Область техники
Изобретение относится к области биотехнологии, генной и белковой инженерии, к нанотехнолическим применениям с использованием жгутиков архей и может быть использовано в построении устройств для диагностики, элементов аккумуляторов, в медицине и в фармацевтической промышленности.
Уровень техники
В настоящее время большие усилия направлены на поиск новых наноматериалов, которые могут быть полезны при разработках широкого спектра устройств, активность работы которых зависит от площади контакта активных зон размещенных на поверхности устройств.
К таким устройствам можно отнести сенсорные датчики [1], компоненты для создания элементов электроники, аккумуляторов, дисплеев [2], покрытие биологических и медицинских устройств, требующих отсутствия агрессивных химических компонентов [3], к таким средствам можно также отнести компоненты покрытий или композиций в составе фармакологических препаратов [4].
Известны изобретения, в которых в качестве элементов покрытий используют биологические материалы, созданные на основе пептидов [4] или вирусов [2, 5-9]. С целью формирования развитой поверхностной структуры используют вирусы, на поверхности которых иммобилизуют как частицы неорганических веществ, так и ионы металлов [5-10]. В практике создания поверхностей с развитой поверхностной структурой особое внимание уделяется вариантам, которые включают белковые структуры, включающие аминокислоты, способные нековалентно связывать ионы металлов [5-16].
Для того чтобы наноструктурированное покрытие удерживалось на поверхности разрабатываемых устройств, используют клеевые соединения [3]. Однако введение клеевых соединений не всегда возможно особенно в медико-биологических применениях. Поэтому наиболее оптимально использовать материалы, которые сами обладают адгезивными свойствами. Выбор биологических материалов для создания наноструктур с развитой поверхностью, обеспечивающих работу и сохранение биологически активных зон, является непростой задачей.
Известны изобретения, в которых для создания развитых поверхностей используются наноматериалы, выполненные на основе вируса М13 [5-10]. Однако вирусные структуры не сохраняют свои свойства в растворах с высокой температурой [5-10].
Известно, что жгутики архей сохраняют свою структуру при высоких температурах [17]. Кроме того, известно, что материалы, выполненные на основе жгутиков, обладают достаточными адгезивными свойствами и могут быть использованы для создания молекулярных клеев. В изобретении [3] рассматриваются вопросы получения клеевых материалов, содержащих, по меньшей мере, один белок, полученный из флагеллинов архей. В заявке показано, что флагеллины архей хорошо прилипают не только к органическим материалам, например к клеточной стенке, но также и к поверхностям, выполненным из неорганических материалов, таких как металлы, пластмассы и кварц. В изобретении указано, что не все флагеллины обладают такой возможностью, так, для H.Salinarum в качестве перечисленных в заявке флагеллинов фигурируют только FlaB1 и FlaB2, которые являются не основными белками жгутика, а выполняют связующую роль между клеточным мотором и протяженной спиральной нитью жгутика, состоящей из А-флагеллинов [17]. Основанием для такого интереса служат научная информация о адгезивной роли некоторых флагеллинов в составе жгутика и последовавший за этим поиск гомологичных последовательностей флагеллинов среди всех архей с известными геномными последовательностями. В качестве основного материала молекулярного клея авторы используют отдельные белки флагеллинов, из которых формируют материал клея. В заявке на изобретение уделяется большое внимание методам выделения целевых флагеллинов из жгутиков, что предполагает необратимое разделение жгутиков на компоненты и разрушение их структуры.
Однако в данном изобретении [3] не раскрыт механизм высокой адгезии флагеллинов к широкому классу неорганических веществ и не раскрыты места на поверхности флагеллинов, которые выполняют роль молекулярного клея. В то же время авторы полагают, что в структурах рассматриваемых флагеллинов могут быть введены замены до 60 аминокислот, причем место и состав замен и вставок, не указаны ни для одного типа флагеллина. Для выяснения возможности таких замен требуется проведение дополнительных исследований свойств флагеллинов, полученных генно-инженерным способом. Указанное изобретение не подтверждает возможность и эффективность применения генно-инженерных флагеллинов для эффективной нековалентной связи на его поверхности ионов металлов.
Целью настоящего изобретения является расширение типов покрытий с развитой поверхностной структурой на основе наноматериалов, на основе которых можно обеспечить нековалентное связывание ионов металлов, наночастиц металлов, полупроводников, с целью формирования поверхности сенсоров, пластин аккумуляторов, нетканых материалов с новыми свойствами.
Другая цель изобретения состоит в получении технического результата, предотвращающего воздействие высоких температур на сохранность поверхностной структуры наноматериала и его адгезивных свойств.
Следующей целью изобретения является повышение эффективности связывания ионов металлов, наночастиц металлов, полупроводников по сравнению с материалами, полученными из архей дикого типа.
Указанные технические результаты обеспечиваются за счет использования в качестве наноматериала рекомбинантных жгутиков флагеллина, причем, по меньшей мере, в один белок флагеллина вводят, по меньшей мере, одну пептидную вставку, аминокислотный состав которой позволяет обеспечить эффективную нековалентную связь с соответствующим, по меньшей мере, одним типом иона металла, наночастицы металла, полупроводника.
Сущность изобретения
Объектами изобретения являются рекомбинантные белки флагеллина А1 и флагеллина А2, используемые для формирования разных типов жгутиков археи Halobacterium salinarum с повышенной активностью связывания ионов металлов, наночастиц металлов, полупроводников, по сравнению с их аналогами дикого типа. Для повышения активности связывания ионов металлов, наночастиц металлов, полупроводников в последовательность флагеллинов дикого типа A1 (SEQ ID NO: 2) и A2 (SEQ ID NO: 3) включают, по меньшей мере, одну пептидную вставку в область между первым и вторым сайтом гликозилирования. Данные области включают последовательность SEQ ID NO: 6 флагеллина А1 и последовательность SEQ ID NO:7 флагеллина А2.
Другими объектами изобретения являются последовательности пептидной вставки, которая может содержать одиночную последовательность длиной от 6 до 60 аминокислот, или вставка содержит короткие последовательно соединенные друг с другом последовательности с общей длиной вставки до 60 аминокислот, при этом количество коротких последовательностей лежит в пределах от 2 до 10.
Последовательность пептидной вставки выбирают таким образом, чтобы она нековалентно связывала ионы металлов, выбираемых из группы, в которую входят: кобальт, ванадий, никель, марганец, железо, кадмий, вольфрам, хром, цирконий, титан, скандий, иттрий, медь, кальций, алюминий, барий, бериллий, магний и стронций, и/или наночастицы металлов, выбираемых из группы, состоящей из золота, платины, серебра, палладия, или наночастицы, обладающие полупроводниковыми свойствами: например ZnS, CdS, а также наночастицы, обладающие магнитными свойствами, например FePt, CoPt.
Следующими объектами изобретения являются плазмидные ДНК на основе плазмиды pNXA для экспрессии разных типов рекомбинантных жгутиков археи Н.Salinarum с повышенной эффективностью связывания ионов металлов, наночастиц металлов, полупроводников, по сравнению с их аналогами дикого типа. В состав плазмид входят А-флагеллиновый оперон Н.Salinarum, содержащий последовательно размещенные области А1 и А2 генов флагеллина по сайтам клонирования XbaI и HindIII, ген устойчивости к ампициллину AmpR, ген устойчивости к новобиоцину NovR, ген галофильной b-галактозидазы bgaH. При этом в зависимости от типа рекомбинантного жгутика плазмиды кодируют: а) рекомбинантный белок флагеллина А1 с пептидной вставкой и рекомбинантный белок, кодирующий флагеллин А2 дикого типа (SEQ ID NO: 3), б) рекомбинантный белок флагеллина А2 с пептидной вставкой и рекомбинантный белок, кодирующий флагеллин А1 дикого типа (SEQ ID NO: 2), в) рекомбинантный белок флагеллина А1 с пептидной вставкой и рекомбинантный белок флагеллина А2 с пептидной вставкой.
Следующими вариантами объектов изобретения являются штаммы на основе Н.Salinarum R1 в качестве продуцентов разных типов рекомбинантных жгутиков археи, содержащих устойчивость к антибиотикам: клетки, полученных путем трансформации клеток Н.Salinarum R1 рекомбинантными плазмидами ДНК pNX, кодирующими а) рекомбинантный белок флагеллина А1 с пептидной вставкой и рекомбинантный белок, кодирующий флагеллин А2 дикого типа (SEQ ID NO: 3), б) рекомбинантный белок флагеллина А2 с пептидной вставкой и рекомбинантный белок, кодирующий флагеллин А1 дикого типа (SEQ ID NO: 2), в) рекомбинантный белок флагеллина А1 с пептидной вставкой и рекомбинантный белок флагеллина А2 с пептидной вставкой.
Следующими вариантами объектов изобретения являются способы получения наноструктурированного материала на основе рекомбинантных жгутиков археи. Указанные способы обеспечивают следующие этапы: а) синтез рекомбинантных жгутиков архей путем культивирования штамма-продуцента, б) выделение жгутиков архей, содержащих пептидные вставки для нековалентного связывания, в) модификацию поверхности жгутиков посредством связывания пептидных вставок с ионами металлов, наночастицами металлов или полупроводников, г) оксидирование ионов металлов, д) промывку, сушку и упаковку полученного наноструктурированного материала.
Следующими вариантами объектов изобретения являются композиции из наноструктурированного материала на основе жгутиков архей, которые выполнены в виде раствора или лиофилизированной формы и включают в свой состав, по меньшей мере, один тип рекомбинантного жгутика, состоящего из а) рекомбинантного белка флагеллина А1 с пептидной вставкой и рекомбинантного белка, кодирующего флагеллин А2 дикого типа (SEQ ID NO: 3), б) рекомбинантного белка флагеллина А2 с пептидной вставкой и рекомбинантного белка, кодирующего флагеллин А1 дикого типа (SEQ ID NO: 2), в) рекомбинантного белка флагеллина А1 с пептидной вставкой и рекомбинантного белка флагеллина А2 с пептидной вставкой. При этом поверхности рекомбинантных жгутиков модифицированы и композиции дополнительно содержат приемлемые добавки для формирования покровного материала. При этом композиция выполнена в виде раствора или лиофилизированной формы и включает приемлемые добавки для формирования покровного материала.
Следующими вариантами объектов изобретения являются покровные материалы для формирования наноструктурированного покрытия пластин, пленок, капсул, текстильного материала, содержащие, по меньшей мере, одну композицию, включающую, по меньшей мере, один тип модифицированного рекомбинантного жгутика, и распределенные, по меньшей мере, на части покровного материала.
Следующими вариантами объектов изобретения являются наборы для формирования изделий из жгутиков архей, содержащие, по меньшей мере, контейнер, композицию рекомбинантных жгутиков архей и упаковочный материал.
Перечень фигур
На Фиг.1 представлена физическая карта плазмиды pNXA.
На Фиг.2 представлена аминокислотная последовательность А1-флагеллина. Последовательности сайтов гликозилирования выделены цветом.
На Фиг.3 представлена аминокислотная последовательность А2-флагеллина. Последовательности сайтов гликозилирования выделены цветом.
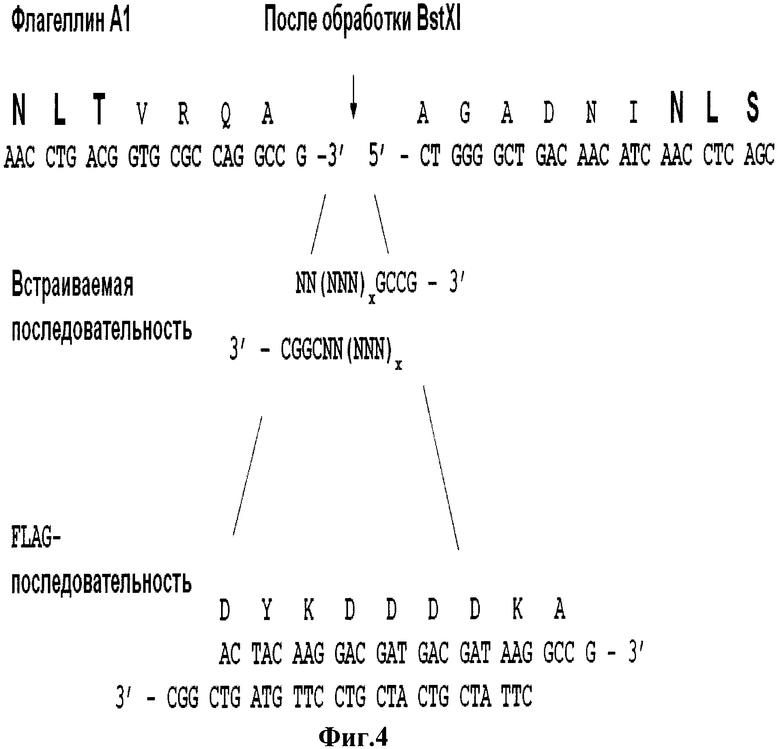
На Фиг.4 представлены нуклеотидная и приведенная в соответствие к ней аминокислотная последовательность А1-флагеллина в области вставок, а также последовательности самих вставок. Последовательности сайтов гликозилирования и последовательность сайта рестрикции BstXI выделены жирным шрифтом.
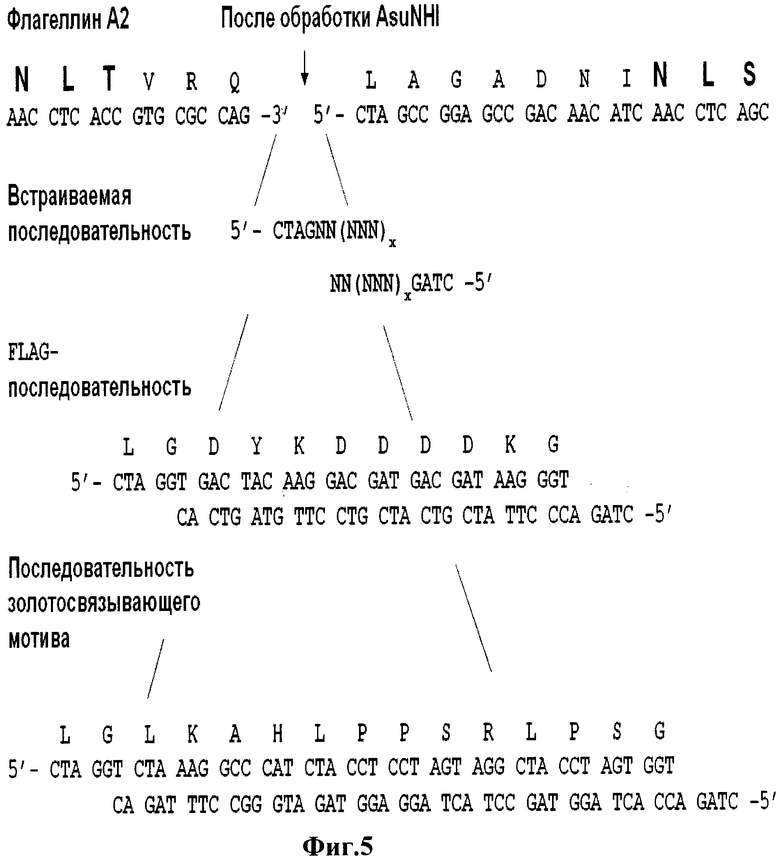
На Фиг.5 представлены нуклеотидная и приведенная в соответствие к ней аминокислотная последовательность А2-флагеллина в области вставок после введения сайта эндонуклеазы рестрикции AluNHI, а также последовательности самих вставок. Последовательности сайтов гликозилирования и последовательность сайта рестрикции AluNHI выделены жирным шрифтом.
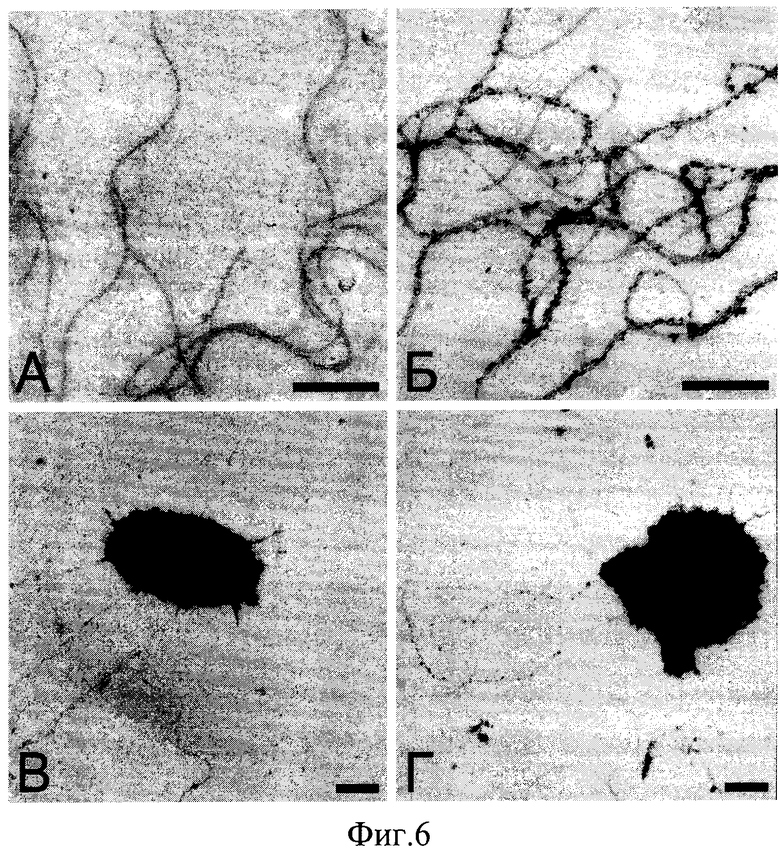
На Фиг.6. представлено электронно-микроскопическое изображение жгутиков Н.salinarum: (А) дикий тип, (Б) модифицированные жгутики со вставкой в A1-флагеллин FLAG-пептида; и клеток Н.salinarum: (В) дикий тип, (Г) FLAG-штамм. Препараты были мечены специфичными антителами и конъюгатами белка А с частицами коллоидного золота (10 нм). Негативное контрастирование уранил-ацетатом. Масштабная линейка - 300 нм.
На Фиг.7. представлено электронно-микроскопическое изображение жгутиков Н.salinarum: (А) дикий тип, (Б-В) модифицированные жгутики со вставкой в A1-флагеллин FLAG-пептида. Жгутики были минерализованы оксидом кобальта (А, Б) и оксидом меди (В). Негативное контрастирование проведено уранил-ацетатом. Масштабная линейка - 300 нм.
Описание
Исследование свойств протяженных жгутиков Н.Salinarum (диаметром 12 нм и длиной несколько микрометров) на основе флагеллинов А1 и А2 «дикого типа» (без генно-инженерных вставок) показало, что жгутики таких флагеллинов обладают адгезивными свойствами к поверхности металлов, пластмасс и других материалов. Однако такие жгутики обладают низкой эффективностью связывания на своей поверхности ионов металлов, что не позволяет использовать флагеллин дикого типа для создания наноматериалов с заданными свойствами. Для получения наноструктурированного материала с заданными свойствами полимерной структуре жгутика, состоящего из А1- и А2-флагеллинов, необходимо придать свойство специфично связывать ионы или наночастицы металлов. Для реализации подобной задачи необходимо выбрать в последовательностях флагеллинов участки, пригодные для включения в них пептидных вставок, с возможностью их последующей модификации ионами металлов. При этом должны быть сохранены структурные механизмы, обеспечивающие полимеризацию флагеллинов А1 и А2 в жгутик, а для проведения эффективной модификации вставок необходимо обеспечить выведение пептидных вставок на внешнюю поверхность сформированного жгутика. В результате получают комбинированный наноструктурированный материал волокнистой структуры, содержащий нити модифицированных жгутиков.
Известно, что флагеллины А1 и А2, формирующие состав жгутиков H.salinarum N, гликозилированы, при этом каждый флагеллин содержит до 3-х сайтов гликозилирования (последовательности NXT, NXS) [17-18].
В процессе изучения свойств жгутиков было обнаружено, что введение коротких пептидных вставок в участки, расположенные между сайтами гликозилирования флагеллинов, не влияет на формирование структуры жгутиков, а сами вставки при синтезе жгутиков остаются экспонированными на внешней поверхности жгутиков, что дает возможность для эффективного нековалентного связывания аминокислот, входящих в состав вставок с ионами и наночастицами метаоллов.
В общем случае способ получения рекомбинантных жгутиков архей состоит из следующих стадий?
A) Выделение ДНК, кодирующей флагелин.
Б) Выбор типа плазмиды и ее генетической конструкции.
B) Выбор последовательности, кодирующей пептид вставки.
Г) Выбор места в аминокислотной последовательности внутреннего фрагмента флагеллина для пептидной вставки, а также выбор эндонуклеазы рестрикции для разрезания последовательности в выбранном месте.
Д) Формирование плазмиды.
Е) Выбор клеток и трансформация клеток полученной плазмидой.
Ж) Выращивание клеток на поверхности агаризированной среды и последующий рост клеток в жидкой среде.
З) Выделение жгутиков из культуральной среды для модификации и последующей минерализации поверхности жгутиков.
И) Модификация жгутиков выбранным ионом металла/или модификация поверхности наночастицами металла и последующая минерализация поверхности жгутиков выбранным веществом.
К) Выделение, сушка и хранение минерализированных жгутиков, предназначенных для формирования материала.
Выделение ДНК, кодирующей флагелин.
Для выделения тотальной ДНК из H.salinarum, а также выделения плазмидной ДНК из Е. coli используют колонки со стеклянным наполнителем фирмы Qiagen (США). ДНК выделяют согласно инструкции фирмы-производителя. Тотальную ДНК H.salinarum используют в качестве матрицы для наработки ПЦР-фрагментов А-флагеллиного оперона с фланкирующими участками.
Выбор типа плазмиды и ее генетической конструкции.
Рекомбинантная плазмидная ДНК pNXA, представленная на Фиг.1, имеет размер молекулы около 11000 п.о. и содержит клонированный А-флагеллиновый оперон Н.Salinarum, содержащий области А1 и А2 генов флагеллина по сайтам клонирования XbaI и HindIII. Данная плазмида получена на основе плазмиды pNX [19] путем клонирования ПЦР фрагмента А-флагеллинового оперона Н.Salinarum, при этом длина клонированного фрагмента составляет около 2400 пар нуклеотидов (SEQ ID NO: 1). Плазмида содержит ген устойчивости к ампициллину (AmpR) и ген устойчивости к новобиоцину (NovR). В плазмиде также содержится ген галофильной b-галактозидазы (bgaH). Ген AmpR и бактериальный сайт инициации репликации позволяют наращивать данную плазмиду и ее модификации в клетках Е. coli. Ген NovR обеспечивает устойчивость трансформированных клеток архей к новобиоцину, продукт гена bgaH позволяет трансформированным клеткам приобретать синюю окраску продуктами разложения коммерческого субстрата X-Gal, что служит дополнительным маркером для подтверждения трансформации. В плазмиде отсутствует галоархейный сайт инициации репликации. Это приводит к тому, что при трансформации плазмидой устойчивость к новобиоцину клетки архей приобретают только в случае встраивания плазмиды в хромосому по механизму гомологичной рекомбинации. В конструкции плазмиды около 1200 п.о. в сумме составляют А1-ген флагеллина, представленный на Фиг.2 (SEQ ID NO: 2), и А2-ген флагеллина, представленный на Фиг.3 (SEQ ID NO: 3), а также левый (около 600 п.о) и правый (400 п.о.) фланкирующие участки А-флагеллинового оперона [26]. Кроме того гены А1 и А2 разделяет межгенный промежуток длиной 11 пар нуклеотидов. Фрагмент ДНК А-флагеллинового оперона наработан праймерами:
5'-GTCGGTGCTCTAGACGTCGGGGATCGG-3' (SEQ ID NO: 4)
5'-TCACGATGCAAAGCTTCCATCAGTACGG-3' (SEQ ID NO: 5)
с помощью ПЦР. Фрагмент клонирован в плазмиду по сайтам рестрикции HindIII и XbaI. В зависимости от вариантов генетической конструкции плазмиды в область А1-гена, либо А2-гена, либо одновременно в области А1 и А2 гена вводят последовательность, кодирующую пептидную вставку.
Выбор области в аминокислотной последовательности флагеллина для пептидной вставки.
Введение пептидных вставок осуществляют в области А1- и А2-флагеллинов, располагающиеся между их первыми двумя сайтами гликозилирования от NLT до NLS. Для A1-флагеллина последовательность области выглядит как VRQAAGADNI (SEQ ID NO: 6), а для А2-флагеллина данная последовательность выглядит как VRQAAGADNI (SEQ ID NO: 7). Допустимо введение вставки в любое место между данными сайтами гликозилирования, от поз. 86 до поз. 96 в области флагеллина А1 (Фиг.2) и между поз. 82 до поз. 92 в области флаггелина А2 (Фиг.3). Предпочтительно вводить вставку в середину области между поз. 89 до поз. 92 для флагеллина А1. Так на Фиг.2 представлена аминокислотная последовательность А1-флагеллина, а последовательности сайтов гликозилирования выделены цветом. Для флагеллина А2 предпочтительно вводить вставку в область между поз. 85 до поз. 88. На Фиг.3 представлена аминокислотная последовательность А2-флагеллина, а последовательности сайтов гликозилирования также выделены цветом. При этом введенная аминокислотная последовательность экспонируется на поверхность сформированного жгутика, что позволяет осуществить его последующую обработку веществами, обладающими сродством к использованной вставке.
Клонирование последовательностей, кодирующих пептидные вставки Клонирование последовательностей, кодирующих выбранный пептид, в области А1- и/или А2-гена осуществляется с использованием стандартных методик, включающих в себя: 1) выбор сайта рестрикции в области генов, соответствующих аминокислотным последовательностям фрагментов белков (SEQ ID NO: 6) и (SEQ ID NO: 7), 2) синтез парных олигонуклеотидов, один из которых кодирует пептидную вставку, а другой комплементарен ему для формирования дуплекса. Также данные олигонуклеотиды должны содержать липкие концы для выбранного сайта рестрикции, 3) Клонирование дуплекса по выбранному сайту. Последовательность пептидной вставки содержит одиночную последовательность длиной от 6 до 60 аминокислот или содержит короткие последовательно соединенные друг с другом последовательности с общей длиной вставки до 60 аминокислот, при этом количество коротких последовательностей лежит в пределах от 2 до 10.
Ниже приведен пример клонирования последовательностей, кодирующих выбранный пептид. Так, например, для А1-гена, подобное клонирование состояло из этапов: 1) выбрали эндонуклеазу рестрикции BstXI, которая разрезает А1-ген флагеллина в месте, соответствующем 90-91 аминокислоте белка, 2) синтезировали олигонуклеотидные последовательности SEQ ID NO: 21, SEQ ID NO: 22, 3) клонировали дуплекс олигонуклеотидов в плазмиду pNXA (Фиг.4). При этом все олигонуклеотидные последовательности, кодирующие выбранный пептид, можно схематически изобразить следующим образом: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 3'-конец для клонирования по сайту рестрикции BstXI: 5'-NN(NNN)xGCCG-3' (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту BstXI: 5'-(NNN)xNNCGGC-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в A1-ген приведена на Фиг.4. Число x определяет число кодонов, кодирующих аминокислоту и может достигать величины от 6 до 60 кодонов для одиночной вставки. Если возникает необходимость введения вставки от поз. 86 до поз. 96 в области флагеллина A1 и между поз. 82 до поз. 92 в области флаггелина А2, но нельзя подобрать подходящий сайт для эндонуклеазы рестрикции, то возможно в данной области изменение нуклеотидной и, соотвественно, аминокислотной последовательности. Подобная замена не приводит к утрате функциональности флагеллина и может при необходимости осуществляться как на А1-, так и на А2-генах флагеллинов, если содержит дополнительные замены аминокислот с сохранением, по меньшей мере, 80%, предпочтительно, по меньшей мере, 90%, более предпочтительно, по меньшей мере, 95%, даже более предпочтительно, по меньшей мере, 99% идентичности последовательности, при этом любая замена аминокислоты является предпочтительно консервативной.
Для А2-гена, подобное клонирование состояло из этапов: 1) проведена замена .в нуклеотидной последовательности А2-гена с составе плазмиды pNXA синтетическими праймерами SEQ ID NO: 23, SEQ ID NO: 24 с использованием QuikChange site-directed mutagenesis kit фирмы Stratagen (поскольку не удалось подобрать подходящей эндонуклеазы рестрикции). Эта замена привела к появлению сайта рестрикции AsuNHI в гене А2, а в соответствующей гену аминокислотной последовательности произошла замена в поз.86 аланина на лейцин SEQ ID NO: 8, 2) синтезированы олигонуклеотидные последовательности, например, SEQ ID NO: 25, SEQ ID NO: 26, 3) дуплекс олигонуклеотидов клонирован в плазмиду pNXA (Фиг.5) При этом все олигонуклеотидные последовательности, кодирующие выбранный пептид, можно схематически изобразить следующим образом: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 5'-конец для клонирования по сайту рестрикции AsuNHI: 5'-CTAGNN(NNN)x-3' (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту AsuNHI: 5'-CTAG(NNN)xNN-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в А2-ген приведена на Фиг.5. Число x определяет число кодонов, кодирующих аминокислоту и может достигать величины от 6 до 60 кодонов для одиночной вставки.
Получение плазмид с тандемно клонированными вставками перечисленных последовательностей и получение штаммов продуцентов на их основе возможно приведет к усилению или улучшению отдельных характеристик получаемого наноматериала.
Если вставка тандемно повторяется, то величина x должна уменьшиться кратно числу повторяющихся вставок.
Выбор последовательности, кодирующей пептид вставки.
Известны пептидные последовательности, которые обладают способностью связывать определенные лиганды [5-13, 15, 16]. В качестве одного из примеров, подтверждающего воспроизводимость и работоспособность способа получения наноструктурированного материала на основе жгутиков архей, создан штамм галофильного археона H. salinarum, синтезирующий жгутики, модифицированные FLAG-пептидом DYKDDDDK (SEQ ID NO: 9). Данный пример (см. пример 1) включает, но не ограничивает других вариантов жгутиков, модифицированных пептидными последовательностями, имеющими высокое сродство к различным лигандам.
Подобным образом могут быть получены модифицированные и другими экспонированными на поверхности вставками флагеллины архей с сохранением протяженной надмолекулярной структуры жгутиков. В свете имеющихся данных целесообразным кажется получение жгутиков Н.Salinarum с модифицированным А1- и/или А2-флагеллином следующими вариантами пептидных последовательностей, которые связывают заряженные ионы, оксиды и наночастицы металлов, фосфат железа, связывают лиганды с полупроводниковыми и магнитными свойствами.
Последовательность тетраглутамина ЕЕЕЕ (SEQ ID NO: 10) связывает положительно заряженные ионы металлов. Жгутики, модифицированные данной последовательностью, могут быть использованы для получения материала для электродов литий-ионных аккумуляторов [5-6]. Кроме того, показано, что данный тетрапептид способен связывать и фосфат железа, также являющийся перспективным материалом для катодов литий-ионных аккумуляторов [20].
Для некоторых вариантов покрытий, использующих наноструктурированные материалы, могут быть использованы жгутики архей с пептидными вставками, связывающими коллоидные наночастицы благородных металлов, или жгутики архей, содержащие два типа вставок, одна из них связывает ионы металла, другая наночастицы благородных металлов. В качестве примера вставки, связывающей наночастицы золота с поверхностью жгутика, служит так называемый золотосвязывающий пептидный мотив LKAHLPPSRLPS (SEQ ID NO: 11) [5-6], который включает, но не ограничивает типы пептидных вставок для связывания наночастиц на поверхности жгутика археи.
Возможно совместное применение двух разных типов пептидных вставок (SEQ ID NO: 11) и (SEQ ID NO: 10), которые, например, совместно экспрессировались для получения бифункциональных частиц модифицированного вируса М13 [5-6].
В качестве пептидной вставки могут быть использованы пептидные последовательности, которые могут связывать лиганды, обеспечивающие полупроводниковые и магнитные свойства. Так пептид с последовательностью CNNPMHQNC (SEQ ID NO: 12) связывает ZnS, пептид с последовательностью SLTPLTTSHLRS (SEQ ID NO: 13) связывает CdS, пептид с последовательностью HNKHLPSTQPLA (SEQ ID NO: 14) связывает FePt, пептид с последовательностью CNAGDHANC (SEQ ID NO: 15) связывает CoPt [10].
Известно, что тетрапептиды RRRR(SEQ ID NO: 16) и RKRK (SEQ ID NO: 17) связывают ионы и оксиды металлов. Пары обладают избирательным сродством к Cu2O и ZnO соответственно [21].
Пептидные структуры, образующие петли, обладают свойствами связывать ионы металлов, и, в зависимости от состава, имеют анионные или катионные свойства. К таким петлям относятся гистидиновые петли "His-loop" (GHHHHHH) (SEQ ID NO: 18), глутамин-аспарагиновые петли "Glu-Asp loop" (DQDQDQG) (SEQ ID NO: 19), аргнинин-лизиновые петли "Arg-Lys loop" (RKRKRKR) (SEQ ID NO: 20) [11].
Варианты плазмид
Для проверки эффективности изобретения на основе плазмиды pNXA были получены следующие варианты плазмид: 1) рекомбинантная плазмидная ДНК pNXA1FLAG, обеспечивающая синтез модифицированного FLAG-пептидом А1-флагеллина и немодифицированного А2-флагеллина, 2) рекомбинантная плазмидная ДНК pNXA1A2FLAG, обеспечивающая синтез модифицированных FLAG-пептидом А1- и А2-флагеллинов, 3) рекомбинантная плазмидная ДНК pNXA1FLAGA2GBP, обеспечивающая синтез модифицированного FLAG-пептидом A1-флагеллина и модифицированного золотосвязывающим пептидом А2-флагеллина в клетках Н. Salinarum R1 при трансформации и встраивании данной плазмиды в хромосому организма. Варианты плазмид и способы их получения приведены в примере 2.
Все перечисленные плазмидные ДНК были получены путем однократной вставки каждой последовательности в указанные места. Однако данные последовательности также могут быть встроены тандемно 2 и более раз, что приведет к удлинению вставленного кодируемого пептида кратно его длине и может привести к синтезу флагеллинов, отличных по свойствам не только по сравнению с нативным, но и перечисленными модификациями флагеллинов.
Выбор клеток и трансформация клеток полученной плазмидой
Полученными плазмидами трансформируют клетки Н.salinarum и выращивают их на поверхности агаризованной среды с антибиотиком. Выросшие колонии клеток пересевают в жидкую среду, и после роста из клеток либо выделяют жгутики, либо отбирают небольшие количества клеток.
Для получения штаммов-продуцентов модифицированных флагеллинов трансформируют клетки Н.Salinarum по приводимой методике [22]. Н.salinarum - это экстремальный галофильный грамнегативный археон с облигатной аэробностью. Естественной средой обитания данного организма является вода Мертвого моря, а оптимальной для роста является экстремально высокая соленость (до 5.5 М NaCl). В лабораторных условиях выращивается на среде следующего состава: 25% NaCl, 0.2% KCl, 2% Mg2SO4 7×H2O, 0.3% Na-цитрат, 0.5% триптон, 0.2% дрожжевой экстракт, рН 7.5 при 37°С [23].
Н.salinarum представляет собой одиночные клетки палочковидной формы, не образующие спор, имеющие лофотрихиальное расположение жгутиков, на стационарной фазе у клеток Н.Salinarum возможна амфитрихиальная локализация. Во всех случаях нити жгутиков образуют пучок, вращение которого создает гидродинамическое усилие, приводящее клетку в движение. В пучок входит 5-10 нитей, представляющих собой полужесткую спираль. При росте Н.salinarum в культуральной среде обнаруживается значительное количество свободных жгутиков.
Взвеси клеток в жидких питальных средах, либо колонии, выросшие на поверхности агаризованной среды, обладают красным цветом за счет присутствия в клеточной стенке бактородопсина.
В России доступен штамм Н.salinarum R1, предоставляемый Всероссийской коллекцией промышленных микроорганизмов (г.Москва). Все полученные штаммы-продуценты на основе Н.Salinarum R1 характеризуются следующими признаками:
- морфологические признаки: клетки палочковидной формы, грамотрицательные, неспороносные,
- культуральные признаки: при росте на агаризованной среде для Н.Salinarum колонии круглые, гладкие, блестящие, край ровный, окрашены в красный цвет. При росте на жидких средах образуют интенсивную ровную муть,
- физико-биологические признаки: клетки растут при температуре от 20°С до 45°С при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы,
- устойчивость к антибиотикам: клетки проявляют устойчивость к новобиоцину (до 0.6 мкг/мл), обусловленную наличием в плазмиде pNXA гена NovR,
- штаммы-продуценты отличаются от штамма-реципиента Н.Salinarum R1 только наличием различных вариантов рекомбинантной плазмидной ДНК pNXA, которая придает ему устойчивость к новобиоцину,
- отсутствие галоархейного сайта инициации репликации приводит к тому, что при трансформации плазмидой устойчивость к новобиоцину клетки архей приобретают только в случае встраивания плазмиды в хромосому по механизму гомологичной рекомбинации,
- штаммы-продуценты помимо встроенного с плазмидой модифицированного А-флагеллинового оперона также содержат в себе нативный А-оперон. Следовательно, в составе жгутиков помимо модифицированных флагеллинов (его наличие показано экспериментально иммуно-электронной микроскопией) могут содержаться и немодифицированные флагеллины.
Следующие этапы получения рекомбинантных жгутиков, такие как а) выращивание клеток на поверхности агаризированной среды и последующий рост клеток в жидкой среде, б) выделение жгутиков из культуральной среды для модификации и последующей минерализации поверхности жгутиков, г) модификация жгутиков выбранным ионом металла и/или модификация поверхности наночастицами металла или полупроводника и затем последующая минерализация поверхности жгутиков выбранным веществом, д) выделение, сушка и хранение минерализированных жгутиков, предназначенных для формирования материала, - приведены в примерах 4-6.
Из приведенных примеров модифицированных жгутиков жгутики со вставкой FLAG-пептида (SEQ ID NO: 9) позволяют детектировать его присутствие на поверхности с помощью специфичных моноклональных антител. С помощью иммуно-электронной микроскопии как выделенного препарата жгутиков, так и фиксированных клеток FLAG-штамма Н.salinarum было показано, что данный пептид присутствует в составе флагеллина, а жгутики специфично метятся антителами. Видно, что модифицированные жгутики сохраняют нативную спиральную структуру и морфологически не отличаются от жгутиков дикого типа, при этом FLAG-пептид доступен для связывания с антителами, то есть находится в составе поверхностного участка полипетидной цепи A1-флагеллина (Фиг.6). Известно, что выведенные на поверхность белков тетраглутаминовые или тетрааспарагиновые последовательности способны эффективно связывать ионы металлов (например, Со2+, Cu2+) [5-13]. Так как в составе FLAG-пептида также имеется 4 последовательных отрицательно заряженных аминокислотных остатка - аспартата, жгутики были проверены на связывание металлических ионов. Петля, образованная восемью остатками, оказалась достаточной для выведения кластера заряженных аминокислот на поверхность жгутика и способной связывать ионы металлов. Затем ионы в присутствии боргидрида натрия переводились в нерастворимые частицы оксидов, прикрепленные к жгутику (Фиг.7). Образовавшиеся структуры напоминают наноструктурированные материалы, полученные другими исследователями на основе вирусных частиц и бактериальных жгутиков. Считается, что подобные материалы, в зависимости от типа связанного лиганда, могут быть использованы в качестве материала для электродов литий-ионных аккумуляторов, различных наносенсоров и катализаторов, получения материалов с магнитными и полупроводниковыми свойствами [3-16].
Примеры
Пример 1. Получение генетических конструкций
Генетические конструкции получают на основе плазмиды pNX [19], содержащей ген устойчивости к ампициллину и ген устойчивости к новобиоцину. Для выделения тотальной ДНК из H.salinarum, а также выделения плазмидной ДНК из Е. coli используют колонки со стеклянным наполнителем фирмы Qiagen (США). ДНК выделяют согласно инструкции фирмы-производителя. Тотальную ДНК Н.salinarum используют в качестве матрицы для наработки ПЦР-фрагментов А-флагеллиного оперона с фланкирующими участками.
С использованием олигонуклеотидных праймеров SEQ ID NO: 4 и SEQ ID NO:5 проводят ПЦР в 50 мкл смеси, состоящей из ПЦР буфера (для Taq ДНК-полимеразы: 57 мМ Трис-HCl, рН 8.8, 16.6 мМ (NH)2SO4, 0.1% Tween-20) с концентрацией MgCl2 от 1.5 до 2.5 мМ, смеси дезоксирибонуклеотидов (в концентрации 0.2 мМ каждого), олигонуклеотидов-праймеров по 20-100 пМ каждого, 2-20 нг плазмидной или 100-200 нг хромосомной ДНК и смеси из 1-2 ед. Taq ДНК-полимеразы и 0.1-0.2 ед. Pfu ДНК-полимеразы. Длительность стадий плавления, отжига и элонгации составляет 10 с, 30 с и 3 мин соответственно. Температуру отжига праймеров определяют по максимальному выходу продуктов, стадию плавления проводят при 95°С, а элонгацию при 72°С, количество циклов - 25.
ПЦР-продукты, а также плазмидную ДНК анализируют методом ДНК-электрофореза в агарозном геле в ТАЕ-буфере согласно [24]. Обработку ДНК эндонуклезами рестрикции (все фирмы Fermentas, Литва) и постановку лигазной реакции с использованием Т4 ДНК-лигазы (Fermentas, Литва) проводят согласно инструкции производителя.
Пример 2. Конструкции плазмид с разными типами пептидных вставок
В качестве клонируемой последовательности использованы различные дуплексы синтетических олигонуклеотидов: (SEQ ID NO: 21), (SEQ ID NO: 22); (SEQ ID NO: 25). (SEQ ID NO: 26); (SEQ ID NO: 27), (SEQ ID NO: 28). Все последовательности в соответствии с комплементарностью попарно отжигают друг на друга.
А) Конструкция плазмиды pNXA1FLAG
В качестве примера, который включает, но не ограничивает другие варианты, в данном примере рассмотрен способ введения вставки, которая кодирует последовательность FLAG-пептида. С этой целью используют праймер с последовательностью SEQ ID NO: 21 и праймер с комплементарной последовательностью SEQ ID NO: 22.
Праймеры отжигают друг на друга с охлаждением их смеси с 80°С до комнатной температуры, после чего проводят их фосфорилирование по 3'-концам полинуклеотидкиназой (Fermentas, Литва) согласно инструкции. Далее проводят клонирование данной синтетической двухцепочечной олигонуклеотидной последовательности, кодирующей FLAG-пептид, путем вставки в А1-ген флагеллина в месте, соответствующем 90-91 аминокислоте белка по сайту рестрикции BstXI, за счет липких концов для данного сайта (Фиг.4), образующегося при отжиге синтетических одноцепочечных олигонуклеотидов (SEQ ID NO: 21) и (SEQ ID NO: 22). Полученной лигазной смесью трансформируют клетки Е. coli согласно [25] и высевают на чашки с агаризованной (1.5%) средой LB с 100 мкг/мл ампициллина. Для наработки плазмидной ДНК выращенную колонию с чашки пересевают в жидкую среду с антибиотиком и после подроста клеток выделяют плазмиду. Полученные плазмиды секвенируют на ДНК-секвенаторе ABI PRIZM 310 (Applied Biosystems, США).
Таким образом, в приведенном примере для клонирования по сайту рестрикции BstXI в A1-генфлагеллина олигонуклеотидов, кодирующих другие вставки, они должны выглядеть следующим образом: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 3'-конец: 5'-NN(NNN)xGCCG-3' (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту BstXI: 5'-(NNN)xNNCGGC-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в А1-ген приведена на Фиг.4. Число x определяет число кодонов, кодирующих аминокислоту и может достигать величины от 6 до 60 кодонов для одиночной вставки.
Б) Конструкция плазмиды pNXA1A2FLAG
Плазмида pNXA1A2FLAG была получена в два этапа: во-первых, созданы условия для вставки в область А2-флагеллинового гена различных последовательностей. Так как сайта рестрикции для BstXI в А2-гене не имеется, а также не удалось подобрать других уникальных сайтов в выбранной области между сайтами гликозилирования, было решено ввести искусственный сайт путем замены нескольких нуклеотидов. Такая замена была осуществлена в плазмиде pNXA с использованием синтетических олигунуклеотидов
5'-CACCGTGCGCCAGCTAGCCGGAGCC-3' (SEQ ID NO: 23) и
5'-GGCTCCGGCTAGCTGGCGCACGGTG-3' (SEQ ID NO: 24)
и набора реактивов QuikChange site-directed mutagenesis kit (фирмы Stratagene). Таким образом, был введен сайт для эндонуклеазы рестрикции AluNHI, приводящей к замене в продукте А2-гена 86 аминокислоты аланина на лейцин. В результате фрагмент флагеллина А2, ограниченный первыми двумя сайтами гликозилирования, после замены выглядит как VRQLAGADNI (SEQ ID NO: 8). Подобная замена не приводит к утрате функциональности флагеллина и может при необходимости осуществляться как на А1-, так и на А2-генах флагеллинов. Количество замен в области из 10 аминокислот между первыми двумя сайтами гликозилирования как в А1, так и А2 флагеллинах может достигать 20% процентов с сохранением, по меньшей мере, 80%, предпочтительно, по меньшей мере, 90%, более предпочтительно, по меньшей мере, 95%, даже более предпочтительно, по меньшей мере, 99% идентичности последовательности, при этом любая замена аминокислоты является предпочтительно консервативной заменой. Далее, в А1-ген в месте, соответствующем 90-91 аминокислоте, и А2-ген в месте, соответствующем 86-87 аминокислоте каждого белка, вставили по синтетической двухцепочечной олигонуклеотидной последовательности, кодирующей FLAG-пептид. Данная последовательность встроена в А1-ген флагеллина по сайту рестрикции BstXI за счет липких концов для данного сайта, образующегося при отжиге синтетических одноцепочечных олигонуклеотидов (SEQ ID NO: 21) и (SEQ ID NO: 22), а в А2-ген флагеллина по сайту рестрикции AluNHI также за счет липких концов для данного сайта, образующегося при отжиге синтетических одноцепочечных олигонуклеотидов (Фиг.4-5).
5'-CTAGGTGACTACAAGGACGATGACGATAAGGGT-3' (SEQ ID NO: 25)
5'-CTAGACCCTTATCGTCATCGTCCTTGTAGTCAC-3' (SEQ ID NO: 26)
Таким образом, в приведенном примере, для клонирования по сайту рестрикции BstXI в A1-генфлагеллина олигонуклеотидов, кодирующих другие вставки, они должны выглядеть следующим образом: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 3'-конец: 5'-NN(NNN)xGCCG-3' (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту BstXI: 5'-(NNN)xNNCGGC-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в А1-ген приведена на Фиг.4. Для вставки в А2-ген: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 5'-конец для клонирования по сайту рестрикции AsuNHI: 5'-CTAGNN(NNN)x-3' (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту AsuNHI: 5'-CTAG(NNN)xNN-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в А2-ген приведена на Фиг.5. Число x определяет число кодонов, кодирующих аминокислоту, и может достигать величины от 6 до 60 кодонов для одиночной вставки.
В) Конструкция плазмиды pNXA1FLAGA2GBP
Плазмида pNXA1FLAGA2GBP также была получена в два этапа: во-первых, аналогично получению плазмиды pNXA1A2FLAG в область А2-флагеллинового гена был введен сайт для эндонуклеазы рестрикции AluNHI, приводящей к замене в продукте А2-гена 86 аминокислоты аланина на лейцин. Далее, в А1-ген в месте, соответствующем 90-91 аминокислоте белка, вставили синтетическую двухцепочечную олигонуклеотидную последовательность, кодирующую FLAG-пептид, а в А2-ген в месте, соответствующем 86-87 аминокислоте белка, синтетическую двухцепочечную олигонуклеотидную последовательность (SEQ ID NO: 11), кодирующую пептид, связывающую наночастицы коллоидного золота (Golden Binding Peptide). Последовательность золотосвязывающего пептидного мотива (SEQ ID NO: 11) определена методом фагового дисплея. Последовательность, кодирующая FLAG-пептид, встроена в А1-ген флагеллина по сайту рестрикции BstXI за счет липких концов для данного сайта, образующегося при отжиге двух синтетических одноцепочечных олигонуклеотидов (SEQ ID NO: 21)и (SEQ ID NO: 22). Последовательность (SEQ ID NO: 11), кодирующую пептид, вставили в А2-ген флагеллина по сайту рестрикции AluNHI также за счет липких концов для данного сайта, образующегося при отжиге синтетических одноцепочечных олигонуклеотидов (Фиг.4-5):
5'-CTAGGTCTAAAGGCCCATCTACCTCCTAGTAGGCTACCTAGTGGT-3'
(SEQ ID NO: 27)
5'-CAGATTTCCGGGTAGATGGAGGATCATCCGATGGATCACCAGATC-3'
(SEQ ID NO: 28).
Таким образом, в приведенном примере, для клонирования по сайту рестрикции BstXI в A1-ген флагеллина олигонуклеотидов, кодирующих другие вставки, они должны выглядеть следующим образом: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 3'-конец: 5'-NN(NNN)xGCCG-3' (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту BstXI: 5'-(NNN)xNNCGGC-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в А1-ген приведена на Фиг.4. Для вставки в А2-ген: первый олигонуклеотид кодирует последовательность вставляемого пептида и также содержит липкий 5'-конец для клонирования по сайту рестрикции AsuNHI: 5'-CTAGNN(NNN)x-3 (где N - стандартное обозначение нуклеотида). Второй олигонуклеотид комплементарен первому для формирования дуплекса и, кроме того, содержит липкий конец для клонирования по сайту AsuNHI: 5'-CTAG(NNN)xNN-3' (где N - стандартное обозначение нуклеотида). Схема способа введения вставки в А2-ген приведена на Фиг.5. Число x определяет число кодонов, кодирующих аминокислоту, и может достигать величины от 6 до 60 кодонов для одиночной вставки.
Пример 3. Получение штаммов продуцентов модифицированных жгутиков Н.salinarum R1
Использован штамм Н.salinarum R1, предоставленный Всероссийской коллекцией промышленных микроорганизмов (г.Москва). Клетки Н.salinarum выращивают на среде следующего состава: 25% NaCl, 0.2% КС1, 2% Mg2S04 7x H20. 0.3% Na-цитрат, 0.5% триптон, 0.2% дрожжевой экстракт, рН 7.5 при 37°С [23].
Штамм получают путем трансформации клеток Н.salinarum с использованием полиэтиленгликоля (ПЭГ) по описанной ранее методике [22] со следующими модификациями. Сферопласты получают отдельно для каждой аликвоты (1.5 мл) клеточной культуры. Клетки осаждают в 1.5 мл пробирках центрифугированием при 4000 g в течение 10 мин и затем ресуспендируют в 150 мкл сферопластирующего раствора. Для получения сферопластов к клеточной суспензии добавляют 15 мкл 0.5 М ЭДТА и выдерживают смесь 10 мин при комнатной температуре. Затем добавляют 10 мкл раствора, содержащего около 5 мкг плазмидной ДНК, и инкубируют смесь в течение 5 мин, после чего добавляют равный объем раствора, содержащего 60% ПЭГ-600 и 40% сферопластирующего раствора (вес/объем). Клетки инкубируют в течение 20 мин при комнатной температуре. Для восстановления клеток к суспензии добавляют 15 мл культуральной среды, содержащей 15% сахарозы, и инкубируют в течение 12-24 часов при 37°С с покачиванием. Селекция трансформантов осуществляют на агаризованной среде, содержащей 15% сахарозы и 0.1-0.2 мкг/мл антибиотика новобиоцина (Sigma, США). Инкубацию осуществляют при 37°С в течение 10-14 сут. Культуры клеток Н.salinarum хранят на чашках Петри на соответствующих средах с 1.5% агаром при при комнатной температуре.
Полученные штаммы культивируются аналогично нетрансформированным клеткам Н.Salinarum R1 за исключением добавки 0.1-0.2 мкг/мл антибиотика новобиоцина
Пример 4. Выделение и очистка жгутиков
Для выделения жгутиков культуру Н.salinarum выращивают в литровых колбах с 200 мл среды до поздней стационарной фазы. Полученную биомассу осаждают центрифугированием при 8000 g в течение 30 мин. К супернатанту, полученному после осаждения клеток, добавляют 4% ПЭГ-6000 и затем осаждают жгутики центрифугированием при 15000 g в течение 45 мин. Далее, для очистки жгутиков используют центрифугирование в градиенте плотности CsCl (12 часов при 55000 об/мин, ротор VTI-80, Beckman, США). Хлористый цезий растворяют в солевой среде до получения плотности 1.36 г/см3. Фракция, содержащая жгутики, образовывала видимую зону в средней части пробирки. Содержимое фракции разводят (в 10 раз) солевой средой, содержащей 25% NaCl, 20 мМ MgSO4, и, далее, жгутики переосаждают центрифугированием при 80000 g в течение 1 часа.
Пример 5. Связывание ионов металлов и минерализация жгутиков.
Связывание ионов металлов проводят аналогично методу, описанному Белчер [5-6] с модификациями. Жгутики при концентрации 0.1 мг/мл выдерживают в течение часа в водном растворе 1.5 М NaCl, содержащем 5 мМ хлорида Co или Cu. Затем к трем объемам раствора жгутиков добавляют один объем раствора, содержащего 1.5 М NaCl и 100 мМ NaBH4, и инкубируют в течение 1 часа до прекращения реакции.
Пример 6. Получение наноструктурированного материала на основе минерализованных жгутиков с модификациями.
После минерализации модифицированных жгутиков (пример. 4) они переходят в легко осаждаемый (4000g, 10 минут) осадок черного цвета (цвет оксидов Co и Cu). Данный осадок отделяется от супернатанта, затем суспендируется в воде и заново осаждается для отмывки от соли. Полученный осадок переносится на инертную подложку, сушится под ваккумом при комнатной температуре в течение суток. Полученный сухой порошок черного цвета хранится при комнатной температуре длительное время.
Пример 7. Получение электронных микрофотографий и исследование жгутиков с помощью иммуно-электронной микроскопии
Для получения электронных микрофотографий использовали микроскоп JEM-100 с «JEOL» (Япония). Для приготовления электронно-микроскопических образцов препараты наносят на электронно-микроскопические медные сетки с формваровой пленкой-подложкой, выдерживают 1 мин и отбирают раствор фильтровальной бумагой, после чего помещают сетку на раствор 2% уранилацетата, выдерживают 30 с, отбирают раствор фильтровальной бумагой и далее высушивают.
Образцы жгутиков сразу наносят на электронно-микроскопические медные сетки с формваровой пленкой-подложкой и выдерживают 5 мин. Для приготовления ИЭМ-препаратов клетки Н.salinarum подвергают предварительной фиксации. Для этого клетки осаждают при 4000 g в течение 5 мин и затем суспендируют в 15% растворе NaCl, к которому добавляют глутаровый альдегид до концентрации 1%, и оставляют на 20 мин. Затем суспензию клеток помещают на электронно-микроскопические медные сетки с формваровой пленкой-подложкой и выдерживают 5 мин, после чего сетки помещают на раствор 0.7% глицина в 5% NaCl для связывания избыточного альдегида. Сетки с образцами жгутиков или фиксированных клеток помещают на блокирующий буфер для ИЭМ (10 мМ Трис-HCl, рН 8, 150 мМ NaCl, 1% БСА) на 30 мин, а затем переносят на тот же буфер с препаратом моноклональных ANTI FLAG М2-антител (Sigma, США) и выдерживают 30 мин. Антитела разводят в 500, 200 и 100 раз. Затем сетки отмывают от антител в блокирующем буфере для ИЭМ 3-4 раза по 1 мин и переносят на тот же буфер с добавлением конъюгатов белка А с частицами коллоидного золота (размер частиц 10 нм, Jannsen Biotech, Бельгия) на 45-60 мин. Конъюгаты разводят в 50, 20 и 10 раз. Далее сетки отмывают блокирующим буфером для ИЭМ без БСА 2-3 раза по 5-10 мин, отбирают раствор фильтровальной бумагой и помещают на раствор 2% уранилацетата для негативного контрастирования. Через 30 с раствор 2% уранилацетата отбирают фильтровальной бумагой, сетки высушивают и исследуют образцы на микроскопе. Наилучшее мечение препаратов жгутиков было достигнуто при разведении антител в 200-100, а конъюгатов в 20-10 раз.
Несмотря на то что настоящее изобретение описывает конкретные и предпочтительные варианты его реализации, следует иметь в виду, что указанные варианты не ограничивают возможные модификации и варианты настоящего изобретения.
Литература
[1] Bock L. et al. Nano-Chem-Fet Based Biosensors. US Pat. Applic. №20080044911 (February 21, 2008).
[2] Iran B.Q. NANO-electronics. US Pat. Applic. №20070285843 (December 13, 2007).
[3] Wirth R. et al. Molecular Glue. US Pat. Applic. №20080305524 (December 11, 2008).
[4] Gazit E. et al. Peptide nanostructures encapsulating a foreign material and method of manufacturing same. US Pat. Applic. №20060079455 (April 13, 2006).
[5] Nam K.T. et al. Virus-Enabled Synthesis and Assembly of Nanowires for Lithium Ion Battery Electrodes. Science, V.312, P.885-888 (2006).
[6] Belcher A.M. et al. Nanoscaling ordering of hybrid materials using genetically engineered mesoscale virus. US Pat. Applic. №20030073104 (April 17, 2003).
[7] Belcher A.M. et al. Multifunctional biomaterials as scaffolds for electronic, optical, magnetic, semiconducting, and biotechnological applications. US Pat. Applic. №20050170336 (August 4, 2005).
[8] Belcher A.M. et al. Biological control ofnanoparticle nucleation, shape and crystal phase US Pat. Applic. №20060275791 (December 7, 2006).
[9] Nam К.Т. et al. Virus scaffold for self-assembled, flexible and light lithium battery. US Pat. Applic. №20060121346 (June 8, 2006).
[10] Мао С. et al., Virus-Based Toolkit for the Directed Synthesis of Magnetic and Semiconducting Nanowires. Science, 2004, V.303, P.213-217.
[11] Kumara M.T. et al., Self-assembly of metal nanoparticles and nanotubes on bioengineered flagella scaffolds. Chem. Mater., 2007, V.19, P.2056-2064.
[12] Yu В. et al., A Novel Biometallic Interface: High Affinity Tip-Associated Binding by Pilin-Derived Protein Nanotubes. J.Bionanosci., 2007, V.1, 73-83.
[13] Audette G.F., Hazes B. Development of protein nanotubes from a multi-purpose biological structure. J.Nanosci. Nanotechnol., 2007, V.7, P.2222-2229.
[14] Атабеков И.Г. Применение вирусных структур в качестве инструментов нанотехнологий. Росс. Нанотехн., 2008, Т.3, Р.132-141.
[15] Mudalige Thilak Kumara et al., Bioengineered Flagella Protein Nanotubes with Cysteine Loops: Self-Assembly and Manipulation in an Optical Trap Nano Lett., Vol.6, No.9, 2006.
[16] Benita Westerlund-Wikstro et al., Functional expression of adhesive peptides as fusions to Escherichia coli flagellin. Protein Engineering vol.10 no.11 pp.1319-1326, 1997.
[17] Thomas A.N. et al., The archaeal flagellum: a different kind of prokaryotic motility structure. FEMS Microbiol. Reviews, 2001, V.25, P.147-174.
[18]] Logan S.M. Flagellar glycosylation - a new component of the motility repertoire? Microbiol., 2006, V.152, P.1249-1262.
[19] Tarasov V.Y., Pyatibratov M.G., Tang S., Dyall-Smith M., Fedorov O.V. // Role of flagellins from A and B loci in flagella formation of Halobacterium salinarum. Mol. Microbiol., 2000, V.35, P.69-78.
[20] Fabricating Genetically Engineered High-Power Lithium Ion Batteries Using Multiple Virus Genes. Yun Jung Lee, Hyunjung Yi, Woo-Jae Kirn, Kisuk Kang, Dong Soo Yun, Michael S. Strano, Gerbrand Ceder, Angela M. Belcher. 2 April 2009/10.1126/science.
[21] Corrine K. Thai et al., Identification and Characterization of Cu2O- and ZnO-Binding Polypeptides by Escherichia coli Cell Surface Display: Toward an Understanding of Metal Oxide Binding. BIOTECHNOLOGY AND BIOENGINEERING, VOL.87, NO. 2, JULY 20, 2004.
[22] Cline S.W. et al., Transformation methods for halophilic archaebacteria. Can. J. Microbiol., 1989, V.35, P.148-152.
[23] Dyall-Smith M. The halohandbook: protocols for halobacterial genetics. 2008 (http://www.haloarchaea.com/resources/halohandbook).
[24] Sambrook J., Russell D.W. Molecular cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 2001.
[25] Inoue H. et al., High efficiency transformation of Escherichia coli with plasmids. Gene, 1990, V.96, P.23-28.
[26] Oesterhelt The system was conceived and implemented within the Dept. of Membrane Biochemistry of the Max-Planck-Institute of Biochemistry, Martinsried, Germany (http://www.halolex.mpg.de/public)/











Изобретение относится к области биотехнологии и может быть использовано для получения наноструктурированного материала на основе рекомбинантных жгутиков архей H. salinarum, связанных с ионами металлов или наночастицами. Выращивают трансформированные рекомбинантной плазмидой клетки архей, выделяют жгутики, содержащие пептидные вставки для связывания с ионами металлов или с наночастицами. Модифицируют поверхность жгутиков посредством связывания пептидных вставок с указанными ионам или наночастицами с последующей промывкой, сушкой и упаковкой полученного материала. Конструкция плазмиды содержит рекомбинантные гены для синтеза флагеллинов А1 и А2, формирующих жгутик. При этом последовательность флагеллина А1 и/или флагеллина А2 содержит пептидную вставку для избирательного связывания ионов металлов или наночастиц, где место пептидной вставки определяют в области между первым и вторым сайтами гликозилирования, расположенной в флагеллине А1 между позицией 86 и позицией 96 SEQ ID NO: 2, а в флагеллине А2 - между позицией 82 и позицией 92 SEQ ID NO: 3. Изобретение позволяет получить наноструктурированный материал с улучшенными адгезивными свойствами, устойчивый по отношению к воздействию высоких температур. 9 з.п. ф-лы, 7 ил., 7 пр. 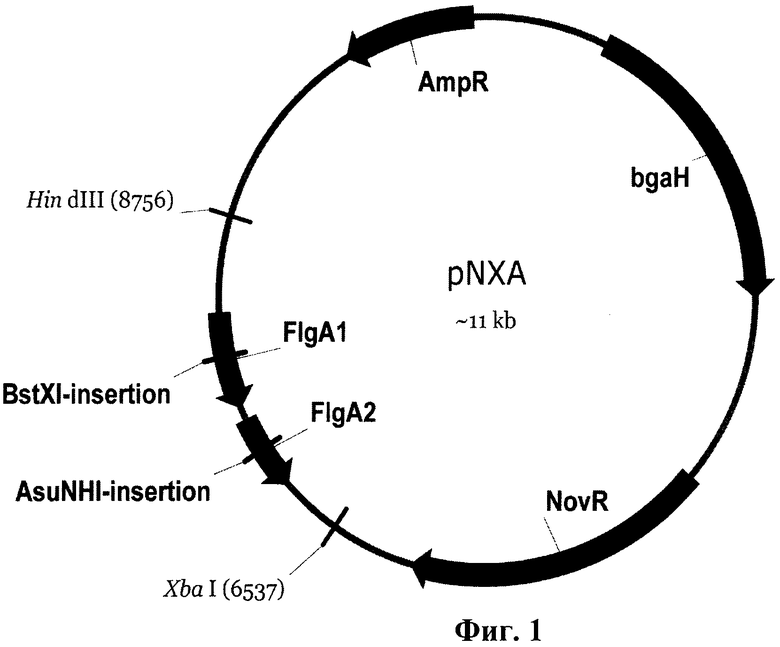
1. Способ получения наноструктурного материала на основе рекомбинантных жгутиков архей Н. salinarum, связанных с ионами металлов или наночастицами, включающий трансформацию клеток архей рекомбинантной плазмидой, выращивание клеток, выделение жгутиков архей, содержащих пептидные вставки для связывания с ионами или наночастицами металлов или с наночастицами с магнитными или полупроводниковыми свойствами, модификацию поверхности жгутиков посредством связывания пептидных вставок с металлическими, полупроводниковыми, магнитными наночастицами или ионами металлов, с возможностью последующей минерализации указанных ионов или наночастиц и промывкой, сушкой и упаковкой полученного наноструктурированного материала, где конструкция плазмиды содержит рекомбинантные гены для синтеза флагеллинов А1 и А2, формирующих жгутик, при этом последовательность флагеллина А1 и/или флагеллина А2 содержит пептидную вставку для избирательного связывания ионов металлов или наночастиц, где место пептидной вставки определяют в области между первым и вторым сайтами гликозилирования, расположенной в флагеллине А1 между позицией 86 и позицией 96 SEQ ID NO: 2, а в флагеллине А2 - между позицией 82 и позицией 92 SEQ ID NO: 3, где пептидная вставка может содержать одиночную последовательность длиной от 6 до 60 аминокислот или содержать короткие последовательно соединенные друг с другом последовательности с общей длиной вставки до 60 аминокислот, при этом количество коротких последовательностей лежит в пределах от 2 до 10.
2. Способ по п.1, отличающийся тем, что генетическую конструкцию рекомбинантной плазмиды получают на основе плазмиды pNXA.
3. Способ по п.1, отличающийся тем, что модификацию поверхности жгутиков осуществляют посредством связывания пептидных вставок с ионами металлов с последующей минерализацией.
4. Способ по п.3, отличающийся тем, что последовательность пептидной вставки выбирают таким образом, чтобы она нековалентно связывала ионы металлов, выбираемых из группы, в которую входят: кобальт, ванадий, никель, марганец, железо, кадмий, вольфрам, хром, цирконий, титан, скандий, иттрий, медь, кальций, алюминий, барий, бериллий, магний и стронций.
5. Способ по п.1, отличающийся тем, что модификацию поверхности жгутиков осуществляют посредством связывания пептидных вставок с металлическими, полупроводниковыми, магнитными наночастицами.
6. Способ по п.5, отличающийся тем, что последовательность пептидной вставки выбирают таким образом, чтобы она нековалентно связывала наночастицы металлов выбираемых из группы, состоящей из золота, платины, серебра, палладия.
7. Способ по п.5, отличающийся тем, что последовательность пептидной вставки выбирают таким образом, чтобы она нековалентно связывала наночастицы, обладающие полупроводниковыми свойствами, входящими в группу: ZnS, CdS.
8. Способ по п.5, отличающийся тем, что последовательность пептидной вставки выбирают таким образом, чтобы она нековалентно связывала наночастицы, обладающие магнитными свойствами, входящими в группу: FePt, CoPt.
9. Способ по п.1, отличающийся тем, что пептидную вставку выбирают из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 9-20.
10. Способ по п.9, отличающийся тем, что пептидную вставку выбирают из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 9 и 11.
| KUMARA M.T | |||
| et al., Self-Assembly of Metal Nanoparticles and Nanotubes on Bioengineered Flagella Scaffolds, Chem | |||
| Mater., 2007, v | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Приспособление для разгонки рельс | 1925 |
|
SU2056A1 |
| DEPLANCHE K et al, Manufacture of stable palladium and gold nanoparticles on native and genetically engineered flagella scaffolds, Biotechnol | |||
| Bioeng., 2008, v | |||
| Приспособление для записи звуковых явлений на светочувствительной поверхности | 1919 |
|
SU101A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Щелевая форсунка | 1925 |
|
SU873A1 |

