Область, к которой относится изобретение
Настоящее изобретение относится к самоорганизующимся пептидным наночастицам, содержащим эпитопы В-клеток и/или эпитопы Т-клеток. Кроме того, изобретение относится к применению таких наночастиц для вакцинации.
Предпосылки создания изобретения
Адаптивная иммунная система имеет два различных вида ответа, гуморальный иммунный ответ и клеточный иммунный ответ. Первый характеризуется антителогенезом, при котором такие антитела связываются с поверхностными эпитопами патогенов, в то время как последний характеризуется цитотоксическими Т-лимфоцитами (CTL), которые уничтожают инфицированные клетки. Оба вида иммунного ответа дополнительно стимулируются клетками Т-хелперов, которые активируют либо В-клетки, которые вырабатывают специфические антитела, связывающие патоген, либо Т-клетки, которые являются направленными против инфицированных Т-клеток.
Специфичность взаимодействия между антителами, которые вырабатываются В-клетками, и патогеном определяется поверхностными структурами патогена, так называемыми эпитопами В-клеток, в то время как специфичность взаимодействия CTL с инфицированными клетками-мишенями осуществляется с помощью эпитопов Т-клеток, которые презентируются на поверхностных молекулах целевой клетки, так называемых молекулах класса I основного комплекса гистосовместимости (МНС I). Этот тип эпитопов Т-клеток (CTL эпитопы) представляет собой фрагменты белков из патогена, которые продуцируются инфицированной клеткой. В завершение, специфичность взаимодействия клеток Т-хелперов с соответствующей В-клеткой или CTL определяется связыванием рецепторных молекул Т-хелперных клеток с другим типом эпитопов Т-клеток (HTL эпитопами), которые презентируются классом II молекул МНС (МНС II) на В-клетках или CTL-клетках.
Связывание антитела с эпитопами В-клеток требует, чтобы эпитоп В-клетки приобретал определенную пространственную структуру, такую же структуру, что и та, которую эпитоп В-клетки имеет в естественной среде, то есть, тогда, когда он находится на поверхности патогена. Эпитоп В-клетки может состоять более чем из одной пептидной цепи и является организованным в пространственную структуру с помощью каркасной структуры белка.
Эпитопы Т-клеток, однако, не требуют наличия определенной пространственной структуры, предпочтительнее они связываются с соответствующей молекулой МНС I или МНС II весьма специфическим образом. Эпитопы CTL урезаются до размера 9 аминокислот в длину для оптимальной презентации молекулами МНС I, в то же время HTL эпитопы осуществляют подобное взаимодействие с молекулами МНС II, но могут быть длиннее, чем 9 аминокислот. В контексте данного изобретения является важным, что связывание эпитопов с молекулами МНС соответствует весьма простым правилам, то есть, только пептиды со специфическими характеристиками будут способными связываться с соответствующей молекулой МНС и, таким образом, являются полезными в качестве эпитопов. Эти характеристики были тщательно исследованы и на основе известных ценных эпитопов, были также разработаны программы прогнозирования, которые являются способными предсказывать с высокой точностью эпитопы, способные связываться с молекулами МНС. Пептидные цепочки, состоящие из нескольких таких эпитопов Т-клеток в линейной пептидной цепи, в настоящее время были сконструированы в качестве кандидатных вакцин.
В общем случае эффективная вакцина должна индуцировать сильный гуморальный, а также сильный клеточный иммунный ответ. Было продемонстрировано, что сильный гуморальный иммунный ответ может быть достигнут с помощью повторяемого антигенного дисплея эпитопов В-клеток. Вирусоподобные частицы (VLP) могут использоваться в качестве эффективного средства для презентации эпитопов В-клеток регулярным, повторяемым и жестким образом, и, таким образом, VLP в настоящее время широко используются для вакцинных конструкций. Другой подход для повторяемого антигенного дисплея был описан в патенте ЕР 1594469 В1. В этом патенте были сконструированы самоорганизующиеся пептидные наночастицы (SAPN), состоящие из тримерных и пентамерных белковых доменов олигомеризации, которые повторяемым образом демонстрируют эпитопы В-клеток на своей поверхности. Эпитопы В-клеток присоединяли на конце доменов олигомеризации для гарантии того, что эпитопы В-клеток являются презентированными на поверхности наночастиц в виде многочисленных копий. Один из наиболее часто встречающихся фрагментов олигомеризации белка представляет собой биспиральный структурный фрагмент, и этот фрагмент может эффективно использоваться для конструирования таких SAPN.
Краткое изложение сущности изобретения
Изобретение относится к самоорганизующимся пептидным наночастицам (SAPN), включающим эпитопы Т-клеток и/или эпитопы В-клеток. В частности, наночастицы в соответствии с изобретением состоят из агрегатов беспрерывной пептидной цепи, включающей два домена олигомеризации, связанные с помощью линкерного сегмента, где один или два домена олигомеризации имеют биспиральную структуру, которая включает эпитопы Т-клеток и/или эпитопы В-клеток в своей пептидной последовательности.
Изобретение также относится к способу вакцинации людей или животных, отличных от человека, при использовании таких самоорганизующихся пептидных наночастиц, включающих эпитопы Т-клеток и/или эпитопы В-клеток.
Краткое описание чертежей
Фигура 1: Структура мышиной МНС II молекулы I-Ad, ковалентно связанной с овальбуминовым пептидом (OVA323-339), который представляет собой HTL эпитоп для I-Ad. MHC II белок показан от вершины С-альфа линии серым цветом. Две спирали, образующие стенки сайтов связывания эпитопа, являются фланкирующими для пептида связывания. Пептид представлен шаростержневой моделью со всеми атомами черным цветом. Пептидный HTL эпитоп в своей связанной форме находится в растянутой конформации, как может быть более четко видно с использованием структуры только одного пептида в нижней части фигуры.
Фигура 2: Схематический рисунок "выровненных единиц" для тримерных и пентамерных доменов олигомеризации [с левой стороны, А)] тримерного и тетрамерного доменов олигомеризации [с правой стороны, В)], соответственно. Количество мономеров (строительные блоки) определяется наименьшим значением общего кратного (LCM) состояний олигомеризации двух доменов олигомеризации D1 и D2 строительных блоков. В выровненных единицах линкерные сегменты всех строительных блоков будут располагаться так близко друг к другу, насколько это является возможным, то есть настолько близко к центру пептидной наночастицы, насколько это является возможным и, таким образом, выровненные единицы подвергаться самосборке (самоорганизации) с образованием сферической наночастицы.
Фигура 3: Элементы внутренней симметрии додекаэдра/икосаэдра. Оси вращательной симметрии (двойная, тройная и пятикратная) показаны в виде линий, обозначенных как 2, 3 и 5. В А) мономерного строительного блока, состоящего из доменов олигомеризации D1 (слева, биспиральный домен с тройной симметрией), линкерного сегмента L (внизу) и домена олигомеризации D2 (справа; биспиральный домен с пятикратной симметрией), показаны так, что элементы внутренней симметрии доменов олигомеризации D1 и D2 накладываются на элементы симметрии многогранника. В В) демонстрируются полные биспиральные домены D1 и D2. Дополнительные объекты симметрии, полученные с помощью элементов тройной и пятикратной вращательной симметрии многогранника, демонстрируются в виде цилиндров, в то время, как исходная молекула демонстрируется как спираль, как представлено в А).
Фигура 4: Динамическое рассеяние света (DLS, А) и просвечивающая электронная микроскопия (ТЕМ, В) самоорганизующихся пептидных наночастиц, образованных из пептидов с последовательностью SEQ ID NO:8, Пример 1. DLS анализ показывает распределение размеров со средним диаметром частицы 32,01 нм и индексом полидисперсности 12,9% (А). ТЕМ изображения (В) показывают наночастицы того же размера, как и определено с помощью DLS.
Фигура 5: Просвечивающая электронная микроскопия (ТЕМ) самоорганизующихся пептидных наночастиц, образованных из пептидов с последовательностью SEQ ID NO:10, Пример 2. ТЕМ изображения показывают наночастицы с размером 25 нм.
Фигура 6: Просвечивающая электронная микроскопия (ТЕМ) самоорганизующихся пептидных наночастиц, образованных из пептидов с последовательностью SEQ ID NO:12, Пример 3. ТЕМ изображения показывают наночастицы с размером приблизительно от 20 до 30 нм.
Фигура 7: Просвечивающая электронная микроскопия (ТЕМ) самоорганизующихся пептидных наночастиц, образованных из пептидов с последовательностями SEQ ID NO:37 (полоска А) и SEQ ID NO:38 (полоска В), для вакцины против гриппа для человека и кур соответственно (Пример 9). ТЕМ изображения показывают наночастицы с размером приблизительно 25 нм.
Фигура 8: Просвечивающая электронная микроскопия (ТЕМ) самоорганизующихся пептидных наночастиц, образованных из пептидов с последовательностью SEQ ID NO:41, Пример 11. ТЕМ изображения показывают наночастицы с размером приблизительно 25 нм.
Подробное описание изобретения
Мономерные строительные блоки
Самоорганизующиеся пептидные наночастицы (SAPN) образуются из совокупности мономерных строительных блоков формулы (I), состоящих из беспрерывной цепи, включающей пептидный домен олигомеризации D1, линкерный сегмент L и пептидный домен олигомеризации D2
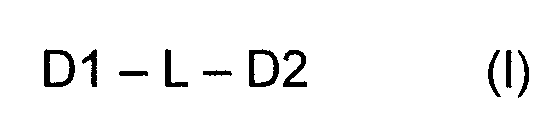 ,
,
где D1 представляет собой синтетический или природный пептид, обладающий тенденцией к образованию олигомеров (D1)m m субъединиц D1, D2 представляет собой синтетический или природный пептид, обладающий тенденцией к образованию олигомеров (D2)n n субъединиц D2, m и n каждый представляет собой цифру от 2 до 10, при условии, что m не является равным n и не является кратным n, а n не является кратным m, L представляет собой связь или короткую линкерную цепь, выбранную из необязательно замещенных атомов углерода, необязательно замещенных атомов азота, атомов кислорода, атомов серы и их комбинаций; либо D1, либо D2, либо оба D1 и D2 являются биспиральными и включают один или более эпитопов Т-клеток и/или эпитоп В-клеток в рамках домена олигомеризации и где D1, D2 и L являются необязательно дополнительно замещенными.
Пептид (или полипептид) представляет собой цепочку или последовательность аминокислот, ковалентно связанных с помощью амидных связей. Пептид может быть природным, природным модифицированным, частично синтетическим или полностью синтетическим. Под природным модифицированным, частично синтетическим или полностью синтетическим понимают тот, который не существует в природе. Термин аминокислота охватывает как существующие в природе аминокислоты, выбранные из 20 эссенциальных природных α-L-аминокислот, синтетических аминокислот, таких как α-D-аминокислоты, 6-аминогексановая кислота, норлейцин, гомоцистеин, или подобные им, а также существующие в природе аминокислоты, которые были модифицированы определенным путем для изменения некоторых свойств, таких, как заряд, например, такие, как фосфосерин или фосфотирозин, или подобные им. В производных аминокислот аминогруппа, образующая амидную связь, является алкилированной, или боковая цепь аминогруппы, группы гидрокси или тиогруппы является алкилированной или ацилированной, или боковая цепь группы карбокси является амидированной или эстерифицированной.
Короткая линкерная цепь L является выбранной из необязательно замещенных атомов углерода, необязательно замещенных атомов азота, атомов кислорода, атомов серы и их комбинаций, предпочтительно содержащих 1-60 атомов, в частности, 1-20 атомов в цепи. Такая короткая линкерная цепь представляет собой, например, цепь полиэтиленокси, цепь сахара или, предпочтительно, пептидную цепь, например, пептидную цепь, состоящую из 1-20 аминокислот, в частности, 1-6 аминокислот.
m и n каждый представляет собой число от 2 до 10, при условии, что m не является равным n и не является кратным n, а n не является кратным m. Предпочтительные комбинации m и n представляют собой комбинации, где m равно 2, а n равно 5, или m равно 3, а n равно 4 или 5, или m равно 4, а n равно 5. Кроме того, предпочтительными комбинациями m и n являются комбинации, в которых m равно 5, а n равно 2, или m равно 4 или 5, а n равно 3, или m равно 5, а n равно 4. Наиболее предпочтительными являются комбинации, в которых m или n равно 5.
Биспиральная нить представляет собой пептидную последовательность с беспрерывной моделью в основном гидрофобных остатков с интервалом в 3 и 4 остатка, которая подвергается самоорганизации с образованием мультимерной группы спиралей, как более подробно объясняется в данной заявке ниже.
"Биспиральная нить, которая включает эпитопы Т-клеток и/или В-клеток" означает, что соответствующий эпитоп является включенным в домен олигомеризации так, что аминокислотные последовательности на N-терминальном и C-терминальном концах эпитопа заставляют эпитоп адаптироваться к конформации, которая все еще является биспиральной наряду со свойствами олигомеризации домена олигомеризации, включающего эпитоп. В частности, "встроенный" исключает случай, когда эпитоп является присоединенным к любому концу биспирализованного домена олигомеризации.
В контексте этого документа термин эпитопы Т-клеток будет использоваться для отнесения как к СТ, так и HTL эпитопам.
Эпитопы Т-клеток связываются с МНС молекулами в растянутой конформации (Фигура 1). Таким образом, включение эпитопов Т-клеток в α-спиральную биспирализованную нить (сравни Фигуру 3А и Фигуру 1) не является обычной задачей белковой инженерии. В данном изобретении продемонстрировано, как эти пептидные последовательности, которые имеют растянутую конформацию, когда связываются с соответствующими молекулами МНС, могут, несмотря на это, встраиваться в α-спиральный биспирализованный домен олигомеризации.
Необязательные заместители D1, D2 и L представляют собой, например, эпитопы В-клеток, нацеливающие остатки или заместители, усиливающие адъювантные свойства наночастицы, такие, как иммуностимуляторная нуклеиновая кислота, предпочтительно, олигодезоксинуклеотид, содержащий дезоксиинозин, олигодезоксинуклеотид, содержащий дезоксиуридин, олигодезоксинуклеотид, содержащий фрагмент CG, или молекулу нуклеиновой кислоты, содержащую инозин и цитидин. Другие заместители, усиливающие адъювантные свойства наночастицы, представляют собой антимикробные пептиды, такие, как катионные пептиды, представляющие собой класс иммуностимуляторных, позитивно заряженных молекул, которые могут способствовать и/или улучшать адаптивный иммунный ответ. Пример такого пептида с иммунопотенциирующими свойствами представляет собой позитивно заряженный искусственный антимикробный пептид KLKLLLLLKLK (SEQ ID NO:63), который индуцирует сильный адаптивный иммунитет, запускаемый специфическим белком типа 2, после иммунизации на основе первичной вакцинации - вторичной прививки (бустеризации). Специфическим нацеливающим остатком считается заместитель, который представляет собой ER-нацеливающий сигнал, то есть, сигнальный пептид, который индуцирует транспорт белка или пептида к эндоплазматическому ретикулуму (ER). Другие необязательные заместители представляют собой, например, ацильную группу, например, ацетил, связанную с аминогруппой, в частности, с N-терминальной аминокислотой, или связь амино со свободной группой карбокси C-терминальной аминокислоты с образованием карбоксамидной группы.
Необязательные заместители, например, те необязательные заместители, которые описаны в данной заявке выше, являются предпочтительно соединенными с приемлемыми аминокислотами ближе к свободному концу домена олигомеризации D1 и/или D2. При самосборке пептидной наночастицы такие заместители будут присутствовать на поверхности SAPN.
В наиболее предпочтительном воплощении заместитель имеет другую пептидную последовательность S1 и/или S2, представляющую собой простое удлинение пептидной цепи D1 - L - D2 на любом из концов или на обоих концах для получения одной комбинированной пептидной последовательности любой из форм S1 - D1 - L - D2, D1 - L - D2 - 82 или S1 - D1 - L - D2 - S2, где S1 и S2 представляют собой пептидные заместители, как определено в данной заявке выше и ниже. Заместители S1 и/или S2 считаются такими, которые предназначены для удлинения последовательности D1 - L - D2 SAPN. Любая такая пептидная последовательность S1 - D1 - L - D2, D1 - L - D2 - S2 или S1 - D1 - L - D2 - S2 может экспрессироваться в рекомбинантной системе экспрессии белка в виде одной молекулы.
Предпочтительный заместитель S1 и/или S2 представляет собой эпитоп В-клетки. Другие рассматриваемые эпитопы В-клеток представляют собой гаптеновые молекулы, такие, как углевод или никотин, которые также присоединяются к концу доменов олигомеризации D1 и/или D2, и, таким образом, будут представляться на поверхности SAPN.
Очевидно, является также возможным присоединять более одного заместителя к доменам олигомеризации D1 и/или D2. Например, рассматривая пептидную последовательность S1 - D1 - L - D2 - S2, можно сказать, что к ней может быть ковалентно присоединен другой заместитель, предпочтительно, в положении, отдаленном от линкерного сегмента L, либо ближе к концам D1 и/или D2, либо где-либо в заместителях S1 и/или S2.
Является также возможным присоединять заместитель к линкерному сегменту L. В этом случае при сборке SAPN заместитель будет располагаться во внутренней полости SAPN.
Тенденция к образованию олигомеров означает, что такие пептиды могут образовывать олигомеры в зависимости от условий, например, при условиях денатурации они могут представлять собой мономеры, в то время, как при физиологических условиях они могут образовывать, например, тримеры. При предварительно определенных условиях они приобретают одно единственное состояние олигомеризации, которое является необходимым для образования наночастиц. Однако их состояние олигомеризации может быть изменено при изменяющихся условиях, например, от димеров до тримеров при увеличивающейся концентрации соли (Burkhard P. и др., Protein Science 2000, 9:2294-2301) или от пентамеров до мономеров при понижении значения рН.
Архитектура строительного блока в соответствии с формулой (I) четко отличается от вирусных капсидных белков. Вирусные капсиды состоят либо из одного единственного белка, который образует 60 олигомеров, либо числа, кратного ему, как например, частицы вируса гепатита В (ЕР 1262555, ЕР 0201416), или более чем из одного белка, который подвергается процессу самоорганизации с образованием вирусного капсида, который может приобретать также другие геометрические структуры, отличные от икосаэдра в зависимости от типа вируса (Fender P. и др., Nature Biotechnology 1997, 15:52-56). Самоорганизующиеся пептидные наночастицы (SAPN) настоящего изобретения также четко отличаются от вирусоподобных частиц, так как они (а) являются сконструированными из белков, отличных от вирусного капсида, и (b) полость внутри наночастицы является слишком малой для того, чтобы разместить ДНК/РНК цельного вирусного генома.
Пептидные домены олигомеризации являются хорошо известными (Burkhard P. и др., Trends Cell Biol 2001, 11:82-88). Наиболее простой домен олигомеризации, вероятно, представляет собой действительно биспирализованный участок фолдинга. Этот участок олигомеризации был продемонстрирован как такой, который представляет собой димер, тример, тетрамер и пентамер. Некоторые примеры представляют собой GCN4 лейциновую застежку, фибритин, тетрабрахион и СОМР, являющиеся димерными, тримерными, тетрамерными и пентамерными биспиральными нитями, соответственно.
Один или оба домена олигомеризации D1 и D2, независимо друг от друга, являются биспирализованными доменами. Биспирализованная нить представляет собой пептидную последовательность с непрерывной моделью в основном гидрофобных остатков с интервалом в 3 и 4 остатка, обычно последовательность, состоящую из семи аминокислот (семичленный повтор) или одиннадцати аминокислот (одиннадцатичленный повтор), который подвергается самоорганизации (сборке) с образованием мультимерной группы спиралей. Биспирализованные нити с последовательностями, включающими в некоторой степени нерегулярное распределение расположенных 3 и 4 остатков, также предполагаются. Гидрофобные остатки представляют собой, в частности, гидрофобные аминокислоты Val, Ile, Leu, Met, Tyr, Phe и Trp. Преимущественно гидрофобный означает, что, по крайней мере, 50% остатков должно быть выбрано из упомянутых гидрофобных аминокислот.
Например, в предпочтительном мономерном строительном блоке формулы (I) D1 и/или D2 представляет собой пептид любой из формул
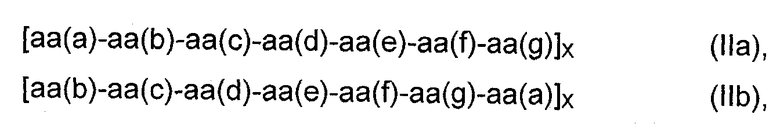

где аа означает аминокислоту или ее производную, аа(а), aa(b), aa(c), aa(d), аа(е), aa(f), и аа(g) являются одинаковыми или различными аминокислотами или их производными, предпочтительно аа(а) и aa(d) являются одинаковыми или различными гидрофобными аминокислотами или их производными; а Х представляет собой число от 2 до 20, предпочтительно 3, 4, 5 или 6.
Гидрофобные аминокислоты представляют собой Val, Ile, Leu, Met, Tyr, Phe и Trp.
Семичленный пептид представляет собой гептапептид формулы аа(а)-аа(b)-aa(c)-aa(d)-aa(e)-aa(f)-aa(g) (IIa) или любую их комбинацию формул от (IIb) до (IIg).
Предпочтительными являются мономерные строительные блоки формулы (I), в которых один или два пептидных домена олигомеризации D1 или D2 представляют собой
(1) пептид в соответствии с любой из формул (IIa) - (IIg), где Х равен 3, и аа(а) и aa(d) являются выбранными из 20 природных α-L-аминокислот, так, что сумма баллов из Таблицы 1 для этих 6 аминокислот составляет, по крайней мере, 14, и такие пептиды включают вплоть до 17 дополнительных групп из 7 членов; или
(2) пептид в соответствии с любой из формул (IIa) - (IIg), где Х равен 3, и аа(а) и aa(d) являются выбранными из 20 природных α-L-аминокислот, так, что сумма баллов из Таблицы 1 для этих 6 аминокислот составляет, по крайней мере 12, при условии, что одна аминокислота аа(а) является заряженной аминокислотой, способной к образованию внутриспиральных солевых мостиков с аминокислотой aa(d) или аа(g) соседней семичленной группы, или что одна аминокислота aa(d) является заряженной аминокислотой, способной к образованию внутриспиральных солевых мостиков с аминокислотой аа(а) или аа(е) соседней семичленной группы, и такие пептиды включают вплоть до двух дополнительных семичленных групп. Заряженная аминокислота, способная к образованию внутриспиральных солевых мостиков с аминокислотой соседней семичленной группы, представляет собой, например, Asp или Glu, если другие аминокислоты представляют собой Lys, Arg или His, или наоборот
Также предпочтительными являются мономерные строительные блоки формулы (I), где один или два пептидных домена олигомеризации D1 или D2 являются выбранными из следующих предпочтительных пептидов:
(11) Пептид в соответствии с любой из формул (IIa) - (IIg), где аа(а) является выбранным из Val, Ile, Leu и Met, и их производных, и aa(d) является выбранным из Leu, Met и Ile, и их производных.
(12) Пептид в соответствии с любой из формул (IIa) - (IIg), где один аа(а) представляет собой Asn, а другие аа(а) являются выбранными из Asn, Ile и Leu, a aa(d) представляет собой Leu. Такой пептид обычно представляет собой домен димеризации (m или n=2).
(13) Пептид в соответствии с любой из формул (IIa) - (IIg), где аа(а) и aa(d) оба представляют собой Leu или оба являются Ile. Такой пептид обычно представляет собой домен тримеризации (m или n=3).
(14) Пептид в соответствии с любой из формул (IIa) - (IIg), где аа(а) и aa(d) оба представляют собой Trp. Такой пептид обычно представляет собой домен пентамеризации (m или n=5).
(15) Пептид в соответствии с любой из формул (IIa) - (IIg), где аа(а) и aa(d) оба представляют собой Phe. Такой пептид обычно представляет собой домен пентамеризации или домен тетрамеризации (m или n=4 или 5).
(16) Пептид в соответствии с любой из формул (IIa) - (IIg), где аа(а) и aa(d) оба представляют собой либо Trp, либо Phe. Такой пептид обычно представляет собой домен пентамеризации (m или n=5).
(17) Пептид в соответствии с любой из формул (IIa) - (IIg), где аа(а) представляет собой либо Leu, либо Ile, и один aa(d) является Gln, а другие aa(d) являются выбранными из Gln, Leu и Met. Такой пептид потенциально может быть доменом пентамеризации (m или n=5).
Другие предпочтительные пептиды представляют собой пептиды (1), (2), (11), (12), (13), (14), (15), (16) и (17), как определено в данной заявке выше, и где дополнительно (21) по крайней мере, один аа(g) является выбранным из Asp и Glu, а аа(е) в следующей семичленной группе представляет собой Lys, Arg или His; и/или
(22) по крайней мере, один аа(g) является выбранным из Lys, Arg и His, а аа(е) в следующей семичленной группе представляет собой Asp или Glu, и/или
(23) по крайней мере, один аа(а - g) является выбранным из Lys, Arg и His, a аа(а - g) с интервалом 3 или 4 аминокислоты в последовательности представляет собой Asp или Glu. Такие пары аминокислот аа(а - g) представляют собой, например, аа(b) и аа(е) или aa(f).
Программы для прогнозирования биспирализованной нити, такие как COILS (http://www.ch.embnet.org/software/COILS_form.html; Gruber М. и др., J. Struct. Biol. 2006, 155(2):140-5) или MULTICOIL (http://groups.csail.mit.edu/cb/multicoil/cgi-bin/multicoil.cgi), могут предсказывать пептидные последовательности, образующие биспирализованную структуру. Таким образом, в предпочтительном мономерном строительном блоке формулы (I) D1 и/или D2 представляют собой пептид, который содержит, по крайней мере, последовательность из двух семичленных повторов, что может быть предсказано с помощью программы прогнозирования биспиральной нити COILS с получением биспиральной нити со степенью вероятности, большей чем 0,9 для всех ее аминокислот, по крайней мере, с размерами одного из окон, которые составляют 14, 21 или 28.
В более предпочтительном мономерном строительном блоке формулы (I), D1 и/или D2 представляет собой пептид, содержащий, по крайней мере, одну последовательность длиной из трех семичленных повторов, которая является предсказанным с помощью программы COILS для прогнозирования биспирализованной нити для создания биспирализованной структуры с вероятностью выше чем 0,9 для всех ее аминокислот, по крайней мере, с размерами одного из окон, которые составляют 14, 21 или 28.
В другом более предпочтительном мономерном строительном блоке формулы (I), D1 и/или D2 представляет собой пептид, содержащий, по крайней мере, две отдельные последовательности длиной из двух семичленных повторов, которая является предсказанным с помощью программы COILS для прогнозирования биспирализованной нити для создания биспирализованной структуры с вероятностью выше чем 0,9 для всех ее аминокислот, по крайней мере, размерами одного из окон, которые составляют 14, 21 или 28.
В другом предпочтительном воплощении один домен олигомеризации D1 или D2 представляет собой домен пентамеризации (m или n=5) СОМР (Malashkevich V.N. и др., Science 1996, 274:761-765) или его производную. Этот домен пентамеризации имеет последовательность LAPQMLRELQETNAALQDVRELLRQQVKQITFLKNTVMECDACG (SEQ ID NO:1). Небольшие модификации этого домена также предусматриваются. Такие модификации могут представлять собой, например, замену аминокислот за пределами пентамера в положениях аа(b), аа(с) или aa(f), предпочтительно в положении aa(f), на Cys с целью образования дисульфидной связи между соседними доменами. Другие модификации поверхностных аминокислот этого домена могут включать замены аминокислот для оптимизации взаимодействий на границе раздела между соседними доменами олигомеризации, такие, как гидрофобные, гидрофильные или ионные взаимодействия или ковалентные связи, подобные дисульфидным мостикам. Также предусматриваются более короткие конструкции этого домена, например, также предусматривается отсутствие C-терминального CDACG фрагмента, в котором цистеины образуют межмолекулярные дисульфидные мостики на C-терминальном конце этого домена пентамеризации. Модификация аминокислот, затрагивающая состояние олигомеризации этого домена, также предусматривается, что приводит, например, к переходу от пентамера к тетрамеру. Другие модификации поверхностных аминокислот этого домена могут включать замены аминокислот (например, на цистеин или лизин) для получения сайтов присоединения для функциональных групп.
В другом предпочтительном воплощении один домен олигомеризации D1 или D2 представляет собой домен пентамеризации (m или n=5) триптофановой застежки (Liu J и др., Proc Natl Acad Sci USA 2004; 101(46):16156-61) или его производную. Этот домен пентамеризации имеет последовательность SSNAKWDQWSSDWQTWNAKWDQWSNDWNAWRSDWQAWKDDWARWNQRWDNWAT (SEQ ID NO:2). Небольшие модификации этого домена также предусматриваются. Такие модификации могут представлять собой, например, замену аминокислот за пределами пентамера в положениях аа(b), аа(с) или aa(f), предпочтительно в положении aa(f), на Cys с целью образования дисульфидной связи между соседними доменами. Другие модификации поверхностных аминокислот этого домена могут включать замены аминокислот для оптимизации взаимодействий на границе раздела между соседними доменами олигомеризации, такие, как гидрофобные, гидрофильные или ионные взаимодействия или ковалентные связи, подобные дисульфидным мостикам. Также предусматриваются более короткие конструкции этого домена. Модификация аминокислот, затрагивающая состояние олигомеризации этого домена, также предусматривается, это приводит, например, к переходу от домена пентамеризации к домену тетрамеризации при замене остатка сердцевины Trp на Phe. Другие мутации остатков сердцевины, такие, как описанные в Примере 10, также рассматриваются, однако, по крайней мере, 70% положений сердцевины аа(а) и aa(d) должны представлять собой либо Trp или другую ароматическую аминокислоту. Другие модификации поверхностных аминокислот этого домена могут включать замены аминокислот (например, на цистеин или лизин) для получения сайтов присоединения для функциональных групп.
В другом предпочтительном воплощении один домен олигомеризации D1 или D2 представляет собой домен тетрамеризации (m или n=4) биспирального домена тетрабрахиона (Stetefeld J. и др., Nature Structural Biology, 2000; 7(9):772-776) или его производную. Этот домен тетрамеризации имеет последовательность IINETADDIVYRLTVIIDDRYESLKNLITLRADRLMIINDNVSTILASG (SEQ ID NO:64). Последовательности биспирализованных структур характеризуются семичленным повтором из семи остатков с 3,4-гидрофобным повтором. Следующая периодичность, которая позволяет остаткам приобретать квази-эквивалентные положения после небольшого количества поворотов, составляет три оборота или 11 остатков. Основываясь на присутствии повторов из 11 остатков, C-терминальный конец поверхностного слоя гликопротеинового тетрабрахиона из гипертермофильной архебактерии Staphylothermus marinus образует биспирализованную структуру правого вращения. Это приводит к образованию стержня на основе тетрамерной спирализованной α-спирали длиной 70 нм, который прикрепляется к клеточной мембране на своем C-терминальном конце. Эта тетрамерная биспирализованная структура содержит серию HTL эпитопов (Пример 9) и, таким образом, является идеально приемлемой в качестве сердцевинного олигомера самоорганизующейся пептидной наночастицы (SAPN).
Еще в одном предпочтительном воплощении один домен олигомеризации D1 или D2 представляет собой домен тримеризации (фолдон) белка бактериофага Т4 фибритина (Tao, Y. и др., Structure 1997, 5:789-798) или его производную. Этот домен тримеризации (m или n=3) имеет последовательность GYIPEAPRDGQAYVRKDGEWVLLSTFL (SEQ ID NO:3). Небольшие модификации этого домена также предусматриваются. Такие модификации могут представлять собой замену Asp 9 на Cys с целью образования дисульфидной связи между соседними доменами. Другие модификации поверхностных аминокислот этого домена могут включать замены остатков для оптимизации взаимодействий на границе раздела между соседними доменами олигомеризации, такие, как гидрофобные, гидрофильные или ионные взаимодействия или ковалентные связи, подобные дисульфидным мостикам. Другие модификации поверхностных аминокислот этого домена могут включать замены аминокислот (например, на цистеин или лизин) для получения сайтов присоединения для функциональных групп.
Наиболее предпочтительными являются биспирализованные последовательности и мономерные строительные блоки, описанные в примерах.
Самоорганизующиеся пептидные наночастицы: выровненные единицы
Самоорганизующиеся пептидные наночастицы (SAPN) образуются из мономерных строительных блоков формулы (I). Если такие строительные блоки собираются вместе, то они образуют так называемые "выровненные единицы". Количество мономерных строительных блоков, которые будут объединяться вместе в такую выровненную единицу, будет определяться наименьшим общим множителем (LCM). Таким образом, если, например, два домена олигомеризации мономерного строительного блока образуют тример (D1)3 (m=3) и пентамер (D2)5 (n=5), то 15 мономеров будет образовывать выровненную единицу (Фигура 2А). Если линкерный сегмент L имеет приемлемую длину, то такая выровненная единица будет собираться в виде сферической пептидной наночастицы. Подобно этому, если два домена олигомеризации D1 и D2 мономерного строительного блока образуют тример (D1)3 (m=3) и тетрамер (D2)4 (n=4), то количество мономеров, которые являются необходимыми для образования выровненной единицы, будет составлять 12 (Фигура 2В).
Поскольку m и n не могут быть равны или кратны друг другу, то наименьший общий множитель (LCM) всегда будет большим, чем m и n.
Самоорганизующиеся пептидные наночастицы (SAPN) могут образовываться путем объединения только одной или более одной выровненной единицы (Таблица 2). Такие SAPN представляют собой топологически закрытые структуры.
Правильные многогранники
Существует пять правильных многогранников: тетраэдр, куб, октаэдр, додекаэдр и икосаэдр. Они обладают различными элементами внутренней вращательной симметрии. Тетраэдр содержит одну ось второго порядка и две оси третьего порядка, куб и октаэдр имеют оси вращательной симметрии второго, третьего и четвертого порядка, а додекаэдр и икосаэдр имеют оси вращательной симметрии второго, третьего и пятого порядка. В кубе пространственная ориентация этих осей является точно такой же, что и в октаэдре, а в додекаэдре и икосаэдре пространственная ориентация этих осей относительно друг друга является в точности одинаковой. Таким образом, для целей создания SAPN в соответствии с изобретением куб и октаэдр, а также подобному этому додекаэдр и икосаэдр, могут считаться идентичными. Куб/октаэдр строятся из 24 идентичных пространственных строительных блоков, в то время, как додекаэдр/икосаэдр строятся из 60 идентичных пространственных строительных блоков (Таблица 2). Эти строительные блоки представляют собой асимметричные единицы (AU) многогранника. Они являются трипирамидами и каждая из граней пирамиды соответствует одной из осей вращательной симметрии, таким образом, эти AU будут нести на своих гранях элементы симметрии второго, третьего, четвертого или пятого порядка, в зависимости от типа многогранника. Если эти элементы симметрии образованы из пептидных доменов олигомеризации, то такие AU конструируются из мономерных строительных блоков, как описано выше. Является достаточным выровнять два домена олигомеризации D1 и D2 вдоль двух осей симметрии AU (Фигура 3). Если эти два домена олигомеризации образуют стабильные олигомеры, то граница раздела симметрии вдоль третьей оси симметрии будет образовываться автоматически, и она может быть стабилизирована путем оптимизации взаимодействий вдоль этой границы раздела, например, гидрофобных, гидрофильных или ионных взаимодействий или ковалентных связей, таких как дисульфидные мостики.
Сборка до самоорганизующихся пептидных наночастиц (SAPN) с правильной симметрией многогранника
Для получения самоорганизующихся пептидных наночастиц (SAPN) с правильной геометрией (додекаэдр, куб), требуется более одной выровненной единицы. Например, для получнения додекаэдра из мономера, содержащего тримерные и пентамерные домены олигомеризации, необходимо 4 выровненные единицы, каждая из которых состоит из 15 мономерных строительных блоков, то есть пептидная наночастица с правильной геометрией будет состоять из 60 мономерных строительных блоков. Необходимые комбинации состояний олигомеризации двух доменов олигомеризации и количество выровненных единиц для образования любого правильного многогранника являются приведенными в Таблице 2.
Будут ли выровненные единицы также подвергаться сборке с образованием правильного многогранника, состоящего более чем из одной выровненной единицы, зависит от геометрического расположения двух доменов олигомеризации D1 и D2 по отношению друг к другу, в частности, от угла между осями вращательной симметрии двух доменов олигомеризации. Этот процесс регулируется с помощью i) взаимодействий на границе раздела между соседними доменами в наночастице, ii) длиной линкерного сегмента L, iii) формой индивидуальных доменов олигомеризации. Этот угол является большим в выровненных единицах по сравнению с размещением в правильном многограннике. Кроме того, этот угол не является идентичным в мономерных строительных блоках, в отличие от правильного многогранника. Если этот угол ограничен малыми значениями правильного многогранника (с помощью гидрофобных, гидрофильных или ионных взаимодействий или ковалентного дисульфидного мостика), и линкерный сегмент L является достаточно коротким, то данное число топологически закрытых выровненных единиц, каждая из которых содержит определенное количество мономерных строительных блоков, будет также дополнительно отжигаться с образованием правильного многогранника (Таблица 2), или включать больше мономерных строительных блоков с образованием наночастицы, в которой отсутствует строгая внутренняя симметрия многогранника.
Если угол между двумя доменами олигомеризации является достаточно малым (даже меньше, чем в правильном многограннике с икосаэдрической симметрией), то большое количество (несколько сотен) пептидных цепей может объединяться в пептидную наночастицу. Этого можно достичь путем замены двух остатков цистеина, которые размещаются на границе между двумя спиралями, как приведено в исходной модели Raman S. и др., Nanomedicine: Nanotechnology, Biology, and Medicine 2006, 2:95-102, и эти остатки будут формировать дисульфидный мостик между двумя спиралями, небольшими остатками аланина, как указано в последовательности SEQ ID NO:33. Угол между двумя спиралями можно делать меньше и, следовательно, более 60 пептидных цепей могут объединяться в виде SAPN. В такой модели SAPN имеет молекулярный вес, который составляет приблизительно 4 Мдальтон, что соответствует приблизительно 330 пептидным цепям (Пример 6).
Эпитопы Т-клеток и эпитопы В-клеток
Поскольку эпитопы Т-клеток - в отличие от эпитопов В-клеток - не нуждаются в представлении на поверхности носителя для того, чтобы вызвать иммунизацию, они могут быть встроены в сердцевину каркасной структуры SAPN, то есть, в биспиразилованную последовательность домена олигомеризации. В настоящем изобретении продемонстрировано, каким образом характеристики МНС связывания эпитопов Т-клеток, которые требуют протяженной конформации для МНС связывания (Фигура 1), могут быть соединены с характеристиками образования биспирализованной структуры, что требует наличия α-спиральной конформации для образования биспирализованной структуры, так чтобы эти эпитопы оба могли быть частью биспирализованной каркасной структуры SAPN, как и быть способными связываться с соответствующими молекулами МНС. Следует отметить, что не все биспирализованные последовательности будут способными связываться с МНС молекулами и не все эпитопы Т-клеток могут быть встроены в биспирализованную структуру. Данное изобретение обеспечивает общие правила, как выбрать приемлемые эпитопы Т-клеток и описывает пути, с помощью которых можно встроить их в конкретный биспирализованный домен олигомеризации, так что эти пептиды будут формировать SAPN. При использовании этих правил широкое разнообразие эпитопов Т-клеток может быть встроено в биспирализованную каркасную структуру SAPN.
В дополнительном аспекте данного изобретения эпитопы В-клеток, которые не являются биспирализованными, встраивают в биспирализованную последовательность SAPN домена олигомеризации путем встраивания их между двумя растянутыми участками биспирализованных сегментов, так, что цельная последовательность действует как один домен олигомеризации. Это представляет особый интерес, поскольку биспирализованная каркасная структура может обеспечить средство для ограничения конформации эпитопа В-клеток конформацией, которая является почти идентичной его нативной конформации.
Источники эпитопов Т-клеток
Для встраивания эпитопов Т-клеток в домен олигомеризации, что в заключение приводит к образованию самоорганизующихся пептидных наночастиц (SAPN), эпитопы Т-клеток могут быть выбраны из различных источников: Например, эпитопы Т-клеток могут быть определены с помощью экспериментальных способов, они являются известными из литературы, они могут быть предсказаны с помощью алгоритмов прогнозирования, которые основываются на существующих белковых последовательностях определенного патогена, или они могут быть de novo сконструированными пептидами или их комбинацией.
Существует большое количество известных эпитопов Т-клеток, доступных из научной литературы. Эти эпитопы Т-клеток могут быть выбраны из определенного патогена (например, как в Примерах 12, 13 и 14), из специфических для рака пептидных последовательностей (например, как в Примере 4), или они могут представлять собой de novo сконструированные пептиды с определенной характеристикой, например, PADRE пептид (патент США 5,736,142), который связывается со многими различными молекулами МНС II, что делает его так называемым промискуитетным (смешанным) эпитопом Т-клеток (например, как в Примере 1). Существуют общедоступными базы данных, которые содержат тысячи различных эпитопов Т-клеток, например, база данных МНС "MHCBN VERSION 4,0" (http://www. imtech.res.in/raghava/mhcbn/index.html) или база данных PDB "Protein Data Bank" ("База данных белковых структур) (http://www.rcsb.org/pdb), или другие.
Является хорошо известным и подробно описанным тот факт, что встраивание HTL эпитопов в другую неиммуногенную пептидную последовательность или присоединение ее к непептидному антигену может сделать их более иммуногенными. PanDR связывающий пептид HTL эпитопа PADRE широко используется в конструировании вакцин для малярии, болезни Альцгеймера и многих других вакцинах.
В соответствии с определением базы данных MHCBN (смотри выше) эпитопы Т-клеток представляют собой пептиды, которые обладают связывающими аффинностями (IC50 значения), меньшими, чем 50000 нМ, с молекулой МНС. Такие пептиды считаются связывающими МНС агентами. В соответствии с этим определением, по данным на август 2006, в Версии 4,0 базы данных MHCBN, следующие данные являются доступными: 20717 связывающих МНС агентов и 4022 не связывающих МНС агентов.
Приемлемые эпитопы Т-клеток могут также быть полученными при использовании алгоритмов прогнозирования. Эти алгоритмы прогнозирования могут либо сканировать существующую белковую последовательность из патогена для эпитопов Т-клеток, или они могут прогнозировать, будет ли de novo сконструированные пептиды связываться с молекулой МНС. Много таких алгоритмов прогнозирования является общедоступными по Интернету. Примеры представляют собой SVRMHCdb (http://svrmhc.umn.edu/SVRMHCdb; J. Wan и др., ВМС Bioinformatics 2006, 7:463), SYFPEITHI (http://www.syfpeithi.de), MHCPred (http://www.jenner.ac.uk/MHCPred), Motif Scanner (http://hcv.lanl.gov/content/immuno/motif_scan/motif_scan) или NetMHCIIpan (http://www.cbs.dtu.dk/services/NetMHCIIpan) для молекулы, связывающей МНС II, и NetMHCpan (http://www.cbs.dtu.dk/services/NetMHCpan) для эпитопов, связывающих МНС I.
HTL эпитопы, которые описаны в данной заявке и являются предпочтительными для конструирования, представляют собой пептидные последовательности, которые либо оцениваются с помощью биофизических способов, либо прогнозируются с помощью NetMHCIIpan для связывания с любой МНС II молекулой со связывающими аффинностями (IC50 значения), лучшими чем 500 нМ. Такие считаются слабыми связывающими агентами. Предпочтительно, когда эти эпитопы оцениваются с помощью биофизических способов или прогнозируются с помощью NetMHCIIpan для связывания с МНС II молекулами со значениями IC50, лучшими чем 500 нМ. Такие считаются сильными связывающими агентами.
CTL эпитопы, которые описаны в данной заявке и являются предпочтительными для конструирования, представляют собой пептидные последовательности, которые либо оцениваются с помощью биофизических способов, либо прогнозируются с помощью NetMHCIIpan для связывания с любой МНС I молекулой со связывающими аффинностями (IC50 значения), лучшими чем 500 нМ. Такие пептиды считаются слабыми связывающими агентами. Предпочтительно, когда эти эпитопы оцениваются с помощью биофизических способов или прогнозируются с помощью NetMHCIIpan для связывания с МНС I молекулами со значениями IC50, лучшими чем 50 нМ. Такие пептиды считаются сильными связывающими агентами.
Места для эпитопов Т-клеток
Эпитопы Т-клеток могут быть встроены в нескольких местах в пределах пептидной последовательности биспирализованного домена олигомеризации D1 и или D2. Для достижения этого частная последовательность с эпитопом Т-клеток должна подчиняться правилам для формирования биспирализованной структуры, а также правилам для связывания МНС. Правила для формирования биспирализованной структуры были подробно приведены выше. Правила связывания с молекулой МНС являются введенными в программы прогнозирования МНС связывания, которые используют сложные алгоритмы для прогнозирования пептидов, связывающих МНС.
Существует много различных молекул HLA, каждая из которых имеет ограничения относительно аминокислот в своей последовательности, которая будет обладать наилучшими связывающими характеристикамми. Связывающие мотивы являются подытоженными в Таблице 3. В этой Таблице мотив демонстрирует x для положений, которые могут иметь любую аминокислоту, и в квадратных скобках (список) аминокислоты, которые могут быть только в определенном положении связывающего мотива.
Многие МНС молекулы имеют весьма подобные связывающие мотивы и, таким образом, могут быть сгруппированы в так называемые HLA супертипы. Связывающие мотивы для этих супертипов являются подытоженными в Таблице 4.
Частота встречаемости определенной аминокислоты в определенном положении эпитопа Т-клетки также может быть подытожена. Для связывания МНС положения 1, 4, 6 и 9 в эпитопе Т-клетки являются наиболее важными. Наиболее предпочтительные остатки в этих положениях приведены в Таблице 5, однако предпочтительность для определенной аминокислоты в этих положениях варьирует в значительной степени для различных молекул МНС.
Таким образом, как было упомянуто выше, связывание определенной аминокислотной последовательности с МНС молекулой может быть более точно предсказано с помощью программ, приведенных выше.
Из Таблицы 5 легко можно увидеть, что, например, наиболее часто встречаемыми аминокислотами в положении 1 и положении 4 являются такие, которые обнаружены в сердцевинных положениях биспирализованного семичленного повтора (обозначено с помощью подчеркивания). Положения 1 и 4 могут накладываться на положения семичленного повтора аа(а) и aa(d). Таким образом, эпитоп Т-клеток с аминокислотой L в положении 1 и аминокислотой V в положении 4 идеально находится в соответствии с биспирализованным пептидом, имеющим те же аминокислоты в сердцевинных положениях аа(а) и aa(d) семичленного повтора. Таким образом, если пептидная последовательность удовлетворяет оба требования, как в отношении ограничения мотива связывания Т-клетки, так и ограничения биспирализованного мотива семичленного повтора, она может быть встроена в домен олигомеризации SAPN. Этого можно достичь для большого количества эпитопов Т-клеток путем подгонки выравнивания пептидной последовательности, так, что мотив связывания Т-клеток перекрывается с мотивом формирования биспирализованной структуры.
Инжиниринг эпитопов Т-клеток в биспирализованную структуру
Конструирование SAPN, которая включает эпитопы Т-клеток в биспирализованном домене олигомеризации SAPN, должно быть осуществлено в три этапа. На первом этапе кандидатный эпитоп Т-клеток должен быть выбран при использовании известных эпитопов Т-клеток из литературных источников или из баз данных, или из предсказанных эпитопов Т-клеток при использовании приемлемых программ для прогнозирования эпитопов. На втором этапе сайт протеосомального расщепления должен быть встроен на C-терминальном конце CTL эпитопов. Это может быть осуществлено при использовании программы прогнозирования для протеосомальных сайтов расщепления PAProc (http://www.paproc2.de/paproc1/paproc1.html; Hadeler K.P. и др., Math. Biosci. 2004, 188:63-79) и модификации остатков, которые непосредственно следуют за желаемым сайтом расщепления. Этот второй этап не требуется для HTL эпитопов. На третьем и наиболее важном этапе последовательность эпитопа Т-клеток должна быть выровнена с биспирализованной последовательностью, так, что это является наиболее совместимым с правилами для образования биспирализованной структуры, как описано выше. Будет ли последовательность со встроенным эпитопом Т-клетки действительно образовывать биспиральную структуру может быть предсказано, а наилучшее выравнивание между последовательностью эпитопа Т-клетки и последовательностью биспирализованного повтора может быть оптимизировано при использовании программ для прогнозирования биспирализованной структуры, таких как COILS (http://www.ch.embnet.org/software/COILS_form.html; Gruber М. и др., J. Struct. Biol. 2006, 155(2):140-5) или MULTICOIL (http://groups.csail.mit.edu/cb/multicoil/cgi-bin/multicoil.cgi), которые являются доступными через Интернет.
Даже тогда, когда невозможно обнаружить приемлемое выравнивание (может быть по той причине, что эпитоп Т-клетки содержит глицин или даже пролин, который не является совместимым с биспирализованной структурой) - эпитоп Т-клетки может быть встроен в домен олигомеризации (смотри Пример 3). В этом случае эпитоп Т-клетки должен быть фланкирован сильными последовательностями, образующими биспирализованную структуру, того же состояния олигомеризации. Это будет либо стабилизировать биспирализованную структуру до достаточной степени, либо, альтернативно, это может генерировать петлеобразную структуру в пределах данного биспирализованного домена олигомеризации. Это является существенно такой же процедурой, как описано в следующем разделе для встраивания эпитопов В-клеток в последовательность биспирализованной сердцевинной структуры SAPN.
Встраивание эпитопов В-клеток в биспирализованную сердцевинную структуру
В частном аспекте настоящего изобретения встраивание в биспирализованную сердцевинную структуру SAPN небольших эпитопов В-клеток, которые не являются α-спирализованными, также предполагается. Это может быть осуществлено с помощью такой же процедуры, что и та, которая описана выше для эпитопов Т-клеток, которые не являются совместимыми с биспирализованной структурой. Структура фибритина Т4 (pdb номер доступа 1аа0 на http://www.rcsb.org/pdb/) содержит две петлевые структуры в пределах своей биспирали. Петли выпячиваются из биспирали между двумя витками спирали, так, что спиральная структура биспирали не прерывается.
В фибритине петли выходят из спирали в положении аа(b) биспирали и снова входят в спираль в положении аа(с) биспиральной последовательности. Одна из петель представляет собой короткий бета-виток, в то время как другая обладает более неправильной структурой петли. Пространство между остатками aa(b) и аа(с) в биспирали является идеально приемлемым для того, чтобы служить в качестве якорных точек для антипараллельного бета-витка пептида. Когда остатки аа(b) или аа(с) или оба из них представляют собой остатки глицина, то это позволяет придавать необходимую гибкость вторичной структуры белка для выхода и возврата альфа-спирали биспирализованной структуры.

В последовательности фибритина, приведенной выше, петлевые структуры представлены курсивом, а остатки в положениях аа(b) и аа(с) (где две петли выходят и вновь возвращаются в спираль) обозначаются путем подчеркивания. Три из этих четырех остатков являются остатками глицина. Используя это в качестве матрицы, эпитоп В-клеток, который обладает антипараллельной конформацией альфа-спирали, может встраиваться в биспиральную сердцевину SAPN. Биспирализованные структуры должны быть в достаточной мере стабильными для того, чтобы позволить осуществить встраивание таких петельных структур, таким образом, такая структура должна быть способной образовывать биспирали с обеих сторон петли. Самая маленькая автономно собирающаяся описанная последовательность биспирали имеет длину, равную двум семичленным повторам. В последовательности, представленной ниже, конец V3 эпитопа из белка gp120 ВИЧ, который представляет собой антипараллельный пептид бета-витка, является встроенным в биспираль сконструированной стабильной биспирализованной структуры с фланкирующими спиралями более чем двух семичленных повторов с обеих сторон. Таковые представляют собой очень стабильные биспирализованные фрагменты, полученные в соответствии с Burkhard P. и др., J Mol Biol 2002, 318:901-910.
Это будет ограничивать конформацию V3 петли в пределах биспирали до антипараллельной конформации бета-витка, что соответствует нативной конформации этого пептида в белке.
Предпочтительная модель
Для конструирования SAPN с самым лучшим иммунологическим профилем для данного частного применения необходимо принять в расчет следующие соображения:
Для CTL эпитопов необходим протеосомальный сайт расщепления на их C-терминальном конце. Эпитопы не должны быть подобными человеческим последовательностям для того, чтобы избежать аутоиммунного ответа - за исключением тех случаев, когда ставится цель вызвать иммунный ответ против пептида человека. Возможные примеры представляют собой специфические для опухоли CTL эпитопы Примера 4.
В соответствии с этим является предпочтительным SAPN, в которой, по крайней мере, один из эпитопов Т-клеток представляет собой CTL эпитоп, и, в частности, в котором последовательность дополнительно содержит протеосомальный сайт расщепления после CTL эпитопа.
Также предпочтительным является SAPN, в которой, по крайней мере, один из эпитопов Т-клеток представляет собой HTL эпитоп, в частности, HTL эпитоп, связывающий pan-DR. Такие HTL эпитопы, связывающие pan-DR, связываются со многими молекулами МНС класса II, как приведено в конце Таблицы 3, и, таким образом, узнаются у большинства здоровых индивидуумов, что является крайне необходимым для получения хорошей вакцины.
Также предпочтительным является SAPN, в которой последовательность D1 - L - D2 содержит серию перекрывающихся эпитопов Т-клеток, если либо D1, либо D2 является тримером (Примеры 7 и 8), тетрамером (Пример 9) или пентамером (Пример 10).
Для эпитопов В-клеток является необходимым, чтобы они были представлены на поверхности SAPN. Они могут быть или могут не быть частью биспирализованной последовательности, то есть, биспираль сама по себе может частично представлять собой эпитоп В-клетки, в зависимости от того является ли часть биспиральной структуры поверхностно доступной. Например, эпитоп В-клетки, состоящий из тримерной биспирали поверхностных белков вирусов, которые содержат оболочку, может быть представлен на поверхности SAPN и быть частью биспирализованной последовательности в одно и то же время. Пример такой модели представлен у Raman S. и др., Nanomedicine: Nanotechnology, Biology, and Medicine 2006; 2:95-102. Биспирали любого состояния олигомеризации в общем случае являются исключительно хорошо приспособленными для того, чтобы презентироваться SAPN специфическим для конформации образом. Биспирали являются избыточными не только в поверхностных белках вирусов, имеющих оболочку, но также, например, в геноме патогена малярии Plasmodium falciparum (Villard V. и др., PLoS ONE 2007; 2(7):e645).
Однако в общем случае эпитопы В-клеток не будут частью биспирализованных доменов олигомеризации или они могут состоять из биспирали и дополнительной части, которая не является биспиралью, как, например, тримерный адгезионный аутотранспортер (ТАА) бактерии, который обладает биспирализованным стержнем и глобулярным доменом головки, как, например, ТАА N. meningitidis.
Особый интерес представляют белки в качестве эпитопов В-клеток, которые сами по себе являются олигомерными, такие, как тримерный гемагглютинин, и тетрамерная нейраминидаза или М2 поверхностные белки вируса гриппа.
Обсуждение модели вакцины против патогена
Такая вакцина предпочтительно содержит все три типа эпитопов, эпитопы В-клеток, HTL и CTL. (1) Является предпочтительным, когда только один (или совсем мало) эпитоп В-клетки будет размещаться на одном из концов пептидных цепей. Таковой будет содержать эпитоп В-клетки на поверхности SAPN в виде повторяемого антигенного дисплея. (2) HTL эпитопы должны обладать промискуитетом в той мере, насколько это возможно. Они не должны обязательно иметь происхождение от патогена, но могут быть пептидами, которые вызывают сильный ответ Т-хелперных клеток. Примером может служить PADRE пептид. Предпочтительно, когда таковые представляют собой эпитопы Т-клеток, которые являются встроенными в D1 - L - D2 сердцевинную последовательность SAPN. (3) CTL эпитопы должны быть специфическими для патогена, они должны содержать C-терминальные протеосомальные сайты расщепления. Поскольку эпитопы Т-клеток не требуют повторяемого антигенного дисплея, то несколько различных эпитопов Т-клеток может быть встроено в один SAPN путем совместной сборки пептидных цепей, которые все имеют одну и ту же наночастицу, формирующую D1 - L - D2 сердцевину, но несут различные эпитопы Т-клеток, которые не являются частью последовательности, формирующей сердцевину, и таким образом, могут не быть встроенными в биспирализованные последовательности.
Подобным образом пептидные цепи, которые несут сигнал нацеливания на ER, то есть, сигнальный пептид, который индуцирует транспорт белка или пептида к эндоплазматическому ретикулуму (ER), могут подвергаться совместной сборке в одну и ту же SAPN для того, чтобы нести CTL эпитопы в ER для собственной презентации молекулами МНС I, поскольку перекрестная презентация не является очень эффективной у людей. Однако не является необходимым, чтобы сигнал нацеливания на ER находился на отдельной пептидной цепи, он может находиться в том же пептиде, что и CTL эпитопы. Приемлемый ER сигнальный пептид может представлять собой, например, ER нацеливающий сигнал (Е3/19K) MRYMILGLLALAAVCSA (SEQ ID NO:6).
Терапевтическая вакцина, направленная на генерацию сильного антителогенеза
Терапевтическая вакцина, направленная на генерацию сильного антителогенеза, является особенно полезной для лечения болезни Альцгеймера, гипертензии, ожирения, наркомании или воспаления. Для такой вакцины предпочтительно используется только один эпитоп В-клетки. Сильный гуморальный ответ благодаря повторяемому антигенному дисплею может быть дополнительно улучшен путем включения одного или более промискуитетных HTL эпитопов в SAPN. Предпочтительно, когда таковые представляют собой эпитопы Т-клеток, которые являются встроенными в D1 - L - D2 сердцевинную последовательность SAPN. Кроме того, должно быть настолько мало CTL эпитопов и они должны быть настолько слабо связанными, насколько это является возможным, - в частности, не против человеческого пептида, для того, чтобы избежать аутоиммунного ответа.
Терапевтическая вакцина для индукции CTL ответа, например, против рака
В этом случае не нужно использовать никакого эпитопа В-клетки. Иммунный ответ против конкретнх CTL эпитопов (например, MAGE-1,2,3; MART-1,2,3; или Her-2/neu, смотри также Пример 4) дополнительно усиливается путем включения одного или более промискуитетных HTL эпитопов в SAPN.
Самоорганизующиеся пептидные наночастицы (SAPN) в качестве адъювантов
SAPN, которая состоит из многочисленных HTL эпитопов, будет индуцировать сильный Т-хелперный иммунный ответ (смотри Пример 2). При использовании в той же дозе с любой другой вакцинной композицией это будет приводить к стимуляции иммунного ответа. Такая SAPN будет представлять собой адъювант без необходимости присутствия какого-либо CTL эпитопа или эпитопа В-клеток. Однако эпитопы В-клеток и CTL эпитопы могут комбинироваться с такой адъювантной SAPN. Кроме того, дополнительные молекулы адъюванта могут быть ковалентно слиты с SAPN в виде заместителя домена олигомеризации D1 или D2 для дополнительной стимуляции адъювантного эффекта SAPN. Особый интерес представляют собой иммуностимуляторные нуклеиновые кислоты, предпочтительно олигодезоксинуклеотид, содержащий дезоксиинозин, олигодезоксинуклеотид, содержащий дезоксиуридин, олигодезоксинуклеотид, содержащий CG мотив, инозин и цитидин, содержащий молекулу нуклеиновой кислоты. Другие иммуностимуляторные молекулы представляют собой, например, антимикробные пептиды, такие, как катионные пептиды, которые представляют собой класс иммуностимуляторов, позитивно заряженные молекулы, которые являются способными усиливать и/или улучшать адаптивные иммунные ответы. Примером такого пептида с иммуностимуляторными свойствами является позитивно заряженный искусственный антимикробный пептид KLKLLLLLKLK (SEQ ID NO:63), который индуцирует сильный специфический для белка типа 2 адаптивный иммунитет после иммунизации на основе первично-вторичной вакцинации.
Предпочтительно, когда антигены в соответствии с изобретением являются выбранными из группы, состоящей из (а) белков, приемлемых для индукции иммунного ответа против раковой клетки; (b) белков или углеводов, приемлемых для индукции иммунного ответа против инфекционных болезней; (с) белков, приемлемых для индукции иммунного ответа против аллергенов; (d) пептидных гормонов, приемлемых для индукции иммунного ответа для лечения заболеваний человека; и (е) молекулы гаптена, приемлемой для индукции иммунного ответа для лечения наркомании и других заболеваний. Пептидные наночастицы, включающие такие белки, их пептидные фрагменты, пептиды, углеводы или гаптены могут быть приемлемыми для индукции иммунного ответа у людей, или также у сельскохозяйственных животных и домашних питомцев.
В одном предпочтительном воплощении изобретения антигены или антигенные детерминанты являются такими, которые полезны для предотвращения инфекционного заболевания. Такое лечение будет полезным для предотвращения большого разнообразия инфекционных заболеваний, которые поражают широкий спектр хозяев, например, людей, животных, отличных от человека, таких, как корова, овца, свинья, собака, кошка, других видов млекопитающих, а также видов, отличных от млекопитающих.
В частности, изобретение относится к SAPN, включающей один из следующих антигенов:
(a) антиген, приемлемый для индукции иммунного ответа против бактерий;
(b) антиген, приемлемый для индукции иммунного ответа против вирусов;
(c) антиген, приемлемый для индукции иммунного ответа против паразитов;
(d) антиген, приемлемый для индукции иммунного ответа против раковой клетки;
(e) антиген, приемлемый для индукции иммунного ответа против аллергенов;
(f) антиген, приемлемый для индукции иммунного ответа против зависимостей;
(g) антиген, приемлемый для индукции иммунного ответа против заболеваний и метаболических расстройств;
(h) антиген, приемлемый для индукции иммунного ответа у сельскохозяйственных животных; и
(i) антиген, приемлемый для индукции иммунного ответа у домашних питомцев.
Поддающиеся лечению инфекционные заболевания являются хорошо известными специалисту в данной области техники. Примеры включают инфекции вирусной, бактериальной или паразитарной этиологии, такие, как следующие заболевания:
амебная дизентерия, сибирская язва, инфекции, вызванные Campylobacter, ветряная оспа, холера, лихорадка Денге, дифтерия, энцефалит, лихорадка Эбола, грипп, японский энцефалит, лейшманиоз, малярия, корь, эпидемический менингит, паротит, внутрибольничные инфекции, коклюш, пневмококковая инфекция, полио (полиомиелит), коревая краснуха, опоясывающий лишай, шистоматоз, столбняк, клещевой вирусный энцефалит, трихомониаз, трипаносомоз, туберкулез, брюшной тиф, ветряная оспа, желтая лихорадка.
В частности, изобретение относится к SAPN, которая включает один из антигенов из приведенных ниже паразитов:
Campylobacter, цитомегаловирус, вирус Эпштейна-Барра, вирус ящура, Haemophilus influenzae типа b, Helicobacter pylori, вирус гепатита, вирус гепатита С, вирус гепатита Е, вирус простого герпеса, вирус иммунодефицита человека, человеческий папилломавирус, Neisseria meningitides, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pneumoniae, респираторно-синцитиальный вирус, ротавирус, круглые черви, анкилостома, вирус болезни западного Нила.
В предпочтительном аспекте изобретения предусматривается композиция для предотвращения и лечения малярии (Пример 11). Жизненный цикл малярийного паразита обеспечивает несколько стадий, на которых вмешательство может приводить к приостановке инфекционного процесса. В жизненном цикле малярийного паразита человек становится инфицированным малярией в результате укуса самки малярийного комара (Anopheles). Комар вводит свой хоботок в хозяина и при этом вводит форму спорозоита Plasmodium falciparum (или vivax), которая присутствует в слюне комара. Возможные белковые и пептидные последовательности, приемлемые для модели пептидной вакцины, могут включать последовательности из следующих белков Plasmodium: MSP-1 (большой полиморфный белок, который экспрессируется на поверхности клетки паразита), MSA1 (основной поверхностный антиген 1 мерозоита 1), CS белок (нативный белок спорозоита), белок весом 35 кДа или белок весом 55 кДа или белок весом195 кДа в соответствии с патентом США 4,735,799, АМА-1 (антиген 1 апикальной мембраны), или LSA (антиген печеночной стадии).
В предпочтительной модели один из эпитопов В-клеток представляет собой последовательность, содержащую от 8 до приблизительно 48 остатков, которые составляют В-клеточный эпитоп белка спорозоита (CS). Этот эпитоп В-клеток представляет собой участок избыточного повтора аминокислотной последовательности NANP для Plamodium falciparum. В предпочтительной модели SAPN этот В-клеточный эпитоп включает от двух до приблизительно пяти повторов последовательности аминокислотных остатков NANP или ее пермутации ANPN, NPNA и PNAN. Соответствующий участок повтора Plasmodium vivax состоит из любой из следующих в высокой степени подобных последовательностей.
В предпочтительной модели один из эпитопов В-клеток представляет собой последовательность, содержащую от 8 до приблизительно 48 остатков в составе любой из этих последовательностей.
Специфические пептидные последовательности для модели SAPN для лечения малярии являются приведенными в трех таблицах, представленных ниже, для эпитопов В-клеток, HTL-эпитопов и CTL-эпитопов.
В Таблице 7 приведены предпочтительные биспирализованные эпитопы В-клеток Р. falciparum (Villard V. и др., PLoS ONE 2007, 2(7):е645 и Agak G.W., Vaccine (2008) 26, 1963-1971. Поскольку для эпитопов В-клеток только поверхностно доступные остатки представляют особую важность для их взаимодействия с рецептором В-клетки и продукции антител, биспирализованные остатки сердцевины в положениях аа(а) и aa(d), которые не являются поверхностными, могут быть модифицированы без изменения способности антигена вызывать образование нейтрализующих антител. Например, замена валина в положении аа(а) изолейцином не будет оказывать воздействия на общие иммунологические свойства биспирализованного эпитопа В-клеток. Таким образом, эти биспирализованные последовательности могут быть искусственно стабилизированы путем оптимизации сердцевинных остатков для наилучшего образования биспирализованной структуры и стабильности (Пример 13) без нарушения их иммунологического потенциала. В соответствии с этим, пептидные эпитопы В-клеток в одном или более из их сердцевинных остатков в положении аа(а) и/или aa(d), наряду со свойствами образования биспирализованной структуры, как описывается более подробно выше, также предусматриваются для этих эпитопов В-клеток.
Таким образом, в предпочтительной модели биспирализованный эпитоп В-клеток с модификациями одного или более своих сердцевинных положений представляет собой пептид, который содержит, по крайней мере, последовательность, которая имеет длину, равную двум семичленным повторам, и которая является спрогнозированной с помощью программы для предсказания биспирализованной структуры COILS, для образования биспирализованной нити с вероятностью более высокой чем 0,9, для всех ее аминокислот, по крайней мере, с одним окном размером 14, 21 или 28.
В другой предпочтительной модели биспирализованный эпитоп В-клеток с модификациями одного или более своих сердцевинных положений представляет собой пептид, который содержит, по крайней мере, одну последовательность длиной в три семичленных повтора, которая является спрогнозированной с помощью программы для предсказания биспирализованной структуры COILS, для образования биспирализованной нити с вероятностью более высокой чем 0,9, для всех ее аминокислот, по крайней мере, с одним окном размером 14, 21 или 28.
В другой предпочтительной модели биспирализованный эпитоп В-клеток с модификациями одного или более своих сердцевинных положений представляет собой пептид, который содержит, по крайней мере, две отдельные последовательности длиной в два семичленных повтора, которая является спрогнозированной с помощью программы для предсказания биспирализованной структуры COILS, для образования биспирализованной нити с вероятностью более высокой чем 0,9, для всех ее аминокислот, по крайней мере, с одним окном размером 14, 21 или 28.
Таблица 8 приводит предпочтительные HTL эпитопы Р. falciparum (Doolan, D.L, The Journal of Immunology, 2000, 165:1123-1137; патент США 5,114,713)
Таблица 9 приводит предпочтительные CTL эпитопы Р. falciparum (патенты США 5,028,425, 5,972,351, 6,663,871)
В другом предпочтительном аспекте изобретения предполагается композиция для предотвращения и лечения ВИЧ (Примеры 5 и 12). Для получения анти-ВИЧ вакцины можно использовать синтетический пептид, способный вызывать выработку специфических для ВИЧ антител, при этом указанный синтетический пептид имеет аминокислотную последовательность функционального эпитопа Т-клетки или эпитопа В-клетки оболочки или gag белка, или gp120, или gp41 ВИЧ-1 для обеспечения иммунного ответа. Особый интерес представляют последовательности в рамках gp41 или gp120, которые могут индуцировать специфические для конформации нейтрализующие антитела, способные препятствовать процессу слияния, подобно известным антителам 2F5 и 4Е10, или из VS-петли gp41 или gp120. Такие последовательности, в основном, локализуются в и поблизости от HR1 и HR2 и участков кластера I и кластера II. Антитела, связывающиеся, например, с биспирализованным тримером gp41, которые индуцируются наночастицами в соответствии с изобретением, включающими этот биспирализованный тример, будут ингибировать образование "шпильки" и, таким образом, слияние вируса. Подобно этому, антитела, образованные против тримерной биспирали вируса Эбола или другого вируса с подобным процессом слияния, будут ингибировать проникновение этих вирусов в клетку.
При использовании высоко консервативных белковых последовательностей ВИЧ, как описывается у Letourneau S. и др., PLoS ONE 2007, 10:е984, CTL эпитопы были спрогнозированы с помощью SVRMHCdb (http://svrmhc.umn.edu/SVRMHCdb; Wan J. и др., ВМС Bioinformatics 2006, 7:463). Эти консервативные белковые последовательности содержат CTL эпитопы, спрогнозированные как такие, которые связываются с HLA молекулами, как приведено в Таблице 10, и представляют собой предпочтительные CTL эпитопы для модели ВИЧ вакцины на основе SAPN. Эти пептидные эпитопы содержат перекрывающиеся в значительной степени последовательности, которые могут быть объединены для создания более длинных пептидных последовательностей, несущих многочисленные CTL эпитопы в одной единственной беспрерывной пептидной цепочке (Таблица 11).
В другом предпочтительном аспекте изобретения предполагается композиция для предотвращения и лечения гриппа. Грипп А кодирует интегральный мембранный белок, М2, гомотетрамер, субъединица которого имеет внешний домен (М2е), состоящий из 23 аминокислотных остатков. Природный М2 белок присутствует в нескольких копиях в вирусной частице и прячется для иммунной системы за массой других поверхностных белков гемагглютинина и нейраминидазы. С другой стороны, он существует в избытке на поверхности мембраны инфицированной вирусом клетки. Последовательность М2е является высоко консервативной. Было продемонстрировано, что М2е презентируется для иммунной системы в виде тетрамера в химерном GNC4-М2е белке и при этом генерирует высоко специфический и протективный гуморальный ииммунный ответ (DeFilette М. и др., J Biol Chem 2008; 283(17):11382-11387).
М2е тетрамер представляет собой высоко консервативный эпитоп В-клетки как для штаммов вируса гриппа человека, так и для птиц (Таблицы 12 и 13). В предпочтительном воплощении настоящего изобретения он может представляться в своей нативной тетрамерной конформации, когда присоединяется к N-терминальному концу тетрамерной биспирализованной структуры из тетрабрахиона (или любой другой тетрамерной биспирализованной структуры) SAPN (Пример 9).
Гемагглютинин вируса гриппа (НА) активируется путем разрезания белкового предшественника на две отдельные пептидные цепи (Steinauer D.S. и др., Virology 1999; 258:1-20). Разрезание предшественника НА молекулы НА0 является необходимым для активации вирусной инфекционности, и распределение активирующих протеаз в хозяйской клетке является одной из детерминант тропизма и, как таковой, патогенности. НА млекопитающих и непатогенных вирусов птиц расщепляются за пределами клетки, что ограничивает их распространение в хозяевах до тканей, в которых имеются приемлемые протеазы. С другой стороны, НА патогенных вирусов подвергается расщеплению внутри клетки с помощью убиквитарно существующих протеаз и, таким образом, обладает способностью инфицировать различные типы клеток и вызывать системные инфекции.
В отличие от М2е последовательности, N-терминальная часть пептида, полученного в результате разрезания, не является высоко консервативной (C-терминальная часть пептида, полученного в результате разрезания, фактически является высоко консервативной). В белке-предшественнике НА пептид, получаемый в результате разрезания, размещается на поверхности, и шесть остатков (три остатка с каждой стороны сайта разрезания), которые размещаются вокруг сайта разрезания, являются наиболее характерными для этой пептидной последовательности (выделены жирным в Таблице 14). В предпочтительной модели SAPN эти шесть остатков представляют эпитоп В-клеток, который может индуцировать антитела, которые при связывании с пептидом могут защищать белок-предшественник НА от такого разрезания.
SAPN являются идеально приемлемыми для представления множества различных последовательностей разрезания, специфических для различных типов НА (Таблица 14) путем совместной сборки пептидов, которые обладают такой же сердцевиной D1 - L - D2, формирующей SAPN, но различными эпитопами В-клеток, присоединенными к ним (Пример 15).
Например, нить гемагглютининового эпитопа В-клеток, включающая последовательности сайтов расщепления Н1, Н2, и Н3 со встроенной аспартатной аминокислотой для того, чтобы сделать последовательность более растворимой и менее основной, будет выглядеть подобно представленной ниже: SIQSRGLFGDIESRGLFGERQTRGIFG (SEQ ID NO:227).
Пептиды с такой же сердцевинной последовательностью, но различными эпитопами В-клеток или последовательностями эпитопов, могут подвергаться совместной сборке с образованием одной SAPN для получения мультивалентного SAPN иммуногена, который возможно включает все или наиболее важные (Н1, Н2, Н3, Н5, Н7 и Н9 для человеческой вакцины) последовательности Таблицы 14 (Пример 15).
В подобном подходе SAPN для вакцины против гриппа, которая состоит из шести пептидных цепей с идентичным ядром и идентичным N-терминальным эпитопом В-клетки М2е и приблизительно 20 CTL эпитопами на C-терминальном конце (три или четыре каждого на пептидную цепь), может подвергаться совместной сборке с образованием одной SAPN (Пример 14). В Таблице 15 приведены предпочтительные консервативные CTL эпитопы, описанные у Parida R. и др., Vaccine 2007, 25:7530-7539. Поскольку сердцевина этих шести пептидных цепей является идентичной, то совместная сборка этих шести пептидных цепей в единственную SAPN позволяет ввести около 20 различных CTL эпитопов в единственную SAPN.
В другом предпочтительном воплощении композиции в соответствии с изобретением представляют собой иммунотерапевтические средства, которые могут использоваться для лечения метаболических расстройств и заболеваний или зависимостей. Наиболее предпочтительными являются иммунотерапевтические средства для лечения болезни Альцгеймера, гипертензии, ожирения, никотиновой и кокаиновой зависимостей.
Аβ-фрагмент (Аβ) представляет собой пептид длиной 42 аминокислоты (Aβ1-42). Поскольку последовательность цельного пептида длиной 42 аминокислоты также содержит CTL эпитопы, которые могут вызывать аутоиммунные реакции, является желательным использовать только более короткие фрагменты этого пептида для конструирования вакцины, такие, как Аβ1-12, или даже такие короткие пептиды, как Аβ1-6 (патент США 7,279,165).
Подобно этому, полноразмерный белок TNFα имеет некоторые ограничения в качестве иммуногена. Локальное перепроизводство провоспалительного TNFα играет важную роль в патогенезе некоторых хронических воспалительных расстройств, включая ревматоидный артрит, псориаз и болезнь Крона. Нейтрализация TNFα моноклональными антителами (mAb, инфликсимаб, адалимумаб) или химерными растворимыми рецепторами (этанерцепт) является эффективной в лечении таких состояний, однако имеет некоторые потенциальные побочные эффекты. Она может индуцировать специфические для аллотипа или Id-специфические антитела, которые могут ограничивать долгосрочную эффективность у многих пациентов. Кроме того, поскольку количество пациентов, подвергающихся лечению, растет, то становится очевидным, что лечение с помощью антагонистов TNFα, в частности, с помощью моноклональных антител, повышает риск оппортунистических инфекций, в частности, таких, вызванных внутриклеточными паразитами, подобными Mycobacterium tuberculosis, Listeria monocytogenes или Histoplasma capsulatum. Иммунизация при использовании более коротких фрагментов TNFα, включающих только остатки 4-23, как было продемонстрировано, дает возможность избежать некоторых из этих проблем (G. Spohn и др., The Journal of Immunology, 2007, 178:7450-7457). Таким образом, иммунизация с помощью TNFα 4-23 представляет собой новую эффективную терапию для ревматоидного артрита и других аутоиммунных заболеваний, что обеспечивает новый уровень безопасности для существующих способов терапии на основе TNFα. Путем селективного нацеливания только растворимой формы TNFα и недостаточного использования трансмембранной формы патогенные эффекты TNFα нейтрализуются с помощью вакцины, в то время как важные функции в отношении хозяйского ответа на внутриклеточные патогены остаются без изменений.
Помимо никотина и кокаина также в качестве молекул гаптена при создании моделей В-клеточной SAPN вакцины для лечения зависимостей могут использоваться следующие соединения: опиаты, марихуана, амфитамины, барбитураты, глутетимид, метиприлон, хлоральгидрат, метаквалон, бензодиазепины, ЛСД, антихолинергические лекарственные средства, антипсихотические лекарственные средства, триптамин, другие психомиметические лекарственные средства, седативные средства, фенциклидин, псилоцибин, летучий нитрит, и другие лекарственные средства, индуцирующие физическую зависимость и/или психологическую зависимость.
В другом предпочтительном воплощении композиции в соответствии с изобретением представляют собой иммунотерапевтические средства, которые могут использоваться для лечения аллергий. Выбор антигенов или антигенных детерминант для композиции и способа лечения аллергий будут хорошо известны специалисту в области медицины для лечения таких расстройств. Характерные примеры антигенов и антигенных детерминант такого типа включают фосфолипазу пчелиного яда А2, Bet v I (аллерген березовой пыльцы), 5 Dol m V (аллерген яда осы пятнистой) и Der p I (клещевой аллерген домашней пыли).
В другом предпочтительном воплощении композиции в соответствии с изобретением представляют собой иммунотерапевтические средства, которые могут использоваться для лечения рака. Основные виды рака для нацеленных моделей вакцин представляют собой следующие: рак мозга, рак молочной железы, рак шейки матки, рак ободочной и прямой кишки, рак пищевода, глиобластому, лейкемию (острая миелогенная и хроническая миелоидная), рак печени, рак легких (немелкоклеточный рак легких, мелкоклеточный рак легких), лимфому (не-Ходжинскую лимфому), меланому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки.
Выбор антигенов или антигенных детерминант для композиции и способа лечения рака будут хорошо известны специалисту в области медицины для лечения таких расстройств. Характерные примеры антигенов и антигенных детерминант такого типа включают следующие: HER2/neu (рак молочной железы), GD2 (нейробластома), EGF-R (злокачественная глиобластома), СЕА (рак медуллярной части щитовидной железы), CD52 (лейкемия), MUC1 (экспрессируется при гематологических злокачественных перерождениях), gp100 белок или продукт гена супрессора опухоли WT1.
В Таблице 17 показаны представляющие интерес специфические для рака эпитопы Т-клеток с релевантным белком происхождения и МНС ограничением.
Наиболее предпочтительными являются биспирализованные последовательности, B-клеточные HTL- и CTL-эпитопы, мономерные строительные блоки, SAPN и модели вакцин, описанные в примерах.
Примеры
Следующие примеры являются полезными для дополнительного объяснения изобретения, но не путем ограничения объема изобретения.
Пример 1: PADRE (P5c-8-Mal)
pan-DR эпитоп PADRE с последовательностью AKFVAAWTLKAAA (SEQ ID NO:296) встраивали в тримерную биспирализованную нить SAPN при использовании следующих критериев модели: остаток аланина (А) обладает сильной способностью к образованию альфа-спиралей. Выравнивание с сердцевинными положениями биспирали является таким, что остатки валина, триптофана и аланина находятся в положении аа(а), aa(d) и снова аа(а) модели семичленного повтора. Оставшуюся часть тримерной биспирали N-терминального и C-терминального концов до HTL эпитопа конструировали так, что последовательность является спрогнозированной как такая, которая образует очень сильную биспираль.

Последовательность этого пептида (SEQ ID NO:7) прогнозировали для образования биспирализованной структуры при использовании программы для прогнозирования COILS. Вероятность формирования биспирали составляет более 95% для всех остатков в последовательности при использовании окна из 21 аминокислоты для предсказания биспирализованной структуры (смотри Таблицу 18 выше). Таким образом, таковая представляет собой предсказанную биспираль, которая также содержит Т-клеточный эпитоп.
Является очень важным, чтобы окно включало только 14 аминокислот, предсказание падает до очень низких значений вероятности (менее чем 2%) в пределах последовательности Т-клеточного эпитопа. Больший размер окна в пределах 21 аминокислоты с высокой предсказанной предрасположенностью последовательности к образованию биспирализованной структуры на протяжении всей последовательности демонстрирует эффект фланкирующих участков на N- и C-терминальных концах T-клеточного эпитопа. Если таковые являются последовательностями, которые сильно благоприятствуют образованию биспирали, то меньшая склонность к благоприятному образованию биспирализованной структуры эпитопа Т-клетки может быть компенсирована, и цельная последовательность будет индуцировать образование биспирализованной структуры даже тогда, когда эпитоп Т-клетки не содержит очень благоприятной биспирализованной последовательности.
Тримерную биспираль с последовательностью SEQ ID NO:7 потом включали в последовательность SAPN SEQ ID NO:8, приведенную ниже, которая состоит из His-метки, пентамерной биспирали СОМР, тримерной биспирали, состоящей из последовательности SEQ ID NO:7, включающей PADRE Т-клеточный эпитоп, и эпитоп В-клетки из CS-белка Plasmodium berghei.

Пептид с этой последовательностью экспрессировали в E.coli и очищали на колонке с никелевой аффинностью с помощью стандартных биотехнологических процедур. Повторную сборку осуществляли в соответствии с Raman S. и др., Nanomedicine: Nanotechnology, Biology, and Medicine 2 (2006) 95-102. Полученную в результате сборки SAPN подвергали анализу на образование наночастицы путем методик динамического рассеяния света (DLS) и просвечивающей электронной микроскопии (ТЕМ). DLS анализ выявил хороший размер распределения со средним диаметром частицы 32,01 нм и показателем полидисперсности 12,9% (Фигура 4А). ТЕМ изображения (Фигура 4В) показывают наночастицы одного и того же размера, как определено с помощью DLS.
Пример 2: Р5с-6-общий
Эта биспирализованная последовательность содержит четыре перекрывающихся HTL эпитопа с последовательностями LEELERSIW, IWMLQQAAA, WMLQQAAAR, и MLQQAAARL, которые предсказываются с помощью алгоритма SVRMHC для связывания с различными молекулами МНС II DQA1*0501, DRB1*0501, DRB5*0101, и DRB1*0401, соответственно, с предсказанными связывающими аффинностями (pIC50 значения) 6,122, 8,067, 6,682, и 6,950, соответственно. Эти HTL эпитопы выравнивают с помощью биспирализованного семичленного повтора так, что они являются спрогнозированными для формирования очень сильной биспирали. Сердцевинные положения аа(а) и aa(d) биспирализованной структуры являются занятыми Leu, Leu, Ile, Leu, Ala и Leu, большинство из них представляют собой очень хорошие остатки для высокой склонности к формированию биспирали, при этом только Ala является в некоторой степени менее благоприятным.

Таким образом, последовательность пептида прогнозируется для образования биспирализованной структуры с помощью программы COILS. Вероятность образования биспирали составляет более чем 99% для всех остатков HTL эпитопов в последовательности (Таблица 19). Сравнение размеров малого и большого окна для прогнозирования биспирали вновь продемонстрировало влияние фланкирующих последовательностей для стабильности биспирали. При размере окна 28 аминокислот цельная последовательность прогнозируется для образования биспирали с вероятностью 100%, в то время, как размер окна, который составляет 14 аминокислот, демонстрирует эффект более низкой склонности к образованию биспирали эпитопов Т-клеток на N-терминальном конце с более низкими значениями предсказания для образования биспирали. Таким образом, для цельной последовательности этот пептид предсказывается как такой, который образует стабильную биспираль, включающую четыре различных HTL эпитопа.
Тримерную биспираль потом включали в SAPN последовательность SEQ ID NO:10, которая приведена ниже и состоит из His-метки, пентамерной биспирали СОМР, тримерной биспирали SEQ ID NO:9 и эпитопа В-клеток из CS-белка Plasmodium berghei.

Пептид с этой последовательностью экспрессировали в E.coli и очищали на колонке с никелевой аффинностью с помощью стандартных биотехнологических процедур. Повторную сборку осуществляли в соответствии с Raman S. и др., Nanomedicine (смотри выше). Полученную в результате сборки SAPN подвергали анализу на образование наночастицы путем методик динамического рассеяния света (DLS) и просвечивающей электронной микроскопии (ТЕМ). DLS анализ выявил хороший размер распределения со средним диаметром частицы 46,96 нм и показателем полидисперсности 8,7%. ТЕМ изображения (Фигура 5) показывают наночастицы с размером приблизительно 30 нм.
Пример 3: Т1с-7 вируса гриппа
Эта биспирализованная последовательность содержит два последовательных HTL эпитопа с последовательностями IRHENRMVL и YKIFKIEKG из белков М1 и нейраминидазы вируса гриппа А, соответственно. С помощью компьютерного алгоритма SVRMHC они были предсказаны как такие, которые сильно связываются с МНС II молекулами DRB1*0405 и DRB1*0401 с предсказанными связывающими аффинностями (pIC50 значения) 8,250 и 6,985, соответственно. Кроме того, в соответствии с Parida R. и др., Vaccine 2007, 25:7530-7539, первый эпитоп Т-клетки IRHENRMVL является предсказанным для связывания со многими другими HLA молекулами, а также такими, как В14, В1510, В2705, В2706, В3909, DP9, DR11, DR12, DR17, DR53 и DRB1, то есть, он представляет собой предсказанный промискуитетный эпитоп.
Наилучшее выравнивание этих HTL эпитопов с биспирализованным семичленным повтором представлено ниже (SEQ ID NO:11). Однако последовательность второго из этих эпитопов содержит неблагоприятный остаток глицина, который в целом действует как остаток, разрушающий спираль. Фланкирующие части биспирализованного тримера представляют собой относительно короткие последовательности, но такие, которые обладают сильной способностью к образованию биспирализованной структуры. Когда используется маленькое окно размером 14 аминокислот в программе предсказания биспиральной структуры COILS, то видно, что биспирализованные структуры предсказываются для обеих сторон эпитопов Т-клеток (смотри Таблицу 20, приведенную ниже). Таким образом, цельная последовательность будет снова действовать в качестве единственной собирающейся единицы и образовывать стабильную биспираль с тримерным состояние олигомеризации.

Тримерную биспираль потом включали в SAPN последовательность SEQ ID NO:12, которая приведена ниже, состоящую из His-метки, пентамерной Trp-застежки, тримерной биспирали, как описано выше, и эпитопа В-клеток из CS-белка Plasmodium berghei.

Пептид с этой последовательностью экспрессировали в Е.coli и очищали на колонке с никелевой аффинностью с помощью стандартных биотехнологических процедур. Повторную сборку осуществляли в соответствии с Raman S. и др., Nanomedicine (смотри выше). Полученную в результате сборки SAPN подвергали анализу на образование наночастицы путем методик динамического рассеяния света (DLS) и просвечивающей электронной микроскопии (ТЕМ). DLS анализ показал в некоторой степени более широкий размер распределения со средним диаметром частицы 57,70 нм и показателем полидисперсности 21,6%. ТЕМ изображения (Фигура 6) показывают наночастицы с размером приблизительно 30-50 нм. Некоторые из них обладают склонностью слипаться вместе, и условия сборки будут требовать небольшого усовершенствования.
Пример 4: Прогнозирование биспиралей для раковых CTL эпитопов
В следующих примерах показано, как специфические для рака эпитопы Т-клеток могут включаться в пептид, образующий SAPN. 19 различных эпитопов Т-клеток собирали в тримерную биспираль SAPN, и соответствующие способности к образованию биспирали подсчитывали для первых 10 из этих последовательностей. Хорошо видно, что способности к образованию биспирали для эпитопов Т-клеток являются ниже, но фланкирующие последовательности с очень высокими способностями к образованию биспирали будут компенсировать и индуцировать образование биспирали на протяжении всей последовательности, подобно тому, как это представлено в Примерах 1-3, приведенных ниже.

Следующие последовательности аналогичным образом демонстрируют высокую способность к образованию биспирали, несмотря на то, что способность к образованию биспирали для эпитопов Т-клеток является более низкой, но фланкирующие последовательности с очень высокими способностями к образованию биспирали будут компенсировать это и индуцировать образование биспирали на протяжении всей последовательности.


Пример 5: ВИЧ-V3
Эта биспирализованная последовательность содержит антипараллельный виток пептида, который представляет собой конец V3 петли gp120 из ВИЧ. Этот пептид представляет собой хорошо известный эпитоп В-клетки ВИЧ. Его встраивали в биспирализованный семичленный повтор с помощью двух глициновых остатков в положениях аа(b) и аа(с) биспирали.
Фланкирующие части биспирализованного тримера представляют собой относительно короткие последовательности, но такие, которые обладают сильной способностью к образованию биспирали. Когда используется маленькое окно размером 14 аминокислот в программе предсказания биспиральной структуры COILS, то можно увидеть, что биспирализованные структуры предсказываются для обеих сторон эпитопов В-клеток (смотри Таблицу 22, приведенную ниже). Таким образом, цельная последовательность будет снова действовать в качестве единственной собирающейся единицы и образовывать стабильную биспираль с тримерным состоянием олигомеризации с выступающим бета-витком пептида.
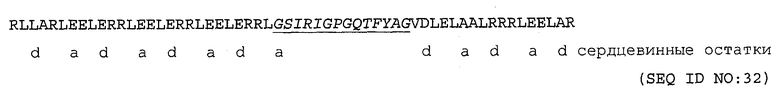
Пример 6: Аналитическое ультрацентрифугирование Р5с
Пептид следующей последовательности (SEQ ID NO:33) рекомбинантно экспрессировали в стандартной экспрессионной системе Е.coli при использовании аффинной схемы очистки с His-меткой:

Эта последовательность является родственной с последовательностями из Примеров 1 и 2 (SEQ ID NO:8 и SEQ ID NO:10), но без C-терминального конца B-клеточного эпитопа. Образовавшаяся SAPN не имеет дисульфидного мостика между двумя спиралями (подчеркнутые остатки 55 и 64, замена двух цистеинов аланином по сравнению с исходной моделью Raman S. и др., Nanomedicine: Nanotechnology, Biology, and Medicine 2006; 2:95-102), но предпочтительно содержит вместо этого меньшую аминокислоту аланин, что позволяет установить меньшие углы между двумя спиралями и, таким образом, в SAPN встраиваются более чем 60 пептидных цепей.
Проверяли три условия для сборки наночастиц из мономерного строительного блока SEQ ID NO:33. Молекулярный вес (MB) SAPN оценивали с помощью аналитического ультрацентрифугирования: пептид растворяли при концентрации 0,42 мг/мл, 0,34 мг/мл и 0,21 мг/мл в 150 мМ NaCl, 20 мМ Трис, рН 7,5. Измеренные значения MB соответствуют SAPN, которая состоит из 330 мономеров, то есть, наночастице с большим количеством мономеров, чем требуется для правильного многогранника с 60 асимметрическими единицами (Таблица 23). Две спирали двух доменов олигомеризации не фиксируются дисульфидной связью в своей относительной ориентации относительно друг друга, а меньшая аминокислота аланин позволяет сделать две спирали ближе и, таким образом, угол между ними становится меньше.
Пример 7: Тримерная биспираль с серией перекрывающихся подсчитанных HTL эпитопов (pan3m)
Приведенная ниже структура представляет собой пример модели тримерной биспирали, которая включает пептидные эпитопы из полимеразы вируса гепатита В, которая может подвергаться измерению в отношении связывающих аффинностей с различными МНС II молекулами (Mizukoshi Е. и др., J Immunol 2004, 173: 5863-5871). Два из этих пептидов были последовательно сконструированы в виде тримерной биспирали в соответствии с принципами, описанными в этом документе.

Измеренные связывающие аффинности пептидов, содержащихся в последовательности LQSLTNLLSSNLSWLSLDVSAAF из полимеразы вируса гепатита В для различных МНС II молекул, являются следующими (связывающие аффинности в нМ приведены в скобках); DRB1*0101 (2), DRB1*0301 (62), DRB1-0401 (10), DRB1*0405 (17), DRB1*0701 (173), DRB1*0802 (598), DRB1*0901 (791), DRB1*1101 (303), DRB1*1201 (397), DRB1*1302 (143), DRB1*1501 (21), DRB3*0101 (5), DRB4-0101 (7).
Последовательность пептида прогнозировали для образования биспирали с помощью программы для предсказания COILS. Вероятность формирования спирали составляет более 98% для всех остатков в последовательности (Таблица 24), таким образом, цельная последовательность является спрогнозированной для образования полностью собранной биспирали.
Пример 8: Тримерная биспираль с серией перекрывающихся предсказанных HTL эпитопов (pan3p)
Приведенная ниже структура представляет собой пример модели тримерной биспирали, которая включает часть HTL эпитопов из последовательности SEQ ID NO:34, описанной у Mizukoshi Е. и др., J Immunol 2004, 173:5863-5871, в комбинации с последовательностью, которая представляет собой часть последовательности HTL эпитопа PADRE.

Приведенные ниже сведения представляют собой связывающие аффинности, как предсказано с помощью алгоритма NetMHCII, эпитопов, содержащихся в последовательности ARFVAAWTLKVREVERELSWLSLDVSAAF, для различных молекул класса МНС II (связывающие аффинности в нМ в скобках): DRB1*0101 (23), DRB1*0401 (72), DRB1*0405 (37), DRB1*0701 (164), DRB1*0802 (462), DRB1*0901 (440), DRB1*1101 (271), DRB1*1302 (303), DRB1*1501 (16), DRB3*0101 (60), DRB4*0101 (51). Эта последовательность содержит в некоторой степени такие же эпитопы, что и последовательность pan3m (SEQ ID NO:34), которая, как было показано, демонстрирует даже более сильное связывание с молекулами МНС II, чем предсказано с помощью программы NetMHC II (Mizukoshi Е. и др., J Immunol 2004, 173:5863-5871).
Последовательность пептида предсказывали для образования биспирали с помощью программы для прогнозирования COILS. Вероятность формирования биспирали составляет более 80% для всех остатков в последовательности (Таблица 25), за исключением 8 остатков. Таким образом, последовательность является спрогнозированной для образования полностью собранной биспирали с C-терминальным концом, расположенным немного в отдалении, который не препятствует образованию SAPN, поскольку большая часть последовательности на N-терминальном конце образует биспираль.
Пример 9: Тетрамерная биспираль с серией перекрывающихся предсказанных HTL эпитопов (BN5c-M2eN)
Последовательность тетрамерной биспирали из тетрабрахиона (Stetefeld J. и др., Nature Structural Biology 2000, 7(9):772-776) характеризуется преимущественным одиннадцатичленным биспиральным повтором, вместо семичленного повтора. Следующая слабо модифицированная последовательность получена из этой тетрамерной биспирали.

Эта тетрамерная биспираль является особенно приемлемой в качестве биспиральной структуры SAPN, поскольку она содержит серию перекрывающихся предсказанных HTL эпитопов. Последовательности эпитопов YRLTVIIDD, LKNLITLRA, LITLRADRL, IINDNVSTLR, INDNVSTLRA и VSTLRALLM предсказываются с помощью алгоритма NetMHCII для связывания с различными молекулами МНС II DRB1*0101, DRB1*0401, DRB1*0404, DRB1*0405, DRB1*0701, DRB1*1101, DRB1*1302, и DRB1*1501, соответственно, с предсказанными связывающими аффинностями (нМ) 3, 48, 78, 162, 243, 478, 12 и 420, соответственно.
Тетрамерная биспираль является также в высокой степени приемлемой для презентации эпитопов В-клеток, таких, как М2е пептид из вируса гриппа, который был создан в соответствующей модели SAPN для человеческой вакцины, с последовательностью (SEQ ID NO:37):

Пептид (SEQ ID NO:37) содержит, начиная от N-терминального конца: His-метку, М2е эпитоп В-клеток из специфического для человека штамма вируса гриппа, тетрамерную биспираль с HTL эпитопами, линкер и тримерную биспираль. Эту последовательность экспрессировали в Е.coli и очищали на колонке с никелевой аффинностью с помощью стандартных биотехнологических процедур. Повторную сборку осуществляли в соответствии с Raman S. и др., Nanomedicine (смотри выше). Полученную в результате сборки SAPN подвергали анализу на образование наночастицы путем методики просвечивающей электронной микроскопии (ТЕМ). ТЕМ изображения (Фигура 7А) показывают наночастицы с размером приблизительно 30 нм.
Кроме того, может быть продемонстрирован специфический для кур пептид М2е из вируса гриппа в своем нативном состоянии олигомеризации и конформации в виде тетрамера на тетрамерной биспирали тетрабрахиона, которую создавали в следующей модели SAPN для животной вакцины с последовательностью (SEQ ID NO:38):

Пептид с этой последовательностью (SEQ ID NO:38) экспрессировали в Е.coli и очищали на колонке с никелевой аффинностью с помощью стандартных биотехнологических процедур. Он содержит, начиная от N-терминального конца: His-метку, М2е эпитоп В-клеток из специфического для кур штамма вируса гриппа, тетрамерную биспираль с HTL эпитопами, линкер и тримерную биспираль. Повторную сборку осуществляли в соответствии с Raman S. и др., Nanomedicine: Nanotechnology, Biology, and Medicine 2006; 2:95-102 с буфером 20 мМ Трис-HCl рН 7,5, 150 мМ NaCl и 5% глицерином. Полученные в результате сборки SAPN подвергали анализу на образование наночастицы путем методик динамического рассеяния света (DLS) и просвечивающей электронной микроскопии (ТЕМ). DLS анализ показал распределение размера со средним значением диаметра частицы 45 нм и показатель полидисперсности 8,9%. ТЕМ изображения (Фигура 7В) продемонстрировали наночастицы того же размера, составляющего 30 нм.
Пример 10: Пентамерная биспираль с серией перекрывающихся предсказанных HTL эпитопов
Эту последовательность прогнозировали для образования α-спирали (Н) с высокой вероятностью (в основном, наиболее высокий показатель 9) в соответствии со вторичной структурой с помощью программы для предсказания PSIPRED (http://bioinf.cs.ucl.ac.uk/psipred/ - The PSIPRED Protein Structure Prediction Server). Поскольку сердцевинные положения аа(а) и aa(d) семичленного повтора биспирали представляют собой, главным образом, остатки триптофана, эта последовательность была спрогнозирована для образования пентамерной биспирали (Liu J и др., Proc Natl Acad Sci USA 2004; 101(46):16156-61).

Последовательность этой пентамерной биспирали содержит частично перекрывающиеся HTL эпитопы с последовательностями FVAAWTLKV, WKIWKSLWK и KSLWKAWRL, которые являются предсказанными с помощью алгоритма NetMHC II для связывания с различными молекулами МНС II DRB1*0101, DRB-T0401, DRB1*0405, DRB1*0701, DRB1*0801, DRB1*0901, DRB1*1101, DRB1*1501 и DRB5*0101, соответственно, с предсказанными связывающими аффинностями (нМ) 24, 73, 13, 120, 42, 596, 396, 6 и 13, соответственно.
Пример 11: Тримерная биспираль с HTL эпитопом Plasmodium falciparum (t811c-9-pf)
Приведенная ниже структура представляет собой пример модели тримерной биспирали, которая включает pan DR связывающий эпитоп из Plasmodium falciparum. Последовательность соответствует 17 C-терминальным аминокислотам с двумя цистеинами, замененными аланинами, которая также является известной как CS.T3 пептид (SEQ ID NO:40) из белка спрозоита CS.

В анализе клеточной пролиферации было показано, что этот CS.T3 пептид обладает pan DR активностью и является стимуляторным для DR1, DR2, DR4, DR5, DRw6, DR7 и DR9 молекул (патент США 5,114,713).
Последовательность этого пептида является предсказанной для образования биспирали при использовании программы для прогнозирования COILS. Вероятность образования биспиральной структуры составляет более 99% для всех остатков этой последовательности (Таблица 26), за исключением последних двух аминокислот. Таким образом, цельная последовательность прогнозируется как такая, которая образует полностью собранную биспираль.
Эту биспираль (SEQ ID NO:40) использовали для модели SAPN со следующей последовательностью (SEQ ID NO:41)

Эта последовательность (SEQ ID NO 41) содержит his-метку, пентамерную триптофановую застежку, линкер, тримерную биспираль SEQ ID NO:40 и B-клеточный эпитоп Plasmodium falciparum, который представляет собой тчетырехчленный повтор (NANP) повторяемой последовательности того же белка спорозоита CS.
Пептид с этой последовательностью экспрессировали в Е.coli и очищали на колонке с никелевой аффинностью с помощью стандартных биотехнологических процедур. Повторную сборку осуществляли в соответствии с Raman S. и др., Nanomedicine (смотри выше). Полученную в результате сборки SAPN подвергали анализу на образование наночастиц путем методик динамического рассеяния света (DLS) и просвечивающей электронной микроскопии (ТЕМ). DLS анализ при рН 6,5 показал распределение размера со средним значением диаметра частицы 44,6 нм и показатель полидисперсности 19,6%. ТЕМ изображения (Фигура 8) показывают наночастицы с размером приблизительно 30 нм.
Пример 12: Вакцина проти ВИЧ: продукт совместной сборки HTL, CTL, В-клеточных эпитопов
Приведенная ниже структура представляет собой модель ВИЧ вакцины. Эти консервативные белковые последовательности содержат CTL эпитопы, спрогнозированные как такие, которые связываются с HLA молекулами, как приведено в Таблице 10.


Первые семь цепей CTL пептида Таблицы 10 конструировали на C-терминальном конце шести пептидных цепей с идентичным ядром и идентичным N-терминальным эпитопом В-клетки и, таким образом, осуществляли совместную сборку с образованием SAPN.
Сердцевина содержит тот же тример, что и в Примере 11, с промискуитетным HTL эпитопом Р. falciparum, связанным с Trp застежкой пентамерной биспирали, которая содержит PADRE HTL эпитоп.
Эпитоп В-клеток представляет собой проксимальный участок мембраны GP41, который обладает нейтрализующим потенциалом, как является очевидным из связывания с моноклональными нейтрализующими антителами 2F5 и 4Е10. Этот эпитоп представляет собой α-спираль в решении и удерживается в α-спиральной конформации за счет того, что является частично сконструированным в виде пентамерной биспирали. В этой модели поверхность доступных остатков (то есть, остатков, которые не являются биспирализованными сердцевинными остатками) является такой, которая связывается с антителом 4Е10.
Пример 13: Вакцина против малярии: продукт совместной сборки ядра на основе HTL-CTL-B-клеточных эпитопов, эпитопов В-клеток и CTL
Приведенная ниже структура представляет собой пример малярийной SAPN вакцины, которая состоит из шести пептидных цепей с идентичной сердцевиной и идентичного C-терминального эпитопа В-клеток, а также 18 CTL эпитопов на N-терминальном конце (по три на каждую пептидную цепь), подвергшихся совместной сборке в одну SAPN.

Сердцевина (подчеркнуто) представляет собой комбинацию тримерной биспирали, которая содержит CTL эпитоп MEKLKELEK и модифицированный эпитоп В-клеток KLRNLEEELHSLRKNLNILNEELEELT (последовательность 27 у Villard V. и др., PLoS ONE 2007, 2(7):е645), а также пентамер, представленный в Примере 10, с отличными свойствами связывания panDR. На C-терминальном конце все шесть пептидных цепей имели идентичный B-клеточный эпитоп, который представляет собой четырехчленный повтор (NANP) повторяемой последовательности белка спорозоита CS из Plasmodium falciparum. На N- терминальном конце содержится 18 различных CTL эпитопов Р. falciparum (патенты США 5,028,425, 5,972,351, 6,663,871) по три различных эпитопа на цепь. CTL-эпитопы разделены с помощью оптмизированных протеосомальных сайтов расщепления (http://www.paproc2.de/paproc1/paproc1.html; Hadeler K.P. и др., Math. Biosci. 2004, 188:63-79). Поскольку сердцевина этих шести пептидных цепей является идентичной, то совместная сборка этих шести пептидных цепей в одну SAPN позволяет встраивать приблизительно 18 различных CTL эпитопов в одну SAPN.
Пример 14: Вакцина против гриппа: продукт совместной сборки HTL-сердцевины, B-клеточного тетрамера, CTL
Приведенная ниже структура представляет собой пример SAPN для вакцины против гриппа, которая состоит из шести пептидных цепей с идентичной сердцевиной и идентичным N-терминальным эпитопом В-клеток М2е и приблизительно из 20 CTL эпитопов на C-терминальном конце (по три или четыре каждого на пептидную цепь), подвергнутых совместной сборке в одну SAPN. Сердцевина (подчеркнуто) представляет собой комбинацию тримера из Примера 7 и тетрамера из Примера 9 с отличными свойствами связывания panDR. Тетрамерный N-терминальный эпитоп В-клеток является таким же, как и в Примере 9. На C-терминальном конце размещаются консервативные CTL эпитопы, как описано у Parida R. и др., Vaccine 2007, 25:7530-7539 (Таблица 15). Поскольку сердцевина этих шести пептидных цепей является идентичной, то совместная сборка этих шести пептидных цепей в одну SAPN позволяет встраивать приблизительно 20 различных CTL эпитопов в одну SAPN.


Пример 15: Вакцина против гриппа: продукт совместной сборки HTL-сердцевины, B-клеточного тетрамера, пептида расщепления
Приведенная ниже структура представляет собой пример SAPN для вакцины против гриппа, которая состоит из трех пептидных цепей с идентичной сердцевиной и идентичным N-терминальным эпитопом В-клеток М2е и девяти эпитопов В-клеток на C-терминальном конце (по три на каждую пептидную цепь), подвергнутых совместной сборке в одну SAPN.

Сердцевина (подчеркнуто) представляет собой комбинацию тримера из Примера 8 и тетрамера из Примера 9 с отличными свойствами связывания panDR. Тетрамерный N-терминальный эпитоп В-клеток является таким же, как и в Примере 9. C-терминальные эпитопы В-клеток представляют собой такие, как в Таблице 8 для Н1, Н2, Н3, Н5 консенсусных 1, Н5 консенсусного 2, Н7 консенсусного 1, Н7 консенсусного 2, Н7 консенсусного 3, Н9 с негативными зарядами между эпитопами для того, чтобы сделать нить B-клеточного эпитопа менее позитивно заряженной.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТИ ПЕПТИДОВ И ПЕПТИДНЫЕ КОМПОЗИЦИИ | 2008 |
|
RU2466737C2 |
| НОВЫЕ ИММУНОГЕННЫЕ ПЕПТИДЫ | 2015 |
|
RU2709711C2 |
| МОДИФИЦИРОВАННЫЕ ЭПИТОПЫ ДЛЯ УСИЛЕНИЯ ОТВЕТОВ CD4+ Т-КЛЕТОК | 2013 |
|
RU2724994C2 |
| ХЕЛПЕРНЫЙ ПЕПТИД РАКОВОГО АНТИГЕНА | 2010 |
|
RU2588442C2 |
| RA АНТИГЕННЫЕ ПЕПТИДЫ | 2004 |
|
RU2359974C2 |
| КОНЪЮГИРОВАННАЯ ВАКЦИНА НА ОСНОВЕ ПЕПТИДА АНТИГЕНА WT1 | 2014 |
|
RU2668560C2 |
| НОВЫЕ ПЕПТИДЫ ДЛЯ ПРИМЕНЕНИЯ В ИММУНОТЕРАПИИ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 1999 |
|
RU2233290C2 |
| ПЕПТИДНЫЙ АНТИГЕН, ПОЛУЧЕНЫЙ ИЗ БЕЛКА Х ВИРУСА ГЕПАТИТА B (ВАРИАНТЫ), PH-ЧУВСТВИТЕЛЬНАЯ ЛИПОСОМА ДЛЯ ИНДУКЦИИ КЛЕТОЧНОГО ИММУНИТЕТА ПРОТИВ ВИРУСА ГЕПАТИТА B | 1998 |
|
RU2189989C2 |
| НОВЫЕ ПРОНИКАЮЩИЕ В КЛЕТКУ ПЕПТИДЫ | 2013 |
|
RU2670488C2 |
| ИСКУССТВЕННЫЕ НЕИЗБИРАТЕЛЬНЫЕ ЭПИТОПЫ КЛЕТОК Т-ХЕЛПЕРОВ КАК ИММУННЫЕ СТИМУЛЯТОРЫ ДЛЯ СИНТЕТИЧЕСКИХ ПЕПТИДНЫХ ИММУНОГЕНОВ | 2019 |
|
RU2839585C2 |























































































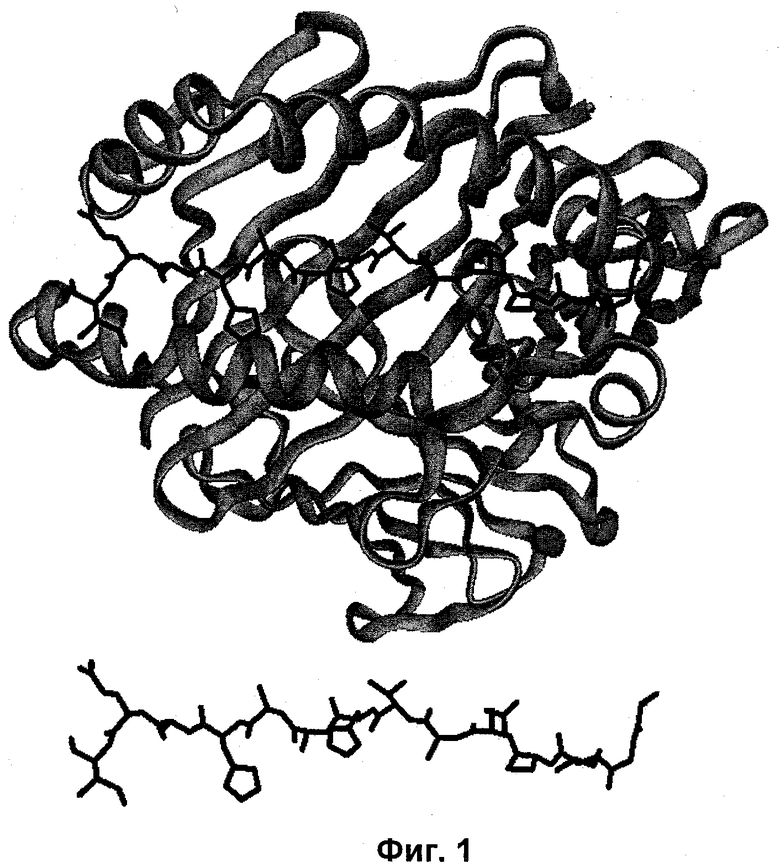
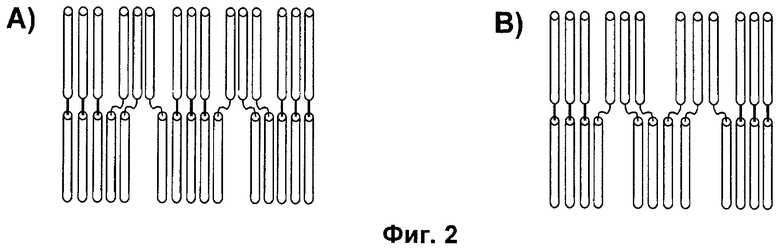



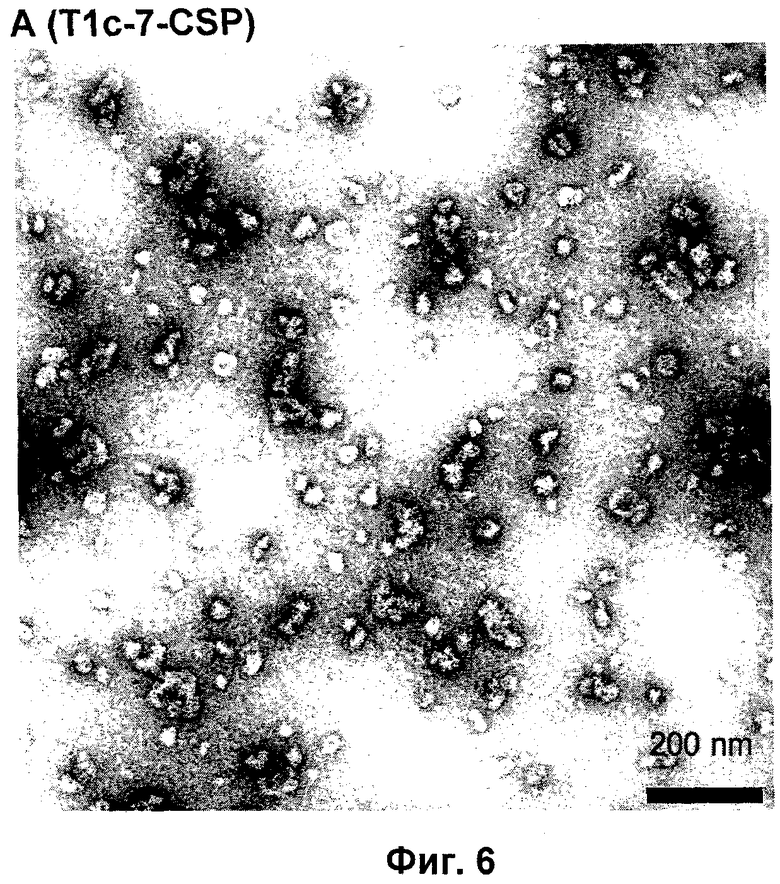
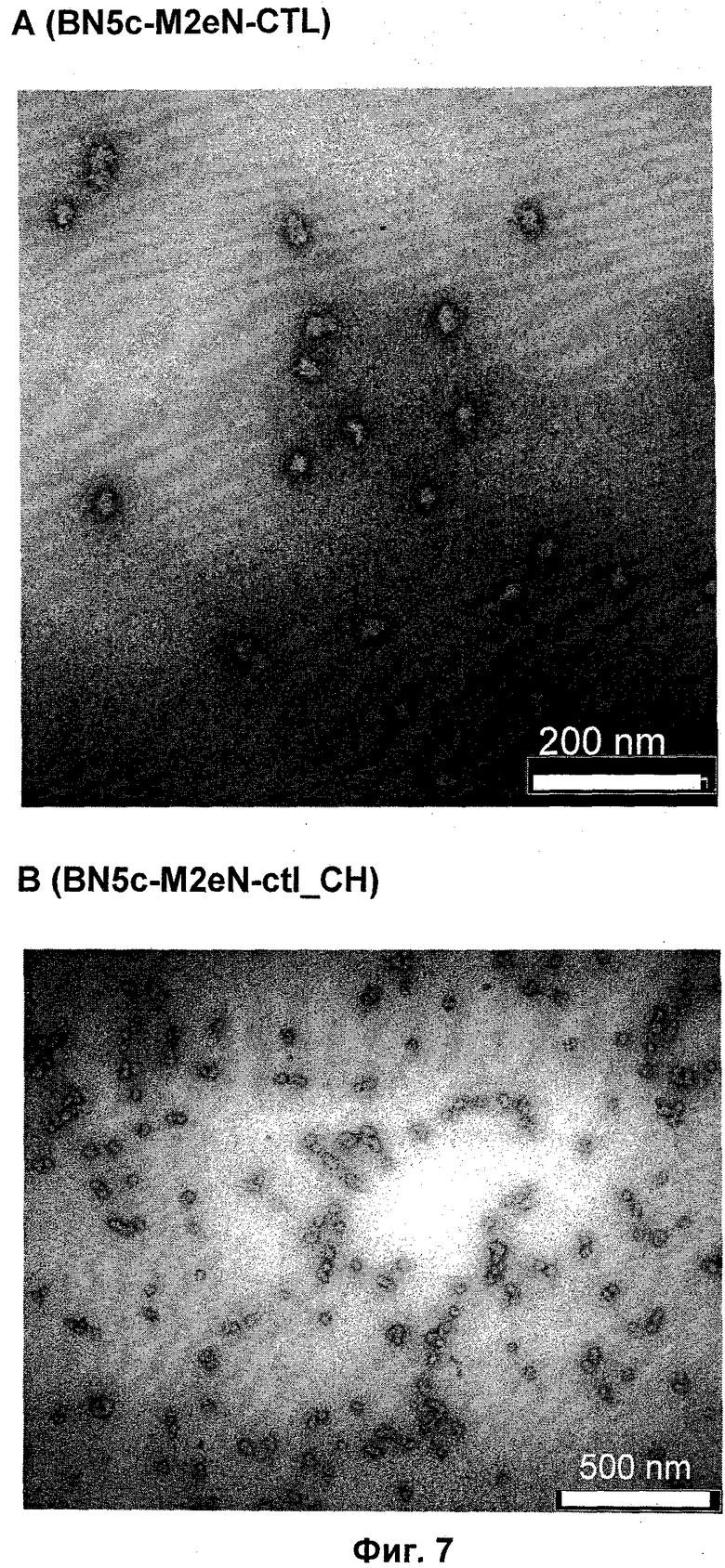
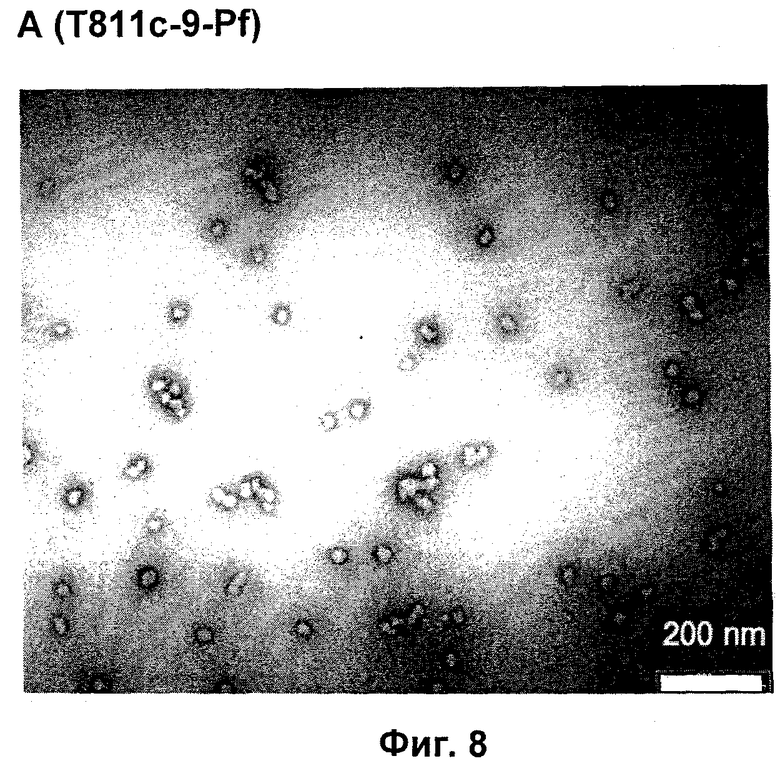
Изобретение относится к биотехнологии. Описываются самоорганизующиеся пептидные наночастицы (SAPN), включающие эпитопы Т-клеток и/или эпитопы В-клеток. Наночастицы в соответствии с изобретением состоят из агрегатов беспрерывной пептидной цепи, включающей два домена олигомеризации, связанные линкерным сегментом, где один или оба домена олигомеризации включают эпитопы Т-клеток и/или эпитопы В-клеток в рамках их пептидной последовательности. Эти наночастицы являются полезными в качестве вакцин и адъювантов. 5 н. и 36 з.п. ф-лы, 8 ил., 26 табл., 15 пр.
1. Самоорганизующаяся пептидная наночастица для индукции иммунного ответа, которая состоит из агрегатов множества строительных блоков формулы (I), состоящих из беспрерывной цепи, включающей пептидный домен олигомеризации D1, линкерный сегмент L и пептидный домен олигомеризации D2
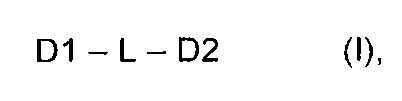
где D1 представляет собой пептид, обладающий тенденцией к образованию олигомеров (D1)m m субъединиц D1, D2 представляет собой пептид, обладающий тенденцией к образованию олигомеров (D2)n n субъединиц D2, m и n каждый представляет собой число от 2 до 10, при условии, что m не является равным n и не является кратным n, а n не является кратным m, L представляет собой связь или короткий линкерный сегмент, выбранные из цепи полиэтиленокси, цепи сахара и пептидной цепи, состоящей из 1-20 аминокислот, либо D1, либо D2, либо оба D1 и D2 являются биспирализованным доменом олигомеризации, который включает по меньшей мере один хелпер Т лимфоцитарного эпитопа (HTL эпитопа) и/или по меньшей мере один цитотоксический Т лимфоцитарный этипоп (HTL эпитоп) в биспирали, и где биспирализованный домен олигомеризации D1 и/или D2 пептида выбран из
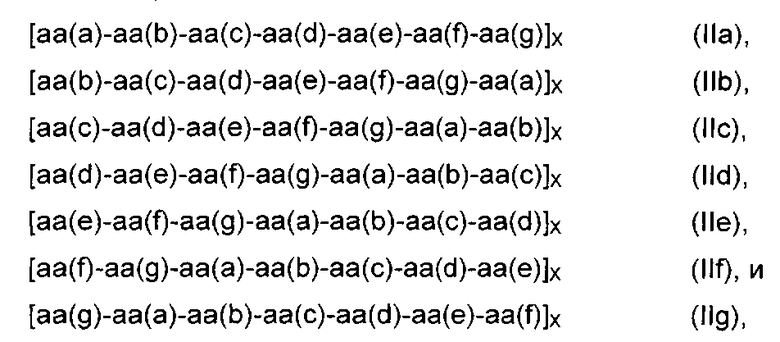
где aa(b), aa(c), aa(e), aa(f) и aa(g) являются одинаковыми или различными аминокислотами или их производными, аа(а) и aa(d) являются одинаковыми или различными гидрофобными аминокислотами или их производными; а X представляет собой число от 2 до 20,
и где D1, D2 и L являются необязательно дополнительно замещенными.
2. Пептидная наночастица по п.1, в которой пептидная цепь D1 на ее N-терминальном конце и/или пептидная цепь D2 на ее C-терминальном конце является замещенной одним или более дополнительными В- и/или Т-клеточными эпитопами, одним или более другими функциональными пептидами или белками, одним или более дополнительными гаптенами или другими функциональными молекулами.
3. Пептидная наночастица по п.2 формул S1-D1-L-D2, D1-L-D2-S2 или S1-D1-L-D2-S2, где S1 и S2 представляют собой пептидные заместители.
4. Пептидная наночастица по п.1, состоящая из идентичных строительных блоков D1-L-D2, где, по крайней мере, один из идентичных строительных блоков несет один или более различных заместителей на N-терминальном конце D1 и/или C-терминальном конце D2.
5. Пептидная наночастица по п.1, в которой один из двух доменов олигомеризации D1 и D2 представляет собой домен пентамеризации триптофановой застежки или его производную.
6. Пептидная наночастица по п.1, в которой один из двух доменов олигомеризации D1 и D2 представляет собой домен тетрамеризации тетрабрахиона или его производную.
7. Пептидная наночастица по п.1, в которой биспираль включает по крайней мере один CTL эпитоп.
8. Пептидная наночастица по п.1, в которой биспираль включает по крайней мере один HTL и/или CTL эпитоп.
9. Пептидная наночастица по п.1, в которой последовательность D1-L-D2 включает серию необязательно перекрывающихся HTL и/или CTL эпитопов.
10. Композиция для вакцинации людей или животных, включающая пептидную наночастицу по пп.1-9.
11. Композиция по п.10, где по крайней мере один из HTL и/или CTL эпитопов встроен в биспираль, или дополнительный B- или T-клеточный эпитоп заместитель выбран из группы, которая состоит из:
(a) антигена, приемлемого для индукции иммунного ответа против бактерий;
(b) антигена, приемлемого для индукции иммунного ответа против вирусов;
(c) антигена, приемлемого для индукции иммунного ответа против паразитов;
(d) антигена, приемлемого для индукции иммунного ответа против раковых клеток;
(e) антигена, приемлемого для индукции иммунного ответа против аллергенов;
(f) антигена, приемлемого для индукции иммунного ответа против зависимостей;
(g) антигена, приемлемого для индукции иммунного ответа против заболеваний и метаболических расстройств;
(h) антигена, приемлемого для индукции иммунного ответа у сельскохозяйственных животных; и
(i) антигена, приемлемого для индукции иммунного ответа у домашних питомцев.
12. Композиция по п.10, где по крайней мере один из HTL и/или CTL эпитопов встроен в биспираль, или дополнительный B- или T-клеточный эпитоп заместитель выбран из белка патогенов, вызывающих заболевания, выбранные из группы, которая состоит из амебной дизентерии, сибирской язвы, инфекции, вызванной Campylobacter, ветряной оспы, холеры, лихорадки Денге, дифтерии, энцефалита, лихорадки Эбола, гриппа, японского энцефалита, лейшманиоза, малярии, кори, эпидемического менингита, паротита, внутрибольничных инфекций, коклюша, пневмококковой инфекции, полио (полиомиелита), коревой краснухи, опоясывающего лишая, шистоматоза, столбняка, клещевого вирусного энцефалита, трихомониаза, трипаносомоза, туберкулеза, брюшного тифа, ветряной оспы, желтой лихорадки.
13. Композиция по п.10, где по крайней мере один из HTL и/или CTL эпитопов встроен в биспираль, или дополнительный B- или T-клеточный эпитоп заместитель выбран из белка патогенов, вызывающих заболевания, выбранные из группы, которая состоит из Campylobacter, цитомегаловируса, вируса Эпштейна-Барра, вируса ящура, Haemophilus influenzae типа b, Helicobacter pylori, вируса гепатита, вируса гепатита С, вируса гепатита Е, вируса простого герпеса, вируса иммунодефицита человека, человеческого папилломавируса, Neisseria meningitides, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pneumoniae, респираторно-синцитиального вируса, ротавируса, круглых червей, анкилостомы, вируса болезни западного Нила.
14. Композиция по п.10, где по крайней мере один из HTL и/или CTL эпитопов встроен в биспираль, или дополнительный B- или T-клеточный эпитоп заместитель выбран из белков вируса гриппа гемагглютинина и/или M2.
15. Композиция по п.14, где М2 эпитоп В-клеток представляет собой тетрамерную форму внеклеточной части этого М2е, присоединенного к N-терминальному концу тетрамерно биспирализованного домена олигомеризации D1.
16. Композиция по п.15, где тетрамерный домен олигомеризации D1 представляет собой тетрамерный биспиральный домен тетрабрахиона или его производную.
17. Композиция по п.10, где по крайней мере один из эпитопов T-клеток является выбранным из группы пептидов вируса гриппа
18. Композиция по п.10, где по крайней мере один из эпитопов B-клеток является выбранным из группы пептидов вируса гриппа, состоящей из
и их фрагментов, включающих, по крайней мере, последовательность шести аминокислот, начиная с третьей аминокислоты последовательностей SEQ ID NO 211-226.
19. Композиция по п.10, где по крайней мере один из эпитопов В-клеток представляет собой последовательность между 8 и 48 остатками, которая составляет B-клеточный эпитоп белка спорозоита Plasmodium falciparum (CS), при этом указанный B-клеточный эпитоп состоит из 2 - приблизительно 12 повторов последовательности аминокислотных остатков Asn-Ala-Asn-Pro или ее пермутаций.
20. Композиция по п.10, где по крайней мере один из эпитопов В-клеток представляет собой последовательность между 8 и 48 остатками, которая составляет B-клеточный эпитоп белка спорозоита Plasmodium falciparum (CS), при этом указанный B-клеточный эпитоп состоит из - приблизительно шести повторов любой из последовательностей аминокислотных остатков, состоящей из
21. Композиция по п.10, где по крайней мере один из эпитопов В-клеток является выбранным из группы пептидов малярии, состоящей из
или их модификаций в одном или более сердцевинных положений aa(a) и/или aa(d).
22. Композиция по п.10, где по крайней мере один из HTL эпитопов является выбранным из группы пептидов малярии, состоящей из
23. Композиция по п.10, где по крайней мере один из CTL эпитопов является выбранным из группы пептидов малярии, состоящей из
24. Композиция по п.10, где по крайней мере один из эпитопов B-клеток представляет собой белок ВИЧ, выбранный из белков gp41, gp120 или gp160.
25. Композиция по п.10, где по крайней мере один из эпитопов B-клеток представляет собой пептид ВИЧ, выбранный из сайтов связывания нейтрализующих антител 2F5 и/или 4E10 gp41.
26. Композиция по п.10, где по крайней мере один из эпитопов В-клеток представляет собой пептид ВИЧ, выбранный из V3-петли gp120 ВИЧ.
27. Композиция по п.10, где по крайней мере один из эпитопов T-клеток является выбранным из группы пептидов ВИЧ, которая состоит из
28. Композиция по п.10, где по крайней мере один из эпитопов T-клеток является выбранным из группы пептидов ВИЧ, которая состоит из
29. Композиция по п.10, в которой эпитоп B-клеток представляет собой антиген, приемлемый для индукции иммунного ответа против зависимостей, выбранных из группы, состоящей из опиатов, марихуаны, амфитаминов, барбитуратов, глутетимида, метиприлона, хлоральгидрата, метаквалона, бензодиазепинов, ЛСД, антихолинергических лекарственных средств, антипсихотических лекарственных средств, триптамина, других психомиметических лекарственных средств, седативных средств, фенциклидина, псилоцибина, летучего нитрита и других лекарственных средств, индуцирующих физическую зависимость и/или психологическую зависимость.
30. Композиция по п.10, в которой по крайней мере один из эпитопов B-клеток представляет собой никотин.
31. Композиция по п.10, в которой по крайней мере один из эпитопов B-клеток представляет собой кокаин.
32. Композиция по п.10, в которой по крайней мере один из B- или T-клеточных эпитопов является выбранным из антигена, приемлемого для индукции иммунного ответа против раковых клеток вида рака, выбранного из рака мозга, рака молочной железы, рака шейки матки, рака ободочной и прямой кишки, рака пищевода, глиобластомы, лейкемии (острой миелогенной и хронической миелоидной), рака печени, рака легких (немелкоклеточного рака легких, мелкоклеточного рака легких), лимфомы (не-Ходжинской лимфомы), меланомы, рака яичника, рака поджелудочной железы, рака предстательной железы, рака почки.
33. Композиция по п.10, которая включает последовательность, выбранную из группы, которая состоит из
34. Композиция по п.10, в которой по крайней мере один из В- или T-клеточных эпитопов является выбранным из группы антигенов, состоящей из
35. Композиция по п.10, в которой по крайней мере один из эпитопов В-клеток представляет собой Aβ-пептид или его фрагмент, включающий, по крайней мере, последовательность из 6 аминокислот, начиная от N-терминальных аминокислот.
36. Композиция по п.10, в которой по крайней мере один из эпитопов B-клеток представляет собой ангиотензин I или ангиотензин II.
37. Композиция по п.10, в которой по крайней мере один из эпитопов B-клеток представляет собой грелин.
38. Композиция по п.10, в которой по крайней мере один из эпитопов B-клеток представляет собой TNFα или его фрагмент, включающий, по крайней мере, последовательность из 20 аминокислот, начиная от четвертой N-терминальной аминокислоты TNFα.
39. Способ вакцинации человека или животного, отличного от человека, который включает введение эффективного количества пептидной наночастицы по пп.1-9 субъекту, который нуждается в такой вакцинации.
40. Способ вакцинации человека или животного, отличного от человека, который включает введение эффективного количества композиции по п.10 субъекту, который нуждается в такой вакцинации
41. Мономерный строительный блок формулы (I), предназначенный для получения самоорганизующейся пептидной наночастицы для индукции иммунного ответа, состоящий из беспрерывной цепи, включающей пептидный домен олигомеризации D1, линкерный сегмент L и пептидный домен олигомеризации D2
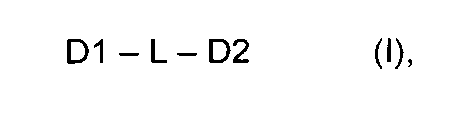
где D1 представляет собой пептид, обладающий тенденцией к образованию олигомеров (D1)m m субъединиц D1, D2 представляет собой пептид, обладающий тенденцией к образованию олигомеров (D2)n n субъединиц D2, m и n каждый представляет собой число от 2 до 10, при условии, что m не является равным n и не является кратным n, а n не является кратным m, L представляет собой связь или короткий линкерный сегмент, выбранные из цепи полиэтиленокси, цепи сахара и пептидной цепи, состоящей из 1-20 аминокислот, либо D1, либо D2, либо оба D1 и D2 являются биспирализованным доменом олигомеризации, который включает по меньшей мере один хелпер T лимфоцитарного эпитопа (HTL эпитопа) и/или по меньшей мере один цитотоксический T лимфоцитарный эпитоп (HTL эпитоп), где биспирализованный домен олигомеризации D1 и/или D2 пептида выбран из

где aa(b), aa(c), aa(e), aa(f) и aa(g) являются одинаковыми или различными аминокислотами или их производными, аа(а) и aa(d) являются одинаковыми или различными гидрофобными аминокислотами или их производными; а X представляет собой число от 2 до 20, и где D1, D2 и L являются необязательно дополнительно замещенными.
| WO 2004071493 A1, 26.08.2004 | |||
| WO 9426903 A2, 24.11.1994 | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| GUILDFORD, GB, vol | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| КОМПОЗИЦИЯ (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ НЕ ПРИРОДНОЙ УПОРЯДОЧЕННОЙ И СОДЕРЖАЩЕЙ ПОВТОРЫ АНТИГЕННОЙ МАТРИЦЫ, СПОСОБ ТЕРАПЕВТИЧЕСКОГО ЛЕЧЕНИЯ И СПОСОБ ИММУНИЗАЦИИ | 1999 |
|
RU2245163C2 |

