Область изобретения
Изобретение относится к области биотехнологии, генной инженерии и может быть использовано для получения антиоксидантного препарата.
Уровень техники
В настоящее время считается общепризнанным, что острые патологические процессы в организме человека сопровождаются мощным окислительным стрессом в различных тканях организма (гиперпродукция активных форм кислорода), который является одним из основных поражающих факторов при развитии острых воспалительных процессов.
Широкий спектр заболеваний, в патогенезе которых важную роль играет окислительный стресс, вынуждает искать пути эффективной нейтрализации активных форм кислорода (АФК), так как в подавляющем числе случаев ситуацию можно существенно скорректировать, используя антиоксидантные препараты. В медицине широко используются лекарственные препараты антиоксидантного действия как природного, так и синтетического происхождения. Однако в большинстве случаев они оказываются относительно малоэффективными и используются как дополнительные лекарственные препараты или как профилактическое средство. Этот факт стимулировал исследования принципиально новых препаратов антиоксидантного действия. Одним из перспективных направлений может быть использование ферментов-антиоксидантов в качестве основы для лекарственных препаратов антиоксидантного действия, так как эффективность этих ферментов на несколько порядков выше, чем антиоксидантная активность широко используемых низкомолекулярных соединений.
К настоящему времени уже имеются экспериментальные данные, свидетельствующие о том, что ферменты-антиоксиданты, в первую очередь пероксиредоксины и супероксиддисмутаза, могут быть перспективны для использования в качестве основного компонента лекарственных препаратов антиоксидантного действия при лечении легочных и сердечно-сосудистых заболеваний, а также при трансплантации и консервации органов.
Известны несколько представителей рекомбинантных белков, используемых в качестве антиоксидантных средств в составе композиций для лечения млекопитающих. Одним из представителей таких белков является пероксиредоксин PrxVI.
Известны: рекомбинантная плазмидная ДНК на основе экспрессионной плазмиды рЕТ-23а, штамм Escherichia coli, трансформированный данной рекомбинантной плазмидной ДНК, - продуцент рекомбинантного белка пероксиредоксина PrxVI [патент RU №2280448, класс А61К 31/38, А61К 38/05, опубл. 10,05.2004 г.]. Известно применение фрагмента пероксиредоксина PrxVI, обладающего высокой эффективностью в составе композиций для лечения млекопитающих при меньших размерах белка [патент США №8003345, Fesenko, et al. Antioxidant pharmaceutical compound, method for producing polypeptide and method of cure, опубл. 23.08.2011].
Известно применение фармацевтической композиции для предотвращения и лечения рестеноза, в состав которой в качестве активного ингредиента входит белок пероксиредоксин Prx2 или генная конструкция для его экспрессии в клетках организма [патент US 7794708, Kang, et al. Pharmaceutical composition for preventing and treating restenosis comprising peroxiredoxin 2 protein as an active ingredient and composition for screening therapeutics of restenosis comprising peroxiredoxin 2 gene or its protein product, and method for screening therapeutics ofrestinosis using said composition, 14.09.2010].
Известен способ лечения болезней сердца путем прямой инъекции в сердце вектора, содержащего нуклеотидную последовательность, кодирующую белок АОР-1 (аналог белка PrxVI), для улучшения продукции белка АОР-1 в сердце, защиты сердца от повреждений и поддержания пульса [патент US 7598228, Hattori et al. Therapeutic methods and agents for diseases associated with decreased expression of АОР-1 gene or AOP-1, 06.10.2009].
Другое направление при создании эффективных антиоксидантных средств относится к попыткам создания антиоксидантов состоящих, например, из двух разных типов белков.
Известны примеры формирования слитых белков, в которых белки или их домены, например, ответственные за транспорт в клетку, были ковалентно связаны с целевыми белками, в качестве которых выступала Cu/Zn супероксиддисмутаза (СОД) [патент США №7306944, Choi S. Park J. Advanced cell-transducing transport domain-target protein-transport domain fusion protein and uses thereof, 2007-12-11].
Известны примеры формирования бифункциональных рекомбинантных белков для млекопитающих, в состав которого входит киназа и эндонуклеаза [патент США №7439061, Kaufman et al. DNA encoding the novel mammalian protein, Irelp, 21.10.2008].
Наиболее близким к техническому решению настоящего изобретения является применение антиоксидантной терапии с использованием комбинированного применения СОД и каталазы (КАТ) в экспериментах по созданию ковалентно связанного биферментного конъюгата СОД-хондроитинсульфат - КАТ. [Maksimenko AV, Golubykh VL, Tischenko EG. The combination of modified antioxidant enzymes for anti-thrombotic protection of the vascular wall: the significance ofcovalent connection ofsuperoxide dismutase and catalase activities. J Pharmacy Pharmacol 2004; 56: 1463-8., А.В.Ваваев, А.В.Максименко. Ферментные антиоксиданты на пути к практической медицине (http://www.consilium-medicum.com/article/18782)]. Для получения биферментного конъюгата, СОД-1 была ковалентно присоединена через гликозаминогликан сосудистой стенки - хондроитинсульфат (ХС) - к КАТ и получен аддукт СОД-ХС-КАТ. Продукт супероксиддисмутазной реакции - перекись водорода - оказывался субстратом для последующей КАТ-трансформации в безопасные в этих условиях воду и молекулярный кислород. Однако данными авторами в качестве пероксидазы была взята каталаза, что существенно ограничило спектр нейтрализуемых гидропероксидов (разрушается только перекись водорода). Сам процесс создания химерного бифункционального фермента был осуществлен химической сшивкой коммерчески доступных ферментов, что привело к существенному уменьшению ферментативной активности каждой составляющей химерного белка. Последний факт связан с отсутствием избирательности при химической сшивке белков, при которой может экранироваться значительная часть активных центров ферментов. Тем не менее, был показан существенный терапевтический эффект данного химерного белка при защите органов от оксидативного стресса, возникающего при ишемии-реперфузии. Показано, что биферментный коньюгат зарекомендовал себя эффективным агентом антитромботического действия.
Одной из задач предлагаемого изобретения является устранение указанных недостатков химической сшивки ферментов за счет конструирования химерной нуклеиновой кислоты, кодирующей бифункциональный белок, содержащий аминокислотную последовательность пероксиредоксина человека (Prx6) и аминокислотную последовательность Mn-супероксиддисмутазы (SOD). При этом белок сохраняет высокую антиоксидантную активность как Prx6, так и Mn-SOD.
Другой задачей предлагаемого изобретения является создание высокопродуктивного штамма-продуцента для получения бифункционального полипептида PSH.
Следующей задачей является применение бифункционального полипептида в медицине, в частности, в составе композиций, используемых при реперфузии сердца.
Поставленные задачи решаются путем создания бифункционального фермента-антиоксиданта PSH (Пероксиредоксин-Супероксиддисмутаз-Гис-таг), который способен нейтрализовать все основные типы активных форм кислорода (супероксид радикалы, неорганические и органические гидропероксиды и пероксинитриты).
В начале создается генная конструкция, кодирующая последовательно оба белка, затем она включается в экспрессирующий вектор рЕТ23(ДЕ3), на основе которого создается штамм Е.coli pET23(PSH), обеспечивающий синтез полипептида PSH
Сущность изобретения
Одним из объектов изобретения является химерная нуклеиновая кислота, кодирующая гибридный белок PSH, включающий человеческий пероксиредоксин Prx6 и марганцевую супероксиддисмутазу MnSOD, обладающий антиоксидантной активностью супероксиддисмутазы и пероксидазы, первичная структура которой описывается аминокислотной последовательностью SEQ ID NO:1.
Другим объектом изобретения является рекомбинантный вектор экспрессии pET23(PSH), содержащий химерную нуклеиновую кислоту, кодирующую белок PSH с аминокислотной последовательностью SEQ ID NO:1, оперативно встроенную в плазмиду pET22b(+) по сайтам рестрикции NdeI и XhoI с С-концевой локализацией полигисгидинов.
Следующим объектом изобретения является гибридный белок PSH для реперфузии сердца, первичная структура которого описывается аминокислотной последовательностью SEQ ID NO:1.
Перечень фигур
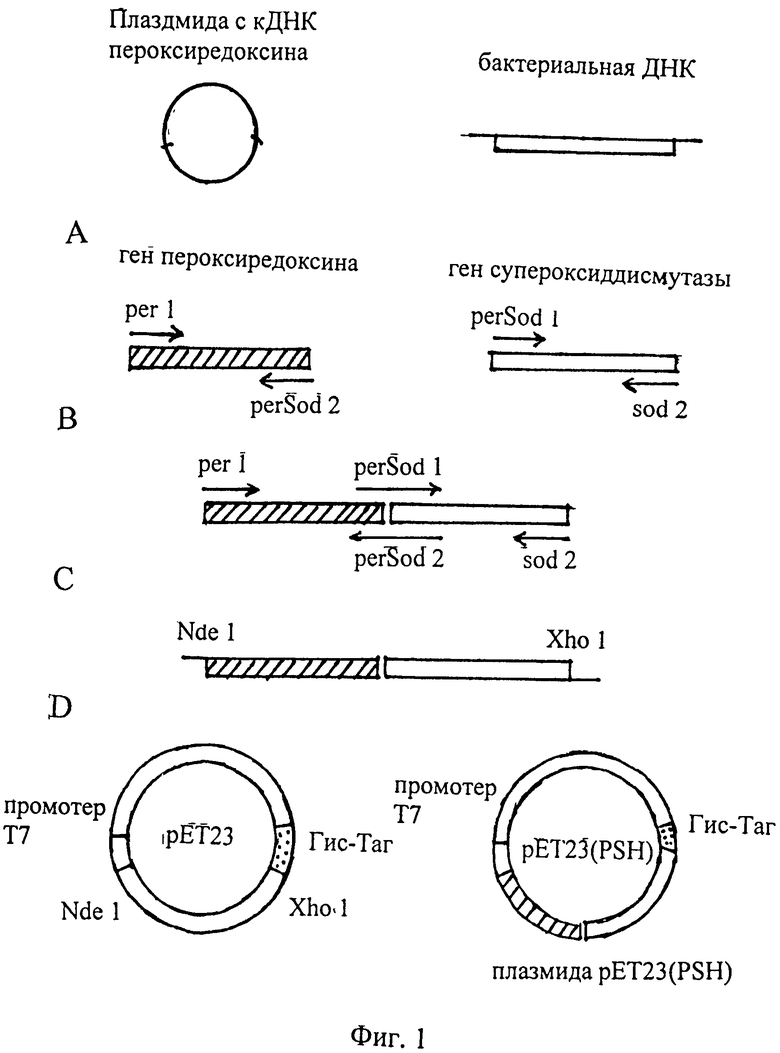
Фиг.1 Получение плазмиды pET23(PSH). Где А - получение генов пероксиредоксина и супероксиддисмутазы; В - сшивание генов ПЦР с праймерами per1 и sod2; С - рестрикция ПЦР фрагмента ферментами NdeI и XhoI; D - встраивание обработанного ПЦР фрагмента в плазмиду рЕТ23.
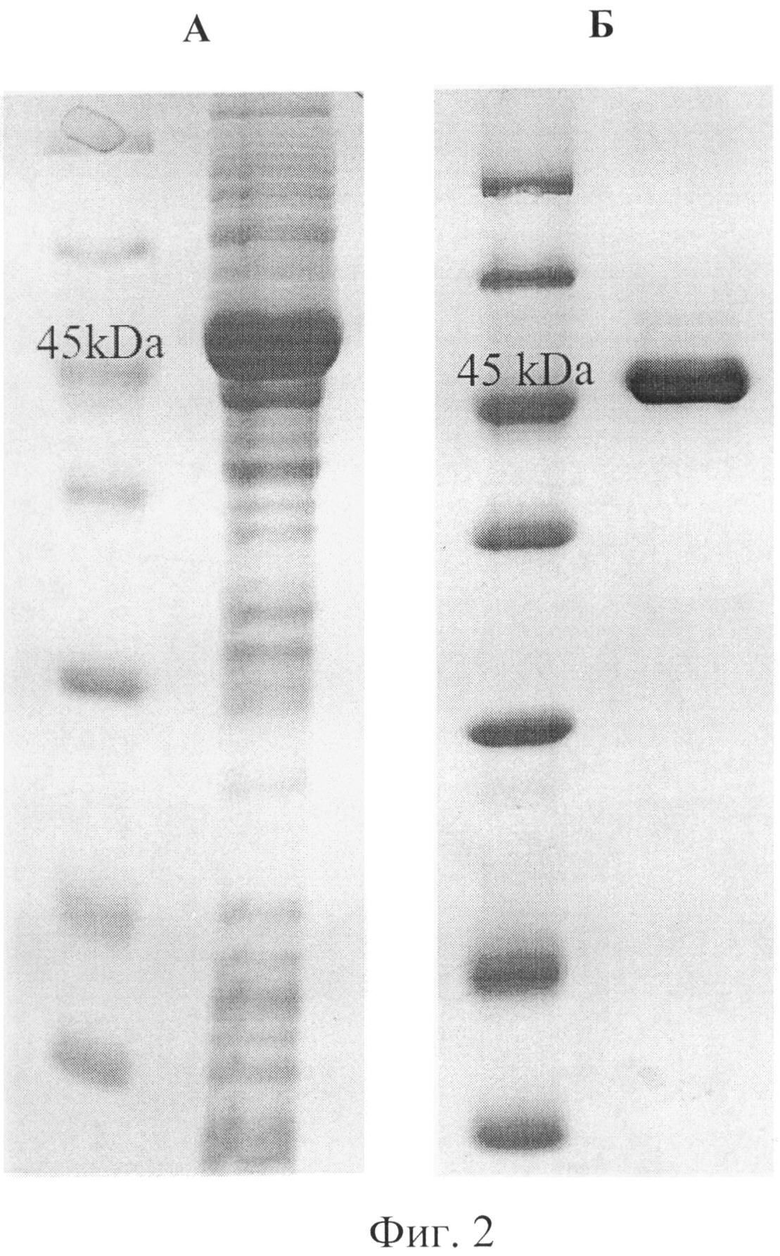
Фиг.2 Получение гибридного белка PSH. А - наработка белка в E.coli BL21(DE3). Б - очистка белка на Ni-NTA агарозе.
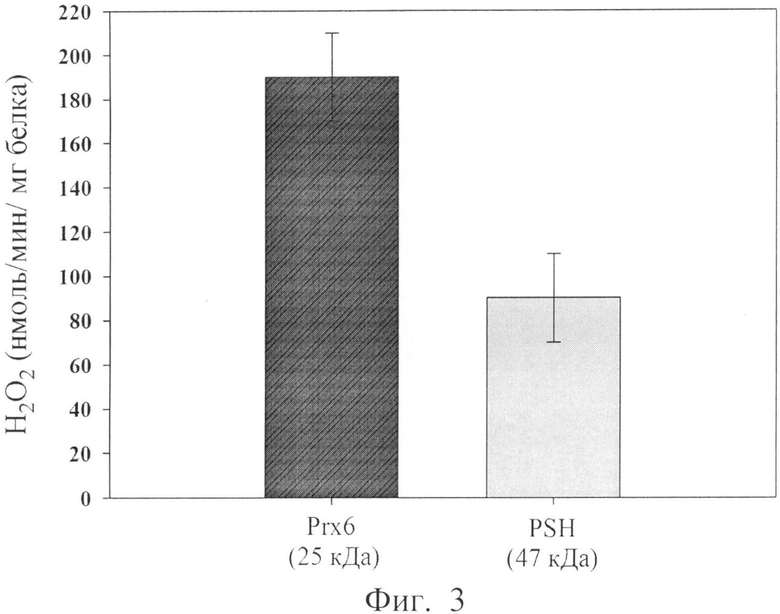
Фиг.3. Пероксидазная активность белка PSH. В качестве контроля взят рекомбинантный пероксиредоксин 6 человека (Prx6).
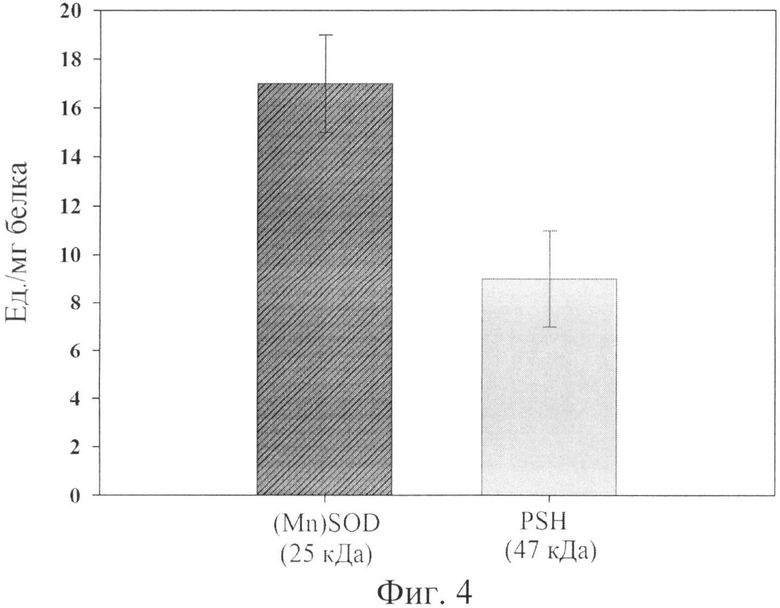
Фиг.4. Супероксиддисмутазная активность белка PSH. В качестве контроля использована супероксиддисмутаза.
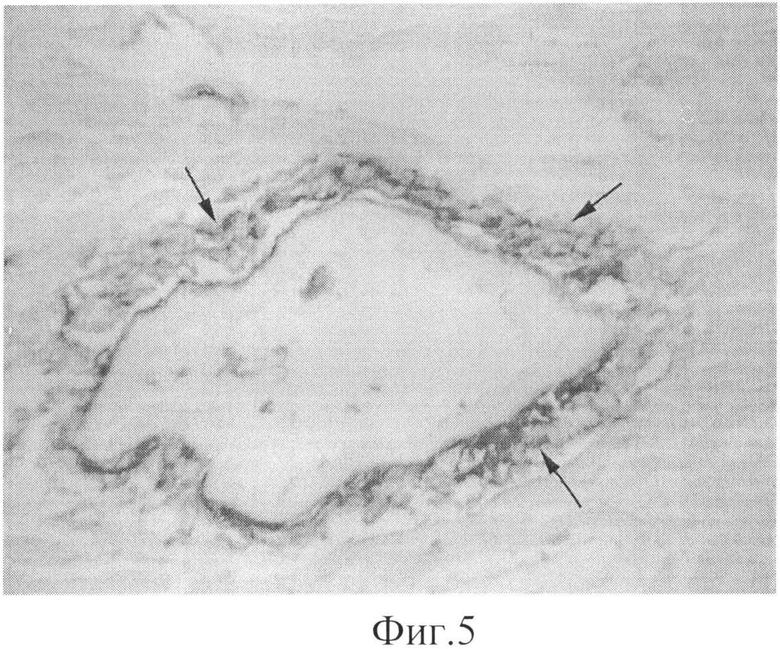
Фиг.5. Локализация эндогенного пероксиредоксина Prx6 в сердце. Увеличение × 1000. Первичные антитела - поликлональные антитела кролика к перокстредоксину 6; вторичные антитела - антитела осла к иммуноглобулинам кролика, конъюгированным с щелочной фосфатазой (USBiological, USA).
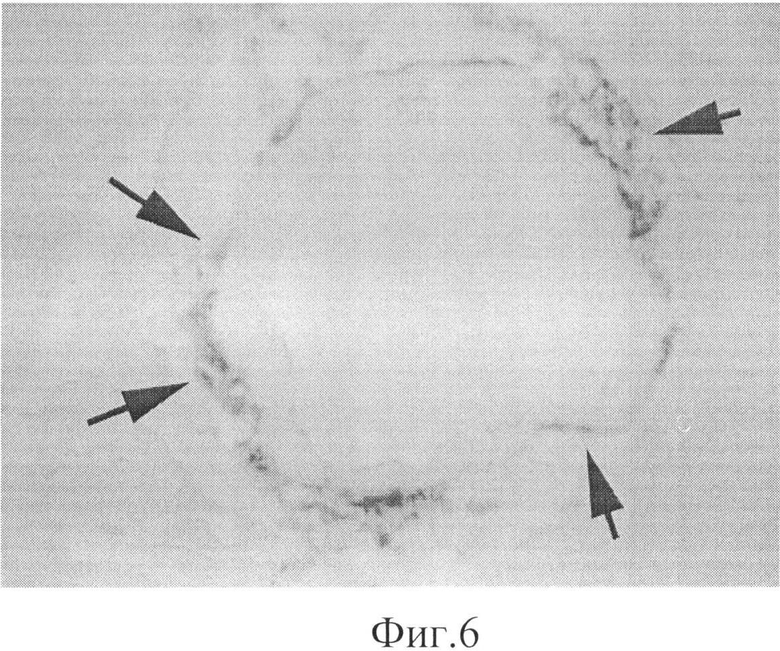
Фиг.6. Локализация экзогенного белка PSH при реперфузии сердца в присутствии перекиси водорода.
Описание
Для решения поставленных задач и получения эффективного полифункционального химерного белка, способного нейтрализовать широкий спектр активных форм кислорода (бифункциональный белок с совмещенными супероксиддисмутазной и пероксидазной активностями), необходимо сделать выбор как между разными типами супероксиддисмутаз, так и различными классами и подклассами пероксидаз.
В качестве первого компонента фермента-антиоксиданта был выбран белок пероксиредоксин (Prx6), обладающий широкой субстратной специфичностью по отношению к гидропероксидам. Пероксиредоксины (Prx1-Prx6) широко представлены в живом мире и входят в семейство неселеновых пероксидаз, осуществляющих разрушение пероксидов как органической, так и неорганической природы. Для катализа Prx используют консервативный остаток цистеина (Cys) в активном центре и не содержат каких-либо окислительно-восстановительных кофакторов (гем, флавины или ионы металлов). Наиболее ярким представителем класса пероксиредоксинов является пероксиредоксин 6 (Prx6), содержащий только один активный остаток цистеина (Cys47). Белок Prx6 обнаружен во всех клетках млекопитающих, однако его наибольшие количества выявляются в цитоплазме эпителиальных клеток: легких, дыхательных путей, желудочно-кишечного тракта и ротовой полости, а также в клетках печени и поджелудочной железы. Prx6 способен восстанавливать широкий спектр пероксидов, включая Н2О2, небольшие органические гидропероксиды, такие как гидропероксид трет-бутила и гидропероксид кумена, гидропероксиды жирных кислот и гидропероксиды фосфолипидов. При этом Prx6 способен восстанавливать как свободные молекулы гидропероксидов жирных кислот, так и в составе фосфолипидов. Пероксидазная активность Prx6 человека составляет от 1.8 до 5 мкмоль/мин/мг белка. В отличие от остальных пероксиредоксинов, Prx6 помимо пероксидазной активности, в кислой среде проявляет активность Са2+ независимой фосфолипазы А2 (iPLA2), которая может играть важную роль в метаболизме фосфолипидов эпителиальных клеток легких [Fisher А.В., Dodia С., Feinstein S.I., Но Y.S. Altered lung phospholipid metabolism in mice with targeted deletion oflysosomal-type phospholipase А2. J. Lipid Res. Vol.46, pp.1248-1256 (2005)].
Для реализации задачи нейтрализации всех основных типов активных форм кислорода, входящих в группу, состоящую из супероксидрадикалов, неорганических и органических гидропероксидов и пероксинитритов, в качестве второго компонента химерного бифункционального белка был выбран один из представителей супероксиддисмутаз - марганецсодержащая супероксиддисмутаза E.coli (SOD2) [Roberts В, Hirst R. Identification and characterisation of a superoxide dismutase and catalase from Mycobacterium ulcerans. J. Med. Microbiol. Vol.45, pp.383-387. (1996)].
Супероксиддисмутазная активность (F) составила ~9 ед./мг белка, что практически соответствует ожидаемой величине ~17 ед./мг белка Mn-SOD E.coli [Roberts В. and Hirst R., 1996], где одна ед. соответствует количеству белка, необходимому для проявления 50% дисмутации супероксидрадикалов.
Неочевидность технического решения по получению бифункционального белка PSH, представленного в настоящем изобретении, состоит в том, что белок пероксиредоксин обладает сложной третичной структурой и сопряжение двух белков СОД2 и Prx6 могло привести к резкому снижению антиоксидантной активности нового комплекса.
Другой неочевидной характеристикой выбранной структуры бифункционального белка PSH является его стабильность и сохранение как пероксидазной, так и супероксиддисмутазной активности, например, при работе в межклеточном пространстве.
Следующей неочевидной характеристикой бифункционального комплекса PSH является его высокая эффективность в качестве компонента, обеспечивающего реперфузии органов млекопитающих.
Создание рекомбинантного химерного бифункционального белка-антиоксиданта проводится генно-инженерными методами путем комбинации генов пероксиредоксина и супероксиддисмутазы. В этом случае связь между белками осуществляется строго контролируемым способом в области нерегулярных участков и практически не влияет на пространственную структуру составляющих структур, что позволяет сохранить ферментативные активности для каждого составляющего фермента.
Для получения генно-инженерной конструкции для экспрессии бифункционального белка PSH выбрана комбинация следующих ферментов-антиоксидантов: человеческий пероксиредоксин Prx6 и марганцевая супероксиддисмутаза (MnSOD) из E.coli.
В общем случае способ получения генно-модифицированного бифункционального белка PSH состоит из следующих стадий:
А) выделяют ДНК, кодирующую пероксиредоксин Prx6,
Б) выделяют ДНК, кодирующую супероксиддисмутазу (MnSOD) из E.coli,
В) выбирают тип плазмиды и ее генетическую конструкцию,
Д) формируют плазмиду,
Е) выбирают клетки и проводят трансформацию клеток полученной плазмидой,
Ж) выращивают клетки на поверхности агаризированной среды и обеспечивают последующий рост клеток в жидкой среде,
З) выделяют генно-модифицированный бифункциональный белок PSH из разрушенных клеток и проводят последующую очистку белка PSH.
На первом этапе получения конструкции составного бифункционального белка PSH выделяют ДНК, кодирующую пероксиредоксип Prx6, и ДНК, кодирующую супероксиддисмутазу (MnSOD) из E.coli.
Ген пероксиредоксина Prx6 получен как ПЦР продукт, используя в качестве матрицы плазмиду, содержащую кДНК гена пероксиредоксина Prx6 [патент США №8003345, Fesenko, et al. Antioxidant pharmaceutical compound, method for producing polypeptide and method of cure, опубл. 23.08. 2011], а также праймер Peri (SEQ ID NO:2)
5'-TTTCATATG CCCGGAGGTCTGCTT;
и праймер Per Sod 2 (SEQ ID NO:3)
5'-CAGGGATGGCAGGGTATAGCTAGGCTGGGTGTGTAGCGG.
Праймер peri (SEQ ID NO:2) содержит в начале последовательность сайта узнавания рестриктазы Nde I (выделена подчеркнутым фрагментом) и начало гена пероксиредоксина Prx6. Таким образом, праймер per1 комплементарен 5' области гена пероксиредоксина 6 человека.
Праймер perSod 2 состоит из двух последовательностей - конец гена пероксиредоксина (выделена подчеркнутым фрагментом) и начало гена супероксиддисмутазы (MnSOD) E.coli.
Вторая часть составного гена была получена как ПЦР продукт с использованием в качестве матрицы ДНК E.coli С и праймера perSod I (SEQ ID NO:4)
5' CCGCTACACACCCCAGCCT AGCTATACCCTGCCATCCCTG
и праймера sod 2 (SEQ ID NO:5)
5' GCGGCACGTTTTGCGGCGAAAAAACTCGAGTTT.
Праймер sod 2 комплементарен 3' области гена марганцевой супероксиддисмутазы E.coli. В праймерах конец гена MnSOD и сайт рестрикции Xho I выделены подчеркнутыми фрагментами.
Режим ПЦР: прогрев 94°С - 2 мин, затем 30 циклов - прогрев 94°С - 30 сек, отжиг праймеров - 58°С - 30 сек, синтез 68°С - 60 сек. В конце ПЦР - достройка 5 мин 68°С. Продукт ПЦР был подвергнут электрофорезу в 0,8% агарозе, и полоса, содержащая нужный фрагмент, была вырезана из геля. ДНК была выделена и очищена с помощью колонки QIAGEN (Германия) согласно инструкции фирмы-производителя.
Очищенные фрагменты были смешаны (по 5 нг в 50 мкл) и гибридизованы (отжиг) 2 мин 95°С затем 5 мин 52°С, и так 5 раз. Затем в реакционную смесь были внесены два праймера peri и sod 2 и была выполнена ПЦР. Режим ПЦР: прогрев 94°С - 2 мин, затем 30 циклов - прогрев 94°С - 30 сек, отжиг праймеров - 58°С - 30 сек, синтез 68°С - 60 сек. В конце ПЦР - достройка 5 мин 68°С. Продукт ПЦР был подвергнут электрофорезу в 0,8% агарозе, и полоса, содержащая нужный фрагмент, была вырезана из геля. ДНК была выделена и очищена с помощью колонки QIAGEN (Германия) согласно инструкции фирмы-производителя. Полученный ДНК фрагмент был обработан рестриктазами NdeI и XhoI и снова подвергнут электрофорезу в 0,8% агарозе. Полоса нужного фрагмента была вырезана и очищена на колонке с целью удаления ферментов рестрикции. Полученный таким образом фрагмент был лигирован в плазмиду рЕТ23, обработанную теми же рестриктазами. Таким путем была получена плазмида pET23(PSH), кодирующая составной белок PSH молекулярной массы 45 кД, аминокислотная последовательность которого приведена в SEQ ID NO:1.
Плазмида содержит: промотор РНК-полимеразы фага Т7, участок инициации репликации (ori) и терминатор транскрипции рибосомального оперона генетический маркер - Ар, детерминирующий устойчивость трансформированных плазмидой клеток E.coli к ампициллину, уникальные сайты узнавания рестрикционными эндонуклеазами, а также последовательность, кодирующую 6 гистидиновых остатков, непосредственно примыкающую к концу гена PSH.
Полученный и очищенный белок PSH применен при проведении реперфузии изолированного сердца. Данный вариант применения белка PSH включает, но не ограничивает других вариантов применения белка PSH.
Белок PSH растворен в растворе хлорида натрия, предпочтительно изотоническом (0.9% NaCl), белок PSH может также растворяться в известных консервирующих растворах, таких как растворы Кребса-Гензелейта, UW, Ст. Томас II, Коллинза, Станфордском и подобных им. Данный раствор может быть использован во время всех фаз трансплантации органов, особенно сердца, включая следующие (но не ограничиваясь ими): 1) изоляцию органа от донора; 2) консервацию органа (хранение и транспортировка в условиях гипотермии) и 3) имплантацию органа реципиенту (раствор для реперфузии).
Для приготовления раствора вначале готовят 10-кратные концентраты всех компонентов (маточный раствор). Для этого в градуированную бутыль емкостью 1 л наливают 700-800 мл бидистиллированной воды и растворяют последовательно (во избежание образования труднорастворимых осадков) следующие химически чистые соли (в граммах). В качестве одного из вариантов в таблице 1 приведен раствор для реперфузии изолированного сердца крысы. Концентрация всех типов белков в реперфузионной жидкости - 0,5 мг/мл.
Для предупреждения выпадения осадка раствор хлористого кальция готовят отдельно и медленно добавляют к раствору остальных солей после полного их растворения. Полученный основной раствор доводят дидистиллированной водой до 1 л, фильтруют через бумажный фильтр и стерилизуют пропусканием через фильтры 0,45 и 0,22 мк.
При реперфузии через аорту изолированного сердца крысы (с принудительной прокачкой раствора Хенкса) в контроле отмечается стабильное сердцебиение сердца с частотой 80-100 ударов/мин. При длительной реперфузии (более часа) частота сердцебиения незначительно уменьшается.
При добавлении в среду Хенкса перекиси водорода (искусственной окислительный стресс) наблюдается быстрое (в течение первых минут) существенное увеличение частоты сердцебиения (до 125-140 ударов/мин). Таким образом, внешне эффект экзогенного введения в кровеносное русло перекиси проявляется в учащении сердцебиения. Однако через 15-30 мин наблюдается прекращение сердцебиения.
При добавлении в перфузионную среду Хенкса рекомбинантного пероксиредоксина 6 наблюдается незначительное увеличение частоты сердцебиения (до 125 ударов/мин Наблюдается более качественное сокращение миокарда. Отмывка введенного пероксиредоксина 6 сохраняет частоту сокращения изолированного сердца при сохранении других параметров. Введение после отмывки пероксиредоксина 6 в раствор Хенкса перекиси водорода практически не влияло на частоту и качество.
При введении в среду Хенкса бифункционального белка PSH наблюдается уменьшение частоты сердцебиения (до 50 ударов/мин). Внешне наблюдается более качественное сокращение миокарда. Отмывка введенного химерного бифункционального белка PSH приводит к увеличению частоты сокращения изолированного сердца до 85-90 ударов/мин при сохранении других параметров. Введение перекиси водорода в раствор Хенкса после отмывки бифункционального белка PSH незначительно увеличивало частоту сердцебиения (до 100 ударов/мин), что соответствовало частоте сердцебиения в контрольных экспериментах.
Полный протокол полученных результатов приведен в таблице 2.
38 (конец)
130 (после H2O2)
90 (после белка)
55 (после H2O2)
76 (после белка)
55 (после H2O2)
Иммуногистохимические исследования показали, что в норме эндогенный пероксиредоксин Prx6 локализуется преимущественно в кровеносных сосудах сердца, (см. фиг.5), причем на внешней стороне сосуда, которая примыкает к соединительной ткани и непосредственно к клеткам миокарда. Действие перекиси водорода приводит к некоторому увеличению количества пероксиредоксинов в сосудах сердца, что, по-видимому, связано с увеличением экспрессии пероксиредоксииа в условиях окислительного стресса.
При введении экзогенных пероксиредоксинов Prx6 и белка PSH при реперфузии сердца через полчаса после начала реперфузии белки также обнаруживаются преимущественно в кровеносных сосудах сердца, причем на внешней стороне сосуда (см. фиг.6), которая примыкает к соединительной ткани и непосредственно к клеткам миокарда. Неочевидным фактом является то, что экзогенные пероксиредоксин Prx6 и белок PSH адсорбируются в тех же местах, в которых преимущественно локализован и эндогенный пероксиредоксин Prx6. Это приводит к существенному повышению антиоксидантного статуса сосудов и прилегающих к сосудам тканей сердца.
Ниже приведены примеры 1-7, которые иллюстрируют этапы формирования плазмиды, трансформации и выращивания клеток, а также очистку белка. Данные примеры включают, но не ограничивают специфическими условиями и параметрами объема изобретения.
Пример 1. Выбор типа плазмиды и ее генетической конструкции и формирование плазмиды
Полученный таким образом ПЦР фрагмент, включающий в себя гены пероксиредоксина 6 и марганцевой супероксиддисмутазы, клонировали в экспрессирующий вектор pET23b по точкам рестрикции NdeI и XhoI.
Проверку конструкций на наличие нуклеотидных замен проводили с помощью секвенирования в фирме Евроген (http://evrogen.ru).
Результаты секвенирования анализировали с помощью программы Chromas и Fasta (http://www.ebi.ac.uk/Tools/sss/fasta/nucleotide.html).
Проведенный анализ полученной конструкции показал, что действительно была получена конструкция, в которой гены пероксиредоксина 6 и супероксиддисмутазы расположены последовательно и не имеют нуклеотидных замен. Полученная конструкция была использована для трансформации экспрессирующего штамма E.coli BL21(PSH) и последующей наработки гибридного белка.
Пример 2. Клонирование генно-инженерных конструкций в бактериальный или дрожжевой геном
2.1. Лигирование
40 нг плазмидной ДНК были смешаны с 40 нг ПЦР-фрагмента в объеме 50 мкл. Добавлено 5 мкл лигазного буфера и 5 ед. лигазы. В контроле вместо фрагмента была просто вода. Лигирование происходило при 18°С в течение 9 часов.
2.2 Трансформация
Бактерии BL21(DE3) предварительно выращивают в течение ночи из единичной колонии. Для трансформации выращивают свежую культуру - ночную культуру разводят в свежей среде 1:100 и выращивают на качалке около 3 часов до появления заметной мутности. 8 мл свежевыращенных бактерий центрифугируют и осадок промывают 0.1 М хлористым кальцием. Окончательно клетки были суспендированы в 0.4 мл 0.1 М CaCl2. Операции проводят строго на холоде (4°С). На холоде смешивают лигазные реакции с 200 мкл бактерий и инкубируют на льду 40 мин, затем переносят в водяной термостат на 41°С на 60 сек, после чего охлаждают и добавляют 800 мкл среды LB и инкубируют на качалке при 37° в течение 1 часа. Делают высевы по 10 и 100 мкл на чашки Петри со средой LB и с добавлением 100 ед./мл ампициллина и 20 ед./мл хлорамфеникола. Чашки инкубируют в течение ночи при 37°С.
2.3 Отбор клонов
В результате было отобрано 3 клона, содержащих вставку нужного размера и экспрессирующийся составной белок. Для этих трех клонов было проведено секвенирование вставки с тем, чтобы убедиться что клонированный составной ген имеет правильную последовательность. Секвенирование было выполнено компанией ЕвроГен с использованием капиллярного секвенатора. В одном случае последовательность гена оказалась с ошибкой, в двух других получено точное совпадение с последовательностью, представленной в GenBank.
Один из этих клонов был выбран для дальнейшей работы. Его нуклеотидная последовательность длиной 1290 п.о. и аминокислотная последовательность длиной 435 аминокислот в экспрессируемом белке PSH приведена в перечне последовательностей SEQ ID NO:1. Молекулярный вес PSH составляет около 47 кДа (Prx6 - около 25 кДа, Mn-SOD - около 22 кДа). Место стыковки гена пероксиредоксина (кончается на GCCT) и гена СОД (начинается с ATGAGC) соответствует позициям 672-673 п.о в перечне последовательностей SEQ ID NO:1.
Пример 3. Проверка эффективности трансформации и генетической стабильности продуцента
Для проверки генетической стабильности ночную культуру клона высевают на агар с антибиотиками и без них и подсчитывают число колоний, выросших на чашках при высеве из одного и того же разведения. Одну из колоний, выросших на агаре с антибиотиками, снова засевают в питательную среду и опять подсчитывают число колоний, выросших на агаре с антибиотиками и без.
Для этого 100 мкл суспензии бактерий из штока, выращенного из отдельной колонии и хранящегося при -75°, были засеяны в 50 мл среды LB. Инкубация продолжалась около 16 часов (ночь) на качалке при 37°. Из полученной суспензии делают контрольные высевы на чашки с антибиотиками (ампициллин 100 ед./мл и хлорамфеникол 20 ед./мл) и остальной материал вносят в 1,8 л среды LB с добавлением тех же антибиотиков в тех же концентрациях. В контрольных высевах проверяют наличие плазмид: одинаковое число колоний на контрольных и чашках с антибиотиками указывает на стабильность штамма и наличие плазмид.
Всего было сделано и проанализировано 8 таких пассажей. Результаты представлены в таблице 3.
Как видно из таблицы 3, на протяжении всех пассажей не обнаружено потери плазмиды, кодирующей устойчивость к амипициллину.
Культуру последнего пассажа проверили на индукцию синтеза белка PSH, и подтверждена нормальная индукция экспрессии белка PSH. Таким образом, штамм Е.coli на протяжении многих генераций сохраняет клонированный ген.
Для проверки сохранности клонированного гена при среднесрочном хранении продуцента свежевыращенная ночная культура PSH выдерживалась в холодильнике при 4°С. Периодически делают высевы на агар с ампициллином (100 ед./мл) и хлорамфениколом (20 ед./мл), а также на агар без антибиотиков из 10(-7) разведения. На следующий день подсчитывают число колоний.
Выращивание штамма производят в ферментере на 2 л при 37° при интенсивной аэрации до достижения мутности около 0,4. Затем вносят ИПТГ до конечной концентрации 1 мМ и продолжают инкубацию еще 2 часа.
Бактерии собирают центрифугированием (4000 об/мин, 20 мин), полученный осадок замораживают при -20°С.
Пример 4. Выделение и очистка белка SPH
Присутствие His-tag в структуре белка PSH упрощает процедуру очистки рекомбинантного белка благодаря тому, что этот домен специфически связывается с Ni-NTA-агарозой.
Осадок биомассы бактерий E.coli ресуспендируют в 25 мл буфера для нанесения на колонку 12 мМ (Трис-HCl, рН 7.8, 10 мМ имидазола) и разрушают ультразвуком при 4°С. Клеточный дебрис удаляют центрифугированием при 14000 об/мин в течение 10 мин, надосадок фильтруют через 0,45 мкм фильтр (Corning, США) и наносят на колонку (BioRad, США) объемом 10 мл с Ni-NTA-агарозой (Invitrogen, США), которая предварительно была уравновешена буфером для нанесения. Инкубацию бактериального лизата с Ni-NTA-агарозой проводят в течение 40 мин при +4°С. Затем, колонку промывают 100 мл раствора: 12 мМ Трис-HCl рН 7.8, 20 мМ имидазола. Элюцию белка проводят 5 мл буфера: 12 мМ Трис-HCl, рН 7.8, 250 мМ имидазола. Белок концентрируют с помощью мембранного концентратора VIVASCIENCE 30.000 MWCO (Sartorius, Германия) и в нем же диализуют против 1×PBS (1.7 мМ KH2PO4, 5.2 мМ Na2HPO4, 150 мМ NaCl, рН 7.4). Чистота белка, оцениваемая по электрофорезу в полиакриламидном геле, составляла около 95%. Обычно получают около 20-40 мг очищенного белка с одной ферментации. Электрофорез белков проводят в денатурирующих условиях (в присутствии SDS) по стандартной методике Laemmli [Laemmli U.K. 1970 Nature, 227, 680-685] на оборудовании Mini Vertical Unit («Amersham», США). В работе используют 5% концентрирующий и 12,5% разрешающий полиакриламидные гели (ПААГ) на Трис-буфере (130 мМ Трис-HCl, рН 6.8, для концентрирующего и 375 мМ Трис-HCl, рН 8.8, для разрешающего), содержащие 0.1% SDS, 0.1% PSA и 0.08% TEMED. Электродным буфером служит раствор: 25 мМ Триса, 250 мМ глицина и 0,1% SDS. Электрофорез в концентрирующем геле проводят при силе тока 15 мА (10-15 мин), затем в разрешающем геле при 25 мА (40-45 мин). Окрашивание геля проводят с помощью Comassie Brilliant Blue R-250 («Fluka», США). На фиг.2 приведены данные электрофореза гибридного белка PSH. Где А - наработка белка в E.coli BL21(DE3). Б - очистка белка на Ni-NTA агарозе.
Пример 5. Определение ферментативных активностей белка SPH
5.1. Пероксидазная активность белка SPH в отношении пероксида водорода и гидропероксида трет-бутила
Пероксидазную активность определяют согласно [Kang S.W., Baines I.С., Rhee S.G. (1998). Characterization of a mammalian peroxiredoxin that contains one conserved cysteine. // J. Biol. Chem. Vol.273, pp.6303-6311] с небольшими модификациями, используя в качестве субстратов гидропероксид водорода (неорганический гидропероксид) и трет-бутил гидропероксид (органический гидропероксид). В качестве отрицательного контроля используют пероксиредоксин 6 человека с заменой остатка цистеина в активном центре на серии (Cys47Ser). Реакционная смесь (150 мкл) содержит 7 мМ фосфатный буфер, рН 7.3, 150 мМ NaCl, 2 мМ ДТТ, 100 мкМ пероксид водорода или 100 мкМ гидропероксид трет-бутила (ГПТБ) и фермент в различных концентрациях. Реакцию проводят в течение 10 мин при 37°С и останавливают, добавляя 50 мкл 0.6 М HCl. Затем добавляют 100 мкл 10 мМ Fe(NH4)2(SO4)2 и 50 мкл 2.5 М KSCN, что приводит к образованию комплексного соединения железа красного цвета. Концентрация пероксида пропорциональна интенсивности окраски, которую измеряют при длине волны 492 нм на приборе Multiscan. Пример такого определения показан на фиг.3 Пероксидазная активность PSH в отношении перекиси водорода составляет 100 нмоль/мин/мг, что примерно вдвое ниже, чем для чистого пероксиредоксина.
5.2. Супероксиддисмутазная активность
Супероксиддисмутазную активность определяют с помощью коммерческого набора Superoxide Dismutase Assay Kit (Calbiochem, США), в соответствии с инструкцией производителя.
Супероксиддисмутазная активность PSH составила около 9 ед./мг белка, см. фиг.4, что практически соответствует ожидаемой величине около 17 ед./мг белка Mn-SOD E.coli [Roberts В. and Hirst R., 1996], где одна ед. соответствует количеству белка, необходимому для проявления 50% дисмутации супероксидрадикалов.
Принимая во внимание, что молекулярный вес PSH около 47 кДа (Prx6 около 25 кДа, Mn-SOD около 22 кДа), в 1 мг PSH примерно в 2 раза меньше молекул Prx6 и Mn-SOD, чем в 1 мг белка Prx6 или Mn-SOD, следовательно, при расчете активности на 1 мг PSH ожидается двукратное снижение активностей. Таким образом, можно заключить, что как пероксидазная, так и супероксиддисмутазная активность сохранилась полностью в составе белка PSH.
Пример 6. Условия культивирования при получении препаративных количеств рекомбинантного белка PSH
При разработке методов культивирования E.coli с экспрессированным белком PSH основное внимание было уделено получению условий, при которых происходит максимальная экспрессия белка SPH при минимальной экспрессии собственных белков E.coli.
Штаммы E.coli:_BL21(DE3)[pRIL] Codon Plus. Генотип: В F-dcm ompT hsdS (rB- mB-) gal λ (DE3) [pRIL CamR] («Stratagen», США). Этот штамм является производной BL21, содержащей плазмиду pRIL, которая определяет устойчивость к хлорамфениколу и содержит 3 гена тРНК, которые узнают кодоны аргинина (R), изолейцина (I) и лейцина (L), характерные для эукариот. Штамм лизогенен по λDE3, который содержит ген Т7 РНК полимеразы, под контролем lacUV5 промотора (индукторы - IPTG, галактоза; репрессор - глюкоза). Эффективность трансформации компетентных клеток (полученных Са2+ методом) до 1×107 трансформантов на 1 мкг плазмидной ДНК pUC19. Этот штамм обычно используют для улучшения синтеза белков эукариот в клетках E.coli. Выращивание в жидкой среде LB+хлорамфеникол (35 µg/ml), на твердой питательной среде LB+агар+хлорамфеникол (35 µg/ml). Для трансформации бактерии BL21(DE3) предварительно выращивают в течение ночи из единичной колонии. Для трансформации выращивают свежую культуру - ночную культуру разводят в свежей среде 1:100 и выращивают на качалке около 3 часов до появления заметной мутности. 8 мл свежевыращенных бактерий центрифугируют и осадок промывают 0.1 М хлористым кальцием. Окончательно клетки были суспендированы в 0.4 мл 0.1 М CaCl2. Операции проводят строго на холоде (4°С). На холоде смешивают лигазные реакции с 200 мкл бактерий и инкубируют на льду 40 мин, затем переносят в водяной термостат на 41°С на 60 сек, после чего охлаждают, добавляют 800 мкл среды LB и инкубируют на качалке при 37° в течение 1 часа. После скрининга трансформантов определяют экспрессию гена PSH. Для этого каждый клон засевают в пробирку Falkone на 50 мл в объеме 10 мл с добавлением антибиотиков и выращивают на качалке до оптической плотности 0,5, после чего вносят ИПТГ до концентрации 1 мМ и продолжают выращивание еще 3 часа. После этого отбирают по 1 мл, центрифугируют при 10000 об/мин в течение 5 мин. Осадок ресуспендируют в 50 мкл буфера и проводят электрофорез. В присутствии ИПТГ происходит индукция экспрессии гена под промотором Т7, и в бактериях нарабатывается белок PSH. Как правило, выход белка достигает около 30% от суммарного растворимого белка, что легко обнаруживается на электрофорезе. В результате было отобрано 3 клона, содержащих вставку нужного размера, и экспрессирующийся ген. Для этих трех клонов было проведено секвенирование гена с тем, чтобы убедиться, что клонированный ген имеет полигистидин и правильную последовательность. Секвенирование было выполнено компанией ЕвроГен с использованием капиллярного секвенатора. В одном случае последовательность гена оказалась с ошибкой, в двух других получено точное совпадение с последовательностью, представленной в GenBank. Один из этих клонов был выбран для дальнейшей работы. В дальнейшем этот клон назван E.coli BL21(PSH).
Пример 7. Применение бифункционального белка PSH при реперфузии сердца
Зажим, перекрывающий поступление перфузионного раствора в канюлю, открывают полностью, чем увеличивают приток перфузирующего раствора в канюлю до 12-15 мл/мин. Затем отрезают легкие. В результате перфузионный и кардиоплегический раствор из аорты поступает в коронарные артерии, питает сердце и выходит через вены сердца в правое предсердие. При сокращении правого предсердия и правого желудочка раствор вытекает через легочный ствол, верхнюю и нижнюю полые вены, омывая сердце.
Введение раствора перекиси водорода в концентрации 200 мкм в кровяное русло резко изменяет ритм сердцебиения. Резко возрастает пульс (в два раза по сравнению с контролем) после введения перекиси. При этом эффект развивается быстро - в течение первой минуты. Дальнейшее проведение реперфузии приводит к прекращению сердцебиения через 30 минут. Таким образом, имеются объективные параметры, позволяющие контролировать состояние сердечной мышцы.
Введение в реперфузионный раствор бифункционального белка PSH в концентрации 0,1-0,5 мг/мл, а также последующая реперфузия и удаление не связавшегося белка из перфузионной среды существенно улучшает состояние сердечной мышцы. При последующем введении в реперфузионный раствор перекиси не наблюдается ее влияния на деятельность сердца. Особо следует отметить, что в данных экспериментах белок PSH не присутствовал в кровеносном русле, так как перед добавлением перекиси водорода в реперфузионный раствор свободный белок PSH был полностью удален отмывкой из сердца. Таким образом, эффект белка PSH обусловлен количеством белка PSH, который адсорбировался в сердце в процессе реперфузии.
Прямые измерения содержания всех исследованных пероксиредокинов, адсорбированных в тканях сердца в процессе реперфузии, показали, что в сердце адсорбируется относительно малое количество пероксиредоксинов. Существенным моментом является тот факт, что наиболее эффективно пероксиредоксины при реперфузии сорбируются в предсердии, а не в желудочке сердца. По-видимому, этим и объясняется сильный и быстрый эффект защиты сердца от действия перекиси водорода (см. ниже).
Как уже отмечалось выше, при реперфузии происходит сорбция белка PSH в разных отделах сердца. В этом случае можно надеяться на существенное повышение антиоксидантного статуса в тканях сердца. В контроле при введении в перфузионный раствор перекиси водорода наблюдается резкое увеличение перекисного окисления липидов в тканях сердца (5-8 раз). Предварительная реперфузия белка PSH приводит к полному блокированию дополнительного перекисного окисления липидов, вызванного перекисью водорода. Таким образом, действительно рекомбинантный белок PSH обеспечивает пролонгированный эффект повышения антиоксидантного статуса органа, по крайней мере, в случае сердца.
Промышленная применимость
Белок PSH, экспрессированный в E.coli, обладает как пероксидазной, так и супероксиддисмутазпой активностями, аналогичными молярными активностями человеческого пероксиредоксина 6 и дрожжевой супероксиддисмутазы (100 нмоль/мин/мг по трет-бутил гидропероксиду и ~9 ед./мг по супероксидрадикалу).
Проверка эффективности трансформации Е.coli показала, что при индукции экспрессии в E.coli рекомбинантного белка PSH с использованием ИПТГ данный белок является мажорным и составляет примерно 30% от всех водорастворимых белков.
Экспрессированный белок PSH является водорастворимым (а не содержится в тельцах включения E.coli), что позволяет существенно упростить процедуру его выделения с использованием аффинной хроматографии.
Таким образом, получен продуцент химерного бифункционального белка PSH с совмещенными пероксидазной и супероксиддисмутазной активностями на основе E.coli. и сам высокоочищенный белок PSH.
Полученный белок PSH способствует эффективному восстановлению сердечной деятельности после длительной ишемии, снижению метаболических расстройств и контрактурных повреждений миокарда.
| название | год | авторы | номер документа |
|---|---|---|---|
| Средство, его применение и способ повышения устойчивости организма млекопитающих к ишемически-реперфузионному поражению тонкого кишечника | 2018 |
|
RU2729788C2 |
| СПОСОБ ПРИМЕНЕНИЯ СРЕДСТВА НА ОСНОВЕ МОДИФИЦИРОВАННОГО ПЕРОКСИРЕДОКСИНА 2 ЧЕЛОВЕКА ДЛЯ КОРРЕКЦИИ ПОСЛЕДСТВИЙ ИШЕМИЧЕСКИ-РЕПЕРФУЗИОННОГО ПОРАЖЕНИЯ ПОЧЕК | 2020 |
|
RU2747121C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, РЕКОМБИНАНТНЫЙ ШТАММ, РЕКОМБИНАНТНЫЙ БЕЛОК ВМР-7 И СПОСОБ ВЫДЕЛЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА В ДИМЕРНОЙ ФОРМЕ | 2012 |
|
RU2499047C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, РЕКОМБИНАНТНЫЙ ШТАММ, РЕКОМБИНАНТНЫЙ БЕЛОК ВМР-2 И СПОСОБ ВЫДЕЛЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА В ДИМЕРНОЙ ФОРМЕ | 2012 |
|
RU2499048C1 |
| РЕКОМБИНАНТНЫЙ БЕЛОК Collbd-BMP-7, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCollbd-BMP-7, ШТАММ Escherichia coli-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА Collbd-BMP-7, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА Collbd-BMP-7 | 2009 |
|
RU2408730C1 |
| Иммуногенная композиция на основе рекомбинантных псевдоаденовирусных частиц, а также на основе белковых антигенов и способ получения иммуногенной композиции | 2018 |
|
RU2691302C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET23-a(+)PrxVIhumΔ178, КОДИРУЮЩАЯ N-КОНЦЕВОЙ ФРАГМЕНТ ПЕРОКСИРЕДОКСИНА VI ЧЕЛОВЕКА, И ШТАММ E.coli BL21/DE3/pET23-a(+)/PrxVIhumΔ178 - ПРОДУЦЕНТ N-КОНЦЕВОГО ФРАГМЕНТА ПЕРОКСИРЕДОКСИНА VI ЧЕЛОВЕКА | 2003 |
|
RU2250262C1 |
| РЕКОМБИНАНТНАЯ ДНК, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНЫЙ ГИБРИДНЫЙ БЕЛОК ОКСИДАЗЫ D-АМИНОКИСЛОТ С ХИТИНСВЯЗЫВАЮЩИМ ДОМЕНОМ (DAOcbd) РЕКОМБИНАНТНАЯ ПЛАЗМИДА pVR1, ОБЕСПЕЧИВАЮЩАЯ ЕГО СИНТЕЗ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli С 41(DE3)/pVR1 - ПРОДУЦЕНТ DAOcbd | 2006 |
|
RU2310688C1 |
| РЕКОМБИНАНТНЫЙ ХИМЕРНЫЙ БЕЛОК SUMO3-apoA-I ДЛЯ ПОЛУЧЕНИЯ ЗРЕЛОГО АПОЛИПОПРОТЕИНА A-I ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ХИМЕРНОГО БЕЛКА SUMO3-apoA-I И СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО АПОЛИПОПРОТЕИНА A-I ЧЕЛОВЕКА | 2015 |
|
RU2605326C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET40CmAP/MBL-T, КОДИРУЮЩАЯ ГИБРИДНЫЙ БИФУНКЦИОНАЛЬНЫЙ ПОЛИПЕПТИД CmAP/MBL-T СО СВОЙСТВАМИ ВЫСОКОАКТИВНОЙ ЩЕЛОЧНОЙ ФОСФАТАЗЫ CMAP И МАННАН-СВЯЗЫВАЮЩЕГО ЛЕКТИНА С-ТИПА MBL-T, РЕКОМБИНАНТНЫЙ ШТАММ E.coli Rosetta(DE3)/pET40CmAP/MBL-T - ПРОДУЦЕНТ ГИБРИДНОГО БИФУНКЦИОНАЛЬНОГО ПОЛИПЕПТИДА CmAP/MBL-T И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2538169C1 |



Изобретение относится к биохимии и представляет собой новый бифункциональный белок PSH, включающий человеческий пероксиредоксин Prx6 и марганцевую супероксиддисмутазу MnSOD, обладающий антиоксидантной активностью супероксиддисмутазы и пероксидазы. Описана также химерная нуклеиновая кислота, кодирующая предложенный белок. Раскрыт способ получения предложенного белка путем культивирования клеток штамма E.coli BL21(PSH), трансформированных сконструированным рекомбинантным вектором экспрессии на основе плазмиды pET22b(+). Изобретение позволяет получить высокой выход белка PSH, обладающего высокой антиоксидантной активностью. 3 н. и 1 з.п. ф-лы, 6 ил., 3 табл., 7 пр.
1. Химерная нуклеиновая кислота, кодирующая гибридный белок PSH, включающий человеческий пероксиредоксин Prx 6 и марганцевую супероксиддисмутазу MnSOD, обладающий антиоксидантной активностью супероксиддисмутазы и пероксидазы, первичная структура которого описывается аминокислотной последовательностью SEQ ID NO:1.
2. Химерная нуклеиновая кислота по п.1, которая получена клонированием отдельных фрагментов генов Prx 6 и MnSOD в реакции ПЦР с применением синтетических олигонуклеотидов SEQ ID NO:2 - SEQ ID NO:5.
3. Рекомбинантный вектор экспрессии pET23(PSH), содержащий химерную нуклеиновую кислоту, кодирующую белок PSH с аминокислотной последовательностью SEQ ID NO:1, оперативно встроенную в плазмиду pET22b(+) по сайтам рестрикции Nde I и Xho I с С-концевой локализацией полигистидинов.
4. Гибридный белок PSH для реперфузии сердца, первичная структура которого описывается аминокислотной последовательностью SEQ ID NO:1.
| MAKSIMENKO AV et.al | |||
| The combination of modified antioxidant enzymes for anti-thrombotic protection of the vascular wall: the significance of covalent connection of superoxide dismutase and catalase activities, J Pharm Pharmacol | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET23-a(+)PrxVIhumΔ178, КОДИРУЮЩАЯ N-КОНЦЕВОЙ ФРАГМЕНТ ПЕРОКСИРЕДОКСИНА VI ЧЕЛОВЕКА, И ШТАММ E.coli BL21/DE3/pET23-a(+)/PrxVIhumΔ178 - ПРОДУЦЕНТ N-КОНЦЕВОГО ФРАГМЕНТА ПЕРОКСИРЕДОКСИНА VI ЧЕЛОВЕКА | 2003 |
|
RU2250262C1 |
| КОМПОЗИЦИЯ С АНТИОКСИДАНТНЫМИ СВОЙСТВАМИ И СПОСОБ ЛЕЧЕНИЯ БОЛЕЗНЕЙ МЛЕКОПИТАЮЩИХ | 2002 |
|
RU2280448C2 |
| US 0007306944 B2, 11.12.2007 | |||

