ОБЛАСТЬ ТЕХНИКИ
Это описание изобретения притязает на приоритет заявки на патент Японии № 2007-181899 (поданной 11 июля 2007).
Настоящее изобретение относится к новым генам фосфатазы фосфатидной кислоты.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Жирные кислоты являются важными компонентами липидов, таких как фосфолипиды и триацилглицерины. Жирные кислоты, содержащие две или более ненасыщенных связей, в совокупности именуются полиненасыщенными жирными кислотами (PUFA) и, как известно, включают арахидоновую кислоту, дигомо-γ-линоленовую кислоту, эйкозапентаеновую кислоту и докозагексаеновую кислоту. Некоторые из этих полиненасыщенных жирных кислот не могут синтезироваться в организме животного, и такие полиненасыщенные жирные кислоты должны заимствоваться в качестве незаменимых жирных кислот из пищевых источников. Полиненасыщенные жирные кислоты широко распространены. В качестве примера, арахидоновую кислоту получают при разложении липидов, экстрагированных из надпочечников и/или печеней животных. Однако количество полиненасыщенных жирных кислот, содержащихся в органах животных, является небольшим, и их невозможно получить в достаточном количестве для больших запасов при простом экстрагировании или выделении из органов животных. По этой причине методы с использованием микроорганизмов были разработаны для получения полиненасыщенных жирных кислот путем культивирования различных микроорганизмов. Известно, что, в первую очередь, микроорганизмы рода Mortierella продуцируют липиды, содержащие арахидоновую кислоту и другие полиненасыщенные жирные кислоты.
Также предпринимались другие попытки для продуцирования полиненасыщенных жирных кислот в растениях. Полиненасыщенные жирные кислоты образуют запасные липиды, такие как триацилглицерины, и, как известно, аккумулируются в клетках микроорганизмов или семенах растений.
Триацилглицерины, являющиеся запасными липидами, продуцируются in vivo следующим образом. Перенос ацильной группы происходит на глицерол-3-фосфат с помощью глицерол-3-фосфат-ацилтрансферазы с образованием лизофосфатидной кислоты (LPA), на которую перенос ацильной группы далее происходит с помощью LPA-ацилтрансферазы с образованием фосфатидной кислоты. Эта фосфатидная кислота, в свою очередь, подвергается дефосфорилированию с помощью фосфатазы фосфатидной кислоты с образованием диацилглицерина, на который затем переносится ацильная группа с помощью диацилглицерол-ацилтрансферазы с образованием триацилглицерина.
В вышеприведенном каскаде реакций фосфатидная кислота (в дальнейшем также именуемая «РА» или «1,2-диацил-sn-глицерол-3-фосфат») служит не только в качестве предшественника триацилглицерина, но также в качестве предшественника для биосинтеза глицерофосфолипидов диацильного типа. В дрожжах или других микроорганизмах РА-цитидилтрансфераза действует на РА и цитидин-5'-трифосфат (СТР) с вызовом синтеза CDP-диацилглицерина (CDP-DG), который в свою очередь используется для биосинтеза различных фосфолипидов.
Как описано выше, известно, что дефосфорилирование РА при биосинтезе диацилглицерина (в дальнейшем также именуемого «DG») катализируется фосфатазой фосфатидной кислоты (Е.С. 3.1.3.4; в дальнейшем также именуемой «РАР»). Известно, что этот фермент РАР присутствует у всех организмов, простирающихся от бактерий до позвоночных животных. Кроме того, у грибов РАР является ключевым ферментом биосинтеза триацилглицеринов (которые являются запасными липидами), начинающегося с РА.
Известно, что дрожжи Saccharomyces cerevisiae, которые являются грибком, имеют два типа РАР (непатентный документ 1). Один фермент является Mg2+-зависимой РАР (РАР1), а другой фермент является Mg2+-независимой РАР (РАР2). В качестве гена, кодирующего первый фермент, был известен ген РАН1. Однако поскольку мутанты pah1Δ обладают РАР1-активностью, существуют другие возможные гены, ответственные за РАР1-активность. С другой стороны, в качестве генов, кодирующих РАР2, были известны гены DPP1 и LPP1, и они в значительной степени ответственны за РАР2-активность у дрожжей. Кодируемые этими генами ферменты имеют широкую субстратную специфичность и, как известно, действуют также на диацилглицеролпирофосфат (DGPP), лизофосфатидную кислоту, сфингооснование-фосфат и изопреноид-фосфат и т.д. с вызовом их дефосфорилирования.
В других грибах было подтверждено присутствие этих гомологов, но нет сообщения об их функции, и т.п.
Было известно, что в липидпродуцирующих грибах, Mortierella alpina, РАР-активностью обладает микросомальная фракция (непатентный документ 2).
Непатентный документ 1: Trends Biochem. Sci., 31(12), 694-699, 2006.
Непатентный документ 2: Biochemical Society Transactions, 28, 707-709, 2000.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ПРОБЛЕМЫ, КОТОРЫЕ ДОЛЖНЫ БЫТЬ РАЗРЕШЕНЫ НАСТОЯЩИМ ИЗОБРЕТЕНИЕМ
Однако ранее описанные гены РАР не были исследованы на их способность к изменению жирнокислотной характеристики композиций жирных кислот, продуцируемых клетками-хозяевами, в которые введены и в которых экспрессируются эти гены РАР. Существует потребность в идентификации нового гена, который позволяет продуцировать жиры и масла с требуемой жирнокислотной характеристикой и/или обогащением требуемыми жирными кислотами при введении в клетки-хозяева или экспрессии в них.
СПОСОБЫ РАЗРЕШЕНИЯ ПРОБЛЕМ
Целью настоящего изобретения является обеспечение белка или нуклеиновой кислоты, которые позволяют продуцировать жиры и масла с требуемой жирнокислотной характеристикой и/или обогащением требуемыми жирными кислотами при экспрессии в клетках-хозяевах или введении в них.
Для достижения вышеприведенной цели авторы настоящего изобретения выполнили большие объемы напряженных работ. Во-первых, был проведен анализ маркерных экспрессируемых последовательностей (EST) на липидпродуцирующем грибе, Mortierella alpina, для получения последовательностей, разделяющих высокую идентичность с известными генами РАР. Для получения полноразмерной открытой рамки считывания (ORF), кодирующей РАР, гены далее клонировали путем отбора из кДНК-библиотеки или ПЦР. В результате предпринятия попытки ввести эти гены в клетки-хозяева с высокой пролиферативной активностью (например, дрожжевые клетки) для продуцирования, тем самым, требуемой композиции жирной кислоты, авторы настоящего изобретения добились успеха в клонировании гена, относящегося к новой РАР, которая делает возможной продукцию композиции жирной кислоты, отличной от смеси, продуцируемой хозяином, не несущим этот ген. Это привело к завершению настоящего изобретения. То есть, настоящее изобретение представляет собой следующие изобретения:
(1) Нуклеиновую кислоту, включающую нуклеотидную последовательность, указанную в любом из подпунктов (а)-(е) ниже:
(а) нуклеотидную последовательность, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и обладающий активностью фосфатазы фосфатидной кислоты;
(b) нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(c) нуклеотидную последовательность, которая состоит из нуклеотидной последовательности, идентичной на 70% или больше нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты;
(d) нуклеотидную последовательность, которая кодирует аминокислотную последовательность, идентичную на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты; или
(e) нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты.
(2) Нуклеиновую кислоту в соответствии с (1) выше, которая включает нуклеотидную последовательность, указанную в любом из подпунктов (а)-(с) ниже:
(а) нуклеотидную последовательность, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-10 аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и обладающий активностью фосфатазы фосфатидной кислоты;
(b) нуклеотидную последовательность, которая поддается гибридизации в условиях использования 2 х SSC при 50οС с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты; или
(c) нуклеотидную последовательность, которая кодирует аминокислотную последовательность, идентичную на 90% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты.
(3) Нуклеиновую кислоту, включающую нуклеотидную последовательность, указанную в любом из подпунктов (а)-(с) ниже, или ее фрагмент:
(а) нуклеотидную последовательность, указанную в SEQ ID NO: 4;
(b) нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2; или
(с) нуклеотидную последовательность, указанную в SEQ ID NO: 1.
(4) Нуклеиновую кислоту, включающую нуклеотидную последовательность, указанную в любом из подпунктов (а)-(е) ниже:
(а) нуклеотидную последовательность, которая кодирует следующий белок:
белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
(b) нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует следующий белок:
белок, который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
(c) нуклеотидную последовательность, которая состоит из нуклеотидной последовательности, идентичной на 70% или больше нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует следующий белок:
белок, который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
(d) нуклеотидную последовательность, которая кодирует следующий белок:
белок, который состоит из аминокислотной последовательности, идентичной на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты; или
(e) нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и которая кодирует следующий белок:
белок, который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
(5) Нуклеиновую кислоту в соответствии с (4) выше, которая включает нуклеотидную последовательность, указанную в любом из подпунктов (а)-(с) ниже:
(а) нуклеотидную последовательность, которая кодирует следующий белок:
белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением 1-10 аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
(b) нуклеотидную последовательность, которая поддается гибридизации в условиях использования 2 х SSC при 50οС с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует следующий белок:
белок, который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты; или
(c) нуклеотидную последовательность, которая кодирует следующий белок:
белок, который состоит из аминокислотной последовательности, идентичной на 90% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты.
(6) Белок, указанный в подпункте (а) или (b) ниже:
(а) белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в SEQ ID NO: 2, и который обладает активностью фосфатазы фосфатидной кислоты; или
(b) белок, который состоит из аминокислотной последовательности, идентичной на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и который обладает активностью фосфатазы фосфатидной кислоты.
(7) Белок, указанный в подпункте (а) или (b) ниже:
(а) белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в SEQ ID NO: 2, и который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, состоящий из указанной выше аминокислотной последовательности, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты; или
(b) белок, который состоит из аминокислотной последовательности, идентичной на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и который обладает способностью к обеспечению жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей i)-vi), указанных ниже, в жирнокислотной характеристике хозяина, экспрессирующего белок, состоящий из указанной выше аминокислотной последовательности, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты.
(8) Белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2.
(9) Рекомбинантный вектор, включающий нуклеиновую кислоту в соответствии с любым из (1)-(5) выше.
(10) Трансформант, трансформированный рекомбинантным вектором в соответствии с (9) выше.
(11) Композиция жирной кислоты, полученная путем культивирования трансформанта в соответствии с (10) выше, в жирнокислотной характеристике которой по крайней мере один или несколько показателей i)-vi), указанных ниже, больше чем в жирнокислотной характеристике продукта культивирования, полученного путем культивирования хозяина, который не трансформирован рекомбинантным вектором в соответствии с (9) выше:
i) содержание олеиновой кислоты;
ii) отношение содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношение содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношение общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношение содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительный показатель содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты.
(12) Способ приготовления композиции жирной кислоты, который включает получение композиции жирной кислоты в соответствии с (11) выше из продукта культивирования, полученного путем культивирования трансформанта в соответствии с (10) выше.
(13) Пищевой продукт, включающий композицию жирной кислоты в соответствии с (11) выше.
ПРЕИМУЩЕСТВА НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
РАР настоящего изобретения предоставляет хозяину возможность продуцировать композицию жирной кислоты жирных кислот, жирнокислотная характеристика которой отличается от жирнокислотной характеристики композиции жирной кислоты, продуцируемой хозяином, не несущим РАР. В результате РАР настоящего изобретения делает возможным обеспечение липидов, имеющих требуемые свойства и действия, и является полезной при применении в пищевых продуктах, косметических средствах, мылах и т.п.
Кроме того, РАР настоящего изобретения позволяет увеличить способность к продукции жирных кислот и запасных липидов и, следовательно, является предпочтительной в качестве средства увеличения выработки полиненасыщенных жирных кислот в микроорганизмах и растениях.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 демонстрируется последовательность кДНК MaPAP1 в соответствии с настоящим изобретением вместе с ее расшифрованной аминокислотной последовательностью.
На фиг. 2 демонстрируется совмещение расшифрованной аминокислотной последовательности MaPAP1p (SEQ ID NO: 2) с аминокислотными последовательностями белков семейства РАР2. На этой фигуре три сегмента с двойным подчеркиванием представляют консенсусные области, консервативные среди ферментов семейства РАР2, а «*» представляет аминокислотный остаток, необходимый для РАР-активности.
ЛУЧШИЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым генам для фосфатазы фосфатидной кислоты, происходящим из рода Mortierella, характеризующейся дефосфорилированием фосфатидной кислоты с образованием диацилглицерина.
Фосфатаза фосфатидной кислоты (РАР) в настоящем изобретении является ферментом, катализирующим реакцию, в которой фосфатидная кислота подвергается дефосфорилированию с образованием диацилглицерина. Субстратом для РАР в настоящем изобретении является, как правило, но без ограничения, фосфатидная кислота.
Нуклеиновые кислоты настоящего изобретения, кодирующие фосфатазу фосфатидной кислоты
Фосфатаза фосфатидной кислоты (РАР) в настоящем изобретении включает MaPAP1. В таблице 1 ниже резюмировано соответствие между кДНК, CDS (кодирующей белок последовательностью), ORF и аминокислотными последовательностями нуклеиновой кислоты, кодирующей MaPAP1.

То есть, последовательности, относящиеся к РАР настоящего изобретения, включают SEQ ID NO: 2 (аминокислотную последовательность MaPAP1), SEQ ID NO: 4 (последовательность, являющуюся областью ORF MaPAP1), SEQ ID NO: 3 (последовательность, являющуюся областью CDS MaPAP1) и SEQ ID NO: 1 (нуклеотидную последовательность кДНК MaPAP1). Среди них SEQ ID NO: 3 соответствует нуклеотидам 105-1223 SEQ ID NO: 1, в то время как SEQ ID NO: 4 соответствует нуклеотидам 105-1220 SEQ ID NO: 1 или нуклеотидам 1-1116 SEQ ID NO: 3.
Нуклеиновые кислоты настоящего изобретения включают одноцепочечные и двухцепочечные ДНК, а также комплементарные им РНК, которые могут или иметь природное происхождение, или быть искусственно приготовлены. ДНК включают, но без ограничения, геномные ДНК, кДНК, соответствующие геномным ДНК, химически синтезированные ДНК, амплифицированные с помощью ПЦР ДНК, а также их комбинации и гибриды ДНК/РНК.
Предпочтительные варианты осуществления нуклеиновых кислот настоящего изобретения включают (а) нуклеотидную последовательность, указанную в SEQ ID NO: 4; (b) нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2; и (с) нуклеотидную последовательность, указанную в SEQ ID NO: 1.
Указанные выше нуклеотидная последовательность, указанная в SEQ ID NO: 4, нуклеотидная последовательность, кодирующая белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и нуклеотидная последовательность, указанная в SEQ ID NO: 1, являются последовательностями, продемонстрированными в таблице 1.
Для получения этих нуклеотидных последовательностей данные, касающиеся нуклеотидных последовательностей: EST или геномных ДНК из организмов, обладающих РАР-активностью, можно использовать для поиска нуклеотидной последовательности, кодирующей белок, обдающий высокой степенью идентичности с известными белками, обладающими РАР-активностью. Предпочтительными организмами, обладающими РАР-активностью, являются липидпродуцирующие грибы, включающие, но без ограничения, M. alpina.
Для анализа EST сначала готовят библиотеку кДНК. В отношении методов приготовления библиотеки кДНК можно привести ссылку на “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001)). В альтернативном случае можно использовать имеющийся в продаже набор для приготовления библиотеки кДНК. Методы приготовления библиотеки кДНК, подходящей для настоящего изобретения, являются, в качестве примера, следующими. А именно, соответствующий штамм M. alpina, липидпродуцирующего гриба, засевают в соответствующую среду и подвергают предварительному культивированию в течение соответствующего периода времени. Условия культивирования, подходящие для этого предварительного культивирования, включают, например, состав среды из 1,8% глюкозы, 1% дрожжевого экстракта и рН 6,0, период культивирования, составляющий 3 дня, и температуру культивированию, составляющую 28οС. Продукт предварительного культивирования затем подвергают основному культивированию при соответствующих условиях. Составом среды, подходящим для основного культивирования, может быть, например, 1,8% глюкозы, 1% соевого порошка, 0,1% оливкового масла, 0,01% Adekanol, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2 .2H2O, 0,05% MgCl2 .6H20 и рН 6,0. Условиями культивирования, подходящими для основного культивирования, могут быть, например, аэробное культивирование с постоянным перемешиванием при 300 оборотах в минуту, 1 vvm (отношение количества подводимого воздуха к емкости сосуда для культивирования), 26οC в течение 8 дней. Во время культивирования можно добавлять подходящее количество глюкозы. В соответствующие моменты времени во время основного культивирования отбирают образцы продукта культивирования, из которых затем собирают клетки для приготовления обшей РНК. Для приготовления общей РНК можно использовать любой известный метод, такой как метод с использованием гуанидина гидрохлорида/CsCl. Результирующую общую РНК можно обработать, используя имеющийся в продаже набор для очистки поли(А)+РНК. Кроме того, библиотеку кДНК можно приготовить с использованием имеющегося в продаже набора. Затем нуклеотидную последовательность любого клона из приготовленной таким образом библиотеки кДНК определяют, используя праймеры, которые сконструированы на основе вектора, для того чтобы сделать возможным определение нуклеотидной последовательности вставки. В результате можно получить EST. Например, при использовании для приготовления библиотеки кДНК набора для клонирования ZAP-cDNA GigapackIII Gold (Stratagene) можно выполнить прямое клонирование.
После анализа с помощью BLASTX оказалось, что ген MaPAP1 настоящего изобретения идентичен на 57,4% нуклеотидной последовательности, кодирующей происходящий из Neurospora crassa гомолог диацилглицеролпирофосфат-фосфатазы (DPP1) (входящий № CAD70721), имеющей наименьший Е-показатель (т.е. демонстрирующей наибольшую идентичность), и идентичен на 59,3% аминокислотной последовательности этого гомолога.
Настоящим изобретением также охватываются нуклеиновые кислоты, функционально эквивалентные нуклеиновой кислоте, включающей вышеприведенную нуклеотидную последовательность, указанную в SEQ ID NO:4 (в дальнейшем также именуемой «нуклеотидная последовательность настоящего изобретения»), или нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO:4 (в дальнейшем также именуемой «аминокислотная последовательность настоящего изобретения»). Предполагается, что выражение «функционально эквивалентный» означает, что белок, кодируемый нуклеотидной последовательностью настоящего изобретения, или белок, состоящий из аминокислотной последовательности настоящего изобретения, обладает РАР-активностью. Помимо этой РАР-активности белок, кодируемый нуклеотидной последовательностью настоящего изобретения, или белок, состоящий из аминокислотной последовательности настоящего изобретения, может обладать способностью к обеспечению жирнокислотной характеристики, в котором гарантируется большая степень по крайней мере одного или нескольких показателей:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
в жирнокислотной характеристике хозяина, экспрессирующего белок, чем в жирнокислотной характеристике хозяина, не экспрессирующего этот белок (такой белок в дальнейшем также именуется «белком, обладающим способностью к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении»).
Конкретным примером является нуклеиновая кислота, включающая нуклеотидную последовательность, кодирующую белок, обладающий способностью к обеспечению жирнокислотной характеристики, которая удовлетворяет по крайней мере одному или нескольким следующим условиям:
i) содержание олеиновой кислоты составляет 46% или больше, предпочтительно 48% или больше, 50% или больше, 52% или больше, 54% или больше;
ii) отношение содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты составляет 7,0, предпочтительно 8,0 или больше, 8,5% или больше;
iii) отношение содержания олеиновой кислоты к содержанию пальмитиновой кислоты составляет 7,5 или больше, предпочтительно 8,0 или больше, 10,0 или больше, 12,0 или больше, 13,0 или больше;
iv) отношение общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты составляет 8,0 или больше, предпочтительно 10,0 или больше, 12,0 или больше, 14,0 или больше, 14,5 или больше;
v) отношение содержания жирных кислот С18 к содержанию жирных кислот С16 составляет 1,1 или больше, предпочтительно 1,2 или больше, 1,3 или больше, 1,4 или больше; и
vi) содержание арахидоновой кислоты составляет 0,45 или больше, предпочтительно 0,50 или больше, 0,53 или больше, 0,55 или больше, содержание дигомо-γ-линоленовой кислоты составляет 0,34 или больше, предпочтительно 0,40 или больше, 0,42 или больше, 0,45 или больше, и/или содержание γ-линоленовой кислоты составляет 0,55 или больше, предпочтительно 0,65 или больше, 0,67 или больше, 0,70 или больше, 0,75 или больше (все вышеприведенные числа указывают на отношение к общему содержанию жирных кислот, когда ген РАР настоящего изобретения экспрессируется в продуцирующих арахидоновую кислоту дрожжевых клетках),
когда указанная выше нуклеотидная последовательность настоящего изобретения встроена в экспрессионный вектор pYE22m (Biosci. Biotech. Biochem., 59. 1221-1228, 1995) и трансформирована в являющегося дрожжами хозяина, штамм EH13-15 Saccharomyces cerevisiae (Appl. Microbiol. Biotechnol., 30, 515-520, 1989), и результирующий трансформант культивируют для сбора клеток, которые затем анализируют на жирные кислоты с помощью способов, описанных в примере 6 ниже. Более предпочтительной является нуклеиновая кислота, включающая нуклеотидную последовательность, кодирующую белок, обладающий как РАР-активностью, так и указанной выше способностью к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении.
Одним из характерных признаков композиции жирной кислоты настоящего изобретения является высокое содержание арахидоновой кислоты. Арахидоновая кислота, субстрат, представляемый химической формулой С20Н32О2 и имеющий молекулярную массу 304,47, является карбоновой кислотой, содержащей 20 атомов углерода и имеющей 4 двойные связи ([20:4(n-6]), и относящейся к члену (n-6)-рядов. Арахидоновая кислота присутствует в качестве важного фосфолипида (в частности, фосфатидилэтаноламина, фосфатидилхолина, фосфатидилинозита) в мембранах клеток животных и содержится в большом количестве в головном мозге. Кроме того, арахидоновая кислота служит в качестве исходного материала для ряда эйкозаноидов (например, простагландина, тромбоксана, лейкотриена), образуемых с помощью каскада реакций с вовлечением арахидоновой кислоты, и также важна в качестве вторичного мессенджера при межклеточной передаче сигналов. С другой стороны, арахидоновая кислота синтезируется из линолевой кислоты в организме животного. Однако, у некоторых животных, в зависимости от их вида и возраста, эта функция не проявляется в степени, достаточной для продуцирования требуемого количества арахидоновой кислоты, или функция продуцирования арахидоновой кислоты отсутствует. Поэтому арахидоновая кислота должна заимствоваться из пищевых источников и может считаться незаменимой жирной кислотой.
Содержание арахидоновой кислоты в композиции жирной кислоты настоящего изобретения можно определить, в качестве примера, следующим образом. А именно, плазмиду для РАР настоящего изобретения встраивают в такой вектор, как pDuraSC или pDura5MCS, описанный в примере 8, и трансформируют в штамм M. alpina. Полученному в результате трансформанту предоставляют возможность экспрессироваться, и его культивируют в соответствии со способами, описанными в примере 8. Полученные таким образом прокультивированные клетки используют для определения содержания жирных кислот в клетках и/или содержания арахидоновой кислоты на единицу среды и т.п. Для анализа содержания арахидоновой кислоты и т.п., например, жирные кислоты в результирующих прокультивированных клетках превращают в метиловые эфиры соответствующих жирных кислот с помощью метода с использованием соляной кислоты/метанола и затем экстрагируют гексаном. После отгонки гексана жирные кислоты анализируют с помощью газовой хроматографии. Согласно этому анализу установлено, что M. alpina, трансформированный РАР настоящего изобретения, демонстрирует не только высокое содержание жирных кислот в клетках, но также высокую продукцию арахидоновой кислоты на единицу среды. Таким образом, композиция жирной кислоты настоящего изобретения, имеющая высокое содержание арахидоновой кислоты, является предпочтительной, поскольку она делает возможным эффективное пополнение арахидоновой кислоты.
Другим характерным признаком композиции жирной кислоты настоящего изобретения является высокое содержание дигомо-γ-линоленовой кислоты. Дигомо-γ-линоленовая кислота (DGLA), субстрат, представляемый химической формулой С20Н34О2 и имеющий молекулярную массу 306,48, является карбоновой кислотой, содержащей 20 атомов углерода и 3 двойные связи ([20:3(n-6]), и относится к члену (n-6)-рядов. DGLA образуется при удлинении γ-линоленовой кислоты ([18:3(n-6]). При добавлении одной или более двойных связей в DGLA образуется арахидоновая кислота.
Еще одним характерным признаком композиции жирной кислоты настоящего изобретения является высокое содержание γ-линоленовой кислоты. γ-Линоленовая кислота, субстрат, представляемый химической формулой С18Н30О2 и имеющий молекулярную массу 278,436, является карбоновой кислотой, содержащей 18 атомов углерода и 3 двойные связи ([18:3(n-6]) и относящейся к члену (n-6)-рядов. Люди обладают способностью к продукции γ-линоленовой кислоты из линолевой кислоты ([18:2(n-6]) через катализируемую Δ6-десатуразой дегидрогенизацию, но они часто получают ее из пищевых продуктов.
Такие нуклеиновые кислоты, которые функционально эквивалентны нуклеиновым кислотам настоящего изобретения, включают нуклеиновую кислоту, включающую нуклеотидную последовательность, указанную в любом из подпунктов (а)-(е) ниже. Следует отметить, что при использовании для описания нуклеотидных последовательностей, перечисленных ниже, выражение «указанная выше активность настоящего изобретения», как предполагается, означает «РАР-активность и/или способность к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении», как определено выше.
(а) Нуклеотидная последовательность, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и обладающий указанной выше активностью настоящего изобретения.
Нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, включают нуклеотидную последовательность, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и обладающий указанной выше активностью настоящего изобретения.
Конкретнее, она является нуклеотидной последовательностью, которая кодирует белок, состоящий из
(i) аминокислотной последовательности с делецией одной или нескольких (предпочтительно одной или нескольких (например, 1-110, 1-100, 1-75, 1-50, 1-30, 1-25, 1-20, 1-15, 1-10, более предпочтительно 1-5)) аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2;
(ii) аминокислотной последовательности с заменой другими аминокислотами одной или нескольких (предпочтительно одной или нескольких (например, 1-110, 1-100, 1-75, 1-50, 1-30, 1-25, 1-20, 1-15, 1-10, более предпочтительно 1-5)) аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2;
(iii) аминокислотной последовательности с добавлением другой одной или нескольких (предпочтительно одной или нескольких (например, 1-110, 1-100, 1-75, 1-50, 1-30, 1-25, 1-20, 1-15, 1-10, более предпочтительно 1-5)) аминокислот в аминокислотную последовательность, указанную в SEQ ID NO: 2; или
(iv) аминокислотной последовательности с любой комбинацией модификаций (i)-(iii), указанных выше, и обладающий указанной выше активностью настоящего изобретения.
Среди указанных выше модификаций замена предпочтительно является консервативной, что означает замену определенного аминокислотного остатка другим остатком, обладающим схожими физическими и химическими характеристиками. Она может быть любой заменой, если она значительно не изменяет структурные характеристики исходной последовательности. Например, возможна любая замена, если замененные аминокислоты не разрушают спиральную структуру исходной последовательности или не разрушают любой другой тип вторичной структуры, характеризующей исходную последовательность.
Консервативную замену обычно вводят при синтезе в биологических системах или химическом пептидном синтезе, предпочтительно при химическом пептидном синтезе. В этом случае замены могут включать не встречающиеся в природе аминокислотные остатки, а также пептидомиметики и инвентированные формы аминокислотных последовательностей, в которых незамещенные области инвертированы.
Аминокислотные остатки классифицированы на следующие группы взаимозаменяемых членов, но без ограничения, и перечислены ниже:
группа A: лейцин, изолейцин, норлейцин, валин, норвалин, аланин, 2-аминомасляная кислота, метионин, О-метилсерин, трет-бутилглицин, трет-бутилаланин и циклогексилаланин;
группа B: аспарагиновая кислота, глютаминовая кислота, изоаспарагиновая кислота, изоглютаминовая кислота, 2-аминоадипиновая кислота и 2-аминосубериновая кислота;
группа C: аспарагин и глютамин;
группа D: лизин, аргинин, орнитин, 2,4-диаминомасляная кислота и 2,3-диаминопропионовая кислота;
группа E: пролин, 3-гидроксипролин и 4-гидроксипролин;
группа F: серин, треонин и гомосерин; и
группа G: фенилаланин и тирозин.
Неконсервативная замена может включать обмен члена одного из указанных выше классов на член из другого класса. В этом случае с целью сохранения биологических функций белков настоящего изобретения предпочтительно, когда во внимание принимают индекс гидрофобности аминокислот (Kyte et al., J. Mol. Biol., 157: 105-131 (1982)).
В случае неконсервативной замены замены аминокислот можно также выполнить на основе гидрофильности.
В описании и чертежах настоящей заявки нуклеотиды, аминокислоты и их сокращения являются нуклеотидами, аминокислотами и их сокращениями, установленными Комиссией по номенклатуре биохимических соединений IUPAC-IUB, или теми, которые обычно используются в данной области техники, например, как описано в Immunology-A Synthesis (второе издание, под редакцией E.S. Golub и D.R. Gren, Sinauer Associates, Sunderland, Massachusetts (1991)). Кроме того, предполагается, что аминокислоты, которые могут иметь оптические изомеры, являются их L-изомером, кроме особо оговоренных случаев.
Стереоизомеры (например, D-аминокислоты) указанных выше аминокислот, не встречающихся в природе аминокислот, таких как α,α-двузамещеннные аминокислоты, N-алкиламинокислоты, молочная кислота и другие необычные аминокислоты, могут также быть элементами, составляющими белки настоящего изобретения.
Следует отметить, что в используемом здесь символическом представлении белков направление влево является направлением к амино-концу, а направление вправо является направлением к карбоксильному концу, в соответствии со стандартным использованием и соглашением.
Подобным образом, кроме особо оговоренных случаев, левый конец одноцепочечных полинуклеотидных последовательностей является 5'-концом, а направление влево в двухцепочечных полинуклеотидных последовательностях именуется 5'-направлением.
Квалифицированные в данной области техники специалисты могли бы сконструировать и приготовить подходящие мутанты описанных здесь белков, используя известные в данной области техники методы. Например, при определении в качестве цели области, которая, по-видимому, менее важна для биологической активности белка настоящего изобретения, можно идентифицировать подходящую область в молекуле белка, структуру которой можно изменить без нарушения биологической активности белка настоящего изобретения. Можно также идентифицировать остатки или области в молекуле, которые являются консервативными между схожими белками. Кроме того, можно ввести консервативные аминокислотные замены в область, которая, по-видимому, важна для биологической активности или структуры белка настоящего изобретения, без нарушения биологической активности или без неблагоприятного воздействия на полипептидную структуру белка. В частности в настоящем изобретении, как указано с помощью двойного подчеркивания на фиг. 2, аминокислотная последовательность (SEQ ID NO: 2) MaPAP1 настоящего изобретения содержит три консенсусные области, соответствующие остаткам 146-154, 202-205 и 260-264, консервативные среди ферментов семейства Mg2+-независимых фосфатаз фосфатидной кислоты (РАР2). В этих трех консенсусных областях, консервативных среди ферментов семейства РАР2, аргинин в домене 1 и гистидины в доменах 2 и 3 известны в качестве аминокислот, необходимых для активности, и эти аминокислоты являются также консервативными аминокислотами в MaPAP1 настоящего изобретения, представленными аргинином в положении 153 и гистидинами в положениях 205 и 260 SEQ ID NO: 2. Указанные выше консенсусные области являются необходимыми для ферментов семейства РАР2, а также важны для РАР настоящего изобретения. Таким образом, мутанты в соответствии с настоящим изобретением никоим образом не ограничиваются, пока сохранены указанные выше консенсусные области.
Квалифицированные в данной области техники специалисты могли бы провести так называемый структурно-функциональный анализ, с помощью которого идентифицируют остатки, в белке настоящего изобретения и в схожем с ним пептиде, которые важны для биологической активности или структуры, и сравнивают аминокислотные остатки между этими двумя пептидами, что позволяет, тем самым, предсказать, какие остатки в белке, схожем с белком настоящего изобретения, являются аминокислотными остатками, соответствующими аминокислотным остаткам, важным для биологической активности или структуры. Кроме того, для предсказанных таким образом аминокислотных остатков можно выбрать замены химически схожими аминокислотами с отбором, тем самым, мутанта, который сохраняет биологическую активность белка настоящего изобретения. Также квалифицированные в данной области техники специалисты могли бы также проанализировать трехмерную структуру и аминокислотную последовательность этого мутантного белка. Полученные таким образом результаты анализа можно в дальнейшем использовать для предсказания расположения аминокислотных остатков относительно трехмерной структуры белка. Поскольку аминокислотные остатки, которые согласно предсказанию находятся на поверхности белка, могут быть вовлечены в важные взаимодействия с другими молекулами, квалифицированные в данной области техники специалисты на основе вышеотмеченных результатов анализа могли бы получить мутант, который не обусловлен заменой тех аминокислотных остатков, которые согласно предсказанию находятся на поверхности белка. Кроме того, квалифицированные в данной области техники специалисты могли бы также получить мутант, имеющий одну аминокислотную замену для любого из аминокислотных остатков, составляющих белок настоящего изобретения. Эти мутанты можно подвергнуть скринингу с помощью любого известного анализа для сбора информации об отдельных мутантах, которая в свою очередь позволяет оценить полезность отдельных аминокислотных остатков, составляющих белок настоящего изобретения, при осуществлении сравнения со случаем, в котором мутант, имеющий замену конкретного аминокислотного остатка, проявляет биологическую активность, меньшую биологической активности белка настоящего изобретения, в котором такой мутант не проявляет биологическую активность, или в котором такой мутант проявляет неподходящую активность, ингибирующую биологическую активность белка настоящего изобретения. Кроме того, на основе информации, собранной из таких обычных экспериментов, квалифицированные в данной области техники специалисты могут легко проанализировать аминокислотные замены, нежелательные в мутантных белках настоящего изобретения, или отдельно, или в комбинации с другими мутациями.
Описанный выше белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, можно приготовить в соответствии с такими методами, как сайт-направленный мутагенез, описанный в “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001)), “Current Protocols in Molecular Biology” (John Wiley & Sons (1987-1997), Kunkel (1985) Proc. Natl. Acad. Sci. USA 82: 488-492, и Kunkel (1988) Method Enzymol. 85: 2763-2766. Приготовление мутанта с такой мутацией, как делеция, замена или добавление аминокислот, можно выполнить, например, с помощью известных способов, таких как способ Kunkel или способ с использованием дуплекса с брешью, используя набор для введения мутаций на основе сайт-направленного мутагенеза, такой как набор для сайт-направленного мутагенеза QuikChangeTM (Stratagene), систему для сайт-направленного мутагенеза GeneTailorTM (Invitrogen) или систему для сайт-направленного мутагенеза TaKaRa (например, Mutan-K, Mutan-Super Express Km; Takara Bio Inc., Япония).
Методы, позволяющие делетировать, заменить или добавить одну или несколько аминокислот в аминокислотных последовательностях белков при сохранении их активности, включают вышеотмеченный сайт-направленный мутагенез, а также другие методы, например, методы обработки гена мутагеном и методы, в которых ген избирательно расщепляют для удаления, замены или добавления выбранного нуклеотида или нуклеотидов, а затем подвергают лигированию.
Предпочтительной нуклеотидной последовательностью, содержащейся в нуклеиновых кислотах настоящего изобретения, является нуклеотидная последовательность, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-10 аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, и обладающий РАР-активностью.
Кроме того, нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, также предпочтительно включают нуклеотидную последовательность, которая кодирует белок, состоящий из аминокислотной последовательности с делецией, заменой или добавлением 1-10 аминокислот в SEQ ID NO: 2 и обладающий указанной выше активностью настоящего изобретения.
Число или места мутаций или модификаций аминокислот в белке настоящего изобретения не ограничены, пока результирующий мутант сохраняет РАР-активность или способность к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении.
РАР-активность в настоящем изобретении или способность к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении, можно определить известным способом. Например, можно привести ссылку на следующий документ: J. Biol. Chem., 273, 14331-14338 (1988).
«РАР-активность» в настоящем изобретении можно определить, в качестве примера, следующим образом. Микросомальную фракцию получают из дрожжевых клеток, трансформированных для экспрессии РАР настоящего изобретения, как описано, например, в J. Bacteriology, 173, 2026-2034 (1991). К реакционному раствору, содержащему 0,5 мМ фосфатидную кислоту и 10 мМ 2-меркаптоэтанол в 50 мМ Tris-HCl (pH 7,5), затем добавляют указанную выше микросомальную фракцию, и реакцию осуществляют при 28οС в течение соответствующего периода времени. Для остановки реакции добавляют смесь хлороформ:метанол, а затем осуществляют экстракцию липидов. Получаемые в результате липиды подвергают фракционированию с помощью тонкослойной хроматографии или других методов, посредством чего можно определить количество образованного диацилглицерина.
Подобным образом, «способность к обеспечению жирнокислотной характеристики, обеспечиваемой РАР», в настоящем изобретении можно определить, в качестве примера, следующим образом. К лиофилизированным клеткам, полученным способом настоящего изобретения для приготовления композиции жирной кислоты, добавляют смесь хлороформ:метанол в соответствующей пропорции и перемешивают, а затем подвергают тепловой обработке в течение соответствующего периода времени. Далее выполняют центрифугирование для отделения клеток и сбора растворителя. Эту процедуру повторяют несколько раз. Затем липиды высушивают подходящим образом, и для растворения липидов добавляют растворитель, такой как хлороформ. Соответствующую аликвоту этого образца обрабатывают с помощью метода с использованием соляной кислоты/метанола для превращения жирных кислот в клетках в соответствующие метиловые эфиры, а затем подвергают экстракции гексаном. После отгонки гексана жирные кислоты анализируют с помощью газовой хроматографии.
(b) Нуклеотидная последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, включают нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения. SEQ ID NO: 4 и РАР-активность являются описанными выше последовательностью и активностью.
Для получения указанной выше нуклеотидной последовательности зонд можно приготовить из соответствующего фрагмента известным квалифицированным в данной области техники специалистам способом, и этот зонд можно использовать в известных методах гибридизации, таких как гибридизация с НК колоний, гибридизация с НК бляшек или Саузерн-блоттинг, для получения нуклеотидной последовательности из библиотеки кДНК, геномной библиотеки или т.п.
В отношении детализированных процедур для методов гибридизации можно привести ссылку на “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001), в частности, части 6-7), “Current Protocols in Molecular Biology” (John Wiley & Sons (1987-1997), в частности, части 6.3-6.4), “DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.” (Oxford University (1995), в частности часть 2.10 в отношении условий гибридизации).
Эффективность гибридизации определяется главным образом условиями гибридизации, более предпочтительно условиями гибридизации и условиями отмывки. Предполагается, что используемый здесь термин «жесткие условия» включает умеренно или в высокой степени жесткие условия.
Конкретнее, умеренно жесткие условия включают, например, условия гибридизации в 1 х SSC - 6 x SSC при 42°С-55°С, более предпочтительно в 1 х SSC - 3 x SSC при 45°С-50°С и наиболее предпочтительно в 2 х SSC при 50°С. В некоторых случаях, например, когда раствор для гибридизации содержит приблизительно 50% формамида, используют температуру, которая на 5°С-15°С ниже указанной выше температуры. Условиями отмывки могут быть 0,5 х SSC - 6 x SSC при 40°С-60°С. Во время гибридизации и отмывки можно обычно добавить 0,05%-0,2% SDS, предпочтительно 0,1% SDS.
В высокой степени жесткие условия включают гибридизацию и/или отмывку при более высокой температуре и/или более низкой концентрации соли по сравнению с умеренно жесткими условиями. Например, условиями гибридизации могут быть 0,1 х SSC - 2 x SSC при 55°С-65°С, более предпочтительно 0,1 х SSC - 1 x SSC при 60°С-65°С и наиболее предпочтительно 0,2 х SSC при 63°С. Условиями отмывки могут быть 0,2 х SSC - 2 x SSC при 50°С-68°С и более предпочтительно 0,2 х SSC при 60°С-65°С.
Условия гибридизации, используемые, в частности, в настоящем изобретении, включают, но без ограничения, предварительную гибридизацию в 5 х SSC, 1% SDS, 50 мМ Tris-HCl (pH 7,5) и 50% формамида при 42οС, инкубацию в течение ночи при 42°С в присутствии зонда для формирования гибридов и последующие три отмывки в 0,2 х SSC, 0,1% SDS при 65°С в течение 20 минут.
Можно также использовать имеющийся в продаже набор для гибридизации, в котором не используется радиоактивное вещество в качестве зонда. Конкретные примеры включают гибридизацию с использованием набора для обнаружения нуклеиновых кислот, меченных DIG, (Roche Diagnostics) или системы прямого мечения и обнаружения ECL (Amersham).
Предпочтительной нуклеотидной последовательностью, находящейся в пределах настоящего изобретения, является нуклеотидная последовательность, которая поддается гибридизации в условиях использования 2 х SSC при 50°С с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий РАР-активностью.
(c) Нуклеотидная последовательность, которая состоит из нуклеотидной последовательности, идентичной на 70% или больше нуклеотидной последовательности, состоящей из SEQ ID NO: 4, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, включают нуклеотидную последовательность, которая состоит из нуклеотидной последовательности, идентичной на по крайней мере 70% или больше последовательности нуклеиновой кислоты, указанной в SEQ ID NO: 4, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Предпочтительные примеры включают нуклеиновые кислоты, включающие нуклеотидную последовательность, которая идентична на по крайней мере 75%, более предпочтительно 80% (например, 85% или более, даже более предпочтительно 90% или более, конкретнее 95%, 98% или 99%) последовательности нуклеиновой кислоты, указанной SEQ ID NO: 4, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Идентичность в процентах между двумя последовательностями нуклеиновых кислот можно определить с помощью визуального исследования и математического расчета, или более предпочтительно используя компьютерную программу для сравнения информации о последовательностях двух нуклеиновых кислот. Компьютерные программы для сравнения последовательностей включают, например, версию 2.2.7 программы BLASTN (Altschul et al. (1990) J. Mol. Biol. 215: 403-410), доступную для использования на Web-сайте National Library of Medicine: http://www.ncbi.nlm.nih.gov/blast/bl2seq/bls.html, или алгоритм WU-BLAST 2.0. Стандартные устанавливаемые по умолчанию параметры для WU-BLAST 2.0 описаны на следующем Интернет-сайте: http://blast.wustl.edu.
(d) Нуклеотидная последовательность, которая кодирует аминокислотную последовательность, идентичную на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, включают нуклеотидную последовательность, которая кодирует аминокислотную последовательность, идентичную на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения. Белками, кодируемыми нуклеиновыми кислотами настоящего изобретения, могут также быть белки, имеющие идентичность с аминокислотной последовательностью MaPAP1, если они функционально эквиваленты белкам, обладающим указанной выше активностью настоящего изобретения.
Конкретные примеры включают аминокислотные последовательности, которые идентичны на 75% или больше, предпочтительно 80% или больше, более предпочтительно 85%, даже более предпочтительно 90% (например, 95%, более предпочтительно 98%) аминокислотной последовательности, указанной в SEQ ID NO: 2.
Предпочтительной нуклеотидной последовательностью, содержащейся в нуклеиновых кислотах настоящего изобретения, является нуклеотидная последовательность, которая кодирует аминокислотную последовательность, идентичную на 90% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения. Более предпочтительной является нуклеотидная последовательность, которая кодирует аминокислотную последовательность, идентичную на 95% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Идентичность в процентах между двумя аминокислотными последовательностями можно определить с помощью визуального исследования и математического расчета. В альтернативном случае идентичность в процентах можно определить, используя компьютерную программу. Примеры такой компьютерной программы включают BLAST, FASTA (Altschul et al., J. Mol. Biol. 215: 403-410 (1990), и ClustalW. В частности, различные условия (параметры) для исследования идентичности с использованием программы BLAST описываются Altschul и др.. (Nucl. Acids Res., 25, p. 3389-3402, 1997) и общедоступны на Web-сайте National Center for Biotechnology Information (NCBI) или DNA Data Bank of Japan (DDBJ) (BLAST Manual, Altschul et al., NCB/NLM/NIH Bethesda, MD 20894; Altschul et al.). Для определения идентичности в процентах можно также использовать такую программу, как программное обеспечение для обработки генетической информации GENETYX версии 7 (Genetyx Corporation, Япония), DINASIS Pro (Япония) или Vector NTI (Infomax).
Определенные методы совмещения аминокислотных последовательностей могут также приводить к совмещению последовательностей в специфической небольшой области, и также можно выявить район с очень высокой идентичностью последовательностей в такой небольшой совмещенной области, даже когда нет значительного родства между использованными полноразмерными последовательностями. Кроме того, в алгоритме BLAST используется матрица замен аминокислот BLOSUM62, и оптимальными параметрами, которые можно использовать, являются следующие параметры: (А) включение фильтра для экранирования сегментов последовательности запроса, которые имеют низкий уровень композиционной сложности, (определяемых с помощью программы SEG Wootton и Federhen (Computer and Chemistry, 1993); смотри также Wootton и Federhen, 1996, “Analysis of compositionally biased regions in sequence databases”, Methods Enzymol., 266: 554-571), или сегментов, состоящих из внутренних повторов с небольшой регулярностью (определяемых с помощью программы XNU Claverie и States (Computers and Chemistry, 1993)), и (В) пороговый уровень статистической значимости для сообщений о совмещениях с последовательностями из базы данных, или Е-показатель (ожидаемая вероятность совмещений, обнаруживаемых чисто случайно, в соответствии со стохастической моделью Karlin и Altschul, 1990; если статистическая значимость, приписанная совмещению, не превышает этот пороговый уровень Е-показателя, сообщения о совмещении не будет).
e) Нуклеотидная последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, включают нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Такой белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и условия гибридизации описаны выше. Нуклеотидные последовательности, содержащиеся в нуклеиновых кислотах настоящего изобретения, включают нуклеотидную последовательность, которая поддается гибридизации в жестких условиях с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и которая кодирует белок, обладающий указанной выше активностью настоящего изобретения.
Нуклеиновые кислоты настоящего изобретения также включают нуклеиновую кислоту, которая включает нуклеотидную последовательность с делецией, заменой или добавлением одного или нескольких нуклеотидов в нуклеотидной последовательности, состоящей из в SEQ ID NO: 4 и кодирующей белок, обладающий указанной выше активностью настоящего изобретения. Конкретнее, можно также использовать нуклеиновую кислоту, которая включает нуклеотидную последовательность, выбираемую из
(i) нуклеотидной последовательности с делецией одного или нескольких (предпочтительно одного или нескольких (например, 1-330, 1-300, 1-250, 1-200, 1-150, 1-100, 1-50, 1-30, 1-25, 1-20, 1-15, 1-10, более предпочтительно 1-5)) нуклеотидов в нуклеотидной последовательности, указанной в SEQ ID NO: 4;
(ii) нуклеотидной последовательности с заменой другими нуклеотидами одного или нескольких (предпочтительно одного или нескольких (например, 1-330, 1-300, 1-250, 1-200, 1-150, 1-100, 1-50, 1-30, 1-25, 1-20, 1-15, 1-10, более предпочтительно 1-5)) нуклеотидов в нуклеотидной последовательности, указанной в SEQ ID NO: 4;
(iii) нуклеотидной последовательности с добавлением другого одного или нескольких (предпочтительно одного или нескольких (например, 1-330, 1-300, 1-250, 1-200, 1-150, 1-100, 1-50, 1-30, 1-25, 1-20, 1-15, 1-10, более предпочтительно 1-5)) нуклеотидов в нуклеотидную последовательность, указанную в SEQ ID NO: 4; или
(iv) нуклеотидной последовательности с любой комбинацией модификаций (i)-(iii), указанных выше, и кодирующей белок, обладающий указанной выше активностью настоящего изобретения.
Предпочтительные варианты нуклеиновых кислот настоящего изобретения также включают нуклеиновую кислоту, включающую нуклеотидную последовательность, указанную в любом из подпунктов (а)-(с) ниже, или ее фрагмент:
(а) нуклеотидную последовательность, указанную в SEQ ID NO: 4;
(b) нуклеотидную последовательность, кодирующую белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2; или
(с) нуклеотидную последовательность, указанную в SEQ ID NO: 1.
Указанные выше (а) нуклеотидная последовательность, указанная в SEQ ID NO: 4, (b) нуклеотидная последовательность, кодирующая белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и (с) нуклеотидная последовательность, указанная в SEQ ID NO: 1, являются последовательностями, продемонстрированными в таблице 1. Фрагменты этих последовательностей, включающие содержащиеся в вышеприведенных нуклеотидных последовательностях области, т.е. ORF, CDS, биологически активную область, область, используемую в качестве праймера, как описывается позже, и область, которая может служить в качестве зонда, могут или иметь природное происхождение, или быть приготовлены искусственно.
Являющиеся фосфатазой фосфатидной кислоты белки настоящего изобретения
Белки настоящего изобретения, которые могут или иметь природное происхождение, или быть приготовлены искусственно, включают белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, и белки, функционально эквивалентные этому белку. Такой белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2, является белком, описанным выше. Предполагается, что «функционально эквивалентные белки» означают белки, обладающие «указанной выше активностью настоящего изобретения», как объяснено в разделе «Нуклеиновые кислоты настоящего изобретения, кодирующие фосфатазу фосфатидной кислоты», представленном выше.
В настоящем изобретении белки, функционально эквивалентные белку, состоящему из аминокислотной последовательности, указанной в SEQ ID NO: 2, включают белок, указанный в (а) или (b) ниже:
(а) белок, который состоит из аминокислотной последовательности с делецией, заменой или добавлением одной или нескольких аминокислот в SEQ ID NO: 2, и который обладает указанной выше активностью настоящего изобретения; или
(b) белок, который состоит из аминокислотной последовательности, идентичной на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, и который обладает указанной выше активностью настоящего изобретения.
Среди вышеупомянутых, аминокислотная последовательность с делецией, заменой или добавлением одной или нескольких аминокислот в SEQ ID NO: 2 или аминокислотная последовательность, идентичная на 70% или больше аминокислотной последовательности, состоящей из SEQ ID NO: 2, является последовательностью, пояснения в отношении которой даны в приведенном выше разделе «Нуклеиновые кислоты настоящего изобретения, кодирующие фосфатазу фосфатидной кислоты». Предполагается, что выражение «белок, который обладает указанной выше активностью настоящего изобретения», также включает мутанты белка, кодируемого нуклеиновой кислотой, включающей нуклеотидную последовательность SEQ ID NO: 4, или мутированные белки с различными модификациями, такими как замена, делеция или добавление одной или нескольких аминокислот в аминокислотной последовательности, указанной в SEQ ID NO: 2, а также модифицированные формы этих белков, боковые цепи аминокислот которых или т.п. модифицированы, и их формы, слитые с другими белками, если эти белки обладают РАР-активностью и/или способностью к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении.
Белки настоящего изобретения могут также быть приготовлены искусственно с помощью методов химического синтеза, таких как метод с использованием Fmoc-защиты (метод с использованием флуоренилметилоксикарбонильной зашиты) и метод с использованием tBoc-защиты (метод с использованием трет-бутилоксикарбонильной защиты). Кроме того, для химического синтеза можно использовать пептидные синтезаторы, доступные от Advanced ChemTech, Perkin Elmer, Pharmacia, Protein Technology Instrument, Synthecell-Vega, PerSeptive, Shimadzu Corporation (Япония) или других производителей.
Клонирование нуклеиновых кислот, кодирующих РАР
Кодирующие РАР нуклеиновые кислоты настоящего изобретения можно клонировать, например, путем отбора из библиотеки кДНК, используя подходящий зонд. Их можно также клонировать путем амплификации с помощью ПЦР с использованием подходящих праймеров и последующего лигирования в подходящий вектор. Полученные таким образом клоны можно в дальнейшем субклонировать в другой вектор.
Например, можно использовать имеющиеся в продаже плазмидные векторы, включающие pBlue-ScriptTM SK(+) (Stratagene), pGEM-T (Promega), pAmp (TM: Gibco-BRL), p-Direct (Clontech) и pCR2.1-TOPO (Invitrogen). В случае использования амплификации с помощью ПЦР праймерами могут быть любые области нуклеотидной последовательности, указанной, например, в SEQ ID NO: 1. В качестве примера, можно использовать следующие праймеры из SEQ ID NO: 1:
D1: 5'-CATGGGTTGCTTCGCGCGCAAGACG-3' (SEQ ID NO: 5) в качестве прямого праймера, и
D2: 5'-CGAAGCCGGCAAAGGCGGCAGTC-3' (SEQ ID NO: 6) в качестве обратного праймера. Затем проводят ПЦР на кДНК, приготовленной из клеток M. alpina, с использованием вышеприведенных праймеров и ДНК-полимеразы или т.п. Хотя эта реакция может быть без труда выполнена квалифицированными в данной области техники специалистами в соответствии, например, с “Molecular Cloning, A Laboratory Manual 3rd ed.” (Cold Spring Harbor Press (2001)), можно, в качестве примера, установить следующие условия для ПЦР в настоящем изобретением:
Температура для денатурации: 90-95°С,
Температура для отжига: 40-60°С,
Температура при элонгации: 60-75°С,
Число циклов: 10 или более.
Получаемые в результате ПЦР-продукты можно очистить известным способом, например, используя набор (например, GENECLEAN (Funakoshi Co., Ltd, Япония), наборы для очистки ПЦР-продуктов QIAquik (QIAGEN), ExoSAP-IT (GE Heathcare Bio-Sciences)), фильтр из DEAE-целлюлозы или мешок для диализа. В случае использования агарозного геля ПЦР-продукты подвергают электрофорезу в агарозном геле, и фрагменты нуклеотидной последовательности вырезают из агарозного геля, а затем очищают с использованием наборов для выделения из геля GENECLEAN (Funakoshi Co., Ltd, Япония) или QIAquik (QIAGEN) или способом замораживания-сдавливания и т.п.
Нуклеотидные последовательности клонированных нуклеиновых кислот можно определить с использованием нуклеотидного секвенатора.
Векторная конструкция для экспрессии РАР и получение трансформанта
Настоящим изобретением также обеспечивается рекомбинантный вектор, включающий нуклеиновую кислоту, кодирующую РАР настоящего изобретения. Настоящим изобретением, кроме того, обеспечивается трансформант, трансформированный указанным выше рекомбинантным вектором.
Такой рекомбинантный вектор и трансформант можно получить следующим образом. А именно, плазмиду, несущую нуклеиновую кислоту, кодирующую РАР настоящего изобретения, расщепляют ферментами рестрикции. Примеры ферментов рестрикции, имеющихся в наличии для использования, включают, но без ограничения, EcoRI, KpnI, BamHI и SalI. За этим расщеплением может следовать затупление концов с использованием Т4-полимеразы. Фрагмент расщепленной нуклеотидной последовательности очищают путем электрофореза в агарозном геле. Этот фрагмент нуклеотидной последовательности можно интегрировать в экспрессионный вектор известным способом с получением вектора для экспрессии РАР. Этот экспрессионный вектор вводят в хозяина для получения трансформанта, который затем предоставляется для экспрессии требуемого белка.
В этом случае типы экспрессионного вектора и хозяина никоим образом не ограничены, если они делают возможной экспрессию требуемого белка. Примеры хозяина включают грибы, бактерии, растения, животные или их клетки. Грибы включают нитчатые грибы, такие как липидпродуцирующий M. alpina, и дрожжевые штаммы, такие как Saccharomyces cerevisiae. Бактерии включают Escherichia coli (E.coli) и Bacillus subtilis. Также растения включают масличные культуры, такие как рапс, соя, хлопчатник, сафлор и лен.
В качестве липидпродуцирующих штаммов можно использовать такие штаммы, которые указаны в MYCOTAXON, Vol. XLIV, No. 2, pp. 257-265 (1992). Конкретные примеры включают микроорганизмы, относящиеся к роду Mortierella, иллюстрируемые микроорганизмами, относящимися к такому подроду Mortierella, как Mortierella elongate IFO8570, Mortierella exigua IFO8571, Mortierella hydrophila IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70, CBS754.68, а также микроорганизмами, относящимися к такому подроду Micromucor, как Mortierella isabellina, CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308, IFO7884, Mortierella nana IFO8190, Mortierella ramanniana IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185, IFO8287, Mortierella vinacea CBS236.82. Особенно предпочтительным является Mortierella alpina.
При использовании в качестве хозяина гриба, желательно, чтобы нуклеиновая кислота настоящего изобретения была самореплицирующейся в хозяине или имела структуру, способную к встраиванию в хромосому гриба. В то же самое время предпочтительно, когда она дополнительно включает промотор и терминатор. При использовании в качестве хозяина M. alpina, примеры экспрессионных векторов включают pD4, pDuraSC и pDura5. Можно использовать любой промотор, если он делает возможной экспрессию в хозяине, и примеры включают промоторы, происходящие из M. alpina, такие как промотор гена гистона Н4.1, промотор гена GAPDH (глицеральдегид-3-фосфат-дегидрогеназы) и промотор гена TEF (фактора элонгации трансляции).
Методы введения рекомбинантного вектора в нитчатые грибы (например, M. alpina) включают электропорацию, способы доставки с использованием сферопластов и частиц, а также прямую микроинъекцию ДНК в ядра. В случае использования ауксотрофного штамма-хозяина штаммы, растущие на селективной среде, в которой отсутствуют питательные вещества, требуемые для штамма-хозяина, можно отобрать с получением, тем самым, трансформированных штаммов. Альтернативно, в случае использования для трансформации маркерного гена, обеспечивающего устойчивость к лекарственному средству, культивирование можно проводить с использованием селективной среды, содержащей лекарственное средство, с получением, тем самым, колоний, устойчивых к лекарственному средству.
При использовании в качестве хозяина дрожжей примеры экспрессионного вектора включают pYE22m. В альтернативном случае можно также использовать имеющиеся в продаже экспрессионные векторы, такие как pYES (Invitrogen) и pESC (Stratagene). Дрожжи-хозяева, подходящие для настоящего изобретения, включают, но без ограничения, штамм EH13-15 Saccharomyces cerevisiae (trp1, MATα). Примеры промоторов, подходящих для использования, включают промоторы, происходящие из дрожжей или т.п., такие как промотор GAPDH, промотор gal1 и промотор gal10.
Методы введения рекомбинантного вектора в дрожжевые клетки включают способ с использованием ацетата лития, электропорацию и способы с использованием сферопластов, а также декстран-опосредованную трансфекцию, преципитацию фосфатом кальция, полибрен-опосредованную трансфекцию, слияние протопластов, инкапсуляцию полинуклеотида(ов) в липосомы и прямую микроинъекцию ДНК в ядра.
При использовании в качестве хозяина бактерии, такой как E. coli, примеры экспрессионного вектора включают pGEX и pUC18, доступные от Pharmacia. Примеры промоторов, имеющихся в наличии для использования, включают промоторы, происходящие из E. coli, фага или т.п., например, промотор trp, промотор lac, промотор PL и промотор PR. Методы введения рекомбинантного вектора в бактерии включают электропорацию и способы с использованием хлорида кальция.
Способ настоящего изобретения для приготовления композиции жирной кислоты
Настоящим изобретением обеспечивается способ приготовления композиции жирной кислоты из указанного выше трансформанта, т.е. способ приготовления композиции жирной кислоты из продукта культивирования, полученного путем культивирования указанного выше трансформанта, конкретнее как описано ниже. Однако способ настоящего изобретения не ограничивается следующим способом и может быть выполнен любым другим способом, как правило, известным.
Для культивирования организмов, трансформированных для экспрессии РАР, можно использовать любую среду, если она является раствором (средой) для культивирования, имеющим подходящий рН и осмотическое давление, а также содержащим питательные вещества, необходимые для роста каждого хозяина, микроэлементы и биоматериалы, такие как сыворотка, или антибиотики. Например, в случае дрожжевых клеток, трансформированных для экспрессии РАР, среду SC-Trp, среду YPD, среду YPD5 или т.п. можно использовать, без ограничения указанными средами. Детализированный состав среды иллюстрируется для среды SC-Trp: 6,7 г дрожжевого азотистого основания без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смеси 1,25 г аденина сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глютаминовой кислоты, 0,6 г гистидина, 1,8 г лейцина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 0,6 г урацила) на литр среды.
Можно использовать любые условия культивирования, если они подходят для роста хозяина и являются адекватными для сохранения образуемого фермента в стабильном состоянии. Конкретнее, можно регулировать отдельные условия, в том числе степень анаэробности, период культивирования, температуру, влажность, статическое культивирование или культивирование при покачивании. Культивирование можно проводить в одних и тех же условиях (одностадийное культивирование) или согласно так называемому двухстадийному или трехстадийному культивированию, используя два или более различных условий культивирования. Для широкомасштабного культивирования предпочтительным является двухстадийное культивирование или культивирование с использованием большего числа стадий из-за присущей им высокой эффективности культивирования.
Для объяснения детализированных процедур для способа приготовления композиции жирной кислоты настоящего изобретения, ниже, в качестве примера, поясняется двухстадийное культивирование, в котором в качестве хозяина используются дрожжи. А именно, на стадии предварительной культуры колонии, полученные выше, засевают в любую среду, описанную выше (например, среду SC-Trp), и культивируют при покачивании при 30οС в течение 2 дней. Затем на стадии основного культивирования среду предварительного культивирования (500 мкл) добавляют к 10 мл среды YPD5 (2% дрожжевого экстракта, 1% полипептона, 5% глюкозы) и культивируют при покачивании при 30°С в течение 2 дней.
Композиции жирной кислоты настоящего изобретения
Настоящим изобретением также обеспечивается композиция жирной кислоты, которая является совокупностью одной или нескольких жирных кислот в клетках, экспрессирующих РАР настоящего изобретения. Жирные кислоты могут быть свободными жирными кислотами или могут быть триглицеридами, фосфолипидами или т.п. В частности, композиция жирной кислоты настоящего изобретения отличается обладанием жирнокислотной характеристики, в которой гарантируется большая степень по крайней мере одного или нескольких показателей:
i) содержания олеиновой кислоты;
ii) отношения содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты;
iii) отношения содержания олеиновой кислоты к содержанию пальмитиновой кислоты;
iv) отношения общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты;
v) отношения содержания жирных кислот С18 к содержанию жирных кислот С16; и
vi) относительного показателя содержания арахидоновой кислоты, дигомо-γ-линоленовой кислоты и/или γ-линоленовой кислоты;
по сравнению с продуктом культивирования, полученным путем культивирования хозяина, который не трансформирован рекомбинантным вектором настоящего изобретения. Предполагается, что под используемым здесь выражением «хозяин, который не трансформирован рекомбинантным вектором настоящего изобретения», имеется в виду, например, хозяин, трансформированный пустым вектором, не несущим какую-либо из нуклеиновых кислот, описанных в разделе «Нуклеиновые кислоты настоящего изобретения, кодирующие фосфатазу фосфатидной кислоты».
Жирные кислоты, содержащиеся в композиции жирной кислоты настоящего изобретения, относятся к линейным или разветвленным монокарбоновым кислотам длинноцепочечных углеводов, включающим, но без ограничения, миристиновую кислоту (тетрадекановую кислоту), (14:0), миристоленовую кислоту (тетрадеценовую кислоту) (14:1), пальмитиновую кислоту (гексадекановую кислоту) (16:0), пальмитолеиновую кислоту (9-гексадеценовую кислоту) (16:1), стеариновую кислоту (октадекановую кислоту) (18:0), олеиновую кислоту (цис-9-октадеценовую кислоту) (18:1(9)), вакценовую кислоту (11-октадеценовую кислоту (18:1(11)), линолевую кислоту (цис,цис-9,12-октадекадиеновую кислоту (18:2(9,12)), α-линоленовую кислоту (9,12,15-октадекатриеновую кислоту) (18:3(9,12,15), γ-линоленовую кислоту (6,9,12-октадекатриеновую кислоту) (18:3(6,9,12), стеаридоновую кислоту (6,9,12,15-октадекатетраеновую кислоту) (18:4(6,9,12,15), арахиновую кислоту (эйкозановую кислоту) (20:0), (8,11-эйкозадиеновую кислоту) (20:2(8,11)), медово-винную кислоту (5,8,11-эйкозатриеновую кислоту) (20:3(5,8,11)), дигомо-γ-линоленовую кислоту (8,11,14-эйкозатриеновую кислоту) (20:3(8,11,14)), арахидоновую кислоту (5,8,11,14-эйкозатетраеновую кислоту) (20:4(5,8,11,14)), эйкозатетраеновую кислоту (8,11,14,17-эйкозатетраеновую кислоту) (20:4(8,11,14,17)), эйкозапентаеновую кислоту (5,8,11,14,17-эйкозапентаеновую кислоту) (20:5(5,8,11,14,17)), бегеновую кислоту (докозановую кислоту) (22:0), (7,10,13,16-докозатетраеновую кислоту) (22:4(7,10,13,16)), (7,10,13,16,19-докозапентаеновую кислоту) (22:5(7,10,13,16,19)), (4,7,10,13,16-докозапентаеновую кислоту) (22:5(4,7,10,13,16)), (4,7,10,13,16,19-докозагексаеновую кислоту) (22:6(4,7,10,13,16,19)), лигноцериновую кислоту (тетрадокозановую кислоту) (24:0), ацетэруковую кислоту (цис-15-тетрадокозановую кислоту) (24:1) и церотиновую кислоту (гексадокозановую кислоту) (26:0). Следует отметить, что вышеприведенные названия веществ являются обычными названиями, установленными Комиссией по номенклатуре биохимических соединений IUPAC, и их систематические названия приводятся в круглых скобках вместе с числами, указывающими на число атомов углерода и положения двойных связей.
Композиция жирной кислоты настоящего изобретения может состоять из любого числа и любого типа жирных кислот, если она является композицией одной или нескольких жирных кислот, выбираемых из перечисленных выше жирных кислот.
Факт получения такой композиции жирной кислоты настоящего изобретения, т.е. факт экспрессии РАР настоящего изобретения, можно подтвердить обычно известным способом, например, по изменению жирнокислотной характеристики при экспрессии РАР в дрожжевых клеткам. А именно, к лиофилизированным клеткам, полученным вышеприведенным способом настоящего изобретения для приготовления композиции жирной кислоты, добавляют смесь хлороформ:метанол в соответствующей пропорции и перемешивают, а затем подвергают тепловой обработке в течение соответствующего периода времени. Далее выполняют центрифугирование для отделения клеток и сбора растворителя. Эту процедуру повторяют несколько раз. Затем липиды высушивают подходящим образом, и для растворения липидов добавляют растворитель, такой как хлороформ. Соответствующую аликвоту этого образца обрабатывают с помощью метода с использованием соляной кислоты/метанола для превращения жирных кислот в клетках в соответствующие метиловые эфиры, а затем подвергают экстракции гексаном. После отгонки гексана жирные кислоты анализируют с помощью газовой хроматографии.
В результате, если получена композиция жирной кислоты, имеющая указанную выше жирнокислотную характеристику, можно определить, что получена композиция жирной кислоты настоящего изобретения.
Пищевые или другие продукты, включающие композиции жирной кислоты настоящего изобретения
Настоящим изобретением также обеспечивается пищевой продукт, включающий указанную выше композицию жирной кислоты. Композицию жирной кислоты настоящего изобретения можно использовать обычным образом для таких целей, как производство пищевых продуктов, содержащих жиры и масла, а также производство промышленных исходных материалов (исходных материалов для косметических средств, лекарственных препаратов (например, наружных препаратов для кожи), мыла и т.п.). Косметические средства (косметические композиции) или лекарственные препараты (фармацевтические композиции) можно приготовить в любой дозированной форме, включающей, но без ограничения, растворы, пасты, гели, сухое вещество или порошки. Подобным образом, возможные формы пищевых продуктов включают фармацевтические композиции, такие как капсулы, а также пищевые продукты, подвергшиеся технологической обработке, такие как обычная жидкая пища, полупереваренная пища для усиленного питания, пища в виде элементных смесей, годные для питья продукты или продукты для энтерального питания, которые включают композицию жирной кислоты настоящего изобретения в смеси с белками, сахарами, жирами, микроэлементами, витаминами, эмульгаторами, вкусовыми ароматизирующими веществами.
Кроме того, примеры пищевого продукта настоящего изобретения включают, но без ограничения, продукты с пищевыми добавками, здоровую пищу, функциональные продукты питания, пищевые продукты для детей, молоко измененного состава для младенцев, молоко измененного состава для недоношенных детей и гериатрические пищевые продукты. Термин «пища» или «пищевой продукт» используется здесь в качестве родового названия годных в пищу материалов в форме сухого вещества, текучих сред, жидкостей или их смесей.
Термин «продукты с пищевыми добавками» относится к пищевым продуктам, обогащенным специфическими пищевыми ингредиентами. Термин «здоровая пища» относится к пищевым продуктам, которые полезны для здоровья, и включают продукты с пищевыми добавками, природные пищевые продукты и диетические пищевые продукты. Термин «функциональные продукты питания» относится к пищевым продуктам для пополнения пищевых ингредиентов, которые помогают организму контролировать функции. Термин «функциональные продукты питания» является синонимом термина «пищевые продукты для заданного целебного применения». Термин пищевые продукты для детей относится к пищевым продуктам, даваемым детям возрастом вплоть до приблизительно 6 лет. Термин «гериатрические пищевые продукты» относится к пищевым продуктам, обработанным для облегчения пищеварения и абсорбции по сравнению с необработанными продуктами. Термин «молоко измененного состава для младенцев» относится к молоку измененного состава, даваемому детям возрастом вплоть до приблизительно одного года. Термин «молоко измененного состава для недоношенных детей» относится к молоку измененного состава, даваемому недоношенным детям до приблизительно 6 месяцев после рождения.
Эти пищевые продукты включают природные пищевые продукты (обработанные жирами и маслами), такие как мясо, рыба и орехи; пищевые продукты, дополненные жирами и маслами во время приготовления (например, китайские продукты питания, китайская лапша, супы), продукты питания, приготовленные с использованием жиров и масел в качестве нагревательных сред (например, температура (хорошо прожаренные рыба и овощи), хорошо прожаренные пищевые продукты, жаренный соевый творог, китайский жареный рис, жареные пирожки, японские жареные лепешки (каринто), пищевые продукты или подвергнутые технологической обработке пищевые продукты на основе жиров и масел, дополненные жирами и маслами во время обработки (например, масло, маргарин, майонез, заправка, шоколад, лапша быстрого приготовления, карамель, бисквиты, пирожные, торт, мороженное), и пищевые продукты, на которые нанесены жиры и масла или которые покрыты ими по окончанию приготовления (например, рисовое сухое печенье, галеты, сладкая галета-паста из соевых бобов). Однако пищевой продукт настоящего изобретения не ограничивается пищевыми продуктами, содержащими жиры и масла, и другие примеры включают продукты сельского хозяйства, такие как хлебобулочные изделия, лапша, приготовленный рис, сладости (например, кондитерские изделия, жевательные резинки, камеди, пластинки, японские сладости), соевый творог и подвергнутые технологической обработке продукты из них, ферментированные пищевые продукты, такие как японское вино из риса (саке), водный раствор лекарства, сладкое вино, используемое в кулинарии (мирин), уксус, соевый соус и мисо (паста из соевых бобов), пищевые продукты животного происхождения, такие как йогурт, ветчина, бекон и колбасные изделия, морепродукты, такие как рыбные лепешки (камабоко), хорошо прожаренные рыбные лепешки (агетен) и рыхлые лепешки из рыбы (ханпен), а также фруктовые напитки, безалкогольные напитки, спортивные напитки, алкогольные напитки и чай.
Способ оценки или отбора штамма, используя кодирующую РАР нуклеиновую кислоту или белок РАР настоящего изобретения
Настоящим изобретением также обеспечивается способ оценки или отбора липидпродуцирующего штамма, используя кодирующую РАР нуклеиновую кислоту или белок РАР настоящего изобретения.
Детали приведены ниже.
(1) Способ оценки
Одним вариантом осуществления настоящего изобретения является способ оценки липидпродуцирующего штамма, используя кодирующую РАР нуклеиновую кислоту или белок РАР настоящего изобретения. В качестве первого примера указанного выше способа оценки настоящего изобретения, липидпродуцирующий испытуемый штамм проверяют на указанную выше активность настоящего изобретения, используя праймеры или зонды, сконструированные на основе нуклеотидной последовательности настоящего изобретения. Общие способы такой оценки известны, и их можно найти, например, в публикации международной заявки на патент № WO01/040514 или JP 8-205900A. Ниже приводится краткое пояснение этой оценки.
Сначала готовят геномную ДНК испытуемого штамма. Для приготовления геномной ДНК можно использовать любой известный метод, такой как способ Герефорда или способ с использованием ацетата калия (смотри, например, Methods in Yeast Genetics, Cold Spring Harbor Laboratory Press, p130 (1990)).
Праймеры или зонды конструируют на основе нуклеотидной последовательности настоящего изобретения, предпочтительно SEQ ID NO: 4. Эти праймеры или зонды могут представлять собой любые области нуклеотидной последовательности настоящего изобретения, и при их конструировании можно использовать известные способы. Число нуклеотидов в полинуклеотиде, используемом в качестве праймера, обычно составляет 10 нуклеотидов или больше, предпочтительно 15-25 нуклеотидов. Подобным образом, число нуклеотидов, подходящее для области, которая будет фланкироваться праймерами, обычно составляет 300-2000 нуклеотидов.
Приготовленные, как указано выше, праймеры или зонды используют для проверки того, содержит ли геном указанного выше испытуемого штамма последовательность, специфичную для нуклеотидной последовательности настоящего изобретения. Последовательность, специфичную для нуклеотидной последовательности настоящего изобретения, можно выявить, используя известные способы. Например, полинуклеотид, включающий часть или всю последовательность, специфичную для нуклеотидной последовательности настоящего изобретения, или полинуклеотид, включающий нуклеотидную последовательность, комплементарную указанной выше нуклеотидной последовательности, используют в качестве одного праймера, а полинуклеотид, включающий часть или всю последовательность, расположенную 5' или 3' от этой последовательности, или полинуклеотид, включающий нуклеотидную последовательность, комплементарную указанной выше нуклеотидной последовательности, используют в качестве другого праймера для амплификации нуклеиновых кислот испытуемого штамма с помощью ПЦР или других способов с последующим определением присутствия или отсутствия продуктов амплификации, молекулярной массы продуктов амплификации и т.п.
Условия ПЦР, подходящие для способа настоящего изобретения, никоим образом не ограничиваются, и можно, в качестве примера, установить следующие условия:
Температура для денатурации: 90-95°С,
Температура для отжига: 40-60°С,
Температура при элонгации: 60-75°С,
Число циклов: 10 или более.
Получаемые в результате продукты реакции можно отделить с помощью электрофореза в агарозном геле или т.п. для определения молекулярной массы продуктов амплификации. Для каждого продукта амплификации затем устанавливают, имеет ли его молекулярная масса величину, достаточную для охвата молекулы нуклеиновой кислоты, соответствующей области, специфичной для нуклеотидной последовательности настоящего изобретения, посредством чего можно предсказать или оценить указанную выше активность настоящего изобретения в испытуемом штамме. Кроме того, если анализируют, как указано выше, нуклеотидные последовательности указанных выше продуктов амплификации, указанную выше активность настоящего изобретения можно предсказать или оценить точнее. Следует отметить, что способы оценки указанной выше активности настоящего изобретения являются описанными выше способами.
В качестве другого примера указанного выше способа оценки настоящего изобретения, испытуемый штамм подвергают культивированию и определяют уровень экспрессии РАР, кодируемой нуклеотидной последовательностью настоящего изобретения (например, SEQ ID NO: 4), посредством чего можно оценить указанную выше активность настоящего изобретения в испытуемом штамме. Следует отметить, что уровень экспрессии РАР можно определить путем культивирования испытуемого штамма в соответствующих условиях и количественного определения мРНК или белка для РАР. Количественное определение мРНК или белка можно выполнить, используя известные способы, например, гибридизацию по Нозерну или количественную ОТ-ПЦР для количественного определения мРНК и Вестерн-блоттинг для количественного определения белка (Current Protocols in Molecular Biology, John Wiley & Sons 1994-2003). Для оценки указанной выше активности можно также использовать определение жирнокислотной характеристики композиции жирной кислоты, образуемой РАР настоящего изобретения. Способы определения жирнокислотной характеристики композиции жирной кислоты являются описанными выше способами.
(2) Способ отбора
Другим вариантом осуществления настоящего изобретения является способ отбора липидпродуцирующего штамма, используя кодирующую РАР нуклеиновую кислоту или белок РАР настоящего изобретения. В качестве примера указанного выше способа отбора настоящего изобретения, испытуемые штаммы подвергают культивированию и определяют уровень экспрессии РАР, кодируемой нуклеотидной последовательностью настоящего изобретения (например, SEQ ID NO: 4), для отбора штамма с требуемым уровнем экспрессии, посредством чего можно отобрать штамм, обладающий требуемой активностью. Альтернативно, заранее определяют стандартный штамм, и этот стандартный штамм и каждый из испытуемых штаммов подвергают культивированию и определяют в них указанный выше уровень экспрессии с последующим сравнением уровня экспрессии между стандартным штаммом и каждым испытуемым штаммом, посредством чего можно отобрать требуемый штамм. Конкретнее, например, стандартный штамм и испытуемые штаммы подвергают культивированию в соответствующих условиях и определяют в них уровни экспрессии для отбора испытуемого штамма, демонстрирующего экспрессию, которая выше или ниже экспрессии в стандартном штамме, посредством чего можно отобрать штамм, обладающий требуемой активность. Примеры требуемой активности включают уровень экспрессии РАР и жирнокислотную характеристику композиции жирной кислоты, образуемой РАР, которые можно определить, как описано выше.
В другом примере указанного выше способа отбора настоящего изобретения испытуемые штаммы подвергают культивированию с отбором штамма, в котором указанная выше активность настоящего изобретения является высокой или низкой, посредством чего можно отобрать штамм, обладающий требуемой активностью. Примеры требуемой активности включают уровень экспрессии РАР и жирнокислотную характеристику композиции жирной кислоты, образуемой РАР, которые можно определить, как описано выше.
Примеры испытуемого штамма или стандартного штамма, имеющиеся в наличии для использования, включают, но без ограничения, штамм, трансформированный указанным выше вектором настоящего изобретения, штамм, модифицированный для подавления экспрессии указанной выше нуклеиновой кислоты настоящего изобретения, штамм, модифицированный с помощью мутагенеза, и штамм, имеющий природную мутацию(и). Следует отметить, что РАР-активность в настоящем изобретении и способность к обеспечению жирнокислотной характеристики, обеспечиваемой РАР в настоящем изобретении, можно определить, например, с помощью способов, приведенных в разделах «Нуклеиновые кислоты настоящего изобретения, кодирующие фосфатазу фосфатидной кислоты» и «Композиции жирной кислоты настоящего изобретения». Мутагенез можно осуществить с помощью, но без ограничения, физических методов, включающих ультрафиолетовое или радиоактивное облучение, или химических методов, включающих обработку таким агентом, как EMS (этилметансульфонат) или N-метил-N-нитрозогуанидин (смотри, например, Yasuji Oshima ed., Biochemistry Experiments vol. 39, Experimental Protocols for Yeast Molecular Genetics, pp. 67-75, Japan Scientific Societies Press).
Штаммы, используемые в настоящем изобретении в качестве стандартного и испытуемых штаммов, включают, но без ограничения, указанные выше липидпродуцирующие штаммы или штаммы дрожжей. Конкретнее, стандартным штаммом или испытуемым штаммом может быть комбинация любых штаммов, относящихся к различным родам или видам, и один или более испытуемых штаммов могут использоваться одновременно.
Настоящее изобретение теперь будет подробнее описано в виде следующих примеров, которые, как предполагается, не ограничивают объем настоящего изобретения.
Пример 1
(1) Анализ EST
Штамм 1S-4 M. alpina засевали в 100 мл среды (1,8% глюкозы, 1% дрожжевого экстракта, рН 6,0) и подвергали предварительному культивированию в течение 3 дней при 28οС. 10л-сосуд для культивирования (Able Co., Tokyo) загружали 5 л среды (1,8% глюкозы, 1% соевого порошка, 0,1% оливкового масла, 0,01% Adekanol, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2 .2H2O, 0,05% MgCl2 .6H20, рН 6,0) и засевали взятым в целом продуктом предварительного культивирования с последующим аэробным культивированием с постоянным перемешиванием при 300 оборотах в минуту, 1 vvm и 26°C в течение 8 дней. В дни 1, 2 и 3 культивирования добавляли глюкозу в количестве, соответствующем 2%, 2% и 1,5%, соответственно. На каждой стадии культивирования (день 1, 2, 3, 6 или 8) получали клетки для приготовления общей РНК с помощью метода, в котором используется гуанидина гидрохлорид/CsCl. Используя набор для очистки Oligotex-dT30<Super>mRNA (Takara Bio Inc., Япония), поли(А)+РНК очищали из общей РНК. Библиотеку кДНК готовили для каждой стадии с использованием набора для синтеза кДНК ZAP (Stratagene) с последующим анализом за один проход последовательности с 5'-конца кДНК (8000 клонов × 5 стадий). Результирующие последовательности группировали.
(2) Поиск гомологов гена РАР
Нуклеотидные последовательности, полученные при анализе EST, искали на фоне аминокислотных последовательностей, зарегистрированных в GENBANK, используя программу поиска гомологии, BLAST, для выделения гомологов гена РАР. В результате среди EST была обнаружена последовательность (SEQ ID NO: 1), которая обладала высокой степенью гомологии с происходящим из Neurospora crassa предполагаемым белком, являющимся гомологом диацилглицеролпирофосфат-фосфатазы (DPP), обладающей РАР-активностью. Контиг, полученный для этой SEQ ID NO: 1 при анализе EST, состоял из 9 последовательностей из библиотеки в день 8, указанной выше в (1) выше.
Пример 2
(1) Клонирование гомолога РАР (MaPAP1)
Для получения фрагмента кДНК, содержащего полноразмерную ORF гомолога РАР, были разработаны следующие праймеры:
D1: 5'-CATGGGTTGCTTCGCGCGCAAGACG-3' (SEQ ID NO: 5),
D2: 5'-CGAAGCCGGCAAAGGCGGCAGTC-3' (SEQ ID NO: 6).
Используя эти праймеры, ПЦР проводили с использованием ExTaq (Takara Bio Inc., Япония) и кДНК в день 8 в качестве матрицы. Получаемый в результате ДНК-фрагмент размером 0,64 т.п.о. был подвергнут клонированию с использованием ТА, используя набор для клонирования TOPO-TA (Invitrogen), для определения нуклеотидной последовательности вставки.
Результаты подтверждали, что был клонирован ДНК-фрагмент, включающий нуклеотидную последовательность, охватывающую нуклеотиды 113-744 SEQ ID NO: 1. Эту плазмиду назвали pCR-2051-P. Затем плазмиду pCR-2051-P использовали в качестве матрицы для проведения ПЦР с теми же праймерами, которые указаны выше. В ПЦР использовали ExTaq (Takara Bio Inc., Япония), но прилагаемую смесь dNTP заменяли раствором для мечения во время ПЦР (Roche Diagnostic) для мечения амплифицируемой ДНК дигоксигенином (DIG), посредством чего готовили зонд для использования при скрининге библиотеки кДНК. Этот зонд использовали для скрининга библиотеки кДНК в день 8.
Устанавливали следующие условия гибридизации:
Буфер: 5 x SSC, 1% SDS, 50 мМ Tris-HCl (pH 7,5), 50% формамида,
Температура: 42°С (в течение ночи)
Отмывка: 0,2 x SSC, 0,1% SDS при 65°С в течение 20 минут (с повтором три раза).
Выявление проводили, используя набор для выявления нуклеиновых кислот, меченных DIG (Roche Diagnostic). Из фаговых клонов, полученных при скринировании, вырезали плазмиду с помощью in vivo вырезания для получения плазмидной ДНК.
В результате скрининга кДНК, как описано выше, установлено, что нуклеотидная последовательность клона с самой длинной вставкой идентична SEQ ID NO: 1. Эту плазмиду назвали pB-PAP1. SEQ ID NO: 1 содержит CDS размером 1119 п.о. (SEQ ID NO: 3), что, тем самым, говорит о том, что была получена последовательность, кодирующая полноразмерный гомолог РАР (фиг. 1). Этот ген назвали геном MaPAP1. Расшифрованная аминокислотная последовательность белка (MaPAP1p), кодируемая этим геном, указана в SEQ ID NO: 2.
(2) Анализ последовательности
Нуклеотидную последовательность гена MaPAP1 и кодируемую им расшифрованную аминокислотную последовательность подвергли поискам гомологии с использованием BLAST и ClustalW на фоне известных последовательностей нуклеиновых кислот и аминокислотных последовательностей. Среди последовательностей, найденных при поиске гомологии с использованием BLAST на фоне аминокислотных последовательностей, зарегистрированных в GENBACK, последовательность, имеющая наименьший Е-показатель, была предполагаемым белком происходящего из Neurospora crassa гомолога DPP1 (входящий № CAD70721). Идентичность по аминокислотной последовательности составляла 59,3%, а идентичность последовательности нуклеиновой кислоты, кодирующей этот белок, составляла 57,4%. Кроме того, гомологии также наблюдались с белками семейства Mg2+-независимых фосфатаз 2 фосфатидной кислоты (РАР2) грибов, растений и животных. Ферменты семейства РАР2 являются мембранными белками и представляют собой тип ферментов, шесть раз пронизывающих мембрану. На фиг. 2 демонстрируется совмещение аминокислотных последовательностей этих белков семейства РАР2 с расшифрованной аминокислотной последовательностью, кодируемой геном MaPAP1, полученным в этом исследовании. Ферменты семейства РАР2 содержат три консенсусные области, и также известны аминокислоты, необходимые для их активности. Как продемонстрировано на фиг. 2, установлено, что в MaPAP1p также сохранены эти консенсусные области (с двойным подчеркиванием на фиг. 1) и остатки, необходимые для активности, т.е. аргинин в домене 1 и гистидины в доменах 2 и 3 (указанные * на фиг. 2).
Пример 3
Конструирование вектора для экспрессии в дрожжах
Для экспрессии MaPAP1 в дрожжевых клетках вектор для экспрессии в дрожжах был сконструирован следующим образом. Сначала pB-PAP1 использовали в качестве матрицы для проведения ПЦР с использованием ExTaq (Takara Bio Inc., Япония), а также праймера D1 (SEQ ID NO: 5) и праймера D3: 5'-CCACTAAACTATGTTCTCAGGC-3' (SEQ ID NO: 7). Получаемый в результате ДНК-фрагмент размером 1,1 т.п.о. был подвергнут клонированию с использованием ТА, используя набор для клонирования TOPO-TA (Invitrogen), и подтверждали его нуклеотидную последовательность. Плазмиду, несущую безошибочную нуклеотидную последовательность CDS (SEQ ID NO:3) MaPAP1, назвали pCR-PAP1. Плазмиду pCR-PAP1 частично расщепляли ферментом рестрикции EcoRI. Фрагмент размером приблизительно 1,1 т.п.о. вырезали из агарозного геля после электрофореза, очищали с использованием набора для очистки ДНК GFX (GE Healthcare BI-Sciences) и лигировали в EcoRI-сайт вектора для экспрессии в дрожжах pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995). Подтверждали ориентацию встроенного фрагмента ДНК, и конструкцию, несущую вставку в такой ориентации, которая обуславливает транскрипцию ORF с промотора GAPDH pYE22m, назвали pYE-MAPAP1.
Пример 4
Трансформация дрожжей
Плазмиду pYE22m или pYE-MAPAP1 использовали для трансформации дрожжевого штамма EH13-15 Saccharomyces cerevisiae (trp1, MATα) (Appl. Microbiol. Biotechnol., 30, 515-520, 1989), используя способ с использованием ацетата лития. Трансформированные штаммы отбирали по способности расти на агаризованной (2% агара) среде SC-Trp, содержащей, на литр, 6,7 г дрожжевого азотистого основания без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смеси 1,25 г аденина сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глютаминовой кислоты, 0,6 г гистидина, 1,8 г лейцина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 0,6 г урацила).
Пример 5
Культивирование дрожжей
Среди штаммов, трансформированных каждым из векторов, любые два штамма (штаммы с-1 и с-2 или штаммы MaPAP1-1 и MaPAP1-2) отбирали и подвергали культивированию в соответствующих условиях.
А именно, на стадии предварительного культивирования петлю каждого дрожжевого штамма засевали с чашки со средой SC-Trp (10 мл) и культивировали с покачиванием при 30°С в течение 2 дней. На стадии основного культивирования среду предварительного культивирования (500 мкл) добавляли к 10 мл среды YPD5 (2% дрожжевого экстракта, 1% полипептона, 5% глюкозы) и культивировали с покачиванием при 30°С в течение 2 дней.
Пример 6
Анализ жирных кислот дрожжевых штаммов
Каждую из сред культивирования дрожжей центрифугировали для сбора клеток. После промывки 10 мл стерилизованной воды клетки снова собирали центрифугированием и лиофилизировали. К лиофилизированным клеткам добавляли смесь хлороформ:метанол (2:1, 4 мл) и энергично перемешивали, а затем подвергали инкубации при 70°С в течение 1 часа. Клетки отделяли с помощью центрифугирования для сбора растворителя. К остающимся клеткам снова добавляли смесь хлороформ:метанол (2:1, 4 мл), и эту же процедуру повторяли для сбора растворителя. После высушивания липидов с использованием SpeedVac для растворения липидов добавляли 2 мл хлороформа. 200 мкл-аликвоту этого образца обрабатывали с помощью метода с использованием соляной кислоты/метанола для превращения жирных кислот в клетках в соответствующие метиловые эфиры, а затем подвергали экстракции гексаном. После отгонки гексана жирные кислоты анализировали с помощью газовой хроматографии.
Полученные результаты представлены в таблице 2.
Таблица 2. Жирнокислотная характеристика трансформированных штаммов (хозяин: ЕН13-15).
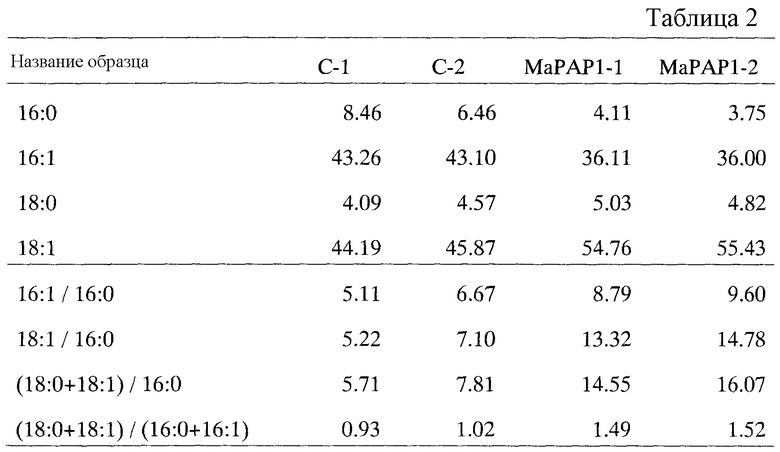
Сравнивали жирнокислотные характеристики дрожжевых штаммов, трансформированных MaPAP1 (MaPAP1-1 и MaPAP1-2), и контрольных дрожжевых штаммов (С-1 или С-2). В трансформированных MaPAP1 дрожжах процентное содержание олеиновой кислоты увеличивалось на 20% или больше по сравнению с контрольными штаммами. Процентное содержание как пальмитиновой кислоты, так и пальмитолеиновой кислоты снижалось по сравнению с контрольными штаммами, в то время как отношение содержания пальмитолеиновой кислоты к содержанию пальмитиновой кислоты слегка увеличивалось по сравнению с контрольными штаммами. Кроме того, в трансформированных MaPAP1 дрожжах увеличивалось как отношение содержания олеиновой кислоты к содержанию пальмитиновой кислоты, так и отношение общего содержания стеариновой и олеиновой кислот к содержанию пальмитиновой кислоты по сравнению с контрольными штаммами. Кроме того, в трансформированных MaPAP1 дрожжах увеличивалось отношение содержания жирных кислот С18 к содержанию жирных кислот С16, что указывает на то, что образовывались жирные кислоты с большими длинами цепей.
То есть, эти результаты указывают на то, что экспрессия гена MaPAP1 позволяет изменить жирнокислотную характеристику.
Пример 7
Анализ экспрессии в продуцирующих арахидоновую кислоту дрожжевых штаммах
(1) Селекция продуцирующих арахидоновую кислоту дрожжевых штаммов
Для осуществления селекции продуцирующих арахидоновую кислоту дрожжевых штаммах (Saccharomyces cerevisiae) конструировали следующие плазмиды.
Сначала кДНК, приготовленную из штамма 1S-4 M. alpina, использовали в качестве матрицы для проведения ПЦР с использованием ExTaq, а также набора праймеров Δ12-f и Δ12-r, Δ6-f и Δ6-r, GLELO-f и GLELO-r или Δ5-f и Δ5-r для амплификации, тем самым, гена десатуразы Δ12 жирных кислот, гена десатуразы Δ6 жирных кислот, гена элонгазы жирных кислот GLELO или гена десатуразы Δ5 жирных кислот в штамме 1S-4 M. alpina.
Эти гены клонировали с использованием набора для клонирования TOPO-TA. Нуклеотидные последовательности клонов подтверждали, и плазмиды, содержащие нуклеотидные последовательности SEQ ID NO: 16-19, назвали плазмидами pCR-MAΔ12DS (содержащей нуклеотидную последовательность SEQ ID NO: 16), pCR-MAΔ62DS (содержащей нуклеотидную последовательность SEQ ID NO: 17), pCR-MAGLELO (содержащей нуклеотидную последовательность SEQ ID NO: 18) и pCR-MAΔ5DS (содержащей нуклеотидную последовательность SEQ ID NO: 19), соответственно.
С другой стороны, HindIII-расщепленный ДНК-фрагмент размером приблизительно 1,2 т.п.о., полученный из плазмиды pURA34 (JP 2001-120276 A), встраивали в HindIII-сайт вектора pUC18, который был расщеплен ферментами рестрикции EcoRI и SphI с последующим затуплением концов и их самолигированием. Клон, в котором EcoRI-сайт вектора был на 5'-конце URA3, назвали pUC-URA3. Подобным образом, SalI- и XhoI-расщепленный ДНК-фрагмент размером приблизительно 2,2 т.п.о., полученный из плазмиды YEp13, встраивали в SalI-сайт вектора pUC18, и клон, в котором EcoRI-сайт вектора был на 5'-конце LUE2, назвали pUC-LEU2.
Затем плазмиду pCR-MAΔ12DS расщепляли ферментом рестрикции HindIII и, после затупления концов, в дальнейшем расщепляли ферментом рестрикции XbaI с получением ДНК-фрагмента размером приблизительно 1,2 т.п.о., в то время как вектор pESC-URA (Stratagene) расщепляли ферментом рестрикции SacI и, после затупления концов, в дальнейшем расщепляли ферментом рестрикции SpeI с получением ДНК-фрагмента размером приблизительно 6,6 т.п.о. Эти ДНК-фрагменты лигировали с получением плазмиды pESC-U-Δ12. Плазмиду pCR-MAΔ6DS расщепляли ферментом рестрикции XbaI и, после затупления концов, в дальнейшем расщепляли ферментом рестрикции HindIII с получением ДНК-фрагмента размером приблизительно 1,6 т.п.о., в то время как плазмиду pESC-U-Δ12 расщепляли ферментом рестрикции SalI и, после затупления концов, в дальнейшем расщепляли ферментом рестрикции HindIII с получением ДНК-фрагмента размером приблизительно 8 т.п.о. Эти ДНК-фрагменты лигировали с получением плазмиды pESC-U-Δ12:Δ6. Эту плазмиду частично расщепляли ферментом рестрикции PvuII, и получаемый в результате фрагмент размером приблизительно 4,2 т.п.о. встраивали в SmaI-сайт pUC-URA3 с получением плазмиды pUC-URA-Δ12:Δ6.
Подобным образом, плазмиду pCR-MAGLELO расщепляли ферментами рестрикции XbaI и SacI с получением ДНК-фрагмента размером приблизительно 0,95 т.п.о., в то время как вектор pESC-LEU (Stratagene) расщепляли ферментами рестрикции XbaI и SacI с получением ДНК-фрагмента размером приблизительно 7,7 т.п.о. Эти ДНК-фрагменты лигировали с получением плазмиды pESC-L-GLELO. Плазмиду pCR-MAΔ5DS расщепляли ферментом рестрикции XbaI и, после затупления концов, в дальнейшем расщепляли ферментом рестрикции HindIII с получением ДНК-фрагмента размером приблизительно 1,3 т.п.о., в то время как плазмиду pESC-L-GLELO расщепляли ферментом рестрикции ApaI и, после затупления концов, в дальнейшем расщепляли ферментом рестрикции HindIII с получением ДНК-фрагмента размером приблизительно 8,7 т.п.о. Эти ДНК-фрагменты лигировали с получением плазмиды pESC-L-GLELO:Δ5. Эту плазмиду расщепляли ферментом рестрикции PvuII, и получаемый в результате фрагмент размером приблизительно 3,2 т.п.о. встраивали в SmaI-сайт pUC-LEU2 с получением плазмиды pUC-LEU-GLELO:Δ5.
Штамм YPH499 S. cerevisiae (Stratagene) котрансформировали плазмидой pUC-URA-Δ12:Δ6 и плазмидой pUC-LEU-GLELO:Δ5. Трансформированные штаммы отбирали по способности расти на агаризованной (2% агара) среде SC-Leu,Ura, содержащей, на литр, 6,7 г дрожжевого азотистого основания без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смеси 1,25 г аденина сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глютаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 1,2 г триптофана). Среди полученных таким образом штаммов один штамм назвали штаммом ARA3-1.
(2) Получение и анализ трансформированных штаммов продуцирующих арахидоновую кислоту дрожжей
Штамм ARA3-1 трансформировали соответственно плазмидами pYE22m и pYE-MAPAP1. Трансформированные штаммы отбирали по способности расти на агаризованной (2% агара) среде SC-Trp,Leu,Ura, содержащей, на литр, 6,7 г дрожжевого азотистого основания без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислотного порошка (смеси 1,25 г аденина сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глютаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина и 6 г треонина). Среди полученных таким образом штаммов для каждой плазмиды отбирали любые 4 штамма.
Каждый их этих штаммов культивировали при 30οС в течение 1 дня в указанной выше жидкой среде SC-Trp,Leu,Ura (10 мл), а затем 1 мл каждой культуры вносили в 10 мл жидкой среды SG-Trp,Leu,Ura, содержащей, на литр, 6,7 г дрожжевого азотистого основания без аминокислот (DIFCO), 20 г галактозы и 1,3 г аминокислотного порошка (смеси 1,25 г аденина сульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глютаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина и 6 г треонина), и культивировали при 15°С в течение 7 дней с последующим анализом жирных кислот в клетках. В таблицах 3 и 4 демонстрируется жирнокислотная характеристика клеток и содержание жира и масла в клетках, соответственно.
Как показано выше, в случае, когда происходящий из M. alpina ген РАР1 в высокой степени экспрессировался в продуцирующих арахидоновую кислоту дрожжевых клетках, внутриклеточное содержание жирных кислот увеличивалось по сравнению с контрольными штаммами. Подобным образом, также увеличивались отношения содержания γ-линоленовой кислоты, DGLA и арахидоновой кислоты к общему содержанию жирных кислот.
Пример 8
Векторная конструкция для экспрессии в M. alpina
Векторами, используемыми для экспрессии в M. alpina, были pDuraSC, который делает возможной экспрессию требуемого гена с промотора GAPDH, и pDura5MCS, который делает возможной экспрессию требуемого гена с промотора гистона.
Для экспрессии РАР1 в клетках M. alpina векторы конструировали следующим образом. Плазмиду pCR-PAP1 расщепляли ферментом рестрикции PstI, а затем частично расщепляли ферментом рестрикции XhoI. Среди получаемых в результате ДНК-фрагментов фрагмент размером приблизительно 1,1 т.п.о. вырезали и встраивали между PstI- и XhoI-сайтами в сайте для множественного клонирования вектора pDuraSC или pDura5MCS. Получаемые в результате конструкции назвали плазмидами pDuraSC-PAP1 и pDura5MCS-PAP1, соответственно.
Получение трансформированных штаммов M. alpina
Ауксотрофный по урацилу штамм Δura-3, полученный из M. alpina, как описано в заявке на патент WO2005/019437 (озаглавленной “Method of Breeding Lipid-Producing Fungus”), использовали в качестве хозяина и трансформировали этими плазмидами с помощью способа доставки частиц. Для отбора трансформированных штаммов использовали агаризованную среду SC (0,5% дрожжевого азотистого основания без аминокислот и сульфата аммония (DIFCO), 0,17% сульфата аммония, 2% глюкозы, 0,002% аденина, 0,003% тирозина, 0,0001% метионина, 0,0002% аргинина, 0,0002% гистидина, 0,0004% лизина, 0,0004% триптофана, 0,0005% треонина, 0,0006% изолейцина, 0,0006% лейцина, 0,0006% фенилаланина и 2% агара).
Оценка трансформантов M. alpina
Каждый получаемый в результате трансформированный штамм засевали в 4 мл среды GY (2% глюкозы, 1% дрожжевого экстракта) и культивировали с покачиванием при 28°С в течение 3 или 4 дней. Клетки собирали с помощью фильтрации, и РНК экстрагировали с использованием заводского набора RNeasy (QIAGEN). Для синтеза кДНК использовали систему SuperScript First-Strand для ОТ-ПЦР (Invitrogen). Для установления экспрессии с введенной конструкции и общей экспрессии каждого гена проводили ОТ-ПЦР с использованием следующих наборов праймеров.
Штаммы, трансформированные плазмидой pDuraSC-PAP1
Использованными для установления экспрессии РАР1 с введенной конструкции праймерами были:
MaGAPDHpfw: CACACCACACATTCAACATC (SEQ ID NO: 20) и
D2: CGAAGCCGGCAAAGGCGGCAGTCG (SEQ ID NO: 21).
Использованными для установления общей экспрессии РАР1 праймерами были:
D1: CATGGGTTGCTTCGCGCGCAAGACG (SEQ ID NO: 22) и
D2.
Штаммы, трансформированные плазмидой pDura5MCS-PAP1
Использованными для установления экспрессии РАР1 с введенной конструкции праймерами были:
PD4P: CGCATCCCGCAAACACACAC (SEQ ID NO: 23) и
D2.
Использованными для установления общей экспрессии РАР1 праймерами были:
D1 и
D2.
Основываясь на результатах указанной выше ОТ-ПЦР, были отобраны трансформанты, демонстрирующие высокий уровень экспрессии каждого гена, как экспрессии с введенной конструкции, так и общей экспрессии: штаммы Gp-PAP1-49 и Hp-PAP1-2 из штаммов, трансформированных плазмидами pDuraSC-PAP1 и pDura5MCS-PAP1, соответственно.
Каждый из этих штаммов засевали в среду GY (4 мл) и культивировали с покачиванием при 125 оборотах в минуту при 28οС. На 4 день культивирования все клетки собирали с помощью фильтрации и лиофилизировали. Часть (приблизительно 10-20 мг) высушенных клеток обрабатывали с помощью метода с использованием соляной кислоты/метанола для превращения жирных кислот в клетках в соответствующие метиловые эфиры, а затем подвергали экстракции гексаном. После отгонки гексана жирные кислоты анализировали с помощью газовой хроматографии. В таблицах 5-7 ниже резюмированы внутриклеточное содержание жирных кислот и продукция арахидоновой кислоты на единицу среды.
Как показано выше, высокий уровень экспрессии гена РАР1 в М. alpina позволял повысить как внутриклеточное содержание жирных кислот, так и отношение содержания арахидоновой кислоты к общему содержанию жирных кислот. Кроме того, также можно было увеличить продукцию арахидоновой кислоты на единицу среды.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ НЕСТАНДАРТНОГО ФОРМАТА
SEQ ID NO:5: праймер
SEQ ID NO:6: праймер
SEQ ID NO:7: праймер
SEQ ID NO:8: праймер
SEQ ID NO:9: праймер
SEQ ID NO:10: праймер
SEQ ID N0:11: праймер
SEQ ID NO:12: праймер
SEQ ID NO:13: праймер
SEQ ID NO:14: праймер
SEQ ID NO:15: праймер
SEQ ID NO:20: праймер
SEQ ID NO:21: праймер
SEQ ID NO:22: праймер
SEQ ID NO:23: праймер















Настоящее изобретение относится к области биотехнологии и может быть использовано в пищевой промышленности. Из Mortierella alpina получен новый ген, который кодирует фосфатазу фосфатидной кислоты (ЕС 3.1.3.4), определена открытая рамка считывания и выведена аминокислотная последовательность фермента, названного MaPAP1. Описано получение рекомбинантного белка путем трансформации клеток-хозяев, где при использовании в этом качестве дрожжевых клеток или клеток липидпродуцирующих грибков обеспечивается не только увеличение общего содержания жирных кислот (ЖК), но и изменение жирнокислотного состава клетки, выражающееся, в частности, в повышении отношения количества арахидоновой кислоты к общему количеству ЖК. 8 н.п. ф-лы, 7 табл., 2 ил., 8 пр.
1. Нуклеиновая кислота, которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты, характеризующаяся нуклеотидной последовательностью, указанной в любом из подпунктов (а)-(е)ниже:
(a) нуклеотидная последовательность, которая кодирует белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO: 2;
(b) нуклеотидная последовательность, которая поддается гибридизации в условиях использования 2 X SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO: 4, при условии, что остатки 457-459, 613-615 и 778-780 SEQ ID NO:4 сохраняются;
(c) нуклеотидная последовательность, которая состоит из нуклеотидной последовательности, идентичной на 95% или больше нуклеотидной последовательности, состоящей из SEQ ID NO:4, при условии, что остатки 457-459, 613-615 и 778-780 SEQ ID NO:4 сохраняются;
(d) нуклеотидная последовательность, которая кодирует аминокислотную последовательность, идентичную на 95% или больше аминокислотной последовательности, состоящей из SEQ ID NO:2, при условии, что остатки 153, 205 и 260 SEQ ID NO:2 сохраняются; или
(е) нуклеотидная последовательность, которая поддается гибридизации в условиях использования 2 X SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO:2, при условии, что остатки 153, 205 и 260 SEQ ID NO:2 сохраняются.
2. Нуклеиновая кислота, которая кодирует белок, обладающий активностью фосфатазы фосфатидной кислоты, характеризующаяся нуклеотидной последовательностью, указанной в любом из подпунктов (а)-(с) ниже:
(a) нуклеотидная последовательность, указанная в SEQ ID NO:4;
(b) нуклеотидная последовательность, кодирующая белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO:2; или
(c) нуклеотидная последовательность, указанная в SEQ ID NO:1.
3. Нуклеиновая кислота, которая кодирует белок, обладающий способностью к обеспечению требуемой жирнокислотной характеристики, которая характеризуется нуклеотидной последовательностью, указанной в любом из подпунктов (а)-(е) ниже:
(a) нуклеотидная последовательность, которая кодирует белок, который состоит из аминокислотной последовательности, указанной в SEQ ID NO:2;
(b) нуклеотидная последовательность, которая поддается гибридизации в условиях использования 2 X SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, состоящей из SEQ ID NO:4, при условии, что остатки 457-459, 613-615 и 778-780 SEQ ID NO:4 сохраняются;
(с) нуклеотидная последовательность, которая состоит из нуклеотидной последовательности, идентичной на 95% или больше нуклеотидной последовательности, состоящей из SEQ ID NO:4, при условии, что остатки 457-459, 613-615 и 778-780 SEQ ID NO:4 сохраняются;
(d) нуклеотидная последовательность, которая кодирует белок, который состоит из аминокислотной последовательности, идентичной на 95% или больше аминокислотной последовательности, состоящей из SEQ ID NO:2, при условии, что остатки 153, 205 и 260 SEQ ID NO:2 сохраняются;
(e) нуклеотидная последовательность, которая поддается гибридизации в условиях использования 2 X SSC при 50°C с нуклеиновой кислотой, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей белок, состоящий из аминокислотной последовательности, указанной в SEQ ID NO:2, при условии, что остатки 153, 205 и 260 SEQ ID NO:2 сохраняются.
4. Белок, обладающий активностью фосфатазы фосфатидной кислоты, указанный в подпункте (а) или (b) ниже:
(a) белок, который состоит из аминокислотной последовательности, показанной в SEQ ID NO:2; или
(b) белок, который состоит из аминокислотной последовательности, идентичной на 95% или больше аминокислотной последовательности, состоящей из SEQ ID NO:2, при условии, что остатки 153, 205 и 260 SEQ ID NO:2 сохраняются.
5. Белок, обладающий способностью к обеспечению требуемой жирнокислотной характеристики, который указан в подпункте (а) или (b) ниже:
(а) белок, который состоит из аминокислотной последовательности, показанной в SEQ ID NO:2; или
(b) белок, который состоит из аминокислотной последовательности, идентичной на 95% или больше аминокислотной последовательности, состоящей из SEQ ID NO:2, при условии, что остатки 153, 205 и 260 SEQ ID NO:2 сохраняются.
6. Белок, обладающий активностью фосфатазы фосфатидной кислоты, состоящий из аминокислотной последовательности, указанной в SEQ ID NO:2.
7. Рекомбинантный вектор, включающий нуклеиновую кислоту по любому из пп.1-3.
8. Трансформант для продуцирования фосфатазы фосфатидной кислоты, трансформированный рекомбинантным вектором по п.7.
| Способ прогнозирования тяжелой формы сальмонеллеза у новорожденных | 1990 |
|
SU1748068A1 |
| PILLAI M | |||
| et al., Biochim | |||
| Biophys | |||
| Acta, 1998, 1393, №1, 128-136 | |||
| ЛОСЬ Д.А | |||
| Успехи биол | |||
| химии, 2001, т.41, 163-198. | |||

