Изобретение относится к микробиологической и медицинской промышленности, генной инженерии, биотехнологии и может быть использовано для получения зрелого аполипопротеина Α-I человека.
Аполипопротеин Α-I (апоА-I) человека является основным белком плазмы крови, участвующим в образовании липопротеинов высокой плотности (ЛПВП). ΑпοΑ-I состоит из одной полипептидной цепи, содержащей 243 аминокислотных остатков, и имеющей молекулярный вес 28,3 кДа (1, 2). ΑпοΑ-I является главным структурным белком частиц ЛПВП и многофункциональным обменным протеином, имеет широкий спектр физиологических функций, участвует в регуляции метаболизма холестерина и регулирует уровень липидов в крови, обладает противовоспалительными, антиокислительными и противоопухолевыми свойствами (3, 4). ΑпοΑ-I может быть использован в терапевтических целях, например, для терапии атеросклероза (5), а также может служить естественным переносчиком лекарств, не взаимодействующим с иммунной и ретикулоэндотелиальной системой (6-9).
ΑпοΑ-I получают из сыворотки крови человека, однако этот способ получения имеет низкий выход, высокую стоимость из-за ограниченных объемов донорской крови, опасности заражения крови и других проблем, что не позволяет достичь промышленных объемов производства, существенно ограничивает применение нативного апоА-I в медицине. Создание штамма продуцента рекомбинантного апоА-I человека методами генной инженерии является лучшим средством для крупномасштабного производства этого белка.
Известно получение рекомбинантного химерного белка апоА-I человека, дополнительно включающего с N-конца аминокислотную последовательность Met-Arg-Gly-Ser-(His)6-Met), в клетках Е. coli (10). Выход целевого белка, обладающего сходными с нативным белком апоА-I человека физико-химическими свойствами, оказался крайне низким и составил 10 мг на литр культуральной среды. Другим недостатком аналога является то, что при очистке апоА-I человека, как и других рекомбинантных терапевтических белков, получаемых в клетках E.coli, остается загрязнение эндотоксинами (липополисахаридами). Известно, что даже небольшие остаточные количества примеси липополисахаридов могут вызвать побочные эффекты в организме хозяина, такие как лихорадка, повреждение тканей и шок (11, 12).
Известно получение рекомбинантного белка апоА-I человека, в клетках штамма продуцента, сконструированного на основе штамма Е. coli Rozetta 2(DE3) и экспрессирующего прокариотического вектора pJExpress401, в составе которого клонирован синтетический ген АроА1_1946/15 с С-концевым участком, кодирующим шестигистидиновую аминокислотную последовательность (13). Достигается удовлетворительная очистка целевого белка с помощью двойной аффинной хроматографии, после которой выход целевого белка составляет около 50 мг/л культуры. Рекомбинантный апоА-I человека по антигенным свойствам и молекулярной массе, с учетом размера С-концевого шестигистидинового тракта, соответствует зрелой форме апоА-I человека. Недостатком известного аналога является низкий выход целевого белка.
Система экспрессии чужеродных генов в дрожжах Pichia pastoris, в отличие от Е. coli, лишена проблемы очистки рекомбинантных белков от липополисахаридов и позволяет получать чужеродные белки, секретируемые в культуральную среду, с большим выходом, чем в Е. coli (14).
Известно использование дрожжей Pichia pastoris для получения рекомбинантного апоА-I человека. Дрожжевой штамм продуцент получен на основе Pichia pastoris штамма Х-33, трансформированного плазмидой pPICZαA:ApoA1_1946/21, содержащей ген зрелого белка апоА-I человека (13). Однако при индукции метанолом экспрессии клонированного гена апоА-I наблюдается выход в культуральную среду как зрелого, так и непроцессированного апоА-I, сопровождаемый лизисом дрожжевых клеток, обусловленным, по-видимому, мембранотропностью рекомбинантного белка апоА-I.
Наиболее близким к заявленному является получение рекомбинантного зрелого белка апоА-I человека с использованием штамма дрожжей Pichia pastoris GS115, полученного путем трансформации вектором рР1С9К, содержащим ген зрелого белка апоА-I человека. После индукции метанолом в ферментере выход секретируемого рекомбинантного белка апоА-I человека был относительно невысоким для системы экспрессии Pichia pastoris и составил 90 мг на литр, что является недостаточным для промышленного производства. Аминокислотная последовательность и молекулярный вес рекомбинантного зрелого белка апоА-I человека были идентичны нативному (15).
Задачей заявляемой группы изобретений является увеличение выхода зрелого белка апоА-I человека, полученного с использованием штамма Pichia pastoris.
Решение поставленной задачи достигается тем, что для получения зрелого белка апоА-I человека используют рекомбинантный химерный белок SUMO3-apoA-I, 343 а.о. с молекулярной массой 39,6 кДа, включающий с N-конца аминокислотную последовательность убиквитин-подобного белка SUMO3 человека (SUMO3) размером 100 а.о. с молекулярной массой 11,5 кДа, слитую с С-конца с аминокислотной последовательностью зрелого белка апоА-I человека размером 243 а.о. с молекулярной массой 28 кДа; для продукции рекомбинантного химерного белка SUMO3-apoA-I используют нативный штамм дрожжей Pichia Pastoris Х-33, полученный путем трансформации линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I, обеспечивающей синтез рекомбинантного химерного белка SUMO3-apoA-I; зрелый белок апоА-I человека получают путем ферментативного гидролиза рекомбинантного химерного белка SUMO3-apoAI с использованием SUMO-специфичной протеазы SP2.
Раскрытие изобретения
Для получения зрелого белка аполипопротеин Α-I человека используют рекомбинантный химерный белок SUMO3-apoA-I (аминокислотная последовательность SEQ ID NO 1) размером 343 а.о. с молекулярной массой 39,6 кДа, включающий с N-конца аминокислотную последовательность убиквитин-подобного белка SUMO3 человека (SUMO3) размером 100 а.о. с молекулярной массой 11,5 кДа, (аминокислотная последовательность SEQ ID NO 2), слитую с С-конца с аминокислотной последовательностью зрелого белка апоА-I человека размером 243 а.о. с молекулярной массой 28 кДа (аминокислотная последовательность SEQ ID NO 3). Данный химерный белок кодируется нуклеотидной последовательностью SEQ ID NO 1 размером 1029 п. н., включающей с 5′ конца нуклеотидную последовательность гена убиквитин-подобного белка SUMO3 человека (нуклеотидная последовательность SEQ ID NO 2), слитую с 3′-конца с нуклеотидной последовательностью гена зрелого белка аполипопротеин A-I человека (нуклеотидная последовательность SEQ ID NO 3).
Штамм дрожжей Pichia pastoris - продуцент рекомбинантного химерного белка SUMO3-apoA-I получен в результате трансформации нативного штамма дрожжей Pichia pastoris Х-33 линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I, обеспечивающей синтез рекомбинантного химерного белка SUMO3-apoA-I.
Способ получения зрелого аполипопротеина Α-I человека включает микробиологический синтез рекомбинантного химерного белка SUMO3-apoA-I в процессе культивирования рекомбинантного штамма дрожжей Pichia pastoris, полученного указанным выше путем, осаждение дрожжевых клеток, отделение супернатанта, осаждение из супернатанта рекомбинантного химерного белка SUMO3-apoA-I, его ресуспендирование и диализ, последующий ферментативный гидролиз SUMO-специфичной протеазой SP2. Эта протеаза распознает аминокислотную последовательность SUMO в N-концевой области химерного белка и осуществляет высокоспецифичный гидролиз пептидной связи между С-концевым аминокислотным остатком SUMO и N-концевым аминокислотным остатком целевого белка в составе химера (16, 17).
Линеаризованную плазмидную ДНК pPsecSUMO3-apoA-I получают в результате обработки рекомбинантной плазмиды pPsecSUMO3-apoA-I (фиг. 1) рестриктазой BstXI, которая осуществляет гидролиз указанной плазмиды в области промотора гена АOХ1.
Для конструирования рекомбинантной плазмиды pPsecSUMO3-apoA-I сначала создают рекомбинантную плазмиду pJ201-apoA-I путем клонирования в коммерческой плазмиде pJ201 синтетического гена зрелого белка апоА-I с составом кодонов, оптимизированным для увеличения экспрессии в клетках P. pastoris. Оптимизацию кодонов осуществляют как с использованием компьютерной программы «Gene designer» (фирма "DNA 2.0", США), так и с помощью введения синонимических замен в часто встречаемые в геноме P. pastoris пары кодонов (18), тормозящих в соответствующих участках трансляцию мРНК. Из известных кодонов, по которым целесообразно проводить оптимизацию, выбрана такая их комбинация, которая в последующем обеспечила высокий уровень экспрессии рекомбинантного химерного белка SUMO3-apoA-I в заявленном штамме дрожжей P. pastoris. Полученный технический результат с очевидностью не вытекает из уровня техники, т.к. выбор кодонов для их оптимизации в целевых генах не всегда приводит к возрастанию эффективности экспрессии данных генов. В результате оптимизации кодонов иногда наблюдается снижение стабильности мРНК и, как следствие этого, снижение количества синтезируемого целевого белка (19).
Рекомбинантную плазмиду pPsecSUMO3-apoA-I получают путем сшивки (лигирования) фрагментов обработанных рестриктазами BsmBI и XbaI рекомбинантных плазмид pJ201-apoA-I и pPsecSUMO3 с помощью ДНК-лигазы фага Т4 (примеры 1-2).
Штамм дрожжей Pichia pastoris - продуцент рекомбинантного химерного белка SUMO3-apoA-I получают путем трансформации компетентных клеток нативного штамма дрожжей Pichia pastoris Х-33 линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I методом электропорации (пример 3).
Для ферментативного гидролиза рекомбинантного химерного белка SUMO3-apoA-I используют коммерческую SUMO-специфичную протеазу SP2 (SUMQstar Protease, Lifesensors, USA) (20) либо SUMO-специфичную протеазу SP2, полученную генно-инженерными методами, например, как описано в примерах 4-6.
Биосинтез рекомбинантного химерного белка SUMO3-apoA-I человека в клетках заявляемого штамма дрожжей Pichia pastoris осуществляют путем индукции культивируемых клеток метанолом (пример 7).
Предложенная группа технических решений позволяет значительно увеличить выход зрелого аполипопротеина Α-I человека до 400 мг/л в сравнении с прототипом и другими аналогами. При этом рекомбинантный зрелый аполипопротеин Α-I человека по молекулярной массе и выведенной аминокислотной последовательности соответствует нативному аполипопротеину Α-I человека, что важно для использования его, как в качестве средства адресной доставки лекарственных средств и биологически активных соединений в клетки органов и тканей человека, так и для получения на его основе лекарственного препарата для лечения атеросклероза.
Достижение заявленного технического результата обусловлено следующим:
1. Получение рекомбинантного штамма дрожжей P. pastoris путем трансформации линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I нативного штамма дрожжей Р. pastoris Х-33.
2. Использование в составе линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I синтетического гена апоА-I человека, полученного с учетом оптимизации выбранных кодонов (199 синонимичных нуклеотидных замен) в гене зрелого белка апоА-I человека, а также дополнительное введение в проектируемый синтетический ген ароА-1 синонимических нуклеотидных замен в трех парах выбранных кодонов, которые часто встречаются в геноме P. pastoris и значительно тормозят в соответствующих участках трансляцию мРНК, что в совокупности способствовало увеличению экспрессии гена.
3. Использование в конструкции гена рекомбинантного химерного белка SUMO3-ароА-1 нуклеотидной последовательности гена, кодирующего убиквитин-подобный белок SUMO3 человека, что способствовало не только увеличению экспрессии гена, секреции и фолдингу рекомбинантного химерного белка SUMO3-apoA-I, увеличению его синтеза в рекомбинантном штамме дрожжей P. pastoris (16), но привело к неизвестному ранее результату: в процессе секреции рекомбинантного химерного белка SUMO3-apoA-I не наблюдается лизиса клеток дрожжей P. pastoris, который происходит в случае прямой экспрессии гена зрелого белка ароА-1 человека в клетках P.pastoris. Это обусловлено, по-видимому, особенностью формирования третичной структуры рекомбинантного химерного белка SUMO3-apoA-I, препятствующей проявлению мембранно-литической активности белка ароА-I. Все это позволило значительно увеличить продукцию рекомбинантного химерного белка SUMO3-apoA-I в заявленном штамме дрожжей Р. pastoris и, соответственно, повысить выход зрелого аполипопротеина A-I человека в заявленном способе его получения.
Перечень фигур, чертежей и иных материалов
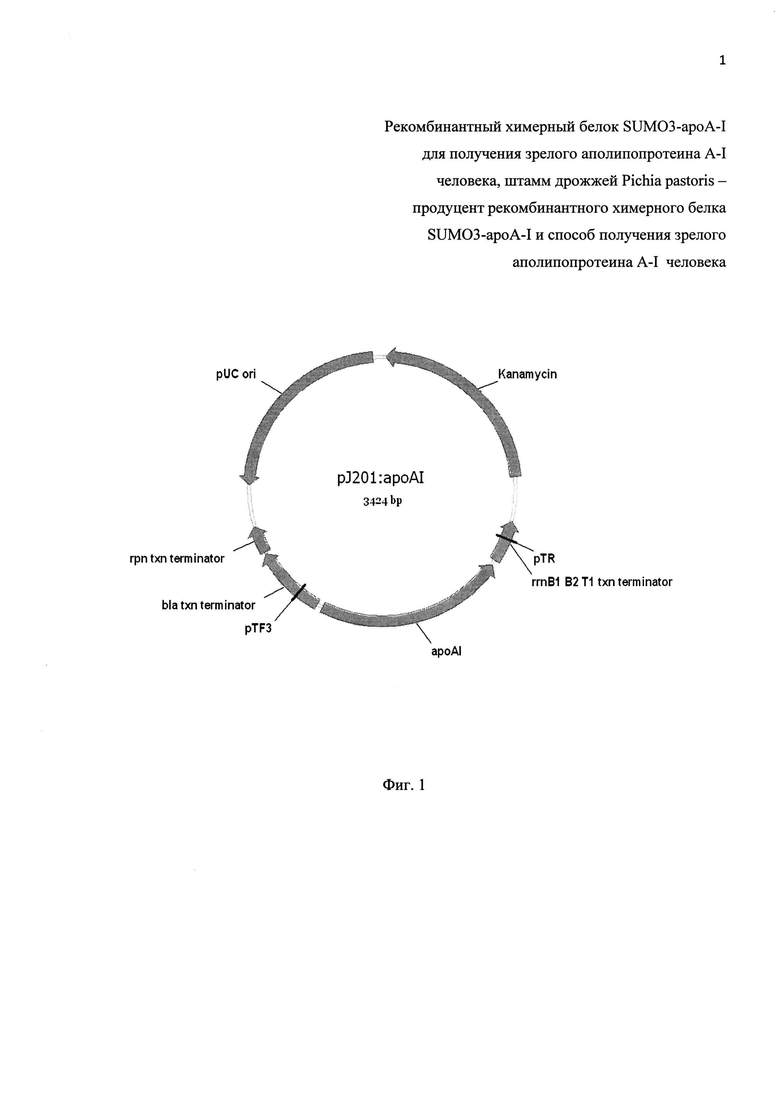
Фиг. 1. Физическая карта рекомбинантной плазмиды pJ201-apoA-I.
pUC ori - область начала репликации; rpn txn terminator, bla txn terminator, rrnB1 B2 txn terminator - терминаторы транскрипции различных генов; Kanamicyn - ген неомицин фосфотрансферазы II, ароА-I - химически синтезированный ген зрелого апоА-I человека.
Фиг. 2. Физическая карта рекомбинантной плазмиды pPsecSUMO3-apoA-I.
pUC origin - область начала репликации; CYC1 transcription terminator - область терминатора транскрипции гена цитохрома C1; Zeo(R) - ген Sh ble, обеспечивающий устойчивость к антибиотику зеоцину; ЕМ7 - прокариотический промотор, TEF1 - эукариотический промотор (фактор элонгации трансляции из Saccharomyces cerevisiae); АОХ1 promoter-5′ область промотора алкоголь оксидазы; АОХ1 transcription terminator-терминатор транскрипции АОХ1; Alpha secretion signal - альфа - сигнал секреции из Saccharomyces cerevisiae; 6Х HisTag - последовательность, кодирующая N-концевую 6-гистидиновую последовательность химерного белка; Human SUMO3 - ген сигнального белка SUMO3 человека; ароА-I - синтетический ген зрелого белка апоА-I человека; MCS - область сайтов для клонирования; 5′АОХ1 и 3′АОХ1 primer - сайты внешних праймеров 5′АОХ1 и 3′АОХ1.
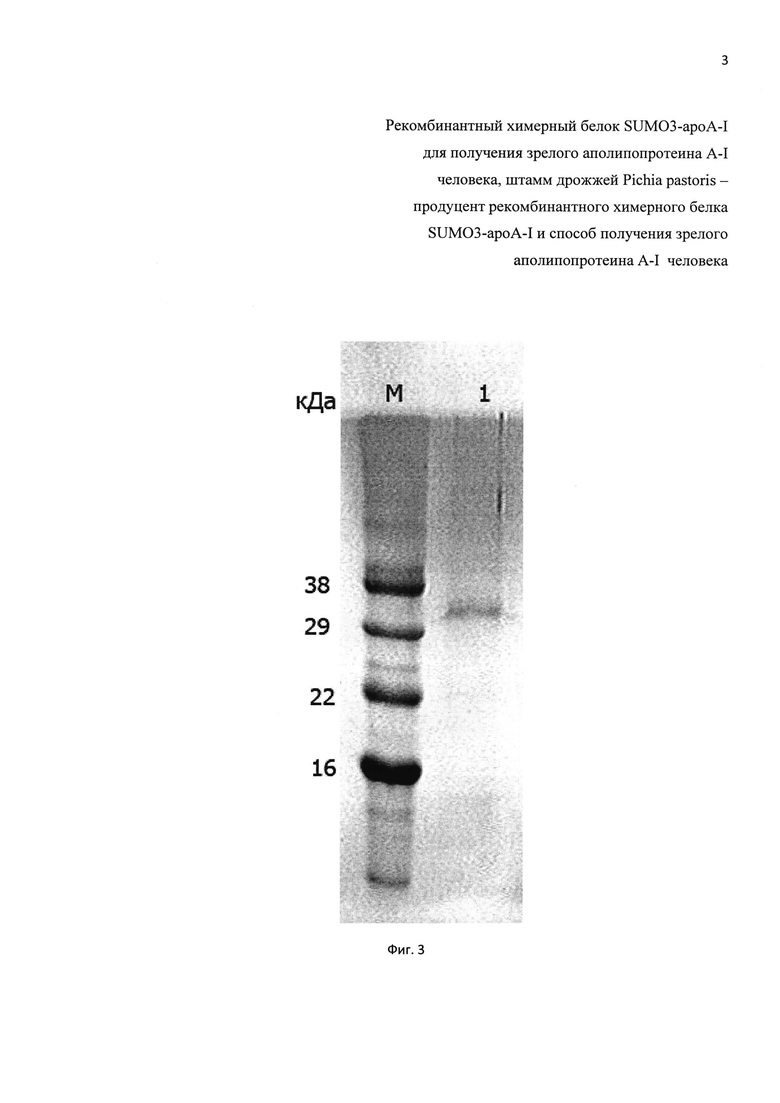
Фиг. 3. Электрофореграмма белков супернатанта, полученного из культуральной жидкости заявляемого продуцента рекомбинантного химерного белка SUMO3-apoA-I на 4-е сутки индукции метанолом (дорожка 1), М - маркер молекулярных масс.
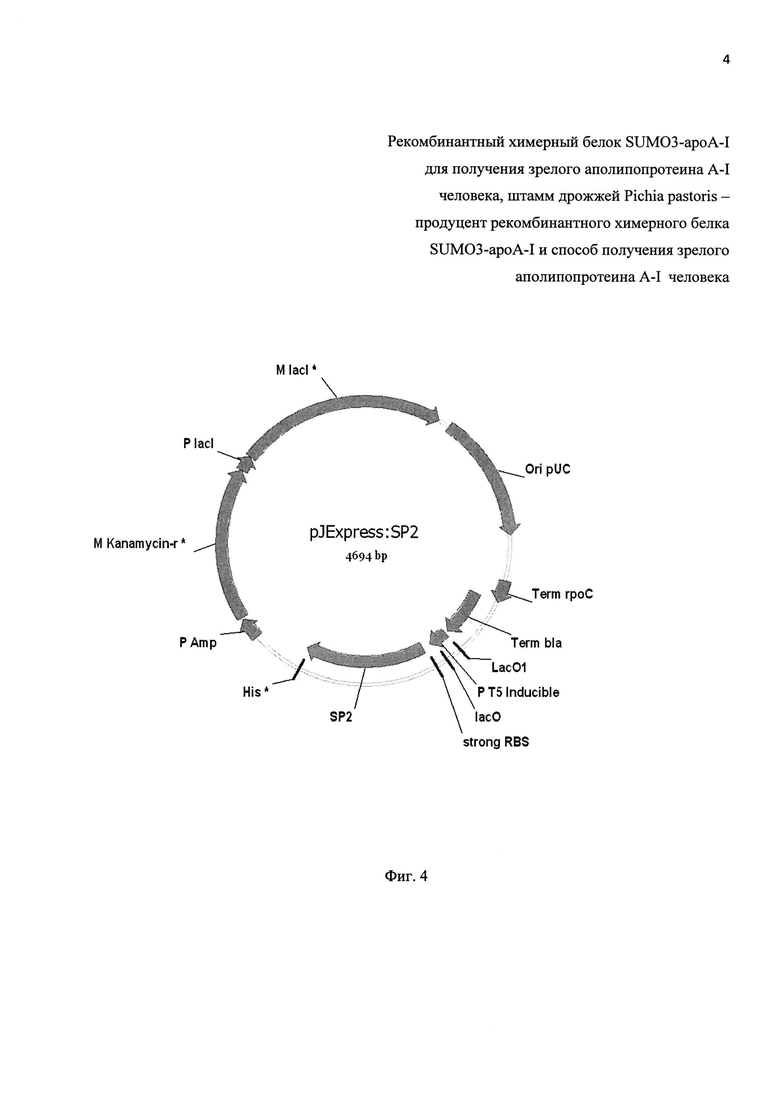
Фиг. 4. Физическая карта рекомбинантной плазмиды pJExpress:SP2.
pUC origin - область начала репликации; Term rpoC - терминатор бета-субъединицы РНК полимеразы Е. coli; Term bla - терминатор гена bla (бета-лактамазы); lac 0, Lac0l - регуляторные гены lac оперона; Р Т5 inducible - промотор из фага Т5; strong RBS -сайт связывания рибосомы; SP2 - синтетический ген протеазы SP2; His* - последовательность, кодирующая N-концевую 6-ти гистидиновую последовательность; Р Amp -; Kanamycin-r*
- ген ген неомицин фосфотрансферазы II; Ρ lacl -; M lacl* - регуляторные гены lac оперона.
Фиг. 5. Электрофореграмма образцов рекомбинантного зрелого белка апоА-I человека, полученного на конечной стадии очистки, и нативного апоА-I человека. 1- рекомбинантный зрелый белок апоА-I человека; 2- нативный белок апоА-I человека.
Таблица. Сравнительный анализ нуклеотидных последовательностей фрагмента нативного гена зрелого апоА-I человека с 1 по 60 нуклеотид (№1) и соответствующей области сконструированного на его основе синтетического гена зрелого белка апоА-I человека после оптимизации кодонов (№2) (приводится после списка использованных источников).
Осуществление изобретения
Пример 1. Оптимизация нуклеотидной последовательности проектируемого синтетического гена зрелого белка Α-I человека для его эффективной экспрессии в клетках Р. pastoris. Клонирование синтетического гена апоА-I человека в составе плазмиды pJ201 в клетках Е. coli.
Дизайн нуклеотидной последовательности проектируемого синтетического гена зрелого белка апоА-I человека разрабатывают на основе известной аминокислотной последовательности нативного аполипопротеина Α-I человека при помощи программы Gene designer (DNA2.0 USA), руководствуясь информацией о кодоновом составе эффективно экспрессирующихся генов Pichia pastoris (21). При проектировании синтетического гена ароА-I человека для проведения оптимизации состава кодонов в гене зрелого белка ароА-I человека выбирают 199 позиций, по которым проводят синонимические нуклеотидные замены, нашедшие отражение в нуклеотидных последовательностях SEQ ID NO 1, SEQ ID NO 3, с целью увеличения экспрессии гена зрелого белка ароА-I человека в P. pastoris. В таблице приведен пример таких замен в 5′-концевом фрагменте гена ароА-I с 1 по 60 нуклеотид.
Кроме того, в проектируемом синтетическом гене ароА-I человека выявляют пары кодонов, часто встречаемых в геноме P. pastoris, которые, по данным (18, 22), значительно тормозят или прерывают в соответствующих участках трансляцию мРНК. Из них выбирают 3 пары кодонов, по которым вводят синонимические нуклеотидные замены: TTCAAG/TTTAAG (позиции: 673-678); AAGAAG/AAGAAA (позиции: 316-321); TTTGAA/TTCGAA (позиции: 97-102). Исследуют наличие в нуклеотидной последовательности мРНК, кодируемой проектируемым синтетическим геном зрелого белка апоА-I человека, нежелательных протяженных участков вторичной структуры с целью предотвращения их образования в конечном варианте конструируемого гена. Предусматривают в проектируемом синтетическом гене наличие необходимых для клонирования сайтов рестрикции при помощи программы Visual Gene Developer (23). Далее проводят химико-ферментативный синтез спроектированного гена зрелого белка апоА-I человека (нуклеотидная последовательность SEQ ID NO 3. Синтезированный ген клонируют в составе плазмиды pJ201 (DNA2.0, USA), в результате чего получают рекомбинантную плазмиду pJ201-apoA-I (фиг. 1).
Пример 2. Конструирование рекомбинантной плазмиды pPsecSUMO3-apoA-I
Конструирование рекомбинантной плазмиды pPsecSUMO3-apoA-I осуществляют с использованием рекомбинантной плазмиды pJ201-apoA-I, синтезированной в примере 1, и коммерческой плазмиды pPsecSUMO3 (Lifesensors, USA) (19).
На матрице рекомбинантной плазмиды pJ201-apoAI методом ПЦР с помощью высокоточной термостабильной полимеразы Phusion (Thermo scientific, USA) амплифицируют нуклеотидную последовательность гена зрелого белка ароА-I человека. Для амплификации используют прямой (Apo-F) и обратный (Apo-R) праймеры, с помощью которых на 5′-концах 2-х цепей ампликона достраивают сайты рестрикции BsmBI и XbaI, соответственно (сайты в нуклеотидных последовательностях праймеров подчеркнуты):
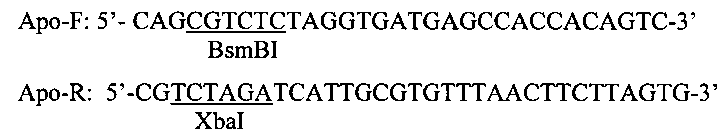
Ампликон, полученный в ходе реакции ПЦР, выделяют при помощи экстракции фенолом/хлороформом с последующим осаждением изопропанолом и гидролизуют рестриктазами BsmBI и XbaI.
Плазмиду pPsecSUMO3 обрабатывают рестриктазой BsmBI и полученную линеаризованную плазмиду pPsecSUMO3 выделяют из агарозного геля при помощи набора GeneJet (Thermo scientific, USA).
Ампликон гена ароА-I человека, гидролизованный рестриктазами BsmBI и XbaI, и линеаризованную плазмиду pPsecSUMO3 «сшивают» (лигируют) с помощью ДНК-лигазы фага Т4. Лигирование проводят 15 мин при +20°С в буферной смеси следующего состава: 66 mM Tris-HCl (рН=7.6 at 25°С), 10 mM MgCl2, 1 mM дитиотреитол, 1 mM АТФ, 7,5% полиэтилен гликоль (PEG 6000). После завершения реакции лигирования проверяют наличие в реакционной смеси «сшитых» между собой фрагментов с помощью ПЦР в присутствии прямого (AOX1-F) и обратного (AOX1-R) праймеров, нуклеотидные последовательности которых представлены ниже:
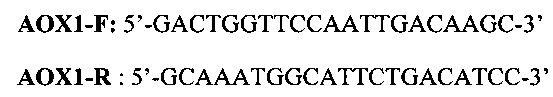
После трансформации клеток E.coli BL 21 (DE3) лигазной смесью получают клоны, содержащие рекомбинантную плазмиду pPsecSUMO3-apoA-I. Отбор клонов, содержащих плазмиду pPsecSUMO3-apoA-I, проводят с помощью ПЦР колоний в присутствии праймеров AOX1-F и AOX1-R. На фиг. 2 приведена физическая карта рекомбинантной плазмиды pPsecSUMO3-apoA-I. Карта включает ген (Sh ble), обусловливающий устойчивость трансформантов E.coli и Pichia pastoris к антибиотику зеоцину, регион репликации pUC, обеспечивающий амплификацию плазмиды pPsecSUMO3-apoA-I в клеках E.coli, нуклеотидные последовательности промотора и терминатора гена алкоголь-оксидазы (АОХ1), участвующие в гомологичной рекомбинации с соответствующими последовательностями гена (АОХ1) в геноме Pichia pastoris и обусловливающих встройку в геном нуклеотидной последовательности, кодирующей рекомбинантный химерный белок SUMO3-apoA-I. Между нуклеотидными последовательностями промотора и терминатора гена АОХ1 находится нуклеотидная последовательность препро-альфа сигнала секреции из пекарских дрожжей Saccharomyces cerevisiae и нуклеотидная последовательность, кодирующая рекомбинантный химерный белок SUMO3-apoA-I (SEQ ID NO 1).
Ген убиквитин-подобного белка SUMO3 человека, входящий в состав рекомбинантной плазмиды pPsecSUMO3-apoA-I, имеет размер 300 п. н. (нуклеотидная последовательность SEQ ID NO 2). Ген зрелого белка апоА-I человека, входящий в состав рекомбинантной плазмиды pPsecSUMO3-apoA-I, имеет размер 729 п. н. (нуклеотидная последовательность SEQ ID NO 3).
Пример 3. Конструирование заявляемого штамма дрожжей Pichia pastoris - продуцента рекомбинантного химерного белка SUMO3-apoA-I
В качестве штамма-реципиента используют штамм дрожжей Pichia pastoris Х-33, содержащий в геноме ген алкоголь-оксидазы АОХ1, нуклеотидные последовательности промотора и терминатора которого содержится так же в рекомбинантной плазмиде pPsecSUMO3-apoA-I. Трансформацию штамма дрожжей Pichia pastoris Х-33 осуществляют с использованием линеаризованной плазмиды pPsecSUMO3-apoA-I, которую получают следующим образом.
Рекомбинантную плазмиду pPsecSUMO3-apoA-I, полученную, как в примере 2, обрабатывают рестриктазой BstXI, которая гидролизует указанную плазмиду в области промотора гена АОХ1, затем проводят очистку полученной линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I фенолом/хлороформом и осаждение изопропанолом. Для трансформации клеток штамма дрожжей Pichia pastoris Х-33 посредством электропорации берут 10 мкг линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I.
Компетентные клетки штамма дрожжей Pichia pastoris Х-33 получают следующим образом.
Индивидуальную колонию клеток штамма дрожжей Pichia pastoris Х-33 инокулируют в 5 мл среды YPD (пептон 2%, дрожжевой экстракт 1%, глюкоза (декстроза) 2%) в 50 мл пробирке и культивируют на качалке при 30°С и 160 об/мин в течение ночи; инокулируют 200 мл свежей среды в 1-литровой колбе 200 мкл ночной культуры. Культивируют в течение ночи на качалке при 30°С и 160 об/мин до оптической плотности (600 нм), равной 1,3-1,5 оптических единиц (о.е.); центрифугируют клетки при 1500×g в течение 5 мин при +4°С. Суспендируют осадок в 200 мл ледяной (0°С) стерильной деионизованной воды. Центрифугируют клетки при 1500×g в течение 5 мин при +4°С, затем ресуспендируют осадок в 100 мл ледяной (0°С) стерильной деионизованной воды. Центрифугируют клетки при 1500×g в течение 5 мин при +4°С, затем ресуспендируют осадок в 8 мл ледяного (0°С) раствора 1М сорбитола. Клетки центрифугируют при 1500×g в течение 5 мин при +4°С, затем ресуспендируют осадок в 400 мкл ледяного (0°С) раствора 1М сорбитола, получив конечный объем примерно 600 мкл. Клетки держат на льду, используют в этот же день. Для трансформации берут аликвоты по 40 мкл суспензии компетентных клеток.
Электропорацию клеток штамма дрожжей Pichia pastoris Х-33 проводят на приборе Gene Puiser (Biorad) при следующих параметрах: кювета 2 мм, напряжение 2000 В, емкость 25 мкф, сопротивление 200 Ом. В кювету, предварительно охлажденную во льду, вносят 40 мкл суспензии компетентных клеток, содержащей 10 мкг линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I. Кювету инкубируют на льду 2 мин, затем дают электроимпульс, сразу же добавляют в кювету 1 мл ледяного раствора 1М сорбитола, переносят клетки в стерильный флакон емкостью 15 мл, инкубируют при 30°С при медленном покачивании 2-3 часа. Затем высевают 100 и 200 мкл суспензии клеток на чашки с агаризованной средой YPD, содержащей антибиотик зеоцин в концентрации 100 мкг/мл.
Колонии трансформантов Pichia pastoris Х-33 появляются на чашках на третьи или четвертые сутки. Отбор клонов проводят, выращивая индивидуальные колонии в пробирках с 5 мл среды GY (пептон 2%, дрожжевой экстракт 1%, глицерин 1%) в течение 2-х суток. Затем в пробирки с культурами ежедневно в течение 4-х суток вносят 10% метанол до конечной концентрации 1%. По окончании индукции биосинтеза белка метанолом пробы центрифугируют при 15000 g в течение 5 минут. Аликвоты по 15 мкл супернатантов анализируют с помощью электрофореза в полиакриламидном геле в денатурирующих условиях (SDS-PAAG). Отбирают клоны, продуцирующие наибольшее количество рекомбинантного химерного белка SUMO3-apoA-I. Для дальнейшей работы отбирают клон, обеспечивающий индуцируемый синтез секретируемого химерного белка SUMO3-apoA-I в количестве около 600 мкг/мл культуральной жидкости. На фиг. 3 приведена электрофореграмма образцов супернатанта культуральной жидкости заявляемого штамма продуцента рекомбинантного химерного белка SUMO3-apoA-I на 4-е сутки индукции метанолом.
При трансформации методом электропорации происходит встраивание линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I в геном дрожжей Pichia pastoris путем гомологичной рекомбинации и образование трансформантов генетически стабильных как в присутствии, так и в отсутствии в среде культивирования селективного антибиотика (14). В результате трансформации и отбора клонов получают заявляемый рекомбинантный штамм дрожжей Pichia pastoris, который при индукции метанолом способен синтезировать и секретировать в культуральную жидкость рекомбинантный химерный белок SUMO3-apoA-I.
Полученный рекомбинантный штамм дрожжей Pichia pastoris характеризуется следующими свойствами:
Морфологические и кулътуралъные характеристики. При росте на плотной среде YPD колонии круглые, гладкие, блестящие, кремовые, край ровный, диаметр колоний 3-6 мм; консистенция пастообразная. Рост в жидкой среде YPD характеризуется ровным помутнением с образованием легкого осадка.
Физиолого-биохимические характеристики. Клетки растут при температуре 4-30°С при оптимуме рН 4,0-6,2. В качестве источника азота используют как минеральные соли в виде смеси, так и органические соединения в виде пептона, дрожжевого экстракта, аминокислот. В качестве источника углерода используют глицерин, глюкозу, метанол.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к зеоцину (до 200 мкг/мл), обусловленную внесением в геном при трансформации линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I гена Sh-ble.
Продуктивность. Заявляемый штамм обеспечивает синтез рекомбинантного химерного белка SUMO3-apoAI в количестве примерно 600 мг на литр культуральной среды.
Пример 4. Клонирование последовательности ДНК, кодирующей SUMO-специфичную протеазу SP2, и конструирование штамма Е. coli - продуцента указанной протеазы.
Вариант синтетического гена SUMO-специфичной протеазы SP2 проектируют на основе известной аминокислотной последовательности протеазы SENP2 человека [база данных UniProt Q9HC62] при помощи программы Gene designer (DNA2.0 USA). При проектировании гена используют те же приемы, которые были описаны выше для расчета последовательности гена ароА-1 человека. В частности, в последовательности гена предусматривают кодоны, обеспечивающие эффективную экспрессию гена в клетках Е. coli, и исключают присутствие в гене наиболее часто встречаемых в геноме Е. coli пар кодонов. Кроме того, с помощью компьютерных программ рассчитывают вторичную структуру мРНК протеазы SP2, кодируемой проектируемым геном, с целью минимизации формирования вторичной структуры в области промотора, рибосомо-связывающего сайта и инициирующего ATG кодона. Синтетический ген SUMO-специфичной протеазы SP2 имеет размер 300 п. н (SEQ ID NO 4). Его получают в составе экспрессирующей прокариотической плазмиды pJExpress (DNA2.0), обеспечивающей индуцируемую экспрессию генов в клетках E.coli под контролем сильного промотора бактериофага Т5 и содержащей ген, обеспечивающий устойчивость к антибиотику канамицину. На фиг. 4 приведена физическая карта рекомбинантной плазмиды pJExpress:SP2.
Пример 5. Получение штамма Е. coli -продуцента SUMO-специфичной протеазы SP2.
Рекомбинантную плазмиду pJExpress:SP2, содержащую ген SUMO-специфичной протеазы SP2, полученную, как в примере 4, используют для трансформации компетентных клеток Е. coli BL 21 (DE3). Колонии трансформантов отбирают на чашках со средой LB (пептон 1%, дрожжевой экстракт 0,5%, NaCl 0,5%) с 30 мкг/мл канамицина.
Пример 6. Биосинтез, выделение и очистка SUMO-специфичной протеазы SP2.
Индивидуальную колонию трансформированных клеток штамма pJExpress:SP2 засевают в пробирку с 2 мл среды LB, содержащей канамицин в концентрации 30 мкг/мл и выращивают в течение 16-18 часов на орбитальном шейкере со скоростью 200 об/мин при температуре 37°С. Ночную культуру переносят в 500 мл свежей среды LB, содержащей канамицин в концентрации 30 мкг/мл. Культивируют в прежних условиях до достижения оптической плотности (600 нм), равной 0,9 оптических единиц (о.е.). К растущей культуре добавляют индуктор IPTG до концентрации 1 mM. Индукцию проводят в течение 12 часов при 30°С на орбитальном шейкере со скоростью 200 об/мин.
Клетки собирают центрифугированием (3500×g) и суспендируют в 8 мл лизирующего буфера следующего состава: 50 мМ Трис-HCl, рН 8,0; 500 мМ NaCl; 6 M гуанидин гидрохлорид; 20 мМ имидазол рН 8,0. Суспензию клеток обрабатывают ультразвуком во льду 6 раз по 30 секунд с перерывами по 1 мин при помощи ультразвукового дезинтегратора. Лизат центрифугируют в течение 15 минут при 3000 g.
2 мл Ni-NTA агарозы (Qiagen) уравновешивают лизирующим буфером и инкубируют с осветленным лизатом в течение часа при медленном переворачивании на ротаторе. Смолу промывают два раза по 4 мл денатурирующего буфера рН 8,0 (8М мочевины; 50 mM Трис-HCl рН 8.0; 500 mM NaCl), два раза по 4 мл денатурирующего буфера рН 6,0 и один раз 4 мл мл денатурирующего буфера рН 5,3. Затем смолу переносят в колонку и проводят элюцию буфером рН 4.0 (8М мочевины; 50 mM Трис-HCl рН 4.0; 500 mM NaCl). В полученный элюат вносят Твин-20 до концентрации 0,1%.
Ренатурацию фермента проводят, последовательно диализуя элюат против буферов, содержащих постепенно снижающуюся концентрацию 6,- 4,- 2,- 0 M мочевины; 50 мМ Трис-HCl рН 8,0; 300 мМ NaCl; 2 мМ дитиотреитола и 0,1% Твин-20.
Пример 7. Биосинтез и выделение рекомбинантного химерного белка SUMO3-ароА-1 человека из клеток заявляемого штамма дрожжей. Pichia pastoris,
Индивидуальную колонию клеток заявленного штамма дрожжей Pichia pastoris переносят в пробирку с 5 мл среды GY (пептон 2%, дрожжевой экстракт 1%, глицерин 1%), выращивают на орбитальном шейкере при 22-25°С и 160 об/мин в течении суток, затем заменяют ватно-марлевую пробку на 2 слоя марли для лучшей аэрации, и продолжают культивирование еще в течение суток. После 2-х суток предварительного роста начинают индукцию метанолом, добавляя по 500 мкл 10% метанола каждый день в течение 4-х суток при 22-25°С. По окончании индукции клетки осаждают центрифугированием при 3500 g в течение 20 мин. при +4°С и отделяют супернатант. Рекомбинантный химерный белок SUMO3-apoA-I человека осаждают из супернатанта, постепенно добавляя в течении 30 мин сульфат аммония до конечной концентрации 60% от насыщения, инкубируют около часа при комнатной температуре и центрифугируют при 14000 об/мин 20 минут при комнатной температуре. Осадок ресуспендируют в фосфатно-солевом буфере (PBS) и проводят диализ против PBS в течение ночи при +4°С. Чистота полученного рекомбинантного химерного белка SUMO3-apoA-I человека составляет не менее 85% по результатам электрофоретического анализа.
Пример 8. Способ получения зрелого аполипопротеина Α-I человека путем ферментативного гидролиза рекомбинантного химерного белка SUMO3-apoA-I SUMO-специфичной протеазой SP2.
Рекомбинантный химерный белок SUMO3-apoA-I, полученный, как в примере 7, инкубируют с SUMO-специфичной протеазой SP2, полученной, как в примере 6 в PBS с 2 mM дитиотреитола в соотношении химерный белок/фермент 1/200 в течение 3-х часов при +30°С. Очистку целевого белка проводят методом аффинной хроматографии в нативных условиях на колонке с Ni-NTA агарозой (Qiagen). По результатам электрофоретического анализа чистота полученного белка составляет не менее 90%.
Полученный зрелый аполипопротеин Α-I человека, размером 243 а.о. имеет молекулярную массу 28 кДа и реагирует с поликлональными антителами к нативному белку апоА-I человека. Как видно на электрофореграмме (фиг. 5), образцы рекомбинантного зрелого аполипопротеина Α-I человека, полученного на конечной стадии очистки, и нативный белок апоА-I человека имеют одинаковую электрофоретическую подвижность.
Выход рекомбинантного зрелого аполипопротеина Α-I человека составляет 400 мг/л.
Список использованных источников
1. Scanu А.М, Edelstein С., Keim Ph. Serum Lipoproteins // The Plasma Proteins, Volume 1 (Second Edition). Academic Press. 1975. P. 317-391.
2. Vitello L.B., Scanu A.M., Studies on human serum high density lipoproteins. Self-association of apolipoprotein A-I in aqueous solutions // J. Biol. Chem. 1976. 251 (4). P. 1131-1136.
3. Brouillette C.G., Anantharamaiah G.M., Engler J.Α., Borhani D.W. Structural models of human apolipoprotein A-I: a critical analysis and review // 2001. Biochim. Biophys. Acta. 1531 (1-2. P.:40-46.
4. Haas M.J., Horani M., Mreyoud Α., Plummer В., Wong N.C., Mooradian A.D. Suppression of apolipoprotein AI gene expression in HepG2 cells by TNF alpha and IL-1 beta // Biochim Biophys Acta. 2003. 1623(2-3). P. 120-128.
5. Smith J.D. Apolipoprotein A-I and its mimetics for the treatment of atherosclerosis // Curr Opin Investig Drugs. 2010. 11(9). P. 989-996. Review.
6. Торховская Т.И., Ипатова O.M., Медведева H.B., Иванов B.C., Иванова Л.И. Липопротеины плазмы крови как переносчики лекарств. Влияние фосфолипидных лекарственных форм // Вестник Российской академии медицинских наук. 2010. №5. С. 42-50.
7. Rensen P.C., de Vrueh R.L., Kuiper J., Bijsterbosch M.K., Biessen E.A., van Berkel T.J. Recombinant lipoproteins: lipoprotein-like lipid particles for drug targeting // Adv Drug Deliv Rev. 2001. 47(2-3). P. 251-76. Review.
8. Lacko A.G., Nair M., Prokai L., McConathy W.J. Prospects and challenges of the development of lipoprotein-based formulations for anti-cancer drugs // Expert Opin. Drug Deliv. 2007. 4(6). P. 665-675. Review.
9. Поляков Л.М. Липопротеины плазмы крови: транспортная система для ксенобиотиков и биологически активных веществ // Сибирский научный медицинский журнал. 1998. Т. 18. №3. С. 23-29.
10. Bergeron J., Frank P.G., Emmanuel F., Latta M., Zhao Y., Sparks D.L., Rassart Ε.,  P., Marcel Y.L. Characterization of human apolipoprotein A-I expressed in Escherichia coli // Biochim Biophys Acta. 1997. 1344(2). P. 139-152
P., Marcel Y.L. Characterization of human apolipoprotein A-I expressed in Escherichia coli // Biochim Biophys Acta. 1997. 1344(2). P. 139-152
11. Mayer H, Rapin AM, Schmidt G, Boman HG. Immunochemical studies on lipopolysaccharides from wild-type and mutants of Escherichia coli K-12. Eur J Biochem. 1976. 66(2). P. 357-368.
12. Petsch D., Anspach F.B. Endotoxin removal from protein solutions // J Biotechnol. 2000. 76(2-3). P. 97-119. Review.
13. Мамаев А.Л., Беклемишев А.Б., 2014, Сибирский научный медицинский журнал, Т. 34. №5. С. 37-42.
14. Cregg J.M. Methods in Molecular Biology: Pichia Protocols / Second Edition. Humana Press, 2007.389. P. 1-10.
15. Feng M.Q., Cai Q.S., Song D.X., Dong J.B., Zhou P. High yield and secretion of recombinant human apolipoprotein AI in Pichia pastoris // Protein Expr Purif. 2006. 46 (2). P. 337-342.
16. Malakhov M.P., Mattern M.R., Malakhova O.A., Drinker M., Weeks S.D., Butt T.R. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins // J. Struct. Funct. Genomics. 2004. 5 (1-2). P. 75-86.
17. Marblestone J.G., Edavettal S.C., Lim Y., Lim P., Zuo X., Butt T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: enhanced expression and solubility with SUMO // Protein Sci. 2006. 15(1). P. 182-189.
18. Hatfield G.W., Roth D.A. Optimizing scaleup yield for protein production: Computationally Optimized DNA Assembly (CODA) and Translation Engineering // Biotechnol Annu Rev. 2007. 13. P. 27-42. Review.
19. Lammertyn E., Van Mellaert L., Bijnens A.P., Joris В.,  J. Codon adjustment to maximise heterologous gene expression in Streptomyces lividans can lead to decreased mRNA stability and protein yield // Mol Gen Genet. 1996.250 (2). P. 223-229.
J. Codon adjustment to maximise heterologous gene expression in Streptomyces lividans can lead to decreased mRNA stability and protein yield // Mol Gen Genet. 1996.250 (2). P. 223-229.
20. Sumo Expression Systems (SUMOstar Protease, Lifesensors, USA), www. lifesensors. com.
21. Bai J., Swartz D.J., Protasevich I.I., Brouillette C.G., Harrell P.M., Hildebrandt E., Gasser В., Mattanovich D., Ward A., Chang G., Urbatsch I.L. A gene optimization strategy that enhances production of fully functional P-glycoprotein in Pichia pastoris // PLoS One. 2011. 6(8). e22577. doi: 10.1371/journal.pone.0022577.
22. Irwin В., Heck J.D., Hatfield G.W. Codon pair utilization biases influence translational elongation step times // J Biol Chem. 1995. 270 (39). P. 22801-22806.
23. Jung S.K., McDonald K. Visual gene developer: a fully programmable bioinformatics software for synthetic gene optimization // BMC Bioinformatics. 2011. 12. P. 340. doi: 10.1186/1471-2105-12-340.






Изобретение относится к генной инженерии и может быть использовано для получения зрелого аполипопротеина A-I человека. Для увеличения выхода зрелого аполипопротеина A-I человека его получают путем ферментативного гидролиза SUMO-специфичной протеазой SP2 рекомбинантного химерного белка SUMO3-апоА-I, синтезированного штаммом дрожжей Pichia pastoris. Указанный штамм сконструирован путем трансформации штамма дрожжей Pichia pastoris Х-33 линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I. Рекомбинантный химерный белок SUMO3-apoA-I включает с N-конца аминокислотную последовательность убиквитин-подобного белка SUMO3 человека, слитую с С-конца с аминокислотной последовательностью зрелого белка апоА-I человека. 3 н.п. ф-лы, 1 табл., 5 ил., 7 пр.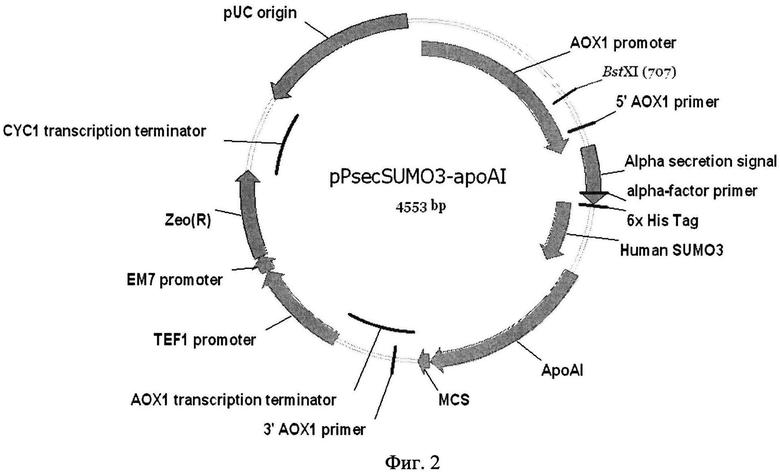
1. Рекомбинантный химерный белок SUMO3-apoA-I, имеющий аминокислотную последовательность SEQ ID NO: 1, предназначенный для получения зрелого аполипопротеина A-I человека.
2. Штамм дрожжей Pichia pastoris X-33/pPsecSUMO3-apoA-I - продуцент рекомбинантного химерного белка SUMO3-apoA-I по п. 1, полученный путем трансформации дрожжей Pichia pastoris Х-33 линеаризованной плазмидной ДНК pPsecSUMO3-apoA-I, включающей в свой состав нуклеотидную последовательность SEQ ID NO: 1, кодирующую рекомбинантный химерный белок SUMO3-apoA-I, при этом указанная плазмидная ДНК характеризуется физической картой, приведенной на фиг. 2.
3. Способ получения зрелого аполипопротеина A-I человека, включающий микробиологический синтез рекомбинантного химерного белка SUMO3-апоА-I по п. 1 в процессе культивирования штамма дрожжей Pichia pastoris X-33/pPsecSUMO3-apoA-I по п. 2, осаждение клеток указанного штамма, отделение супернатанта, осаждение из него рекомбинантного химерного белка SUMO3-apoA-I, его ресуспендирование и диализ, последующий ферментативный гидролиз SUMO-специфичной протеазой SP2.
| МАМАЕВ А.Л | |||
| и др., КЛОНИРОВАНИЕ И АНАЛИЗ ЭКСПРЕССИИ СИНТЕТИЧЕСКИХ ГЕНОВ АПОЛИПОПРОТЕИНА А-I ЧЕЛОВЕКА В КЛЕТКАХ ESCHERICHIA COLI И МЕТИЛОТРОФНЫХ ДРОЖЖЕЙ PICHIA PASTORIS, БЮЛЛЕТЕНЬ СО РАМН, т | |||
| Нивелир для отсчетов без перемещения наблюдателя при нивелировании из средины | 1921 |
|
SU34A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| WANG X.J | |||
| et al., Recombinant production of the antimicrobial peptide NZ17074 in Pichia pastoris using SUMO3 as a fusion partner, Lett | |||
| Appl | |||
| Microbiol., 2014, v.59, n.1, p.71-78 | |||
| RU 2013108962 A, 10.09.2014. | |||

