Изобретение относится к молекулярной биологии, биотехнологии, препаративной биохимии, медицине и может быть использовано в качестве средства для профилактики гриппа, вызываемого существующими штаммами гриппа, а также возможными реассортантами.
Термин «гибридный белок» в данном тексте обозначает белок, полученный в результате экспрессии рекомбинантной молекулы ДНК, в которой соединены друг с другом в одной рамке считывания кодирующие участки нескольких разных генов.
Ежегодно вирус гриппа поражает от 5 до 15 процентов населения, вызывая острое заболевание дыхательных путей. Ежегодно разрабатываются и широко применяются вакцины против сезонного гриппа. Однако существует опасность пандемического распространения отдельных штаммов вируса гриппа, комбинация генов которых ранее не встречалась. Такие вирусы могут быть более агрессивны и могут характеризоваться высокой долей осложнений и смертельных случаев. В 2009 году в мире сложилась неблагоприятная ситуация, связанная с распространением высокопатогенного вируса гриппа A(H1N1)pdm/09. Заболевания были вызваны вирусом гриппа с новыми антигенными детерминантами. Особую тревогу у специалистов вызвал тот факт, что новый вирус являлся реассортантом вирусов, циркулирующих среди животных (свиней) и среди людей. По данным ВОЗ, в сравнении с коэффициентом инфицирования сезонным гриппом лиц, имевших контакты с больными людьми (от 5% до 15%), аналогичный коэффициент в отношении A(H1N1) pdm/09 оценивался в пределах от 22% до 33%.
Поверхностный белок вируса гриппа гемагглютинин (НА) представляет особый интерес для разработки вакцин. НА отвечает за связывание вируса с рецепторами клетки и таким образом определяет чувствительность организма к заболеванию. Мутации в НА обычно ведут к появлению вирусов с высокими эпидемическими, иногда пандемическими потенциями.
В-клеточный и Т-клеточный иммунный ответ развивается на антигенные детерминанты белков вируса - эпитопы. Подход к иммунизации против вирусных инфекций, а конкретно вируса гриппа, с использованием аминокислотных последовательностей, состоящих из совокупности эпитопов вируса, перспективен для развития поливалентного иммунного ответа на вирус и позволяет создать универсальную вакцину для предотвращения как сезонных вспышек, так и пандемии. Использование антигенных детерминант различных субтипов вируса гриппа в одной конструкции облегчает получение поливалентной вакцины и позволяет создавать эффективные вакцины против новых штаммов вирусов.
Уровень техники
Создание универсальной вакцины, способной обеспечить защиту от существующих и возможных реассортантных штаммов вируса гриппа, является необходимостью. Использование вакцин на основе рекомбинантных белков позволяет избежать рисков, связанных с введением вируса в организм, пусть и инактивированного. В настоящее время предпринимаются попытки создания такой универсальной безопасной вакцины.
Известно изобретение, по которому ген, кодирующий мультиэпитопный гемагглютинин вируса гриппа птиц, в составе векторной конструкции используется для продукции в растениях рекомбинантной мультиэпитопной вакцины (RU 2008139004 А, 01.10.2008). В патенте US 2009106864 (2009) и RU 2008139004 авторы использовали консенсусную последовательность гемагглютинина птичьего гриппа субтипа А, оптимизированную по кодонам для экспрессии в растениях. В патенте JP 2009102416 (2009) для экспрессии гемагглютинина вирусов гриппа А и В авторы использовали бакуловирусные конструкции в клетках насекомых. Недостатками подобных подходов является использование последовательности белка (гемагглютинина) только одного субтипа вируса гриппа (H5N1), что ограничивает спектр активностей вакцины. Также экспрессия в растениях и в клетках насекомых обеспечивает низкий выход белка по сравнению с экспрессией в Е. coli.
Известны гибридные полипептиды, включающие последовательность, имеющую большой процент соответствия поверхностному протеину НА вируса гриппа А с пятью иммунодоминантными антигенными участками (RU 92004487, 10.12.1992). Одно из предложенных применений таких полипептидов - для производства вакцин против соответствующих антигенов, для лечения и профилактики (включая иммунотерапевтические методы) в случае заражения такими антигенами. Однако не указано, гемагглютинин какого/каких субтипов используется. При разработке универсальной вакцины следует учитывать антигенные детерминанты гемагглютинина и также вируса гриппа В, т.к. иммуногенность вакцины ограничена теми штаммами, последовательности которых использованы.
Известна поливалентная вакцина от гриппа, содержащая поливалентный противогриппозный антиген, адъювант, агент, обеспечивающий проникновение вакцины и приемлемый эксципиент (CN101450209, 31.12.2008). В качестве антигенной компоненты заявлены любые из H1-H16 и N1-N9. Также известна поливалентная вакцина против гриппа, антигенной компонентой которой являются поверхностные белки вирусов A/H1N1, A/H3N2, гемагглютинин В-типа (CN1 01524538, 26.03.2009), адъювант - глицерол или гидроксид алюминия. Использование и приемлемого адъюванта, и антигенных детерминант в гибридной конструкции и только одного типа белка - гемагглютинина - позволит упростить и удешевить процесс производства вакцины, поскольку отпадает необходимость дополнительного подбора условий и контроля качества производства компонента.
Известна поливалентная вакцина на основе убитых вирусов, действующим веществом которой являются антигены A/H1N1, H3N2, В, H5N1 и A (H1N1)pdm/09 (CN 101732711, 31.12.2009). Указанные антигены получают выделением из убитых вирусов, что говорит о возможных проблемах при масштабном производстве вакцины, связанных с использованием вирусов как таковых, в особенности вирусов гриппа (например, необходимо создание условий работы с легко передающимися высокопатогенными штаммами вируса). Кроме того, указанные антигены являются отдельными белками, преимущества использования гибридной конструкции приведены выше.
Прототипом изобретения является смесь флагелл, содержащих, как минимум, 4 эпитопа белков вируса гриппа, взаимодействующих с человеческими клетками, причем каждый экспрессирован индивидуально в флагеллине Salmonella (WO 0032228, 30.11.1998). Указанные эпитопы - из группы, состоящей из: (i) одного эпитопа гемагглютинина, взаимодействующего с В-клетками, (ii) одного эпитопа гемагглютинина (НА) или нуклеопротеина (NP), которые могут связываться с молекулами МНС для представления рецепторам на Т-хелперах и (iii) хотя бы двух эпитопов нуклеопротеина (NP) или матриксного белка (М), которые рестриктируются МНС антигенами, превалирующими у представителей белой европеоидной расы и связываются с цитотоксическими лимфоцитами (CTL).
Флагеллин (FHC), взаимодействуя с Toll-like receptor-5 (TLR-5), стимулирует созревание макрофагов и дендритных клеток - антиген-презентирующих клеток, что приводит к выработке иммунного ответа (Mc Dermott P.F..High-affinity interaction between Gram-negative flagellin and a cell surface polypeptide results in human monocyte activation. Infect. Immun. - 2000. - V. 68. - p.:5525-5529; Means Т.К. et al. The Toll-like receptor 5 stimulus bacterial flagellin induces maturation and chemokine production in human dendritic cells. J.Immunol. - 2003. - V. 170. - p.:5165-5175).
На данный момент флагеллин является одним из наиболее перспективных и хорошо изученных адъювантов нового поколения. Результаты исследований показывают, что рекомбинантные белки, вводимые с флагеллином, имеют повышенные иммуногенные и антигенные характеристики. Ответы на них регистрируются в более короткие сроки и вызывают более сильный клеточный и гуморальный иммунный ответ (Balaram, 2008).
Недостаток данной разработки в многокомпонентности иммунизирующей смеси. Использование слитого белка, содержащего В-, Т-клеточные эпитопы разных субтипов вирусов гриппа А и В и флагеллин, позволит обеспечить сильный иммунный ответ при меньших затратах на производство. Кроме того, целесообразно использование также лишь компонентов флагеллина. Во флагеллине обнаружены два рецептор-активирующих участка в терминальных областях (а.о. 79-117 и а.о. 408-439) (Tonyia, 2001).
TLR-5 экспрессирован на клетках врожденного иммунитета, на эпителиальных и эндотелиальных клетках (Sebastiani G. et al. Cloning and characterization of the murine Toll-like receptor 5(Tlr5) gene: sequence and mRNA expression studies in Salmonella-susceptibleMOLF/Eimice. Genomics. - 2000. - V. 64. - p.230-240; Zarember K.A. and Godowski P.J. Tissue expression of human Toll-like receptors and differential regulation of Toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokines. J.Immunol. - 2002. - V.168. - p.554-561; Delneste, 2007). Ввиду этого целесообразно использование для иммунизации поверхности слизистой, что значительно облегчает доставку иммуногена.
Раскрытие изобретения
Предложена вакцина на основе высокоочищенного гибридного белка (SEQ ID NO:1), включающего фрагменты белков H1 (281 - 461 а.о.), Н3 (466 - 505 а.о.), Н5 (510 - 549 а.о.) вируса гриппа А, а также фрагмент гемагглютинина вируса гриппа В (554 -593 а.о.), а также компоненты флагеллина - FliC1 (1-169 а.о.) и FliC2 (182-276 а.о.) в качестве адъюванта, компоненты соединены гибкими мостиками. Представленные фрагменты белков представляют собой консервативные части гемагглютининов H1, Н3, Н5 и В, к которым в процессе естественной инфекции образуются специфические антитела, перекрестно реагирующие с гомологичными эпитопами среди различных штаммов вирусов гриппа А и В. Использование эпитопов нескольких белков позволяет увеличить эффективность вакцины, а использование гибких мостиков между эпитопами позволяет сохранить правильную пространственную укладку белка и, соотвественно, обеспечивает полноценное функционирование каждого эпитопа.
На основании выбранной аминокислотной последовательности были рассчитаны нуклеотидные последовательности соответствующих генов с учетом частоты встречаемости ко донов в белок-кодирующих генах Е. coli, при этом кодоны выбирались с учетом уменьшения dG соответствующих мРНК.
Технический результат от использования изобретения главным образом состоит увеличении валентности вакцины против гриппа: после прививания вакциной, заявленной в изобретении, у человека вырабатывается иммунитет к различным субтипам вирусов гриппа и А, и В, существующим, а также тем, которые могут появиться в результате реассортации любых штаммов вирусов гриппа. После вакцинации в организме вырабатываются антитела на эпитопы гемагглютинина от разных штаммов вируса гриппа. Таким образом формируется способность вырабатывать антитела в ответ на попадание в организм вируса гриппа. В дальнейшем, при попадании в организм любого вируса гриппа, иммунный ответ будет быстро вырабатываться, что выразится в том, что удастся избежать заражения, либо заболевание пройдет в легкой форме. Такой подход к вакцинации позволит элиминировать данное заболевание.
Технический результат от кодонной оптимизации нуклеотидной последовательности выражается в увеличении выхода белка в процессе производства.
Кроме того, использование в качестве адъюванта компонентов флагеллина - нетоксичного агента - позволит усилить вырабатывающийся на вакцину иммунный ответ.
Технический результат выражается в удешевлении и сокращении сроков производства вакцины, за счет производства лишь одного компонента - гибридного белка - взамен производства множества отдельных компонентов вакцинной смеси (прототип).
Для получения в соответствии с настоящим изобретением гибридного белка могут быть использованы стандартные методы молекулярной биологии и микробиологии, известные специалистам в данной области техники. Такие методы полно представлены в научной литературе.
Описание чертежей
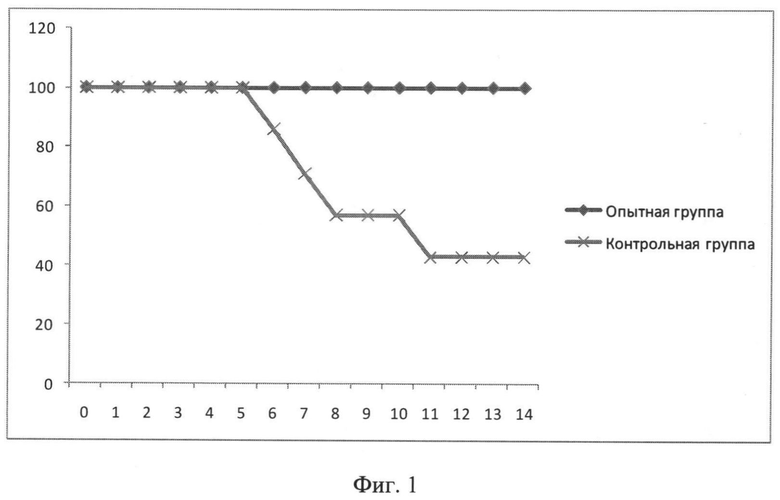
Фиг.1. Динамика выживаемости мышей после инфицирования 1 LD/50 вируса A/PR/8/34 (H1N1). На оси абсцисс отмечены дни после инфицирования, на оси ординат - процент выживших мышей.
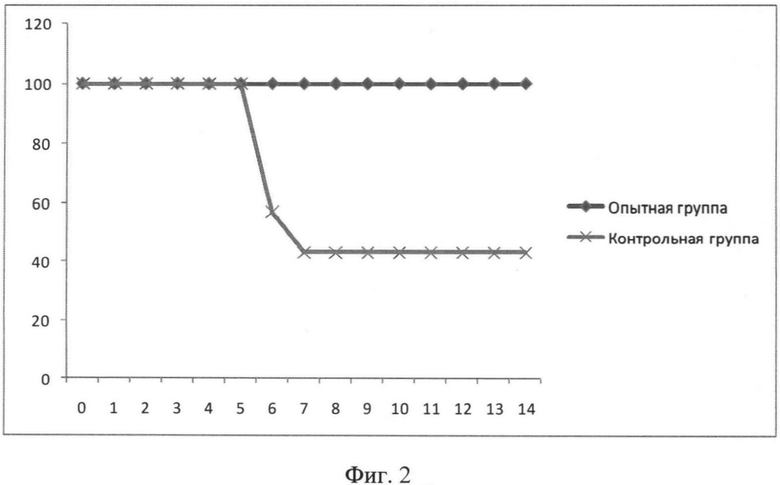
Фиг.2. Динамика выживаемости мышей после инфицирования 1 LD/50 вируса A/California/07/09(H1N1). На оси абсцисс отмечены дни после инфицирования, на оси ординат - процент выживших мышей.
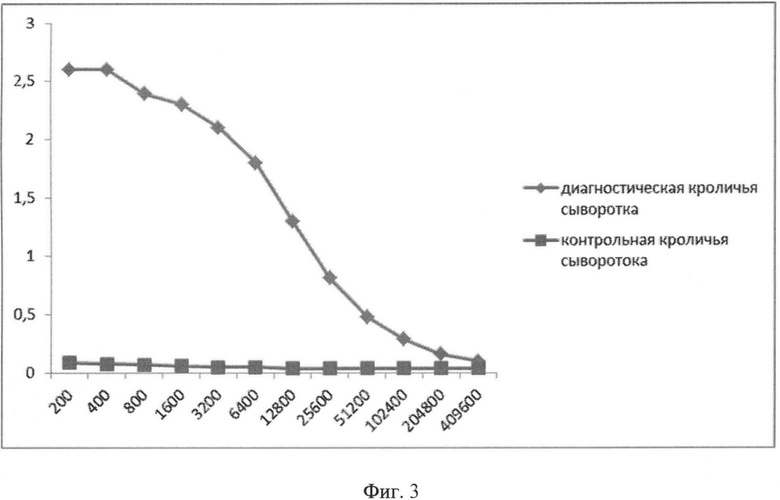
Фиг.3. Сывороточные IgG к вирусу гриппа B/Florida/04/06. По оси абсцисс - разведение сывороток (обратные величины). По оси ординат - оптическая плотность при длине волны 450 нм. Вирус сорбировали в концентрации 2 мкг/мл.
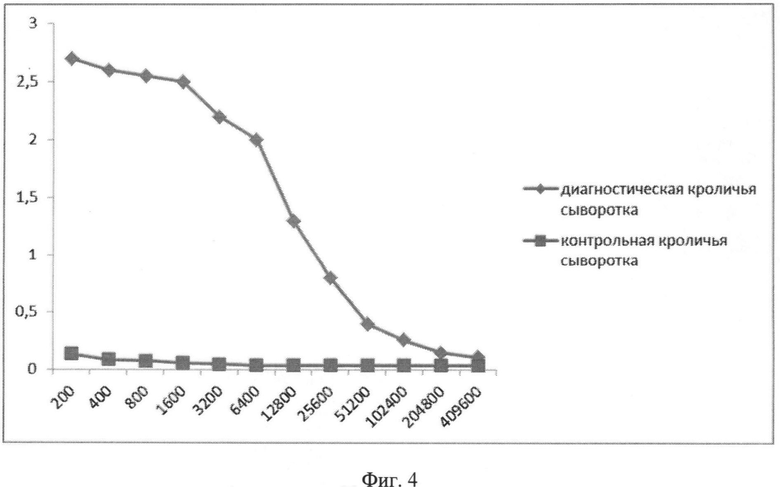
Фиг.4. Сывороточные IgG к гибридному белку. По оси абсцисс - разведение сывороток (обратные величины). По оси ординат - оптическая плотность при длине волны 450 нм. Рекомбинантный белок сорбировали в концентрации 3 мкг/мл.
Осуществление настоящего изобретения.
Пример 1. Моделирование гибридного белка
Спланированный гибридный полипептид является сложным мультидоменным белком (6 доменов: FliC1, FliC2, H1, Н3, Н5, В). Для моделирования мультидоменных белков были произведены следующие действия.
1. Определение границ доменов.
2. Построение модели целого белка для определения ориентации доменов.
3. Построение моделей для каждого домена (с использованием образцов 3D структур и ab initio).
4. Докинг моделей с использованием модели целого белка.
В спланированном гибридном полипептиде два домена имели образцы, а четыре нуждались в ab initio моделировании, кроме этого, в ab initio моделировании требовалось сформировать гибкие мостики между доменами.
Для получения более приближенных к реальности результатов в автоматическом режиме использовали алгоритм I-Tasser, признанный лучшим на последних трех CASP (Critical Assessment of protein Structure Prediction) - соревнованиях по моделированию белков. Данный анализ проводился в течение пяти дней. Однако даже с использованием данного мощного алгоритма получение адекватных данных для мультидоменного белка с необходимостью ab initio моделирования доменов и их границ не полностью достоверно (70%).
Для получения более точных данных разбили белок на используемые домены, провели их моделирование с использованеием I-Tasser и далее провели их докинг.
Смоделированный гибридный белок состоит из 593 а.о., представлена его аминокислотная последовательность - SEQ ID NO:1. Анализ аминокислотной последовательности данного белка с помощью программы ProtParam () показал, что гибридный белок стабилен и имеет молекулярную массу 63,6 кДа, pI 6,2.
Пример 2. Создание нуклеотидной последовательности, кодирующей гибридный белок
Перевели аминокислотную последовательность гибридного белка, включающего фрагменты FliC1, FliC2, H1, Н3, Н5, В, в нуклеотидную (1785 п.н.), оптимизировав последнюю для экспрессии в клетках E.coli (SEQ ID NO:2).
Синтез данной нуклеотидной последовательности осуществляли путем удлиннения взаимоперекрывающихся олигонуклеотидов согласно описанным методам (Majumder, 1992). Олигонуклеотиды представляли собой фрагменты гибридного гена длиной около 70 нуклеотидов со взаимоперекрывающимися участками длиной около 20 нуклеотидов. Основные требования к праймерам заключались в том, что их длина не должна была превышать 60 нуклеотидов, а участки гибридизации должны были быть не меньше 20 нуклеотидов. Кроме того, на концах олигонуклеотидов не должно было быть длинных участков с повторяющимися G или С. В ряде случаев подбор оптимальных праймеров осуществляли эмпирически путем сдвига праймера по отношеню к матрице или изменения длины праймера на 3-6 нуклеотидов. В общей сложности для синтеза гибридного гена длиной 1785 п.н. было использовано 65 праймеров. Синтезированные фрагменты по 300 п.н. выделяли с помощью гель-электрофореза и клонировали в плазмидном векторе pGEM-T Easy. Клонирование осуществляли с использованием рестрикционных сайтов Kpn1, SacII, EcoRV, BamHI или посредством „тупых" концов. После секвенирования фрагменты амплифицировали, после чего соединяли в нуклеотидную последовательность гибридного белка путем их сплавления методом полимеразной цепной реакции (ПЦР). После заключительного этапа синтеза гибридного гена путем лигирования фрагментов искусственный ген клонировали в вектор pGEM-T по рестрикционным сайтам KpnI и SacI. Полученный ген был фланкирован дополнительными рестрикционными сайтами EcoRI на 5'-конце и XhoI на 3'-конце. Далее искусственный ген переклонировали в экспрессионный вектор рЕТ24а по рестрикционным сайтам EcoRI и XhoI.
Пример 3. Создание плазмидной ДНК, кодирующей гибридный белок
По методике, описанной в примере 2, получали нуклеотидную последовательность гибридного белка для создания вакцины от гриппа.
Полученный ген клонировали в плазмиде рЕТ24а для последующей экспрессии. Для этого проводили реакцию лигирования гена и вектора рЕТ24а, с использованием соответствующего буфера и лигазы, при +20°C в течение 2 часов.
Смесь прогревали при +95°C в течение 10 мин и очищали от солей диализом на нитроцелюлозных фильтрах с диаметром пор 0,025 мкм (Millipore, США). Диализ проводили против раствора, содержащего 0,5 мМ ЭДТА в 10% глицерине, в течение 10 мин.
Пример 4. Создание штамма E.coli для амплификации плазмидной ДНК, содержащей гибридный ген
По методике, описанной в примере 3, получали нуклеотидную последовательность белка для создания вакцины от гриппа и клонировали ее в плазмиде рЕТ24а. Полученной плазмидой трансформировали клетки Е. coli штамма DH10B/R (Gibko BRL, США) с генотипом F-mcrA Δ(mrr-hsdRMS-mcrBC) φ0dlacZΔM 15 ΔlacX74 deoR recA1 endA1 araD139 Δ(ara, leu)769 galU galKλ- rpsL nupG методом электропорации.
После трансформации клетки инкубировали в SOC-среде (2% бакто-триптон, 0.5% дрожжевой экстракт, 10 мМ NaCl, 2.5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) в течение 40 мин при +37°C.
С помощью скрининга клеток E.coli на наличие плазмид на селективной среде, содержащей LB-arap, 100 мкг/мл канамицина, отобрали колонии клеток E.coli - штамм E.coli для амплификации плазмидной ДНК, содержащей гибридный ген.
Из выросших клонов выделяли плазмидную ДНК с использованием набора Wizard Minipreps DNA Purification System (Promega, США).
Очищенную плазмидную ДНК проверяли с помощью рестрикционного анализа и секвенирования. В ходе работы были отобраны клоны, содержащие фрагменты ДНК требуемого размера в составе плазмиды, из которых такие плазмиды были выделены для дальнейшей индукции экспрессии гена.
Пример 5. Создание штамма E.coli - продуцента гибридного белка
По методике, описанной в примере 4, получали нуклеотидную последовательность белка для создания вакцины от гриппа и клонировали ее в плазмиде рЕТ24а, амплифицировали полученную плазмиду в клетках E.coli штамма DH10B/R с последующим ее выделением.
Для экспрессии белка использовали клетки E.coli штамма BL21 Star (DE3) (Invitrogen, USA), с генотипом F- ompT hsdSB (rB-mB-) gal dcm rne 131 (DE3), содержащие в геноме λDE3 лизоген и мутацию rne 131. Мутированный ген rne (rne131) кодирует усеченную форму РНКазы Е, что уменьшает внутриклеточное разрушение мРНК, приводя к увеличению ее ферментативной стабильности. lon- и ompT-мутации по генам протеаз позволяют получать непротеолизированные рекомбинантные белки в больших количествах.
Подготавливали клетки Е. coli штамма BL 21 с генотипом F- ompT hsdSB (rB-mB-) gal dcm rne 131 (DE3) следующим образом. Инкубировали клетки при +37°C в течение ночи в 5 мл L-бульона, содержащего 1% триптон, 1% дрожжевой экстракт и 1% натрий хлористый. Разводили культуру свежим L-бульоном в 50-100 раз и выращивали на качалке при +37°C до оптической плотности 0,2-0,3 при длине волны 590 нм. При достижении оптической плотности более 0,3 культуру разводили свежим L-бульоном до оптической плотности 0,1 и растили 30 мин. Переносили 100 мл культуры в стерильную центрифужную пробирку и осаждали клетки при +4°C на 5000g в течение 10 мин. Супернатант сливали, клетки ресуспендировали в деионизованной воде в исходном объеме с последующим центрифугированием. Процедуру отмывки повторяли трижды. После отмывки осадок клеток ресуспендировали в малом объеме деионизованной воды и центрифугировали 30 сек при 5000 об/мин на микроцентрифуге.
Трансформацию компетентных клеток осуществляли методом электропорации. Для этого 1 мкл плазмидной ДНК добавляли к 12 мкл компетентных клеток, перемешивали и проводили электропорацию на генераторе высоковольтных импульсов ГВИ-1 (СПбГТУ, Санкт-Петербург) в стерильных ячейках при электрическом импульсе напряженностью 10 кВ/см длительностью 4 мсек.
После трансформации клетки инкубировали в SOC-среде (2% бакто-триптон, 0,5% дрожжевой экстракт, 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) в течение 40 мин при +37°C. 10-100 мкл клеточной суспензии высеивали на селективную LB-среду (Gibko BRL, США), содержащую канамицин (100 мкг/мл), для отбора клонов, содержащих плазмиды (штаммов-продуцентов).
Полученная после трансформации компетентных клеток штаммов Е. coli плазмида обеспечивала высокий уровень биосинтеза рекомбинантного белка, закодированного в ней.
Пример 6. Получение гибридного белка для создания вакцины от гриппа в клетках E.coli индукцией синтеза белка 0.2% лактозой по методу Штудиера
По методике, описанной в примере 5, получали нуклеотидную последовательность гибридного белка для создания вакцины от гриппа и клонировали ее в плазмиде рЕТ24а, амплифицировали полученную плазмиду в клетках E.coli штамма DH10B/R с последующим ее выделением, трансформировали ею клетки E.coli штамма BL21 для последующей индукции экспрессии целевого гена.
Для культивирования полученных штаммов-продуцентов использовали стандартную агаризованную LB-среду, содержащую канамицин в концентрации 100 мкг/мл и глюкозу в концентрации 1% для блокирования неспецифической экспрессии.
Индукцию экспрессии проводили при достижении культурой клеток оптической плотности 0.6-0.8 оптических единиц при длине волны 600 нм.
В качестве индуктора использовали 0.2% лактозу (Studier, 2005).
Для автоиндукции экспрессии по методу Штудиера (Studier, 2005) использовали среду PYP-5052, состоящую из 1% пептона (Gibco, США), 0.5% дрожжевого экстракта (Gibco, США), 50 мМ Na2HPO4, 50 мМ K2HPO4, 25 мМ (NH4)2SO4, 2 мМ MgSO4, 0.5% глицерола, 0.05% глюкозы и 0.2% лактозы.
В среду PYP-5052, содержащую канамицин в концентрации 100 мкг/мл, инокулировали единичную колония штамма-продуцента. Ферментаци проводили при +37°C в термостатированном шейкере роторного типа при 250 об/ мин в течение 20 часов до отсутствия существенного изменения ОП600 за 1 час. Отбирали аликвоту клеток на анализ экспрессии гена, кодирующего вакцинный белок, методом электрофореза в ПААГ, а оставшуюся биомассу осаждали центрифугированием при 9000g.
Белок выделяли из клеток E.coli посредством лизиса клеток. Клетки ресуспендировали в лизирующем буфере, содержащем 20 мМ трис-HCl рН 7,5, 5 мМ ЭДТА и 1 мМ феноксиметилсульфонилфторид, из расчета на 1 г клеток 5-7 мл буфера. Суспензию клеток обрабатывали ультразвуком 7 раз по 30 сек с интервалом в 30 сек (частота ультразвука составляет 22 кГц). Лизат центрифугировали 10 мин при +4°C, 5000 g. Надосадочную жидкость сливали, к осадку добавляли раствор 1 М мочевины из расчета 10 мл на 1 г клеток, интенсивно перемешивали. Повторяли центрифугирование. Супернатант сливали, осадок ресуспендировали в растворе 2М мочевины того же объема. Повторяли центрифугирование. Супернатант сливали.
Полученный препарат содержал по данным SDS-PAGE (PolyAcrylamide Gel Electrophoresis with Sodium dodecyl sulfate) около 98% гибридного белка в концентрации 1 мг/мл.
Условия выделения и очистки подбирались экспериментальным путем и могут варьировать в известных среднему специалисту в этой области значениях.
Пример 7. Получение очищенного препарата гибридного белка
К раствору гибридного белка добавляли MgCl2 до концентрации 6 мМ и проводили очистку гибридного белка с использованием метода иммобилизованной металлоаффинной хроматографии (ИМАХ) с использованием сорбента Ni-НТУ сефарозы. Связывание с данным сорбентом происходит за счет 6 остатков гистидина, имеющихся на N-конце полученного рекомбинантного белка. Белковый раствор фильтровали через ПВДФ фильтр с диаметром пор 0,22 мкн, к полученному фильтрату добавляли сорбент Ni-НТУ сефарозу, уравновешенную буфером для рефолдинга, из расчета, что 1 мл сорбента связывает не более 40 мг белка, и проводили связывание при постоянном перемешивании в течение 2 часов. Сорбент осаждали центрифугированием и в минимальном объеме наносили на гравитационную хроматографическую колонку, содержащую 2 мл сорбента.
Элюцию проводили ступенчатым градиентом имидазола от 20 до 300 с шагом в 50% от предыдущего, при этом на каждую ступеньку приходился объем элюирующего раствора, равный 5-20 объемам колонки. Выход и чистоту белка контролировали методом диск-электрофореза и измерением концентрации белка по методу Бредфорд.
Для определения содержания эндотоксинов приготавливали стандартный стоковый раствор эндотоксина с концентрацией 4000 EU на мл, полученный раствор был стабилен при хранении при +4°C в течение, по крайней мере, 2-х недель. Серию разведений стокового раствора эндотоксина с шагом в 2 раза приготавливали с использованием свободной от эндотоксинов воды в стерильных полистиреновых пробирках, заклеивающихся свободным от эндотоксина парафильмом. Готовили серию разведений образца субстанции с шагом концентрации 5, а после предварительного определения содержания эндотоксина - с шагом концентрации 2.
На дно каждого эппендорфа добавляли образец, воду или эндотоксин в 100 мкл, после чего добавляли 100 мкл лизата ЛАЛ-реактива и инкубировали в течение 1 часа при 37°C на водяной бане. Результаты оценивали по наличию или отсутствию плотного тромба на дне пробирки путем переворачивания пробирки.
Было показано, что в исследуемом образце отсутствуют факторы, ингибирующие образование тромба. Гель-тромб образовался при 1/8 разведении образца, т.е. при чувствительности метода 0,03 EU/мл, в образце с концентрацией 50 мкг/мл содержится менее 100EU.
Конформацию рекомбинантного гибридного белка при его синтезе в клетках E.coli определяли методом диск-электрофореза в ПААГ клеток E.coli, разрушенных с использованием ультразвука после проведения индукции экспрессии, при этом анализировали осадок и супернатант, образовавшиеся после осаждения клеточных обломков. В результате денситометрического анализа было показано, что рекомбинантный белок, синтезированный в клетках E.coli штаммов-продуцентов при индукции экспрессии 0.2% лактозой, на 100% находится в супернатанте, т.е. растворимой фракции.
Культивирование бактерий при различных температурах: 20°C, 30°C и 37°C, не вызывало изменения растворимости белка.
После проведения диск-электрофореза в ПААГ лизатов клеток E.coli после автоиндукции экспрессии 0.2% лактозой был осуществляли анализ уровня экспрессии целевого белка в клетках E.coli методом денситометрирования. В результате денситометрического анализа было показано, что в клетках E.coli гибридный белок накапливается в количестве 43% от общего клеточного белка.
Полученный уровень экспрессии не изменялся в штамме-продуценте в течение 6 пассажей, что свидетельствует о стабильном характере экспрессии анализируемого гена.
Пример 8. Получение рекомбинантного гибридного белка с использованием культивирования клеток E.coli с непрерывным добавлением питательных субстратов, предпочтительно глюкозы и дрожжевого экстракта
При культивировании штамма Escherichia coli для получения гибридного белка использовали метод периодических культур с подпиткой. Данный метод применяется для предотвращения негативных последствий лимита субстрата, при этом субстрат или другие необходимые компоненты добавляются либо непрерывно, либо по сигналу от какого-либо датчика. Для оптимизации выхода продуктов, выделяемых в среду, важно усилить биосинтетическую способность клеток бактерий, а метод культивирования с подпиткой позволяет продлить вторую фазу роста и повысить выход внеклеточных метаболитов. Данный метод можно использовать в случае потенциально токсичного субстрата (рекомбинантные белки токсичны для клеток E.coli), так как при этом его концентрация в среде будет поддерживаться на низком уровне.
Ограничение скорости поглощения субстрата скоростью его доставки оказывается способом преодоления «катаболитной репрессии» образования продукта.
Культура с подпиткой оказалась наиболее эффективным путем для достижения высокой плотности клеток и высокой продуктивности.
Штамм E.coli стерильно пересевали на скошенные косяки ЭДТА-содержащего агара и выдерживали в термостате 5 суток. Для получения инокулята осуществляли смыв культуры с косяков, засевали в жидкую среду с ЭДТА и культивировали 3-4 суток. После инокулят в количестве 10 мл переносили в стерильные 750-мл колбы с 200 мл стерильной жидкой среды и культивировали в течение 10 суток на качалке при 150-200 об/мин при температуре 28°C - 30°C.
Твердая питательная среда использовалась для получения свежей культуры штамма.
Жидкую питательную среду применяли для получения посевного материала и для периодического культивирования. Приготовление жидкой питательной среды
l)MgSO4·7H20 10 мл/л
2) CaCl2·2 H2O 20 мл/л
3) KH2PO4+NaH2PO4·12 H2O 10 мл/л
4) EDTA 10 мл/л
5) Микроэлементы 1 мл/л
(доводили до рН=4,2)+5,6 г/л ЭДТА:
FeCl2·4H2O 1,5 г/л
Н3ВО3 0,06 г/л
MnCl2·6 H2O 0,1 г/л
CaCl2·6H2O 0,12 г/л
ZnCl2 0,07 г/л
NiCl2·6 H2O 0,025 г/л
CuCl2·2H2O 0,015 г/л
Na2MoCl4 0,025 г/л
6) Витамины
Пиридоксин 20 мг
Тиамин 10 мг
Рибофлавин 10 мг
Никотиновая кислота 10 мг
Р - аминобензойная кислота 10 мг
Липоевая кислота 10 мг
Никотинамид 10 мг
Витамин В12 10 мг
Биотин 4 мг
Фолиевая кислота 4 мг
Все компоненты растворяли в 200 мл воды, стерилизовали при 0,5 атм. 30 минут и добавляли в жидкую питательную среду в количестве 1 мл/л.
Компоненты питательных сред взвешивали на технических и аналитических электронных весах и растворяли в дистиллированной воде. Опыт по приготовлению питательных сред показал, что ее удобно готовить из заранее стерилизованных концентрированных растворов.
Твердую питательную среду готовили как жидкую, с добавлением 3% агара.
Оптическая плотность (ОП600) культур клеток E.coli составила 9 О.Е. Клетки осаждали центрифугированием при 13000 g в течение 6 мин при температуре 10°C. Осадок клеток взвешивали, он составил 2.1 гр.
Клетки ресуспендировали в 30 мл буфера (50 мМ ТрисHCl, рН, 50 мМ ЭДТА, 20 мМ L-цистеин, рН8.6) и разрушали с использованием ультразвука (время озвучивания - 10 мин, время импульса - 30 сек, время паузы между импульсами - 30 сек, амплитуда - 70%). После разрушения клеток тельца включения осаждали с использованием центрифугирования при 30000g в течение 20 мин при температуре 10°C, вес сырого остатка телец включения из клеток составил 1.45 гр. Осажденные тельца включения отмывали с использованием последовательной смены нескольких буферов по следующей схеме:
1. Осадок телец включения ресуспендировали в 10 мл (последняя отмывка 15 мл) буфера для отмывки.
2. Ресуспендированные тельца включения перемешивали на горизонтальном шейкере в течение 1 часа при комнатной температуре.
3. Тельца включения осаждали центрифугированием при 30000g в течение 20 мин при температуре 10°C.
После пяти отмывок сырой вес телец включения составил 0.7 г.
Для солюбилизации белка из отмытых телец включения использовали 18 мл раствора (9 М мочевина, 2 мМ ЭДТА, 50 мМ ТрисHCl, рН8.6). Солюбилизированные тельца включения центрифугировали при 30000g в течение 20 мин при температуре 10°C. Образовавшийся супернатант переносили в новые фальконы и использовали для рефолдинга.
Рефолдинг проводили методом 10-кратного разведения солюбилизированных телец включения в буфере для рефолдинга при температуре +4°C по каплям. К раствору рефолдированного белка добавляли MgCl2 до 6 мМ и использовали для очистки белка метод иммобилизованной металлоаффинной хроматографии (ИМАХ) с использованием сорбента Ni-НТУ сефарозы. Белковый раствор фильтровали через ПВДФ фильтр с диаметром пор 0,22 микрона и к полученному фильтрату добавляли сорбент Ni-НТУ сефарозу, уравновешенный буфером для рефолдинга, из расчета, что 1 мл сорбента связывает не более 40 мг белка, после чего проводили связывание при постоянном перемешивании в течение 2 часов. Сорбент осаждали центрифугированием и в минимальном объеме наносили на гравитационную хроматографическую колонку, содержащую 2 мл сорбента. Элюцию проводили ступенчатым градиентом имидазола (20, 40, 100, 150, 200 и 300 мМ), при этом на каждую ступеньку приходился объем элюирующего раствора, равный 10 объемам колонки.
С использованием данного метода очистки был получен 41 мг гибридного белка с чистотой 97%.
Пример 9. Получение гибридного белка, очищенного перед растворением удалением растворимых клеточных компонентов, включающих ДНК, РНК, белки, липополисахариды посредством промывания буферным раствором, содержащим детергент гуанидингидрохлорид
После проведения ферментации и разрушения клеток E.coli, отмывки телец включения (см. Пример 8) проводили солюбилизацию белка 18 мл раствора (8 М GuHCl, 2 мМ ЭДТА, 50 мМ ТрисHCl, рН8.6), содержащего гуанидингидрохлорид. Солюбилизированные тельца включения центрифугировали при 30000g в течение 20 мин при температуре 10°C. Образовавшийся супернатант переносили в новые фальконы и использовали для рефолдинга.
Далее проводили рефолдинг рекомбинантного белка и их очистку (см. Пример 8). В результате очистки был получен 41 мг гибридного белка с чистотой 98%.
Пример 10. Определение иммуногенности препарата белка
Проводили оценку иммуногенности препарата полученного белка. Иммуногенность - это способность антигена инициировать иммунную систему к формированию эффекторов, нейтрализующих антигенную чужеродность. Чтобы спровоцировать иммунный ответ, антиген должен обладать иммуногенностью. Следует подчеркнуть, что иммуногенность - комплексная характеристика, которая зависит от свойств самого антигена, пути его введения и способа иммунизации.
Иммунизацию мышей проводили внутрибрюшинно, вводили 20 мкг гибридного белка. Иммунизацию проводили двукратно с интервалом в 2 недели. Животные были разделены на опытные и контрольные группы по 5-6 мышей в каждой.
Образцы крови получали от 5-6 мышей каждой группы через 2 недели после второй иммунизации из хвостовой вены. Для получения сыворотки кровь инкубировали в течение 30 минут при температуре 37°C. После образования сгустков крови образцы помещали на поверхность льда и охлаждали в течение 1-го часа с последующим центрифугированием в течение 15 минут при 400g. Сыворотку крови от мышей каждой группы пулировали и замораживали при температуре минус 20С.
Титры антител в сыворотках иммунизированных мышей определяли с помощью иммуноферментного анализа (ИФА). ИФА проводили общепринятым методом. Использовали 96-луночные планшеты (Greiner, Германия), на которых сорбировался гибридный белок (в карбонатном буфере, рН 9,5-9,6), выдерживали ночь при 4°C.
Гибридный белок денатурировали следующим образом. К образцу белка добавляли детергент Tween-20 до конечной концентрации 1% (w/v), инкубировали на водяной бане в течение 1 часа при 37°C. Далее образец центрифугировали 1 час при 200°C и 2000 g, отбирали супернатант, содержащий гибридный белок. От детергента избавлялись при помощи Detergent-OUTtmMicro Kit (Millipore), свободный от детергента образец концентрировали на установке SpeedVac до первоначального объема. Дополнительную обработку конечного препарата проводили 8М мочевиной в присутствии дитиотриэтанола (0,02M) с последующим диализом в течение ночи против карбонатного буфера (рН=8,5).
Планшет обрабатывали блокирующим буфером (0,01 М ФСБ, рН=7,2-7,4 с 5% ЭТС) в течение 1 часа при комнатной температуре, отмывали 3 раза ФБР с Tween-20. В лунки планшета добавляли 100 мкл 2-кратных разведений сыворотки (начиная с 1:200) в блокирующем буфере, инкубировали 1 час при комнатной температуре. Сыворотку исследовали в дубликате. В качестве конъюгата использовали кроличьи поликлональные антимышиные IgG (Abeam, UK) в разведении 1:8000, меченные пероксидазой хрена. В качестве субстрата использовали ТМБ. Учет реакции проводили при длине волны 450 нм. За титр принимали наибольшее разведение сыворотки, которое дает оптическую плотность, по крайней мере, в 2 раза больше, чем сыворотка неиммунизированных мышей в том же разведении.
Полученные результаты свидетельствуют о высокой иммуногенности полученного белка. При иммунизации гибридным белком в крови мышей выявлялись антитела к антигену, титр 51 200.
Таким образом, было показано формирование у мышей, иммунизированных гибридным белком, сильного иммунного ответа на него.
Пример 11. Протективность иммунного ответа, вызываемого гибридным белком, против различных штаммов вирусов гриппа
В настоящем исследовании были использованы мыши линии Balb/c (самки), 7-8 недель (массой 16-18 г), полученные из Учреждения Российской академии наук Института биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова РАН (Филиал) Питомник лабораторных животных «Пущино». Лабораторные животные были клинически здоровы и свободны от гельминтов (ветеринарное свидетельство 250 №0187942 от 27 ноября 2012 г.) Животных содержали в виварии ФГБУ "НИИ гриппа» Минздрава России в соответствии с действующими санитарными правилами.
Иммунизацию мышей проводили внутрибрюшинно, используя 20 мкг гибридного белка. Иммунизацию проводили двукратно с интервалом в 2 недели.
Для изучения протективности гибридного белка на модели летальной гриппозной инфекции использовали штаммы вируса гриппа A/California/07/2009 (H1N1) и A/PR/8/34 (H1N1), адаптированные к мышам. Титрование вирусов для определения одной дозы, вызывающей 50% гибели животных, проводили на мышах линии Balb/c (самки, возраст 6-7 недель). Замороженный вирус оттаивали, определяли мышиную летальную дозу (вызывающую 50% гибель) путем заражения мышей десятикратными разведениями вируса (по 4 мыши на разведение). Наблюдение за гибелью мышей вели в течение 14 суток после заражения. Титр вируса определяли по методу Рида и Менча. Для заражения мышей вирус вводили интраназально в дозе 1 LD/50 (доза, вызывающая гибель 50% мышей) и 5 LD/50 (доза, вызывающая гибель 90% мышей) по 50 мкл/мышь под легким эфирным наркозом. После заражения проводили ежедневное наблюдение за животными в течение 14 дней.
Гибель мышей начиналась на 6 день, в контрольной группе выживаемость мышей составила 43%. В опытной группе наблюдалась стопроцентная выживаемость мышей (Фиг.1).
Гибель мышей начиналась на 6 день. В контрольной группе выживаемость мышей составила 42%. В опытной группе наблюдалась стопроцентная выживаемость мышей (Фиг.2).
Пример 12. Формирование поливалентного иммунного ответа к различным штаммам вирусов гриппа А и В после иммунизации мышей гибридным белком
В настоящем исследовании были использованы мыши линии Balb/c (самки), 7-8 недель (массой 16-18 г), полученные из Учреждения Российской академии наук Института биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова РАН (Филиал) Питомник лабораторных животных «Пущино». Лабораторные животные были клинически здоровы и свободны от гельминтов (ветеринарное свидетельство 250 №0187942 от 27 ноября 2012 г.) Животных содержали в виварии ФГБУ "НИИ гриппа» Минздрава России в соответствии с действующими санитарными правилами.
Иммунизацию мышей проводили внутрибрюшинно, используя 20 мкг/200 мкл гибридного белка. Иммунизацию проводили двукратно с интервалом в 2 недели. Животные были разделены на опытные и контрольные группы по 5-6 мышей в каждой.
Образцы крови получали от 5-6 мышей каждой группы через 2 недели после второй иммунизации из хвостовой вены. Для получения сыворотки кровь инкубировали в течение 30 минут при температуре 37°C. После образования сгустков крови образцы помещали на поверхность льда и охлаждали в течение 1-го часа с последующим центрифугированием в течение 15 минут при 400g. Сыворотку крови от мышей каждой группы пулировали и замораживали при температуре минус 20°C.
Титры антител в сыворотках иммунизированных мышей определяли с помощью иммуноферментного анализа (ИФА). ИФА проводили общепринятым методом. Использовали 96-луночные планшеты (Greiner, Германия), на которых сорбировали антигены штаммов A/California/07/09 (H1N1), A/PR/8/34 (H1N1), A/Perth/16/09 (H3N2), A/Chicken/Kurgan/05/2005 (H5N1), в концентрации 2 мкг/мл (в карбонатном буфере, рН 9,5-9,6), выдерживали ночь при 4°C.
Гибридный белок денатурировали следующим образом. К образцу добавляли детергент Tween-20 до конечной концентрации 1% (w/v), инкубировали на водяной бане в течение 1 часа при 37°C. Далее образец центрифугировали 1 час при 20°C и 2000 g, отбирали супернатант, содержащий гибридный белок. От детергента избавлялись при помощи Detergent-OUTtmMicro Kit (Millipore), свободный от детергента образец концентрировали на установке SpeedVac до первоначального объема. Дополнительную обработку конечного препарата проводили 8М мочевиной в присутствии дитиотриэтанола (0,02 М) с последующим диализом в течение ночи против карбонатного буфера (рН=8,5).
Планшеты обрабатывали блокирующим буфером (0,01 М ФСБ, рН=7,2-7,4 с 5%ЭТС) в течение 1 часа при комнатной температуре, отмывали 3 раза ФБР с твином. В лунки планшета добавляли 100 мкл 2-кратных разведений сывороток (начиная с 1:200) в блокирующем буфере, инкубировали 1 час при комнатной температуре. Все сыворотки исследовались в дубликатах. В качестве конъюгата использовали кроличьи поликлональные антимышиные IgG (Abeam, UK) в разведении 1:8000, меченные пероксидазой хрена. В качестве субстрата использовали ТМБ. Учет реакции проводили при длине волны 450 нм. За титр принимали наибольшее разведение сыворотки, которое дает оптическую плотность, по крайней мере, в 2 раза больше, чем сыворотка неиммунизированных мышей в том же разведении.
При иммунизации гибридным белком в крови мышей выявлялись антитела ко всем исследованным штаммам вируса гриппа субтипа А в диапазоне от 1:3200 в случае H3N2 A/Perth/16/09 до 1:51200 в случае H1N1 A/California/07/09.
Таким образом, было показано формирование у мышей, иммунизированных гибридным белком, поливалентного иммунного ответа к различным штаммам вируса гриппа А и В.
Пример 13. Способность антител, образующихся после иммунизации животных убитым вирусом гриппа В, связываться с гибридным белком
В исследовании использовались кролики породы шиншилла, весом 2 - 2,5 кг, полученные из питомника лабораторных животных Российской Академии медицинских наук, пос. Рапполово Ленинградской области. Животных содержали в виварии НИИ гриппа СЗО РАМН в соответствии с действующими санитарными правилами.
На кроликах проведено изучение способности антител, образующихся после иммунизации животных убитым вирусом гриппа В, связываться с гибридным белком. Кроликов иммунизировали двукратно с интервалом 1 месяц, подкожно, убитым вирусом гриппа B/Florida/04/ с адъювантом Фрейнда в дозе 109 БОЭ. Кровь брали из ушной вены через 1,5 месяца после второй иммунизации. В ИФА (использовали Anti Rabbit Ig-HRP conjugate, Sigma-Aldrich, A 6154 в разведении - 1:5000) оценивали титры сывороточных антител (IgG) к вирусу гриппа B/Florida/04/06, гибридному белку, в качестве контроля использовали сыворотку крови неиммунизированных кроликов. Результаты проведенного исследования показывают, что антитела к поверхностным белкам вируса гриппа B/Florida/04/06 способны связываться с гибридным белком, титры антител к гибридному белку составили 1:204 000.
В ИФА (использовали Anti Rabbit Ig-HRP conjugate, Sigma-Aldrich, A 6154 в разведении - 1:5000) оценивали титры сывороточных антител (IgG) к вирусу гриппа B/Florida/04/06, рекомбинантному белку, входящему в состав вакцины, в качестве контроля использовали сыворотку крови неиммунизированных кроликов. Результаты представлены на Фиг.3 и Фиг.4. Титры антител в ИФА к штамму вируса гриппа B/Florida/04/06 и гибридному белку составили 409 600 и 204 800 соответственно.
Таким образом, показана иммунологическая поливалентность гибридного белка против вирусов гриппа А и В.


Изобретение относится к области биотехнологии и медицины и касается вакцины против гриппа, вызываемого известными штаммами вирусов гриппа А и В, а также возможными реассортантами. Предложенная поливалентная вакцина против гриппа получена на основе гибридного белка, включающего фрагменты белков Н1, Н3 и Н5 вируса гриппа А, фрагмент гемагглютинина вируса гриппа В и компоненты флагеллина - FliC1 и FliC2 (SEQ ID NO:1), соединенные гибкими мостиками. Кодирующая белок нуклеотидная последовательность (SEQ ID NO:2) оптимизирована для высокоэффективной экспрессии в клетках Escherichia coli. Использование охарактеризованной вакцины позволит обеспечить универсальную защиту от гриппа. 2 з.п. ф-лы, 4 ил., 1 табл., 13 пр.
1. Поливалентная вакцина против гриппа на основе гибридного белка (SEQ ID NO:1), включающего фрагменты белков H1 (281 - 461 а.о.), Н3 (466 - 505 а.о.), Н5 (510 - 549 а.о.) вируса гриппа А, а также фрагмент гемагглютинина вируса гриппа В (554 - 593 а.о.), а также компоненты флагеллина - FliC1 (1-169 а.о.) и FliC2 (182-276 а.о.) в качестве адъюванта, соединенные гибкими мостиками.
2. Поливалентная вакцина против гриппа по п.1, характеризующаяся тем, что гибридный белок закодирован в нуклеотидной последовательности SEQ ID NO:2, оптимизированной по кодонному составу для эффективной экспрессии в клетках Escherichia coli.
3. Поливалентная вакцина против гриппа по п.1, характеризующаяся тем, что получена способом, включающим создание рекомбинантной ДНК, кодирующей гибридный белок по п.1, введение такой ДНК в векторную конструкцию, обеспечивающую высокий уровень ее экспрессии в клетках бактерии Escherichia coli, введение данной векторной конструкции в клетки бактерий Escherichia coli, продукцию гибридного белка в указанном организме, его выделение, очистку и смешивание с физиологически приемлемым носителем.
| WO 2000032228 A2, 08.06.2000 | |||
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| US 20120064117 A1, 15.03.2012 | |||
| US 20100297174 A1, 25.11.2010 | |||

