ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области биологии и относится к антителу к SIRPα.
УРОВЕНЬ ТЕХНИКИ
SIRPα представляет собой трансмембранный белок, экспрессируемый на поверхности миелоидных клеток, таких как макрофаги, моноциты, дендритные клетки, гранулоциты, и является членом суперсемейства иммуноглобулинов (IgSF). SIRPα состоит из N-концевой внеклеточной области, трансмембранной области и C-концевой внутриклеточной области. Внеклеточная область содержит три домена IgSF (N-конец представляет собой домен Ig-V), и внутриклеточная область содержит иммунорецепторный ингибиторный мотив на основе тирозина (ITIM). Основным лигандом SIRPα является CD47, который представляет собой группу трансмембранных гликопротеинов, которые широко экспрессированы в нормальных тканях и в поражениях. Связывание CD47 с SIRPα на миелоидных клетках, таких как макрофаги, приводит к фосфорилированию ITIM во внутриклеточной области SIRPα, что в свою очередь привлекает и активирует фосфатазы SHP-1 и SHP-2, ингибируя тем самым фагоцитоз макрофагами посредством нисходящей передачи сигналов. Нормальные ткани высвобождают сигнал «не ешь меня» в качестве механизма самозащиты, экспрессируя CD47 и индуцируя передачу сигналов CD47-SIRPα.
Обнаружено, что CD47 сверхэкспрессируется почти на всех опухолевых клетках. Опухолевые клетки избегают надзора иммунных клеток, посылая сигнал «не ешь меня» посредством связывания CD47 с SIRPα. С клинической точки зрения уровни экспрессии CD47 тесно связаны с неблагоприятным прогнозом у пациентов. Исследования in vitro и in vivo также продемонстрировали, что блокирование CD47/SIRPα может способствовать фагоцитозу опухолевых клеток и ингибировать рост опухоли в моделях на животных. Упомянутое выше свидетельствует, что нацеливание на CD47/SIRPα можно применять в качестве нового подхода для разработки противоопухолевых иммунотерапевтических средств. Учитывая, что CD47 широко экспрессируется на нормальных клетках или тканях, особенно на эритроцитах и тромбоцитах, нацеливание на CD47 может вызвать гематологическую токсичность. Кроме того, в дополнение к SIRPα, CD47 также может взаимодействовать с TSP-1 и интегрином. CD47 участвует в различных сигнальных путях, и нацеливание на CD47 может привести ко многим проблемам с безопасностью. Таким образом, разработка антител к SIRPα для блокирования сигнального пути CD47/SIRPα может быть более эффективной стратегией лечения рака.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предназначено для обеспечения нового антитела к SIRPα, которое может специфично связываться с SIRPα и является очень перспективным для лечения опухолей.
Согласно первому аспекту настоящего изобретения предложено антитело к SIRPα, которое связывается с SIRPα или его фрагментом, или его антигенсвязывающий фрагмент. Антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую три CDR, CDR1 VH, CDR2 VH и CDR3 VH, и вариабельную область легкой цепи, содержащую три CDR, CDR1 VL, CDR2 VL и CDR3 VL; причем
CDR1 VH содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 3, 13, 23 или 33, CDR2 VH содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 4, 14, 24 или 34, CDR3 VH содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 5, 15, 25 или 35, CDR1 VL содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 8, 18, 28 или 38, CDR2 VL содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 9, 19, 29 или 39, и CDR3 VL содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 10, 20, 30 или 40.
Согласно предпочтительным вариантам реализации антитело содержит CDR1 VH, CDR2 VH и CDR3 VH и CDR1 VL, CDR2 VL и CDR3 VL, причем
аминокислотная последовательность CDR1 VH представлена в SEQ ID NO: 3;
аминокислотная последовательность CDR2 VH представлена в SEQ ID NO: 4;
аминокислотная последовательность CDR3 VH представлена в SEQ ID NO: 5;
аминокислотная последовательность CDR1 VL представлена в SEQ ID NO: 8;
аминокислотная последовательность CDR2 VL представлена в SEQ ID NO: 9; и
аминокислотная последовательность CDR3 VL представлена в SEQ ID NO: 10.
Согласно предпочтительным вариантам реализации антитело содержит CDR1 VH, CDR2 VH и CDR3 VH и CDR1 VL, CDR2 VL и CDR3 VL, причем
аминокислотная последовательность CDR1 VH представлена в SEQ ID NO: 13;
аминокислотная последовательность CDR2 VH представлена в SEQ ID NO: 14;
аминокислотная последовательность CDR3 VH представлена в SEQ ID NO: 15;
аминокислотная последовательность CDR1 VL представлена в SEQ ID NO: 18;
аминокислотная последовательность CDR2 VL представлена в SEQ ID NO: 19; и
аминокислотная последовательность CDR3 VL представлена в SEQ ID NO: 20.
Согласно предпочтительным вариантам реализации антитело содержит CDR1 VH, CDR2 VH и CDR3 VH и CDR1 VL, CDR2 VL и CDR3 VL, причем
аминокислотная последовательность CDR1 VH представлена в SEQ ID NO: 23;
аминокислотная последовательность CDR2 VH представлена в SEQ ID NO: 24;
аминокислотная последовательность CDR3 VH представлена в SEQ ID NO: 25;
аминокислотная последовательность CDR1 VL представлена в SEQ ID NO: 28;
аминокислотная последовательность CDR2 VL представлена в SEQ ID NO: 29; и
аминокислотная последовательность CDR3 VL представлена в SEQ ID NO: 30.
Согласно предпочтительным вариантам реализации антитело содержит CDR1 VH, CDR2 VH и CDR3 VH и CDR1 VL, CDR2 VL и CDR3 VL, причем
аминокислотная последовательность CDR1 VH представлена в SEQ ID NO: 33;
аминокислотная последовательность CDR2 VH представлена в SEQ ID NO: 34;
аминокислотная последовательность CDR3 VH представлена в SEQ ID NO: 35;
аминокислотная последовательность CDR1 VL представлена в SEQ ID NO: 38;
аминокислотная последовательность CDR2 VL представлена в SEQ ID NO: 39; и
аминокислотная последовательность CDR3 VL представлена в SEQ ID NO: 40.
Согласно некоторым вариантам реализации аминокислотная последовательность вариабельной области тяжелой цепи представлена в SEQ ID NO: 1, 11, 21 или 31, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью.
Согласно некоторым вариантам реализации аминокислотная последовательность вариабельной области легкой цепи представлена в SEQ ID NO: 6, 16, 26 или 36, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью.
Согласно определенным предпочтительным вариантам реализации три CDR в вариабельной области тяжелой цепи и/или три CDR в вариабельной области легкой цепи определены в соответствии с системой нумерации Kabat или Chothia.
Согласно некоторым вариантам реализации антитело или его антигенсвязывающий фрагмент, раскрытый в настоящем документе, могут дополнительно содержать одну или более из константной области тяжелой цепи, константной области легкой цепи и области Fc. Согласно другим предпочтительным вариантам реализации константная область легкой цепи представляет собой константную область λ-цепи или κ-цепи. Согласно некоторым предпочтительным вариантам реализации антитело или его антигенсвязывающий фрагмент принадлежит к типу IgG1, IgG2, IgG3 или IgG4.
Согласно некоторым вариантам реализации антитело или его антигенсвязывающий фрагмент представляет собой химерное антитело или гуманизированное антитело, или его антигенсвязывающий фрагмент.
Согласно второму аспекту предложена молекула нуклеиновой кислоты, содержащая нуклеотиды, кодирующие антитело или его антигенсвязывающий фрагмент, раскрытый в настоящем документе. Согласно некоторым вариантам реализации молекула нуклеиновой кислоты кодирует вариабельную область тяжелой цепи и/или вариабельную область легкой цепи антитела или его антигенсвязывающего фрагмента.
Согласно предпочтительным вариантам реализации молекула нуклеиновой кислоты кодирует вариабельную область тяжелой цепи, и соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 2, 12, 22 или 32, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью. Согласно другим предпочтительным вариантам реализации молекула нуклеиновой кислоты кодирует вариабельную область легкой цепи, и соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 7, 17, 27 или 37, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью.
Согласно третьему аспекту настоящего изобретения предложен биоматериал, который представляет собой:
(1) вектор, клетку-хозяина, микроорганизм или т.п., содержащий молекулу нуклеиновой кислоты, раскрытую в настоящем документе; или
(2) продукт экспрессии, суспензию, супернатант или тому подобное указанного выше (1).
Специалисты в данной области техники смогут легко выбрать и получить векторы, клетки-хозяева или микроорганизмы, содержащие кодирующую последовательность антитела в соответствии с аминокислотной последовательностью антитела, и могут знать, как культивировать такие клетки-хозяева или микроорганизмы для получения соответствующих продуктов экспрессии, суспензий, супернатантов и т. д., чтобы получить соответствующее антитело. Упомянутые методики являются обычными в данной области техники.
Согласно четвертому аспекту предложена композиция, содержащая антитело или его антигенсвязывающий фрагмент, раскрытый в настоящем документе; предпочтительно композиция представляет собой фармацевтическую композицию, дополнительно содержащую фармацевтически приемлемый носитель.
Согласно пятому аспекту предложен способ получения антитела или его антигенсвязывающего фрагмента, раскрытого в настоящем документе, включающий: культивирование указанной выше клетки-хозяина для экспрессии антитела или его антигенсвязывающего фрагмента и выделение антитела или его антигенсвязывающего фрагмента.
Согласно шестому аспекту предложено применение антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, биоматериала или композиции, раскрытых в настоящем документе, в получении лекарственного средства для лечения опухоли; предпочтительно опухоль представляет собой CD47-положительную опухоль; более предпочтительно опухоль представляет собой гематологическую опухоль или солидную опухоль, такую как лейкоз, лимфома, рак мочевого пузыря, рак молочной железы, рак головы и шеи, рак желудка, меланома, рак поджелудочной железы, колоректальный рак, рак пищевода, рак печени, рак почки, рак легкого, рак предстательной железы, рак яичника, рак щитовидной железы, нейроглиома и другие солидные опухоли.
Согласно седьмому аспекту предложено применение антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, биоматериала или композиции, раскрытых в настоящем документе, в получении состава, который блокирует связывание SIRPα с CD47.
Согласно восьмому аспекту предложено применение антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, биоматериала или композиции, раскрытых в настоящем документе, в комбинации с одним или более дополнительными противораковыми терапевтическими агентами в получении лекарственного средства для лечения опухоли; предпочтительно опухоль представляет собой CD47-положительную опухоль.
Согласно некоторым предпочтительным вариантам реализации дополнительный противораковый терапевтический агент включает, но не ограничивается ими, химиотерапевтический агент, радиотерапевтический агент и биомакромолекулярное лекарственное средство. Также предпочтительно биомакромолекулярное лекарственное средство представляет собой лекарственное средство на основе моноклонального антитела, нацеленное на поверхностный антиген опухолевых клеток, включая антитело к CD20 (такое как зуберитамаб или ритуксимаб), цетуксимаб и трастузумаб.
Согласно девятому аспекту предложен способ лечения опухоли, причем указанная опухоль представляет собой CD47-положительную опухоль; предпочтительно опухоль представляет собой гематологическую опухоль или солидную опухоль, такую как лейкоз, лимфома, рак мочевого пузыря, рак молочной железы, рак головы и шеи, рак желудка, меланома, рак поджелудочной железы, колоректальный рак, рак пищевода, рак печени, рак почки, рак легкого, рак предстательной железы, рак яичника, рак щитовидной железы, нейроглиома и другие солидные опухоли; способ включает введение индивидууму, нуждающемуся в этом, антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, биоматериала или композиции, раскрытых в настоящем документе.
Согласно десятому аспекту предложен способ блокирования связывания SIRPα с CD47, включающий применение антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, биоматериала или композиции, раскрытых в настоящем документе.
Согласно одиннадцатому аспекту предложен способ лечения опухоли с помощью комбинированной терапии, причем указанная опухоль представляет собой CD47-положительную опухоль; указанный способ включает введение индивидууму, нуждающемуся в этом, антитела или его антигенсвязывающего фрагмента, молекулы нуклеиновой кислоты, биоматериала или композиции, раскрытых в настоящем документе, и одного или более дополнительных противораковых терапевтических агентов; предпочтительно дополнительный противораковый терапевтический агент включает, но не ограничивается ими, химиотерапевтический агент, радиотерапевтический агент и биомакромолекулярное лекарственное средство; более предпочтительно биомакромолекулярное лекарственное средство представляет собой лекарственное средство на основе моноклонального антитела, нацеленное на поверхностный антиген опухолевых клеток, включая антитело к CD20 (такое как зуберитамаб или ритуксимаб), цетуксимаб и трастузумаб.
Антитело к SIRPα или его антигенсвязывающий фрагмент, раскрытый в настоящем документе, представляет собой блокирующее антитело, которое способно эффективно блокировать связывание SIRPα с CD47, тем самым эффективно способствуя фагоцитозу опухолевых клеток макрофагом, эффективно ингибируя рост опухолей, и обладает хорошей фармацевтической перспективой.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 показано блокирующее действие мышиных моноклональных антител на связывание SIRPα с CD47.
На фиг. 2 показано связывание гуманизированных моноклональных антител с SIRPα.
На фиг. 3 показано блокирующее действие гуманизированных моноклональных антител на связывание SIRPα с CD47.
На фиг. 4A показано блокирующее действие антитела №14 и KWAR23 на связывание SIRPα (V1) с CD47.
На фиг. 4B показано блокирующее действие антитела №14, 18D5 и 1H9 на связывание SIRPα (V1) с CD47.
На фиг. 4C показано блокирующее действие антитела №14, KWAR23, 18D5 и 1H9 на связывание SIRPα (V2) с CD47.
На фиг. 4D показано блокирующее действие антитела №14, KWAR23, 18D5 и 1H9 на связывание SIRPα (V8) с CD47.
На фиг. 5 показана кинетика связывания антитела №14 и KWAR23 с рекомбинантным белком SIRPα.
На фиг. 6А показано стимулирование индуцированного антителом к CD20 фагоцитоза антителом №14; На фиг. 6B показано стимулирование индуцированного антителом к HER2 фагоцитоза антителом №14.
На фиг. 7 показана противоопухолевая эффективность антитела №14 в модели опухоли MC38.
На фиг. 8 показано, что антитело №14 и антитело к CD20 могут синергически ингибировать рост опухоли.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение будет проиллюстрировано со ссылкой на следующие конкретные примеры. Реагенты и инструменты, использованные в следующих процедурах, являются общепринятыми в данной области техники и коммерчески доступными, если не указано иное; все использованные способы являются общепринятыми в данной области техники, и специалисты в данной области техники, несомненно, могут реализовать способы и получить соответствующие результаты в соответствии с описанием примеров.
Определения:
В настоящем изобретении термин «антитело» представляет собой иммуноглобулин, который может специфично распознавать и связывать антиген, и охватывает множество структур антител, включая, но не ограничиваясь ими, моноклональные антитела, поликлональные антитела, биспецифичные антитела или фрагменты антител, синтезированные антитела, рекомбинантно продуцируемые антитела, человеческие антитела, антитела нечеловеческого происхождения (например, мышиные антитела), гуманизированные антитела, химерные антитела и внутриклеточные антитела и фрагменты антител, такие как, но не ограничиваясь перечисленными, фрагменты Fab, фрагменты Fab', фрагменты F(ab')2, фрагменты Fv, фрагменты Fd, фрагменты Fd', одноцепочечные Fv (scFv), одноцепочечные Fab (scFab) или антигенсвязывающие фрагменты любого из указанных выше антител. Антитела, предложенные в настоящем документе, включают членов любого класса (например, IgG, IgM, IgD, IgE, IgA или IgY), любого подкласса (например, IgG1, IgG2, IgG3, IgG4, IgA1 или IgA2) или типа (например, IgG2a или IgG2b) иммуноглобулина. Согласно предпочтительным вариантам реализации антитело, раскрытое в настоящем документе, представляет собой мышиное антитело, химерное антитело или гуманизированное антитело.
В настоящем документе «фрагмент антитела» или «антигенсвязывающий фрагмент» антитела относится к любой части полноразмерного антитела, которая меньше полной длины, но содержит по меньшей мере часть вариабельной области антитела, которая связывается с антигеном (например, один или более CDR и/или один или более связывающих сайтов антитела), и, таким образом, сохраняет специфичность связывания, а также по меньшей мере часть способности к специфичному связыванию полноразмерного антитела. Таким образом, антигенсвязывающий фрагмент относится к фрагменту антитела, который содержит антигенсвязывающую часть, которая связывается с тем же антигеном, что и антитело, из которого получен фрагмент антитела. Фрагменты антител включают производные антител, полученные путем ферментной обработки полноразмерных антител, а также синтезированные производные, например, рекомбинантно полученные производные. Антитела включают фрагменты антител. Примеры фрагментов антител включают, но не ограничиваются ими, Fab, Fab', F(ab')2, одноцепочечный Fv (scFv), Fv, dsFv, диатела, фрагменты Fd и Fd' и другие фрагменты, включая модифицированные фрагменты (см., например, Methods in Molecular Biology, Vol 207: Recombinant Antibodies for Cancer Therapy Methods and Protocols (2003); Chapter 1; p 3-25, Kipriyanov). Фрагмент может содержать множество цепей, связанных друг с другом, например, дисульфидной связью и/или пептидным линкером. Фрагмент антитела обычно содержит по меньшей мере или примерно 50 аминокислот и, как правило, по меньшей мере или примерно 200 аминокислот. Антигенсвязывающий фрагмент включает любой фрагмент антитела, который при вставке в каркас антитела (например, путем замены соответствующей области) дает антитело, которое иммуноспецифично связывается с антигеном.
В настоящем документе «обычное антитело» относится к антителу, содержащему две тяжелые цепи (которые могут быть обозначены как H и H'), две легкие цепи (которые могут быть обозначены как L и L') и два антигенсвязывающих сайта, причем каждая тяжелая цепь может представлять собой полноразмерную тяжелую цепь иммуноглобулина или любую ее функциональную область, которая сохраняет антигенсвязывающую способность (например, тяжелая цепь включает, но не ограничивается этим, цепь VH, цепь VH-CH1 и цепь VH-CH1-CH2-CH3), и каждая легкая цепь может представлять собой полноразмерную легкую цепь или любую ее функциональную область (например, легкая цепь включает, но не ограничивается этим, цепь VL и цепь VL-CL). Каждая тяжелая цепь (H или H') спарена с одной легкой цепью (L или L', соответственно).
В настоящем документе полноразмерное антитело представляет собой антитело, имеющее две полноразмерные тяжелые цепи (например, VH-CH1-CH2-CH3) и две полноразмерные легкие цепи (VL-CL) и шарнирную область, например, антитело, естественно продуцируемое В-клеткой, секретирующей антитело, и синтетически продуцируемое антитело, имеющее такие же домены.
В настоящем документе dsFv относится к Fv со сконструированной межмолекулярной дисульфидной связью, которая стабилизирует пару VH-VL.
В настоящем документе фрагмент Fab представляет собой фрагмент антитела, полученный путем расщепления полноразмерного иммуноглобулина папаином, или фрагмент той же структуры, полученный синтетическим способом, например, рекомбинантным способом. Фрагмент Fab содержит легкую цепь (содержащую VL и CL) и другую цепь, содержащую вариабельный домен тяжелой цепи (VH) и один константный домен тяжелой цепи (CH1).
В настоящей заявке фрагмент F(ab')2 представляет собой фрагмент антитела, полученный путем расщепления иммуноглобулина пепсином при рН 4,0-4,5, или фрагмент той же структуры, полученный синтетическим способом, например, рекомбинантным способом. Фрагмент F(ab')2 по существу содержит два фрагмента Fab, причем каждый фрагмент тяжелой цепи содержит несколько дополнительных аминокислот, включая цистеины, которые образуют дисульфидную связь, связывающую два фрагмента.
В настоящем документе фрагмент Fab' представляет собой фрагмент, который содержит половину фрагмента F(ab')2 (одну тяжелую цепь и одну легкую цепь).
В настоящем документе фрагмент ScFv относится к фрагменту антитела, содержащему вариабельную область легкой цепи (VL) и вариабельную область тяжелой цепи (VH), ковалентно связанные в любом порядке полипептидным линкером.
Термин «вариабельная область» относится к домену тяжелой цепи или легкой цепи антитела, который распознает и специфично связывается с эпитопом антигена, который содержит аминокислотную последовательность, которая варьируется между различными антителами. Каждая легкая цепь или каждая тяжелая цепь имеет домен вариабельной области VL или VH, соответственно. Вариабельные домены обеспечивают антигенную специфичность и, следовательно, отвечают за распознавание антигена. Каждая вариабельная область содержит CDR и каркасные участки (FR), и CDR являются частями домена антигенсвязывающего сайта.
Участок CDR или «определяющий комплементарность участок» относится к участку в вариабельной области антитела, который является гипервариабельным по последовательности и образует петлю с определенной структурой и/или содержит контактирующие с антигеном остатки аминокислот. CDR в первую очередь отвечают за связывание антитела с эпитопом антигена и определяют специфичность антитела. В конкретной аминокислотной последовательности вариабельной области тяжелой цепи или вариабельной области легкой цепи конкретные аминокислотные последовательности CDR определяют с использованием любой одной или комбинации многих хорошо известных схем нумерации, включая, например, Kabat, Contact, AbM и Chothia. CDR антитела согласно настоящему изобретению могут быть определены в соответствии с любой схемой в данной области техники или их комбинацией.
В настоящем документе «гипервариабельный участок», «определяющий комплементарность участок» и «CDR» могут быть использованы взаимозаменяемо и относятся к одной из множества частей в каждой вариабельной области, которые совместно образуют антигенсвязывающие сайты антитела. Каждый домен вариабельной области содержит три CDR, обозначенные CDR1, CDR2 и CDR3. Например, домен вариабельной области легкой цепи содержит три CDR, обозначенные CDR1 VL, CDR2 VL и CDR3 VL; домен вариабельной области тяжелой цепи содержит три CDR, обозначенные H CDR1 (или CDR1 VH), H CDR2 (или CDR2 VH) и H CDR3 (или CDR3 VH). Три CDR в вариабельной области не являются смежными вдоль линейной аминокислотной последовательности, но находятся близко в уложенном полипептиде. CDR расположены в пределах петель параллельных цепей, которые соединяют β-лист вариабельного домена. Как описано в настоящем документе, CDR известны специалистам в данной области техники и могут быть идентифицированы специалистами в данной области техники на основе нумерации Kabat или Chotnia (см., например, Kabat, E.A. et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, AIH Publication No. 91-3242, и Chothia, C. et al. (1987) J. Mol. Biol. 196:901-917).
В настоящем документе каркасный участок (FR) представляет собой домен в пределах домена вариабельной области антитела, который расположен в пределах β-листа; участки FR являются более консервативными по отношению к гипервариабельным участкам с точки зрения аминокислотной последовательности.
В настоящем документе домен «константной области» представляет собой домен в тяжелой цепи или легкой цепи антитела, который содержит аминокислотную последовательность, которая является более консервативной относительно аминокислотной последовательности домена вариабельной области. В молекулах обычных полноразмерных антител каждая легкая цепь имеет один домен константной области легкой цепи (CL), и каждая тяжелая цепь содержит один или более доменов константной области тяжелой цепи (CH), включая CH1, CH2, CH3 и CH4. Константная область антитела может выполнять эффекторные функции, такие как, но не ограничиваясь перечисленными, устранение антигенов, патогенов и токсинов, с которыми антитело специфично связывается, например, путем взаимодействия с различными клетками, биомолекулами и тканями.
В настоящем документе функциональная область домена VH представляет собой по меньшей мере часть интактного домена VH, которая сохраняет по меньшей мере часть специфичности связывания интактного домена VH (например, путем сохранения одного или более CDR интактного домена VH), так что функциональная область домена VH связывается с антигеном отдельно или в комбинации с другим доменом антитела (например, доменом VL) или его областью. Примерные функциональные области доменов VH представляют собой области, содержащие CDR1, CDR2 и/или CDR3 доменов VH.
В настоящем документе функциональная область домена VL представляет собой по меньшей мере часть интактного домена VL, которая сохраняет по меньшей мере часть специфичности связывания интактного домена VL (например, путем сохранения одного или более CDR интактного домена VL), так что функциональная область домена VL связывается с антигеном отдельно или в комбинации с другим доменом антитела (например, доменом VH) или его областью. Примерные функциональные области доменов VL представляют собой области, содержащие CDR1, CDR2 и/или CDR3 доменов VL.
В настоящем документе термины «полинуклеотид» и «молекула нуклеиновой кислоты» относятся к олигомеру или полимеру, содержащему по меньшей мере два связанных нуклеотида или производных нуклеотида, включая дезоксирибонуклеиновую кислоту (ДНК) и рибонуклеиновую кислоту (РНК), которые обычно связаны друг с другом фосфодиэфирными связями.
В настоящем документе выделенная молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая выделена из других молекул нуклеиновой кислоты, присутствующих в природном источнике молекул нуклеиновой кислоты. «Выделенная» молекула нуклеиновой кислоты, такая как молекула кДНК, может быть по существу свободной от другого клеточного материала или культуральной среды при ее получении с помощью рекомбинантных методик, или по существу свободной от химических предшественников или других химических компонентов при ее химическом синтезе. Примерные выделенные молекулы нуклеиновых кислот, предложенные в настоящем документе, включают выделенные молекулы нуклеиновых кислот, кодирующие антитела или антигенсвязывающие фрагменты, предложенные в настоящем документе.
«Идентичность» последовательности имеет значение, хорошо известное в данной области техники, и процент идентичности последовательностей между двумя молекулами нуклеиновой кислоты, пептидными молекулами или областями может быть рассчитан с использованием методик, раскрытых в данной области техники. Идентичность последовательности может быть измерена по всей длине полинуклеотида или полипептида или вдоль области молекулы. (См., например, Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991). Хотя существует множество способов измерения идентичности между двумя полинуклеотидами или полипептидами, термин «идентичность» хорошо известен специалистам (Carrillo, H. & Lipman, D., SIAM J Applied Math 48:1073 (1988)).
В настоящем документе «экспрессия» относится к способу получения полипептида путем транскрипции и трансляции полинуклеотида. Уровень экспрессии полипептида можно оценить с помощью любого способа, известного в данной области техники, включая, например, способы определения количества полипептида, продуцируемого в клетке-хозяине. Такие способы могут включать, но не ограничиваются перечисленными, количественное определение полипептидов в клеточных лизатах с помощью ИФА, окрашивания Кумасси синим после гель-электрофореза, анализа белка по Лоури и анализа белка по Брэдфорд.
В настоящем документе «клетка-хозяин» представляет собой клетку, которая применяется для получения, поддержания, репликации и амплификации вектора. Клетку-хозяина также можно применять для экспрессии полипептида, кодируемого вектором. При делении клетки-хозяина нуклеиновая кислота в векторе реплицируется и, таким образом, амплифицируется. Клетка-хозяин может представлять собой эукариотическую клетку или прокариотическую клетку. Подходящие клетки-хозяева включают, но не ограничиваются перечисленными, клетки CHO, клетки HeLa и клетки HEK, такие как клетки HEK 293.
В настоящем документе «вектор» представляет собой реплицируемую нуклеиновую кислоту, из которой может быть экспрессирован один или более гетерологичных белков при трансформации вектора в подходящую клетку-хозяина. Векторы включают те, в которые может быть введена нуклеиновая кислота, кодирующая полипептид или его фрагмент, как правило, путем расщепления ферментами рестрикции и лигирования. Векторы также включают векторы, содержащие нуклеиновую кислоту, кодирующую полипептид. Векторы используют для введения нуклеиновой кислоты, кодирующей полипептид, в клетки-хозяева для амплификации нуклеиновой кислоты или для экспрессии/экспонирования полипептида, кодируемого нуклеиновой кислотой. Векторы обычно находятся в свободном состоянии, но могут быть сконструированы для интеграции гена или его части в хромосомы генома. Также предусмотрены искусственные хромосомные векторы, такие как искусственные векторы дрожжей и искусственные хромосомы млекопитающих. Специалистам в данной области техники хорошо известно о выборе и применении таких носителей.
В настоящем документе «вектор экспрессии» включает векторы, способные экспрессировать ДНК, функционально связанную с регуляторными последовательностями, такими как промоторные области, способными влиять на экспрессию таких фрагментов ДНК. Такие дополнительные фрагменты могут включать последовательности промотора и терминатора и необязательно могут включать одну или более точек начала репликации, один или более селективных маркеров, энхансеров, сигналов полиаденилирования и тому подобное. Векторы экспрессии обычно получены из плазмид или вирусной ДНК или могут содержать их элементы. Таким образом, вектор экспрессии относится к рекомбинантной конструкции ДНК или РНК, такой как плазмида, фаг, рекомбинантный вирус или другие векторы, которые при введении в соответствующую клетку-хозяина вызывают экспрессию клонированной ДНК. Подходящие векторы экспрессии хорошо известны специалистам в данной области техники и включают векторы экспрессии, реплицируемые в эукариотических клетках и/или прокариотических клетках, а также векторы экспрессии, которые поддерживаются в свободном состоянии, или векторы экспрессии, интегрированные в геном клетки-хозяина.
В настоящем документе «лечение» индивидуума, имеющего заболевание или состояние, означает, что симптомы индивидуума частично или полностью ослаблены или остаются неизменными после лечения. Таким образом, лечение включает предотвращение, лечение и/или излечение. Предотвращение относится к предотвращению основных заболеваний и/или предотвращению ухудшения симптомов или прогрессирования заболевания. Лечение также включает любое фармацевтическое применение любого антитела или его антигенсвязывающего фрагмента, а также композиции, предложенной в настоящем документе.
В настоящем документе «эффективность» означает эффект, возникший в результате лечения индивидуума, который изменяет, как правило, уменьшает или улучшает симптомы заболевания или состояния или излечивает заболевание или состояние.
В настоящем документе «терапевтически эффективное количество» или «терапевтически эффективная доза» относится к количеству вещества, соединения, материала или композиции, содержащей соединение, которое по меньшей мере является достаточным для достижения терапевтического эффекта после введения субъекту. Таким образом, это количество, необходимое для предотвращения, излечения, улучшения, блокирования или частичного блокирования симптомов заболевания или состояния. Также в настоящем документе «профилактически эффективное количество» или «профилактически эффективная доза» относится к количеству вещества, соединения, материала или композиции, содержащей соединение, которое при введении субъекту будет обладать желаемым профилактическим эффектом, например, предотвращение или отсрочка возникновения или рецидива заболевания или состояния, или уменьшение возможности возникновения или рецидива заболевания или состояния. Полная профилактически эффективная доза не обязательно достигается введением одной дозы, но может быть достигнута после введения ряда доз. Таким образом, профилактически эффективное количество может быть введено посредством одной или более доз.
В настоящем документе термин «индивидуум» относится к млекопитающему, такому как человек.
Антитело к SIRPα, раскрытое в настоящем документе
Согласно настоящему изобретению предложено антитело к SIRPα, обладающее высокой аффинностью в отношении человеческого белка SIRPα. Антитело может эффективно ингибировать связывание SIRPα с его лигандом CD47, тем самым блокируя нижележащую передачу сигналов CD47/SIRPα, стимулируя фагоцитоз опухолевых клеток макрофагами и дополнительно осуществляя устранение опухолевых клеток. При связывании с белком SIRPα на поверхности клетки антитело к SIRPα, раскрытое в настоящем документе, может вызывать эндоцитоз, что приводит к снижению экспрессии белка SIRPα на поверхности клетки, тем самым дополнительно снижая передачу сигналов CD47/SIRPα.
Антитело к SIRPα или его антигенсвязывающий фрагмент, раскрытый в настоящем документе, содержит замену, вставку или делецию. Антитело к SIRPα, раскрытое в настоящем документе, содержит модификацию вариабельной области легкой цепи, вариабельной области тяжелой цепи, легкой цепи или тяжелой цепи, и их модифицированная аминокислотная последовательность отличается от аминокислотной последовательности, из которой получено антитело. Например, аминокислотная последовательность, полученная из одного и того же конкретного белка, может быть подобна исходной последовательности, например, может иметь определенный процент идентичности. Например, она может иметь 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% идентичности с исходной последовательностью.
В настоящем документе «идентичность» относится к проценту идентичных оснований (или аминокислот) в двух сравниваемых последовательностях при выравнивании последовательностей двух пептидов или двух молекул нуклеиновой кислоты. Такое выравнивание и процент гомологии или идентичности последовательности можно определить с применением программного обеспечения, известного в данной области техники, такого как описанное в Ausubel et al., eds., (2007), Current Protocols in Molecular Biology. Предпочтительно выравнивание выполняют с использованием параметров по умолчанию. Одним из примеров программ для выравнивания является BLAST с использованием параметров по умолчанию. В частности, программа BLASTN или BLASTP.
Согласно определенным вариантам реализации в область Fc антитела, предложенного в настоящем документе, может быть введена одна или более аминокислотных модификаций с созданием тем самым варианта Fc. Вариант Fc может содержать последовательность области Fc человека, содержащую аминокислотную модификацию в одном или более аминокислотных положениях.
Подходящие «антитела и их антигенсвязывающие фрагменты» в настоящем изобретении включают, но не ограничиваются перечисленными, поликлональные, моноклональные, моновалентные, биспецифичные, мультиспецифичные, рекомбинантные, гетерологичные, химерные, гуманизированные или деиммуногенные антитела или фрагменты Fab, фрагменты Fab', фрагменты F(ab')2, одноцепочечные антитела, нанотела и эпитопсвязывающие фрагменты любого из указанных выше.
Согласно некоторым вариантам реализации антитело, раскрытое в настоящем документе, может быть моноспецифичным, биспецифичным или полиспецифичным. Антитело к SIRPα может быть лигировано с другим антителом или фрагментом антитела с получением биспецифичного или мультиспецифичного антитела со второй или большим числом видов специфичности связывания.
Согласно определенным вариантам реализации антитело может быть дополнительно модифицировано для добавления функциональных компонентов. Подходящие фрагменты для дериватизации антитела включают, но не ограничиваются перечисленными, ПЭГ, декстран, белки, липиды, терапевтические агенты и токсины. Антитело может быть модифицировано путем фосфорилирования, ацетилирования, гликозилирования, пегилирования, амидирования, лигирования с другими белками или т.п.
Лечение опухоли и применение антитела
Антитело к SIRPα и его антигенсвязывающий фрагмент и содержащую их фармацевтическую композицию, предложенные в настоящем документе, можно применять для диагностики, прогнозирования, лечения или ингибирования разных видов рака. Настоящее изобретение относится к способу лечения рака у субъекта путем введения антитела к SIRPα или его фрагмента, раскрытого в настоящем документе, субъекту, нуждающемуся в этом. Терапевтические соединения, раскрытые в настоящем документе, включают, но не ограничиваются перечисленными, антитела, раскрытые в настоящем документе (включая варианты и производные, раскрытые в настоящем документе), и нуклеиновые кислоты или полинуклеотиды, кодирующие антитела, раскрытые в настоящем документе (включая варианты и производные, раскрытые в настоящем документе).
Согласно настоящему изобретению также предложена комбинированная терапия, включающая комбинированное применение антитела к SIRPα, раскрытого в настоящем документе, и по меньшей мере одного дополнительного терапевтического агента, включая, но не ограничиваясь перечисленными, химиотерапевтический агент, радиотерапевтический агент и биомакромолекулярное лекарственное средство. Согласно одному варианту реализации биомакромолекулярное лекарственное средство представляет собой лекарственное средство на основе моноклонального антитела, нацеленное на поверхностный антиген опухолевых клеток, включая антитело к CD20 (такое как зуберитамаб или ритуксимаб), цетуксимаб и трастузумаб.
Антитело, раскрытое в настоящем документе (и любые дополнительные терапевтические агенты), можно вводить любым подходящим способом, включая, но не ограничиваясь перечисленными, внутрибрюшинную, внутривенную, подкожную, интраназальную и внутримышечную инъекцию. Антитело и его вариант или композицию можно вводить любым удобным путем, например, с помощью болюсной инъекции или инфузии, или всасывания через эпителий или слизистую оболочку кожи.
Последовательности антитела к SIRPα, приведенные в настоящем документе в качестве примера
Пример 1: Получение мышиных моноклональных антител к SIRPα
В данном примере описано получение мышиных моноклональных антител к человеческому SIRPα с применением гибридомной технологии. Внеклеточную область белка SIRPα (Uniprot: P78324) экспрессировали как иммуноген. Fc-метку IgG1 человека сливали с C-концом аминокислотной последовательности (Glu31-Arg370) внеклеточной области белка SIRPα. Затем ген клонировали в вектор V152 (поставленный Huabio) с получением вектора V152-SIRPα ECD-Fc. Вектор временно трансфицировали в клетки 293 (поставленные ATCC). Через пять дней супернатант клеточной культуры собирали и очищали с помощью белка A (производитель: Solarbio, № по каталогу I8090). Для получения мышиных моноклональных антител к человеческому SIRPα мышей BALB/c в возрасте 4 недель (поставленных Huabio) иммунизировали 100 мкг белка SIRPα. В дни 14 и 28 после первой иммунизации иммунизированных мышей повторно иммунизировали 50 мкг белка SIRPα. Сывороточный титр у иммунизированных мышей измеряли с помощью ИФА. 96-луночные планшеты покрывали SIRPα (1 мкг/мл) в ФСБ (производитель: ZSGB-BIO, № по каталогу ZLI-9062) при 4°C в течение ночи. Планшеты блокировали 1% блокирующим раствором БСА-ФСБ при 37°C в течение 1 часа и промывали ФСБТ. Разведения сыворотки от иммунизированных мышей добавляли в планшеты, и планшеты инкубировали в течение 1 часа при 37°C. Планшеты промывали и добавляли ПХ-меченное козье антитело к мышиному IgG (производитель: Abcam, № по каталогу Ab205719, разведение 1:10000). После 0,5 ч инкубации при 37°C планшеты промывали и добавляли раствор TMB (производитель: InnoReagents, № по каталогу TMB-S-00). После 5 мин инкубации при комнатной температуре в темноте реакцию останавливали добавлением 2 н. H2SO4. Поглощение считывали при 450 нм на считывающем устройстве для микропланшетов. Мышей с достаточными титрами антитела к SIRPα иммунизировали 50 мкг белка SIRPα в качестве бустерной дозы на день 42. Через 3-5 дней мышей умерщвляли. Клетки селезенки собирали, промывали 2-3 раза основной средой IMDM (производитель: BasalMedia, № по каталогу L610KJ) путем центрифугирования, а затем смешивали с клетками миеломы мыши SP2/0 (поставлены Huabio) в соотношении 1:1. ПЭГ (производитель: Roche, № по каталогу 25771700) добавляли к перемешанным клеткам, и смесь осторожно перемешивали и оставляли постоять при 37°С в течение 30 с. Слитые клетки разводили в среде для отбора IMEM, содержащей 15% фетальной бычьей сыворотки (производитель: Tianhang Biotechnology, № по каталогу 11011-8611) и 1× HAT (производитель: Sigma, № по каталогу H0262-1VL), добавляли в 96-луночный планшет для культивирования клеток при 200 мкл/лунку и инкубировали в инкубаторе с 5% СО2 при 37°С. Через 10-14 дней антитело к SIRPα в супернатанте гибридомных клеток детектировали с помощью ИФА. Одиннадцать различных гибридомных клонов, 10F11-5-6, 1B6-1-1, 27A11-1-8, 14B11-5-4, 31D4-4-5, 7C2-3-8, 30C1-6-4, 4A3-5-1, 6H1-8-6, 9A12-5 и 10F4-15, идентифицировали и подвергали дальнейшему анализу.
Пример 2: Связывание мышиных антител с SIRPα
Способность связывания мышиных антител с белком SIRPα детектировали с помощью ИФА. Конкретный способ соответствует тому, который описан в примере 1. Мышиные антитела 3-кратно последовательно разводили ФСБ из 1 мкг/мл и получали в общей сложности семь градиентов концентрации. Результаты представлены в Таблице 5.
Связывание мышиных моноклональных антител с мембранным SIRPα на моноцитах миелоидного лейкоза человека (THP-1, поставленных Банком клеток Китайской академии наук), эндогенно экспрессирующих SIRPα, анализировали с помощью проточной цитометрии (FACS). Клетки THP-1 инкубировали совместно с мышиными моноклональными антителами в различных концентрациях (3-кратное последовательное разведение из 1 мкг/мл, в общей сложности семь градиентов концентрации) в течение 30 мин при 4°С. Клетки дважды промывали перед добавлением FITC-меченного козьего антитела к мышиному IgG (производитель: Jackson, № по каталогу 115-095-003, разведение 1:1000), а затем инкубировали при 4°С в течение 30 мин в темноте. После двукратной промывки клетки детектировали на проточном цитометре BD C6. Результаты представлены в Таблице 6.
Пример 3: Блокирующее действие мышиных антител к SIRPα на связывание SIRPα с CD47
В соответствии с результатами Примера 2, при комбинировании способности связывания мышиного моноклонального антитела к SIRPα и способности связывания с клетками THP-1 МАТ №4, МАТ №11, МАТ №13 и МАТ №14 представляют собой молекулы с наилучшей активностью. Таким образом, МАТ №4, МАТ №11, МАТ №13 и МАТ №14 отобрали для определения того, могут ли эти мышиные МАТ блокировать связывание SIRPα с CD47. 96-луночные планшеты покрывали CD47-Fc (производитель: Acrobiosystems, № по каталогу CD7-H5256) при 4°C в течение ночи. Планшеты промывали и блокировали 1% блокирующим раствором БСА-ФСБТ. После 1 ч инкубации при 37°C планшеты промывали, и в планшеты добавляли антитела в различных концентрациях (3-кратное разведение из 66,7 нМ, в общей сложности семь градиентов концентрации) и меченный биотином SIRPα (производитель: Acrobiosystems, № по каталогу CDA-H82F2). После 1 ч инкубации при 37°C планшеты промывали и добавляли меченный ПХ стрептавидин (производитель: Abcam, № по каталогу: ab7403). После 0,5 ч инкубации при 37°C планшеты промывали и добавляли раствор TMB. После 0,5 ч инкубации при комнатной температуре в темноте реакцию останавливали добавлением 2 н. H2SO4. Поглощение считывали при 450 нм на считывающем устройстве для микропланшетов. В качестве контроля использовали IgG (производитель: GenScript). Результаты, как показано на фиг. 1, демонстрируют, что все указанные мышиные антитела могут ингибировать связывание SIRPα с CD47 в различной степени.
Пример 4: Гуманизация мышиных антител
На основании результатов Примера 2 и Примера 3 для гуманизации отобрали МАТ №4, МАТ № 11, МАТ № 13 и МАТ №14. Вариабельные области тяжелой и легкой цепей этих антител сравнивали с доступной базой данных последовательностей генов IgG человека для выявления оптимальных совпадений с последовательностями генов Ig зародышевой линии человека. В частности, тяжелые цепи IgG зародышевой линии человека, отобранные для МАТ №4, МАТ №11, МАТ №13 и МАТ №14, представляли собой IGHV1-69-2*01, IGHV2-26*01, IGHV7-4-1*01 и IGHV4-31*02, соответственно, и легкие цепи IgG зародышевой линии человека, отобранные для МАТ №4, МАТ №11, МАТ №13 и МАТ №14, представляли собой IGKV4-1*01, IGKV1-9*01, IGKV4-1*01 и IGKV4-1*011, соответственно. CDR тяжелой цепи МАТ №4, МАТ №11, МАТ №13 и МАТ №14 прививали в каркасный участок тяжелых цепей IgG зародышевой линии человека, и CDR легкой цепи МАТ №4, МАТ №11, МАТ №13 и МАТ №14 прививали в каркасный участок легких цепей IgG зародышевой линии человека. Новые гуманизированные молекулы обозначали №4, №11, №13 и №14, соответственно. Последовательности вариабельной области тяжелой цепи, вариабельной области легкой цепи и участков CDR сконструированных гуманизированных антител представлены в Таблице 3. Последовательности тяжелой и легкой цепей антитела №14 представлены в Таблице 4. Константные области тяжелой и легкой цепей антител №4, №11 и №13 соответствуют константным областям антитела №14. Полные аминокислотные последовательности тяжелых и легких цепей антител №4, №11 и №13 известны специалистам в данной области техники на основании информации, раскрытой выше.
Антитела, раскрытые в настоящем документе, экспрессировали в клетках 293 (поставленных ATCC) и очищали. Кодирующие последовательности тяжелой и легкой цепей антитела клонировали в вектор V152 (поставленный Huabio). Вектор V152, несущий кодирующие последовательности тяжелых и легких цепей антител, переносили в клетки 293 с использованием реагента для трансфекции ПЭИ (производитель: Polyscience, № по каталогу 23966-2). Плазмидную ДНК и реагент для трансфекции готовили в шкафу биологической безопасности. Плазмидную ДНК и ПЭИ (массовое соотношение = 0,15:1,75) равномерно смешивали, а затем оставляли на 10 мин. Смесь осторожно добавляли к 293 клеткам и осторожно перемешивали. После культивирования клеток при 37°С и 5% CO2 в течение 5 дней супернатант клеточной культуры центрифугировали при 3000 об./мин в течение 10 мин. Супернатант очищали с помощью белка A (производитель: Solarbio, № по каталогу I8090), чтобы чистота антитела превышала 95%.
Пример 5: Связывание гуманизированных антител с белком SIRPα
Связывание гуманизированных антител с белком SIRPα детектировали с помощью ИФА. 96-луночные планшеты покрывали SIRPα (производитель: Acrobiosystems, № по каталогу SIA-H5225) (1 мкг/мл) в ФСБ при 4°C в течение ночи. Затем планшеты блокировали 1% блокирующим раствором БСА-ФСБ в течение 1 часа при температуре 37°C. Планшеты промывали ФСБТ. Гуманизированные антитела разводили до различных концентраций (3-кратное последовательное разведение из 66,7 нМ, в общей сложности 11 градиентов концентрации) и добавляли к планшетам. После 1 ч инкубации при 37°C планшеты промывали и добавляли ПХ-меченное козье антитело к IgG человека (производитель: Abcam, № по каталогу Ab98595, разведение 1:10000). После 0,5 ч инкубации при 37°C планшеты промывали и добавляли раствор TMB. После 5 мин инкубации при комнатной температуре в темноте реакцию останавливали добавлением 2 н. H2SO4. Поглощение считывали при 450 нм на считывающем устройстве для микропланшетов. Результаты показаны на Фиг. 2. Значения EC50 для связывания с SIRPα антител №4, №11, №13 и №14 составили 1,833 нМ, 0,4642 нМ, 0,9517 нМ и 0,6831 нМ, соответственно, это указывает на то, что все эти гуманизированные молекулы обладают высокой активностью связывания с SIRPα.
Пример 6: Блокирующее действие гуманизированных антител к SIRPα на связывание SIRPα с CD47
Блокирующее действие гуманизированных антител к SIRPα №4, №11, №13 и №14 на связывание SIRPα с CD47 тестировали, как описано в примере 3. Антитела 3-кратно последовательно разводили из 66,7 нМ и получали в общей сложности 11 градиентов концентрации. Результаты показаны на Фиг. 3. Значения IC50 антител №4, №11, №13 и №14 составили 6,03 нМ, 0,2086 нМ, 1,5 нМ и 0,1315 нМ, соответственно, это свидетельствует о том, что все указанные гуманизированные антитела могут эффективно блокировать связывание SIRPα с CD47, при этом антитела №11 и №14 оказывают более сильное блокирующее действие, чем два других гуманизированных антитела.
В качестве контролей использовали KWAR23, 18D5 и 1H9. KWAR23 получали в соответствии с последовательностями, раскрытыми в US2018037652 (SEQ ID NO: 1 и SEQ ID NO: 2), 18D5 получали в соответствии с последовательностями, раскрытыми в US20190127477 (SEQ ID NO: 24 и SEQ ID NO: 31), и 1H9 получали в соответствии с последовательностями, раскрытыми в US20190119396 (SEQ ID NO: 7 и SEQ ID NO: 8). Поскольку человеческий белок SIRPα является высокополиморфным во внеклеточной области, а распространенные генотипы включают V1, V2 и V8, провели дополнительное сравнение блокирующего действия антитела №14 и вышеуказанных контрольных антител на связывание распространенных генотипов SIRPα с CD47 (в других примерах все белки SIRPα без указанного генотипа относятся к генотипу V1).
Блокирующее действие различных антител на связывание SIRPα (V1) с CD47 тестировали, как описано в примере 3. Антитела 2-кратно последовательно разводили из 83,4 нМ и получали в общей сложности девять градиентов концентрации. Результаты показаны на Фиг. 4А. Значение IC50 антитела №14 составило 3,404 нМ, и значение IC50 KWAR23 составило 13,54 нМ, это указывает на то, что антитело №14 оказывает более сильное блокирующее действие на связывание SIRPα (V1) с CD47, чем KWAR23. На фиг. 4B значения IC50 антитела №14, 18D5 и 1H9 составили 4,4 нМ, 4,1 нМ и 4,2 нМ, соответственно, это указывает на то, что блокирующее действие антитела №14, 18D5 и 1H9 на связывание SIRPα (V1) с CD47 было подобным.
Блокирующее действие различных антител на связывание SIRPα (V2) с CD47 определяли способом, описанным в примере 3, в котором использованный меченный биотином SIRPα представлял собой SIRPα (V2) (производитель: Acrobiosystems, № по каталогу SI2-H82W8). Антитела 3-кратно последовательно разводили из 200 нМ и получали в общей сложности девять градиентов концентрации. Результаты показаны на Фиг. 4C. Антитело №14, KWAR23 и 1H9 может блокировать связывание SIRPα (V2) с CD47, в то время как 18D5 не может блокировать связывание SIRPα (V2) с CD47. Блокирующая активность антитела №14 сильнее, чем у 1H9. Значение IC50 антитела №14 составило 7,8 нМ, и значение IC50 KWAR23 составило 6,4 нМ, это указывает на то, что антитело №14 и KWAR23 обладают сходной блокирующей активностью в отношении SIRPα (V2).
Блокирующее действие различных антител на связывание SIRPα (V8) с CD47 определяли методом, описанным в примере 3, в котором использованный меченный биотином SIRPα представлял собой SIRPα (V8) (производитель: Acrobiosystems, № по каталогу SI8-H82W6). Антитела 3-кратно последовательно разводили из 200 нМ и получали в общей сложности девять градиентов концентрации. Результаты показаны на Фиг. 4D. Антитело №14, KWAR23 и 1H9 могут блокировать связывание SIRPα (V8) с CD47, в то время как 18D5 не может блокировать связывание SIRPα (V8) с CD47. Блокирующая активность антитела №14 сильнее, чем у 1H9. Значение IC50 антитела №14 составило 11,5 нМ, и значение IC50 KWAR23 составило 10,8 нМ, это указывает на то, что антитело №14 и KWAR23 обладают сходной блокирующей активностью в отношении SIRPα (V8).
Согласно приведенным выше результатам антитело №14 может блокировать связывание распространенных генотипов SIRPα (V1, V2 и V8) с CD47 и обладает более сильной блокирующей активностью в отношении SIRPα (V1), чем KWAR23, и более сильной блокирующей активностью в отношении SIRPα (V2) и SIRPα (V8), чем 18D5 и 1H9.
Пример 7. Кинетика связывания антител с SIRPα
Кинетику связывания антитела с антигеном определяли с использованием системы Octet Red96 (производитель: ForteBio), оснащенной биосенсором H1S1K (производитель: ForteBio). Биосенсор использовали для непосредственного захвата антигена, а затем сразу же погружали в образец аналита (антитела). Эксперимент состоял из пяти процедур: 1, базовая линия (100 с), 2, загрузка (захват антигена SIRPα) (500 с), 3, базовая линия (100 с), 4, ассоциация (связывание с антителом) (500 с), 5, диссоциация (диссоциация антитела) (1000 с). После завершения теста сенсор регенерировали путем чередования замачивания в буфере для регенерации (глицин, pH 1,5) и буфере для нейтрализации (ФСБ) в течение 5 секунд, и в общей сложности выполняли пять циклов для регенерации сенсора. Рабочий буфер в этом эксперименте представлял собой ФСБ.
Обработка образца: антиген, белок SIRPα (производитель: Acrobiosystems, № по каталогу SIA-H5225), разводили до рабочей концентрации 2,5 мкг/мл с использованием рабочего буфера, а образцы аналита (антитело №14 и KWAR23) последовательно разводили до пяти рабочих концентраций: 1 мкг/мл, 0,5 мкг/мл, 0,25 мкг/мл, 0,125 мкг/мл и 0,0625 мкг/мл. Для анализа данных значения сигнала отклика (сигнал образца связанного аналита с вычтенным сигналом холостого образца аналита) рассчитывали с использованием анализа данных Octet (версия 7.0 или последняя), и данные аппроксимировали с использованием модели ассоциации 1:1.
Диаграмма аппроксимации кинетики связывания представлена на фиг. 5, и константы ассоциации, диссоциации и равновесия представлены в таблице 7. Можно видеть, что антитело №14 обладает очень высокой аффинностью, которая сравнима с аффинностью антитела для положительного контроля KWAR23.
Пример 8: Стимулирующее действие антител к SIRPα, раскрытых в настоящем документе, на фагоцитоз опухолевых клеток макрофагами
Клетки МКПК (производитель: Allcells, № по каталогу PB004F-C) культивировали в основной среде 1640 (производитель: Gibco, № по каталогу 22400-089) при 37°C и 5% CO2 в течение 2-3 ч. Несвязанные клетки удаляли с помощью пипетки и добавляли индуцирующую среду (80 нг/мл M-CSF) для индукции. Вносили свежую среду, содержащую достаточное количество цитокинов, каждые 3 дня. Клетки культивировали в течение 7 дней с получением макрофагов. Клетки Raji (поставленные Банком клеток Китайской академии наук) флуоресцентно метили в соответствии с инструкцией для реагента CFSE (производитель: Abcam, № по каталогу AB113853). Меченые клетки-мишени Raji и вышеуказанные дифференцированные макрофаги смешивали равномерно в соотношении 3:1, и добавляли антитело к CD20 (зуберитамаб, CAS RN: 2251143-19-6, WHO Drug Information, Vol. 33, No.4, 2019, 914-915; концентрация: 0,05 мкг/мл, 0,5 мкг/мл; производитель: BioRay), комбинацию антитела к CD20 (концентрация: 0,05 мкг/мл, 0,5 мкг/мл) и антитела №14 (концентрация: 10 мкг/мл) или комбинацию антитела к CD20 (концентрация: 0,05 мкг/мл, 0,5 мкг/мл) и антитела KWAR23 (концентрация: 10 мкг/мл). Клетки культивировали при температуре 37°C и 5% CO2 в течение 4 ч. Затем клетки промывали ФСБ и добавляли APC-конъюгированное антитело к CD14 (производитель: BD Biosciences, № по каталогу 555399). После 30 мин инкубации при 4°C в темноте клетки промывали и анализировали с помощью проточной цитометрии. Скорость фагоцитоза (АЗКФ) вычисляли по следующей формуле: скорость фагоцитоза (%) = соотношение (APC + CFSE) положительных клеток/соотношение APC-положительных клеток × 100%. На фиг. 6A показано, что антитело №14 может усиливать фагоцитоз клеток Raji, индуцированный антителом к CD20.
Также изучали стимулирующий эффект комбинации антитела №14 и антитела к Her2 на фагоцитоз макрофагами клеток SK-BR-3, сверхэкспрессирующих Her2. В соответствии с способом, описанным выше, меченные клетки SK-BR-3 (поставленные Банком клеток Китайской академии наук) и дифференцированные макрофаги смешивали равномерно в соотношении 2:1, и добавляли антитело к Her2 (концентрация: 0,5 мкг/мл; производитель: BioRay) или комбинацию антитела к Her2 (концентрация: 0,5 мкг/мл) и антитела №14 (концентрация: 10 мкг/мл). Клетки культивировали при 37°С и 5% CO2 в течение 2 ч. Скорость фагоцитоза анализировали и рассчитывали с помощью проточной цитометрии. На фиг. 6B показано, что антитело №14 может усиливать фагоцитоз клеток SK-BR-3, индуцированный антителом к Her2.
Пример 9: Антитело к SIRPα, раскрытое в настоящем документе, индуцирует эндоцитоз SIRPα на поверхности клеток
Клетки THP-1 (поставленные Банком клеток Китайской академии наук) инкубировали совместно с 10 мкг/мл антитела при 4°С в течение 30 мин. Клетки дважды промывали и разделяли на две части. Одну часть инкубировали при 37°С в течение 4 ч, а другую часть дополнительно инкубировали при 4°С в качестве контроля. Клетки дважды промывали и добавляли FITC-меченное козье антитело к IgG человека (производитель: Abcam, № по каталогу ab97224). После 30 мин инкубации при 4°C в темноте клетки промывали и затем анализировали с помощью проточной цитометрии. Скорость эндоцитоза рассчитывали по уменьшению флуоресценции клеточной поверхности при 37°С по сравнению с 4°С. Скорость эндоцитоза вычисляли по следующей формуле: скорость эндоцитоза (%) = (интенсивность флуоресценции MFI при 4°C - интенсивность флуоресценции MFI при 37°C)/интенсивность флуоресценции MFI при 4°C × 100%.
В таблице 8 показано, что скорость эндоцитоза для антитела №14 составляла 52,9%, в то время как скорость эндоцитоза для KWAR23 составляла 34,7%, это указывает на то, что антитело №14 более эффективно снижает уровень экспрессии SIRPα на клеточной поверхности.
Пример 10: Ингибирование роста опухоли, индуцированное антителом к SIRPα, раскрытым в настоящем документе
Противоопухолевую эффективность антитела №14 оценивали в модели рака толстой кишки hCD47-MC38 на мышах. Клетки hCD47-MC38 (поставленные Shanghai Model Organisms) инокулировали мышам C57BL/6 с двойной гуманизацией по hSIRPα/hCD47 (поставленные Shanghai Model Organisms). Когда объем опухоли достиг примерно 150 мм3, мышам вводили антитело №14 (200 мкг/мышь) путем внутрибрюшинной инъекции дважды в неделю, и контрольная группа получала эквивалентные количества ФСБ. Результаты показаны на Фиг. 7. Антитело №14 может непрерывно ингибировать рост опухоли по сравнению с контрольной группой.
Также оценивали противоопухолевую эффективность комбинации антитела №14 и антитела к CD20 (зуберитамаб, поставленный компанией BioRay) в модели лимфомы. Клетки Raji-Luc (поставленные Biocytogen) инокулировали мышам B-NDG-hSIRPα (поставленным Biocytogen) через хвостовые вены. Когда средняя интенсивность сигнала визуализации опухоли достигала примерно 1×106 имп./с, мышам вводили контрольный IgG (поставленный Biocytogen, 200 мкг/мышь), только антитело №14 (200 мкг/мышь), только антитело к CD20 (2 мкг/мышь) или антитело №14 (200 мкг/мышь) плюс антитело к CD20 (2 мкг/мышь) в указанных дозах. Как показано на фиг. 8, антитело №14 и антитело к CD20 обладают синергической противоопухолевой эффективностью.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> BioRay Pharmaceutical Co., Ltd.
Hisun Biopharmaceutical Co., Ltd.
<120> АНТИТЕЛО К SIRPα И ЕГО ПРИМЕНЕНИЕ
<130> ESP1V230753ZX-RU
<150> CN 202011464010.2
<151> 2020-12-11
<160> 42
<170> PatentIn, версия 3.5
<210> 1
<211> 113
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VH
<400> 1
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Arg Gly Ser Gly Phe Asn Ile Lys Asp Tyr
20 25 30
Tyr Ile Gln Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asp Pro Glu Asn Gly Asp Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Tyr Ala Lys Gly Pro Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
100 105 110
Ser
<210> 2
<211> 339
<212> ДНК
<213> искусственная последовательность
<220>
<223> #4 VH ДНК
<400> 2
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggcgccac cgtgaagatc 60
tcctgccgcg gctccggctt caacatcaag gactactaca tccagtgggt gcagcaggcc 120
cccggcaagg gcctggagtg gatgggctgg atcgaccccg agaacggcga caccaagtac 180
gcccccaagt tccagggccg cgtgaccatc accgccgaca cctccaccga caccgcctac 240
atggagctgt cctccctgcg ctccgaggac accgccgtgt actactgcta cgccaagggc 300
ccctactggg gccagggcac caccctgacc gtctcgagc 339
<210> 3
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VH-CDR1
<400> 3
Asp Tyr Tyr Ile Gln
1 5
<210> 4
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VH-CDR2
<400> 4
Trp Ile Asp Pro Glu Asn Gly Asp Thr Lys Tyr Ala Pro Lys Phe Gln
1 5 10 15
Gly
<210> 5
<211> 4
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VH-CDR3
<400> 5
Lys Gly Pro Tyr
1
<210> 6
<211> 113
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VL
<400> 6
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 7
<211> 339
<212> ДНК
<213> искусственная последовательность
<220>
<223> #4 VL ДНК
<400> 7
gacatcgtga tgacccagtc ccccgactcc ctggccgtgt ccctgggcga gcgcgccacc 60
atcaactgca agtcctccca gtccctgttc tactcctcca accagaagaa cttcctggcc 120
tggtaccagc agaagcccgg ccagcccccc aagctgctga tctactgggc ctccacccgc 180
gagtccggcg tgcccgaccg cttcaccggc tccggctccg gcaccgactt caccctgacc 240
atctcctccg tgaaggccga ggacctggcc gtgtactact gccagcagta ctactcctac 300
ccccccacct tcggcggcgg caccaagctc gagataaaa 339
<210> 8
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VL-CDR1
<400> 8
Lys Ser Ser Gln Ser Leu Phe Tyr Ser Ser Asn Gln Lys Asn Phe Leu
1 5 10 15
Ala
<210> 9
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VL-CDR2
<400> 9
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 10
<211> 9
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #4 VL-CDR3
<400> 10
Gln Gln Tyr Tyr Ser Tyr Pro Pro Thr
1 5
<210> 11
<211> 118
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VH
<400> 11
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser His
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Ser Asp Gly Ser Thr Thr Tyr Asn Ser Thr Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg His Gly Asn Tyr His Tyr Asn Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 12
<211> 354
<212> ДНК
<213> искусственная последовательность
<220>
<223> #11 VH ДНК
<400> 12
caggtgaccc tgaaggagtc cggccccgtg ctggtgaagc ccaccgagac cctgaccctg 60
acctgcaccg tgtccggctt ctccctgtcc tcccacggcg tgcactggat ccgccagccc 120
cccggcaagg ccctggagtg gctggccgtg atctggtccg acggctccac cacctacaac 180
tccaccctga agtcccgcct gaccatctcc aaggacacct ccaagtccca ggtggtgctg 240
accatgacca acatggaccc cgtggacacc gccacctact actgcgcccg ccacggcaac 300
taccactaca acatggacta ctggggccag ggcaccctgg tgaccgtctc gagc 354
<210> 13
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VH-CDR1
<400> 13
Ser His Gly Val His
1 5
<210> 14
<211> 16
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VH-CDR2
<400> 14
Val Ile Trp Ser Asp Gly Ser Thr Thr Tyr Asn Ser Thr Leu Lys Ser
1 5 10 15
<210> 15
<211> 10
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VH-CDR3
<400> 15
His Gly Asn Tyr His Tyr Asn Met Asp Tyr
1 5 10
<210> 16
<211> 108
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VL
<400> 16
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Ser Arg Val Asn Ser Gly
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Thr Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Tyr Pro
85 90 95
Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 17
<211> 324
<212> ДНК
<213> искусственная последовательность
<220>
<223> #11 VL ДНК
<400> 17
gacatccagc tgacccagtc cccctccttc ctgtccgcct ccgtgggcga ccgcgtgacc 60
atcacctgcc gcacctcctc ccgcgtgaac tccggctacc tgcactggta ccagcagaag 120
cccggcaagg cccccaagct gctgatctac tccacctcca ccctggcctc cggcgtgccc 180
tcccgcttct ccggctccgg ctccggcacc gagttcaccc tgaccatctc ctccctgcag 240
cccgaggact tcgccaccta ctactgccag cagtactcct cctacccctg gaccttcggc 300
ggcggcacca agctcgagat aaaa 324
<210> 18
<211> 12
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VL-CDR1
<400> 18
Arg Thr Ser Ser Arg Val Asn Ser Gly Tyr Leu His
1 5 10
<210> 19
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VL-CDR2
<400> 19
Ser Thr Ser Thr Leu Ala Ser
1 5
<210> 20
<211> 9
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #11 VL-CDR3
<400> 20
Gln Gln Tyr Ser Ser Tyr Pro Trp Thr
1 5
<210> 21
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VH
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Cys Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Gly Pro Leu Tyr Arg Tyr Asp Gly Tyr Gly Leu Glu Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 22
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> #13 VH ДНК
<400> 22
caggtgcagc tggtgcagtc cggctccgag ctgaagaagc ccggcgcctc cgtgaaggtg 60
tcctgcaagg cctccggcta caccttcacc gactactcca tccactgggt gcgccaggcc 120
cccggccagg gcctggagtg gatgggctgg atcaacaccg agaccggcga gcccacctac 180
gccgacgact tcaagggccg cttcgtgttc tccctggaca cctccgtgtc caccgcctac 240
ctgcagatct gctccctgaa ggccgaggac accgccgtgt actactgctc ccgcggcccc 300
ctgtaccgct acgacggcta cggcctggag tactggggcc agggcaccct ggtgaccgtc 360
tcgagc 366
<210> 23
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VH-CDR1
<400> 23
Asp Tyr Ser Ile His
1 5
<210> 24
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VH-CDR2
<400> 24
Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 25
<211> 13
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VH-CDR3
<400> 25
Gly Pro Leu Tyr Arg Tyr Asp Gly Tyr Gly Leu Glu Tyr
1 5 10
<210> 26
<211> 107
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VL
<400> 26
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Cys Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Glu Asp Val Ala Val Tyr Tyr Cys His Gln Asp Tyr Ile Ser Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 27
<211> 321
<212> ДНК
<213> искусственная последовательность
<220>
<223> #13 VL ДНК
<400> 27
gacatcgtga tgacccagtc ccccgactcc ctggccgtgt ccctgggcga gcgcgccacc 60
atcaactgca aggcctccca gtccgtgtcc aacgacgtgg cctggtacca gcagaagccc 120
ggccagcccc ccaagctgct gatctactac gcctccaacc gctgcaccgg cgtgcccgac 180
cgcttctccg gctccggctc cggcaccgac ttcaccctga ccatctcctc cctgcaggcc 240
gaggacgtgg ccgtgtacta ctgccaccag gactacatct ccccctacac cttcggcggc 300
ggcaccaagc tcgagataaa a 321
<210> 28
<211> 11
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VL-CDR1
<400> 28
Lys Ala Ser Gln Ser Val Ser Asn Asp Val Ala
1 5 10
<210> 29
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VL-CDR2
<400> 29
Tyr Ala Ser Asn Arg Cys Thr
1 5
<210> 30
<211> 9
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #13 VL-CDR3
<400> 30
His Gln Asp Tyr Ile Ser Pro Tyr Thr
1 5
<210> 31
<211> 118
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VH
<400> 31
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Tyr Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Asp Gly Ser Arg Tyr Asn Asn Pro Ser Leu
50 55 60
Lys Asn Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Glu Tyr Ala Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 32
<211> 354
<212> ДНК
<213> искусственная последовательность
<220>
<223> #14 VH ДНК
<400> 32
caggtgcagc tgcaggagag cggaccagga ctggtgaagc ctagccagac actgtctctg 60
acctgcacag tgagcggcgg ctctatctcc agcggctact attggtcttg gatcagacag 120
cacccaggca agggcctgga gtggatcggc tacatctcct atgatggcag caggtataac 180
aatccttccc tgaagaaccg ggtgaccatc tctgtggaca catccaagaa tcagttcagc 240
ctgaagctgt cttccgtgac cgccgctgat acagccgtgt actattgcgc tcgcgaggag 300
tacgccaact atttcgctta ctggggccag ggcaccacag tgaccgtgag ctct 354
<210> 33
<211> 6
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VH-CDR1
<400> 33
Ser Gly Tyr Tyr Trp Ser
1 5
<210> 34
<211> 16
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VH-CDR2
<400> 34
Tyr Ile Ser Tyr Asp Gly Ser Arg Tyr Asn Asn Pro Ser Leu Lys Asn
1 5 10 15
<210> 35
<211> 9
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VH-CDR3
<400> 35
Glu Glu Tyr Ala Asn Tyr Phe Ala Tyr
1 5
<210> 36
<211> 113
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VL
<400> 36
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 37
<211> 339
<212> ДНК
<213> искусственная последовательность
<220>
<223> #14 VL ДНК
<400> 37
gatatcgtga tgacccagtc tcctgactcc ctggccgtga gcctgggcga gagagctaca 60
atcaactgta agtccagcca gtctctgttc tactcttcca accagaagaa ttttctggcc 120
tggtatcagc agaagcccgg ccagccccct aagctgctga tctactgggc tagcaccaga 180
gagtctggag tgcctgaccg cttcaccgga tccggaagcg gaacagactt caccctgaca 240
atcagctctg tgaaggccga ggatctggcc gtgtactatt gccagcagta ctattcttat 300
ccacccacct tcggccaggg cacaaagctc gagatcaag 339
<210> 38
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VL-CDR1
<400> 38
Lys Ser Ser Gln Ser Leu Phe Tyr Ser Ser Asn Gln Lys Asn Phe Leu
1 5 10 15
Ala
<210> 39
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VL-CDR2
<400> 39
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 40
<211> 9
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 VL-CDR3
<400> 40
Gln Gln Tyr Tyr Ser Tyr Pro Pro Thr
1 5
<210> 41
<211> 448
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 HC
<400> 41
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Tyr Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Asp Gly Ser Arg Tyr Asn Asn Pro Ser Leu
50 55 60
Lys Asn Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Glu Tyr Ala Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 42
<211> 220
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> #14 LC
<400> 42
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНСВЯЗЫВАЮЩИЙ ПОЛИПЕПТИД, СВЯЗЫВАЮЩИЙСЯ С CD47, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2833294C2 |
| АНТИ-SIRPα АНТИТЕЛА | 2018 |
|
RU2771174C2 |
| СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2812280C1 |
| СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2773758C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2020 |
|
RU2814713C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2016 |
|
RU2732924C2 |
| АНТИТЕЛО ДЛЯ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА 4 | 2017 |
|
RU2774446C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2019 |
|
RU2796019C2 |
| УСОВЕРШЕНСТВОВАННЫЕ ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2019 |
|
RU2810748C2 |
| АНТИТЕЛО ПРОТИВ TNFR2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2793165C1 |
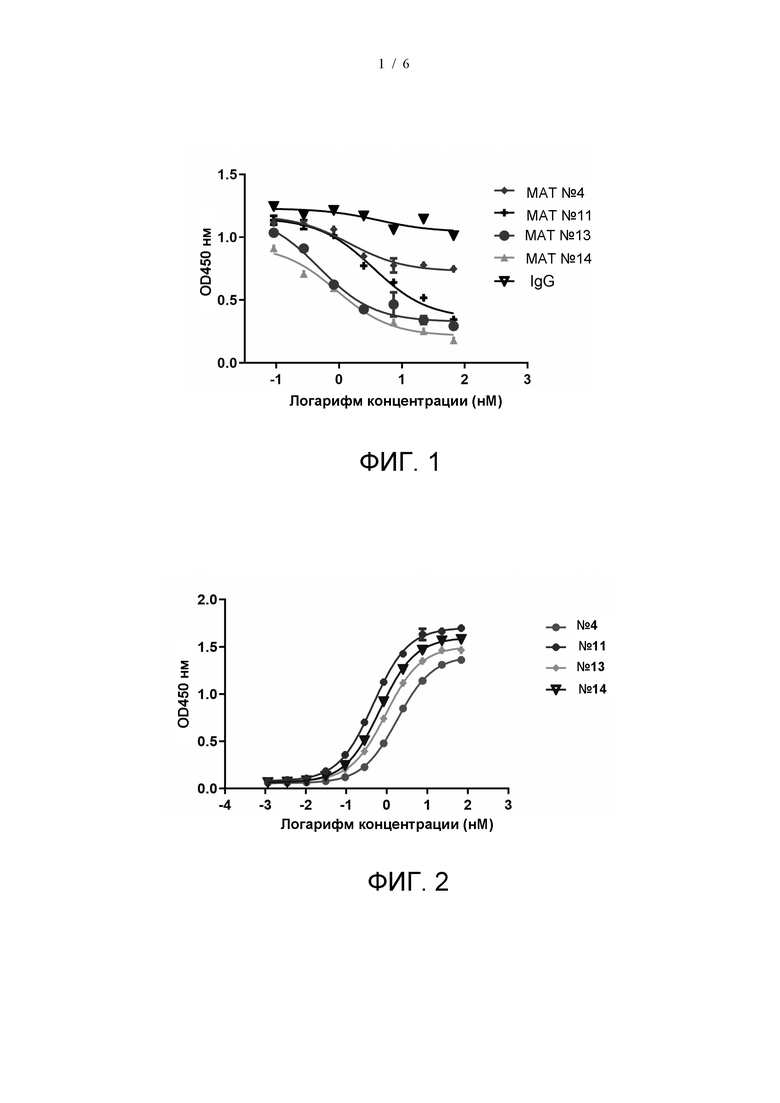
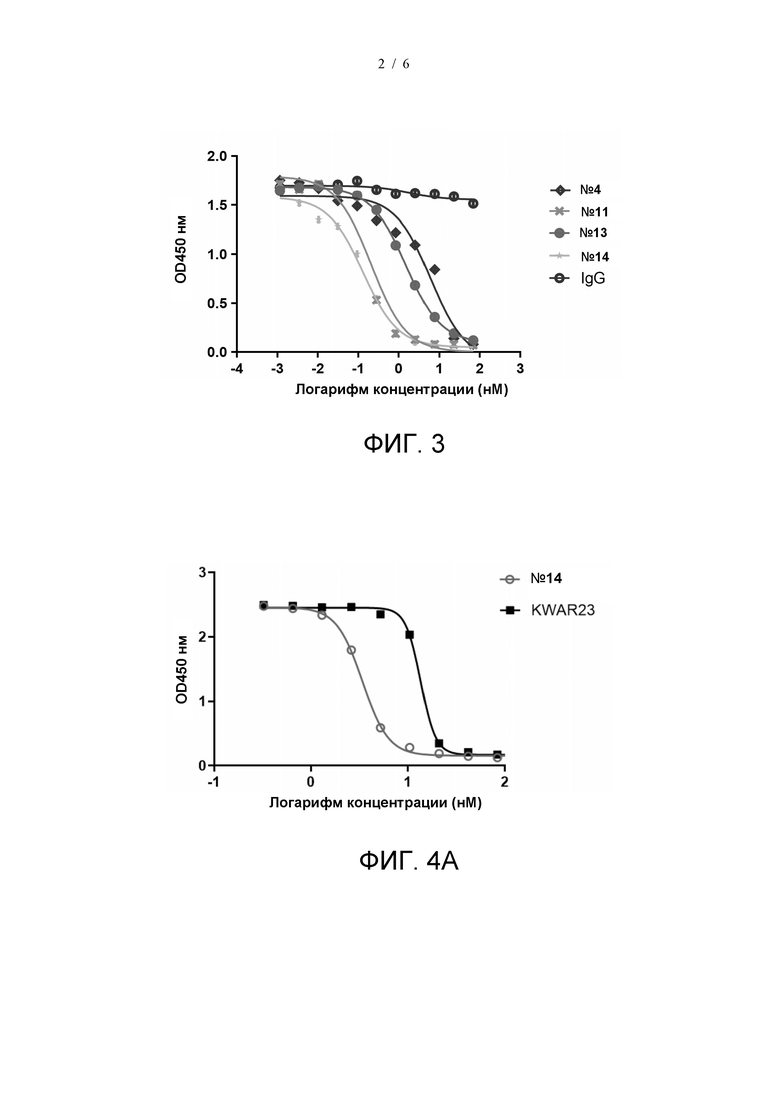
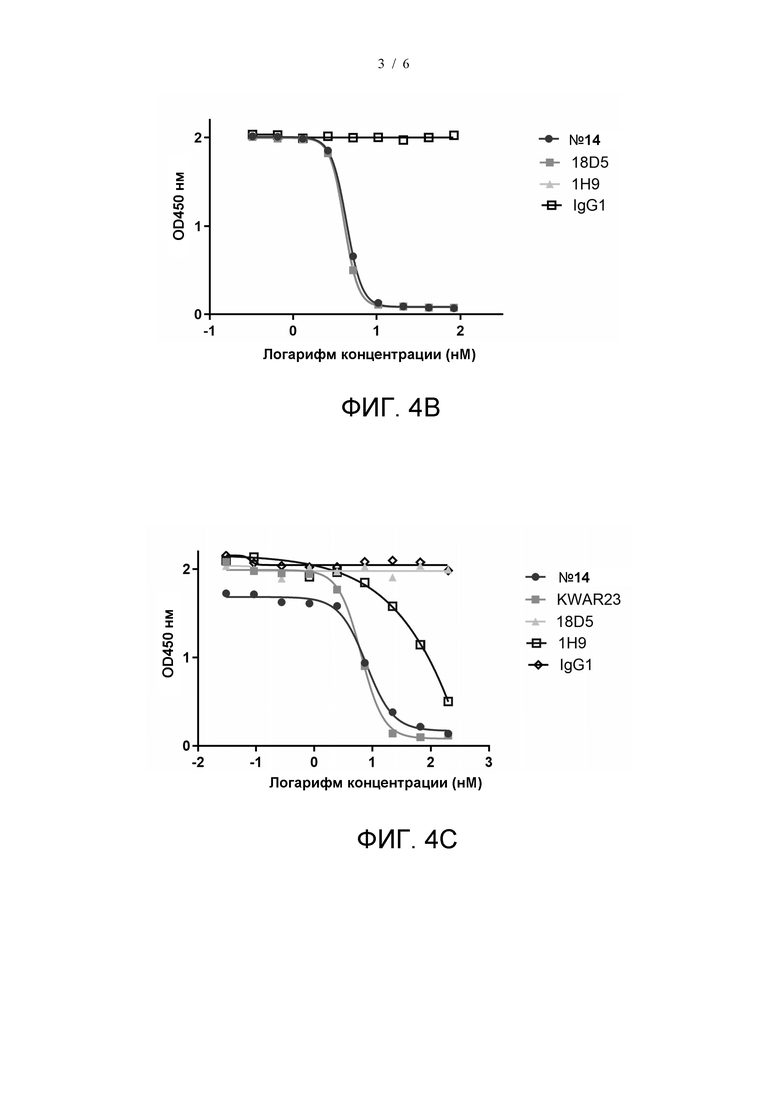
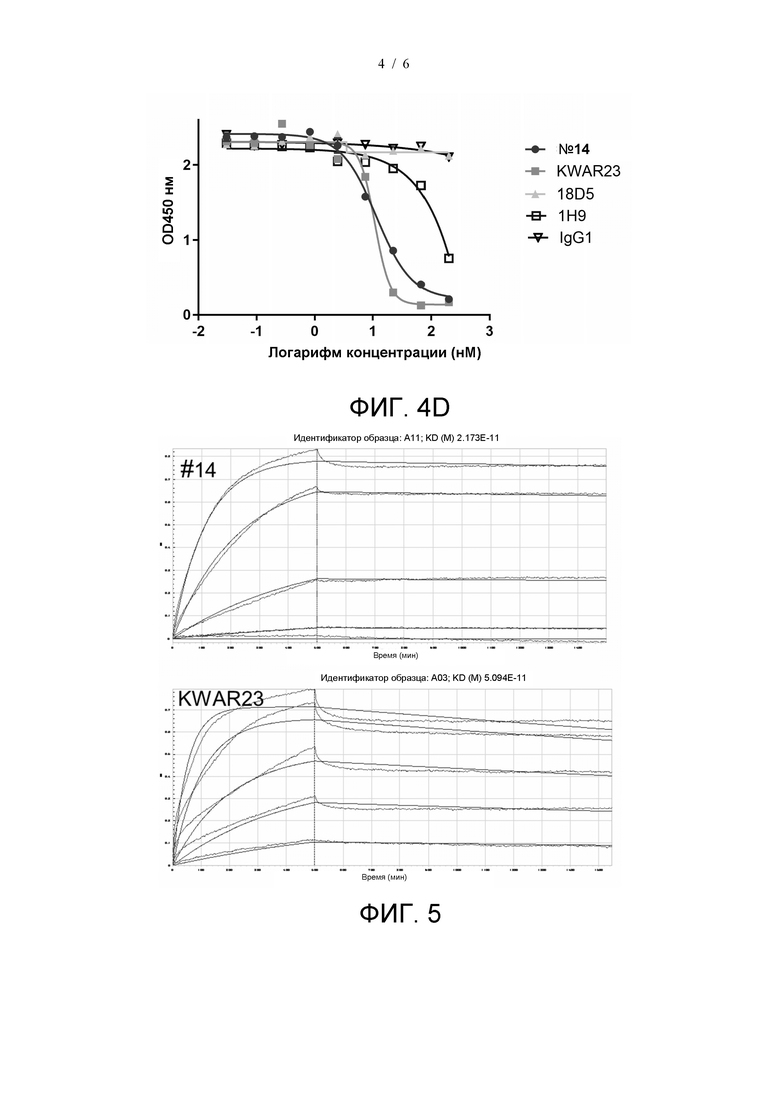
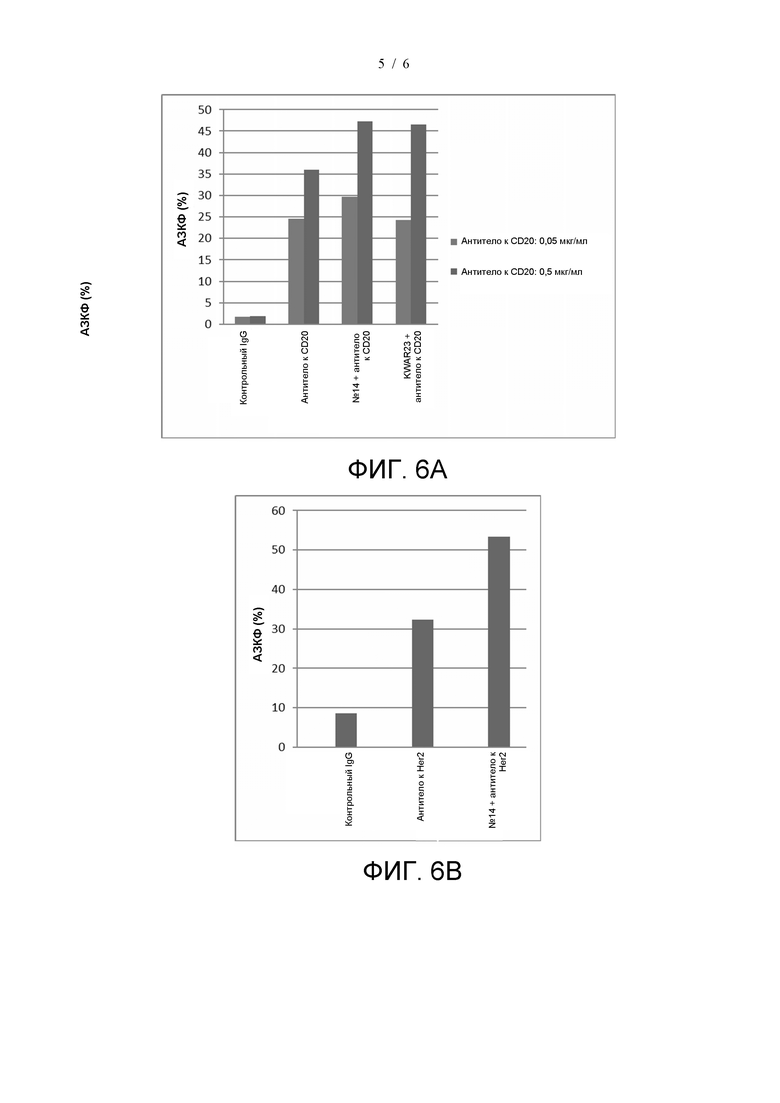
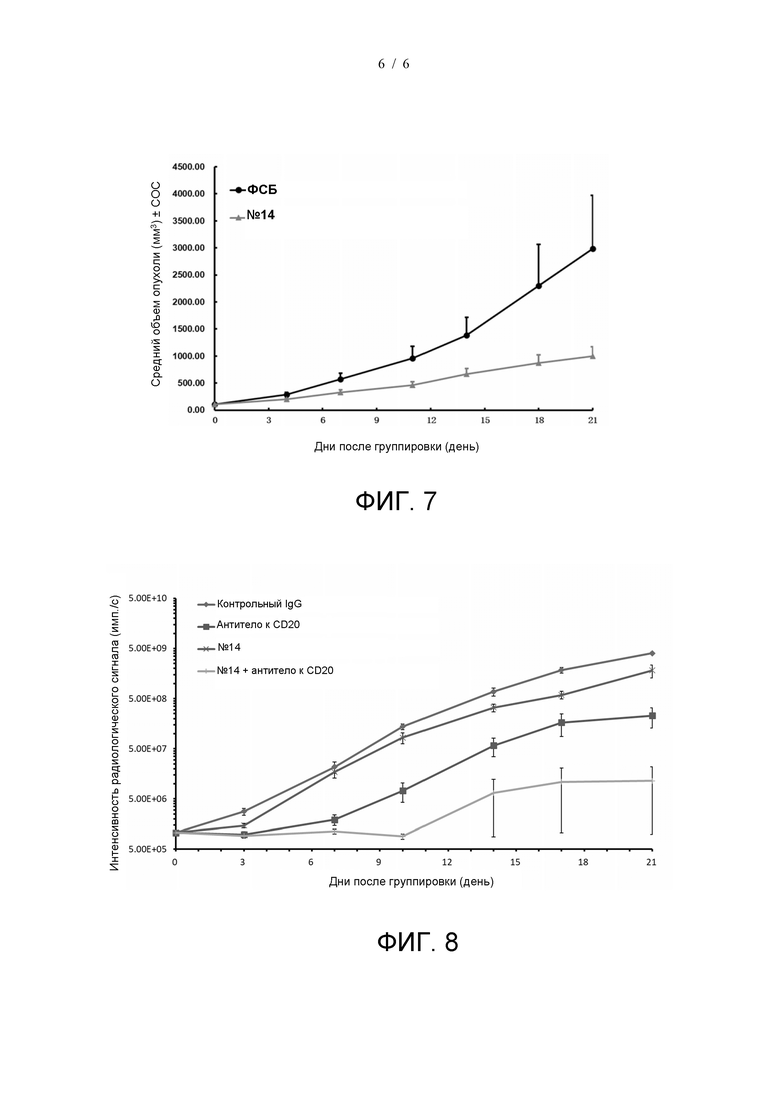
Изобретение относится к области биотехнологии. Предложены антитело к SIRPα или его антигенсвязывающий фрагмент, способные эффективно блокировать связывание SIRPα и CD47, тем самым эффективно стимулируя фагоцитоз опухолевых клеток макрофагами, эффективно ингибируя рост опухолей, и обладающие хорошей фармацевтической перспективой. Также изобретение относится к нуклеиновой кислоте, кодирующей указанные антитело или его фрагмент, способу получения антитела или его фрагмента, фармацевтической композиции, а также к применению антитела или его фрагмента для лечения опухоли. 7 н. и 15 з.п. ф-лы, 12 ил., 8 табл., 10 пр.
1. Антитело к SIRPα или его антигенсвязывающий фрагмент, содержащие: вариабельную область тяжелой цепи, содержащую три определяющих комплементарность участка (CDR), CDR1 VH, CDR2 VH и CDR3 VH, и вариабельную область легкой цепи, содержащую три CDR, CDR1 VL, CDR2 VL и CDR3 VL, причем
аминокислотная последовательность указанного CDR1 VH представлена в SEQ ID NO: 3;
аминокислотная последовательность указанного CDR2 VH представлена в SEQ ID NO: 4;
аминокислотная последовательность указанного CDR3 VH представлена в SEQ ID NO: 5;
аминокислотная последовательность указанного CDR1 VL представлена в SEQ ID NO: 8;
аминокислотная последовательность указанного CDR2 VL представлена в SEQ ID NO: 9; и
аминокислотная последовательность указанного CDR3 VL представлена в SEQ ID NO: 10;
или
аминокислотная последовательность указанного CDR1 VH представлена в SEQ ID NO: 13;
аминокислотная последовательность указанного CDR2 VH представлена в SEQ ID NO: 14;
аминокислотная последовательность указанного CDR3 VH представлена в SEQ ID NO: 15;
аминокислотная последовательность указанного CDR1 VL представлена в SEQ ID NO: 18;
аминокислотная последовательность указанного CDR2 VL представлена в SEQ ID NO: 19; и
аминокислотная последовательность указанного CDR3 VL представлена в SEQ ID NO: 20;
или
аминокислотная последовательность указанного CDR1 VH представлена в SEQ ID NO: 23;
аминокислотная последовательность указанного CDR2 VH представлена в SEQ ID NO: 24;
аминокислотная последовательность указанного CDR3 VH представлена в SEQ ID NO: 25;
аминокислотная последовательность указанного CDR1 VL представлена в SEQ ID NO: 28;
аминокислотная последовательность указанного CDR2 VL представлена в SEQ ID NO: 29; и
аминокислотная последовательность указанного CDR3 VL представлена в SEQ ID NO: 30;
или
аминокислотная последовательность указанного CDR1 VH представлена в SEQ ID NO: 33;
аминокислотная последовательность указанного CDR2 VH представлена в SEQ ID NO: 34;
аминокислотная последовательность указанного CDR3 VH представлена в SEQ ID NO: 35;
аминокислотная последовательность указанного CDR1 VL представлена в SEQ ID NO: 38;
аминокислотная последовательность указанного CDR2 VL представлена в SEQ ID NO: 39; и
аминокислотная последовательность указанного CDR3 VL представлена в SEQ ID NO: 40.
2. Антитело к SIRPα или его антигенсвязывающий фрагмент по п. 1, отличающиеся тем, что указанная аминокислотная последовательность вариабельной области тяжелой цепи представлена в SEQ ID NO: 1, 11, 21 или 31, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью; указанная аминокислотная последовательность вариабельной области легкой цепи представлена в SEQ ID NO: 6, 16, 26 или 36, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью.
3. Антитело к SIRPα или его антигенсвязывающий фрагмент по п. 1 или 2, дополнительно содержащие одно или более из константной области тяжелой цепи, константной области легкой цепи и области Fc.
4. Антитело к SIRPα или его антигенсвязывающий фрагмент по п. 3, отличающиеся тем, что указанная константная область легкой цепи представляет собой константную область λ-цепи или κ-цепи.
5. Антитело к SIRPα или его антигенсвязывающий фрагмент по п. 3, отличающиеся тем, что указанное антитело или его антигенсвязывающий фрагмент относится к типу IgG1, IgG2, IgG3 или IgG4.
6. Антитело к SIRPα или его антигенсвязывающий фрагмент по п. 3, отличающиеся тем, что указанное антитело или его антигенсвязывающий фрагмент представляет собой химерное антитело или гуманизированное антитело, или его антигенсвязывающий фрагмент.
7. Молекула нуклеиновой кислоты, содержащая нуклеотиды, кодирующие антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6.
8. Молекула нуклеиновой кислоты по п. 7, отличающаяся тем, что указанная молекула нуклеиновой кислоты кодирует вариабельную область тяжелой цепи и/или вариабельную область легкой цепи указанного антитела или его антигенсвязывающего фрагмента.
9. Молекула нуклеиновой кислоты по п. 8, отличающаяся тем, что указанная молекула нуклеиновой кислоты кодирует вариабельную область тяжелой цепи, и соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 2, 12, 22 или 32, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью; или указанная молекула нуклеиновой кислоты кодирует вариабельную область легкой цепи, и соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 7, 17, 27 или 37, или имеет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с указанной последовательностью.
10. Фармацевтическая композиция для лечения опухоли, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6 и фармацевтически приемлемый носитель.
11. Способ получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6, включающий: культивирование клетки-хозяина, содержащей молекулу нуклеиновой кислоты по любому из пп. 7-9, для экспрессии указанного антитела или его антигенсвязывающего фрагмента и выделение указанного антитела или его антигенсвязывающего фрагмента.
12. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6 или композиции по п. 10 в получении лекарственного средства для лечения опухоли.
13. Применение по п. 12, отличающееся тем, что указанная опухоль представляет собой CD47-положительную опухоль.
14. Применение по п. 12, отличающееся тем, что указанная опухоль представляет собой гематологическую опухоль или солидную опухоль.
15. Применение по п. 14, отличающееся тем, что указанная опухоль представляет собой лейкоз, лимфому, рак мочевого пузыря, рак молочной железы, рак головы и шеи, рак желудка, меланому, рак поджелудочной железы, колоректальный рак, рак пищевода, рак печени, рак почки, рак легкого, рак предстательной железы, рак яичника, рак щитовидной железы, нейроглиому и другие солидные опухоли.
16. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6 или композиции по п. 10 в получении состава, который блокирует связывание SIRPα с CD47.
17. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6 или композиции по п. 10 в комбинации с одним или более дополнительными противораковыми терапевтическими агентами в получении лекарственного средства для лечения опухоли.
18. Применение по п. 17, отличающееся тем, что указанная опухоль представляет собой CD47-положительную опухоль.
19. Применение по п. 17, отличающееся тем, что указанный дополнительный противораковый терапевтический агент включает, но не ограничивается перечисленными, химиотерапевтический агент, радиотерапевтический агент и биомакромолекулярное лекарственное средство.
20. Применение по п. 19, отличающееся тем, что указанное биомакромолекулярное лекарственное средство представляет собой лекарственное средство на основе моноклонального антитела, нацеленное на поверхностный антиген опухолевых клеток.
21. Применение по п. 20, отличающееся тем, что указанное лекарственное средство на основе моноклонального антитела представляет собой антитело к CD20, цетуксимаб или трастузумаб.
22. Применение по п. 21, отличающееся тем, что указанное антитело к CD20 представляет собой зуберитамаб или ритуксимаб.
| WO 2018190719 A2, 18.10.2018 | |||
| TADAHIKO YANAGITA et al | |||
| Anti-SIRPα antibodies as a potential new tool for cancer immunotherapy, JCI Insight, 2017 Jan 12; 2(1): e89140 | |||
| VOETS E | |||
| et al | |||
| Железобетонная шпала | 1924 |
|
SU1805A1 |

