Настоящее изобретение относится к моноклональному антителу человека, специфичному в отношении альфа-токсина S. aureus, гибридоме, продуцирующей указанное антитело, нуклеиновым кислотам, кодирующим его, а также к клеткам-хозяевам, трансфецированным указанными нуклеиновыми кислотами. Кроме того, настоящее изобретение относится к способам получения указанного моноклонального антитела. Дополнительно, настоящее изобретение относится к фармацевтическим композициям, содержащим по меньшей мере одно антитело или по меньшей мере одну нуклеиновую кислоту, кодирующую указанное антитело. Настоящее изобретение также относится к применению указанного моноклонального антитела для лечения или предотвращения образования абсцесса.
Золотистый стафилококк (S. aureus) - это факультативно анаэробная грамположительная сферическая бактерия, которая считается оппортунистическим патогеном. S. aureus обычно колонизирует нос, кожу и слизистую оболочку желудочно-кишечного тракта здоровых людей. В любой момент времени приблизительно 20-30% населения инфицированы S. aureus. У здоровых людей эти бактерии часто вызывают вторичные инфекции, такие как прыщи и фурункулы. Как правило, слизистые оболочки и эпидермальные барьеры (кожа) защищают от инфицирования S. aureus. Нарушение целостности этих естественных барьеров в результате повреждений, таких как ожоги, травмы или хирургические операции, значительно увеличивает риск инфицирования и может стать причиной тяжелых и/или системных заболеваний. Болезни, которые подавляют иммунную систему (например, сахарный диабет, поздние стадии почечной недостаточности, рак, СПИД и другие вирусные инфекции), а также иммуносупрессивные виды терапии, например, такие как облучение, химиотерапия и трансплантационная терапия, увеличивают риск инфекции. Оппортунистические инфекции, вызванные S. aureus, могут стать довольно серьезными и индуцировать развитие эндокардита, бактериемии, остеомиелита и образование абсцессов, которые, в свою очередь, могут быть причиной серьезных заболеваний и смерти. Стафилококковые инфекции могут быть разделены на локализованные инфекции, такие как пневмония, и клинически более сложные стафилококковые инфекции, такие как сепсис и образование абсцессов, вызванное диссеминацией патогенна в отдаленный орган.
Во всем мире S. aureus является главной причиной сепсиса, инфекций кожи, мягких тканей, а также нижних дыхательных путей. На протяжении многих лет количество случаев заболеваемости как внутрибольничными, так и внебольничными инфекциями неуклонно растет. Помимо этого, лечение подобных инфекций осложнилось появлением штаммов с множественной лекарственной устойчивостью. В развитых странах, таких как США, устойчивость метициллин-резистентных штаммов золотистого стафилококка (МРЗС) к антибиотикам β-лактамного ряда является серьезной проблемой в больницах и других медицинских учреждениях. Важно, что заболеваемость всеми инвазивными инфекциями, вызываемыми МРЗС, включая внебольничные инфекции, является высокой по сравнению с другими бактериальными патогенами, и 20% этих инфекций приводят к смерти. Дополнительно, появление приобретенной устойчивости к ванкомицину дополнительно ограничило возможности лечения тяжелых стафилококковых инфекций.
S. aureus имеет разнообразный арсенал факторов вирулентности, вносящих вклад в патогенез заболевания. Их можно грубо разделить на поверхностные белки и белки, секретируемые во внешнюю среду. Поверхностные белки включают как структурные компоненты клеточной стенки бактерий, такие как пептидогликан и липотейхоевые кислоты, так и поверхностные белки, преимущественно экспрессируемые в течение фазы экспоненциального роста, включая белок А, фибронектин-связывающий белок и фактор слипания. Секретируемые белки, как правило, выделяются из бактериальных клеток во время стационарной фазы роста бактерий и включают несколько токсинов, таких как альфа-токсин (также известный как альфа-гемолизин), энтеротоксин В, лейкоцидины (включая лейкоцидин Пантон-Валентина, ПВЛ), липазу и протеазу V8. Однако, несмотря на большой объем знаний о биохимических и молекулярных свойствах этих токсинов, их точная роль в патогенезе инфекций, вызванных S. aureus, не ясна до конца.
Экспериментальные подтверждения и эпидемиологические данные позволяют предположить, что альфа-токсин может быть вовлечен в патогенез пневмонии наряду с другими цитотоксинами (Mс Elroy et al., 1999). Предполагают, что альфа-токсин связывается с поверхностными рецепторами восприимчивых клеток-хозяев и таким образом прикрепляется к поверхности клетки. Это событие способствует олигомеризации токсина в гептамерную структуру, формирующую раннюю пору, с последующим встраиванием β-цилиндрической структуры с диаметром поры 2 нм в плазматическую мембрану. Образование поры приводит к потере целостности мембраны, дестабилизирует клетки и, в конечном итоге, инициирует апоптоз и лизис клетки. К образованию поры, индуцированному альфа-токсином, особенно чувствительны такие клетки, как лимфоциты, макрофаги, альвеолярные эпителиальные клетки, легочный эндотелий и эритроциты. В то же время гранулоциты и фибробласты, по-видимому, устойчивы к лизису (Mc Elroy et al., 1999).
Точная роль альфа-токсина в воспалительной реакции и индукции врожденного иммунного ответа на бактериальные инфекции понятна не полностью. S. aureus экспрессирует ряд других факторов вирулентности, и на сегодняшний день вклад каждого фактора вирулентности в проявление болезни понятен не до конца и представляет собой задачу для разработки профилактики и лечения клинически сложных инфекций, вызванных золотистым стафилококком.
Как известно, альфа-токсин является одним из факторов вирулентности при развитии у хозяина инфекции, индуцированной S. aureus, и в ряде исследований подчеркивалась важность альфа-токсина в течении болезни, например, инстилляция очищенного альфа-токсина кролику или в легочные ткани крысы вызывает сосудистую утечку и легочную гипертензию, которую связывают с высвобождением различных сигнальных молекул (например, фосфатидилинозитола, оксида азота, простагландинов и тромбоксана А2). В литературе было показано, что невосприимчивость к альфа-токсину защищает от вредного воздействия токсинов, однако разработка вакцин против альфа-токсина остается серьезной проблемой.
Wardenburg и Schneewind (2008) показали, что тяжесть болезни легких у мышей коррелирует с уровнями альфа-токсина, вырабатываемого определенным изолятом золотистого стафилококка. Кроме того, авторы показали, что иммунизация против варианта альфа-токсина, неспособного к формированию поры, индуцирует развитие невосприимчивости к пневмонии, вызываемой S. aureus. Эти данные согласуются с результатами исследования той же группы, демонстрирующими, что альфа-токсин важен для патогенеза внебольничной пневмонии, вызываемой (метициллин-резистентный золотистый стафилококк). В другом эксперименте авторы показали, что антитела к альфа-токсину также защищали эпителиальные клетки легких человека от S. aureus-индуцированного лизиса (Wardenburg and Schneewind (2008)).
Хотя эти результаты показывают, что альфа-токсин участвует в разрушении тканей легких, остается неясным, является ли смерть животных в описанных выше экспериментах результатом прямого разрушения клеток легких токсином или чрезмерной воспалительной реакции или обеих причин. Пассивный перенос антител к альфа-токсину значительно снижал уровень циркулирующего интерлейкина 1β-цитокина, который, как известно, сопутствует острому повреждению легких. Таким образом, логично заключить, что воспалительный ответ может вносить свой вклад в повреждение легких, опосредуемое альфа-токсином.
При локализованной инфекции, такой как пневмония у людей, у приблизительно 40% пациентов с пневмонией, вызванной S. aureus, развиваются инфекции кровяного русла и диссеминированное поражение. Распространение бактериальной инфекции может привести к общему заражению крови и обсеменению удаленного органа. Инфекция кровяного русла может привести к септицемии, быстро прогрессирующему и часто летальному осложнению инфекций, обусловленных S. aureus.
Распространение инфекции S.aureus также часто наблюдается у модельных животных с пневмонией, вызванной S. aureus, причем также у приблизительно 40% животных развивается диссеминированная бактериемия вследствие повреждения тканей и проникновения инфекции через слои эпителия в кровяное русло и лимфатическую ткань. Тем не менее, диссеминация во многом зависит от генетического фона используемой линии животных, а также потенциала врожденной иммунной системы, такого как активация нейтрофилов для контроля роста, например, животные линии C57B/L, характеризующейся истощением нейтрофилов, очень восприимчивы к инфицированию почек золотистым стафилококком, в то время как иммунокомпетентные животные устойчивы к инфекциям. Напротив, животные линии А/J очень чувствительны к инфекциям, в основном вследствие задержанного поступления нейтрофилов в почки (von Kockritz-Blickwede, 2008).
Хотя данные о структуре и функции белков S.aureus стали более полными, разработка эффективной вакцины остается трудной задачей.
Была сделана попытка безопасно обеспечить иммунитет к альфа-токсину и бактериям S.aureus с использованием композиций, содержащих комбинацию антител, которые специфично связываются с антигеном - альфа-токсином S.aureus, и антител, которые специфично связываются с другим бактериальным антигеном (WO 2007/145689). Эти композиции хотя и включают количества антител, которые сами по себе не являются эффективными, нейтрализуют инфекции и/или обеспечивают защиту от инфекции в результате синергетической активности комбинации антител.
Было показано, что защитная эффективность указанной комбинации антител, нейтрализующих токсин S.aureus, и опсонических антител через 72 часа после инфицирования изолятом S.aureus выше, чем защитный эффект иммунизации только нейтрализующими или только опсоническими антителами. Комбинация опсонических и нейтрализующих токсин антител продемонстрировала защитный эффект, предотвращая инфицирование кожи и мягких тканей, а также диссеминацию органов. Тем не менее, раскрытых в заявке на патент нейтрализующих анти-альфа-токсин антител самих по себе не достаточно для предотвращения диссеминации органа/образования абсцесса или нейтрализации инфекции.
Другая попытка была сделана Heveker с соавт. (1994а, 1994b), который описывает нейтрализующие моноклональные антитела человека и мыши, направленные против альфа-токсина S.aureus. Моноклональное антитело человека подтипа IgG/лямбда охарактеризовано через последовательность и демонстрируют нейтрализацию.
Описанная Heveker (1994а) линия клеток гибридомы человека, продуцирующая антитела к альфа-токсину, была изолирована с использованием периферических лейкоцитов крови здорового добровольца, ранее иммунизированного испытательной вакциной, содержащей альфа-анатоксин S.aureus. Поскольку альфа-анатоксин, использованный в исследовании Heveker, представляет собой химически модифицированный альфа-токсин, можно предположить, что изменение действительно делает антигенные детерминанты менее иммуногенными или даже неиммуногенными. Следовательно, этот подход, как таковой, не может приводить к образованию такого же эффективного иммунитета, как это было продемонстрировано для других бактериальных токсинов, таких как токсин холеры: токсоид вакцины стимулировал образование анти-токсиновых антител, которые не вызывали развитие невосприимчивости к инфекциям (Levine (1983)).
В литературе идентифицированы различные факторы, являющиеся важнейшими факторами вирулентности для образования абсцесса, такие как токсины, пептидогликаны, внеклеточные факторы и ферменты. Предположение о возможной роли альфа-токсина в образовании абсцессов было высказано Kapral et al. (1980). Сообщалось, что альфа-токсин резко накапливается в абсцессе после его созревания, при этом не было продемонстрировано, является ли альфа-токсин необходимым для образования абсцесса. Вторая публикация, сделанная Adlam et al. (1977), отрицала ключевую роль альфа-токсина в образовании абсцесса. Авторы показали, что альфа-токсин играет ключевую роль в распространении геморрагической формы инфекционного мастита кроликов (blue-breast), наблюдаемого при естественных вспышках заболевания. Они воспроизвели в лаборатории клиническую картину, используя два штамма стафилококков, не являющихся близкими. Высокие титры циркулирующих анти-альфа-токсиновых антител привели к формированию защиты от этой летальной формы мастита. Таким образом, нейтрализующий титр антител может предотвращать летальный исход, изменяя клиническую картину в сторону состояния, характеризующегося менее тяжелыми абсцессами. Тем не менее, нейтрализация альфа-токсина не влияла/не предотвращала образование абсцесса у кроликов. В более поздней публикации Kielian et al. (2001) исследовали роль альфа-токсина в образовании абсцесса в мозге мышей. В эксперименте авторы имплантировали штамм дикого типа S. aureus и его мутанты в ткани мозга лобных долей и оценили способность каждого штамма вызывать абсцессы мозга. Авторы использовали штаммы, мутантные в локусах, связанных с экспрессией известных факторов вирулентности, например мутанты по локусу sarA и agr, каждый из которых участвует в общей регуляции важных факторов вирулентности. Так как альфа-токсин находится под контролем регуляторной системы sarA/agr, авторы также включили в свои эксперименты штамм, мутантный по альфа-токсину. Экспериментальные данные показали, что репликация штаммов бактерий, мутантных по альфа-токсину или локусу sarA/agr, снизила вирулентность бактериальных клеток при инъецировании в череп по сравнению с изогенным контрольным штаммом RN6390, что привело в результате к снижению количества бактерий и уменьшению обнаруживаемых очагов воспаления в мозге животных по сравнению с большими, хорошо сформированными абсцессами у мышей, получавших изогенный штамм.
Тем не менее, в экспериментальной модели абсцесса мозга мутантные штаммы были не полностью авирулентны и поэтому нельзя исключить того, что помимо альфа-токсина, дополнительный фактор или факторы могут играть важную роль/роли в образовании абсцесса мозга.
Роль альфа-токсина в образовании абсцесса была оценена в других экспериментальных условиях, как это изложено Schwan et al. (2003) в анализе локальной, системной и абсцесс-формирующей инфекционной стафилококковой модели. Авторы заметили, что с течением времени негемолитические штаммы золотистого стафилококка стали более распространенными в мышиных моделях абсцесса и моделях ран, но не в тканях органов, связанных с системными инфекциями. Например, в смешанной инфекции с использованием всех вариантов штамма золотистого стафилококка RN6390 (гипергемолитический, гемолитический и негемолитический) в модели абсцесса, гипергемолитическая группа заметно снизилась на 7 день после заражения, а негемолитическая популяция значительно увеличилась. Секвенирование некоторых мутантов, полученных путем направленного мутагенеза, показало наличие мутаций в гене agrC или в межгенной области agrA-agrC, которые приводили к снижению активности как альфа-токсина, так и дельта-токсина. При анализе определенных мутантных штаммов на активность agr (agr-) или альфа-токсина (hla-) в абсцессе, ране и системной модели инфекции, agr-мутантный штамм и hla-мутантный штамм не показали каких-либо различий общего количества бактерий в абсцессах мышей на 4-й день по сравнению с исходными штаммами дикого типа (RN6390). То же справедливо для локальных инфекций (модель раны), в то время как в модели системной инфекции произошла значительная очистка hla- и agr-мутантного штамма. Результат явно указывает на важность альфа-токсина в системных инфекциях, но не в локальных инфекциях или в образовании абсцесса. Действительно, смешанные инфекции, вызванные штаммами hla-мутанта и дикого типа, показали в модели абсцесса небольшое преобладание популяции hla-мутанта над штаммом дикого типа. Далее, авторы заключили, что мутации agr вызывают снижение экспрессии альфа- и дельта-токсинов, которые, в свою очередь, способствуют преимущественному росту этой agr-мутантной группы в смешанной популяции клеток S. aureus, присутствующей в абсцессах и ранах. Результаты явно противоречат результатам, описанным Kielian et al., где отсутствие альфа-токсина снижало бактериальную вирулентность. Поэтому роль альфа-токсина в образовании абсцесса не ясна.
В целом, отсутствуют свидетельства, указывающие на единственный фактор вирулентности как основную движущую силу в образовании абсцесса. Поэтому исследование было сосредоточено на обнаружении дополнительных факторов, которые не полностью контролируется золотистым стафилококком, таких как факторы окружающей среды или определенных структурных участков, как общих ключевых факторов в формировании абсцесса. Например, последние данные о факторах вирулентности, влияющих на формирование абсцессов, указывают на влияние несвязанных двухвалентных ионов металлов, таких как Мn2+и Са2+, в образовании абсцесса и размножении бактерий в нем. Хелатирование ионов металлов у животных ингибирует образование абсцессов в печени и подавляет размножение S. aureus в абсцессе (Corbin, 2008). С другой стороны, Tzianabos et al. (2001) предположил, что для образования патологических структур, например абсцессов в тканях, такой организм, как S. aureus, нуждается в факторах вирулентности, присутствующих на поверхности бактериальной клетки. Авторы показали, что штаммы, тесно ассоциированные с клиническими случаями формирования абсцессов, могут иметь один или несколько полисахаридов, связанных с клеточной стенкой и содержащих цвиттерионные участки в молекуле (химическое соединение, общий заряд которого равен нулю, но при этом электрически нейтральная молекула несет формально положительные и отрицательные заряды на различных атомах). При отсутствии цвиттерионного участка абсцесс не формируется. Авторы пришли к выводу, что эти полисахаридные полимеры могут модулировать образование абсцесса организмом. Кроме того, они представили доказательства участия в этом процессе не только основных полисахаридов СР5 и СР8, но и липотейхоевой кислоты (LTA), дополнительного хорошо известного фактора вирулентности, присутствующего в клеточной стенке. Они также идентифицировали цвиттерионный участок в молекуле липотейхоевой кислоты и, таким образом, расширили свою гипотезу формирования абсцесса, включив в нее наличие цвиттерионного участка в любом факторе вирулентности, являющемся ключевым при формировании абсцесса.
Основываясь на результатах, указывающих на то, что различные факторы способствуют формированию абсцесса, индуцируемому S.aureus, специалист в данной области не может ожидать, что нейтрализация только одного фактора позволит предотвратить образование абсцесса.
Таким образом, задачей изобретения является создание средств и способов для профилактики и лечения клинически сложного инфекционного заболевания, вызванного S.aureus, такого как образование абсцесса.
Соответственно, одной технической задачей, лежащей в основе настоящего изобретения, является создание моноклонального антитела, специфичного к альфа-токсину S. aureus, причем антитело обладает in vivo защитной способностью против клинически сложного инфекционного заболевания, вызванного S.aureus, такого как образование абсцесса.
Указанная техническая задача решается при помощи моноклонального антитела, определенного ниже.
Согласно настоящему изобретению предложено моноклональное антитело, обозначенное 243-4, и специфичное в отношении альфа-токсина S. aureus, причем вариабельная область легкой цепи антитела содержит по меньшей мере одну из SEQ ID NO: 1 в участке CDR1, SEQ ID NO: 2 в участке CDR2 и SEQ ID NO: 3 в участке CDR3, и вариабельная область легкой цепи антитела содержит по меньшей мере одну из SEQ ID NO:1 в участке CDR1, SEQ ID NO:2 в участке CDR2 и SEQ ID NO:3 в участке CDR3, и вариабельная область тяжелой цепи антитела содержит по меньшей мере одну из SEQ ID NO:4 в участке CDR1, SEQ ID NO:5 в участке CDR2 и SEQ ID NO:6 в участке CDR3, либо фрагмент или мутеин указанного антитела, способный связывать альфа-токсин S. aureus, причем мутеин моноклонального антитела несет по меньшей мере одну консервативную замену в любом из участков CDR тяжелой или легкой цепи антитела.
В соответствии с предпочтительным вариантом реализации согласно настоящему изобретению предложено человеческое моноклональное антитело, специфичное для альфа-токсина S. aureus, где вариабельная область легкой цепи антитела состоит из SEQ ID NO:1 в участке CDR1, SEQ ID:2 в участке CDR2 и SEQ ID NO:3 в участке CDR3, и где вариабельная область тяжелой цепи антитела содержит SEQ ID NO:4 в участке CDR1, SEQ ID NO:5 в участке CDR2 и SEQ ID NO:6 в участке CDR3, либо их фрагмент или мутеин, способный связывать альфа-токсин S. aureus, где мутеин моноклонального антитела несет по меньшей мере одну консервативную замену в любом из участков CDR (гипервариабельных участков) тяжелой или легкой цепи антитела.
Неожиданно было обнаружено, что моноклональные антитела согласно настоящему изобретению демонстрируют высокую способность препятствовать образованию абсцесса. Предотвращение развития абсцесса было продемонстрировано на модели с использованием почек мышей путем введения моноклонального антитела человека, специфичного к альфа-токсину, согласно настоящему изобретению. Судя по строению токсина, который является скорее секретируемым белком, а не компонентом (полисахаридом) клеточной стенки, любой прямой бактерицидный эффект, такой как уничтожение бактериальной клетки, или непрямая эффекторная функция, опосредуемая иммунной системой, например опсоно-фагоцитарное уничтожение патогена, обусловленное активацией системы комплемента, могут быть исключены и не являются причиной отсутствия образования абсцесса.
В настоящем описании термин «моноклональное антитело» включает любое моноклональное антитело человека или его часть, независимо от источника получения. Предпочтительным является полностью человеческое моноклональное антитело. Предпочтительным является получение моноклонального антитела при помощи гибридомы. Гибридома может быть гибридомой клеток млекопитающих, например мышиных, крупного рогатого скота или человека. Предпочтительной является гибридома человеческого происхождения. Для получения моноклонального антитела человека пригодны гибридомы, полученные путем осуществления слияния B-клетки человека с клеткой миеломы или гетеромиеломы.
B-клетки человека могут быть получены путем иммунизации здорового субъекта или путем отбора выздоравливающих пациентов с последующим отборам у них образцов крои, из которых могут быть выделены В-клетки. выделение B-клеток из образцов крови может быть осуществлено известными способами (см., например. Current Protocols in Immunology. Chapter 7.1. Isolation of whole mononuclear cells from peripheral blood and cord blood. Изд. Wiley & sons, Eds. J.C. Coligan et al.). Слияние B-клетки человека с клеткой миеломы или гетеромиеломы с получением гибридомы осуществляют в соответствии со стандартными методиками согласно классическому подходу Köler и Milstein. К пригодным клеткам миеломы относятся производные P3X63, такие как P3X63Ag8. 653 (ATCC CRL-1580) или SP2/Q (АТСС CRL-1646). Пригодные клетки гетеромиеломы включают, например, F3B6 (ATCC HB-8785). Полученные гибридомы могут быть подвергнуты отбору в соответствии с известными процедурами. Гибридомы культивируют в соответствующей культуральной среде и выделяют антитело из супернатанта.
Моноклональное антитело также может быть получено методами генной инженерии и, в частности, прививкой сегментов CDR, определенных в формуле изобретения, на доступные моноклональные антитела,
путем замены участка CDR основного антитела на сегменты специфичных CDR, как определено формуле изобретения.
Термин «участок CDR» обозначает гипервариабельный участок антитела, то есть участок, определяющий специфичность антитела к определенному антигену. Три участка CDR (CDR1-CDR3) как на легкой, так и на тяжелой цепи, отвечают за связывание антигена.
Участок CDR имеет следующую локализацию в тяжелой цепи:
Участок CDR1: аминокислоты с 26 по 33 в экзоне VH,
Участок CDR2: аминокислоты с 51 по 58 в экзоне VH,
CDR3 участок: аминокислоты с 97 до 110 в экзоне VH.
Положение участков CDR не зависит от класса антитела, то есть IgM, IgA или IgG.
Участок CDR имеет следующую локализацию в легкой цепи лямбда-типа:
Участок CDR1: аминокислоты с 26 по 33 в экзоне Vλ,
Участок CDR2: аминокислоты с 51 по 58 в экзоне Vλ,
CDR3 участок: аминокислоты с 90 до 110 в экзоне Vλ.
Сопоставление (выравнивание) аминокислотных последовательностей экзонов VH, Vχ и Vλ может быть получено из базы данных V Base (http://imqt.cines.fr/IMGT_vquest/share/textes/).
Термин «фрагмент» обозначает любой фрагмент антитела, способный связываться с альфа-токсином S. aureus. Фрагмент имеет длину не менее 10 аминокислот, предпочтительно 20, наиболее предпочтительно 50 аминокислот. Кроме того, предпочтительно, если фрагмент включает связывающий участок антитела. В предпочтительном случае фрагмент является Fab, F(ab')2, одноцепочечным или доменным антителом. Наиболее предпочтительным является фрагмент Fab, или F(ab')2, или их смесь. Фрагменты антитела могут быть гликозилированы, например содержать углеводные остатки в вариабельных областях антитела.
Соответственно, настоящее изобретение дополнительно обеспечивает моноклональное антитело, как определено в настоящем описании, причем антитело представлено Fab, F(ab')2, одиночной цепью или доменным фрагментом антитела.
Термин «мутеин» включает любые мутеины моноклонального антитела, отличающиеся добавлением, удалением и/или заменой по меньшей мере одной аминокислоты. Предпочтительным вариантом реализации является наличие в мутеине моноклонального антитела по меньшей мере одной консервативной замены в любом из участков CDR тяжелой или легкой цепи антитела, как указано в формуле изобретения. Наиболее предпочтительным вариантом реализации является наличие в мутеине не более 5, не более 4, предпочтительно не более 3 и особенно предпочтительно не более 2 консервативных замен. Способность фрагмента или мутеина антитела связываться с альфа-токсином S. aureus определяют прямым иммуноферментным анализом, как это описано в разделе «Примеры»: очищенный альфа-токсин иммобилизуют на твердой фазе планшетов для ИФА. Фрагменты антитела или мутеины антител инкубируют с иммобилизованным альфа-токсином, после чего связанные антитела или их мутеины визуализируют с помощью соответствующего вторичного антитела, конъюгированного с ферментом.
Термин «консервативная замена» обозначает замену одной аминокислоты, принадлежащей к определенной физико-химической группе, на аминокислоту, принадлежащую к той же физико-химической группе. Физико-химические группы определяются следующим образом:
Физико-химическая группа неполярных аминокислот включает в себя: глицин, аланин, валин, лейцин, изолейцин, метионин, пролин, фенилаланин и триптофан. Группа аминокислот, имеющих незаряженные полярные боковые цепи, состоит из аспарагина, глутамина, тирозина, цистеина и цистина. Физико-химическая группа аминокислот с положительно заряженной полярной боковой цепью состоит из лизина, аргинина и гистидина. Физико-химическая группа аминокислот с отрицательно заряженной полярной боковой цепью состоит из аспарагиновой и глутаминовой кислот и их карбоксилат-анионов, называемых также аспартат и глутамат.
Согласно другому варианту реализации настоящее изобретение обеспечивает моноклональное антитело, специфичное к альфа-токсину S. aureus, в котором вариабельная область легкой цепи антитела имеет аминокислотную последовательность SEQ ID NO:7, и вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:8, или фрагменты указанного антитела, способные связываться с альфа-токсином S. aureus, либо вариант указанного антитела, способный связываться с альфа-токсином S. aureus, в котором аминокислотная последовательность вариабельной области легкой цепи антитела по меньшей мере на 85% идентична SEQ ID NO:7 и аминокислотная последовательность вариабельной области тяжелой цепи антитела по меньшей мере на 85% идентична SEQ ID NO:8. В настоящем описании термин «вариант» относится к полипептиду, в котором аминокислотная последовательность демонстрирует определенную степень идентичности с аминокислотной последовательностью, приведенной в перечне последовательностей.
Термин «% идентичность», известный специалистам в данной области, обозначает степень схожести между двумя или более полипептидными молекулами, которая определяется соответствием последовательностей. Процент «идентичности» вычисляется на основании процента идентичных участков в двух или более последовательностях с учетом пропусков или других особенностей последовательностей.
Процент идентичности близких полипептидов может быть определен с помощью известных методик. Как правило, используют специальные компьютерные программы с алгоритмами, учитывающими особые требования. Предпочтительные методики для определения идентичности сначала генерируют наибольшее сходство между изучаемыми последовательностями. Компьютерные программы для определения идентичности между двумя последовательностями включают, но не ограничиваются, пакет программ GCG, в том числе GAP (Devereux J et al., (1984); Genetics Computer Group, Университет Висконсина, Мэдисон (Висконсин); BLASTP, BLASTN и FASTA (Altschul S et al., (1990)). Программа BLAST X может быть получена в Национальном центре биотехнологической информации (NCBI) и из других источников (Справочник BLAST, Altschul S et al., NCB NLM NIH Бетесда, Мэриленд, 20894; Altschul S et al., 1990). Для определения процента идентичности может быть использован хорошо известный алгоритм Смита-Уотермана.
Ниже представлены предпочтительные параметры для сравнения последовательностей:
Алгоритм: Needleman S.В. и Wunsch C.D. (1970),
Матрица сравнения: BLOSUM62 из Henikoff C. и Henikoff J.G. (1992),
Штраф за пропуск в последовательности: 12,
Штраф за длину пропуска в последовательности: 2.
Программа GAP также подходит для использования с указанными параметрами. Указанные выше параметры являются стандартными (параметры по умолчанию) для сравнения аминокислотных последовательностей, в которых пробелы на концах не уменьшают значение идентичности. Когда очень короткие последовательности сравнивают с базовой последовательностью, дополнительно может потребоваться увеличение значения математического ожидания до 100000, а в некоторых случаях уменьшение длины слова (размер слова) до 2.
Могут быть использованы дополнительные алгоритмы модели, штраф за создание разрыва, штраф за удлинение разрыва и матрицы сравнения, включая указанные в "Справочнике программ", пакет Висконсина, версия 9, сентябрь 1997 года. Выбор будет зависеть от сравнения, которое должно быть сделано, и в дальнейшем от того, выполняется ли сравнение между парами последовательностей, в этом случае предпочтительными являются GAP или Best Fit, или между одной последовательностью и большой базой данных по последовательностям, в этом случае предпочтительными являются FASTA и BLAST. Сходство в 85%, определенное при помощи указанных алгоритмов, описывается как 85% идентичность. То же самое применимо к более высокой степени идентичности.
В предпочтительном варианте реализации, варианты согласно настоящему изобретению имеют идентичность, равную 85% или больше, предпочтительно 90% или больше, и наиболее предпочтительно 95% и больше.
В соответствии с предпочтительным вариантом реализации настоящего изобретения, моноклональное антитело является антителом человека. В настоящем описании термин «человеческий» означает, что моноклональное антитело человека практически не содержит аминокислотных последовательностей чужеродных видов, в предпочтительном варианте моноклональное антитело человека (человеческое моноклональное антитело) полностью состоит только из аминокислотной последовательности человека.
Легкая цепь моноклонального антитела согласно настоящему изобретению может быть каппа- или лямбда-типа.
В предпочтительном варианте реализации настоящего изобретения легкая цепь принадлежит к лямбда-типу. Легкая цепь может быть представлена природной цепью, включая природные перестройки, генетически модифицированной или синтетической легкой цепью.
Тяжелая цепь моноклонального антитела согласно настоящему изобретению может быть выбрана из изотипов IgM, IgA, или IgG, предпочтительно IgG.
В соответствии с другим предпочтительным вариантом реализации изобретения тяжелая цепь моноклонального антитела принадлежит IgG.
В настоящем описании термин «способный связываться» относится к связыванию, которое происходит между антителом и распознанным антигеном, к которому направлено данное антитело. Этот тип связывания является специфичным, в отличие от неспецифичного, которое происходит в отсутствие антигена.
Антитела, способные связываться с альфа-токсином, получают с использованием гибридомной технологии, в которой В-клетки представляют собой В-клетки млекопитающих, например мыши, крупного рогатого скота или человека. Предпочтительным вариантом В-клеток являются В-клетки человека. Помимо этого, моноклональное антитело, способное связываться с альфа-токсином, может быть получено путем CDR-пересадки участков CDR, как указано в формуле изобретения, на доступные моноклональные антитела, с получением моноклонального антитела, специфичного в отношении альфа-токсина S. aureus согласно настоящему изобретению.
Согласно следующему варианту реализации в изобретении предложено моноклональное антитело, способное связываться с альфа-токсином S. aureus, которое может быть получено из В-клеток млекопитающих, например мышей, крупного рогатого скота или человека, предпочтительным вариантом являются человеческие В-клетки или гибридомы, полученные путем слияния указанных В-клеток человека с клетками миеломы или гетеромиеломы.
Согласно следующему варианту реализации изобретения предложена гибридома, способная продуцировать моноклональные антитела, связывающиеся с альфа-токсином S. aureus, определенным в настоящей заявке.
В настоящем описании термин «альфа-токсин» относится к бактериальному белку, вырабатываемому S. aureus. После связывания с поверхностью клетки-хозяина альфа-токсин подвергается олигомеризации до гептамера. Образование поры является основной причиной апоптоза и лизиса клеток. Для потенциальной защиты от патогена фундаментальное значение имеет способность моноклонального антитела связываться как с мономерной, так и с олигомерной формой альфа-токсина, продуцируемого S. aureus.
В соответствии с другим предпочтительным вариантом реализации изобретения моноклональное антитело согласно настоящему изобретению способно специфично связывать мономерные и олигомерные формы альфа-токсина S. aureus. В соответствии с другим вариантом реализации моноклональное антитело согласно настоящему изобретению или его фрагмент или мутеин способны специфично связываться как с мономерной, так и олигомерной формой альфа-токсина S. aureus или с обеими формами.
В настоящем описании термин «олигомерные формы» включает форму, отличную от мономерной формы альфа-токсина, такую как димерная, тримерная, тетрамерная, пентамерная, гексамерная, гептамерная и т.д., или полимерные формы, такие как гептамерная форма альфа-токсина, предшествующая формированию поры.
В соответствии с другим предпочтительным вариантом реализации моноклональное антитело согласно настоящему изобретению модифицировано по N-концу, внутренней области или С-концу. Модификации включают ди-, олиго- или полимеризацию мономерной формы, например путем поперечного сшивания с использованием дициклогексилкарбодиимида. Полученные таким образом ди-, олиго- или полимеры могут быть разделены гель-фильтрацией. Дальнейшие модификации включают модификации боковой цепи, например изменения остатков ε-амино-лизина или амино- и карбокси-концевые модификации, соответственно. Дальнейшие модификации включают посттрансляционные модификации, например гликозилирование и/или частичное или полное дегликозилирование белков и образование дисульфидных связей. Антитела могут также быть конъюгированы с меткой, такой как ферментная, люминесцентная или радиоактивная метки. Предпочтительный вариант модификации выбирают по меньшей мере из одного: олигомеризации, гликозилирования или конъюгации с лекарственным препаратом или меткой.
Кроме того, в настоящем изобретении предложены нуклеиновые кислоты, кодирующие соответственно тяжелую и легкую цепи моноклонального антитела настоящего изобретения. Нуклеиновые кислоты могут быть представлены естественной нуклеиновой кислотой либо получены из зародышевой линии или из перестроек, происходящих в В-клетках, также нуклеиновые кислоты могут быть синтетическими. Синтетические нуклеиновые кислоты также включают нуклеиновые кислоты, имеющие модифицированные внутринуклеозидные связи, включая фосфотиоэфиры для увеличения устойчивости нуклеиновых кислот к разрушению. Нуклеиновые кислоты могут быть получены методами генной инженерии или полностью посредством синтеза из нуклеотидов.
В настоящем изобретении дополнительно предложены векторы, содержащие по меньшей мере одну нуклеиновую кислоту, кодирующую легкую цепь моноклонального антитела согласно настоящему изобретению и/или по меньшей мере одну нуклеиновую кислоту, кодирующую тяжелую цепь моноклонального антитела согласно настоящему изобретению. Нуклеиновые кислоты могут быть строены в один и тот же вектор или могут присутствовать в форме бинарных векторов. Вектор предпочтительно содержит промотор, функционально связанный с нуклеиновой кислотой для облегчения экспрессии нуклеиновой кислоты, кодирующей легкую и/или тяжелую цепи.
В предпочтительном варианте вектор дополнительно содержит ориджин для репликации и поддержания в клетке-хозяине. Вектор может также включать нуклеотидную последовательность, кодирующую сигнальную последовательность, расположенную на 5'-конце нуклеиновой кислоты, кодирующей легкую или тяжелую цепи. Сигнальная последовательность может способствовать секреции кодируемой цепи в среду.
В предпочтительном варианте вектор получен из аденовирусов, вирусов вакцины, бакуловирусов, SV 40 вируса, ретровирусов, вирусов растений или бактериофагов, таких как производные лямбда или М13. Наиболее предпочтительным является вектор, содержащий константные области тяжелых и легких цепей Ig человека, такой как интегрированная векторная система для экспрессии иммуноглобулинов в клетках эукариот, описанная Persic et al., 1987.
Кроме того, в настоящем изобретении предложены клетки-хозяева, содержащие вектор и/или нуклеиновую кислоту и пригодные для экспрессии вектора. В области техники, к которой относится данное изобретение, известны многочисленные прокариотические и эукариотические системы экспрессии, причем эукариотические клетки-хозяева, такие как дрожжевые клетки, клетки насекомых, клетки растений и клетки млекопитающих, такие как линии НЕK293, PerC6, СНО, COS или HeLa и их производные, являются предпочтительными. Особенно предпочтительными являются продуцирующие линии клеток человека. В предпочтительном случае трансфецированные клетки-хозяева выделяют продуцируемые антитела в культуральную среду. Если достигается внутриклеточная экспрессия, то осуществляют ренатурацию в соответствии со стандартными методиками, например описанными Benetti et al., 1998.
Моноклональные антитела согласно настоящему изобретению выделяют из лимфоцитов крови выздоравливающего больного, и в результате получают антитела, очищенные и отобранные естественным путем, имеющие высокую аффинность, для нейтрализации и эффективной защиты от инфекций.
Настоящее изобретение также обеспечивает способы получения моноклонального антитела. В одном из вариантов реализации моноклональное антитело получают путем культивирования описанной выше гибридомы. Продуцированное моноклональное антитело секретируется в надосадочную жидкость и может быть очищено от нее посредством обычных методов хроматографии.
В альтернативном варианте моноклональное антитело продуцируется клетками-хозяевами, содержащими вектор согласно настоящему изобретению, путем культивирования клеток-хозяев в условиях, пригодных для рекомбинантной экспрессии кодируемой цепи антитела. В предпочтительном варианте клетка-хозяин содержит по меньшей мере одну нуклеиновую кислоту, кодирующую легкую цепь, и по меньшей мере одну нуклеиновую кислоту, кодирующую тяжелую цепь, и обеспечивает сборку моноклонального антитела с получением трехмерной пространственной структуры, соответствующей трехмерной структуре моноклонального антитела, продуцируемого млекопитающими, предпочтительно В-клетками человека. Если легкая цепь продуцируется отдельно от тяжелой цепи, обе цепи могут быть очищены, а затем собраны с получением моноклонального антитела, имеющего трехмерную структуру моноклонального антитела, по существу сходную со структурой антител, вырабатываемых млекопитающими, предпочтительно В-клетками человека.
Моноклональное антитело также может быть получено путем рекомбинантной экспрессии кодируемой легкой и/или тяжелой цепи, где нуклеиновая кислота получена путем выделения нуклеиновых кислот, кодирующих моноклональное антитело, известным способом и последующим встраиванием последовательности нуклеиновой кислоты, кодирующей CDR участки, в выделенную нуклеиновую кислоту, как определено в формуле настоящего изобретения.
В настоящем изобретении также предложены фармацевтические композиции, содержащие по меньшей мере одно моноклональное антитело и/или по меньшей мере одну нуклеиновую кислоту, кодирующую легкую и/или тяжелую цепь моноклонального антитела.
Фармацевтическая композиция может дополнительно содержать антибиотики, такие как стрептомицин, пенициллин и ванкомицин и т.д., предпочтительно связанные с моноклональным антителом согласно настоящему изобретению.
Фармацевтические композиции содержат моноклональное антитело в интервале доз 0,1-100 мг/кг массы тела.
Фармацевтические композиции можно вводить любым известным способом, например внутривенно, внутримышечно, внутрикожно, подкожно, внутрибрюшинно, местно, интраназально или в форме вдыхаемого спрея.
В предпочтительном варианте реализации изобретения фармацевтические композиции применяют для профилактики или лечения образования абсцесса в органе пациента-млекопитающего, такого как крупный рогатый скот, свинья, кошка, собака, лошадь, человек. В предпочтительном варианте реализации изобретения фармацевтические композиции применяют для людей. В другом варианте реализации изобретения образование абсцесса вызвано стафилококковой инфекцией.
Кроме того, S. aureus-индуцированная инфекция, которую лечат с помощью фармацевтической композиции согласно настоящему изобретению, может быть инфекцией молочной железы, такой как мастит.
Соответственно, в настоящем изобретении предложено применение моноклонального антитела или нуклеиновой кислоты, кодирующей вариабельную область легкой цепи и/или тяжелой цепи согласно настоящему изобретению, для изготовления фармацевтической композиции для профилактики или лечения образования абсцесса в органе млекопитающего, предпочтительно человека.
В предпочтительном варианте реализации изобретения фармацевтические композиции, моноклональное антитело или нуклеиновая кислота, кодирующая вариабельную область легкой или тяжелой цепи, согласно настоящему изобретению применяют для профилактики или лечения абсцесса в органе, таком как почки, сердце, печень, желчный пузырь, поджелудочная железа, тонкий кишечник, толстый кишечник, легкие, мозг, кожа, глаза, лимфатические ткани и селезенка. В предпочтительном варианте реализации изобретения лечение будет направлено на брюшной абсцесс. Соответственно, орган брюшной полости, который будет подвергаться лечению - это печень, желчный пузырь, селезенка, поджелудочная железа, почки, тонкий или толстый кишечник.
В настоящем описании термин «образование абсцесса» относится к образованию абсцесса в органе, например, почке, сердце, печени, желчном пузыре, поджелудочной железе, тонком или толстом кишечнике, легких, мозге, коже, глазах, лимфатических тканях и селезенке. В настоящем описании термин «абсцесс» означает скопление гноя, который накапливается из-за инфекционного процесса (как правило, вызывается бактериями или паразитами) в образованной тканью полости. Токсины, выделяемые размножающимися бактериями, разрушают клетки и вызывают воспалительный ответ, который привлекает в эту область большое количество лимфоцитов и увеличивает площадь локального кровотока. Лейкоциты разрушают мертвые ткани и поглощают бактерии путем фагоцитоза. Из разрушенных тканей, мертвых бактерий и лейкоцитов, а также накопившейся внеклеточной жидкости образуется мутный зеленый или желтоватый гной. Для абсцесса характерна инкапсуляция стенкой, которая образуется здоровыми соседними клетками, пытающимися изолировать гной от близлежащих структур. Это - защитная реакция ткани для приостановки распространения инфекционных материалов в другие части тела. Абсцессы могут развиваться в любом типе плотных тканей, но чаще всего на поверхности кожи (там, где они могут проявляться как поверхностные пустулы (фурункулы) или глубокие кожные абсцессы), в легких, мозге, почках и миндалинах. Главные осложнения - это распространение содержимого абсцесса, например обсеменение органа, на соседние или удаленные ткани и обширное локальное отмирание тканей (гангрена). Образование абсцесса определяют путем оценки бактериальной нагрузки в органе.
В настоящем описании термин «брюшной абсцесс» относится к абсцессу органа брюшной полости. Брюшная полость - это полость тела, которая содержит большую часть внутренних органов, она располагается ниже (или инфериорно) грудной полости и выше полости малого таза. Она является частью абдоминально-тазовой полости. Органы брюшной полости включают желудок, печень, желчный пузырь, селезенку, поджелудочную железу, тонкий и толстый кишечник, почки.
В настоящем описании термин «обсеменение органа» означает распространение живых бактерий из локального очага инфекции в удаленные ткани и органы. Обсеменение органа характеризуется наличием живых, инфицирующих бактериальных клеток в здоровой ткани без образования макроскопических инкапсулированных колоний бактерий.
В настоящем описании термин «бактериальная нагрузка» определяется как количество живых бактериальных клеток в пределах анатомически хорошо определенной ткани, и выражается как количество бактериальных клеток, формирующих колонии на плотной питательное среде, такой как агар. Чтобы оценить бактериальную нагрузку в органе, его извлекают хирургическим путем из окружающих тканей, после чего в стерильных условиях ткани органа измельчают в стерильном физиологическом растворе для разрушения структуры тканей и отделения бактериальных клеток от ткани млекопитающих. Определенное количество клеточной суспензии (или ее серийные разведения в стерильном физиологическом растворе) наносят на плотную питательную среду для бактерий. Бактериальную нагрузку выражают как число «колониеобразующих единиц» в почке (например, КОЕ/почку).
В настоящем изобретении также предложен тестовый набор для диагностики инфекций, вызванных S. aureus, содержащий по меньшей мере одно моноклональное антитело согласно настоящему изобретению и, возможно, дополнительные компоненты, пригодные для проведения диагностического теста.
Ингредиенты, пригодные для проведения диагностического теста, - это, например, буферный раствор с осмоляльностью, варьирующей от 280 до 320 мОсм/л, и значением рН в диапазоне от 6 до 8; буферный раствор, содержащий хелаторы; буферный раствор, содержащий моновалентные или двухвалентные катионы с общей концентрацией катионов в буфере от 0,02 М до 2,0 М и/или буферный раствор, содержащий человеческую сыворотку или сыворотку животного происхождения с концентрацией от 0,01% до 20%.
Тестовый набор применим для специфичной надежной диагностики инфекций, вызванных S.aureus. Количественное определение может быть основано на стандартном ИФА в жидкой фазе или твердофазном ИФА. Детектирование может быть прямым или косвенным, как хорошо известно в области техники, к которой относится данное изобретение, причем антитела могут быть факультативно конъюгированы с ферментной, флуоресцентной или радиоактивной меткой.
Соответственно, настоящее изобретение также обеспечивает применение по меньшей мере одного моноклонального антитела согласно настоящему изобретению для детектирования связывания с альфа-токсином в образце. Связывание антитела согласно настоящему изобретению с альфа-токсином может быть обнаружено с использованием, например, вторичных антител козы к IgG человека, конъюгированных с пероксидазой хрена.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГУРА 1
На фигуре 1 показаны последовательность ДНК и аминокислотная последовательность вариабельной области тяжелой цепи моноклонального антитела человека 243-4. Участок CDR1 антитела 243-4 располагается в положениях с 26 по 33, Участок CDR2 антитела 243-4 располагается в положениях с 51 по 58, а участок CDR3 антитела 243-4 располагается в положениях с 97 по 110.
ФИГУРА 2
На фигуре 2 показаны последовательность ДНК и аминокислотная последовательность вариабельной области легкой цепи моноклонального антитела человека 243-4. Участок CDR1 антитела 243-4 располагается в положениях с 26 по 33, Участок CDR2 антитела 243-4 располагается в положениях 51 по 53, а участок CDR3 антитела 243-4 располагается в положениях с 90 по 101.
ФИГУРА 3
На фигуре 3 показана антигенная специфичность моноклонального антитела человека 243-4. Антигенную специфичность антитела 243-4 оценивали по связыванию с панелью бактериальных токсинов в твердофазном иммуноферментном анализе ELISA. Анализ ELISA осуществляли в планшетах для микротитрования, покрытых очищенными токсинами. После 24-часовой инкубации при комнатной температуре планшеты для микротитрования блокировали бычьим сывороточным альбумином. Затем, используя вторичные антитела козы к IgG человека, конъюгированные с пероксидазой хрена, определяли связывание моноклонального антитела 243-4 с иммобилизованными токсинами. Моноклональное антитело человека 243-4 связывалось с альфа-токсином S. aureus значительно сильнее, по сравнению с другими испытуемыми токсинами.
ФИГУРА 4
На фигуре 4 показаны результаты анализа связывания моноклонального антитела человека 243-4 с альфа-токсином эпидемических штаммов S. aureus методом вестерн-блот. Осуществляли мониторинг выделения альфа-токсина двенадцатью эпидемическими штаммами S. aureus в стационарной фазе роста бактериальной культуры. После культивирования нормализованный бактериальный супернатант и очищенный альфа-токсин загружали на ДСН-ПААГ гель и осуществляли электроблоттинг. После блокирования нитроцеллюлозную мембрану инкубировали с очищенным моноклональным антителом человека 243-4.
Продуцирование и распознавание мономерной и/или гептамерной формы альфа-токсина из каждого исследованного эпидемического штамма подтверждали по связыванию моноклонального антитела человека 243-4.
На фигуре 4 М обозначает размер маркера, цифры 1-12 представляют эпидемические штаммы S. aureus, и αТох обозначает очищенный альфа-токсин.
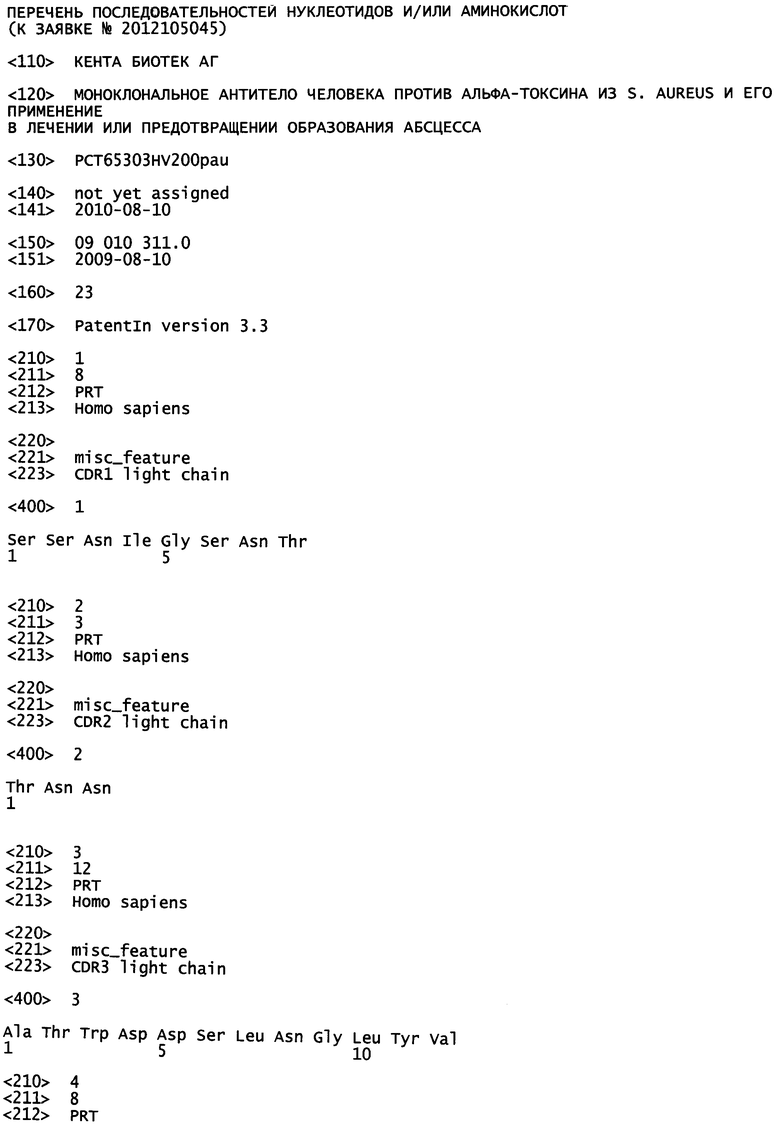
ФИГУРА 5
На фигуре 5 показано определение аффинности моноклонального антитела человека 243-4 в анализе BIAcore. Кинетику связывания моноклонального антитела человека 243-4 анализировали с использованием системы BIAcore 2000. Различные концентрации альфа-токсина загружали в проточную ячейку с иммобилизованным на чипе моноклональным антителом человека 243-4. Для расчета константы диссоциации антитела регистрировали фазы ассоциации и диссоциации. Кинетические данные оценивали путем общей аппроксимации с использованием программного обеспечения BIAevaluation 4.1.
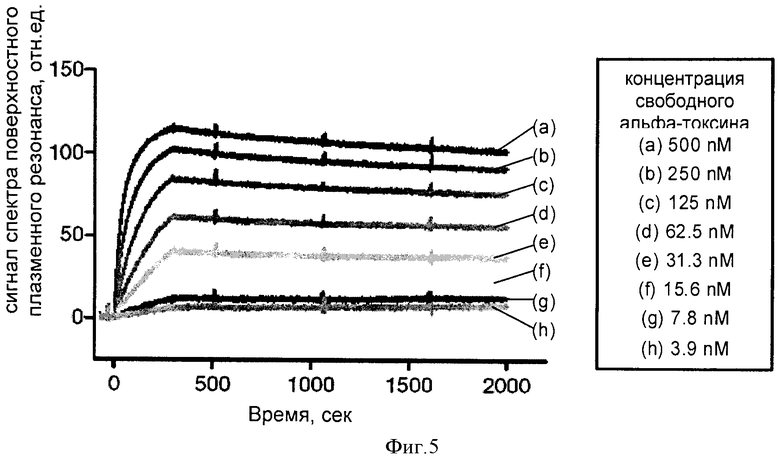
ФИГУРА 6
На фигуре 6 показана нейтрализация альфа-токсина моноклональным антителом человека 243-4 в модели повреждения альвеолярных клеток человека на культуре ткани. Эпителиальные клетки человека А549 культивировали с альфа-токсином на протяжении 16 часов в присутствии либо антитела контроля изотипа или моноклонального антитела 243-4. После этого клетки исследовали с использованием количественного анализа лактатдегидрогеназы (ЛДГ), который определяет наличие клеток, поврежденных альфа-токсином и показывает степень их защиты, которая может быть достигнута при применении антитела. Для интерпретации результатов за 100% принимали степень лизиса клеток при их инкубации с самой высокой концентрацией альфа-токсина. Степень лизиса клеток, обработанных только альфа-токсином, зависела от его концентрации. Сходные результаты были получены для клеток, обработанных альфа-токсином и меченым антителом. Эти данные указывают на отсутствие защитного эффекта антитела контроля изотипа. Инкубация альвеолярных клеток легкого человека с моноклональным антителом человека 243-4, напротив, защищала их от лизиса, индуцированного альфа-токсином. Были проведены три независимых эксперимента, каждый из которых подтвердил защитные свойства антитела 243-4.
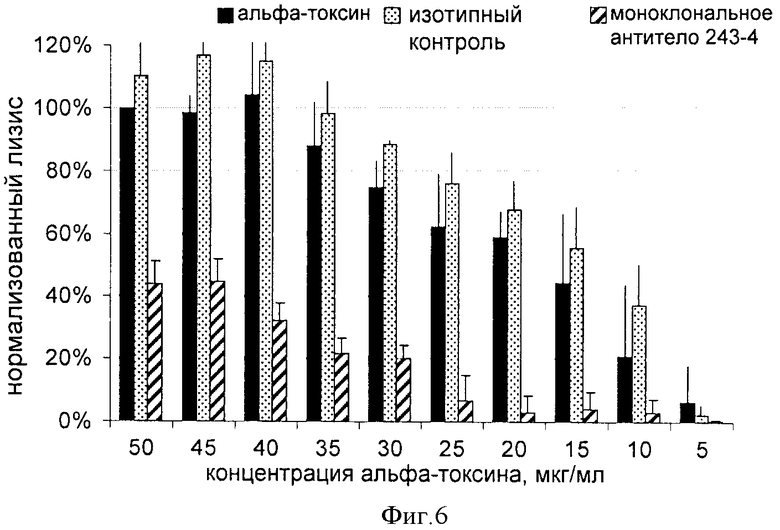
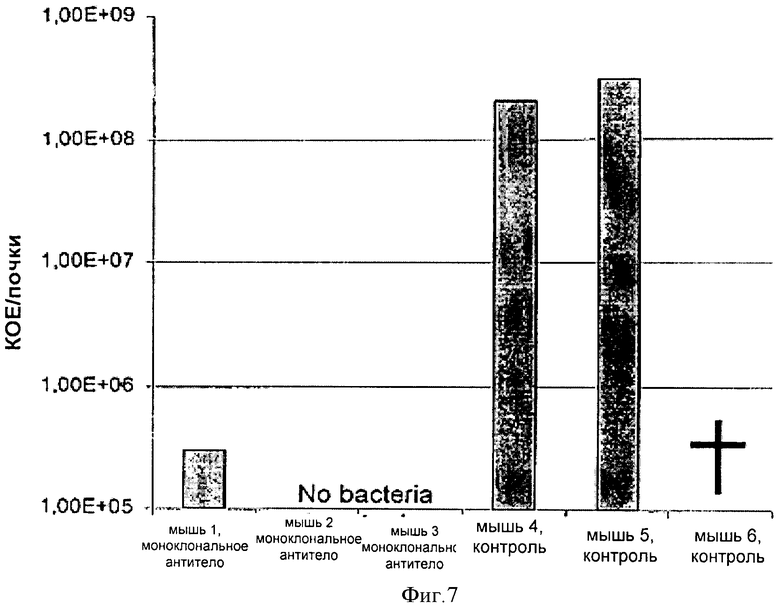
ФИГУРА 7
На фигуре 7 показан защитный эффект моноклонального антитела человека 243-4, введенного через катетер в верхней полой вене, модельным мышам с полиорганной инфекцией. Через 24 часа после установки катетера мышам с помощью катетера вводили 1х107 КОЕ штамма S. aureus US300 и 7,5 мг/кг моноклонального антитела человека 243-4 либо физиологического раствора. Через два дня мышам в группе лечения вводили вторую дозу антитела (5 мг/кг), а мыши из контрольной группы получали только физиологический раствор. Через пять дней после операции мышей подвергали эвтаназии для контроля бактериальной нагрузки почек и образования в них абсцесса. У мышей, иммунизированных моноклональным антителом 243-4, наблюдали значительно более низкую бактериальную нагрузку и отсутствие абсцессов в почках, а у контрольных мышей наблюдали высокую бактериальную нагрузка почек и выраженное образование абсцессов.
Следующие примеры иллюстрируют изобретение, но не ограничивают объем настоящего изобретения. Дальнейшие варианты будут очевидны для специалиста в данной области при изучении описания и с учетом общеизвестных данных.
ПРИМЕРЫ
Пример 1
Последовательность ДНК и аминокислотная последовательность антитела 243-4
Специфичность антител обусловлена последовательностями ДНК и аминокислот, соответственно. Были определены последовательности нуклеотидов ДНК вариабельной области тяжелой и легкой цепи. Для выделения РНК суспензию клеток гибридомы (5х105) осаждали центрифугированием и гомогенизировали в колонках Qiashredder (#79654, Qiagen). Затем мРНК выделяли из осадка гомогенизированных клеток гибридомы при помощи RNeasy-Kit (#74124, Qiagen) в соответствии с инструкцией производителя. кДНК синтезировали на основе изолированной мРНК путем обратной транскрипции с использованием обратной транскриптазы Superscript II (# 18064-022, Invitrogen). Гены 243-4 антитела амплифицировали из синтезированной кДНК с использованием набора Advantage 2 PCR Kit (#639206, Clontech) в соответствии с инструкцией производителя. Специфическую амплификацию генов антитела обеспечивали путем применения комбинации праймеров, предназначенных для амплификации вариабельной области модифицированного IgG человека (Welschof и соавт., 1995). Для амплификации вариабельных областей тяжелой (VH) и легкой цепи (VL) использовали набор прямых цепь-специфичных праймеров в комбинации с одним обратным праймером, который специфично присоединяется в константном домене тяжелой или легкой цепи (амплификация VH тяжелой цепи IgG в сочетании с VH1, VH2 и VH3; амплификация VL легкой цепи λ-типа в сочетании с VLλ1 VLλ 2/5, VLλ 3 VLλ 4a, VLλ 4b и VLλ 6; см. таблицу 1). Амплификаты, полученные в результате ПЦР, были клонированы в плазмиду pCR4-ТОРО набора для секвенирования ТОРО ТА Cloning Kit (# K457540, Invitrogen), и очищенная плазмидная ДНК была отправлена на секвенирование (Microsynth, Balgach, Швейцария) с применением плазмид-специфичных праймеров из ТОРО Cloning Kit (T3 и Т7, см. таблицу 1). Полученные последовательности ДНК обрабатывали и исследовали их первичную структуру с помощью пакета программ для анализа результатов клонирования (# 875-501-1787, Scientific and Educational Software). В результате анализа первичной структуры была получена консенсусная последовательность, которую затем анализировали с использованием V Base - базы данных всех зародышевых последовательностей человеческих вариабельных областей (http://imqt.cines.fr/IMGT_vquest/share/textes/). На основе результатов анализа первоначального секвенирования были сконструированы дополнительные праймеры, специфичные для тяжелой и легкой цепи (VL-atox as и VH-atoc as, см. таблицу 1). Указанные праймеры применяли для подтверждения идентифицированной последовательности антитела в участках гибридизации ранее использованных комбинаций праймеров.
Полученные указанным способом гены антител использовали для секвенирования, как описано выше и как показано на Фигурах 1 и 2.
Основания, указанные в скобках, показывают замены в предшествующем положении, число - процентную долю заменены нуклеотидов.
Пример 2:
Антигенная специфичность моноклонального антитела человека 243-4 (ELISA)
Антигенную специфичность антитела 243-4 оценивали по связыванию с панелью бактериальных токсинов (альфа-токсин: №120, List biological Laboratories, все другие токсины: собственного производства, Kenta Biotech AG). Анализ проводили методом твердофазного иммуноферментного анализа (ELISA) в планшетах для микротитрования (#439454, Nunc MaxiSorp), покрытых очищенными токсинами в концентрации 1 мкг/мл. После 24 ч инкубации при комнатной температуре планшеты для микротитрования блокировали в течение 2 ч 0,5% бычьим сывороточным альбумином. Связывание моноклонального антитела человека 243-4 (1 мкг/мл) с иммобилизованными токсинами определяли, используя вторичные козы к IgG человека, конъюгированные с пероксидазой хрена (HRP), в разведении 1:2000 (# 62-8420, Zymed Laboratories, Invitrogen). Реакцию останавливали НСl.
Оптическую плотность регистрировали на устройстве для считывания ELISA при 490 нм, используя программу Softmax Pro®, как показано на фигуре 3.
Пример 3:
Связывание с альфа-токсином эпидемических штаммов S. aureus в анализе методом вестерн-блот
Выделение альфа-токсина из двенадцати эпидемических S. aureus контролировали через 16 часов роста в среде BHI (# 255003, Becton Dickinson) при 37°С.
Штаммы получены из немецкого справочного центра по золотистому стафилококку (Институт Роберта Коха, Вернигероде) и представляют наиболее распространенные в настоящее время эпидемические штаммы S. aureus, вызывающие заболевания. Некоторые из этих штаммов выделяют меньше альфа-токсина по сравнению с другими, что обуславливает различную интенсивность сигнала.
Различные генотипы исследованных штаммов показаны в таблице 2. После культивирования бактерии осаждали центрифугированием, затем оптическую плотность супернатанта культуры нормировали к значению OD600=0,6 исходной бактериальной культуры. 25 мкл каждого супернатанта загружали на 4-20% ДСН-ПААГ гель (#ЕС60252, Invitrogen) и подвергали электроблоттингу в течение 1 часа. Один микрограмм очищенного альфа-токсина (#120, List biological Laboratories) загружали и исследовали параллельно с контролем. После блокирования в 5% сухом молоке в течение 1 ч нитроцеллюлозную мембрану (#LC2000, Invitrogen) инкубировали с 50 мкг/мл очищенного моноклонального антитела человека 243-4. Связывание антитела 243-4 с альфа-токсином детектировали с использованием вторичных антител козы к IgG человека, конъюгированных с пероксидазой хрена, в разведении 1:2000 (# 62-8420, Zymed Laboratories, Invitrogen), как показано на фигуре 4.
*1) Мультилокусное типирование последовательности.
*2) Клональный комплекс.
*3) Анализ вариабельной повторяющейся области гена белка A (spa).
Пример 4:
Определение аффинности (с использованием BIAcore)
Спектр поверхностного плазмонного резонанса измеряли с помощью системы BIAcore 2000 (BIAcore). Все эксперименты проводили в 20 мМ буфере MOPS, pH 7,0, 150 мМ NaCl и 0,1 мг/мл БСА. Сначала антитела козы к IgG человека (# 81-7100, Zymed Laboratories, Invitrogen) иммобилизовали на чипе СМ5 (BIAcore) с использованием реакции связывания аминов до примерно 13 200 отн. ед., как описано в руководстве BIAapplications. В дополнение к первичному ковалентно связанному покрытию антитело 243-4 иммобилизовали на сенсорном чипе посредством взаимодействия с предварительно иммобилизованным антителом к человеческому IgG, в результате чего сформировали дополнительный уровень иммобилизации, составляющий примерно 240 отн. ед. Для определения кинетики реакции антиген-антитело ячейку перфузировали растворами альфа-токсина с увеличивающимися концентрациями (3,9 нМ, 7,8 нМ, 15,62 нМ, 31,25 нМ, 62,5 нМ, 125 нМ, 250 нМ и 500 нМ, #120, List biological Laboratories) при скорости потока 50 мкл/мин. После каждого цикла измерений (5 мин ассоциации, затем 30 мин диссоциации) комплекс антитело-антиген восстанавливали посредством регенерации поверхности 10 мМ глицина-НСl при рН 1,7. Для вычисления константы диссоциации фазы ассоциации и диссоциации антитела 243-4 регистрировали, оценивали при помощи общей аппроксимации с использованием программного обеспечения BIAevaluation 4.1 (BIAcore AB, как показано на фигуре 5). Для в анализе путем общей аппроксимации учитывали только указанные концентрации антигена, что позволило произвести анализ, используя модель связывания Ленгмюра, давшую соотношение 1:1 (<125 нм, таблица 2), как описано в руководстве BIAcore.
Пример 5:
Модель острого повреждения легких на культуре клеток альвеолярного эпителия человека
Культуру клеток альвеолярного эпителия человека А549 высевали в среду RPMI (#R0883, Sigma-Aldrich) при плотности 3х105 клеток на лунку. Одновременно с этим серию растворов с увеличивающейся концентрацией альфа-токсина (5 мкг/мл - 50 мкг/мл, #120, List biological Laboratories) предварительно инкубировали только со средой, 20 мкг/мл антитела для контроля изотипа (IgG1 λ человека, очищенный белок миеломы; #I 5029, Sigma-Aldrich) или 20 мкг/мл очищенного моноклонального антитела 243-4. Через 4 ч инкубации при 37°С альфа-токсин или раствор альфа-токсин-антитело добавляли к клеткам и инкубацию продолжали еще 16 часов. После этого состояние клеток анализировали, используя набор для количественного определения лактатдегидрогеназы (ЛДГ) (#04744934001, Roche), который позволяет оценить высвобождение ЛДГ из клеток в культуральную среду, как показано на фигуре 6.
Пример 6:
Модель полиорганной инфекции на мышах
Самок мышей линии BALB/C (Charles River, Шульзфельд, Германия) весом 27-31 г акклиматизировали в течение 14 дней перед операцией. Мышей получали от поставщика, при этом было указано, что поставляемые мыши свободны от патогенов. Для размещения катетера мышей анестезировали, вводя ксилазин внутрибрюшинно (8 мг/кг массы тела)/кетамин (100 мг/кг массы тела). Для размещения одного однопросветного полиэтиленового катетера в верхней полой вене на левой стороне обритой шеи был сделан минимальный горизонтальный разрез кожи (внешний диаметр 0,6 мм, Föhr Medical Instruments, Зеехайм, Германия). Через двадцать четыре часа после установки мышам через катетер вводили 1×107 КОЕ штамма S. aureus US 300 (в 100 мкл) и 7,5 мг/кг очищенного антитела 243-4 либо физиологический раствор (в 50 мкл). Два дня спустя мышам получающей лечение группы вводили вторую дозу антитела (5 мг/кг), тогда как мыши из контрольной группы получали только физиологический раствор. Через пять дней после операции мыши были подвергнуты эвтаназии для контроля бактериальной нагрузки и формирования абсцесса в почках. Для этих целей почки асептически извлекали из тел животных, подвергнутых эвтаназии, и гомогенизировали в солевом растворе. Перед извлечением органа проверяли расположение катетера в верхней полой вене. Перед гомогенизацией орган макроскопически обследовали на наличие абсцесса. После этого серийные разведения гомогената органа культивировали на чашках МРК в течение 48 ч при температуре 37°С. Число колониеобразующих единиц (КОЕ) рассчитывали и фиксировали как КОЕ/почку, как показано на фигуре 7.
Ссылки:
Adlam С. et al., 1977: Effect of Immunization with Highly purified Alpha- and Beta-Toxins on Staphylococcal Mastitis in Rabbits. Infect and Immun. 17: 250-256.
Altschul S. et al., 1990: Basic local alignment search tool. J. Mol. Biol. 215: 403.
Benetti P.H. et al., 1998: Expression and characterization of the recombinant catalytic subunit of casein kinase II from the yeast Yarrowia lipolytica in Escherichia coli. Protein Expr Purif(13):283-290.
Corbin B.D. et al., 2008: Metal Chelation and Inhibition of Bacterial Growth in Tissue Abscesses. Science 319, 962-965.
DeLeo F.R. and Otto M., 2008: An antidote for Staphylococcus aureus pneumonia. J Exp Med 05(2): 271-274.
Devereux J et al., 1984: A comprehensive set of sequence analysis programs for the VAX Nucleic Acids Research 12 (12): 387.
Goode R.L., Baldwin J.N., 1974. Comparison of purified alpha-toxins from various strains of Staphylococcus aureus. Appl Microbiol. 28(1):86-90.
Henikoff S. and Henikoff J.G., 1992: Amino acid substitution matrices from protein blocks. Proc Natl Acad Sci USA. 1992 Nov 15; 89(22):10915-9.
Heveker N. et al., 1994a: A human monoclonal antibody with the capacity to neutralize Staphylococcus aureus alpha-toxin. Hum. Antibod. Hybridomas 5: 18-24.
Heveker N. et al., 1994b: Characterization of Neutralizing monoclonal Antibodies directed against Staphylococcus aureus alpha-toxin. Hybridoma 13: 263-270.
Kapral F.A. et al., 1980. Formation of Intraperitoneal Abscesses by Staphylococcus aureus. Infect and Immun. 30: 204-211.
Kielian Т. et al., 2001: Diminished Virulence of an Alpha-Toxin Mutant of Staphylococcus aureus in Experimental Brain Abscesses; Infect and Immun. (69), 6902-6911.
Levine MM. et al., 1983: New Knowledge on Pathogenesis of Bacterial Enteric Infections as Applied to Vaccine Development. Microbiol. Reviews (47), 510-550.
McElroy MC et al., 1999: Alpha-toxin damages the air-blood barrier of the lung in a rat model of Staphylococcus aureus induced pneumonia. Infect and Immun. 67, 5541-5544.
Needleman S.B. Wunsch C.D.,1970: A general method applicable to the search for similarities in the amino acid sequence of two proteins. J. Mol. Biol. 48: 443.
Persic L. et al., 1997: An integrated vector system for the eukaryotic expression of antibodies or their fragments after selection from phage display libraries. Gene 10;187(1):9-18.
Schwan W.R. et al., 2003: Loss of hemolysin expression in Staphylococcus aureus agr mutant correlates with selective survival during mixed infections in murine abscesses and wounds. FEMS Imm and Med Microbiol 28,23-28.
Tzianabos A.O. et al., 2001: Structural rationale for the modulation of abscess formation by Staphylococcus aureus capsular polysaccharides. PNAS 98: 9365-9370.
Von Köckrick-Blickwede M. et al., 2008: Immunological Mechanisms Underlying the Genetic Predisposition to Severe Staphylococcus aureus Infection in the Mouse Model. The American Journal of Pathology 173 (6), 1657-1668.
Wardenburg, J.B., and Schneewind O. 2008: Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 205:287-294.
Wardenburg, J.B. et al., 2007: Poring over pores: alpha-hemolysin and Panton-Valentine leukocidin in Staphylococcus aureus pneumonia. Nat. Med. 13:1405-1406.
Welschof M. et al.; 1995: Amino acid sequence based PCR primers for amplification of rearranged human heavy and light chain immunoglobulin variable region genes. J Immunol Methods. 179(2):203-14.
WO 2007/145689 Use of Alpha-Toxin for treating and preventing Staphylococcus infections.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, КОТОРЫЕ СПЕЦИФИЧНО СВЯЗЫВАЮТ АЛЬФА-ТОКСИН STAPHYLOCOCCUS AUREUS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620065C2 |
| АНТИТЕЛА ПРОТИВ СТЕНОЧНОЙ ТЕЙХОЕВОЙ КИСЛОТЫ И ИХ КОНЪЮГАТЫ | 2014 |
|
RU2687044C2 |
| АНТИТЕЛО, НАПРАВЛЕННОЕ ПРОТИВ ФАКТОРА СЛИПАНИЯ А (ClfA) S. aureus | 2019 |
|
RU2818805C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ГРИБКОВЫХ И БАКТЕРИАЛЬНЫХ ПАТОГЕНОВ | 2014 |
|
RU2717306C2 |
| КОМБИНАЦИИ АНТИТЕЛ К STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2804815C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ, ВЫЗВАННЫХ Staphylococcus aureus | 2015 |
|
RU2636780C1 |
| ЛЕЙКОЦИДИНЫ STAPHYLOCOCCUS AUREUS, ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2677140C1 |
| ЛЕЙКОЦИДИНЫ STAPHYLOCOCCUS AUREUS, ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2795545C2 |
| ЛЕЙКОЦИДИНЫ STAPHYLOCOCCUS AUREUS, ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2644237C2 |
| СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ СТАФИЛОКОККА ЗОЛОТИСТОГО И СВЯЗАННЫХ С НИМИ НАРУШЕНИЙ | 2012 |
|
RU2635462C2 |









Изобретение относится к биохимии, в частности к моноклональному антителу человека, специфичному к альфа-токсину S. aureus. Дополнительно настоящее изобретение относится к фармацевтическим композициям для лечения или предотвращения образования абсцесса в органе, содержащим по меньшей мере одно антитело или одну нуклеиновую кислоту, кодирующую указанное антитело. Изобретение позволяет расширить ассортимент антител, специфичных к альфа-токсину S. aureus. 10 н. и 13 з.п. ф-лы, 7 ил., 4 табл., 6 пр.
1. Моноклональное антитело, специфичное к альфа-токсину S. aureus, характеризующееся тем, что вариабельная область легкой цепи антитела содержит SEQ ID NO:1 в участке CDR1, SEQ ID NO:2 в участке CDR2 и SEQ ID NO:3 в участке CDR3, и при этом вариабельная область тяжелой цепи антитела содержит SEQ ID NO:4 в участке CDR1, SEQ ID NO:5 в участке CDR2 и SEQ ID NO:6 в участке CDR3, или фрагмент, или мутеин указанного антитела, способные связывать альфа-токсин S. aureus, причем мутеин моноклонального антитела несет по меньшей мере одну консервативную замену в одном из участков CDR в тяжелой или легкой цепи.
2. Моноклональное антитело по п.1, в котором вариабельная область легкой цепи антитела имеет аминокислотную последовательность SEQ ID NO:7 и вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:8, или фрагмент указанного антитела, способный связывать альфа-токсин S. aureus, или вариант указанного антитела, способный связывать альфа-токсин S. aureus, причем аминокислотная последовательность вариабельной области легкой цепи антитела по меньшей мере на 85% идентична SEQ ID NO:7, и аминокислотная последовательность вариабельной области тяжелой цепи антитела по меньшей мере на 85% идентична SEQ ID NO:8.
3. Моноклональное антитело по п.1, причем фрагмент антитела представляет собой Fab, F(ab′)2, одноцепочечное или доменное антитело.
4. Моноклональное антитело по п.1, причем антитело представляет собой антитело человека.
5. Моноклональное антитело по п.1, причем легкая цепь принадлежит к типу лямбда.
6. Моноклональное антитело по п.1, отличающееся тем, что тяжелая цепь принадлежит к типу IgG.
7. Моноклональное антитело по п.1, причем антитело способно специфично связываться с мономерной и олигомерной формами альфа-токсина S. aureus.
8. Моноклональное антитело по п.1, причем антитело содержит N-концевые, внутренние или C-концевые модификации.
9. Моноклональное антитело по п.8, причем модификация выбрана из по меньшей мере из одной из олигомеризации, гликозилирования или конъюгации с лекарственным средством или меткой.
10. Моноклональное антитело по любому из пп.1-9, получаемое из В-клеток млекопитающего или гибридомы, полученной путем слияния указанных В-клеток млекопитающего с клетками миеломы или гетеромиеломы.
11. Нуклеиновая кислота, кодирующая легкую цепь моноклонального антитела по любому из пп.1-10.
12. Нуклеиновая кислота, кодирующая тяжелую цепь моноклонального антитела по любому из пп.1-10.
13. Вектор для экспрессии антитела согласно любому из пп.1-10 или фрагмента указанного антитела, содержащий по меньшей мере одну нуклеиновую кислоту, кодирующую легкую цепь по п.11, или по меньшей мере одну нуклеиновую кислоту, кодирующую тяжелую цепь по п.12, или по меньшей мере одну нуклеиновую кислоту, кодирующую легкую цепь по п.11, и по меньшей мере одну нуклеиновую кислоту, кодирующую тяжелую цепь по п.12.
14. Вектор по п.13, причем указанный вектор дополнительно содержит промотор, функционально связанный с нуклеиновой кислотой и усиливающий ее экспрессию.
15. Клетка-хозяин для получения антитела согласно любому из пп.1-10 или фрагмента указанного антитела, содержащая вектор по п.14 или нуклеиновую кислоту по п.11 или 12.
16. Фармацевтическая композиция для лечения или предотвращения образования абсцесса в органе, содержащая по меньшей мере одно моноклональное антитело по пп.1-10, или по меньшей мере одну нуклеиновую кислоту по п.11 и по меньшей мере одну нуклеиновую кислоту по п.12 и фармацевтически приемлемый носитель или ингредиент.
17. Применение моноклонального антитела по любому из пп.1-9 для профилактики и лечения образования абсцесса в органе.
18. Применение нуклеиновой кислоты по п.11 и нуклеиновой кислоты п.12 для получения фармацевтической композиции для профилактики и лечения образования абсцесса в органе.
19. Применение по п.17 или 18, причем абсцесс в органе представляет собой брюшной абсцесс.
20. Применение по любому из пп.17 или 18, причем орган представляет собой почку, сердце, печень, легкое, мозг, кожу и селезенку.
21. Применение по любому из пп.17 или 18, причем формирование абсцесса обусловлено инфекцией S. aureus.
22. Тест-набор для диагностики инфекции S.aureus в образце, включающий по меньшей мере одно моноклональное антитело по любому из пп.1-10 или нуклеиновую кислоту по п.11 и нуклеиновую кислоту по п.12.
23. Применение по меньшей мере одного моноклонального антитела по любому из пп.1-10 для детектирования связывания с альфа-токсином в образце.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| BLOMQVIST L et.al | |||
| Production and characterization of monoclonal antibodies against Staphilococcus aureus alpha-toxin, Taxicon, vol.26, no3, p.265-273, 1988 | |||
| US 6288214, 11.09.2001. | |||

