ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет согласно заявкам на патент США №62/463353 (подана 24 февраля 2017 г.; на рассмотрении) и 62/597594 (подана 12 декабря 2017 г.; на рассмотрении), которые полностью включены в настоящий документ посредством ссылки.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка включает один или более перечней последовательностей в соответствии с §1.821 раздела 37 Свода федеральных правил США и далее, которые раскрыты на машиночитаемом носителе (название файла: 1301_0149PCT_ST25.txt, создан 11 февраля 2018 г. и имеет размер 309094 байта), данный файл полностью включен в настоящий документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение относится к связывающим молекулам, которые имеют один или более эпитопсвязывающих сайтов, специфичных для эпитопа CD137, и один или более эпитопсвязывающих сайтов, специфичных для эпитопа опухолевого антигена («ТА») (например, «CD137×TA связывающая молекула»). Согласно одному варианту реализации настоящего изобретения такие CD137×TA связывающие молекулы будут представлять собой биспецифические молекулы, в частности, биспецифические четырехвалентные диатела, которые состоят из двух, трех, четырех или более четырех полипептидных цепей и имеют два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа CD137, и два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа ТА. Согласно другому варианту такие CD137×TA связывающие молекулы будут представлять собой биспецифические молекулы, в частности, биспецифические трехвалентные связывающие молекулы, состоящие из трех или более полипептидных цепей и имеющие один или два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа CD137, и один или два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа ТА. CD137×TA связывающие молекулы согласно настоящему изобретению способны одновременно связываться с CD137 и ТА. Настоящее изобретение относится к фармацевтическим композициям, которые содержат любые такие CD137×TA связывающие молекулы. Настоящее изобретение дополнительно относится к способам применения таких молекул в лечении рака и других заболеваний и состояний. Согласно настоящему изобретению также предложены новые CD137-связывающие молекулы и HER2/neu-связывающие молекулы, а также их производные и варианты их применения.
УРОВЕНЬ ТЕХНИКИ
[0004] CD137 (также известный как 4-1ВВ и «член 9 суперсемейства рецепторов TNF» («TNFRSF9»)) представляет собой костимулирующий рецептор, который является членом суперсемейства рецепторов фактора некроза опухолей, опосредующий СD28-зависимую и независимую костимуляцию Т-клеток (Vinay, D.S. and Kwon, B.S. (1998) «Role of 4-1BB in immune responses», Semin Immunol. 10:481-489; Bartkowiak, T. et al. (2015) «4-1BB Agonists: Multi-Potent Potentiators Of Tumor Immunity», Frontiers Oncol. 5:117; pp. 1-16; So, Т., et al. (2008) «Immune Regulation And Control Of Regulatory T Cells By OX40 And 4-1BB», Cytokine & Growth Factor Rev. 19:253-262; Croft, M. (2009) «The Role Of TNF Superfamily Members In T-Cell Function And Diseases», Nat. Rev. Immunol. 9:271-285; Yonezawa, A. et al. (2015) «Boosting cancer immunotherapy with Anti-CD137 antibody therapy», Clin. Cancer Res. 21(14):3113-3120; Li, S.Y. et al. (2013) immunotherapy Of Melanoma With The Immunecostimulatory Monoclonal Antibodies Targeting CD137», Clin. Pharmacol. 5:47-53; Vinay, D.S. et al. (2012) «Immunotherapy Of Cancer With 4-1BB», Mol. Cancer Ther. 11:1062-1070; Houot, R. et al. (2012) «Boosting Antibody-Dependent Cellular Cytotoxicity Against Tumor Cells With A CD137 Stimulatory Antibody», Oncoimmunology. 1:957-958; Kwon, B.S. et al. (1989) «cDNA Sequences Of Two Inducible T-Cell Genes», Proc. Natl. Acad. Sci. (U.S.A.) 86:1963-1967; Chen, L. et al. (2013) (Molecular Mechanisms Of T Cell Co-Stimulation And Coinhibition», Nat. Rev. Immunol. 13:227-242; Yao S. et al. «Advances In Targeting Cell Surface Signaling Molecules For Immune Modulation», Nat. Rev. Drug Discov. 12:130-146).
[0005] CD137 индуцируемо экспрессируется T-клетками, природными клетками-киллерами (NK), дендритными клетками (ДК), В-клетками и другими клетками иммунной системы (Vinay, D.S. et al. (2015) «Therapeutic Potential Of Anti-CD137 (4-1BB) Monoclonal Antibodies, Expert Opinion On Therapeutic Targets», DOI:10.1517/14728222.2016.1091448; pp. 1-14; Wang, C. et al. (2009) «Immune Regulation By 4-1BB And 4-1BBL: Complexities And Challenges», Immunol. Rev. 229:192-215; Sallin, M.A. et al. (2014) «The Anti-Lymphoma Activities Of Anti-CD137 Monoclonal Antibodies Are Enhanced In FcγRIII-/- Mice», Cancer Immunol, Immunother. 63:947-958; Melero, I. et al. (2008) «Multilayered Action Mechanisms Of CD137 (4-1BB)-Targeted Immunotherapies», Trends Pharmacol Sci. 29:383-390; Ramakrishna, V. et al. (2015) «Characterization Of The Human T Cell Response To In Vitro CD27 Costimulation With Varlilumab», J. Immunother. Canc. 2:37; pp. 1-13). Белок состоит из 255 аминокислотного белка, имеющего короткую N-концевую цитоплазматическую часть, транс мембранную область и внеклеточный домен, который имеет 3 богатых цистеином мотива (Schwarz, Н. et al. (1993) «А Receptor Induced By Lymphocyte Activation (ILA): A New Member Of The Human Nerve-Growth-Factor / Tumor-Necrosis-Factor Receptor Family», Gene 134:295-298).
[00061 Лигирование CD137 его лигандом CD137L (4-1BBL; TNFSF9), который в основном, хотя и не исключительно, экспрессируется на антигенпрезентирующих клетках (АПК), вызывает различные ответы Т-клеток, такие как размножение клеток, повышенная секреция цитокинов и предотвращение гибели клеток, индуцированной активацией (Qian, Y. et al. (2015) «CD137 Ligand-Mediated Reverse Signaling Inhibits Proliferation And Induces Apoptosis In Non-Small Cell Lung Cancer», Med. Oncol. 32:44; pp. 1-10); Sallin, M.A. et al. (2014) «The Anti-Lymphoma Activities Of Anti-CD137 Monoclonal Antibodies Are Enhanced In FcγRIII-/- Mice», Cancer Immunol. Immunother. 63:947-958; Lee, S.W. et al. (2009) «4-1BB As A Therapeutic Target For Human Diseases, Adv. Exp. Med. Biol. 647:120-129; Thum, E. et al. (2009) «CD137, Implications In Immunity And Potential For Therapy», Front. Biosci. (Landmark Ed). 14:4173-4188; Wang, C. et al. (2009) «Immune Regulation By 4-1BB And 4-1BBL: Complexities And Challenges», Immunol. Rev. 229(1):192-215; Long, A.H. et al. (2015) «4-1ВВ Costimulation Ameliorates T Cell Exhaustion Induced By Tonic Signaling Of Chimeric Antigen Receptors», Nature Med. 21(6):581; pp. 1-13). Таким образом, такое лигирование служит для активации иммунной системы. Однако цис-взаимодействия между CD137 и CD137L также эффективно подавляют экспрессию CD137L (Kwon, В. (2015) «Is CD137 Ligand (CD137L) Signaling a Fine Tuner of Immune Responses?», Immune Network. 15(3): 121-124). Соответственно, функция лиганда CD137 заключается в контроле степени и кинетики СВ137-опосредуемой активации иммунной системы (Kwon, В. (2015) «Is CD137 Ligand (CD137L) Signaling a Fine Tuner of Immune Responses?», Immune Network. 15(3): 121-124; Shuford WW et al. (1997) «4-1BB Costimulatory Signals Preferentially Induce CD8+ T Cell Proliferation And Lead To The Amplification In Vivo Of Cytotoxic T Cell Responses», J. Exp. Med. 186:47-55).
[0007] Примечательно, что CD137, экспрессируемый на человеческих NK-клетках, становится активным после связывания с противоопухолевыми антителами (т.е. антителами, которые связывают опухолевый антиген («ТА»)), которые стали связанными с опухолевыми клетками (Houot, R. et al. (2012) «Boosting Antibody-Dependent Cellular Cytotoxicity Against Tumor Cells With A CD137 Stimulatory Antibody», Oncoimmunology. 1:957-958; Kohrt, H.E. et al. (2014) «Targeting CD137 Enhances The Efficacy Of Cetuximab», J. Clin. Invest. 124(4):2668-2682; Lin, W. et al. (2008) «Fc-Dependent Expression Of CD137 On Human NK Cells: Insights Into «Agonistic» Effects Of Anti-CD137 Monoclonal Antibodies», Blood 112(3):699-707; Mittal, P. et al. (2015) «Tumor-Unrelated CD4 T Cell Help Augments CD134 Plus CD137 Dual Costimulation Tumor Therapy», J. Immunol. Nov 11. pii: 1502032; pp. 1-14;  , A.R. et al. (2015) «Cancer Immunotherapy With Immunomodulatory Anti-CD137 And Anti-PD-1 Monoclonal Antibodies Requires Batf3-Dependent Dendritic Cells», Cancer Discov. Oct 22. pii: CD-15-0510; pp. 1-28; Wei, H. et al. (2014) «Dual Targeting Of CD137 Co-Stimulatory And PD-1 Co-Inhibitory Molecules For Ovarian Cancer Immunotherapy», OncoImmunology 3:e28248; pp. 1-3; Seo, S.K. et al. (2004) «4-1BB-Mediated Immunotherapy Of Rheumatoid Arthritis», Nat. Med. 10:1088-1094).
, A.R. et al. (2015) «Cancer Immunotherapy With Immunomodulatory Anti-CD137 And Anti-PD-1 Monoclonal Antibodies Requires Batf3-Dependent Dendritic Cells», Cancer Discov. Oct 22. pii: CD-15-0510; pp. 1-28; Wei, H. et al. (2014) «Dual Targeting Of CD137 Co-Stimulatory And PD-1 Co-Inhibitory Molecules For Ovarian Cancer Immunotherapy», OncoImmunology 3:e28248; pp. 1-3; Seo, S.K. et al. (2004) «4-1BB-Mediated Immunotherapy Of Rheumatoid Arthritis», Nat. Med. 10:1088-1094).
[0008] Такие утверждения легли в основу предположения, что антитела, которые являются иммуноспецифичными для CD137, можно применять для активации иммунной системы и обеспечения терапии рака (Melero I. et al. (1997) «Monoclonal Antibodies Against The 4-1BB T-Cell Activation Molecule Eradicate Established Tumors», Nat Med. 3:682-385; Sun, Y. et al. (2002) «Со stimulatory Molecule-Targeted Antibody Therapy Of A Spontaneous Autoimmune Disease», Nature Med. 8:1405-1413; Kammer, G.M. et al. (2002) «Immunotherapy Tackles Lupus», Nat. Med. 8(12): 1356-1358; Foell, J. et al. (2003) «CD137 Costimulatory T Cell Receptor Engagement Reverses Acute Disease In Lupus-Prone NZB x NZW F1 Mice», J. Clin. Invest. 111(10):1505-1518; Mittler, R.S. et al. (2004) «Anti-CD137 Antibodies In The Treatment Of Autoimmune Disease And Cancer», Immunol. Res. 29(1-3): 197-208; Foell, J.L. et al. (2004) «Engagement of The CD137 (4-1BB) Costimulatory Molecule Inhibits And Reverses The Autoimmune Process In Collagen-Induced Arthritis And Establishes Lasting Disease Resistance», Immunology 113(1):89-98; Sytwu, H.K. et al. (2003) «Anti-4-1BB-Based Immunotherapy For Autoimmune Diabetes: Lessons From A Transgenic Non-Obese Diabetic (NOD) Model», J. Autoimmun. 21(3):247-254; Hernandez-Chacon JA et al. (2011) «Costimulation Through The CD137/4-1BB Pathway Protects Human Melanoma Tumor-Infiltrating Lymphocytes From Activation Induced Cell Death And Enhances Antitumor Effector Function», J. Immunother. 34:236-250; Morales-Kastresana, A. et al. (2014) «Combinations Of Immunostimulatory Antibodies With Synergistic Effects Against Spontaneous Cancer», OncoImmunology 3:2, e27812, pp. 1-4; Sanmamed, M.F. et al. (2015) «Agonists of Co-stimulation in Cancer Immunotherapy Directed Against CD137, OX40, GITR, CD27, CD28, and ICOS», Seminars Oncol. 42(4):640-655; Tongu, M. et al. (2015) «Intermittent Chemotherapy Can Retain The Therapeutic Potential Of Anti-CD137 Antibody During The Late Tumor-Bearing State», Cancer Sci. 106(1):9-17; Takeda, K. et al. (2007) «Combination Antibody-Based Cancer Immunotherapy», Cancer Sci. 98(9): 1297-1302). Антитела к CD137 раскрыты в патентах США №№2014/0274909; 2013/0280265; 2013/0273078; 2013/0071403; 2012/0058047; 2011/0104049; 2011/0097313; 2008/0166336; 2008/0019905; 2006/0188439; 2006/0182744; 2006/0121030; и 2003/0223989.
[0009] Однако, несмотря на все такие достижения предшествующего уровня техники, остается потребность в улучшенных композициях, способных более активно направлять иммунную систему организма для атаки раковых клеток или клеток, инфицированных патогеном, в частности, при более низких терапевтических концентрациях. Несмотря на то, что адаптивная иммунная система может быть эффективным механизмом защиты против рака и заболевания, ее функционирование часто затрудняется механизмами подавления/уклонения от иммунитета в микросреде опухоли, опосредуемыми сниженной/отсутствующей костимулирующей активностью CD137. Кроме того, коингибирующие молекулы, экспрессируемые опухолевыми клетками, иммунными клетками и стромальными клетками в среде опухоли, могут доминантно ослаблять ответы Т-клеток против раковых клеток.
[00010] Как подробно описано ниже, настоящее изобретение удовлетворяет эту потребность, обеспечивая CD137×TA связывающие молекулы. Такие биспецифические молекулы способны связываться с опухолевыми антигенами, которые экспрессируются на поверхностях опухолевых клеток, и колокализовать CD137-экспрессирующие NK-клетки с такими опухолевыми клетками. Такая колокализация активирует NK-клетки так, чтобы стимулировать активацию или постоянную активацию иммунной системы (например, стимулирование ответа цитотоксических Т-клеток против опухолевых клеток). Эти признаки позволяют применять такие биспецифические молекулы при стимуляции иммунной системы и, в частности, в лечении рака и ассоциированных с патогенами заболеваний и состояний. Настоящее изобретение относится к указанным и другим целям.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[00011] Настоящее изобретение относится к связывающим молекулам, которые имеют один или более эпитопсвязывающих сайтов, специфичных для эпитопа CD137, и один или более эпитопсвязывающих сайтов, специфичных для эпитопа опухолевого антигена («ТА») (например, «CD137×TA связывающая молекула»). Согласно одному варианту реализации настоящего изобретения такие CD137×TA связывающие молекулы будут представлять собой биспецифические молекулы, в частности, биспецифические четырехвалентные диатела, которые состоят из двух, трех, четырех или более четырех полипептидных цепей и имеют два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа CD137, и два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа ТА. Согласно другому варианту такие CD137×TA связывающие молекулы будут представлять собой биспецифические молекулы, в частности, биспецифические трехвалентные связывающие молекулы, состоящие из трех или более полипептидных цепей и имеющие один или два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа CD137, и один или два эпитопсвязывающих сайта каждый из которых специфичен для эпитопа ТА. CD137×TA связывающие молекулы согласно настоящему изобретению способны одновременно связываться с CD137 и ТА. Настоящее изобретение относится к фармацевтическим композициям, которые содержат любые такие CD137×TA связывающие молекулы. Настоящее изобретение дополнительно относится к способам применения таких молекул в лечении рака и других заболеваний и состояний. Настоящее изобретение также относится к новым CD137-связывающим молекулам и HER2/neu-связывающим молекулам, а также их производным и вариантам их применения.
[00012] Настоящее изобретение относится к CD137×TA связывающим молекулам, которые являются одновалентными в том отношении, что они способны связываться только с одной копией эпитопа CD137 и только с одной копией эпитопа ТА, но являются биспецифическими в том отношении, что одно такое диатело способно одновременно связываться с эпитопом CD137 и с эпитопом ТА. Однако настоящее изобретение, в частности, относится к CD137×TA связывающим молекулам, которые состоят из полипептидных цепей, которые гетеродимерно связаны друг с другом с образованием двух сайтов связывания, каждый из которых специфичен для эпитопа CD137, и двух сайтов связывания, каждый из которых специфичен для эпитопа ТА. Такие предпочтительные CD137×TA связывающие молекулы согласно настоящему изобретению называют «биспецифическими четырехвалентными». Настоящее изобретение также относится, в частности, к CD137×TA связывающим молекулам, которые состоят из полипептидных цепей, которые гетеродимерно связаны друг с другом с образованием двух сайтов связывания, каждый из которых специфичен для эпитопа CD137, и одного сайта связывания, который специфичен для эпитопа ТА. Такие предпочтительные CD137×TA связывающие молекулы согласно настоящему изобретению называют «биспецифическими трехвалентными».
[00013] Согласно настоящему изобретению предложены CD137×TA связывающие молекулы, которые содержат три полипептидные цепи («первую», «вторую» и «третью» полипептидные цепи), причем первая и вторая полипептидные цепи ковалентно связаны друг с другом, и первая и третья полипептидные цепи ковалентно связаны друг с другом. Предпочтительные CD137×TA связывающие молекулы согласно настоящему изобретению содержат четыре полипептидные цепи («первую», «вторую», «третью» и «четвертую» полипептидные цепи), причем первая и вторая полипептидные цепи ковалентно связаны друг с другом, третья и четвертая полипептидные цепи ковалентно связаны друг с другом, и первая и третья полипептидные цепи ковалентно связаны друг с другом. Предпочтительными также являются CD137×TA связывающие молекулы согласно настоящему изобретению, содержащие пять полипептидных цепей («первую», «вторую», «третью», «четвертую» и «пятую» полипептидные цепи), причем первая и вторая полипептидные цепи ковалентно связаны друг с другом, третья и четвертая полипептидные цепи ковалентно связаны друг с другом, третья и пятая полипептидные цепи ковалентно связаны друг с другом, и первая и третья полипептидные цепи ковалентно связаны друг с другом.
[00014] Более конкретно согласно настоящему изобретению предложена CD137×TA связывающая молекула, причем указанная связывающая молекула способна специфично связываться с эпитопом CD137 и эпитопом опухолевого антигена (ТА), и при этом указанная CD137×TA связывающая молекула содержит первый вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и первый вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; и при этом:
(A) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL15 МАВ-3 к CD137 (SEQ ID NO: 222); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(B) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL14 МАВ-3 к CD137 (SEQ ID NO: 221); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(C) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL11 МАВ-3 к CD137 (SEQ ID NO: 218); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(D) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL10 МАВ-3 к CD137 (SEQ ID NO: 217); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(E) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL6 МАВ-3 к CD137 (SEQ ID NO: 213); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(F) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL4 МАВ-3 к CD137 (SEQ ID NO: 211); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(G) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-3 к CD137 (SEQ ID NO: 74);
(H) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-4 к CD137 (SEQ ID NO: 91); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-4 к CD137 (SEQ ID NO: 90);
(I) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-5 к CD137 (SEQ ID NO: 97); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-5 к CD137 (SEQ ID NO: 96);
(J) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1A МАВ-3 к CD137 (SEQ ID NO: 83);
(K) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(L) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1C МАВ-3 к CD137 (SEQ ID NO: 85); или
(М) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1D МАВ-3 к CD137 (SEQ ID NO: 86).
[00015] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой первый вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 77); или
(B) hMAB-4 к CD137 (SEQ ID NO: 92);
и/или в которой первый вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 82); или
(B) hMAB-4 к CD137 (SEQ ID NO: 93).
[00016] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой первый вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1E hMAB-3 к CD137 (SEQ ID NO: 208);
(B) VH1B hMAB-3 к CD137 (SEQ ID NO: 84);
(C) VH1A hMAB-3 к CD137 (SEQ ID NO: 83);
(D) VH1 hMAB-3 к CD137 (SEQ ID NO: 76);
(E) VH1C hMAB-3 к CD137 (SEQ ID NO: 85);
(F) VH1D hMAB-3 к CD137 (SEQ ID NO: 86);
(G) VH1F hMAB-3 к CD137 (SEQ ID NO: 209);
(H) VH1G hMAB-3 к CD137 (SEQ ID NO: 210); или
(I) VH1 hMAB-4 к CD137 (SEQ ID NO: 92)
[00017] Настоящее изобретение также относится к таким CD137×TA связывающим молекулам, в которых первый вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL15 hMAB-3 к CD137 (SEQ ID NO: 222);
(B) VL14 hMAB-3 к CD137 (SEQ ID NO: 221);
(C) VL1 hMAB-3 к CD137 (SEQ ID NO: 87);
(D) VL2 hMAB-3 к CD137 (SEQ ID NO: 88);
(E) VL3 hMAB-3 к CD137 (SEQ ID NO: 89);
(F) VL4 hMAB-3 к CD137 (SEQ ID NO: 211);
(G) VL5 hMAB-3 к CD137 (SEQ ID NO: 212);
(H) VL6 hMAB-3 к CD137 (SEQ ID NO: 213);
(I) VL7 hMAB-3 к CD137 (SEQ ID NO: 214);
(J) VL8 hMAB-3 к CD137 (SEQ ID NO: 215);
(K) VL9 hMAB-3 к CD137 (SEQ ID NO: 216);
(L) VL10 hMAB-3 к CD137 (SEQ ID NO: 217);
(M) VL11 hMAB-3 к CD137 (SEQ ID NO: 218);
(N) VL12 hMAB-3 к CD137 (SEQ ID NO: 219);
(O) VL13 hMAB-3 к CD137 (SEQ ID NO: 220);
(P) VL1 hMAB-4 к CD137 (SEQ ID NO: 94); или
(Q) VL2 hMAB-4 к CD137 (SEQ ID NO: 95).
[00018] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой опухолевый антиген (ТА) выбран из группы опухолевых антигенов, состоящей из: 19.9; онкофетального белка 5Т4; антигена 4.2; А33; AFP; ALCAM; BAGE; бета-катенина; СА125; карбоксипептидазы М; B1; CD5; CD19; CD20; CD22; CD23; CD25; CD27; CD30; CD33; CD36; CD46; CD52; CD79a/CD79b; CD123; CD317; СЕА; СЕАСАМ5; СЕАСАМ6; СО-43; СО-514; CTLA-1; CTLA-4; цитокератина 8; ряда Е1; ЭФР-Р; рецептора эфрина; Erb; F3; FC10.2; GAGE GD2; GD3; GD49; GM2; GM3; GICA 19-9; gp37; gp75; gp100; HER-2/neu; антигена CD20 В-лимфомы человека; антигена жировых глобул человеческого молока; Е6 папилломавируса человека/Е7 папилломавируса человека; HMW-MAA; антигена I; ITGB6; IL13Rα2; JAM-3; KID3; KID31; общего антигена карциномы KS 1/4; KS 1/4; KSA; L6; L20; LEA; LUCA-2; M1:22:25:8; М18; М39; MAGE; MART; Myl; MUC-1; MUM-1; N-ацетилглюкозаминилтрансферазы; неогликопротеина; NS-10; OFA-1; OFA-2; онкостатина M; p15; PSA; PSMA; РЕМА; PIPA; простатической кислой фосфатазы; R24; ROR1; SSEA-1; SSEA-3; SSEA-4; sTn; пептида, происходящего из Т-клеточного рецептора; TAG-72; TL5; рецептора TNF-α; рецептора TNF-β; рецептора TNF-γ; TRA-1-85; рецептора трансферрина; TSTA; и VEGF-R.
[00019] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой опухолевый антиген (ТА) выбран из опухолевых антигенов из таблицы 1 и, в частности, в которой опухолевый антиген (ТА) представляет собой: HER2/neu, EphA2 или 5Т4.
[00020] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой опухолевый антиген (ТА) представляет собой HER2/neu, причем указанная CD137×TA связывающая молекула содержит второй вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и первый вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; и при этом
(A) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-1 к HER2 (SEQ ID NO: 63); и
(B) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-1 к HER2 (SEQ ID NO: 62).
[00021] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой:
(A) (1) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(2) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(3) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL3 hMAB-1 к HER2 (SEQ ID NO: 69); и
(B) (1) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(2) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(3) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH3 hMAB-1 к HER2 (SEQ ID NO: 66).
[00022] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой второй вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(B) VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(C) VH3 hMAB-1 к HER2 (SEQ ID NO: 66).
[00023] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой второй вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(B) VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(C) VL3 hMAB-1 к HER2 (SEQ ID NO: 69).
[00024] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой опухолевый антиген (ТА) представляет собой 5Т4, причем указанная CD137×TA связывающая молекула содержит второй вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и первый вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; и при этом:
(I) (A) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-1 к 5Т4 (SEQ ID NO: 135); и
(В) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-1 к 5Т4 (SEQ ID NO: 134); или
(II) (A) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-2 к 5Т4 (SEQ ID NO: 137); и
(В) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-2 к 5Т4 (SEQ ID NO: 136).
[00025] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой второй вариабельный домен тяжелой цепи содержит аминокислотную последовательность: VH1 МАВ-1 (SEQ ID NO: 135).
[00026] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой второй вариабельный домен легкой цепи содержит аминокислотную последовательность: VH1 МАВ-1 (SEQ ID NO: 136).
[00027] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, причем указанная молекула представляет собой биспецифическое четырехвалентное Fc-несущее диатело, содержащее первую, вторую, третью и четвертую полипептидные цепи, при этом указанные полипептидные цепи образуют ковалентно связанный комплекс.
[00028] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой указанный опухолевый антиген (ТА) представляет собой HER2/neu и при этом:
(I) (А) первая и третья полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 100; и
(В) вторая и четвертая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 101;
или
(II) (А) первая и третья полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 102; и
(В) вторая и четвертая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 103.
[00029] Настоящее изобретение также относится к таким CD137×TA связывающим молекулам, причем указанная молекула является биспецифической и четырехвалентной и содержит первую, вторую, третью, четвертую и пятую полипептидные цепи, при этом указанные полипептидные цепи образуют ковалентно связанный комплекс.
[00030] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой опухолевый антиген (ТА) представляет собой HER2/neu и при этом:
(I) (А) первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 104;
(B) вторая и пятая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 105;
(C) третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 106; и
(D) четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 107;
или
(II) (А) первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 104;
(B) вторая и пятая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 105;
(C) третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117 или SEQ ID NO: 118; и
(D) четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122 или SEQ ID NO: 123.
[00031] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, причем указанная молекула является биспецифической и трехвалентной и содержит первую, вторую, третью и четвертую полипептидные цепи, при этом указанные полипептидные цепи образуют ковалентно связанный комплекс.
[00032] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой указанный опухолевый антиген (ТА) представляет собой HER2/neu, и при этом:
(А) указанная первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195 или SEQ ID NO: 196.
(B) указанная вторая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200 или SEQ ID NO: 201;
(C) указанная третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 104; и
(D) указанная четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 105.
[00033] Настоящее изобретение также относится к такой CD137×TA связывающей молекуле, в которой указанный опухолевый антиген (ТА) представляет собой 5Т4, и при этом:
(A) указанная первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195, SEQ ID NO: 196 или SEQ ID NO: 229;
(B) указанная вторая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200, SEQ ID NO: 201 или SEQ ID NO: 230;
(C) указанная третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 231; и
(D) указанная четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 232.
[00034] Настоящее изобретение также относится к фармацевтической композиции, содержащей любую из описанных выше CD137×TA связывающих молекул и физиологически приемлемый носитель.
[00035] Настоящее изобретение также относится к применению любой из описанных выше CD137×TA связывающих молекул или такой фармацевтической композиции в лечении заболевания или состояния, ассоциированного или характеризующегося экспрессией опухолевого антигена (ТА), и при этом, в частности, заболевание или состояние, ассоциированное или характеризующееся экспрессией опухолевого антигена (ТА), представляет собой рак.
[00036] Настоящее изобретение также относится к СВ137-связывающей молекуле, которая содержит вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; причем:
(A) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL15 МАВ-3 к CD137 (SEQ ID NO: 222); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(B) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL14 МАВ-3 к CD137 (SEQ ID NO: 221); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(C) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL11 МАВ-3 к CD137 (SEQ ID NO: 218); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(D) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL10 МАВ-3 к CD137 (SEQ ID NO: 217); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(E) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL6 МАВ-3 к CD137 (SEQ ID NO: 213); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(F) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL4 МАВ-3 к CD137 (SEQ ID NO: 211); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(G) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-3 к CD137 (SEQ ID NO: 74);
(H) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-4 к CD137 (SEQ ID NO: 91); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-4 к CD137 (SEQ ID NO: 90);
(I) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-5 к CD137 (SEQ ID NO: 97); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-5 к CD137 (SEQ ID NO: 96);
(J) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1A МАВ-3 к CD137 (SEQ ID NO: 83);
(K) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(L) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1C МАВ-3 к CD137 (SEQ ID NO: 85); или
(М) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1D МАВ-3 к CD137 (SEQ ID NO: 86).
[00037] Настоящее изобретение также относится к варианту реализации такой CD137-связывающей молекулы, в которой вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 77); или
(B) hMAB-4 к CD137 (SEQ ID NO: 92).
[00038] Настоящее изобретение также относится к варианту реализации такой CD137-связывающей молекулы, в которой вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 82); или
(B) hMAB-4 к CD137 (SEQ ID NO: 93).
[00039] Настоящее изобретение также относится к варианту реализации таких CD137-связывающих молекул, в которых вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1 hMAB-3 к CD137 (SEQ ID NO: 76);
(B) VH1A hMAB-3 к CD137 (SEQ ID NO: 83);
(C) VH1B hMAB-3 к CD137 (SEQ ID NO: 84);
(D) VH1C hMAB-3 к CD137 (SEQ ID NO: 85);
(E) VH1D hMAB-3 к CD137 (SEQ ID NO: 86);
(F) VH1E hMAB-3 к CD137 (SEQ ID NO: 208);
(G) VH1F hMAB-3 к CD137 (SEQ ID NO: 209);
(H) VH1G hMAB-3 к CD137 (SEQ ID NO: 210); или
(I) VH1 hMAB-4 к CD137 (SEQ ID NO: 92).
[00040] Настоящее изобретение также относится к варианту реализации таких CD137-связывающих молекул, в которых вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL15 hMAB-3 к CD137 (SEQ ID NO: 222);
(B) VL14 hMAB-3 к CD137 (SEQ ID NO: 221);
(C) VL1 hMAB-3 к CD137 (SEQ ID NO: 87);
(D) VL2 hMAB-3 к CD137 (SEQ ID NO: 88);
(E) VL3 hMAB-3 к CD137 (SEQ ID NO: 89);
(F) VL4 hMAB-3 к CD137 (SEQ ID NO: 211);
(G) VL5 hMAB-3 к CD137 (SEQ ID NO: 212);
(H) VL6 hMAB-3 к CD137 (SEQ ID NO: 213);
(I) VL7 hMAB-3 к CD137 (SEQ ID NO: 214);
(J) VL8 hMAB-3 к CD137 (SEQ ID NO: 215);
(K) VL9 hMAB-3 к CD137 (SEQ ID NO: 216);
(L) VL10 hMAB-3 к CD137 (SEQ ID NO: 217);
(M) VL11 hMAB-3 к CD137 (SEQ ID NO: 218);
(N) VL12 hMAB-3 к CD137 (SEQ ID NO: 219);
(O) VL13 hMAB-3 к CD137 (SEQ ID NO: 220);
(P) VL1 hMAB-4 к CD137 (SEQ ID NO: 94); или
(Q) VL2 hMAB-4 к CD137 (SEQ ID NO: 95).
[00041] Настоящее изобретение также относится к варианту реализации таких CD137-связывающих молекул, в которых указанная молекула представляет собой антитело или его антигенсвязывающий фрагмент.
[00042] Настоящее изобретение также относится к фармацевтической композиции, содержащей любую из описанных выше CD137-связывающих молекул и физиологически приемлемый носитель.
[00043] Настоящее изобретение также относится к применению любой из описанных выше CD137-связывающих молекул или такой фармацевтической композиции в лечении заболевания или состояния, ассоциированного с подавленной иммунной системой или характеризующегося экспрессией опухолевого антигена (ТА).
[00044] Настоящее изобретение также относится к такому применению, при котором состояние, ассоциированное с подавленной иммунной системой или характеризующееся экспрессией опухолевого антигена (ТА), представляет собой рак.
[00045] Настоящее изобретение также относится к HER2/neu-связывающей молекуле, которая содержит вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; причем:
(A) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-1 к HER2 (SEQ ID NO: 63); и
(B) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-1 к HER2 (SEQ ID NO: 62).
[00046] Настоящее изобретение также относится к варианту реализации таких HER2/neu-связывающих, молекул, причем:
(A) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(2) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(3) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL3 hMAB-1 к HER2 (SEQ ID NO: 69);
и
(B) (1) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(3) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH3 hMAB-1 к HER2 (SEQ ID NO: 66).
[00047] Настоящее изобретение также относится к варианту реализации таких HER2/neu-связывающих молекул, в которых вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(B) VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(С) VH3 hMAB-1 к HER2 (SEQ ID NO: 66)
[00048] Настоящее изобретение также относится к варианту реализации таких HER2/neu-связывающих молекул, в которых вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(B) VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(C) VL3 hMAB-1 к HER2 (SEQ ID NO: 69).
[00049] Настоящее изобретение также относится к варианту реализации таких HER2/neu-связывающих молекул, в которых молекула представляет собой антитело или его антигенсвязывающий фрагмент.
[00050] Настоящее изобретение также относится к фармацевтической композиции, содержащей любую из описанных выше HER2/neu-связывающих молекул и физиологически приемлемый носитель.
[00051] Настоящее изобретение также относится к применению любой из описанных выше HER2/neu-связывающих молекул или такой фармацевтической композиции в лечении заболевания или состояния, ассоциированного или характеризующегося экспрессией HER2/neu, и при этом, в частности, состояние, ассоциированное или характеризующееся экспрессией HER2/neu, представляет собой рак.
[00052] Настоящее изобретение также относится к способам повышения активности агента, нацеленного на опухоль, включающим введение такого агента, нацеленного на опухоль, в комбинации с любой из описанных выше CD137×TA связывающих молекул или фармацевтической композицией, содержащей их. Настоящее изобретение также относится к таким способам, дополнительно включающим введение ингибитора контрольной точки иммунного ответа PD-1/PD-L1, и при этом, в частности, такой ингибитор контрольной точки иммунного ответа представляет собой антитело к PD-1 или антитело к PD-L1. Настоящее изобретение относится, в частности, к таким способам, в которых агент, нацеленный на опухоль, представляет собой антитело, эпитопсвязывающий фрагмент антитела или агент, который опосредует перенаправленное уничтожение клетки-мишени Т-клетками.
[00053] Настоящее изобретение также относится к способам лечения заболевания или состояния, ассоциированного с подавленной иммунной системой или характеризующегося экспрессией опухолевого антигена (ТА), включающим введение субъекту, нуждающемуся в этом, любой из описанных выше CD137×TA связывающих молекул или фармацевтической композиции, содержащей их. В частности, настоящее изобретение относится к таким способам, в которых состояние, ассоциированное с подавленной иммунной системой или характеризующееся экспрессией опухолевого антигена (ТА), представляет собой рак. Настоящее изобретение также относится к таким способам, дополнительно включающим введение агента, нацеленного на опухоль, и при этом, в частности, агент, нацеленный на опухоль, представляет собой антитело, эпитопсвязывающий фрагмент антитела или агент, который опосредует перенаправленное уничтожение клетки-мишени Т-клетками. Настоящее изобретение также относится к таким способам, дополнительно включающим введение ингибитора контрольной точки иммунного ответа PD-1/PD-L1, и при этом, в частности, такой ингибитор контрольной точки иммунного ответа представляет собой антитело к PD-1 или антитело к PD-L1.
[00054] Настоящее изобретение также относится к описанным выше вариантам применения и способам, в которых рак выбран из группы, состоящей из: острого миелолейкоза, опухоли надпочечников, рака, ассоциированного со СПИДом, альвеолярной саркомы мягких тканей, астроцитарной опухоли, рака мочевого пузыря, рака костей, рака головного и спинного мозга, метастатической опухоли головного мозга, рака молочной железы, опухолей сонных артерий, рака шейки матки, хондросаркомы, хордомы, хромофобной карциномы почек, светлоклеточной карциномы, рака толстой кишки, колоректального рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, экстраскелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии костей, рака желчного пузыря или желчных протоков, рака желудка, гестационной трофобластической болезни, герминогенной опухоли, рака головы и шеи, гепатоцеллюлярной карциномы, глиобластомы, опухоли из островковых клеток, саркомы Капоши, рака почек, лейкоза, липомы/доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легких, медуллобластомы, меланомы, менингиомы, злокачественной мезотелиомы, множественной эндокринной неоплазии, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринных опухолей, не мелкоклеточного рака легких, рака яичников, рака поджелудочной железы, рака глотки, папиллярной карциномы щитовидной железы, опухоли околощитовидной железы, педиатрического рака, опухоли оболочки периферических нервов, феохромоцитомы, опухоли гипофиза, рака предстательной железы, постериорной увеальной меланомы, редкого гематологического заболевания, карциномы почек, метастатического рака почек, рабдоидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, рака яичек, рака тимуса, тимомы, метастатического рака щитовидной железы и рака матки.
[00055] В частности, настоящее изобретение относится к описанным выше вариантам применения и способам, в которых рак выбран из группы, состоящей из: рака мочевого пузыря, рака молочной железы, колоректального рака, рака желудка, глиобластомы, рака почек, рака легких, меланомы, нейробластомы, рака яичников, рака поджелудочной железы, рака глотки, рака предстательной железы, карциномы почек, рабдомиосаркомы и плоскоклеточного рака головы и шеи (SCCHN).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[00056] На Фигурах 1А-1В представлены схемы типичного ковалентно связанного диатела, имеющего два эпитопсвязывающих сайта, состоящих из двух полипептидных цепей, каждая из которых имеет домен, способствующий образованию гетеродимера, содержащий Е-спираль или К-спираль (альтернативные домены, способствующие образованию гетеродимера, представлены ниже). Остаток цистеина может присутствовать в линкере (Фигура 1А) и/или в домене, способствующем образованию гетеродимера (Фигура 1В). Домены VL и VH, которые распознают один и тот же эпитоп, показаны с использованием одного и того же шаблона затенения или заливки. Волнистая линия 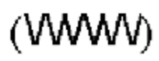 на этой и на всех Фигурах, на которых представлены схематические изображения доменов связывающей молекулы, представляет один или более необязательных доменов, способствующих образованию гетеродимера, которые предпочтительно присутствуют.
на этой и на всех Фигурах, на которых представлены схематические изображения доменов связывающей молекулы, представляет один или более необязательных доменов, способствующих образованию гетеродимера, которые предпочтительно присутствуют.
[00057] На Фигуре 2 представлена схема типичной молекулы ковалентно связанного диатела, имеющей два эпитопсвязывающих сайта, состоящих из двух полипептидных цепей, каждая из которых имеет домен СН2 и СН3, так, что объединившиеся цепи образуют полную область Fc или ее часть. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с использованием одного и того же шаблона затенения или заливки.
[00058] На Фигурах 3А-3Е представлены схемы, показывающие типичные ковалентно связанные четырехвалентные диатела, имеющие четыре эпитопсвязывающих сайта, состоящих из двух пар полипептидных цепей (т.е. в общей сложности из четырех полипептидных цепей). Одна полипептидная цепь каждой пары имеет домен СН2 и СН3, так, что объединившиеся цепи образуют полную область Fc или ее часть. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с использованием одного и того же шаблона затенения или заливки. Две пары полипептидных цепей могут быть одинаковыми. Согласно таким вариантам реализации, в которых две пары полипептидных цепей являются одинаковыми, и домены VL и VH распознают разные эпитопы (как показано на Фигурах 3А-3В), полученная молекула имеет четыре эпитопсвязывающих сайта и является биспецифической и двухвалентной по отношению к каждому связанному эпитопу. Согласно таким вариантам реализации, в которых домены VL и VH распознают один и тот же эпитоп (например, на обеих цепях применяются одни и те же CDR домена VL и одни и те же CDR домена VH), полученная молекула имеет четыре эпитопсвязывающих сайта и является моноспецифической и четырехвалентной в отношении одного эпитопа. Согласно другому варианту две пары полипептидов могут различаться. Согласно таким вариантам реализации, в которых две пары полипептидных цепей различаются, и домены VL и VH каждой пары полипептидов распознают разные эпитопы (как показано различными затенениями и шаблонами на Фигуре 3С), полученная молекула имеет четыре эпитопсвязывающих сайта и является тетраспецифической и одновалентной по отношению к каждому связанному эпитопу. На Фигуре 3А показано диатело, содержащее область Fc, которое содержит пептидный домен, способствующий образованию гетеродимера, содержащий остаток цистеина. На Фигуре 3В показано диатело, содержащее область Fc, которое содержит домены, способствующие образованию гетеродимера, содержащие Е-спираль и К-спираль, а также содержащие остаток цистеина и линкер (с необязательным остатком цистеина). На Фигуре 3С показано диатело, содержащее область Fc, которое содержит домены антитела СН1 и CL. На Фигурах 3D-3E проиллюстрировано, как отбор связывающих доменов, показанных на Фигуре 3В, может привести к получению CD137×TA связывающей молекулы, имеющей два сайта связывания, специфичных для эпитопа CD137, и два сайта связывания, специфичных для эпитопа ТА. На Фигурах 3D-3E проиллюстрировано, как можно отобрать домены для получения CD137×TA связывающих молекул, имеющих различающиеся ориентации (т.е. на Фигуре 3D домен VL CD137 применяется в качестве домена VL1 связывающей молекулы, домен VH CD137 применяется в качестве домена VH1 связывающей молекулы, домен VL ТА применяется в качестве домена VL2 связывающей молекулы и домен VH ТА применяется в качестве домена VH2 связывающей молекулы). Напротив, на Фигуре 3Е домен VL ТА применяется в качестве домена VL1 связывающей молекулы, домен VH ТА применяется в качестве домена VH1 связывающей молекулы, домен VL CD137 применяется в качестве домена VL2 связывающей молекулы и домен VH CD137 применяется в качестве домена VH2 связывающей молекулы. Как представлено ниже, сайты связывания VL/VH, образованные за счет объединения полипептидных цепей, могут быть одинаковыми или различными, чтобы обеспечить четырехвалентное связывание, которое является моноспецифическим, биспецифическим, триспецифическим или тетраспецифическим.
[00059] На Фигуре 4А и 4В представлены схемы типичной молекулы ковалентно связанного диатела, имеющей два эпитопсвязывающих сайта, состоящих из трех полипептидных цепей. Две из полипептидных цепей имеют домен СН2 и СН3 так, что объединившиеся цепи образуют полную область Fc или ее часть. Полипептидные цепи, содержащие домен VL и VH, дополнительно содержат домен, способствующий образованию гетеродимера. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с использованием одного и того же шаблона затенения или заливки.
[00060] На Фигуре 5A-5D представлены схемы типичной ковалентно связанной связывающей молекулы, имеющей четыре эпитопсвязывающих сайта, состоящих из пяти полипептидных цепей. На Фигуре 5А показана общая структура такой CD137×TA связывающей молекулы. Две из полипептидных цепей имеют домен СН2 и СН3 так, что объединившиеся цепи образуют область Fc, которая содержит полную область Fc или ее часть. Полипептидные цепи, содержащие соединенные домены VL и VH, дополнительно содержат домен, способствующий образованию гетеродимера. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с использованием одного и того же шаблона затенения или заливки. На Фигуре 5В показана структура альтернативной предпочтительной CD137×TA связывающей молекулы, в которой вариабельные домены, показанные на Фигуре 5А, отобраны для получения конечной CD137×TA связывающей молекулы, которая имеет два связывающих домена, не характерных для диатела, специфичных для иллюстративного ТА, HER2/neu, и два связывающих домена, характерных для диатела, специфичных для CD137. На Фигуре 5С показана структура альтернативной предпочтительной CD137×TA связывающей молекулы, в которой вариабельные домены, показанные на Фигуре 5А, были отобраны для получения конечной CD137×TA связывающей молекулы, которая имеет два связывающих домена, не характерных для диатела, специфичных для CD137, и два связывающих домена, характерных для диатела, специфичных для HER2/neu. На Фигуре 5D показана структура альтернативной предпочтительной CD137×TA связывающей молекулы, в которой вариабельные домены, показанные на Фигуре 5А, были отобраны для получения конечной CD137×TA связывающей молекулы, которая имеет два связывающих домена, не характерных для диатела, специфичных для эпитопа CD137, один связывающий домен, характерный для диатела, специфичный для эпитопа HER2/neu, и второй связывающий домен, характерный для диатела, специфичный для эпитопа CD137. Такие эпитопы CD137 могут быть одинаковыми или различными. Следует понимать, что при правильном отборе связывающих доменов, показанных на Фигуре 5А, любые три из связывающих доменов могли бы быть отобраны для связывания эпитопа CD137. Аналогичным образом, любые три из связывающих доменов могли бы быть отобраны для связывания эпитопа HER2/neu. Как представлено ниже, сайты связывания VL/VH, образованные за счет объединения полипептидных цепей, могут быть одинаковыми или различными, чтобы обеспечить четырехвалентное связывание, которое является моноспецифическим, биспецифическим, триспецифическим или тетраспецифическим.
[00061] На Фигуре 6A-6F представлены схемы типичных трехвалентных связывающих молекул, содержащих область Fc, имеющих три эпитопсвязывающих сайта. На Фигуре 6А схематично проиллюстрированы домены трехвалентных связывающих молекул, содержащих два связывающих домена, характерных для диатела, и связывающий домен Fab-типа, имеющих разные ориентации доменов, в которых связывающие домены, характерные для диатела, являются С-концевыми по отношению к области Fc. На Фигурах 6В-6С показана структура иллюстративной предпочтительной CD137×TA связывающей молекулы, в которой вариабельные домены, показанные на Фигурах 6А, были отобраны для получения конечной CD137×TA связывающей молекулы, которая имеет связывающие домены, не характерные для диатела, специфичные для CD137, связывающий домен, характерный для диатела, который специфичен для иллюстративного ТА, HER2/neu, и второй связывающий домен, характерный для диатела, который специфичен для CD137. На Фигуре 6D схематично проиллюстрированы домены трехвалентных связывающих молекул, содержащих два связывающих домена, характерных для диатела, и связывающий домен Fab-типа, имеющих разные ориентации доменов, в которых связывающие домены, характерные для диатела, являются С-концевыми по отношению к области Fc. Молекулы на Фигурах 6A-6D содержат четыре цепи. На Фигурах 6Е и 6F, соответственно, схематически проиллюстрированы домены трехвалентных связывающих молекул, содержащих два связывающих домена, характерных для диатела, которые являются N-концевыми по отношению к области Fc, и связывающий домен Fab-типа, в котором легкая цепь и тяжелая цепь соединены за счет полипептидного спейсера, или связывающий домен scFv-типа. Трехвалентные связывающие молекулы на Фигурах 6G и 6Н, соответственно, схематически иллюстрируют домены трехвалентных связывающих молекул, содержащих два связывающих домена, характерных для диатела, которые являются С-концевыми по отношению к области Fc, и связывающий домен Fab-типа, в котором легкая цепь и тяжелая цепь соединены за счет полипептидного спейсера, или связывающий домен scFv-типа. Трехвалентные связывающие молекулы на Фигурах 6Е-6Н содержат три цепи. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с использованием одного и того же шаблона затенения или заливки.
[00062] На Фигуре 7 показаны кривые связывания антител к CD137, урелумаба и утомилумаба (МАВ-1 к CD137 и МАВ-2 к CD137, соответственно), и нескольких новых химерных моноклональных антител (МАТ) к CD137 (chMAB-3 к CD137; chMAB-4 к CD137 и chMAB-5 к CD137) с активированными CD8+ Т-клетками.
[00063] На Фигуре 8 показана индукция секреции ИЛ-2 общей популяцией Т-клеток через 72 часа после стимуляции с применением нанесенного на подложку и высушенного антитела к CD3 (3 мкг/мл-50 мкг/мл) в присутствии антител к CD137, урелумаба или утомилумаба (МАВ-1 к CD137 или МАВ-2 к CD137, соответственно), или новых химерных антител к CD137 (chMAB-3 к CD137, chMAB-4 к CD137 или chMAB-5 к CD137), которые были сшиты с 4× hFc F(ab)'2. Сшитые антитела (Ab+αhFc) применяли в концентрации 0,1, 1,0 или 10 мкг/мл. На графике также представлены следующие контроли: стимулированная общая популяция Т-клеток, обработанных антителом для контроля изотипа, только hFc F(ab)'2 или необработанные клетки.
[00064] На Фигуре 9 показана индукция секреции ИФН-γ общей популяцией Т-клеток через 72 часа после стимуляции гранулами, направленными против CD3, в присутствии сшитых химерных антител к CD137 (chMAB-3 к CD137 или chMAB-5 к CD137) (1 мкг/мл антитела к CD-137 + 4 мкг/мл hFc F(ab)'2) ± клетки-мишени JIMT-1 (HER2/neu++), в присутствии лиганда (1 мкг/мл CD-137L-His) или без него. На графике также представлены следующие контрольные образцы: стимулированная общая популяция Т-клеток ± клетки JIMT-1, ± hFc F(ab)'2, а также необработанная/нестимулированная общая популяция Т-клеток и клеток JIMT-1.
[00065] На Фигурах 10A-10В показана способность CD137×TA связывающих молекул связываться с CD137 активированных CD8+ Т-клеток, согласно результатам измерения методом флуоресцентно-активированной сортировки клеток (FACS) с использованием FITC-меченых CD8 и АФЦ-меченой αhFc. Фигура 10А: DART-B и DART-С (содержащие домены hMAB-3 к CD137 и hMAB-1 к HER2), DART-D и DART-Е (содержащие домены МАВ-3 к CD137 и hMAB-1 к HER2) и контрольные молекулы DART-3 (содержащая домены hMAB-1 к HER2 и варианты доменов паливизумаба) и DART-6 (содержащая варианты доменов паливизумаба и домены МАВ-3 к CD137). Фигура 10В: DART-D (содержащая домены МАВ-3 к CD137 и hMAB-1 к HER2), DART-F (содержащая домены МАВ-4 к CD137 и hMAB-1 к HER2), DART-1 и DART-4 (содержащие домены МАВ-1 к CD137 и hMAB-1 к HER2), DART-2 и DART-5 (содержащие домены МАВ-2 к CD137 и hMAB-1 к HER2) и контрольная связывающая молекула DART-6 (содержащая домены МАВ-3 к CD137 и варианты доменов паливизумаба).
[00066] На Фигурах 11А-11 В показана способность CD137×TA связывающих молекул связываться с иллюстративным ТА, HER2/neu, на поверхности клеток-мишеней рака желудка N87 (HER2+++), согласно результатам измерения средней интенсивности флуоресценции (MFI). Фигура 11А: DART-В, DART-С, DART-D, DART-E и контрольные связывающие молекулы DART-3 и DART-6. Фигура 11B: DART-D, DART-F, DART-1, DART-2, DART-4, DART-5 и DART-6.
[00067] На Фигурах 12А-12В показана способность CD137×TA связывающих молекул DART-B, DART-D и DART-G, а также контрольных молекул DART-3 и DART-6 связываться с CD137, экспрессированным модифицированными клетками СНО (Фигура 12А), и CD137, экспрессированным активированными CD8+ Т-клетками (Фигура 12В).
[00068] На Фигурах 13А-13В показана способность CD137×TA связывающих молекул DART-B, DART-D и DART-G, а также контрольных молекул DART-3 и DART-6 связываться с иллюстративным ТА, HER2/neu, экспрессированным клетками-мишенями рака желудка N87 (HER2+++) (Фигура 13А) и клетками-мишенями JIMT-1 (HER2++) (Фигура 13В).
[00069] На Фигурах 14А-14В представлены результаты первого типичного анализа способности CD137×TA связывающих молекул опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (на примере высвобождения ИФН-γ). На Фигурах показаны результаты для DART-1, DART-2, DART-4, DART-5, DART-A, DART-D, DART-E и DART-F и контрольных молекул, DART-3 и DART-6, испытанных в присутствии HER2/neu-экспрессирующих K клеток-мишеней N87 (HER2+++) (Фигура 14А) или JIMT-1 (HER2++) (Фигура 14В), костимулирующая активность не была обнаружена при применении HER2/neu-отрицательных клеток-мишеней Hs700T или в отсутствие клеток-мишеней.
[00070] На Фигурах 15А-15В представлены результаты второго типичного анализа способности CD137×TA связывающих молекул опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (на примере высвобождения ИФН-γ). На Фигурах показаны результаты для DART-1, DART-2, DART-4, DART-5, DART-B, DART-D и DART-G, а также контрольных молекул DART-3 и DART-6, испытанных в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (HER2+++) (Фигура 15А) или JIMT-1 (HER2++) (Фигура 15В), костимулирующая активность не была обнаружена при применении HER2/neu-отрицательных клеток-мишеней Hs700T или в отсутствие клеток-мишеней.
[00071] На Фигурах 16А и 16В показана способность CD137×TA связывающих молекул опосредовать зависимую от дозы и мишени передачу сигналов с участием пути NF/kB в CD137-экспрессирующей линии репортерных клеток (Jurkat-NF-κB-Luc), о чем свидетельствует увеличение экспрессии люциферазы. На Фигурах показаны результаты для DART-G и контрольных связывающих молекул DART-3 и DART-6 на CD137-экспрессирующих клетках Jurkat-NF-κB-Luc, культивированных совместно с клетками JIMT-1, экспрессирующими HER2/neu (Фигура 16А), или HER2/neu-отрицательными клетками KG-1 (Фигура 16В).
[00072] На Фигуре 17, панели А-О, показана способность CD137×TA связывающих молекул повышать пролиферацию Т-клеток в совместной культуре с ТА-экспрессирующими клетками-мишенями. CFSE-меченые человеческие Т-клетки ± субоптимальная стимуляция αCD3/αCD28 культивировали совместно с клетками-мишенями HER2/neu-high N87 (Фигура 17, панели А-С и J-K), клетками-мишенями HER2/neu-low MCF-7 (Фигура 17, панели D-F и L-M) или в отсутствие клеток-мишеней (Фигура 17, панели G-I и N-O) в присутствии DART-А (Фигура 17, панели A, D и G), DART-3 (Фигура 17, панели В, Е и Н), DART-6 (Фигура 17, панели С, F и I) или без молекул (Фигура 17, панели J, L и N) и контролировали пролиферацию Т-клеток.
[00073] На Фигурах 18А-18В показана способность CD137×TA-связывающей молекулы DART-1 усиливать уничтожение клеток-мишеней N87 за счет АЗКЦ, опосредуемой антителом к ТА, маргетуксимабом, согласно результатам измерения ассоциированной с клетками активности люциферазы (Фигура 18А), и усиливать опосредуемую маргитуксимабом активацию NK-клеток, измеренную на основании экспрессии CD69 (Фигура 18В).
[00074] На Фигурах 19А-19В показаны результаты типичного анализа способности CD137×TA связывающей молекулы DART-7 и контрольной молекулы DART-8 опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (на примере высвобождения ИФН-γ). Высвобождение цитокинов измеряли в присутствии EphA2-экспрессирующих клеток-мишеней Hs700T (Фигура 19А) или EphA2-отрицательных клеток-мишеней Hs700T (EphA2.KO) (Фигура 19В) или в отсутствие клеток-мишеней. На графике также представлены следующие контрольные образцы: стимулированная общая популяция Т-клеток + клетки-мишени и необработанная/нестимулированная общая популяция Т-клеток.
[00075] На Фигурах 20А-20В показана способность производных DART-G, имеющих оптимизированные CDRH3, связываться с CD137, экспрессированным на поверхности модифицированных клеток СНО (Фигура 20А) или CD8+ Т-клеток (Фигура 20В).
[00076] На Фигурах 21А-21С показаны результаты типичного анализа способности CD137×TA связывающих молекул DART-B, DART-D, DART-G, DART-G2, DART-G3 и DART-G4, а также контрольных молекул DART-3 и DART-6, опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (на примере высвобождения ИФН-γ). Высвобождение цитокинов измеряли в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (Фигура 21А), клеток-мишеней JIMT-1 (Фигура 21В) или клеток-мишеней MDA-231 (Фигура 21С) или без клеток-мишеней.
[00077] На Фигуре 22 показаны результаты типичного анализа способности CD137×TA связывающих молекул DART-B, DART-G, DART-G1, DART-G2, DART-G3 и DART-G4, а также контрольных молекул DART-3 и DART-6, опосредовать перенаправленное уничтожение клеток-мишеней N87 (HER2+++), измеренное на основании ассоциированной с клетками активности люциферазы.
[00078] На Фигурах 23А-23С показана способность CD137×TA связывающих молекул связывать разные клетки, экспрессирующие иллюстративный ТА, HER2/neu, согласно результатам измерения средней интенсивности флуоресценции (MFI). Фигура 23А: связывание с клетками рака желудка N87 (HER2+++). Фигура 23В: связывание с клетками карциномы молочной железы JIMT-1 (HER2++). Фигура 23С: связывание с клетками рака молочной железы MCF-7 (HER2+). Связывание для TRIDENT-А, TRIDENT-A2, TRIDENT-A3, TRIDENT-A4, DART-G, DART-G2, DART-G3, DART-G4 и контрольных связывающих молекул TRIDENT-1 и TRIDENT-2 показано на обеих панелях.
[00079] На Фигурах 24А-24В показана способность CD137×TA связывающих молекул связываться с CD137, экспрессированным активированными CD8+ Т-клетками. Фигура 24А: TRIDENT-A, TRIDENT-A2, TRIDENT-A3, TRIDENT-A4 и контрольные связывающие молекулы TRIDENT-1 и TRIDENT-2. Фигура 24В: DART-G, DART-G2, DART-G3, DART-G4 и контрольные связывающие молекулы TRIDENT-1 и TRIDENT-2.
[00080] На Фигурах 25А-25С показана способность CD137×TA связывающих молекул опосредовать конъюгацию клетка-клетка между клетками СНО, экспрессирующими CD137, и клетками, экспрессирующими иллюстративный ТА, HER2/neu, FACS. Фигура 25А: конъюгация клетка-клетка с клетками рака желудка N87 (HER2+++). Фигура 25В: конъюгация клетка-клетка с клетками карциномы молочной железы JIMT-1 (HER2++). Фигура 25С: конъюгация клетка-клетка с клетками рака молочной железы MCF-7 (HER2+). Активность TRIDENT-A, TRIDENT-A2, TRIDENT-АЗ, TRIDENT-A4, DART-G4 и контрольной связывающей молекулы TRIDENT-2 показана на обеих панелях.
[00081] На Фигурах 26А-26С показаны результаты типичного анализа способности CD137×TA связывающих молекул TRIDENT-A, TRIDENT-A2, TRIDENT-A3, TRIDENT-A4, DART-G, DART-G2, DART-G3, DART-G4 и контрольных связывающих молекул TRIDENT-1 и TRIDENT-2 опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (на примере высвобождения ИФН-γ). Высвобождение цитокинов измеряли в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (Фигура 26А), клеток-мишеней JIMT-1 (Фигура 26В) или без клеток-мишеней (Фигура 26С).
[00082] На Фигуре 27 показана способность примерной CD137×TA связывающей молекулы (TRIDENT-A2) усиливать опосредуемое TA×CD3 диателом перенаправленное уничтожение клеток-мишеней Colo205/Luc Т-клетками, измеренное с помощью ассоциированной с клетками люциферазы, в сравнении с контрольной молекулой (TRIDENT-1).
[00083] На Фигурах 28А-28С показана экспрессия маркеров Т-клеток, присутствующих в 1 мг опухоли, отобранном после обработки контрольной средой ( •); TRIDENT-A2  TA×CD3 диателом
TA×CD3 диателом  комбинацией TRIDENT-A2 и диатела TA×CD3
комбинацией TRIDENT-A2 и диатела TA×CD3  МАТ к PD-1
МАТ к PD-1  или комбинацией TRIDENT-A2, диатела TA×CD3 и МАТ к PD-1. Фигура 28А: экспрессия CD4. Фигура 28В: экспрессия CD8. Фигура 28С: экспрессия CD69. Фигура 28D: экспрессия PD-1.
или комбинацией TRIDENT-A2, диатела TA×CD3 и МАТ к PD-1. Фигура 28А: экспрессия CD4. Фигура 28В: экспрессия CD8. Фигура 28С: экспрессия CD69. Фигура 28D: экспрессия PD-1.
[00084] На Фигуре 29 показана способность контрольной среды (•); TRIDENT-А2  TA×CD3 диатела
TA×CD3 диатела  комбинации TRIDENT-A2 и TA×CD3 диатела
комбинации TRIDENT-A2 и TA×CD3 диатела  МАТ к PD-1 и комбинации TRIDENT-А2, диатела и МАТ к PD-1 ингибировать рост опухоли или размножение клеток карциномы яичников в МКПК-восстановленной ксенотрансплантатной модели у мышей.
МАТ к PD-1 и комбинации TRIDENT-А2, диатела и МАТ к PD-1 ингибировать рост опухоли или размножение клеток карциномы яичников в МКПК-восстановленной ксенотрансплантатной модели у мышей.
[00085] На Фигуре 30 показана способность CD137×TA связывающих молекул TRIDENT-A2, TRIDENT-B2 и контрольных связывающих молекул TRIDENT-1 и TRIDENT-2 опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками яванской макаки (на примере высвобождения ИФН-γ). Высвобождение цитокинов измеряли в присутствии клеток-мишеней JIMT-1, экспрессирующих 5Т4 и HER2/neu.
[00086] На Фигурах 31А-31В показаны аминокислотные последовательности VH1B hMAB-3 к CD137 (Фигура 30А, SEQ ID NO: 84) и VL3 hMAB-3 к CD137 (Фигура 30В, SEQ ID NO: 89). Остатки CDR подчеркнуты. Положения замен заключены в рамку и номера согласно Kabat указаны стрелками; последовательная нумерация аминокислотных остатков указана над последовательностями.
[00087] На Фигурах 32А-32В показана способность антител к CD137, содержащих деиммунизированные домены VH и VL, связываться с CD137 активированных CD4+ Т-клеток или CD8+ Т-клеток, согласно результатам измерения методом FACS с использованием V510-меченых CD4+ и αhFc АФЦ (Фигура 32А) или FITC-меченых CD8 и αhFc АФЦ (Фигура 32В).
[00088] На Фигурах 33А-33В показана индукция ИФН-γ общей популяцией Т-клеток через 72 часа после стимуляции нанесенным на подложку и высушенным антителом к CD3 (3 мкг/мл-50 мкг/мл) в присутствии оптимизированных/деиммунизированных антител к CD137, МАВ-3 к CD137 (1b.3), МАВ-3 к CD137 (1Е.15) и МАВ-3 к CD137 (1G.15), в отсутствие сшивания (Фигура 33А) или сшитых с 4× hFc F(ab)'2 (Фигура 33В).
[00089] На Фигурах 34А-34С показана способность CD137×TA связывающих молекул TRIDENT-В2, TRIDENT-B5 и контрольных молекул TRIDENT-1, TRIDENT-3 и TRIDENT-4 связываться с CD137 на поверхности активированных CD8+ Т-клеток, согласно результатам измерения методом FACS с использованием FITC-меченых CD8 и αhFc АФЦ (Фигура 34А), и с иллюстративным ТА, 5Т4, на поверхности клеток-мишеней JIMT-1 (5T4hi) (Фигура 34В) или SKOV-3 (5Т4lo) (Фигура 34С).
[00090] На Фигурах 35А-35С показана способность CD137×TA связывающих молекул опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (на примере высвобождения ИФН-γ). На Фигурах показаны результаты для TRIDENT-B2, TRIDENT-В5 и контрольных молекул TRIDENT-1, TRIDENT-3 и TRIDENT-4, испытанных в присутствии 5Т4-экспрессирующих клеток-мишеней JIMT-1 (5T4hi) (Фигура 35А) или SKOV-3 (5T4lo) (Фигура 35В), костимулирующая активность не была обнаружена в отсутствие клеток-мишеней (Фигура 35С).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00091] Настоящее изобретение относится к связывающим молекулам, которые имеют один или более эпитопсвязывающих сайтов, специфичных для эпитопа CD137, и один или более эпитопсвязывающих сайтов, специфичных для эпитопа опухолевого антигена («ТА») (например, «CD137×TA связывающая молекула»). Согласно одному варианту реализации настоящего изобретения такие CD137×TA связывающие молекулы будут представлять собой биспецифические молекулы, в частности, биспецифические четырехвалентные диатела, которые состоят из двух, трех, четырех или более четырех полипептидных цепей и имеют два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа CD137, и два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа ТА. Согласно другому варианту такие CD137×TA связывающие молекулы будут представлять собой биспецифические молекулы, в частности, биспецифические трехвалентные связывающие молекулы, состоящие из трех или более полипептидных цепей и имеющие один или два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа CD137, и один или два эпитопсвязывающих сайта, каждый из которых специфичен для эпитопа ТА. CD137×TA связывающие молекулы согласно настоящему изобретению способны одновременно связываться с CD137 и ТА. Настоящее изобретение относится к фармацевтическим композициям, которые содержат любые такие CD137×TA связывающие молекулы. Настоящее изобретение дополнительно относится к способам применения таких молекул в лечении рака и других заболеваний и состояний. Согласно настоящему изобретению также предложены новые CD137-связывающие молекулы и HER2/neu-связывающие молекулы, а также их производные и варианты их применения.
I. Антитела и другие связывающие молекулы
[00092] Антитела представляют собой молекулы иммуноглобулинов, способные специфично связываться с областью-мишенью («эпитопом») молекулы, такой как углевод, полинуклеотид, липид, полипептид и т.д. («антиген»), за счет по меньшей мере одного «эпитопсвязывающего сайта», расположенного в вариабельной области молекулы иммуноглобулина. В настоящем документе термины «антитело» и «антитела» относятся к моноклональным антителам, полиспецифическим антителам, человеческим антителам, гуманизированным антителам, синтетическим антителам, химерным антителам, поликлональным антителам, верблюжьим антителам, одноцепочечным Fv (scFv), одноцепочечным антителам, фрагментам Fab, фрагментам F(ab'), дисульфидсвязанным биспецифическим Fv (sdFv), интрателам и эпитопсвязывающим фрагментам любого из указанных выше. В частности, термин «антитело» включает молекулы иммуноглобулинов и иммунологически активные фрагменты молекул иммуноглобулинов, т.е. молекулы, которые содержат эпитопсвязывающий сайт. Молекулы иммуноглобулинов могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса. Антитела способны «иммуноспецифично связываться» с полипептидом или белком или небелковой молекулой благодаря присутствию на такой молекуле определенного домена, фрагмента или конформации («эпитоп»). В настоящем документе термин «эпитопсвязывающий фрагмент антитела» предназначен для обозначения части антитела, способной иммуноспецифично связываться с эпитопом. В настоящем документе термин охватывает фрагменты (такие как Fab, Fab', F(ab')2, Fv) и одноцепочечный фрагмент (scFv), а также эпитопсвязывающий домен диатела. В настоящем документе антитело или его эпитопсвязывающий фрагмент, о которых говорят, что они «иммуноспецифично» связывают область другой молекулы (т.е. эпитоп), если они реагируют или ассоциируются чаще, быстрее, более длительно и/или с большей аффинностью или авидностью с этим эпитопом по сравнению с альтернативными эпитопами. Из этого определения также понятно, что, например, антитело или его эпитопсвязывающий фрагмент, который иммуноспецифично связывается с первой мишенью, может специфично или предпочтительно связываться со второй мишенью или может не связываться с ней. Эпитопсодержащая молекула может обладать иммуногенной активностью так, что она вызывает выработку антител у животного; такие молекулы называются «антигены». Природные антитела способны связываться только с одним типом эпитопов (т.е. они являются «моноспецифическими»), хотя они могут связывать несколько копий эпитопа этого типа (т.е. проявлять «двухвалентность» или «поливалентность»).
[00093] Термин «моноклональное антитело» относится к гомогенной популяции антител, в которой моноклональное антитело состоит из аминокислот (природных или неприродных), которые участвуют в селективном связывании антигена. Моноклональные антитела являются высокоспецифичными и направлены против одного эпитопа (или антигенного сайта). Термин «моноклональное антитело» охватывает не только интактные моноклональные антитела и полноразмерные моноклональные антитела, но также их фрагменты (такие как Fab, Fab', F(ab')2, Fv), одноцепочечные фрагменты (scFv), их мутантов, гибридные белки, содержащие часть антитела, гуманизированные моноклональные антитела, химерные моноклональные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена с требуемой специфичностью и способностью связываться с антигеном. Этот термин не является ограничивающим в отношении источника антитела или способа его получения (например, с помощью гибридомы, отбора фагов, рекомбинантной экспрессии, трансгенных животных и т.д.). Термин включает полные иммуноглобулины, а также фрагменты и т.д., описанные выше под определением «антитело». Способы получения моноклональных антител известны в данной области техники. Одним из способов, которые можно применять, является способ Kohler, G. et al. (1975) «Continuous Cultures Of Fused Cells Secreting Antibody Of Predefined Specificity», Nature 256:495-497 или его модификация. Как правило, моноклональные антитела вырабатываются у мышей, крыс или кроликов. Антитела получают путем иммунизации животного иммуногенным количеством клеток, клеточных экстрактов или белковых препаратов, которые содержат желательный эпитоп. Иммуноген может представлять собой, но не ограничивается указанными, первичные клетки, культивируемые клеточные линии, раковые клетки, белки, пептиды, нуклеиновые кислоты или ткани. Клетки, применяемые для иммунизации, можно культивировать в течение некоторого периода времени (например, по меньшей мере 24 часа) перед их применением в качестве иммуногена. Клетки можно применять в качестве иммуногенов сами по себе или в комбинации с неденатурирующим адъювантом, таким как Ribi (см., например, Jennings, V.M. (1995) «Review of Selected Adjuvants Used in Antibody Productions, ILAR J. 37(3): 119-125). В целом клетки необходимо поддерживать в интактном и предпочтительно жизнеспособном состоянии при применении в качестве иммуногенов. Интактные клетки могут обеспечить лучшее детектирование антигенов иммунизированным животным, чем разрушенные клетки. Применение денатурирующих или агрессивных адъювантов, например, адъюванта Фрейнда, может привести к разрыву клеток и, следовательно, не рекомендуется. Иммуноген можно вводить несколько раз с периодическими интервалами, такими как два раза в неделю или один раз в неделю, или можно вводить так, чтобы поддерживать жизнеспособность у животного (например, в тканевом рекомбинанте). Согласно другому варианту существующие моноклональные антитела и любые другие эквивалентные антитела, которые являются иммуноспецифичными для желательного патогенного эпитопа, могут быть секвенированы и получены рекомбинантно с помощью любого способа, известного в данной области техники. Согласно одному варианту реализации настоящего изобретения такое антитело секвенируют, и полинуклеотидную последовательность затем клонируют в вектор для экспрессии или размножения. Последовательность, кодирующую антитело, представляющее интерес, можно поддерживать в векторе в клетке-хозяине, и клетка-хозяин затем может быть размножена и заморожена для будущего применения. Полинуклеотидную последовательность таких антител можно применять для генетической манипуляции, чтобы получить моноспецифические или полиспецифические (например, биспецифические, триспецифические и тетраспецифические) молекулы согласно настоящему изобретению, а также оптимизированные по аффинности антитела, химерные антитела, гуманизированные антитела и/или канинизированные антитела, чтобы улучшить аффинность или другие характеристики антитела. Общий принцип гуманизации антитела включает сохранение основной последовательности эпитопсвязывающей части антитела и замену оставшейся части антитела из вида, отличного от человека, последовательностями человеческих антител. Существует четыре основных этапа гуманизации моноклонального антитела. Они включают: (1) определение нуклеотидной и/или предсказанной аминокислотной последовательности вариабельных доменов легкой и тяжелой цепей исходного антитела; (2) конструирование гуманизированного антитела или канинизированного антитела, т.е. принятие решения о том, какой(ие) каркасный(ые) участок (участки) антитела применять в процессе гуманизации или канинизации; (3) применение современных методологий/методик гуманизации или канинизации; и (4) трансфекцию и экспрессию гуманизированного или канинизированного антитела (см., например, патенты США №4816567; 5807715; 5866692; и 6331415).
[00094] В последние несколько десятилетий наблюдается возобновление интереса к терапевтическому потенциалу антител, и антитела стали одним из ведущих классов биотехнологических лекарственных препаратов (Chan, С.Е. et at. (2009) «The Use Of Antibodies In The Treatment Of Infectious Diseases», Singapore Med. J. 50(7):663-666). Более 200 препаратов на основе антител были одобрены для применения или находятся на этапе разработки.
А. Общие структурные признаки антител
[00095] Основная структурная единица природных иммуноглобулинов (например, IgG) представляет собой тетрамер, состоящий из двух более коротких «легких цепей», образующих комплекс с двумя более длинными «тяжелыми цепями», и обычно экспрессируется в виде гликопротеина массой приблизительно 150000 Да. Каждая цепь состоит из амино-концевой («N-концевой») части, которая содержит «вариабельный домен», и карбокси-концевой («С-концевой») части, которая содержит по меньшей мере один «константный домен». Легкая цепь IgG состоит из одного «вариабельного домена легкой цепи» («VL») и одного «константного домена легкой цепи» («CL»). Таким образом, структура легких цепей молекулы IgG представляет собой n-VL-CL-c (где n и с представляют, соответственно, N-конец и С-конец полипептида). Тяжелая цепь IgG состоит из одного «вариабельного домена тяжелой цепи» («VH»), трех «константных доменов тяжелой цепи» («СН1», «СН2» и «СН3») и «шарнирной» области («Н»), расположенной между доменами СН1 и СН2. Таким образом, структура тяжелой цепи IgG представляет собой n-VH-CH1-H-CH2-CH3-c (где n и с представляют, соответственно, N-конец и С-конец полипептида). Способность интактного немодифицированного антитела (например, антитела IgG) связывать эпитоп антигена зависит от присутствия и последовательностей вариабельных доменов.
1. Константные домены
(a) Константный домен легкой цепи
[00096] Предпочтительный домен CL представляет собой каппа-домен CL человеческого IgG. Аминокислотная последовательность примерного человеческого каппа-домена CL представляет собой (SEQ ID NO: 1):

[00097] Согласно другому варианту примерный домен CL представляет собой лямбда-домен человеческого IgG. Аминокислотная последовательность примерного человеческого лямбда-домена CL представляет собой (SEQ ID NO: 2):

(b) Домены CH1 тяжелой цепи
[00098] Примерный домен CH1 представляет собой домен СН1 человеческого IgG1. Аминокислотная последовательность примерного домена СН1 человеческого IgG1 представляет собой (SEQ ID NO: 3):

[00099] Примерный домен CH1 представляет собой домен CH1 человеческого IgG2. Аминокислотная последовательность примерного домена СН1 человеческого IgG2 представляет собой (SEQ ID NO: 4):
[000100] Примерный домен CH1 представляет собой домен CH1 человеческого IgG3. Аминокислотная последовательность примерного домена СН1 человеческого IgG3
представляет собой (SEQ ID NO: 5):

[000101] Примерный домен CH1 представляет собой домен СН1 человеческого IgG4. Аминокислотная последовательность примерного домена СН1 человеческого IgG4 представляет собой (SEQ ID NO: 6):

(c) Шарнирные области тяжелой цепи
[000102] Примерная шарнирная область представляет собой шарнирную область человеческого IgG1. Аминокислотная последовательность примерной шарнирной области человеческого IgG1 представляет собой (SEQ ID NO: 7):

[000103] Другой пример шарнирной области представляет собой шарнирную область человеческого IgG2. Аминокислотная последовательность примерной шарнирной области человеческого IgG2 представляет собой (SEQ ID NO: 8):

[000104] Другая примерная шарнирная область представляет собой шарнирную область человеческого IgG3. Аминокислотная последовательность примерной шарнирной области человеческого IgG3 представляет собой (SEQ ID NO: 9):

[000105] Другая примерная шарнирная область представляет собой шарнирную область человеческого IgG4. Аминокислотная последовательность примерной шарнирной области человеческого IgG4 представляет собой (SEQ ID NO: 10):

[000106] Как описано в настоящем документе, шарнирная область IgG4 может содержать стабилизирующую мутацию, такую как замена S228P (пронумерованную с использованием индекса ЕС, как изложено в Kabat). Аминокислотная последовательность примерной стабилизированной шарнирной области IgG4 представляет собой (SEQ ID NO: 11):

(d) Домены СН2 и СН3 тяжелой цепи
[000107] Домены СН2 и СН3 двух тяжелых цепей взаимодействуют с образованием «области Fc» антител IgG, которая распознается клеточными рецепторами Fc, включая, но не ограничиваясь указанными, рецепторы Fc-гамма (FcγR). В настоящем документе термин «область Fc» используется для определения С-концевой области тяжелой цепи IgG. Часть области Fc (включая часть, которая охватывает всю область Fc) называется в настоящем документе «домен Fc». Область Fc, о которой говорят, что она относится к определенному изотипу, классу или подклассу IgG, если ее аминокислотная последовательность наиболее гомологична этому изотипу по сравнению с другими изотипами IgG. Было показано, что в дополнение к известным вариантам применения антител в диагностике их можно применять в качестве терапевтических агентов.
[000108] Аминокислотная последовательность домена СН2-СН3 примерного человеческого IgG1 представляет собой (SEQ ID NO: 12):

пронумерованную с использованием индекса ЕС, как изложено в Kabat, где X представляет собой лизин (K) или отсутствует.
[000109] Аминокислотная последовательность домена СН2-СН3 примерного человеческого IgG2 представляет собой (SEQ ID NO: 13):


пронумерованную с использованием индекса ЕС, как изложено в Kabat, где X представляет собой лизин (K) или отсутствует.
[000110] Аминокислотная последовательность домена СН2-СН3 примерного человеческого IgG3 представляет собой (SEQ ID NO: 14):

пронумерованную с использованием индекса ЕС, как изложено в Kabat, где X представляет собой лизин (K) или отсутствует.
[000111] Аминокислотная последовательность домена СН2-СН3 примерного человеческого IgG4 представляет собой (SEQ ID NO: 15):


пронумерованную с использованием индекса ЕС, как изложено в Kabat, где X представляет собой лизин (K) или отсутствует.
[000112] На всем протяжении настоящего описания нумерация остатков в константной области тяжелой цепи IgG соответствует нумерации с использованием индекса ЕС, как изложено в Kabat et al., SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST, 5th Ed. Public Health Service, NH1, MD (1991) («Kabat»), в явной форме включенной в настоящий документ посредством ссылки. Термин «индекс ЕС, как изложено в Kabat» относится к нумерации константных доменов человеческого антитела IgG1 ЕС.
[000113] Полиморфизмы наблюдали в ряде различных положений в константных областях антитела (например, в положениях Fc, включая, но не ограничиваясь указанными, положения 270, 272, 312, 315, 356 и 358, в соответствии с системой нумерации с использованием индекса ЕС, изложенной в Кабат), и, соответственно, могут существовать небольшие различия между представленной последовательностью и последовательностями, известными из предшествующего уровня техники. Хорошо известны полиморфные формы иммуноглобулинов человека. В настоящее время известно 18 GM-аллотипов: G1m (1, 2, 3, 17) или G1m (а, х, f, z), G2m (23) или G2m (n), G3m (5, 6, 10, 11, 13, 14, 15, 16, 21, 24, 26, 27, 28) или G3m (b1, с3, b3, b0, b3, b4, s, t, g1, c5, u, v, g5) (Lefranc, et al., «The Human IgG Subclasses: Molecular Analysis Of Structure, Function And Regulation)). Pergamon, Oxford, pp. 43-78 (1990); Lefranc, G. et al., 1979, Hum. Genet.: 50, 199-211). Конкретно предусмотрено, что антитела согласно настоящему изобретению могут включать любой аллотип, изоаллотип или гаплотип любого гена иммуноглобулина и не ограничены аллотипом, изоаллотипом или гаплотипом последовательностей, обеспеченных в настоящем документе. Помимо этого в некоторых системах экспрессии С-концевой остаток аминокислоты (выделенный жирным шрифтом) домена СН3 может быть удален после трансляции. Соответственно, С-концевой остаток домена СН3 представляет собой необязательный остаток аминокислоты в CD137×TA связывающих молекулах согласно настоящему изобретению. Настоящее изобретение конкретно охватывает CD137×TA связывающие молекулы, лишенные С-концевого остатка домена СН3. Настоящее изобретение также конкретно охватывает такие конструкции, содержащие С-концевой остаток лизина домена СН3.
2. Вариабельные домены
[000114] Вариабельные домены молекулы IgG состоят из трех «гипервариабельных участков» («CDR»), которые содержат аминокислотные остатки антитела, которые будут вступать в контакт с эпитопом, а также промежуточные сегменты, отличные от CDR, называемые «каркасные участки» («FR»), которые в целом поддерживают структуру и определяют расположение петель CDR, чтобы обеспечить такое контактирование (несмотря на то, что некоторые каркасные остатки также могут связываться с эпитопом). Таким образом, домены VL и VH имеют структуру n-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4-c. Аминокислотные последовательности CDR определяют, сможет ли антитело связываться с конкретным эпитопом. Взаимодействие легкой цепи антитела с тяжелой цепью антитела и, в частности, взаимодействие их доменов VL и VH, образует эпитопсвязывающий сайт антитела.
[000115] Аминокислоты из вариабельных доменов зрелых тяжелых и легких цепей иммуноглобулинов обозначены положением аминокислоты в цепи. Kabat (SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST, 5th Ed. Public Health Service, NH1, MD (1991)) описал многочисленные аминокислотные последовательности для антител, идентифицировал консенсусную аминокислотную последовательность для каждой подгруппы и присвоил номер остатка для каждой аминокислоты, и CDR и FR идентифицированы, как определено Kabat (следует понимать, что CDRH1, как определено Chothia, С.& Lesk, А.М. ((1987) «Canonical Structures For The Hypervariable Regions Of Immunoglobulins», J. Mol. Biol. 196:901-917), начинается на пять остатков ранее). Схема нумерации Kabat может быть расширена на антитела, не включенные в его сборник, путем сопоставления рассматриваемого антитела с одной из консенсусных последовательностей в Kabat по отношению к консервативным аминокислотам. Этот способ присвоения номеров остатков стал стандартным в данной области техники и позволяет легко идентифицировать аминокислоты в эквивалентных положениях в разных антителах, включая химерные или гуманизированные варианты. Например, аминокислота в положении 50 легкой цепи антитела человека занимает положение, эквивалентное таковому для аминокислоты в положении 50 легкой цепи мышиного антитела.
[000116] Полипептиды, которые представляют собой (или могут служить в качестве них) первый, второй и третий CDR легкой цепи антитела обозначены в настоящем документе, соответственно, как: домен CDRL1, домен CDRL2 и домен CDRL3. Аналогичным образом, полипептиды, которые представляют собой (или могут служить в качестве них) первый, второй и третий CDR тяжелой цепи антитела обозначены в настоящем документе, соответственно, как: домен CDRH1, домен CDRH2 и домен CDRH3. Таким образом, термины домен CDRL1, домен CDRL2, домен CDRL3, домен CDRH1, домен CDRH2 и домен CDRH3 относятся к полипептидам, которые при включении в белок придают этому белку способность связываться со специфическим эпитопом, независимо от того, является ли такой белок антителом, имеющим легкие и тяжелые цепи, или диателом или одноцепочечной связывающей молекулой (например, scFv, BiTe и т.д.), или представляет собой белок другого типа. Соответственно, в настоящем документе термин «эпитопсвязывающий фрагмент» обозначает фрагмент молекулы, способный иммуноспецифично связываться с эпитопом. Эпитопсвязывающий фрагмент может содержать любые 1, 2, 3, 4 или 5 доменов CDR антитела или может содержать все 6 из доменов CDR антитела и, несмотря на то, что он способен иммуноспецифично связываться с таким эпитопом, может проявлять иммуноспецифичность, аффинность или селективность в отношении такого эпитопа, которые отличаются от аналогичных показателей такого антитела. Однако предпочтительно эпитопсвязывающий фрагмент будет содержать все 6 из доменов CDR такого антитела. Эпитопсвязывающий фрагмент антитела может представлять собой одну полипептидную цепь (например, scFv) или может содержать две или более полипептидных цепей, каждая из которых имеет амино- конец и карбокси-конец (например, диатело, фрагмент Fab, фрагмент Fab2 и т.д.). Если специально не указано иное, порядок доменов белковых молекул, описанных в настоящем документе, имеет направление «от N-конца к С-концу».
[000117] Эпитопсвязывающий сайт может содержать либо полный вариабельный домен, гибридизованный с константными доменами, либо только гипервариабельные участки (CDR) такого вариабельного домена, привитые на соответствующие каркасные участки. Эпитопсвязывающие сайты могут быть дикого типа или могут быть модифицированы одной или более заменами аминокислот. Это исключает константную область в качестве иммуногена у человека, однако сохраняется возможность иммунного ответа на чужеродный вариабельный домен (LoBuglio, A.F. et al. (1989) «Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Responses, Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224). Другой подход сосредоточен не только на обеспечении константных областей человеческого происхождения, но и на модификации вариабельных доменов, чтобы как можно более точно изменить их на человеческую форму. Известно, что вариабельные домены тяжелых и легких цепей содержат три гипервариабельных участка (CDR), которые изменяются в зависимости от рассматриваемых антигенов и определяют способность к связыванию, фланкированных четырьмя каркасными участками (FR), которые являются относительно консервативными у данного вида и которые предположительно обеспечивают каркас для CDR. При получении антител из вида, отличного от человека, по отношению к определенному антигену, вариабельные домены могут быть «преобразованы» или «гуманизированы» путем прививки CDR, полученных из антител из видов, отличных от человека, на FR, присутствующие в человеческом антителе, подлежащем модификации. Применение упомянутого подхода к различным антителам было описано Sato, K. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) «Reshaping Human Antibodies for Therapy», Nature 332:323-327; Verhoeyen, M. et al. (1988) «Reshaping Human Antibodies: Grafting An Antilysozyme Activity», Science 239:1534-1536; Kettleborough, C.A. et al. (1991) «Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformations», Protein Engineering 4:773-3783; Maeda, H. et al. (1991) «Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity», Human Antibodies Hybridoma 2:124-134; Gorman, S.D. et al. (1991) «Reshaping A Therapeutic CD4 Antibody», Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) «Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo», Bio/Technology 9:266-271; Co, M.S. et al. (1991) «Humanized Antibodies For Antiviral Therapy», Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) «Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy», Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; и Co, M.S. et al. (1992) «Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen», J. Immunol. 148:1149-1154. Согласно некоторым вариантам реализации настоящего изобретения гуманизированные антитела сохраняют все последовательности CDR (например, гуманизированное мышиное антитело, которое содержит все шесть CDR из мышиных антител). Согласно другим вариантам реализации настоящего изобретения гуманизированные антитела содержат один или более CDR (один, два, три, четыре, пять или шесть), последовательности которых отличаются в сравнении с исходным антителом.
В. Гуманизация антител
[000118] В частности, настоящее изобретение охватывает связывающие молекулы (включая антитела и диатела), которые содержат домен VL и/или VH гуманизированного антитела. Термин «гуманизированное» антитело относится к химерной молекуле, обычно полученной с использованием рекомбинантных методик, содержащей эпитопсвязывающий сайт иммуноглобулина из вида, отличного от человека, и остальную иммуноглобулиновую структуру молекулы, которая основана на структуре и/или последовательности человеческого иммуноглобулина. Полинуклеотидная последовательность вариабельных доменов указанных антител может быть использована для генетической манипуляции, чтобы получить указанные производные и улучшить аффинность или другие характеристики указанных антител. Общий принцип гуманизации антитела включает сохранение основной последовательности эпитопсвязывающей части антитела с заменой оставшейся части антитела из вида, отличного от человека, последовательностями человеческих антител. Существуют четыре общих этапа гуманизации моноклонального антитела. Указанные этапы включают: (1) определение нуклеотидной и предсказанной аминокислотной последовательности исходных вариабельных доменов легких и тяжелых цепей антитела, (2) конструирование гуманизированного антитела или канинизированного антитела, т.е. принимается решение о том, какой каркасный участок антитела будет применяться в процессе гуманизации или канинизации, (3) современные методологии/методики гуманизации или канинизации, и (4) трансфекцию и экспрессию гуманизированного антитела. См., например, патенты США №№4816567; 5807715; 5866692; и 6331415.
[000119] Был описан ряд молекул гуманизированных антител, содержащих эпитопсвязывающий сайт, происходящий из иммуноглобулина из вида, отличного от человека, включая химерные антитела, содержащие вариабельный домен грызуна или модифицированный вариабельный домен грызуна, и связанные с ними гипервариабельные участки (CDR), гибридизованные с человеческими константными доменами (см. например, Winter et al. (1991) «Man-made Antibodies», Nature 349:293-299; Lobuglio et al. (1989) «Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Responses», Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224 (1989), Shaw et al. (1987) «Characterization Of A Mouse/Human Chimeric Monoclonal Antibody (17-1A) To A Colon Cancer Tumor-Associated Antigen», J. Immunol. 138:4534-4538, и Brown et al. (1987) «Tumor-Specific Genetically Engineered Murine/Human Chimeric Monoclonal Antibody», Cancer Res. 47:3577-3583). В других ссылках описаны CDR грызунов, привитые в поддерживающий каркасный участок человека (FR), перед гибридизацией с соответствующим константным доменом человека (см. например, Riechmann, L. et al. (1988) «Reshaping Human Antibodies for Therapy», Nature 332:323-327; Verhoeyen, M. et al. (1988) «Reshaping Human Antibodies: Grafting An Antilysozyme Activity», Science 239:1534- 1536; и Jones et al. (1986) «Replacing The Complementarity-Determining Regions In A Human Antibody With Those From A Mouse», Nature 321:522-525). В другой ссылке описаны CDR грызунов, поддерживаемые каркасными участками грызунов после рекомбинации поверхностных остатков. См., например, публикацию европейского патента №519596. Эти «гуманизированные» молекулы предназначены для минимизации нежелательного иммунологического ответа на молекулы антител грызунов к белкам человека, что ограничивает продолжительность и эффективность терапевтического применения указанных фрагментов у реципиентов-людей. Другие способы гуманизации антител, которые также можно применять, раскрыты Daugherty et al. (1991) «Polymerase Chain Reaction Facilitates The Cloning, CDR-Grafting, And Rapid Expression Of A Murine Monoclonal Antibody Directed Against The CD18 Component Of Leukocyte Integrins», Nucl. Acids Res. 19:2471-2476, и в патентах США №№6180377; 6054297; 5997867; и 5866692.
[000120] Несмотря на такие успехи, получение стабильных, функциональных гетер од имерных немоноспецифических диател, оптимизированных для терапевтического применения, может быть дополнительно улучшено с помощью тщательного рассмотрения и размещения доменов, применяемых в полипептидных цепях. Соответственно, настоящее изобретение относится к обеспечению специфических полипептидов, которые специально разработаны для образования, за счет ковалентных связей, стабильных и терапевтически пригодных гетеродимерных диател и гетеродимерных Fc-диател, которые способны одновременно связывать CD137 и ТА.
С. Биспецифические антитела, полиспецифические диатела и диатела DART®
[000121] Как указано выше, природные антитела способны связываться только с одним типом эпитопов, хотя они могут связывать несколько копий этого типа. В данной области техники существует потребность в получении биспецифических антител, и был разработан широкий спектр форматов рекомбинантных биспецифических антител для получения таких биспецифических антител (см., например, РСТ публикации №№ WO 2008/003116, WO 2009/132876, WO 2008/003103, WO 2007/146968, WO 2009/018386, WO 2012/009544, WO 2013/070565). В большинстве таких подходов применяют линкерные пептиды, чтобы гибридизовать дополнительный связывающий фрагмент (например, scFv, VL, VH и т.д.) с центральной частью антитела или в ее пределах (IgA, IgD, IgE, IgG или IgM) или гибридизовать несколько связывающих частей антител друг с другом (например, два фрагмента Fab или scFv). В других форматах применяют линкерные пептиды, чтобы гибридизовать связывающий белок (например, scFv, VL, VH и т.д.) с доменом димеризации, таким как домен СН2-СН3 или альтернативными полипептидами (WO 2005/070966, WO 2006/107786А, WO 2006/107617А, WO 2007/046893). Как правило, такие подходы включают компромиссные варианты и приемлемые варианты. Например, в РСТ публикациях WO 2013/174873, WO 2011/133886 и WO 2010/136172 раскрыто, что применение линкеров может вызвать проблемы в терапевтических условиях, и описано триспецифическое антитело, в котором домены CL и СН1 передвинуты с их соответствующих природных положений, и домены VL и VH были диверсифицированы (WO 2008/027236, WO 2010/108127), чтобы придать им способность связываться более чем с одним антигеном. Соответственно, в молекулах, раскрытых в указанных документах, специфичность связывания изменена в пользу способности связываться с дополнительными видами антигенов. В РСТ публикациях WO 2013/163427 и WO 2013/119903 раскрыта модификация домена СН2 для включения аддукта гибридного белка, содержащего связывающий домен. В документе отмечено, что домен СН2, вероятно, играет лишь минимальную роль в качестве посредника эффекторной функции. В РСТ публикациях WO 2010/028797, WO2010028796 и WO 2010/028795 раскрыты рекомбинантные антитела, области Fc которых были заменены дополнительными доменами VL и VH так, чтобы получить трехвалентные связывающие молекулы. В РСТ публикациях WO 2003/025018 и WO2003012069 раскрыты рекомбинантные диатела, отдельные цепи которых содержат домены scFv. В РСТ публикации WO 2013/006544 раскрыты поливалентные молекулы Fab, которые синтезируют как одну полипептидную цепь и затем подвергают протеолизу с получением гетеродимерных структур. Соответственно, в молекулах, раскрытых в указанных документах, все или некоторые функциональные возможности выступать посредниками эффекторной функции изменены в пользу способности связываться с дополнительными типами антигенов. В РСТ публикациях WO 2014/022540, WO 2013/003652, WO 2012/162583, WO 2012/156430, WO 2011/086091, WO 2008/024188, WO 2007/024715, WO 2007/075270, WO 1998/002463, WO 1992/022583 и WO 1991/003493 раскрыты способы добавления дополнительных связывающих доменов или функциональных групп к антителу или части антитела (например, добавление диатела к легкой цепи антитела или добавление дополнительных доменов VL и VH к легким и тяжелым цепям антитела, или добавление гетерологичного гибридного белка, или связывание нескольких доменов Fab друг с другом). Соответственно, в молекулах, раскрытых в указанных документах, нативная структура антител изменена в пользу способности связываться с дополнительными типами антигенов.
[000122] В данной области техники также упомянута возможность получения диател, которые отличаются от указанных природных антител своей способностью связывать два или более различных типов эпитопов (т.е. проявляющих биспецифичность или полиспецифичность в дополнение к двухвалентности или поливалентности) (см., например, Holliger et al. (1993) «'Diabodies': Small Bivalent And Bispecific Antibody Fragments», Proc. Natl. Acad. Sci. (U.S.A.) 90:6444-6448; US 2004/0058400 (Hollinger et al.); US 2004/0220388 (Mertens et al.); Alt et al. (1999) FEBS Lett. 454(1-2):90-94; Lu, D. et al. (2005) «A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity», J. Biol. Chem. 280(20): 19665-19672; WO 02/02781 (Mertens et al.); Olafsen, T. et al. (2004) «Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications», Protein Eng Des Sel. 17(1):21-27; Wu, A. et al. (2001) «Multimerization Of A Chimeric Anti-CD20 Single Chain Fv-Fv Fusion Protein Is Mediated Through Variable Domain Exchange», Protein Engineering 14(2):1025-1033; Asano et al. (2004) «A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain», Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) «Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System», Protein Eng. 13(8):583-588; Baeuerle, P.A. et al. (2009) «Bispecific T cell Engaging Antibodies For Cancer Therapy», Cancer Res. 69(12):4941-4944).
[000123] Обеспечение немоноспецифических «диател» представляет значительное преимущество по сравнению с антителами: способность колигировать и колокализовать клетки, которые экспрессируют различные эпитопы. Таким образом, биспецифические диатела имеют широкое применение, включая терапию и иммунодиагностику. Биспецифичность обеспечивает большую гибкость в разработке и модификации диател для различных вариантов применения, обеспечивая повышенную авидность в отношении мультимерных антигенов, сшивание различных антигенов и направленное нацеливание на конкретные типы клеток, которое основано на присутствии обоих антигенов-мишеней. Благодаря своей двухвалентности, низким скоростям диссоциации и быстрому клиренсу из системы кровообращения (для диател небольшого размера около или ниже -50 кДа) молекулы диател, известные в данной области техники, также нашли особое применение в области визуализации опухолей (Fitzgerald et al. (1997) «Improved Tumour Targeting By Disulphide Stabilized Diabodies Expressed In Pichia pastoris», Protein Eng. 10:1221). Особое значение имеет колигирование различных клеток, например, сшивание цитотоксических Т-клеток с опухолевыми клетками (Staerz et al. (1985) «Hybrid Antibodies Can Target Sites For Attack By T Cells», Nature 314:628-631, и Holliger et al. (1996) «Specific Killing Of Lymphoma Cells By Cytotoxic T cells Mediated By A Bispecific Diabody», Protein Eng. 9:299-305), чтобы колокализовать Т-клетки в очагах опухолевых клеток.
[000124] В качестве альтернативы нацеливанию таких диател на связывание с Т-клетками эпитопсвязывающие домены диатела могут быть направлены к поверхностной детерминанте В-клетки, такой как CD19, CD20, CD22, CD30, CD37, CD40 и CD74 (Moore, P.A. et al. (2011) «Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T cell Killing Of B-Cell Lymphomas», Blood 117(17):4542-4551; Cheson, B.D. et al. (2008) «Monoclonal Antibody Therapy For B-Cell Non-Hodgkiris Lymphomas», N. Engl. J. Med. 359(6):613-626; Castillo, J. et al. (2008) «Newer Monoclonal Antibodies For Hematological Malignanciess», Exp. Hematol. 36(7):755-768). Во многих исследованиях также было обнаружено, что связывание диатела с детерминантами эффекторных клеток, например, с рецепторами Fcγ (FcγR), активирует эффекторные клетки (Holliger et al. (1996) «Specific Killing Of Lymphoma Cells By Cytotoxic T cells Mediated By A Bispeciftc Diabody», Protein Eng. 9:299-305; Holliger et al. (1999) «Carcinoembryonic Antigen (CEA)-Speciftc T cell Activation In Colon Carcinoma Induced By Anti-CD3 x Anti-CEA Bispeciftc Diabodies And B7 x Anti-CEA Bispeciftc Fusion Proteins», Cancer Res. 59:2909-2916; WO 2006/113665; WO 2008/157379; WO 2010/080538; WO 2012/018687; WO 2012/162068). Обычно активация эффекторных клеток запускается в результате связывания антигенсвязанного антитела с эффекторной клеткой за счет взаимодействия домен Fc-FcγR; соответственно, в этом отношении молекулы диатела могут проявлять Ig-подобную функциональность, независимо от того, содержат ли они домен Fc (например, как определено в любом анализе эффекторной функции, известном в данной области техники или приведенном в настоящем документе в качестве примера (например, анализ АЗКЦ)). За счет перекрестного связывания опухолевых и эффекторных клеток диатело не только сближает эффекторную клетку с опухолевой клеткой, но и приводит к эффективному уничтожению опухоли (см., например, Cao et al. (2003) «Bispecific Antibody Conjugates In Therapeutics», Adv. Drug. Deliv. Rev. 55:171-197).
[000125] Однако вышеупомянутые преимущества сопряжены со значительными затратами. Для образования указанных немоноспецифических диател необходима успешная сборка двух или более отдельных и различных полипептидов (т.е. такой процесс образования требует, чтобы диатела были образованы за счет гетеродимеризации различных типов полипептидных цепей). Этот факт отличается от ситуации с моноспецифическими диателами, которые образуются за счет гомодимеризации идентичных полипептидных цепей. Поскольку по меньшей мере два различных полипептида (т.е. две полипептидные молекулы) должны быть обеспечены для образования немоноспецифического диатела и поскольку гомодимеризация таких полипептидов приводит к получению неактивных молекул (Takemura, S. et al. (2000) «Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System», Protein Eng. 13(8):583-588), получение указанных полипептидов должно быть выполнено так, чтобы предотвратить ковалентное связывание между полипептидами одного и того же типа (т.е., чтобы свести к минимуму их гомодимеризацию) (Takemura, S. et al. (2000) «Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System», Protein Eng. 13(8):583-588). В этой связи в данной области техники было предложена нековалентная связь таких полипептидов (см., например, Olafsen et al. (2004) «Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications», Prot. Engr. Des. Sel. 17:21-27; Asano et al. (2004) «A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domains», Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) «Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System», Protein Eng. 13(8):583-588; Lu, D. et al. (2005) «A Fully Human Recombinant IgG - Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity», J. Biol. Chem. 280(20): 19665-19672).
[000126] Однако в данной области техники установлено, что биспецифические диатела, состоящие из нековалентно связанных полипептидов, являются нестабильными и легко диссоциируют на нефункциональные мономеры с одной полипептидной цепью (см., например, Lu, D. et al. (2005) «A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity», J. Biol. Chem. 280(20): 19665-19672).
[000127] В свете этой проблемы в данной области техники удалось разработать стабильные, ковалентно связанные гетеродимерные немоноспецифические диатела, названные диатела DART®, см., например, Chichili, G.R. et al. (2015) «А CD3xCD123 Bispecific DART For Redirecting Host T Cells To Myelogenous Leukemia: Preclinical Activity And Safety In Nonhuman Primates», Sci. Transl. Med. 7(289):289ra82; Johnson, S. et al. (2010) «Effector Cell Recruitment With Novel Fv-Based Dual-Affinity Re-Targeting Protein Leads To Potent Tumor Cytolysis And In Vivo B-Cell Depletion», J. Molec. Biol. 399(3):436-449; Veri, M.C. et al. (2010) «Therapeutic Control Of В Cell Activation Via Recruitment Of Fcgamma Receptor IIB (CD32B) Inhibitory Function With A Novel Bispecific Antibody Scaffold», Arthritis Rheum. 62(7): 1933-1943; Moore, P.A. et al. (2011) «Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T cell Killing Of B-Cell Lymphoma», Blood 117(17):4542-4551; патенты США Ms 8044180; 8133982; 8187593; 8193318; 8530627; 8669349; 8778339; 8784808; 8795667; 8802091; 8802093; 8946387; 8968730; и 8993730; публикации патентов США №№2009/0060910; 2010/0174053; 2011/0081347; 2011/0097323; 2011/0117089; 2012/0009186; 2012/0034221; 2012/0141476; 2012/0294796; 2013/0149236; 2013/0295121; 2014/0017237; и 2014/0099318; публикации европейских патентов ЕР 1868650; ЕР 2158221; ЕР 2247304; ЕР 2252631; ЕР 2282770; ЕР 2328934; ЕР 2376109; ЕР 2542256; ЕР 2601216; ЕР 2714079; ЕР 2714733; ЕР 2786762; ЕР 2839842; ЕР 2840091; и РСТ публикации №№ WO 2006/113665; WO 2008/157379; WO 2010/027797; WO 2010/033279; WO 2010/080538; WO 2011/109400; WO 2012/018687; WO 2012/162067; WO 2012/162068; WO 2014/159940; WO 2015/021089; WO 2015/026892; и WO 2015/026894). Такие диатела содержат два или более ковалентно связанных полипептидов и включают конструирование одного или более остатков цистеина в каждой из применяемых полипептидных молекул. Например, было показано, что добавление остатка цистеина к С-концу указанных конструкций обеспечивает образование дисульфидной связи между полипептидными цепями, стабилизируя полученный гетеродимер без отрицательного влияния на характеристики связывания двухвалентной молекулы.
[000128] Простейшее диатело DART® содержит две полипептидные цепи, каждая из которых содержит три домена (Фигура 1). Первая полипептидная цепь содержит: (i) первый домен, который содержит связывающую область из вариабельного домена легкой цепи первого иммуноглобулина (VL1), (ii) второй домен, который содержит связывающую область из вариабельного домена тяжелой цепи второго иммуноглобулина (VH2), и (iii) третий домен, который способствует гетеродимеризации (домен, способствующий образованию гетеродимера) со второй полипептидной цепью диатела и образованию ковалентной связи первой полипептидной цепи со второй полипептидной цепью диатела. Вторая полипептидная цепь содержит комплементарный первый домен (домен VL2), комплементарный второй домен (домен VH1) и третий домен, который объединяется с третьим доменом первой полипептидной цепи, чтобы способствовать гетеродимеризации (домен, способствующий образованию гетеродимера) и образованию ковалентной связи с первой полипептидной цепью. Такие молекулы являются стабильными, эффективными и обладают способностью одновременно связываться с двумя или более антигенами. Согласно одному варианту реализации настоящего изобретения каждый из третьих доменов первой и второй полипептидных цепей содержит остаток цистеина («©»), который служит для связывания полипептидов вместе за счет дисульфидной связи. Третий домен одной или обеих полипептидных цепей может дополнительно содержать последовательность домена СН2-СН3 так, что соединение двух полипептидов диатела образует область Fc, которая способна связываться с рецептором Fc клеток (таких как В-лимфоциты, дендритные клетки, природные клетки-киллеры, макрофаги, нейтрофилы, эозинофилы, базофилы и тучные клетки). Было описано много вариантов таких молекул (см., например, публикации патентов США №№2013-0295121; 2010-0174053; 2007-0004909; 2009-0060910; публикации европейских патентов №№ЕР 2714079; ЕР 2601216; ЕР 2376109; ЕР 2158221 и РСТ публикации №№WO 2012/162068; WO 2012/018687; WO 2010/080538; WO 2006/113665) и обеспечено в настоящем документе.
II. Компоненты предпочтительных CD137×TA связывающих молекул согласно настоящему изобретению
[000129] CD137×TA связывающие молекулы согласно настоящему изобретению состоят из полипептидов и могут состоять из двух, трех, четырех или более четырех полипептидных цепей. В настоящем документе термин «состоять из» является неограничивающим, таким образом, CD137×TA связывающие молекулы согласно настоящему изобретению, которые состоят из двух полипептидных цепей, могут иметь дополнительные полипептидные цепи. Такие цепи могут иметь ту же последовательность, что и другая полипептидная цепь связывающей молекулы, или их последовательность может отличаться от любой другой полипептидной цепи связывающей молекулы.
А. Предпочтительные «линкерные» пептиды
[000130] Полипептиды CD137×TA связывающих молекул согласно настоящему изобретению содержат домены, которым предшествуют, после которых следуют и/или соединяют их друг с другом «линкерные» пептиды, такие как линкер 1, линкер 2, линкер 3 и т.д. Несмотря на то, что в настоящем изобретении применяются определенные предпочтительные «линкерные» пептиды, в соответствии с представленными в настоящем документе идеями, альтернативные линкеры могут быть легко выявлены и применены для получения CD137×TA связывающих молекул.
[000131] Наиболее предпочтительно длина линкера 1, который разделяет такие домены VL и VH полипептидной цепи, выбрана, чтобы по существу или полностью предотвращать связывание таких доменов VL и VH друг с другом (например, 12 или менее остатков аминокислот в длину). Таким образом, домены VL1 и VH2 первой полипептидной цепи по существу или полностью не способны связываться друг с другом и не образуют эпитопсвязывающий сайт, который способен по существу связываться либо с первым, либо со вторым антигеном. Аналогичным образом, домены VL2 и VH1 второй полипептидной цепи по существу или полностью неспособны связываться друг с другом и не образуют эпитопсвязывающий сайт, который способен по существу связываться либо с первым, либо со вторым антигеном. Предпочтительный промежуточный спейсерный пептид (линкер 1) имеет последовательность (SEQ ID NO: 16):  которая является слишком короткой, чтоб обеспечить образование комплекса между доменами VL и VH одной и той же полипептидной цепи, в отличие от более длинного промежуточного спейсерного пептида, который применяется для получения молекул scFv (например,
которая является слишком короткой, чтоб обеспечить образование комплекса между доменами VL и VH одной и той же полипептидной цепи, в отличие от более длинного промежуточного спейсерного пептида, который применяется для получения молекул scFv (например,  (SEQ ID NO: 17)).
(SEQ ID NO: 17)).
[000132] Линкер 2 предназначен для отделения домена VH полипептидной цепи от необязательно присутствующего домена, способствующего образованию гетеродимера, этой полипептидной цепи. Любой из различных линкеров можно применять в качестве линкера 2. Предпочтительная последовательность для такого линкера 2 имеет аминокислотную последовательность:  (SEQ ID NO: 18), которая имеет остаток цистеина, который можно применять для ковалентной связи первой и второй полипептидных цепей друг с другом за счет дисульфидной связи, или
(SEQ ID NO: 18), которая имеет остаток цистеина, который можно применять для ковалентной связи первой и второй полипептидных цепей друг с другом за счет дисульфидной связи, или  (SEQ ID NO: 19), которая происходит из домена CH1 IgG. Поскольку линкер 2,
(SEQ ID NO: 19), которая происходит из домена CH1 IgG. Поскольку линкер 2,  (SEQ ID NO: 19), не имеет такого цистеина, применение такого линкера 2 предпочтительно связано с применением цистеинсодержащего домена, способствующего образованию гетеродимера, такого как Е-спираль, имеющая последовательность SEQ ID NO: 38, или K-спираль, имеющая последовательность SEQ ID NO: 39 (см. ниже).
(SEQ ID NO: 19), не имеет такого цистеина, применение такого линкера 2 предпочтительно связано с применением цистеинсодержащего домена, способствующего образованию гетеродимера, такого как Е-спираль, имеющая последовательность SEQ ID NO: 38, или K-спираль, имеющая последовательность SEQ ID NO: 39 (см. ниже).
[000133] Одна из функций линкера 3 заключается в отделении домена, способствующего образованию гетеродимера, полипептидной цепи от домена Fc этой полипептидной цепи. Вторая функция заключается в создании цистеинсодержащего полипептидного домена. Любой из различных линкеров можно применять в качестве линкера 3. Предпочтительная последовательность для такого линкера 3 имеет аминокислотную последовательность:  (SEQ ID NO: 20). Другая предпочтительная последовательность для линкера 3 имеет аминокислотную последовательность:
(SEQ ID NO: 20). Другая предпочтительная последовательность для линкера 3 имеет аминокислотную последовательность: (SEQ ID NO: 21).
(SEQ ID NO: 21).
[000134] Функция линкера 4 заключается в отделении С-конца доменов СН2-СН3 области Fc («домен Fc») от N-конца домена VL. Любой из различных линкеров можно применять в качестве линкера 4. Предпочтительная последовательность для такого линкера 4 имеет аминокислотную последовательность: (SEQ ID NO: 22) или аминокислотную последовательность
(SEQ ID NO: 22) или аминокислотную последовательность (SEQ ID NO: 23) или аминокислотную последовательность
(SEQ ID NO: 23) или аминокислотную последовательность (SEQ ID NO: 24).
(SEQ ID NO: 24).
[000135] Молекулы, содержащие область Fc, согласно настоящему изобретению могут включать дополнительные промежуточные спейсерные пептиды (линкеры), обычно такие линкеры будут включены между доменом, способствующим образованию гетеродимера (например, с Е-спиралью или K-спиралью), и доменом СН2-СН3 и/или между доменом СН2-СН3 и вариабельным доменом (т.е. VH или VL). Как правило, дополнительные линкеры будут содержать 3-20 остатков аминокислот и могут необязательно содержать полную шарнирную область IgG (предпочтительно цистеинсодержащую часть шарнирной области IgG) или ее часть. Линкеры, которые можно применять в молекулах биспецифических диател, содержащих область Fc, согласно настоящему изобретению, включают:  (SEQ ID NO: 19),
(SEQ ID NO: 19),  (SEQ ID NO: 20),
(SEQ ID NO: 20),  (SEQ ID NO: 22),
(SEQ ID NO: 22),  (SEQ ID NO: 23),
(SEQ ID NO: 23),  (SEQ ID NO: 24),
(SEQ ID NO: 24),  (SEQ ID NO: 25),
(SEQ ID NO: 25),  (SEQ ID NO: 26),
(SEQ ID NO: 26),  (SEQ ID NO: 27),
(SEQ ID NO: 27),  (SEQ ID NO: 28),
(SEQ ID NO: 28),  (SEQ ID NO: 29).
(SEQ ID NO: 29). (SEQ ID NO: 27) можно применять вместо
(SEQ ID NO: 27) можно применять вместо  или
или  для простоты клонирования. Помимо этого непосредственно после аминокислот
для простоты клонирования. Помимо этого непосредственно после аминокислот  или
или  (SEQ ID NO: 27) может быть расположена
(SEQ ID NO: 27) может быть расположена  (SEQ ID NO: 20) с образованием альтернативных линкеров:
(SEQ ID NO: 20) с образованием альтернативных линкеров:  (SEQ ID NO: 21); и
(SEQ ID NO: 21); и  (SEQ ID NO: 30). Биспецифические молекулы, содержащие область Fc, согласно настоящему изобретению могут включать шарнирную область IgG, такую как шарнирная область IgG человеческого антитела IgG1, IgG2, IgG3 или IgG4, или ее часть.
(SEQ ID NO: 30). Биспецифические молекулы, содержащие область Fc, согласно настоящему изобретению могут включать шарнирную область IgG, такую как шарнирная область IgG человеческого антитела IgG1, IgG2, IgG3 или IgG4, или ее часть.
В. Предпочтительные домены, способствующие образованию гетеродимера
[000136] Как указано выше, образование CD137×TA связывающих молекул согласно настоящему изобретению включает сборку двух или более различных полипептидных цепей (т.е. гетеродимеризацию). Образование гетеродимеров первой и второй полипептидных цепей может быть обусловлено включением «доменов, способствующих образованию гетеродимера». Домены, способствующие образованию гетеродимера, могут представлять собой домен шарнирной области IgG (или полипептид, происходящий из шарнирной области, такой как, например,  (SEQ ID NO: 31),
(SEQ ID NO: 31),  (SEQ ID NO: 32) или
(SEQ ID NO: 32) или (SEQ ID NO: 33) на одной полипептидной цепи и домен CL (или полипептид, происходящий из домена CL, такой как, например,
(SEQ ID NO: 33) на одной полипептидной цепи и домен CL (или полипептид, происходящий из домена CL, такой как, например,  (SEQ ID NO: 34) или
(SEQ ID NO: 34) или  (SEQ ID NO: 35) на другой полипептидной цепи (US 2007/0004909).
(SEQ ID NO: 35) на другой полипептидной цепи (US 2007/0004909).
[000137] Однако более предпочтительно домены, способствующие образованию гетеродимера, согласно настоящему изобретению будут содержать тандемно повторяющиеся спиральные домены противоположного заряда, например, спиральные домены «Е-спираль» (SEQ ID NO: 36: 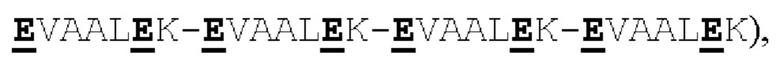 остатки глутамата в которых будут образовывать отрицательный заряд при рН=7, тогда как другой из доменов, способствующих образованию гетеродимера, будет содержать четыре тандемных домена «K-спираль» (SEQ ID NO: 37:
остатки глутамата в которых будут образовывать отрицательный заряд при рН=7, тогда как другой из доменов, способствующих образованию гетеродимера, будет содержать четыре тандемных домена «K-спираль» (SEQ ID NO: 37:
 ), остатки лизина в которых будут образовывать положительный заряд при рН=7. Присутствие таких заряженных доменов способствует объединению первого и второго полипептидов и, соответственно, способствует гетеродимеризации. Согласно другому предпочтительному варианту реализации настоящего изобретения применяется домен, способствующий образованию гетеродимера, в котором один из четырех тандемных спиральных доменов «Е-спираль», имеющий последовательность SEQ ID NO: 36, был модифицирован для включения остатка цистеина:
), остатки лизина в которых будут образовывать положительный заряд при рН=7. Присутствие таких заряженных доменов способствует объединению первого и второго полипептидов и, соответственно, способствует гетеродимеризации. Согласно другому предпочтительному варианту реализации настоящего изобретения применяется домен, способствующий образованию гетеродимера, в котором один из четырех тандемных спиральных доменов «Е-спираль», имеющий последовательность SEQ ID NO: 36, был модифицирован для включения остатка цистеина: 
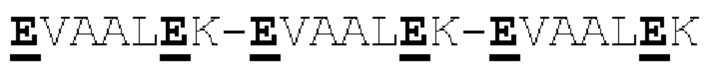 (SEQ ID NO: 38). Аналогичным образом, согласно другому предпочтительному варианту реализации настоящего изобретения применяется домен, способствующий образованию гетеродимера, в котором один из четырех спиральных доменов «K-спираль», имеющий последовательность SEQ ID NO: 37, был модифицирован для включения остатка цистеина:
(SEQ ID NO: 38). Аналогичным образом, согласно другому предпочтительному варианту реализации настоящего изобретения применяется домен, способствующий образованию гетеродимера, в котором один из четырех спиральных доменов «K-спираль», имеющий последовательность SEQ ID NO: 37, был модифицирован для включения остатка цистеина: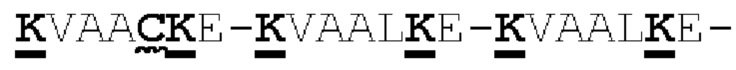
 (SEQ ID NO: 39).
(SEQ ID NO: 39).
C. Ковалентное связывание полипептидных цепей
[000138] CD137×TA связывающие молекулы согласно настоящему изобретению модифицированы так, что пары их полипептидных цепей ковалентно связываются друг с другом за счет одного или более остатков цистеина, расположенных по всей длине, с получением ковалентно связанного молекулярного комплекса. Такие остатки цистеина могут быть введены в промежуточный линкер, который отделяет домены VL и VH полипептидов. Согласно другому варианту линкер 2 или линкер 3, или альтернативный линкер, могут содержать остаток цистеина. Наиболее предпочтительно один или более спиральных доменов из содержащего спираль домена, способствующего образованию гетеродимера, будут содержать замену аминокислоты, которая включает остаток цистеина, как в SEQ ID NO: 38 или SEQ ID NO: 39.
D. Предпочтительные домены Fc
[000139] Домен Fc из Fc-несущей CD137×TA связывающей молекулы согласно настоящему изобретению может содержать полную область Fc (например, полную область Fc IgG) или только фрагмент полной области Fc. Таким образом, домен Fc из Fc-несущих CD137×TA связывающих молекул согласно настоящему изобретению может содержать часть или весь домен СН2 и/или часть или весь домен СН3 полной области Fc или может содержать вариант последовательности СН2 и/или вариант последовательности СН3 (который может содержать, например, одну или более вставок и/или одну или более делеций по отношению к доменам СН2 или СН3 полной области Fc). Домен Fc биспецифических Fc-несущих диател согласно настоящему изобретению может содержать участки полипептида, отличные от области Fc, или может содержать части областей Fc, которые в природных условиях не являются полными, или может содержать неприродные ориентации доменов СН2 и/или СН3 (такие как, например, два домена СН2 или два домена СН3, или, в направлении от N-конца к С-концу, домен СН3, соединенный с доменом СН2, и т.д.).
[000140] Несмотря на то, что домен Fc из Fc-несущей CD137×TA связывающей молекулы согласно настоящему изобретению может содержать аминокислотную последовательность природного домена Fc, предпочтительно домены СН2-СН3, которые образуют такой домен Fc, содержат одну или более замен так, чтобы полученный домен Fc обладал уменьшенным (например, менее чем 50%, менее чем 40%, менее чем 30%, менее чем 20% или менее чем 10% связывания, проявляемого такой молекулой, содержащей домен Fc с аминокислотной последовательностью природной области Fc) или по существу не детектируемым связыванием с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по сравнению со связыванием, проявляемым областью Fc дикого типа). Варианты Fc и мутантные формы, способные опосредовать такое измененное связывание, хорошо известны в данной области техники и включают замены аминокислот в одном или более положениях, выбранных из группы, состоящей из: 234, 235, 265 и 297, при этом указанная нумерация соответствует нумерации с использованием индекса ЕС, как изложено в Kabat (см., например, патент США №5624821, включенный в настоящий документ посредством ссылки). Согласно одному варианту реализации настоящего изобретения домен СН2-СН3 первой и/или третьей полипептидных цепей Fc-несущих молекул согласно настоящему изобретению включает любые 1, 2, 3 или 4 из замен: L234A, L235A, D265A, N297Q и N297G. Согласно другому варианту применяется домен СН2-СН3 природной области Fc, который исходно проявляет уменьшенное (или по существу отсутствующее) связывание с FcγRIIIA (CD16а) и/или сниженную эффекторную функцию (по отношению к связыванию и эффекторной функции, проявляемой областью Fc IgG1 дикого типа (SEQ ID NO: 12)). Согласно конкретному варианту реализации настоящего изобретения Fc-несущие молекулы согласно настоящему изобретению содержат область Fc IgG2 (SEQ ID NO: 13) или область Fc IgG4 (SEQ ID NO: 15). При применении области Fc IgG4 настоящее изобретение также охватывает введение стабилизирующей мутации, такой как замена S228P в шарнирной области, описанной выше (см., например, SEQ ID NO: 11).
[000141] Согласно предпочтительному варианту реализации настоящего изобретения применяемый домен СН2-СН3 Fc-несущих CD137×TA связывающих молекул согласно настоящему изобретению включает замену аланином в положении 234 и замену аланином в положении 235, при этом указанная нумерация соответствует нумерации с использованием индекса ЕС, как и изложено Kabat (SEQ ID NO: 40):

где X представляет собой лизин (K) или отсутствует.
[000142] Период полужизни в сыворотке белков, содержащих области Fc, может быть увеличен за счет увеличения аффинности связывания области Fc с FcRn. В настоящем документе термин «период полужизни» означает фармакокинетическое свойство молекулы, которое является мерой среднего времени существования молекул после их введения. Период полужизни может быть выражен как время, необходимое для удаления пятидесяти процентов (50%) известного количества молекулы из тела субъекта (например, пациента-человека или другого млекопитающего) или его конкретного компартмента, например, при измерении в сыворотке крови, т.е. период полужизни в кровотоке, или в других тканях. Обычно увеличение периода полужизни приводит к увеличению среднего времени удержания (МРТ) в кровотоке для введенной молекулы.
[000143] Согласно некоторым вариантам реализации настоящего изобретения Fc- несущие CD137×TA связывающие молекулы согласно настоящему изобретению содержат вариант области Fc, причем указанный вариант области Fc содержит по меньшей мере одну аминокислотную модификацию относительно области Fc дикого типа так, что указанная молекула имеет увеличенный период полужизни (по сравнению с молекулой, содержащей область Fc дикого типа). Согласно некоторым вариантам реализации настоящего изобретения Fc-несущие CD137×TA связывающие молекулы согласно настоящему изобретению содержат вариант области Fc IgG, причем указанный вариант области Fc содержит замену аминокислоты, увеличивающую период полужизни. В данной области техники известны многочисленные замены аминокислот, способные увеличивать период полужизни Fc-несущей молекулы, см., например, замены аминокислот, описанные в патентах США №№6277375, 7083784; 7217797, 8088376; публикациях США №№2002/0147311; 2007/0148164; и РСТ публикациях №№WO 98/23289; WO 2009/058492; и WO 2010/033279, которые полностью включены в настоящий документ посредством ссылки. Fc-несущая CD137×TA связывающая молекула, имеющая увеличенный период полужизни, может содержать две или более замен, выбранных из: T250Q, M252Y, S254T, Т256Е, K288D, T307Q, V308P, A378V, M428L, N434A, Н435К и Y436I, при этом указанная нумерация соответствует нумерации с использованием индекса ЕС, как изложено в Kabat.
[000144] В частности, применяемый домен СН2-СН3 может содержать замены:
(A) M252Y, S254T и Т256Е;
(B) M252Y и S254T;
(C) M252Y и Т256Е;
(D) T250Q и M428L;
(E) T307Q и N434A;
(F) A378V и N434A;
(G) N434A и Y436I;
(H) V308P и N434A; или
(I) K288D и H435K,
при этом указанная нумерация соответствует нумерации с использованием индекса ЕС, как изложено в Kabat.
[000145] Предпочтительная последовательность для доменов СН2 и СН3 содержит тройную замену аминокислот: M252Y/S254T/T256E (YTE), которая значительно увеличивает время полужизни в сыворотке (Dall'Acqua, W.F. et al. (2006) «Properties of Human IgGs Engineered for Enhanced Binding to the Neonatal Fc Receptor (FcRn)», J. Biol. Chem. 281(33):23514-23524), как в SEQ ID NO: 41 или SEQ ID NO: 42, которые представляют собой варианты домена СН2-СН3 IgG1, или как в SEQ ID NO: 43, которая представляет собой вариант домена СН2-СН3 IgG4: SEQ ID NO: 41:

где X представляет собой лизин (K) или отсутствует.
SEQ ID NO: 42:

где X представляет собой лизин (K) или отсутствует.
SEQ ID NO: 43:

где X представляет собой лизин (K) или отсутствует.
[000146] Настоящее изобретение также охватывает Fc-несущие CD137×TA связывающие молекулы, содержащие варианты домена Fc, которые проявляют измененную эффекторную функцию, измененный период полужизни в сыворотке, измененную стабильность, измененную восприимчивость к клеточным ферментам или измененную эффекторную функцию, согласно результатам NK-зависимого или макрофагозависимого анализа и т.д. Модификации домена Fc, выявленные как изменяющие эффекторную функцию, известны в данной области техники, включая модификации, которые увеличивают связывание с активирующими рецепторами (например, FcγRIIA (CD16A)) и уменьшают связывание с ингибирующими рецепторами (например, FcγRIIB (CD32B)) (см., например, Stavenhagen, J.B. et al. (2007) «Fc Optimization Of Therapeutic Antibodies Enhances Their Ability To Kill Tumor Cells In Vitro And Controls Tumor Expansion In Vivo Via Low-Affinity Activating Fcgamma Receptors», Cancer Res. 57(18):8882-8890). Примерные варианты доменов Fc человеческого IgG1 со сниженным связыванием с CD32B и/или увеличенным связыванием с CD16A содержат замены F243L, R292P, Y300L, V305I или P296L. Эти замены аминокислот могут присутствовать в домене Fc человеческого IgG1 в любой комбинации. Согласно одному варианту реализации настоящего изобретения вариант домена Fc человеческого IgG1 содержит замену F243L, R292P и Y300L, причем указанная нумерация соответствует нумерации с использованием индекса ЕС, как изложено в Kabat. Согласно другому варианту реализации настоящего изобретения вариант домена Fc человеческого IgG1 содержит замену F243L, R292P, Y300L, V305I и P296L, причем указанная нумерация соответствует нумерации с использованием индекса ЕС, как изложено в Kabat.
[000147] Домены СН2 и/или СН3 CD137×TA связывающих молекул согласно настоящему изобретению не обязательно должны иметь идентичные последовательности и преимущественно модифицированы, чтобы способствовать гетеродимеризации между двумя СН2-СН3-несущими полипептидными цепями. Например, замена аминокислоты (предпочтительно замена аминокислотой, содержащей объемную боковую группу, образующую «выступ», например, триптофаном), может быть введена в домен СН2 или СН3 так, что стерическое влияние предотвратит взаимодействие с доменом, мутированным аналогичным образом, и приведет к спариванию мутированного домена с доменом, в который была введена комплементарная или содействующая мутация, т.е. «впадина» (например, замена глицином). Такие группы мутаций могут быть введены в любую пару полипептидов, содержащихся в биспецифической Fc-содержащей молекуле диатела, и, кроме того, введены в любую часть полипептидных цепей указанной пары. Способы модификации белков для стимулирования гетеродимеризации по отношению к гомодимеризации хорошо известны в данной области техники, в частности, касательно модификации молекул, подобных иммуноглобулинам, и охватываются настоящим изобретением (см., например, Ridgway et al. (1996) «'Knobs-Into-Holes' Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization», Protein Engr. 9:617-621, Atwell et al. (1997) «Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library», J. Mol. Biol. 270: 26-35, и Xie et al. (2005) «A New Format Of Bispecific Antibody: Highly Efficient Heterodimerization, Expression And Tumor Cell Lysis», J. Immunol. Methods 296:95-101; каждый из указанных документы полностью включен в настоящий документ посредством ссылки). Согласно одному варианту реализации настоящего изобретения «выступ» вводят в домены СН2-СН3 первой полипептидной цепи, и «впадину» вводят в домены СН2-СН3 третьей полипептидной цепи. Таким образом, «выступ» поможет предотвратить гомодимеризацию двух молекул первой полипептидной цепи за счет их доменов СН2 и/или СН3. Поскольку третья полипептидная цепь этого варианта реализации предпочтительно содержит замену «впадина», она будет способна гетеродимеризоваться с первой полипептидной цепью, а также гомодимеризоваться сама с собой (однако такая гомодимеризация не приводит к образованию молекулы, имеющей эпитопсвязывающие сайты). Предпочтительный «выступ» создают путем модификации нативного домена Fc IgG для включения модификации T366W. Предпочтительную «впадину» создают путем модификации нативного домена Fc IgG для включения модификации T366S, L368A и Y407V. Чтобы облегчить очистку гомодимера третьей полипептидной цепи от готового биспецифического Fc-несущего диатела, содержащего гетеродимеры первой и третьей полипептидных цепей, сайт связывания белка А доменов СН2 и СН3 третьей полипептидной цепи предпочтительно мутируют путем замены аминокислоты в положении 435 (H435R). Таким образом, гомодимер третьей полипептидной цепи не будет связываться с белком А, тогда как правильно собранное биспецифическое Fc-несущее диатело будет сохранять свою способность связывать белок А за счет сайта связывания белка А на первой полипептидной цепи.
[0001481 SEQ ID NO: 44, SEQ ID NO: 45 и SEQ ID NO: 46 обеспечивают примерные предпочтительные последовательности для «несущих выступы» доменов СН2 и СН3, которые можно применять в CD137×TA связывающих молекулах согласно настоящему изобретению:
SEQ ID NO: 44:

где X представляет собой лизин (K) или отсутствует,
SEQ ID NO: 45:

где X представляет собой лизин (K) или отсутствует,
SEQ ID NO: 46:

где X представляет собой лизин (K) или отсутствует.
[0001491 SEQ ID NO: 47, SEQ ID NO: 48 и SEQ ID NO: 49 обеспечивают примерные предпочтительные последовательности для «несущих впадину» доменов СН2 и СН3, которые можно применять в CD137×TA связывающих молекулах согласно настоящему изобретению:
SEQ ID NO: 47:

где X представляет собой лизин (K) или отсутствует,
SEQ ID NO: 48:

где X представляет собой лизин (K) или отсутствует,
SEQ ID NO: 49:

где X представляет собой лизин (K) или отсутствует.
[000150] Как будет отмечено, домены СН2-СН3 из SEQ ID NO: 45 и 48 представляют собой домены IgG4, тогда как домены СН2-СН3 из SEQ ID NO: 44, 46, 47 и 49 представляют собой домены IgG1. SEQ ID NO: 44, 46, 47 и 49 содержат замену аланином в положении 234 и замену аланином в положении 235 и, соответственно, образуют домен Fc, который проявляет уменьшенное (или по существу отсутствующее) связывание с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по сравнению со связыванием, проявляемым областью Fc дикого типа (SEQ ID NO: 12). Кроме того, настоящее изобретение конкретно охватывает конструкции CD137×TA связывающих молекул, в которых отсутствует вышеуказанный С-концевой остаток лизина.
[000151] Согласно варианту реализации, описанному выше, первая полипептидная цепь будет иметь «несущую выступ» последовательность СН2-СН3, такую как последовательность SEQ ID NO: 44. Однако, как будет понятно, «несущий впадину» домен СН2-СН3 (например, SEQ ID NO: 47) мог применяться в первой полипептидной цепи, и в этом случае «несущий выступ» домен СН2-СН3 (например, SEQ ID NO: 44) будет применяться в третьей полипептидной цепи.
Е. Альбуминсвязывающий домен
[000152] Как раскрыто в WO 2012/018687, для того чтобы улучшить фармакокинетические свойства диател в условиях in vivo, диатело может быть модифицировано, чтобы содержать полипептидную часть сывороточного связывающего белка на одном или более концах диатела. Такие соображения также применимы к триспецифическим связывающим молекулам согласно настоящему изобретению. Наиболее предпочтительно, если желательно включение полипептидной части сывороточного связывающего белка в триспецифические связывающие молекулы согласно настоящему изобретению, такая полипептидная часть будет встроена на С-конце одной из полипептидных цепей триспецифической связывающей молекулы.
[000153] Альбумин является самым распространенным белком в плазме крови и имеет период полужизни 19 дней у человека. Альбумин имеет несколько сайтов связывания небольших молекул, которые позволяют ему нековалентно связываться с другими белками и тем самым продлевать их периоды полужизни в сыворотке крови. Альбуминсвязывающий домен 3 (ABD3) белка G штамма Streptococcus G148 состоит из 46 остатков аминокислот, образующих стабильный трехвинтовой жгут, и обладает широкой альбуминсвязывающей специфичностью (Johansson, M.U. et al. (2002) «Structure, Specificity, And Mode Of Interaction For Bacterial Albumin-Binding Modules», J. Biol. Chem. 277(10):8114-8120). Соответственно, особенно предпочтительная полипептидная часть сывороточного связывающего белка для улучшения фармакокинетических свойств диатела в условиях in vivo представляет собой альбуминсвязывающий домен (ABD) из стрептококкового белка G и более предпочтительно альбуминсвязывающий домен 3 (ABD3) белка G штамма Streptococcus dysgalactiae G148 (SEQ ID NO: 50): 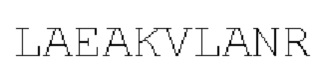

[000154] Как раскрыто в WO 2012/162068 (который включен в настоящую заявку посредством ссылки), «деиммунизированные» варианты, имеющие последовательность SEQ ID NO: 50, способны ослаблять или устранять связывание с ГКГС класса II. Исходя из результатов для комбинированных мутаций, следующие комбинации замен рассматривают как предпочтительные замены для получения указанного деиммунизированного альбуминсвязывающего домена: 66S/70S +71A; 66S/70S +79А; 64A/65A/71A+66S; 64A/65A/71A+66D; 64А/65А/71А+66Е; 64A/65A/79A+66S; 64A/65A/79A+66D; 64А/65А/79А+66Е. Варианты ABD содержат модификации L64A, I65A и D79A или модификации N66S, T70S и D79A. Варианты деиммунизированного ABD, имеющие аминокислотную последовательность SEQ ID NO: 51, 52 или 53, являются особенно предпочтительными, поскольку указанные деиммунизированные альбуминсвязывающие домены проявляют по существу связывание дикого типа, обеспечивая при этом ослабленное связывание с ГКГС класса II:
SEQ ID NO: NO:51:
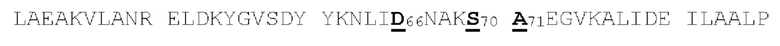
SEQ ID NO: NO:52:

SEQ ID NO: NO:53:

[000155] Несмотря на то, что такие альбуминсвязывающие домены могут быть включены в любую из полипептидных цепей триспецифических связывающих молекул согласно настоящему изобретению, предпочтительно расположить такой домен в направлении С-конца относительно домена Е-спирали (или K-спирали) первой или третьей полипептидной цепи (за счет линкера, который находится между доменом Е-спирали (или K-спирали) и альбуминсвязывающим доменом (который предпочтительно представляет собой деиммунизированный альбуминсвязывающий домен)). Предпочтительная последовательность для такого линкера представляет собой SEQ ID NO: 26: GGGS.
F. Предпочтительные опухолевые антигены (ТА) и примерные вариабельные домены
[000156] CD137×TA связывающие молекулы согласно настоящему изобретению содержат по меньшей мере один эпитопсвязывающий сайт, специфичный для эпитопа опухолевого антигена. Примерные опухолевые антигены («ТА»), которые могут быть связаны CD137×TA связывающими молекулами согласно настоящему изобретению, включают, но не ограничиваются указанными: антиген рака толстой кишки 19.9; онкофетальный белок 5Т4; антиген 4.2 муцинов при раке желудка; антиген колоректальной карциномы А33 (Almqvist, Y. 2006, Nucl Med Biol. Nov; 33(8):991-998); ADAM-9 (публикация патента США 2006/0172350, РСТ публикация WO 06/084075); онкофетальный антиген альфа-фетопротеин, AFP (Malaguarnera, G. et al. (2010) «Serum markers of hepatocellular carcinoma», Dig. Dis. Sci. 55(10):2744-2755); ALCAM (PCT публикация WO 03/093443); BAGE (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); бета-катенин (Prange W. et al. 2003 J Pathol. 201(2):250-9); CA125 (Bast, R.C. Jr. et al. 2005 Int J Gynecol Cancer 15 Suppl 3:274-81); карбоксипептидазу M (публикация патента США 2006/0166291); Bl (Egloff, A.M. et al. 2006, Cancer Res. 66(1):6-9); CD5 (Calin, G.A et al. 2006 Semin Oncol. 33(2): 167-73; CD19 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4): 139-48); CD20 (Thomas, D.A et al. 2006 Hematol Oncol Clin North Am. 20(5): 1125-36); CD20 (Cang, S. et al. (2012) «Novel CD20 Monoclonal Antibodies For Lymphoma Therapy», J. Hematol. Oncol. 5:64 pp. 1-9); CD22 (Kreitman, R.J. 2006 AAPS J. 18; 8(3):E532-51); CD23 (Rosati, S. et al. 2005 Curr Top Microbiol Immunol. 5;294:91-107); CD25 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4): 139-48); CD27 (Bataille, R. 2006 Haematologica 91(9): 1234-40); CD28 (Bataille, R. 2006 Haematologica 91(9): 1234-40); CD30 (Muta, H. et al. (2013) «CD30: From Basic Research To Cancer Therapy», Immunol. Res. 57(1-3): 151-158); CD33 (Walter, R.B. et al. (2012) «Acute myeloid leukemia stem cells and CD33-targeted immunotherapy», Blood 119(26):6198-6208); CD36 (Ge, Y. 2005 Lab Hematol. 11(1):31-7); CD40/CD154 (Messmer, D. et al. 2005 Ann N Y Acad Sci. 1062:51-60); CD45 (Jurcic, J.G. 2005 Curr Oncol Rep. 7(5):339-46); CD56 (Bataille, R. 2006 Haematologica 91(9): 1234-40); CD46 (патент США №7148038; РСТ публикация №WO 03/032814; Russell, S. et al. (2004) «CD46: A Complement Regulator And Pathogen Receptor That Mediates Links Between Innate And Acquired Immune Function», Tissue Antigens 64(2): 111-118); CD52 (Hoelzer, D. et al. (2013) «Targeted therapy with monoclonal antibodies in acute lymphoblastic leukemia», Curr. Opin. Oncol. 25(6):701-706); CD79a/CD79b (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48; Chu, P.G. et al. 2001 Appl Immunohistochem Mol Morphol. 9(2):97-106); CD103 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4): 139-48); CD123 (Taussig, D. et al. (2005) «Hematopoietic Stem Cells Express Multiple Myeloid Markers: Implications For The Origin And Targeted Therapy Of Acute Myeloid Leukemia», Blood 106:4086-4092); CD317 (Palma, G. et al. (2012) «Plasmacytoids Dendritic Cells Are A Therapeutic Target In Anticancer Immunity», Biochim. Biophys. Acta. 1826(2):407-414; CDK4 (Lee, Y.M. et al. 2006 Cell Cycle 5(18):2110-4); CEA (карциноэмбриональный антиген Mathelin, С.2006 Gynecol Obstet Fertil. 34(7-8):638-46; Tellez-Avila, F.I. et al. 2005 Rev Invest Clin. 57(6):814-9); CEACAM5 и CEACAM6 (РСТ публикация №WO 2011/034660; Zheng, С. et al. (2011) «A Novel Anti- CEACAM5 Monoclonal Antibody, CC4, Suppresses Colorectal Tumor Growth and Enhances NK Cells-Mediated Tumor Immunity», PLoS One 6(6):e21146, pp. 1-11); C017-1A (Adkins, J.C. et al. (1998) «Edrecolomab (Monoclonal Antibody 17-1А)», Drugs 56(4):619-626); CO-43 (антиген группы крови Leb) и CO-514 (антиген группы крови Lea) (Garratty, G. (1995) «Blood Group Antigens As Tumor Markers, Parasitic/Bacterial/Viral Receptors, And Their Association With Immunologically Important Proteins», Immunol. Invest. 24(l-2):213-232; CTLA-1 и CTLA-4 (Peggs, K.S. et al. 2006 Curr Opin Immunol. 18(2):206-13); цитокератин 8 (РСТ публикация №WO 03/024191); антиген D1.1 (Dao, Т. et al. (2009) «Identification Of A Human Cyclin D1-Derived Peptide That Induces Human Cytotoxic CD4 T Cells», PLoS One. 4(8):e6730); DR5 (Abdulghani, J. et al. (2010) «TRAIL Receptor Signaling And Therapeutics», Expert Opin. Ther. Targets 14(10):1091-1108; Andera, L. (2009) «Signaling Activated By The Death Receptors Of The TNFR Family», Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 153(3): 173-180; Carlo-Stella, C. et al. (2007) «Targeting TRAIL Agonistic Receptors for Cancer Therapy», Clin, Cancer 13(8):2313-2317; Chaudhari, B.R. et al. (2006) «Following the TRAIL to Apoptosis», Immunologic Res. 35(3):249-262); ряд E1 (группа крови В); ЭФР-Р (рецептор эпидермального фактора роста, Adenis, A. et al. 2003 Bull Cancer. 90 Spec No:S228-32); рецепторы эфрина (и, в частности, EphA2 (патент США №7569672, РСТ публикация WO 06/084226); Erb (ErbB1, ErbB3, ErbB4, Zhou, Н. et al. 2002 Oncogene 21(57):8732-8740; Rimon, E. et al. 2004 bit J Oncol. 24(5): 1325-1338); антиген аденокарциномы легких F3 (Greulich, H. et al. (2012) «Functional analysis of receptor tyrosine kinase mutations in lung cancer identifies oncogenic extracellular domain mutations of ERBB2», Proc. Natl. Acad. Sci. (U.S.A.) 109(36):14476-14481); антиген FC10.2 (Loveless, W. et al. (1990) «Developmental Patterning Of The Carbohydrate Antigen FC10.2 During Early Embryogenesis In The Chick», Development 108(1):97-106); GAGE (GAGE-1; GAGE-2; Akcakanat, A. et al. 2006 Int J Cancer. 118(1): 123-128); GD2/GD3/GD49/GM2/GM3 (Livingston, P.O. et at. 2005 Cancer Immunol Immunother. 54(10): 1018-25); GICA 19-9 (Herlyn et al. (1982) «Monoclonal Antibody Detection Of A Circulating Tumor-Associated Antigen. I. Presence Of Antigen In Sera Of Patients With Colorectal, Gastric, And Pancreatic Carcinoma», J. Clin. Immunol. 2:135-140); gp37 (Т-клеточный антиген лейкоза человека, Bhattacharya-Chatterjee et al. (1988) «Idiotype Vaccines Against Human T Cell Leukemia. II. Generation And Characterization Of A Monoclonal Idiotype Cascade (Ab1, Ab2, and Ab3)», J. Immunol. 141:1398-1403); gp75 (антиген меланомы; Vijayasardahl et al. (1990) «The Melanoma Antigen Gp75 Is The Human Homologue Of The Mouse В (Brown) Locus Gene Product», J. Exp. Med. 171(4): 1375-1380); gp100 (Lotem, M. et al. 2006 J Immunother. 29(6):616-27); HER2/neu (Kumar, Pal S et al. 2006 Semin Oncol. 33(4):386-91); антиген CD20 В-клеточной лимфомы человека (Reff et al. (1994) «Depletion Of В Cells In Vivo By A Chimeric Mouse Human Monoclonal Antibody To CD20», Blood 83:435-445); антиген глобулы молочного жира человека; Е6 папилломавируса человека/Е7 папилломавируса человека (DiMaio, D. et al. 2006 Adv Virus Res. 66:125-59); HMW-MAA (высокомолекулярный антиген меланомы) (Natali et al. (1987) «Immunohistochemical Detection Of Antigen In Human Primary And Metastatic Melanomas By The Monoclonal Antibody 140.240 And Its Possible Prognostic Significance», Cancer 59:55-63; Mittelman et al. (1990) «Active Specific Immunotherapy In Patients With Melanoma. A Clinical Trial With Mouse Antiidiotypic Monoclonal Antibodies Elicited With Syngeneic Anti-High-Molecular-Weight-Melanoma-Associated Antigen Monoclonal Antibodies», J. Clin. Invest. 86:2136-2144); антиген I (дифференцировочный антиген) (Feizi (1985) «Demonstration By Monoclonal Antibodies That Carbohydrate Structures Of Glycoproteins And Glycolipids Are Onco-Developmental Antigens», Nature 314:53-57), такой как I(Ma), обнаруженный при аденокарциномах желудка; интегрин альфа-V-бета-6 интегрин-β6 (ITGB6) (РСТ публикация № WO 03/087340); JAM-3 (РСТ публикация № WO 06/084078); рецептор интерлейкина-13 α2 (IL13Rα2) (Bodhinayake, I. et al. (2014) «Targeting A Heterogeneous Tumor: The Promise Of The Interleukin-13 Receptor α2», Neurosurgery 75(2):N18-9); JAM-3 (РСТ публикация WO 06/084078); KID3 (РСТ публикация WO 05/028498); KID31 (PCT публикация WO 06/076584); общий антиген карциномы KS 1/4 (Perez et al. (1989) «Isolation And Characterization Of A cDNA Encoding The Ks1/4 Epithelial Carcinoma Marker», J. Immunol. 142:3662-3667;  et al. (1991) «Bispecific-Monoclonal-Antibody-Directed Lysis Of Ovarian Carcinoma Cells By Activated Human T Lymphocytes», Cancer Immunol. Immunother. 33(4):210-216; Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); общий антиген карциномы KS 1/4 (Perez et al. (1989) «Isolation And Characterization Of A cDNA Encoding The KsI/4 Epithelial Carcinoma Marker», J. Immunol. 142:3662-3667; et al. (1991) «Bispecific-Monoclonal-Antibody-Directed Lysis Of Ovarian Carcinoma Cells By Activated Human T Lymphocytes», Cancer Immunol. Immunother. 33(4):210-216; Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); KSA (17-1A) (Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); антигены карциномы легких человека L6 и L20 (
et al. (1991) «Bispecific-Monoclonal-Antibody-Directed Lysis Of Ovarian Carcinoma Cells By Activated Human T Lymphocytes», Cancer Immunol. Immunother. 33(4):210-216; Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); общий антиген карциномы KS 1/4 (Perez et al. (1989) «Isolation And Characterization Of A cDNA Encoding The KsI/4 Epithelial Carcinoma Marker», J. Immunol. 142:3662-3667; et al. (1991) «Bispecific-Monoclonal-Antibody-Directed Lysis Of Ovarian Carcinoma Cells By Activated Human T Lymphocytes», Cancer Immunol. Immunother. 33(4):210-216; Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); KSA (17-1A) (Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); антигены карциномы легких человека L6 и L20 ( et al. (1986) «Monoclonal Mouse Antibodies Raised Against Human Lung Carcinoma», Cancer Res. 46:3917-3923); LEA (
et al. (1986) «Monoclonal Mouse Antibodies Raised Against Human Lung Carcinoma», Cancer Res. 46:3917-3923); LEA ( , N. et al. (2012) «Sialyl Lewis x expression in cervical scrapes of premalignant lesions», J. Biosci. 37(6):999-1004); LUCA-2 (публикация патента США №2006/0172349; PCT публикация WO 06/083852); M1:22:25:8; М18, М39 (Cambier, L. et al. (2012) «М19 Modulates Skeletal Muscle Differentiation And Insulin Secretion In Pancreatic B-Cells Through Modulation Of Respiratory Chain Activity», PLoS One 7(2):e31815; Pui, C.H. et al. (1991) «Characterization of childhood acute leukemia with multiple myeloid and lymphoid markers at diagnosis and at relapse», Blood 78(5): 1327-1337); MAGE (MAGE-1; MAGE-3; (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); MART (Kounalakis, N. et al. 2005 Curr Oncol Rep.7(5):377-82; Myl, MUC-1 (Mathelin, C. 2006 Gynecol Obstet Fertil. 34(7-8):638-46); MUM-1 (Castelli, C. et al. 2000 J Cell Physiol. 182(3):323-31); N-ацетилглюкозаминилтрансферазу (Dennis, J.W. 1999 Biochim Biophys Acta. 6; 1473(l):21-34); неогликопротеин (Legendre, H. et al. (2004) «Prognostic Stratification Of Dukes В Colon Cancer By A Neoglycoprotein», Int. J. Oncol. 25(2):269-276); NS-10; OFA-1 и OFA-2 (Takahashi, M. (1984) «A Study On Clinical Significance Of Oncofetal Antigen-1 In Gynecologic Tumors», Nihon Sanka Fujinka Gakkai Zasshi. 36(12):2613-2618); онкостатин M (рецептор онкостатина бета) (патент США №7572896, PCT публикация WO 06/084092); р15 (Gil, J. et al. 2006 Nat Rev Mol Cell Biol. 7(9):667-77); PSA (простатический специфический антиген; Cracco, C.M. et al. 2005 Minerva Urol Nefrol. 57(4):301-11); PSMA (Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); РЕМА (антиген полиморфного эпителиального муцина) (Chu, N.J. et al. (2015) «Nonviral Oncogenic Antigens and the Inflammatory Signals Driving Early Cancer Development as Targets for Cancer Immunoprevention», Clin. Cancer Res. 21(7): 1549-1557); PIPA (патент США №7405061, PCT публикация WO 04/043239); простатическую кислую фосфатазу (Tailor et al. (1990) «Nucleotide Sequence Of Human Prostatic Acid Phosphatase Determined From A Full-Length cDNA Clone», Nucl. Acids Res. 18(16):4928); R24 (Zhou, M. et al. (2008) «Constitutive Overexpression Of A Novel 21 Kda Protein By Hodgkin Lymphoma And Aggressive Non-Hodgkin Lymphomas», Mol. Cancer 7:12); ROR1 (патент США №5843749); Rabbani, H. et al. (2010) «Expression Of ROR1 In Patients With Renal Cancer-A Potential Diagnostic Marker», Iran Biomed. J. 14(3):77-82); сфинголипиды (Hakomori, S. (1998) «Cancer-Associated Glycosphingolipid Antigens: Their Structure, Organization, And Function», Acta Anat. (Basel) 161(1-4):79-90); SSEA-1, SSEA-3 и SSEA-4 (Muramatsu, T. et al. (2004) «Carbohydrate Antigens Expressed On Stem Cells And Early Embryonic Cells», Glycoconj. J. 21(1-2):41-45); sTn (Holmberg, L.A. 2001 Expert Opin Biol Ther. 1(5):881-91); происходящий из Т-клеточного рецептора пептид (Edelson (1998) «Cutaneous T-Cell Lymphoma: A Model For Selective Immunotherapy», Cancer J Sci Am. 4:62-71); T5A7 (Hogg, R.J. et al. (1991) «A monoclonal antibody exhibiting reactivity with both X-hapten- and lactose- bearing glycolipids», Tissue Antigens 37(1):33-38); TAG-72 (Yokota et al. (1992) «Rapid Tumor Penetration Of A Single-Chain Fv And Comparison With Other Immunoglobulin Forms», Cancer Res. 52:3402-3408); TL5 (группа крови A) (Gooi, H.C. et al. (1983) «Monoclonal antibody reactive with the human epidermal-growth-factor receptor recognizes the blood-group- A antigen», Biosci. Rep. 3(11): 1045-1052); рецептор TNF (рецептор TNF-α, рецептор TNF-β или рецептор TNF-γ (van Horssen, R. et al. 2006 Oncologist. 11(4):397-408; Gardnerova, M. et al. 2000 Curr Drug Targets. 1(4):327-64); TRA-1-85 (группа крови H) (Williams, B.P. et al. (1988) «Biochemical and genetic analysis of the OKa blood group antigen», Immunogenetics 27(5):322-329); рецептор трансферрина (патент США №7572895, РСТ публикация WO №05/121179); TSTA, опухолеспецифичный трансплантационный антиген ( et al. (1985) {(Monoclonal Antibodies To Cell Surface Antigens Shared By Chemically Induced Mouse Bladder Carcinomas», Cancer. Res. 45:2210-2188); VEGF-R (O'Dwyer. P.J. 2006 Oncologist. 11(9):992-8); и гаптен Y, Ley (Durrant, L.G. et al. (1989) ((Development Of An ELISA To Detect Early Local Relapse Of Colorectal Саnсеr», Br. J. Cancer 60(4): 533-537).
, N. et al. (2012) «Sialyl Lewis x expression in cervical scrapes of premalignant lesions», J. Biosci. 37(6):999-1004); LUCA-2 (публикация патента США №2006/0172349; PCT публикация WO 06/083852); M1:22:25:8; М18, М39 (Cambier, L. et al. (2012) «М19 Modulates Skeletal Muscle Differentiation And Insulin Secretion In Pancreatic B-Cells Through Modulation Of Respiratory Chain Activity», PLoS One 7(2):e31815; Pui, C.H. et al. (1991) «Characterization of childhood acute leukemia with multiple myeloid and lymphoid markers at diagnosis and at relapse», Blood 78(5): 1327-1337); MAGE (MAGE-1; MAGE-3; (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); MART (Kounalakis, N. et al. 2005 Curr Oncol Rep.7(5):377-82; Myl, MUC-1 (Mathelin, C. 2006 Gynecol Obstet Fertil. 34(7-8):638-46); MUM-1 (Castelli, C. et al. 2000 J Cell Physiol. 182(3):323-31); N-ацетилглюкозаминилтрансферазу (Dennis, J.W. 1999 Biochim Biophys Acta. 6; 1473(l):21-34); неогликопротеин (Legendre, H. et al. (2004) «Prognostic Stratification Of Dukes В Colon Cancer By A Neoglycoprotein», Int. J. Oncol. 25(2):269-276); NS-10; OFA-1 и OFA-2 (Takahashi, M. (1984) «A Study On Clinical Significance Of Oncofetal Antigen-1 In Gynecologic Tumors», Nihon Sanka Fujinka Gakkai Zasshi. 36(12):2613-2618); онкостатин M (рецептор онкостатина бета) (патент США №7572896, PCT публикация WO 06/084092); р15 (Gil, J. et al. 2006 Nat Rev Mol Cell Biol. 7(9):667-77); PSA (простатический специфический антиген; Cracco, C.M. et al. 2005 Minerva Urol Nefrol. 57(4):301-11); PSMA (Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); РЕМА (антиген полиморфного эпителиального муцина) (Chu, N.J. et al. (2015) «Nonviral Oncogenic Antigens and the Inflammatory Signals Driving Early Cancer Development as Targets for Cancer Immunoprevention», Clin. Cancer Res. 21(7): 1549-1557); PIPA (патент США №7405061, PCT публикация WO 04/043239); простатическую кислую фосфатазу (Tailor et al. (1990) «Nucleotide Sequence Of Human Prostatic Acid Phosphatase Determined From A Full-Length cDNA Clone», Nucl. Acids Res. 18(16):4928); R24 (Zhou, M. et al. (2008) «Constitutive Overexpression Of A Novel 21 Kda Protein By Hodgkin Lymphoma And Aggressive Non-Hodgkin Lymphomas», Mol. Cancer 7:12); ROR1 (патент США №5843749); Rabbani, H. et al. (2010) «Expression Of ROR1 In Patients With Renal Cancer-A Potential Diagnostic Marker», Iran Biomed. J. 14(3):77-82); сфинголипиды (Hakomori, S. (1998) «Cancer-Associated Glycosphingolipid Antigens: Their Structure, Organization, And Function», Acta Anat. (Basel) 161(1-4):79-90); SSEA-1, SSEA-3 и SSEA-4 (Muramatsu, T. et al. (2004) «Carbohydrate Antigens Expressed On Stem Cells And Early Embryonic Cells», Glycoconj. J. 21(1-2):41-45); sTn (Holmberg, L.A. 2001 Expert Opin Biol Ther. 1(5):881-91); происходящий из Т-клеточного рецептора пептид (Edelson (1998) «Cutaneous T-Cell Lymphoma: A Model For Selective Immunotherapy», Cancer J Sci Am. 4:62-71); T5A7 (Hogg, R.J. et al. (1991) «A monoclonal antibody exhibiting reactivity with both X-hapten- and lactose- bearing glycolipids», Tissue Antigens 37(1):33-38); TAG-72 (Yokota et al. (1992) «Rapid Tumor Penetration Of A Single-Chain Fv And Comparison With Other Immunoglobulin Forms», Cancer Res. 52:3402-3408); TL5 (группа крови A) (Gooi, H.C. et al. (1983) «Monoclonal antibody reactive with the human epidermal-growth-factor receptor recognizes the blood-group- A antigen», Biosci. Rep. 3(11): 1045-1052); рецептор TNF (рецептор TNF-α, рецептор TNF-β или рецептор TNF-γ (van Horssen, R. et al. 2006 Oncologist. 11(4):397-408; Gardnerova, M. et al. 2000 Curr Drug Targets. 1(4):327-64); TRA-1-85 (группа крови H) (Williams, B.P. et al. (1988) «Biochemical and genetic analysis of the OKa blood group antigen», Immunogenetics 27(5):322-329); рецептор трансферрина (патент США №7572895, РСТ публикация WO №05/121179); TSTA, опухолеспецифичный трансплантационный антиген ( et al. (1985) {(Monoclonal Antibodies To Cell Surface Antigens Shared By Chemically Induced Mouse Bladder Carcinomas», Cancer. Res. 45:2210-2188); VEGF-R (O'Dwyer. P.J. 2006 Oncologist. 11(9):992-8); и гаптен Y, Ley (Durrant, L.G. et al. (1989) ((Development Of An ELISA To Detect Early Local Relapse Of Colorectal Саnсеr», Br. J. Cancer 60(4): 533-537).
[000157] Антитела, которые распознают такие опухолевые антигены, известны в данной области техники или могут быть получены с использованием хорошо известных способов, включая те, которые описаны в WO 2002/014870. Примерные антитела, которые содержат домены VL и VH, способные связываться с опухолевым антигеном, и последовательности или полипептидные цепи которых, соответственно, можно применять при конструировании CD137×TA связывающих молекул согласно настоящему изобретению, перечислены в таблице 1. Примерные домены VH и VL для антител, связывающихся с несколькими опухолевыми антигенами, представлены ниже.


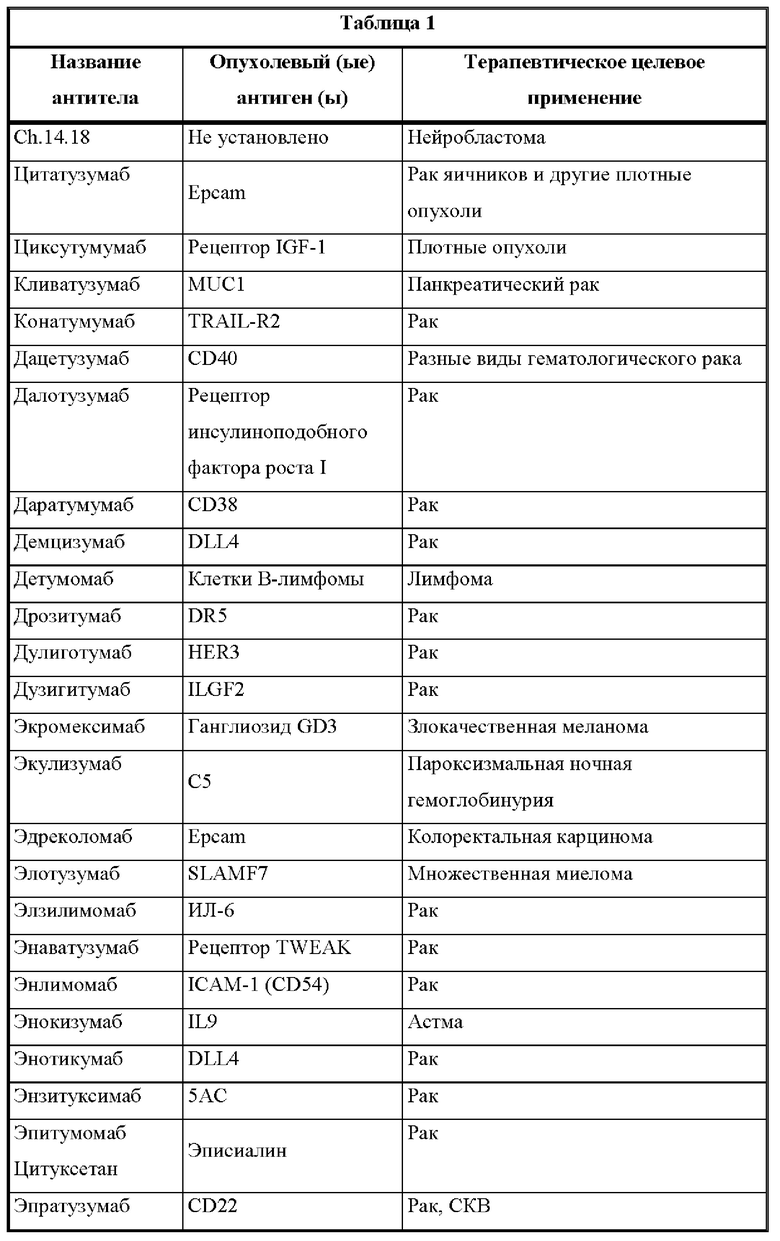
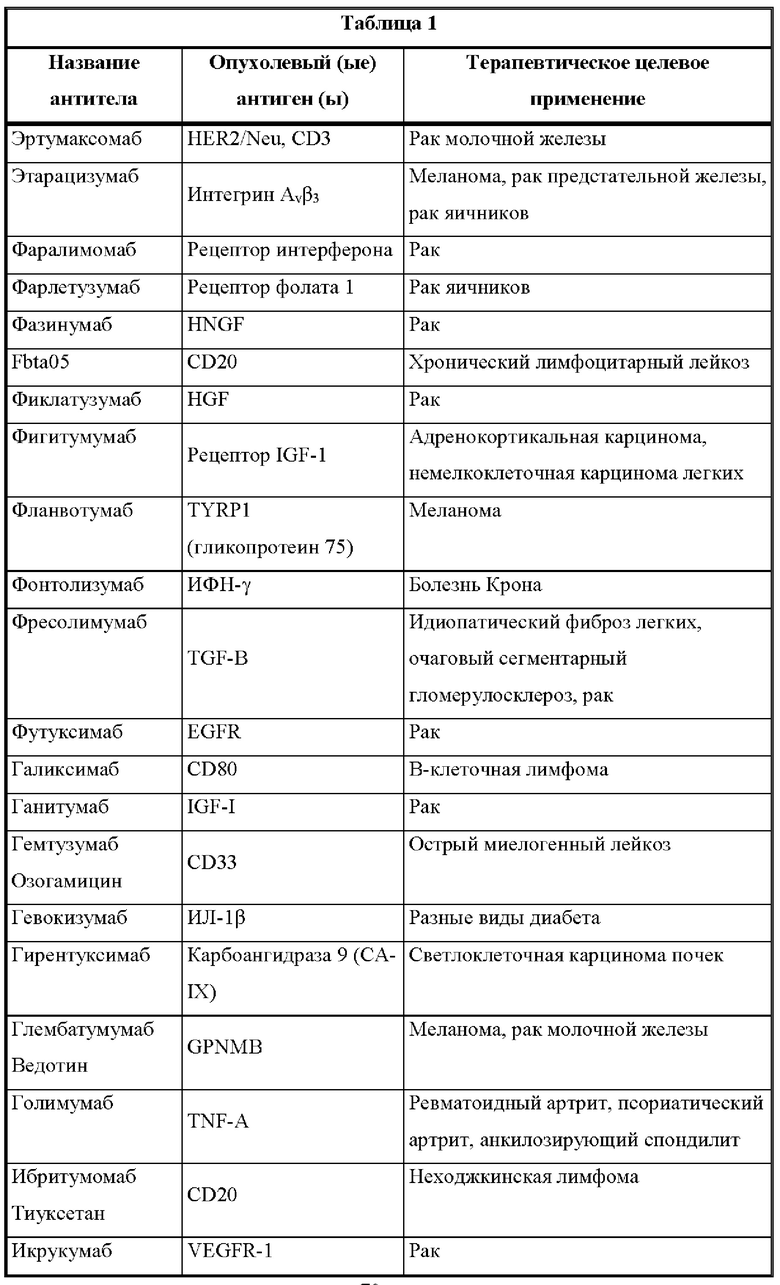

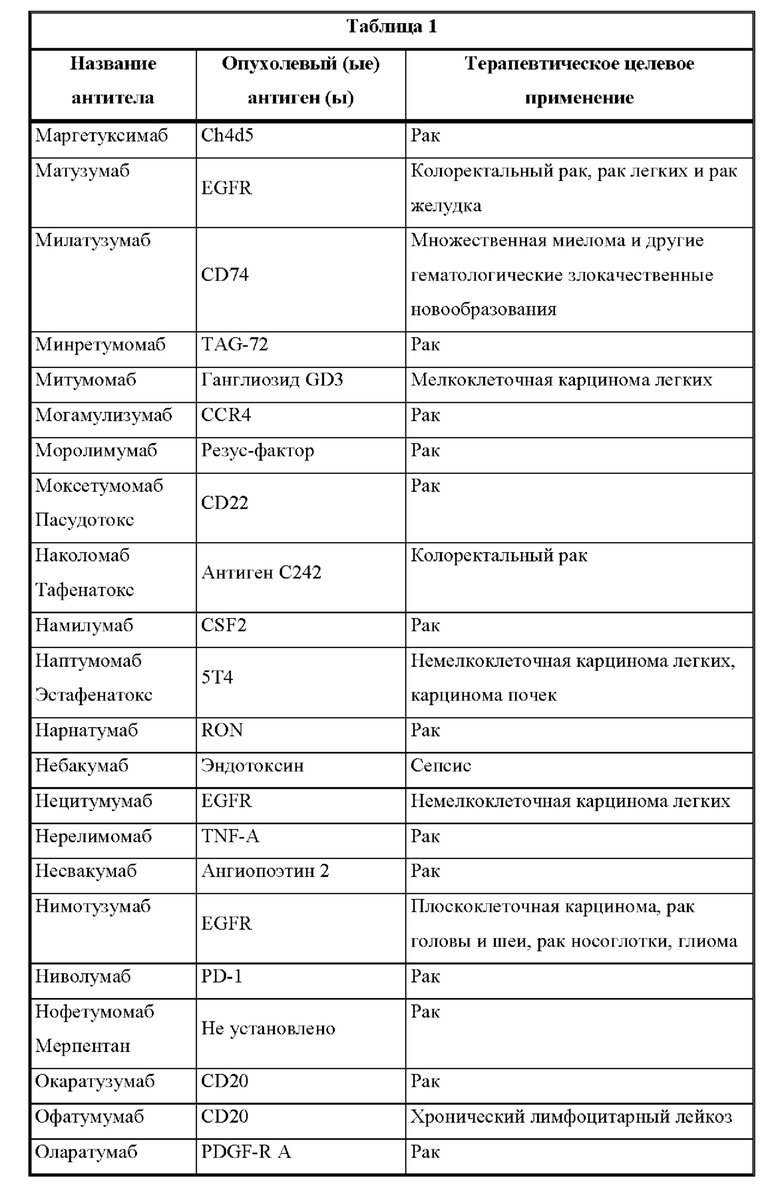



1. Антитела, которые связываются с HER2/neu
[000158] HER2/neu представляет собой рецепторный белок массой 185 кДа, который первоначально был выявлен как продукт трансформирующего гена из нейробластом химически обработанных крыс. HER2/neu интенсивно изучали из-за его роли при некоторых карциномах человека и в процессе развития млекопитающих (Hynes et al. (1994) «The Biology of erbB-2/neu/HER-2 and its Role in Cancer», Biochim. Biophys. Acta 1198:165-184; Dougall et al. (1994) «The neu-Oncogene: Signal Transduction Pathways, Transformation Mechanisms and Evolving Therapies», Oncogene 9:2109-2123; Lee et al. (1995) «Requirement for Neuregulin Receptor erbB2 in Neural and Cardiac Development», Nature 378:394-398).
[000159] Эпитопсвязывающий сайт любого антитела к HER2/neu можно применять в соответствии с настоящим изобретением, и принципы настоящего изобретения проиллюстрированы в отношении опухолевого антигена HER2/neu. Примерные антитела, которые связывают человеческий HER2/neu, включают «маргетуксимаб», «трастузумаб» и «пертузумаб». Маргетуксимаб (также известный как MGAH22; per. номер CAS 1350624-75-7, см., например, патент США №8802093) представляет собой оптимизированное по Fc моноклональное антитело, которое связывается с HER2/neu и опосредует повышенную активность АЗКЦ. Трастузумаб (также известный как rhuMAB4D5 и продаваемый как герцептин (Herceptin®); рег. номер CAS 180288-69-1; см. патент США №5821337) представляет собой гуманизированный вариант антитела 4D5, имеющий константные области IgG1/каппа. Пертузумаб (также известный как rhuMAB2C4 и продаваемый как перджета (Perjeta™); рег. номер CAS 380610-27-5; см., например, WO 2001/000245) представляет собой гуманизированный вариант антитела 2С4, имеющий константные области IgG1/каппа. Также обеспечены дополнительные антитела к HER2/neu.
(а) Маргетуксимаб
[000160] Аминокислотная последовательность домена VH маргетуксимаба представляет собой (SEQ ID NO: 54) (остатки CDRH выделены подчеркиванием):

[000161] Аминокислотная последовательность домена VL маргетуксимаба представляет собой (SEQ ID NO: 55) (остатки CDRL выделены подчеркиванием):


[000162] Аминокислотные последовательности полных тяжелых и легких цепей маргетуксимаба известны в данной области техники (см., например, WHO Drug Information, 2014, Recommended INN: List 71, 28(1):93-94).
(b) Трастузумаб
[000163] Аминокислотная последовательность домена VH трастузумаба представляет собой (SEQ ID NO: 56) (остатки CDRH выделены подчеркиванием):

[000164] Аминокислотная последовательность домена VL трастузумаба представляет собой (SEQ ID NO: 57) (остатки CDRL выделены подчеркиванием):

(c) Пертузумаб
[000165] Аминокислотная последовательность домена VH пертузумаба представляет собой (SEQ ID NO: 58) (остатки CDRH выделены подчеркиванием):

[000166] Аминокислотная последовательность домена VL пертузумаба представляет собой (SEQ ID NO: 59) (остатки CDRL выделены подчеркиванием):

(d) MAB-1 к HER2
[000167] Антитело MAB-1 к HER2 представляет собой мышиное моноклональное антитело к HER2/neu, которое связывает эпитоп HER2/neu, который отличается от эпитопа, распознаваемого маргетуксимабом, трастузумабом и пертузумабом. Аминокислотная последовательность домена VH МАВ-1 к HER2 (называемого в настоящем документе VH МАВ-1 к HER2) представляет собой (SEQ ID NO: 60) (остатки CDRH выделены подчеркиванием):


[000168] Аминокислотные последовательности CDRH из VH MAB-1 к HER2 представляют собой:
CDRH1 (SEQ ID NO: 177): NYGMN
CDRH2 (SEQ ID NO: 178): WINTNIGEPTYTEEFKG
CDRH3 (SEQ ID NO: 179): DDGYGNRVSY
[000169] Аминокислотная последовательность домена VL MAB-1 к HER2 (называемого в настоящем документе VL МАВ-1 к HER2) представляет собой (SEQ ID NO: 61) (остатки CDRL выделены подчеркиванием):

[000170] Аминокислотные последовательности CDRL из VL МАВ-1 к HER2 представляют собой:
CDRL1 (SEQ ID NO: 180): KASQDINSYLS
CDRL2 (SEQ ID NO: 181): RANRLVD
CDRL3 (SEQ ID NO: 182): LQHDEFPWT
(e) Гуманизированное MAB-1 к HER2
[000171] Антитело MAB-1 к HER2 было гуманизировано с образованием антитела hMAB-1 к HER2. Аминокислотная последовательность домена VH такого гуманизированного антитела (VH hMAB-1 к HER2) представляет собой (SEQ ID NO: 62) (остатки CDRH выделены подчеркиванием):

где: X1 представляет собой D или Е и Х2 представляет собой G или I.
[000172] Аминокислотная последовательность домена VL такого гуманизированного антитела (VL hMAB-1 к HER2) представляет собой (SEQ ID NO: 63) (остатки CDRL выделены подчеркиванием):

где: Х3 представляет собой N или S; Х4 представляет собой S, Т или N; Х5 представляет собой V или Q и Х6 представляет собой D, Е или S.
[000173] Были выделены три варианта доменов VH hMAB-1 к HER2: VH1 hMAB-1 к HER2, VH2 hMAB-1 к HER2 и VH3 hMAB-1 к HER2. Аминокислотные последовательности таких вариантов доменов VH hMAB-1 к HER2 представлены ниже.
[000174] Аминокислотная последовательность VH1 hMAB-1 к HER2 представляет собой (SEQ ID NO: 64) (остатки CDRH выделены подчеркиванием; следует отметить, что второй и третий остатки CDRH3 представляют собой D и G, соответственно)

[000175] Аминокислотная последовательность VH2 hMAB-1 к HER2 представляет собой (SEQ ID NO: 65) (остатки CDRH выделены подчеркиванием; следует отметить, что второй и третий остатки CDRH3 представляют собой Е и G, соответственно):

[000176] Аминокислотная последовательность VH3 hMAB-1 к HER2 представляет собой (SEQ ID NO: 66) (остатки CDRH выделены подчеркиванием; следует отметить, что второй и третий остатки CDRH3 представляют собой D и I, соответственно):

[000177] Таким образом, аминокислотные последовательности CDRH1 из VH1 hMAB-1 к HER2, VH2 hMAB-1 к HER2 и VH3 hMAB-1 к HER2 являются идентичными (NYGMN; SEQ ID NO: 177) и аминокислотные последовательности CDRH2 из VH1 hMAB-1 к HER2, VH2 hMAB-1 к HER2 и VH3 hMAB-1 к HER2 являются идентичными (SEQ ID NO: 178). Однако аминокислотные последовательности CDRH3 из VH1 hMAB-1 к HER2, VH2 hMAB-1 к HER2 и VH3 hMAB-1 к HER2 различаются:
CDRH3 из VH1 hMAB-1 к HER2 (SEQ ID NO: 183): DDGYGNRVSY
CDRH3 из VH2 hMAB-1 к HER2 (SEQ ID NO: 184): DEGYGNRVSY
CDRH3 из VH3 hMAB-1 к HER2 (SEQ ID NO: 185): DDIYGNRVSY
[000178] Были выделены три варианта доменов VL hMAB-1 к HER2: VL1 hMAB-1 к HER2, VL2 hMAB-1 к HER2 и VL3 hMAB-1 к HER2. Аминокислотные последовательности таких вариантов доменов VH hMAB-1 к HER2 представлены ниже.
[000179] Аминокислотная последовательность VL1 hMAB-1 к HER2 представляет собой (SEQ ID NO: 67) (остатки CDRL выделены подчеркиванием; следует отметить, что седьмой и восьмой остатки CDRL1 представляют собой N и S, соответственно, и что шестой и седьмой остатки CDRL2 представляют собой V и D, соответственно):

[000180] Аминокислотная последовательность VL2 hMAB-1 к HER2 представляет собой (SEQ ID NO: 68) (остатки CDRL выделены подчеркиванием; следует отметить, что седьмой и восьмой остатки CDRL1 представляют собой N и Т, соответственно, и что шестой и седьмой остатки CDRL2 представляют собой V и Е, соответственно):

[000181] Аминокислотная последовательность VL3 hMAB-1 к HER2 представляет собой (SEQ ID NO: 69) (остатки CDRL выделены подчеркиванием; следует отметить, что седьмой и восьмой остатки CDRL1 представляют собой S и N, соответственно, и что шестой и седьмой остатки CDRL2 представляют собой Q и S, соответственно):

[000182] Таким образом, аминокислотные последовательности CDRL3 из VL1 hMAB-1 к HER2, VL2 hMAB-1 к HER2 и VL3 hMAB-1 к HER2 являются идентичными (LQHDEFPWT; SEQ ID NO: 182). Однако аминокислотные последовательности CDRL1 и CDRL2 из VL1 hMAB-1 к HER2, VL2 hMAB-1 к HER2 и VL3 hMAB-1 к HER2 различаются:
CDRL1 из VL1 hMAB-1 к HER2 (SEQ ID NO: 186): KASQDINSYLS
CDRL1 из VL2 hMAB-1 к HER2 (SEQ ID NO: 187): KASQDINTYLS
CDRL1 из VL3 hMAB-1 к HER2 (SEQ ID NO: 188): KASQDISNYLS
CDRL2 из VL1 hMAB-1 к HER2 (SEQ ID NO: 189): RANRLVD
CDRL2 из VL2 hMAB-1 к HER2 (SEQ ID NO: 190): RANRLVE
CDRL2 из VL3 hMAB-1 к HER2 (SEQ ID NO: 191): RANRLQS
[000183] Любой из таких гуманизированных доменов VH и VL hMAB-1 к HER2, включая любые домены, включенные в общую(ие) последовательность(и) доменов VH и/или VL hMAB-1 к HER2, представленных выше, можно применять для образования антитела, диатела или связывающей молекулы, способных связывать Her2/neu.
(f) Другие антитела к HER2/neu
[000184] В дополнение к указанным выше предпочтительным связывающим молекулам, направленным против HER2/neu, согласно настоящему изобретению предусмотрено применение любой из следующих Her-2-связывающих молекул: 1.44.1; 1.140; 1.43; 1.14.1; 1.100.1; 1.96; 1.18.1; 1.20; 1.39; 1.24; и 1.71.3 (патенты США №№8350011; 8858942; и РСТ публикация патента WO 2008/019290); F5 и С1 (патенты США №№7892554; 8173424; 8974792; и РСТ публикация патента WO 99/55367); а также Her-2-связывающих молекул из публикаций патентов США 2011/0097323, 2013/017114, 2014/0328836, 2016/0130360 и 2016/0257761, и РСТ публикации патента WO 2011/147986), все из этих публикаций включены в настоящий документ посредством ссылки на раскрытие в них связывающих молекул, направленных против HER2/neu.
[000185] Настоящее изобретение, в частности, включает и охватывает CD137×HER2/neu связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH любого из маргетуксимаба, трастузумаба, пертузумаба, hMAB-1 к HER2 или любого из других антител к HER2/neu, обеспеченных в настоящем документе; и более предпочтительно они содержат 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH таких моноклональных антител к HER2/neu.
2. Антитела, которые связываются с EphA2
[000186] Рецепторная тирозинкиназа, рецептор 2 эфрина типа A (EphA2), обычно экспрессируется в местах межклеточного контакта в эпителиальных тканях взрослых, однако недавние исследования показали, что она также сверхэкспрессируется в различных типах эпителиальных карцином, причем наибольший уровень экспрессии EphA2 наблюдается в метастатических поражениях. Высокие уровни экспрессии EphA2 были обнаружены при широком спектре разных видов рака и во многих опухолевых клеточных линиях, включая рак предстательной железы, рак молочной железы, немелкоклеточный рак легких и меланому (Xu, J. et al. (2014) «High EphA2 Protein Expression In Renal Cell Carcinoma Is Associated With A Poor Disease Outcome», Oncol. Lett. Aug 2014; 8(2): 687-692; Miao, B. et al. (2014) «EphA2 is a Mediator ofVemurafenib Resistance and a Novel Therapeutic Target in Melanoma», Cancer Discov. pii: CD-14-0295). EphA2, по-видимому, является не только маркером рака, но, очевидно, постоянно сверхэкспрессируется и функционально изменяется при многих видах рака человека (Chen, P. et al. (2014) «EphA2 Enhances The Proliferation And Invasion Ability Of LnCap Prostate Cancer Cells», Oncol. Lett. 8(1):41-46).
Эпитопсвязывающий сайт любого антитела к EphA2 можно применять в соответствии с настоящим изобретением. Ниже представлены несколько примерных мышиных антител к EphA2, гуманизированные производные таких антител являются особенно предпочтительными.
(a) МАВ-1 к EphA2
[000187] Антитело МАВ-1 к EphA2 представляет собой мышиное моноклональное антитело к EphA2. Аминокислотная последовательность домена VH МАВ-1 к EphA2 представляет собой (SEQ ID NO: 128) (остатки CDR выделены подчеркиванием):

[000188] Аминокислотная последовательность домена VL МАВ-1 к EphA2 представляет собой (SEQ ID NO: 129) (остатки CDR выделены подчеркиванием):

(b) МАВ-2 к EphA2
[000189] Антитело МАВ-2 к EphA2 представляет собой мышиное моноклональное антитело к EphA2. Аминокислотная последовательность домена VH МАВ-2 к EphA2 представляет собой (SEQ ID NO: 130) (остатки CDR выделены подчеркиванием):

[000190] Аминокислотная последовательность домена VL МАВ-2 к EphA2 представляет собой (SEQ ID NO: 131) (остатки CDR выделены подчеркиванием):

(c) МАВ-3 к EphA2
[000191] Антитело МАВ-3 к EphA2 представляет собой мышиное моноклональное антитело к EphA2. Аминокислотная последовательность домена VH МАВ-3 к EphA2 представляет собой (SEQ ID NO: 132) (остатки CDR выделены подчеркиванием):

[000192] Аминокислотная последовательность домена VL МАВ-3 к EphA2 представляет собой (SEQ ID NO: 133) (остатки CDR выделены подчеркиванием):

(d) Другие антитела к EphA2
[000193] В дополнение к указанным выше антителам к EphA2 согласно настоящему изобретению предусмотрено применение любого из следующих антител к EphA2: SPL1, LUCA19, SG5 или LUCA40 (см. РСТ публикацию патента WO 2006/084226); В13 (см. патент США №7101976); D7 (см. патент США №7192698); В-233 и ЕА2 (см. РСТ публикацию патента WO 2003/094859).
[000194] Настоящее изобретение, в частности, включает и охватывает CD137×EphA2 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH моноклональных антител к EphA2, МАВ-1 к EphA2, МАВ-2 к EphA2 и МАВ-3 к EphA2.
3. Антитела, которые связываются с 5Т4
[000195] Онкофетальный белок, 5Т4, представляет собой ассоциированный с опухолью белок, который экспонирован на клеточной мембране многих видов карциномы, включая карциному почек, карциному толстой кишки, карциному предстательной железы, карциному легких и острый лимфобластный лейкоз (см. Boghaert, E.R. et al. (2008) «The Oncofetal Protein, 5T4, Is A Suitable Target For Antibody-Guided Anti-Cancer Chemotherapy With Calicheamicin», Int. J. Oncol. 32(1):221-234; Eisen, T. et al. (2014) «Naptumomab Estafenatox: Targeted Immunotherapy with a Novel Immunotoxin», Curr. Oncol. Rep. 16:370, pp. 1-6). Эпитопсвязывающий сайт любого антитела к 5Т4 можно применять в соответствии с настоящим изобретением. Ниже представлены два примерных антитела к 5Т4, «МАВ-1 к 5Т4» и «МАВ-2 к 5Т4». Дополнительные антитела к 5Т4 описаны в данной области техники (см., например, патенты США №№8084249; 8409577; 8759495; 8409577; РСТ публикации №№WO 2013/041687; WO 2014/137931; WO 2016/022939).
(а) МАВ-1 к 5Т4
[000196] Аминокислотная последовательность домена VH МАВ-1 к 5Т4 представляет собой (SEQ ID NO: 134) (остатки CDR выделены подчеркиванием):

[000197] Аминокислотная последовательность домена VL МАВ-1 к 5Т4 представляет собой (SEQ ID NO: 135) (остатки CDR выделены подчеркиванием):

(b) MAB-2 к 5T4
[000198] Аминокислотная последовательность домена VH МАВ-2 к 5T4 представляет собой (SEQ ID NO: 136) (остатки CDR выделены подчеркиванием):

[000199] Аминокислотная последовательность домена VL MAB-2 к 5T4 представляет собой (SEQ ID NO: 137) (остатки CDR выделены подчеркиванием):

[000200] Настоящее изобретение, в частности, включает и охватывает CD137×5T4 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH моноклональных антител к 5Т4, МАВ-1 к 5Т4 или МАВ-2 к 5Т4, или любого из антител к 5Т4, обеспеченных в WO 2007/106744; WO 2013/041687 или WO 2015/184203.
4. Антитела, которые связываются с В7-Н3
[000201] В7-Н3 представляет собой опухолевый антиген, который сверхэкспрессируется на широком спектре типов плотных опухолей и является членом семейства В7 молекул, которые участвуют в иммунной регуляции (см. патент США №8802091; US 2014/0328750; US 2013/0149236; Loo, D. et al. (2012) «Development Of An Fc-Enhanced Anti-B7-H3 Monoclonal Antibody With Potent Antitumor Activity», Clin. Cancer Res. 18(14):3834-3845). В частности, несколько независимых исследований показали, что клетки злокачественных опухолей человека (например, опухолевые клетки нейробластом и рака желудка, рака яичников и немелкоклеточного рака легких) проявляют существенное повышение экспрессии белка В7-Н3 и что такая повышенная экспрессия была связана с повышенной степенью тяжести заболевания (Zang, X. et al. (2007) «TheB7 Family And Cancer Therapy: Costimulation And Coinhibition», Clin. Cancer Res. 13:5271-5279), это свидетельствует о том, что В7-Н3 используется опухолями как путь уклонения от надзора иммунной системы (Hofmeyer, K. et al. (2008) «The Contrasting Role Qf B7-H3», Proc. Natl. Acad. Sci. (U.S.A.) 105(30): 10277-10278).
[000202] Эпитопсвязывающий сайт любого антитела к В7-Н3 можно применять в соответствии с настоящим изобретением. Одним примерным антителом, которое связывает человеческий В7-Н3, является «эноблитузумаб». Эноблитузумаб (также известный как MGA271; рег. номер CAS 1353485-38-7; см., например, патент США №8802091) представляет собой Fc-оптимизированное моноклональное антитело, которое связывается с В7-Н3 и обеспечивает повышенную активность АЗКЦ. Также представлены дополнительные примерные антитела к В7-Н3.
(a) Эноблитузумаб
[000203] Аминокислотная последовательность домена VH эноблитузумаба представляет собой (SEQ ID NO: 138) (остатки CDRH выделены подчеркиванием):

[000204] Аминокислотная последовательность VL домена эноблитузумаба представляет собой (SEQ ID NO: 139) (остатки CDRL выделены подчеркиванием):

(b) BRCA69D
[000205] Антитело BRCA69D представляет собой мышиное моноклональное антитело к В7-Н3. Аминокислотная последовательность домена VH BRCA69D (SEQ ID NO: 140) представлена ниже (остатки CDRH выделены подчеркиванием).

[000206] Аминокислотная последовательность домена VL BRCA69D (SEQ ID NO: 141) представлена ниже (остатки CDRL выделены подчеркиванием).

(c) Гуманизированное BRCA69D
[000207] Антитело BRCA69D было гуманизировано с получением двух вариантов доменов VH, VH1 hBRCA69D и VH2 hBRCA69D; и двух вариантов доменов VL, VL1 hBRCA69D и VL2 hBRCA69D, которые можно применять в любой комбинации VH/VL для получения функционального гуманизированного связывающего домена. Аминокислотные последовательности таких гуманизированных вариантов представлены ниже.
[000208] Аминокислотная последовательность домена VH из VH1 hBRCA69D представляет собой (SEQ ID NO: 142) (остатки CDRH выделены подчеркиванием):

[000209] Аминокислотная последовательность домена VH из VH2 hBRCA69D представляет собой (SEQ ID NO: 143) (остатки CDRH выделены подчеркиванием):

[000210] Аминокислотная последовательность домена VL из VL1 hBRCA69D представляет собой (SEQ ID NO: 144) (остатки CDRL выделены подчеркиванием).

[000211] Аминокислотная последовательность домена VL из VL2 hBRCA69D представляет собой (SEQ ID NO: 145) (остатки CDRL выделены подчеркиванием).

(d) PRCA157
[000212] Антитело PRCA157 представляет собой мышиное моноклональное антитело к В7-Н3. Аминокислотная последовательность домена VH PRCA157 представляет собой (SEQ ID NO: 146) (остатки CDRH выделены подчеркиванием).

[000213] Аминокислотная последовательность домена VL PRCA157 представляет собой (SEQ ID NO: 147) (остатки CDRH выделены подчеркиванием).

(e) Гуманизированное антитело PRCA157
[000214] Антитело PRCA157 было гуманизировано с получением одного гуманизированного домена VH, VH1 hPRCA157, и одного гуманизированного домена VL, VL1 hPRCA157, для получения функционального гуманизированного связывающего домена. Аминокислотные последовательности гуманизированного PRCA157 представлены ниже.
[000215] Аминокислотная последовательность домена VH из VH1 hPRCA157 представляет собой (SEQ ID NO: 202) (остатки CDRH выделены подчеркиванием):

[000216] Аминокислотная последовательность домена VL из VL1 hPRCA157 представляет собой (SEQ ID NO: 203) (остатки CDRL выделены подчеркиванием):

(f) Другие антитела к B7-H3
[000217] В дополнение к указанным выше предпочтительным связывающим молекулам, направленным против В7-Н3, согласно настоящему изобретению предусмотрено применение любой из следующих связывающих молекул, направленных против В7-Н3: LUCA1; BLA8; РА20; или SKN2 (см. патенты США №№7527969; 8779098 и РСТ публикацию патента WO 2004/001381); М30; сМ30; M30-H1-L1; М30-Н1-L2; M30-H1-L3; M30-H1-L4; M30-H1-L5; M30-H1-L6; M30-H1-L7; M30-H4-L1; М30-H4-L2; M30-H4-L3; и M30-H4-L4 (см. публикацию патента США 2013/0078234 и РСТ публикацию патента WO 2012/147713); и 8Н9 (см. патенты США №№7666424; 7737258; 7740845; 8148154; 8414892; 8501471; 9062110; публикацию патента США 2010/0143245 и РСТ публикацию патента WO 2008/116219).
[000218] Настоящее изобретение, в частности, включает и охватывает CD137×B7-H3 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH любого из BRCA69D, гуманизированного BRCA69D, PRCA157, гуманизированного PRCA157 или эноблитузумаба или любого другого антитела к В7-Н3, обеспеченного в настоящем документе; и более предпочтительно имеют 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH таких моноклональных антител к В7-Н3.
5. Антитела, которые связываются с GpA33
[000219] Трансмембранный гликопротеин А33 массой 43 кДа (gpA33) экспрессируется в >95% случаев всех колоректальных карцином (Heath, J.K. et al. (1997) «The Human А33 Antigen Is A Transmembrane Glycoprotein And A Novel Member Of The Immunoglobulin Superfamily», Proc. Natl. Acad. Sci. (U.S.A.) 94(2):469-474; Ritter, G. et al. (1997) «Characterization Of Posttranslational Modifications Of Human A33 Antigen, A Novel Palmitoylated Surface Glycoprotein Of Human Gastrointestinal Epitheliums, Biochem. Biophys. Res. Commun. 236(3):682-686; Wong, N.A. et al. (2006) «ЕрСАМ and gpA33 Are Markers Of Barrett's Metaplasias, J. Clin. Pathol. 59(3):260-263). Эпитопсвязывающий сайт любого антитела к gpA33 можно применять в соответствии с настоящим изобретением. Примерное антитело к gpA33 («МАВ-1 к gpA33») представлено ниже.
[000220] Аминокислотная последовательность домена VH МАВ-1 к gpA33 представляет собой (SEQ ID NO: 148) (остатки CDR выделены подчеркиванием):

[000221] Аминокислотная последовательность домена VL МАВ-1 к gpA33 представляет собой (SEQ ID NO: 149) (остатки CDR выделены подчеркиванием):

[000222] Настоящее изобретение, в частности, включает и охватывает CD137×gpA33 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH моноклональных антител к gpA33 МАВ-1 к gpA33 или любого из моноклональных антител к gpA33, обеспеченных в WO 2015/026894.
6. Антитела, которые связываются с СЕАСАМ5 и СЕАСАМ6
[000223] Было обнаружено, что родственные карциноэмбриональному антигену молекулы клеточной адгезии 5 и 6 (СЕАСАМ5) и 6 (СЕАСАМ6) ассоциированы с различными типами рака, включая медуллярный рак щитовидной железы, колоректальный рак, рак поджелудочной железы, гепатоцеллюлярную карциному, рак желудка, рак легких, разные виды рака головы и шеи, рак мочевого пузыря, рак предстательной железы, рак матки, рак эндометрия, рак молочной железы, гематопоэтический рак, лейкоз и рак яичников (РСТ публикация №WO 2011/034660), и в частности, колоректальный рак, рак желудочно-кишечного тракта, рак поджелудочной железы, немелкоклеточный рак легких (NSCL), карциному молочной железы, карциному щитовидной железы, карциному желудка, карциному яичников и карциному матки (Zheng, С. et al. (2011) «А Novel Anti-CEACAM5 Monoclonal Antibody, CC4, Suppresses Colorectal Tumor Growth and Enhances NK Cells-Mediated Tumor Immunity», PLoS One 6(6):e21146, pp. 1-11). Было обнаружено, что CEACAM5 сверхэкспрессируется в 90% случаев рака желудочно-кишечного тракта, колоректального рака и рака поджелудочной железы, в 70% клеток немелкоклеточного рака легких и в 50% случаев рака молочной железы (Thompson, J.A. et al. (1991) «Carcinoembryonic Antigen Gene Family: Molecular Biology And Clinical Perspectives», J. Clin. Lab. Anal. 5:344-366). Сверхэкспрессированная родственная карциноэмбриональному антигену молекула клеточной адгезии 6 (СЕАСАМ6) играет важную роль в инвазии и метастазировании различных видов рака человека, включая медуллярный рак щитовидной железы, колоректальный рак, рак поджелудочной железы, гепатоцеллюлярную карциному, рак желудка, рак легких, разные виды рака головы и шеи, рак мочевого пузыря, рак предстательной железы, рак матки, рак эндометрия, рак молочной железы, гематопоэтический рак, лейкоз и рак яичников (РСТ публикация №WO 2011/034660; Deng, X., et al. (2014) «Expression Profiling Of CEACAM6 Associated With The Tumorigenesis And Progression In Gastric Adenocarcinomas, Genet. Mol. Res. 13(3):7686-7697; Cameron, S. et al. (2012) «Focal Overexpression Of CEACAM6 Contributes To Enhanced Tumourigenesis In Head And Neck Cancer Via Suppression Of Apoptosis», Mol. Cancer 11:74, pp. 1-11; Chapin, C. et al. (2012) «Distribution And Surfactant Association Of Carcinoembryonic Cell Adhesion Molecule 6 In Human Lung», Amer. J. Physiol. Lung Cell. Mol. Physiol. 302(2):L216-L25; Riley, C.J. et al. (2009) «Design And Activity Of A Murine And Humanized Anti-CEACAM6 Single-Chain Variable Fragment In The Treatment Of Pancreatic Cancer», Cancer Res. 69(5):1933-1940; Lewis-Wambi, J.S. et al. (2008) «Overexpression Of CEACAM6 Promotes Migration And Invasion Of Oestrogen-Deprived Breast Cancer Cells», Eur. J. Cancer 44(12): 1770-1779; Blumenthal, R.D. et al. (2007) «Expression Patterns Of CEACAM5 And CEACAM6 In Primary And Metastatic Cancers», BMC Cancer. 7:2, pp. 1-15). Эпитопсвязывающий сайт любого антитела к СЕАСАМ5/СЕАСАМ6 можно применять в соответствии с настоящим изобретением. Примерные антитела к СЕАСАМ5/СЕАСАМ6 представлены ниже,
(а) 16С3
[000224] Аминокислотная последовательность домена VH гуманизированного антитела к СЕАСАМ5/СЕАСАМ6 16С3 (ЕР 2585476) представляет собой (SEQ ID NO: 150) (остатки CDR выделены подчеркиванием):

[000225] Аминокислотная последовательность домена VL гуманизированного антитела к СЕАСАМ5/СЕАСАМ6 16С3 (ЕР 2585476) представляет собой (SEQ ID NO: 151) (остатки CDR выделены подчеркиванием):

(b) hMN15
[000226] Аминокислотная последовательность домена VH гуманизированного антитела к СЕАСАМ5/СЕАСАМ6 hMN15 (WO 2011/034660) представляет собой (SEQ ID NO: 152) (остатки CDR выделены подчеркиванием):

[000227] Аминокислотная последовательность домена VL гуманизированного антитела к СЕАСАМ5/СЕАСАМ6 hMN15 (WO 2011/034660) представляет собой (SEQ ID NO: 153) (остатки CDR выделены подчеркиванием):

[000228] Настоящее изобретение, в частности, включает и охватывает CD137×CEACAM5/CEACAM6 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH моноклональных антител к СЕАСАМ5/СЕАСАМ6 16С3 или hMN15.
7. Антитела, которые связываются с CD19
[000229] CD19 (поверхностный антиген В-лимфоцитов В4, номер доступа в Genbank М28170) представляет собой компонент комплекса В-клеточного рецептора (BCR) и является положительным регулятором передачи сигналов с участием В-клеток, который модулирует порог активации В-клеток и гуморального иммунитета. CD19 является одним из наиболее повсеместно экспрессируемых антигенов в линии В-клеток и экспрессируется на >95% злокачественных В-клеточных опухолей, включая острый лимфобластный лейкоз (ОЛЛ), хронический лимфоцитарный лейкоз (ХЛЛ) и неходжкинскую лимфому (NHL). В частности, экспрессия CD19 поддерживается на В-клеточных лимфомах, которые становятся устойчивыми к терапии, направленной против CD20 (Davis et al. (1999) «Therapy of B-cell Lymphoma With Anti-CD20 Antibodies Can Result In The Loss Of CD20 Antigen Expression.» Clin Cancer Res, 5:611-615, 1999). CD19 также был предложен в качестве мишени для лечения аутоиммунных заболеваний (Tedder (2009) «CD19: А Promising В Cell Target For Rheumatoid Arthritis», Nat. Rev. Rheumatol. 5:572-577).
[000230] Примерное антитело, которое связывается с человеческим CD19 и которое можно применять в настоящем изобретении, представляет собой антитело к CD19, раскрытое в WO 2016/048938 (называемое в настоящем документе «МАВ-1 к CD19»).
[000231] Аминокислотная последовательность домена VH МАВ-1 к CD19 (SEQ ID NO: 204) представлена ниже (остатки CDRH выделены подчеркиванием):

[000232] Аминокислотная последовательность домена VL МАВ-1 к CD19 (SEQ ID NO: 205) представлена ниже (остатки CDRL выделены подчеркиванием):

[000233] Настоящее изобретение, в частности, включает и охватывает CD137×CD19 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH моноклонального антитела к CD19 МАВ-1 к CD19 или любого из антител к CD19, раскрытых в патенте США US7112324, или присутствующих в блинатумомабе (блинцито (BLINCYTO®); аминокислотная последовательность может быть найдена в WHO Drug Information, 2009, Recommended INN: List 62, 23(3):240-241) и дувортуксизумабе (известном как MGD011; аминокислотная последовательность может быть найдена в WHO Drug Information, 2016, Proposed INN: List 116, 30(4):627-629). 8. Антитела, которые связываются с CD123
[000234] CD123 (альфа-субъединица рецептора интерлейкина 3, IL-3Ra) представляет собой молекулу массой 40 кДа и является частью рецепторного комплекса интерлейкина 3 (Stomski, F.C. et al. (1996) «Human Interleukin-3 (IL-3) Induces Disulftde-Linked IL-3 Receptor Alpha- And Beta-Chain Heterodimerization, Which Is Required For Receptor Activation But Not High-Affinity Bindings Mol. Cell. Biol. 16(6):3035-3046). Интерлейкин 3 (ИЛ-3) запускает раннюю дифференцировку мультипотентных стволовых клеток в клетки эритроидных, миелоидных и лимфоидных предшественников. Сообщалось, что CD123 сверхэкспрессируется на злокачественных клетках при широком спектре гематологических злокачественных новообразований, включая острый миелолейкоз (ОМЛ) и миелодиспластический синдром (МДС) ( , L. et al. (2001) «Interleukin-3 Receptor Alpha Chain (CD123) Is Widely Expressed In Hematologic Malignancies», Haematologica 86(12):1261-1269). Сверхэкспрессия CD123 ассоциирована с более неблагоприятным прогнозом при ОМЛ (Tettamanti, M.S. et al. (2013) «Targeting Of Acute Myeloid Leukaemia By Cytokine-Induced Killer Cells Redirected With A Novel CD123-Specific Chimeric Antigen Receptors», Br. J. Haematol. 161:389-401).
, L. et al. (2001) «Interleukin-3 Receptor Alpha Chain (CD123) Is Widely Expressed In Hematologic Malignancies», Haematologica 86(12):1261-1269). Сверхэкспрессия CD123 ассоциирована с более неблагоприятным прогнозом при ОМЛ (Tettamanti, M.S. et al. (2013) «Targeting Of Acute Myeloid Leukaemia By Cytokine-Induced Killer Cells Redirected With A Novel CD123-Specific Chimeric Antigen Receptors», Br. J. Haematol. 161:389-401).
[000235] Примерное антитело, которое связывается с человеческим CD123 и которое можно применять в настоящем изобретении, представляет собой «МАВ-1 к CD123» (см., например, РСТ публикацию патента WO 2015/026892).
[000236] Аминокислотная последовательность домена VH МАВ-1 к CD123 (SEQ ID NO: 206) представлена ниже (остатки CDRH выделены подчеркиванием):

[000237] Аминокислотная последовательность домена VL МАВ-1 к CD123 (SEQ ID NO: 207) представлена ниже (остатки CDRL выделены подчеркиванием):

[000238] Настоящее изобретение, в частности, включает и охватывает CD137×CD123 связывающие молекулы, которые содержат домен VL и/или VH, и/или 1, 2 или все 3 из CDRL области VL и/или 1, 2 или все 3 из CDRH домена VH моноклонального антитела к CD123, МАВ-1 к CD123, или любого из антител к CD123, раскрытых в US 2017/081424 и WO 2016/036937, или присутствующих в JNJ-63709178 (Johnson & Johnson, см. также WO 2016/036937) и XmAb 14045 (Xencor, см. также US 2017/081424).
G. Предпочтительные вариабельные домены CD137
[000239] Эпитопсвязывающий сайт любого антитела к CD137 можно применять в соответствии с настоящим изобретением. Примерные антитела, которые связывают человеческий CD137, включают «урелумаб» и «утомилумаб», которые в настоящее время проходят оценку в клинических исследованиях с участием человека. Урелумаб (также известный как BMS-663513, обозначенный в настоящем документе «МАВ-1 к CD137») представляет собой полностью человеческое моноклональное антитело, имеющее константные области IgG4/каппа (см. патент США №8137667). Утомилумаб (также известный как PF-05082566, обозначенный в настоящем документе «МАВ-2 к CD137») представляет собой полностью человеческое моноклональное антитело, имеющее константные области IgC2/лямбда (см. патент США №8337850). Дополнительные примерные антитела, которые являются иммуноспецифичными для человеческого CD137 (обозначены «МАВ-3 к CD137», «МАВ-4 к CD137» и «МАВ-5 к CD137»), представлены ниже.
1. МАВ-1 к CD137
[000240] Аминокислотная последовательность домена VH МАВ-1 к CD137 (VH МАВ-1 к CD137) представляет собой (SEQ ID NO: 70) (остатки CDRH выделены подчеркиванием):

[000241] Аминокислотная последовательность домена VL МАВ-1 к CD137 (VL МАВ-1 к CD137) представляет собой (SEQ ID NO: 71) (остатки CDRL выделены подчеркиванием):

2. МАВ-2 к CD137
[000242] Аминокислотная последовательность домена VH МАВ-2 к CD137 (VH к МАВ-2 к CD137) представляет собой (SEQ ID NO: 72) (остатки CDRH выделены подчеркиванием):

[000243] Аминокислотная последовательность домена VL МАВ-2 к CD137 (VL к МАВ-2 к CD137) представляет собой (SEQ ID NO: 73) (остатки CDRL выделены подчеркиванием):

3. МАВ-3 к CD137
[000244] МАВ-3 к CD137 представляет собой новое мышиное моноклональное антитело. Аминокислотная последовательность домена VH МАВ-3 к CD137 (VH МАВ-3 к CD137) представляет собой (SEQ ID NO: 74) (остатки CDRH выделены подчеркиванием):

[000245] Аминокислотные последовательности CDRH из VH МАВ-3 к CD137 представляют собой:
CDRH1 (SEQ ID NO: 154): SYWIN
CDRH2 (SEQ ID NO: 155): NIYPSDSYTNYNQKFKD
CDRH3 (SEQ ID NO: 156): DYGSSYSFDY
[000246] Аминокислотная последовательность домена VL МАВ-3 к CD137 (VL МАВ-3 к CD137) представляет собой (SEQ ID NO: 75) (остатки CDRL выделены подчеркиванием):

[000247] Аминокислотные последовательности CDRL из VL МАВ-3 к CD137 представляют собой:
CDRL1 (SEQ ID NO: 157): RPSQDISNYLN
CDRL2 (SEQ ID NO: 158): YTSRLRS
CDRL3 (SEQ ID NO: 159): QQGDTLPYT
4. hMAB-3 к CD137
[000248] Антитело МАВ-3 к CD137 было гуманизировано с образованием антитела hMAB-3 к CD137. Аминокислотная последовательность домена VH такого гуманизированного антитела (VH1 hMAB-3 к CD137) представляет собой (SEQ ID NO: 76) (остатки CDRH выделены подчеркиванием):

[0002491 Домен VH гуманизированного антитела hMAB-3 к CD137 (VH1 hMAB-3 к CD137) был оптимизирован для получения домена VH, имеющего аминокислотную последовательность SEQ ID NO: 77:
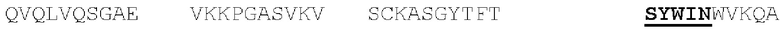
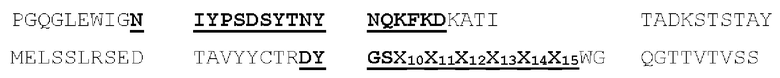
где: X10X11X12X13X14X15 представляют собой:
AYSFHP (1A; SEQ ID NO: 78), или
AYSMST (1B; SEQ ID NO: 79), или
AYSYSL (1С; SEQ ID NO: 80), или
SYSYNV (1D; SEQ ID NO: 81).
[000250] Как указано выше, соответствующая исходная последовательность (присутствующая в SEQ ID NO: 74) представляет собой SYSFDY (SEQ ID NO: 160).
[000251] Домен VL гуманизированного антитела hMAB-3 к CD137 (VL hMAB-3 к CD137) был оптимизирован для получения домена VL, имеющего аминокислотную последовательность SEQ ID NO: 82:

где: X7 представляет собой D или G; X8 представляет собой G или K.
[000252] Были выделены четыре варианта доменов VH1 hMAB-3 к CD137: VH1A hMAB-3 к CD137, VH1B hMAB-3 к CD137, VH1C hMAB-3 к CD137 и VH1D hMAB-3 к CD137. Аминокислотные последовательности таких вариантов доменов VH1 hMAB-3 к CD137 представлены ниже.
[000253] Аминокислотная последовательность VH1A hMAB-3 к CD137 представляет собой (SEQ ID NO: 83) (остатки CDRH выделены подчеркиванием; следует отметить, что остатки 5-10 CDR3H представляют собой AYSFHP (SEQ ID NO: 78):

[000254] Аминокислотная последовательность VH1B hMAB-3 к CD137 представляет собой (SEQ ID NO: 84) (остатки CDRH выделены подчеркиванием; следует отметить, что остатки 5-10 CDR3H представляют собой AYSMST (SEQ ID NO: 79):

[000255] Аминокислотная последовательность VH1C hMAB-3 к CD137 представляет собой (SEQ ID NO: 85) (остатки CDRH выделены подчеркиванием; следует отметить, что остатки 5-10 CDR3H представляют собой AYSYSL (SEQ ID NO: 80):

[000256] Аминокислотная последовательность VH1D hMAB-3 к CD137 представляет собой (SEQ ID NO: 86) (остатки CDRH выделены подчеркиванием; следует отметить, что остатки 5-10 CDR3H представляют собой SYSYNV (ID; SEQ ID NO: 81):

[000257] Таким образом, аминокислотные последовательности CDRH1 из VH1A hMAB-3 к CD137, VH1B hMAB-3 к CD137, VH1C hMAB-3 к CD137 и VH1D hMAB-3 к CD137 являются идентичными (SEQ ID NO: 154), и аминокислотные последовательности CDRH2 из VH1A hMAB-3 к CD137, VH1B hMAB-3 к CD137, VH1C hMAB-3 к CD137 и VH1D hMAB-3 к CD137 являются идентичными (SEQ ID NO: 155). Однако аминокислотные последовательности CDRH3 из VH1A hMAB-3 к CD137, VH1B hMAB-3 к CD137, VH1C hMAB-3 к CD137 и VH1D hMAB-3 к CD137 различаются:
CDRH3 из VH1A hMAB-3 к CD137 (SEQ ID NO: 161): DYGSAYSFHP
CDRH3 из VH1B hMAB-3 к CD137 (SEQ ID NO: 162): DYGSAYSMST
CDRH3 из VH1C hMAB-3 к CD137 (SEQ ID NO: 163): DYGSAYSYSL
CDRH3 из VH1D hMAB-3 к CD137 (SEQ ID NO: 164): DYGSSYSYNV
[000258] Были выделены три варианта доменов VL hMAB-3 к CD137: VL1 hMAB-3 к CD137, VL2 hMAB-3 к CD137 и VL3 hMAB-3 к CD137. Аминокислотные последовательности таких вариантов доменов VL hMAB-3 к CD137 представлены ниже.
[000259] Аминокислотная последовательность VL1 hMAB-3 к CD137 представляет собой (SEQ ID NO: 87) (остатки CDRL выделены подчеркиванием; следует отметить, что остатки 41 и 42 представляют собой D и G, соответственно):

[000260] Аминокислотная последовательность VL2 hMAB-3 к CD137 представляет собой (SEQ ID NO: 88) (остатки CDRL выделены подчеркиванием; следует отметить, что остатки 41 и 42 оба представляют собой G):
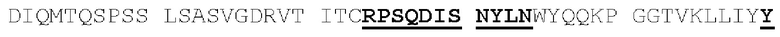

[000261] Аминокислотная последовательность VL3 hMAB-3 к CD137 представляет собой (SEQ ID NO: 89) (остатки CDRL выделены подчеркиванием; следует отметить, что остатки 41 и 42 представляют собой D и K, соответственно):

[000262] Домен VH из VH1B hMAB-3 к CD137 был деиммунизирован, как описано в примерах ниже. Аминокислотные последовательности трех полученных деиммунизированных доменов VH, обозначенных VH1E hMAB-3 к CD137, VH1F hMAB-3 к CD137 и VH1G hMAB-3 к CD137, имеющих замену аминокислотного остатка в положении 38 и/или 48 согласно Kabat, представлены ниже. Конкретно предусмотрено, что выявленные замены R38K и/или I48A могут быть включены в любой из раскрытых доменов VH hMAB-3 к CD137. Согласно конкретному варианту реализации настоящего изобретения замена аминокислоты K38R включена в SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85 или SEQ ID NO: 86.
[000263] Аминокислотная последовательность VH1E hMAB-3 к CD137, содержащая K38R, представляет собой (SEQ ID NO: 208) (остатки CDRH выделены подчеркиванием; замена выделена двойным подчеркиванием):

[000264] Аминокислотная последовательность VH1F hMAB-3 к CD137, содержащая I48A, представляет собой (SEQ ID NO: 209) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000265] Аминокислотная последовательность VH1G hMAB-3 к CD137, содержащая K38R/I48A (SEQ ID NO: 210) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):
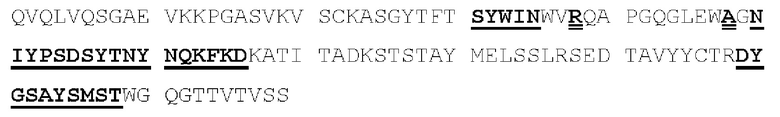
[000266] Домен VL из VL3 hMAB-3 к CD137 был деиммунизирован, как описано в примерах ниже. Аминокислотные последовательности двенадцати из полученных деиммунизированных доменов VL, обозначенных VL4-VL15 hMAB-3 к CD137, имеющих замену остатка аминокислоты в положении 24, 25, 44, 48, 52 и/или 54 согласно Kabat, представлены ниже. Конкретно предусмотрено, что выявленные замены, R24Q, Р25А, V44A, I48A, S52T, S52G и/или L54A, могут быть включены в любой из раскрытых доменов VL hMAB-3 к CD137, представленных выше, с получением деиммунизированного домена VL. Согласно конкретному варианту реализации настоящего изобретения замены аминокислот R24Q, Р25А, I48A, S52G и L54A включены в SEQ ID NO: 82, SEQ ID NO: 87, SEQ ID NO: 88 или SEQ ID NO: 89. Согласно другому конкретному варианту реализации замены аминокислот R24Q, Р25А, 148A, S52T и L54A включены в SEQ ID NO: 82, SEQ ID NO: 87, SEQ ID NO: 88 или SEQ ID NO: 89.
[000267] Аминокислотная последовательность VL4 hMAB-3 к CD137, содержащая R24Q/P25A, представляет собой (SEQ ID NO: 211) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000268] Аминокислотная последовательность VL5 hMAB-3 к CD137, содержащая V44A, представляет собой (SEQ ID NO: 212) (остатки CDRH выделены подчеркиванием; замена выделена двойным подчеркиванием):

[000269] Аминокислотная последовательность VL6 hMAB-3 к CD137, содержащая L54A, представляет собой (SEQ ID NO: 213) (остатки CDRH выделены подчеркиванием; замена выделена двойным подчеркиванием):

[000270] Аминокислотная последовательность VL7 hMAB-3к CD137, содержащая R24Q/P25A/V44A, представляет собой (SEQ ID NO: 214) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

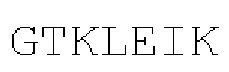
[000271] Аминокислотная последовательность VL8 hMAB-3к CD137, содержащая R24Q/P25A/V44A/L54A, представляет собой (SEQ ID NO: 215) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000272] Аминокислотная последовательность VL9 hMAB-3 к CD137, содержащая R24Q/P25A/L54A, представляет собой (SEQ ID NO: 216) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000273] Аминокислотная последовательность VL10 hMAB-3 к CD137, содержащая S52T, представляет собой (SEQ ID NO: 217) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000274] Аминокислотная последовательность VL11 hMAB-3 к CD137, содержащая S52G, представляет собой (SEQ ID NO: 218) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000275] Аминокислотная последовательность VL12 hMAB-3 к CD137, содержащая I48A/S52T, представляет собой (SEQ ID NO: 219) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000276] Аминокислотная последовательность VL13 hMAB-3 к CD137, содержащая I48A/S52G, представляет собой (SEQ ID NO: 220) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000277] Аминокислотная последовательность VL14 hMAB-3к CD137, содержащая R24Q/P25A/S52T/L54A, представляет собой (SEQ ID NO: 221) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000278] Аминокислотная последовательность VL15 hMAB-3к CD137, содержащая R24Q/P25A/S52G/L54A, представляет собой (SEQ ID NO: 222) (остатки CDRH выделены подчеркиванием; замены выделены двойным подчеркиванием):

[000279] Таким образом, аминокислотные последовательности CDRL3 из VL3-VL15 hMAB-3 к CD137 являются идентичными (QQGDTLPYT; SEQ ID NO: 159). Однако аминокислотные последовательности CDRL1 и/или CDRL2 из VL4 hMAB-3 к CD137 и VL6-VL15 hMAB-3 к CD137 различаются:
[000280] CDRL1 из VL4, VL7, VL8, VL9, VL14 и VL15 hMAB-3 к CD137 (SEQ ID NO: 223): 
CDRL2 из VL6, VL8 и VL9 hMAB-3 к CD137 (SEQ ID NO: 224): 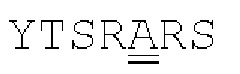
CDRL2 из VL10 и VL12 hMAB-3 к CD137 (SEQ ID NO: 225): 
CDRL2 из VL11 и VL13 hMAB-3 к CD137 (SEQ ID NO: 226): 
CDRL2 из VL14 hMAB-3 к CD137 (SEQ ID NO: 227): 
CDRL2 из VL15 hMAB-3 к CD137 (SEQ ID NO: 228): 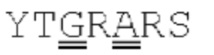
[000281] CDR, домен VL и/или домен VH любого из таких гуманизированных, оптимизированных и/или деиммунизированных доменов VH и VL hMAB-3 к CD137, включая любые включенные в общую(ие) последовательность(и) доменов VH и/или VL hMAB-3 к CD137, представленных выше, можно применять для образования антитела, диатела или связывающей молекулы, способной связывать CD137.
5. МАВ-4 к CD137
[000282] МАВ-4 к CD137 представляет собой новое мышиное моноклональное антитело. Аминокислотная последовательность домена VH МАВ-4 к CD137 (VH МАВ-4 к CD137) представляет собой (SEQ ID NO: 90) (остатки CDRH выделены подчеркиванием):

[000283] Аминокислотные последовательности CDRH из VH MAB-4 к CD137 представляют собой:
CDRH1 (SEQ ID NO: 165): SYWIN
CDRH2 (SEQ ID NO: 166): NIYPSDSYTNYDQKFKD
CDRH3 (SEQ ID NO: 167): SGEYGKIGYYAMDY
[000284] Аминокислотная последовательность домена VL MAB-4 к CD137 (VL MAB-4 к CD137) представляет собой (SEQ ID NO: 91) (остатки CDRL выделены подчеркиванием):

[000285] Аминокислотные последовательности CDRL из VL MAB-4 к CD137 представляют собой:
CDRL1 (SEQ ID NO: 168): RASQDISNYLN
CDRL2 (SEQ ID NO: 169): YTSRLHS
CDRL3 (SEQ ID NO: 170): QQGNTLPYT
6. hMAB-4 к CD137
[000286] Антитело MAB-4 к CD137 было гуманизировано с образованием антитела hMAB-4 к CD137. Аминокислотная последовательность домена VH гуманизированного антитела hMAB-4 к CD137 (VH1 hMAB-4 к CD137) представляет собой (SEQ ID NO: 92) (остатки CDRH выделены подчеркиванием):

[000287] Домен VL гуманизированного антитела hMAB-4 к CD137 был гуманизирован и оптимизирован для получения домена VL (VL hMAB-3 к CD137), имеющего аминокислотную последовательность SEQ ID NO: 93 (остатки CDRL выделены подчеркиванием):


где Х9 представляет собой F или Y.
[000288] Были выделены два варианта доменов VL hMAB-4 к CD137: VL1 hMAB-4 к CD137 и VL2 hMAB-4 к CD137. Аминокислотные последовательности таких вариантов доменов VL hMAB-4 к CD137 представлены ниже.
[000289] Аминокислотная последовательность VL1 hMAB-4 к CD137 представляет собой (SEQ ID NO: 94) (остатки CDRL выделены подчеркиванием; следует отметить, что остаток 87 представляет собой F):

[000290] Аминокислотная последовательность VL2 hMAB-4 к CD137 представляет собой (SEQ ID NO: 95) (остатки CDRL выделены подчеркиванием; следует отметить, что остаток 87 представляет собой Y):

7. MAB-5 к CD137
[000291] MAB-5 к CD137 представляет собой новое мышиное моноклональное антитело. Аминокислотная последовательность домена VH МАВ-5 к CD137 (VH МАВ-5 к CD137) представляет собой (SEQ ID NO: 96) (остатки CDRH выделены подчеркиванием):

[000292] Аминокислотные последовательности CDRH из VH МАВ-5 к CD137 представляют собой:
CDRH1 (SEQ ID NO: 171): SYDIS
CDRH2 (SEQ ID NO: 172): VWTGGGTNYNSAFMS
CDRH3 (SEQ ID NO: 173): VDY
[000293] Аминокислотная последовательность домена VL MAB-5 к CD137 (VL MAB-5 к CD137) представляет собой (SEQ ID NO: 97) (остатки CDRL выделены подчеркиванием):


[000294] Аминокислотные последовательности CDRL из VL MAB-5 к CD137 представляют собой:
CDRL1 (SEQ ID NO: 174): RSSQSLVHSNGNTYLH
CDRL2 (SEQ ID NO: 175): KVSNRFS
CDRL3 (SEQ ID NO: 176): SQSTHVPWT
III. CD137×TA связывающие молекулы согласно настоящему изобретению
[000295] Настоящее изобретение, в частности, относится к трехвалентным CD137×TA связывающим молекулам, которые содержат домен Fc, Fc-несущим CD137×TA диателам DART®, способным одновременно связываться с CD137 и ТА, и другим Fc-несущим CD137×TA связывающим молекулам, способным одновременно связываться с CD137 и ТА. Настоящее изобретение также относится к применению таких молекул в лечении рака и других заболеваний и состояний. Несмотря на то, что неоптимизированные CD137×TA связывающие молекулы являются полностью функциональными, также как и в случае улучшений, достигнутых в экспрессии генов за счет оптимизации кодонов (см., например, Grosjean, Н. et al. (1982) «Preferential Codon Usage In Prokaryotic Genes: The Optimal Codon-Anticodon Interaction Energy And The Selective Codon Usage In Efficiently Expressed Genes» Gene 18(3): 199-209), существует возможность дополнительного улучшения стабильности и/или функции CD137×TA связывающих молекул согласно настоящему изобретению путем модификации или оптимизации их последовательности.
A. CD137×TA Fc-несущие диатела DART®
[000296] Настоящее изобретение, в частности, охватывает широкий спектр Fc-несущих диател DART®, способных одновременно связываться с CD137 и ТА. Примерные CD137×TA Fc-несущие диатела DART® описаны ниже.
[000297] Согласно одному варианту реализации настоящего изобретения такие Fc-несущие антитела будут содержать две полипептидные цепи. Как показано на Фигуре 2, первая из таких полипептидных цепей может содержать, в направлении от N-конца к С-концу, N-конец, вариабельный домен легкой цепи (VL), способный связываться с эпитопом «первого» антигена (VL1) (CD137 или ТА), вариабельный домен тяжелой цепи (VH), способный связываться с эпитопом «второго» антигена (VH2) (ТА, если VL1 был выбран для связывания с эпитопом CD137; CD137, если VL1 был выбран для связывания с эпитопом ТА), цистеинсодержащий линкер, домен СН2-СН3 и С-конец. Вторая из таких полипептидных цепей может содержать, в направлении от N-конца к С-концу, N-конец, вариабельный домен легкой цепи (VL), способный связываться с эпитопом «второго» антигена (VL2) (ТА, если первым антигеном был CD137; CD137, если первым антигеном был ТА), вариабельный домен тяжелой цепи (VH), способный связываться с эпитопом «второго» антигена (VH2) (ТА, если VL2 был выбран для связывания с эпитопом CD137; CD137, если VL2 был выбран для связывания с эпитопом ТА), цистеинсодержащий линкер, домен СН2-СН3 и С-конец. Промежуточный линкерный пептид (линкер 1) отделяет вариабельный домен легкой цепи (VL1 или VL2) от вариабельного домена тяжелой цепи (VH1 или VH2).
[000298] Согласно другому варианту реализации настоящего изобретения Fc-несущие антитела согласно настоящему изобретению содержат три полипептидные цепи и изображены на Фигурах 4А-4В. Первая полипептидная цепь такого диатела содержит четыре домена: (i) VL1-содержащий домен, (ii) VH2-содержащий домен, (iii) домен, который способствует гетеродимеризации («домен, способствующий образованию гетеродимера») и образованию ковалентной связи со второй полипептидной цепью диатела, и (iv) домен, содержащий последовательность СН2-СН3. Вторая полипептидная цепь таких Fc-содержащих диател содержит: (i) VL2-содержащий домен, (ii) VH1-содержащий домен и (iii) домен, который способствует гетеродимеризации («домен, способствующий образованию гетеродимера») и образованию ковалентной связи с первой полипептидной цепью диатела. Третья полипептидная цепь таких Fc-несущих диател содержит последовательность СН2-СН3. Таким образом, первая и вторая полипептидные цепи таких Fc-несущих диател объединяются с образованием сайта связывания VL1/VH1, который способен связываться с первым эпитопом (1), а также сайта связывания VL2/VH2, который способен связываться со вторым эпитопом (2). Предпочтительные Fc-несущие диатела согласно настоящему изобретению представляют собой CD137×TA биспецифические диатела, которые способны связываться с «первым эпитопом», который может представлять собой либо CD137, либо ТА, и «вторым эпитопом», который представляет собой ТА, если первым эпитопом является CD137, и CD137, если первым эпитопом является ТА. Первый и второй полипептиды связаны друг с другом за счет одной или более дисульфидных связей с участием остатков цистеина в их соответствующих линкерах и/или третьих доменах. Примечательно, что первая и третья полипептидные цепи образуют комплекс друг с другом с образованием домена Fc, который стабилизирован за счет дисульфидной связи. Такие диатела обладают повышенной активностью. Предпочтительные Fc-несущие диатела согласно настоящему изобретению могут иметь одну из двух ориентации (таблица 2):

[000299] Как показано на Фигуре 4А, в первом варианте реализации, первая из таких трех полипептидных цепей может содержать, в направлении от N-конца к С-концу, N-конец, вариабельный домен легкой цепи (VL), способный связываться с эпитопом «первого» антигена (VL1) (либо CD137, либо ТА), вариабельный домен тяжелой цепи (VH), способный связываться с эпитопом «второго» антигена (VH2) (ТА в качестве первого антигена, если вторым антигеном являлся CD137; CD137 в качестве первого антигена, если вторым антигеном являлся ТА), цистеинсодержащий домен, домен, способствующий образованию гетеродимера, второй цистеинсодержащий домен, домен СН2-СН3 и С-конец. Промежуточный линкерный пептид (линкер 1) отделяет вариабельный домен легкой цепи (VL1) от вариабельного домена тяжелой цепи (VH2). Предпочтительно вариабельный домен тяжелой цепи (VH1) соединен с доменом, способствующим образованию гетеродимера, промежуточным линкерным пептидом (линкер 2). Согласно предпочтительному варианту реализации CD137×TA биспецифического Fc-несущего диатела С-конец домена, способствующего образованию гетеродимера, соединен с доменами СН2-СН3 области Fc («домен Fc») с помощью промежуточного линкерного пептида (линкер 3) или промежуточного пептидного спейсера (линкер 3). Таким образом, наиболее предпочтительно первая из трех полипептидных цепей будет содержать, в направлении от N-конца к С-концу: VL1 - линкер 1 - VH2 - линкер 2 - домен, способствующий образованию гетеродимера, - линкер 3 - домен Fc.
[000300] Вторая из таких трех полипептидных цепей будет содержать, в направлении от N-конца к С-концу: N-конец, вариабельный домен легкой цепи (VL), способный связываться с эпитопом «второго» антигена (VL2), вариабельный домен тяжелой цепи (VH), способный связываться с эпитопом «первого» антигена (VH1), цистеинсодержащий домен, домен, способствующий образованию гетеродимера, и С-конец. Промежуточный линкерный пептид (линкер 1) отделяет вариабельный домен легкой цепи (VL2) от вариабельного домена тяжелой цепи (VH1). Предпочтительно вариабельный домен тяжелой цепи (VH1) соединен с доменом, способствующим образованию гетеродимера, промежуточным линкерным пептидом (линкер 2). Таким образом, наиболее предпочтительно вторая из трех полипептидных цепей будет содержать, в направлении от N-конца к С-концу: VL2 - линкер 1 - VH1 - линкер 2 - домен, способствующий образованию гетеродимера.
[000301] Третья из таких трех полипептидных цепей будет содержать, в направлении от N-конца к С-концу, цистеинсодержащий пептид (такой как линкер 3) и домены СН2-СН3 области Fc («домен Fc»). Поскольку третья полипептидная цепь не содержит домен VL или домен VH, третья полипептидная цепь может быть идентична в двух или более различных CD137×TA биспецифических Fc-несущих диателах согласно настоящему изобретению.
[000302] Согласно другому варианту, как показано на Фигуре 4В, во втором варианте реализации, первая из таких трех полипептидных цепей будет содержать, в направлении от N-конца к С-концу, N-конец, цистеинсодержащий пептид (такой как линкер 3), домены СН2-СН3 области Fc («домен Fc»), промежуточный спейсерный пептид (линкер 4), вариабельный домен легкой цепи (VL), способный связываться с эпитопом «первого» антигена (VL1) (либо CD137, либо ТА), вариабельный домен тяжелой цепи (VH), способный связываться с эпитопом «второго» антигена (VH2) (ТА, если первым антигеном являлся CD137; CD137, если первым антигеном являлся ТА), цистеинсодержащий линкер, домен, способствующий образованию гетеродимера, и С-конец. Промежуточный линкерный пептид (линкер 1) отделяет вариабельный домен легкой цепи (VL1) от вариабельного домена тяжелой цепи (VH2). Предпочтительно вариабельный домен тяжелой цепи (VH1) соединен с доменом, способствующим образованию гетеродимера, с помощью промежуточного линкерного пептида (линкер 2). Наиболее предпочтительно, в такой альтернативной ориентации, первая из трех полипептидных цепей, соответственно, будет содержать, в направлении от N-конца к С-концу: линкер 3 - домен Fc - линкер 4 - VL1 - линкер 1 - VH2 - линкер 2 - домен, способствующий образованию гетеродимера. Вторая и третья полипептидные цепи такого альтернативного диатела могут быть идентичны второй и третьей полипептидным цепям из первого варианта реализации.
[000303] В каждом из приведенных выше вариантов реализации вариабельный домен легкой цепи первой полипептидной цепи (VL1) координировано отобран так, чтобы он мог взаимодействовать с вариабельным доменом тяжелой цепи второй полипептидной цепи (VH1) с образованием функционального эпитопсвязывающего сайта, который способен иммуноспецифично связывать эпитоп первого антигена (т.е. либо ТА, либо CD137). Аналогичным образом, вариабельный домен легкой цепи второй полипептидной цепи (VL2) координировано отобран так, чтобы он мог взаимодействовать с вариабельным доменом тяжелой цепи первой полипептидной цепи (VH2) с образованием функционального эпитопсвязывающего сайта, который способен иммуноспецифично связывать эпитоп второго антигена (т.е. либо ТА, либо CD137). Таким образом, отбор вариабельных доменов легкой цепи и вариабельных доменов тяжелой цепи координируется таким образом, что две полипептидные цепи совместно содержат эпитопсвязывающие сайты, способные связываться с CD137 и ТА.
[000304] Предпочтительные Fc-несущие диатела согласно настоящему изобретению представляют собой ковалентно связанные четырехвалентные диатела, имеющие четыре эпитопсвязывающих сайта, которые содержат четыре полипептидные цепи, и изображены на Фигурах 3А-3С. Первая и третья полипептидные цепи такого диатела содержат: (i) VL1-содержащий домен, (ii) VH2-содержащий домен, (iii) домен, способствующий образованию гетеродимера, и (iv) домен, содержащий последовательность СН2-СН3. Вторая и четвертая полипептидные цепи содержат: (i) VL2-содержащий домен, (ii) VH1-содержащий домен, и (iii) домен, способствующий образованию гетеродимера, причем домены, способствующие образованию гетеродимера, способствуют димеризации и ковалентному связыванию первой/третьей полипептидных цепей со второй/четвертой полипептидными цепями. Предпочтительно домены VH соединены с доменами, способствующими образованию гетеродимера, с помощью промежуточных линкерных пептидов (линкер 2). Согласно предпочтительному варианту реализации CD137×TA биспецифического Fc-несущего диатела С-конец домена, способствующего образованию гетеродимера, первой полипептидной цепи соединен с доменами СН2-СН3 с помощью промежуточного линкерного пептида (линкер 3) или промежуточного спейсерного пептида (линкер 3). Домены VL и/или VH третьей и четвертой полипептидных цепей и домены VL и/или VH первой и второй полипептидных цепей могут быть одинаковыми или различными, чтобы обеспечить четырехвалентное связывание, которое является либо моноспецифическим, либо биспецифическим, либо тетраспецифическим. Условные сокращения «VL3» и «VH3» обозначают, соответственно, вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связывают «третий» эпитоп такого диатела. Аналогичным образом, условные сокращения «VL4» и «VH4» обозначают, соответственно, вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связывают «четвертый» эпитоп такого диатела. Общая структура полипептидных цепей типичных четырехцепочечных биспецифических диател, содержащих область Fc, согласно настоящему изобретению представлена в таблице 3:

[000305] Согласно предпочтительным вариантам реализации CD137×TA связывающие молекулы согласно настоящему изобретению являются биспецифическими, четырехвалентными (т.е. обладают четырьмя эпитопсвязывающими сайтами) Fc-несущими диателами, которые состоят в общей сложности из четырех полипептидных цепей (Фигура 3А-3С). CD137×TA связывающие молекулы согласно настоящему изобретению представляют собой биспецифические, четырехвалентные Fc-несущие диатела, которые содержат два эпитопсвязывающих сайта, иммуноспецифичных для CD137 (которые могут быть способны связываться с одним и тем же эпитопом CD137 или с различными эпитопами CD137), и два эпитопсвязывающих сайта, иммуноспецифичных для опухолевого антигена (которые могут быть способны связываться с одним и тем же эпитопом ТА или с различными эпитопами ТА или с различными эпитопами разных ТА).
[000306] Дополнительные предпочтительные Fc-несущие диатела согласно настоящему изобретению содержат пять полипептидных цепей и изображены на Фигуре 5A-5D. Первая полипептидная цепь такого диатела содержит: (i) VH1-содержащий домен, (ii) CH1-содержащий домен, и (iii) домен, содержащий последовательность СН2-СН3. Первая полипептидная цепь может представлять собой тяжелую цепь антитела, которое содержит VH1 и константную область тяжелой цепи. Вторая и пятая полипептидные цепи такого диатела содержат: (i) VL1-содержащий домен, и (ii) CL-содержащий домен. Вторая и/или пятая полипептидные цепи такого диатела могут представлять собой легкие цепи антитела, которое содержит VL1, комплементарный VH1 первой/третьей полипептидной цепи. Первая, вторая и/или пятая полипептидные цепи могут быть выделены из природного антитела. Согласно другому варианту они могут быть сконструированы рекомбинантно. Согласно предпочтительному варианту реализации настоящего изобретения вторая и пятая полипептидные цепи имеют одинаковую аминокислотную последовательность. Третья полипептидная цепь такого диатела содержит: (i) VH1-содержащий домен, (ii) CH1-содержащий домен, (iii) домен, содержащий последовательность СН2-СН3, (iv) VL2-содержащий домен, (v) VH3-содержащий домен, и (vi) домен, способствующий образованию гетеродимера, причем домены, способствующие образованию гетеродимера, способствуют димеризации третьей цепи с четвертой цепью. Четвертый полипептид таких диател содержит: (i) VL3-содержащий домен, (ii) VH2-содержащий домен, и (iii) домен, который способствует гетеродимеризации и образованию ковалентной связи с третьей полипептидной цепью диатела. Предпочтительно С-конец VH3- и VH2-содержащих доменов третьей и четвертой полипептидных цепей соединен с доменом, способствующим образованию гетеродимера, с помощью промежуточного линкерного пептида (линкер 2), и С-конец доменов СН2-СН3 третьей полипептидной цепи соединен с VL2-содержащим доменом с помощью промежуточного линкерного пептида (линкер 4).
[000307] Соответственно, первая и вторая, а также третья и пятая полипептидные цепи таких диател объединяются с образованием двух сайтов связывания VL1/VH1, способных связывать первый эпитоп. Третья и четвертая полипептидные цепи таких диател объединяются с образованием сайта связывания VL2/VH2, который способен связываться со вторым эпитопом, а также сайта связывания VL3/VH3, который способен связываться с третьим эпитопом. Первый и третий полипептиды связаны друг с другом за счет дисульфидной связи с участием остатков цистеина в их соответствующих константных областях. Примечательно, что первая и третья полипептидные цепи образуют комплекс друг с другом с образованием области Fc. Такие биспецифические диатела обладают повышенной активностью. На Фигурах 5A-5D проиллюстрирована структура таких диател. Следует понимать, что домены VL1/VH1, VL2/VH2 и VL3/VH3 могут быть одинаковыми или различными, чтобы обеспечить связывание, которое является моноспецифическим, биспецифическим или триспецифическим. Однако, как предусмотрено в настоящем документе, эти домены предпочтительно отобраны так, чтобы связывать CD137 и ТА.
[000308] Домены VL и VH полипептидных цепей отбирают так, чтобы они образовывали сайты связывания VL/VH, специфичные для желательного эпитопа. Сайты связывания VL/VH, образованные за счет объединения полипептидных цепей, могут быть одинаковыми или различными, чтобы обеспечить четырехвалентное связывание, которое является моноспецифическим, биспецифическим, триспецифическим или тетраспецифическим. В частности, домены VL и VH могут быть отобраны так, что биспецифическое диатело может содержать два сайта связывания для первого эпитопа и два сайта связывания для второго эпитопа или три сайта связывания для первого эпитопа и один сайт связывания для второго эпитопа, или два сайта связывания для первого эпитопа, один сайт связывания для второго эпитопа и один сайт связывания для третьего эпитопа (как изображено на Фигурах 5A-5D). Общая структура полипептидных цепей типичных пятицепочечных диател, содержащих область Fc, согласно настоящему изобретению представлена в таблице 4:

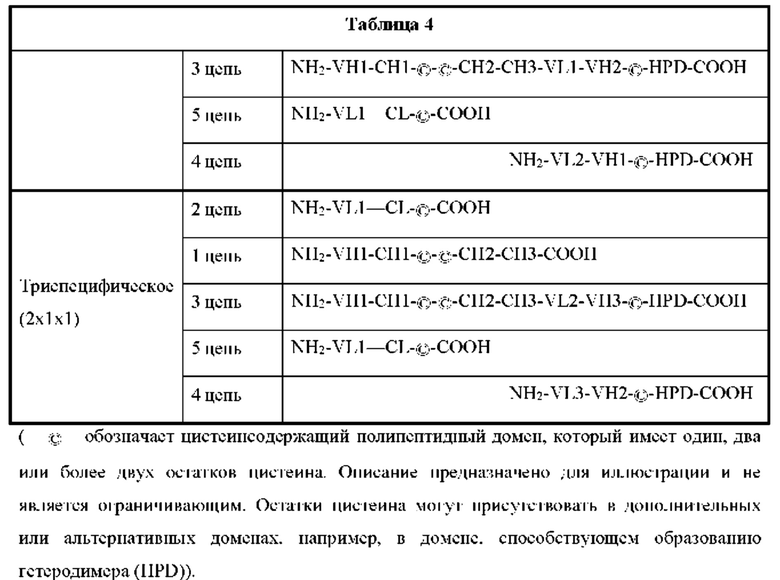
[000309] Согласно предпочтительному варианту реализации настоящего изобретения CD137×TA связывающие молекулы согласно настоящему изобретению являются биспецифическими, четырехвалентными (т.е. обладают четырьмя эпитопсвязывающими сайтами) Fc-несущими диателами, которые состоят в общей сложности из пяти полипептидных цепей, имеющих два эпитопсвязывающих сайта, иммуноспецифичных для CD137 (которые могут быть способны связываться с одним и тем же эпитопом CD137 или с различными эпитопами CD137), и два эпитопсвязывающих сайта, иммуноспецифичных для ТА (которые могут быть способны связываться с одним и тем же эпитопом ТА или с различными эпитопами ТА или с различными эпитопами разных ТА). Согласно другому варианту реализации настоящего изобретения CD137×TA связывающие молекулы согласно настоящему изобретению представляют собой биспецифические, четырехвалентные Fc-несущие диатела, которые содержат три эпитопсвязывающих сайта, иммуноспецифичных для CD137 (которые могут быть способны связываться с одним и тем же эпитопом CD137 или с двумя или тремя различными эпитопами CD137), и один эпитопсвязывающий сайт, специфичный для ТА.
В. Трехвалентные CD137×TA связывающие молекулы
[000310] Согласно одному варианту реализации настоящего изобретения CD137×TA связывающие молекулы согласно настоящему изобретению являются трехвалентными и будут содержать первый эпитопсвязывающий сайт (например, VL1 и VH1), второй эпитопсвязывающий сайт (например, VL2 и VH2) и третий эпитопсвязывающий сайт (например, VL3 и VH3) и, соответственно, будут способны связываться с эпитопом ТА, эпитопом CD137 и третьим эпитопом, причем третий эпитоп может представлять собой:
(a) тот же или другой эпитоп ТА;
(b) тот же или другой эпитоп CD137; или
(c) эпитоп другого ТА.
[000311] Предпочтительно такие «трехвалентные CD137×TA связывающие молекулы» согласно настоящему изобретению будут содержать два эпитопсвязывающих сайта для эпитопа CD137 (эти эпитопы могут быть одинаковыми или различными) и один эпитопсвязывающий сайт для эпитопа ТА.
[000312] В целом такие трехвалентные CD137×TA связывающие молекулы согласно настоящему изобретению состоят из трех, четырех, пяти или более пяти полипептидных цепей, которые благодаря одной или более дисульфидным связям между парами таких полипептидов образуют ковалентно связанный молекулярный комплекс, который содержит «связывающий домен, характерный для диатела» и «связывающий домен, не характерный для диатела».
[000313] «Связывающий домен, характерный для диатела» представляет собой эпитопсвязывающий домен диатела и, в частности, диатела DART®. Термины «диатело» и «диатело DART®» обсуждались выше. Связывающий домен «не характерным для диатела» предназначен для обозначения связывающего домена, который не имеет структуры связывающего домена, характерного для диатела. Предпочтительно связывающий домен, не характерный для диатела, представляет собой связывающий домен Fab-типа или связывающий домен scFv-типа. В настоящем документе термин «связывающий домен Fab-типа» относится к эпитопсвязывающему домену, который образуется в результате взаимодействия домена VL легкой цепи иммуноглобулина и комплементарного домена VH тяжелой цепи иммуноглобулина. Связывающие домены Fab-типа отличаются от связывающего домена, характерного для диатела, тем, что две полипептидные цепи, которые образуют связывающий домен Fab-типа, содержат только один эпитопсвязывающий домен, тогда как две полипептидные цепи, которые образуют связывающий домен, характерный для диатела, содержат по меньшей мере два эпитопсвязывающих домена. Связывающие домены scFv-типа отличаются от связывающего домена, характерного для диатела, тем, что домены VL и VH одной и той же полипептидной цепи взаимодействуют с образованием эпитопсвязывающего домена. Таким образом, в настоящем документе связывающие домены Fab-типа и связывающие домены scFv-типа отличаются от связывающего домена, характерного для диатела.
[000314] Соответственно, трехвалентные CD137×TA связывающие молекулы согласно настоящему изобретению предпочтительно содержат:
(i) «первый» эпитопсвязывающий домен, который способен иммуноспецифично связываться с «первым» эпитопом;
(ii) «второй» эпитопсвязывающий домен, который способен иммуноспецифично связываться со «вторым» эпитопом;
(iii) «третий» эпитопсвязывающий домен, который способен иммуноспецифично связываться с «третьим» эпитопом; и
(iv) домен Fc, который образован за счет объединения двух доменов СН2-СН3 друг с другом;
причем:
(A) «первый» эпитопсвязывающий домен и «второй» эпитопсвязывающий домен оба представляют собой «связывающие домены, характерные для диатела»;
(B) «третий» эпитопсвязывающий домен представляет собой связывающий домен, не характерный для диатела; и
(C) один из таких «первого», «второго» или «третьего» эпитопсвязывающих доменов связывает эпитоп ТА, и другой из таких «первого», «второго» или «третьего» эпитопсвязывающих доменов связывает эпитоп CD137;
[000315] Эпитоп, который связан оставшимся эпитопсвязывающим доменом, может представлять собой любой желательный эпитоп, предпочтительно эпитоп CD137. Такой эпитоп может быть идентичен или может отличаться от эпитопа CD137, связываемого другими эпитопсвязывающими доменами молекулы.
[000316] На Фигуре 6А-6Н схематически представлены домены предпочтительных трехвалентных CD137×TA связывающих молекул. На Фигурах 6A-6D схематически проиллюстрированы домены предпочтительных трехвалентных CD137×TA связывающих молекул, которые состоят из ковалентного комплекса четырех полипептидных цепей и имеют один сайт связывания, не характерный для диатела (VL3/VH3 и, соответственно, являются одновалентными для такого эпитопа), и два сайта связывания, характерных для диатела (VL1/VH1 и VL2/VH2 и, соответственно, являются одновалентными для каждого из таких эпитопов). На Фигурах 6Е-6Н схематически показаны домены предпочтительных трехвалентных CD137×TA связывающих молекул, которые состоят из ковалентного комплекса трех полипептидных цепей и имеют один сайт связывания, не характерный для диатела (VL3/VH3 и, соответственно, являются одновалентными для такого эпитопа), и два сайта связывания, характерных для диатела (VL1/VH1 и VL2/VH2 и, соответственно, являются одновалентными для каждого из таких эпитопов). Сайт связывания, не характерный для диатела, показанный на Фигурах 6А-6Н, представляет собой связывающий домен Fab-типа на Фигурах 6A-6D и представляет собой связывающий домен scFv-типа на Фигурах 6Е-6Н. Как указано ниже, сайты связывания VL/VH, образованные за счет объединения полипептидных цепей, могут быть одинаковыми или различными, чтобы обеспечить трехвалентное связывание, которое является моноспецифическим, биспецифическим или триспецифическим.
IV Примерные CD137×TA связывающие молекулы
[000317] Согласно настоящему изобретению предложены CD137×TA связывающие молекулы, которые представляют собой биспецифические четырехвалентные Fc-несущие диатела, способные одновременно и специфично связываться с CD137 и с ТА. Как указано выше, CD137×TA связывающие молекулы согласно настоящему изобретению могут содержать четыре или пять полипептидных цепей. Полипептидные цепи семи примерных CD137×TA связывающих молекул, способных связываться с CD137 и ТА, HER2/neu, представлены ниже (обозначены «DART-A», «DART-B», «DART-C», «DART-D», «DART-E», «DART-F», «DART-G», «DART-G1», «DART-G2», «DART-G3» и «DART-G4», соответственно). Согласно настоящему изобретению также предложены CD137×TA связывающие молекулы, которые представляют собой биспецифические трехвалентные связывающие молекулы, способные одновременно и специфично связываться с CD137 и с ТА. Как указано выше, трехвалентные CD137×TA связывающие молекулы согласно настоящему изобретению могут содержать четыре полипептидные цепи. Полипептидные цепи примерной CD137×TA триспецифической связывающей молекулы, способной связываться с CD137 и ТА, HER2/neu, и ее оптимизированные варианты представлены ниже (обозначены «TRIDENT-A», «TRIDENT-A1», «TRIDENT-A2», «TRIDENT-А3», «TRIDENT-A5», «TRIDENT-B», «TRIDENT-B2» и «TRIDENT-В5», соответственно).
A. DART-A
[000318] DART-A состоит из четырех полипептидных цепей, из которых первая и третья полипептидные цепи являются идентичными, и вторая и четвертая полипептидные цепи являются идентичными (см. Фигуру 3В).
[000319] Первая и третья полипептидные цепи DART-А содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с HER2/neu (VLHER2/neu) (VL2 hMAB-1 к HER2 (SEQ ID NO: 68)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связываться с CD137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 38), линкер (LEPKSADKTHTCPPCP (SEQ ID NO: 30)), домен CH2-CH3 примерного человеческого IgG1, у которого по существу отсутствует эффекторная функция (SEQ ID NO: 40), и С-конец.
[000320] Аминокислотная последовательность первой и третьей полипептидных цепей DART-А представляет собой (SEQ ID NO: 98):

[000321] Соответственно, первая и третья полипептидные цепи DART-А состоят из: SEQ ID NO: 68 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 18 - SEQ ID NO: 38 - SEQ ID NO: 30 - SEQ ID NO: 40.
[000322] Вторая и четвертая полипептидные цепи DART-А содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с CD137 (VLCD137 (VL1 hMAB-3 к CD137 (SEQ ID NO: 87)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu (VH1 hMAB-1 к HER2, SEQ ID NO: 64)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO: 39)), и С-конец.
[000323] Соответственно, вторая и четвертая полипептидные цепи DART-А состоят из: SEQ ID NO: 87 - SEQ ID NO: 16 - SEQ ID NO: 64 - SEQ ID NO: 18 - SEQ ID NO: 39.
[000324] Аминокислотная последовательность второй и четвертой полипептидных цепей DART-А представляет собой (SEQ ID NO: 99):

В. DART-В
[000325] DART-B состоит из четырех полипептидных цепей, из которых первая и третья полипептидные цепи являются идентичными, и вторая и четвертая полипептидные цепи являются идентичными (см. Фигуру 3В).
[000326] Первая и третья полипептидные цепи DART-B содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с HER2/neu (VLHER2/neu) (VL3 hMAB-1 к HER2 (SEQ ID NO: 69)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связываться с CD137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) (EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 38)), линкер (LEPKSADKTHTCPPCP (SEQ ID NO: 30)), домен CH2-CH3 примерного человеческого IgG1, у которого по существу отсутствует эффекторная функция (SEQ ID NO: 40), и С-конец.
[000327] Соответственно, первая и третья полипептидные цепи DART-B состоят из: SEQ ID NO: 69 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 18 - SEQ ID NO: 38 - SEQ ID NO: 30 - SEQ ID NO: 40.
[000328] Аминокислотная последовательность первой и третьей полипептидных цепей DART-B представляет собой (SEQ ID NO: 100):


[000329] Вторая и четвертая полипептидные цепи DART-B содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с CD137 (VLCD137 (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu (VH1 hMAB-1 к HER2, SEQ ID NO: 64)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO: 39)), и С-конец.
[000330] Соответственно, вторая и четвертая полипептидные цепи DART-B состоят из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 64 - SEQ ID NO: 18 - SEQ ID NO: 39.
[000331] Аминокислотная последовательность второй и четвертой полипептидных цепей DART-B представляет собой (SEQ ID NO: 101):

С. DART-С
[000332] DART-C состоит из четырех полипептидных цепей, из которых первая и третья полипептидные цепи являются идентичными, и вторая и четвертая полипептидные цепи являются идентичными (см. Фигура 3В).
[000333] Первая и третья полипептидные цепи DART-С содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с CD137 (VLCD137 (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) (EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 38)), линкер (LEPKSADKTHTCPPCP (SEQ ID NO: 30)), домен СН2-СН3 примерного человеческого IgG1, у которого по существу отсутствует эффекторная функция (SEQ ID NO: 40), и С-конец.
[000334] Соответственно, первая и третья полипептидные цепи DART-С состоят из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 64 - SEQ ID NO: 18 - SEQ ID NO: 38 - SEQ ID NO: 30 - SEQ ID NO: 40.
[000335] Аминокислотная последовательность первой и третьей полипептидных цепей DART-C представляет собой (SEQ ID NO: 102):

[000336] Вторая и четвертая полипептидные цепи DART-C содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с HER2/neu (VLHER2/neu (VL3 hMAB-1 к HER2, SEQ ID NO: 69)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137 (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO: 39)), и С-конец.
[000337] Соответственно, вторая и четвертая полипептидные цепи DART-С состоят из: SEQ ID NO: 69 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 18 - SEQ ID NO: 39.
[000338] Аминокислотная последовательность второй и четвертой полипептидных цепей DART-C представляет собой (SEQ ID NO: 103):


D. DART-D
[000339] DART-D состоит из пяти полипептидных цепей, из которых вторая и пятая являются идентичными (см. Фигуру 5А, причем VL2/VH2 идентичны VL3/VH3, и Фигуру 5В).
[000340] Первая полипептидная цепь DART-D содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), домен CH1 человеческого IgG1 (SEQ ID NO: 3), шарнирную область человеческого IgG1 (SEQ ID NO: 7) и «несущий впадину» домен СН2 и домен СНЗ (SEQ ID NO: 47).
[000341] Соответственно, первая полипептидная цепь DART-D состоит из: SEQ ID NO: 64 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 47.
[000342] Аминокислотная последовательность первой полипептидной цепи DART-D представляет собой (SEQ ID NO: 104):

[000343] Вторая и пятая полипептидные цепи DART-D содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с HER2/neu (VLHER2/neu) (VL3 hMAB-1 к HER2 (SEQ ID NO: 69)), каппа-домен CL человеческого IgG (SEQ ID NO: 1) и С-конец.
[000344] Соответственно, вторая и пятая полипептидные цепи DART-D состоят из: SEQ ID NO: 69 - SEQ ID NO: 1.
[000345] Аминокислотная последовательности второй и пятой полипептидных цепей DART-D представляет собой (SEQ ID NO: 105):


[000346] Третья полипептидная цепь DART-D содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), домен CH1 человеческого IgG1 (SEQ ID NO: 3), шарнирную область человеческого IgG1 (SEQ ID NO: 7), «несущий выступ» домен CH2 и домен СН3 (SEQ ID NO: 44), промежуточный линкерный пептид (GGGSGGGSGGG (SEQ ID NO: 24)), домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL МАВ-3 к CD137 (SEQ ID NO: 75)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD 137 (VHCD137) (VH МАВ-3 к CD137 (SEQ ID NO: 74)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) (EVAALEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 36)), и С-конец.
[000347] Соответственно, третья полипептидная цепь DART-D состоит из: SEQ ID NO: 64 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 44- SEQ ID NO: 24 - SEQ ID NO: 75 - SEQ ID NO: 16 - SEQ ID NO: 74- SEQ ID NO: 18 - SEQ ID NO: 36.
[000348] Аминокислотная последовательность третьей полипептидной цепи DART-D представляет собой (SEQ ID NO: 106):


[000349] Четвертая полипептидная цепь DART-D содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL МАВ-3 к CD137 (SEQ ID NO: 75)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH МАВ-3 к CD137 (SEQ ID NO: 74)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAALKE-KVAALKE-KVAALKE-KVAALKE) (SEQ ID NO: 37), и С-конец.
[000350] Соответственно, четвертая полипептидная цепь DART-D состоит из: SEQ ID NO: 75 - SEQ ID NO: 16 - SEQ ID NO: 74 - SEQ ID NO: 18 - SEQ ID NO: 37.
[000351] Аминокислотная последовательность четвертой полипептидной цепи DART-D представляет собой (SEQ ID NO: 107):

Е. DART-E
[000352] DART-E состоит из пяти полипептидных цепей, из которых вторая и пятая являются идентичными (см. Фигуру 5А, причем VL2/VH2 идентичны VL3/VH3, и Фигуру 5С).
[000353] Первая полипептидная цепь DART-E содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с CD 137 (VHCD137) (VH МАВ-3 к CD137 (SEQ ID NO: 74)), домен CH1 IgG1 (SEQ ID NO: 3), шарнирный домен IgG1 (SEQ ID NO: 7), «несущий впадину» домен CH2 и домен СН3 (SEQ ID NO: 47) и С-конец.
[000354] Соответственно, первая полипептидная цепь DART-E состоит из: SEQ ID NO: 74 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 47.
[000355] Аминокислотная последовательность первой полипептидной цепи DART-E представляет собой (SEQ ID NO: 108):


[000356] Вторая и пятая полипептидные цепи DART-E содержат, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с CD137 (VLCD137u) (VL МАВ-3 к CD137 (SEQ ID NO: 75)), каппа-домен CL человеческого IgG (SEQ ID NO: 1) и С-конец.
[000357] Соответственно, вторая и пятая полипептидные цепи DART-E состоят из: SEQ ID NO: 75 - SEQ ID NO: 1
[000358] Аминокислотная последовательность второй и пятой полипептидных цепей DART-E представляет собой (SEQ ID NO: 109):

[000359] Третья полипептидная цепь DART-E содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с CD137 (VHCD137) (VH МАВ-3 к CD137 (SEQ ID NO: 74)), домен CH1 человеческого IgG1 (SEQ ID NO: 3), шарнирную область человеческого IgG1 (SEQ ID NO: 7), «несущий выступ» домен CH2 и домен СН3 (SEQ ID NO: 44), промежуточный линкерный пептид (GGGSGGGSGGG (SEQ ID NO: 24)), домен VL моноклонального антитела, способного связывать HER2/neu (VLHER2/neu) (VL3 hMAB-1 к HER2 (SEQ ID NO: 69)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) (EVAALEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 36)), и С-конец.
[000360] Соответственно, третья полипептидная цепь DART-E состоит из: SEQ ID NO: 74 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 44 - SEQ ID NO: 24 - SEQ ID NO: 69 - SEQ ID NO: 16 - SEQ ID NO: 64 - SEQ ID NO: 18 - SEQ ID NO: 36
[000361] Аминокислотная последовательность третьих полипептидных цепей DART-Е представляет собой (SEQ ID NO: 110):

[000362] Четвертая полипептидная цепь DART-E содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать HER2/neu (VLHER2/neu) (VL3 hMAB-1 к HER2 (SEQ ID NO: 69)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAALKE-KVAALKE-KVAALKE-KVAALKE) (SEQ ID NO: 37), и С-конец.
[000363] Соответственно, четвертая полипептидная цепь DART-E состоит из: SEQ ID NO: 69 - SEQ ID NO: 16 - SEQ ID NO: 64 - SEQ ID NO: 18- SEQ ID NO: 37.
[000364] Аминокислотная последовательность четвертой полипептидной цепи DART-E представляет собой (SEQ ID NO: 111):


F. DART-F
[000365] DART-F состоит из пяти полипептидных цепей, из которых вторая и пятая являются идентичными (см. Фигуру 5А, причем VL2/VH2 идентичны VL3/VH3, и Фигуру 5В).
[000366] Первая полипептидная цепь DART-F идентична первой полипептидной цепи DART-D (SEQ ID NO: 104).
[000367] Вторая и пятая полипептидные цепи DART-F идентичны второй и пятой полипептидным цепям DART-D (SEQ ID NO: 105).
[000368] Третья полипептидная цепь DART-F содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), домен CH1 IgG1 (SEQ ID NO: 3), шарнирный домен IgG1 (SEQ ID NO: 7), «несущий выступ» домен CH2 и домен СН3 (SEQ ID NO: 44), промежуточный линкерный пептид (GGGSGGGSGGG (SEQ ID NO: 24)), домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL МАВ-4 к CD137 (SEQ ID NO: 91)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH МАВ-4 к CD137 (SEQ ID NO: 90)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) (EVAALEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 36)), и С-конец.
[000369] Соответственно, третья полипептидная цепь DART-F состоит из: SEQ ID NO: 64 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 44 - SEQ ID NO: 24 - SEQ ID NO: 91 - SEQ ID NO: 16 - SEQ ID NO: 90 - SEQ ID NO: 18 - SEQ ID NO: 36
[000370] Аминокислотная последовательность третьей полипептидной цепи DART-F представляет собой (SEQ ID NO: 112):


[000371] Четвертая полипептидная цепь DART-F содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL МАВ-4 к CD137 (SEQ ID NO: 91)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH МАВ-4 к CD137 (SEQ ID NO: 90)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAALKE-KVAALKE-KVAALKE-KVAALKE) (SEQ ID NO: 37), и С-конец.
[000372] Соответственно, четвертая полипептидная цепь DART-F состоит из: SEQ ID NO: 91 - SEQ ID NO: 16 - SEQ ID NO: 90 - SEQ ID NO: 18 - SEQ ID NO: 37.
[000373] Аминокислотная последовательность четвертой полипептидной цепи DART-F представляет собой (SEQ ID NO: 113):

G. DART-G
[000374] DART-G состоит из пяти полипептидных цепей, из которых вторая и пятая являются идентичными (см. Фигуру 5А, причем VL2/VH2 идентичны VL3/VH3, и Фигуру 5В).
[000375] Первая полипептидная цепь DART-G идентична первой полипептидной цепи DART-D (SEQ ID NO: 104).
[000376] Вторая и пятая полипептидные цепи DART-G идентичны второй и пятой полипептидным цепям DART-D (SEQ ID NO: 105).
[000377] Третья полипептидная цепь DART-G содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), домен CH1 IgG1 (SEQ ID NO: 3), шарнирный домен IgG1 (SEQ ID NO: 7), «несущий выступ» домен CH2 и домен СН3 (SEQ ID NO: 44), промежуточный линкерный пептид (GGGSGGGSGGG (SEQ ID NO: 24)), домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептидный (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (Е-спираль) (EVAALEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 36)), и С-конец.
[000378] Соответственно, третья полипептидная цепь DART-G состоит из: SEQ ID NO: 64 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 44 - SEQ ID NO: 24 - SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 18 - SEQ ID NO: 36.
[000379] Аминокислотная последовательность третьей полипептидной цепи DART-G представляет собой (SEQ ID NO: 114):

[000380] Можно применять альтернативные варианты третьей полипептидной цепи DART-G, в которых аминокислотные остатки SEQ ID NO: 76 (домен VH моноклонального антитела, способного связывать CD137 (VHCD137)) заменены аминокислотными остатками SEQ ID NO: 83 (VH1A hMAB-3 к CD137), SEQ ID NO: 84 (VH1B hMAB-3 к CD137), SEQ ID NO: 85 (VH1C hMAB-3 к CD137) или SEQ ID NO: 86 (VH1D hMAB-3 к CD137)). Оптимизированные молекулы, содержащие такие полипептидные цепи, описаны ниже.
Четвертая полипептидная цепь DART-G содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; GGCGGG (SEQ ID NO: 18)), домен, способствующий образованию гетеродимера (K-спираль) (KVAALKE-KVAALKE -KVAALKE-KVAALKE) (SEQ ID NO: 37)), и С-конец.
[000381] Соответственно, четвертая полипептидная цепь DART-G состоит из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 18 - SEQ ID NO: 37.
[000382] Аминокислотная последовательность четвертой полипептидной цепи DART-G представляет собой (SEQ ID NO: 119):

[000383] Можно применять альтернативные варианты четвертой полипептидной цепи DART-G, в которых аминокислотные остатки SEQ ID NO: 76 (домен VH моноклонального антитела, способного связывать CD137 (VHCD137)) заменены аминокислотными остатками SEQ ID NO: 83 (VH1A hMAB-3 к CD137), SEQ ID NO: 84 (VH1B hMAB-3 к CD137), SEQ ID NO: 85 (VH1C hMAB-3 к CD137) или SEQ ID NO: 86 (VH1D hMAB-3 к CD137)). Оптимизированные молекулы, содержащие такие полипептидные цепи, описаны ниже.
Н. Оптимизированное DART-G
[000384] Оптимизированные варианты DART-G, обозначенные «DART-G1», «DART-G2», «DART-G3» и «DART-G4», состоят из пяти полипептидных цепей и содержат HER2/neu-связывающие домены антитела hMAB-1 к HER2 (1.3) и CD137-связывающие домены любого антитела hMAB-3 к CD137 (1A.3)-(1D.3).
[000385] Первая полипептидная цепь таких оптимизированных вариантов DART-G имеет ту же аминокислотную последовательность, что и первая полипептидная цепь DART-G (SEQ ID NO: 104)
[000386] Вторая и пятая полипептидные цепи таких оптимизированных вариантов DART-G имеют ту же аминокислотную последовательность, что и вторая и пятая полипептидные цепи DART-G (SEQ ID NO: 105).
[000387] Третья и четвертая полипептидные цепи таких оптимизированных вариантов DART-G имеют аминокислотные последовательности SEQ ID NO: 115 и SEQ ID NO: 120 (DART-G1); SEQ ID NO: 116 и SEQ ID NO: 121 (DART-G2); SEQ ID NO: 117 и SEQ ID NO: 122 (DART-G3); или SEQ ID NO: 118 и SEQ ID NO: 123 (DART-G4), как указано ниже.
[000388] Аминокислотная последовательность третьей полипептидной цепи оптимизированного DART-G1, содержащая SEQ ID NO: 83 (VH1A hMAB-3 к CD137), представляет собой SEQ ID NO: 115:

[000389] Аминокислотная последовательность третьей полипептидной цепи оптимизированного DART-G2, содержащая SEQ ID NO: 84 (VH1B hMAB-3 к CD137), представляет собой SEQ ID NO: 116:

[000390] Аминокислотная последовательность третьей полипептидной цепи оптимизированного DART-G3, содержащая SEQ ID NO: 85 (VH1C hMAB-3 к CD137), представляет собой SEQ ID NO: 117:

[000391] Аминокислотная последовательность третьей полипептидной цепи оптимизированного DART-G4, содержащая SEQ ID NO: 86 (VH1D hMAB-3 к CD137), представляет собой SEQ ID NO: 118:

[000392] Аминокислотная последовательность четвертой полипептидной цепи оптимизированного DART-G1, содержащая SEQ ID NO: 83 (VH1A hMAB-3 к CD137), представляет собой SEQ ID NO: 120:

[000393] Аминокислотная последовательность четвертой полипептидной цепи оптимизированного DART-G2, содержащая SEQ ID NO: 84 (VH1B hMAB-3 к CD137), представляет собой SEQ ID NO: 121:

[000394] Аминокислотная последовательность четвертой полипептидной цепи оптимизированного DART-G3, содержащая SEQ ID NO: 85 (VH1C hMAB-3 к CD137), представляет собой SEQ ID NO: 122:

[000395] Аминокислотная последовательность четвертой полипептидной цепи оптимизированного DART-G4, содержащая SEQ ID NO: 86 (VH1D hMAB-3 к CD137), представляет собой SEQ ID NO: 123:

I. TRIDENT-A
[000396] TRIDENT-A представляет собой трехвалентную CD137×CD137×TA связывающую молекулу, имеющую два сайта связывания CD 137 и один сайт связывания для примерного ТА, HER2/neu. TRIDENT-A состоит из четырех полипептидных цепей (см. Фигуру 6А, причем VL1/VH1 (сайт А) идентичны VL2/VH2 (сайт В) и связывают CD 137, и VL3/VH3 (сайт С) связывают HER2/neu)).
[000397] Первая полипептидная цепь TRIDENT-A содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD 137 (VLCD137) (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD 137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; ASTKG (SEQ ID NO: 19)), цистеинсодержащий домен, способствующий образованию гетеродимера (Е-спираль) (EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 38)), промежуточный линкерный пептид (GGGDKTHTCPPCP (SEQ ID NO: 21)), «несущий выступ» домен СН2 и домен СН3 (SEQ ID NO: 44) и С-конец.
[000398] Соответственно, первая полипептидная цепь TRIDENT-A состоит из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 19 - SEQ ID NO: 38 - SEQ ID NO: 21 - SEQ ID NO: 44.
[000399] Аминокислотная последовательность первой полипептидной цепи TRIDENT-A представляет собой (SEQ ID NO: 192):

[000400] Вторая полипептидная цепь TRIDENT-А содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD 137 (VLCD137) (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD 137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; ASTKG (SEQ ID NO: 19)), цистеинс о держащий домен, способствующий образованию гетеродимера (K-спираль) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO: 39)), и С-конец.
[000401] Соответственно, вторая полипептидная цепь TRIDENT-А состоит из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 19 - SEQ ID NO: 39
[000402] Аминокислотная последовательность второй полипептидной цепи TRIDENT-A представляет собой (SEQ ID NO: 197):

[000403] Можно применять альтернативную первую и вторую полипептидные цепи TRIDENT-A, в которых аминокислотные остатки SEQ ID NO: 76 (домен VH моноклонального антитела, способного связывать CD137 (VHCD137)) заменены аминокислотными остатками SEQ ID NO: 83 (VH1A hMAB-3 к CD137), SEQ ID NO: 84 (VH1B hMAB-3 к CD137), SEQ ID NO: 85 (VH1C hMAB-3 к CD137), SEQ ID NO: 86 (VH1D hMAB-3 к CD137), SEQ ID NO: 208 (VH1E hMAB-3 к CD137), SEQ ID NO: 209 (VH1F hMAB-3 к CD137) или SEQ ID NO: 210 (VH1G hMAB-3 к CD137) и/или аминокислотные остатки SEQ ID NO: 89 (домен VL моноклонального антитела, способного связывать CD 137 (VHCD137)) заменены аминокислотными остатками SEQ ID NO: 211 (VL4 hMAB-3 к CD137), SEQ ID NO: 212 (VL5 hMAB-3 к CD137), SEQ ID NO: 213 (VL6 hMAB-3 к CD137), SEQ ID NO: 214 (VL7 hMAB-3 к CD137), SEQ ID NO: 215 (VL8 hMAB-3 к CD137), SEQ ID NO: 216 (VL9 hMAB-3 к CD137), SEQ ID NO: 217 (VL10 hMAB-3 к CD137), SEQ ID NO: 218 (VL11 hMAB-3 к CD137), SEQ ID NO: 219 (VL12 hMAB-3 к CD137), SEQ ID NO: 220 (VL13 hMAB-3 к CD137), SEQ ID NO: 221 (VL14 hMAB-3 к CD137) или SEQ ID NO: 222 (VL15 hMAB-3 к CD137) Оптимизированные/деиммунизированные молекулы, содержащие множество таких полипептидных цепей, описаны ниже.
[000404] Третья полипептидная цепь TRIDENT-A содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с HER2/neu (VHHER2/neu) (VH1 hMAB-1 к HER2 (SEQ ID NO: 64)), домен CH1 человеческого IgG1 (SEQ ID NO: 3), шарнирную область человеческого IgG1 (SEQ ID NO: 7) и «несущий впадину» домен СН2 и домен СН3 (SEQ ID NO: 47). Соответственно, третья полипептидная цепь TRIDENT-A состоит из: SEQ ID NO: 64 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 47 и имеет ту же аминокислотную последовательность, что и первая полипептидная цепь DART-D (SEQ ID NO: 104), представленная выше.
[000405] Четвертая полипептидная цепь TRIDENT-A содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с HER2/neu (VLHER2/neu) (VL3 hMAB-1 к HER2 (SEQ ID NO: 69)), каппа-домен CL человеческого IgG (SEQ ID NO: 1) и С-конец. Соответственно, четвертая полипептидная цепь TRIDENT-A состоит из: SEQ ID NO: 69 - SEQ ID NO: 1 и имеет ту же аминокислотную последовательность, что и вторая, и пятая полипептидные цепи DART-D (SEQ ID NO: 105), предоставленные выше.
1. Оптимизированные варианты TRIDENT-A
[000406] Оптимизированные варианты TRIDENT-A, обозначенные «TRIDENT-A1», «TRIDENT-A2», «TRIDENT-А3» и «TRIDENT-A4», состоят из четырех полипептидных цепей и содержат HER2/neu-связывающие домены антитела hMAB-1 к HER2 (1.3) и CD137-связывающие домены любого антитела hMAB-3 к CD137 (1A.3)-(1D.3).
[000407] Третья полипептидная цепь такого оптимизированного TRIDENT-А имеет ту же аминокислотную последовательность, что и третья полипептидная цепь TRIDENT-А, которая, как отмечено выше, идентична первой полипептидной цепи DART-D (SEQ ID NO: 104).
[000408] Четвертая полипептидная цепь такого оптимизированного TRIDENT-А имеет ту же аминокислотную последовательность, что и четвертая полипептидная цепь TRIDENT-A, которая, как отмечено выше, идентична второй и пятой полипептидным цепям DART-D (SEQ ID NO: 105).
[000409] Первая и вторая полипептидные цепи таких оптимизированных вариантов TRIDENT-A имеют аминокислотную последовательность SEQ ID NO: 193 и SEQ ID NO: 198 (TRIDENT-A1); SEQ ID NO: 194 и SEQ ID NO: 199 (TRIDENT-A2); SEQ ID NO: 195 и SEQ ID NO: 200 (TRIDENT-A3); или SEQ ID NO: 196 и SEQ ID NO: 201 (TRIDENT-A4), представленные ниже.
[000410] Аминокислотная последовательность первой полипептидной цепи TRIDENT-A1, содержащая SEQ ID NO: 83 (VH1A hMAB-3 к CD137), представляет собой SEQ ID NO: 193:

[000411] Аминокислотная последовательность первой полипептидной цепи TRIDENT-A2, содержащая SEQ ID NO: 84 (VH1B hMAB-3 к CD137), представляет собой SEQ ID NO: 194:


[000412] Аминокислотная последовательность первой полипептидной цепи TRIDENT-А3, содержащая SEQ ID NO: 85 (VH1C hMAB-3 к CD137), представляет собой SEQ ID NO: 195:

[000413] Аминокислотная последовательность первой полипептидной цепи TRIDENT-A4, содержащая SEQ ID NO: 86 (VH1D hMAB-3 к CD137), представляет собой SEQ ID NO: 196:

[000414] Аминокислотная последовательность второй полипептидной цепи TRIDENT-A1, содержащая SEQ ID NO: 83 (VH1A hMAB-3 к CD137), представляет собой SEQ ID NO: 198:


[000415] Аминокислотная последовательность второй полипептидной цепи TRIDENT-A2, содержащая SEQ ID NO: 84 (VH1B hMAB-3 к CD137), представляет собой SEQ ID NO: 199:

[000416] Аминокислотная последовательность второй полипептидной цепи TRIDENT-А3, содержащая SEQ ID NO: 85 (VH1C hMAB-3 к CD137), представляет собой SEQ ID NO: 200:

[000417] Аминокислотная последовательность второй полипептидной цепи TRIDENT-A4, содержащая SEQ ID NO: 86 (VH1D hMAB-3 к CD137), представляет собой SEQ ID NO: 201:

2. Деиммунизированные варианты TRIDENT-A
[000418] Деиммунизированные варианты TRIDENT-A состоят из четырех полипептидных цепей и содержат HER2/neu-связывающие домены антитела hMAB-1 к HER2 (1.3) и домен VH CD 137 любого из VH1E-VH1G hMAB-3 к CD137 и домен VL CD 137 любого из VL4-VL15 hMAB-3 к CD137.
[000419] Примерный деиммунизированный вариант TRIDENT-A, обозначенный «TRIDENT-A5», состоит из четырех полипептидных цепей и содержит HER2/neu-связывающие домены антитела hMAB-1 к HER2 (1.3) и CD137-связывающие домены любого антитела hMAB-3 к CD137 (1Е.15).
[000420] Аминокислотная последовательность первой полипептидной цепи TRIDENT-A5 (содержащая VH1E hMAB-3 к CD137 и VL15 hMAB-3 к CD137), представляет собой (SEQ ID NO: 229):

[000421] Аминокислотная последовательность второй полипептидной цепи TRIDENT-A5 (содержащая VH1E hMAB-3 к CD137 и VL15 hMAB-3 к CD137) представляет собой SEQ ID NO: 230:

[000422] Третья полипептидная цепь такого деиммунизированного TRIDENT-А5 имеет ту же аминокислотную последовательность, что и третья полипептидная цепь TRIDENT-A, которая, как отмечено выше, идентична первой полипептидной цепи DART-D (SEQ ID NO: 104).
[000423] Четвертая полипептидная цепь такого деиммунизированного TRIDENT-A5 имеет ту же аминокислотную последовательность, что и четвертая полипептидная цепь TRIDENT-A, которая, как указано выше, идентичная второй и пятой полипептидным цепям DART-D (SEQ ID NO: 105).
J. TRIDENT-B
[000424] TRIDENT-B представляет собой трехвалентную CD137×CD137×TA связывающую молекулу, имеющую два сайта связывания CD137 и один сайт связывания для примерного ТА, 5Т4. TRIDENT-B состоит из четырех полипептидных цепей (см. Фигуру 6А, причем VL1/VH1 (сайт А) идентичны VL2/VH2 (сайт В) и связывают CD137, и VL3/VH3 (сайт С) связывают 5Т4).
[000425] Первая полипептидная цепь TRIDENT-B содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; ASTKG (SEQ ID NO: 19)), цистеинсодержащий домен, способствующий образованию гетеродимера (Е-спираль) (EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO: 38)), промежуточный линкерный пептид (GGGDKTHTCPPCP (SEQ ID NO: 21)), «несущий выступ» домен СН2 и домен СН3 (SEQ ID NO: 44) и С-конец. Соответственно, первая полипептидная цепь TRIDENT-B состоит из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 19 - SEQ ID NO: 38 - SEQ ID NO: 21 - SEQ ID NO: 44 и имеет ту же аминокислотную последовательность, что и первая полипептидная цепь TRIDENT-A (SEQ ID NO: 192), представленная выше.
[000426] Вторая полипептидная цепь TRIDENT-B содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связывать CD137 (VLCD137) (VL3 hMAB-3 к CD137 (SEQ ID NO: 89)), промежуточный линкерный пептид (линкер 1; GGGSGGGG (SEQ ID NO: 16)), домен VH моноклонального антитела, способного связывать CD137 (VHCD137) (VH1 hMAB-3 к CD137 (SEQ ID NO: 76)), промежуточный линкерный пептид (линкер 2; ASTKG (SEQ ID NO: 19)), цистеинсодержащий домен, способствующий образованию гетеродимера (K-спираль) (KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO: 39)), и С-конец. Соответственно, вторая полипептидная цепь TRIDENT-B состоит из: SEQ ID NO: 89 - SEQ ID NO: 16 - SEQ ID NO: 76 - SEQ ID NO: 19 - SEQ ID NO: 39 и имеет ту же аминокислотную последовательность, что и вторая полипептидная цепь TRIDENT-A (SEQ ID NO: 197), предоставленная выше.
[000427] Можно применять альтернативные варианты первой и второй полипептидных цепей TRIDENT-B, в которых аминокислотные остатки SEQ ID NO: 76 (домен VH моноклонального антитела, способного связывать CD137 (VHCD137)) заменены аминокислотными остатками SEQ ID NO: 83 (VH1A hMAB-3 к CD137), SEQ ID NO: 84 (VH1B hMAB-3 к CD137), SEQ ID NO: 85 (VH1C hMAB-3 к CD137), SEQ ID NO: 86 (VH1D hMAB-3 к CD137), SEQ ID NO: 208 (VH1E hMAB-3 к CD137), SEQ ID NO: 209 (VH1F hMAB-3 к CD137) или SEQ ID NO: 210 (VH1G hMAB-3 к CD137) и/или аминокислотные остатки SEQ ID NO: 89 (домен VL моноклонального антитела, способного связывать CD137 (VHCD137)) заменены аминокислотными остатками SEQ ID NO: 211 (VL4 hMAB-3 к CD137), SEQ ID NO: 212 (VL5 hMAB-3 к CD137), SEQ ID NO: 213 (VL6 hMAB-3 к CD137), SEQ ID NO: 214 (VL7 hMAB-3 к CD137), SEQ ID NO: 215 (VL8 hMAB-3 к CD137), SEQ ID NO: 216 (VL9 hMAB-3 к CD137), SEQ ID NO: 217 (VL10 hMAB-3 к CD137), SEQ ID NO: 218 (VL11 hMAB-3 к CD137), SEQ ID NO: 219 (VL12 hMAB-3 к CD137), SEQ ID NO: 220 (VL13 hMAB-3 к CD137), SEQ ID NO: 221 (VL14 hMAB-3 к CD137) или SEQ ID NO: 222 (VL15 hMAB-3 к CD137). Оптимизированные/деиммунизированные молекулы, содержащие множество таких полипептидных цепей, описаны в настоящем документе и имеют те же аминокислотные последовательности, что и первая и вторая полипептидные цепи оптимизированных/деиммунизированных молекул TRIDENT-A (например, TRIDENT-А1 TRIDENT-A5).
[000428] Третья полипептидная цепь TRIDENT-B содержит, в направлении от N-конца к С-концу, N-конец, домен VH моноклонального антитела, способного связываться с 5Т4 (VH5T4) (VH1 hMAB-1 к 5Т4 (SEQ ID NO: 134)), домен CH1 человеческого IgG1 (SEQ ID NO: 3), шарнирную область человеческого IgG1 (SEQ ID NO: 7) и «несущий впадину» домен СН2 и домен СН3 (SEQ ID NO: 47). Соответственно, первая полипептидная цепь TRIDENT-B состоит из: SEQ ID NO: 134 - SEQ ID NO: 3 - SEQ ID NO: 7 - SEQ ID NO: 47
[000429] Аминокислотная последовательность третьей полипептидной цепи TRIDENT-B представляет собой (SEQ ID NO: 231):


[000430] Четвертая полипептидная цепь TRIDENT-В содержит, в направлении от N-конца к С-концу, N-конец, домен VL моноклонального антитела, способного связываться с 5Т4 (VL5T4) (VL1 hMAB-1 к 5Т4 (SEQ ID NO: 135)), каппа-домен CL человеческого IgG (SEQ ID NO: 1) и С-конец. Соответственно, четвертая полипептидная цепь TRIDENT-B состоит из: SEQ ID NO: 135 - SEQ ID NO: 1.
[000431] Аминокислотная последовательность четвертой полипептидной цепи TRIDENT-B представляет собой (SEQ ID NO: 232):

1. Оптимизированные и деиммунизированные варианты TRIDENT-B
[000432] Оптимизированные и деиммунизированные варианты TRIDENT-B состоят из четырех полипептидных цепей и содержат 5Т4-связывающие домены антитела hMAB-1 к 5Т4, а также домен VH CD137 любого из VH1A-VH1G hMAB-3 к CD137 и домен VL CD137 любого из VL3-VL15 hMAB-3 к CD137.
[000433] Примерные оптимизированные и деиммунизированные варианты TRIDENT-B, обозначенные «TRIDENT-B2» и «TRIDENT-B5», состоят из четырех полипептидных цепей и содержат 5Т4-связывающие домены антитела hMAB-1 к 5Т4 и CD137-связывающие домены антитела hMAB-3 к CD137 (1 В.3) и hMAB-3 к CD137 (1Е.15), соответственно.
[000434] Первая полипептидная цепь TRIDENT-В2 имеет ту же аминокислотную последовательность, что и первая полипептидная цепь TRIDENT-A2 (SEQ ID NO: 194), представленная выше.
[000435] Вторая полипептидная цепь TRIDENT-B2 имеет ту же аминокислотную последовательность, что и вторая полипептидная цепь TRIDENT-А2 (SEQ ID NO: 199), представленная выше.
[000436] Первая полипептидная цепь TRIDENT-В5 имеет ту же аминокислотную последовательность, что и первая полипептидная цепь TRIDENT-A5 (SEQ ID NO: 229), представленная выше.
[000437] Вторая полипептидная цепь TRIDENT-B5 имеет ту же аминокислотную последовательность, что и второй полипептид TRIDENT-A5 (SEQ ID NO: 230), представленный выше.
[000438] Третьи полипептидные цепи как примерных оптимизированных вариантов, так и деиммунизированных вариантов TRIDENT-B имеют ту же аминокислотную последовательность, что и третья полипептидная цепь TRIDENT-В (SEQ ID NO: 231).
[000439] Четвертая полипептидная цепь как примерных оптимизированных вариантов, так и деиммунизированных вариантов TRIDENT-B имеет ту же аминокислотную последовательность, что и четвертая полипептидная цепь TRIDENT-B (SEQ ID NO: 232).
K. Альтернативные CD137×TA связывающие молекулы
[000440] Как будет понятно в свете настоящего описания, дополнительные CD137×TA связывающие молекулы, имеющие общую структуру любой из приведенных выше примерных молекул и содержащие сайт связывания для альтернативного опухолевого антигена и/или имеющие оптимизированный/деиммунизированный сайт связывания CD137, могут быть сконструированы с применением доменов VL и VH альтернативных антител к опухолевому антигену вместо доменов VL и VH антител к HER2/neu или антител к 5Т4 и/или доменов VL и VH любого из оптимизированных/деиммунизированных антител к CD137, раскрытых в настоящем документе. Аналогичным образом, как предусмотрено в настоящем документе, альтернативные CD137×TA связывающие молекулы также могут быть сконструированы с включением альтернативных линкеров и/или доменов, способствующих образованию гетеродимера.
V. Контрольные молекулы
[000441] Для того чтобы продемонстрировать в большей степени свойства CD137×TA связывающих молекул согласно настоящему изобретению, ниже представлены контрольные антитела, домены VL и VH которых можно применять для получения контрольных Fc-несущих диател и других контрольных связывающих молекул.
[000442] Антитело к флуоресцеину 4-4-20 (Gruber, М. et al. (1994) ((Efficient Tumor Cell Lysis Mediated By A Bispecific Single Chain Antibody Expressed In Escherichia coli», J. Immunol. 152(11):5368-5374; Bedzyk, W.D. et al. (1989) «Comparison Of Variable Region Primary Structures Within An Anti-Fluorescein Idiotype Family», J. Biol. Chem. 264(3): 1565-1569) представляет собой подходящее контрольное антитело, диатела. Аминокислотные последовательности вариабельного легкого и вариабельного тяжелого доменов антитела к флуоресцеину 4-4-20 представлены далее:
[000443] Аминокислотная последовательность домена VH антитела к флуоресцеину 4-4-20 (SEQ ID NO: 124) представлена ниже (остатки CDRH выделены подчеркиванием):
EVKLDETGGG LVQPGRPMKL SCVASGFTFS DYWMNWVRQS PEKGLEWVAQ IRNKPYNYET YYSDSVKGRF TISRDDSKSS VYLQMNNLRV EDMGIYYCTG SYYGMDYWGQ GTSVTVSS
[000444] Аминокислотная последовательность домена VL антитела к флуоресцеину 4-4-20 (SEQ ID NO: 125) представлена ниже (остатки CDRL выделены подчеркиванием):
DWMTQTPFS LPVSLGDQAS ISCRSSQSLV HSNGNTYLRW YLQKPGQSPK VLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP WTFGGGTKLE IK
[000445] Паливизумаб (см., например, Protein Data Bank (PDB) ID No. 2HWZ) представляет собой гуманизированное моноклональное антитело (IgG), направленное против эпитопа в антигенном сайте А белка F РСВ, и представляет собой подходящее контрольное антитело, домены VL и VH которого можно применять для получения контрольных диател и других контрольных связывающих молекул. Альтернативные антитела к гликопротеину F РСВ включают мотавизумаб (см., например, PDB ID No. 3IXT) и вариант паливизумаба, модифицированный для удаления остатков цистеина из CDR1 легкой цепи. Вариант паливизумаба применяли для получения контрольных молекул, представленных ниже.
[000446] Аминокислотная последовательность домена VH варианта паливизумаба (SEQ ID NO: 126) представлена ниже (остатки CDRH выделены подчеркиванием):
QVTLRESGPA LVKPTQTLTL TCTFSGFSLS TSGMSVGWIR QPPGKALEWL ADIWWDDKKD YNPSLKSRLT ISKDTSKNQV VLKVTNMDPA DTATYYCARS MITNWYFDVW GAGTTVTVSS
[000447] Аминокислотная последовательность домена VL варианта паливизумаба (SEQ ID NO: 127) представлена ниже (остатки CDRL выделены подчеркиванием):
DIQMTQSPST LSASVGDRVT ITCRASQSVG YMHWYQQKPG KAPKLLIYDT SKIASGVPSR FSGSGSGTEF TLTISSLQPD DFATYYCFQG SGYPFTFGGG TKLEIK
VI. Обобщенное описание CD137×TA связывающих и контрольных молекул
[000448] В таблице 5 обобщены характерные признаки доменов DART-А - DART-G, TRIDENT-A-A5:

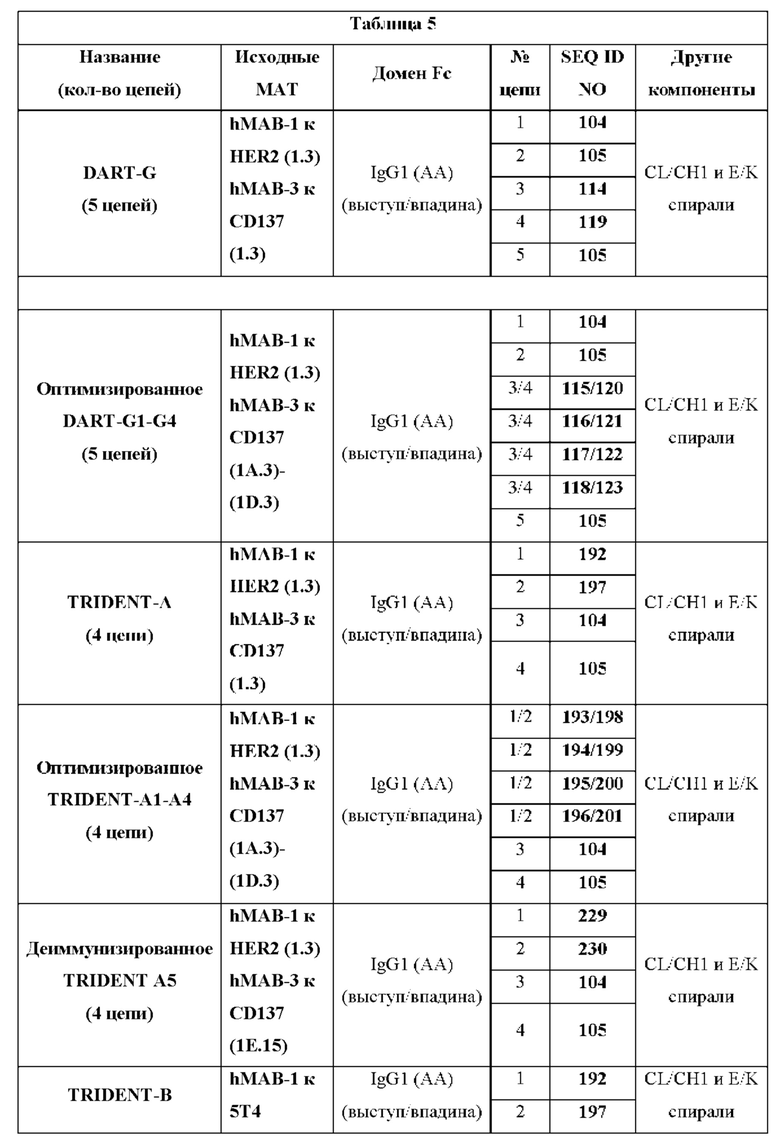

[000449] В таблице 6 представлены характерные признаки дополнительных молекул DART и TRIDENT, которые были получены:



VII. Фармацевтические композиции
[000450] Композиции согласно настоящему изобретению включают нерасфасованные лекарственные композиции, которые можно применять при изготовлении фармацевтических композиций (например, неочищенных или нестерильных композиций) и фармацевтических композиций (т.е. композиций, которые пригодны для введения субъекту или пациенту), которые можно применять при получении стандартных лекарственных форм. Такие композиции содержат CD137×TA связывающую молекулу согласно настоящему изобретению или комбинацию таких агентов и фармацевтически приемлемый носитель. Предпочтительно композиции согласно настоящему изобретению содержат профилактически или терапевтически эффективное количество CD137×TA биспецифического Fc-несущего диатела согласно настоящему изобретению и фармацевтически приемлемый носитель.
[000451] Настоящее изобретение также охватывает фармацевтические композиции, содержащие CD137×TA связывающие молекулы согласно настоящему изобретению и одну или более дополнительных молекул, которые эффективны при стимуляции иммунного ответа (например, ингибитор контрольной точки иммунного ответа) и/или в комбинации с одной или более дополнительными молекулами, которые специфично связывают опухолевый антиген (например, опухолеспецифичное моноклональное антитело или диатело), которые специфичны для по меньшей мере одного конкретного опухолевого антигена (ТА, как описано выше), и фармацевтически приемлемый носитель.
[000452] Согласно конкретному варианту реализации настоящего изобретения термин «фармацевтически приемлемый» означает одобренный регулирующим органом федерального правительства или правительства штата или перечисленный в фармакопее США или другой общепризнанной фармакопее для применения у животных и, в частности, у человека. Термин «носитель» относится к разбавителю, адъюванту (например, адъюванту Фрейнда (полному и неполному), вспомогательному веществу или среде, с которыми вводят терапевтическое средство. Такими фармацевтическими носителями могут быть стерильные жидкости. Водные носители, такие как солевые растворы, водные растворы декстрозы и глицерина являются предпочтительными, если фармацевтическую композицию вводят внутривенно.
[000453] Обычно ингредиенты композиций согласно настоящему изобретению поставляются либо по отдельности, либо смешанными в виде стандартной лекарственной формы, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметически закрытом контейнере, таком как ампула или саше, с указанием количества активного агента. Если композиция предназначена для введения путем инфузии, она может распределяться с флаконом для инфузии, содержащим стерильную воду или физиологический раствор фармацевтического качества. Если композиция предназначена для введения путем инъекции, может быть обеспечена ампула стерильной воды для инъекций или физиологического раствора для смешивания ингредиентов перед введением.
[000454] Согласно настоящему изобретению также предложена фармацевтическая упаковка или набор, содержащий один или более контейнеров, содержащих CD137×TA связывающую молекулу согласно настоящему изобретению, по отдельности или с другими агентами, предпочтительно с фармацевтически приемлемым носителем. Помимо этого один или более других профилактических или терапевтических агентов, которые можно применять для лечения заболевания, также могут быть включены в фармацевтическую упаковку или набор. Согласно настоящему изобретению также предложена фармацевтическая упаковка или набор, содержащий один или более контейнеров, заполненных одним или более ингредиентами фармацевтических композиций согласно настоящему изобретению. При желании с таким (и) контейнером (ами) может быть связано уведомление в форме, предписанной правительственным агентством, регулирующим изготовление, применение или продажу фармацевтических препаратов или биологических продуктов, причем это уведомление отражает одобрение этим агентством изготовления, применения или продажи для введения человеку.
[000455] Набор может содержать CD137×TA связывающую молекулу согласно настоящему изобретению. Набор может дополнительно содержать один или более других профилактических и/или терапевтических агентов, которые можно применять для лечения рака, в одном или более контейнерах; и/или набор может дополнительно содержать одно или более цитотоксических антител, которые связывают один или более опухолевых антигенов (ТА). Согласно определенным вариантам реализации другой профилактический или терапевтический агент представляет собой химиотерапевтическое средство. Согласно другим вариантам реализации профилактическое или терапевтическое средство представляет собой биологическое или гормональное терапевтическое средство.
VIII. Способы введения
[000456] Композиции согласно настоящему изобретению могут быть обеспечены для лечения, профилактики и улучшения одного или более симптомов, ассоциированных с раком или другим заболеванием, или нарушением, путем введения субъекту эффективного количества молекулы согласно настоящему изобретению или фармацевтической композиции, содержащей молекулу согласно настоящему изобретению. Согласно предпочтительному аспекту такие композиции являются по существу чистыми (т.е. по существу не содержат вещества, которые ограничивают их действие или вызывают нежелательные побочные эффекты). Согласно конкретному варианту реализации настоящего изобретения субъект представляет собой животное, предпочтительно млекопитающее, такое как млекопитающее, отличное от примата (например, бычьих, лошадиных, кошачьих, собачьих, грызунов и т.д.), или примата (например, обезьяну, такую как яванская макака, человек и т.д.). В предпочтительном варианте реализации субъект представляет собой человека.
[000457] Различные системы доставки известны и могут применяться для введения молекул и композиций согласно настоящему изобретению, например, инкапсуляция в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать антитело или гибридный белок, опосредуемый рецепторами эндоцитоз (см., например, Wu et al. (1987) «Receptor-Mediated In Vitro Gene Transformation By A Soluble DNA Carrier System», J. Biol. Chem. 262:4429-4432), конструирование нуклеиновой кислоты как части ретровирусного или другого вектора и т.д.
[000458] Способы введения молекулы согласно настоящему изобретению включают, но не ограничиваются указанными, парентеральное введение (например, внутрикожное, внутримышечное, внутрибрюшинное, внутривенное и подкожное), эпидуральное и через слизистые оболочки (например, интраназальный и пероральный пути). Согласно конкретному варианту реализации настоящего изобретения CD137×TA связывающие молекулы согласно настоящему изобретению вводят внутримышечно, внутривенно или подкожно. Композиции можно вводить любым удобным путем, например, путем инфузии или болюсной инъекции, путем всасывания через эпителиальные или слизистые оболочки (например, слизистую оболочку полости рта, слизистую оболочку прямой кишки и слизистую оболочку кишечника и т.д.) и можно вводить вместе с другими биологически активными агентами. Введение может быть системным или локальным.
[000459] Согласно настоящему изобретению также предложены CD137×TA связывающие молекулы согласно настоящему изобретению, упакованные в герметически закрытый контейнер, такой как ампула или саше, с указанием количества молекулы. Согласно одному варианту реализации настоящего изобретения CD137×TA связывающие молекулы согласно настоящему изобретению поставляются в виде сухого стерилизованного лиофилизированного порошка или безводного концентрата в герметически закрытом контейнере и могут быть восстановлены, например, с применением воды или солевого раствора до соответствующей концентрации для введения субъекту. Предпочтительно CD137×TA связывающие молекулы согласно настоящему изобретению поставляются в виде сухого стерильного лиофилизированного порошка в герметически закрытом контейнере.
[000460] Лиофилизированные CD137×TA связывающие молекулы согласно настоящему изобретению следует хранить при температуре от 2 до 8°С в их оригинальном контейнере, и молекулы следует вводить в течение 12 часов, предпочтительно в течение 6 часов, в течение 5 часов, в течение 3 часов или в течение 1 часа после восстановления. В альтернативном варианте реализации CD137×TA связывающие молекулы согласно настоящему изобретению поставляются в жидкой форме в герметически закрытом контейнере с указанием количества и концентрации молекулы, гибридного белка или конъюгированной молекулы. Предпочтительно жидкая форма CD137×TA связывающих молекул согласно настоящему изобретению поставляется в герметически закрытом контейнере.
[000461] Количество композиции согласно настоящему изобретению, которое будет эффективным в лечении, предотвращении или улучшении одного или более симптомов, ассоциированных с нарушением, может быть определено с помощью стандартных клинических методик. Точная доза, которая будет применяться в составе, также будет зависеть от пути введения и серьезности состояния и должна определяться в соответствии с суждением практикующего врача и обстоятельствами каждого пациента. Эффективные дозы могут быть экстраполированы из кривых доза-ответ, полученных из тестовых систем в условиях in vitro или в моделях на животных.
[000462] В настоящем документе термин «эффективное количество» фармацевтической композиции, в одном варианте реализации, представляет собой количество, достаточное для достижения благоприятных или желательных результатов, включая, но не ограничиваясь указанными, клинические результаты, такие как уменьшение симптомов, вызванных заболеванием, ослабление симптома заболевания (например, пролиферации раковых клеток, наличия опухоли, метастазов опухоли и т.д.), чтобы повысить качество жизни тех, кто страдает этим заболеванием, уменьшение дозы других лекарственных средств, необходимых для лечения заболевания, усиление эффекта другого лекарственного средства, например, за счет нацеливания и/или интернализации, задержку прогрессирования заболевания и/или увеличение продолжительности выживания индивидуумов.
[000463] Такое эффективное количество можно вводить один или несколько раз. Для целей данного изобретения эффективное количество лекарственного препарата, соединения или фармацевтической композиции представляет собой количество, достаточное для уменьшения пролиферации (или эффекта) присутствующего вируса и для уменьшения и/или задержки развития заболевания (например, рака), будь то непосредственно или косвенно. Согласно некоторым вариантам реализации настоящего изобретения эффективное количество лекарственного препарата, соединения или фармацевтической композиции может быть достигнуто в сочетании с другим лекарственным препаратом, соединением или фармацевтической композицией, но не обязательно. Таким образом, «эффективное количество» может рассматриваться в отношении введения одного или более химиотерапевтических агентов, и один агент может считаться введенным в эффективном количестве, если, в сочетании с одним или более другими агентами, может быть достигнут или достигается желательный результат. Несмотря на то, что индивидуальные потребности различаются, определение оптимальных диапазонов эффективных количеств каждого компонента находится в пределах квалификации в данной области техники.
[000464] Для CD137×TA связывающих молекул, охватываемых настоящим изобретением, вводимую пациенту дозу предпочтительно определяют на основании массы тела (кг) субъекта-реципиента. Вводимая доза, как правило, составляет от приблизительно 0,01 мкг/кг до приблизительно 150 мг/кг или более от массы тела субъекта.
[000465] Дозировка и частота введения CD137×TA связывающих молекул согласно настоящему изобретению может быть уменьшена или изменена путем усиления поглощения и проникновения в ткани CD137×TA связывающих молекул с помощью таких модификаций как, например, липидирование.
[000466] Дозировка CD137×TA связывающих молекул согласно настоящему изобретению, вводимых пациенту, может быть рассчитана для применения в качестве терапии одним агентом. Согласно другому варианту CD137×TA связывающие молекулы согласно настоящему изобретению применяют в комбинации с другими терапевтическими композициями так, что вводимая пациенту дозировка ниже, чем при применении указанных молекул в качестве терапии одним агентом.
[000467] Лечение субъекта с применением терапевтически или профилактически эффективного количества CD137×TA связывающих молекул согласно настоящему изобретению может включать однократное введение или предпочтительно может включать несколько введений. В предпочтительном примере субъекта лечат с применением такого диатела один раз в неделю, один раз в две недели (т.е. один раз каждые две недели) или один раз каждые три недели в течение от 1 до 52 недель. Фармацевтические композиции согласно настоящему изобретению можно вводить один раз в сутки, два раза в сутки или три раза в сутки. Согласно другому варианту фармацевтические композиции можно вводить один раз в неделю, два раза в неделю, один раз каждые две недели, один раз в месяц, один раз каждые шесть недель, один раз каждые два месяца, два раза в год или один раз в год. Также следует понимать, что эффективная дозировка молекул, применяемых для лечения, может увеличиваться или уменьшаться в ходе конкретного лечения.
IX. Применение композиций согласно настоящему изобретению.
[000468] CD137×TA связывающие молекулы согласно настоящему изобретению способны связывать Т-клетки (АПК) (например, за счет связывания с CD 137, экспрессируемым на поверхности таких Т-клеток) и способны связывать ТА-экспрессирующие опухолевые клетки (например, за счет связывания с ТА, экспрессируемым на поверхности таких опухолевых клеток). Таким образом, CD137×TA связывающие молекулы согласно настоящему изобретению способны ко локализовать Т-клетки с опухолевыми клетками, экспрессирующими ТА, и, соответственно, могут применяться для лечения любого заболевания или состояния, ассоциированного или характеризующегося экспрессией ТА. Таким образом, но не ограничиваясь этим, фармацевтические композиции, содержащие такие молекулы, можно применять для диагностики или лечения видов рака, которые экспрессируют ТА, включая такие виды рака, которые характеризуются наличием раковой клетки, включая, но не ограничиваясь раковой клеткой, острого миелолейкоза, опухоли надпочечников, СПИД-ассоциированного рака, альвеолярной саркомы мягких тканей, астроцитарной опухоли, рака мочевого пузыря, рака костей, рака головного и спинного мозга, метастатической опухоли головного мозга, рака молочной железы, опухолей сонных артерий, рака шейки матки, хондросаркомы, хордомы, хромофобной карциномы почек, светлоклеточной карциномы, рака толстой кишки, колоректального рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, экстраскелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии кости, рака желчного пузыря или желчного протока, рака желудка, гестационного трофобластического заболевания, герминогенной опухоли, рака головы и шеи, гепатоцеллюлярной карциномы, глиобластомы, опухоли из островковых клеток, саркомы Капоши, рака почек, лейкоза, липомы/доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легких, медуллобластомы, меланомы, менингиомы, злокачественной мезотелиомы, множественной эндокринной неоплазии, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринных опухолей, немелкоклеточного рака легких, рака яичников, рака поджелудочной железы, рака глотки, папиллярной карциномы щитовидной железы, опухоли околощитовидной железы, педиатрического рака, опухоли оболочек периферических нервов, феохромоцитомы, опухоли гипофиза, рака предстательной железы, постериорной увеальной меланомы, редкого гематологического заболевания, карциномы почек, метастатического рака почек, рабдоидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, рака яичек, рака тимуса, тимомы, метастатического рака щитовидной железы или рака матки.
[000469] В частности, CD137×TA связывающие молекулы согласно настоящему изобретению можно применять для лечения разных видов плоскоклеточного рака головы и шеи (SCCHN), разных видов рака мочевого пузыря, разных видов рака молочной железы, разных видов колоректального рака, разных видов рака желудка, глиобластом, разных видов рака почек, разных видов рака легких, включая немелкоклеточный рак легких (NSCLC), меланом, разных видов рака яичников, разных видов рака поджелудочной железы, разных видов рака глотки, разных видов рака предстательной железы, карцином почек и опухолей из небольших округлых синих клеток у детей, включая нейробластомы и рабдомиосаркомы, каждая из которых экспрессирует ТА на высоком уровне.
[000470] CD137×TA связывающие молекулы согласно настоящему изобретению можно дополнительно применять при изготовлении лекарственных средств для лечения описанных выше состояний.
[000471] Как продемонстрировано в настоящем документе, CD137×TA связывающие молекулы согласно настоящему изобретению повышают активность агентов, нацеленных на опухоль. В соответствии с этим CD137×TA связывающие молекулы согласно настоящему изобретению можно дополнительно применять в комбинации с другими агентами, нацеленными на опухоль, включая, но не ограничиваясь указанными, антитело, антигенсвязывающий фрагмент антитела (например, scFv, Fab, F(ab)2 и т.д., TandAb и т.д.), полиспецифическую связывающую молекулу (например, диатело, биспецифическое антитело, трехвалентную связывающую молекулу и т.д.), способные связывать желательный ТА. Конкретно предусмотрено, что агент, нацеленный на опухоль, может связывать тот же или другой ТА, что и CD137×TA связывающая молекула, применяемая в таких комбинациях. В конкретных вариантах реализации агент, нацеленный на опухоль, представляет собой полиспецифическую молекулу, которая связывается с ТА и эпитопом, экспрессируемым на Т-клетках, включая, например, CD3 и/или CD8. Примерные агенты, нацеленные на опухоль, включают, но не ограничиваются указанными, молекулы, которые связываются с ТА и CD3 («TA×CD3»). Примерные TA×CD3 связывающие молекулы (например, биспецифические антитела, молекулы DART®, молекулы BiTe®, TandAb и т.д.) и способы их получения, которые можно применять в таких комбинациях, хорошо известны в данной области техники (см., например, WO 2013026835, WO 2013158856, WO 2014047231; WO 2014110601; WO 2014131711; WO 2015/026894; WO 2015026892; WO 2015/184203; WO 2016/036937; WO 2016182751; WO 2017091656; WO 2017/142928; WO 2017118675).
[000472] Как предусмотрено в настоящем документе, применение CD137×TA связывающих молекул согласно настоящему изобретению в комбинации с агентом, нацеленным на опухоль (например, TA×CD3 связывающей молекулой), может приводить к активации ингибирующего иммунного модулятора, белка запрограммированной гибели клеток 1 («PD-1», также известного как «CD279»). PD-1 опосредует ингибирование иммунной системы, связывая PD-L1 и PD-L2 (также известные как В7-Н1 и B7-DC) (Flies, D.B. et at. (2007) «The New B7s: Playing a Pivotal Role in Tumor Immunity», J. Immunother. 30(3):251-260; патенты США №№6803192; 7794710; публикации заявок на патент США №№2005/0059051; 2009/0055944; 2009/0274666; 2009/0313687; РСТ публикации №WO 01/39722; WO 02/086083). Однако, как показано в настоящем документе, дополнительное добавление агента, который ингибирует ингибирующую активность PD-1 («ингибитор контрольной точки иммунного ответа PD-1/PD-L1»), подавляет экспрессию PD-1 и дополнительно повышает активность CD137×TA и агентов, нацеленных на опухоль, таких как TA×CD3 связывающие молекулы. В частности, настоящее изобретение охватывает ингибиторы контрольной точки иммунного ответа PD-1/PD-L1, содержащие эпитопсвязывающий сайт антитела, которое связывает PD-1.
[000473] Соответственно, CD137×TA связывающие молекулы согласно настоящему изобретению можно дополнительно применять в комбинации с другими агентами, нацеленными на опухоль, в дополнительной комбинации с ингибитором контрольной точки иммунного ответа PD-1/PD-L1. Ингибиторы контрольной точки иммунного ответа PD-1/PD-L1 включают, но не ограничиваются указанными, антитело, антигенсвязывающий фрагмент антитела (например, scFv, Fab, F(ab)2 и т.д., TandAb и т.д.), полиспецифическую связывающую молекулу (например, диатело, биспецифическое антитело, трехвалентную связывающую молекулу и т.д.), способную связываться с PD-1 и/или PD-L1. Примерные ингибиторы контрольной точки иммунного ответа PD-1/PD-L1 и способы их изготовления, которые можно применять в таких комбинациях, хорошо известны в данной области техники (см., например, патенты США №№9617338; 9273135; 9062112; 8981063; 8779108; 8609089; 8552154; 8460927; 8008449; публикации патентов США №№2015/0197571; 2016/0075782; 2016/0159905; 2016/0319019; 2017/0044259; РСТ публикации патентов №№ WO 2004/056875; WO 2006/121168; WO 2008/156712; WO 2012/135408; WO 2012/145549; WO 2013/014668; WO 2014/055897; WO 2013/079174; WO 2014/179664; WO 2014/194302; WO 2015/109124; WO 2015/112800; WO 2015/112805; WO 2016/000619; WO 2016007235; WO 2016/061142; WO 2016/111645; WO 2016/210262; WO 2016/014688; WO 2016/077397; WO 2017/019846; WO 2017/079112; WO 2017/087547; и WO 2017/106656).
[000474] При применении таких комбинаций конкретно предусмотрено, что одна или более молекул могут быть введены субъекту «одновременно» (например, CD137×TA связывающая молекула может быть введена одновременно с введением TA×CD3 связывающей молекулы и/или ингибитора контрольной точки иммунного ответа PD-1/PD-L1) и/или что одну или более молекул можно вводить «последовательно» (например, вводят CD137×TA связывающую молекулу и позднее вводят TA×CD3 связывающую молекулу и/или ингибитор контрольной точки иммунного ответа PD-1/PD-L1, или наоборот).
X. Конкретные варианты реализации изобретения
[000475] Настоящее изобретение было описано выше в общих чертах, оно будет более понятно со ссылкой на следующие пронумерованные варианты реализации («Е»), которые представлены в качестве иллюстрации и не предназначены для ограничения настоящего изобретения, если не указано иное:
E1. CD137×TA связывающая молекула, которая характеризуется тем, что такая связывающая молекула способна специфично связываться с эпитопом CD137 и эпитопом опухолевого антигена (ТА) и такая CD137×TA связывающая молекула содержит первый вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и первый вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; и при этом
(A) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL15 МАВ-3 к CD137 (SEQ ID NO: 222); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(B) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL14 МАВ-3 к CD137 (SEQ ID NO: 221); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(C) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL11 МАВ-3 к CD137 (SEQ ID NO: 218); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(D) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL10 МАВ-3 к CD137 (SEQ ID NO: 217); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(E) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL6 МАВ-3 к CD137 (SEQ ID NO: 213); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(F) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL4 МАВ-3 к CD137 (SEQ ID NO: 211); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(G) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-3 к CD137 (SEQ ID NO: 74);
(H) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-4 к CD137 (SEQ ID NO: 91); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-4 к CD137 (SEQ ID NO: 90);
(I) (1) C CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-5 к CD137 (SEQ ID NO: 97); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-5 к CD137 (SEQ ID NO: 96);
(J) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1A МАВ-3 к CD137 (SEQ ID NO: 83);
(K) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(L) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1C МАВ-3 к CD137 (SEQ ID NO: 85); или
(М) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1D МАВ-3 к CD137 (SEQ ID NO: 86).
Е2. CD137×TA связывающая молекула по п. Е1, которая характеризуется тем, что указанный первый вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) ЬМАВ-3 к CD137 (SEQ ID NO: 77); или
(B) hMAB-4 к CD137 (SEQ ID NO: 92).
Е3. CD137×TA связывающая молекула по п. Е1 или Е2, которая характеризуется тем, что указанный первый вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 82); или
(B) hMAB-4 к CD137 (SEQ ID NO: 93).
Е4. CD137×TA связывающая молекула по любому из пп. Е1-Е3, которая характеризуется тем, что указанный первый вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1E hMAB-3 к CD137 (SEQ ID NO: 208);
(B) VH1B hMAB-3 к CD137 (SEQ ID NO: 84);
(C) VH1A hMAB-3 к CD137 (SEQ ID NO: 83);
(D) VH1 hMAB-3 к CD137 (SEQ ID NO: 76);
(E) VH1C hMAB-3 к CD137 (SEQ ID NO: 85);
(F) VH1D hMAB-3 к CD137 (SEQ ID NO: 86);
(G) VH1F hMAB-3 к CD137 (SEQ ID NO: 209);
(H) VH1G hMAB-3 к CD137 (SEQ ID NO: 210); или
(I) VH1 hMAB-4 к CD137 (SEQ ID NO: 92).
E5. CD137×TA связывающая молекула по любому из пп. Е1-Е4, которая характеризуется тем, что указанный первый вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL15 hMAB-3 к CD137 (SEQ ID NO: 222);
(B) VL14 hMAB-3 к CD137 (SEQ ID NO: 221);
(C) VL1 hMAB-3 к CD137 (SEQ ID NO: 87);
(D) VL2 hMAB-3 к CD137 (SEQ ID NO: 88);
(E) VL3 hMAB-3 к CD137 (SEQ ID NO: 89);
(F) VL4 hMAB-3 к CD137 (SEQ ID NO: 211);
(G) VL5 hMAB-3 к CD137 (SEQ ID NO: 212);
(H) VL6 hMAB-3 к CD137 (SEQ ID NO: 213);
(I) VL7 hMAB-3 к CD137 (SEQ ID NO: 214);
(J) VL8 hMAB-3 к CD137 (SEQ ID NO: 215);
(K) VL9 hMAB-3 к CD137 (SEQ ID NO: 216);
(L) VL10 hMAB-3 к CD137 (SEQ ID NO: 217);
(M) VL11 hMAB-3 к CD137 (SEQ ID NO: 218);
(N) VL12 hMAB-3 к CD137 (SEQ ID NO: 219);
(O) VL13 hMAB-3 к CD137 (SEQ ID NO: 220);
(P) VL1 hMAB-4 к CD137 (SEQ ID NO: 94); или
(Q) VL2 hMAB-4 к CD137 (SEQ ID NO: 95).
E6. CD137×TA связывающая молекула по любому из пп. Е1-Е5, которая характеризуется тем, что указанный опухолевый антиген (ТА) выбран из группы опухолевых антигенов, состоящей из: 19.9; онкофетального белка 5Т4; антигена 4.2; А33; AFP; ALCAM; BAGE; бета-катенина; СА125; карбоксипептидазы М; B1;CD5; CD19; CD20; CD22; CD23; CD25; CD27; CD30; CD33; CD36; CD46; CD52; CD79a/CD79b; CD123; CD317; СЕА; CEACAM5; СЕАСАМ6; СО-43; СО-514; CTLA-1; CTLA-4; цитокератина 8; ряда Е1; ЭФР-Р; рецептора эфрина; Erb; F3; FC10.2; GAGE GD2; GD3; GD49; GM2; GM3; GICA 19-9; gp37; gp75; gp100; HER-2/neu; антигена CD20 В-лимфомы человека; антигена жировых глобул человеческого молока; Е6 папилломавируса человека/Е7 папилломавируса человека; HMW-MAA; антигена I; ITGB6; IL13Rα2; JAM-3; KID3; KID31; общего антигена карциномы KS 1/4; KS 1/4; KSA; L6; L20; LEA; LUCA-2; M1:22:25:8; М18; М39; MAGE; MART; My1; MUC-1; MUM-1; N-ацетилглюкозаминилтрансферазы; неогликопротеина; NS-10; OFA-1; OFA-2; онкостатина M; p15; PSA; PSMA; РЕМА; PIPA; простатической кислой фосфатазы; R24; ROR1; SSEA-1; SSEA-3; SSEA-4; sTn; пептида, происходящего из рецептора Т-клеток; TAG-72; TL5; рецептора TNF-α; рецептора TNF-β; рецептора TNF-γ; TRA-1-85; рецептора трансферрина; TSTA; и VEGF-R.
Е7. CD137×TA связывающая молекула по любому из пп. Е1-Е5, которая характеризуется тем, что указанный опухолевый антиген (ТА) выбран из опухолевых антигенов в таблице 1.
Е8. CD137×TA связывающая молекула по любому из пп. Е1-Е7, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой: HER2/neu, EphA2 или 5Т4.
Е9. CD137×TA связывающая молекула по п. Е8, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой HER2/neu и CD137×TA связывающая молекула содержит второй вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и первый вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; и при этом:
(A) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-1 к HER2 (SEQ ID NO: 63); и
(B) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-1 к HER2 (SEQ ID NO: 62).
Е10. CD137×TA связывающая молекула по п. Е9, в которой:
(A) (1) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(2) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(3) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL3 hMAB-1 к HER2 (SEQ ID NO: 69);
и
(B) (1) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(2) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(3) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH3 hMAB-1 к HER2 (SEQ ID NO: 66).
E11. CD137×TA связывающая молекула по п.ЕЮ, которая характеризуется тем, что указанный второй вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(B) VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(C) VH3 hMAB-1 к HER2 (SEQ ID NO: 66).
E12. CD137×TA связывающая молекула по пп. Е10 или Е11, которая характеризуется тем, что указанный второй вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(B) VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(C) VL3 hMAB-1 к HER2 (SEQ ID NO: 69).
E13. CD137×TA связывающая молекула по п. Е8, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой 5Т4 и CD137×TA связывающая молекула содержит второй вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3 , и первый вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; и при этом:
(I) (A) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-1 к 5Т4 (SEQ ID NO: 135); и
(В) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-1 к 5Т4 (SEQ ID NO: 134); или
(II) (A) CDRL1, CDRL2 и CDRL3 второго вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-2 к 5Т4 (SEQ ID NO: 137); и
(В) CDRH1, CDRH2 и CDRH3 второго вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-2 к 5Т4 (SEQ ID NO: 136).
Е14. CD137×TA связывающая молекула по п. Е13, которая характеризуется тем, что указанный второй вариабельный домен тяжелой цепи содержит аминокислотную последовательность: VH1 МАВ-1 (SEQ ID NO: 135).
Е15. CD137×TA связывающая молекула по п. Е13 или Е14, которая характеризуется тем, что указанный второй вариабельный домен легкой цепи содержит аминокислотную последовательность: VH1 МАВ-1 (SEQ ID NO: 136).
Е16. CD137×TA связывающая молекула по любому из пп. Е1-Е15, причем указанная молекула представляет собой биспецифическое четырехвалентное Fc-несущее диатело, содержащее первую, вторую, третью и четвертую полипептидные цепи, при этом указанные полипептидные цепи образуют ковалентно связанный комплекс.
Е17. CD137×TA связывающая молекула по п. Е16, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой HER2/neu и при этом:
(I) (А) первая и третья полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 100; и
(В) вторая и четвертая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 101; или
(II) (А) первая и третья полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 102; и
(В) вторая и четвертая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 103.
Е18. CD137×TA связывающая молекула по любому из пп. Е1-Е15, причем указанная молекула является биспецифической и четырехвалентной и содержит первую, вторую, третью, четвертую и пятую полипептидные цепи, при этом указанные полипептидные цепи образуют ковалентно связанный комплекс.
Е19. CD137×TA связывающая молекула по п. Е15, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой HER2/neu и при этом:
(I) (А) первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 104;
(B) вторая и пятая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 105;
(C) третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 106; и
(D) четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 107;
или
(II) (А) первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 104;
(B) вторая и пятая полипептидные цепи имеют аминокислотную последовательность SEQ ID NO: 105;
(C) третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117 или SEQ ID NO: 118; и
(D) четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 119, SEQ ID NO: 120, SEQ ID NO: 121, SEQ ID NO: 122 или SEQ ID NO: 123.
E20. CD137×TA связывающая молекула по любому из пп. Е1-15, причем указанная молекула является биспецифической и трехвалентной и содержит первую, вторую, третью и четвертую полипептидные цепи, при этом указанные полипептидные цепи образуют ковалентно связанный комплекс.
Е21. CD137×TA связывающая молекула по п. Е20, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой HER2/neu и при этом:
(A) указанная первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195, SEQ ID NO: 196 или SEQ ID NO: 229;
(B) указанная вторая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200, SEQ ID NO: 201 или SEQ ID NO: 230;
(C) указанная третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 104; и
(D) указанная четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 105
Е23. CD137×TA связывающая молекула по п. Е20, которая характеризуется тем, что указанный опухолевый антиген (ТА) представляет собой 5Т4 и при этом:
(A) указанная первая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 192, SEQ ID NO: 193, SEQ ID NO: 194, SEQ ID NO: 195, SEQ ID NO: 196 или SEQ ID NO: 229;
(B) указанная вторая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 197, SEQ ID NO: 198, SEQ ID NO: 199, SEQ ID NO: 200, SEQ ID NO: 201 или SEQ ID NO: 230;
(C) указанная третья полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 231; и
(D) указанная четвертая полипептидная цепь имеет аминокислотную последовательность SEQ ID NO: 232.
Е24. Фармацевтическая композиция, содержащая CD137×TA связывающую молекулу по любому из пп. Е1-Е23 и физиологически приемлемый носитель.
Е25. Применение CD137×TA связывающей молекулы по любому из пп. Е1-Е23 или фармацевтической композиции по п. Е24 в лечении заболевания или состояния, ассоциированного или характеризующегося экспрессией опухолевого антигена (ТА).
Е26. Применение по п. Е25, которое характеризуется тем, что указанное заболевание или состояние, ассоциированное или характеризующееся экспрессией опухолевого антигена (ТА), представляет собой рак.
Е27. CD137-связывающая молекула, которая содержит вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; причем:
(A) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL15 МАВ-3 к CD137 (SEQ ID NO: 222); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(B) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL14 МАВ-3 к CD137 (SEQ ID NO: 221); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(C) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL11 МАВ-3 к CD137 (SEQ ID NO: 218); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(D) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL10 МАВ-3 к CD137 (SEQ ID NO: 217); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(E) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL6 МАВ-3 к CD137 (SEQ ID NO: 213); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(F) (1) указанные CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL4 МАВ-3 к CD137 (SEQ ID NO: 211); и
(2) указанные CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(G) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-3 к CD137 (SEQ ID NO: 74);
(H) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-4 к CD137 (SEQ ID NO: 91); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-4 к CD137 (SEQ ID NO: 90);
(I) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-5 к CD137 (SEQ ID NO: 97); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-5 к CD137 (SEQ ID NO: 96);
(J) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1A МАВ-3 к CD137 (SEQ ID NO: 83);
(K) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1B МАВ-3 к CD137 (SEQ ID NO: 84);
(L) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1C МАВ-3 к CD137 (SEQ ID NO: 85); или
(М) (1) CDRL1, CDRL2 и CDRL3 первого вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-3 к CD137 (SEQ ID NO: 75); и
(2) CDRH1, CDRH2 и CDRH3 первого вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1D МАВ-3 к CD137 (SEQ ID NO: 86).
Е28. CD137-связывающая молекула по п. Е27, которая характеризуется тем, что указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 77); или
(B) hMAB-4 к CD137 (SEQ ID NO: 92).
Е29. CD137-связывающая молекула по п. Е27 или Е28, которая характеризуется тем, что указанный вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) hMAB-3 к CD137 (SEQ ID NO: 82); или
(B) hMAB-4 к CD137 (SEQ ID NO: 93).
Е30. CD137-связывающая молекула по любому из пп. Е27-Е29, которая характеризуется тем, что указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1 hMAB-3 к CD137 (SEQ ID NO: 76);
(B) VH1A hMAB-3 к CD137 (SEQ ID NO: 83);
(C) VH1B hMAB-3 к CD137 (SEQ ID NO: 84);
(D) VH1C hMAB-3 к CD137 (SEQ ID NO: 85);
(E) VH1D hMAB-3 к CD137 (SEQ ID NO: 86);
(F) VH1E hMAB-3 к CD137 (SEQ ID NO: 208);
(G) VH1F hMAB-3 к CD137 (SEQ ID NO: 209);
(H) VH1G hMAB-3 к CD137 (SEQ ID NO: 210); или
(I) VH1 hMAB-4 к CD137 (SEQ ID NO: 92)
Е31. CD137-связывающая молекула по любому из пп. Е27-Е30, которая характеризуется тем, что указанный вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL15 hMAB-3 к CD137 (SEQ ID NO: 222);
(B) VL14 hMAB-3 к CD137 (SEQ ID NO: 221);
(C) VL1 hMAB-3 к CD137 (SEQ ID NO: 87);
(D) VL2 hMAB-3 к CD137 (SEQ ID NO: 88);
(E) VL3 hMAB-3 к CD137 (SEQ ID NO: 89);
(F) VL4 hMAB-3 к CD137 (SEQ ID NO: 211);
(G) VL5 hMAB-3 к CD137 (SEQ ID NO: 212);
(H) VL6 hMAB-3 к CD137 (SEQ ID NO: 213);
(I) VL7 hMAB-3 к CD137 (SEQ ID NO: 214);
(J) VL8 hMAB-3 к CD137 (SEQ ID NO: 215);
(K) VL9 hMAB-3 к CD137 (SEQ ID NO: 216);
(L) VL10 hMAB-3 к CD137 (SEQ ID NO: 217);
(M) VL11 hMAB-3 к CD137 (SEQ ID NO: 218);
(N) VL12 hMAB-3 к CD137 (SEQ ID NO: 219);
(O) VL13 hMAB-3 к CD137 (SEQ ID NO: 220);
(P) VL1 hMAB-4 к CD137 (SEQ ID NO: 94); или
(Q) VL2 hMAB-4 к CD137 (SEQ ID NO: 95).
E32. CD137-связывающая молекула по любому из пп. Е27-31, причем указанная молекула представляет собой антитело или его антигенсвязывающий фрагмент.
Е33. Фармацевтическая композиция, содержащая CD137-связывающую молекулу по любому из пп. Е27-Е32 и физиологически приемлемый носитель.
Е34. Применение CD137-связывающей молекулы по любому из пп. Е27-Е31 или фармацевтической композиции по п. Е33 в лечении заболевания или состояния, ассоциированного с подавленной иммунной системой или характеризующегося экспрессией опухолевого антигена (ТА).
Е35. Применение по п. Е34, которое характеризуется тем, что указанное состояние, ассоциированное с подавленной иммунной системой или характеризующееся экспрессией опухолевого антигена (ТА), представляет собой рак.
Е36. HER2/neu-связывающая молекула, которая содержит вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; причем:
(A) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL МАВ-1 к HER2 (SEQ ID NO: 63); и
(B) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH МАВ-1 к HER2 (SEQ ID NO: 62).
Е37. HER2/neu-связывающая молекула по п. Е36, в которой:
(A) (1) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(2) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(3) CDRL1, CDRL2 и CDRL3 вариабельного домена легкой цепи представляют собой CDR легкой цепи VL3 hMAB-1 к HER2 (SEQ ID NO: 69);
и
(B) (1) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(2) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(3) CDRH1, CDRH2 и CDRH3 вариабельного домена тяжелой цепи представляют собой CDR тяжелой цепи VH3 hMAB-1 к HER2 (SEQ ID NO: 66).
Е38. HER2/neu-связывающая молекула по п. Е337, которая характеризуется тем, что указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность:
(A) VH1 hMAB-1 к HER2 (SEQ ID NO: 64);
(B) VH2 hMAB-1 к HER2 (SEQ ID NO: 65); или
(C) VH3 hMAB-1 к HER2 (SEQ ID NO: 66)
E39. HER2/neu-связывающая молекула по пп. E37-E38, которая характеризуется тем, что указанный вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL1 hMAB-1 к HER2 (SEQ ID NO: 67);
(B) VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(C) VL3 hMAB-1 к HER2 (SEQ ID NO: 69)
E40. HER2/neu-связывающая молекула по любому из пп. E36-E39, которая характеризуется тем, что указанная молекула представляет собой антитело или его антигенсвязывающий фрагмент.
Е41. Фармацевтическая композиция, содержащая HER2/neu-связывающую молекулу по любому из пп. Е36-Е40 и физиологически приемлемый носитель.
Е42. Применение HER2/heu-связывающей молекулы по любому из пп. Е33-Е37 или фармацевтической композиции по п. Е41 в лечении заболевания или состояния, ассоциированного или характеризующегося экспрессией HER2/neu.
Е43. Применение по п. Е42, которое характеризуется тем, что указанное состояние, ассоциированное или характеризующееся экспрессией HER2/neu, представляет собой рак.
Е44. Способ повышения активности агента, нацеленного на опухоль, включающий введение указанного агента, нацеленного на опухоль, в комбинации с CD137×TA связывающей молекулой по любому из пп. Е1-Е23 или фармацевтической композицией по варианту реализации Е24.
Е45. Способ лечения заболевания или состояния, ассоциированного с подавленной иммунной системой или характеризующегося экспрессией опухолевого антигена (ТА), включающий введение субъекту, нуждающемуся в этом, CD137×TA связывающей молекулы по любому из пп. Е1-Е23 или фармацевтической композиции по п. Е24.
Е46. Способ по п.Е45, который характеризуется тем, что указанное состояние, ассоциированное с подавленной иммунной системой или характеризующееся экспрессией опухолевого антигена (ТА), представляет собой рак.
Е47. Способ по п. Е45 или Е46, дополнительно включающий введение агента, нацеленного на опухоль.
Е48. Способ по любому из пп. Е44-Е47, дополнительно включающий введение ингибитора контрольной точки иммунного ответа PD-1/PD-L1.
Е49. Способ по п. Е44 или Е47, который характеризуется тем, что указанный агент, нацеленный на опухоль, представляет собой антитело, эпитопсвязывающий фрагмент антитела или агент, который опосредует перенаправленное уничтожение клетки-мишени Т-клетками.
Е50. Способ по п. Е48 или Е49, который характеризуется тем, что указанный ингибитор контрольной точки иммунного ответа PD-1/PD-L1 представляет собой антитело к PD-1 или антитело к PD-L1.
Е51. Способ по любому из пп. Е48-Е50, который характеризуется тем, что указанный ингибитор контрольной точки иммунного ответа PD-1/PD-L1 содержит домены VH и VL антитела, отобранного из тех, которые перечислены в таблице 7: 


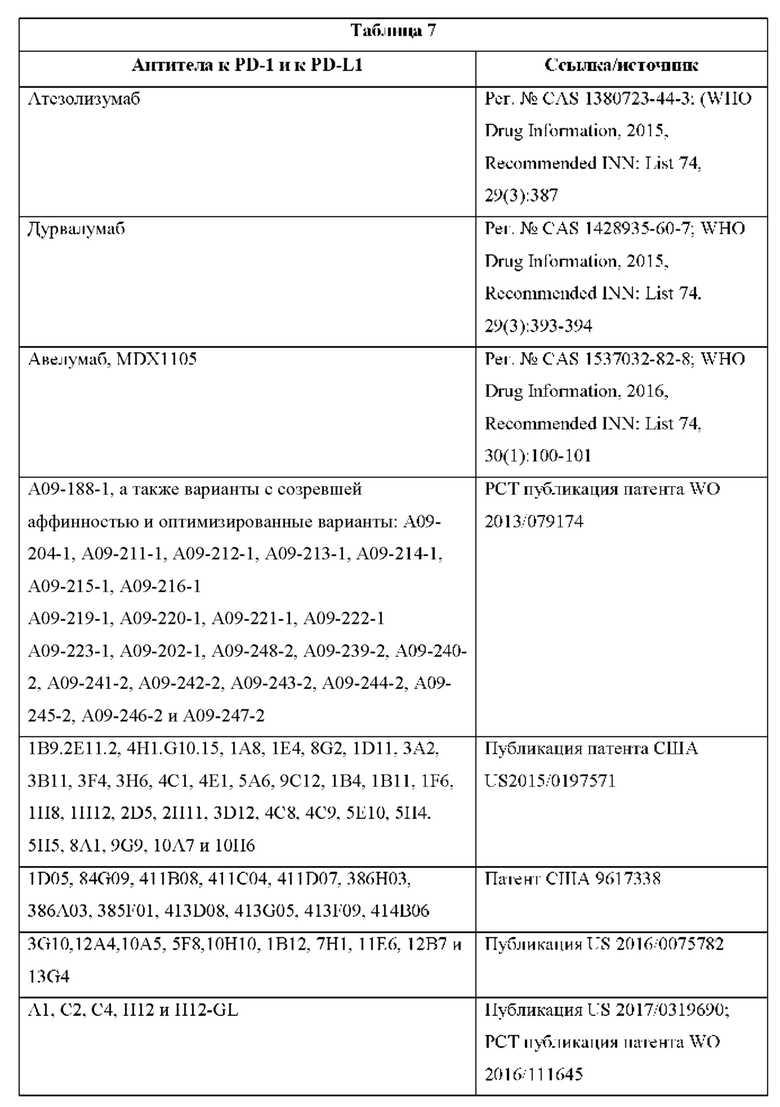

Е52. Применение по пп. Е26, Е35, Е42 или способ по любому из пп. Е46-Е51, которые характеризуются тем, что указанный рак выбран из группы, состоящей из: острого миелолейкоза, опухоли надпочечников, СПИД-ассоциированного рака, альвеолярной саркомы мягких тканей, астроцитарной опухоли, рака мочевого пузыря, рака костей, рака головного и спинного мозга, метастатической опухоли головного мозга, рака молочной железы, опухолей сонных артерий, рака шейки матки, хондросаркомы, хордомы, хромофобной карциномы почек, светлоклеточной карциномы, рака толстой кишки, колоректального рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, экстраскелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии кости, рака желчного пузыря или желчного протока, рака желудка, гестационного трофобластического заболевания, герминогенной опухоли, рака головы и шеи, гепатоцеллюлярной карциномы, глиобластомы, опухоли из островковых клеток, саркомы Капоши, рака почек, лейкоза, липомы/доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легких, медуллобластомы, меланомы, менингиомы, злокачественной мезотелиомы, множественной эндокринной неоплазии, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринных опухолей, немелкоклеточного рака легких, рака яичников, рака поджелудочной железы, рака глотки, папиллярной карциномы щитовидной железы, опухоли околощитовидной железы, педиатрического рака, опухоли оболочек периферических нервов, феохромоцитомы, опухоли гипофиза, рака предстательной железы, постериорной увеальной меланомы, редкого гематологического заболевания, карциномы почек, метастатического рака почек, рабдоидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, рака яичек, рака тимуса, тимомы, метастатического рака щитовидной железы или рака матки
Е53. Применение по пп. Е26, Е35, Е42 или способ по любому из пп. Е46-Е51, которые характеризуются тем, что указанный рак выбран из группы, состоящей из: рака мочевого пузыря, рака молочной железы, колоректального рака, рака желудка, глиобластомы, рака почек, рака легких, меланомы, нейробластомы, рака яичников, рака поджелудочной железы, рака глотки, рака предстательной железы, карциномы почек, рабдомиосаркомы и плоскоклеточного рака головы и шеи (SCCHN).
ПРИМЕРЫ
[000476] Следующие примеры иллюстрируют различные способы для композиций в способах диагностики или лечения согласно настоящему изобретению. Примеры предназначены для иллюстрации и никоим образом не ограничивают объем настоящего изобретения.
Пример 1
Гуманизация МАВ-1 к HER2
[0004771 В РСТ публикации WO 2001/036005 раскрыто антитело к HER2/neu, обозначенное в упомянутом документе «8Н11», однако представленные в нем последовательности доменов VL и VH являются неточными. Антитело 8Н11 связывается с эпитопом HER2/neu, который отличается от эпитопа, связываемого трастузумабом, маргетуксимабом и пертузумабом.
[000478] Были установлены корректные последовательности для доменов VH и VL антитела к HER2/neu, 8Н11 (чтобы обеспечить VH МАВ-1 к HER2 (SEQ ID NO: 60) и VL МАВ-1 к HER2 (SEQ ID NO: 61)), и затем проводили несколько раундов гуманизации для того чтобы получить описанный выше гуманизированный домен VH «VH1 hMAB-1 к HER2» (SEQ ID NO: 64) и гуманизированный домен VL «VL1 hMAB-1 к HER2» (SEQ ID NO: 67). В ходе этих действий в доменах VH1 hMAB-1 к HER2 и VL1 hMAB-1 к HER2 было выявлено несколько возможных недостатков последовательностей, и были сделаны замены, которые устраняли эти недостатки при сохранении аффинности связывания. В частности:
(1) сайт изомеризации аспарагиновой кислоты был выявлен в положении 96 согласно Kabat домена VH1 hMAB-1 к HER2 и был скорректирован с помощью либо замены D96E, либо замены G97I, как в VH2 hMAB-1 к HER2 (SEQ ID NO: 65) и VH3 hMAB-1 к HER2 (SEQ ID NO: 66), соответственно;
(2) возможный сайт дезамидирования аспарагина был выявлен в положении 30 согласно Kabat домена VL1 hMAB-1 к HER2 и был скорректирован с помощью либо замены N30S, либо замены S31T, как в VL3 hMAB-1 к HER2 (SEQ ID NO: 69) и VL2 hMAB-1 к HER2 (SEQ ID NO: 68), соответственно;
(3) возможный сайт изомеризации аспарагиновой кислоты был выявлен в положении D56 согласно Kabat домена VL1 hMAB-1 к HER2 и был скорректирован с помощью замены: V55Q, D56E или D56S, как в VL3 hMAB-1 к HER2 (SEQ ID NO: 69) и VL2 hMAB-1 к HER2 (SEQ ID NO: 68).
[000479] Гуманизированные вариабельные домены тяжелой и легкой цепей hMAB-1 к HER2 можно применять в любой комбинации, и определенные комбинации гуманизированных цепей упоминаются со ссылкой на конкретные домены VH/VL. Например, гуманизированное антитело, содержащее VH1 hMAB-1 к HER2 и VL3 hMAB-1 к HER2, называется в настоящем документе «hMAB-1 к HER2 (1.3)».
[000480] Гуманизированные варианты отбирали так, чтобы получить улучшенную аффинность связывания по сравнению с исходным мышиным антителом, и, как описано, они могут быть дополнительно модифицированы для устранения недостатков последовательностей (например, сайтов изомеризации, дезамидирования), как описано выше. Аффинность связывания нескольких комбинаций гуманизированных доменов VH и VL исследовали с помощью BIACORE®. Соответственно, внеклеточную часть человеческого HER2, гибридизованную с гистидинсодержащим пептидом («huHER2»), пропускали над поверхностью, покрытой иммобилизованным антителом. В общих чертах, каждую испытываемую молекулу захватывали на поверхность, покрытую козьим F(ab)2 к человеческой области Fc, и затем инкубировали в присутствии различных концентраций (6,25 нМ - 50 нМ) huHER2. Кинетику связывания определяли с помощью анализа связывания BIACORE® (нормированная модель связывания Ленгмюра в соотношении 1:1). Рассчитанные ka, kd и KD из этих исследований представлены в таблице 8.

[000481] В отдельном исследовании кинетику связывания CD137×TA биспецифического диатела, DART-В (описанного выше), определяли для связывания с человеческим HER2 и HER2 яванской макаки по существу, как описано выше. Рассчитанные ka, kd и KD из этих исследований представлены в таблице 9.

Пример 2
Выделение и характеристика мышиных моноклональных антител к CD137
[000482] Панель мышиных моноклональных антител, специфичных для человеческого CD137, подвергали скринингу для выявления антител, обладающих высокой аффинностью связывания. Три антитела отбирали для дальнейшего исследования. Для таких оценок очищенную общую популяцию Т-клеток (см. выше) стимулировали гранулами, направленными против CD3/CD28 (клетки:гранулы = 1:1), в течение 72 часов в присутствии ИЛ-2 (100 Ед/мл). По 100 мкл активированных Т-клеток (1,0×106 клеток/мл) и 100 мкл последовательно разведенного (5- или 10-кратные разведения) испытываемого изделия (антитела или биспецифического диатела) добавляли в каждую лунку планшета (ов) для микротитрования, смешивали и инкубировали при комнатной температуре в течение 45 мин. Клетки промывали буфером для FACS и затем в каждую лунку добавляли вторичное антитело (АФЦ-конъюгированное антитело к человеческой области Fc); смешивали и инкубировали при комнатной температуре в течение 30 мин. Затем клетки промывали буфером для FACS, и в каждую лунку добавляли антитела к поверхностным маркерам Т-клеток (антитело к CD4, меченое V510, и антитело к CD8, меченое FITC); смешивали и инкубировали при комнатной температуре в течение 30 мин. Клетки промывали и ресуспендировали в 100 мкл буфера для FACS и анализировали методом проточной цитометрии (BD LSR Fortessa или FACSCanto II) для сбора клеточных событий. Анализ данных проводили с помощью FloJo v10.
[000483] Было обнаружено, что два таких отобранных антитела, МАВ-3 к CD137 и МАВ-4 к CD137, проявляют сильное связывание с человеческим CD137 и способны блокировать связывание между CD137 и его природными лигандами. Третье из таких отобранных антител, МАВ-5 к CD137, проявляло более низкое связывание с человеческим CD137 и не было способно блокировать связывание между CD 137 и его природными лигандами.
[000484] Получали химерные антитела, имеющие области Fc человеческого IgGl и мышиные домены VH/VL (обозначенные: chMAB-1 к CD137, chMAB-2 к CD137, chMAB-3 к CD137, chMAB-4 к CD137 и chMAB-5 к CD137, соответственно), и испытывали их способность связываться с активированными CD8+ Т-клетками. Как показано на Фигуре 7, отобранные антитела проявляли диапазон величин аффинности связывания.
[000485] Биннинг эпитопов выполняли с помощью исследований перекрестной конкуренции. Результаты этих исследований и исследований связывания лигандов свидетельствуют о том, что:
(1) МАВ-2 к CD137, МАВ-3 к CD137 и МАВ-4 к CD137 конкурируют с CD137L за связывание с CD137, однако МАВ-2 к CD137 распознает эпитоп CD137, который отличается от эпитопа, распознаваемого МАВ-1 к CD137, МАВ-3 к CD137, МАВ-4 к CD137 и МАВ-5 к CD137;
(2) МАВ-1 к CD137 и МАВ-5 к CD137 не конкурируют с CD137L за связывание с CD137;
(3) МАВ-3 к CD137 и МАВ-4 к CD137 проявляют конкуренцию с МАВ-1 к CD137, но связывают другой эпитоп, о чем свидетельствует их способностью конкурировать за связывание лиганда;
(4) МАВ-5 к CD137 распознает эпитоп, который отличается от эпитопов, распознаваемых МАВ-1 к CD137, МАВ-2 к CD137, МАВ-3 к CD137 или МАВ-4 к CD137.
[000486] Также испытывали способность химерных антител индуцировать высвобождение цитокинов (например, ИФН-γ, TNF-α) из Т-клеток (т.е. их агонистическую активность) в отсутствие лиганда. Для таких оценок очищенную общую популяцию Т-клеток (см. выше) ресуспендировали в средах для анализа и помещали в инкубатор для тканевых культур на ночь. Опухолевые клетки-мишени (включая ЛМТ-1, N87) получали из культуры. После промывки клеточную массу ресуспендировали в культуральных средах при плотности клеток 1,0×105 клеток/мл. По 104 клеток затем добавляли в каждую лунку белого планшета с плоским дном, помещенного в инкубатор для тканевых культур на ночь. На следующий день измеряли плотность и жизнеспособность общей популяции покоящихся человеческих Т-клеток на основании исключения трипанового синего с использованием счетчика Beckman Coulter Vi-Cell и доводили до плотности 2×106 клеток/мл. Опухолевые клетки-мишени, предварительно высеянные на планшет, промывали один раз средами для анализа.
[000487] Для анализов высвобождения цитокинов: 50 мкл последовательно разведенного испытываемого изделия (антитела (+/- сшивание с (Fab)'2 к человеческой Fc), диатела, трехвалентные молекулы и т.д.), 50 мкл предварительно промытых Dynabead αCD3 (REF 11151D; Invitrogen от Thermo Fisher Scientific) в количестве 2×106 гранул/мл, 50 мкл/лунку общей популяции человеческих Т-клеток в количестве 2×106 клеток/мл и 50 мкл сред для анализа по отдельности или содержащих 1,6 мкг/мл αCD28 (каталожный номер 555725; BD Pharmigen) добавляли в каждую лунку аналитического планшета. Конечный объем в каждой лунке в планшете составлял 200 мкл. Для тех контрольных лунок, которые не содержали испытываемое изделие или гранулы αCD3, добавляли среды для анализа, чтобы довести общий объем до 200 мкл, и планшеты инкубировали в течение 72 часов в инкубаторе для тканевых культур. Супернатанты затем собирали из каждой лунки и измеряли высвобожденные цитокинов ИФН-γ, TNF-α и ИЛ-2R с использованием набора для ИФА цитокинов от R&D System (Human IL-2 DuoSet ELISA (каталожный номер: DY202), Human IFN-gamma DuoSet ELISA (каталожный номер: DY285) и Human TNF-alpha DuoSet ELISA (каталожный номер: DY210) или аналогичных коммерческих реагентов. Способы проведения ИФА были предоставлены с наборами. Для анализа данных использовали Microsoft Excel и SoftMax Pro, чтобы экстраполировать уровни цитокинов, которые были представлены в графической форме с помощью Prism.
[000488] Как показано на Фигуре 8, каждое из МАВ-1 к CD137, chMAB-3 к CD137 и chMAB-4 к CD137 проявляло сильную агонистическую активность, МАВ-2 к CD137 проявляло меньшую агонистическую активность, и МАВ-5 к CD137 не проявляло агонистическую активность в отсутствие лиганда.
[000489] Также испытывали способность химерных антител chMAB-3 к CD137 и chMAB-4 к CD137 индуцировать высвобождение цитокинов (например, ИФН-γ, TNF-α) из общей популяции Т-клеток в отсутствие и в присутствии лиганда и/или клеток-мишеней. Общую популяцию Т-клеток человека очищали из донорских МКПК с использованием набора Dynabeads Untouched Human T Cells (Invitrogen, каталожный номер 11344D) в соответствии с протоколом изготовителя. Результаты исследования высвобождения ИФН-γ представлены на Фигуре 9. На Фигуре 9 показана индукция ИФН-γ общей популяцией Т-клеток через 72 часа после стимуляции гранулами, покрытыми антителом к CD3, в присутствии или в отсутствие 1 мкг/мл химерных антител к CD137 (chMAB-3 к CD137 или chMAB-5 к CD137) и в присутствии или в отсутствие 1 мкг/мл CD-137L-His и 4 мкг/мл hFc F(ab)'2. На Фигуре показано, что индукция ИФН-γ происходила в присутствии или в отсутствие CD137-His, и что большую индукцию наблюдали для ЛМТ-1/общей популяции Т-клеток по сравнению с той, которая наблюдалась только для общей популяции Т-клеток. Аналогичные результаты получали для TNF-α.
[000490] В присутствии как Т-клеток, так и клеток-мишеней, или только Т-клеток, chMAB-3 к CD137 проявляло сильную агонистическую активность, которая была сопоставимой в присутствии или в отсутствие лиганда. Напротив, chMAB-5 к CD137 проявляло минимальную агонистическую активность в отсутствие лиганда и значительно повышенную агонистическую активность в присутствии лиганда.
Пример 3
Гуманизация мышиных моноклональных антител к CD137
[000491] Антитела МАВ-3 к CD137 и МАВ-4 к CD137 отбирали для гуманизации. Проводили четыре раунда гуманизации МАВ-3 к CD137 с получением одного гуманизированного домена VH, обозначенного «VH1 hMAB-3 к CD137» (SEQ ID NO: 76), и трех гуманизированных доменов VL, обозначенных «VL1 hMAB-3 к CD137» (SEQ ID NO: 87), «VL2 hMAB-3 к CD137»(SEQ ID NO: 88) и «VL3 hMAB-3 к CD137» (SEQ ID NO: 89).
[000492] Проводили три раунда гуманизации МАВ-4 к CD137 с получением одного гуманизированного домена VH, обозначенного «VH1 hMAB-4 к CD137» (SEQ ID NO: 92), и двух гуманизированных доменов VL, обозначенных «VL1 hMAB-4 к CD137» (SEQ ID NO: 94) и «VL2 hMAB-4 к CD137» (SEQ ID NO: 95).
[000493] Гуманизированные вариабельные домены тяжелой и легкой цепей конкретного антитела к CD137 (например, МАВ-3 к CD137) можно применять в любой комбинации, и определенные комбинации гуманизированных цепей упоминаются со ссылкой на конкретные домены VH/VL, например, гуманизированное антитело, содержащее VH1 hMAB-3 к CD137 и VL3 hMAB-3 к CD137, называется в настоящем документе «hMAB-3 к CD137 (1.3)».
[000494] Величины аффинности связывания химерных антител, имеющих мышиные вариабельные домены, и полностью гуманизированных антител исследовали с помощью Biacore.
[000495] Соответственно, внеклеточную часть человеческого CD137, гибридизованную с гистидинс о держащим пептидом («huCD137»), пропускали над поверхностью, покрытой иммобилизованным антителом. В общих чертах, каждую испытываемую молекулу захватывали на поверхность, покрытую козьим F(ab)2 к человеческой Fc, и затем инкубировали в присутствии различных концентраций (25 нМ и 100 нМ) huCD137. Кинетику связывания определяли с помощью анализа связывания BIACORE® (нормированная модель связывания Ленгмюра в соотношении 1:1). Рассчитанные ka, kd и KD из этих исследований представлены в таблице 10 и свидетельствуют о том, что гуманизированные MAT проявляли приблизительно 2-кратное снижение аффинности связывания по сравнению с мышиными антителами.

Пример 4
Характеристика CD137×TA связывающих молекул
[000496] Как описано выше, получали ряд примерных CD137×TA связывающих молекул, способных связываться с CD 137 и примерным ТА, HER2/neu. В частности, получали шесть биспецифических диател (обозначенных «DART-A», «DART-B» и «DART-C», «DART-D», «DART-E» и «DART-G»), содержащих домены МАВ-3 к CD137 или hMAB-3 к CD137, одно биспецифическое диатело (обозначенное «DART-F»), содержащее домены МАВ-4 к CD137, два биспецифических диатела (обозначенных «DART-1» и «DART-4»), содержащих домены МАВ-1 к CD1370, и два биспецифических диатела (обозначенных «DART-2» и «DART-5»), содержащих домены МАВ-2 к CD137, каждое из которых содержало домены hMAB-1 к HER2. DART-A, DART-B, DART-C, DART-1 и DART-2 имеют общую структуру, показанную на Фигуре 3 В (DART-А, DART-B, DART-1 и DART-2 имеют структуру, показанную на Фигуре 3Е; DART-C имеет структуру, показанную на Фигуре 3D). DART-D, DART-E, DART-F, DART-G, DART-4 и DART-5 имеют общую структуру, показанную на Фигуре 5А (DART-D, DART-F, DART-G, DART-4 и DART-5 имеют структуру, показанную на Фигуре 5В; DART-E имеет структуру, показанную на Фигуре 5С).
[000497] Получали другую примерную CD137×TA связывающую молекулу, способную связываться с CD137 и примерным ТА, EphA2 (обозначена «DART-7»). DART-7 представляет собой биспецифическое диатело, имеющее структуру, представленную на Фигуре 3 В, содержащее МАВ-1 к CD137 и гуманизированные домены МАВ-3 к EphA2.
[000498] Помимо этого получали три контрольные молекулы, «DART-3», содержащую hMAB-1 к HER2 и варианты доменов паливизумаба (HER2×RSV), «DART-6», содержащую варианты доменов паливизумаба и домены МАВ-3 к CD137 (RSV×CD137), и «DART-8», содержащую гуманизированное МАВ-3 к EphA2 и варианты доменов паливизумаба (EphA2×RSV).
[000499] CD137×TA связывающие молекулы, имеющие два сайта связывания для CD137 и 2 сайта связывания для HER2/neu, и контрольная молекула CD137×RSV (DART-6) связывались с CD137, присутствующим на поверхности активированных Т-клеток. Напротив, контрольная молекула HER2/neu×RSV (DART-3) не проявляла какое-либо связывание. В этом анализе DART-1 проявляла самое высокое связывание, и DART-5 проявляла самое низкое связывание, при этом остальные испытанные молекулы (DART-В, DART-C, DART-D, DART-E, DART-F, DART-2, DART-4 и DART-6) проявляли сопоставимое связывание (Фигура 10). Относительные положения доменов, связывающих HER2/neu и CD137, оказывали минимальное влияние на связывание CD137 или не оказывали влияния в этом анализе для молекул, имеющих структуру, представленную на Фигуре 3 В (сравните DART-B/DART-C), или структуру, представленную на Фигуре 5В-5С (сравните DART-D/DART-E).
[000500] Аналогичным образом, оценивали способность CD137×TA связывающих молекул связываться с HER2/neu, присутствующим на поверхности клеток-мишеней рака желудка N87 (Фигуры 11А-11В). Контрольная молекула RSV×CD137 (DART-6) не проявляла какое-либо связывание. Относительные положения доменов, связывающих HER2/neu и CD137, оказывали минимальное влияние на связывание HER2/neu или не оказывали влияния в этом анализе для молекул, имеющих структуру, представленную на Фигуре 3 В (сравните DART-B/DART-C). Напротив, DART-E проявляла уменьшенное связывание HER2/neu по сравнению с DART-D, это указывает на возможность существования позиционного эффекта на связывание HER2/neu для молекул, имеющих структуру, представленную на Фигуре 5В-5С, или на то, что связывание, опосредуемое конкретно HER2/neu-связывающими доменами hMAB-1 к HER2 (1.3), проявляет позиционный эффект.
[000501] Дополнительно оценивали способность CD137×TA связывающих молекул, DART-В, DART-D, DART-G и контрольных молекул DART-3 и DART-6 связываться с CD137, экспрессируемым клетками СНО (Фигура 12А) и активированными Т-клетками (Фигура 12В).
[000502] Дополнительно оценивали способность CD137×TA связывающих молекул, DART-В, DART-D, DART-G и контрольных молекул DART-3 и DART-6 связываться с HER2/neu, экспрессируемым на клетках N87 (Фигура 13А) и клетках JIMT-1 (Фигура 13В). Для такой оценки в каждую лунку аналитического (их) планшета (ов) для микротитрования добавляли по 100 мкл клеток-мишеней, экспрессирующих HER2/neu (1,0×106 клеток/мл), и 100 мкл последовательно разведенного (5- или 10-кратные разведения) испытываемого изделия (биспецифическое диатело), смешивали и инкубировали при комнатной температуре в течение 45 мин. Клетки промывали буфером для FACS и затем в каждую лунку добавляли вторичное антитело (АФЦ-конъюгированное антитело к человеческой области Fc); смешивали и инкубировали при комнатной температуре в течение 30 мин. Клетки промывали и ресуспендировали в 100 мкл буфера для FACS и анализировали методом проточной цитометрии (BD LSR Fortessa или FACSCanto II) для сбора клеточных событий. Анализ данных выполняли с помощью FloJo v10. HER2×CD137 связывающие молекулы, имеющие структуру, представленную на Фигуре 3 В, проявляли незначительно лучшее связывание с HER2/neu, чем молекулы, имеющие структуру, представленную на Фигуре 5А-5С (сравните DART-B/DART-D). Контрольная молекула RSV×CD137 (DART-6) не проявляла какое-либо связывание. CD137×TA биспецифические диатела, имеющие мышиный или гуманизированный сайт связывания CD 137, проявляли сходное связывание с HER2/neu (сравните DART-D/DART-G).
[000503] На Фигурах 14А-14В представлены результаты первого типичного анализа способности CD137×TA связывающих молекул опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (выполненного по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ) в присутствии или в отсутствие клеток-мишеней, экспрессирующих примерный ТА, HER2/neu. На Фигурах показаны результаты исследований DART-1, DART-2, DART-3, DART-4, DART-5, DART-6, DART-A, DART-D, DART-E и DART-F (с использованием описанных выше протоколов) в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (3+) (Фигура 14А) или JIMT-1 (2+) (Фигура 14В), HER2/neu-отрицательных клеток-мишеней Hs700T или без клеток-мишеней. Результаты показывают, что контрольные молекулы DART-3 и DART-6 не проявляли костимулирующую активность. CD137×TA связывающие молекулы не проявляли какую-либо наблюдаемую костимулирующую активность в отсутствие клеток-мишеней или с HER2/neu-отрицательными клетками-мишенями. DART-D и DART-F, содержащие новые антитела к CD137 (МАВ-3 и МАВ-4 к CD137, соответственно), проявляли самую высокую костимулирующую активность в присутствии любой линии клеток-мишеней. DART-E (содержащая замененные домены HER2/neu и CD137) проявляла более низкую костимулирующую активность, в частности, в присутствии клеток-мишеней N87. Сходные закономерности наблюдали для ИЛ-2 и TNF-α.
[000504] На Фигурах 15А-15В представлены результаты второго типичного анализа способности CD137×TA связывающих молекул опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (выполненном по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ). На Фигурах показаны результаты испытаний DART-1, DART-2, DART-3, DART-4, DART-5, DART-6, DART-B, DART-D и DART-G (с использованием описанных выше протоколов) в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (Фигура 15А) или JIMT-1 (Фигура 15В), HER2/neu-отрицательных клеток-мишеней Hs700T или без клеток-мишеней.
[000505] Результаты показывают, что контрольные молекулы DART-3 и DART-6 не проявляли костимулирующую активность. CD137×TA связывающие молекулы не проявляли какую-либо наблюдаемую костимулирующую активность в отсутствие клеток-мишеней или с HER2/neu-отрицательными клетками-мишенями. DART-D и DART-G, содержащие новое антитело к CD137, МАВ-3 к CD137 (мышиное и гуманизированное, соответственно), проявляли самую высокую костимулирующую активность в присутствии любой линии клеток-мишеней. DART-G проявляла самую высокую костимулирующую активность, несмотря на то, что она проявляла незначительно более низкое связывание с поверхностью клеток, экспрессирующих CD137. Как подробно описано выше, DART-B и DART-G содержат идентичные сайты связывания HER2 и CD137, но имеют структуры, показанные на Фигуре 3В и Фигуре 5В, соответственно. DART-G проявляла более высокую костимулирующую активность, чем DART-B, в присутствии любой линии клеток-мишеней.
[000506] Дополнительно оценивали способность CD137×TA связывающей молекулы, DART-G, и контрольных молекул, DART-3 и DART-6, опосредовать дозозависимую передачу сигнала Т-клетками с участием пути NF/κВ в репортерной клеточной линии, экспрессирующей CD137 (Jurkat-NF-κB-Luc), в присутствии клеток-мишеней, положительных или отрицательных в отношении примерного ТА, HER2/neu. В общих чертах, репортерные клетки Jurkat-NF-κB-Luc, сверхэкспрессирующие CD137, культивировали совместно с клетками JIMT-1, экспрессирующими HER2/neu (Фигура 16А), или с отрицательными по HER2/neu клетками KG-1 (Фигура 16В) в присутствии повышающихся концентраций DART-G, DART-3 или DART-6. После инкубации передачу сигнала измеряли на основании люминесценции, используя относительную световую единицу люминесценции (RLU) в качестве регистрируемого показателя. Результаты показывают, что CD137×TA связывающие молекулы, такие как DART-G, опосредуют дозозависимую передачу сигнала Т-клетками, которая зависит от присутствия ТА-экспрессирующих клеток-мишеней.
[000507] Дополнительно оценивали способность CD137×TA связывающей молекулы, DART-А, и контрольных молекул, DART-3 и DART-6, опосредовать повышенную пролиферацию Т-клеток в присутствии клеток, экспрессирующих различные уровни примерного ТА, HER2/Neu. В общих чертах, CFSE-меченые человеческие Т-клетки ± субоптимальная стимуляция αCD3/αCD28 культивировали совместно с клетками-мишенями HER2/neu-high N87 (Фигура 17, панели А-С и J-K), клетками-мишенями HER2/neu-low MCF-7 (Фигура 17, панели D-F и L-M) или культивировали по отдельности (Фигура 17, панели G-I и N-O) в присутствии DART-А (Фигура 17, панели A, D и G), DART-3 (Фигура 17, В, Е и Н), DART-6 (Фигура 17, панели С, F и I) или без молекул (Фигура 17, панели J, L и N) и контролировали пролиферацию Т-клеток. Результаты показывают, что CD137×TA связывающие молекулы, такие как DART-А, повышают пролиферацию Т-клеток. Такая повышенная пролиферация Т-клеток зависит от экспрессии ТА и коррелирует с уровнями экспрессии ТА.
[000508] Маргетуксимаб представляет собой Fc-оптимизированное моноклональное антитело к HER2/neu, которое может индуцировать повышенную антителозависимую клеточную цитотоксичность после связывания опухолевой клетки-мишени. Как отмечено выше, МАВ-1 к HER2 связывается с эпитопом HER2/neu, который отличается от того, который связывается маргетуксимабом. Активность антитела к ТА, маргетуксимаба, направленную на уничтожение клеток-мишеней, исследовали в комбинации с CD137×TA связывающей молекулой, DART-1, или контрольной молекулой, DART-3 (HER2×RSV). В общих чертах, клетки-мишени N87, которые были модифицированы для экспрессии репортерного гена люциферазы (luc) (клетки N87/GFP/Luc), инкубировали с очищенными эффекторными NK-клетками при соотношении эффектор : мишень (Е : Т) 2:1 с повышающимися концентрациями маргетуксимаба в комбинации с фиксированной концентрацией (0,1 мкг/мл) DART-1 или DART-3 в течение 72 часов, и цитотоксичность определяли с помощью люминесцентного (LUM) анализа, в котором измеряли клеточную активность люциферазы клеток-мишеней, используя относительную световую единицу люминесценции (RLU) в качестве регистрируемого показателя (Фигура 18А). Помимо этого оценивали экспрессию маркера активации, CD69, NK-клетками (CD3-/CD56+) из этого анализа (Фигура 18В). Результаты этого исследования демонстрируют, что CD137×TA связывающая молекула, DART-1, повышает как опосредуемую маргитуксимабом активность АЗКЦ против клеток-мишеней N87/Luc, так и опосредуемую маргитуксимабом активацию CD69 на NK-клетках по сравнению с контрольной молекулой, способной связываться только с HER2/neu.
[000509] Исследовали способность CD137×TA связывающей молекулы, обозначенной «DART-7», которая связывает иллюстративный ТА, EphA2, опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (выполненном по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ). CD137×TA связывающую молекулу DART-7 и контрольную молекулу DART-8 испытывали (используя описанные выше протоколы) в присутствии EphA2-экспрессирующих клеток-мишеней Hs700T (Фигура 19А) или EphA2-отрицательных клеток-мишеней Hs700T (EphA2.KO) (Фигура 19В), или без клеток-мишеней. Результаты показывают, что CD137×TA связывающая молекула DART-7 проявляла костимулирующую активность в присутствии EphA2-экспрессирующих клеток, но не проявляла какую-либо наблюдаемую костимулирующую активность в отсутствие клеток-мишеней или с EphA2-отрицательными клетками-мишенями. Контрольная молекула DART-8 не проявляла костимулирующую активность.
[000510] Костимулирующую активность CD137×TA связывающих молекул на популяциях Т-клеток исследовали, оценивая долю Т-клеток центральной памяти (Tcm) и Т-клеток эффекторной памяти (Tem). В общих чертах, человеческие Т-клетки культивировали совместно с EphA2-положительными клетками-мишенями аденокарциномы толстой кишки Соlо205 в течение 5 дней при субоптимальных условиях стимуляции в присутствии CD137×TA связывающей молекулы DART-7 или контрольных молекул DART-8 или DART-6. После совместного культивирования процент клеток Tcm (CCR7+CD45RA-) и Tem (CCR7-CD45RA-) определяли в селективно пропущенных подгруппах CD4+ и CD8+. Результаты, обобщенные в таблице 11, показывают, что наблюдается существенное увеличение доли CD8+ Tcm и типов Tem-клеток и что это увеличение требует связывания как CD137, так и ТА (например, EphA2). Эти результаты указывают на то, что CD137×TA связывающие молекулы индуцируют существенное увеличение доли клеток CD8+ Tcm и Tem в присутствии соответствующих клеток, экспрессирующих опухолевый антиген. Нацеленный Т-клеточный агонизм, проявляемый CD137×TA связывающими молекулами, может позволить индуцировать активацию CD137, заякоренного на опухолевых клетках, при этом ограничивая системную активацию иммунных клеток и связанные побочные эффекты.
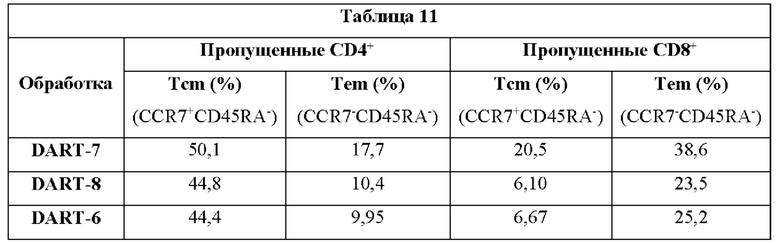
[000511] Помимо этого конструировали и испытывали CD137×TA связывающие молекулы, имеющие один сайт связывания для CD137 и один или более сайтов связывания для HER2/neu (см., например. Фигуру 4А, 5А и 6А). Такие молекулы являются одновалентными для CD137 и проявляют уменьшенное связывание с активированными Т-клетками, как и ожидалось, учитывая их уменьшенную авидность. Такие молекулы проявили различное связывание с клетками-мишенями N87. Однако ни одна из одновалентных для CD137 молекул не проявляла какую-либо костимулирующую активность в анализах высвобождения цитокинов Т-клетками. Это наблюдение указывает на то, что для ко стимулирующей активности в этих анализах могут потребоваться по меньшей мере два сайта связывания CD137.
Пример 5
Оптимизация гуманизированного МАВ-3 к CD137
[000512] Как отмечено выше, гуманизация МАВ-3 к CD137 привела приблизительно к 2-кратной потере аффинности связывания (см., например, таблицу 9 выше). Оптимизацию применяли для выявления клонов гуманизированного МАВ-3 к CD137, имеющих сопоставимую или лучшую аффинность связывания, чем исходное мышиное антитело. В общих чертах, случайный мутагенез использовали для введения замен в домены CDRH3 тяжелой цепи (положения 99-102 согласно Kabat) hMAB-3 к CD137 (1.3). Чтобы выявить клоны, имеющие повышенное связывание с CD137, применяли три раунда селекционного скрининга с повышением жесткости условий. 48 клонов отбирали в раундах 2 и 3 и оценивали их аффинность в качестве диател (имеющих структуру, показанную на Фигуре 5В). В таблице 12 представлено выравнивание аминокислотной последовательности остатков 99-102 согласно Kabat CDRH3 из четырех клонов, отобранных на основании повышенного связывания с hCD137.

[000513] Аминокислотные последовательности этих клонов включали в 5-цепочечное диатело DART-G для получения оптимизированных молекул DART-G (обозначенных DART-G1, DART-G2; DART-G3 и DART-G4). Аффинность связывания DART-G2, DART-G3 и DART-G4 оценивали с помощью BIAcore (таблица 11). Для такой оценки His-меченый растворимый человеческий CD137 (huCD137) или CD137 яванской макаки (cynoCD137) (содержащий внеклеточную часть CD137 человека или яванской макаки, гибридизованную с гистидинсодержащим пептидом) пропускали над поверхностью, покрытой иммобилизованным антителом. В общих чертах, каждую испытываемую молекулу захватывали на поверхность, покрытую козьим F(ab)2 к человеческой Fc, и затем инкубировали в присутствии различных концентраций (6,25-100 нМ) huCD137 или cynoCD137. Кинетику связывания определяли с помощью анализа связывания BIACORE® (нормированная модель связывания Ленгмюра в соотношении 1:1). Рассчитанные ka, kd и KD из этих исследований представлены в таблице 13.

[000514] Способность диател связывать CD137 оценивали, как описано выше. Результаты этого исследования представлены на Фигурах 20А-20В. Как отмечено выше, DART-G, имеющая сайты связывания hMAB-3 к CD137 (1.3) для CD137, проявляла незначительно более низкое связывание по сравнению с конструкциями, имеющими такую же структуру, но содержащими сайты связывания CD137 мышиного МАВ-3 к CD137 (DART-6). HER2×CD137 диатела, содержащие оптимизированные домены VH hMAB-3 к CD137, проявляли более высокое связывание, при этом относительная активность связывания молекул с человеческим CD137 была следующей: DART-G4 > DART-G3 > DART-G2 ≈ DART-6 > DART-G. DART-G и оптимизированные варианты проявляли практически идентичное связывание с HER2 на поверхности множества типов клеток-мишеней, включая клетки N87, JIMT-1 и MDA-MB231.
[000515] На Фигурах 21А-21С показаны результаты типичного анализа способности DART-3, DART-6, DART-B, DART-D, DART-G, DART-G2, DART-G3 и DART-G4 опосредовать ко стимулирующую активность в анализе высвобождения цитокинов Т-клетками (выполненном по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ). Высвобождение цитокинов измеряли, как описано выше, в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (Фигура 21А), клеток-мишеней JIMT-1 (Фигура 21В) или клеток-мишеней MDA-231 (Фигура 21C), или без клеток-мишеней.
[000516] Результаты показывают, что контрольные диатела DART-3 и DART-6 не проявляли костимулирующую активность. CD137×TA связывающие молекулы не проявляли какую-либо наблюдаемую костимулирующую активность в отсутствие клеток-мишеней или с HER2/neu-отрицательными клетками-мишенями. DART-G, содержащая новое гуманизированное антитело к CD137, hMAB-3 к CD137, проявляла самую высокую костимулирующую активность в присутствии любой клеточной линии-мишени. TNF-α и ИЛ-2 показали сходные профили высвобождения.
[000517] Было обнаружено, что костимулирующая активность оптимизированных вариантов DART-G2, DART-G3 и DART-G4 обратно пропорциональна повышенному связыванию с CD137, экспрессированным на поверхности клеток, это указывает на то, что более высокая аффинность связывания, по-видимому, не коррелирует непосредственно с костимулирующей активностью в этом анализе. Однако, как показано ниже, оптимизированные варианты имеют сопоставимую активность в других функциональных анализах. Эти характерные признаки делают оптимизированные по аффинности варианты особенно подходящими для детектирования экспрессии CD137 и для диагностических анализов, в которых измеряют экспрессию CD137, в дополнение к их применению в качестве костимулирующих агентов.
[000518] Исследовали способность CD137×TA связывающих молекул опосредовать перенаправленное уничтожение опухолевых клеток-мишеней клетками. В общих чертах, клетки-мишени N87/GFP/Luc (модифицированные для экспрессии репортерного гена люциферазы (luc)) инкубировали с общей популяцией Т-клеток в условиях субоптимальной стимуляции в течение 72 часов в отсутствие или в присутствии DART-В, DART-G, DART-G1, DART-G2, DART-G3, DART-G4 или контрольных молекул DART-3 или DART-6 (каждая по 0,001, 0,01, 0,1 и 1 мкг/мл) по существу, как описано выше для анализов костимуляции. В конце инкубации цитотоксичность определяли с помощью люминесцентного (LUM) анализа, в котором измеряли клеточную активность люциферазы клеток-мишеней, используя относительную световую единицу люминесценции (RLU) в качестве регистрируемого показателя (Фигура 22). Каждая из исследованных CD137×TA связывающих молекул проявляла некоторую степень активности в отношении перенаправленного уничтожения клеток, причем DART-G проявляла наибольшую активность, и DART-B проявляла наименьшую активность. Напротив, контрольные молекулы не проявляли детектируемую активность в отношении перенаправленного уничтожения клеток. Результаты этого исследования демонстрируют, что CD137×TA связывающие молекулы способны опосредовать перенаправленное уничтожение опухолевых клеток.
Пример 6
Характеристика трехвалентных CD137×TA связывающих молекул
[000519] Как отмечено выше, CD137×TA связывающие молекулы, имеющие один сайт связывания для CD137 и один или более сайтов связывания для HER2/neu, не проявляли какую-либо костимулирующую активность в анализах высвобождения цитокинов Т-клетками. Это наблюдение указывает на то, что для костимулирующей активности могут потребоваться по меньшей мере два сайта связывания CD137. Чтобы ответить на этот вопрос, получали набор примерных трехвалентных CD137×TA связывающих молекул, способных связываться с CD137 и с примерным ТА, HER2/neu. Эти молекулы включали четыре биспецифические трехвалентные связывающие молекулы «TRIDENT-A», «TRIDENT-A2», «TRIDENT-A3» и «TRIDENT-A4», соответственно, каждая из которых содержит два домена hMAB-3 к CD137 и один домен hMAB-1 к HER2 (CD137×CD137×HER2). Структура и последовательности этих примерных CD137×TA связывающих молекул подробно представлены выше. Помимо этого получали две контрольные молекулы «TRIDENT-1», содержащую один вариант домена паливизумаба и два оптимизированных домена hMAB-3 к CD137 (CD137×CD137×RSV), и «TRIDENT-2», содержащую один домен hMAB-1 к HER2, один домен 4-4-20 и один вариант домена паливизумаба (RSV×FITC×HER2). Трехвалентные молекулы имеют общую структуру, показанную на Фигуре 6А, и были получены и охарактеризованы, как подробно описано ниже, для определения их способности связываться с HER2/neu, присутствующим на поверхности мишени N87(3+), JIMT-1(2+) и MCF-7(1+) с помощью анализа FACS, по существу, как описано выше. Также оценивали CD137×TA связывающие молекулы, имеющие два сайта связывания для CD137 и два сайта связывания для HER2/neu (DART-G, DART-G2, DART-G3 и DART-G4).
[000520] Как показано на Фигуре 23А-23С, молекулы, которые содержат один или два HER2/neu-связывающих домена, связывались со всеми тремя типами клеток-мишеней. Следует отметить, что кривые связывания молекул, имеющих два сайта связывания HER2/neu (DART-G, DART-G2, DART-G3 и DART-G4), быстрее достигают насыщения, в частности, на клетках, имеющих высокие уровни HER2/neu на клеточной поверхности. Это указывает на то, что некоторые молекулы проявляют двухвалентное связывание (т.е. связывание двух молекул HER2/neu на поверхности). Двухвалентное связывание более вероятно в присутствии высоких концентраций лиганда-мишени.
[000521] CD137×TA связывающие молекулы, имеющие два сайта связывания для CD 137 и один сайт связывания для HER2/neu (TRIDENT-A, TRIDENT-A2, TRIDENT-A3 и TRIDENT-A4), контрольные молекулы CD137×CD137×RSV и TA×FITC×RSV (TRIDENT-1 и TRIDENT-2) и CD137×TA связывающие молекулы, имеющие два сайта связывания для CD137 и 2 сайта связывания для HER2/neu (DART-G, DART-G2, DART-G3 и DART-G4), также оценивали с помощью анализа FACS по существу, как описано в примере 2 выше, для определения их способности связывать CD137, присутствующий на поверхности активированных CD4+ (Фигура 24А) и CD8+ (Фигура 24В) Т-клеток. Молекулы, которые содержали два CD137-связывающих домена, проявляли сильное связывание с активированными Т-клетками. Улучшенное связывание наблюдали для примерных HER2×CD137 диател, содержащих оптимизированные домены VH hMAB-3 к CD137, описанные выше. Как и ожидалось, контрольная молекула TA×FITC×RSV (TRIDENT-2) не проявляла какое-либо связывание с активированными Т-клетками.
[000522] В другом исследовании оценивали способность CD137×TA связывающих молекул, имеющих два сайта связывания для CD137 и один сайт связывания для HER2/neu (TRIDENT-A, TRIDENT-A2, TRIDENT-A3 и TRIDENT-A4), CD137×TA связывающей молекулы, имеющей два сайта связывания для CD137 и два сайта связывания для HER2/neu (DART-G4), и контрольной молекулы TA×FITC×RSV (TRIDENT-2) опосредовать конъюгацию клетка-клетка между клетками, экспрессирующими CD137, и клетками-мишенями, экспрессирующими HER2/neu. Для такой оценки 1,5×104 CD137-экспрессирующих клеток СНО, меченых РKН26, инкубировали совместно с 1,5×104 HER2/neu-экспрессирующих клеток-мишеней (N87 (уровень экспрессии HER2: 3+), JIMT-1 (уровень экспрессии HER2: 2+) или MCF-7 (уровень экспрессии HER2: 1+), меченых CFSE, в соотношении 1:1 в присутствии последовательно разведенного (10-кратные разведения) испытываемого изделия в течение 30 мин. при комнатной температуре. Затем образцы анализировали методом проточной цитометрии, чтобы определить процент конъюгированных клеток (CFSE+/PKH26+) в качестве регистрируемого показателя для конъюгации клетка-клетка. Как показано на Фигуре 25А-25С, молекулы, которые содержали по меньшей мере один HER2/neu-связывающий домен и два CD137-связывающих домена, могли опосредовать конъюгацию клетка-клетка, в то время как контрольная молекула была неактивной. Для всех активных молекул активность конъюгации клетка-клетка коррелировала с уровнем экспрессии HER2/neu.
[000523] В другом исследовании оценивали способность CD137×TA связывающих молекул, имеющих два сайта связывания для CD137 и один сайт связывания для HER2/neu (TRIDENT-A, TRIDENT-A2, TRIDENT-A3 и TRIDENT-A4), контрольных молекул CD137×RSV и TA×FITC×RSV (TRIDENT-1 и TRIDENT-2) и CD137×TA связывающих молекул, имеющих два сайта связывания для CD137 и два сайта связывания для HER2/neu (DART-G, DART-G2, DART-G3 и DART-G4), опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками (выполненном по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ) в присутствии или в отсутствие клеток-мишеней, экспрессирующих примерный ТА, HER2/neu. Высвобождение цитокинов измеряли, как описано выше, в присутствии HER2/neu-экспрессирующих клеток-мишеней N87 (Фигура 26А), клеток-мишеней JIMT-1 (Фигура 26В) или без клеток-мишеней (Фигура 26С).
[000524] Результаты показывают, что примерные CD137×TA связывающие молекулы проявляют костимулирующую активность в присутствии любой линии клеток-мишеней, причем активность коррелирует с уровнем экспрессии HER2/neu. TRIDENT-A, TRIDENT-A2, TRIDENT-A3 и TRIDENT-A4 (каждая из которых имеет два сайта связывания для CD137 и один сайт связывания для HER2/neu) проявляли более высокую костимулирующую активность в отношении клеток JIMT-1 (HER2++), чем DART-G, DART-G2, DART-G3 или DART-G4 (с двумя сайтами связывания для CD137 и 2 сайтами связывания для HER2/neu). CD137×TA связывающие молекулы не проявляли какую-либо наблюдаемую костимулирующую активность в отсутствие клеток-мишеней или с HER2/neu-отрицательными клетками-мишенями. Контрольные молекулы TRIDENT-1 и TRIDENT-2 не проявляли костимулирующую активность.
Пример 7
CD137×TA связывающие молекулы повышают активацию Т-клеток
[000525] Как описано выше, CD137×TA связывающие молекулы согласно настоящему изобретению могут повышать размножение и активацию Т-клеток (см., например. Фигуры 17А-17O) и эффекторную функцию NK (АЗКЦ), опосредуемую нацеленным на опухоль агентом (см., например, повышение опосредуемой маргетуксимабом АЗКЦ, Фигура 18А-18В). Известно, что биспецифические молекулы TA×CD3 (например, антитела DART, молекулы BiTE® и т.д.), которые связываются с ТА и с CD3, опосредуют перенаправленное уничтожение клеток-мишеней, экспрессирующих ТА, Т-клетками и стимулируют размножение и активацию Т-клеток (см., например, РСТ публикации №WO 2017/030926; WO 2016/048938; и WO 2015/026892). Способность CD137×TA связывающих молекул согласно настоящему изобретению повышать активацию Т-клеток и перенаправленное уничтожение Т-клетками, опосредуемое такими агентами, нацеленными на опухоль, оценивали в нескольких исследованиях с использованием примерных TA×CD3 биспецифических молекул.
[000526] В одном из таких исследований способность примерной CD137×TA связывающей молекулы TRIDENT-A4 усиливать перенаправленное уничтожение Т-клетками, опосредуемое TA×CD3 диателом, которое связывается с иллюстративным ТА gpA33, оценивали с использованием анализа цитотоксических Т-лимфоцитов (CTL) на основе люциферазы. В общих чертах, общую популяцию Т-клеток инкубировали с клетками Со1о205, модифицированными для экспрессии репортерного гена люциферазы (luc) (Colo205/luc), при соотношении эффектор : мишень (Е : Т) 3:1 в присутствии TRIDENT-A4 или контрольной молекулы TRIDENT-1 в комбинации с TA×CD3 диателом или контрольным Irrel×CD3 биспецифическим диателом (содержащим нецелевой связывающий домен (4-4-20 к флуоресцеину) вместо gpA33). В конце инкубации цитотоксичность определяли с помощью люминесцентного (LUM) анализа, в котором измеряли клеточную активность люциферазы клеток-мишеней, используя относительную световую единицу люминесценции (RLU) в качестве регистрируемого показателя (Фигура 27). Результаты этого исследования демонстрируют, что CD137×TA связывающие молекулы согласно настоящему изобретению способны повышать перенаправленное уничтожение Т-клетками, опосредуемое TA×CD3 биспецифическими молекулами.
[000527] В другом исследовании способность примерной CD137×TA связывающей молекулы TRIDENT-A2 повышать размножение Т-клеток и противоопухолевую активность в комбинации in vivo оценивали в модели карциномы яичников SK-OV3 (экспрессирующей ТА HER2/neu и 5Т4). В этом исследовании активность TRIDENT-A2 исследовали по отдельности или в комбинации с TA×CD3 диателом, которое связывается с иллюстративным ТА, 5Т4 (5T4×CD3), и необязательно в дополнительной комбинации с MAT к PD-1 (MAT 7 к чPD-1 (1.2) IgG4 (Р), см., например, SEQ ID NO: 264 и 266 из РСТ публикации WO 2017/019846).
[000528] В общих чертах, свежеизолированные МКПК вводили путем ретроорбитальной инъекции мышам NSG.MHCI-/- (по 7 самок мышей в группе для контроля опухолей; по 3 в группе для профилирования Т-клеток и опухолей) на 0 день исследования. Клетки SK-OV3 (5×106) смешивали в соотношении 1:1 с матригелем и вводили подкожно на 0 день исследования. На 7 день исследования мыши получали антитело к CD4, ОKТ4 (начальная доза 10 мг/кг подкожно, и затем 5 мг/кг в/б два раза в неделю), чтобы устранить CD4+ Т-клетки. Начиная с 22 дня, мыши получали внутривенные (в/в) инъекции: контрольной среды; TRIDENT-A2 (2,5 мг/кг, один раз в неделю); TA×CD3 диатела (0,01 мг/кг, два раза в неделю); комбинацию TRIDENT-A2 (2,5 мг/кг, один раз в неделю) и TA×CD3 диатела (0,01 мг/кг, два раза в неделю); MAT к PD-1 (5 мг/кг один раз в неделю); или комбинацию TRIDENT-A2 (2,5 мг/кг; один раз в неделю), TA×CD3 диатела (0,01 мг/кг, два раза в неделю) и MAT к PD-1 (5 мг/кг, один раз в неделю).
[000529] Т-клеточные маркеры оценивали с помощью анализа FACS через 7 дней после введения испытываемых изделий, и рост опухоли контролировали в течение всего исследования. Экспрессию Т-клеточных маркеров CD4, CD8, CD69 и PD-1, присутствующих в 1 мг опухоли, образец которой отбирали на 7 день после обработки, представляли в графической форме на Фигуре 28А, Фигуре 28В, Фигуре 28С и Фигуре 28D, соответственно. Эти данные показывают, что CD137×TA связывающая молекула TRIDENT-A2 повышает активацию и размножение Т-клеток (на примере увеличения экспрессии CD4, CD8 и CD69), опосредуемые TA×CD3 диателом. Экспрессия молекулы контрольной точки иммунного ответа PD-1 повышалась при обработке TA×CD3 диателом по отдельности. Обработка комбинацией CD137×TA связывающей молекулы TRIDENT-A2 и TA×CD3 диатела дополнительно повышала экспрессию молекулы контрольной точки иммунного ответа PD-1, наблюдаемую при обработке только TA×CD3 диателом, и это свидетельствует о том, что активность комбинации может быть повышена с помощью добавления ингибитора контрольной точки иммунного ответа. Действительно, добавление MAT к PD-1, примерного ингибитора контрольной точки иммунного ответа PD-1/PD-L1, к комбинации TRIDENT-A2/TA×CD3 диатела ингибировало повышение экспрессии PD-1 и дополнительно повышало активацию и пролиферацию Т-клеток.
[000530] Рост опухоли в ходе исследования представлен в графической форме на Фигуре 29, на которой показано, что CD137×TA связывающая молекула TRIDENT-A2 повышает противоопухолевую активность TA×CD3 диатела. Помимо этого добавление ингибитора контрольной точки иммунного ответа PD-1/PD-L1, MAT к PD-1, к комбинации еще более повышало противоопухолевую активность комбинации. Результаты этого исследования демонстрируют, что CD137×TA связывающие молекулы согласно настоящему изобретению, в частности, те, которые имеют два сайта связывания для CD137 и один сайт связывания для ТА, повышают активацию и пролиферацию Т-клеток, а также уничтожение Т-клетками клеток-мишеней в комбинации с агентом, нацеленным на опухоль. Соответственно, CD137×TA связывающие молекулы согласно настоящему изобретению можно преимущественно применять в комбинации с агентами, нацеленными на опухоль, в частности, с теми, которые опосредуют активацию Т-клеток, активацию NK-клеток и/или стимулируют экспрессию CD137 в таких клетках. Кроме того, эти исследования демонстрируют, что добавление ингибитора контрольной точки иммунного ответа PD-1/PD-L1, такого как антитело к PD-1 или антитело к PD-L1, может дополнительно повышать активацию и пролиферацию Т-клеток, а также уничтожение клеток-мишеней Т-клетками.
Пример 8
Исследования стабильности
[000531] Оценивали температуру плавления (Tm) и другие параметры стабильности примерных трехвалентных CD137×TA связывающих молекул TRIDENT-А (содержащей hMAB-3 к CD137 (1.3)) и оптимизированных вариантов TRIDENT-A1 (содержащей hMAB-3 к CD137 (1А.3)), TRIDENT-A2 (содержащей hMAB-3 к CD137 (1 В.3)), TRIDENT-A3 (содержащей hMAB-3 к CD137 (1С.3)) и TRIDENT-A4 (содержащей hMAB-3 к CD137 (1D.3)). Tm очищенного материала (>95% согласно результатам определения методом гель-проникающей хроматографии (ГХ) высокого давления (ВЭГХ)) оценивали с помощью дифференциальной сканирующей калориметрии (DSC). В общих чертах, степень чистоты измеряли методом ВЭЖХ-ГХ с использованием Agilent 1260 Infinity II HPLC, соединенного с колонкой Waters Acquity UPLC ВЕН (200 , 1,7 мкм, 4,6×150 мм). Подвижный буфер представлял собой 20 мМ фосфата натрия, 0,5 М NaCl, 0,02% азида натрия, рН=7,2. Скорость потока составляла 0,4 мл/мин в течение 5 минут. Процент чистоты определяли по % площади кривой поглощения для пика мономера при 280 нм. Измерения термостабильности выполняли на MicroCal VP-Capillary DSC (Malvern Instruments, Inc.) при температуре 15-95°C с линейным увеличением 1°С/мин.
, 1,7 мкм, 4,6×150 мм). Подвижный буфер представлял собой 20 мМ фосфата натрия, 0,5 М NaCl, 0,02% азида натрия, рН=7,2. Скорость потока составляла 0,4 мл/мин в течение 5 минут. Процент чистоты определяли по % площади кривой поглощения для пика мономера при 280 нм. Измерения термостабильности выполняли на MicroCal VP-Capillary DSC (Malvern Instruments, Inc.) при температуре 15-95°C с линейным увеличением 1°С/мин.
[000532] Чтобы оценить другие параметры стабильности, очищенные CD137×TA связывающие молекулы (1 мг/мл в ФСБ, рН=7,2, если не указано иное) подвергали оценке в стрессовых условиях, повторно оценивали с помощью ВЭГХ и сравнивали значения со значениями, полученными для молекулы, не подвергнутой стрессу. Использовали следующие стрессовые условия: три цикла замораживания (1 час при -80°С) с последующим оттаиванием; затем хранение в течение 2 и 7 дней при 25°С или 40°С; затем хранение в течение 0, 2, 7 дней при 25°С в 20 мМ ацетате натрия, рН=5,5; хранение в течение 0, 2, 7 дней при 25°С в 10 мМ гистидине НСl, рН=6,5. Результаты этих исследований приведены в таблице 14.

[000533] Начальная точка Tm и Tm CD137-связывающих областей (Tm1 находятся в оптимальном диапазоне для TRIDENT-A и TRIDENT-A2. Однако начальная точка Tm и Tm1 ниже как для TRIDENT-A3, так и для TRIDENT-A4. Только TRIDENT-A2 была стабильной при 40°С (2 день и 7 день) в ФСБ, остальные молекулы были чувствительны к этим условиям хранения. TRIDENT-A и TRIDENT-A2 обе были стабильны в ацетате, рН=5,5, тогда как TRIDENT-A3 и TRIDENT-A4 обе были чувствительны к инкубации с ацетатом, рН=5,5. Все испытанные молекулы были стабильны в гистидине при рН=6,5. Основываясь на результатах этих исследований, ранжирование по стабильности в испытываемых буферах является следующим TRIDENT-A2 > TRIDENT-A > TRIDENT-А3 > TRIDENT-A4.
Пример 9
Костимулирующая активность с Т-клетками яванской макаки
[000534] Как отмечено выше, молекулы, содержащие связывающие домены МАВ-3 к CD137 (или их гуманизированные, оптимизированные варианты), связывают как CD137 человека, так и CD137 яванской макаки. Типичные CD137×TA связывающие молекулы, TRIDENT-A2 и TRIDENT-B2, имеющие два сайта связывания для CD137 и один сайт связывания для HER2/neu или 5Т4, оценивали для определения их способности опосредовать костимулирующую активность в анализе высвобождения цитокинов Т-клетками яванской макаки. Структура и последовательности этих примерных CD137×TA связывающих молекул подробно представлены выше. Также были включены контрольные молекулы TRIDENT-1 и TRIDENT-2. Анализ высвобождения цитокинов выполняли по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ, в присутствии или в отсутствие клеток-мишеней, за исключением того, что использовали общую популяцию Т-клеток яванской макаки. Для этих исследований применяли клетки-мишени JIMT-1, экспрессирующие HER2/neu и 5Т4.
[000535] Результаты представлены на Фигуре 30 и показывают, что контрольные молекулы TRIDENT-1 и TRIDENT-2 не проявляли костимулирующую активность. В то время как CD137×TA связывающие молекулы TRIDENT-A2 и TRIDENT-B2 (содержащие гуманизированное/оптимизированное антитело к CD137, hMAB-3 к CD137 (1В.3)) обе проявляли костимулирующую активность, зависящую от дозы.
Пример 10
Деиммунизация CD137
[000536] Анализ hMAB-3 к CD137 (1В.3) in silico выявил два потенциальных пептида, связывающих ГКГС класса II (Т-клеточные эпитопы), в домене VH и три потенциальных пептида, связывающих ГКГС класса II, в домене VL. Панель аминокислотных замен hMAB-3 к CD137 (1В.3) исследовали для выявления замен (которые могут представлять собой единичные аминокислотные замены или группы замен) для устранения/снижения потенциальной иммуногенности выявленных Т-клеточных эпитопов. В частности, замены аминокислот вводили в двух положениях (остатки 38 и 48 согласно Kabat) для деиммунизации VH1B hMAB-3 к CD137 и не более чем в шести положениях (остатки 24, 15, 44, 48, 52 и 54 согласно Kabat) для деиммунизации VL3 hMAB-3 к CD137. Замененные остатки аминокислот в доменах VH и VL заключены в рамку, и нумерация согласно Kabat указана стрелками на Фигурах 31А и 31В, соответственно. Получали антитела IgG1 (имеющие домен Fc 234A/235A (SEQ ID NO: 40)), содержащие варианты VH и/или VL hMAB-3 к CD137 (1В.3), имеющие различные замены в этих положениях, по отдельности и/или в комбинации, и характеризовали их способность связывать CD137 с помощью Attana Cell A200 QCM. В общих чертах, hMAB-3 к CD137 (1В.3) и деиммунизированные варианты независимо захватывали на сенсорном чипе Attana, покрытом кроличьим поликлональным антителом к Fc человеческого IgG, и растворимый гибридный человеческий белок CD137, несущий His-метку, пропускали над чипом в концентрации 11,1, 33,3, 100 и 300 мМ. Сенсограммы деиммунизированных вариантов сравнивали с таковыми hMAB-3 к CD137 (1В.3) и оценивали. Антитела также были охарактеризованы по их термостабильности с помощью DSC по существу, как описано выше в примере 8. Результаты для ряда таких вариантов обобщены в таблице 15, аминокислотные последовательности этих вариантов представлены выше.
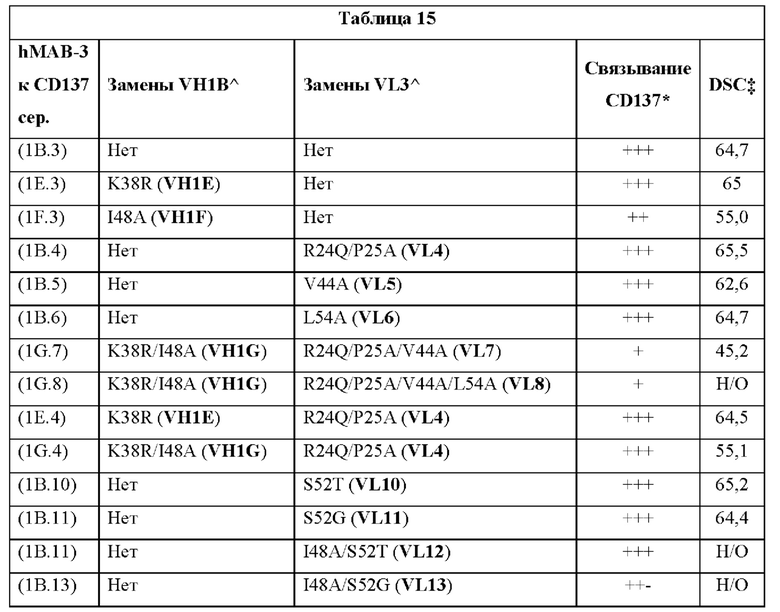

[000537] Было выявлено большое количество деиммунизированных вариантов, которые сохраняют как активность связывания CD137, так и термическую стабильность. Большинство деиммунизированных вариантов, содержащих замену I48A домена VH, проявляли некоторое снижение как связывания CD137, так и начальной точки Tm.
[000538] Способность деиммунизированных hMAB-3 к CD137 (1Е.15) и hMAB-3 к CD137 (1G.15), а также исходного гуманизированного hMAB-3 к CD137 (1В.3) связываться с активированными CD4+ и CD8+ Т-клетками испытывали по существу, как описано выше. Как показано на Фигурах 32А-32В, кривая связывания деиммунизированного антитела hMAB-3 к CD137 (1Е.15) не изменилась, в то время как hMAB-3 к CD137 (1G.15) проявляло незначительно сниженное связывание как с CD4+, так и с CD8+ Т-клетками.
[0005391 Агонистическую активность hMAB-3 к CD137 (1Е.15) и hMAB-3 к CD137 (1G.15) в условиях in vitro оценивали в анализе высвобождения цитокинов Т-клетками (выполненном по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ) в отсутствие сшивания или с ним. Ни одно из антител на основе hMAB-3 к CD137 не проявило активность в отсутствие сшивания (Фигура 33А), при этом сшитое hMAB-3 к CD137 (1Е.15) проявляло агонистическую активность, сходную с таковой hMAB-3 к CD137 (1В.3), в то время как hMAB-3 к CD137 (1G.15) проявляло незначительное снижение агонистической активности, вероятно, вследствие сниженного связывания с CD137 (Фигура 33В). Как сообщалось выше, МАВ-1 к CD137 проявляло агонистическую активность в присутствии и в отсутствие сшивания.
Пример 11
Трехвалентные CD137×TA связывающие молекулы, содержащие деиммунизированные CD137-связывающие домены
[000540] Как описано выше, домены VH и VL hMAB-3 к CD137 гуманизировали, оптимизировали и деиммунизировали. Примерные биспецифические четырехвалентные и/или трехвалентные CD137×TA связывающие молекулы получали путем включения таких доменов VH и VL. Одну такую биспецифическую трехвалентную связывающую молекулу «TRIDENT-B5», содержащую CD137-связывающие домены hMAB-3 к CD137 (1Е.15), получали и оценивали в ряде анализов. Структура и последовательность этой примерной CD137×TA связывающей молекулы подробно представлена выше. В эти исследования включали TRIDENT-B2 и одну или более контрольных молекул: TRIDENT-1, TRIDENT-2, TRIDENT-3, TRIDENT-4.
[000541] Оценивали способность TRIDENT-B2, TRIDENT-B5 и контрольных молекул TRIDENT-1, TRIDENT-3 и TRIDENT-4 связываться с активированными CD8+ Т-клетками и/или 5Т4, присутствующим на поверхности клеток-мишеней (высоко экспрессирующие клетки карциномы молочной железы JIMT-1 и слабо экспрессирующие клетки карциномы яичника SKOV-3), по существу, как описано выше. Как показано на Фигуре 34А, все молекулы, содержащие два CD137-связывающих домена (TRIDENT-B2, TRIDENT-B5 и контрольные молекулы TRIDENT-1 и TRIDENT-4), проявляли сходное связывание с активированными CD8+ Т-клетками, в то время как контрольная молекула, не содержащая CD137-связывающие домены (TRIDENT-3), не связывалась. Аналогичным образом, все молекулы, имеющие 5Т4-связывающий домен (TRIDENT-B2, TRIDENT-B5 и контрольная TRIDENT-3), проявляли сходное сильное связывание с клетками-мишенями, экспрессирующими высокие уровни 5Т4 (JIMT-1, Фигура 34В), и более слабое связывание к клеткам-мишеням, экспрессирующим низкие уровни 5Т4 (SKOV-3, Фигура 34С), в то время как контрольные молекулы, не содержащие 5Т4-связывающий домен (TRIDENT-1 и TRIDENT-4), не связывались ни с одним типом клеток-мишеней, экспрессирующих 5Т4.
[000542] Агонистическую активность TRIDENT-B2, TRIDENT-B5 и контрольных молекул TRIDENT-1, TRIDENT-3 и TRIDENT-4 в условиях in vitro оценивали в анализе высвобождения цитокинов Т-клетками (выполненном по существу, как описано в примере 2 выше, и на примере высвобождения ИФН-γ) в присутствии или в отсутствие клеток-мишеней, экспрессирующих различные уровни примерного ТА, клеток 5Т4 (JIMT-1 (5Т4hi) или клеток SKOV-3 (5Т4lo).
[000543] Как показано на Фигурах 35А-35С, TRIDENT-1, TRIDENT-4 (не содержащая 5Т4-связывающий домен) не проявляли костимулирующую активность. Также было обнаружено, что TRIDENT-3 (не содержащая CD137-связывающие домены) не обладает какой-либо костимулирующей активностью. TRIDENT-B2 и TRIDENT-B5 проявляли сходную дозозависимую костимулирующую активность в присутствии клеток-мишеней JIMT-1, экспрессирующих высокие уровни 5Т4 (Фигура 35А), но незначительную или отсутствующую костимулирующую активность в присутствии клеток-мишеней SKOV-3, экспрессирующих низкие уровни 5Т4 (Фигура 35В), или в отсутствие клеток-мишеней (Фигура 35С).
[000544] Результаты этих исследований показывают, что CD137×TA связывающие молекулы, содержащие деиммунизированные домены VH и VL hMAB-3 к CD137 (1Е.15), проявляют сопоставимые профили связывания и костимулирующей активности, как и CD137×TA связывающие молекулы, содержащие оптимизированные домены VH и VL hMAB-3 к CD137 (1В.3).
[000545] Все публикации и патенты, упомянутые в этом описании, включены в настоящий документ посредством ссылки в той же степени, как если бы каждая отдельная публикация или заявка на патент была конкретно и индивидуально указана для полного включения посредством ссылки. Несмотря на то, что настоящее изобретение было описано применительно к его конкретным вариантам реализации, следует понимать, что в него могут быть внесены дополнительные модификации и подразумевается, что настоящая заявка охватывает любые изменения, варианты применения или адаптации настоящего изобретения в целом в соответствии с принципами настоящего изобретения, и включая такие отступления от настоящего изобретения, которые соответствуют известной или обычной практике в области техники, к которой относится настоящее изобретение, и которые могут быть применены к существенным признакам, изложенным выше в настоящем документе.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| ТРИСПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, КОТОРЫЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАЮТ АНТИГЕНЫ МНОЖЕСТВА ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2718692C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| КОВАЛЕНТНЫЕ ДИАНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2593720C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2008 |
|
RU2535992C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2733315C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ВЫЗЫВАЮЩИЕ КЛЕТОЧНУЮ АКТИВАЦИЮ | 2019 |
|
RU2770620C1 |
| СПОСОБЫ ПРИМЕНЕНИЯ CD32B X CD79B-СВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРИ ЛЕЧЕНИИ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ И РАССТРОЙСТВ | 2017 |
|
RU2772434C2 |
| КОВАЛЕНТНЫЕ ДИАНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2744176C2 |
| ВЫСОКОАФФИННЫЕ АНТИТЕЛА К CD3 И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2831840C2 |


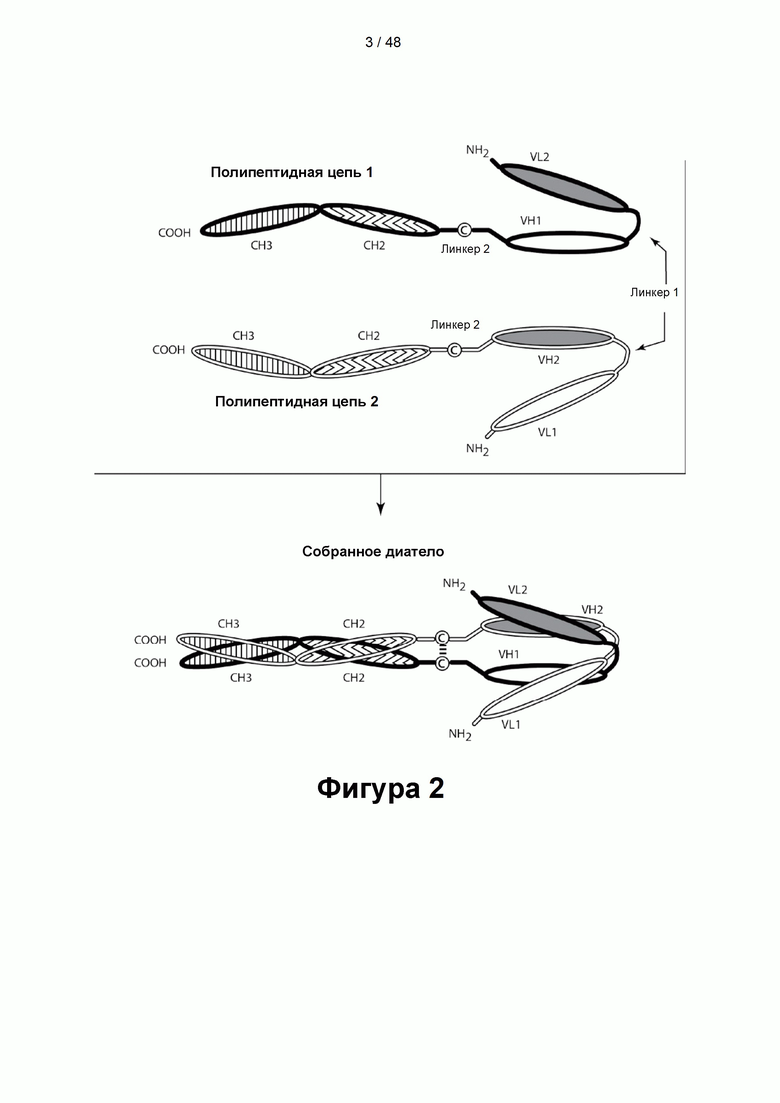

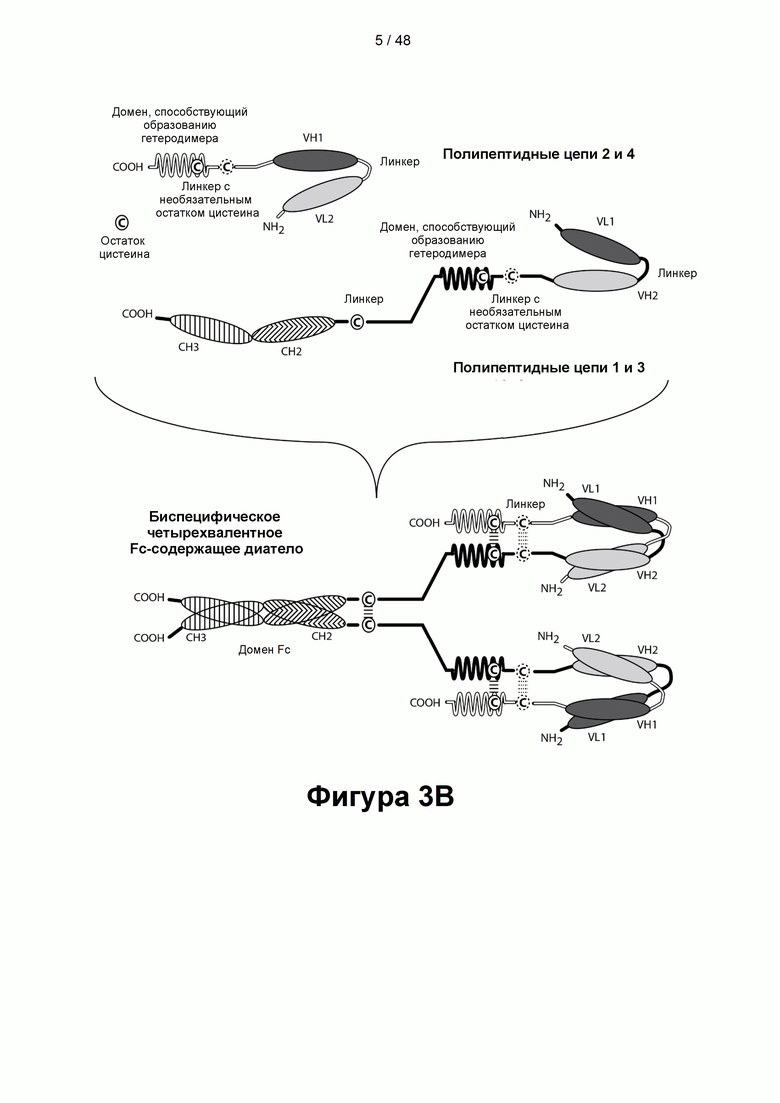
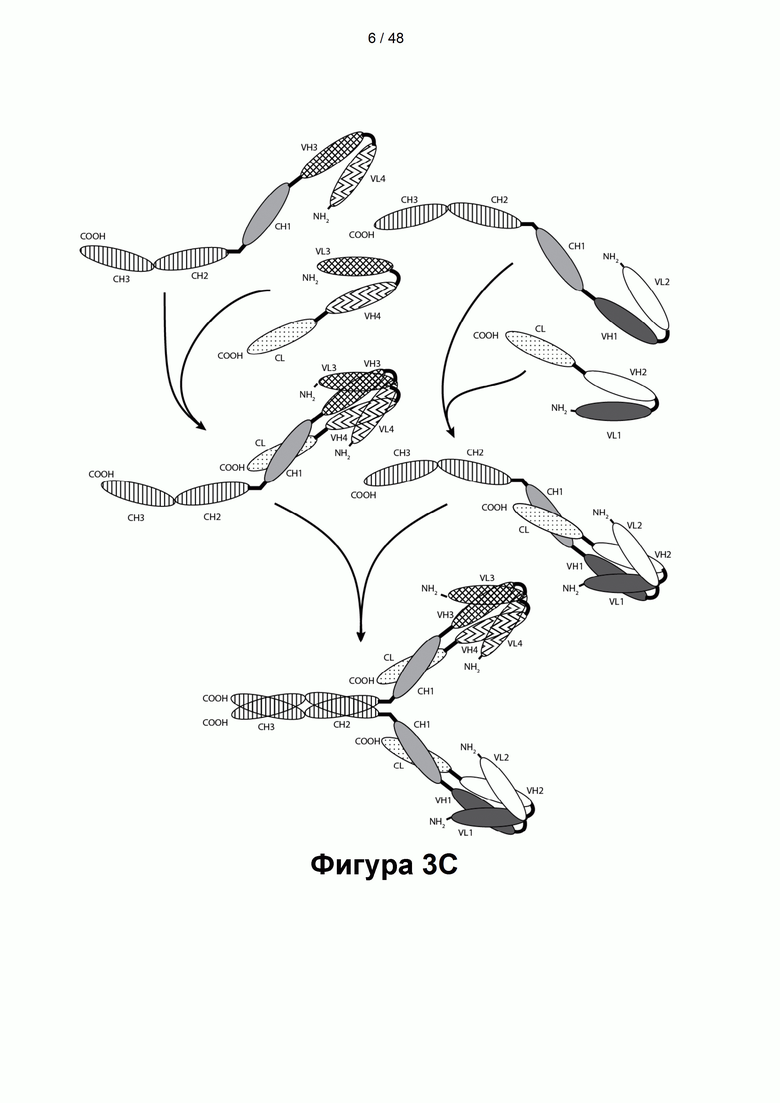
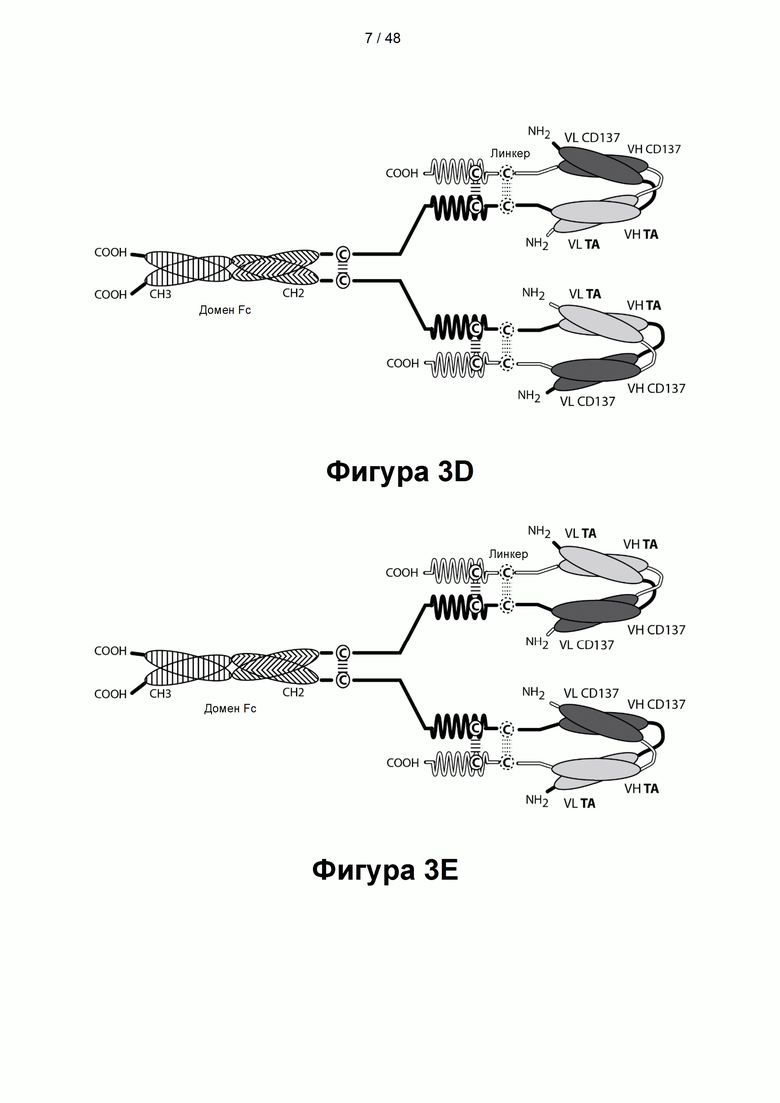
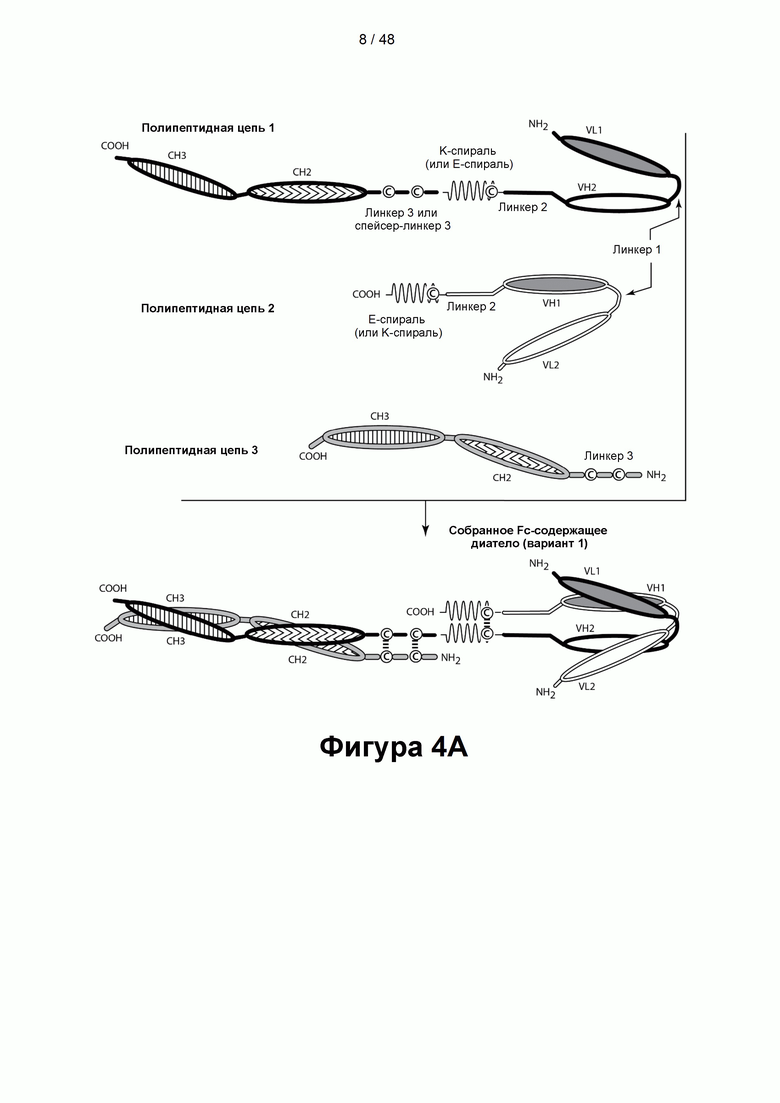




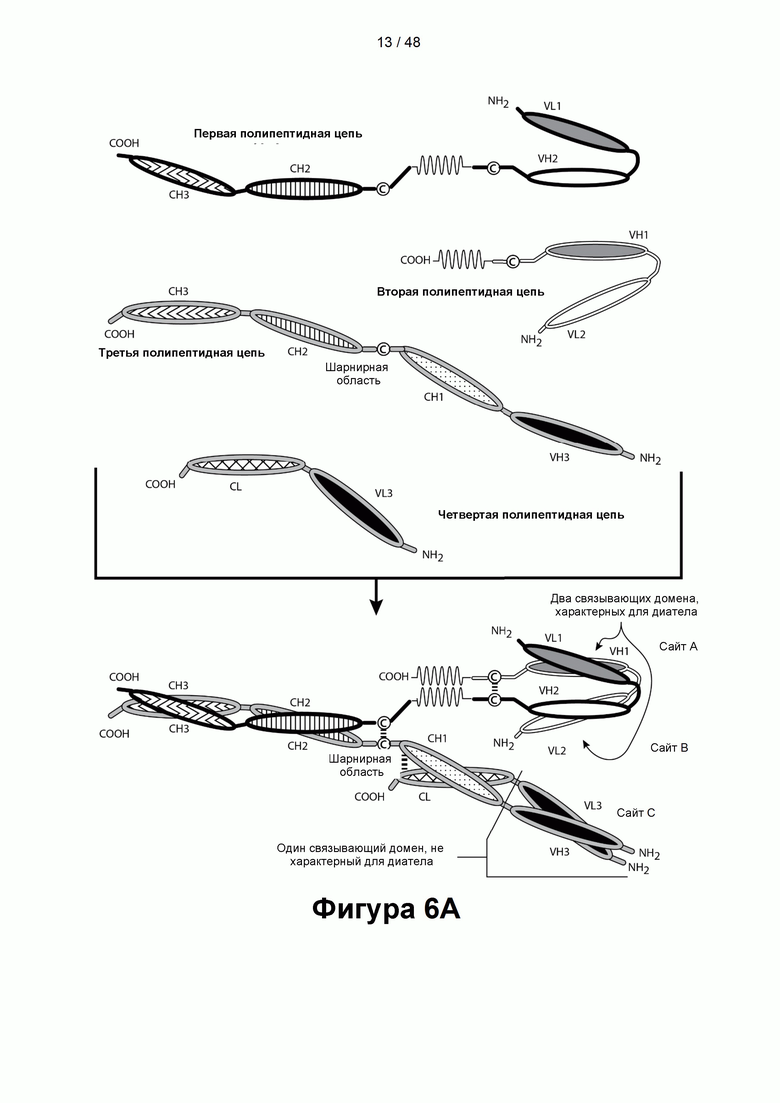




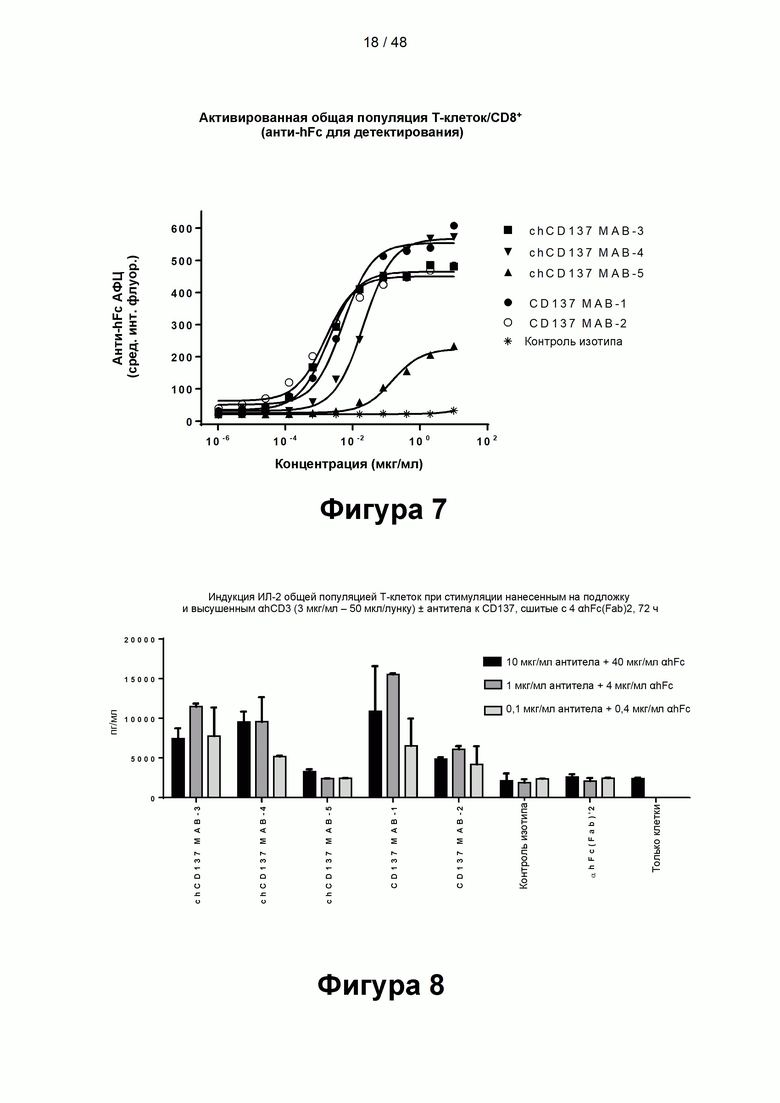

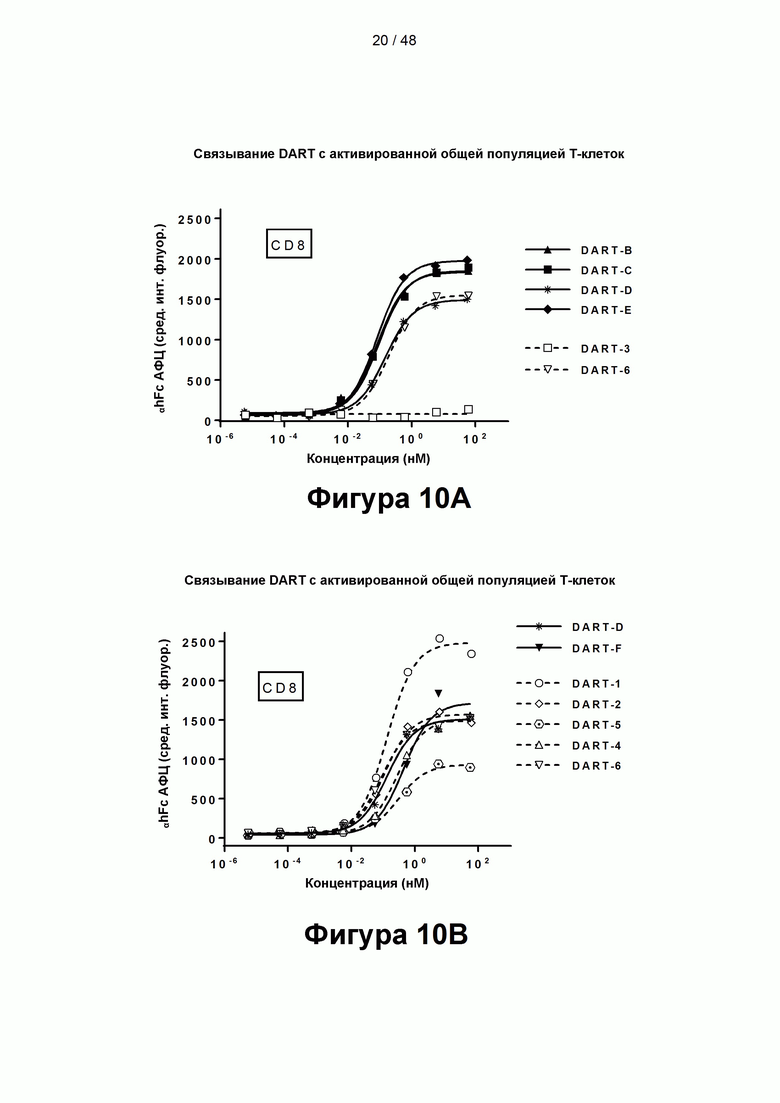


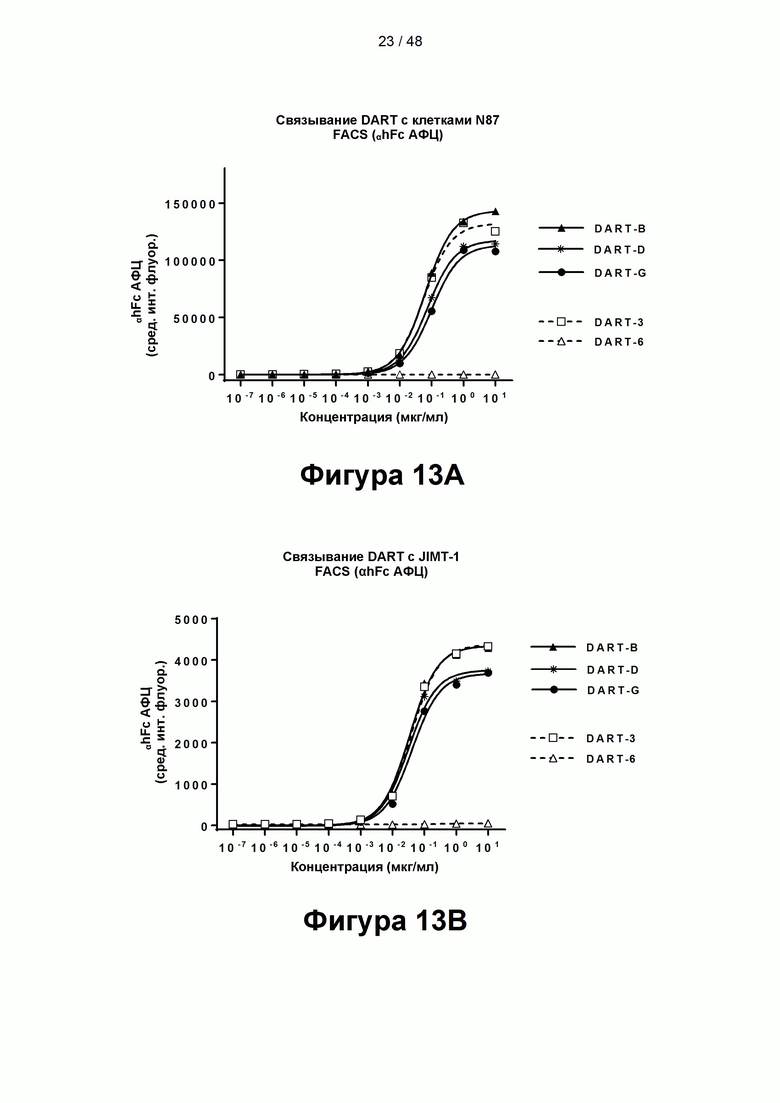




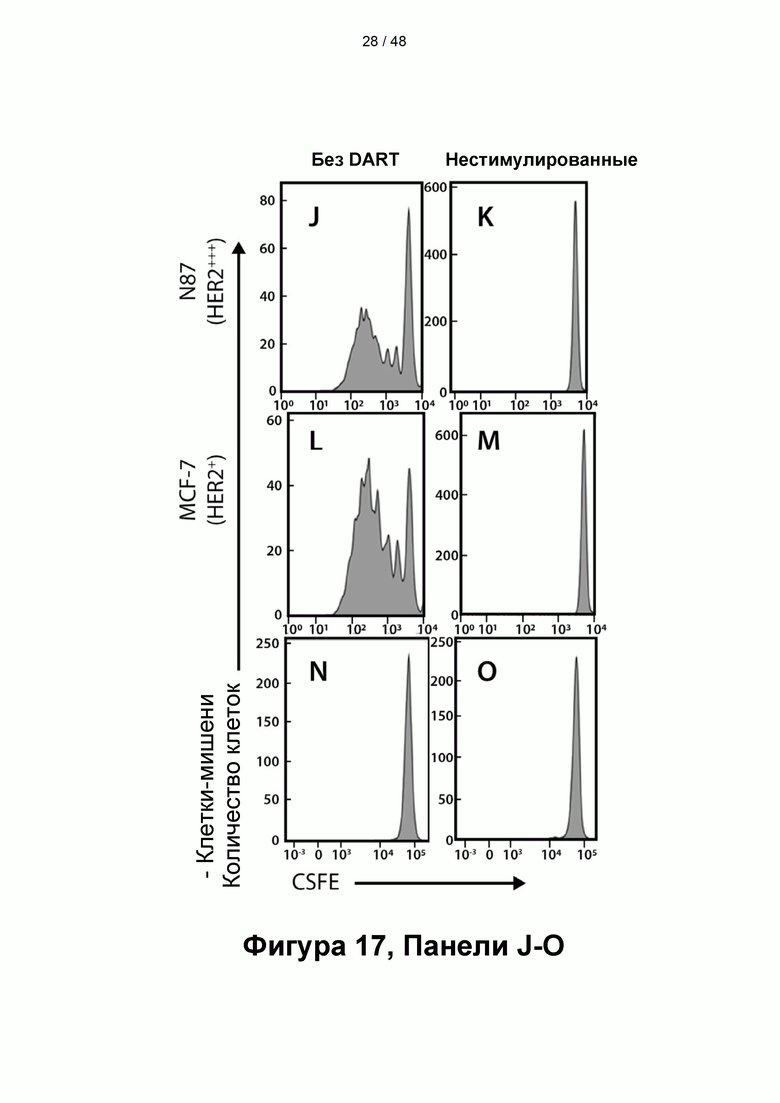




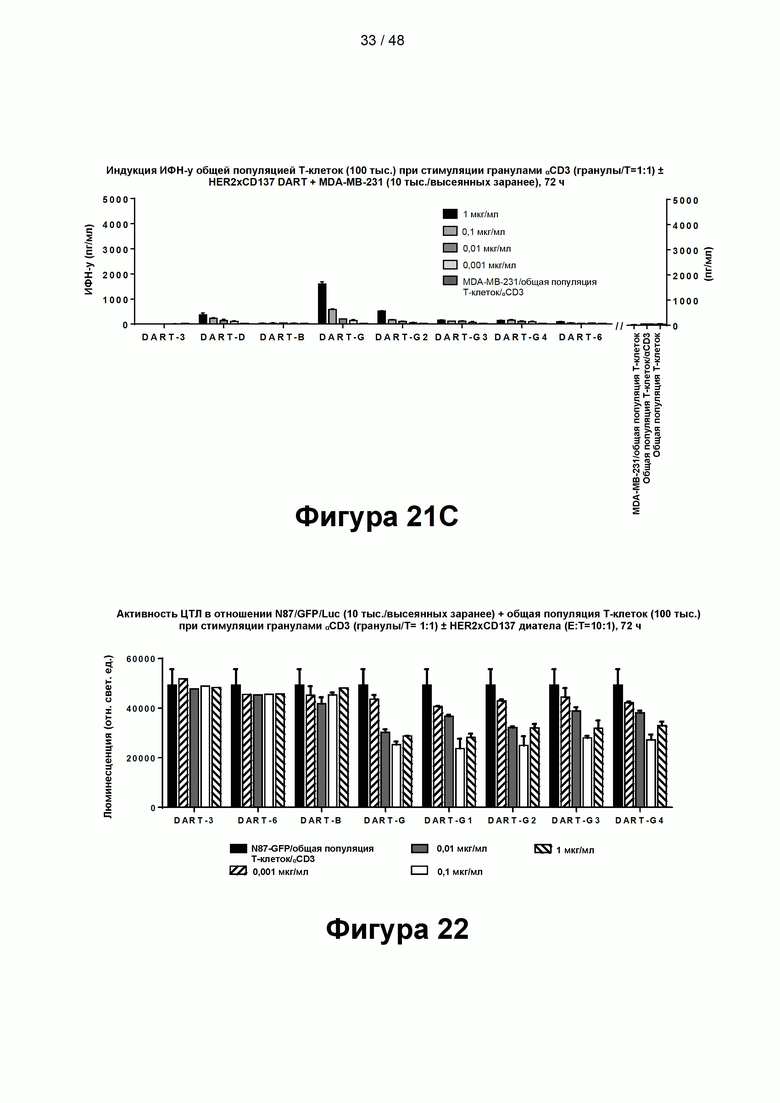


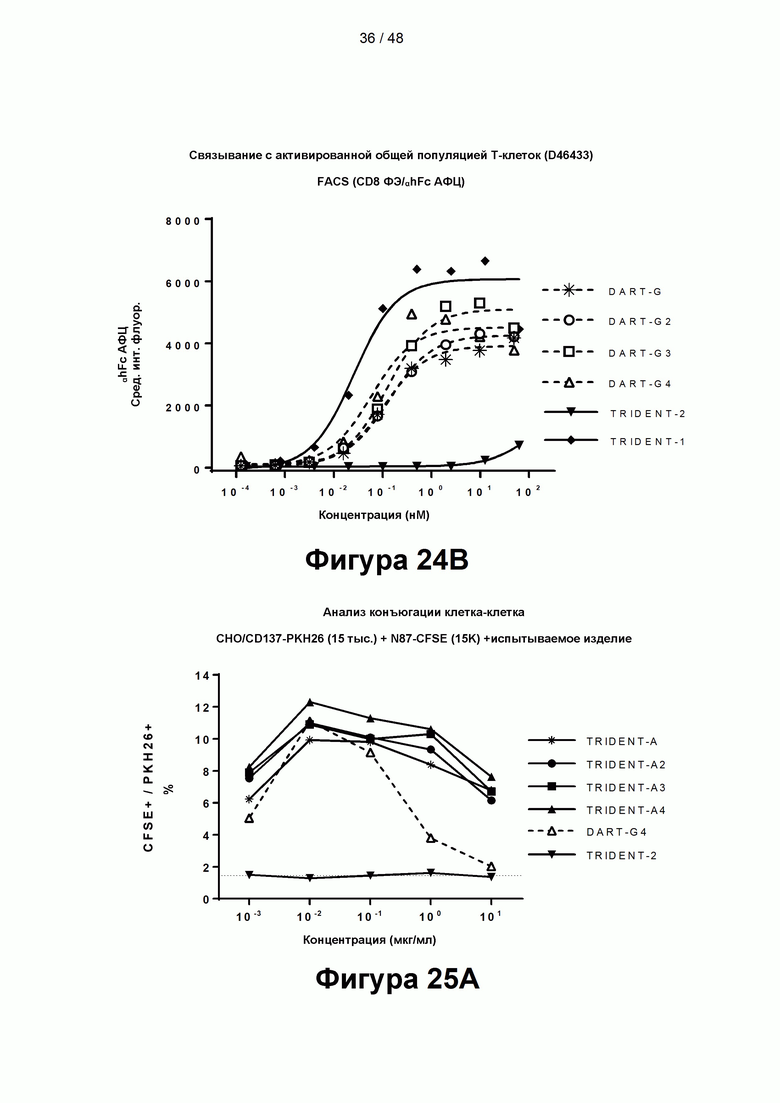

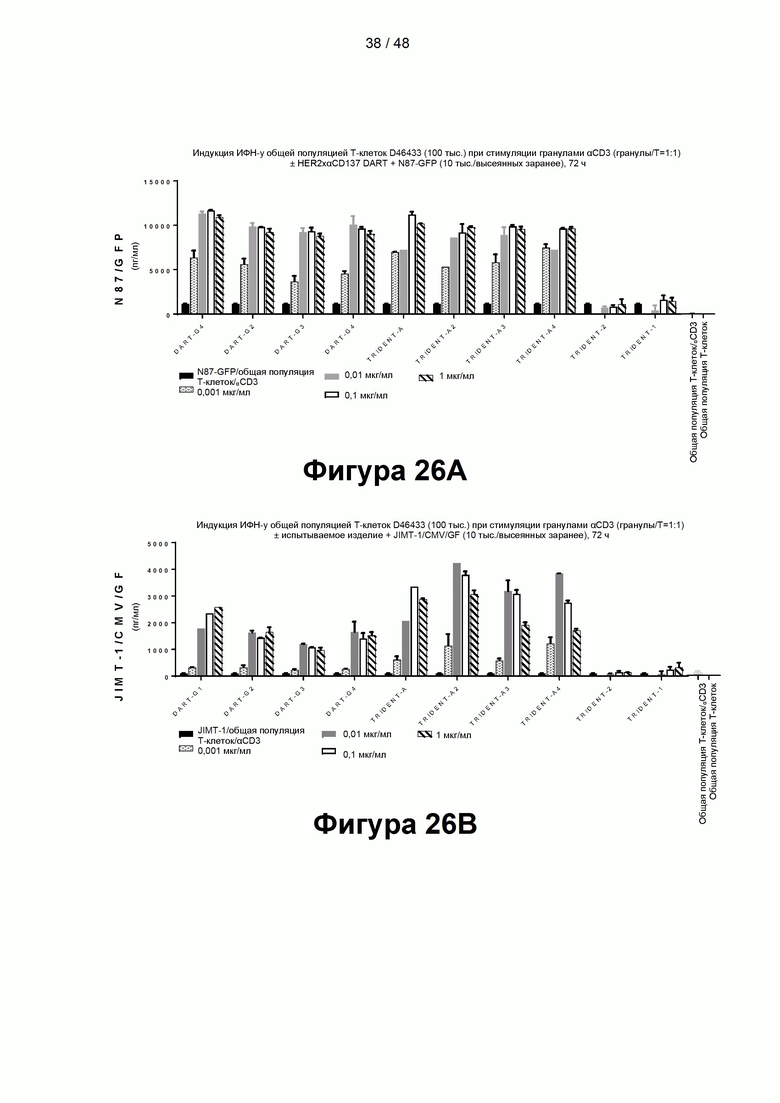



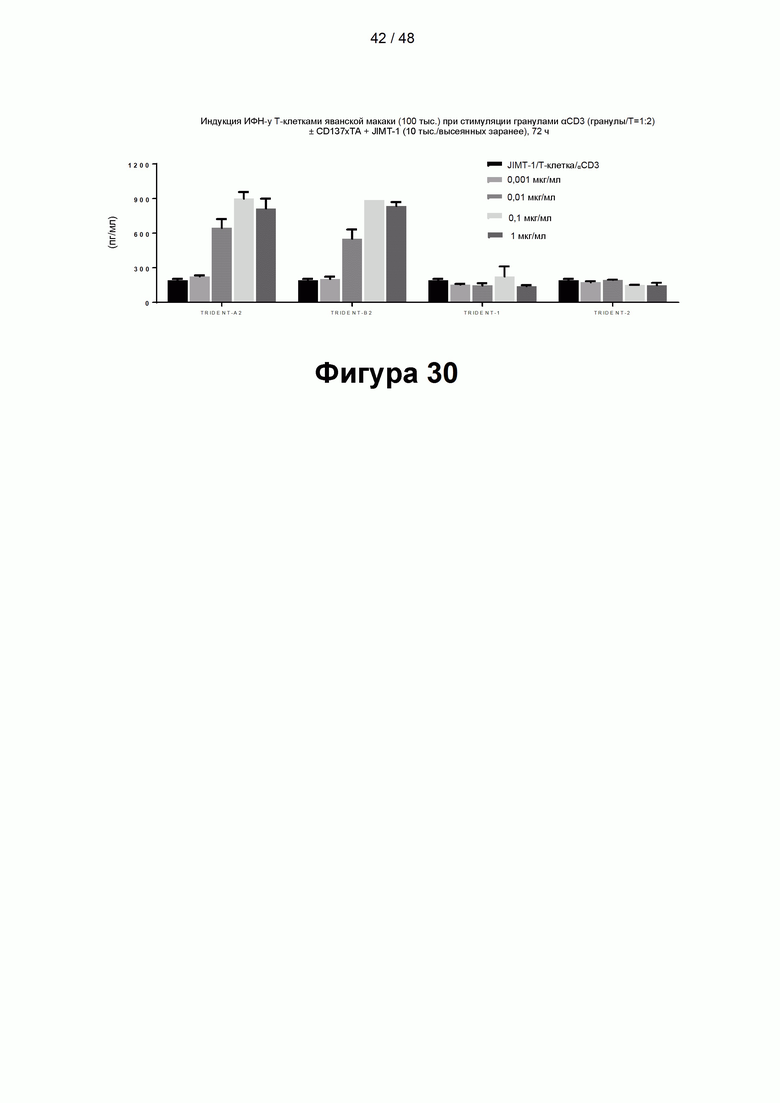

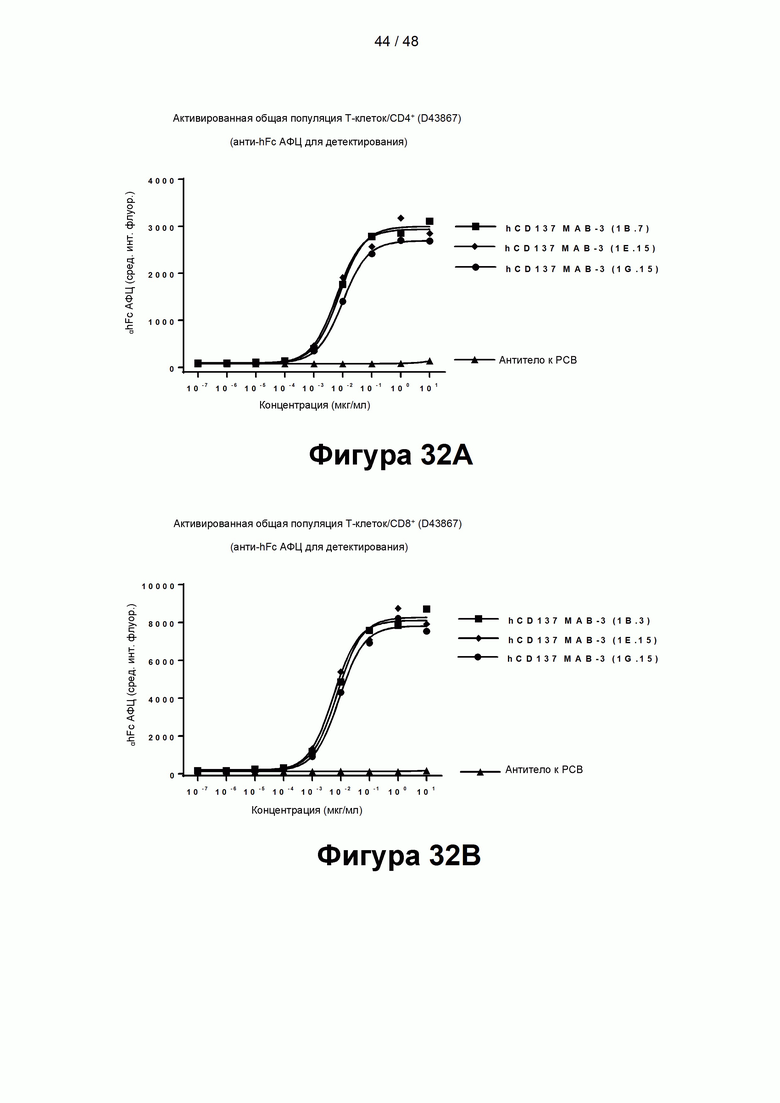


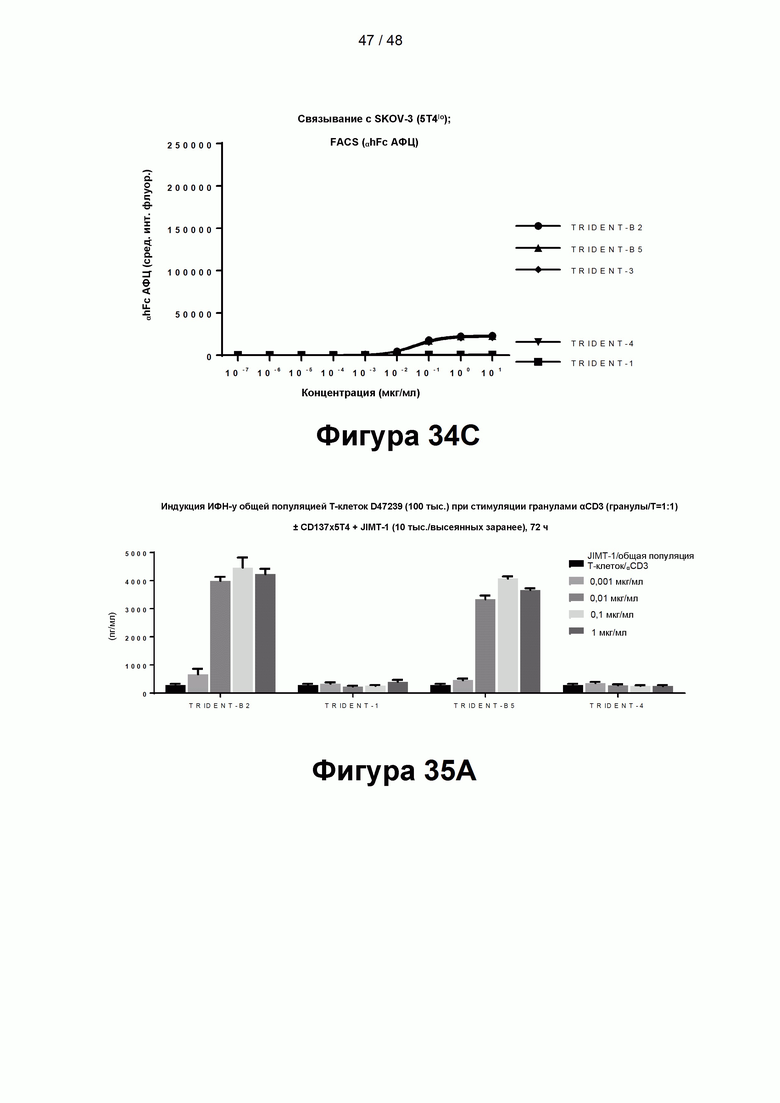

Изобретение относится к области биотехнологии. Предложена новая HER2/neu-связывающая молекула. Также предложены фармацевтическая композиция, содержащая указанные связывающие молекулы, и применение указанных молекул для лечения заболеваний и состояний, ассоциированных с экспрессией HER2/Neu, таких как рак. Изобретение обеспечивает получение связывающих молекул с улучшенной аффинностью связывания с HER2 и улучшенной стабильностью. 3 н. и 9 з.п. ф-лы, 81 ил., 15 табл., 11 пр.
1. HER2/neu-связывающая молекула, содержащая вариабельный домен легкой цепи, который содержит CDRL1, CDRL2 и CDRL3, и вариабельный домен тяжелой цепи, который содержит CDRH1, CDRH2 и CDRH3; при этом:
(A) (1) указанный вариабельный домен легкой цепи содержит CDRL1, содержащий аминокислотную последовательность SEQ ID NO:187, CDRL2, содержащий аминокислотную последовательность SEQ ID NO:190, и CDRL3, содержащий аминокислотную последовательность SEQ ID NO:182; или
(2) указанный вариабельный домен легкой цепи содержит CDRL1, содержащий аминокислотную последовательность SEQ ID NO:188, CDRL2, содержащий аминокислотную последовательность SEQ ID NO:191, и CDRL3, содержащий аминокислотную последовательность SEQ ID NO:182;
и
(B) указанный вариабельный домен тяжелой цепи содержит CDRH1, содержащий аминокислотную последовательность SEQ ID NO:177, CDRH2, содержащий аминокислотную последовательность SEQ ID NO:178, и CDRH3, содержащий аминокислотную последовательность SEQ ID NO:179.
2. HER2/neu-связывающая молекула по п. 1, отличающаяся тем, что указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность VH1 hMAB-1 к HER2 (SEQ ID NO: 64).
3. HER2/neu-связывающая молекула по п. 1 или 2, отличающаяся тем, что указанный вариабельный домен легкой цепи содержит аминокислотную последовательность:
(A) VL2 hMAB-1 к HER2 (SEQ ID NO: 68); или
(B) VL3 hMAB-1 к HER2 (SEQ ID NO: 69).
4. HER2/neu-связывающая молекула по п. 3, отличающаяся тем, что
указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность VH1 hMAB-1 к HER2 (SEQ ID NO: 64); и
указанный вариабельный домен легкой цепи содержит аминокислотную последовательность VL2 hMAB-1 к HER2 (SEQ ID NO: 68).
5. HER2/neu-связывающая молекула по п. 3, отличающаяся тем, что
указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность VH1 hMAB-1 к HER2 (SEQ ID NO: 64); и
указанный вариабельный домен легкой цепи содержит аминокислотную последовательность VL3 hMAB-1 к HER2 (SEQ ID NO: 69).
6. HER2/neu-связывающая молекула по любому из пп. 1-5, отличающаяся тем, что указанная молекула представляет собой антитело или его антигенсвязывающий фрагмент.
7. Фармацевтическая композиция, содержащая HER2/neu-связывающую молекулу по любому из пп. 1-6 и физиологически приемлемый носитель, для лечения заболевания или состояния, ассоциированного с или характеризующегося экспрессией HER2/Neu.
8. Фармацевтическая композиция по п. 7, отличающаяся тем, что указанное состояние, ассоциированное с или характеризующееся экспрессией HER2/neu, представляет собой рак.
9. Фармацевтическая композиция по п. 8, отличающаяся тем, что указанный рак выбран из группы, состоящей из: рака мочевого пузыря, рака молочной железы, колоректального рака, рака желудка, глиобластомы, рака почек, рака легких, меланомы, нейробластомы, рака яичников, рака поджелудочной железы, рака глотки, рака предстательной железы, почечно-клеточной карциномы, рабдомиосаркомы и плоскоклеточного рака головы и шеи (SCCHN).
10. Применение HER2/neu-связывающей молекулы по любому из пп. 1-6 или фармацевтической композиции по п. 7 в лечении заболевания или состояния, ассоциированного с или характеризующегося экспрессией HER2/Neu.
11. Применение по п. 10, отличающееся тем, что указанное состояние, ассоциированное с или характеризующееся экспрессией HER2/neu, представляет собой рак.
12. Применение по п. 11, отличающееся тем, что указанный рак выбран из группы, состоящей из: рака мочевого пузыря, рака молочной железы, колоректального рака, рака желудка, глиобластомы, рака почек, рака легких, меланомы, нейробластомы, рака яичников, рака поджелудочной железы, рака глотки, рака предстательной железы, почечно-клеточной карциномы, рабдомиосаркомы и плоскоклеточного рака головы и шеи (SCCHN).
| EP 2998319 A1, 23.03.2016 | |||
| Способ дубления жестких кож | 1960 |
|
SU136005A1 |
| WONG D.J.L., HURVITZ S.A., Recent advances in the development of anti-HER2 antibodies and antibody-drug conjugates, Ann Transl Med, 2014, vol.2(12):122 | |||
| СОКОЛОВА Е.А | |||
| и др., Рекомбинантный иммунотоксин 4D5scFv-PE40 для таргетной терапии HER2-положительных опухолей, ACTA NATURAE, 2015, Том 7, | |||
