Область техники, к которой относится изобретение
Настоящее изобретение относится к новым лизофосфолипид-ацилтрансферазам.
Уровень техники
Биосинтез полиненасыщенных жирных кислот
Жирные кислоты являются главными компонентами липидов, таких как фосфолипиды и триацилглицерины. Жирные кислоты, содержащие две или более ненасыщенных связей, обобщенно называются полиненасыщенными жирными кислотами (PUFA), и они, как известно, включают арахидоновую кислоту, дигомо-γ-линоленовую кислоту, эйкозапентаеновую кислоту, докозагексаеновую кислоту и т.д. Сообщается о различных физиологических активностях этих жирных кислот (непатентный документ 1).
Предполагается, что такие полиненасыщенные жирные кислоты найдут применение в различных областях, но некоторые из них не могут быть синтезированы in vivo у животных. Это привело к развитию способов получения полиненасыщенных жирных кислот культивированием различных микроорганизмов. Также были предприняты попытки производства полиненасыщенных жирных кислот в растениях. Известно, что в подобных случаях полиненасыщенные жирные кислоты накапливаются в качестве компонентов запасных липидов, таких как триацилглицерины, например, в микробных клетках или семенах растений.
Среди полиненасыщенных жирных кислот привлекла внимание арахидоновая кислота в качестве промежуточного метаболита в синтезе простагландинов, лейкотриенов и подобного, и было предпринято множество попыток для ее использования в качестве вещества для функциональных продуктов питания и медикаментов. Кроме того, арахидоновая кислота содержится в грудном молоке, таким образом, она важна для роста младенцев, особенно для развития длины плода и мозга, и, таким образом, она также привлекает внимание с точки зрения питания как необходимый компонент для роста младенцев, наряду с DHA (докозагексаеновая кислота).
Арахидоновая кислота биосинтезируется по пути, показанном на фиг.1. В частности, арахидоновая кислота продуцируется через несколько стадий элонгации цепи и десатурации из пальмитиновой кислоты, генерированной синтезом жирных кислот de novo. В этом пути элонгаза и Δ9 десатураза воздействуют на ацил-CoA. С другой стороны известно, что Δ12 десатураза, Δ6 десатураза и Δ5 десатураза воздействуют на ацильные группы фосфолипидов, таких как фосфатидилхолин (непатентный документ 2). Таким образом, требуется ацильный перенос между ацил-CoA и фосфолипидами в биосинтезе PUFA, таких как арахидоновая кислота. Не ограничиваясь биосинтезом PUFA, замена только жирных кислот после биосинтеза фосфолипидов известна как “ремоделирование” фосфолипидов, и лизофосфолипид-ацилтрансферазы (здесь и далее в данном описании называемые “LPLAT”) известны, как принимающие участие в этой реакции (непатентный документ 3).
Биосинтез триацилглицеринов
Среди запасных липидов триацилглицерины синтезируются in vivo следующим образом. Глицерин-3-фосфат ацилируется глицерин-3-фосфат-ацилтрансферазой (здесь и далее в данном описании иногда называемой “GPAT”) по гидроксильной группе в положении 1 (α-положение) с образованием лизофосфатидной кислоты (здесь и далее в данном описании иногда называемой “LPA”). LPA представляет собой лизофосфолипид, содержащий только одну ацильную группу, и он ацилируется ацилтрансферазой лизофосфатидной кислоты (здесь и далее в данном описании иногда называемой “LPAAT”) с образованием фосфатидной кислоты (здесь и далее в данном описании иногда называемой “PA”). PA дефосфорилируется фосфатазой фосфатидной кислоты с образованием диацилглицерина, который, в свою очередь, ацилируется диацилглицерин-ацилтрансферазой (здесь и далее в данном описании иногда называемой “DGAT”) с образованием триацилглицерина. Известно, что ацил-CoA: холестерин-ацилтрансфераза (здесь и далее в данном описании иногда называемая “ACAT”) и лизофосфатидилхолин-ацилтрансфераза (здесь и далее в данном описании иногда называемая “LPCAT”) и подобные опосредованно вовлечены в биосинтез триацилглицеринов.
Биосинтез фосфолипидов
PA, продуцированные из LPA, путем воздействия LPAAT, как описано выше, служат в качестве предшественника в биосинтезе различных фосфолипидов. Например, важные фосфолипиды, такие как фосфатидилэтаноламин (PE), фосфатидилхолин (PC), фосфатидилсерин (PS), фосфатидилинозитол (PI) и фосфатидилглицерин (PG) биосинтезируются из PA. Таким образом, PA является не только промежуточным продуктом в липидном синтезе, но также является внутриклеточным и межклеточным липидным медиатором, обладающим очень широким диапазоном биологических и фармакологических эффектов, таких как клеточная пролиферация, агрегация тромбоцитов, сокращение гладкой мускулатуры, повышение раковой инвазии и т.д.
Лизофосфолипид-ацилтрансферазы
Как описано выше, предполагается, что LPLAT вовлечены в биосинтез PUFA. LPLAT, совместно называемые ферментами, обладающими активностью введения ацильной группы в лизофосфолипиды, и включают ферменты, имеющие различные названия, основанные на специфичности к субстрату, т.е. молекулярным видам лизофосфолипидов, используемым в качестве субстрата. Одним из примеров являются LPAAT, которые принимают участие в синтезе триацилглицеринов и фосфолипидов при использовании LPA в качестве субстрата. Другие лизофосфолипиды, на которые воздействуют LPLAT, включают лизофосфатидилхолин (LPC), лизофосфатидилсерин (LPS), лизофосфатидилэтаноламин (LPE), лизофосфатидилинозитол (LPI) и т.д. Таким образом, ферментами называют LPAAT, LPCAT, лизофосфатидилсерин-ацилтрансферазу (LPSAT), лизофосфатидилинозитол-ацилтрансферазу (LPIAT) и подобные, на основании молекулярных видов, на которые они воздействуют. Каждый фермент может специфично воздействовать на один лизофосфолипид или конкретные сложные лизофосфолипиды. Например, LPLAT, называемые LPAAT, включают такие, которые воздействуют не только на LPA, но также на LPC, LPE и т.д.
Основанная на профиле последовательностей классификация лизофосфолипид-ацилтрансфераз
LPLAT классифицируются как глицерофосфолипид-ацилтрансферазы. Глицерофосфолипид-ацилтрансферазы делятся на три группы, исходя из сравнения аминокислотных последовательностей, т.е. семейство LPAAT, семейство MBOAT (мембранно-связанная O-ацилтрансфераза) и семейство DGAT2 (непатентный документ 5). Ферменты, принадлежащие к семейству LPAAT, обычно характеризуются мембранно-связанным доменом и последовательно сохраненным мотивом (LPAAT мотив). Ферменты, принадлежащие к членам семейства LPAAT, включают LPAAT, GPAT и т.д. Ферменты, включенные в семейство MBOAT, обычно характеризуются мембранно-связанным доменом. Известно, что семейство MBOAT включает DGAT, ACAT и подобное в дополнение к LPLAT. Полагают, что некоторые ферменты у животных или подобных, принадлежащие к семейству MBOAT, являются ответственными за реакцию ремоделирования, ответственную за синтез мембранных фосфолипидов.
Сообщается о LPLAT у широкого спектра организмов от одноклеточных организмов, таких как бактерии и дрожжи, до более высокоорганизованных организмов, таких как млекопитающие. В дрожжах (Saccharomyces cerevisiae), принадлежащих к грибам, SLC1 (YDL052C) и SLC4 (YOR175C) (в данном описании иногда называемые “ALE1” или “LPT1”), известны как гены мембранно-связанных LPLAT (непатентный документ 5). Известно, что у животных существуют составные гомологи LPLAT, включая ответственные за реакцию воздействия на LPA в системе триглицеридного синтеза de novo PA и ответственные за фосфолипидное ремоделирование (непатентный документ 6).
В липид-продуцирующем грибе Mortierell alpin (здесь и далее в данном описании иногда называемая “M. alpina”), были получены четыре LPLAT, при этом все они принадлежат к семейству LPAAT (патентные документы 1-3). Тем не менее, нет сведений о том, что какие-либо LPLAT, принадлежащие к семейству MBOAT, были получены из M. alpina.
Список цитированной литературы
Патентные документы
Патентный документ 1: Международная публикация № WO2004/087902
Патентный документ 2: Опубликованная патентная заявка США № US2006/0094090
Патентный документ 3: Международная публикация № WO2008/146745
Непатентные документы
Непатентный документ 1: Lipids, 39, 1147 (2004)
Непатентный документ 2: J.B.C., 278(37), 35115-35126, (2003)
Непатентный документ 3: J.B.C., 276(29), 26745-26752, (2001)
Непатентный документ 4: Proc. Natl. Acad. Sci., 105(8), 2830-2835, (2008)
Непатентный документ 5: J.B.C., 282(42), 30845-30855, (2007)
Непатентный документ 6: J.B.C., 284(1), 1-5, (2009)
Непатентный документ 7: Trends Biochem. Sci., 25, 111-112, (2000)
Непатентный документ 8: Journal of lipid research 2009 R80035 JLR200v1
Сущность изобретения
Технические задачи
Как описано выше, фосфолипидное ремоделирование незаменимо в биосинтезе PUFA, таких как арахидоновая кислота, и LPLAT могут быть вовлечены в эту реакцию. Однако до настоящего времени было известно, что гомологи LPAAT обладают недостатком, в котором количественное соотношение PUFA в общем количестве жирных кислот не может быть достаточно высоким, даже если они были перенесены и экспрессированы в организме-хозяине. Следовательно, необходимо идентифицировать новую нуклеиновую кислоту и белок, которые могут достаточно увеличить количественное соотношение PUFA в общем количестве жирных кислот в хозяине при переносе и экспрессии хозяину. Так же существует необходимость идентифицировать нуклеиновую кислоту и белок, способные продуцировать жиры с высоким содержанием полезных в промышленном отношении жирных кислот, и разработать способ, посредством которого полезные жирные кислоты могут быть произведены или содержание полезных жирных кислот может быть повышено посредством их использования.
Решение задач
Задачей настоящего изобретения является обеспечение белков и нуклеиновых кислот, способных продуцировать полезные жиры посредством их экспрессии в клетке-хозяине для влияния на липидный метаболизм хозяина или для повышения содержания требуемых жирных кислот.
Биосинтез PUFA, таких как арахидоновая кислота, фосфолипидное ремоделирование является необходимым. Липид-продуцирующий гриб M. alpina может накапливать большое количество полезных PUFA, таких как арахидоновая кислота, однако не была получена какая-либо ацилтрансфераза из M. Alpina, принадлежащая к семейству MBOAT, вовлеченному в ремоделирование липидов у животных или тому подобное. Авторы обнаружили это и тщательно исследовали для достижения указанной выше задачи, в результате, авторы получили кДНК, кодирующую фермент, принадлежащий к семейству MBOAT из M. alpina. Дополнительно, авторы предприняли попытку продукции композиции жирной кислоты трансформированием полученной в результате кДНК в высоко пролиферативную клетку-хозяина, такую как дрожжи, для обнаружения того, что клетка-хозяин может продуцировать различные композиции жирных кислот, особенно композицию жирной кислоты, обладающую высоким содержанием арахидоновой кислоты, по сравнению с композицией жирной кислоты, продуцированной хозяевами, трансформированными векторами, содержащими нуклеиновые кислоты, кодирующие известные LPAAT, полученные из M. alpina. Таким образом, авторам удалось клонировать гены новых LPLAT, отличных от известных LPAAT, и завершить настоящее изобретение.
Таким образом, настоящее изобретение обеспечивает следующие аспекты.
(1) Нуклеиновую кислоту по любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы;
(b) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью лизофосфолипид-ацилтрансферазы;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 80% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирующую белок, обладающий активностью лизофосфолипид-ацилтрансферазы;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы; и
(e) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью лизофосфолипид-ацилтрансферазы.
(2) Нуклеиновую кислоту по пункту (1), которая соответствует любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из варианта аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, где аминокислоты 1-50 удалены, замещены или добавлены, и обладающий активностью лизофосфолипид-ацилтрансферазы;
(b) нуклеиновая кислота, которая гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью лизофосфолипид-ацилтрансферазы;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 90% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирующую белок, обладающий активностью лизофосфолипид-ацилтрансферазы;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 90% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы; и
(e) нуклеиновая кислота, которая гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью лизофосфолипид-ацилтрансферазы.
(3) Нуклеиновую кислоту по любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором;
(b) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 80% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирующая белок, обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором; и
(e) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
(4) Нуклеиновую кислоту по пункту (3), которая соответствует любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-50 в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором;
(b) нуклеиновая кислота, которая гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 90% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирующую белок, обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 90% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором; и
(e) нуклеиновая кислота, которая гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
(5) Нуклеиновую кислоту по любому из пунктов (1)-(4), где кодируемый белок принадлежит семейству мембранно-связанных O-ацилтрансфераз.
(6) Нуклеиновую кислоту по любому из нижеследующих пунктов (a)-(d):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:1 или 6, или ее частичную последовательность;
(b) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, или ее частичную последовательность;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:4 или 9, или ее частичную последовательность; и
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:5 или 10, или ее частичную последовательность.
(7) Белок по любому из нижеследующих пунктов (a) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в варианте аминокислотной последовательности SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы.
(8) Белок по пункту (7), который соответствует нижеследующему пункту (a) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-50 в аминокислотной последовательности SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы; или
(b) белок, состоящий из аминокислотной последовательности, обладающей 90% или более идентичностью с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью лизофосфолипид-ацилтрансферазы.
(9) Белок по нижеследующему пункту (a) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую аминокислотную последовательность, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую аминокислотную последовательность, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
(10) Белок по пункту (9), который соответствует нижеследующему пункту (a) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-50 в аминокислотной последовательности SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую аминокислотную последовательность, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 90% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую аминокислотную последовательность, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
(11) Белок по любому из пунктов (7)-(10), который принадлежит семейству мембранно-связанных O-ацилтрансфераз.
(12) Белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7.
(13) Рекомбинантный вектор, содержащий нуклеиновую кислоту по любому из пунктов (1)-(6).
(14) Клетку, трансформированную рекомбинантным вектором по пункту (13).
(15) Композицию жирных кислот, полученную культивированием трансформированной клетки по пункту (14), где количественное соотношение арахидоновой кислоты в композиционном соотношении жирных кислот в указанной композиции жирных кислот выше, чем количественное соотношение арахидоновой кислоты в композиции жирных кислот, полученной культивированием нетрансформированного хозяина.
(16) Способ получения композиции жирных кислот, включающий сбор композиции жирных кислот по пункту (15) из культур трансформированной клетки по пункту (14).
(17) Пищевой продукт, содержащий композицию жирной кислоты по пункту (15).
(18) Способ применения рекомбинантного вектора по пункту (13) для повышения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном вектором, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
(19) Нуклеиновую кислоту по любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и участвующая в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL;
(b) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая содержит нуклеотидную последовательность, кодирующую белок, участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 80% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирующая белок, участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL; и
(e) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая содержит нуклеотидную последовательность, кодирующую белок, участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL.
(20) Белок по нижеследующему пункту (a) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7, и участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL.
Преимущественные эффекты изобретения
LPLAT настоящего изобретения позволяют усовершенствование возможности производства жирных кислот, таких как арахидоновая кислота и/или запасные липиды, и, следовательно, предпочтительны в качестве средств для улучшения производительности полиненасыщенных жирных кислот в микроорганизмах и растениях. Таким образом, они могут обеспечить липиды, обладающие требуемыми характеристиками или свойствами, так что они могут быть эффективно применены для использования в пищевых продуктах, косметических средствах, лекарственных препаратах, мылах и т.д.
Краткое описание фигур
Фиг.1 является схематической диаграммой, демонстрирующей биосинтетический путь арахидоновой кислоты. Аббревиатуры на фиг.1 имеют следующие значения: PL - фосфолипид; CoA - кофермент A; DS - десатураза (фермент десатураза жирных кислот); GLELO - элонгаза жирных кислот; 18:0 - стеарильная группа; 18:1 - олеоильная группа; 18:2 - лильноильная группа; 18:3(n-6) - γ- лильнолеильная группа; DGLA - дигомо-γ-лильнолеильная группа; ARA - арахидоильная группа.
Фиг.2 демонстрирует полную длину последовательности кДНК (SEQ ID NO:4) LPLAT5 из M. alpina штамма 1S-4 и аминокислотную последовательность (SEQ ID NO:2), полученную из нее.
Фиг.3 демонстрирует полную длину последовательности кДНК (SEQ ID NO:9) LPLAT6 из M. alpina штамма 1S-4 и аминокислотную последовательность (SEQ ID NO:7), полученную из нее.
На фиг.4 продемонстрировано сравнение между геномной последовательностью (SEQ ID NO:5) и ORF последовательностью. (SEQ ID NO:1) LPLAT5 из M. alpina штамма 1S-4.
Фиг.4B демонстрирует сравнение между геномной последовательностью (SEQ ID NO:5) и ORF последовательностью (SEQ ID NO:1) LPLAT5 из M. alpina штамма 1S-4.
Фиг.4C демонстрирует сравнение между геномной последовательностью (SEQ ID NO:5) и ORF последовательностью (SEQ ID NO:1) LPLAT5 из M. alpina штамма 1S-4.
Фиг.5 демонстрирует сравнение между геномной последовательностью (SEQ ID NO:10) и ORF последовательностью (SEQ ID NO:6) LPLAT6 из M. alpina штамма 1S-4.
Фиг.5B демонстрирует сравнение между геномной последовательностью (SEQ ID NO:10) и ORF последовательностью (SEQ ID NO:6) LPLAT6 из M. alpina штамма 1S-4.
Фиг.5C демонстрирует сравнение между геномной последовательностью (SEQ ID NO:10) и ORF последовательностью (SEQ ID NO:6) LPLAT6 из M. alpina штамма 1S-4.
Фиг.6 является графиком, демонстрирующим состав композиции полиненасыщенных жирных кислоты в клетках при подавлении экспрессии LPLAT6 или Δ5 десатуразы жирных кислот в M. alpina. На фиг.6, аббревиатуры имеют следующие значения: GLA - γ-линоленовая кислота; DGLA - дигомо-γ-линоленовая кислота; ARA - арахидоновая кислота.
Фиг.7 является графиком, демонстрирующим состав композиции полиненасыщенных жирных кислот в триацилглицериновой фракции, когда подавлена экспрессия LPLAT6 или Δ5 десатуразы жирных кислот M. alpina. На фиг.7, аббревиатуры имеют следующие значения: GLA - γ-линоленовая кислота; DGLA - дигомо-γ-линоленовая кислота; ARA - арахидоновая кислота.
Фиг.8 является графиком, демонстрирующим состав композиции полиненасыщенных жирных кислот в фосфолипидной фракции, когда подавлена экспрессия LPLAT6 или Δ5 десатуразы жирных кислот в M. alpina. На фиг.8, аббревиатуры имеют следующие значения: GLA - γ-линоленовая кислота; DGLA - дигомо-γ-линоленовая кислота; ARA - арахидоновая кислота.
Описание вариантов осуществления
Настоящее изобретение относится к новым лизофосфолипид-ацилтрансферазам (“LPLAT”) рода Mortierell, характеризующихся переносом ацильной группы между ацил-CoA и фосфолипидами в биосинтетическом продуцировании арахидоновой кислоты. Белки настоящего изобретения могут воздействовать на лизофосфолипиды. Донором ацильной группы, как правило, является, ацил-CoA, но не ограничивается этим.
Варианты осуществления настоящего изобретения более конкретно описаны ниже.
Нуклеиновые кислоты, кодирующие лизофосфолипид-ацилтрансферазы настоящего изобретения
Лизофосфолипид-ацилтрансферазы (LPLAT), кодируемые нуклеиновыми кислотами настоящего изобретения, в качестве конкретных примеров включают LPLAT 5 и 6. В отличие от композиции жирных кислот, продуцируемых хозяевами, экспрессирующими известные LPAAT из M. alpina, LPLAT5 и 6 могут продуцировать композиции жирных кислот, характеризующиеся высоким количественным соотношением арахидоновой кислоты, как пояснено в приведенных ниже примерах. Следовательно, LPLAT настоящего изобретения преимущественно продуцируют арахидоновую кислоту с очень высокой эффективностью по сравнению с известными LPAAT из M. alpina.
Взаимосвязь кДНК, CDS, ORF нуклеиновых кислот, кодирующих LPLAT5 и LPLAT6 настоящего изобретения, и аминокислотных последовательностей объединена в приведенной ниже таблице 1.
Таким образом, последовательности, относящиеся к LPLAT5 настоящего изобретения, включают SEQ ID NO:1, представляющую последовательность ORF области LPLAT5; SEQ ID NO:2, представляющую аминокислотную последовательность LPLAT5; SEQ ID NO:3, представляющую последовательность CDS области LPLAT5; SEQ ID NO:4, представляющую нуклеотидную последовательность кДНК; и SEQ ID NO:5, представляющую геномную последовательность. Более конкретно, нуклеотиды 161-1693 SEQ ID NO:4, представляющие последовательность кДНК LPLAT5, соответствуют CDS (SEQ ID NO:3), и нуклеотиды 161-1690 соответствуют ORF (SEQ ID NO:1). Последовательность кДНК LPLAT5 и полученная из нее аминокислотная последовательность показаны на фиг.2. Геномная последовательность (SEQ ID NO:5) LPLAT5 содержит две интронных и экзонную области, соответствующие нуклеотидам 1-314, 461-587 и 668-1759 SEQ ID NO:5.
Подобным образом, последовательности, относящиеся к LPLAT6 настоящего изобретения, включают SEQ ID NO:6, представляющую последовательность ORF области LPLAT6; SEQ ID NO:7, представляющую аминокислотную последовательность LPLAT6; SEQ ID NO:8, представляющую последовательность CDS области LPLAT6; SEQ ID NO:9, представляющую нуклеотидную последовательность кДНК; и SEQ ID NO:10, представляющую геномную последовательность. Более конкретно, нуклеотиды 38-1759 SEQ ID NO:9, представляющие последовательность кДНК LPLAT6, соответствуют CDS (SEQ ID NO:8), и нуклеотиды 38-1756 соответствуют ORF (SEQ ID NO:6). Последовательность кДНК LPLAT6 и полученная из нее аминокислотная последовательность приведены на фиг.3. Геномная последовательность (SEQ ID NO:10) LPLAT6 содержит одну интронную и экзонную области, соответствующие нуклеотидам 1-1095 и 1318-1944 SEQ ID NO:10.
Нуклеиновые кислоты настоящего изобретения включают одноцепочечные и двухцепочечные ДНК, а также их комплементы РНК, и могут быть как природного происхождения, так и искусственно полученными. ДНК включают, но не ограничиваются ими, геномные ДНК, кДНК, соответствующие геномным ДНК, химически синтезированные ДНК, амплифицированные с помощью ПЦР ДНК и их комбинации, например, такие как ДНК/РНК гибриды.
Предпочтительные варианты нуклеиновых кислот настоящего изобретения включают (a) нуклеиновую кислоту, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:1 или 6; (b) нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7; (c) нуклеиновую кислоту, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:4 или 9; или (d) нуклеиновую кислоту, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:5 или 10, и т.д.
Для получения указанных выше нуклеотидных последовательностей, данные о нуклеотидной последовательности EST или геномной ДНК из организма, обладающего LPLAT активностью, могут быть также исследованы на наличие нуклеотидных последовательностей, кодирующих белки, распределяющих высокую идентичность с известными белками, обладающими LPLAT активностью. Организм, обладающий LPLAT активностью, предпочтительно является липид-продуцирующим грибом, таким как, но без ограничения, M. alpina.
Для проведения EST анализа, сначала создают библиотеку кДНК. Методики создания библиотеки кДНК можно найти в “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001)). Также могут быть использованы коммерчески доступные наборы для создания библиотеки кДНК. Методикой создания библиотеки кДНК, подходящей для настоящего изобретения, является, например, такая, как описано ниже. По этой методике подходящий штамм липид-продуцирующего гриба M. alpina инокулируют в соответствующую среду и предварительно культивируют в течение соответствующего периода времени. Культуру собирают в соответствующие моменты времени во время основного культивирования и получают клетки для получения тотальной РНК. Тотальная РНК может быть получена с использованием общеизвестных технологий, таких как гуанидингидрохлоридный/CsCl способ. Поли(A)+РНК может быть очищена из полученной в итоге тотальной РНК с использованием коммерчески доступного набора. Библиотека кДНК также может быть создана с использованием коммерчески доступного набора. Затем могут быть получены EST путем установления нуклеотидной последовательности любых клонов из созданной библиотеки кДНК, с использованием праймеров, сконструированных для того, чтобы дать возможность секвенированию вставки в векторе. Например, направленное клонирование может быть выполнено, когда библиотека кДНК была создана с использованием набора для клонирования ZAP-cDNA GigapackIII Gold (STRATAGENE).
В результате анализа гомологии SEQ ID NO:1 и 6 с использованием BLASTX против аминокислотных последовательностей, размещенных в GenBank, аминокислотная последовательность, полученная из SEQ ID NO:1, демонстрирует гомологию с LPLAT гомологами из грибов, и аминокислотная последовательность, полученная из SEQ ID NO:6, демонстрирует гомологию с LPLAT гомологами из животных. Идентичность нуклеотидной последовательности и идентичность аминокислотной последовательности с последовательностью, демонстрирующей наибольшую идентичность с ORF каждой последовательности, были определены с помощью clustalW, выявившей, что гомолог лизофосфолипид-ацилтрансферазы из Schizosaccharomyces pombe (GI:161085648) продемонстрировал самое низкое E-значение или наибольшую идентичность с SEQ ID NO:1, и идентичность нуклеотидной последовательности и идентичность аминокислотной последовательности в ORF составили 43,2% и 33,3%, соответственно. Подобным образом, предполагаемый белок из Xenopus laevis (GI:56788919) продемонстрировал наибольшую идентичность с SEQ ID NO:6, и идентичность нуклеотидной последовательности и идентичность аминокислотной последовательности в ORF составили 41,2% и 28,6%, соответственно.
Идентичность нуклеотидной последовательности и идентичность аминокислотной последовательности в ORF между LPLAT5 и LPLAT6 составляют 40,0% и 19,1%, соответственно.
Настоящее изобретение также охватывает нуклеиновые кислоты, функционально эквивалентные нуклеиновым кислотам, которые содержат нуклеотидные последовательности, показанные в SEQ ID NO:1 и 6 выше (в данном описании иногда называемые “нуклеотидными последовательностями настоящего изобретения”) и нуклеотидные последовательности, кодирующие белки, состоящие из аминокислотных последовательностей, показанных в SEQ ID NO:2 и 7 (в данном описании иногда называемые “аминокислотными последовательностями настоящего изобретения”). Выражение “функционально эквивалентный” означает, что белок, кодируемый нуклеотидной последовательностью настоящего изобретения, и белок, состоящий из аминокислотной последовательности настоящего изобретения, обладают “активностью лизофосфолипид-ацилтрансферазы (LPLAT активность)”, “активностью увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую белок настоящего изобретения, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором” (здесь и далее в данном описании иногда называемой “активностью увеличения количественного соотношения арахидоновой кислоты”) и/или “активностью, участвующей в одном или более преобразованиях, выбранных из группы, состоящей из преобразования из 18:1-CoA в 18:1-PL, преобразования из 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразования из DGLA-CoA в DGLA-PL (здесь и далее в данном описании иногда называемой “активностью, участвующей в биосинтетическом пути арахидоновой кислоты”)”. Как правило, это означает, что белки имеют активность сходную с активностью LPLAT5 и/или 6.
“Лизофосфолипид-ацилтрансферазная (LPLAT) активность” настоящего изобретения относится к активности переноса ацильной группы между ацил-CoA и лизофосфолипидом. “Лизофосфолипид” относится к фосфолипиду, в котором удалена одна ацильная группа. В используемом в данном описании значении, лизофосфолипиды включают, но не ограничиваются ими, лизофосфатидную кислоту (LPA), лизофосфатидилхолин (LPC), лизофосфатидилсерин (LPS), лизофосфатидилэтаноламин (LPE), лизофосфатидилинозитол (LPI) и т.д.
LPLAT настоящего изобретения могут специфично воздействовать на один лизофосфолипид или конкретные сложные лизофосфолипиды.
LPLAT активность настоящего изобретения может быть проанализирована известными способами, включая, например, способ, описанный в J.B.C., 282(47), 34288-34298 (2007).
“Активность увеличения количественного соотношения арахидоновой кислоты” настоящего изобретения относится к активности увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту настоящего изобретения, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором, как описано выше. В частности, это относится к активности увеличения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот хозяина, трансформированного рекомбинантным вектором, содержащим нуклеиновую кислоту, которая содержит нуклеотидную последовательность настоящего изобретения, или нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности настоящего изобретения, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором. Активность может быть проанализирована известными способами включающими, например, трансформацию экспрессионного вектора pYE22m, содержащего нуклеотидную последовательность настоящего изобретения или подобное, в рекомбинантного дрожжевого хозяина Saccharomyces cerevisiae, способного продуцировать арахидоновую кислоту при помощи введения и экспрессии гена десатуразы Δ12 жирных кислот, десатуразы Δ6 жирных кислот, гена элонгазы жирных кислот GLELO и гена десатуразы Δ5 жирных кислот; культивирование полученного в результате трансформанта; сбор клеток, выросших в культуре; и подвергание их жирно-кислотному анализу методикой, описанной в приведенных ниже примерах.
“Активность, участвующая в биосинтетическом пути арахидоновой кислоты”, настоящего изобретения относится к активности, участвующей в преобразовании из 18:1-CoA в 18:1-PL, преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и/или преобразовании из DGLA-CoA в DGLA-PL. Как правило, активность относится к активности, участвующей в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA, и/или преобразовании из DGLA-CoA из DGLA-PL. В данном случае, 18:1 - представляет олеоильную группу, 18:3(n-6) - представляет γ-лилнолеильную группу, DGLA - представляет дигомо-γ-лилнолеильную группу, PL представляет фосфолипид, и CoA представляет кофермент A, соответственно. Следовательно, DGLA-CoA относится к ацил-CoA, содержащему дигомо-γ-лилнолеильную группу, и DGLA-PL относится к фосфолипиду, содержащему, например, дигомо-γ-лилнолеильную группу. Активность, участвующая в биосинтетическом пути арахидоновой кислоты, может быть определена путем наблюдения за преобразованием каждого исходного субстрата в произведенный субстрат. Альтернативно, активность может быть определена при наблюдении за тем, что белок настоящего изобретения недостаточно экспрессирован в хозяине или клетке, трансформированной рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую белок настоящего изобретения, или экспрессия белка подавлена в клетке, способной продуцировать арахидоновую кислоту. Например, активность может быть определена путем анализа композиционного соотношения жирных кислот в хозяине или в клетке, сверхэкспрессирующей белок настоящего изобретения, или в хозяине или в клетке с недостаточной экспрессией белка настоящего изобретения, и наблюдения за изменениями в композиционном соотношении жирных кислот для оценки преобразования из каждого исходного субстрата в произведенный субстрат методикой описанной в приведенных ниже примерах.
Более предпочтительно, нуклеотидные последовательности настоящего изобретения или подобные являются нуклеиновыми кислотами, которые содержат нуклеотидную последовательность, кодирующую белок, обладающий LPLAT активностью, активностью увеличения количественного соотношения арахидоновой кислоты и/или активностью, участвующей в биосинтетическом пути арахидоновой кислоты.
Еще более предпочтительно, лизофосфолипид-ацилтрансферазы (LPLAT), кодируемые нуклеиновыми кислотами настоящего изобретения, относятся к ферментам, принадлежащим к семейству мембранно-связанных O-ацилтрансфераз (MBOAT) среди LPLAT.
“Семейство MBOAT” относится к семейству, к которому относится белок с PFAM номером доступа PF03062, и относится к группе ферментов, имеющих трансмембранный домен в аминокислотной последовательности глицерофосфолипид-ацилтрансфераз. PFAM (http://pfam.sanger.ac.uk/) относится к базе данных профилей, полученных выравниваниями белковых семейств, обеспеченными Sanger Institute. Каждый профиль составлен из одинаковых последовательностей и проанализирован скрытой марковской моделью. Семейство белков, к которому принадлежит требуемый белок, может быть исследовано с использованием ключевых слов, последовательностей нуклеиновых кислот, кодирующих белок, аминокислотной последовательности белка, номера доступа и тому подобное, в дополнение к названию представляющего интерес белка. Поиск с использованием последовательностей нуклеиновых кислот, кодирующих LPLAT, полученных по настоящему изобретению, или аминокислотных последовательностей LPLAT выявляет то, что белки принадлежат семейству MBOAT с номером доступа PF03062. Кроме того, ферменты, принадлежащие к семейству MBOAT, в основном имеют сохраненный остаток гистидина в активном центре, такой как, например, остаток гистидина в положении 317 в аминокислотной последовательности LPLAT5, и остаток гистидина в положении 456 в аминокислотной последовательности LPLAT6.
В отличие от LPAAT семейства MBOAT семейство не содержит мотив LPAAT. Мотив LPAAT относится к сохраненному мотиву “HXXXXD (HX4D)”, встречающемуся в четырех сайтах в аминокислотных последовательностях белков LPAAT, описанных в патентном документе 3. Например, мотив LPAAT встречается в аминокислотных остатках 115-120 SEQ ID NO:2 в патентном документе 3 в LPAAT3 и в аминокислотных остатках 115-120 SEQ ID NO:4 в LPAAT4, которые получены из липид-продуцирующего гриба M. alpina, описанного в патентном документе 3. Однако LPLAT белки настоящего изобретения не содержат такой мотив.
В M. alpina до настоящего времени были обнаружены четыре LPLAT (патентные документы 1-3), но не был обнаружен LPLAT фермент, принадлежащий к семейству MBOAT. Таким образом, наиболее предпочтительно LPLAT настоящего изобретения принадлежат к семейству MBOAT и обладают указанной выше активностью настоящего изобретения.
Нуклеиновые кислоты, функционально эквивалентные нуклеиновым кислотам настоящего изобретения, как описано выше, включают нуклеиновую кислоту, которая содержит нуклеотидную последовательность по любому из нижеследующих пунктов (a)-(e). Как использовано при ссылках на нуклеотидные последовательности в данном описании выражение “указанная выше активность настоящего изобретения” относится к “LPLAT активности, активности увеличения количественного соотношения арахидоновой кислоты и/или активности, участвующей в биосинтетическом пути арахидоновой кислоты”, определенным выше.
(a) Нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения.
Нуклеиновая кислота настоящего изобретения содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения. “Указанная выше активность настоящего изобретения” является такой, как определено выше.
В частности, нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой и/или добавлением одной или более (предпочтительно одной или нескольких (например, 1-400, 1-200, 1-100, 1-50, 1-30, 1-25, 1-20, 1-15, более предпочтительно 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1)) аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7; и обладающий указанной выше активностью настоящего изобретения. В данном случае, выражение “аминокислотная последовательность с делецией, заменой и/или добавлением” означает, что одна или более аминокислот удалены, замещены и/или добавлены в одном или нескольких произвольных положениях в одной и той же аминокислотной последовательности. Две или более делеций, замен и/или добавок могут встречаться одновременно, но число делеций, замен и/или добавок, как правило, предпочтительно мало.
В указанных выше модификациях замена предпочтительно является консервативной. Консервативная замена относится к замене определенного аминокислотного остатка другим остатком, обладающим сходными физико-химическими свойствами, и может быть любой заменой, которая по существу не воздействует на структурные свойства первоначальной последовательности, например, это может любая замена, при условии, что замененные аминокислоты не разрушают спираль, присутствующую в первоначальной последовательности, или не разрушают любые другие типы вторичной структуры, характерные для первоначальной последовательности.
Консервативную замену, как правило, вводят синтезом в биологических системах или химическим пептидным синтезом, предпочтительно химическим пептидным синтезом. Заместители могут включать неприродные аминокислотные остатки, а также пептидомиметики, и обращенные или инвертированные формы аминокислотных последовательностей, в которых незамещенные участки реверсированы или инвертированы.
Неограничивающий список групп аминокислотных остатков, которые могут быть заменены друг другом, представлен ниже.
Группа A: лейцин, изолейцин, норлейцин, валин, норвалин, аланин, 2-аминобутановая кислота, метионин, O-метилсерин, т-бутилглицин, трет-бутилаланин и циклогексилаланин;
Группа B: аспарагиновая кислота, глутаминовая кислота, изоаспарагиновая кислота, изоглутаминовая кислота, 2-аминоадипиновая кислота и 2-аминопробковая кислота;
Группа C: аспарагин и глутамин;
Группа D: лизин, аргинин, орнитин, 2,4-диаминобутановая кислота и 2,3-диаминопропионовая кислота;
Группа E: пролин, 3-гидроксипролин и 4-гидроксипролин;
Группа F: серин, треонин и гомосерин; и
Группа G: фенилаланин и тирозин.
Неконсервативная замена может включать замену члена одной из указанных выше групп членом другой группы, в этом случае предпочтительно должны быть учтены индексы гидрофобности аминокислот (аминокислотные индексы гидрофобности) для того, чтобы сохранить биологические функции белков настоящего изобретения (Kyte et al., J. Mol. Biol., 157:105-131(1982)).
Неконсервативная замена также может включать аминокислотную замену на основании гидрофильности.
В описании и чертежах данного описания нуклеотид и аминокислотные названия и аббревиатуры основаны на IUPAC-IUB Commission on Biochemical Nomenclature или на традиционно используемых в данной области методиках, как описано, например, в Immunology--A Synthesis (second edition, edited by E.S. Golub and D.R. Gren, Sinauer Associates, Sunderland, Massachusetts (1991)). Любые оптические изомеры аминокислот, которые могут существовать, относятся к L-изомерам, если не указано иное.
Стереоизомеры указанных выше аминокислот, такие как D-аминокислоты, неприродные аминокислоты, такие как α,α-дизамещеные аминокислоты, N-алкиламинокислоты, молочная кислота и другие неканонические аминокислоты, также могут быть компонентами белков настоящего изобретения.
В данном описании амино-конец белков записан слева, и карбокси-конец белков справа, в соответствии со стандартным использованием и договоренностью в данной области. Подобным образом, 5'-конец одноцепочечных полинуклеотидных последовательностей записан слева и 5'-конец одной из цепей двухцепочечных полинуклеотидных последовательностей обычно записан слева, если специально не указано иное.
Специалист в данной области сможет сконструировать и генерировать подходящие варианты белков, описанных в данном описании, с использованием известных в данной области технологий. Например, он сможет идентифицировать подходящие участки молекулы белка, которые могут быть структурно изменены без нарушения биологической активности белка настоящего изобретения, мечением участков, которые не считаются важными для биологической активности белка настоящего изобретения. Также, специалист сможет идентифицировать консервативные у сходных белков остатки и области. Кроме того, специалист сможет ввести консервативные аминокислотные замены в участки, которые могут быть важными для биологической активности или структуры белка настоящего изобретения, без нарушения биологической активности и без неблагоприятного воздействия на полипептидную структуру белка.
Специалист в данной области может провести так называемые структурно-функциональные исследования, идентифицируя остатки в пептиде, сходном с пептидом белка настоящего изобретения, которые важны для биологической активности или структуры белка настоящего изобретения, и сравнивая аминокислотные остатки в двух пептидах, чтобы предсказать, какие остатки в белке, сходном с белком настоящего изобретения, являются аминокислотными остатками, которые соответствуют аминокислотным остаткам, которые важны для биологической активности или структуры. Дополнительно, специалист может отобрать варианты, которые сохраняют биологическую активность белка настоящего изобретения выбором химически сходных аминокислотных замен для таких предсказанных аминокислотных остатков. Специалист в данной области может также проанализировать трехмерную структуру и аминокислотную последовательность вариантов белка. С точки зрения аналитических результатов, специалист сможет дополнительно предсказать выравнивание аминокислотных остатков с учетом трехмерной структуры белка. На основании аналитических результатов, как описано выше, специалист в данной области также может генерировать варианты, не содержащие изменений аминокислотных остатков, по отношению к которым предсказано расположение на поверхности белка, так как такие остатки могут участвовать в важных взаимодействиях с другими молекулами. Кроме того, специалист в данной области может генерировать варианты, содержащие единичную аминокислотную замену среди аминокислотных остатков, составляющих белок настоящего изобретения. Варианты могут быть отсортированы известными анализами для накопления сведений об отдельных вариантах. В результате специалист может оценить практическую значимость замен отдельных аминокислотных остатков, составляющих белок настоящего изобретения, путем сравнения вариантов, содержащих изменение определенного аминокислотного остатка, чтобы оценить, демонстрируют ли они пониженную биологическую активность по сравнению с биологической активностью белка настоящего изобретения или не демонстрируют такую биологическую активность, или они демонстрируют нежелательную активность, подавляющую биологическую активность белка настоящего изобретения. Кроме того, на основании информации, накопленной из таких стандартных экспериментов, специалист в данной области сможет без труда проанализировать нежелательные аминокислотные замены для вариантов белка настоящего изобретения как отдельно, так и в сочетании с другими мутациями.
Как описано выше, белки, состоящие из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотную последовательность, показанную в SEQ ID NO:2 или 7, могут быть получены такими технологиями, как сайт-направленный мутагенез, описанный в Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001)); “Current Protocols in Molecular Biology” (John Wiley & Sons (1987-1997); Kunkel (1985) Proc. Natl. Acad. Sci. USA 82: 488-92; Kunkel (1988) Method. Enzymol. 85: 2763-6, и т.д. Получение таких вариантов, содержащих аминокислотные делеции, замены или добавки или тому подобное, может быть осуществлено известными методиками, такими как например, способ Кункеля или содержащий двухцепочечные разрывы способ, с использованием набора для введения мутаций, основанного на сайт-направленном мутагенезе, такого как например, набор для мутагенеза QuikChangeTM Site-Directed (Stratagene), GeneTailorTM Site-Directed Mutagenesis System (Invitrogen) или TaKaRa Site-Directed Mutagenesis System (Mutan-K, Mutan-Super Express Km, и т.д.; Takara Bio Inc.).
В дополнение к сайт-направленному мутагенезу, приведенному выше, технологии введения делеций, замен или добавлений одной или более аминокислот в аминокислотные последовательности белков с сохранением, при этом, их активности включают обработку гена мутагеном и селективное расщепление гена для удаления, замены или добавления выбранного нуклеотида с последующим лигированием.
Нуклеиновая кислота настоящего изобретения предпочтительно содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-50 в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения. Если указанная выше активность настоящего изобретения сохраняется, то нет ограничений на число или сайты аминокислотных замен или модификаций в белках настоящего изобретения. Способ определения указанной выше активности настоящего изобретения является таким, как описано выше.
(b) Нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6 и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеиновая кислота настоящего изобретения гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и содержит нуклеотидную последовательность, кодирующую белок, обладающий указанной выше активностью настоящего изобретения. “Приведенная выше активность настоящего изобретения” является такой, как описано выше.
Указанная выше нуклеотидная последовательность может быть получена из библиотеки кДНК и геномной библиотеки или подобного известными технологиями гибридизации, такими как гибридизация колоний, гибридизация бляшек, или блоттингом по Саузерну с использованием зонда, приготовленной из соответствующего фрагмента способом, известным специалисту в данной области.
Подробные методики гибридизации можно найти в “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001); особенно разделы 6-7); “Current Protocols in Molecular Biology” (John Wiley & Sons (1987-1997); особенно разделы 6.3-6.4); “DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.” (Oxford University (1995); особенно раздел 2,10 для условий гибридизации), и т.д.
Жесткость условий гибридизации первоначально определяется условиями гибридизации, более предпочтительно условиями гибридизации и условиями промывки. В используемом в данном описании значении, “жесткие условия” включают умеренно или крайне жесткие условия.
В частности, умеренно жесткие условия включают, например, условия гибридизации 1×SSC-6×SSC при 42°C-55°C, более предпочтительно 1×SSC-3×SSC при 45°C-50°C, наиболее предпочтительно 2×SSC при 50°C. Если гибридизационный раствор содержит, например, около 50% формамида, то используются температуры на 5-15°C ниже обозначенной ранее температуры. Условия промывки предполагают 0,5×SSC-6×SSC при 40°C-60°C. Во время гибридизации и промывки может быть добавлено, как правило, 0,05%-0,2%, предпочтительно около 0,1% SDS.
Крайне жесткие (очень жесткие) условия включают гибридизацию и/или промывку при повышенных температурах и/или пониженных концентрациях солей, по сравнению с умеренно жесткими условиями. Например, условия гибридизации предполагают 0,1×SSC-2×SSC при 55°C-65°C, более предпочтительно 0,1×SSC-1×SSC при 60°C-65°C, наиболее предпочтительно 0,2×SSC при 63°C. Условия промывки предполагают 0,2×SSC-2×SSC при 50°C-68°C, более предпочтительно 0,2×SSC при 60-65°C.
Условия гибридизации, конкретно используемые в настоящем изобретении, включают, например, но не ограничиваются ими, предварительную гибридизацию в условиях 5×SSC, 1% SDS, 50 мМ Tris-HCl (pH 7,5) и 50% формамида при 42°C, с последующей гибридизацией с зондами при 42°C в течение ночи и затем трехкратной промывкой в условиях 0,2×SSC, 0,1% SDS при 65°C в течение 20 минут.
Также могут быть использованы коммерчески доступные наборы для гибридизации без использования радиоактивного зонда. В частности, гибридизация может быть выполнена с использованием набора для детектирования нуклеиновых кислот DIG (Roche Diagnostics) или системы ECL для прямого мечения и детектирования (Amersham) и т.д.
Нуклеиновая кислота, включенная в настоящее изобретение, предпочтительно гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и содержит нуклеотидную последовательность, кодирующую белок, обладающий указанной выше активностью настоящего изобретения.
(c) Нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 80% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирующую белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеиновая кислота настоящего изобретения содержит нуклеотидную последовательность, обладающую по меньшей мере 80% идентичностью с нуклеотидной последовательностью, показанной в SEQ ID NO:1 или 6, и кодирующую белок, обладающий указанной выше активностью настоящего изобретения. “Приведенная выше активность настоящего изобретения” является такой, как описано выше.
Предпочтительно, нуклеиновая кислота содержит нуклеотидную последовательность, обладающую по меньшей мере 80%, более предпочтительно 85%, еще более предпочтительно 90% (например, 92% или более, еще более предпочтительно 95% или более, даже 97%, 98% или 99%) идентичностью с нуклеотидной последовательностью, показанной в SEQ ID NO:1 или 6, и кодирующую белок, обладающий указанной выше активностью настоящего изобретения.
Процент идентичности между двумя последовательностями нуклеиновых кислот может быть установлен визуальным контролем и математическим расчетом или предпочтительно сравнением информации о последовательности двух нуклеиновых кислот с использованием компьютерной программы. Компьютерные программы сравнения последовательностей включают, например, программу BLASTN (Altschul et al. (1990) J. Mol. Biol. 215: 403-10) версия 2.2.7, доступную на интернет-сайте U.S. National Library of Medicine: http://www.ncbi.nlm.nih.gov/blast/bl2seq/bls.htмл, или WU-BLAST 2.0 algorithm и т.д. Установки стандартных параметров для WU-BLAST 2.0 доступны на следующем интернет-сайте: http://blast.wustl.edu.
(d) Нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения.
Нуклеиновая кислота настоящего изобретения содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения. “Приведенная выше активность настоящего изобретения” является такой, как описано выше.
В частности, аминокислотная последовательность обладает 80% или более, предпочтительно 85% или более, более предпочтительно 90%, еще более предпочтительно 95% или более, даже более предпочтительно 97% (например, 98%, даже 99%) или более идентичностью с аминокислотной последовательностью SEQ ID NO:2 или 7 или подобной.
Нуклеиновая кислота настоящего изобретения предпочтительно содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения. Более предпочтительно, нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 98% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения.
Процент идентичности между двумя аминокислотными последовательностями может быть установлен визуальным контролем и математическим расчетом. Альтернативно, процент идентичности может быть установлен с использованием компьютерной программы. Такие компьютерные программы включают, например, BLAST, FAST (Altschul et al., J. Mol. Biol., 215:403-410 (1990)) и ClustalW и т.д. В частности, различные условия (параметры) для поиска идентичности программой BLAST описаны Altschul et al. (Nucl. Acids. Res., 25, p.3389-3402, 1997) и находятся в открытом доступе на интернет-сайте National Center for Biotechnology Information (NCBI) или DNA Data Bank of Japan (DDBJ) (BLAST Manual, Altschul et al., NCB/NLM/NIH Bethesda, MD 20894; Altschul et al.). Процент идентичности также может быть установлен при использовании программ для обработки генетических сведений, таких как GENETYX Ver.7 (Genetyx), DNASIS Pro (Hitachisoft), Vector NTI (Infomax) и т.д.
Некоторые схемы выравнивания для выравнивания аминокислотных последовательностей могут привести к сопоставлению даже специфических коротких областей последовательностей, и таким образом возможно детектировать область с очень высокой идентичностью последовательности в таком малом участке выравнивания, даже если отсутствует существенное сходство между полными длинами используемых последовательностей. Кроме того, BLAST алгоритм может использовать BLOSUM62 матрицу аминокислотных замен и по желанию следующие параметры: (A) включение фильтра для маскировки сегментов сомнительной последовательности, которая имеет низкий уровень композиционной сложности (как определено SEG program of Wootton and Federhen (Computers и Chemistry, 1993); также см. Wootton и Federhen, 1996, “Analysis of compositionally biased regions in sequence databases,” Methods Enzymol., 266: 554-71) или сегментов, состоящих из коротких периодичных внутренних повторов (как определено XNU program of Claverie and States (Computers and Chemistry, 1993)), и (B) статистическая значимость порога для сообщения о сопоставлениях по сравнению с базой данных последовательностей или E-оценка (ожидаемая вероятность того, что сопоставления будут обнаружены только в пределах шансов, соответствующих вероятностной модели Karlin и Altschul, 1990; если статистическая значимость, приписываемая сопоставлению, больше порога E-оценки, то о сопоставлении не будет сообщено).
(e) Нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая содержит нуклеотидную последовательность, кодирующую белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеиновая кислота настоящего изобретения гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и содержит нуклеотидную последовательность, кодирующую белок, обладающий указанной выше активностью настоящего изобретения.
“Приведенная выше активность настоящего изобретения” и условия гибридизация являются такими, как описано выше.
Дополнительно, нуклеиновые кислоты настоящего изобретения также включают нуклеиновую кислоту, которая содержит нуклеотидную последовательность с делецией, заменой или добавлением одного или более нуклеотидов в нуклеотидную последовательность, содержащую SEQ ID NO:1 или 6, и кодирующую белок, обладающий указанной выше активностью настоящего изобретения. В частности, также возможно использовать нуклеиновую кислоту, которая содержит нуклеотидную последовательность с делецией, заменой или добавлением одного или более (предпочтительно одного или нескольких (например, 1-1500, 1-1000, 1-500, 1-300, 1-250, 1-200, 1-150, 1-100, 1-50, 1-30, 1-25, 1-20, 1-15, более предпочтительно 10, 9, 8, 7, 6, 5, 4, 3, 2, или 1)) нуклеотидов в нуклеотидную последовательность, показанную в SEQ ID NO:1 или 6, и кодирующую белок, обладающий указанной выше активностью настоящего изобретения. В используемом в данном описании значении, выражение “нуклеотидная последовательность с делецией, заменой или добавкой” означает, что один или более нуклеотидов удалены, заменены и/или добавлены в одном или нескольких произвольных положениях в одной и той же нуклеотидной последовательности. Две или более делеций, замен и/или добавок могут встречаться одновременно, но, как правило, число делеций, замен и/или добавок предпочтительно мало.
Предпочтительные варианты нуклеиновых кислот настоящего изобретения также включают нуклеиновую кислоту по любому из нижеследующих пунктов (a)-(d):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:1 или 6, или ее частичную последовательность;
(b) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность, показанную в SEQ ID NO:2 или 7, или ее частичную последовательность;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:4 или 9, или ее частичную последовательность;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:5 или 10, или ее частичную последовательность.
Нуклеиновые кислоты, определенные как (a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:1 или 6; (b) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7; и (c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, показанную в SEQ ID NO:4 или 9, являются такими, как описано выше. Частичная последовательность указанных выше последовательностей представляет собой области, содержащиеся в указанных выше нуклеотидных последовательностях включая ORF, CDS, биологически активные области, области, используемые в качестве праймеров, как описано ниже, и области, которые могут быть зондами, и могут быть природного происхождения или искусственно полученными.
Нуклеиновые кислоты настоящего изобретения предпочтительно являются нуклеиновыми кислотами, кодирующими белок, принадлежащий к семейству мембранно-связанных O-ацилтрансфераз. “Семейство мембранно-связанных O-ацилтрансфераз” является таким, как описано выше.
Нуклеиновые кислоты настоящего изобретения также включают:
(1) нуклеиновую кислоту по любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую 80% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7;
(e) нуклеиновая кислота, которая гибридизуется в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7; и
(2) нуклеиновую кислоту по пункту (1), которая соответствует любому из нижеследующих пунктов (a)-(e):
(a) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-50 в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6;
(c) нуклеиновая кислота, которая содержит нуклеотидную последовательность, обладающую идентичностью 90% или более с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6;
(d) нуклеиновая кислота, которая содержит нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, обладающей идентичностью 90% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7;
(e) нуклеиновая кислота, которая гибридизуется в условиях 2×SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7.
Лизофосфолипид-ацилтрансферазные белки настоящего изобретения
Белки настоящего изобретения характеризуются тем, что они обладают “активностью лизофосфолипид-ацилтрансферазы (LPLAT активность)”, “активностью увеличения количественного соотношения арахидоновой кислоты” и/или “активностью, участвующей в биосинтетическом пути арахидоновой кислоты”. Белки настоящего изобретения могут быть природного происхождения или искусственно полученными.
Белки настоящего изобретения предпочтительно являются LPLAT5 и LPLAT6, состоящими из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7. Дополнительно, настоящее изобретение также охватывает варианты LPLAT5 и LPLAT6, т.е. варианты, удовлетворяющие критериям: обладают “активностью лизофосфолипид-ацилтрансферазы (LPLAT активность)”, “активностью увеличения количественного соотношения арахидоновой кислоты” и/или “активностью, участвующей в биосинтетическом пути арахидоновой кислоты”.
“Активность лизофосфолипид-ацилтрансферазы”, “активность увеличения количественного соотношения арахидоновой кислоты” и “активность, участвующая в биосинтетическом пути арахидоновой кислоты” являются такими, как описано выше в разделе “Нуклеиновые кислоты, кодирующие лизофосфолипид-ацилтрансферазы настоящего изобретения”. В используемом в данном описании значении, “приведенная выше активность настоящего изобретения” далее относится к “LPLAT активности, активности увеличения количественного соотношения арахидоновой кислоты и/или активности, участвующей в биосинтетическом пути арахидоновой кислоты”, определенным выше.
Белки настоящего изобретения включают белок по нижеследующему пункту (a) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения;
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7, и обладающий указанной выше активностью настоящего изобретения.
Определения “аминокислотная последовательность с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности” и “идентичность 80% или более” имеют значения, описанные выше в разделе “Нуклеиновые кислоты, кодирующие лизофосфолипид-ацилтрансферазы настоящего изобретения”.
Белки настоящего изобретения также включают вариант белка, кодируемый нуклеиновой кислотой, содержащей нуклеотидную последовательность SEQ ID NO:1 или 6, или белок аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, или модифицированный иным способом, или модифицированный белок, имеющий модифицированную боковую аминокислотную цепь, или являющийся белком слияния с другим белком, и обладающий указанной выше активностью настоящего изобретения.
Белки настоящего изобретения могут быть искусственно получены методами химического синтеза, такими как способ Fmoc (флуоренилметилоксикарбонильный способ) и способ tBoc (трет-бутилоксикарбонильный способ). Они также могут быть химически синтезированы с использованием пептидного синтезатора, доступного от Advanced ChemTech, Perkin Elmer, Pharmacia, Protein Technology Instrument, Synthecell-Vega, PerSeptive, Shimadzu Corporation или подобных.
Кроме того, белки настоящего изобретения предпочтительно являются белками, принадлежащими к семейству мембранно-связанных O-ацилтрансфераз. Определение “семейство мембранно-связанных O-ацилтрансфераз” или тому подобное является таким, как разъяснено выше в разделе “Нуклеиновые кислоты, кодирующие лизофосфолипид-ацилтрансферазы настоящего изобретения”.
Белки настоящего изобретения также включают:
(1) белок по нижеследующему пункту (a) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или более аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7;
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 80% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7;
(2) белок по пункту (1), который соответствует нижеследующему пункту (a) или (b):
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-50 в аминокислотной последовательности SEQ ID NO:2 или 7;
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 90% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
Клонирование нуклеиновых кислот настоящего изобретения
Нуклеиновые кислоты, кодирующие LPLAT белки настоящего изобретения, могут быть клонированы, например, с помощью скрининга из библиотеки кДНК с использованием соответствующих зондов. Они также могут быть клонированы амплификацией с использованием ПЦР с соответствующими праймерами с последующим лигированием в соответствующий вектор. Полученный таким образом клон может быть дополнительно субклонирован в другой вектор.
Например, могут быть использованы коммерчески доступные плазмидные векторы, такие как pBlue-ScriptTM SK (+) (Stratagene), pGEM-T (Promega), pAmp (TM: Gibco-BRL), p-Direct (Clontech) и pCR2.1-TOPO (Invitrogen). Для амплификации с использованием ПЦР, любые участки нуклеотидной последовательности, показанной в SEQ ID NO:1 или 6 и подобной, могут быть использованы в качестве праймеров, включая, например, праймеры, показанные в примере 1 ниже. ПЦР осуществляют добавлением указанных выше праймеров и термоустойчивой ДНК полимеразы или тому подобное, для воздействия на кДНК, полученную из клеток M. alpina. Указанная выше методика может быть без труда выполнена специалистом в данной области в соответствии с “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001)) или тому подобное.
Полученный таким образом ПЦР продукт может быть очищен с использованием известных способов. Например, способы очистки включают использование наборов, таких как GENECLEAN (Funakoshi Co., Ltd.), QIAquick PCR purification (QIAGEN), ExoSAP-IT (GE Healthcare Bio-Sciences); или использование целлюлозных фильтров DEAE или диализных трубок и т.д. Если используют агарозный гель, то фрагменты ДНК подвергают электрофорезу в агарозном геле и фрагменты ДНК вырезают из агарозного геля, с последующей очисткой наборами GENECLEAN (Funakoshi Co., Ltd.), QIAquick Gel extraction (QIAGEN), способом замораживания-сдавливания и т.д.
Нуклеотидные последовательности клонированных нуклеиновых кислот могут быть установлены c использованием нуклеотидного секвенатора.
Конструирование экспрессионных векторов настоящего изобретения и получение трансформированных клеток
Настоящее изобретение также обеспечивает рекомбинантные векторы, содержащие нуклеиновую кислоту, кодирующую LPLAT белок настоящего изобретения. Настоящее изобретение дополнительно обеспечивает клетки, трансформированные рекомбинантными векторами.
Такие рекомбинантные векторы и трансформанты могут быть получены следующим образом. Плазмиду, несущую нуклеиновую кислоту, кодирующую LPLAT белок настоящего изобретения, обрабатывают эндонуклеазами рестрикции. Используемые эндонуклеазы рестрикции включают, например, но без ограничения, EcoRI, KpnI, BamHI и SalI, и т.д. Концы плазмиды могут быть затуплены обработкой T4 полимеразой. Обработанный фрагмент ДНК очищают электрофорезом в агарозном геле. Полученный фрагмент ДНК может быть встроен в экспрессионный вектор известным способом, посредством чего получают вектор для экспрессии LPLAT белка. Полученный экспрессионный вектор трансформируют в хозяина для получения трансформанта, который используется для экспрессии требуемого белка.
Экспрессионный вектор и хозяин в данном описании не ограничены особым образом, при условии, что требуемый белок может быть экспрессирован, и подходящие хозяева включают, например, грибы, бактерии, растения и животных или их клетки. Грибы включают волокнистые грибы, такие как липид-продуцирующий гриб M. alpina, дрожжи, такие как S. cerevisiae (Saccharomyces cerevisiae) и т.д. Бактерии включают Escherichi coli, Bacillus subtilis и т.д. Дополнительно, растения включают продуцирующие масло растения, такие как рапс, соевые бобы, хлопчатник, сафлор и лен.
Липид-продуцирующие грибы, которые могут быть использованы, включают, например, штаммы, описанные в MYCOTAXON, Vol. XLIV, NO. 2, pp. 257-265 (1992), особенно микроорганизмы, принадлежащие к роду Mortierella, включая микроорганизмы, принадлежащие к подроду Mortierella, такие как Mortierella elongata (M. elongata) IFO8570, Mortierella exigua (M. exigua) IFO8571, Mortierella hygrophila (M. hygrophila) IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS 219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70, CBS754.68, или микроорганизмы, принадлежащие к подроду Micromucor, такие как Mortierella isabellina (M. isabellina) CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308, IFO7884, Mortierella nana (M. nana) IFO8190, Mortierella ramanniana (M. ramanniana) IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185, IFO8287, Mortierella vinacea (M. vinacea) CBS236.82. Среди прочих предпочтителен M. alpin.
При использовании в качестве хозяина гриба вектор предпочтительно имеет структуру, которая позволяет нуклеиновой кислоте настоящего изобретения самореплицироваться в хозяине или встраиваться в хромосому гриба. Также, вектор предпочтительно содержит промотор и терминатор. При использовании в качестве хозяина M. alpina экспрессионным вектором может быть, например, pD4, pDuraSC, pDura5 или подобные. Может быть использован любой промотор, который может быть экспрессирован в хозяине, включая полученные из M. alpina промоторы, такие как промотор гена гистона H4.1, промотор гена GAPDH (глицеральдегид-3-фосфат-дегидрогеназа) и промотор гена TEF (фактор элонгации трансляции).
Технологии трансформирования рекомбинантного вектора в волокнистые грибы, такие как M. alpin, включают электропорацию, сферопластный способ, доставку частиц и непосредственную микроинъекцию ДНК в ядро, и т.д. При использовании штамма ауксотрофного хозяина трансформированные штаммы могут быть получены отбором штаммов, растущих на селективной среде без незаменимых для организма питательных веществ. При использовании для трансформации маркерного гена устойчивости к лекарственному средству могут быть получены культивированием в селективной среде, содержащей лекарственное средство, колонии клеток, демонстрирующие устойчивость к лекарственному средству.
При использовании дрожжей в качестве хозяина, экспрессионным вектором может быть, например, pYE22m или тому подобное. Также могут быть использованы коммерчески доступные дрожжевые экспрессионные векторы, такие как pYES (Invitrogen) и pESC (STRATAGENE). Дрожжевые хозяева, подходящие для настоящего изобретения, включают, но не ограничиваются ими, S. cerevisiae штамма EH13-15 (trp1, MATα) и т.д. Используемые промоторы включают, например, полученные из дрожжей или подобные, такие как промотор GAPDH, промотор GAL1 и промотор GAL10.
Технологии трансформирования рекомбинантного вектора в дрожжи включают, например, литийацетатный способ, электропорацию, сферопластный способ, декстран-опосредованную трансфекцию, кальцийфосфатную преципитацию, полибрен-опосредованную трансфекцию, слияние протопластов, инкапсуляцию (одного или нескольких) полинуклеотида (ов) в липосомы и непосредственную микроинъекцию ДНК в ядро, и т.д.
При использовании в качестве хозяина бактерии, такой как E. coli, экспрессионным вектором может быть, например, pGEX, pUC18 или подобные, доступные от Pharmacia. Промоторы, которые могут быть использованы, включают полученные из E. coli, бактериофаги и подобные, такие как, например, trp промотор, lac промотор, PL промотор и PR промотор. Технологии трансформирования рекомбинантного вектора в бактерии включают, например, электропорацию и кальцийхлоридный способ.
Способы получения композиций жирных кислот настоящего изобретения
Настоящее изобретение обеспечивает способы получения композиции жирных кислот из трансформированных клеток, описанных выше, т.е. способы получения композиции жирных кислот из культур трансформированной клетки. В частности, она может быть получена по методике, описанной ниже. Однако способы настоящего изобретения не ограничиваются приведенными ниже методиками, а также могут быть осуществлены с использованием других общеизвестных методик.
Любая жидкая среда (культуральная среда) может быть использована для культивирования организма, экспрессирующего белок настоящего изобретения, при условии, что она имеет соответствующие pH и осмотическое давление и содержит питательные вещества, требуемые для роста каждого хозяина, следовые элементы и биологические вещества, такие как сыворотка или антибиотики. Например, культуральные среды, которые могут быть использованы для дрожжевых клеток, трансформированных для экспрессии LPLAT 5 и 6, включают, но без ограничения, SC-Trp, Leu, Ura среду, YPD среду, YPD5 среду и подобные.
Могут быть использованы любые культуральные условия, подходящие для роста хозяина и для устойчивого поддержания уровня генерированного фермента, и в частности, могут быть установлены отдельные условия, включая анаэробность, инкубационный период, температуру, влажность, статичное культивирование или культивирование при встряхивании и т.д. Культивирование может быть осуществлено в одних и тех же условиях (одностадийное культивирование) или может быть, так называемым двухстадийным или трехстадийным культивированием с использованием двух или более различных культуральных условий, но двухстадийное культивирование и подобные предпочтительны для большого масштаба культивирования из-за высокой эффективности культивирования.
Композиции жирных кислот настоящего изобретения
Настоящее изобретение также предоставляет композиции жирных кислот, содержащие совокупность одной или более жирных кислот в клетке, экспрессирующей LPLAT белок настоящего изобретения, характеризующиеся тем, что количественное соотношение арахидоновой кислоты в композиционном соотношении жирных кислот в композиции жирных кислот выше, чем количественное соотношение арахидоновой кислоты в композиции жирных кислот, полученных культивированием нетрансформированных хозяев. Предпочтительно настоящее изобретение предоставляет композиции жирных кислот, полученные культивированием трансформированных клеток, экспрессирующих LPLAT5 и 6 настоящего изобретения. В приведенных ниже примерах, количественное соотношение арахидоновой кислоты в дрожжах, продуцирующих арахидоновую кислоту, трансформированных LPLAT5 или 6, повышено по меньшей мере в 1,5 раза по сравнению с количественным соотношением арахидоновой кислоты в контрольной композиции жирных кислот.
Жирные кислоты могут быть в виде свободных жирных кислот или в виде сложных триглицеридов, фосфолипидов или подобных.
Жирные кислоты, содержащиеся в композиции жирных кислот настоящего изобретения, являются линейными или разветвленными монокарбоновыми кислотами с длинными углеводными цепями, включая, например, но, не ограничиваясь ими, миристиновую кислоту (тетрадекановая кислота) (14:0), миристоленовую кислоту (тетрадеценовая кислота) (14:1), пальмитиновую кислоту (гексадекановая кислота) (16:0), пальмитолиновую кислоту (9-гексадеценовая кислота) (16:1), стеариновую кислоту (октадекановая кислота) (18:0), олеиновую кислоту (цис-9-октадеценовая кислота) (18:1 (9) или иногда называемая просто 18:1), вакценовую кислоту (11-октадеценовая кислота) (18:1 (11)), линолевую кислоту (цис, цис-9,12-октадекадиеновая кислота) (18:2 (9,12) или иногда называемая просто 18:2), α-линоленовую кислоту (9,12,15-октадекатриеновая кислота) (18:3 (9,12,15)), γ-линоленовую кислоту (6,9,12-октадекатриеновая кислота) (18:3 (6,9,12), GLA или иногда называемая 18:3(n-6)), стеаридоновую кислоту (6,9,12,15-октадекатетраеновая кислота) (18:4 (6,9,12,15)), арахиновую кислоту (эйкозановая кислота) (20:0), (8,11-эйкозадиеновая кислота) (20:2 (8,11)), медовую кислоту (5,8,11-эйкозатриеновая кислота) (20:3 (5,8,11)), дигомо-γ-линоленовую кислоту (8,11,14-эйкозатриеновая кислота) (20:3 (8,11,14) или иногда называемая DGLA), арахидоновую кислоту (5,8,11,14-эйкозатетраеновая кислота) (20:4 (5,8,11,14) или иногда называемая ALA), эйкозатетраеновую кислоту (8,11,14,17-эйкозатетраеновая кислота) (20:4 (8,11,14,17)), эйкозапентаеновую кислоту (5,8,11,14,17-эйкозапентаеновая кислота) (20:5 (5,8,11,14,17)), бегеновую кислоту (докозановая кислота) (22:0), (7,10,13,16-докозатетраеновая кислота) (22:4 (7,10,13,16)), (7,10,13,16,19-докозапентаеновая кислота) (22:5 (7,10,13,16,19)), (4,7,10,13,16-докозапентаеновая кислота) (22:5 (4,7,10,13,16)), (4,7,10,13,16,19-докозагексаеновая кислота) (22:6 (4,7,10,13,16,19)), лигноцериновую кислоту (тетрадокозановая кислота) (24:0), ацетэруковую кислоту (цис-15-тетрадокозановая кислота) (24:1), церотиновую кислоту (гексадокозановая кислота) (26:0) и т.д. Химические названия, представленные выше, являются общепринятыми названиями, получившими определение в соответствии с IUPAC Biochemical Nomenclature, и после каждого следует в круглых скобках систематическое название и затем количество атомов углерода и число и положения двойных связей.
Композиция жирных кислот настоящего изобретения может быть составлена из любого числа и любого типа жирных кислот, при условии, что она содержит комбинацию одной или более жирных кислот, перечисленных выше.
Лиофилизированные клетки, полученные способами получения композиций жирных кислот настоящего изобретения, описанными выше, встряхивают со смесью хлороформ/метанол, приготовленной в подходящем соотношении, и затем нагревают в течение подходящего периода времени. Дополнительно разделение клеток центрифугированием и замену растворителя повторяют несколько раз. Затем липиды сушат подходящим способом и затем растворяют в растворителе, таком как хлороформ. Собирают аликвоту этого образца, и жирные кислоты в клетках преобразуют в метиловые эфиры с использованием метанольной HCl, затем экстрагируют гексаном, отгоняют гексан, и остаток анализируют газовой хроматографией.
Количественное соотношение арахидоновой кислоты в композиционном соотношении жирных кислот композиции жирных кислот, полученной культивированием клетки, трансформированной рекомбинантным вектором, содержащим нуклеиновую кислоту настоящего изобретения выше, чем количественное соотношение арахидоновой кислоты в известных LPLAT композициях жирных кислот. Это объясняется тем фактом, что LPLAT настоящего изобретения могут увеличить преобразование жирных кислот, для которого требуется ацильный перенос от ацил-CoA к фосфолипидам или от фосфолипидов к CoA. В частности, количественное соотношение арахидоновой кислоты в композиции жирных кислот, продуцируемых дрожжами, продуцирующими арахидоновую кислоту (S. cerevisiae), экспрессируя LPLAT5 и LPLAT6, в соответствии с предпочтительными вариантами осуществления настоящего изобретения увеличивается, как дополнительно описано в приведенных ниже примерах. В данном случае, было обнаружено, что LPLAT5 участвует в преобразовании из 18:1-CoA в 18:1-PL, преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразовании из DGLA-CoA в DGLA-PL, и что LPLAT6 участвует в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразовании из DGLA-CoA в DGLA-PL.
Как описано в приведенных ниже примерах, также обнаружено, что LPLAT6, участвует в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразовании из DGLA-CoA в DGLA-PL в M. alpina.
Следовательно, настоящее изобретение также обеспечивает способ применения рекомбинантного вектора для повышения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном вектором, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
Пищевые или другие продукты, содержащие композиции жирных кислот настоящего изобретения
Настоящее изобретение также предоставляет пищевые продукты, содержащие указанные выше композиции жирных кислот. Композиции жирных кислот настоящего изобретения могут быть использованы на регулярной основе для производства пищевых продуктов и сырья для промышленности, содержащих жиры и масла (сырьевые вещества для косметических средств, лекарственные препараты (например, лекарственные средства для проблемной кожи), мыла и т.д.) или для других целей. Косметические средства (композиции) или лекарственные препараты (композиции) могут быть представлены в любой форме, включая, но, не ограничиваясь ими, раствор, мазь, гель, твердое вещество, порошок или подобные. Пищевые продукты могут также быть представлены в форме фармацевтической рецептуры, такой как капсула, или продовольствия, подвергшегося технологической обработке, такого как натуральное жидкое питание, бесшлаковое питание, питание элементными смесями, питательный напиток или энтеральное питание, при этом формула содержит композицию жирных кислот настоящего изобретения в комбинации с белками, сахарами, жирами, следовыми элементами, витаминами, эмульгаторами, ароматизирующими веществами и т.д.
Другие примеры пищевых продуктов настоящего изобретения включают, но не ограничиваются ими, диетические добавки, продукты здорового питания, функциональные продукты питания, питание для детей, модифицированное молоко для младенцев, модифицированное молоко для недоношенных младенцев, гериатрическое питание и т.д. Выражение «пищевые продукты» в используемом в данном описании обобщающем значении относится к съедобным продуктам в форме твердого вещества, флюида, жидкости или смеси таковых.
Выражение «диетические добавки» относится к пищевым продуктам, обогащенным специфическими питательными ингредиентами. Выражение «продукты здорового питания» относится к пищевым продуктам, которые, как известно, являются полезными или благотворно влияющими на здоровье, и включают диетические добавки, природные пищевые продукты, диетические пищевые продукты и т.д. Выражение «функциональные продукты питания» относится к пищевым продуктам для доставки питательных ингредиентов, обладающих функциями физиологического регулирования, и эти продукты могут также быть названы пищевыми продуктами для определенного использования с воздействием на здоровье. Выражение «питание для детей» относится к пищевым продуктам, предназначенным для детей до примерно 6 лет. Выражение «гериатрическое питание» относится к пищевым продуктам, обработанным для облегчения переваривания и всасывания по сравнению с необработанными пищевыми продуктами. Выражение «модифицированное молоко для младенцев» относится к модифицированному молоку, предназначенному для детей до примерно года. Выражение «модифицированное молоко для недоношенных младенцев» относится к модифицированному молоку, предназначенному для недоношенных младенцев до примерно 6 месяцев.
Пищевые продукты включают природные пищевые продукты, такие как мясо, рыба, орехи (обработанные с жирами и маслами); пищевые продукты, приготовленные с жирами и маслами, такие как китайские пищевые продукты, китайская лапша, супы; пищевые продукты, использующие жиры и масла в качестве теплоносителей, такие как темпура (жареные во фритюре рыба и овощи), жареные во фритюре пищевые продукты, покрытые панировочными сухарями, жареный фасолевый творог, китайский жареный рис, пончики, Karinto (Японское жареное печенье из теста); пищевые продукты на масляной и жировой основе или пищевые продукты, произведенные с жирами и маслами, такие как сливочное масло, маргарин, майонез, заправка для салата, шоколад, лапша быстрого приготовления, карамель, бисквиты, печенье, кекс, мороженное; и пищевые продукты, политые или покрытые жирами и маслами во время завершения приготовления, такие как рисовые крекеры, жесткие бисквиты, сладкий фасолевый пастообразный хлеб. Тем не менее, пищевые продукты настоящего изобретения не ограничиваются жиро- и маслосодержащими пищевыми продуктами, а также включают сельскохозяйственные пищевые продукты, подвергшиеся технологической обработке, такие как хлеб, лапша, приготовленный рис, сладости (конфеты, жевательные резинки, мармелад, таблетки, Японские сладости), фасолевый творог и произведенные из него продукты; ферментированные пищевые продукты, такие как сакэ (Японские рисовое вино), лекарственный ликер, мирин (сладкий херес для готовки), уксус, соевый соус и мисо (соевая фасолевая паста); продукция животноводства, такая как йогурт, ветчина, бекон и сосиски; подвергшиеся обработке морепродукты, такие как Kamaboko (рыбный брикет), Ageten (жареный во фритюре рыбный брикет) и Hanpen (толстый рыбный брикет); и фруктовые напитки, безалкогольные напитки, спортивные напитки, спиртные напитки, чай и тому подобное.
Способ оценки или выбора штаммов с использованием нуклеиновых кислот, кодирующих LPLAT белки или LPLAT белки настоящего изобретения
Настоящее изобретение также предоставляет способы оценки или выбора липид-продуцирующих штаммов с использованием нуклеиновых кислот, кодирующих LPLAT белки или LPLAT белки настоящего изобретения. Способы более конкретно описаны ниже.
(1) Способы оценки
Один из вариантов осуществления настоящего изобретения заключается в способе оценки липид-продуцирующего штамма с использованием нуклеиновой кислоты, кодирующей LPLAT белок или LPLAT белок настоящего изобретения. Способ оценки настоящего изобретения может содержать оценку тестируемого липид-продуцирующего штамма на указанную выше активность настоящего изобретения с использованием праймера или зонда, сконструированных на основании нуклеотидной последовательности настоящего изобретения. Общие методики для такого способа оценки известны и описаны, например, в WO01/040514 или JPA HEI 8-205900 A. Данный способ оценки коротко пояснен ниже.
Сначала получают геном тестируемого штамма. Могут быть использованы любые известные способы получения, такие как способ Hereford или калийацетатный способ (см., например, Methods in Yeast Genetics, Cold Spring Harbor Laboratory Press, p130 (1990)).
Праймер или зонд конструируют на основании нуклеотидной последовательности настоящего изобретения, предпочтительно SEQ ID NO:1 или 6. Праймер или зонд могут быть сконструированы, исходя из любой области нуклеотидной последовательности настоящего изобретения, с использованием известных методик. Число нуклеотидов в полинуклеотиде, используемом в качестве праймера, составляет, как правило, 10 или более, предпочтительно 15-25. Как правило, число нуклеотидов, соответствующее области, которая должна быть фланкирована праймерами, обычно составляет 300-2000.
Праймер или зонд, полученные выше, используют для оценки того, содержит или нет геном указанного выше тестируемого штамма последовательность, характерную для нуклеотидной последовательности настоящего изобретения. Последовательность, характерная для нуклеотидной последовательности настоящего изобретения, может быть обнаружена при использовании известных методик. Например, полинуклеотид, содержащий часть или всю последовательность, специфическую для нуклеотидной последовательности настоящего изобретения, или полинуклеотид, содержащий нуклеотидную последовательность, комплементарную указанной выше нуклеотидной последовательности, используют в качестве одного праймера, и полинуклеотид, содержащий часть или всю последовательность, расположенную перед или расположенную после этой последовательности, или полинуклеотид, содержащий нуклеотидную последовательность, комплементарную указанной выше нуклеотидной последовательности, используют в качестве другого праймера для амплификации нуклеиновой кислоты тестируемого штамма с помощью ПЦР или подобного, устанавливая, таким образом, присутствие или отсутствие амплифицированного продукта, молекулярную массу амплифицированного продукта и т.д.
Условия ПЦР, подходящие для способа настоящего изобретения, особым образом не ограничиваются. Полученный в результате реакции продукт, т.е. амплифицированный продукт, может быть разделен электрофорезом в агарозном геле или подобным для определения молекулярной массы амплифицированного продукта. Таким образом, может быть предсказана или оценена указанная выше активность настоящего изобретения в тестируемом штамме путем определения того, достаточна или нет молекулярная масса амплифицированного продукта для молекулы нуклеиновой кислоты, соответствующей области, характерной для нуклеотидной последовательности настоящего изобретения. Кроме того, указанная выше активность настоящего изобретения может быть более точно предсказана или оценена анализом нуклеотидной последовательности амплифицированного продукта способом, описанным выше или подобным. Способ оценки указанной выше активности настоящего изобретения является таким, как описано выше.
Альтернативно, способ оценки настоящего изобретения может включать культивирование тестируемого штамма и установление уровня экспрессии LPLAT белка, кодируемого нуклеотидной последовательностью настоящего изобретения, такой как SEQ ID NO:1 или 6, оценивая, таким образом, тестируемый штамм на указанную выше активность настоящего изобретения. Уровень экспрессии LPLAT белка может быть установлен культивированием тестируемого штамма в соответствующих условиях и количественным определением мРНК LPLAT белка или данного белка. Определение количества мРНК или белка может быть выполнено с использованием известных методик. Количественный анализ мРНК может быть выполнен, например, нозерн-гибридизацией или количественной ПЦР в реальном времени, тогда как количественный анализ белка может быть выполнен, например, вестерн-блоттингом (Current Protocols in Molecular Biology, John Wiley & 1994-2003).
(2) Способы отбора
Другой вариант осуществления настоящего изобретения заключается в способе отбора липид-продуцирующего штамма с использованием нуклеиновой кислоты, кодирующей LPLAT белок или LPLAT белок настоящего изобретения. Способ отбора по настоящему изобретению может включать культивирование тестируемых штаммов и определение уровня экспрессии LPLAT белка, кодируемого нуклеотидной последовательностью настоящего изобретения, такой как SEQ ID NO:1 или 6, для выбора штамма, имеющего требуемый уровень экспрессии, тем самым, может быть выбран штамм, обладающий требуемой активностью. Альтернативно, способ может включать заранее определенный конкретный штамм, отдельное культивирование конкретного штамма и тестируемых штаммов, установление указанного выше уровня экспрессии в каждом штамме и сравнение уровней экспрессии между конкретным штаммом и каждым тестируемым штаммом, тем самым, может быть выбран требуемый штамм. В частности, штамм, обладающий требуемой активностью, может быть выбран культивированием конкретного штамма и тестируемых штаммов в соответствующих условиях, установлением уровня экспрессии в каждом штамме и выбором тестируемого штамма, демонстрирующего, например, более высокий или более низкий уровень экспрессии, чем уровень экспрессии конкретного штамма. Требуемая активность может быть определена установлением уровня экспрессии LPLAT белка, как описано выше.
Альтернативно, способ отбора настоящего изобретения может включать культивирование тестируемых штаммов и отбор штамма, демонстрирующего более высокий или более низкий уровень указанной выше активности настоящего изобретения, тем самым, может быть выбран штамм, обладающий требуемой активностью. Требуемая активность может быть определена установлением уровня экспрессии LPLAT белка, как описано выше.
Примеры тестируемых штаммов или конкретных штаммов, которые могут быть использованы, например, включают, но не ограничиваются ими, штамм, трансформированный указанным выше вектором настоящего изобретения, штамм с подавленной экспрессией указанных выше нуклеиновых кислот настоящего изобретения, мутагенизированный штамм, мутированный естественным образом штамм и т.д. Технологии мутагенеза включают, но не ограничивается ими, физические способы, такие как УФ или радиоактивное облучение, и химические способы, такие как химические методы обработки EMS (этилметансульфонат), N-метил-N-нитрозогуанидином или подобным (см., например, Yasuji Oshima ed., Biochemistry Experiments vol. 39, Experimental Protocols for Yeast Molecular Genetics, pp. 67-75, Japan Scientific Societies Press).
Штаммы, используемые в качестве конкретных и тестируемых штаммов настоящего изобретения, включают, но не ограничиваются ими, липид-продуцирующие грибы или дрожжи, перечисленные выше. В частности, конкретные и тестируемые штаммы могут быть комбинацией любых штаммов, принадлежащих к различным родам или видам, и один или более тестируемых штаммов могут быть использованы одновременно.
Следующие примеры дополнительно иллюстрируют настоящее изобретение. Тем не менее, следует понимать, что настоящее изобретение не ограничивается приведенными ниже примерами.
ПРИМЕРЫ
[Пример 1]
Геномный анализ M. alpina
M. alpina штамма 1S-4 инокулировали в 100 мл среды GY2:1 (2% глюкозы, 1% дрожжевого экстракта, pH 6,0) и культивировали при встряхивании в течение 2 дней при 28°C. Клетки собирали фильтрованием, с получением геномной ДНК, используя DNeasy (QIAGEN).
Нуклеотидную последовательность геномной ДНК определяли, используя Roche454GS FLX Standard. Это подразумевало две серии секвенирования последовательности фрагмента библиотеки и три серии сопряженного парного секвенирования библиотеки. Полученные в результате нуклеотидные последовательности составляли в 300 суперконтигов.
Создание библиотек кДНК
M. alpina штамма 1S-4 инокулировали в 100 мл среды (1,8% глюкозы, 1% дрожжевого экстракта, pH 6,0) и предварительно культивировали в течение 3 дней при 28°C. Общее количество предварительной культуры инокулировали в 5 л среды (1,8% глюкозы, 1% порошка соевых бобов, 0,1% оливкового масла, 0,01% адеканола, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2∙2H2O, 0,05% МgCl2∙6H2O, pH 6,0) в 10 л культуральном сосуде (Able Co., Tokyo) и инкубировали при аэрации и встряхивании в условиях 300 об/мин, 1 vvm, 26°C в течение 8 дней. В инкубационные дни 1, 2 и 3 добавляли глюкозу в количествах, эквивалентных 2%, 2% и 1,5%, соответственно. На каждой стадии инкубационных дней 1, 2, 3, 6 и 8, клетки собирали, с получением тотальной РНК гуанидингидрохлоридным/CsCl способом. Поли(A)+РНК очищали из тотальной РНК с использованием набора для очистки Oligotex-dT30 <Super> mRNA (Takara Bio Inc.). Библиотеку кДНК на каждой стадии создавали с использованием набора для клонирования ZAP-cDNA GigapackIII Gold (STRATAGENE).
Поиск гомологов SCL4 из S. cerevisiae
Суперконтиги, содержащие последовательности, показанные в SEQ ID NO:5 и SEQ ID NO:10, идентифицировали tblastn анализом аминокислотной последовательности, полученной из SLC4 (YOR175c), кодирующей LPLAT MBOAT семейства S. cerevisiae (PfamPFO3062) по отношению к геномной нуклеотидной последовательности M. alpina 1S-4.
Получение зонда
Для клонирования кДНК, соответствующих SEQ ID NO:5 и SEQ ID NO:10, получали следующие праймеры (таблица 2).
В качестве матрицы использовали библиотеки кДНК для проведения ПЦР с ExTaq (Takara Bio Inc.) и указанными выше праймерами в следующих сочетаниях: MaLPAAT5-1F/MaLPAAT5-3R и MaLPAAT6-2F/MaLPAAT5-3R. Полученные таким образом фрагменты ДНК клонировали с использованием набора для клонирования TOPO-TA (INVITROGEN), с получением плазмиды, содержащей нуклеотиды 195-931 SEQ ID NO:4, обозначенной pCR-LPLAT5-P, и плазмиды, содержащей нуклеотиды 766-1414 SEQ ID NO:9, обозначенной pCR-LPLAT6-P. Затем полученные плазмиды использовали в качестве матрицы для проведения ПЦР с указанными выше праймерами. Для реакции использовали ExTaq (Takara Bio Inc.), но вместо смеси dNTP использовали ПЦР раствор для мечения (Roche Diagnostics), содержащий набор для получения зонда, меченного дигоксигенином (DIG) из амплифицированной ДНК. Полученный зонд использовали для сортировки библиотек кДНК.
Условия гибридизации следующие.
Буфер: 5×SSC, 1% SDS, 50 мМ Tris-HCl (pH 7,5), 50% формамида;
Температура: 42°C (в течение ночи);
Условия промывки: 3 раза в растворе 0,2×SSC, 0,1% SDS (65°C) в течение 20 минут.
Детектирование выполняли с использованием набора для детектирования нуклеиновой кислоты DIG (Roche Diagnostics). Плазмиды вырезали in vivo вырезанием сегмента ДНК из клонов бактериофагов, полученных скринингом, с получением каждой плазмидной ДНК. Плазмиду, имевшую самую длинную вставку среди полученных скринингом с зондом 1 LPLAT5, обозначали pB-LPLAT5, и плазмиду, имевшую самую длинную вставку среди полученных скринингом с зондом 1 LPLAT6, обозначали pB-LPLAT6. Нуклеотидной последовательностью вставки плазмиды pB-LPLAT5, т.е. кДНК LPLAT5, была SEQ ID NO:4, тогда как нуклеотидной последовательностью вставки плазмиды pB-LPLAT6, т.е. кДНК LPLAT6, была SEQ ID NO:9.
Анализ последовательности
Последовательность кДНК LPLAT5, т.е. SEQ ID NO:4, содержала CDS, состоящую из нуклеотидов 161-1693 (SEQ ID NO:3), и ORF, состоящую из нуклеотидов 161-1690 (SEQ ID NO:1). Последовательность кДНК LPLAT5 и полученная из нее аминокислотная последовательность изображены на фиг.2.
С другой стороны, последовательность кДНК LPLAT6, т.е. SEQ ID NO:9, содержала CDS, состоящую из нуклеотидов 38-1759 (SEQ ID NO:8), и ORF, состоящую из нуклеотидов 38-1756 (SEQ ID NO:6). Последовательность кДНК LPLAT6 и полученная из нее аминокислотная последовательность изображены на фиг.3.
SEQ ID NO:1 и SEQ ID NO:6 подвергали анализу гомологии с использованием BLASTX по отношению к аминокислотным последовательностям, внесенным в GENEBANK. Аминокислотная последовательность, полученная из SEQ ID NO:1, продемонстрировала гомологию с гомологами LPLAT из грибов, тогда как аминокислотная последовательность, полученная из SEQ ID NO:6, продемонстрировала гомологию с гомологами LPLAT из животных. Аминокислотные последовательности, продемонстрировавшие самое низкое E-значение или наибольшую идентичность с каждый последовательностью, были такими, как указано ниже. Идентичность нуклеотидной последовательности и идентичность аминокислотной последовательности с последовательностью, демонстрирующей наибольшую идентичность с ORF каждой последовательности, определяли с помощью clustalW, и о них также сообщается ниже.
SEQ ID NO:1 имела 43,2% идентичность нуклеотидной последовательности и 33,3% идентичность аминокислотной последовательности в ORF с гомологом лизофосфолипид-ацилтрансферазы из Schizosaccharomyces pombe (GI:161085648). С другой стороны, SEQ ID NO:6 имела 41,2% идентичность нуклеотидной последовательности и 28,6% идентичность аминокислотной последовательности в ORF с предполагаемым белком из Xenopus laevis (GI:56788919). Идентичность нуклеотидной последовательности и идентичность аминокислотной последовательности в ORF между LPLAT и LPLAT6 составляют 40,0% и 19,1%, соответственно.
Геномные последовательности, содержащие CDS LPLAT5 (SEQ ID NO:3) и CDS LPLAT6 (SEQ ID NO:8), описаны в SEQ ID NO:5 и SEQ ID NO:10, соответственно. SEQ ID NO:5 содержала два интрона и экзоны, соответствующие нуклеотидам 1-314, 461-587, и 668-1759. С другой стороны, SEQ ID NO:10 содержала один интрон и экзон, соответствующие нуклеотидам 1-1095 и 1318-1944. Фиг.4 иллюстрирует выравнивание между геномной последовательностью и ORF последовательностью LPLAT5, и фиг.5 иллюстрирует выравнивание между геномной последовательностью и ORF последовательностью LPLAT6.
[Пример 2]
Конструирование дрожжевых экспрессионных векторов
Для того чтобы экспрессировать LPLAT5 и LPLAT6 в дрожжах, векторы конструировали следующим образом.
Используя pBLPLAT5 в качестве матрицы, выполняли ПЦР с ExTaq (Takara Bio) и праймером Eco-MaLPLAT5-F(SEQ ID NO:15):
GAATTCATGCTAAACTCATTCTTCGGGGACGC и
праймером Xho-MaLPLAT5-R(SEQ ID NO:16):
CTCGAGTTACAGCGTCTTGATTTTAACTGCAGC.
Полученные фрагменты ДНК клонировали с использованием набора для клонирования TOPO-TA (INVITROGEN), и определяли нуклеотидную последовательность вставки, с получением плазмиды, имеющей соответствующую нуклеотидную последовательность, обозначенную pCR-LPLAT5. Фрагмент ДНК около 1,6 т.п.н., полученный обработкой полученной плазмиды эндонуклеазами рестрикции EcoRI и XhoI, встраивали по EcoRI-SalI сайтам дрожжевого экспрессионного вектора pYE22m (Appl. Microbiol. Biotechnol., 30, 515-520, 1989) для генерирования плазмиды pYE-MALPLAT5.
С другой стороны, фрагмент ДНК 1,9 т.п.н., полученный обработкой pBLPLAT6 эндонуклеазами рестрикции EcoRI и KpnI, встраивали по EcoRI-KpnI сайтам дрожжевого экспрессирующего вектора pYE22m для генерирования плазмиды pYE-LPLAT6.
Экспрессия в продуцирующих арахидоновую кислоту дрожжах
(1) Селекция продуцирующих арахидоновую кислоту дрожжей
Для селекции продуцирующих арахидоновую кислоту дрожжей (S. cerevisiae) были сконструированы следующие плазмиды.
Первоначально выполнена ПЦР с использованием кДНК, полученной из M. alpina штамма 1S-4, в качестве матрицы с ExTaq и следующим набором праймеров: Δ12-f/Δ12-r, Δ6-f/Δ6-r, GLELO-f/GLELO-r или Δ5-f/Δ5-r для амплификации гена Δ12 десатуразы жирных кислот, гена Δ6 десатуразы жирных кислот, гена GLELO элонгазы жирных кислот и гена Δ5 десатуразы жирных кислот M. alpina штамма 1S-4.
Δ12-f: TCTAGAATGGCACCTCCCAACACTATTG (SEQ ID NO:17)
Δ12-r: AAGCTTTTACTTCTTGAAAAAGACCACGTC (SEQ ID NO:18)
Δ6-f: TCTAGAATGGCTGCTGCTCCCAGTGTGAG (SEQ ID NO:19)
Δ6-r: AAGCTTTTACTGTGCCTTGCCCATCTTGG (SEQ ID NO:20)
GLELO-f: TCTAGAATGGAGTCGATTGCGCAATTCC (SEQ ID NO:21)
GLELO-r: GAGCTCTTACTGCAACTTCCTTGCCTTCTC (SEQ ID NO:22)
Δ5-f: TCTAGAATGGGTGCGGACACAGGAAAAACC (SEQ ID NO:23)
Δ5-r: AAGCTTTTACTCTTCCTTGGGACGAAGACC (SEQ ID NO:24).
Гены клонировали с использованием набора для клонирования TOPO-TA. Были определены нуклеотидные последовательности, и клоны, содержавшие нуклеотидные последовательности, были обозначены как плазмиды pCR-MAΔ12DS (содержащая нуклеотидную последовательность SEQ ID NO:25), pCR-MAΔ6DS (содержащая нуклеотидную последовательность SEQ ID NO:26), pCR-MAGLELO (содержащая нуклеотидную последовательность SEQ ID NO:27) и pCR-MAΔ5DS (содержащая нуклеотидную последовательность SEQ ID NO:28).
Фрагмент ДНК около 1,2 т.п.н., полученный обработкой плазмиды pURA34 (JPA 2001-120276 A) эндонуклеазами рестрикции HindIII, встраивали по HindIII сайту вектора, полученного обработкой вектора pUC18 эндонуклеазами рестрикции EcoRI и SphI с последующим затуплением концов и лигированием для генерирования клона, обозначенного pUC-URA3, с EcoRI сайтом вектора на 5'-конце URA3. Фрагмент ДНК около 2,2 т.п.н., полученный обработкой YEp13 эндонуклеазами рестрикции SalI и XhoI, встраивали по SalI сайту вектора pUC18 для генерирования клона, обозначенного pUC-LEU2, с EcoRI сайтом вектора на 5'-конце LEU2.
Затем фрагмент ДНК около 1,2 т.п.н., полученный обработкой плазмиды pCR-MAΔ12DS эндонуклеазами рестрикции HindIII с последующим затуплением концов и дополнительной обработкой эндонуклеазой рестрикции XbaI, лигировали во фрагмент ДНК около 6,6 т.п.н., полученный обработкой вектора pESC-UR (STRATAGENE) эндонуклеазой рестрикции SacI с последующим затуплением концов и дополнительной обработкой эндонуклеазой рестрикции SpeI для генерирования плазмиды pESC-U-Δ12. Фрагмент ДНК около 1,6 т.п.н., полученный обработкой плазмиды pCR-MAΔ6DS эндонуклеазами рестрикции XbaI с последующим затуплением концов и дополнительной обработкой эндонуклеазой рестрикции HindIII, лигировали во фрагмент ДНК около 8 т.п.н., полученный обработкой плазмиды pESC-U-Δ12 эндонуклеазой рестрикции SalI с последующим затуплением концов и дополнительной обработкой эндонуклеазой рестрикции HindIII для генерирования плазмиды pESC-U-Δ12:Δ6. Фрагмент около 4,2 т.п.н., полученный частичным расщеплением pESC-U-Δ12:Δ6 эндонуклеазой рестрикции PvuII встраивали по SmaI сайту pUC-URA3 для генерирования плазмиды pUC-URA-Δ12:Δ6.
Фрагмент ДНК около 0,95 т.п.н., полученный обработкой плазмиды pCR-MAGLELO эндонуклеазами рестрикции XbaI и SacI, лигировали во фрагмент ДНК около 7,7 т.п.н., полученный обработкой вектора pESC-LEU (STRATAGENE) эндонуклеазами рестрикции XbaI и SacI для генерирования плазмиды pESC-L-GLELO. Фрагмент ДНК около 1,3 т.п.н., полученный обработкой плазмиды pCR-MAΔ5DS эндонуклеазой рестрикции XbaI с последующим затуплением концов и дополнительной обработкой эндонуклеазой рестрикции HindIII, лигировали во фрагмент ДНК около 8,7 т.п.н., полученный обработкой плазмиды pESC-L-GLELO эндонуклеазой рестрикции ApaI с последующим затуплением концов и дополнительной обработкой эндонуклеазой рестрикции HindIII, для генерирования плазмиды pESC-L-GLELO:Δ5. Фрагмент около 3,2 т.п.н., полученный обработкой полученной плазмиды эндонуклеазой рестрикции PvuII встраивали по SmaI сайту pUC-LEU2 для генерирования плазмиды pUC-LEU-GLELO:Δ5. S. cerevisiae штамма YPH499 (STRATAGENE) трансформировали совместно с плазмидой pUC-URA-Δ12:Δ6 и плазмидой pUC-LEU-GLELO:Δ5. Трансформированные штаммы выбирали на основании жизнеспособности на SC-Leu, Ura агарозной среде (2% агара), содержащей на литр 6,7 г дрожжевой азотистой основы без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 1,2 г триптофана). Произвольно один из выбранных штаммов обозначали ARA3-1 штаммом. Этот штамм может продуцировать арахидоновую кислоту посредством экспрессии гена Δ12 десатуразы жирных кислот, гена Δ6 десатуразы жирных кислот, гена GLELO элонгазы жирных кислот и гена Δ5 десатуразы жирных кислот из-под GAL1/10 промотора при культивировании в содержащей галактозу среде.
(2) Трансформация продуцирующих арахидоновую кислоту дрожжей
Штамм ARA3-1 трансформировали плазмидами pYE22m, pYE-MALPLAT5 и pYE-MALPLAT6. Трансформированные штаммы отбирали по жизнеспособности на SC-Trp, Leu, Ura агарозной среде (2% агара) содержащей на литр 6,7 г дрожжевой азотистой основы без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина и 6 г треонина).
Три произвольных штамма, трансформированных каждой плазмидой, использовали для последующего культивирования. В следующих таблицах 3-8 контроль представляет штаммы, трансформированные плазмидой pYE22m, LPLAT5 представляет штаммы, трансформированные плазмидой pYE-MALPLAT5, и LPLAT6 представляет штаммы, трансформированные плазмидой pYE-MALPLAT6.
(3) Культивирование в среде без жирных кислот
Трансформированные ранее штаммы культивировали при встряхивании в 10 мл жидкой среды SC-Trp, Leu, Ura при 30°C в течение 1 дня, и 1 мл культур инкубировали при встряхивании в 10 мл SG-Trp, Leu, Ura жидкой среды, содержащей на литр 6,7 г дрожжевой азотистой основы без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина и 6 г треонина) при 15°C в течение 6 дней. Клетки собирали, промывали водой, затем лиофилизировали и подвергали жирно-кислотному анализу.
Результаты представлены в таблице 3.
Композиционное соотношение жирных кислот в дрожжевых клетках, экспрессирующих каждый ген (%)
- культивировано в среде без жирных кислот
На основании результатов из таблицы 3 определяли процент преобразования жирной кислоты в другую жирную кислоту по синтетическому пути арахидоновой кислоты. Например, преобразование 18:2→18:3(n-6) определяется следующим образом:
Процент преобразования=(18:3(n-6)+DGLA+ARA)/(18:2+18:3(n-6)+DGLA+ARA)×100
Результаты представлены в таблице 4.
Преобразования жирных кислот в синтетическом пути арахидоновой кислоты (%)
- культивировано в среде без жирных кислот -
Как показано в таблицах 3 и 4, количественное соотношение арахидоновой кислоты в общем количестве жирных кислот повысилось в 1,5 раза в LPLAT5-экспрессирующих штаммах и в 2,3 раза в LPLAT6-экспрессирующих штаммах по сравнению с контролем. Были рассмотрены преобразования жирных кислот в биосинтетическом пути арахидоновой кислоты, при этом было обнаружено, что преобразования 18:1→18:2, 18:3(n-6)→DGLA и DGLA→ARA были повышены в LPLAT5-экспрессирующих штаммах, тогда как в LPLAT6-экспрессирующих штаммах было существенно повышено преобразование DGLA→ARA. Для таких преобразований требовался ацильный перенос от ацил-CoA к фосфолипидам или от фосфолипидов к CoA, как показано на фиг.1, при этом предполагается, что LPLAT5 и LPLAT6 вовлечены в такие преобразования.
(4) Культивирование в среде, содержащей линолевую кислоту
Трансформированные штаммы культивировали при встряхивании в 10 мл жидкой среды SC-Trp, Leu, Ura при 30°C в течение 1 дня, и 1 мл культуры инокулировали в 10 мл жидкой среды SC-Trp, Leu, Ura, содержащей 5 мг/мл линолевой кислоты и 0,1% Triton X-100, и инкубировали при встряхивании при 15°C в течение 6 дней. Клетки собирали, промывали водой, затем лиофилизировали и подвергали жирно-кислотному анализу. Результаты представлены в таблице 5.
Композиционное соотношение жирных кислот в дрожжевых клетках, экспрессирующих каждый ген (%)
- культивировано в среде, содержащей линолевую кислоту -
На основании результатов в таблице 5 определяли процент преобразования жирной кислоты в другую жирную кислоту в синтетическом пути арахидоновой кислоты. Результаты представлены в таблице 6.
Преобразование жирных кислот в синтетическом пути арахидоновой кислоты (%)
Как показано в таблице 5, количественное соотношение арахидоновой кислоты в общем количестве жирных кислот повысилось в 2 раза в LPLAT5-экспрессирующих штаммах и в 3,5 раза в LPLAT6-экспрессирующих штаммах по сравнению с контролем. В LPLAT5-экспрессирующих штаммах, количественное соотношение при добавлении линолевой кислоты повысилось по сравнению с контролем. Были рассмотрены преобразования жирных кислот в синтетическом пути арахидоновой кислоты (таблица 6), при этом было обнаружено, что преобразования 18:3(n-6)→DGLA и DGLA→ARA повысились как в LPLAT5-экспрессирующих штаммах, так и в LPLAT6-экспрессирующих штаммах, особенно значительно в LPLAT6-экспрессирующих штаммах.
(5) Культивирование в среде, содержащей γ-линоленовую кислоту
Трансформированные штаммы культивировали при встряхивании в 10 мл жидкой среды SC-Trp, Leu, Ura при 30°C в течение 1 дня, и по 1 мл культур инокулировали в 10 мл жидкой среды SC-Trp, Leu, Ura, содержащей 5 мг/мл γ-линоленовой кислоты и 0,1% Triton X-100, и инкубировали при встряхивании при 15°C в течение 6 дней. Клетки собирали, промывали водой, затем лиофилизировали и подвергали жирно-кислотному анализу. Результаты представлены в таблице 7.
Композиционное соотношение жирных кислот в дрожжевых клетках, экспрессирующих каждый ген (%)
- культивировано в среде, содержащей γ-линоленовую кислоту -
На основании результатов в таблице 7 определяли преобразование жирной кислоты в другую жирную кислоту в синтетическом пути арахидоновой кислоты (таблица 8).
Преобразование жирных кислот, идущих после γ-линоленовой кислоты, в синтетическом пути арахидоновой кислоты (%)
- культивировано в среде, содержащей γ-линоленовую кислоту -
Как представлено в таблице 7, количественное соотношение γ-линоленовой кислоты, добавленной к общему количеству жирных кислот, повысилось в LPLAT5-экспрессирующих штаммах. Однако процентное содержание идущих после продуктов дигомо-γ-линоленовой кислоты и арахидоновой кислоты не увеличилось (таблица 8). Напротив, количественное соотношение арахидоновой кислоты в общем количестве жирных кислот повысилось в 2,8 раза по сравнению с контролем, и преобразование DGLA→ARA значительно повысилось в LPLAT6-экспрессирующих штаммах (таблица 8).
Эти результаты показывают, что LPLAT5 и LPLAT6 могут повысить преобразование жирных кислот, для которых требуется ацильный перенос от ацил-CoA к фосфолипидам или от фосфолипидов к CoA. Было предположено участие LPLAT5 в преобразовании из 18:1-CoA в 18:1-PL, преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразовании из DGLA-CoA в DGLA-PL. С другой стороны, было предположено участие LPLAT6 в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразовании из DGLA-CoA в DGLA-PL.
[Пример 3]
Функциональный анализ LPLAT6 в M. alpina
Создание экспрессионных векторов Mortierella
Для использования в качестве адаптеров синтезировали следующие олигонуклеотиды.
A-1: GATCCGGCGCGCCGCGGCCGCTCTAGAGTCGACGGCGCGCC (SEQ ID NO:29)
A-2: AGCTTGGCGCGCCGTCGACTCTAGAGCGGCCGCGGCGCGCCG (SEQ ID NO:30).
A-1 и A-2 гибридизовали и лигировали во фрагмент, полученный обработкой плазмиды pUC18 эндонуклеазами рестрикции EcoRI и HindIII для генерирования pUC18-R.
С использованием в качестве матрицы геномной ДНК или плазмиды, полученной из M. alpina штамма 1S-4, каждый фрагмент ДНК амплифицировали с помощью ПЦР, используя ExTaq (Takara Bio) со следующим набором праймеров, и клонировали при использовании набора для клонирования TOPO-T (Invitrogen).
В частности, геномную ДНК использовали в качестве матрицы для амплификации геномной ДНК около 2 т.п.н., содержащей URA5 ген, используя набор праймеров:
праймер URA5g-F1: GTCGACCATGACAAGTTTGC (SEQ ID NO:31) и
праймер URA5g-R1: GTCGACTGGAAGACGAGCACG (SEQ ID NO:32);
для амплификации GAPDH промотора около 0,9 т.п.н., используя набор праймеров:
праймер GAPDHp-F1: GTCGACGATCACGTCGGGTGATGAGTTG (SEQ ID NO:33),
праймер GAPDHp-R1: TCTAGAGATGTTGAATGTGTGGTGTGTG (SEQ ID NO:34);
и для амплификации GAPDH терминатора около 0,5 т.п.н., используя набор праймеров:
праймер GAPDHт-F1: GCGGCCGCTAAGAAAAGGGAGTGAATCGC (SEQ ID NO:35) и
праймер GAPDHт-R1: GGATCCGGCGCGCCGATCCATGCACGGGTCCTTCTC (SEQ ID NO:36).
Плазмиду pB-LPLAT6 использовали в качестве матрицы для амплификации CDS около 1,6 т.п.н. гена LPLAT6, используя набор праймеров:
праймер XbaI-LPLAT6-F1: TCTAGAATGGAGGCACTCTTGCACCAGG (SEQ ID NO:37) и
праймер NotI-LPLAT6-R1: GCGGCCGCTTACTCAGTCTTGACAGACTTG (SEQ ID NO:38);
и для амплификации 3'-фрагмента около 0,7 т.п.н. CDS гена LPLAT6, используя набор праймеров:
праймер EcoRV-LPLAT6-F2: GATATCGGGTAAAGCCTTCCTGGAACG (SEQ ID NO:39) и
праймер XbaI-LPLAT6-R2: TCTAGATTACTCAGTCTTGACAGACTTGGATCG (SEQ ID NO:40).
Аналогично, плазмиду pCR-MAΔ5DS использовали в качестве матрицы для амплификации CDS около 1,3 т.п.н. гена Δ5 десатуразы жирных кислот, используя набор праймеров:
праймер XbaI-Δ5DS-F1: TCTAGAATGGGTGCGGACACAGGAAAAAC (SEQ ID NO:41) и
праймер NotI-Δ5DS-R1: GCGGCCGCTTACTCTTCCTTGGGACGAAG (SEQ ID NO:42);
и для амплификации 3'-фрагмента около 0,5 т.п.н. CDS гена Δ5 десатуразы жирных кислот, используя набор праймеров:
праймер NdeI-Δ5DS-R2: TCTAGATTACTCTTCCTTGGGACGAAG (SEQ ID NO:43) и
праймер XbaI-Δ5DS-F2: CATATGCATCCAGGACATCAACATCTTG (SEQ ID NO:44).
По сайтам эндонуклеаз рестрикции EcoRI/NotI плазмиды pUC18-R встраивали фрагмент, вырезанный такими же эндонуклеазами рестрикции из GAPDH терминатора, для генерирования плазмиды pUC-GAPDHt. Впоследствии, плазмиду pUC-GAPDHt расщепляли эндонуклеазами рестрикции XbaI и SalI, и встраивали фрагмент, вырезанный такими же эндонуклеазами рестрикции из GAPDH промотора, для генерирования плазмиды pUC-GAPDHpt. Плазмиду pUC-GAPDHpt расщепляли эндонуклеазой рестрикции SalI и встраивали фрагмент, расщепленный такими же эндонуклеазами рестрикции из геномной ДНК, содержащей URA5 ген. Подтверждали ориентацию вставок и выбирали вектор, содержащий ген URA5, встроенный в такой же ориентации, как для сайтов эндонуклеаз рестрикции EcoRI→HindIII, и обозначали как плазмида pDUraRSC.
Плазмиду pDUraRSC расщепляли эндонуклеазами рестрикции XbaI и NotI, и фрагмент ДНК, вырезанный такими же эндонуклеазами рестрикции из CDS гена LPLAT6, встраивали для генерирования плазмиды pDUraRSC-LPLAT6. Фрагмент ДНК около 7 т.п.н., полученный расщеплением плазмиды pDUraRSC-LPLAT6 эндонуклеазами рестрикции EcoRV и XbaI, лигировали во фрагмент ДНК, вырезанный такими же эндонуклеазами рестрикции из 3'-фрагмента около 0,7 т.п.н. CDS гена LPLAT6 для генерирования плазмиды pDUraRSC-LPLAT6-RNAi.
Конструирование векторов с подавленной экспрессией Δ5DS (RNAi)
Плазмиду pDUraRSC расщепляли эндонуклеазами рестрикции XbaI и NotI, и фрагмент ДНК вырезали такими же эндонуклеазами рестрикции из CDS гена Δ5 десатуразы жирных кислот для генерирования плазмиды pDUraRSC-Δ5DS. Фрагмент ДНК около 1,2 т.п.н., полученный расщеплением плазмиды pDUraRSC-Δ5DS эндонуклеазами рестрикции EcoRI и NdeI лигировали во фрагмент ДНК около 5,5 т.п.н., полученный расщеплением эндонуклеазами рестрикции XbaI и EcoRI, и фрагмент вырезали эндонуклеазами рестрикции NdeI и XbaI из 3'-фрагмента CDS гена Δ5 десатуразы жирных кислот около 0,5 т.п.н. для генерирования плазмиды pDUraRSC-Δ5DS-S-RNAi.
Выработка трансформированных штаммов M. alpina
Урацил-зависимый ауксотрофный штамм Δura-3, полученный из M. alpina, в соответствии со способом, описанным в патентном документе (WO2005/019437, названный “Method of Breeding Lipid-Producing Fungus”), в плазмиде pDUraRSC-LPLAT6-RNAi или в плазмиде pDUraRSC-Δ5DS-RNAi использовали в качестве хозяина и трансформировали способом доставки частиц. Для отбора трансформированных штаммов использовали агарозную среду SC (0,5% дрожжевой азотистой основы без аминокислот и аммонийсульфатом (Difco), 0,17% аммонийсульфата, 2% глюкозы, 0,002% аденина, 0,003% тирозина, 0,0001% метионина, 0,0002% аргинина, 0,0002% гистидина, 0,0004% лизина, 0,0004% триптофана, 0,0005% треонина, 0,0006% изолейцина, 0,0006% лейцина, 0,0006% фенилаланина и 2% агара).
Оценка трансформированных штаммов M. alpina
Около 50 штаммов, трансформированных каждой плазмидой, инокулировали в 4 мл среды GY (2% глюкозы, 1% дрожжевого экстракта, pH 6,0) и культивировали при встряхивании при 28°C в течение 4 дней. В конце культивирования клетки собирали фильтрованием и лиофилизировали. Часть лиофилизированных клеток (около 10-20 мг) собирали, и жирные кислоты в клетках преобразовывали в метиловые эфиры с использованием метанольной HCl, затем экстрагировали гексаном, гексан отгоняли и остаток подвергали жирно-кислотному анализу газовой хроматографией. Среди штаммов, трансформированных разными плазмидами, отбирали имеющие более высокое количественное соотношение дигомо-γ-линоленовой кислоты, чем количественное соотношение арахидоновой кислоты, т.е. LPLAT6-D#6 (трансформированный плазмидой pDUraRSC-LPLAT6-RNAi) и Δ5DS-D#45 (трансформированный плазмидой pDUraRSC-Δ5DS-RNAi).
Два штамма и контроль (M. alpina дикого типа штамма 1S-4) культивировали при встряхивании в 4 мл среды GY при 28°C в течение 4 дней. В конце культивирования клетки собирали фильтрованием и лиофилизировали. Часть лиофилизированных клеток (около 10-20 мг) собирали и разрушали механически. Клетки выдерживали в 4 мл смеси хлороформ-метанол (2:1) при 70°C в течение 1 часа при периодическом встряхивании и затем центрифугировали для сбора супернатанта. Оставшиеся клетки выдерживали еще раз в 4 мл смеси хлороформ-метанол (2:1) при 70°C в течение 1 часа при периодическом встряхивании и затем центрифугировали для сбора супернатанта, который смешивали с предыдущим супернатантом. Липиды сушили в центрифужном концентраторе SpeedVac и растворяли в 5 мл хлороформа. Один мл раствора сушили, как описано выше, и жирные кислоты преобразовывали в метиловые эфиры с использованием метанольной HCl и подвергали жирно-кислотному анализу. В то же время, 2 мл раствора хлороформа также сушили, как описано выше, растворяли в небольшом количестве хлороформа и весь раствор подвергали тонкослойной хроматографии следующим образом. Липиды распределяли тонкослойной хроматографией на 60 силикагелевых пластинах (Merck), элюировали смесью гексан:диэтиловый эфир:уксусная кислота 70:30:1. Пластины опрыскивали водным раствором, содержащим 0,015% примулин, 80% ацетона (примулин раствор), и липиды визуализировали при УФ облучении, затем триацилглицериновые фракции (TG) и фосфолипидные (PL) фракции отмечали карандашом, силикагель в отмеченных участках снимали и собирали в пробирки. Жирные кислоты преобразовывали в метиловые эфиры с использованием метанольной HCl и подвергали жирно-кислотному анализу газовой хроматографией. Таким образом, жирные кислоты преобразовывали в метиловые эфиры путем взаимодействия с 1 мл дихлорметана и 2 мл 10% метанольной HCl при 50°C в течение 3 часов. Затем добавляли 4 мл гексана и 1 мл воды, раствор интенсивно перемешивали, затем центрифугировали и верхний слой собирали. Растворитель отгоняли в SpeedVac, остаток растворяли в ацетонитриле и подвергали жирно-кислотному анализу газовой хроматографией. Результаты показаны на фиг.6-8.
Фиг.6 демонстрирует количественное соотношение в композиции полиненасыщенных жирных кислот во всех липидах, экстрагированных смесью хлороформ-метанол (2:1). В отличие от контроля, содержащего высокое количественное соотношение арахидоновой кислоты, штамм LPLAT6-D#6 и штамм Δ5DS-D#45 продемонстрировали сопоставимое количественное соотношение дигомо-γ-линоленовой кислоты и арахидоновой кислоты из-за ингибирования преобразования из дигомо-γ-линоленовой кислоты в арахидоновую кислоту. Фиг.7 демонстрирует состав композиции полиненасыщенных жирных кислот в триацилглицеринах, составляющих большую часть липидов в клетках. Аналогично составу композиции во всех липидах в клетках, штамм LPLAT6-D#6 и штамм Δ5DS-D#45 продемонстрировали более высокое количественное соотношение дигомо-γ-линоленовой кислоты по сравнению с контролем. Тем не менее, состав композиции жирных кислот фосфолипидной фракции, показанный на фиг.8, значительно различался между штаммом LPLAT6-D#6 и штаммом Δ5DS-D#45. В частности, штамм Δ5DS-D#45 продемонстрировал высокое количественное соотношение дигомо-γ-линоленовой кислоты, тогда как штамм LPLAT6-D#6 продемонстрировал высокое количественное соотношение арахидоновой кислоты, но ниже контроля, а также продемонстрировал высокое количественное соотношение γ-линоленовой кислоты по сравнению с контролем и штаммом Δ5DS-D#45.
Предполагается, что биосинтетический путь арахидоновой кислоты в M. alpina происходит, как показано на фиг.1. Эксперименты, описанные выше, также однозначно показывают, что Δ5 десатураза жирных кислот воздействует на DGLA-PL с продуцированием арахидоновой кислоты. Напротив, 18:3(n-6)-PL накапливалась в штаммах с подавленной экспрессией LPLAT6. Количественное соотношение DGLA в TG фракции повысилось, но не наблюдалось существенного увеличения количественного соотношения DGLA в PL фракциях. Эти результаты однозначно показывают, что LPLAT6 обуславливает преобразование 18:3(n-6)-PL в 18:3(n-6)-CoA и преобразование DGLA-CoA в DGLA-PL в M. alpina.
Свободный текст списка последовательностей
SEQ ID NO:11: праймер
SEQ ID NO:12: праймер
SEQ ID NO:13: праймер
SEQ ID NO:14: праймер
SEQ ID NO:15: праймер
SEQ ID NO:16: праймер
SEQ ID NO:17: праймер
SEQ ID NO:18: праймер
SEQ ID NO:19: праймер
SEQ ID NO:20: праймер
SEQ ID NO:21: праймер
SEQ ID NO:22: праймер
SEQ ID NO:23: праймер
SEQ ID NO:24: праймер
SEQ ID NO:29: адаптер A-1
SEQ ID NO:30: адаптер A-2
SEQ ID NO:31: праймер URA5g-F1
SEQ ID NO:32: праймер URA5g-R1
SEQ ID NO:33: праймер GAPDHp-F1
SEQ ID NO:34: праймер GAPDHp-R1
SEQ ID NO:35: праймер GAPDHt-F1
SEQ ID NO:36: праймер GAPDHt-R1
SEQ ID NO:37: праймер XbaI-LPLAT6-F1
SEQ ID NO:38: праймер NotI-LPLAT6-R1
SEQ ID NO:39: праймер EcoRV-LPLAT6-F2
SEQ ID NO:40: праймер XbaI-LPLAT6-R2
SEQ ID NO:41: праймер XbaI-Δ5DS-F1
SEQ ID NO:42: праймер NotI-Δ5DS-R1
SEQ ID NO:43: праймер NdeI-Δ5DS-F1
SEQ ID NO:44: праймер XbaI-Δ5DS-R1

























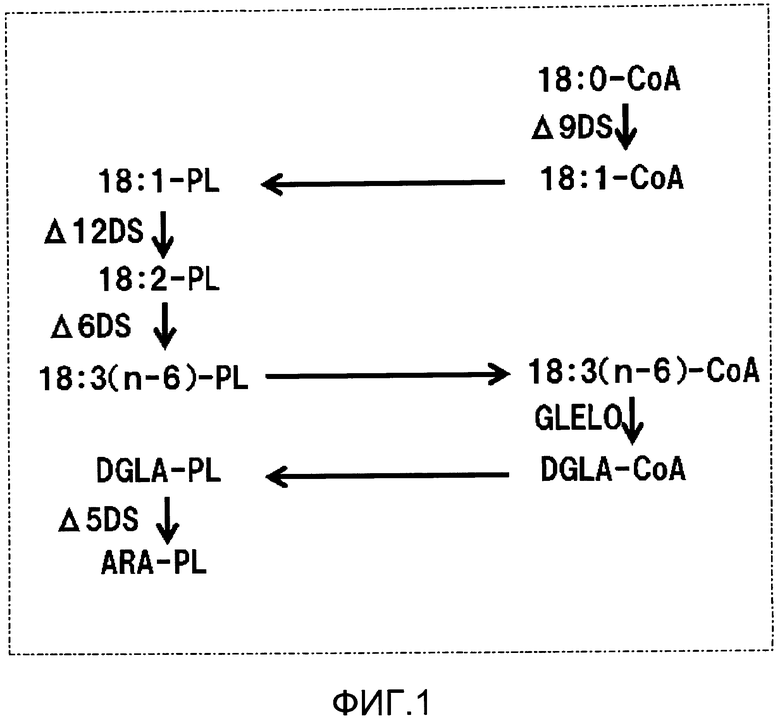







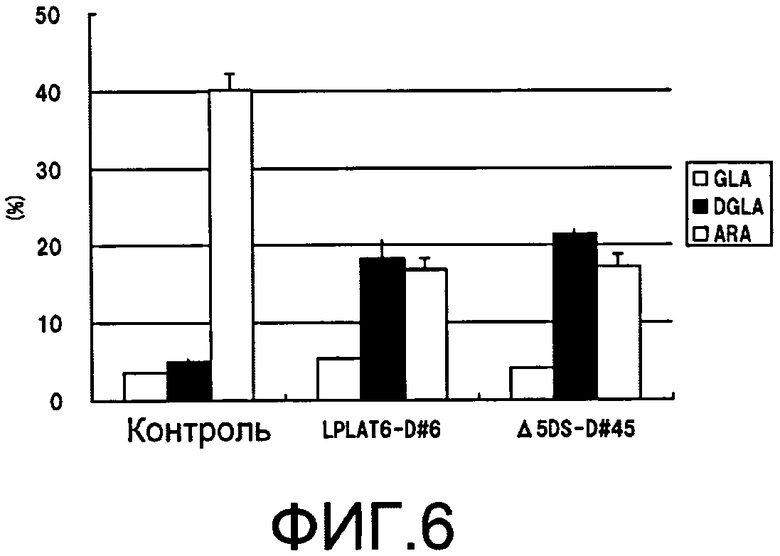
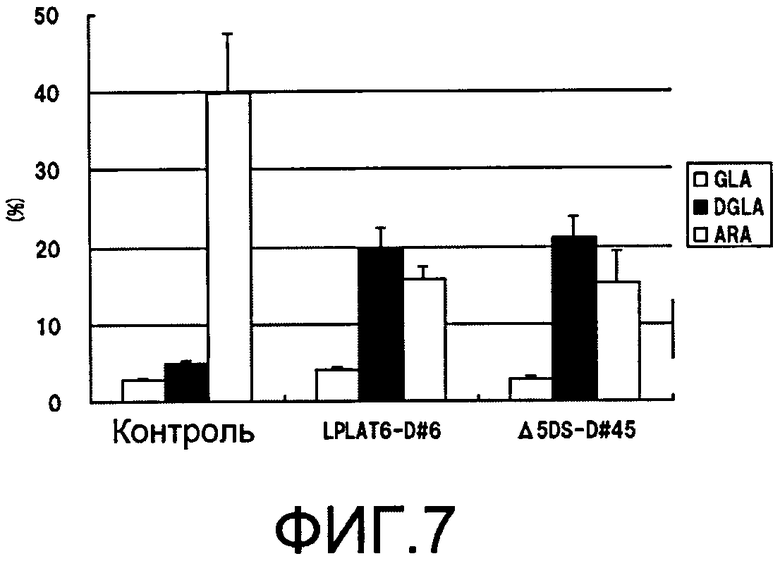
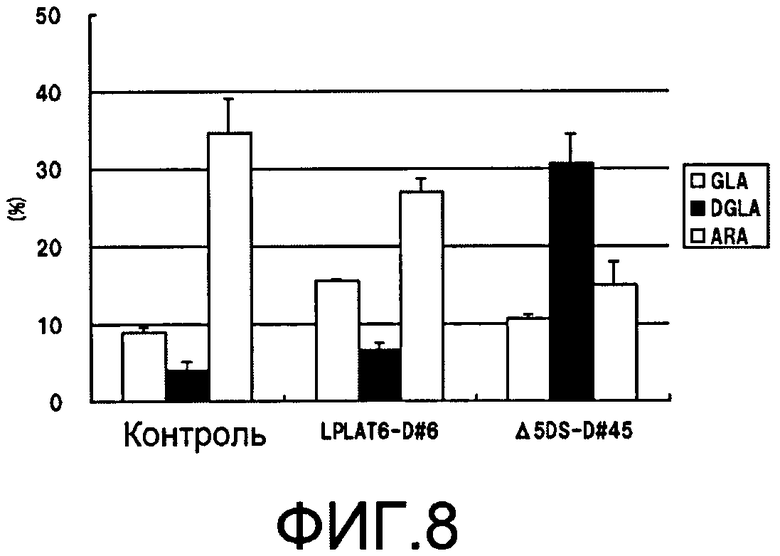
Изобретение относится к области биотехнологии. Представлены вариантные белки лизофосфолипид-ацилтрансферазы, катализирующие реакцию превращения 18:3(n-6)-PL в 18:3(n-6)-CoA и/или DGLA-CoA в DGLA-PL, имеющие аминокислотные последовательности, по меньшей мере на 95% идентичные SEQ ID NO:2 и 7, представленным в описании. Кроме того, раскрыты нуклеиновые кислоты, кодирующие указанные белки. Также представлены рекомбинантный вектор экспрессии, содержащий указанные нуклеиновые кислоты, и клетка-хозяин для получения композиции жирных кислот, трансформированная указанным вектором. Предложено применение указанного вектора для повышения количественного соотношения арахидоновой кислоты (ARA) в композиции жирных кислот в хозяине. Описан способ получения композиции жирных кислот, включающий культивирование клетки-хозяина для получения композиции жирных кислот с повышенным количеством арахидоновой кислоты по сравнению с ее количеством в композиции жирных кислот, полученной культивированием нетрансформированного хозяина, и сбор композиции жирных кислот из культур трансформированной клетки. Изобретение позволяет получить композицию жирных кислот с повышенным содержанием арахидоновой кислоты. 12 н. и 6 з.п. ф-лы, 8 ил., 8 табл., 3 пр.
1. Нуклеиновая кислота, кодирующая белок, обладающий активностью лизофосфолипид-ацилтрансферазы, выбранная из любой согласно (а)-(е) ниже:
(a) нуклеиновая кислота, кодирующая указанный белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-25 аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая гибридизуется в очень жестких условиях, включающих условия гибридизации 0,2 х SSC при 63°С и условия промывки 0,2 х SSC при 60-65°С, с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая кодирует указанный белок;
(c) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, обладающей 95% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирует указанный белок;
(d) нуклеиновая кислота, кодирующая указанный белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7; и
(e) нуклеиновая кислота, которая гибридизуется в очень жестких условиях, включающих условия гибридизации 0,2 х SSC при 63°С и условия промывки 0,2 х SSC при 60-65°С, с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая кодирует указанный белок.
2. Нуклеиновая кислота по п.1, выбранная из (а)-(с) ниже:
(а) нуклеиновая кислота, которая кодирует указанный белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-10 в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, обладающей 98% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирует белок;
(c) нуклеиновая кислота, которая кодирует указанный белок, состоящий из аминокислотной последовательности, обладающей идентичностью 98% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
3. Нуклеиновая кислота, кодирующая белок, обеспечивающий увеличенное соотношение арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим указанную нуклеиновую кислоту, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором, выбранная из любой согласно (а)-(е) ниже:
(а) нуклеиновая кислота, которая кодирует указанный белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-25 аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая гибридизуется в очень жестких условиях, включающих условия гибридизации 0,2 х SSC при 63°С и условия промывки 0,2 х SSC при 60-65°С, с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая кодирует указанный белок;
(c) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, обладающей 95% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирует указанный белок;
(d) нуклеиновая кислота, которая кодирует белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7; и
(e) нуклеиновая кислота, которая гибридизуется в очень жестких условиях, включающих условия гибридизации 0,2 х SSC при 63°С и условия промывки 0,2 х SSC при 60-65°С, с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая кодирует указанный белок.
4. Нуклеиновая кислота по п.3, выбранная из (а)-(с):
(a) нуклеиновая кислота, которая кодирует указанный белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-10 в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, обладающей 98% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирует указанный белок; и
(c) нуклеиновая кислота, которая кодирует белок, состоящий из аминокислотной последовательности, обладающей идентичностью 98% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
5. Нуклеиновая кислота по любому из пп.1-4, где кодируемый белок принадлежит семейству мембранно-связанных О-ацилтрансфераз.
6. Нуклеиновая кислота, кодирующая белок, обладающий активностью лизофосфолипид-ацилтрансферазы, выбранная из любой согласно (a)-(d) ниже:
(a) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, показанной в SEQ ID NO:1 или 6;
(b) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(c) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, показанной в SEQ ID NO:4 или 9; и
(d) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, показанной в SEQ ID NO:5 или 10.
7. Белок, обладающий активностью лизофосфолипид- ацилтрансферазы, выбранный из (а) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-25 аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
8. Белок по п.7, выбранный из (а) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-10 в аминокислотной последовательности SEQ ID NO:2 или 7; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 98% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
9. Белок, обеспечивающий увеличенное количественное соотношение арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном рекомбинантным вектором, содержащим нуклеиновую кислоту, кодирующую аминокислотную последовательность, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором, выбранный из (а) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-25 аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
10. Белок по п.9, который выбран из (а) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением аминокислот 1-10 в аминокислотной последовательности SEQ ID NO:2 или 7; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 98% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
11. Белок по любому из пп.7-10, который принадлежит семейству мембранно-связанных О-ацилтрансфераз.
12. Белок, обладающий активностью лизофосфолипид-ацилтрансферазы, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7.
13. Рекомбинантный вектор экспрессии, содержащий нуклеиновую кислоту по любому из пп.1-6.
14. Клетка для получения композиции жирных кислот, трансформированная рекомбинантным вектором по п.13.
15. Способ получения композиции жирных кислот, включающий культивирование трансформированной клетки по п.14 для получения композиции жирных кислот, где количественное соотношение арахидоновой кислоты в композиционном соотношении жирных кислот в указанной композиции жирных кислот выше, чем количественное соотношение арахидоновой кислоты в композиции жирных кислот, полученной культивированием нетрансформированного хозяина, и сбор композиции жирных кислот из культур трансформированной клетки по п.14.
16. Применение рекомбинантного вектора по п.13 для повышения количественного соотношения арахидоновой кислоты в композиционном соотношении жирных кислот в хозяине, трансформированном вектором, по сравнению с количественным соотношением в композиционном соотношении жирных кислот в хозяине, который не был трансформирован вектором.
17. Нуклеиновая кислота, кодирующая белок, участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-СоА и/или преобразовании из DGLA-CoA в DGLA-PL, выбранная из (a)-(e) ниже:
(a) нуклеиновая кислота, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-25 аминокислот в аминокислотной последовательности, показанной в SEQ ID NO:2 или 7;
(b) нуклеиновая кислота, которая гибридизуется в очень жестких условиях, включающих условия гибридизации 0,2 х SSC при 63°С и условия промывки 0,2 х SSC при 60-65°С, с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:1 или 6, и которая кодирует указанный белок;
(c) нуклеиновая кислота, которая состоит из нуклеотидной последовательности, обладающей 95% или более идентичностью с нуклеотидной последовательностью, состоящей из SEQ ID NO:1 или 6, и кодирует указанный белок;
(d) нуклеиновая кислота, которая кодирует белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7; и
(e) нуклеиновая кислота, которая гибридизуется в очень жестких условиях, включающих условия гибридизации 0,2 х SSC при 63°С и условия промывки 0,2 х SSC при 60-65°С, с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, показанной в SEQ ID NO:2 или 7, и которая кодирует указанный белок.
18. Белок, участвующий в преобразовании из 18:3(n-6)-PL в 18:3(n-6)-СоА и/или преобразовании из DGLA-CoA в DGLA-PL, выбранный из (а) или (b) ниже:
(a) белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-25 аминокислот в аминокислотной последовательности SEQ ID NO:2 или 7; или
(b) белок, состоящий из аминокислотной последовательности, обладающей идентичностью 95% или более с аминокислотной последовательностью, состоящей из SEQ ID NO:2 или 7.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| US 7198937 B2, 03.04.2007 | |||
| СПОСОБ ПРОИЗВОДСТВА КОНСЕРВОВ "СОЛЯНКА С МЯСОМ" | 2015 |
|
RU2577006C1 |
| ДОБАВКА ДЛЯ УЛУЧШЕНИЯ ОСНОВНОГО ВКУСА, СОДЕРЖАЩАЯ ВЫСОКОНЕНАСЫЩЕННУЮ ЖИРНУЮ КИСЛОТУ С ДЛИННОЙ ЦЕПЬЮ И/ИЛИ ЕЕ СЛОЖНЫЙ ЭФИР, И СОДЕРЖАЩАЯ ЕЕ КОМПОЗИЦИЯ НА ОСНОВЕ РАСТИТЕЛЬНОГО ЖИРА И МАСЛА | 2003 |
|
RU2333675C2 |

