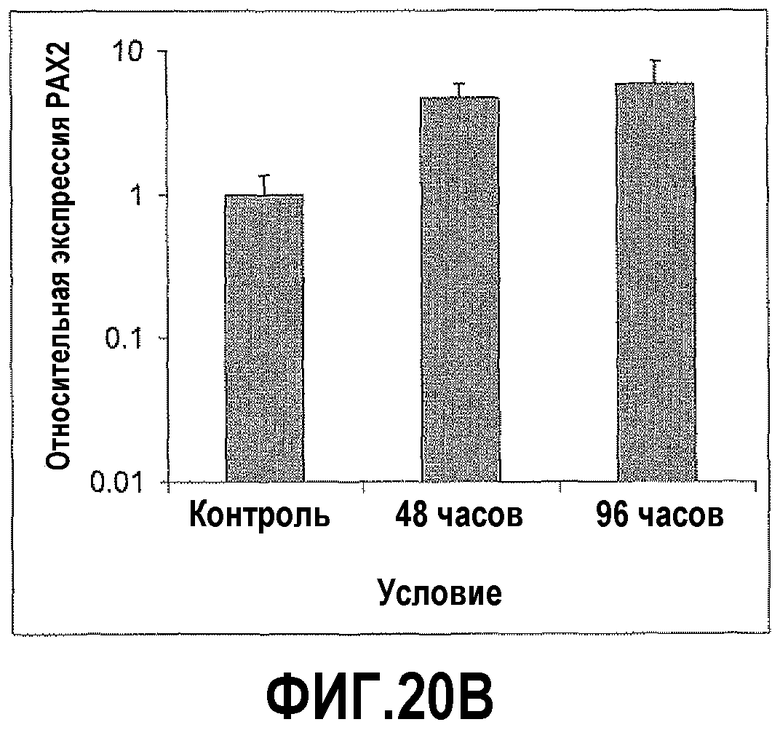
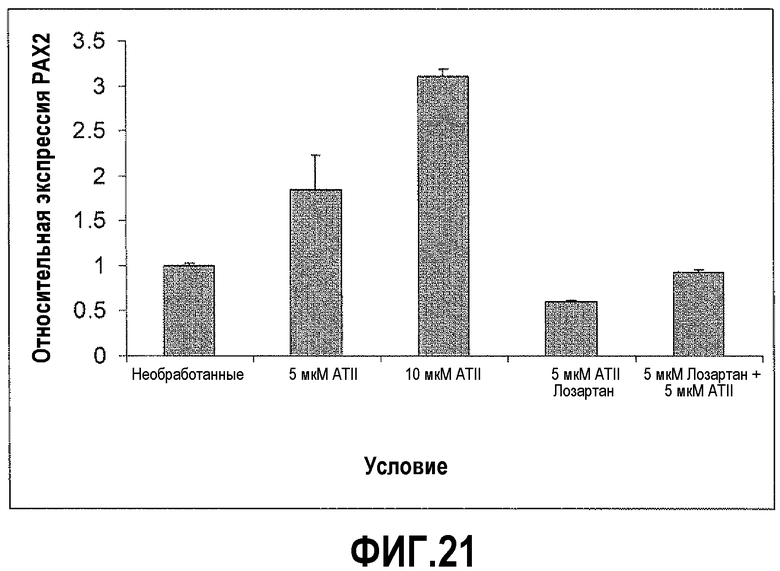
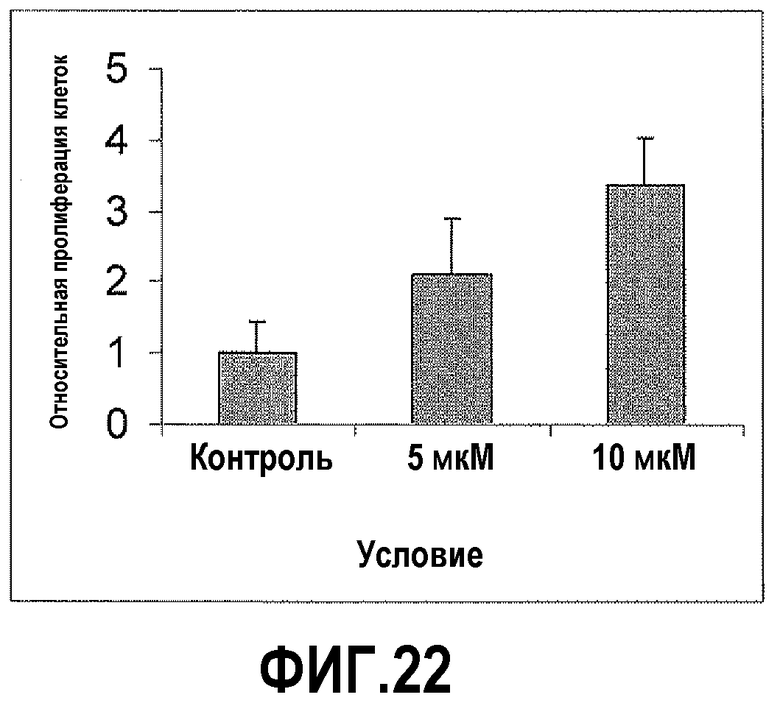
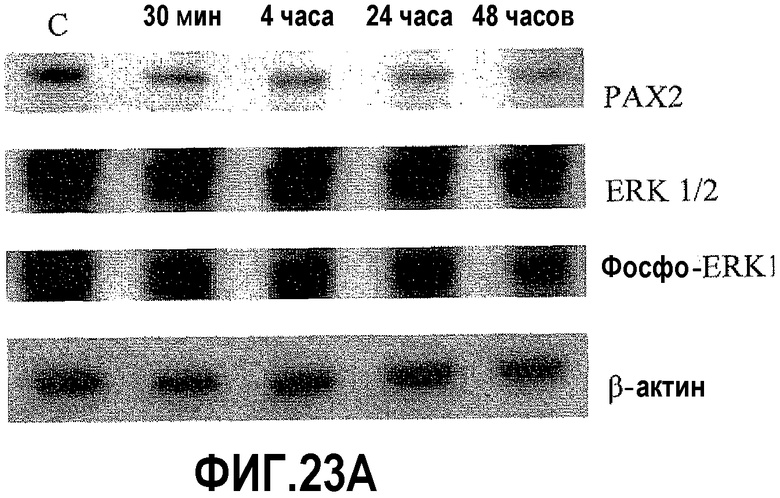
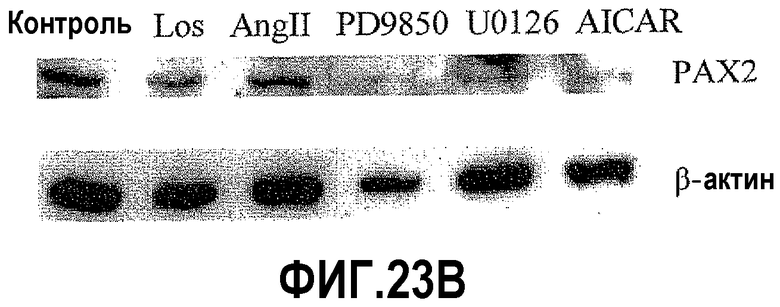


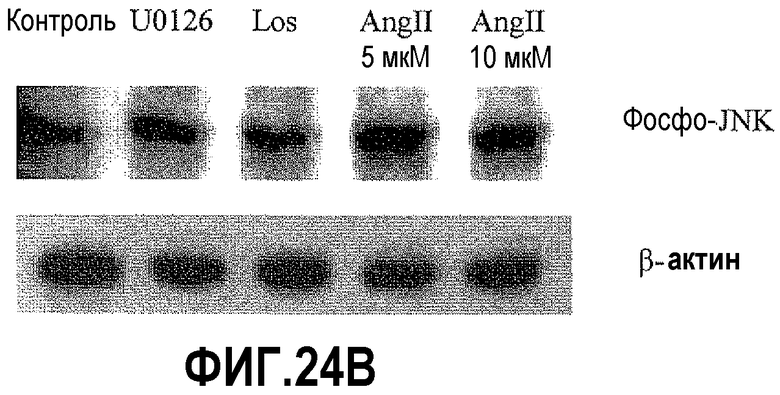
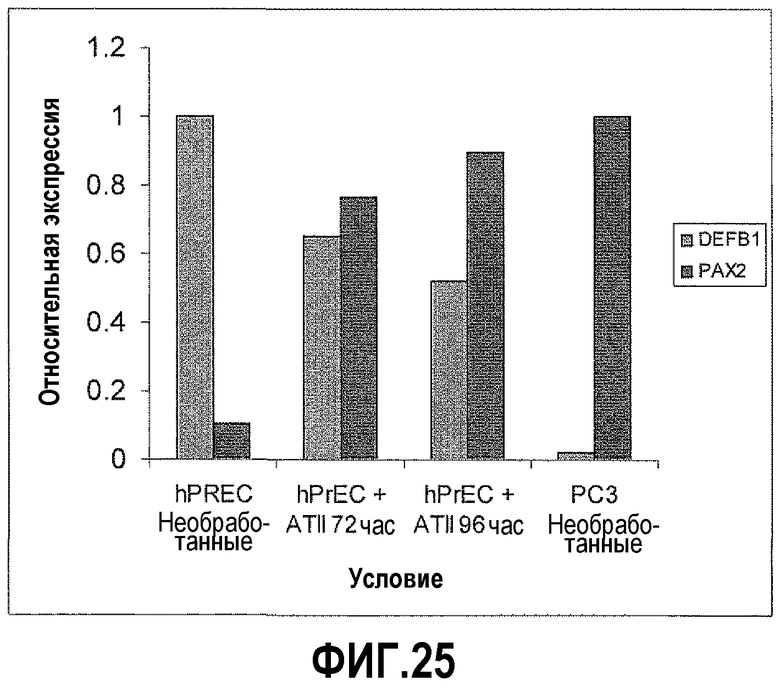
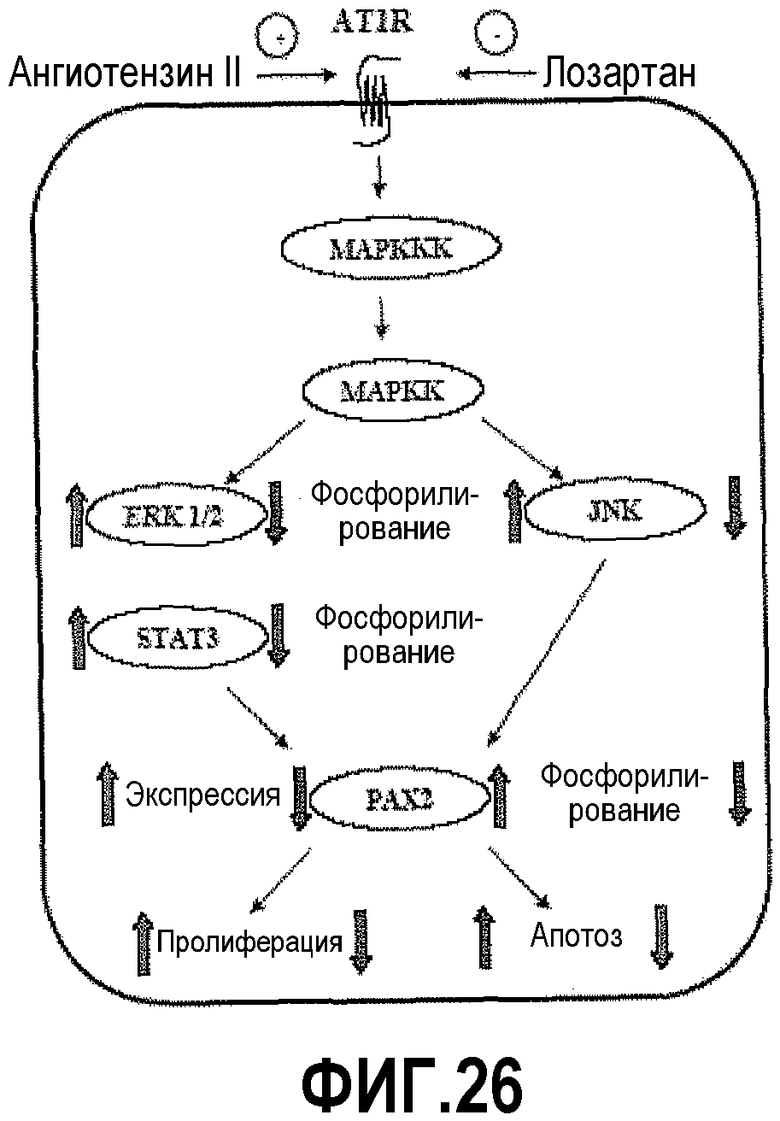
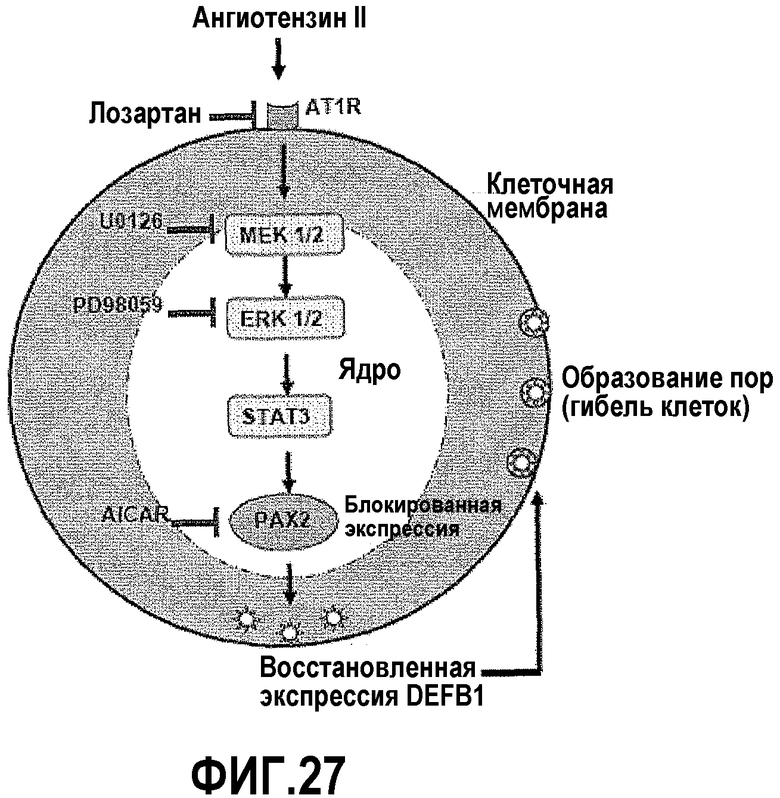
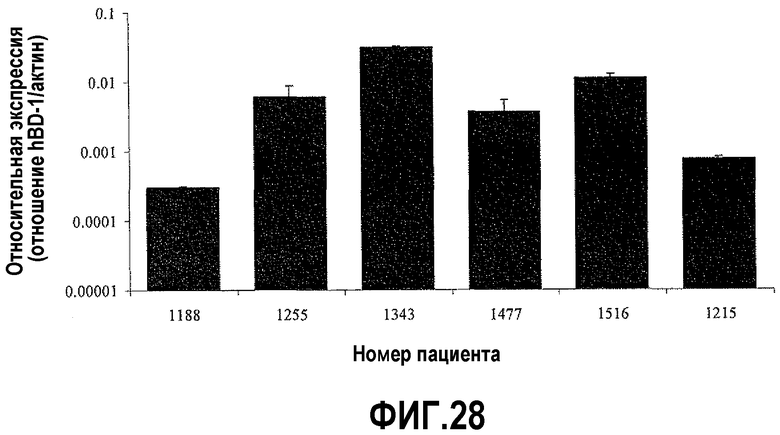
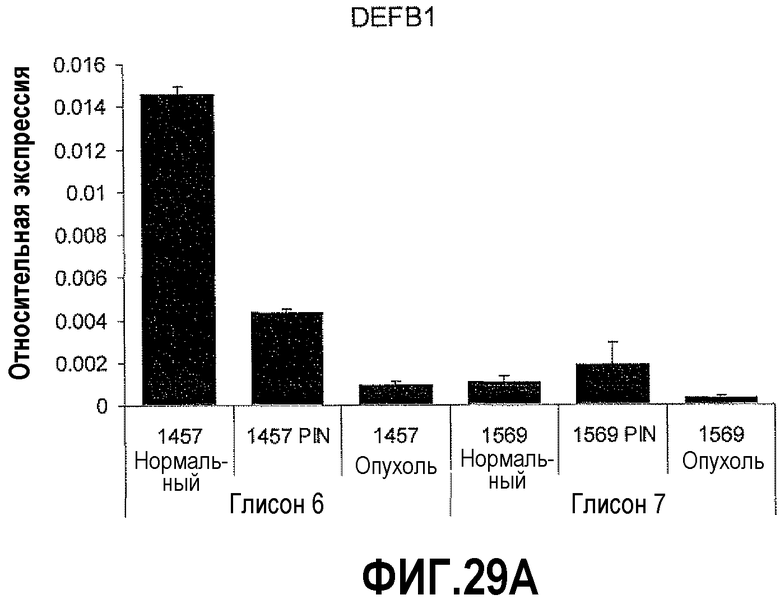
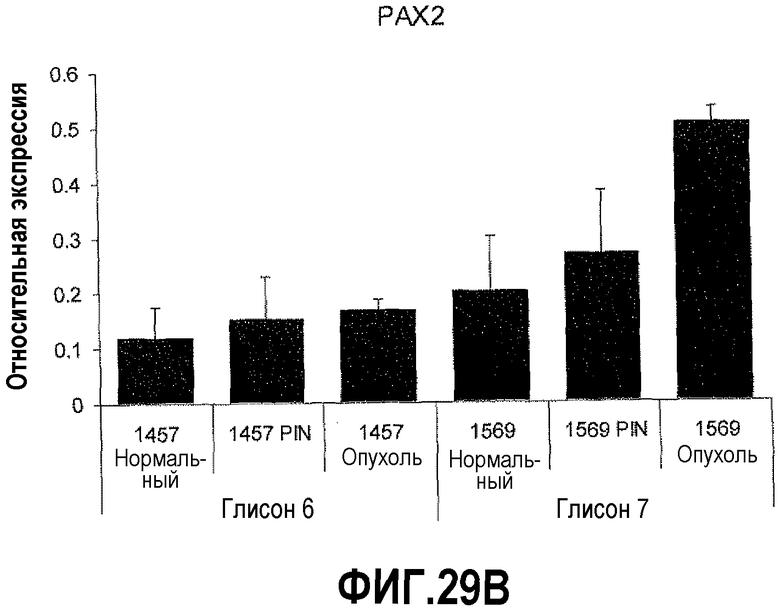
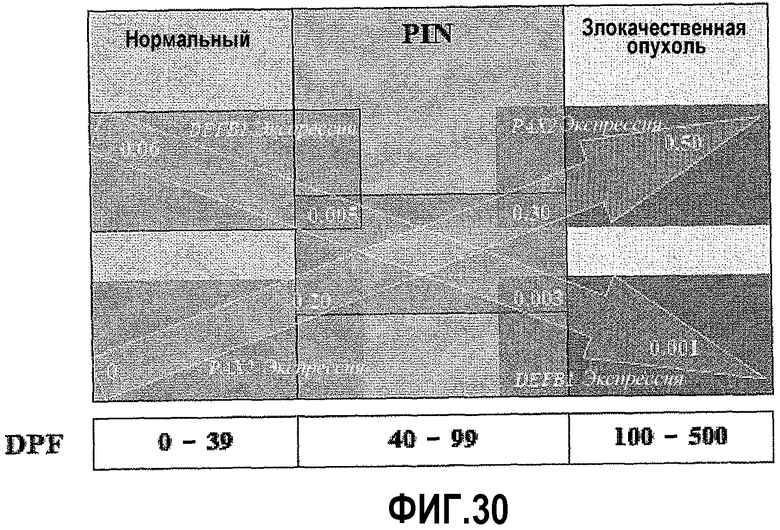
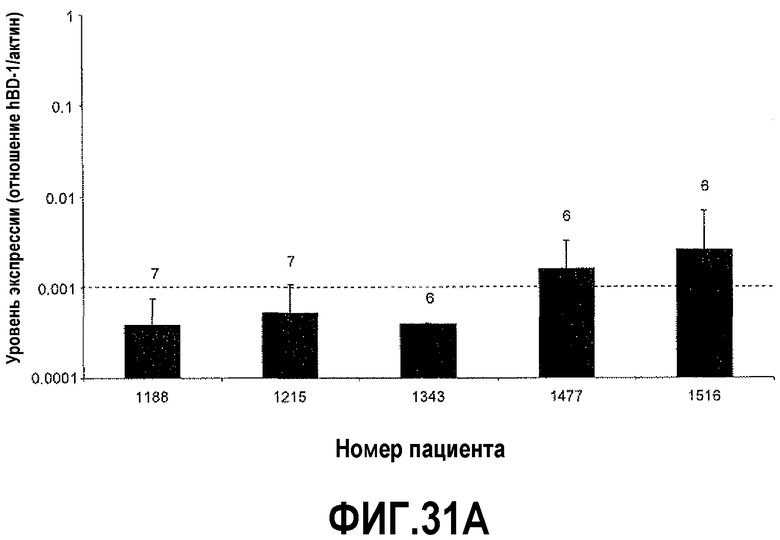
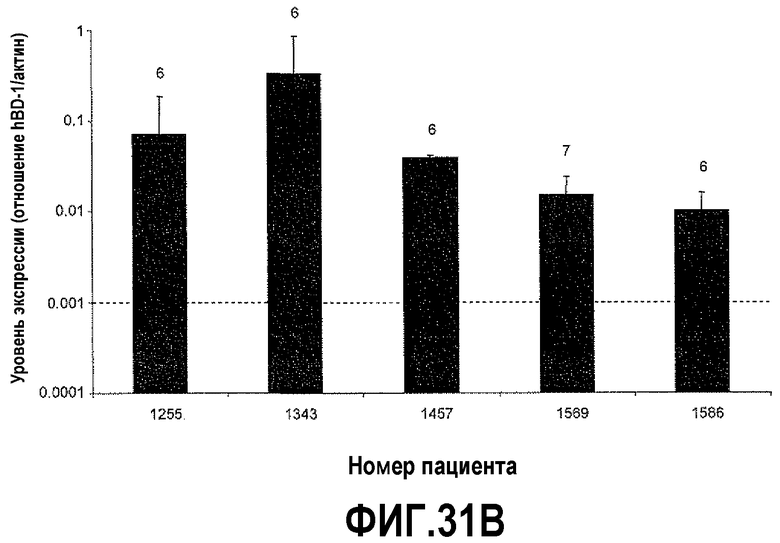
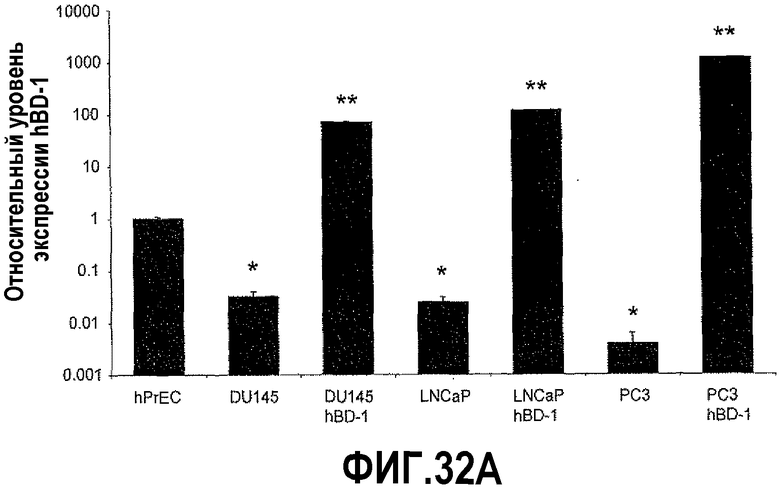
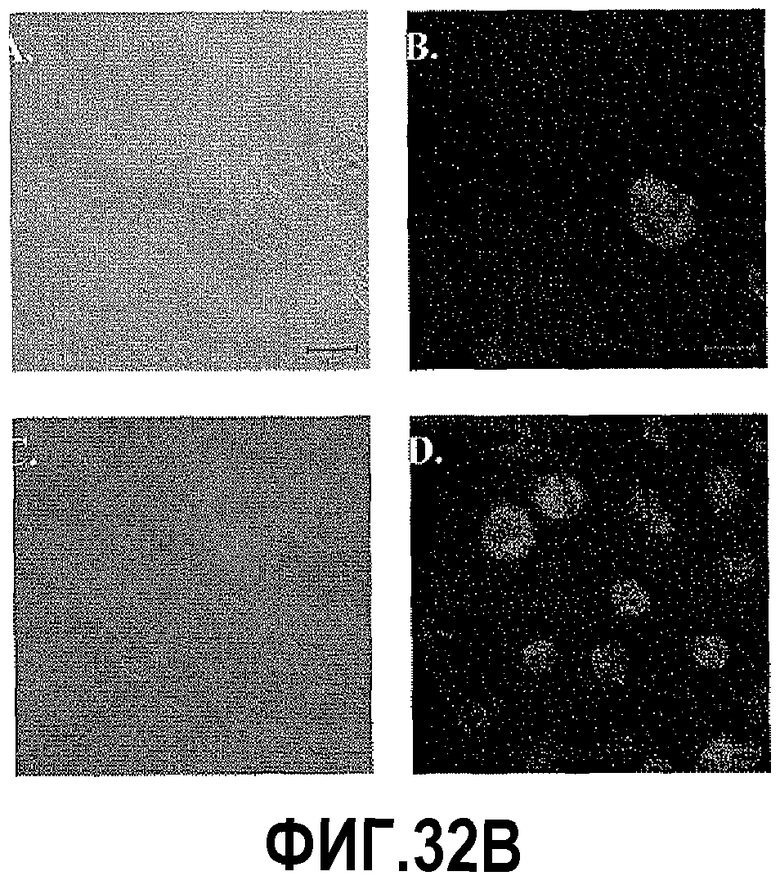
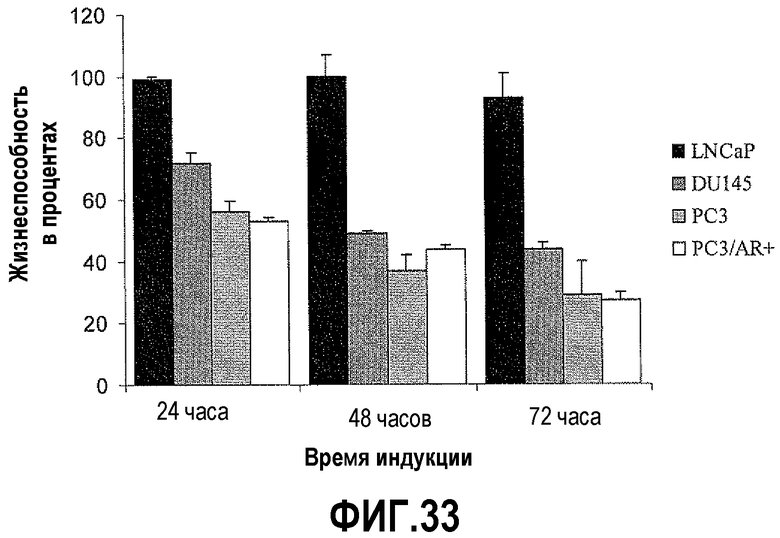
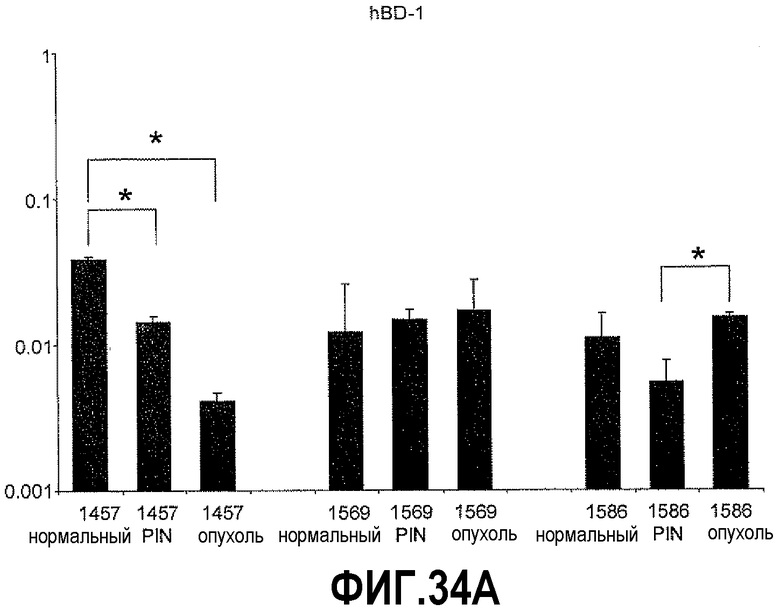
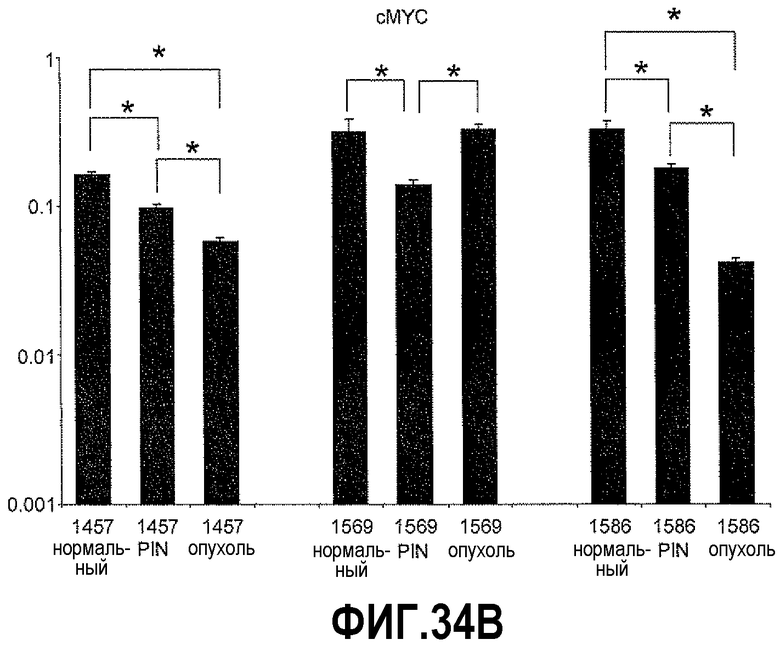
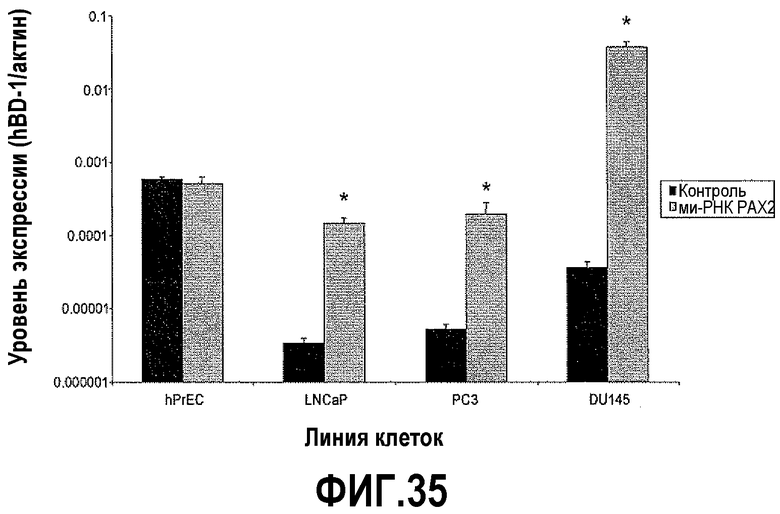
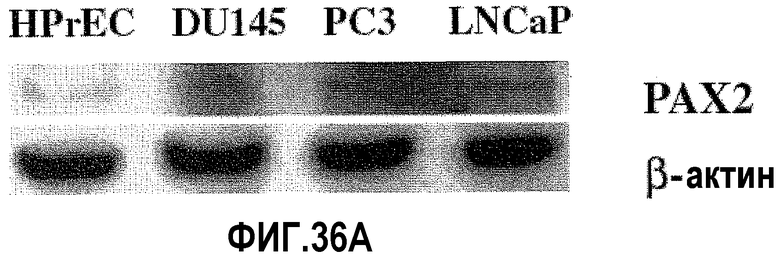
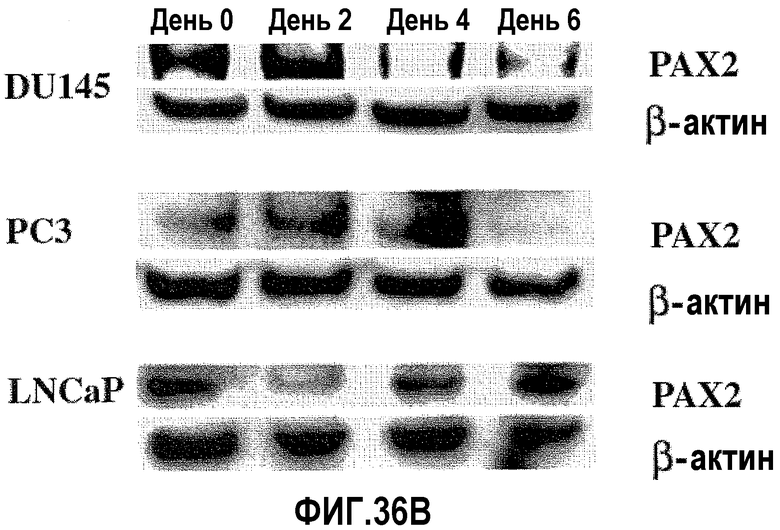
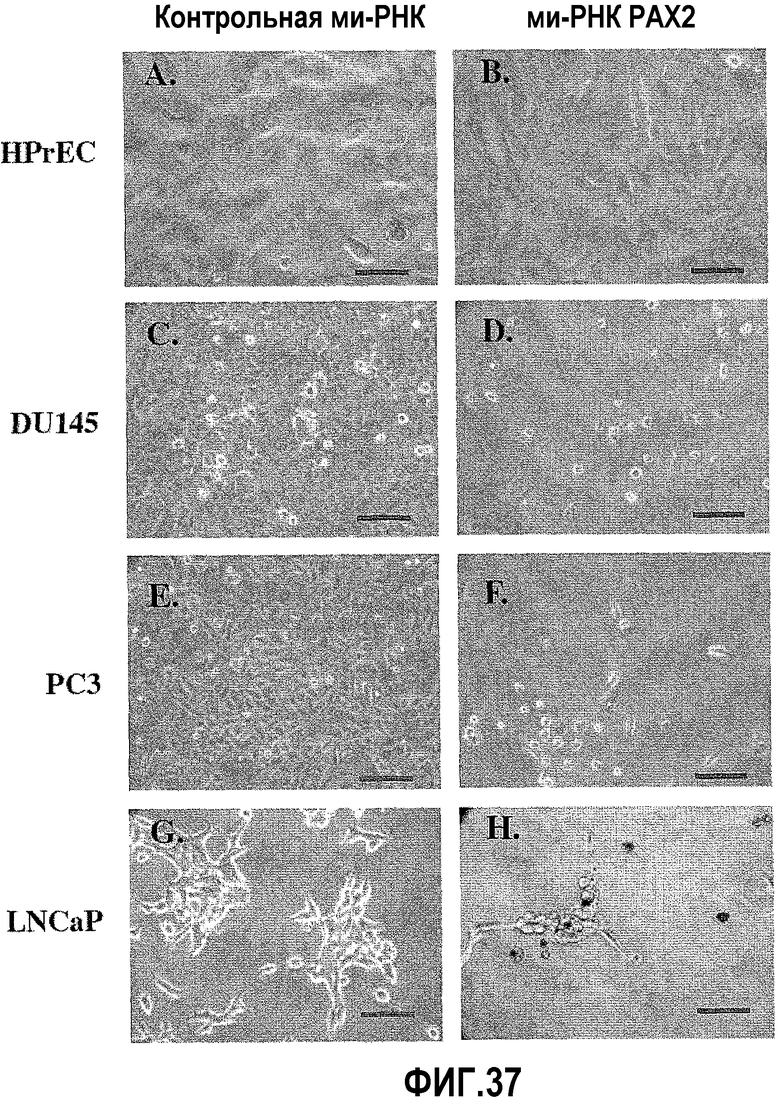
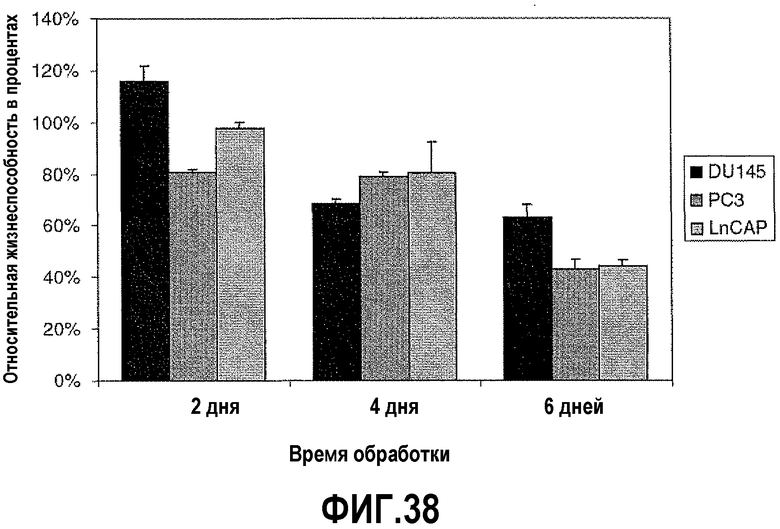
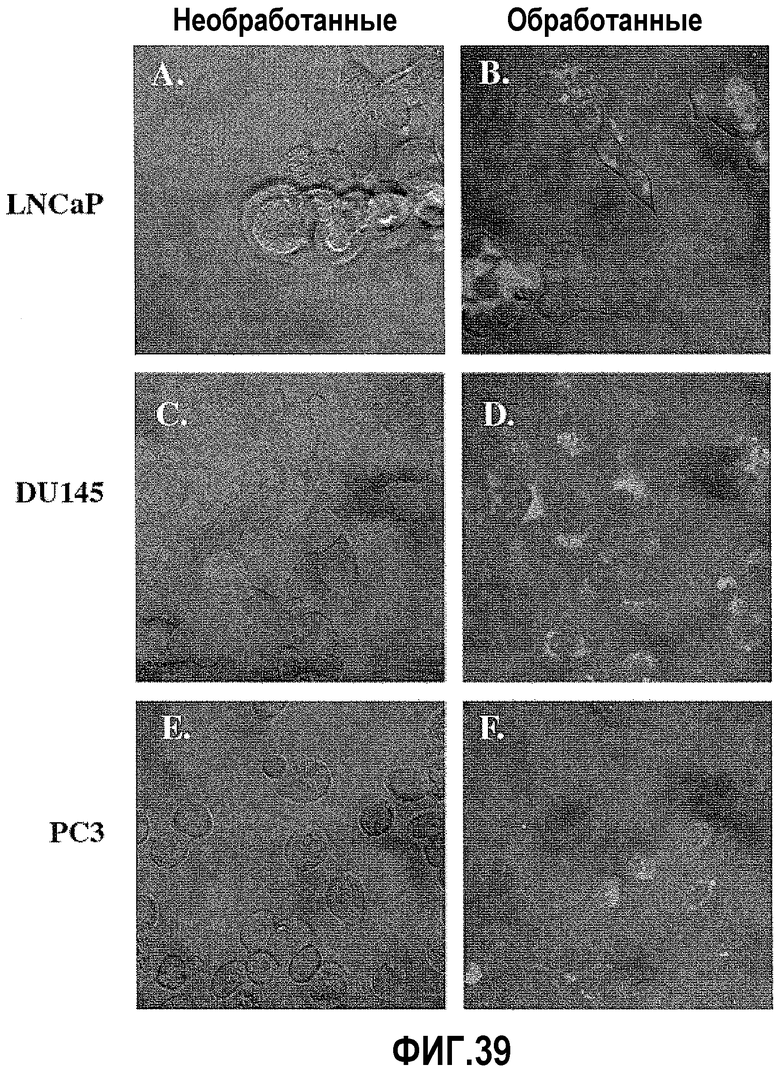
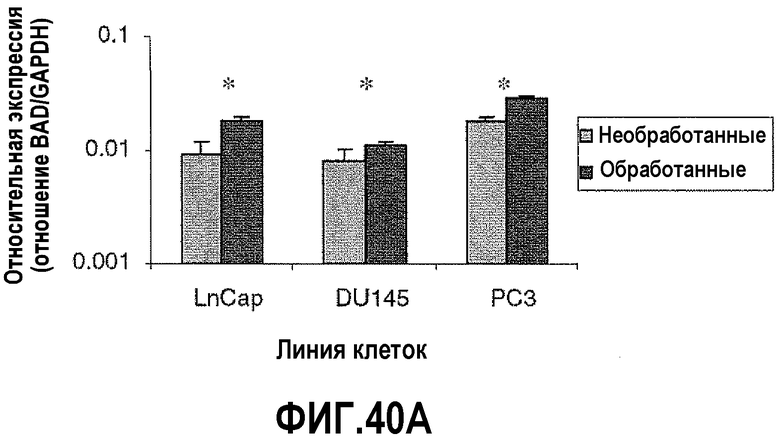
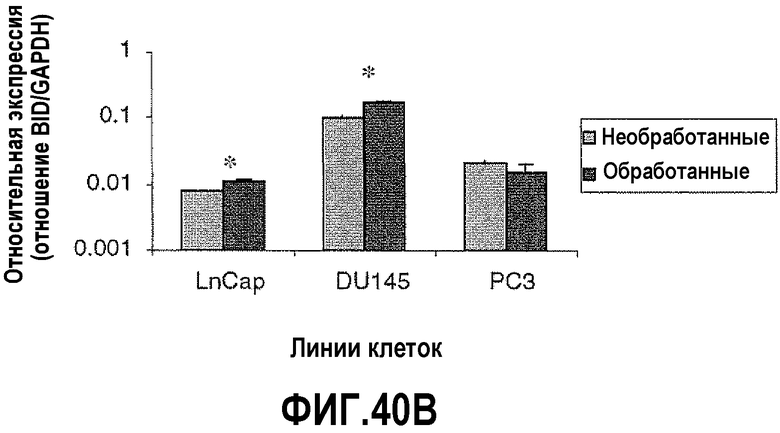
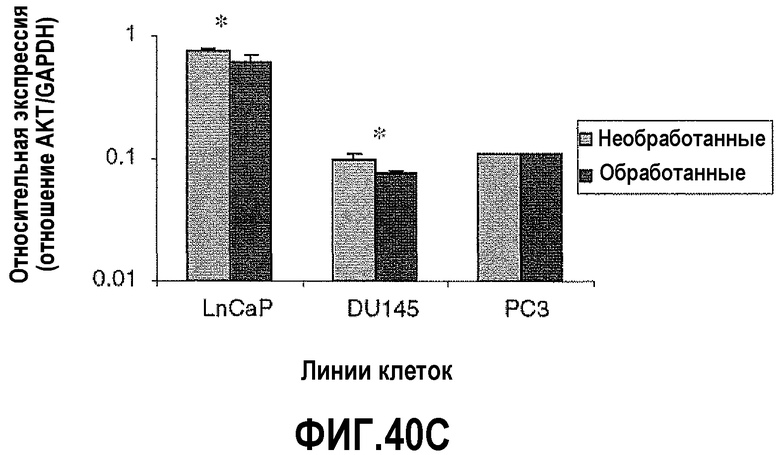
Изобретение относится к области медицины, генетической инженерии и молекулярной биологии. Предложен способ мониторинга рака молочной железы у субъекта. Способ включает определение отношения экспрессии гена парного бокса 2 к экспрессии гена бета-дефензина-1 (PAX2-к-DEFB1) в клетках, полученных из молочной железы субъекта, при этом отношение экспрессии PAX2-к-DEFB1 коррелирует с состояниями молочной железы. Также предложен набор для мониторинга рака молочной железы. Изобретение может быть использовано в медицине при лечении и мониторинге рака. 3 н. и 7 з. п. ф-лы, 40 ил., 8 табл., 17 пр.
1. Способ мониторинга рака молочной железы у субъекта, включающий:
определение отношения экспрессии гена парного бокса 2 (PAX2) к уровню экспрессии гена бета-дефензина-1 (DEFB1) для получения отношения экспрессии (PAX2-к-DEFB1) в клетках, полученных из молочной железы субъекта,
в котором отношение экспрессии PAX2-к-DEFB1 100:1 или выше является показателем наличия рака молочной железы у субъекта, и отношение экспрессии PAX2-к-DEFB1 менее чем 100:1 является показателем наличия незлокачественного или предзлокачественного состояния молочной железы у субъекта.
2. Способ по п.1, в котором предзлокачественным состоянием молочной железы является внутриэпителиальная неоплазия молочной железы.
3. Способ по п.1, в котором стадия определения дополнительно включает:
определение уровня экспрессии гена PAX2 по сравнению с уровнем экспрессии контрольного гена для получения нормального уровня экспрессии PAX2;
определение уровня экспрессии гена DEFB1 по сравнению с уровнем экспрессии того же контрольного гена для получения нормального уровня экспрессии DEFB1; и
определение отношения экспрессии PAX2-к-DEFB1 на основе уровней экспрессии PAX2 и DEFB1.
4. Способ по п.3, в котором уровни экспрессии генов PAX2, DEFB1 и контрольного гена определяют количественной ОТ-ПЦР в реальном времени и/или с использованием микроматрицы для определения экспрессии.
5. Способ по п.3, в котором контрольным геном является ген глицеральдегид-3-фосфатдегидрогеназы (GAPDH).
6. Способ по п.1, дополнительно включающий определение статуса рецептора эстрогена/рецептора прогестерона/рецептора эпидермального фактора роста человека 2 (ER/PR/HER2) в клетках, полученных из ткани молочной железы.
7. Набор для мониторинга рака молочной железы у субъекта, содержащий:
одну или несколько пар олигонуклеотидных праймеров для выявления экспрессии PAX2 в образце ткани;
одну или несколько пар олигонуклеотидных праймеров для выявления экспрессии DEFB1 в образце ткани; и
инструкции о том, как определить отношение экспрессии PAX2-к-DEFB1 в образце ткани с использованием праймеров,
в котором одна или несколько пар олигонуклеотидных праймеров для выявления экспрессии PAX2, содержат пару олигонуклеотидных праймеров, выбранную из группы, состоящей из последовательностей SEQ ID NO: 43 и 47, SEQ ID NO: 44 и 48 и SEQ ID NO: 45 и 49, и/или
в котором одна или несколько пар олигонуклеотидных праймеров для выявления экспрессии DEFB1 содержат последовательности SEQ ID NO: 35 и 37, и/или
в котором набор дополнительно содержит одну или несколько пар контрольных олигонуклеотидных праймеров.
8. Набор по п.7, в котором одна или несколько пар контрольных олигонуклеотидных праймеров содержат олигонуклеотидные праймеры для выявления экспрессии β-актина, при этом необязательно
олигонуклеотидные праймеры для выявления экспрессии β-актина содержат последовательности SEQ ID NO: 34 и 36, и/или
в котором одна или несколько пар контрольных олигонуклеотидных праймеров содержат олигонуклеотидные праймеры для выявления экспрессии GAPDH, при этом необязательно
олигонуклеотидные праймеры для выявления экспрессии GAPDH содержат последовательности SEQ ID NO: 42 и 46.
9. Набор по п.7, дополнительно содержащий один или несколько реагентов для ПЦР-реакции и/или для экстракции РНК.
10. Набор для мониторинга рака молочной железы у субъекта, содержащий:
микроматрицу олигонуклеотидов, имеющую олигонуклеотидные зонды для выявления экспрессии PAX2 и DEFB1;
инструкции о том, как определить отношение экспрессии PAX2-к-DEFB1 в образце ткани с использованием микроматрицы олигонуклеотидов; и необязательно
дополнительно содержащий реагенты для экстракции РНК из образца ткани.
| СПОСОБ ДИАГНОСТИКИ НА ОСНОВЕ РАСТВОРА | 2003 |
|
RU2315312C2 |
| US 20070099209 A1, 03.05.2007 | |||
| СПОСОБ ДИАГНОСТИКИ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2340896C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ТЕРАПИИ И ДИАГНОСТИКИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2001 |
|
RU2344831C2 |

