Область применения
Изобретение относится к использованию антивоспалительного цитокина интерлейкина-10 (ИЛ-10) у больных с нейродегенеративными нарушениями, вызванными гипоксией, ишемией, судорожной активностью, механической травмой. Изобретение также относится к фармакологической композиции, содержащей интерлейкин-10.
Уровень техники
Одной из главных задач нейрофармакологии является разработка нового класса лекарственных средств, которые в отличие от ныне существующих препаратов, узконаправленного действия, обладали бы широким спектром своего лечебного воздействия. Известно, что различные по своей этиологии нейродегенеративные процессы в мозге, вызванные, в частности, ишемией/гипоксией, судорожной активностью, механической травмой, инфекциями, такими заболеваниями ЦНС, как болезни Паркинсона и Альцгеймера, а также аутоиммунными заболеваниями, характеризуются рядом сходных патогенетических механизмов, включающих локальную активацию микроглии и астроцитов, повышенную продукцию потенциально нейротоксических провоспалительных цитокинов, воспалительный ответ, активацию иммунной системы, экзитотоксичность и гипервозбудимость нейронов (Gendelman H.E. J. of NeuroVirology, 2002, v. 8, 474-479; Qian L., Block M.L., J. of Pharmacology and Experimental Therapeutics, 2006, v. 319, 44-52; Gupta Y.K. and Chauhan A. Indian J. Medical Research, 2011, v. 133, 15-26; Li G., Bauer S., Nowak M., J. of Seizure, 2011, v. 20, 249-256; Yirmiya R., Goshen I. Brain, Behavior, and Immunity, 2011, v. 25, 181-213). Острый воспалительный ответ рассматривается как механизм врожденного иммунитета, который направлен на защиту организма от повреждающих воздействий. Однако при хроническом воспалении макрофаги иммунной системы выделяют в области повреждения токсические вещества, включающие провоспалительные цитокины и свободные радикалы кислорода, которые приводят к разрушению клеток в поврежденной области мозга. Для прерывания такого хронического воспалительного процесса в разных тканях организма существуют разные механизмы регуляции воспалительного ответа, ключевую роль в которых играют антивоспалительные цитокины, обладающие одновременно иммуномодулирующими, антивоспалительными и трофическими функциями. Известно, что ЦНС, являющаяся иммуннопривилегированной областью, изолирована от клеток и медиаторов иммунной системы с помощью гематоэнцефалического барьера. Тем не менее выяснилось, что она содержит свои собственные иммуннорегуляторные клетки, включающие эндотелиальные клетки, микроглию, астроциты и олигодендроциты (Gendelman H.E. J. of NeuroVirology, 2002, v. 8, 474-479). Особую роль здесь играют микроглия и астроциты, которые оказывают как повреждающее, так и нейропротектирующее действие при реализации воспалительного ответа в локальной области мозга. В норме глиальные клетки способны процессировать антигены, экспрессировать молекулы главного комплекса гистосовместимости классов I и II, регулировать продукцию и захват экзитотоксинов, секретировать про- и антивоспалительные цитокины. Эти клетки активируются стимулами окружающей среды и участвуют в процессах нейрональной пластичности, формировании памяти и нейрогенезе (Yirmiya R., Goshen I. Brain, Behavior, and Immunity, 2011, v. 25, 181-213). Однако в условиях, когда иммунная система активируется ишемией/гипоксией, судорожной активностью нейронов, механической травмой или инфекциями, также как и сильным психофизиологическим стрессом, микроглия и астроциты изменяют свою морфологию и функциональные свойства и начинают продуцировать большие количества потенциально нейротоксических провоспалительных цитокинов, что приводит к нарушению процессов нейрональной пластичности, формирования памяти, нейрогенеза и в конечном итоге гибели центральных нейронов. Согласно современным представлениям повреждающие эффекты вышеназванных патогенных стимулов опосредованы провоспалительными цитокинами, продуцируемыми резидентными клетками мозга, прежде всего микроглией и астроцитами (Rothwell N.J. J. of Physiology, 1999, v. 514.1, 3-17). Известно, что индуцируемая провоспалительными цитокинами гибель нейронов мозга может быть опосредована следующими патогенетическими механизмами: 1. активацией фагоцитарной NADPH оксидазы (РНОХ), обнаруженной в микроглии, 2. активацией индуцибельной формы синтазы оксида азота (iNOS) в глии и 3. фагоцитозом нейронов микроглией.
При повреждениях мозга, связанных с такими нейродегенеративными заболеваниями, как болезнь Паркинсона, болезнь Альцгеймера, аутоиммунные заболевания, а также при механической травме, гипоксии/ишемии или инфекциях, экспрессия таких провоспалительных цитокинов, как интерлейкин-1, интерлейкин-6 и фактор некроза опухоли-альфа, в мозге резко возрастает, что может приводить к развитию хронического воспалительные ответа и последующей гибели клеток мозга (Rothwell N.J. J. of Physiology, 1999, v. 514.1, 3-17).
Ишемия/гипоксия
Известно, что патогенетические механизмы нейродегенеративных заболеваний мозга имеют мультифакторный характер. Например, во время ишемического инсульта повреждение клеток мозга происходит в три фазы: 1 фаза связана с прерыванием кровообращения и обеспечения клеток кислородом и питательными веществами; 2 фаза обусловлена массивным высвобождением возбуждающего нейромедиатора глутамата (экзитотоксичность) и 3 фаза гибели связана с хроническим воспалением, индуцируемым высвобождением провоспалительных цитокинов.
В обзоре Gupta Y.K. and Chauhan A. Indian J. Medical Research, 2011, v. 133, 15-26, отмечается, что до настоящего времени клинические испытания различных нейроактивных веществ как нейропротекторов при ишемии/гипоксии, а было протестировано большое число терапевтических агентов, включая антагонисты рецепторов глутамата, блокаторы кальциевых каналов, антиоксидантов и т.д., не дали результатов. К настоящему времени из новых препаратов за рубежом одобрен только рекомбинантный активатор тканевого плазминогена (rtPA) для лечения ишемического инсульта. Недостатком rtPA является то, что лечение им имеет узкое терапевтическое окно (около 3 часов) и повышает риск внутримозговых кровоизлияний. Однако отмечается, что использование в качестве нейропротекторов антивоспалительных веществ может быть многообещающим. Это заключение подтверждается, в частности, на моделях фокальной ишемии на крысах, где было обнаружено, что антивоспалительный цитокин ИЛ-10, введенный в латеральный желудочек мозга или хвостовую вену, редуцировал зону инфаркта мозга посла окклюзии правой срединной артерии (Spera P.A., Ellison J.A., Nauroscience Letters, 1998, v. 251(3), 189-192). Кроме того, было показано что, повышенная экспрессия ИЛ-10 повышает сопротивляемость клеток мозга мышей к фокальной ишемии мозга.
Судорожные расстройства
Известно, что гипоксия/ишемия, механическая травма и инфекции, индуцирующие в мозге локальные воспалительные процессы и процессы апоптоза, часто приводят к последующему развитию судорожной активности в нейронах, что, в свою очередь, повышает риск у человека заболевания эпилепсиями (Ravizza Т., Balosso S., Vezzani A. Neuroscience Letters, 2011, DOL10.1016 / J. neulet. 2011.02.040; Fiala M., Avagyan H., Merino J.J., Pathophysiology, 2012, DOI: 1016/ J.pathophys. 2012.02.003). Эти наблюдения указывают на то, что такие медиаторы воспалительных процессов, как провоспалительные цитокины, принимают участие в механизмах эпилептогенеза. Известны две линии доказательств участия провоспалительных цитокинов в механизмах эпилептогенеза: 1. во время индукции судорожной активности в локальных областях мозга уровень этих иммуномодуляторов возрастает и 2. фармакологическая блокада специфических провоспалительных сигнальных каскадов в клетках мозга оказывает антиэпилептогенное действие (Ravizza Т., Balosso S., Neuroscience Letters, 2011, DOI:10.1016 / J. neulet. 2011.02.040). По мнению некоторых авторов, хроническое воспаление является фундаментальным механизмом хронической эпилепсии (Rodgers K.M., Hutchinson M.R., J. of Brain, 2009, v. 132, 2478-2486). Надо отметить, что в настоящее время все существующие антиэпилептические средства направлены на устранение симптомов заболевания, а не самих патогенетических механизмов, лежащих в их основе. Имеются основания считать, что фармакологическая блокада воспалительных процессов в мозге может быть ключом к созданию нового класса антиэпилептических препаратов, направленных на устранение развития патогенетических механизмов эпилепсии.
Механическая травма
Механическое повреждение нейронов центральной и периферической нервной системы сопровождается повышением экспрессии таких провоспалительных цитокинов, как ИЛ-1 и фактор некроза опухоли-альфа (Knoblach S.M., Faden A.I., Experimental Neurology, 1998, v. 153(1), 143-151; Sawada Т., Sano M., Neuroscience Letters, 2007, v. 417, 55-60). Причем антивоспалительный цитокин ИЛ-10, введенный как до, так и после травмы, оказывает явно выраженное нейропротектирующее действие.
Инфекции
Локальное приложение липополисахарида (компонента оболочки грамотрицательных бактерий), имитирующее реакцию центральной нервной системы на инфекционный агент, к неокортексу мозга крысы приводило к развитию эпилептиформной активности в области введения (Rodgers K.M., Hutchinson M.R., Brain, 2009, v. 132, 2478-2486). Причем этот проконвульсивный ответ был опосредован рецепторами к провоспалительному цитокину ИЛ-1. Липополисахарид вызывает также гибель нейронов в первичной нейроглиальной культуре клеток среднего мозга, индуцируя продукцию провоспалительного цитокина фактора некроза опухоли-альфа (Qian L., Block M.L., J. of Pharmacology and Experimental Therapeutics, 2006, v. 319, 44-52). Антивоспалительный цитокин ИЛ-10 (3-30 нг / мл) оказывает нейропротектирующее действие на вызванную липополисахаридом гибель нейронов.
Используемые в настоящее время в клинике стероидные и нестероидные противовоспалительные препараты имеют ряд существенных недостатков при лечении нейродегенеративных заболеваний мозга. Так, такие стероидные препараты, как гюкокортикоиды, при их хроническом применении сами по себе оказывают повреждающее действие на нейроны и нарушают когнитивные функции мозга. Показано, что гюкокортикоиды ухудшают выживаемость нейронов ЦНС после ишемии, механической травмы, судорожной активности, HIV-1 инфекции и токсического действия бета-амилоида (Gendelman H.E. J. of NeuroVirology, 2002, v. 8, 474-479). Нестероидные противовоспалительные препараты также имеют ряд существенных недостатков при лечении нейродегенеративных заболеваний мозга. Большинство препаратов этой группы являются неселективными ингибиторами фермента циклооксигеназы, отвечающего за выработку простагландинов и тромбоксана из арахидоновой кислоты. В свою очередь, простагландины оказывают провоспалительное действие. Поскольку патогенетические механизмы нейродегенеративных заболеваний мозга имеют мультифакторный характер, то узконаправленный механизм действия этих препаратов далеко не всегда способен устранять последствия хронического воспалительного процесса. Кроме того, препараты этой группы имеют ряд побочных нежелательных эффектов прежде всего на сердечно-сосудистую систему, желудочно-кишечный тракт и ряд других органов организма (Yirmiya R., Goshen I. Brain, Behavior, and Immunity, 2011, v. 25, 181-213).
Одним из перспективных подходов для разработки нового класса нейропротекторов может быть использование эндогенных антивоспалительных цитокинов, обладающих одновременно иммуномодулирующими, антивоспалительными и трофическими свойствами. Это их действие связано, прежде всего, со способностью ингибировать образование провоспалительных цитокинов. Типичными представителями эндогенных антивоспалительных цитокинов, которые продуцируются резидентными клетками мозга являются интерлейкин-10 (ИЛ-10) и трансформирующий фактор роста-бета (ТФР-бета). Оба эти антивоспалительных цитокина оказывают явно выраженное нейропротектирующее действие на разных моделях нейрологических нарушений у животных. Тем не менее сравнение влияния ИЛ-10 и ТФР-бета на продукцию индуцируемых липополисахаридом провоспалительных цитокинов (ИЛ-6 и фактора некроза опухоли-альфа) и оксида азота в сочетанной культуре астроцитов и клеток микроглии показало, что в отличие от ТФР-бета, который подавлял продукцию только оксида азота, ИЛ-10 подавлял образование как оксида азота, так и провоспалительных цитокинов (Ledeboer A., Breve J.J, Glia, 2000, v. 30(2), 134-142). Таким образом, по сравнению с ТФР-бета антивоспалительный цитокин ИЛ-10 оказывал более сильный депрессирующий эффект на продукцию провоспалительных цитокинов в культуре глиальных клеток мозга.
ИЛ-10, первоначально названный фактором ингибирования синтеза цитокинов, является плейотропным цитокином, модулирующим различные внутриклеточные сигнальные пути. В зависимости от того, какие пути активируются, ИЛ-10 может регулировать иммунный ответ, воспалительные процессы, дифференцировку, а также пролиферацию или выживание различных клеток, включая клетки мозга в ответ на повреждающие воздействия.
Функционально активные рецепторы к антивоспалительному цитокину ИЛ-10 обнаруживаются на астроцитах, олигодендроцитах и микроглии в мозге, причем как астроциты, так и микроглия способны синтезировать этот цитокин (Strle К., Zhou J.H., Crit. Rev. Immunol., 2001, v. 21(5), 427-449). На периферии и в центральной нервной системе млекопитающих ИЛ-10 обнаруживается как нековалентно связанный гомодимер (35 кДа у мыши и 36 кДа у человека). Он обладает иммуномодулирующими, антивоспалительными и трофическими свойствами. ИЛ-10 способен специфически и с высоким сродством связываться со своими рецепторами, расположенными на плазматической мембране различных клеток, стимулируя рост стволовых клеток и тимоцитов, а также дифференцировку В клеток. Рецепторный комплекс к ИЛ-10 содержит две разные субъединиц: IL-10Rα и IL-10Rβ. Бета-субъединица выполняет вспомогательную функцию и необходима для инициации сигнальной трансдукции сигнала при активации ИЛ-10 рецептора.
Сущность изобретения
Предметом настоящего изобретения является применение интерлейкина-10 для лечения нейродегенеративных повреждений мозга, вызванных гипоксией, судорожной активностью нейронов и механической травмой.
Другим предметом настоящего изобретения является создание фармацевтической композиции, которая облегчает негативные последствия нейродегенеративных повреждений мозга, вызванных гипоксией, судорожной активностью нейронов и механической травмой.
Данный метод может быть использован у больных с нарушенными функциями ЦНС, подвергнувшихся действию гипоксии или ишемии после инсульта, механической травмы, хронических инфекций или аутоиммунных заболеваний с целью коррекции или ослабления симптомов повреждения нейронов мозга. Степень повреждения нейронов ЦНС может быть оценена по нарушению когнитивных функций и/или склонности нейронов к развитию судорожной активности.
Для использования ИЛ-10 по настоящему изобретению для терапевтического лечения (включая профилактическое лечение) млекопитающих, включая человека, его обычно формулируют в фармацевтическую композицию. В соответствии с этим аспектом, изобретение относится к фармацевтической композиции, содержащей соединение по настоящему изобретению в сочетании с фармацевтически приемлемым носителем, разбавителем или наполнителем.
Типичную композицию получают смешиванием соединения по настоящему изобретению и носителя, разбавителя или наполнителя. Подходящие носители, разбавители и наполнители хорошо известны специалисту в данной области. Растворители обычно выбирают исходя из их безопасности (GRAS), признанной специалистам в данной области, при введении млекопитающему. Обычно, безопасные растворители представляют собой нетоксичные водные растворители, такие как вода и другие нетоксичные растворители, которые растворимы или способны смешиваться с водой. Поскольку ИЛ-10 является хорошо растворимым белком, то в качестве подходящего растворителя может быть использован физиологический раствор или раствор Кребса-Рингера.
Композиция может также содержать одну или несколько буферов, стабилизаторы, поверхностно-активные вещества, увлажняющие вещества, эмульгаторы, отдушки, ароматизаторы, технологические добавки и другие известные добавки, обеспечивающие хорошую презентацию лекарственного средства или способствующие получению фармацевтического продукта.
Композиция может быть получена путем смешивания при температуре окружающей среды при соответствующем рН и при желаемой степени чистоты, с физиологическими приемлемыми носителями, то есть носителями, которые являются нетоксичными для реципиента при используемых дозах и концентрациях. рН может находиться в пределах около 6 до около 8.
Соединение по настоящему изобретению для использования здесь является предпочтительно стерильным. В частности, композиции, применяемые для введения in vivo, должны быть стерильными. Такая стерилизация легко достигается фильтрованием через стерилизующие фильтрационные мембраны.
ИЛ-10 в соединении по изобретению обычно может быть в виде твердой композиции, лиофилизированной композиции или в виде водного раствора. ИЛ-10 или его аналоги могут вводиться больным внутривенно, интерназально или интрацеребровентрикулярно в физиологических концентрациях, зависящих от типа и тяжести повреждения.
Вещества могут вводиться в однократной или многократных дозах. ИЛ-10 может быть очищен из естественных источников или получен с помощью ДНК-рекомбинантной технологии.
В качестве общего суждения, концентрации ИЛ-10 или его синтетических биологически активных аналогов в мозге должны быть достаточны для того, чтобы оказывать лечебное действие на данное нейрологическое заболевание. Можно вводить один ИЛ-10 или совместно с другими терапевтическими агентами, включающими факторы роста, антиоксиданты, антибактериальные или антивирусные препараты. Приемлемые дозы ИЛ-10 при центральном введении могут быть от 1 нг/мл до 1 мкг/мл, а при системном введении - между 1-15 мкг на 100 г веса.
Изобретение поддерживается экспериментальными данными (примерами), в которых было найдено следующее:
(1) Антивоспалительный цитокин ИЛ-10 оказывает противогипоксическое действие на нейроны мозга;
(2) Антивоспалительный цитокин ИЛ-10 устраняет развитие потенциально эпилептогенной постгипоксической гипервозбудимости в нейронах мозга;
(3) По сравнению с большими концентрациями ИЛ-10 его низкие концентрации более эффективно улучшают способность этого антивоспалительного цитокина оказывать нейропротектирующее действие на развитие постгипоксической гипервозбудимости в центральных нейронах;
(4) Антивоспалительный цитокин ИЛ-10 повышает порог развития судорожной активности в мозге у свободноподвижных животных.
Экспериментальные данные приведены в прилагаемых примерах и фигурах, на которых демонстрируется:
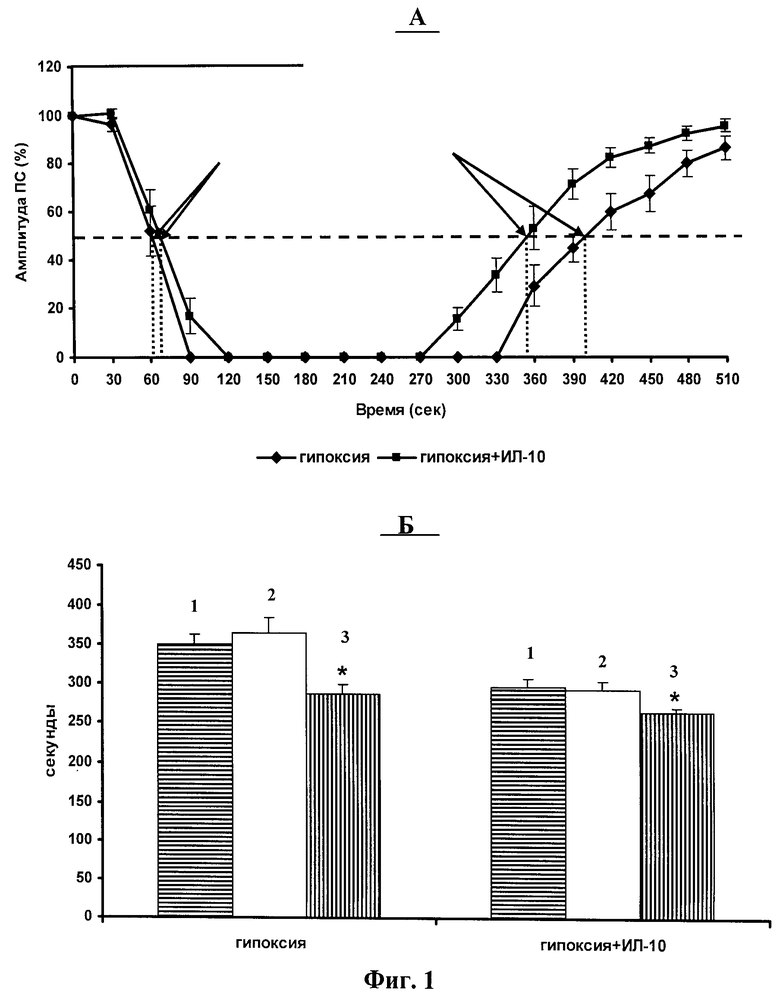
фиг.1 - Влияние ИЛ-10 на вызванное эпизодом гипоксии угнетение амплитуды популяционного спайка (ПС). А - динамика изменения амплитуды ПС во время эпизода гипоксии и гипоксии+ИЛ-10. Б - длительность (в секундах) угнетающего действия гипоксии и гипоксии+ИЛ-10 на амплитуду ПС в 1, 2 и 3 эпизодах гипоксии.
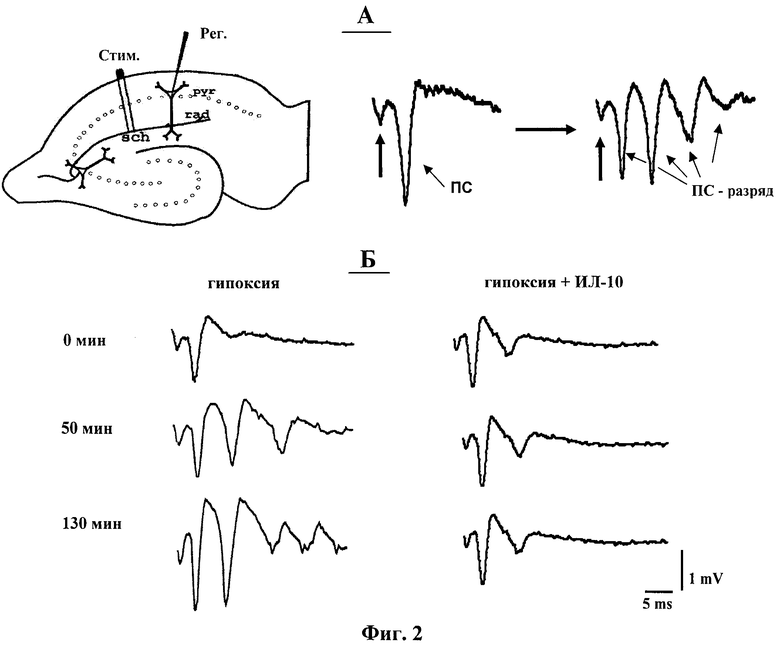
фиг.2 - Схема расположения электродов в срезе гиппокампа и типичные формы пре- и постгипоксических ПС-ответов. А: слева - локализация стимулирующего (Стим.) и регистрирующего (Рег.) электродов соответственно в радиальном (rad) и пирамидном (pyr) слоях поля СА1 срезов гиппокампа; Sch - коллатерали Шафера/комиссуральные волокна; справа - форма ПС в норме и при его трансформации в эпилептиформный паттерн после эпизодов гипоксии. Б: типичные формы ПС-ответов, регистрируемых до (0) и через 50 и 130 минут после эпизодов гипоксии. Слева - после эпизодов гипоксии; справа - после эпизодов гипоксии на фоне ИЛ-10 (1 нг/мл).
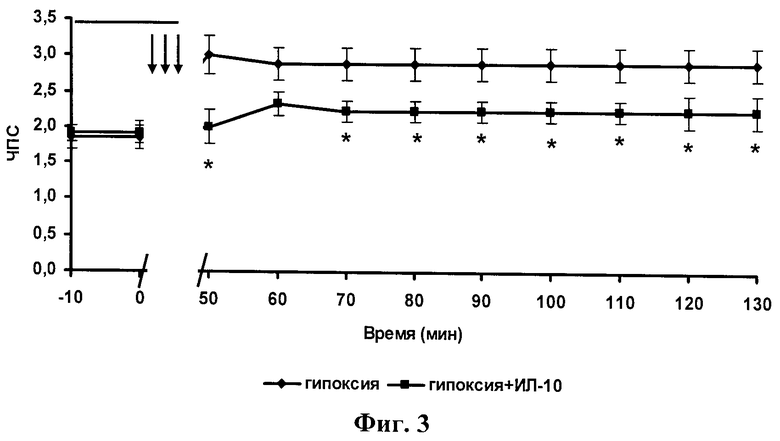
фиг.3 - Динамика изменения числа популяционных спайков в разряде (ЧПС) при действии эпизодов гипоксии без и на фоне ИЛ-10 (1 нг/мл). Звездочками отмечены постгипоксические значения после эпизодов гипоксии на фоне ИЛ-10, отличающиеся достоверно от значений после эпизодов гипоксии без ИЛ-10. Прямая линия на графике отмечает время приложения ИЛ-10. Три стрелки - три 3-минутных эпизода гипоксии.
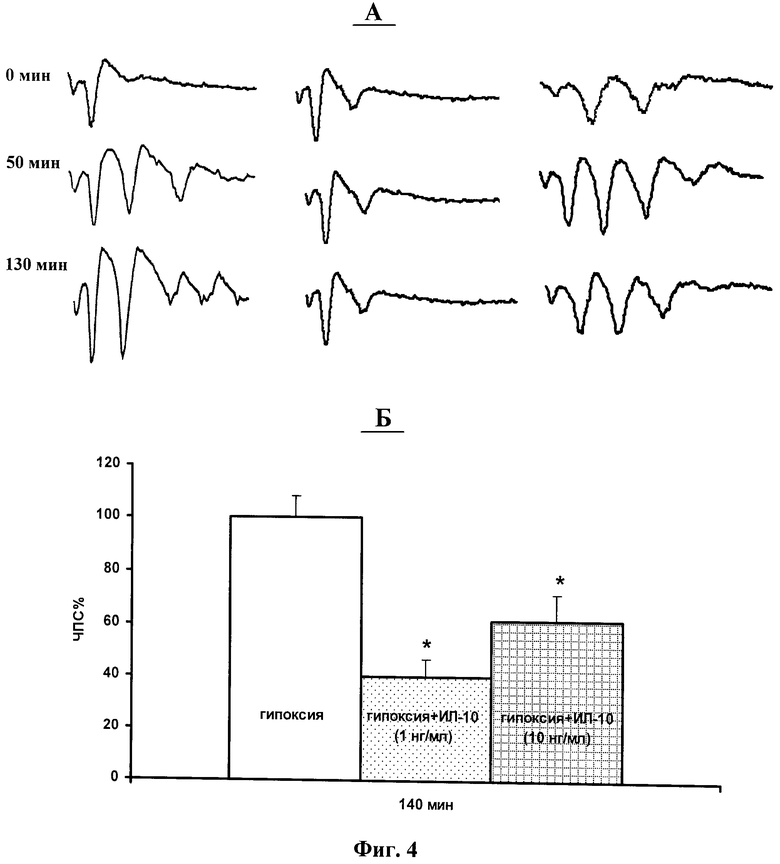
фиг.4 - А - типичные формы ПС-ответов, регистрируемых до (0) и через 50 и 130 минут после эпизодов гипоксии. Слева - после эпизодов гипоксии; в центре - после эпизодов гипоксии на фоне ИЛ-10 (1 нг / мл); справа - после эпизодов гипоксии на фоне ИЛ-10 (10 нг/мл). Б - концентрационная зависимость модулирующего действие ИЛ-10 на величину значения ЧПС через 140 минут после окончания эпизодов гипоксии в процентах относительно их прегипоксических значений.
фиг.5 - Динамика изменения параметров: порога генерации дополнительного спайка (ПГДС) (А) и ЧПС (Б) до (-20 - 0 минут) и после трех эпизодов гипоксии (40-140 минут) (↓↓↓). Значения параметра ПГДС представлены как процент от его прегипоксических значений (100%); значения параметра ЧПС представлены в абсолютных величинах. Одной и двумя звездочками отмечены соответственно постгипоксические значения после трех эпизодов гипоксии на фоне ИЛ-10 (10 нг / мл) или на фоне ИЛ-10 (1 нг / мл), отличающихся достоверно от значений после трех эпизодов гипоксии. Линия на графике указывает время приложения ИЛ-10.
фиг.6 - BrdU-меченые клетки в области имплантированного электрода через 11 дней после имплантации. А и С - организация тканевого рубца (Scar) в двух последовательных 50 мкм срезах гиппокампа мозга крысы при низком увеличении. В и D - те же отображения, но при большом увеличении, соответственно для А и С. Е - детализированная структура BrdU-меченых клеток на отображении D, где видна высокая митотическая активность клеток в области механической травмы. SPGL - супрагранулярный слой; DG - зубчатая фасция; bv - кровеносные сосуды/капилляры; fis -fissure; CA3/CA4 - поля гиппокампа; hilus - хилус гиппокампа.
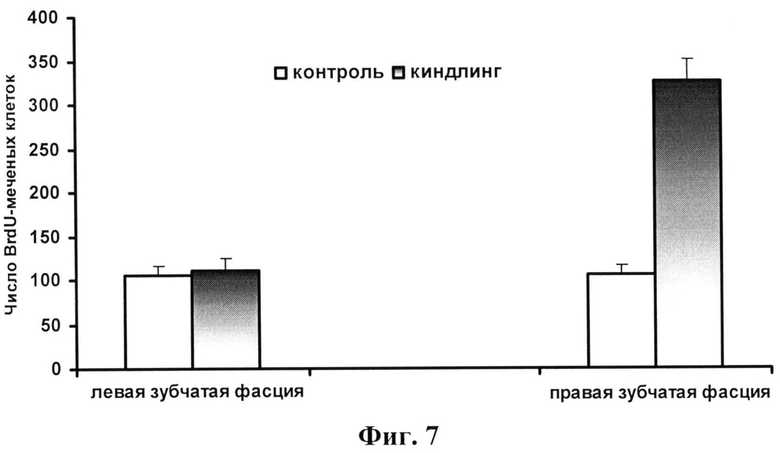
фиг.7 - Число BrdU-меченых клеток в правой (область стимуляции) и левой (контралатеральная область, где не производилась стимуляция и не были имплантированы электроды) зубчатой фасции гиппокампальной формации у контрольной и киндлинговой групп животных. Наблюдаются достоверные (Р=0,01) отличия в числе BrdU-меченых клеток для правой зубчатой фасции у контрольной и киндлинговой групп.
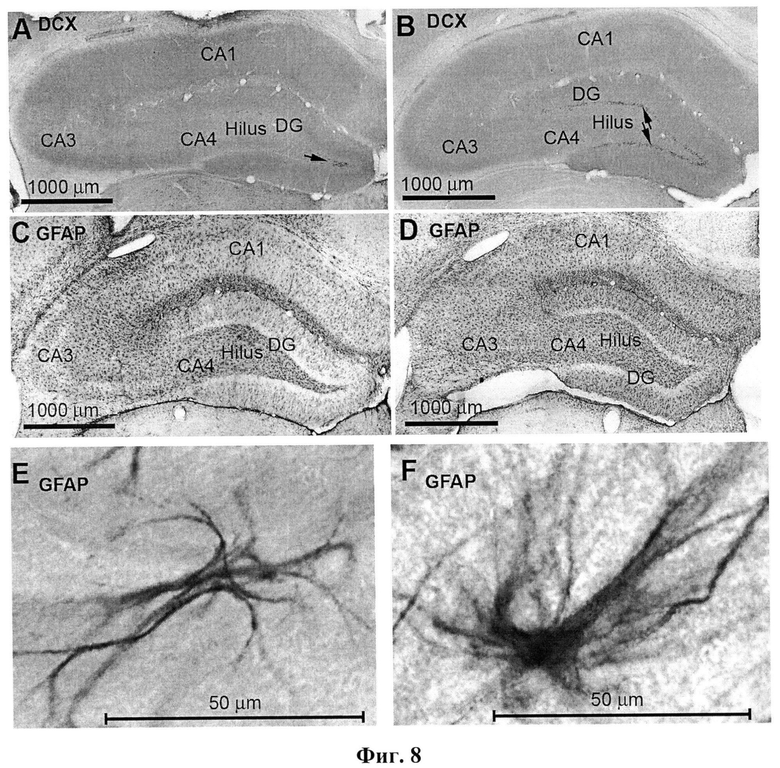
фиг.8 - DCX- и GFAP-меченые клетки в гиппокампе крыс, подвергнутых эпилептогенной киндлинговой стимуляции. А и С - левый (нестимулируемый) гиппокамп. В и D - правый (стимулируемый) гиппокампа за пределами рубца. Е - GFAP-меченые клетки в левом (нестимулируемом) гиппокампе. F - GFAP-меченые клетки в правом (стимулируемом) гиппокампе. Обозначения сокращений такое же, как на рисунке 6.
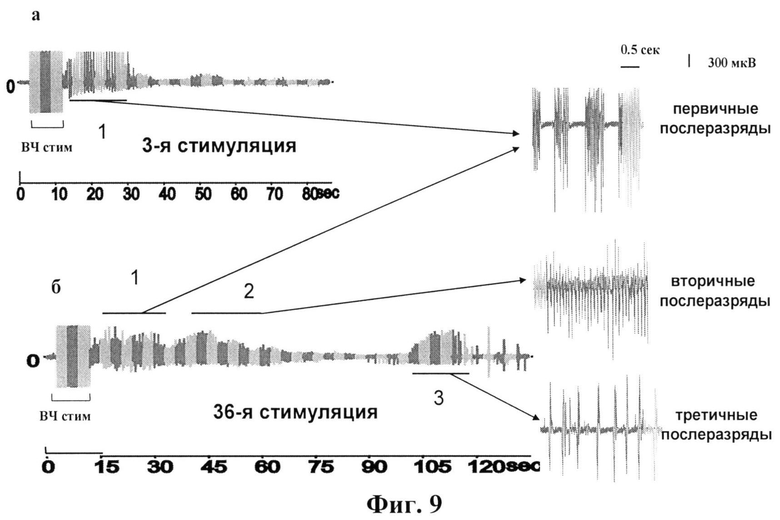
фиг.9 - Примеры ЭЭГ-послеразрядов во время парциального электрического киндлинга дорсального гиппокампа у свободноподвижных крыс после первых трех стимуляций (3 стимуляция) и 36 стимуляций (36 стимуляция). Период стимуляции показан на ЭЭГ-ответе как ВЧ стим. Цифрами 1, 2 и 3 показаны соответственно первичный, вторичный и третичный компоненты послеразрядов.
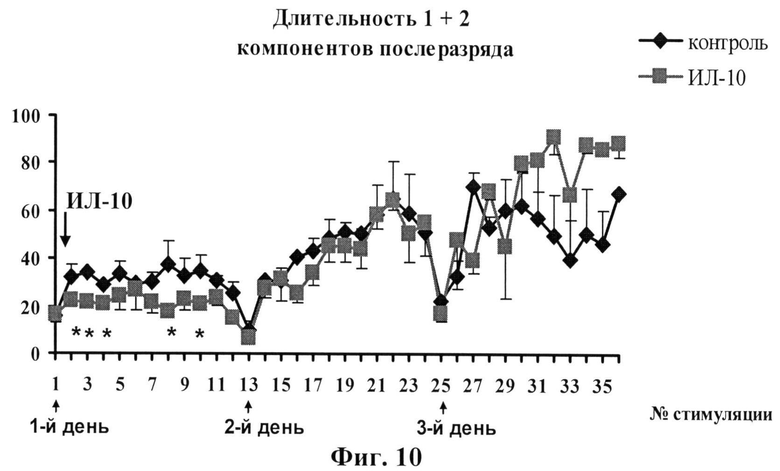
фиг.10 - Влияние однократного внутригиппокампального введения ИЛ-10 (1 нг/5 мкл) на развитие эпилептиформных разрядов при электрическом киндлинге дорсального гиппокампа. На графике показана зависимость суммы первых двух компонентов (1+2) ЭЭГ-послеразрядов от числа стимуляций. ИЛ-10 вводили за 1 час до начала киндлинговой стимуляции только в 1 день стимуляции.
Сущность и преимущества заявляемого изобретения иллюстрируются следующими примерами.
Пример 1. Антивоспалительный цитокин ИЛ-10 оказывает противогипоксическое действие на нейроны мозга
Работа осуществлялась с соблюдением принципов гуманного обращения с животными (директива Европейского сообщества 86/609/ЕС). В экспериментах использовали двухмесячных самцов крыс Вистар. Срезы гиппокампа толщиной 250-300 мкм помещались в камеру погруженного типа и суперфузировались раствором Кребса-Рингера следующего состава (мМ): NaCl - 124, KCl - 3, MgSO4 - 2, CaCl2 - 2, NaHCO3 -26, KH2PO4 - 1,24, глюкоза - 10, О2/CO2 - 95%/5% (рН 7,4), при температуре раствора 32°С и скорости его протока 2,4 мл/мин. Регистрацию активности начинали через 5 часов от начала суперфузии. Популяционные спайки (ПС) регистрировались от пирамидных нейронов поля СА1 гиппокампа в ответ на электрическую тестирующую стимуляцию коллатералей Шаффера до, во время и после предъявления трех трехминутных эпизодов гипоксии, разделенных 10-минутными интервалами (фигура 2А). Эффективность гипоксического эпизода угнетать амплитуду ПС оценивалась по следующей формуле: Гэфф=Тв г-Тд г, где Гэфф - эффективность эпизода гипоксии угнетать амплитуду ПС (в секундах); Тд г - время, через которое амплитуда ПС снижается до 50% от ее прегипоксических значений при предъявлении эпизода гипоксии; Тв г - время, через которое амплитуда ПС восстанавливается до 50% от ее прегипоксических значений после окончания действия гипоксии. Электрическая активность нейронов измерялась в течение 6 часов после выделения среза гиппокампа на серии из 7 одиночных стимулов градуально увеличивающейся интенсивности от пороговой (для возникновения ПС) до максимальной, при которой не происходило дальнейшего увеличения амплитуды ПС. Частота импульсов в серии была 0,1 Гц, частота серий - 1 серия / 20 минут. Гипоксические эпизоды предъявлялись при следующих условиях: (1) без добавления в среду ИЛ-10; (2) через 10 минут после добавления в среду ИЛ-10 в концентрации 1 нг / мл. ИЛ-10 растворялся в среде суперфузии срезов гиппокампа. Общее время суперфузии с ИЛ-10 составляло 40 минут. ИЛ-10 удалялся из среды одновременно с окончанием последнего эпизода гипоксии. Кроме того, в отдельной серии ИЛ-10 (1 - 10 нг/мл) в разной концентрации добавлялся к срезам гиппокампа в отсутствии эпизодов гипоксии. Ввод вызванных электрических ответов в компьютер, а также их обработка в режиме off-line осуществлялись с помощью соответствующего программного обеспечения. Данные представляли в виде средняя±ошибка средней. Оценку достоверности эффектов ИЛ-10 в "гипоксических" срезах гиппокампа без приложения и при приложении ИЛ-10 осуществляли посредством сравнения параметров активности методом ANOVA (post-hoc Scheffe's test) для одних и тех же моментов времени регистрации активности. Для оценки достоверности эффектов ИЛ-10 на параметры активности в "негипоксических" (контрольных) срезах гиппокампа использовали парный t-тест Стьюдента. При значениях Р<0,05 различия считались достоверными.
На фигуре 1 и в таблице 1 показано модулирующее влияние ИЛ-10 на вызванное эпизодом гипоксии угнетение амплитуды ПС, регистрируемых в поле СА1 гиппокампа. Антивоспалительный цитокин снижал эффективность гипоксии, угнетая амплитуду ПС (фигура 1А). На фигуре 1Б и в таблице 1 показано влияние ИЛ-10 на длительность угнетающего действия гипоксии во время 1, 2 и 3 эпизодов гипоксии. Можно видеть, что ИЛ-10 снижал длительность угнетающего действия гипоксии на амплитуду ПС во всех предъявляемых эпизодах. Полученные результаты показали, что антивоспалительный цитокин ИЛ-10 в концентрации 1 нг/мл достоверно снижает эффективность гипоксии угнетать активность пирамидных нейронов поля СА1 гиппокампа во время гипоксического эпизода.
(Р=0,516*)
(Р=0,025*)
(Р=0,003**)
(Р=0,003*)
(Р=0,94**)
Пример 2. Антивоспалительный цитокин ИЛ-10 устраняет развитие потенциально эпилептогенной постгипоксической гипервозбудимости в нейронах мозга
В примере 2 выделение переживающих срезов гиппокампа крысы, а также регистрацию и измерение биоэлектрической активности производили также, как и в примере 1. Оценка развития постгипоксической гипервозбудимости нейронов в срезах гиппокампа, проявляющейся в форме эпилептиформных ПС-разрядов, производилась по разработанной в нашей лаборатории оригинальной методике. Измерялись два параметра такой активности: (1) порог стимулирующего тока (мкА), при котором появляются разряды ПС в ответ на одиночный теститующий стимул (ПГДС) и (2) число ПС в разряде (ЧПС) на стимул, интенсивность которого приводила к появлению второго ПС в ответе до добавления в среду суперфузии срезов гиппокампа ИЛ-10 и предъявления эпизодов гипоксии. Электрическая активность нейронов измерялась в течение 6 часов после выделения среза гиппокампа на серии из 7 одиночных стимулов градуально увеличивающейся интенсивности от пороговой (для возникновения ПС) до максимальной, при которой не происходило дальнейшего увеличения амплитуды ПС.
Из фигуры 2 видно, что ИЛ-10 в концентрации 1 нг/мл устранял развитие эпилептиформных ПС-разрядов, индуцируемых кратковременными эпизодами гипоксии. На фигуре 3 показана динамика параметра ЧПС после действия эпизодов гипоксии и эпизодов гипоксии на фоне ИЛ-10. Эти результаты показали, что антивоспалительный цитокин ИЛ-10 в концентрации 1 нг/мл устраняет развитие эпилептической постгипоксической гипервозбудимости в пирамидных нейронах поля СА1 гиппокампа.
Пример 3. По сравнению с большими концентрациями ИЛ-10 его низкие концентрации более эффективно оказывют нейропротектирующее действие на развитие постгипоксической гипервозбудимости в центральных нейронах
В примере 3 выделение переживающих срезов гиппокампа мозга крысы, а также регистрацию и измерение биоэлектрической активности производили также, как и в примерах 1 и 2. Целью данной работы было исследование влияние ИЛ-10 в концентрациях 1 и 10 нг/мл на развитие вызванных кратковременными эпизодами гипоксии эпилептиформных разрядов в пирамидных нейронах поля СА1 в срезах гиппокампа крысы. ИЛ-10 в концентрациях 1 нг/мл или 10 нг/мл добавляли в суперфузионную среду за 10 минут до гипоксических эпизодов и оставляли в среде в течение всего времени предъявления эпизодов гипоксии. Общее время суперфузии среза гиппокампа с ИЛ-10 составляло 40 минут. В контрольных экспериментах (без предъявления эпизодов гипоксии) ИЛ-10 в различных концентрациях также апплицировался в течение 40 минут в те же интервалы времени суперфузии срезов гиппокампа, как и в опытах с гипоксическими эпизодами.
На фигурах 4 и 5 показано, что ИЛ-10 в концентрациях 1 и 10 нг/мл устранял развитие эпилептиформных ПС-разрядов, индуцируемых кратковременными эпизодами гипоксии. Однако нейропротектирующее действие ИЛ-10 в концентрации 1 нг/мл было более эффективным. Таким образом эти результаты обнаружили, что ИЛ-10 в концентрации 1 нг/мл полностью устраняет индуцируемое эпизодами гипоксии развитие эпилептиформной активности, тогда как в концентрации 10 нг/мл его нейропротектирующий эффект был менее выражен. Это указывает на то, что более низкие дозы ИЛ-10 оказывают более эффективное нейропротектирующее действие в отношении способности эпизодов гипоксии индуцировать развитие постгипоксической гипервозбудимости в центральных нейронах.
Пример 4. Антивоспалительный цитокин ИЛ-10 повышает порог развития судорожной активности в мозге у свободноподвижных животных
В этих работах использовалась киндлинговая модель развития судорожной активности: парциальный электрический киндлинг (раскачка) дорсального гиппокампа и регистрация электроэнцефалограммы (ЭЭГ) в фокусе развития судорожной активности в гиппокампе у свободноподвижных крыс. Использовались самцы крыс Вистар массой 320-350 грамм. Работа осуществлялась с соблюдением принципов гуманного обращения с животными (директива Европейского сообщества 86/609/ЕС). Под нембуталовым наркозом (60 мг/кг, в.б.) крысам имплантировались три свитые нихромовые проволочки диаметром 0,1 мм, две из которых являлись биполярным стимулирующим электродом, а третья - монополярным электродом для регистрации электрографических послеразрядов (ПР) в ответ на электрическую стимуляцию той же области гиппокампа. Нихромовые проволочки приклеивались к направляющей для введения инъекционной канюли, кончик которой располагался на 0,2 мм выше кончика электродов. Индифферентным электродом был винт, вкрученный в назальные части черепа. Электроды вводились в правый дорсальный гиппокамп по координатами: 3 мм каудальнее брегмы, 3,7 мм латеральное средней линии и 3 мм ниже твердой мозговой оболочки. Они фиксировались к костям черепа зубным цементом, подпаивались к разъемам для последующей электрической стимуляции и ЭЭГ-регистрации в условиях свободного поведения животных. Морфологический контроль показал, что кончики электродов локализованы в области апикальных дендритов поля СА1. Через неделю после операции животные случайным образом разделялись на две группы: (1) контрольную, в которой парциальный электрический киндлинг вырабатывался после однократного введения 5 мкл стерильного раствора 0,9% NaCl в область электродов, и (2) экспериментальную, в которой парциальный электрический киндлинг вырабатывался после однократного введения 5 мкл раствора ИЛ-10 (1 нг) в стерильном 0,9% NaCl. Электрическая стимуляция начиналась через 1 час после введения растворов. Использовался протокол быстрого киндлинга. Крысы обеих групп получали 12 стимуляций в день через каждые 30 минут в течение трех дней. Каждая стимуляция состояла из 9-секундной пачки прямоугольных импульсов длительностью 0,1 мс и частотой 20 Гц. Интенсивность стимула (200-400 мкА) была пороговой для индукции электрографических ПР длительностью не менее 15-25 секунд при первых стимуляциях. ПР идентифицировались как иктальная (судорожная) электрографическая спайковая активность с амплитудой по крайней мере в 2-3 раза выше значений ЭЭГ до стимуляции и начинающейся примерно через 1-2 секунды после окончания электрической стимуляции. ЭЭГ активность регистрировалась в режиме on-line аналоговым ЭЭГ усилителем и через аналого-цифровой преобразователь вводилась в компьютер с соответствующим программным обеспечением для последующего анализа структуры и длительности электрографических ПР в режиме off-line. Кроме длительности ПР, тяжесть судорог оценивалась также по модифицированной поведенческой шкале Racine: стадия 1 - отряхивание мокрой собаки (ОМС); 2 - ОМС+жевание; 3 - клонус передних конечностей (КПК); 4 - КПК+вставание на задние лапы и 5 - КПК с вставанием на задние лапы и падение на спину. Поскольку в данной работе использовалась процедура быстрого парциального киндлинга, то электрическая стимуляция заканчивалась, если в поведении животных возникали проявления генерализованных поведенческих судорог, характерных для 4 и 5 стадий. Методики приготовления срезов мозга для световой и электронной микроскопии, методы BrdU (5-бром-2-деоксиуридин; маркер пролиферирующих клеток) - иммунногистохимии, DCX (doublecortin, маркер мигрирующих вновь образованных нейронов) - иммунногистохимии и GFAP (маркер глиального фибриллярного кислого белка) - иммунногистохимии, а также метод трехмерной реконструкции синаптических структур описаны в работе (Kraev I.V., Neuroscience, 2009, v. 162(2), 254-267).
Полученные результаты показали следующее. Механическая травма, вызванная имплантацией электродов в ЦНС, сама по себе без всякой стимуляции приводила к увеличению BrdU-меченых клеток в области образовавшего рубца. На фигуре 6 показаны три типа BrdU-меченых клеток через 11 дней после имплантации электродов в зубчатую фасцию мозга крысы: 1. астроциты (черные стрелки), 2. созревающие нейроны и микроглия/астроциты (красные стрелки) и 3. эндотелиальные клетки (синие стрелки). Однако эпилептогенная киндлинговая стимуляция приводила к дополнительному увеличению числа BrdU-меченых клеток как в области имплантированных электродов, так и в отдаленных областях мозга в фокусе развития судорожной активности.
В зубчатой фасции гиппокампа в локусе электрической раскачивающей стимуляции по морфологическим критериям было выделено 6 типов дендритных шипиков: стволовые, пеньковые, тонкие, разветвленные тонкие, грибовидные и разветвленные грибовидные. По сравнению с нормальными животными у животных после киндлинга выявлялось достоверное увеличение пропорции тонких дендритных шипиков и снижение пропорции грибовидных шипиков (уменьшение примерно в два раза). Киндлинговая стимуляция приводила также к достоверному увеличению числа BrdU-меченых клеток в зоне развития фокальной судорожной активности (фигура 7), а также числа DCX- и GFAP-меченых клеток (фигура 8). Таким образом, киндлинговая эпилептогенная стимуляция сопровождалась пролиферацией и активацией глиальных клеток, нейрогенезом и структурными перестройками синаптических структур в фокусе стимуляции процессов, лежащих в основе развития судорожной активности в мозге. Пример ЭЭГ-послеразрядов и изменение их структуры во время электрического киндлинга дорзального гиппокампа у свободноподвижных крыс показаны на фигуре 9.
Структура ПР состояла из трех компонентов, возникающих во временной последовательности: (1) спайковые разряды, сгруппированные в пачки длительностью 200-500 мс и разделенные интервалом 200-500 мс (первичный компонент ПР); (2) за первичным компонентом следовали непрерывные одиночные спайковые разряды (вторичный компонент ПР) и (3) за вторичным компонентом через несколько секунд следовали одиночные спайковые разряды, разделенные 200-500 мс интервалами (третичный компонент ПР). Общая длительность этих компонентов (сумма 1, 2 и 3 компонентов) ПР в ответ на первые стимуляции не превышала 15-25 сек, однако после 36 стимуляций она увеличивалась до 80-120 секунд. На фигуре 10 показана зависимость длительности суммы 1 и 2 компонентов ПР (иктальные или судорожные компоненты) от числа стимуляций в контроле и после однократного введения ИЛ-10 (1 нг/5 мкл) в область стимуляции. Во время первых 12 стимуляций (1 день стимуляции) наблюдались только фокальные судороги (поведенческие стадии 1 и 2) у обеих групп животных. При этом общая длительность электрографических ПР в области стимуляции, отражающая эффективность развития фокальных судорог, у контрольной группы животных (введение 0,9% NaCl) составляла в среднем 45 секунд. Тогда как у животных, которым вводили ИЛ-10, длительность этих разрядов была достоверно ниже (примерно 23 секунды). На второй день стимуляций (без введения ИЛ-10) изменения в длительностях электрографических ПР и тяжести поведенческих судорог не отличались у сравниваемых групп животных. К концу третьего дня стимуляции (без введения ИЛ-10) общая длительность ПР достигала 90-120 секунд, а тяжесть поведенческих судорог - 3-4 стадии. Как видно из рисунка 10, на третий день стимуляции у крыс обеих групп наблюдалось статистически недостоверное увеличение длительности ПР. Таким образом, анализ длительностей отдельных компонентов ПР в процессе раскачивающей стимуляции обнаружил, что введение ИЛ-10 приводит к достоверному снижению суммарной длительности первичного и вторичного компонентов в течение первых шести часов после введения цитокина (первый день стимуляции) по сравнению с контролем. Изменения в длительности третичного компонента ПР (интериктального или межсудорожного) после введения ИЛ-10 не наблюдалось.
Таким образом, результаты проведенных исследований показали, что однократное внутригиппокампальное введение ИЛ-10 в концентрации 1 нг/5 мкл угнетает развитие судорожной активности (фокальных судорог) в области стимуляции при парциальном киндлинге у свободноподвижных животных в течение первых часов после введения. Однако ИЛ-10 не влиял ни на длительность «интериктального» компонента фокальных послеразрядов, ни на тяжесть поведенческих судорог. Полученные результаты указывают, что антивоспалительный цитокин ИЛ-10 в использованной концентрации, помимо своего анти-гипоксического действия, оказывает протектирующее влияние на инициацию «иктального», но не «интериктального», компонента судорожной активности в нейронах гиппокампа.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЦЕНКИ КОМПЛЕКСНОЙ ЗАЩИТЫ КЛЕТОК ПРИ НЕЙРОВОСПАЛЕНИИ НА МОДЕЛИ IN VITRO ПРИ ВОЗДЕЙСТВИИ АГОНИСТОМ ПАР1 | 2022 |
|
RU2796524C1 |
| СПОСОБ ЧАСТИЧНОГО ВОССТАНОВЛЕНИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ НЕЙРОННЫХ СЕТЕЙ IN VITRO В УСЛОВИЯХ ИХ ЗНАЧИТЕЛЬНОГО ПОВРЕЖДЕНИЯ | 2015 |
|
RU2594065C1 |
| Способ лечения нейропатической боли | 2021 |
|
RU2780139C1 |
| ПРИМЕНЕНИЕ ПРЕПАРАТА "СЕМАКС" В КАЧЕСТВЕ СРЕДСТВА АКТИВАЦИИ АДАПТИВНОГО ИММУНИТЕТА | 2015 |
|
RU2584234C1 |
| НОВОЕ ПРИМЕНЕНИЕ | 2004 |
|
RU2416426C2 |
| ПРИМЕНЕНИЕ СТВОЛОВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, И ИХ КОМПОЗИЦИЙ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ (ВАРИАНТЫ) | 2017 |
|
RU2742828C2 |
| Способ диагностики нейроиммунных нарушений у детей раннего возраста с эпилепсией | 2021 |
|
RU2767271C1 |
| МОДУЛЯЦИЯ ПУТЕЙ ОБМЕНА ПРОСТАГЛАНДИНА/ЦИКЛООКСИГЕНАЗЫ | 2007 |
|
RU2448712C2 |
| 7-ГИДРОКСИЭПИАНДРОСТЕРОН, ОБЛАДАЮЩИЙ НЕЙРОПРОТЕКТИВНОЙ АКТИВНОСТЬЮ | 2001 |
|
RU2307654C2 |
| Способ прогнозирования возникновения неонатальных судорог у новорожденных из группы высокого перинатального риска | 2019 |
|
RU2722022C1 |


Изобретение относится к области медицины и фармации и касается применения интерлейкина-10 для лечения нейродегенеративных повреждений мозга, вызванных гипоксией/ишемией, судорожной активностью нейронов и механической травмой. 4 пр., 10 ил.
Применение интерлейкина-10 для лечения нейродегенеративных повреждений мозга, вызванных гипоксией/ишемией, судорожной активностью нейронов и механической травмой.
| С.Г.Левин, О.В.Годухин: "Протектирующее действие интерлейкина-10 на развитие эпилептиформной активности, вызванной кратковременными эпизодами гипоксии, в срезах гиппокампа крыс" | |||
| ЖУРНАЛ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ, 2006, том 56, N3, с.379-383 | |||
| О.В.Годухин, С.Г.Левин, Е.Ю.Парнышкова: "Влияние интерлейкина-10 на развитие эпилептиформной активности в |

