Область техники
Изобретение относится к области генной инженерии и касается способа детекции и выделения ДНК, кодирующей пептид, который обладает секреторной активностью.
Предпосылки
К настоящему времени гены, кодирующие гормоны или факторы роста, были выделены и использованы для получения большого числа рекомбинантных белков, имеющих коммерческое значение в качестве лекарственных средств. Большинство из них являются секреторными белками. Следовательно, выделение гена, кодирующего новый секреторный белок, является исключительно важным этапом в разработке новых лекарственных средств. С учетом этого и были разработаны способы выделения гена, кодирующего секреторный белок. Например, Honjo et al. разработали способ (нерассмотренная опубликованная Японская патентная заявка №Hei 6-315380), основывающийся на использовании того свойства, что секреторные белки включают сигнальную последовательность, которая обеспечивает белкам, экспрессированным внутри клеток, способность перемещаться на клеточную поверхность. В этом способе сигнальная последовательность α-субъединицы рецептора интерлейкина-2-человека, являющейся секреторным белком, замещается коротким фрагментом кДНК, который соответствует 5'-концевой последовательности мРНК клетки-мишени или ткани-мишени с получением библиотеки, которую затем вносят в клетки. Среди полученных клонов рецептор IL-2 экспрессируется на поверхности клеток тех клонов, в которых имеется сигнальная последовательность, но не в тех, где она отсутствует. Присутствие сигнальной последовательности, таким образом, может быть выявлено с использованием антитела, специфичного по отношению к рецептору IL-2.
В Генетическом институте Кембриджского ун-та (Genetics Inst. Inc.: Camdridge, MA, США) была разработана более чувствительная система, основанная на использовании метаболического фермента дрожжей (патент США №5536637). Этот метаболический фермент дрожжей, называемый инвертазой, является секретируемым ферментом, катализирующим расщепление сахарозы в культуральной среде с образованием глюкозы и фруктозы в системе переноса энергии. Мутантный штамм, который не способен секретировать этот фермент, не может расти в культуральной среде, содержащей сахарозу в качестве единственного источника углерода, отсутствие глюкозы. В этом способе, основывающемся на данном явлении, ген инвертазы лигируют на кДНК с получением библиотеки, которую затем вносят в клетки мутантного штамма дрожжей, лишенного гена инвертазы. Клоны, несущие сигнальный пептид, выделяют путем отбора клонов, способных расти в среде, содержащей только сахарозу в качестве источника углерода.
Однако недостатки способа, разработанного Honjo et аl., связаны с наличием трудоемких этапов, которые необходимы для отбора позитивных клонов, что обусловливается использованием антитела. Более того, чувствительность детекции в этом способе очень низка. Способу Генетического института (Inc.) также свойственна проблемность, определяемая тем, что клон, характеризующийся низкой эффективностью секреции в дрожжах, выделить не удается. Кроме того, эти способы позволяют выявлять только короткие ДНК из-за вероятной потери антигенных свойств или каталитической активности в случаях, когда репортерный белок слит с крупным полипептидом. Более того, эти способы не позволяют обнаружить мембранные белки II-го типа, у которых N-концевая часть “опущена” внутрь клетки, а С-концевая часть находится вне клетки.
Описание изобретения
Настоящее изобретение представляет способ, предназначенный для выяснения того, кодирует ли анализируемая кДНК пептид, обладающий способностью секретироваться, или нет. Оно также представляет способ выделения кДНК, кодирующей пептид, обладающий секреторной активностью, который позволяет использовать кДНК, кодирующую кодирующий сегмент длинного белка.
Белки, такие как рецепторы цитокинов, перемещаются на клеточную поверхность, где димеризуются в процессе связывания с соответствующими лигандами и обусловливают пролиферацию клеток. Способность белков к такому перемещению (т.е. способность секретироваться) на клеточную поверхность, как известно, зависит от наличия сигнальной последовательности. Авторы настоящей заявки представляют способ выяснения того, обладает ли желаемый пептид секреторной активностью, путем отщепления сигнальной последовательности (или иного дополнительного внеклеточного сегмента) от белков и замещения его химерным белком, включающим желательный пептид, экспрессирования химерного белка в клетках и тестирования пролиферационной активности этих клеток. Если такой пептид обладает способностью секретироваться, то химерный белок будет перемещаться на поверхность клетки, димеризоваться на ней и тем самым побуждать пролиферацию клеток. Напротив, если такой пептид не обладает секреторной активностью, то химерный белок не сможет перемещаться на поверхность клеток и индуцировать клеточную пролиферацию. Следовательно, способность химерного пептида к секреции может быть протестирована с помощью простой оценки клеточной пролиферации, используемой в качестве индекса. Более того, заявители делают возможным проведение позитивного скрининга пептида на его секреторную активность путем отбора пролиферирующих клеток. Следовательно, в настоящем изобретении используется рецептор тромбопоэтина (mpl) в качестве белка, который способствует пролиферации клеток за счет перемещения на клеточную поверхность и димеризации, и представляется способ детекции и выделения пептида, проявляющего способность секретироваться.
Конкретно заявители получили ДНК, кодирующую белок mpl человека, лишенный способности секретироваться вследствие удаления сигнального сегмента секреции и большей части внеклеточного домена из состава активной по конститутивному типу формы mpl, что было подтверждено в настоящем изобретении (mpl изменен таким образом, чтобы быть способным обусловливать автономную пролиферационную способность у зависимой от IL-3 клеточной линии за счет передачи сигнала в отсутствие лиганда: Blood, 88, 1399-1406, 1996). Затем эту ДНК лигировали на предназначенную для тестирования кДНК, на ДНК, кодирующую известный секретируемый белок, или на ДНК, кодирующую секретируемый пептид, из состава которого сигнальный сегмент удален. Полученные в результате химерные гены экспрессировали в клетках и определяли параметры пролиферационной активности этих клеток. Полученные результаты показывают, что ДНК, кодирующая известный секретируемый белок, использовавшаяся в качестве тест-кДНК, индуцировала пролиферацию клеток, в то время как пролиферация клеток не выявлялась в случае с ДНК, кодирующей секретируемый белок, из состава которого был предварительно удален сигнальный сегмент. В этом случае заявители обнаружили, что разработанная таким образом система может быть использована для простого выявления и выделения ДНК, кодирующей пептид, обладающий секреторной активностью и включающий участок, кодирующий длинный пептид при том, что в качестве индекса используется показатель клеточной пролиферации. Действительно, был осуществлен скрининг, приведший к успешному выявлению и выделению молекул ДНК, кодирующих секретируемые белки, включая мембранные белки I-го типа и мембранные белки II-го типа.
Таким образом, настоящее изобретение представляет:
(1) пептид, способный индуцировать пролиферацию клеток по механизму димеризации на клеточной поверхности и утративший способность секретироваться;
(2) пептид, описанный в п.(1), при том, что этот пептид является производным от рецептора цитокина;
(3) пептид, описанный в п.(1), при том, что этот пептид является производным от белка mpl;
(4) пептид, описанный в пп.(2) или (3), при том, что этот пептид является лиганднезависимым;
(5) пептид, описанный в п.(1), при том, что этот пептид включает аминокислотную последовательность SEQ ID NO 4;
(6) ДНК, кодирующая пептид, описанный в любом из пп.(1)-(5);
(7) вектор, включающий ДНК, описанную в п.(6), и сайт клонирования для кДНК по 5'-верхнему участку ДНК;
(8) вектор, описанный в п.(7), при том, что этот вектор является производным от ретровируса;
(9) вектор, описанный в пп.(7) или (8), при том, что кДНК встроена в 5'-верхний участок ДНК по п.(6);
(10) клетка, несущая вектор, описанный в п.(9);
(11) клетка, описанная в п.(10), при том, что эта клетка является клеткой млекопитающего;
(12) способ определения того, обладает ли тестируемый пептид, кодируемый кДНК, способностью секретироваться, при том, что этот способ включает:
(a) лигирование тестируемой кДНК в состав вектора по п.(7);
(b) внесение вектора, полученного на этапе (а), в клетку;
(c) культивирование трансформанта, полученного на этапе (b), и
выявление способности клеток v-пролиферации;
(13) способ выделения кДНК, кодирующей пептид, обладающий способностью секретироваться, при том, что данный способ включает:
(a) лигирование библиотеки кДНК в состав вектора по п.(7);
(b) внесение вектора, полученного на этапе (а), в клетку;
(c) культивирование трансформанта, полученного на этапе (b), и
выявление способности клеток к пролиферации;
(d) отбор позитивной клетки, которая принимается как клетка, обладающая способностью к пролиферации на этапе (с), и выделение кДНК из упомянутой клетки;
(14) способ в соответствии с описанным в пп.(12) или (13), при том, что вектор является ретровирусным, а клетка, в которую предполагается вносить упомянутый вектор, является клеткой млекопитающего;
(15) кДНК, кодирующая пептид, обладающий способностью секретироваться, которая была выделена с применением способа (13);
(16) пептид, кодируемый кДНК в соответствии с описанным в (15).
Настоящее изобретение представляет способ детекции кДНК, кодирующей пептид, обладающий способностью секретироваться. Этот детекционный способ базируется на использовании ДНК, кодирующей пептид, способный индуцировать пролиферацию клеток по механизму димеризации на клеточной поверхности и утративший секреторную активность, для целей выявления пептида, обладающего способностью секретироваться. В данном тексте выражение “пептид, способный индуцировать пролиферацию клеток по механизму димеризации на клеточной поверхности” охватывает белок mpl (Proc. Natl.Acad.Sci.USA, 89, 5640-5644 [1992]), α-цепь или β-цепь рецептора колониестимулирующего фактора гранулоцитов/макрофагов (GM-CSF) (Blood, 83, 2802 [1994), рецептор эритропоэтина (Nature, 348, 647 [1989], рецептор c-kit (Blood, 85, 790 [1995]) и белок neu (Nature, 339, 230 [1989], но тем самым перечень не ограничивается. В способе по настоящему изобретению кДНК конструируется таким образом, чтобы она кодировала указанные выше пептиды, способность секретироваться которых устранена. Способность секретироваться обычно устраняется путем делегирования участка сигнальной последовательности. Например, сигнальная последовательность белка mpl человека является участком, соответствующим 1-25-м позициям аминокислотной последовательности этого белка (Proc.Natl.Acad. Sci.USA, 89, 5640-5644 [1992]), в то время как в аминокислотной последовательности β-цепи рецептора GM-CSF человека он соответствует участку 1-48-й аминокислот (Proc.Natl.Acad.Sci.USA, 87, 9655-9659 [1990]). Предпочтительно внеклеточный домен также делегируют из состава такого пептида.
Пептид, кодируемый сконструированной кДНК, предпочтительно является лиганднезависимым (если такой пептид является лигандзависимым, то он может утрачивать способность связывать лиганд и становиться неактивным после формирования химерного белка). Способ создания лиганднезависимого пептида связан, например, с внесением мутации в состав такого пептида. В случае с белком mpl замена серина на аспарагин в 498-м положении может снять зависимость от лиганда - тромбопоэтина (Blood, 88, 1399-1406 [1996]). Белок mpl, использовавшийся в представляемом настоящим изобретением способе, предпочтительно включает аминокислотную последовательность, показанную в SEQ ID NO 4.
ДНК, полученная в соответствии с описанным выше, встраивается в состав подходящего экспрессирующего вектора. Тип экспрессирующего вектора ничем не ограничивается, а предпочтительно он является ретровирусным вектором, который может быть внесен в различные клетки при высоком уровне эффективности вирусной инфекции и обеспечит стабильную экспрессию ДНК, встроенной в состав вектора в этих клетках. Примеры ретровирусных векторов включают векторы, сконструированные для библиотек кДНК, такие как pBabeX (Proc.Natl. Acad.Sci.USA, 92, 9146-9150 [1995] или рМХ (Ехр.Hematol., 24, 324-329 [1996]). Также могут быть использованы вирусные векторы, такие как аденовирусы, вирус Эпштейна-Барр и папилломавирус, или плазмидные векторы, такие как pEF-BOS (Nucl.Acids Res., 18 [1990]) и pcD SRα296 (Mol.Cell.Biol., Jan., 466-472 [1988]). Экспрессирующие векторы должны включать сайт клонирования для предназначенной к тестированию по секреторной способности кодируемого продукта кДНК, расположенный с 5'-стороны от указанной ДНК-вставки, с целью обеспечения экспрессии химерного белка. Способы создания сайта клонирования известны специалистам в данной области техники.
Далее, сформированный вектор лигируют на кДНК, тестирование которой будет осуществляться. Тестируемую кДНК лигируют в 5'-верхней части “ДНК, кодирующей пептид, способный индуцировать пролиферацию клеток по механизму димеризации на клеточной поверхности”, которая встроена в состав данного вектора. Тестируемая кДНК может быть любой кДНК, кодирующей пептид, способность секретироваться которого должна быть протестирована. Тестируемая кДНК может быть лигирована в состав вектора в соответствии со стандартными методами. Например, метод лигирования основывается на использовании ДНК-лигазы фага Т4 с помощью линкера (T.Maniatis: Molecular Cloning).
Сформированный вектор затем вносят в клетку. Типы клеток, в которые вносится данный вектор, ничем не ограничиваются и включают цитокинзависимую пролиферирующую клетку, такую как Ba/F3, OTT-1, FDCP-1 и TF-1. Векторы могут быть внесены в клетки с применением стандартных методов, включая липофекцию, кальцийфосфатный метод, метод с DEAE-декстраном и электропорацию. В методе внесения, опосредуемого инфекционными ретровирусами, вектор вносят в упаковочные клетки, где он встраивается в вирусные частицы. Этот вектор может быть внесен с применением стандартных методов, таких как кальцийфосфатный метод и липофекция. Например, в качестве упаковочных клеток могут быть использованы такие клетки, как BOSC23, Bing (Proc. Natl. Acad. Sci. USA, 90, 8392-8396 [1993]) или клетки NX-E и NX-A (G.P.Nolan, 1998, Immunity, 8, 461-471).
Далее, полученные таким образом трансформанты культивируют и тестируют по их пролиферационной активности. В случае, когда белок, кодируемый встроенной в данный вектор кДНК, экспрессируется в виде химерного белка, включающего лиганднезависимый активный рецептор цитокина, данный трансформант культивируют в среде, не содержащей цитокин (т.е. лиганд), от которого клетка зависит. Если выявляется существенная пролиферация клеток, то тестируемая кДНК определяется как “позитивный клон”, кодирующий пептид, обладающий способностью секретироваться. С другой стороны, если существенная клеточная пролиферация не выявляется, то эта кДНК определяется как “негативный клон”, который кодирует пептид, не обладающий способностью секретироваться. В случае, когда белок, кодируемый встроенной кДНК, экспрессирован в виде химерного белка, включающего лигандзависимый рецептор цитокина, такой трансформант культивируют в присутствие соответствующего лиганда. Если выявляется существенная клеточная пролиферация, что оценивают путем сравнения с отрицательным контролем в отсутствие соответствующего лиганда (если это является необходимым), тестируемая кДНК определяется как “позитивный клон”. Другие условия культивирования трансформантов могут быть подходящим образом выбраны специалистом в данной области техники, учитывая тип клеток, в которые данный вектор встраивают, а также природу химерного белка, который будет экспрессирован в них.
Также настоящее изобретение касается способа выделения кДНК, кодирующей пептид, который обладает способностью секретироваться. В этом способе библиотеку кДНК лигируют в состав вектора вместо описанной выше тестируемой кДНК, который затем используют для выявления кДНК, кодирующей пептид, обладающий способностью секретироваться. В одном конкретном варианте настоящего изобретения кДНК, полученные в результате использования случайной затравки, лигировали через BstXI-адаптор и встраивали между двумя сайтами узнавания рестриктазами BstXI: один из этих сайтов относится к вектору, а другой сайт встраивается в состав участка, кодирующего внеклеточный участок процессинга активного mpl. Источники библиотеки кДНК не ограничиваются каким-либо одним источником, но могут представлять собой клетку или ткань, из которых может быть выделен желательный пептид, обладающий способностью секретироваться. Для конструирования библиотеки кДНК могут быть использованы различные стандартные методы. В представляемом способе клетки, для которых признана способность к пролиферации, отбирают из клеток, в которые была внесена библиотека кДНК; кДНК, содержащаяся в отобранных клетках, как предполагается кодирует пептид, обладающий способностью секретироваться кДНК может быть выделена из клеток, пролиферация которых может быть выявлена, например, путем экстрагирования геномной ДНК или РНК, амплифицирования интересующей кДНК с помощью ПЦР с использованием затравок, сформированных так, чтобы охватывать сайты клонирования (в случае с РНК - после их превращения в ДНК с использованием обратной транскриптазы), и выделения полученных продуктов.
И в случае с полноразмерной выделенной кДНК или ее фрагментом, и в случае с кДНК, кодирующей новый секретируемый пептид, все они могут быть проанализированы путем сравнения последовательности кДНК с последовательностями известных белков, имеющимися в базах данных. Если кДНК не является полноразмерной, ее используют для скрининга вторичной библиотеки кДНК с целью выделения полноразмерной кДНК. Вторичная библиотека кДНК может быть сконструирована с применением методов, известных специалистам в данной области техники, таких, которые описаны в научной литературе (Sambrook et al., 1989, “Molecular Cloning: A Laboratory Manual”, 2-d ed., Cold Spring Harbor Lab. Press, NY).
кДНК, кодирующая пептид, обладающий способностью секретироваться, выделенная с помощью способа по настоящему изобретению, может быть использована для выработки рекомбинантного белка, который будет применим в качестве лекарственного средства или в генотерапии соответствующих заболеваний. Рекомбинантный белок, кодируемый выделенной кДНК, может быть получен с применением методов, известных в данной области техники. Например, кДНК вносят в состав подходящего вектора, таких как pED (Kaufman et al., 1991, Nucl. Acids Res., 19, 4484-4490), pEF-BOS (Mizushima et al., 1990, Nucl. Acids Res., 18, 5322), pXM, рJ13 и pJL4 (Gough et al., 1985, EMBO J., 4, 645-653), вектор вносят в подходящую клетку-хозяина, полученный трансформант культивируют так, чтобы обеспечить экспрессию рекомбинантного белка, и полученный рекомбинантный белок очищают.
Краткое описание чертежей
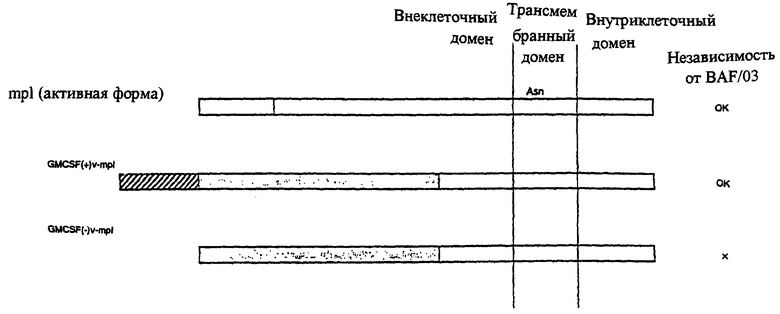
На чертеже 1 схематически показаны пептиды, применяемые для детекции способности секретироваться, и результаты детекции цитокиннезависимой пролиферационной способности клеток линии BAF/03 по параметрам экспрессии пептидов.
Наилучший путь осуществления изобретения
Настоящее изобретение подробно описано ниже со ссылкой на примеры, которые, тем не менее, приведены, не имея целью какое-либо его ограничение.
Пример 1
Конструирование вектора
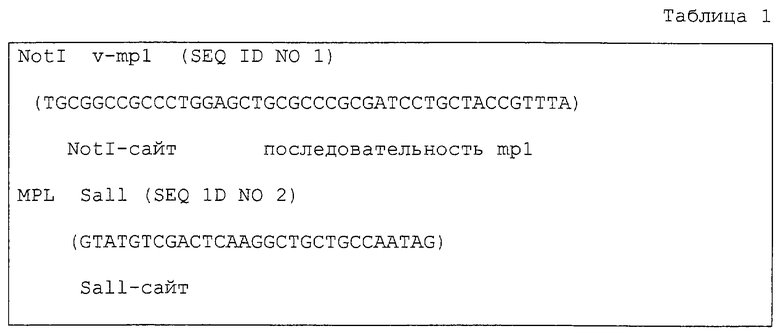
С использованием вируса миелопролиферативного лейкоза мышей ген env лигировали на ген mpl мыши, включающий внеклеточный сегмент, состоящий из 56 аминокислот - от трансмембранного домена до N-конца, весь трансмембранный домен и внутриклеточный домен. ПЦР проводили с целью получения кДНК, кодирующей соответствующий участок mpl человека, который простирается от 449-го кодона (лейцин) до стоп-кодона (636-й), включая NotI-сайт, находящийся непосредственно перед кодоном лейцина-449 (вставка единственного нуклеотида), а также Sall-сайт непосредственно за стоп-кодоном так, чтобы он находился в пределах рамки кДНК GM-CSF, показанной далее. Конструкцию “pBabeX MPLM” (Blood, 88, 1399-1406 [1996]), в которую клонировали кДНК активного mpl человека, использовали в качестве матрицы. Использовавшиеся затравки приведены в табл. 1.
ПЦР осуществляли в реакционной смеси, содержащей 10 мкг/мл матричной ДНК, по 1 мкМ каждой из затравок, 50 ед./мл ДНК-полимеразы KOD (Toyobo), 1 мМ MgCl2, 0/2 мМ смеси дНТФ, 120 мМ Трис-НСl (рН 8), 10 мМ КСl, 6 мМ (NH4)2SO4, 0,1% Тритон Х-100 и 10 мкг/мл бычьего сывороточного альбумина с применением оборудования GeneAmpPCR (Perkin Elmer) при следующих параметрах: денатурация при 98°С в течение 60 секунд, затем 25 циклов при 98°С в течение 15 секунд, при 60°С в течение 10 секунд и при 74°С в течение 30 секунд. Полученные ПЦР-продукты анализировали электрофоретически в агарозном геле и кусочки геля, содержащие интересующие фрагменты длиной 600 нуклеотидов, вырезали для последующего экстрагирования ДНК. Затем ДНК фосфорилировали по 5'-концам с использованием полинуклеотидкиназы Т4 (Toyobo) и лигировали с использованием ДНК-лигазы Т4 (Toyobo) в состав вектора pBluescript SK(-) (Stratagene), который предварительно обрабатывали рестриктазой Smal (TaKaRa Shuzo) и бактериальной щелочной фосфатазой (TaKaRa Shuzo). Нуклеотидную последовательность кДНК активного белка mpl, встроенной в состав полученной плазмиды, проверяли с использованием секвенатора ABI PRISM 310 (Perkin Elmer). Эту плазмиду обрабатывали рестриктазами NotI (TaKaRa Shuzo) и SalI (TaKaRa Shuzo) и разделяли электрофоретически в агарозном геле с целью выделения 600-нуклеотидного фрагмента. Этот фрагмент лигировали в состав плазмиды рМХ (Proc.Natl.Acad. Sci. USA, 92, 9146-9150 [1995]), которую также расщепляли рестриктазами NorI и SalI, обрабатывали бактериальной щелочной фосфатазой и очищали методом электрофореза в агарозном геле с использованием ДНК-лигазы Т4, получая в результате конструкцию “pMX-mplM”. Плазмида pMXv-mplM включает кДНК, кодирующую активный белок mpl, не обладающий способностью секретироваться. Нуклеотидная последовательность вставки кДНК и аминокислотная последовательность пептида, кодируемого этой кДНК, показаны в SEQ ID NO 3 и 4, соответственно.
Далее, для получения кДНК GM-CSF человека, в которой стоп-кодон заменен на рестрикционный NotI-сайт, ПЦР осуществляли с использованием pcDSRα 298 hGM-CSF (Proc.Natl.Acad.Sci.USA, 82, 4360-4364 [1985]), включающей кДНК GM-CSF человека в качестве матрицы. Использовавшиеся затравки показаны в табл. 2.

Конструкции EcoGM формировали таким образом, чтобы они включали кодон инициации трансляции ATG вместо серина (17-й), а в случаях конструкций EcoGMss, EcoGM EcoRI-сайт и консенсусную последовательность Козака (J.Cell Biol., 108, 29 [1989]) непосредственно перед кодоном ATG. Пары затравок для конструкций EcoGMss и GM Not были использованы в методе ПЦР с целью амплификации GM-CSF, несущей сигнальную последовательность, а конструкциии EcoGM и GM Not были использованы с целью амплификации GM-CSF, лишенного сигнальной последовательности. ПЦР осуществляли в соответствии с описанным выше, за исключением того, что для отжига использовали температуру 55°С, а полученные продукты клонировали в состав вектора pBluescript SK(-). Нуклеотидную последовательность вставок ДНК проверяли с использованием секвенатора ABI PRISM 310 (Perkin Elmer). Затем полученные плазмиды расщепляли рестриктазами EcoRI (TaKaRa) и NotI и встраивали по сайгам EcoRI/NotI в состав конструкции рМХ v-mplM в соответствии с описанным выше, в результате чего были получены конструкции "рМХ GM(+) v-mplM" и "рМХ GM(-) v-mplM". Конструкции "рМХ GM(+) v-mplM" и " рМХ GM(-) v-mplM" кодируют химерный белок, объединяющий С-концевую часть активного белка mpl, начиная от 449-го остатка (лейцин), и весь полипептид GM-CSF, включая и не включая сигнальную последовательность, соответственно. Нуклеотидные последовательности входящих в их состав вставок кДНК показаны в SEQ ID NO 8 и 10, а аминокислотные последовательности белков, кодируемых этими кДНК, показаны в SEQ 1D NO 9 И 11.
Пример 2
Вирусная инфекция
Каждую из названных выше плазмид вносили в упаковочную клеточную линию BOSC23 (Proc.Natl.Acad.Sci.USA, 90, 8392-8396 [1993]) с использованием метода липофекции: LipofectAMINE (Life Technologies). Клетки BOSC23 высевали на 6-см чашки (Corning), содержащие модифицированную по Дальбекко среду Игла (DMEM; Nissui Pharm.) с 10% плодной телячьей сывороткой (ПТС: JRH Biosciences). Спустя 6 часов инкубации клетки промывали средой OPT I-MEM I со сниженным содержанием сыворотки (Life Technologies). Отдельно липофектамин (18 мкл) разводили в 200 мкл среды OPTI-MEM I и смешивали с 3 мкг образцов каждой плазмиды, разведенных в свою очередь в 200 мкл OPTI-MTM I. Полученные в результате смеси оставляли при комнатной температуре на 15 минут, смешивали с 1,6 мл среды OPTI-MTM I и затем добавляли к клеткам. Через 5 часов 2 мл к клеткам добавляли 2 мл среды DMEM, содержащей 20% ПТС, которые затем инкубировали еще в течение 19 часов. Затем культуральную среду заменяли на 3 мл DMEM, содержащей 10% ПТС и надосадочную фракцию культуры выделяли через 24 часа. Интерлейкин-3 мыши (IL-3) и 10 мкг/мл полибрена (гексадитметринбромид: Sigma) добавляли к культуральной надосадочной фракции, содержащей рекомбинантный вирус, и клетки Ba/F3 суспендировали в ней с целью обеспечения их инфицирования. Через 24 часа реакции инфицирования клетки промывали дважды средой RPMI-1640 (Nissui Pharm.), содержащей 10% ПТС без мышиного IL-3, и продолжали культивирование в той же самой среде.
Клетки, несущие химерный белок, между полноразмерным GM-CSF, включающим сигнальную последовательность, и активным участком mpl (производным от конструкции рМХ GM(+)v-mplM), росли в отсутствие интерлейкина-3 так же, как в случае присутствия активного mpl, обладающего способностью секретироваться. Напротив, клетки, несущие химерный белок, между GM-CSF без сигнальной последовательности и активным белком mpl (конструкция рМХ GM(+)v-mplM), не могли расти - так же, как и контрольные клетки Ba/F3, в которые не вносили вектор, экспрессирующий химерный белок (фиг.1).
Пример 3
Скрининг
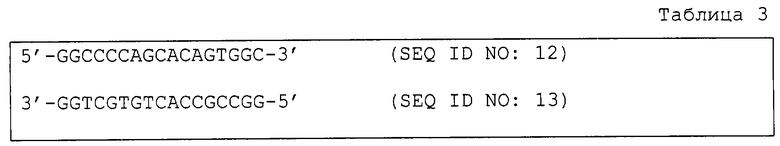
Были синтезированы следующие олигонуклеотиды (табл. 3), 5'-концы которых были фосфорилированы с использованием полинуклеотидкиназы Т 4. Эти олигонуклеотиды смешивали и денатурировали при 95°С и затем отжигали их при постепенно понижающейся до 40°С температуре с получением в результате кассеты ДНК.
Конструкцию рМХ GM(-) v-mplM, которую расщепляли рестриктазой NotI (TaKaRa) и обрабатывали бактериальной щелочной фосфатазой, смешивали с полученной кассетой и лигировали на нее с использованием ДНК-лигазы Т4. Проверяемая путем секвенирования ДНК ориентация кассеты в полученной в результате плазмиде должна была быть направлена от BstXI-сайта к NotI-сайту (рМХ GM(-) v-mplM). Пул тотальной РНК готовили на материале линии нейробластных клеток MNS70 крысы с использованием реагента TRIZOL (Gibco BRL) и пропускали через колонку, содержащую олиго-дТ фрагменты (Pharmacia), с получением полиаденилированных РНК. Двухцепочечные кДНК синтезировали с использованием метода случайных гексамеров в системе Super-Script Choice System (Gibco BRL). У кДНК “затупляли” концы, лигировали ее на адаптор с BstXI-сайтами (Invitrogen) и фракционировали с использованием вращающейся колонки SizeSep 400 Spun Column (Pharmacia). Затем кДНК смешивали и лигировали в состав плазмиды рМХ GM(-)v-mplM, которую расщепляли рестриктазой BstXI (TaKaRa) и обрабатывали бактериальной щелочной фосфатазой, с использованием ДНК-лигазы Т4. Полученную ДНК вносили в клетки E.coli штамма DH10B (Gibco BRL) методом электропорации с использованием прибора Gene Pulser (BioRad) с получением библиотеки кДНК.
Плазмиды экстрагировали из рекомбинантных клеток E.coli, несущих библиотеку кДНК, и очищали на колонке JET-star (Genomed). Библиотечные плазмиды вносили в упакованные клетки линии BOSC23 с использованием липофектамина в соответствии с описанным выше. К культуральному надосадочному слою, содержащему рекомбинантные вирусы, добавляли мышиный интерлейкин-3 (10 нг/мл) и полибрен (10 мкг/мл) (гексадиметринбромид: Sigma), а клетки Ba/F3 суспендировали в нем для обеспечения их инфицирования. Через 24 часа периода инфицирования клетки дважды промывали фосфатным буфером и далее культивировали в среде RPMI-1640, содержащей 10% ПТС. Геномную ДНК получали из тех клонов, которые росли в отсутствие IL-3, а ПЦР проводили с использованием затравок, сконструированных таким образом, чтобы включить сайт встраивания кДНК, что обеспечит выделение кДНК-фрагментов.

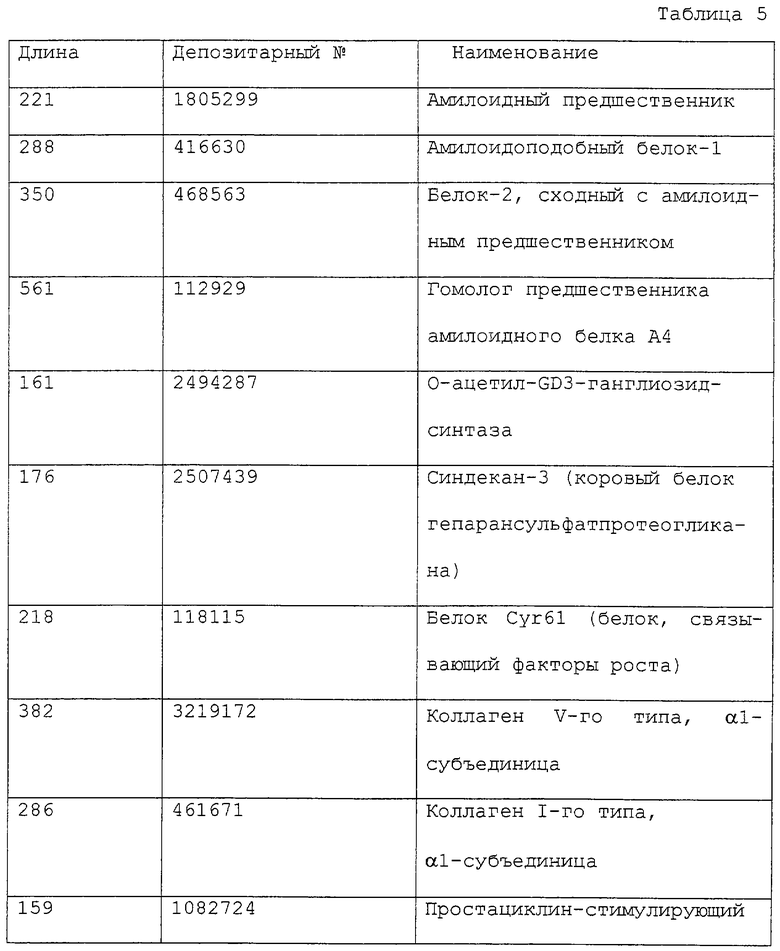
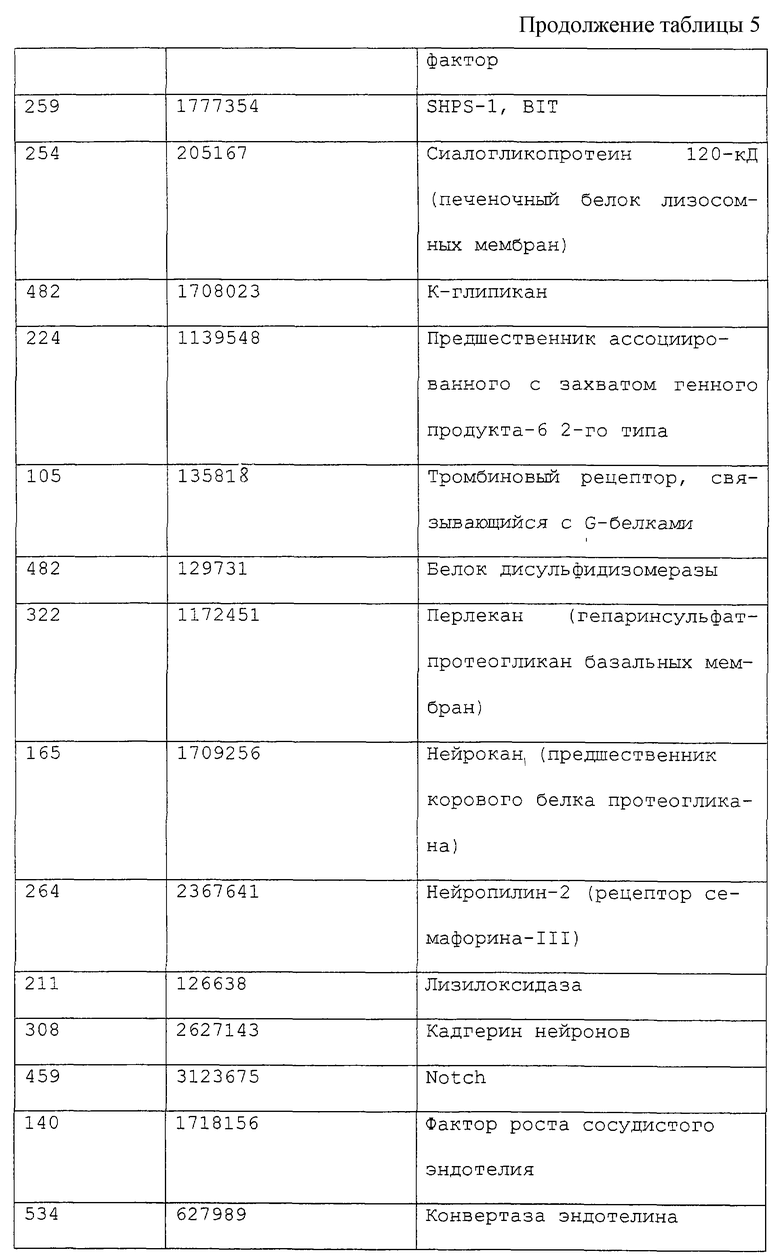
ПЦР проводили при объеме реакционной смеси 50 мкл, содержащей 500 нг геномной ДНК, 500 пкМ каждой из затравок, 2,5 ед. Taq-полимеразы TaKaRa LA (TaKaRa), 2,5 mm MgCl2, 0,3 мМ смеси дНТФ и соответствующий буфер, с использованием амплификатора GeneAmp PCR System 2400 по следующей схеме: денатурация при 98°С в течение 1 минуты, затем 30 циклов при 98°С в течение 20 секунд и 68°С в течение 2 минут. Полученные ПЦР-продукты разделяли электрофоретически в агарозном геле и вырезали участки геля, содержащие амплифицированные фрагменты, с последующей очисткой ДНК. Были определены нуклеотидные последовательности ДНК-фрагментов из 190 полученных в результате клонов, а для 150 клонов было установлено присутствие кДНК, кодирующих известный секретируемый белок или мембранный белок, или их участки. Для 40 других клонов было установлено кодирование неизвестных секретируемых белков. Некоторые из полученных с помощью такого подхода секретируемых белков показаны в табл. 5, в которой обозначение “длина” указывает на размер открытой кодирующей рамки, имеющейся в составе выделенного кДНК-фрагмента, выраженный в числе кодируемых аминокислотных остатков. Средняя длина клонов, кодирующих известные секретируемые белки, составила 273 аминокислоты. Обозначение “депозитарный номер” указывает на тот номер, под которым было произведено депонирование в базу данных GenBank. Необходимо отметить, что “фоновый” показатель (т.е. частота ошибок) для заявляемого способа - по сути частота выявления кДНК, кодирующей несекретируемые белки, или кДНК, которые встроены в обратной ориентации, - составляет 1% и менее.

Промышленное применение
Настоящее изобретение представляет способ детекции и выделения кДНК, кодирующей секретируемый пептид на основе использования пептида, способствующего пролиферации клеток по механизму его димеризации на клеточной поверхности при отсутствии у него способности секретироваться. Поскольку данный способ в качестве детекционного показателя (индекса) использует наличие клеточной пролиферации, то он оказывается простым и чувствительным. Более того, по сравнению со стандартными способами, которые позволят выявлять лишь короткие фрагменты ДНК, заявляемый способ позволяет детектировать и выделять кДНК, включающие более длинные кодирующие участки, что тем самым обеспечивает получение большего количества информации уже по первым выделенным клонам. Кроме того, данный способ обеспечивает детекцию и выделение секретируемых белков, включающие мембранные белки I-го и II-го типов.

















Изобретение относится к биотехнологии, в частности к генной инженерии, и может быть использовано для детекции и выделения ДНК, кодирующей пептид, который обладает секреторной активностью. ДНК, кодирующую белок mpI человека, лишенный сигнальной последовательности, лигируют в векторе с тестируемой кДНК, кодирующей пептид. Химерный ген экспрессируют в клетках для оценки способности к клеточной пролиферации. По способности к пролиферации клеток определяют присутствие сигнального пептида в тестируемой кДНК. ДНК, кодирующие секретируемые белки, выявляют путем конструирования библиотеки кДНК с последующим ее скринингом. Изобретение позволяет выявлять секретируемые белки, в том числе, мембранные белки I-го и II-го типа. 5 с. и 6 з.п. ф-лы, 1 ил., 5 табл.
| ISABELLE VIGON et al | |||
| Molecular cloning and characterization of MPL, the human homolog of the v-mpI oncogene... | |||
| Proc | |||
| Nat | |||
| Acad | |||
| Sci | |||
| USA, 1992, v.89, p.5640-5644 | |||
| KAZUHIRO HAYASHIDA et al | |||
| Molecular cloning of a second subunit of the receptor for human granulocyte-macrophage colony-stimulating factor... | |||
| Proc | |||
| Nat | |||
| Acad | |||
| Sci | |||
| USA, 1990, v.87, p.9655-9659 | |||
| MAYUMI ONISHI et al | |||
| Identification of an oncogenic form of the Thrombopoietin receptor MPL... | |||
| Blood | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |

