Данная заявка представляет собой международную заявку РСТ, в которой испрашивается приоритет по предварительной заявке США с серийным номером 62/782 253, поданной 19 декабря 2018 г., которая в полном объеме включена в данный документ посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
Иммунные ответы требуют кооперативного взаимодействия между антигенпрезентирующими клетками и клетками Т-хелперами (Th). Выявление эффективного ответа антител требует, чтобы антигенпрезентирующие клетки распознавали целевой антигенный сайт иммуногена субъекта и чтобы клетки Т-хелперы распознавали эпитоп клеток Т-хелперов. Обычно Т-хелперный эпитоп на иммуногене субъекта отличается от его В-клеточного эпитопа (ов) или эпитопа (ов) родственных эффекторных Т-клеток (например, цитотоксических Т-лимфоцитов или CTL). В-клеточные эпитопы или эпитопы родственных эффекторных Т-клеток представляют собой сайты на желаемом целевом иммуногене, который распознается В-клетками и родственными клетками, что приводит к продукции антител или цитокинов против желаемого целевого сайта. Естественная конформация мишени определяет сайт, с которым непосредственно связывается антитело или родственная эффекторная Т-клетка. Для индукции Th-клеточного ответа требуется, чтобы рецептор Th-клетки распознал комплекс на мембране антигенпрезентирующей клетки, который образуется между процессированным пептидным фрагментом целевого белка и ассоциированным с ним главным комплексом гистосовместимости (MHC) класса II. Таким образом, для Th-клеточного ответа необходимы пептидный процессинг целевого белка и трехстороннее распознавание. Трехкомпонентный комплекс трудно определить, потому что 1) критические контактные остатки MHC класса II по-разному расположены на разных пептидах, связывающих MHC (Th-эпитопы); 2) разные пептиды, связывающие MHC, имеют разную длину и разные аминокислотные последовательности; и 3) молекулы MHC класса II могут сильно различаться в зависимости от генотипа хозяина. Иммунологическая реактивность на конкретный Th-эпитоп частично определяется генами MHC хозяина, а реактивность Th-эпитопов различается среди отдельных индивидуумов в популяции. Th-эпитопы, которые являются реактивными у разных видов и индивидуумов (то есть неизбирательные Th-эпитопы) в пределах одного вида, трудно идентифицировать.
Для каждого составляющего этапа стадии распознавания Т-клеток требуется множество факторов, таких как соответствующий пептидный процессинг клеткой, процессирующей антиген, презентация пептида генетически детерминированной молекулой MHC класса II и распознавание молекулы MHC или пептидного комплекса рецептором на Th-клетках. Требования к распознаванию неизбирательного Th-эпитопа для обеспечения полиспецифической реактивности могут быть трудными для определения.
Следует понимать, что для индукции антител и родственных цитокинов против иммунных ответов иммуноген должен включать как В-клеточный эпитоп/эпитоп эффекторной Т-клетки, так и детерминанту (детерминанты) Th-клеток. Обычно белок-носитель (например, гемоцианин лимфы улитки; KLH) связывают с иммуногеном-мишенью для обеспечения Th ответа с целью повышения иммуногенности иммуногена-мишени. Однако применение больших белков-носителей для повышения иммуногенности иммуногена-мишени имеет много недостатков. В частности, трудно производить четко определенные, безопасные и эффективные конъюгаты пептид-белок-носитель, потому что (а) химическое связывание включает реакции, которые могут привести к гетерогенности по размеру и составу, например, конъюгация с глутаральдегидом (Borras-Cuesta et al., Eur J Immunol, 1987; 17: 1213-1215); (b) белок-носитель создает потенциал для нежелательных иммунных реакций, таких как аллергические и аутоиммунные реакции (Bixler et al., WO 89/06974); (c) большой белок-носитель пептида вызывает нерелевантные иммунные ответы, преимущественно неправильно направленные на белок-носитель, а не на сайт-мишень (Cease et al., Proc Natl Acad Sci USA, 1987; 84: 4249-4253); и (d) белок-носитель также создает возможность супрессии эпитопов у хозяина, который ранее был иммунизирован иммуногеном, содержащим тот же белок-носитель. Когда хозяина впоследствии иммунизируют другим иммуногеном, в котором тот же белок-носитель связан с другим гаптеном, результирующий иммунный ответ усиливается для белка-носителя, но подавляется для гаптена (Schutze et al., J Immunol, 1985; 135: 2319-2322).
Чтобы избежать описанных выше рисков, желательно вызвать помощь Т-клеток без применения традиционных белков-носителей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В данном изобретении предлагаются неизбирательные эпитопы искусственных клеток Т-хелперов (Th) для стимуляции функциональных сайт-направленных антител против антигена-мишени для профилактического и терапевтического применения. Данное изобретение также относится к пептидным иммуногенным конструкциям, которые содержат Th-эпитопы, композициям, содержащим Th-эпитопы, способам создания и применения Th-эпитопов и антителам, продуцируемым пептидными иммуногенными конструкциями, содержащими Th-эпитопы.
Описанные в данном документе искусственные эпитопы Т-хелперов (Th) могут быть связаны с В-клеточными эпитопом (ами) и/или эпитопом (ами) эффекторных Т-клеток («целевой антигенный сайт») через необязательный спейсер для получения конструкции пептидного иммуногена. Описанные Th-эпитопы придают пептидному иммуногену способность индуцировать сильный иммунный ответ, опосредованный клетками Т-хелперами, с продуцированием высокого уровня антител и/или клеточных ответов против целевого антигенного сайта. Описанные пептидные иммуногенные конструкции обеспечивают выгодную замену больших белков-носителей и сайтов Т-хелперов, происходящих от патогенов, в пептидных иммуногенах с описанными искусственными Th-эпитопами, специально разработанными для улучшения иммуногенности целевого антигенного сайта. Относительно короткие пептидные иммуногенные конструкции, содержащие описанные Th-эпитопы, вызывают высокий уровень антител и/или цитокинов, связанных с эффекторными клетками, к конкретному целевому антигенному сайту, не вызывая значительного воспалительного ответа или иммунного ответа против Th-эпитопа.
Иммунный ответ, вызываемый пептидными иммуногенными конструкциями (включая титры антител, Cmax, начало продукции антител, продолжительность ответа и т. д.), можно модулировать путем варьирования: (a) выбора Th-эпитопа, который химически связан с B-клеточным эпитопом, (b) длины B-клеточного эпитопа, (c) адъюванта, который применяется в составе, содержащем пептидную иммуногенную конструкцию, и/или (d) схемы введения, включая дозу для каждой иммунизации, а также моменты времени введения прайм- и буст-дозы для каждой иммунизации. Следовательно, специфические иммунные ответы на целевые антигенные сайты могут быть разработаны с применением описанных Th-эпитопов, которые могут облегчить адаптацию персонализированного терапевтического лечения к индивидуальным характеристикам любого пациента или субъекта.
Пептидные иммуногенные конструкции, содержащие описанные искусственные эпитопы Th по данному изобретению, могут быть представлены следующими формулами:
(A)n-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
{(A)n-(Th)p-(B)o-(целевой антигенный сайт)-(B)o-(Th)p-(A)n-X}m
где:
каждый A независимо представляет собой аминокислоту;
каждый B независимо представляет собой гетерологичный спейсер;
каждый Th независимо представляет собой искусственный Th-эпитоп;
Целевой антигенный сайт представляет собой В-клеточный эпитоп, эпитоп CTL, пептидный гаптен, непептидный гаптен или их иммунологически реактивный аналог;
X представляет собой аминокислоту, α-COOH или α-CONH2;
n равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10;
m равно 1, 2, 3 или 4.
o равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
p равно 0, 1, 2, 3 или 4;
Примером пептидного гаптена в качестве целевого антигенного сайта являются аминокислоты 1-14 бета-амилоидного (Aβ) белка (Aβ1-14) (SEQ ID NO: 56). Примеры непептидного гаптена включают ассоциированный с опухолью углеводный антиген (TACA - англ.: tumor associated carbohydrate antigen) или низкомолекулярное лекарственное средство.
Данное изобретение также относится к композициям, которые содержат пептидные иммуногенные конструкции, содержащие искусственные Th-эпитопы. Такие композиции способны вызывать у иммунизированного хозяина ответы антител на желаемый целевой антигенный сайт. Целевой антигенный сайт может происходить из патогенных организмов, аутоантигенов, которые в норме являются невосприимчивыми, или мишеней, ассоциированных с опухолью.
Описанные композиции являются пригодными для многих разнообразных медицинских и ветеринарных применений, включая вакцины для обеспечения защитного иммунитета от инфекционного заболевания, иммунотерапии для лечения расстройств, возникающих в результате нарушения нормальных физиологических процессов, иммунотерапии для лечения рака, а также агенты для желательного вмешательства и модификации нормальных физиологических процессов.
Некоторые из целевых антигенов, которые могут быть ковалентно связаны с Th-эпитопами по данному изобретению, включают части: бета-амилоида (Aβ) для лечения болезни Альцгеймера, альфа-синуклеина (α-Syn) для лечения болезни Паркинсона, внеклеточного ближайшего к мембране домена мембраносвязанного IgE (или EMPD IgE) для лечения аллергического заболевания, Tau для лечения таупатий, включая болезнь Альцгеймера, и интерлейкина-31 (IL-31) для лечения атопического дерматита, и это лишь некоторые из них. Более конкретно, указанные целевые антигены включают Aβ1-14 (как описано в патенте США № 9 102 752), α-Syn111-132 (как описано в Международной заявке РСТ № PCT/US2018/037938), IgE EMPD1-39 (как описано в Международной заявке РСТ № PCT/US2017/069174), Tau379-408 (как описано в Международной заявке РСТ № PCT/US2018/057840), и IL-3197-144 (как описано в Международной заявке РСТ № PCT/US2018/065025) и те целевые антигенные сайты, которые описаны Таблице 3A и Таблице 3B.
В данном изобретении также предлагаются способы предотвращения и/или лечения заболевания или патологического состояния у субъекта путем введения пептидной иммуногенной конструкции (включающей описанный искусственный Th-эпитоп и антиген-презентирующий эпитоп) субъекту, нуждающемуся в этом. В некоторых вариантах осуществления пептидные иммуногенные конструкции вызывают иммуногенный воспалительный ответ у субъекта, который по меньшей мере примерно в 3-раза ниже, чем иммуногенный воспалительный ответ положительного контроля, как продемонстрировано в Примере 12.
Данное изобретение также относится к антителам, продуцируемым пептидными иммуногенными конструкциями, содержащими описанные искусственные Th-эпитопы. Антитела, продуцируемые пептидными иммуногенными конструкциями, являются высокоспецифичными к целевому антигенному сайту, а не к искусственным Th-эпитопам.
Ссылки:
Каждый патент, публикация и непатентный документ, цитируемые в данной заявке, включены в данный документ посредством ссылки в полном объеме, как если бы каждый их них был включен посредством ссылки отдельно.
1. BORRAS-CUESTA, F., et al., “Engineering of immunogenic peptides by co‐linear synthesis of determinants recognized by B and T cells”, Eur. J. Immunol., 17:1213-1215 (1987).
2. CEASE, K.B., et al., “Helper T-cell antigenic site identification in the acquired immunodeficiency syndrome virus gp120 envelope protein and induction of immunity in mice to the native protein using a 16-residue synthetic peptide”, Proc. Natl. Acad. Sci. USA, 84:4249-4253 (1987).
3. CHIANG, H-L., et al., “A novel synthetic bipartite carrier protein for developing glycotope-based vaccines”, Vaccine, 30(52):7573-7581 (2012).
4. DANISHEFSKY, S.J., et al., “Development of Globo-H Cancer Vaccine”, Acc. Chem. Res.48(3):643-652 (2015).
5. HUANG, C.Y., et al., “Carbohydrate microarray for profiling the antibodies interacting with Globo H tumor antigen”, Proc. Natl. Acad. Sci. USA 103(1):15-20 (2006).
6. JACQUES, S., et al., “Chemoenzymatic synthesis of GM3 and GM2 gangliosides containing a truncated ceramide functionalized for glycoconjugate synthesis and solid phase applications”, J. Am. Chem. Soc., 134(10):4521-4 (2012).
7. KATIAL, R.K., et al., “Cytokine Production in Cell Culture by Peripheral Blood Mononuclear Cells from Immunocompetent Hosts”, Clin. Diag. Lab. Immunol., 5(1):78-81 (1998).
8. KUDUK, S.D., et al., “Synthetic and Immunological Studies on Clustered Modes of Mucin-Related Tn and TF O-Linked Antigens: The Preparation of a Glycopeptide-Based Vaccine for Clinical Trials against Prostate Cancer”; J. Am. Chem. Soc., 120(48):12474-85 (1998).
9. KUZNIK, G., et al., “Chemical and enzymatic synthesis of high-affinity selectin ligands”, Bioorganic & Medicinal Chemistry Letters, 7(5):577-580 (1997).
10. MEISTER, G.E., et al., “Two novel T cell epitope prediction algorithms based on MHC-binding motifs; comparison of predicted and published epitopes from Mycobacterium tuberculosis and HIV protein sequences”, Vaccine, 13(6):581-591 (1995).
11. PARTIDOS, C.D., et al., “Immune responses in mice following immunization with chimeric synthetic peptides representing B and T cell epitopes of measles virus proteins”, J. of Gen. Virology, 72:1293-1299 (1991).
12. ROTHBARD, J.B., et al., “A sequence pattern common to T cell epitopes”, EMBO J., 7(1):93-101 (1988).
13. SCHUTZE, M.P., et al., “Carrier-induced epitopic suppression, a major issue for future synthetic vaccines”, J. Immunol., 135(4):2319-2322 (1985).
14. TOYOKUNI, T., et al., “Synthetic carbohydrate vaccines: synthesis and immunogenicity of Tn antigen conjugates”, Bioorg. Med. Chem., 11:1119-32 (1994).
15. WO1989/06974 by BIXLER, et al., “T-Cell Epitope As Carriers Molecule For Conjugate Vaccines” (1989-08-10).
16. WO1995/011998 by WANG, et al., “Structured Synthetic Antigen Libraries As Diagnostics, Vaccines And Therapeutics” (1995-05-04).
17. WO1999/066957 by WANG, “Artificial T Helper Cell Epitopes As Immune Stimulators For Synthetic Peptide Immunogens” (1999-12-29) и соответствующий патент США № 6 713 301 (2004-03-30).
18. Патент США № 5 912 176, выданный WANG, “Antibodies against a host cell antigen complex for pre and post exposure protection from infection by HIV” (1999-06-15), и связанные с ним патенты США №№ 5 961 976 (1999-10-05) и 6 090 388 (2000-07-18).
19. Патент США № 6 025 468, выданный WANG, “Artificial T helper cell epitopes as immune stimulators for synthetic peptide immunogens including immunogenic LHRH peptides” (2000-02-15), и связанные с ним патенты США №№ 6 228 987 (2001-05-08) и 6 559 282 (2003-05-06).
20. Патент США № 6 048 538, выданный WANG, et al., “Peptides derived from the non-structural proteins of foot and mouth disease virus as diagnostic reagents” (2000-04-11) и связанный с ним патент США № 6 107 021 (2000-08- 22).
21. Патент США № 6 811 782, выданный WANG, et al., “Peptide composition as immunogen for the treatment of allergy” (2004-11-02), и связанный с ним патент США № 7 648 701 (2010-01-19).
22. Патент США № 6 906 169, выданный WANG, “Immunogenic peptide composition comprising measles virus Fprotein Thelper cell epitope (MUFThl-16) and N-terminus of β-amyloid peptide” (2005-06-14), и связанные с ним патенты США №№ 7 951 909 (2011-05-31) и 8 232 373 (2012-07-31).
23. Патент США № 9 102 752, выданный WANG, “Peptide vaccine for prevention and immunotherapy of dementia of the Alzheimer's type” (2015-08-11).
24. Публикация США № 2013/0236487, выданная WANG, et al., “Designer Peptide-Based PCV2 Vaccine” (2013-09-12).
25. Публикация США № 2014/0335118, выданная WANG, “Synthetic Peptide-Based Marker Vaccine And Diagnostic System For Effective Control Of Porcine Reproductive And Respiratory Syndrome (PRRS)” (2014-11-13).
26. Публикация США № 2015/0306203, выданная WANG, “Synthetic Peptide-Based Emergency Vaccine Against Foot And Mouth Disease (FMD)” (2015-10-29).
27. Публикация США № 2017/0216418, выданная WANG, et al., “Immunogenic LHRH Composition And Use Thereof In Pigs” (2017-08-03).
28. Международная заявка РСТ № PCT/US2017/069174, выданная WANG, “Peptide Immunogens and Formulations Thereof Targeting Membrane-Bound IgE for Treatment of IgE Mediated Allergic Diseases” (подана 31 декабря 2017 г.).
29. Международная заявка РСТ № PCT/US2018/037938, выданная WANG, “Peptide Immunogens From the C-Terminal End of Alpha-Synuclein Protein and Formulations Thereof for Treatment of Synucleinopathies” (подана 15 июня 2018 г.)
30. Международная заявка РСТ № PCT/US2018/057840, выданная WANG, “Tau Peptide Immunogen Constructs” (подана 26 октября 2018 г.).
31. Международная заявка РСТ № PCT/US2018/065025, выданная WANG, “Peptide Immunogens of IL-31 and Formulations Thereof for the Treatment and/or Prevention of Atopic Dermatitis” (подана 11 декабря 2018 г.).
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигуры 1A и 1B: Схемы, демонстрирующие типовый состав и особенности платформы эпитопов Т-хелперных клеток, описанной в данном документе. Фигура 1A представляет собой схематическое обобщение компонентов, которые могут быть включены в составы с пептидными иммуногенами, содержащими носители Th-эпитопов, включая UBITh®. На Фигуре 1B обобщены некоторые особенности и технические преимущества платформы эпитопов Т-хелперных клеток, описанной в данном документе, включая UBITh®.
Фигура 2:График, иллюстрирующий теоретические результаты, которые могут быть получены с применением платформы эпитопов Т-хелперных клеток, описанной в данном документе.
Фигура 3: Повышение иммуногенности пептидных иммуногенных конструкций с помощью выбранных индивидуальных пептидов Th-эпитопа с короткими неиммуногенными пептидами B-эпитопа, полученными из альфа-синуклеина (верхняя панель) и IgE EMPD (нижняя панель). Цифры под гистограммой для альфа-синуклеина (верхняя панель) соответствуют конструкциям пептидного иммуногена, описанным в Таблице 5. ;а числа под гистограммой для IgE EMPD (нижняя панель) соответствуют пептидным иммуногенным конструкциям, описанным в Таблице 6.
Фигуры 4A и 4B: Графики, иллюстрирующие титры антител против α-синуклеина с помощью ИФА, полученные после иммунизации морских свинок смесью трех отдельных пептидных иммуногенных конструкций, содержащих α-Syn (G111-G132), IgE-EMPD (G1-C39) и IL-6 (C73-C83), ковалентно связанных с отдельными Th-эпитопами, продемонстрированными в Таблице 8. Конкретные иммуногенные конструкции α-синуклеинового пептида, оцененные на этих графиках, обобщены в Таблице 5. Фигура 4А содержит данные о титрах антител для всех оцениваемых пептидных иммуногенных конструкций. Фигура 4B содержит подмножество данных, продемонстрированных на Фигуре 4A, чтобы подчеркнуть, что различные пептидные иммуногенные конструкции способны достигать одинаковых значений Cmax с разной скоростью.
Фигура 5: График, иллюстрирующий титры антител анти-IgE EMPD с помощью ИФА, полученные после иммунизации морских свинок смесью трех отдельных пептидных иммуногенных конструкций, содержащих α-Syn (G111-G132), IgE-EMPD (G1-C39) и IL-6 (C73-C83), ковалентно связанных с отдельными Th-эпитопами, продемонстрированными в Таблице 8. Конкретные IgE EMPD-пептидные иммуногенные конструкции, оцененные на этих графиках, обобщены в Таблице 6.
Фигура 6: График, иллюстрирующий титры антител анти-IL-6 с помощью ИФА, полученные после иммунизации морских свинок смесью трех отдельных пептидных иммуногенных конструкций, содержащих α-Syn (G111-G132), IgE-EMPD (G1-C39) и IL-6 (C73 -C83), ковалентно связанных с отдельными Th-эпитопами, продемонстрированными в Таблице 8. Конкретные IL-6-пептидные иммуногенные конструкции, оцененные на этих графиках, обобщены в Таблице 7.
Фигура 7:Графики, иллюстрирующие титры антител анти-DPR (дипептидный повтор) с помощью ИФА, полученные после иммунизации морских свинок пептидными иммуногенными конструкциями, продемонстрированными в Таблице 9.
Фигура 8: График, иллюстрирующийтитры анти-Aβ1-28 с помощью ИФА, полученные после иммунизации морских свинок различными количествами вакцины Aβ (UB-311), которая содержит два пептидных иммуногена Aβ1-14-εK-KKK-MvF5 Th (SEQ ID NO: 67) и Aβ1-14-εK-HBsAg3 Th (SEQ ID NO: 68).
Фигура 9: График, иллюстрирующий титры анти-Aβ1-28 с помощью ИФА, полученные после иммунизации морских свинок различными праймовыми и бустерными дозами вакцины Aβ (UB-311), которая содержит два пептидных иммуногена Aβ1-14-εK-KKK-MvF5 Th (SEQ ID NO: 67) и Aβ1-14-εK-HBsAg3 Th (SEQ ID NO: 68).
Фигура 10: Графики, иллюстрирующие титры антител анти-Aβ1-28 с помощью ИФА, полученные после иммунизации людей вакциной Aβ (UB-311) в течение трехмесячной схемы бустерной вакцинации (верхняя панель) или шестимесячноой схемы бустерной вакцинации (нижняя панель) с применением Aβ вакцины (UB-311). В рамке на каждом графике выделены средние титры для всех субъектов-людей, участвовавших в исследовании.
Фигуры 11A и 11B:Графики, иллюстрирующие титры антител анти-LHRH и концентрацию тестостерона, полученные после иммунизации свиней различными количествами смеси иммуногенных конструкций LHRH пептида, приведенных в Таблице 10 (SEQ ID NO: 239-241) с применением MONTANIDE ISA50V в качестве адъюванта. На Фигуре 11A продемонстрированы титры антител и концентрации тестостерона, полученные после иммунизации крыс составом LHRH в количестве 100 мкг. На Фигуре 11В продемонстрированы титры антител и концентрации тестостерона, полученные после иммунизации крыс составом LHRH в количестве 300 мкг.
Фигура 12: График, иллюстрирующий титры антител анти-IL-6 с помощью ИФА, полученные после иммунизации крыс различными количествами иммуногенной конструкции IL-6 пептида SEQ ID NO: 243 или контрольных плацебо в составах, содержащих различные адъюванты (то есть MONTANIDE ISA51 или ADJUPHOS).
Фигуры 13A и 13B: Графики, иллюстрирующие титры антител анти-IgE-EMPD, полученные после иммунизации макак различными количествами иммуногенной конструкции IgE-EMPD пептида SEQ ID NO: 178. На Фигуре 13A продемонстрированы титры антител, полученные с применением ADJUPHOS в качестве адъюванта, составленного в виде стабилизированного иммуностимулирующего комплекса с применением CpG3. На Фигуре 13В продемонстрированы титры антител, полученные с применением MONTANIDE ISA51 в качестве адъюванта, составленного в виде стабилизированного иммуностимулирующего комплекса с применением CpG3.
Фигуры 14A и 14B: Графики, иллюстрирующие титры антител анти-LHRH и концентрацию тестостерона, полученные после иммунизации свиней различными количествами смеси иммуногенных конструкций LHRH пептида, приведенных в Таблице 10 (SEQ ID NO: 239-241) с применением разных адъювантов. На Фигуре 14A продемонстрированы титры антител, полученные с применением Emulsigen D в качестве адъюванта. На Фигуре 14В продемонстрированы титры антител, полученные с применением MONTANIDE ISA50V в качестве адъюванта.
Фигура 15: Обнаружение неизбирательных и искусственных Т-клеток, отвечающих на Th-пептид, в наивных мононуклеарных клетках периферической крови нормальных доноров.
Фигура 16: Структуры ассоциированных с опухолью углеводных антигенов (TACA): GD3, GD2, Глобо-H, GM2, Fucosyl GM1, PSA, Ley, Lex, SLex, SLea и STn.
Фигуры с 17 по 21:Иллюстративная поэтапная схема синтеза пептида UBITh®, конъюгированного с гликаном, с применением схемы твердофазного пептидного синтеза.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В данном изобретении предлагаются неизбирательные эпитопы искусственных клеток Т-хелперов (Th) для стимуляции функциональных сайт-направленных антител против антигена-мишени для профилактического и терапевтического применения. Данное изобретение также относится к пептидным иммуногенным конструкциям, которые содержат Th-эпитопы, композициям, содержащим Th-эпитопы, способам создания и применения Th-эпитопов и антителам, продуцируемым пептидными иммуногенными конструкциями, содержащими Th-эпитопы.
Описанные в данном документе искусственные эпитопы Т-хелперов (Th) могут быть связаны с В-клеточным эпитопом и/или эпитопом эффекторных Т-клеток (например, цитотоксические Т-клетки; CTL) («целевой антигенный сайт») через необязательный спейсер для получения конструкции пептидного иммуногена. Описанные Th-эпитопы придают пептидному иммуногену способность индуцировать сильный иммунный ответ, опосредованный клетками Т-хелперами, с продуцированием высокого уровня антител и/или клеточных ответов (например, цитокинами) против целевого антигенного сайта для терапевтических эффектов. Описанные пептидные иммуногенные конструкции обеспечивают выгодную замену больших белков-носителей и сайтов Т-хелперов, происходящих от патогенов, в пептидных иммуногенах с описанными искусственными Th-эпитопами, специально разработанными для улучшения иммуногенности целевого антигенного сайта. Относительно короткие пептидные иммуногенные конструкции, содержащие описанные Th-эпитопы, обуславливают высокий уровень антител и/или цитокинов, связанных с эффекторными клетками, к конкретному целевому антигенному сайту, не вызывая значительного воспалительного ответа или иммунного ответа против Th-эпитопа.
Иммунный ответ, вызываемый пептидными иммуногенными конструкциями (включая титры антител, Cmax, начало продукции антител, продолжительность ответа и т. д.), можно модулировать путем варьирования: (a) выбора Th-эпитопа, который химически связан с B-клеточным эпитопом, (b) длины B-клеточного эпитопа, (c) адъюванта, который применяется в составе, содержащем пептидную иммуногенную конструкцию, и/или (d) схемы введения, включая дозу для каждой иммунизации, а также моменты времени введения прайм- и буст-дозы для каждой иммунизации. Следовательно, специфические иммунные ответы на целевые антигенные сайты могут быть разработаны с применением описанных Th-эпитопов, которые могут облегчить адаптацию персонализированного терапевтического лечения к индивидуальным характеристикам любого пациента или субъекта.
Описанные пептидные иммуногенные конструкции, содержащие искусственные Th-эпитопы, способны вызывать у иммунизированного хозяина ответы антител и/или цитокинов против желаемого целевого антигенного сайта. Целевой антигенный сайт может представлять собой конкретный белок, углевод, связанный с раковым антигеном, низкомолекулярное лекарственное соединение или любую аминокислотную последовательность любого целевого пептида или белка. В некоторых вариантах осуществления данного изобретения описаны искусственные Th-эпитопы, которые можно применять для получения пептидных иммуногенов, которые вызывают антитела, нацеленные на амилоид β(Aβ), капсидный белок ящура (FMD), гликопротеин вируса свиного репродуктивного и респираторного синдрома (PRRSV), гормон, стимулирующий высвобождение лютеинизирующего гормона (LHRH), и любые другие пептидные или белковые последовательности.
В определенных вариантах осуществления целевой антигенный сайт взят из аутоантигена или ассоциированного с опухолью неоантигена-мишени, который в норме является «иммуномолчащим» (например, Aβ, тау, альфа-синуклеин, дипептидный белок, IgE EMPD, IL-6, CGRP, амилин, IL-31, неоантигены и др.). Неограничивающие репрезентативные последовательности аутоантигенов и ассоциированных с опухолью неоантигенных сайтов приведены в Таблице 3A. В некоторых вариантах осуществления целевой антигенный сайт взят из патогенного организма (например, FMDV, PRRSV, CSFV, ВИЧ, HSV и т. д.). Неограничивающие репрезентативные последовательности патогенных антигенных сайтов приведены в Таблице 3B.
Пептиды или целевой антигенный сайт по данному изобретению могут быть применимы в медицине и ветеринарии. Например, пептидные иммуногенные конструкции, содержащие описанные искусственные Th-эпитопы, можно применять в композициях вакцин для обеспечения защитного иммунитета от инфекционных заболеваний или нейродегенеративных заболеваний или в фармацевтических композициях для лечения расстройств, возникающих в результате нарушения нормальных физиологических процессов, иммунотерапии для лечения рака, диабета 2 типа, или в качестве агентов, вмешивающихся в нормальные физиологические процессы.
Общие сведения
Заголовки разделов, используемые в данном описании, предназначены только для организационных целей и не должны толковаться как ограничивающие описанный предмет. Все ссылки или части ссылок, процитированные в этой заявке, прямо включены в данный документ посредством ссылки в полном объеме для любых целей.
Если не указано иное, все употребляемые в данном документе технические и научные термины имеют то же значение, которое, как правило, подразумевается обычным специалистом в данной области техники, к которой относится настоящее изобретение. Формы единственного числа включают ссылки на формы множественного числа, если в контексте явно не указано иное. Подобным образом, слово «или» предназначено для охватывания «и», если в контексте явно не указано иное. Следовательно, фраза «содержащий A или B» означает включение A, или B, или A и B. Кроме того, следует понимать, что все размеры аминокислот, а также все значения молекулярной массы или молекулярных масс, приведенные для полипептидов, являются приблизительными и представлены для описания. Хотя способы и материалы, подобные или эквивалентные описанным в данном документе, можно применять на практике или при тестировании описанного способа, пригодные способы и материалы описаны ниже. Все публикации, заявки на патенты, патенты и другие ссылки, упомянутые в данном документе, полностью включены в данный документ посредством ссылки. В случае конфликта приоритет будет иметь данное описание, включая объяснения терминов. Кроме того, материалы, способы и примеры являются иллюстративными и не носят ограничительного характера.
Пептидные иммуногенные конструкции
Термин «пептидный иммуноген» или «пептидная иммуногенная конструкция» в контексте данного описания относится к молекулам, содержащим искусственные Th-эпитопы, ковалентно связанные с целевым антигенным сайтом, с гетерологичным спейсером или без него, посредством ковалентных связей (например, обычная пептидная связь или тиоэфир), так, чтобы образовать один пептид большего размера. Обычно пептидные иммуногенные конструкции содержат (а) гетерологичный неизбирательный искусственный Th-эпитоп; (b) целевой антигенный сайт, такой как В-клеточный эпитоп или эпитоп эффекторных Т-клеток (например, CTL); и (c) необязательный гетерологичный спейсер.
Присутствие неизбирательного искусственного Th-эпитопа в пептидном иммуногене может вызвать сильный опосредованный Th-клетками иммунный ответ и высокий уровень антител, направленных на целевой антигенный сайт у животного после иммунизации пептидным иммуногеном. Описанные пептидные иммуногенные конструкции обеспечивают выгодную замену больших белков-носителей и сайтов Т-хелперов, происходящих от патогенов, в пептидных иммуногенах с описанными искусственными Th-эпитопами, специально разработанными для улучшения иммуногенности целевого антигенного сайта. Относительно короткие пептидные иммуногенные конструкции, содержащие описанные Th-эпитопы, вызывают высокий уровень антител и/или цитокинов, связанных с эффекторными клетками, к конкретному целевому антигенному сайту, не вызывая значительного воспалительного ответа или иммунного ответа против Th-эпитопа.
Пептидные иммуногенные конструкции, содержащие описанные искусственные эпитопы Th по данному изобретению, могут быть представлены следующими формулами:
(A)n-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
{(A)n-(Th)p-(B)o-(целевой антигенный сайт)-(B)o-(Th)p-(A)n-X}m
где:
каждый A независимо представляет собой аминокислоту;
каждый B независимо представляет собой гетерологичный спейсер;
каждый Th независимо представляет собой искусственный Th-эпитоп;
Целевой антигенный сайт представляет собой В-клеточный эпитоп, эпитоп CTL, пептидный гаптен, непептидный гаптен или их иммунологически реактивный аналог;
X представляет собой аминокислоту, α-COOH или α-CONH2;
n равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10;
m равно 1, 2, 3 или 4.
o равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
p равно 0, 1, 2, 3 или 4;
Пептидные иммуногены согласно данному описанию могут содержать от около 20 до около 100 аминокислот. В некоторых вариантах осуществления пептидная иммуногенная конструкция содержит около 20, около 25, около 30, около 35, около 40, около 45, примерно 50, около 55, около 60, около 65, около 70, около 75, около 80, около 85, около 90, около 95 или около100 аминокислотных остатков.
Различные компоненты описанной иммуногенной конструкции пептида IL-31 описаны ниже.
А - Аминокислота
Каждая A в иммуногенных пептидах по данному описанию независимо представляет собой гетерологичную аминокислотную последовательность.
Термин «гетерологичный» в контексте данного описания относится к аминокислотной последовательности, которая не является частью или не гомологична аминокислотной последовательности целевого антигенного сайта дикого типа (например, В-клеточного эпитопа). Таким образом, гетерологичная аминокислотная последовательность A содержит аминокислотную последовательность, которая не встречается в природе в белке или пептиде целевого антигенного сайта. Поскольку последовательность компонента А гетерологична целевому антигенному сайту, природная аминокислотная последовательность целевого антигенного сайта не удлиняется ни в N-концевом, ни в С-концевом направлениях, когда компонент A ковалентно связан с целевым антигенным сайтом.
В некоторых вариантах осуществления каждая A независимо представляет собой не встречающуюся в природе или встречающуюся в природе аминокислоту.
Природные аминокислоты включают аланин, аргинин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин.
Не встречающиеся в природе аминокислоты включают, помимо прочего, ε-N-лизин, ß-аланин, орнитин, норлейцин, норвалин, гидроксипролин, тироксин, γ-амино масляную кислоту, гомосерин, цитруллин, аминобензойную кислоту, 6-аминокапроновую кислоту (Aca; 6-аминогексановую кислоту), гидроксипролин, меркаптопропионовую кислоту (MPA), 3-нитро-тирозин, пироглутаминовую кислоту и тому подобное
В некоторых вариантах осуществления n равно 0, что указывает на то, что в этом положении формулы аминокислота не добавлена. В других вариантах осуществления n равно 1 и выбирается из любой природной или неприродной аминокислоты. В определенных вариантах осуществления n равно больше единицы, и каждый A независимо представляет собой одну и ту же аминокислоту. В некоторых вариантах осуществления n равно больше единицы, и каждый A независимо представляет собой отличающуюся аминокислоту.
B - необязательный гетерологичный спейсер
Каждый B в иммуногенном пептиде по данному описанию является необязательным гетерологичным спейсером. Необязательный гетерологичный спейсер компонента B независимо представляет собой аминокислоту, -NHCH(X)CH2SCH2CO-, -NHCH(X)CH2SCH2CO(εN)Lys-, -NHCH(X)CH2S-сукцинимидил(εN)Lys-, -NHCH(X)CH2S-(сукцинимидил)- и/или любую их комбинацию. Спейсер может содержать один или больее количество природных или не встречающихся в природе аминокислотных остатков, как описано выше для компонента А.
Как обсуждалось выше, термин «гетерологичный» относится к аминокислоте, которая не является частью или не гомологична аминокислотной последовательности целевого антигенного сайта дикого типа (например, В-клеточного эпитопа). Таким образом, когда спейсер представляет собой аминокислоту, спейсер содержит аминокислотную последовательность, которая не встречается в природе в белке или пептиде целевого антигенного сайта. Поскольку последовательность компонента В гетерологична целевому антигенному сайту, природная аминокислотная последовательность целевого антигенного сайта не удлиняется ни в N-концевом, ни в С-концевом направлениях, когда компонент В ковалентно связан с целевым антигенным сайтом.
Спейсер может быть гибким шарнирным спейсером для улучшения разделения Th-эпитопа и целевого антигенного сайта. В некоторых вариантах осуществления гибкая шарнирная последовательность может быть богатой пролином. В некоторых вариантах осуществления гибкий шарнир имеет последовательность Pro-Pro-Xaa-Pro-Xaa-Pro (SEQ ID NO: 55), которая моделируется на основе гибкой шарнирной области, обнаруженной в тяжелых цепях иммуноглобулина. Хаа в нем может быть любой аминокислотой. В некоторых вариантах осуществления Xaa представляет собой аспарагиновую кислоту. В некоторых вариантах осуществления конформационное разделение, обеспечиваемое спейсером, может обеспечить более эффективные взаимодействия между презентованным пептидным иммуногеном и соответствующими Th-клетками и B-клетками. Иммунные ответы на Th-эпитоп могут быть усилены, чтобы обеспечить улучшенную иммунную реактивность.
Когда o˃1, каждый B независимо является одинаковым или отличающимся. В некоторых вариантах осуществления B представляет собой Gly-Gly, Pro-Pro-Xaa-Pro-Xaa-Pro (SEQ ID NO: 55), εNLys, εNLys-Lys-Lys-Lys (SEQ ID NO: 53), Lys-Lys-Lys-εNLys (SEQ ID NO: 54), Lys-Lys-Lys, -NHCH(X)CH2SCH2CO-, -NHCH(X)CH2SCH2CO(εNLys)-, -NHCH(X)CH2S-сукцинимидил-εNLys-, or -NHCH(X)CH2S-(сукцинимидил)-, и/или любую их комбинацию. Типовые гетерологичные спейсеры приведены в Таблице 2.
Целевой антигенный сайт
Целевой антигенный сайт может включать любую аминокислотную последовательность любого целевого пептида или белка, включая чужеродные или собственные пептиды или белки, эпитоп В-клеток, эпитоп CTL, пептидный гаптен, непептидный гаптен или их иммунологически реактивный аналог. Целевой антигенный сайт может представлять собой конкретный белок, углевод, связанный с раковым антигеном, низкомолекулярное лекарственное соединение или любую аминокислотную последовательность любого целевого пептида или белка. В некоторых вариантах осуществления данного изобретения описаны искусственные Th-эпитопы, которые можно применять для получения пептидных иммуногенов, которые вызывают антитела, нацеленные на амилоид β(Aβ), капсидный белок ящура (FMD), гликопротеин вируса свиного репродуктивного и респираторного синдрома (PRRSV), гормон, стимулирующий высвобождение лютеинизирующего гормона (LHRH), и любые другие пептидные или белковые последовательности.
В определенных вариантах осуществления целевой антигенный сайт взят из аутоантигена или ассоциированного с опухолью неоантигена-мишени, который в норме является «иммуномолчащим» (например, Aβ, тау, альфа-синуклеин, дипептидный белок, IgE EMPD, IL-6, CGRP, амилин, IL-31, неоантигены и др.). Неограничивающие репрезентативные последовательности аутоантигенов и ассоциированных с опухолью неоантигенных сайтов приведены в Таблице 3A. В некоторых вариантах осуществления целевой антигенный сайт взят из патогенного организма (например, FMDV, PRRSV, CSFV, ВИЧ, HSV и т. д.). Неограничивающие репрезентативные последовательности патогенных антигенных сайтов приведены в Таблице 3B.
В конкретных вариантах осуществления целевой антигенный сайт происходит из частей гормона, высвобождающего лютеинизирующий гормон (LHRH) (например, патенты США № 6 025 468, 6 228 987, 6 559 282 и публикация США № US2017/0216418); амилоида β (Aβ) (например, патенты США № 6 906 169, 7 951 909, 8 232 373 и 9 102 752); капсидного белка ящура (например, патенты США № 6 048 538, 6 107 021 и публикация США № 2015/0306203); эпитопов вирионов ВИЧ для профилактики и лечения ВИЧ-инфекции (например, патенты США № 5 912 176, 5 961 976 и 6 090 388); капсидного белка цирковируса свиней типа 2 (PCV2) (например, публикация США № 2013/0236487), гликопротеина вируса свиного репродуктивного и респираторного синдрома (PRRSV) (например, публикация США № 2014/0335118), IgE (например, патенты США № 7648701 и 6811782), альфа-синуклеина (α-Syn) (Международная заявка PCT № PCT/US2018/037938), ближайшего к мембране внеклеточного домена мембраносвязанного IgE (или EMPD IgE) (Международная заявка РСТ № PCT/US2017/069174), Tau (Международная заявка PCT № PCT/US2018/057840), и интерлейкина-31 (IL-31) (Международная заявка PCT № PCT/US2018/065025), антигена CS плазмодия для профилактики малярии; CETP для профилактики и лечения атеросклероза; IAPP (амилин) для профилактики и лечения диабета 2 типа и любого другого пептида или белковой последовательности. Все патенты и патентные публикации включены в данный документ в полном объеме посредством ссылок.
В других вариантах осуществления целевой антигенный сайт представляет собой непептидный гаптен, включая ассоциированные с опухолью углеводные антигены (TACA) и низкомолекулярное лекарственное соединение. Примеры TACA включают GD3, GD2, Глобо-H, GM2, Фукозил GM1, GM2, PSA, Ley, Lex, SLex, SLea, Tn, TF и STn, как дополнительно обсуждается в Примере 11 и продемонстрировано на Фигурах 16 и 17.
Th - T-хелперный эпитоп
Неизбирательный искусственный эпитоп клеток Т-хелперов (Th) в пептидной иммуногенной конструкции усиливает иммуногенность целевого антигенного сайта, что способствует продукции специфических антител с высоким титром, направленных против оптимизированного целевого В-клеточного эпитопа, за счет рационального дизайна.
В данном контексте термин «неизбирательный» относится к Th-эпитопу, который является реактивным у разных видов и у индивидуумов одного вида.
Термин «искусственный», применяемый в связи с эпитопами Th, относится к аминокислотным последовательностям, которые не встречаются в природе. Соответственно, искусственные Th-эпитопы по данномуу изобретению имеют последовательности, гетерологичные по отношению к целевому антигенному сайту. Как обсуждалось выше, термин «гетерологичный» относится к аминокислотной последовательности, которая получена из аминокислотной последовательности, которая не является частью или не гомологична последовательности целевого антигенного сайта дикого типа. Таким образом, гетерологичный Th-эпитоп представляет собой Th-эпитоп, полученный из аминокислотной последовательности, которая не встречается в природе в целевом антигенном сайте. Поскольку Th-эпитоп гетерологичен целевому антигенному сайту, природная аминокислотная последовательность целевого антигенного сайта не удлиняется ни в N-концевом, ни в C-концевом направлениях, когда гетерологичный Th-эпитоп ковалентно связан с целевым антигенным сайтом.
Th-эпитоп может иметь аминокислотную последовательность, полученную от любого вида (например, человека, свиньи, крупного рогатого скота, собаки, крысы, мыши, морских свинок и т. д.). Th-эпитоп может также иметь мотивы неизбирательного связывания с молекулами MHC класса II множества видов. В некоторых вариантах осуществления Th-эпитоп содержит множественные неизбирательные мотивы связывания MHC класса II для обеспечения максимальной активации Т-хелперов, которая приводит к инициации и регуляции иммунных ответов. Предпочтительно, Th-эпитоп сам по себе является иммуномолчащим, то есть. небольшое количество антител, генерируемых пептидными иммуногенными конструкциями, если они вообще есть, будут направлены на Th-эпитоп, что позволяет получить очень сфокусированный иммунный ответ, направленный на целевой антигенный сайт.
Th-эпитопы могут иметь размер от около 15 до около 50 аминокислотных остатков. В некоторых вариантах осуществления Th-эпитопы могут иметь около 15, около 20, около 25, около 30, около 35, около 40, около 45 или около 50 аминокислотных остатков. Th-эпитопы могут иметь общие структурные особенности и специфические опознавательные последовательности. В некоторых вариантах осуществления Th-эпитопы имеют амфипатические спирали, то есть альфа-спиральные структуры с гидрофобными аминокислотными остатками, доминирующими на одной поверхности спирали, и заряженными и полярными остатками, доминирующими на окружающих поверхностях.
Th-эпитопы и описания из публикации WO 1999/066957 и соответствующего патента США № 6 713 301 включены в данное описание в полном объеме посредством ссылки.
Неизбирательная Th-детерминанта может быть эффективной для усиления слабо иммуногенного пептида. Хорошо модифицированные химерные пептиды неизбирательных Th/B-клеточных эпитопов могут вызывать Th-ответы с гуморальными иммунными ответами, нацеленными на сайт B-клеток у большинства представителей генетически разнообразной популяции. В некоторых вариантах осуществления Th-клетки могут быть доставлены к целевому пептидному антигену путем ковалентного связывания пептида-носителя с хорошо изученной неизбирательной Th-детерминантой.
Неизбирательные Th-эпитопы могут содержать дополнительные первичные аминокислотные паттерны. В некоторых вариантах осуществления неизбирательные Th-эпитопы могут содержать последовательность Ротбарда, в которой неизбирательный Th-эпитоп содержит заряженный остаток (например, -Gly-), за которым следуют два-три гидрофобных остатка, за которыми следует заряженный или полярный остаток (Rothbard and Taylor, EMBO J, 1988; 7:93-101). Неизбирательные Th-эпитопы могут подчиняться правилу 1, 4, 5, 8, в котором за положительно заряженным остатком следуют гидрофобные остатки в четвертом, пятом и восьмом положениях, что соответствует амфипатической спирали, имеющей положения 1, 4, 5 и 8, расположенные на той же поверхности. В некоторых вариантах осуществления профиль 1, 4, 5, 8 гидрофобных, заряженных и полярных аминокислот может повторяться внутри одного Th-эпитопа. В некоторых вариантах осуществления неизбирательный Т-клеточный эпитоп может содержать по меньшей мере одну из последовательности Ротбарда или эпитопа, который подчиняется правилу 1, 4, 5, 8. В других вариантах осуществления Th-эпитоп содержит более одной последовательности Ротбарда.
Неизбирательные Th-эпитопы, происходящие из патогенов, включают: поверхностный Th-клеточный эпитоп гепатита B (HBsAg Th), Th-клеточный эпитоп ядерного антигена гепатита B (HBc Th), Th-клеточный эпитоп токсина коклюша (PT Th), Th-клеточный эпитоп столбнячного токсина (TT Th), Th-клеточный эпитоп белка F вируса кори (MVF Th), Th-клеточный эпитоп главного белка внешней мембраны Chlamydia trachomatis (CT Th), Th-клеточный эпитоп токсина дифтерии (DT Th), Th-клеточный эпитоп циркумспорозоитного белка Plasmodium falciparum (PF Th), Th-клеточный эпитоп триозофосфат-изомеразы Schistosoma mansoni(SM Th) и Th-клеточный эпитоп TraT Escherichia coli (TraT Th), Clostridium tetani, Bordetella pertussis, холерный токсин, MP1 гриппа, NSP1 гриппа, вирус Эпштейна-Барр (EBV), цитомегаловирус человека (HCMV), но не ограничиваются ими. Примеры Th-эпитопов, применяемых в данном изобретении, приведены в Таблице 1.
В некоторых вариантах осуществления Th-эпитопы по данному описанию могут быть комбинаторными Th-эпитопами, содержащими смесь пептидов, содержащих схожие аминокислотные последовательности. Структурированные библиотеки синтетических антигенов (SSAL - англ.: Structured synthetic antigen libraries), также называемые комбинаторными искусственными Th-эпитопами, содержат множество Th-эпитопов с аминокислотными последовательностями, организованными вокруг структурного каркаса инвариантных остатков с заменами в определенных положениях. Последовательности эпитопов SSAL определяются путем сохранения относительно инвариантных остатков и изменения других остатков для обеспечения распознавания различных рестрикционных элементов MHC. Последовательности эпитопов SSAL могут быть определены путем выравнивания первичной аминокислотной последовательности неизбирательного Th, выбора и сохранения остатков, ответственных за уникальную структуру Th-пептида в качестве структурного каркаса, и изменения оставшихся остатков в соответствии с известными рестрикционными элементами MHC. Инвариантные и вариабельные положения с предпочтительными аминокислотами рестрикционных элементов MHC могут применяться для получения MHC-связывающих мотивов, которые можно применять для конструирования SSAL Th-эпитопов.
Гетерологичные Th-эпитопные пептиды, представленные в виде комбинаторной последовательности, содержат смесь аминокислотных остатков, представленных в определенных положениях в пределах пептидного остова на основе вариабельных остатков гомологов для этого конкретного пептида. В некоторых вариантах осуществления последовательности библиотеки Th-эпитопов сконструированы для сохранения структурных мотивов неизбирательного Th-эпитопа и для приспособления реактивности к более широкому диапазону гаплотипов. В некоторых вариантах осуществления членом SSAL может быть вырожденный Th-эпитоп SSAL1 Th1, смоделированный на основе неизбирательного эпитопа, взятого из белка F вируса кори (например, SEQ ID NO: 1-5). В других вариантах осуществления представителем SSAL может быть вырожденный Th-эпитоп SSAL2 Th2, смоделированный на основе неизбирательного эпитопа, взятого из HBsAg1 (например, SEQ ID NO: 19-24).
Общее количество пептидов, присутствующих в смеси комбинаторных искусственных Th-эпитопов (или SSAL) после синтеза, может быть рассчитано путем перемножения количества вариантов, доступных в каждом вариабельном положении, друг на друга. Например, SEQ ID NO: 16 представляет собой комбинацию из 32 различных пептидов, потому что она содержит 5 вариабельных положений, где каждое вариабельное положение имеет вариант из 2 различных остатков (то есть. 2×2x2×2x2=25=32). Аналогичным образом SEQ ID NO: 5 представляет собой комбинацию 524288 различных пептидов (т.е., 2×4x2×4x2×4x4×4x2×4x2×4=25×47=524288). Комбинаторные последовательности искусственного Th-эпитопа включают (а) смесь всех пептидов, охваченных вариабельными последовательностями, и (b) каждый отдельный пептид, содержащий единственную последовательность в пределах комбинации.
В некоторых вариантах осуществления заряженный остаток Glu или Asp может быть добавлен в положение 1 для увеличения заряда, окружающего гидрофобную поверхность Th. В некоторых вариантах осуществления гидрофобная поверхность амфипатической спирали может поддерживаться гидрофобными остатками в 2, 5, 8, 9, 10, 13 и 16. В некоторых вариантах осуществления аминокислотные остатки в 2, 5, 8, 9, 10 и 13 можно изменять, чтобы обеспечить сторону со способностью связываться с широким диапазоном рестрикционных элементов MHС. В некоторых вариантах осуществления изменение аминокислотных остатков может увеличить диапазон иммунологической реактивности искусственных Th-эпитопов.
Искусственные Th-эпитопы могут включать все свойства и особенности известных неизбирательных Th-эпитопов. В некоторых вариантах осуществления искусственные Th-эпитопы являются членами SSAL. В некоторых вариантах осуществления искусственный Th-сайт может быть объединен с пептидными последовательностями, взятыми из аутоантигенов и чужеродных антигенов, для обеспечения усиленных гуморальных иммунных ответов на сайт-специфические мишени. В некоторых вариантах осуществления искусственный Th-эпитоп иммуногена может обеспечивать эффективные и безопасные гуморальные иммунные ответы, проявлять высокую иммуногенную потенцию и демонстрировать полиспецифическую реактивную ответную реакцию.
Также предложены идеализированные искусственные Th-эпитопы. Эти идеализированные искусственные Th-эпитопы смоделированы на основе двух известных природных Th-эпитопов и прототипов пептидов SSAL, описанных в публикации WO 95/11998. SSALS включает в себя комбинаторные мотивы связывания молекулы MHC (Meister et al., 1995), предназначенные для вызова полиспецифических иммунных ответов среди представителей генетически разнообразной популяции. Прототипы пептидов SSAL были сконструированы на основе Th-эпитопов вируса кори и антигенов вируса гепатита B, модифицированных путем введения множественных MHC-связывающих мотивов. Конструкты других Th-эпитопов были смоделированы по образцу других известных Th-эпитопов путем упрощения, добавления и/или модификации множественных MHC-связывающих мотивов для получения серии новых искусственных Th-эпитопов. Неизбирательные искусственные Th-сайты были включены в синтетические пептидные иммуногены, несущие множество целевых антигенных сайтов. Полученные химерные пептиды были способны стимулировать эффективные гуморальные иммунные ответы на целевые антигенные сайты.
Прототип искусственного эпитопа клеток Т-хелперов (Th), продемонстрированный вТаблице 1 как «SSAL1 Th1», смесь четырех пептидов (SEQ ID NO: 1-4), представляет собой идеализированный Th-эпитоп, смоделированный на основе неизбирательного Th-эпитопа белка F вируса кори (Partidos et al. 1991). Модельный Th-эпитоп, продемонстрированный в Таблице 1 как «MVF Th (UBITh®5)» (SEQ ID NO: 6), соответствует остаткам 288-302 белка F вируса кори. MVF Th (SEQ ID NO: 6) был модифицирован до прототипа SSAL1 Th1 (SEQ ID NO: 1-4) путем добавления заряженного остатка Glu/Asp в положение 1 для увеличения заряда, окружающего гидрофобную поверхность эпитопа; добавления или сохранения заряженных остатков или Gly в положениях 4, 6, 12 и 14; и добавления или сохранения заряженного остатка или Gly в положениях 7 и 11 в соответствии с «правилом Ротбарда». Гидрофобная поверхность Th-эпитопа содержит остатки в положениях 2, 5, 8, 9, 10, 13 и 16. Гидрофобные остатки, обычно связанные с неизбирательными эпитопами, были заменены в этих положениях для получения комбинаторных Th-эпитопов SSAL, SSAL1 Th1 (SEQ ID NO: 1-4). Другой важной особенностью прототипа SSAL1 Th1 (SEQ ID NO: 1-4) является то, что положения 1 и 4 не полностью повторяются как палиндром с обеих сторон от положения 9, чтобы имитировать MHC-связывающий мотив. Этот палиндромный профиль «1, 4, 9» SSAL1 Th1 был дополнительно модифицирован в SEQ ID NO: 2 (Таблица 1), чтобы более точно отражать последовательность исходной MvF модели Th (SEQ ID NO: 6).
Комбинаторные искусственные Th-эпитопы можно упростить, чтобы получить серию эпитопов с одной последовательностью. Например, комбинаторная последовательность SEQ ID NO: 5 может быть упрощена до Th-эпитопов с одной последовательностью, представленных SEQ ID NO: 1-4. Эти Th-эпитопы с одной последовательностью могут быть связаны с целевыми антигенными сайтами для обеспечения повышенной иммуногенности.
В некоторых вариантах осуществления иммуногенность Th-эпитопов может быть улучшена путем удлинения N-конца с помощью неполярной и полярной незаряженной аминокислоты, например, Ile и Ser, и удлинения C-конца с помощью заряженной и гидрофобной аминокислоты, например, Lys и Phe. Кроме того, добавление остатка лизина или множественных остатков лизина (например, KKK) к Th-эпитопам может улучшить растворимость пептида в воде. Дальнейшие модификации включали замену С-конца общим MHC-связывающим мотивом AxTxIL (Meister et al, 1995).
Искусственный Th-эпитоп может быть известным природным Th-эпитопом или прототипом пептида SSAL. В некоторых вариантах осуществления Th-эпитоп из SSAL может включать комбинаторные мотивы связывания молекулы MHC, предназначенные для вызова полиспецифических иммунных ответов среди членов генетически разнообразной популяции. В некоторых вариантах осуществления прототип пептида SSAL может быть сконструирован на основе Th-эпитопов вируса кори и антигенов вируса гепатита B, модифицированных путем введения множественных MHC-связывающих мотивов. В некоторых вариантах осуществления искусственный Th-эпитоп может упрощать, добавлять и/или модифицировать множественные MHC-связывающие мотивы для получения серии новых искусственных Th-эпитопов. В некоторых вариантах осуществления недавно адаптированные неизбирательные искусственные Th-сайты могут быть включены в синтетические пептидные иммуногены, несущие множество целевых антигенных сайтов. В некоторых вариантах осуществления полученные химерные пептиды могут стимулировать эффективные гуморальные иммунные ответы на целевые антигенные сайты.
Искусственные Th-эпитопы по настоящему описанию могут быть смежными последовательностями природных или неприродных аминокислот, которые содержат сайт связывания молекул MHC класса II. В некоторых вариантах осуществления искусственный Th-эпитоп может усиливать или стимулировать гуморальный иммунный ответ на целевой антигенный сайт. В некоторых вариантах осуществления Th-эпитоп может состоять из непрерывных или прерывистых аминокислотных сегментов. В некоторых вариантах осуществления не каждая аминокислота Th-эпитопа участвует в распознавании MHC. В некоторых вариантах осуществления Th-эпитопы по изобретению могут содержать иммунологически функциональные гомологи, такие как иммуностимулирующие гомологи, перекрестно-реактивные гомологи и их сегменты. В некоторых вариантах осуществления функциональные Th-гомологи могут дополнительно содержать консервативные замены, добавления, делеции и вставки 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных остатков и обеспечивать Th-стимулирующую функцию Th-эпитопа.
Th-эпитопы могут быть прикреплены непосредственно к целевому сайту. В некоторых вариантах осуществления Th-эпитопы могут быть прикреплены к целевому сайту посредством необязательного гетерологичного спейсера, например, пептидного спейсера, такого как Gly-Gly или (ε-N) Lys. Спейсер физически отделяет Th-эпитоп от B-клеточного эпитопа и может нарушить образование любых искусственных вторичных структур, созданных связыванием Th-эпитопа или функционального гомолога с целевым антигенным сайтом, тем самым устраняя любое вмешательство в Th- и/или В-клеточные ответы.
Th-эпитопы включают идеализированные искусственные Th-эпитопы и комбинаторные идеализированные искусственные Th-эпитопы, как показано в Таблице 1. В некоторых вариантах осуществления Th-эпитоп представляет собой неизбирательный эпитоп клеток Th с SEQ ID NO: 1-52, любой его гомолог, и/или любой его иммунологический аналог. Th-эпитопы также включают иммунологические аналоги Th-эпитопов. Иммунологические аналоги Th включают иммуномодулирующие аналоги, перекрестно-реактивные аналоги и сегменты любого из этих Th-эпитопов, которые достаточны для усиления или стимуляции иммунного ответа на целевой антигенный сайт.
Функциональные иммунологические аналоги Th-эпитопных пептидов также эффективны и включены как часть данного изобретения. Функциональные иммунологические аналоги Th могут включать консервативные замены, добавления, делеции и вставки от одного до около пяти аминокислотных остатков в Th-эпитопе, которые существенно не изменяют Th-стимулирующую функцию Th-эпитопа. Консервативные замены, добавления и вставки могут быть выполнены с природными или неприродными аминокислотами, как описано выше для целевого антигенного сайта. Таблица 1 идентифицирует другой вариант функционального аналога пептида Th-эпитопа. В частности, SEQ ID NO: 6 и 7 MvF1 и MvF2 Th являются функциональными аналогами SEQ ID NO: 16 и 17 MvF4 и MvF5 в том смысле, что они различаются в аминокислотной рамке по делеции (SEQ ID NO: 6 и 7 ) или включению (SEQ ID NO: 16 и 17) двух аминокислот каждая на N- и C-концах. Различия между этими двумя сериями аналогичных последовательностей не влияют на функцию Th-эпитопов, содержащихся в этих последовательностях. Следовательно, функциональные иммунологические Th-аналоги включают несколько версий Th-эпитопа, полученного из слитого белка вируса кори MvF1-4 Ths (SEQ ID NO: 6-18) и из поверхностного белка гепатита HBsAg 1-3 Ths (SEQ ID NO: 19-31).
Th-эпитоп в конструкте пептидного иммуногена может быть ковалентно связан либо на N-, либо на C-конце целевого антигенного сайта с образованием химерного Th/B-клеточного сайта пептидного иммуногена. В некоторых вариантах осуществления Th-эпитоп может быть ковалентно присоединен к целевому антигенному сайту посредством химического связывания или посредством прямого синтеза. В некоторых вариантах осуществления Th-эпитоп ковалентно связан с N-концом целевого антигенного сайта. В других вариантах осуществления Th-эпитоп ковалентно связан с С-концом целевого антигенного сайта. В некоторых вариантах осуществления более одного Th-эпитопа ковалентно связаны с целевым антигенным сайтом. Когда более чем один Th-эпитоп связан с целевым антигенным сайтом, каждый Th-эпитоп может иметь одинаковую аминокислотную последовательность или разные аминокислотные последовательности. Кроме того, когда более чем один Th-эпитоп связан с целевым антигенным сайтом, Th-эпитопы могут быть расположены в любом порядке. Например, Th-эпитопы могут быть последовательно связаны с N-концом целевого антигенного сайта или могут быть последовательно связаны с C-концом целевого антигенного сайта, или Th-эпитоп может быть ковалентно связан с N- концом целевого антигенного сайта, в то время как отдельный Th-эпитоп ковалентно связан с C-концом целевого антигенного сайта. В расположении Th-эпитопов отсутствуют ограничения по отношению к целевому антигенному сайту.
В некоторых вариантах осуществления Th-эпитоп непосредственно ковалентно связан с целевым антигенным сайтом. В других вариантах осуществления Th-эпитоп ковалентно связан с целевым антигенным сайтом через гетерологичный спейсер, более подробно описанный ниже.
Способы синтеза
Пептидные иммуногены по данному описанию можно синтезировать химическими способами. В некоторых вариантах осуществления пептидные иммуногены по данному описанию могут быть синтезированы с использованием твердофазного пептидного синтеза. В некоторых вариантах осуществления пептиды по данному изобретению синтезируют с применением автоматизированного твердофазного пептидного синтеза Меррифилда с применением t-Boc или Fmoc для защиты α-NH2 или аминокислот боковой цепи.
Гетерологичные Th-эпитопные пептиды, представленные в виде комбинаторной последовательности содержат смесь аминокислотных остатков, представленных в определенных положениях в пределах пептидного остова на основе вариабельных остатков гомологов для этого конкретного пептида. Комбинация комбинаторных пептидов может быть синтезирована в одном процессе путем добавления смеси обозначенных защищенных аминокислот вместо одной конкретной аминокислоты в определенное положение во время процесса синтеза. Такие комбинаторные сборки гетерологичных пептидов Th-эпитопа могут обеспечивать широкий охват Th-эпитопов для животных, имеющих разнообразный генетический фон. Репрезентативные комбинаторные последовательности гетерологичных пептидов Th-эпитопа включают SEQ ID NO: 5, 10, 13, 16, 24 и 27, которые приведены в Таблице 1. Th-эпитопные пептиды согласно данному изобретению обеспечивают широкую реактивность и иммуногенность для животных и пациентов из генетически разнообразных популяций.
Представляет интерес тот факт, что несоответствия и/или ошибки, которые могут быть внесены во время синтеза Th-эпитопа, B-клеточного эпитопа и/или пептидного иммуногенного конструкта, содержащих Th-эпитоп и B-клеточный эпитоп, чаще всего не препятствуют или не предотвращают желаемый иммунный ответ у животного, которое подвергается лечению. Фактически, несоответствия/ошибки, которые могут быть внесены во время синтеза пептида, генерируют множественные аналоги пептидов вместе с целевым синтезом пептидов. Эти аналоги могут включать вставку, делецию, замену аминокислоты, и преждевременную терминацию. Как описано выше, такие пептидные аналоги применимы в пептидных препаратах в качестве факторов, влияющих на антигенность и иммуногенность, при применении в иммунологических целях либо в качестве твердофазного антигена для целей иммунодиагностики, либо в качестве иммуногенов для целей вакцинации.
Пептидные иммуногенные конструкты, содержащие Th-эпитопы, продуцируются одновременно в одном твердофазном пептидном синтезе в тандеме с целевым антигенным сайтом. Th-эпитопы также включают иммунологические аналоги Th-эпитопов. Иммунологические аналоги Th включают иммуномодулирующие аналоги, перекрестно-реактивные аналоги и сегменты любого из этих Th-эпитопов, которые достаточны для усиления или стимуляции иммунного ответа на целевой антигенный сайт.
После полной сборки желаемого пептидного иммуногена твердофазную смолу можно обработать, чтобы отщепить пептид от смолы и удалить функциональные группы на боковых цепях аминокислот. Свободный пептид можно очистить с помощью ВЭЖХ и охарактеризовать биохимически. В некоторых вариантах осуществления свободные пептиды характеризуют биохимически с помощью аминокислотного анализа. В некоторых вариантах осуществления свободные пептиды характеризуют биохимически с использованием последовательности пептида. В некоторых вариантах осуществления свободные пептиды характеризуют биохимически с помощью масс-спектрометрии.
Пептидные иммуногены по данному изобретению могут быть синтезированы с применением галогенацетилированных и цистеинилированных пептидов посредством образования тиоэфирной связи. В некоторых вариантах осуществления цистеин может быть добавлен к С-концу Th-содержащего пептида, а тиоловая группа остатка цистеина может быть применена для образования ковалентной связи с электрофильной группой, такой как Nα-хлорацетил-модифицированная группа или дериватизированная малеимидом α- или ε-NH2-группа остатка лизина. Полученное синтетическое промежуточное соединение может быть присоединено к N-концу пептида целевого антигенного сайта.
Более длинные синтетические пептидные конъюгаты могут быть синтезированы с помощью методик клонирования нуклеиновых кислот. В некоторых вариантах осуществления Th-эпитопы по данному изобретению можно синтезировать путем экспрессии рекомбинантной ДНК и РНК. Для конструирования гена, экспрессирующего Th-пептид целевого антигенного сайта по данному изобретению, аминокислотная последовательность может быть обратно транслирована в последовательность нуклеиновой кислоты. В некоторых вариантах осуществления аминокислотная последовательность обратно транслируется в последовательность нуклеиновой кислоты с применением оптимизированных кодонов для организма, в котором будет экспрессироваться ген. Может быть создан ген, кодирующий пептид. В некоторых вариантах осуществления ген, кодирующий пептид, может быть получен путем синтеза перекрывающихся олигонуклеотидов, которые кодируют пептид и необходимые регуляторные элементы. Синтетический ген можно собрать и вставить в желаемый вектор экспрессии.
Синтетические последовательности нуклеиновых кислот по данному описанию могут включать последовательности нуклеиновых кислот, которые кодируют Th-эпитопы по данному изобретению, пептиды, содержащие Th-эпитопы, их иммунологически функциональные гомологи, и конструкты нуклеиновых кислот, характеризующиеся изменениями в некодирующих последовательностях, которые не изменяют иммуногенные свойства пептида или кодируемого Th-эпитопа. Синтетический ген можно вставить в подходящий вектор клонирования, и рекомбинанты могут быть получены и охарактеризованы. Th-эпитопы и пептиды, содержащие Th-эпитопы, могут затем экспрессироваться в условиях, подходящих для выбранной системы экспрессии и хозяина. Th-эпитоп или пептид можно очистить и охарактеризовать.
Фармацевтические композиции
В данном описании также описаны фармацевтические композиции, содержащие пептидные иммуногены по данному изобретению. В некоторых вариантах осуществления фармацевтическая композиция по данному описанию может применяться в качестве фармацевтически приемлемой системы доставки для введения пептидных иммуногенов. В некоторых вариантах осуществления фармацевтическая композиция по данному описанию может содержать иммунологически эффективное количество одного или более пептидных иммуногенов.
Пептидные иммуногены по данному изобретению могут быть составлены в виде иммуногенных композиций. В некоторых вариантах осуществления иммуногенная композиция может включать адъюванты, эмульгаторы, фармацевтически приемлемые носители или другие ингредиенты, обычно входящие в состав вакцинных композиций. Адъюванты или эмульгаторы, которые можно применять в данном изобретении, включают квасцы, неполный адъювант Фрейнда (IFA), липосин, сапонин, сквален, L121, эмульсиген, монофосфориллипид A (MPL), диметилдиоктадециламмонийбромид (DDA), QS21 и ISA 720, ISA 51, ISA 35, ISA 206 и другие эффективные адъюванты и эмульгаторы. В некоторых вариантах осуществления композиция по данному изобретению может быть составлена для немедленного высвобождения. В некоторых вариантах осуществления композиция по данному изобретению может быть составлена для пролонгированного высвобождения.
Адъюванты, используемые в фармацевтической композиции, могут включать масла, соли алюминия, виросомы, фосфат алюминия (например, ADJU-PHOS®), гидроксид алюминия (например, ALHYDROGEL®), липосин, сапонин, сквален, L121, Emulsigen®, монофосфориллипид A (MPL), QS21, ISA 35, ISA 206, ISA50V, ISA51, ISA 720, а также другие адъюванты и эмульгаторы.
В некоторых вариантах осуществления фармацевтическая композиция содержит Montanide™ ISA 51 (масляная адъювантная композиция, состоящая из растительного масла и маннида олеата для получения водно-масляных эмульсий), TWEEN® 80 (также известный как: Полисорбат 80 или Полиоксиэтилен (20) сорбитан моноолеат), CpG-олигонуклеотид и/или любую их комбинацию. В других вариантах осуществления фармацевтическая композиция представляет собой эмульсию «вода в масле в воде» (то есть w/o/w) с ЭМУЛЬСИГЕНОМ или ЭМУЛЬСИГЕНОМ D в качестве адъюванта.
Фигура 1A представляет собой схематическое обобщение компонентов, которые могут быть включены в составы с пептидными иммуногенами, содержащими носители Th-эпитопов, включая UBITh®. Типовый состав, продемонстрированный на Фигуре 1A, содержит две отдельные пептидные иммуногенные конструкции. Каждая пептидная иммуногенная конструкция содержит (а) индивидуальный функциональный В-клеточный эпитоп, который предназначен для выработки высоконаправленных антител против желаемых эпитопов, (b) линкер, который помогает оптимизировать презентацию В-клеточного эпитопа иммунной системе для повышения иммуногенности и (c) Th-эпитоп, который сам по себе является иммуностимулирующим, но помогает вызвать устойчивый ответ на В-клеточный эпитоп. Указанный состав также содержит фармацевтически приемлемый адъювант или носитель, пригодный для конкретного применения, в котором будет использоваться указанная композиция. Кроме того, композиция может быть составлена в виде стабилизированного иммуностимулирующего комплекса с применением олигонуклеотида CpG (дополнительно описанного ниже).
На Фигуре 1B обобщены некоторые особенности и технические преимущества платформы эпитопов Т-хелперных клеток, описанной в данном документе, включая UBITh®. Например, платформа Th-эпитопа, описанная в данном документе, характеризуется длительным действием, но обратима в отсутствие бустерной дозы. Платформа Th-эпитопа генерирует высокоспецифичные антитела, направленные к B-клеточному эпитопу, с небольшим количеством антител, если таковые имеются, к линкеру или последовательностям Th-эпитопа. Кроме того, платформа Th-эпитопа может применяться с широким спектром B-клеточных эпитопов, что позволяет создавать бесконечные комбинации пептидных иммуногенных конструкций.
В некоторых вариантах осуществления композиция составлена для применения в качестве вакцины. Вакцинную композицию можно вводить любым удобным путем, включая подкожное, пероральное, внутримышечное, внутрибрюшинное, парентеральное или энтеральное введение. В некоторых вариантах осуществления иммуногены вводят в однократной дозе. В некоторых вариантах осуществления иммуногены вводят в нескольких дозах.
Фармацевтические композиции могут быть приготовлены в виде впрыскиваемых лекарственных средств, или в виде жидких растворов, или суспензий. Жидкие носители, содержащие пептидную иммуногенную конструкцию tau, также можно приготовить перед инъекцией. Фармацевтическую композицию можно вводить любым пригодным для этого способом, например, п/к, в/в, п/о, в/м, интраназально, перорально, подкожно и т. д., и в любом пригодном устройстве доставки. В определенных вариантах осуществления фармацевтическую композицию составляют для внутривенного, подкожного, внутрикожного или внутримышечного введения. Также могут быть приготовлены фармацевтические композиции, пригодные для других способов введения, включая пероральное и интраназальное применение.
Композиция по данному изобретению может содержать эффективное количество одного или большего количества пептидных иммуногенов и фармацевтически приемлемый носитель. В некоторых вариантах осуществления композиция в пригодной единичной дозированной форме может содержать от около 0,5 мкг до около 1 мг пептидного иммуногена на кг веса тела субъекта. В некоторых вариантах осуществления композиция в пригодной единичной дозированной форме может содержать около 10 мкг, около 20 мкг, около 30 мкг, около 40 мкг, около 50 мкг, около 60 мкг, около 70 мкг, около 80 мкг, около 90 мкг, около 100 мкг, около 200 мкг, около 300 мкг, около 400 мкг, около 500 мкг, около 600 мкг, около 700 мкг, около 800 мкг, около 900 мкг или около 1000 мкг пептидного иммуногена на кг веса тела субъекта. В некоторых вариантах осуществления композиция в пригодной дозированной форме может содержать около 100 мкг, около 150 мкг, около 200 мкг, около 250 мкг, около 300 мкг, около 350 мкг, около 400 мкг, около 450 мкг или около 500 мкг пептидного иммуногена на кг веса тела субъекта. В некоторых вариантах осуществления композиция в пригодной единичной дозированной форме может содержать от около 0,5 мкг до около 1 мг пептидного иммуногена на кг веса тела субъекта. В некоторых вариантах осуществления композиция в пригодной единичной дозированной форме может содержать около 10 мкг, около 20 мкг, около 30 мкг, около 40 мкг, около 50 мкг, около 60 мкг, около 70 мкг, около 80 мкг, около 90 мкг, около 100 мкг, около 200 мкг, около 300 мкг, около 400 мкг, около 500 мкг, около 600 мкг, около 700 мкг, около 800 мкг, около 900 мкг или около 1000 мкг пептидного иммуногена. В некоторых вариантах осуществления композиция в пригодной дозированной форме может содержать около 100 мкг, около 150 мкг, около 200 мкг, около 250 мкг, около 300 мкг, около 350 мкг, около 400 мкг, около 450 мкг или около 500 мкг пептидного иммуногена.
При доставке несколькими дозами композиция может быть разделена на соответствующее количество на дозу. В некоторых вариантах осуществления доза составляет от около 0,2 мг до около 2,5 мг. В некоторых вариантах осуществления доза составляет около 1 мг. В некоторых вариантах осуществления доза составляет около 1 мг и вводится путем инъекции. В некоторых вариантах осуществления доза составляет около 1 мг и вводится внутримышечно. В некоторых вариантах осуществления за дозой может следовать повторная (бустерная) доза. Дозировки могут быть оптимизированы в зависимости от возраста, веса и общего состояния здоровья пациента.
Вакцины, содержащие смеси пептидных иммуногенов, могут обеспечивать повышенную иммуноэффективность в более широкой популяции. В некоторых вариантах осуществления смесь пептидных иммуногенов содержит Th-сайты, полученные из MVF Th и HBsAg Th. В некоторых вариантах осуществления вакцины, содержащие смеси пептидных иммуногенов, могут обеспечивать улучшенный иммунный ответ на целевой антигенный сайт.
Иммунный ответ на конъюгаты Th/целевой антигенный сайт можно улучшить путем доставки посредством захвата в или на биоразлагаемых микрочастицах. В некоторых вариантах осуществления пептидные иммуногены можно инкапсулировать с адъювантом или без него, и такие микрочастицы могут нести иммуностимулирующий адъювант. В некоторых вариантах осуществления микрочастицы можно вводить совместно с пептидными иммуногенами для усиления иммунных ответов.
Иммуностимулирующие комплексы
Данное изобретение также относится к фармацевтическим композициям, содержащим пептидную иммуногенную конструкцию tau в форме иммуностимулирующего комплекса с олигонуклеотидом CpG. Такие иммуностимулирующие комплексы специально приспособлены для действия в качестве адъюванта и стабилизатора пептидного иммуногена. Иммуностимулирующие комплексы находятся в форме частиц, которые могут эффективно презентировать пептидный иммуноген tau клеткам иммунной системы, вызывая иммунный ответ. Иммуностимулирующие комплексы могут быть приготовлены в виде суспензии для парентерального введения. Иммуностимулирующие комплексы также могут быть составлены в форме в/м эмульсий, в виде суспензии в комбинации с минеральной солью или с желирующим полимером in situ для эффективной доставки пептидного иммуногена tau к клеткам иммунной системы хозяина после парентерального введения.
Стабилизированный иммуностимулирующий комплекс может быть образован путем комплексирования пептидного иммуногенного конструкта tau с анионной молекулой, олигонуклеотидом, полинуклеотидом или их комбинациями посредством электростатической ассоциации. Стабилизированный иммуностимулирующий комплекс может быть включен в фармацевтическую композицию в качестве системы доставки иммуногена.
В некоторых вариантах осуществления пептидный иммуногенный конструкт tau сконструирован с возможностью содержания катионной части, которая положительно заряжена при рН в диапазоне от 5,0 до 8,0. Общий заряд катионной части пептидного иммуногенного конструкта tau или смеси конструктов рассчитывают, назначая заряд +1 для каждого лизина (K), аргинина (R) или гистидина (H), заряд -1 для каждой аспарагиновой кислоты (D) или глутаминовой кислоты (E) и заряд 0 для другой аминокислоты в пределах последовательности. Заряды суммируются в пределах катионной части пептидной иммуногенной конструкции tau и выражаются как общий средний заряд. Пригодный пептидный иммуноген имеет катионную часть с чистым средним положительным зарядом +1. Предпочтительно пептидный иммуноген имеет чистый положительный заряд в диапазоне, превышающем +2. В некоторых вариантах осуществления катионная часть пептидной иммуногенной конструкции tau представляет собой гетерологичный спейсер. В определенных вариантах осуществления катионная часть пептидного иммуногенного конструкта tau имеет заряд +4, когда последовательность спейсера представляет собой (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 53) или Lys-Lys-Lys-ε-N-Lys (SEQ ID NO: 54).
«Анионная молекула», как описано в данном документе, относится к любой молекуле, которая имеет отрицательный заряд при pH в диапазоне 5,0-8,0. В некоторых вариантах осуществления анионная молекула представляет собой олигомер или полимер. Чистый отрицательный заряд олигомера или полимера рассчитывается путем присвоения заряда -1 для каждой фосфодиэфирной или фосфоротиоатной группы в олигомере. Пригодным анионным олигонуклеотидом является одноцепочечная молекула ДНК с 8-64 нуклеотидными основаниями с числом повторов CpG-мотива в диапазоне от 1 до 10. Предпочтительно иммуностимулирующие одноцепочечные молекулы CpG ДНК содержат 18-48 нуклеотидных оснований с числом повторов мотива CpG в диапазоне от 3 до 8.
Более предпочтительно анионный олигонуклеотид представлен формулой: 5' X1CGX2 3', где C и G являются неметилированными; а X1 выбран из группы, состоящей из A (аденина), G (гуанина) и T (тимина); и Х2 представляет собой С (цитозин) или Т (тимин). Или анионный олигонуклеотид представлен формулой: 5 '(X3)2CG (X4)2 3', где C и G неметилированы; и X3 выбран из группы, состоящей из A, T или G; и X4 представляет собой C или T. В некоторых вариантах реализации олигонуклеотид CpG может быть CpG1 (SEQ ID NO: 146), CpG2 (SEQ ID NO: 147) или CpG3 (SEQ ID NO: 148).
Полученный иммуностимулирующий комплекс имеет форму частиц с размером обычно в диапазоне от 1 до 50 микрон и является функцией многих факторов, включая стехиометрию относительного заряда и молекулярную массу взаимодействующих частиц. Иммуностимулирующий комплекс в виде частиц имеет то преимущество, что обеспечивает адъювантацию и активацию специфических иммунных ответов in vivo. Кроме того, стабилизированный иммуностимулирующий комплекс является пригодным для приготовления фармацевтических композиций различными способами, включая водно-масляные эмульсии, суспензии минеральных солей и полимерные гели.
Применения
Пептидные иммуногены, содержащие искусственные Th-эпитопы согласно данному описанию, могут быть полезны в целях медицинского и ветеринарного применения. В некоторых вариантах осуществления пептидные иммуногены по данному изобретению можно применять в качестве вакцин для обеспечения защитного иммунитета от инфекционного заболевания, иммунотерапии для лечения расстройств, возникающих в результате нарушения нормальных физиологических процессов, в качестве иммунотерапевтических средств для лечения рака и в качестве агентов для вмешательства или изменения нормальных физиологических процессов.
Искусственные Th-эпитопы по данному описанию могут вызывать иммунный ответ в сочетании с целевыми В-клеточными эпитопами различных микроорганизмов, белков или пептидов. В некоторых вариантах осуществления искусственные Th-эпитопы по данному описанию могут быть связаны с одним целевым антигенным сайтом. В некоторых вариантах осуществления искусственный Th-эпитоп по данному описанию может быть связан с двумя целевыми антигенными сайтами.
Искусственные Th-эпитопы по данному описанию могут быть связаны с целевыми антигенными сайтами для предотвращения и/или лечения различных заболеваний и патологических состояний. В некоторых вариантах осуществления композиция по данному изобретению может применяться для профилактики и/или лечения нейродегенеративных заболеваний, инфекционных заболеваний, артериосклероза, рака предстательной железы, профилактики неприятного запаха кабана, иммунокастрации животных, лечения эндометриоза, рака молочной железы и других гинекологических раковых заболеваний, вызванных гонадными стероидными гормонами, а также для контрацепции у мужчин и женщин. Например, искусственные Th-эпитопы могут быть связаны с антигенными сайтами следующих белков:
a. Соматостатина для стимуляции роста сельскохозяйственных животных.
b. IgE для лечения аллергических заболеваний.
c. Рецептора CD4 Th-клеток для лечения и/или профилактики инфекций вируса иммунодефицита человека (ВИЧ) и иммунных расстройств.
d. Капсидного белка вируса ящура (FMD) для профилактики FMD.
e. Эпитопов вирионов ВИЧ для профилактики и лечения ВИЧ-инфекций.
f. Циркумспорозоитного антигена Plasmodium falciparum для профилактики и лечения малярии.
g. CETP для профилактики и лечения артериосклероза.
h. Aβ для лечения или вакцинации от болезни Альцгеймера.
i. Альфа-синуклеина для лечения или вакцинации от болезни Паркинсона.
j. Tau для лечения и вакцинации от таупатий, включая болезнь Альцгеймера.
k. IL-31 для лечения атопического дерматита.
l. CGRP для профилактики и лечения мигрени.
m. IAPP (амилин) для профилактики и лечения диабета 2 типа.
Было обнаружено, что применение гетерологичных искусственных Th-эпитопов особенно важно для нацеливания на белки, участвующие в патогенезе нейродегенеративных заболеваний (например, Aβ, альфа-синуклеин, Tau). В частности, пептидные иммуногены, которые содержат эндогенные Th-эпитопы целевых нейродегенеративных белков, могут вызывать воспаление головного мозга при введении субъекту. Напротив, пептидные иммуногенные конструкции, которые содержат гетерологичный искусственный Th-эпитоп, похожий на антигенный сайт нейродегенеративного белка, не вызывают воспаления головного мозга.
Фигура 2 представляет собой график, иллюстрирующий теоретические результаты, которые могут быть получены с применением платформы Th-эпитопов, описанной в данной заявке. Приведенный график демонстрирует, что пептидные иммуногенные конструкции, содержащие В-клеточные эпитопы, конъюгированные с Th-эпитопами, характеризуются быстрым началом реакции, быстро достигая Cmax. Cmax находится в терапевтическом диапазоне, который находится между минимальной эффективной концентрацией (MEC) и минимальной терапевтической концентрацией (MTC). Примеры, которые следуют ниже, демонстрируют, что Cmax, продолжительность действия, время начала реакции и tmax можно контролировать и/или регулировать путем варьирования применяемого Th-эпитопа, дозы пептидной иммуногенной конструкции и адъюванта.
Амилоид β
Полагают, что пептид Aβ является причиной возникновения и прогрессирования болезни Альцгеймера. Предполагается, что токсические формы олигомеров Aβ и фибрилл Aβ ответственны за гибель синапсов и нейронов, которая приводит к развитию болезни Альцгеймера и деменции. Успешная терапия, модифицирующая заболевание, для болезни Альцгеймера может включать средства, которые влияют на локализацию Aβ в головном мозге.
Пептидный иммуноген по данному описанию может содержать Th-клеточные эпитопы и пептиды, нацеленные на Aβ. В некоторых вариантах осуществления Th-клеточный эпитоп представляет собой Th1 или Th2. В некоторых вариантах осуществления пептидный иммуноген может содержать Th1 и Th2. Пептид, нацеленный на Aβ, или B-клеточные эпитопы могут представлять собой Aβ1-14, Aβ1-16, Aβ1-28, Aβ17-42 или Aβ1-42. В некоторых вариантах осуществления пептид, нацеленный на Aβ, представляет собой Aβ1-14. В данном контексте термин Aβx-y обозначает последовательность Aβ от аминокислоты x до аминокислоты y полноразмерного белка Aβ дикого типа.
Пептидный иммуноген по настоящему описанию может содержать более одного пептида, нацеленного на Aβ. В некоторых вариантах осуществления пептидный иммуноген может содержать два пептида, нацеленных на Aβ. В некоторых вариантах осуществления пептидный иммуноген может содержать один пептид Aβ1-14 и один пептид Aβ1-42. В некоторых вариантах осуществления пептидный иммуноген может содержать два пептида, нацеленных на Aβ1-14. В некоторых вариантах осуществления пептидный иммуноген может содержать два пептида, нацеленных на Aβ1-14, каждый из которых связан с разными Th-клеточными эпитопами в виде химерного пептида.
В данном описании также предложены пептидные вакцины Aβ1-14, содержащие два пептида, нацеленных на Aβ1-14, каждый из которых связан с разными Th-клеточными эпитопами в виде химерного пептида. В некоторых вариантах осуществления химерный пептид Aβ1-14 может быть составлен в систему доставки с Th1-смещением для минимизации воспалительной реактивности Т-клеток. В некоторых вариантах осуществления химерный пептид Aβ1-14 может быть составлен в систему доставки с Th2-смещением для минимизации воспалительной реактивности Т-клеток.
Конкретные варианты осуществления данного изобретения
(1) Неизбирательный искусственый эпитоп Т-хелперных клеток (Th), выбранный из группы, состоящей из SEQ ID NO: 32-52.
(2) Пептидная иммуногенная конструкция, представленная следующими формулами:
(A)n-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
{(A)n-(Th)p-(B)o-(целевой антигенный сайт)-(B)o-(Th)p-(A)n-X}m
где:
каждый A независимо представляет собой аминокислоту;
каждый B независимо представляет собой гетерологичный спейсер;
каждый Th независимо представляет собой неизбирательный искусственный Th-эпитоп по п. (1);
целевой антигенный сайт представляет собой В-клеточный эпитоп из белка чужеродного антигена, белка аутоантигена или его иммунологически реактивного аналога;
X представляет собой аминокислоту, α-COOH или α-CONH2;
n равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10;
m равно 1, 2, 3 или 4; а также
o равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
p равно 0, 1, 2, 3 или 4;
(3) Пептидная иммуногенная конструкция по п. (2), отличающаяся тем, что целевой антигенный сайт представляет собой В-клеточный эпитоп из белка чужеродного антигена, выбранного из группы, состоящей из капсидного белка вируса ящура (FMD), гликопротеина вируса репродуктивного и респираторного синдрома свиней (PRRSV), вируса классической чумы свиней (CSFV), вируса иммунодефицита человека (ВИЧ) и вируса простого герпеса (HSV).
(4) Пептидная иммуногенная конструкция по п. (2), отличающаяся тем, что целевой антигенный сайт представляет собой В-клеточный эпитоп из аутоантигенного белка, выбранного из группы, состоящей из:
(a) пептида Aβ, имеющего аминокислотную последовательность SEQ ID NO: 56, 57, 58, 59 или 60;
(b) пептида альфа-Syn, имеющего аминокислотную последовательность SEQ ID NO: 61;
(c) пептида IgE EMPD, имеющего аминокислотную последовательность SEQ ID NO: 62;
(d) пептида тау, имеющего аминокислотную последовательность SEQ ID NO: 63, 69, 70 или 71;
(e) пептида IL-31, имеющего аминокислотную последовательность SEQ ID NO: 64 или 72; а также
(f) пептида IL-6, имеющего аминокислотную последовательность SEQ ID NO: 145.
(5) Пептидная иммуногенная конструкция по п. (2), отличающаяся тем, что гетерологичный спейсер компонента B выбран из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N) Lys, ε-N-Lys-Lys-Lys-Lys ( SEQ ID NO: 53), Lys-Lys-Lys-εNLys (SEQ ID NO: 54), Gly-Gly, Pro-Pro-Xaa-Pro-Xaa-Pro (SEQ ID NO: 55) и любой их комбинации.
(6) Пептидная иммуногенная конструкция по п. (2), отличающаяся тем, что гетерологичный спейсер выбран из группы, состоящей из (α, ε-N) Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 53) и Lys-Lys-Lys-εNLys (SEQ ID NO: 54).
(7) Фармацевтическая композиция, содержащая пептидную иммуногенную конструкцию по п. (2).
(8) Способ предотвращения и/или лечения заболевания, патологического состояния или болезни у субъекта, включающий введение указанному субъекту фармацевтически эффективного количества фармацевтической композиции по (7).
(9) Способ по п. (8), отличающийся тем, что целевой антигенный сайт представляет собой В-клеточный эпитоп из белка чужеродного антигена, выбранного из группы, состоящей из капсидного белка вируса ящура (FMD), гликопротеина вируса репродуктивного и респираторного синдрома свиней (PRRSV), вируса классической чумы свиней (CSFV), вируса иммунодефицита человека (ВИЧ) и вируса простого герпеса (HSV).
(10) Способ по п. (8), отличающийся тем, что целевой антигенный сайт представляет собой В-клеточный эпитоп из аутоантигенного белка, выбранного из группы, состоящей из:
(a) пептида Aβ, имеющего аминокислотную последовательность SEQ ID NO: 56, 57, 58, 59 или 60;
(b) пептида альфа-Syn, имеющего аминокислотную последовательность SEQ ID NO: 61;
(c) пептида IgE EMPD, имеющего аминокислотную последовательность SEQ ID NO: 62;
(d) пептида тау, имеющего аминокислотную последовательность SEQ ID NO: 63, 69, 70 или 71;
(e) пептида IL-31, имеющего аминокислотную последовательность SEQ ID NO: 64 или 72; и
(f) пептида IL-6, имеющего аминокислотную последовательность SEQ ID NO: 145.
(11) Пептидная иммуногенная конструкция, представленная следующими формулами:
(A)n-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(A)n-X
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X
или
{(A)n-(Th)p-(B)o-(целевой антигенный сайт)-(B)o-(Th)p-(A)n-X}m
где:
каждый A независимо представляет собой аминокислоту;
каждый B независимо представляет собой гетерологичный спейсер;
каждый Th независимо представляет собой неизбирательный искусственный Th-эпитоп, выбранный из группы, состоящей из SEQ ID NO: 1-52;
Целевой антигенный сайт представляет собой эпитоп CTL, ассоциированный с опухолью углеводный антиген (TACA), В-клеточный эпитоп из неоантигена, низкомолекулярное лекарственное средство или его иммунологически реактивный аналог;
X представляет собой аминокислоту, α-COOH или α-CONH2;
n равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10;
m равно 1, 2, 3 или 4; а также
o равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
p равно 0, 1, 2, 3 или 4;
(12) Пептидный иммуноген по п. (11), отличающийся тем, что целевой антигенный сайт представляет собой эпитоп CTL, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 76-144.
(13) Пептидный иммуноген по п. (12), отличающийся тем, что целевой антигенный сайт представляет собой эпитоп CTL из ВИЧ, выбранный из группы, состоящей из SEQ ID NO: 76-82.
(14) Пептидный иммуноген по п. (12), отличающийся тем, что целевой антигенный сайт представляет собой эпитоп CTL из HSV, выбранный из группы, состоящей из SEQ ID NO: 83-106.
(15) Пептидный иммуноген по п. (12), отличающийся тем, что целевой антигенный сайт представляет собой эпитоп CTL из FMDV, выбранный из группы, состоящей из SEQ ID NO: 107-123.
(16) Пептидный иммуноген по п. (12), отличающийся тем, что целевой антигенный сайт представляет собой эпитоп CTL из PRRSV, выбранный из группы, состоящей из SEQ ID NO: 124-142.
(17) Пептидный иммуноген по п. (12), отличающийся тем, что целевой антигенный сайт представляет собой эпитоп CTL из CSFV, выбранный из группы, состоящей из SEQ ID NO: 143-144.
(18) Пептидный иммуноген по п. (11), отличающийся тем, что целевой антигенный сайт представляет собой TACA, выбранный из группы, состоящей из GD3, GD2, Глобо-H, GM2, Фукозил GM1, GM2, PSA, Ley, Lex, SLex, SLea, Tn, TF и STn.
(19) Пептидный иммуноген по п. (11), отличающийся тем, что целевой антигенный сайт представляет собой В-клеточный эпитоп из неоантигена, выбранного из группы, состоящей из SEQ ID NO: 73-75.
(20) Пептидный иммуноген по п. (11), отличающийся тем, что целевой антигенный сайт представляет собой низкомолекулярное лекарственное средство.
(21) Пептидная иммуногенная конструкция по п. (11), отличающаяся тем, что гетерологичный спейсер компонента B выбран из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N) Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 53), Lys-Lys-Lys-εNLys (SEQ ID NO: 54), Gly-Gly, Pro-Pro-Xaa-Pro-Xaa-Pro (SEQ ID NO: 55) и любой их комбинации.
(22) Пептидная иммуногенная конструкция по п. (11), отличающаяся тем, что гетерологичный спейсер выбран из группы, состоящей из (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 53), и Lys-Lys-Lys-εNLys (SEQ ID NO: 54).
(23) Фармацевтическая композиция, содержащая пептидную иммуногенную конструкцию по п. (11).
(24) Способ предотвращения и/или лечения заболевания, патологического состояния или болезни у субъекта, включающий введение указанному субъекту фармацевтически эффективного количества фармацевтической композиции по п. (23).
(25) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой ВИЧ-инфекцию, и при этом целевой антигенный сайт представляет собой CTL-эпитоп из ВИЧ, выбранный из группы, состоящей из SEQ ID NO: 76-82.
(26) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой HSV-инфекцию, и при этом целевой антигенный сайт представляет собой CTL-эпитоп из HSV, выбранный из группы, состоящей из SEQ ID NO: 83-106.
(27) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой FMDV-инфекцию, и при этом целевой антигенный сайт представляет собой CTL-эпитоп из FMDV, выбранный из группы, состоящей из SEQ ID NO: 107-123.
(28) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой PRRSV-инфекцию, и при этом целевой антигенный сайт представляет собой CTL-эпитоп из PRRSV, выбранный из группы, состоящей из SEQ ID NO: 124-142.
(29) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой CSFV-инфекцию, и при этом целевой антигенный сайт представляет собой CTL-эпитоп из CSFV, выбранный из группы, состоящей из SEQ ID NO: 143-144.
(30) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой CSFV-инфекцию, и при этом целевой антигенный сайт представляет собой CTL-эпитоп из CSFV, выбранный из группы, состоящей из SEQ ID NO: 143-144.
(31) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой рак и при этом целевой антигенный сайт представляет собой TACA, выбранный из группы, состоящей из GD3, GD2, Глобо-H, GM2, Фукозил GM1 , GM2, PSA, Ley, Lex, SLex, SLea, Tn, TF и STn.
(32) Способ по п. (24), отличающийся тем, что указанное заболевание, патологическое состояние или болезнь представляет собой рак и при этом целевой антигенный сайт представляет собой В-клеточный эпитоп из неоантигена, выбранного из группы, состоящей из SEQ ID NO: 73-75 .
(33) Способ направления иммунного ответа в организме субъекта, включающий:
(a) приготовление более чем одной пептидной иммуногенной конструкции по п. (11), в которой целевой антигенный сайт остается константным, а Th-эпитоп отличается в каждой пептидной иммуногенной конструкции;
(b) приготовление более чем одной фармацевтической композиции, каждая из которых содержит одну из пептидных иммуногенных конструкций, полученных в п. (а), и фармацевтически приемлемый адъювант или носитель;
(c) введение каждой из фармацевтических композиций, приготовленных в п. (b), различным субъектам;
(d) мониторинг иммунного ответа у каждого из субъектов; а также
(e) выбор фармацевтической композиции, которая вызывает желаемый иммунный ответ.
(34) Способ по п. (33), отличающийся тем, что фармацевтически приемлемый адъювант или носитель в каждой из фармацевтических композиций является одинаковым.
(35) Способ по п. (33), отличающийся тем, что фармацевтически приемлемый адъювант или носитель в каждой из фармацевтических композиций является разным.
ПРИМЕР 1
Приготовление пептидов и пептидных иммуногенных конструкций
Пептиды, в том числе пептидные иммуногенные конструкции, синтезировали с помощью автоматизированного твердофазного синтеза, очищали с помощью препаративной ВЭЖХ и характеризовали с помощью ионизации лазерной десорбцией - время-пролетной масс-спектрометрии с применением матрицы (MALDI-TOF), аминокислотного анализа и обращенно-фазовой ВЭЖХ.
Вакцина Aβ (UB-311) содержит два пептидных иммуногена, каждый с N-концевым пептидом Aβ1-14, синтетически связанным через аминокислотный спейсер с различными пептидами Th-клеточного эпитопа (эпитопами UBITh®), полученными из двух патогеных белков: поверхностного антигена гепатита B и слитого белка вируса кори. В частности, пептидный иммуноген, связанный со слитым белком вируса кори, представлял собой Aβ1-14-εK-KKK-MvF5 Th (SEQ ID NO: 67), а пептидный иммуноген, связанный с поверхностным антигеном гепатита B, представлял собой Aβ1-14-εK-HBsAg3 Th (SEQ ID NO: 68).
UB-311 был составлен в квасцы-содержащей Th2-смещенной системе доставки и содержал пептиды Aβ1-14-εK-HBsAg3 и Aβ1-14-εK-KKK-MvF5 Th в эквимолярном соотношении. Два иммуногена Aβ смешивали с полианионным олигодезоксинуклеотидом CpG (ODN) с образованием стабильных иммуностимулирующих комплексов из частиц микронного размера. Минеральную соль алюминия (ADJU-PHOS®) добавляли к конечному составу вместе с хлоридом натрия для придания тоничности и 0,25% 2-феноксиэтанолом в качестве консерванта.
Последовательности нескольких иллюстративных целевых антигенных сайтов (В-клеточных и CTL-эпитопов) приведены в таблицах 3A и 3B соответственно Последовательности нескольких типовых пептидных иммуногенных конструкций, содержащих Aβ1-14, крысиный IL-672-82, и IgE-EMPD1-39, в качестве целевых антигенных сайтов, приведены в Таблице 4. Последовательности пептидных иммуногенных конструкций, содержащих α-синуклеин111-132, ковалентно связанный с различными Th-эпитопами, приведены в Таблице 5. Последовательности пептидных иммуногенных конструкций, содержащих IgE-EMPDG1-C39, ковалентно связанный с различными Th-эпитопами, приведены в Таблице 6. Последовательности пептидных иммуногенных конструкций, содержащих IL-673-83 человека, ковалентно связанный с различными Th-эпитопами, приведены в Таблице 7. Последовательности пептидных иммуногенных конструкций, содержащих последовательности дипептидных повторов (DPR), ковалентно связанных с UBITh®1, приведены в Таблице 9. Последовательности пептидных иммуногенных конструкций, содержащих LHRH, ковалентно связанный с различными Th-эпитопами, приведены в Таблице 10.
ПРИМЕР 2
Ранжирование гетерологичных Т-хелперных эпитопов, происходящих от патогенов, и их включение вдизайн пептидных иммуногенных конструкций альфа-синуклеина 111-132, IgE EMPD1-39и IL-673-83 для повышения иммуногенности этих выбранных пептидов В-эпитопа
a. Синтез пептидного иммуногена
Три коротких пептида В-клетчного эпитопа из альфа-синуклеина (AA 111-132; SEQ ID NO: 61); IgE EMPD (AA 1-39; SEQ ID NO: 62); и IL-6 (AA 73-83; SEQ ID NO: 145), которые были подробно охарактеризованы по их функциональным свойствам, применяли в качестве репрезентативных целевых антигенных сайтов. Эти три В-клеточных эпитопа были превращены в пептидные иммуногенные конструкции в соответствии с формулой, приведенной ниже, для оценки способности репрезентативных неизбирательных искусственных Th-эпитопов (выбранных из SEQ ID NO: 1-52) придавать иммуногенность трем отдельным целевым антигенным сайтам:
(Th)m-(B)o-(Целевой антигенный сайт)-X
где:
Th был выбран из искусственных Th-эпитопов, описанных в данном документе, и m было равно 1;
(B)o представлял собой спейсер, имеющий аминокислотную последовательность SEQ ID NO: 53 или 54;
Целевой антигенный сайт представлял собой В-клеточный эпитоп SEQ ID NO: 61, 62 или 145; а также
X представлял собой аминокислоту -CONH2.
Аминокислотные последовательности полученных пептидных иммуногенных конструкций α-синуклеина, IgE EMPD и IL-6 приведены в Таблицах 5, 6 и 7соответственно.
b. Составы, содержащие пептидные иммуногенные конструкции и иммунизации
Репрезентативное исследование иммуногенности было проведено на морских свинках для оценки относительной эффективности соответствующих гетерологичных Т-хелперных эпитопов, приведенных в Таблице 1. После получения различных пептидных иммуногенов α-синуклеина, IgE EMPD и IL-6, конструкции, содержащие одинаковые последовательности Th-эпитопа, смешивали вместе в соотношении 1:1:1 , как продемонстрировано в Таблице 8. Например, конструкции α-синуклеина, IgE EMPD и IL-6, содержащие эпитоп UBITH®1 (SEQ ID NO: 149, 178 и 207), смешивали вместе, чтобы получить состав № 1, продемонстрированный в Таблице 8. Смесь иммуногенных конструкций α-синуклеина, IgE EMPD и пептида IL-6 смешивали с адъювантом MONTANIDE ISA50V2 и затем составляли в виде стабилизированного иммуностимулирующего комплекса с применением олигонуклеотида CpG3. Каждый из 29 составов, продемонстрированных в Таблице 8, содержал в общей сложности 135 мкг пептидов (45 мкг на пептид) в объеме 0,5 мл.
Составы вводили морским свинкам (по 3 на группу) через 0, 3 и 6 недель после начальной иммунизации (wpi - англ.: weeks post initial immunization) посредством внутримышечной (в/м) инъекции. Образцы сыворотки отбирали в моменты времени 0, 3, 6 и 8 wpi для оценки уровней титра антител.
c. Результаты иммуногенности
Результаты, полученные через 8 недель после начальной иммунизации (8 wpi), использовали для ранжирования различных пептидных иммуногенных конструкций α-синуклеина (Фигура 3, верхняя панель), IgE (Фигура 3, нижняя панель) и IL-6 (Таблица 15). Графики, содержащие данные об иммуногенности, полученные в ходе исследования для α-синуклеина, продемонстрированы на Фигуре 4A; данные для IgE-EMPD продемонстрированы на Фигуре 5; и данные для IL-6продемонстрированы на Фигуре 6.
Все Th-эпитопы были способны повышать иммуногенность трех коротких пептидов В-клеточного эпитопа в различной степени. В частности, Th-эпитопы : KKKMvF3 Th (SEQ ID NO: 13), Clostridium tetani TT2 Th (SEQ ID NO: 36), EBV EBNA-1 Th (SEQ ID NO: 42), MvF5 Th; UBITh®1 (SEQ ID NO: 17), EBV BHRF1 Th (SEQ ID NO: 41), MvF4 Th; UBITh®3 (SEQ ID NO: 16) и холерный токсин Th (SEQ ID NO: 33) усиливали иммуногенность пептида α-синуклеина (SEQ ID NO: 61) в большей степени, чем другие Th-эпитопы (верхняя панельФигуры 3 иФигура 4A). Эти пептидные иммуногенные конструкции представлены составами №№ 13, 22, 21, 01, 19, 03 и 11, соответственно, в Таблице 5.
Для пептида IgE EMPD (SEQ ID NO: 62) Th-эпитопы Clostridium tetani TT4 Th (SEQ ID NO: 38), UBITh®1 (SEQ ID NO: 17), UBITh®3 (SEQ ID NO: 16) , HBsAg1 Th; SSAL2 Th2 (SEQ ID NO: 24), KKKMvF3 Th (SEQ ID NO: 13), Clostridium tetani TT2 Th (SEQ ID NO: 36), холерный токсин Th (SEQ ID NO: 33), EBV BHRF1 Th (SEQ ID NO : 41) и HBsAg3 Th; UBITH®2 (SEQ ID NO: 28) усиливали иммуногенность IgE EMPD в большей степени, чем другие Th-эпитопы (нижняя панель Фигуры 3и Фигура 5). Эти пептидные иммуногенные конструкции представлены составами №№ 24, 01, 03, 14, 13, 22, 11, 19 и 02, соответственно, в Таблице 6.
Для циклического пептида IL-673-83 (SEQ ID NO: 145) Th-эпитопы HBsAg3 Th; UBITH®2 (SEQ ID NO: 28), UBITh®1 (SEQ ID NO: 17), UBITh®3 (SEQ ID NO: 16), Clostridium tetani TT1 Th (SEQ ID NO: 34) и Clostridium tetani TT4 Th (SEQ ID NO: 38), как было обнаружено, наиболее эффективно усиливали результирующую иммуногенность IL-6 (Таблица 15 и Фигура 6). Эти пептидные иммуногенные конструкции представлены составами №№ 02, 01, 03, 20 и 24, соответственно в Таблице 7.
Эти результаты демонстрируют, что различная иммуногенность по отношению к отдельным целевым В-клеточным эпитопам может быть получена при прмименении различных искусственных Th-эпитопов, описанных в данном документе. Тщательная калибровка иммуногенности для каждой пептидной иммуногенной конструкции у разных видов, включая приматов, необходима для обеспечения окончательного отбора Th-пептида и успеха в разработке окончательных вакцинных композиций.
c. Скорость в зависимости от Cmax
Дальнейший анализ данных иммуногенности пептидных иммуногенных конструкций α-синуклеина, ковалентно связанных с различными Th-эпитопами, демонстрирует, что конкретная Cmax может достигаться с разными скоростями в зависимости от того, какой Th-эпитоп применяется. Фигура 4B содержит подмножество данных иммуногенности, представленных на Фигуре 4A. В частности, в морских свинок, иммунизированных составами № 13 и 21, которые содержат Th-эпитопы KKKMvF3 Th (SEQ ID NO: 13) и EBV EBNA-1 Th (SEQ ID NO: 42), ковалентно связанные с пептидом α-синуклеина (SEQ ID NO: 61) достигается то же значение Cmax в момент времени 8 wpi, но с разной скоростью. В частности, пептидная иммуногенная конструкция α-синуклеина, содержащая KKKMvF3 Th (SEQ ID NO: 13), достигает своего значения Cmax быстрее, чем пептидная иммуногенная конструкция α-синуклеина, содержащая EBV EBNA-1 Th (SEQ ID NO: 42). Аналогичным образом пептидная иммуногенная конструкция α-синуклеина, содержащая UBITh®1 (SEQ ID NO: 17), достигает своего значения Cmax быстрее, чем пептидная иммуногенная конструкция α-синуклеина, содержащая EBV BHRF1 Th (SEQ ID NO: 41); и пептидная иммуногенная конструкция α-синуклеина, содержащая Clostridium tetani TT1 Th (SEQ ID NO: 34), достигает своего значения Cmax быстрее, чем пептидная иммуногенная конструкция α-синуклеина, содержащая Influenza MP1_1 Th (SEQ ID NO: 48). Результаты, приведенные на Фигуре 4B, демонстрируют, что выбор Th-эпитопа может влиять на скорость, с которой титры антител достигают своего значения Cmax.
d. Резюме
Результаты этого эксперимента демонстрируют, что иммунный ответ, вызываемый пептидными иммуногенными конструкциями (включая титры антител, Cmax, начало продукции антител, продолжительность ответа и т. д.), можно модулировать путем выбора Th-эпитопа, который химически связан с В-клеточным эпитопом. Следовательно, специфические иммунные ответы на целевые антигенные сайты могут быть разработаны путем изменения Th-эпитопа, который конъюгирован с B-клеточным эпитопом в пептидной иммуногенной конструкции, что может облегчить адаптацию персонализированного медицинского применения к индивидуальным характеристикам любого пациента или субъекта.
Пример 3
ИММУНОГЕННОСТЬ ПЕПТИДНЫХ ИММУНОГЕННЫХ КОНСТРУКЦИЙ МОЖЕТ ИЗМЕНЯТЬСЯ В ЗАВИСИМОСТИ ОТ ДЛИНЫ ПОСЛЕДОВАТЕЛЬНОСТИ В-КЛЕТОЧНОГО ЭПИТОПА
Иммунизация и оценка иммуногенных конструкций пептидов с тремя дипептидными повторами (DPR) подробно описаны ниже.
a. Иммунизация и сбор сыворотки
Были получены три пептидные иммуногенные конструкции DPR, имеющие аминокислотные последовательности SEQ ID NO: 236, 237 и 238, как продемонстрировано в Таблице 9. Каждый пептидный иммуноген составляли в MONTANIDE™ ISA51 и CpG для иммунизации морских свинок в дозе 400 мкг/ml в качестве первичной иммунизации и 100 мкг/ml в качестве бустерной дозы через 3, 6, 9 и 12 недель после инъекции (WPI), по 3 морские свинки на группу.
b. Оценка титров антител
ИФА выполняли для оценки иммуногенности созданных пептидных иммуногенных конструкций DPR. Пептиды В-клеточного эпитопа DPR или пептидные иммуногенные конструкции применяли для покрытия лунок планшета, которые служили нацеливающими пептидами. Иммунную сыворотку морской свинки разводили от 1:100 до 1:100 000 путем 10-кратных серийных разведений. Титр исследуемой сыворотки, выраженный как Log10, рассчитывали с помощью линейного регрессионного анализа A450 нм с порогом A450, установленным на 0,5. Все пептидные иммуногены индуцировали сильные титры иммуногенности против пептидов В-клеточного эпитопа, нанесенных в лунки планшета.
c. ИФА на основе пептидов для анализа специфичности антител
ИФА для оценки образцов иммунной сыворотки проводили, как описано ниже.
Лунки 96-луночных планшетов покрывали по отдельности в течение 1 часа при температуре 37°С 100 мкл той же пептидной иммуногенной конструкции DPR, применяемой для иммунизации животного (например, SEQ ID NO: 236, 237, или 238), в концентрации 2 мкг/мл в буфере 10 мМ NaHCO3, pH 9,5.
d. Оценка реактивности антител к DPR с помощью тестов ИФА.
Покрытые пептидом лунки инкубировали с 250 мкл 3% по массе желатина в PBS при 37°С в течение 1 часа , чтобы блокировать неспецифические сайты связывания белков, с последующими тремя промывками с применением PBS, содержащим 0,05% по объему TWEEN® 20 и сушили. Анализируемую сыворотку разбавляли 1:20 (если не указано иное) с применением PBS, содержащим 20% по объему нормальной козьей сыворотки, 1% по весу желатина и 0,05% по объему TWEEN® 20. Сто микролитров (100 мкл) разбавленных образцов (например, сыворотки, плазмы) добавляли в каждую из лунок и оставляли реагировать в течение 60 минут при 37°C. Затем лунки промывали шесть раз 0,05% по объему TWEEN® 20 в PBS, чтобы удалить несвязанные антитела. Специфические козьи анти-IgG, конъюгированные с пероксидазой хрена (HRP) (например, мыши, морской свинки или человека), применяли в качестве меченого индикатора для связывания с комплексом антитело/пептидный антиген, образованным в положительных лунках. Сто микролитров козьих анти-IgG, меченных пероксидазой, в предварительно заданном оптимальном разведении и в 1% по объему нормальной козьей сыворотки с 0,05% по объему TWEEN® 20 в PBS, добавляли в каждую лунку и инкубировали при 37°С в течение еще 30 минут. Лунки промывали шесть раз 0,05% по объему TWEEN® 20 в PBS для удаления несвязанного антитела и оставляли для реакции со 100 мкл смеси субстратов, содержащей 0,04% по массе 3’, 3’, 5’, 5’-тетраметилбензидин (TMB) и 0,12% по объему перекиси водорода в натрийцитратном буфере еще на 15 минут. Эту смесь субстратов применяли для обнаружения пероксидазной метки путем образования окрашенного продукта. Реакции останавливали добавлением 100 мкл 1,0 M H2SO4 и определяли оптическую плотность при 450 нм (A450). Для определения титров антител у иммунизированных животных, которые получали различные пептидные иммуногены, полученные из DPR, тестировали10-кратные серийные разведения сывороток от 1:100 до 1:10000, и титр тестируемой сыворотки, выраженный как Log10, рассчитівали с помощью линейного регрессионного анализа A450 с пороговым значением A450, установленным на 0,5.
e. Оценка иммуногенности
Преиммунные и иммунные образцы сыворотки от животных собирали в соответствии с экспериментальными протоколами иммунизации и нагревали при 56°С в течение 30 минут для инактивации факторов комплемента сыворотки. После введения фармацевтической композиции, содержащей пептидную иммуногенную конструкцию DPR, образцы крови получали в соответствии с протоколами и оценивали их иммуногенность против конкретного целевого сайта (ов). Тестировали серийно разведенные сыворотки, и положительные титры выражали как Log10 обратного разведения. Иммуногенность конкретной фармацевтической композиции оценивали по ее способности вызывать В-клеточный иммунный ответ с высоким титром, направленный против желаемой эпитопной специфичности в целевом антигене, при сохранении реактивности антител по отношению к Th-эпитопу от низкой до незначительной, используемого для обеспечения усиления желаемых ответов В-клеток.
f. Иммуноанализ на предмет уровня DPR в иммунной сыворотке мышей
Уровни DPR в сыворотке мышей, получавших пептидные иммуногены, полученные из DPR, измеряли с помощью сэндвич-ИФА (Cloud-clon, SEB222Mu) с применением антител анти-DPR в качестве захватывающего антитела и меченного биотином антитела анти-DPR в качестве антитела для обнаружения. Вкратце, антитело иммобилизовали на 96-луночных планшетах в концентрации 100 нг/лунку в покрывающем буфере (15 мМ Na2CO3, 35 мМ NaHCO3, pH 9,6) и инкубировали при 4°C в течение ночи. Покрытые лунки блокировали с применением раствора для разведения в количестве 200 мкл/лунку (0,5% BSA, 0,05% TWEEN®-20, 0,02% ProClin 300 в PBS) при комнатной температуре в течение 1 часа. Планшеты промывали 3 раза промывочным буфером 200 мкл/лунку (PBS с 0,05% TWEEN®-20). Очищенный рекомбинантный DPR применяли для построения стандартной кривой (диапазон от 156 до 1250 нг/мл путем 2-кратного серийного разведения) в растворе для разведения с 5% сывороткой мышей. Пятьдесят микролитров (50 мкл) разведенных сывороток (1:20) и стандарты добавляли в покрытые лунки. Инкубацию проводили при комнатной температуре в течение 1 часа. Все аспирировали и промывали 6 раз промывочным буфером в количестве 200 мкл. мкл/лунку. Захваченный DPR инкубировали со 100 мкл раствора антител для обнаружения (50 нг/мл меченного биотином HP6029 в растворе для разведения) при комнатной температуре в течение 1 часа. Затем связанный биотин-HP6029 обнаруживали с применением стрептавидина поли-HRP (разведение 1:10 000, Thermo Pierce) в течение 1 часа (100 мкл/лунку). Все лунки аспирировали и промывали 6 раз промывочным буфером 200 мкл/лунку и реакцию останавливали добавлением 1M H2SO4 100 мкл/лунку. Стандартную кривую генерировали с помощью программного обеспечения SoftMax Pro (Molecular Devices) для создания подгонки четырехпараметрической логистической кривой и применяли для расчета концентраций DPR во всех тестируемых образцах. Критерий Стьюдента применяли для сравнения данных с помощью программного обеспечения Prism.
g. Очистка антител анти-DPR
Антитела анти-DPR очищали из сывороток, собранных через 3-15 недель после инъекции (WPI) у морских свинок или мышей, иммунизированных пептидными иммуногенными конструкциями DPR, содержащими пептиды с различными последовательностями (SEQ ID NO: 236, 237 или 238), с применением аффинной колонки (Thermo Scientific, Rockford). Вкратце, после уравновешивания буфера (0,1 М фосфат и 0,15 М хлорид натрия, pH 7,2) 400 мкл сыворотки добавляли на колонку Nab Protein G Spin с последующим вращательным перемешиванием в течение 10 мин и центрифугированием при 5800 x g в течение 1 мин. Колонку промывали связывающим буфером (400 мкл) трижды. Затем в спин-колонку добавляли элюирующий буфер (400 мкл, 0,1 М глицина, pH 2,0) для элюирования антител после центрифугирования при 5800 x g в течение 1 мин. Элюированные антитела смешивали с буфером для нейтрализации (400 мкл, 0,1 М Трис, pH 8,0), и концентрации этих очищенных антител измеряли с помощью Nan-Drop при OD280 с BSA (бычий сывороточный альбумин) в качестве стандарта.
h. Результаты
Титр иммуногенности против пептидов DPR или пептидных иммуногенов из иммунизированной сыворотки морской свинки оценивали с помощью ИФА.
На Фигуре 7 продемонстрированы свойства антисыворотки в течение 15-недельного периода у морских свинок, иммунизированных 3 различными иммуногенными конструкциями пептида DPR. Антисыворотку морских свинок от 0, 3, 6, 9, 12 и 15 wpi разводили 10-кратным серийным разведением. Планшеты для ИФА покрывали пептидами DPR или пептидными иммуногенами. Титр исследуемой сыворотки, выраженный как Log10, рассчитывали с помощью линейного регрессионного анализа A450 нм с порогом A450, установленным на 0,5.
Данные ИФА для пептидных иммуногенных конструкций DPR, содержащих пептиды поли-GA (SEQ ID NO: 236, 237 и 238), затем наносили в виде графиков, продемонстрированных на Фигуре 7, где тот же пептидный иммуноген, который применяли для иммунизации животного, был связан с планшетом ИФА для анализа. Все иммуногенные конструкции DPR продемонстрировали высокую иммуногенность по отношению к соответствующим пептидам DPR или пептидным иммуногенам. Результаты ИФА показали, что в каждой группе до иммунизации на неделе 0 не наблюдалось определяемого титра антител. Полученные данные демонстрируют, что длина дипептидных повторов в пептидных иммуногенных конструкциях DPR может оказывать умеренное влияние на титры антител. В частности, конструкции с 10, 15 и 25 повторами поли-GA (SEQ ID NO: 236, 237 и 238 соответственно) обладают различной иммуногенностью по сравнению друг с другом, как продемонстрировано на Фигуре 7.
Представляет интерес тот факт, что длина пептида В-клеточного эпитопа может влиять на профиль иммуногенности пептидной иммуногенной конструкции. На Фигуре 7 (левый график) продемонстрировано, что иммуногенность пептидной иммуногенной конструкции GA10 (SEQ ID NO: 236) неуклонно возрастает с течением времени при регулярных бустерных дозах, тогда как на Фигуре 7 (средний график) продемонстрировано, что иммуногенность пептидной иммуногенной конструкции GA15 (SEQ ID NO: 237) имеет пики около 9 wpi перед снижением, а Фигура 7 (правый график) демонстрирует, что иммуногенностьпептидной иммуногенной конструкции GA25 (SEQ ID NO: 238) стабильно увеличивается с течением времени при регулярных бустерных дозах.
Результаты на Фигуре 7 демонстрируют, что иммунный ответ может зависеть от длины В-клеточного эпитопа, который применяется в пептидной иммуногенной конструкции.
i. Резюме
Результаты этого эксперимента демонстрируют, что иммунный ответ, вызываемый пептидными иммуногенными конструкциями (включая титры антител, Cmax, начало продукции антител, продолжительность ответа и т. д.), можно модулировать длиной В-клеточного эпитопа, применяемого в пептидной иммуногенной конструкции. Следовательно, специфические иммунные ответы на целевые антигенные сайты могут быть разработаны путем изменения длины В-клеточного эпитопа в пептидной иммуногенной конструкции, что может облегчить адаптацию персонализированного медицинского применения к индивидуальным характеристикам любого пациента или субъекта.
Пример 4
ИММУНОГЕННОСТЬ ПЕПТИДНЫХ ИММУНОГЕННЫХ КОНСТРУКЦИЙ МОЖЕТ ИЗМЕНИТЬСЯ В ЗАВИСИМОСТИ ОТ КОЛИЧЕСТВА ВВОДИМОГО ПЕПТИДА И СХЕМЫ ВВЕДЕНИЯ
Иммунизация и оценка различных доз и схемы введения пептидных иммуногенных конструкций подробно описаны ниже.
a. Вакцина UB-311 (иммуногены пептида Aβ)
Вакцина Aβ (UB-311) содержит два пептидных иммуногена, каждый с N-концевым пептидом Aβ1-14, синтетически связанным через аминокислотный спейсер с различными пептидами Th-клеточного эпитопа (эпитопами UBITh®), полученными из двух патогеных белков: поверхностного антигена гепатита B и слитого белка вируса кори. В частности, пептидный иммуноген, связанный со слитым белком вируса кори, представлял собой Aβ1-14-εK-KKK-MvF5 Th (SEQ ID NO: 67), а пептидный иммуноген, связанный с поверхностным антигеном гепатита B, представлял собой Aβ1-14-εK-HBsAg3 Th (SEQ ID NO: 68).
UB-311 был составлен в квасцы-содержащей Th2-смещенной системе доставки и содержал пептиды Aβ1-14-εK-HBsAg3 и Aβ1-14-εK-KKK-MvF5 Th в эквимолярном соотношении. Два иммуногена Aβ смешивали с полианионным олигодезоксинуклеотидом CpG (ODN) с образованием стабильных иммуностимулирующих комплексов из частиц микронного размера. Минеральную соль алюминия (ADJU-PHOS®) добавляли к конечному составу вместе с хлоридом натрия для придания тоничности и 0,25% 2-феноксиэтанолом в качестве консерванта.
b. Различные дозы UB-311 для морских свинок.
Вакцину Aβ (UB-311) вводили морским свинкам в моменты времени 0, 3 и 6 wpi в дозах, содержащих в общей сложности 0 мкг, 1 мкг, 3мкг, 10мкг, 30мкг, 100мкг, 300мкг, 600 мкг и 1000 мкг пептидных иммуногенных конструкций. Образцы сыворотки отбирали в моменты времени 0, 3, 5 и 7 wpi для оценки уровней титра антител.
Фигура 8 представляет собой график, на котором приведены результаты титров антител, полученные для каждой дозы иммунизации. Полученные результаты демонстрируют, что количество пептида, применяемого при иммунизации, может оказывать заметное влияние на титры антител; однако оптимальный технический эффект следует оценивать для каждой дозы отдельно. В частности, Фигура 8 демонстрирует, что увеличение дозы вводимого пептида с 0 мкг до 600 мкг напрямую соответствует повышению иммуногенности. Однако, когда вводили 1000 мкг пептидного иммуногена, иммуногенность фактически могла снижаться по сравнению с более низкими дозами пептидной иммуногенной конструкций (по сравнению с 300 мкг и 600 мкг с 1000 мкг). Следовательно, оптимальная иммуногенность, полученная с помощью пептидной иммуногенной конструкций, не является линейной, и ее следует тщательно оценивать для каждой пептидной иммуногенной конструкций отдельно.
c. Различные схемы введения UB-311 для морских свинок могут влиять на иммуногенность.
Схемы введения вакцины Aβ (UB-311) оценивали для того, чтобы определить, может ли количество пептидной иммуногенной конструкции, вводимой в качестве прайм-дозы и бустер-дозы, влиять на общую иммуногенность композиции.
Вакцину UB-311 вводили морским свинкам в моменты времени 0, 3 и 6 wpi с различными вводимыми прайм- и бустер-дозами. В частности, одна группа животных получала прайм-дозу 100 мкг UB-311 в момент времени неделя 0 wpi и 2 бустер-дозы по 400 мкг UB-311; тогда как вторая группа животных получала прайм-дозу 400 мкг UB-311 в момент времени неделя 0 wpi и 2 бустер-дозы по 100 мкг UB-311. Результаты этого эксперимента приведены на Фигуре 9.
На Фигуре 9 продемонстрировано, что схема введения может влиять на иммуногенность (Cmax и продолжительность) композиции UB-311. В частности, животные, которые получали низкую дозу в качестве прайм-введения (100 мкг композиции UB-311) и высокую дозу в качестве буст-введения (400 мкг композиции UB-311), достигали более высокого и длительного показателя Cmax по сравнению с животными, которые получали высокую дозу в качестве прайм-введения (400 мкг композиции UB-311) и более низкую дозу в качестве буст-введения (100 мкг композиции UB-311).
Результаты этого исследования демонстрируют, что схема введения может влиять на иммуногенность пептидных иммуногенных конструкций.
На Фигуре 10 представлен дополнительный пример того, что схема введения может влиять на иммуногенность пептидных иммуногенных конструкций. В частности, Фигура 10 (верхняя панель) демонстрирует титры антител анти-Aβ1-28 с помощью ИФА, полученные после иммунизации людей вакциной Aβ в количестве 300 мкг (UB-311) в течение трехмесячной схемы бустерной вакцинации (верхняя панель) или шестимесячноой схемы бустерной вакцинации (нижняя панель) с применением Aβ вакцины (UB-311). В рамке на каждом графике выделены средние титры для всех субъектов-людей, участвовавших в исследовании.
Результаты на Фигуре 10 демонстрируют, что, в зависимости от схемы введения, применяемой для субъектов, может быть достигнут другой профиль иммуногенности.
d. Различные схемы введения пептидных иммуногенных конструкций LHRH у крыс.
Схема введения составов, содержащих различные количества пептидных иммуногенных конструкций LHRH, оценивали для определения того, может ли общее количество пептидной иммуногенной конструкции влиять на иммуногенность составов.
В частности, трех крыс иммунизировали композицией LHRH, содержащей 3 пептида, продемонстрированные в Таблице 10. Одну группу крыс иммунизировали 3 пептидными иммуногенами LHRH в дозе 100 мкг; тогда как вторую группу крыс иммунизировали 3 пептидными иммуногенами LHRH в дозе 300 мкг. Оценивали иммуногенность и концентрации тестостерона, что представлено на Фигурах 11A и 11B.
На Фигуре 11A продемонстрированы титры антител и концентрации тестостерона, полученные после иммунизации крыс составом LHRH в дозе 100 мкг. На Фигуре 11В продемонстрированы титры антител и концентрации тестостерона, полученные после иммунизации крыс составом LHRH в дозе 300 мкг. Фигуры 11A и 11B демонстрируют, что для пептидных иммуногенных конструкций LHRH более высокая доза в 300 мкг приводит к более высоким титрам анти-LHRH и более высокому снижению концентраций тестостерона по сравнению с более низкой дозой пептидных иммуногенных конструкций LHRH.
Таким образом, результаты на Фигурах 11A и 11B демонстрируют, что существует прямая корреляция между уровнями дозы пептидных иммуногенных конструкций и достигнутым техническим эффектом.
e. Резюме
Результаты этого эксперимента демонстрируют, что иммунный ответ, вызываемый пептидными иммуногенными конструкциями (включая титры антител, Cmax, начало продукции антител, продолжительность ответа и т. д.), можно модулировать с помощью применения различных схем введения. Следовательно, специфические иммунные ответы на целевые антигенные сайты могут быть разработаны путем изменения схемы введения для пациента или субъекта, что может облегчить адаптацию персонализированного терапевтического лечения к индивидуальным характеристикам любого пациента или субъекта.
Пример 5
ИММУНОГЕННОСТЬ ПЕПТИДНЫХ ИММУНОГЕННЫХ КОНСТРУКЦИЙ МОЖЕТ ИЗМЕНЯТЬСЯ В ЗАВИСИМОСТИ ОТ ПРИМЕНЯЕМОГО АДЪЮВАНТА
Иммунизацию и оценку пептидных иммуногенных конструкций IL-6, IgE EMPD и LHRH, составленных в различных адъювантах, оценивали, как описано ниже.
a. Пептидные иммуногенные конструкции IL-6
Оценивали иммуногенность составов, содержащих пептидную иммуногенную конструкцию IL-6 с применением различных адъювантов. Исследование POC на крысах CIA продемонстрировало, что разработанные пептидные иммуногенные конструкции с высокой иммуногенностью и терапевтической эффективностью, направленной на IL-6-индуцированный патогенез, предполагают потенциальное иммунотерапевтическое применение при ревматоидном артрите и других аутоиммунных заболеваниях. Последующие исследования были сосредоточены на оптимизации пептидных иммуногенных конструкций и выборе адъювантов, а также на определении дозы у крыс CIA Lewis.
MONTANIDE ISA 51 и ADJU-PHOS как разные адъюванты, содержащие один и тот же пептидный иммуноген (SEQ ID NO: 243) плюс CpG, соответственно, оценивали в исследовании иммунизации крыс CIA. Пять крыс, определенных в каждую из 5 групп, получали один из двух составов адъюванта, в общей сложности 10 групп для этих двух различных адъювантов. Всем животным в группах лечения внутримышечно вводили разные дозы 5, 15, 45, 150 мкг в 0,5 мл в прайм- и буст-режиме в день -7, 7, 14, 21 и 28 с клиническим наблюдением до дня 35. Две разные группы плацебо с адъювантом без пептидного иммуногена получали инъекции только с адъювантами-носителями в составе.
Титр анти-IL-6 измеряли с помощью ИФА относительно рекомбинантного белка IL-6 крысы, нанесенного в лунки планшета. Полученные результаты продемонстрировали, что ни в одной из двух групп плацебо, которым вводили два разных адъюванта-носителя, не было обнаружено определяемых титров антител анти-IL-6, в то время как все группы лечения, иммунизированные иммуногенной конструкцией IL-6 (SEQ ID NO: 243) с обоими адъювантными составами, генерировали антитела против крысиного IL-6 в ИФА. В целом результат продемонстрировал, что наблюдалась зависимость от дозы, особенно для групп с составом ISA 51 (Фигура 12). Состав ISA 51 индуцировал более высокий иммунный ответ, чем состав ADJUPHOS, у иммунизированных крыс, при этом иммуногенность составов ISA 51 имела значения Log10 более 4 для всех доз, соответственно, по сравнению с иммуногенностью ниже 4 (log10) для составов, приготовленных с адъювантом ADJUPHOS. .
Фигура 12 иллюстрирует кинетику ответа антител в течение 43-дневного периода у крыс, иммунизированных различными дозами SEQ ID NO: 243, приготовленными с применением либо ISA 51/CpG, либоr ADJU-PHOS/CpG. Планшеты для ИФА покрывали рекомбинантным крысиным IL-6. Сыворотку разводили от 1:100 до 1:4,19×108 путем 4-кратных серийных разведений. Титр исследуемой сыворотки, выраженный как Log10 рассчитывали путем включения порогового значения 0,45 в четырехпараметрическую логистическую кривую для каждого образца сыворотки с нелинейной регрессией.
Результаты этого эксперимента демонстрируют, что выбор адъюванта может иметь значительное техническое влияние на иммуногенность пептидной иммуногенной конструкции.
b. Пептидные иммуногенные конструкции IgE EMPD
Оценивали иммуногенность составов, содержащих пептидную иммуногенную конструкцию IgE EMPD с применением различных адъювантов. Исследование POC на макаках продемонстрировало, что разработанные пептидные иммуногенные конструкции с высокой иммуногенностью и терапевтической эффективностью, направленной на IgE EMPD-индуцированный патогенез, предполагают потенциальное иммунотерапевтическое применение. Последующие исследования были сосредоточены на оптимизации пептидных иммуногенных конструкций и выборе адъювантов с применением пептидных иммуногенных конструкций IgE EMPD.
ADJU-PHOS и MONTANIDE ISA 51 как разные адъюванты, содержащие один и тот же пептидный иммуноген IgE EMPD (SEQ ID NO: 178) плюс CpG, соответственно, оценивали в исследовании иммунизации макак.
На Фигурах 13A и 13B приведены графики, иллюстрирующие титры антител анти-IgE-EMPD, полученные после иммунизации макак различными количествами пептидной иммуногенной конструкции IgE-EMPD SEQ ID NO: 178 в различных адъювантах. На Фигуре 13A продемонстрированы титры антител, полученные с применением ADJUPHOS в качестве адъюванта, составленного в виде стабилизированного иммуностимулирующего комплекса с применением CpG3; тогда как на Фигуре 13B продемонстрированы титры антител, полученные с применением MONTANIDE ISA51 в качестве адъюванта, составленного в виде стабилизированного иммуностимулирующего комплекса с применением CpG3.
Результаты этих экспериментов демонстрируют, что пептидные иммуногенные конструкции IgE EMPD, составленные в различных адъювантах, являются более иммуногенными при применении с MONTANTIDE ISA51 по сравнению с составами, содержащими ADJUPHOS (Фигуры 13A и 13B). Следовательно, применение различных адъювантов может обуславливать различный технический эффект (иммуногенность) пептидных иммуногенных конструкций.
c. Пептидные иммуногенные конструкции LHRH
Оценивали иммуногенность составов, содержащих пептидные иммуногенные конструкции LHRHс применением различных адъювантов. Исследование POC у свиней продемонстрировало, что разработанные пептидные иммуногенные конструкции с высокой иммуногенностью и терапевтической эффективностью, направленной на LHRH-индуцированный патогенез, предполагают потенциальное иммунотерапевтическое применение. Последующие исследования были сосредоточены на оптимизации пептидных иммуногенных конструкций и выборе адъювантов с применением пептидных иммуногенных конструкций LHRH.
Emulsigen D и MONTANIDE ISA50V в качестве различных адъювантов, содержащих одинаковые пептидные иммуногены LHRH (SEQ ID NO: 239-241) при одинаковой концентрации, оценивали в исследовании иммунизации свиней.
На Фигурах 14A и 14B приведены графики, иллюстрирующие титры антител анти-LHRH, полученные после иммунизации свиней различными количествами пептидной иммуногенной конструкции LHRH SEQ ID NO: 239-241 в различных адъювантах. На Фигуре 14A продемонстрированы титры антител, полученные с применением Emulsigen D в качестве адъюванта; тогда как на Фигуре 14B продемонстрированы титры антител, полученные с применением MONTANIDE ISA50V в качестве адъюванта.
Результаты этих экспериментов демонстрируют, что пептидные иммуногенные конструкции LHRH, составленные в различных адъювантах, имеют разный технический эффект (снижение концентрации тестостерона) при составлении с MONTANTIDE ISA50V по сравнению с составами, содержащими Emulsigen D (Фигуры 15A и 14B). Следовательно, применение различных адъювантов может обуславливать различный технический эффект (продолжительность эффекта, то есть иммунокастрацию в данном случае) пептидных иммуногенных конструкций.
d. Резюме
Результаты этого эксперимента демонстрируют, что иммунный ответ, вызываемый пептидными иммуногенными конструкциями (включая титры антител, Cmax, начало продукции антител, продолжительность ответа и т. д.), можно модулировать путем выбора адъюванта, применяемого в составе, содержащем пептидные иммуногенные конструкции в той же концентрации. Следовательно, специфические иммунные ответы на целевые антигенные сайты могут быть разработаны путем либо изменения Th-эпитопа, который химически связан с B-клеточнм эпитопом, либо адъюванта, применяемого в составе, что может облегчить адаптацию персонализированного медицинского применения к индивидуальным характеристикам любого пациента или субъекта.
ПРИМЕР 6
Исключительная иммуногенность пептидных иммуногенных конструкций у морских свинок, которые нацелены на пептиды Aβeta, но не на Th-эпитопы.
Шесть морских свинок были иммунизированы на неделях 0 и 4 пептидными иммуногенными конструкциями Aβ1-14-εK-KKK-MvF5 (SEQ ID NO: 67) и Aβ1-14-εK-HBsAg3 (SEQ ID NO: 68), составленными вместе в эквимолярном соотношении. На неделе 8 у животных брали кровь и собирали образцы сыворотки для определения титров антител против пептида Aβ и против Th-эпитопа (log10) с помощью ИФА. Гуморальный иммунный ответ всех 6 морских свинок специфически нацеливался на пептид Aβ1-42, а не на два искусственных Th-эпитопа (MvF5 Th и HBsAg3 Th), как продемонстрировано в Таблице 11.
ПРИМЕР 7
Клеточный иммунный ответ в культурах мононуклеарных клеток периферической крови (PBMC) павианов и макак, иммунизированных вакциной UB-311
Мононуклеарные клетки периферической крови (PBMC) павианов и макак Cynomolgus, иммунизированных UB-311, выделяли центрифугированием в градиенте фиколла-гипака. Для индуцированной пептидом пролиферации и продукции цитокинов, клетки (2×105 на лунку) культивировали отдельно или с добавлением отдельных пептидных доменов (включая Aβ1-14, Aβ1-42, UBITh® и нерелевантный пептид). Митогены (PHA, PWM, Con A) применяли в качестве положительного контроля (10 мкг/мл при 1% об./об. культуры). На день 6 добавляли 1 мкКи 3H-тимидина (3H-TdR) в каждую из трех лунок повторной культуры. После 18 ч инкубации клетки собирали и определяли включение 3H-TdR. Индекс стимуляции (S.I.) представляет собой количество импульсов (англ.: cpm) в минуту в присутствии антигена, деленное на количество импульсов в минуту в отсутствие антигена; S.I.> 3,0 считался значимым.
Анализы цитокинов (IL-2, IL-6, IL-10, IL-13, TNFα, IFNγ) из культур PMBC макака Cynomolgus выполняли только на аликвотах культуральной среды или в присутствии пептидных доменов или митогенов. Наборы сэндвич-ИФА для цитокинов, специфичных для обезьян (U-CyTech Biosciences, Утрехт, Нидерланды), применяли для определения концентрации отдельных цитокинов, следуя инструкциям набора.
PBMC выделяли из цельной крови, взятой у макак на неделях 15, 21 и 25,5. Выделенные PBMC культивировали в присутствии различных пептидов Aβ (Aβ1-14 и Aβ1-42).
При добавлении пептида Aβ1-14 к культуральной среде никаких реакций пролиферации лимфоцитов не наблюдалось. Однако при добавлении пептида Aβ1-42 к культурам PBMC были обнаружены положительные реакции пролиферации.
Образцы PBMC, взятые на неделе 15, 21 и 25,5, также тестировали на секрецию цитокинов в присутствии пептидов Aβ или митогена PHA. Как продемонстрировано в Таблице 12, три цитокина (IL-2, IL-6, TNFα) показали детектируемую секрецию в ответ на полноразмерный пептид Aβ1-42, но не на пептид Aβ1-14; усиление секреции цитокинов не было обнаружено в образцах, обработанных вакциной UBITh® AD, по сравнению с образцами, обработанными плацебо-вакциной. Три других цитокина (IL-10, IL-13, IFNγ), протестированные в присутствии пептидов Aβ, были ниже предела обнаружения анализа во всех культурах PBMC.
Макаки были иммунизированы вакциной UB-311, имеющей только N-концевые Aβ1-14-пептидные иммуногены с чужеродными Т-хелперными эпитопами, без пептидного домена Aβ17-42, что указывает на положительные результаты пролиферации, отмеченные в культурах PBMC в присутствии пептида Aβ1-42, не были связаны с ответом на вакцину UB-311 , а скорее являлись фоновым ответом на нативный полноразмерный Aβ.
Эти результаты подтверждают безопасность вакцины UB-311 , которая содержит только Aβ1-14 и чужеродные Т-хелперные эпитопы, показывая, что она не вызывает потенциально воспалительных клеточно-опосредованных иммунных ответов против аутоантигенов на нативные полноразмерные пептиды Aβ у здоровых макак. Напротив, нежелательные явления, ассоциированные с энцефалитом, в клинических исследованиях вакцины AN-1792 были отчасти приписаны включению Т-клеточных эпитопов в мономерный или фибриллярный/агрегированный иммуноген Aβ1-42 этой вакцины.
ПРИМЕР 8
Анализ пролиферации лимфоцитов и цитокиновый анализ PBMC от пациентов с болезнью Альцгеймера, иммунизированных вакциной UB311.
Мононуклеарные клетки периферической крови (PBMC) пациентов с болезнью Альцгеймера выделяли центрифугированием в градиенте фиколла-гипака. Для индуцированной пептидом пролиферации и продукции цитокинов, клетки (2,5×105 на лунку) культивировали в трех повторностях отдельно или с добавлением отдельных пептидных доменов (в конечной концентрации 10 мкг/мл), включая Aβ1-14 (SEQ ID NO: 56), Aβ1-16 (SEQ ID NO: 57), Aβ1-28 (SEQ ID NO: 59), Aβ17-42 (SEQ ID NO: 58), Aβ1-42 (SEQ ID NO: 60) и нерелевантный 38-мерный пептид (p1412). Культуры инкубировали при 37°C с 5% CO2 в течение 72 часов, а затем из каждой лунки удаляли 100 мкл супернатанта и замораживали при -70°C для анализа цитокинов. Десять мкл культуральной среды, содержащей 0,5 мкКи 3H-тимидина (3H-TdR, Amersham, № по кат. TRK637) добавляли в каждую лунку и инкубировали в течение 18 часов с последующим обнаружением включения радиоизотопа с помощью жидкостной сцинтилляционной детекции. Митоген фитогемагглютинин (PHA) применяли в качестве положительного контроля пролиферации лимфоцитов. Клетки, культивированные отдельно без пептида Aβ или митогена PHA, применяли в качестве отрицательного и положительного контролей. Индекс стимуляции (SI) рассчитывали как среднее количество импульсов в минуту (cpm) для трех повторностей экспериментальных культур с пептидом Aβ, деленное на среднее количество импульсов в минуту для трех повторностей культур отрицательного контроля; SI > 3,0 рассматривался как значительный ответ пролиферации.
a. Анализ пролиферации
Образцы мононуклеарных клеток периферической крови выделяли из цельной крови, взятой на 0-й неделе (исходный уровень) и 16-й неделе (через 4 недели после третьей дозы) от пациентов с болезнью Альцгеймера, вакцинированных вакциной UB-311, а затем культивировали в отсутствии или в присутствии различных пептидов Aβ. Как продемонстрировано в Таблице 13, при добавлении в культуральную среду Aβ1-14, других пептидов Aβ или p1412 (нерелевантный контрольный пептид) значительного пролиферативного ответа лимфоцитов не наблюдалось. Как и ожидалось, при добавлении митогена PHA к культуральной среде отмечались положительные пролиферативные ответы. Наличие аналогичных ответов на PHA до и после иммунизации UB-311 (p=0,87) предполагает отсутствие значительного изменения иммунных функций исследуемых субъектов (Таблица 13).
Статистический анализ. Различия в пролиферации лимфоцитов между неделями 0 и 16 исследовали с помощью парного t-критерия. Уровни статистической значимости определяли с помощью двусторонних кртитериев (p < 0,05). Для всех статистических анализов применяли R версию 2.14.1.
b. Анализ цитокинов
Анализы цитокинов (IL-2, IL-6, IL-10, TNFα, IFNγ) из культур PBMC выполняли только на аликвотах культуральной среды или в присутствии доменов пептида Aβ или РНА. С целью определения концентраций (пг/мл) отдельных цитокинов применяли наборы сэндвич-ИФА для цитокинов человека (U-CyTech Biosciences, Утрехт, Нидерланды) в соответствии с инструкциями производителя (Clin Diag Lab Immunol. 5(1):78-81 (1998)).
Образцы PBMC, взятые у пациентов с болезнью Альцгеймера, получавших вакцину UB-311, на неделе 0 и 16, также тестировали на секрецию цитокинов либо только с клетками (отрицательный контроль), либо в присутствии пептидов Aβ, p1412 (нерелевантный пептид) или митогена PHA (положительный контроль) после культивирования в течение 3 дней. Поддающийся количественной оценке диапазон набора составляет от 5 до 320 пг/мл. Любая измеренная концентрация ниже 5 пг/мл или выше 320 пг/мл была обозначена как ниже предела количественного определения (BQL -below quantification limit) или выше предела количественного определения (AQL -above quantification limit), соответственно. Однако по статистическим соображениям BQL или AQL были заменены нижним (5 пг/мл) или верхним (320 пг/мл) количественным пределом, соответственно. Средние концентрации каждого цитокина на неделях 0 и 16 приведены в Таблице 14. Как и ожидалось, отмечалось значительное увеличение продукции цитокинов в присутствии PHA, положительного контроля, за исключением IL-2. Продукция цитокинов в ответ на стимуляцию Aβ1-14 или другими пептидами Aβ наблюдалась на исходном уровне (неделя 0) и на неделе 16, но большинство значений оказались сходными с соответствующими отрицательными контролями (только клетки).
Чтобы оценить изменение клеточно-опосредованного иммунного ответа после иммунизации, сравнивали изменение средних концентраций цитокинов от исходного уровня до недели 16 с таковым для отрицательных контролей и исследовали с помощью парного критерия знаковых рангов Уилкоксона. Четыре цитокина (IFN-γ, IL-6, IL-10, TNF-α) продемонстрировали заметное увеличение секреции в ответ на полноразмерный пептид Aβ1-42; это наблюдение может быть связано с конформационными эпитопами агрегатов Aβ1-42 . Усиление секреции цитокинов не было обнаружено в ответ на Aβ1-14 или другие пептиды Aβ.
c. Резюме
Вакцина UB-311 содержит два пептидных иммуногена, каждый с N-концевым пептидом Aβ1-14, синтетически связанным с эпитопами Th MvF5 и Th HBsAg3, соответственно. Данные анализа пролиферации лимфоцитов и цитокинов in vitro применяли для оценки влияния иммунизации вакциной UB-311 на клеточный иммунный ответ. При добавлении в культуральную среду пептида Aβ1-14 или любых других пептидов Aβ не наблюдалось реакции пролиферации лимфоцитов, как продемонстрировано в Таблице 13. Усиление секреции цитокинов лимфоцитами пациентов, иммунизированных вакциной UB-311, не была обнаружено при лечении Aβ1-14 и другими пептидами Aβ, за исключением Aβ1-42, который вызывал заметное увеличение четырех цитокинов (IFN-γ, IL-6, IL-10, TNF-α) после иммунизации UB-311 на неделе 16 по сравнению с уровнями на неделе 0 до лечения (Таблица 14). Увеличение высвобождения цитокинов за счет Т-клеточного ответа Th2-типа, скорее всего, не связано с ответом на вакцину UB-311, поскольку усиление регуляции только с Aβ1-14 не обнаружено. Предполагается, что ответ на Aβ1-42 является фоновым ответом на нативный Aβ, который может быть связан с нативными Т-хелперными эпитопами, идентифицированными на Aβ1-42. Наблюдалось отсутствие продукции IL-2 в ответ на PHA, что согласуется с результатами, полученными Katial RK, et al. в Clin Diagn Lab Immunol 1998; 5:78-81, в аналогичных экспериментальных условиях с нормальными РВМС человека. В заключение, эти результаты продемонстрировали, что вакцина UB-311 не вызвала потенциально воспалительных клеточно-опосредованных иммунных ответов против аутоантигенов у пациентов с болезнью Альцгеймера от легкой до умеренной степени, которые участвовали в клинических исследованиях фазы I, что дополнительно демонстрирует безопасность вакцины UB-311.
ПРИМЕР 9
НЕИЗБИРАТЕЛЬНЫЕ ИСКУССТВЕННЫЕ КЛЕТКИ, ОТВЕЧАЮЩИЕ НА Th, МОГУТ БЫТЬ ОБНАРУЖЕНЫ В НАИВНЫХ МОНОНУКЛЕАРНЫХ КЛЕТКАХ ПЕРИФЕРИЧЕСКОЙ КРОВИ (PMBC) У ЗДОРОВЫХ ДОНОРОВ КРОВИ С УМЕРЕННЫМ ИММУНОГЕННЫМ ВОСПАЛИТЕЛЬНЫМ ОТВЕТОМ ПО СРАВНЕНИЮ С ОТРИЦАТЕЛЬНЫМ КОНТРОЛЕМ
Анализ ELISpot применяли для обнаружения неизбирательных искусственных клеток, отвечающих на Th, в наивных мононуклеарных клетках периферической крови у здоровых доноров крови для оценки их способности вызывать воспалительные ответы по сравнению с мощным митогеном фитогемагглютинином (PHA) и отрицательным контролем.
В анализах ELISpot применяли методику твердофазного иммуноферментного анализа (ИФА). Для обнаружения активации Т-клеток в качестве аналита определяли IFN-γ или родственный цитокин. Либо моноклональные, либо поликлональные антитела, специфичные для выбранного аналита, предварительно наносили на микропланшет с PVDF (поливинилидендифторидом). Соответствующим образом стимулированные клетки вносили пипеткой в лунки, и микропланшет помещали в инкубатор при 37°C в увлажненной атмосфере CO2 на определенный период времени. В течение этого периода инкубации иммобилизованное антитело в непосредственной близости от секретирующих клеток связывается с секретируемым аналитом. После отмывания любых клеток и несвязавшихся веществ в лунки добавляли биотинилированное поликлональное антитело, специфичное для выбранного аналита. После отмывания для удаления любого несвязанного биотинилированного антитела добавляли щелочную фосфатазу, конъюгированную со стрептавидином. Затем несвязанный фермент удаляли отмыванием и добавляли раствор субстрата (BCIP/NBT). Образовывался сине-черный осадок, который проявлялся в виде пятен в местах локализации цитокинов, причем каждое отдельное пятно представляет собой отдельную клетку, секретирующую аналит. Пятна подсчитывали с помощью автоматизированной системы считывания ELISpot или вручную с помощью стереомикроскопа.
В проведенном in vitro исследовании в качестве положительного контроля применяли PHA в концентрации 10 мкг/мл. Пептиды UBITh®1 (SEQ ID NO: 17) и UBITh®5 (SEQ ID NO: 6) тестировали на количество отвечающих клеток, присутствующих в мононуклеарных клетках периферической крови у обычных здоровых доноров крови. Смесь неизбирательных искусственных пептидов Th-эпитопа с SEQ ID NO: 33-52 получали в качестве другого положительного контроля. Среду применяли отдельно в качестве отрицательного контроля в стандартных условиях культивирования клеток для стимуляции Т-клеток. Вкратце, 100 мкл/лунку PBMC (2×105 клеток), стимулированных митогеном (PHA при 10 мкг/мл) или Th-антигеном (UBITh®1, UBITh®5 или смесь мульти-Ths при 10 мкг/мл), инкубировали при 37°C в инкубаторе с CO2 на протяжении 48 часов. Из лунок/планшетов собирали супернатант. Клетки в планшетах отмывали и обрабатывали для обнаружения целевого аналита IFN-γ.
Как продемонстрировано на Фигуре 15, репрезентативные доноры 1, 2 и 3 были протестированы на предмет их клеток, отвечающих на пептиды неизбирательных искусственных эпитопов UBITh®1 или UBITh®5. Подавляющее количество IFN-γ по ELISPOT всегда выявлялось наивным донором (PBMC, культивированные с PHA; слишком много для подсчета), тогда как PBMC, культивированные в контрольной среде, давали фоновое количество IFN-γ по ELISPOT от 5 до 50. Умеренные количества по ELISPOT были обнаружены для наивных донорных PBMC, культивированных с UBITh®1 или UBITh®5, от 20 до около 120. Смесь мульти-Th-пептидов с SEQ ID NO: 33-52 также культивировали с наивными донорными PBMC для сравнения количеством по ELISPOT от 20 до около 300, как ожидалось. Такие ответы на стимуляцию, вызванные пептидами UBITh®1 или UBITh®5, примерно в 3-5 раз превышают показатели отрицательного контроля.
Таким образом, неизбирательные искусственные клетки, отвечающие на Th, могут быть легко обнаружены в наивных донорных PBMC, которые готовы к развитию иммунных ответов, чтобы помочь продукции B-клеточных антител и соответствующим эффекторным T-клеточным ответам путем секретирования сигнатурных цитокинов. IFN-γ был использован в качестве одного примера, чтобы проиллюстрировать такую стимулирующую природу этих пептидов Th-эпитопа. Однако такие стимулирующие воспалительные ответы являются достаточно умеренными, чтобы вызвать подходящие ответы эффекторных клеток (В-клеток для продукции антител, цитотоксических Т-клеток для уничтожения целевых антигенных клеток) так, чтобы не вызывать нежелательные воспалительные патофизиологические ответы во время процесса вакцинации.
ПРИМЕР 10
ПЕПТИДНЫЕ ИММУНОГЕННЫЕ КОНСТРУКЦИИ НА ОСНОВЕ НЕОЭПИТОПА И ИХ СОСТАВЫ ДЛЯ ПЕРСОНАЛИЗИРОВАННОЙ ИММУНОТЕРАПИИ РАКА
Мы находимся в разгаре Т-клеточной революции в лечении рака. За последние несколько лет новые методы лечения, такие как ингибиторы иммунных контрольных точек (ICI - immune checkpoint inhibitors) и адоптивная клеточная терапия, например, химерные антигенные рецепторы (CAR-T), дали новую надежду миллионам людей, страдающих раком. Препараты ICI помогают преодолевать естественные барьеры, обусловленные раком и заключающиеся в том, что наша собственная иммунная система и, в частности, наши Т-клетки не могли бороться с раком. Например, терапия CAR-T конструирует Т-клетки пациента, чтобы вызвать иммунный ответ против определенных опухолевых клеток. Эти новые методы лечения представляют собой методы лечения будущего, разработанные для «обучения» Т-лимфоцитов атаковать опухоли с еще большей точностью.
Растет интерес относительно персонализированных или неоантигенных вакцин против рака как способа дать новую надежду миллионам людей, страдающих от рака. Неоантигены представляют собой персонализированные опухолевые мутации, которые иммунная система считает чужеродными. Таким образом, персонализированная вакцина нацелена на эти неоантигены, которые обучают иммунную систему находить и уничтожать опухоль.
Для эффективной идентификации истинных неоантигенов широко применялись биоинформатические, протеомные, клеточные анализы и нестандартные методы. Варианты РНК со сдвигом рамки считывания (FS), образованные INDEL, то есть короткие вставки и удаления в микросателлитах и неправильный сплайсинг экзонов, являются богатым источником высокоиммуногенных неоантигенов.
Матрицы, содержащие все возможные (например, 400K) пептиды, полученные из опухоли FS (сдвиг рамки считывания), можно применять для обнаружения реактивности антител из одной капли крови пациента, что обеспечивает замечательный инструмент для диагностики рака и идентификации CTL-эпитопов неоантигена.
Типирование MHC может быть выполнено с помощью RNAseq, в то время как NetMHC и NetMHCpan можно применять для прогнозирования аффинности связывания неоэнтигена.
В независимом от HLA анализе in vitro полипептиды, представляющие каждую идентифицированную мутацию опухоли пациента, доставляются индивидуально в их собственные антигенпрезентирующие клетки (APC), которые затем обрабатываются и представляют пептиды на поверхности клетки, где они распознаются различными эффекторными Т-клетками. Если Т-клетка распознает пептид и связывается с ним, будет запускаться цитокиновый ответ. Истинный антиген измеряется и определяется как «хороший» (то есть стимулирующий) или «плохой» (то есть ингибирующий). Фактические антигены, на которые отвечают Т-клетки пациента - как CD4⁺ (Т-хелперы), так и CD8⁺ (Т-клетки-киллеры), могут быть идентифицированы и выбраны в качестве неоэпитопа для создания неоэпитопных пептидных иммуногенных конструкций.
Таким образом, путем включения эмпирически подтвержденных неоантигенов, на которые пациенты уже имеют ответы, разрабатывается персонализированная противораковая вакцина, к которой иммунная система пациентов уже праймирована.
Вкратце, неоэпитопный пептид, производный от конкретного эмпирически подтвержденного неоантигена, включающего неоэпитоп B или CTL, можно сделать высокоиммуногенным с применением принципов конструирования, описанных в данном документе При участии неизбирательных искусственных Th-эпитопов, ковалентно связанных с выбранным неоэпитопом, такая иммуногенная конструкция неоэпитопного пептида может способствовать индукции и поддержанию антител, направленных на неоантигены, и индукции эффекторных CTL функций, а также генерации B- и CTL- клеток памяти, что приводит к устойчивым B- и CTL- ответам и устойчивому противоопухолевому иммунитету.
Репрезентативные общедоступные неоантигены с выбранными последовательностями целевых эпитопов для неоантигенов меланомы, вариант Histone3 H3.3K27M для глиомы и KRAS (мутант с G на D) для колоректального рака, продемонстрированы как SEQ ID NO: 73, 74 и 75, соответственно, в Таблице 3А.
Разработанные неоэпитопные пептидные иммуногенные конструкции и их составы по данному изобретению могут быть дополнительно оценены в клинических исследованиях на предмет безопасности, иммуногенности и эффективности, которые состоят из трех частей:
1) Исследование безопасности и иммуногенности при монотерапии у онкологических пациентов без признаков заболевания, но с высоким риском рецидива.
2) Исследование безопасности, иммуногенности и эффективности неоэпитопной вакцины в комбинации с одобренным FDA ингибитором иммунных контрольных точек у пациентов с распространенными или метастатическими солидными опухолями.
3) Исследование безопасности, иммуногенности и эффективности неоэпитопной вакцины при монотерапии у пациентов с рецидивирующими или рефрактерными солидными опухолями, которые не ответили на лечение или у которых рак прогрессировал после лечения ингибитором иммунных контрольных точек.
Соответствующие критериям пациенты, завершившие лечение (например, хирургическая резекция, неоадъювантная и/или адъювантная химиотерапия и/или лучевая терапия) меланомы кожи, немелкоклеточного рака легкого (NSCLC), плоскоклеточного рака головы и шеи (SCCHN), или уротелиальной карциномы и не демонстрирующие признаков заболевания при КТ или МРТ, могут получать эту иммунотерапию против рака.
ПРИМЕР 11
АССОЦИИРОВАННЫЕ С ОПУХОЛЬЮ УГЛЕВОДНЫЕ АНТИГЕНЫ (TACA) - ИММУНОГЕННЫЕ КОНСТРУКЦИИ В-ЭПИТОПОВ И ИХ СОСТАВЫ ДЛЯ ИММУНОТЕРАПИИ РАКА
Опухолевые клетки характеризуются профилями аберрантного гликозилирования, которые приводят к гетерогенности, усечению и сверхэкспрессии поверхностных олигосахаридов. Три основные категории структур сахаридов были идентифицированы как потенциальные ассоциированные с опухолью углеводные антигены (TACA), продемонстрированные как GD3, GD2, Глобо-H, GM2, Фукозил GM1, PSA, Ley, Lex, SLex, SLeaи STn. как продемонстрировано на Фигуре 16.
(1) Связанный с муцином о-гликан: Tn, TF и STn
(2) Глико-сфинголипиды: включая ганглиозид GM3, GM2, GD2, GD3, фукозил GM1 и нейтральный глобозид глобол H
(3) Антигены группы крови: SLe x, Le y Le x, SLe a и Le y
Глобол H экспрессируется на поверхности клеток рака молочной железы в виде гликолипида и является привлекательным онкомаркером. Гекса-сахарид глобола H был синтезирован с применением подхода гликальной сборки для синтеза олигосахаридов. Глоба H служит в качестве B-гаптена и может иметь функциональную группу на восстанавливающих концах, чтобы обеспечить ковалентную иммобилизацию с помощью пептидов Th хелперов, приведенных в Таблице 2, с образованием иммуногенных пептидных конструкций. Использование описанных Th-эпитопов в этом типе применения намного более универсально, чем другие подходы с использованием KLH или других обычных белков-носителей, как описано ранее Danishefsky и Livingston (website: glycopedia.eu/Hetero-TACA-vaccines-based-on-protein-carriers).
N-конец каждого полипептида или первичных аминов в боковой цепи остатков лизина (K) белков доступны в качестве мишеней для сшивающих линкерных реагентов N-гидроксисукцинимидного (NHS) типа при pH 7-9 с образованием стабильных амидных связей вдоль с высвобождением уходящей группы N-гидроксисукцинимида. Кроме того, сложный эфир m-малеимидобензоил-N-гидроксисукцинимида (MBS) (H.L. Chiang, et al. Vaccine. 2012 30(52), 7573-7581), сложный эфир p-нитрофенила (PNP) (S.J. Danishefsky, et al. Acc.. Chem. Res. 2015, 48 (3), 643-652) вместе с линкерами разной длины цепи также могут служить важными линкерами конъюгации TACA для Th-хелперных пептидов, приведенных в Таблице 2.
Искусственные Th эпитопы по данному изобретению можно применять в композиции противораковой вакцины, содержащей: (а) иммуногенную композицию, содержащую гликан, по существу, Глобол H или его иммуногенный фрагмент; (b) иммуногенный фрагмент, ковалентно связанный с неизбирательным искусственным пептидом Т-хелперного эпитопа, приведенным в Таблице 2. Пептиды Th-эпитопа, приведенные в Таблице 2 (например, UBITh®), индивидуально получают поэтапно путем твердофазного пептидного синтеза (SPPS) и химии Fmoc со снятием защиты/связыванием от С-конца к N-концу. Последовательность целевого пептида была сконструирована соответствующим образом.
Гликан может быть связан с искусственным пептидом Th-эпитопа, со спейсером или без него, посредством образования амидной связи непосредственно на твердофазной смоле, при этом реакция связывания контролируют с помощью теста Кайзера. Для этого связывания можно применять две стратегии: (1) получение производных гликана с активирующей уходящей группой с последующим связыванием с Th-пептидом, соединенным со смолой и связанным со спейсером, на N-конце которого имеется свободный амин, или (2) превращение связанной со смолой N-концевой аминогруппы в активирующий сложный эфир с последующей реакцией присоединения к гликану. Прямое присоединение гликана к Th-пептиду, соединенному со смолой и связанному со спейсером, могло бы обеспечить более эффективную реакцию присоединения к активирующей группе на связанной со смолой свободной аминогруппе с последующим высвобождением связанного гликанового пептида из смолы с помощью стандартной реакции расщепления без смолы. Стерические препятствия между двумя свободными большими молекулами, такими как полисахариды и длинноцепочечный пептид, могут затруднить реакцию присоединения гликана и пептида и, таким образом, сделать ее менее эффективной и с низким выходом.
В некоторых аспектах T-хелперный носитель, связанный с B-гаптеном, представляет собой связанные со спейсером Th-пептиды, описанные в Таблице 2.
В некоторых вариантах осуществления линкер представляет собой p-нитрофенильный линкер, N-гидроксисукцинимидный линкер, p-нитрофениловый сложный эфир (сложный эфир PNP) или N-гидроксисукцинимидный сложный эфир (сложный эфир NHS ). Сложные эфиры NHS могут реагировать с первичными аминами при pH 7-9 с образованием стабильных амидных связей вместе с высвобождением уходящей группы N-гидроксисукцинимида.
В этом примере используются следующие сокращения: PNP: p-нитрофениловый эфир; NPC: N-нитрофенилхлорформиат; DSS (дисукцинимидил суберат); и NHS: N-гидроксисукцинимид; MBS: m-малеимидобензоил-N-гидроксисукцинимидный сложный эфир.
Чтобы продемонстрировать различные варианты осуществления данного изобретения включены следующие стадии химических реакций и приготовлений (Фигуры с 17 по 21).
1. Получение активированного UBITh® с p-нитрофенильной (PNP) группой
Носитель искусственного Th-эпитопа (например, UBITh®) можно синтезировать с помощью автоматизированного твердофазного синтеза и химии Fmoc. После снятия Fmoc-защиты N-концевой аминокислоты на удлиненной пептидной цепи свободная аминогруппа может быть преобразована в активную 4-нитрофенильную группу обработкой 4-нитрофенилхлорформиатом (NPC, 10 экв.) в растворе DMF, содержащем 10% триэтиламин. Полученную смесь "пептид-смола" можно промыть раствором DCM для удаления реагента и остатка 4-нтриофенола. Желаемый активированный UBITh® с p-нитрофенильной группой может быть получен для дальнейшего конъюгирования с гликаном.
2. Получение активированного UBITh® с N-гидроксисукцинимидной (NHS) группой
Пептидный носитель UBITh® (например, UBITh®) можно синтезировать с помощью автоматизированного твердофазного синтеза и химии Fmoc. После снятия Fmoc защиты N-концевой аминокислоты на удлиненной пептидной цепи свободная аминогруппа может быть преобразована в активный N-гидроксисукцинимид путем обработки DSS в растворе DMF. Полученную смесь "пептид-смола" можно промыть раствором DCM для удаления остатков реагента. Желаемый активированный UBITh® с N-гидроксисукцинимидной группой может быть получен для дальнейшего конъюгирования с гликаном.
3. Синтез пептида UBITh®, конъюгированного с Глобол H
Приготовление гексасахаридных аналогов Глобол H с концевой аминогруппой (2) может быть осуществлено согласно стратегии однореакторного синтеза (C.Y. Huang, et al., Proc. Natl Acad Sci USA 2006, 103, 15-20). Вкратце, раствор Глобол H можно ввести в активированную смолу UBITh® и затем осторожно перемешать в течение 3 часов. После отщепления от смолы и полного снятия защиты неочищенный пептид UBITh®, конъюгированный с Глобол H, может быть очищен с помощью препаративной высокоэффективной жидкостной хроматографии (ВЭЖХ) и охарактеризован с помощью времяпролетной масс-спектрометрии с лазерной ионизацией и десорбцией из жидкой матрицы (MALDI-TOF) и анализа обращенно-фазовой ВЭЖХ.
4. Получение Глобол Н активированного эфира.
Глобол H гексиламин (2) можно растворить в безводном растворе DMF. Затем можно добавить сложный диэфир p-нитрофениладипата и перемешать в течение 2-4 часов при комнатной температуре. Мониторинг реакции можно осуществлять с помощью TLC и теста Кайзера, чтобы проверить исчезновение свободной аминогруппы. Растворитель DMF можно удалить при пониженном давлении без нагревания, а затем полученный остаток можно трижды экстрагировать дихлорметаном и водой с 0,5% уксусной кислотой. Полученный водный раствор можно сконцентрировать и очистить с помощью хроматографии на колонке с обращенной фазой (RP-C18) (изократическое элюирование с MeOH/H2O с 1% уксусной кислотой).
5. Получение активированного эфира GM3.
Синтез и очистка аналогов ганглизидов GM3 были описаны ранее (Jacques S, et al., J. Am. Chem. Soc., 2012 134(10):4521-4). GM3 аналоги амина X (4,5 мг; 5,2 мкмоль) можно растворить в диметилформамиде (1,5 мл) и добавить триэтиламин (3,0 экв.). Затем можно добавить сложный диэфир p-нитрофениладипата (10,0 экв.). Реакция может быть завершена по данным TLC (CH2Cl2-MeOH-H2O-AcOH; 4 : 5 : 1 : 0.5). Значение pH реакции можно довести до 5,0 с помощью уксусной кислоты с последующим совместным выпариванием с толуолом (3х). Остаток можно сконцентрировать, и полученный остаток можно очистить с помощью ВЭЖХ (полупрепаративная колонка Beckman C18-силикагель) с применением градиента MeOH - H2O, содержащего 1% уксусную кислоту. Спектр 1H ЯМР может быть получен в CD3OD.
6. Получение Tn активированного эфира.
Сахарид Tn можно синтезировать на основании предыдущих публикаций (T. Toyokuni, et al., Bioorg Med Chem. 1994, 11, 1119-32; and S.D. Scott, et al., J. Am. Chem. Soc., 1998, 120(48), 12474-85). Аналог 6 (ммоль), NHS (160 мг, 1,39 ммоль) и EDC (268 мг, 1,40 ммоль) в сухом CH2Cl2 (25 мл) можно перемешивать при комнатной температуре в течение 1 часа. Смесь можно промыть предварительно охлажденной H2O (3 х 30 мл), высушить (Na2SO4) и сконцентрировать с получением производных сукцинимид-эфира (7) в виде бесцветного сиропа.
7. Получение активного сложного эфира сиалила Льюиса x (sLe x) (9)
Получение производного sLex тетрасахарида (8) может быть осуществлено с помощью опубликованной синтетической стратегии (G. Kuznik, Bioorganic & Medicinal Chemistry Letters, 7(5):577-580, 1997). Соединение (8) добавляют к раствору NPC (2 экв.) в DMF/DCM, нагревают до комнатной температуры и перемешивают еще 30 мин. Концентрированную неочищенную смесь можно промыть DCM и водным раствором. Полученный водный раствор можно обработать при пониженном давлении и очистить на колонке RP C18.
8. Общая процедура получения гликококъюгатов.
Стандартную реакцию присоединения отдельных гликанов к отдельным Th-пептидам с включенными в них спейсерами, связанными со смолой, проводят по стандартным процедурам присоединения с образованием пептидной связи, осуществляемым с помощью обычного твердофазного пептидного синтезатора (ов) посредством реакции карбодиимидного присоединения.
Пептид со связанным с смолой гликаном может быть удален из смолы, выделен, осажден и лиофилизирован в соответствии со стандартной процедурой синтеза пептидов.
Носитель эпитопов Т-клеточного пептида (например, UBITh®) можно синтезировать с помощью автоматизированного твердофазного синтеза и химии Fmoc. После снятия Fmoc защиты N-концевой аминокислоты на удлиненной пептидной цепи свободная аминогруппа может стать доступной для дальнейшей реакции конъюгации. Синтетические сахаридоные аналоги Глобо H, Tn, GM3, SLexи т.д. с активирующей модификацией уходящей группы можно растворить в растворе DMF и добавить в систему SPPS для реакции с N-концевым амином пептида UBITh®. Мониторинг реакции можно проводить с помощью теста Кайзера. После отщепления от смолы и полного снятия защиты, конъюгированный с сахаридами пептид UBITh® может быть очищен с помощью препаративной высокоэффективной жидкостной хроматографии (ВЭЖХ) и охарактеризован с помощью времяпролетной масс-спектрометрии с лазерной ионизацией и десорбцией из жидкой матрицы (MALDI-TOF) и анализа обращенно-фазовой ВЭЖХ.
Таким образом, в этом примере подробно описана эффективная конъюгация отдельных гликанов с отдельными пептидами Th-хелперного эпитопа, приведенными в Таблице 2, чтобы было представление, как эффективно приготовить такие углеводно-пептидные иммуногенные конструкции для последующих вакцинных составов. Эти углеводные вакцинные составы с применением искусственных Th-эпитопов могут обеспечивать сфокусированные ответы антител, направленные на целевые углеводные антигены, обычно присутствующие на раковых клетках, что позволяет проводить иммунотерапию пациентов с раком в клинических протоколах.
ПРИМЕР 12
УСИЛЕНИЕ ФУНКЦИЙ ЭФФЕКТОРНЫХ Т-КЛЕТОК ПРИ СВЯЗИ НЕИЗБИРАТЕЛЬНЫХ ИСКУССТВЕННЫХ Т-ХЕЛПЕРНЫХ ЭПИТОПОВ С ЭФФЕКТОРНЫМИ КЛЕТОЧНЫМИ ЭПИТОПАМИ (например, CTL ЭПИТОПАМИ) И ИХ СОСТАВЫ ДЛЯ РАЗРАБОТКИ УНИВЕРСАЛЬНЫХ Т-КЛЕТОЧНЫХ ВАКЦИН ПРОТИВ ВИРУСНЫХ ИНФЕКЦИЙ
Вступление
Т-хелперные клетки несут поверхностный маркер CD4 и экспрессируют поверхностный рецептор, известный как рецептор Т-клеток, состоящий из полипептидного гетеродимера (обозначенного, например, α/β). Т-хелперные клетки распознают вирусные пептиды в ассоциации с белком MHC класса II, обычно на поверхности антигенпрезентирующей клетки (APC). Эти взаимодействия приводят к активации, пролиферации и дифференцировке Т-хелперных клеток, обеспечивая достаточно высокую аффинность связывания.
Т-хелперные клетки (CD4+ T-клетки) обеспечивают растворимые медиаторы и взаимодействия рецептор-лиганд с клетками как врожденной, так и адаптивной иммунной системы, которые запускают и модулируют их эффекторную функцию. Эти клетки представляют собой гетерогенную популяцию, и на сегодняшний день охарактеризовано несколько подмножеств, включая Th1, Th2, Th17 и Т-фолликулярные хелперные (Tfh) клетки. Также известны регуляторные CD4+ Т-клетки (Treg), которые подавляют рост и функцию подмножеств Т-хелперных клеток и цитотоксических клеток.
Каждый тип эффекторных Т-клеток контролируется ключевым регулятором транскрипции, экспрессирует определенный набор молекул клеточной поверхности и секретирует «сигнатурные» цитокины, которые вместе способствуют специфической роли этой субпопуляции Т-клеток в одном из звеньев иммунной системы.
Клетки Tfh отличаются от других подмножеств хелперов своей уникальной способностью самонаводиться на B-клеточные фолликулы и оказывать помощь антиген-специфическим B-клеткам, которые подвергаются соматической гипермутации (SHM) своих генов Ig V области и изменяют свою аффинность к антигену. Клетки Tfh секретируют цитокин IL-21, который необходим для дифференцировки В-клеток и развития высокоаффинных, ответов антител против вирусов с переключением изотипа. Сигналы, опосредованные Tfh, обеспечивают отбор В-клеток с более высокой аффинностью к иммунизирующему антигену, которые затем могут дифференцироваться, чтобы стать долгоживущими плазматическими клетками или В-клетками памяти. Из-за возможности появления самореактивных клонов B-клеток в процессе SHM и продолжительности жизни выбранных клонов чрезвычайно важно, чтобы существовали строгие механизмы толерантности для контроля доставки положительных сигналов отбора от Tfh-клеток к В-клеткам.
Клетки Th1 в основном участвуют в усилении цитотоксического ответа. Эти клетки способствуют клеточному ответу на вирусную инфекцию, стимулируя созревание цитотоксических предшественников Т-клеток, частично за счет секреции цитокинов IL-2 и IFN-γ. Клетки Th1 также секретируют фактор некроза опухоли (TNF), опосредуют реакции гиперчувствительности замедленного типа и способствуют выработке антител IgG2a. Клетки Th1 значительно усиливают иммунный ответ, активируя макрофаги и другие Т-клетки в месте вирусной инфекции. Этот ответ является основой реакций гиперчувствительности замедленного типа, которые являются признанной частью патогенеза многих вирусных инфекций.
Другие клетки Th, включая клетки Th2 и Th17, также способствуют иммунному ответу против вирусной инфекции, способствуя воспалению или образованию специфических изотипов антител.
Некоторые Т-клетки могут подавлять ответы других Т-клеток и/или В-клеток. Известно отдельное подмножество CD4+ T-клеток, называемое подмножеством регуляторных Т-клеток (T-reg). Существует два основных типа T-reg: (1) tTreg, которые продуцируются в тимусе во время отрицательного отбора и, как полагают, в основном участвуют в борьбе с аутоиммунными заболеваниями; и (2) iTreg, которые индуцируются во время иммунных ответов и участвуют в прекращении иммунных ответов и возвращении иммунной системы к гомеостазу. Клетки T-reg также могут помочь поддерживать баланс между защитой и иммуноопосредованной патологией.
Влияние Т-хелперов на пептидную вакцинацию огромно. CD4+ Т-хелперные клетки после активации могут обеспечивать сильные устойчивые CD8+ Т-клеточные ответы за счет включения когнатных Т-хелперных эпитопов. Принимая во внимание местную иммуномодулирующую функцию CD4 Т-клеток, предпочтительно активировать когнатную помощь в ткани-мишени, производной от антигенов целевого вируса или опухоли. Чужеродные антигены и даже ассоциированные с опухолью белки часто содержат иммуногенные участки (Th-эпитопы), которые функционируют как горячие точки для иммунной системы. Пептидная иммуногенная конструкция, разработанная и описанная в данном изобретении, посредством ковалентного связывания выбранных неизбирательных искусственных Th-эпитопов с целевыми B-клетками или эффекторными Т-клетками (например, эпитопами CTL) может облегчить тесные взаимодействия APC, CD4 и CD8 T-клеток для индукции оптимальных и защитных CD8 T-клеточных ответов.
Примеры пептидов эпитопа CTL в качестве целевых антигенных сайтов для включения в вирусоспецифические универсальные Т-клеточные вакцины
1. Компонент CTL в вакцине против ВИЧ:
Несмотря на антиретровирусную терапию (АРТ), вирус иммунодефицита человека (ВИЧ)-1 сохраняется в стабильном латентном резервуаре, в первую очередь в CD4+ Т-клетках памяти, находящихся в состоянии покоя. Этот резервуар представляет собой серьезный барьер на пути излечения от ВИЧ-1-инфекции. С целью очищения этого резервуара, фармакологическая реактивация латентного ВИЧ-1 была протестирована как in vitro, так и in vivo. Ключевой нерешенный вопрос заключается в том, могут ли вирус-специфические иммунные механизмы, включая цитотоксические Т-лимфоциты (CTL), очистить инфицированные клетки у пациентов, получавших АРТ, после реверсии латентного периода. После обширного анализа данных, CTL, которые могли распознавать эпитопы немутантного латентного ВИЧ-1 у всех протестированных хронически инфицированных пациентов, были идентифицированы со специфическими пептидами, репрезентативными для таких эпитопов CTL, как продемонстрировано в Таблице 3B (SEQ ID NO: 76-82), включенными в разработку универсальной Т-клеточной вакцины против ВИЧ. Хронически инфицированные пациенты сохраняют вирусоспецифический CTL-ответ широкого спектра. Ожидается, что соответствующее усиление этого ответа с помощью этой универсальной Т-клеточной вакцины против ВИЧ, включающей эти пептиды CTL-эпитопа с ковалентной специфеческой связью с неизбирательными искусственными эпитопами Th по данному изобретению (SEQ ID NO: 1-52), приведет к устранению латентного резервуара.
2. Компонент CTL в вакцине против HSV:
Вирус простого герпеса поражает значительную часть населения мира и вызывает латентную инфекцию, при которой вирусный геном сохраняется в сенсорных нейронах без образования вирионов. Периодическая реактивация вируса из этого латентного состояния приводит к поражениям, которые могут поражать поверхности слизистой оболочки рта и губ, половых путей и роговицу глаза, и реже - кожу и головной мозг. HSV-2 может быть смертельным для новорожденных, которые заразились через родовые пути; HSV-1 инфекции роговицы являются ведущей инфекционной причиной слепоты; а HSV-1 инфекции головного мозга составляют примерно четверть случаев вирусного энцефалита, который может быть смертельным. Вакцины HSV-1, которые прошли клинические исследования, были в первую очередь разработаны для продукции антител и оказались в значительной степени неэффективными. Данные свидетельствуют о значительной роли CD8+ T-клеток в контроле HSV-инфекций как у мышей, так и у людей.
HSV типа 1 (HSV-1) экспрессирует свои гены последовательно как немедленно-ранний (α), ранний (β), поздний (γ1) и истинно поздний (γ2), при этом синтез вирусной ДНК является абсолютным условием только для экспрессии гена γ2. Гликопротеин B (gB) белка γ1 содержит сильно иммунодоминантный CD8+ T-клеточный эпитоп (gB498-505), который распознается 50% как CD8+ эффекторных Т-клеток в остро инфицированных ганглиях тройничного нерва (TG), так и CD8+ Т-клеткок памяти в латентно инфицированных TG.
Благодаря получению обширных данных и тщательному их анализу, полный репертуар HSV-специфических CD8+ T-клеток у мышей C57BL/6 рассматривался при разработке дизайна вакцины против HSV, содержащей CTL. Кроме того, было обнаружено, что различные наборы HSV-1 gB эпитопов распознаются CD4+ T-клетками от индивидуумов, имеющих симптоми, по сравнению с бессимптомным течением. Среди них, gB166-180, gB661-675 и gB666-680 были нацелены на CD4+ CTL, лизирующие аутологичный HSV-1- и вирус осповакцины (экспрессирующий gB [VVgB])-инфицированные LCL. gB 166-180 и gB 666-680, по-видимому, распознаются преимущественно CD4+ T-клетками от HSV-1-серопозитивных здоровых «бессимптомных» индивидуумов, в то время как gB661-675, по-видимому, распознается преимущественно CD4+ T-клетками от индивидуумов с выраженными симптомами. Эффективная иммунотерапевтическая вакцина против герпеса исключила бы потенциальный «симптоматический» эпитоп gB661-675. Кроме того, также были идентифицированы три эпитопа VP11/12 CD8+, которые на высоком уровне распознаются у бессимптомных индивидуумов, которые, как было обнаружено, вызывают сильный защитный иммунитет в «гуманизированной» модели офтальмогерпеса у трансгенных мышей HLA-A*02:01.
Ряд HSV CTL эпитопов был идентифицирован с помощью специфических пептидов, репрезентативных для таких эпитопов CTL, как продемонстрировано в Таблице 3B (SEQ ID NO: 83-106), включенных в конструкцию по данному изобретению для разработки универсальной мультиэпитопной Т-клеточной вакцины против HSV .
3. Универсальные Т-клеточные вакцины против FMDV, PRRSV и CSFV в свиноводстве.
Вирус ящура (FMDV), вирус репродуктивного и респираторного синдрома свиней (PRRSV) и вирус классической чумы свиней (CSFV) являются патогенами, крайне негативно влияющими на отрасль свиноводства. Разработка эффективных вакцин против этих патогенов имеет практическое значение в свиноводстве.
Хотя нейтрализующие антитела, индуцированные после вакцинации, очень эффективны в борьбе с болезнями и передачей вирусов, они не обеспечивают защиту от перекрестных подтипов и могут стать неэффективными из-за антигенных изменений. Клеточные иммунные ответы, особенно продукция цитотоксических Т-лимфоцитов (CTL), привлекают большое внимание из-за их потенциала в разработке эффективных пептидных вакцин с перекрестной защитой против различных вирусов. Например, пептиды эпитопа CTL можно применять для разработки вакцин против гриппа человека с перекрестной защитой, включая рекомбинантный вирусный вектор и пептидную вакцину; пептид эпитопа CTL, идентифицированный для O-серотипа FMDV, имел перекрестную реактивность с другими серотипами FMDV. Однако большая часть анализов была ограничена конкретными вирусными белками и позволила идентифицировать лишь несколько эпитопов CTL.
После обширного сбора данных, разработки дизайна, синтеза, трудоемких и длительных процедур анализа иммуногенности и функциональности, оценка больших наборов разработанных пептидов CTL позволила валидировать выбранные вирусоспецифические эпитопы CTL, полученные из различных вирусных белков FMDV, PRRSV и CSFV. Выбранные пептиды CTL, представляющие эти эпитопы, приведены в Таблице 3B с SEQ ID NO: 107-145.
В наш процесс отбора и идентификации был интегрирован биоинформатический поток для анализа вирусных последовательностей свиней для решения нескольких проблем: (1) генетическая изменчивость, (2) неполный скрининг определенных поверхностных белков и (3) неправильный прогноз, основанный на антигенах лейкоцитов, полученных не от свиней. Объединение этих пептидов эпитопа CTL с надлежащим связыванием неизбирательных искусственных пептидов Th-эпитопа, приводящее к созданию пептидных иммуногенных конструкций по данному изобретению, привело бы к разработке T-клеточных вакцин с устойчивой памятью и долгосрочными ответами CTL. Коммерческая разработка таких высокоточных эффективных вакцин для свиней на основе пептидов против FMDV, PRRSV, CSFV и других вирусных инфекций, которые часто имеют разрушительное влияние на свиноводство, будет иметь первостепенное значение для этой отрасли животноводства.
ТАБЛИЦА 1
Аминокислотные последовательности Th-эпитопов, полученных из патогенных белков, включая идеализированные искусственные Th-эпитопы, для применения в разработке пептидных иммуногенных конструкций
(SSAL1-Th1)
EI EIR III RIE I
V V VVV V V
F F FFF F F
XXSXXXGXXXHXXXGX
2
3
4
5
T RT TR T
ISIXEIXXVIVXXIEXILF
9
10
T RT TR T
KKKISIXEIXXVIVXXIEXILF
12
13
T RT TR
ISIXEIXXVIVXXIETILF
15
16
(SSAL2 Th2)
RRRIKII RII I L IR
VRVV VV V I V
F FF FF F V F
F
XXXXXXXTXXXTXPXSXX
20
21
22
23
24
FFLL L ITTI
KKKXXXXTRIXTIXXXXD
26
27
ТАБЛИЦА 2
Примеры необязательных гетерологичных спейсеров и CpG-олигонуклеотидов
аланин, аргинин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин
ε-N-лизин, ß-аланин, орнитин, норлейцин, норвалин, гидроксипролин, тироксин, γ-амино масляную кислоту, гомосерин, цитруллин, аминобензойную кислоту, 6-аминокапроновую кислоту (Aca; 6-аминогексановую кислоту), гидроксипролин, меркаптопропионовую кислоту (MPA), 3-нитро-тирозин, пироглутаминовую кислоту и тому подобное
-NHCH(X)CH2SCH2CO(εN)Lys-,
-NHCH(X)CH2S-сукцинимидил(εN)Lys-,
-NHCH(X)CH2S-(сукцинимидил)-
(полностью фосфоротиоатная)
(полностью фосфоротиоатная)
(полностью фосфоротиоатная)
ТАБЛИЦА 3A
Примеры целевых антигенных сайтов (В-клеточные эпитопы)
(Получено из GenBank: NP_000336)
(Получено из GenBank: AGF19246.1)
(Получено из Uniprot C7G0W1-1; GenBank: BAH97742.1)
для глиомы
(МУТАНТ от G к D)
* Аминокислоты, замененные цистеином, подчеркнуты.
ТАБЛИЦА 3B
Примеры целевых антигенных сайтов (CTL эпитопы)
ТАБЛИЦА 4
Типовые пептидные иммуногенные конструкции
T RT TR
FFLL L ITTI
T RT TR
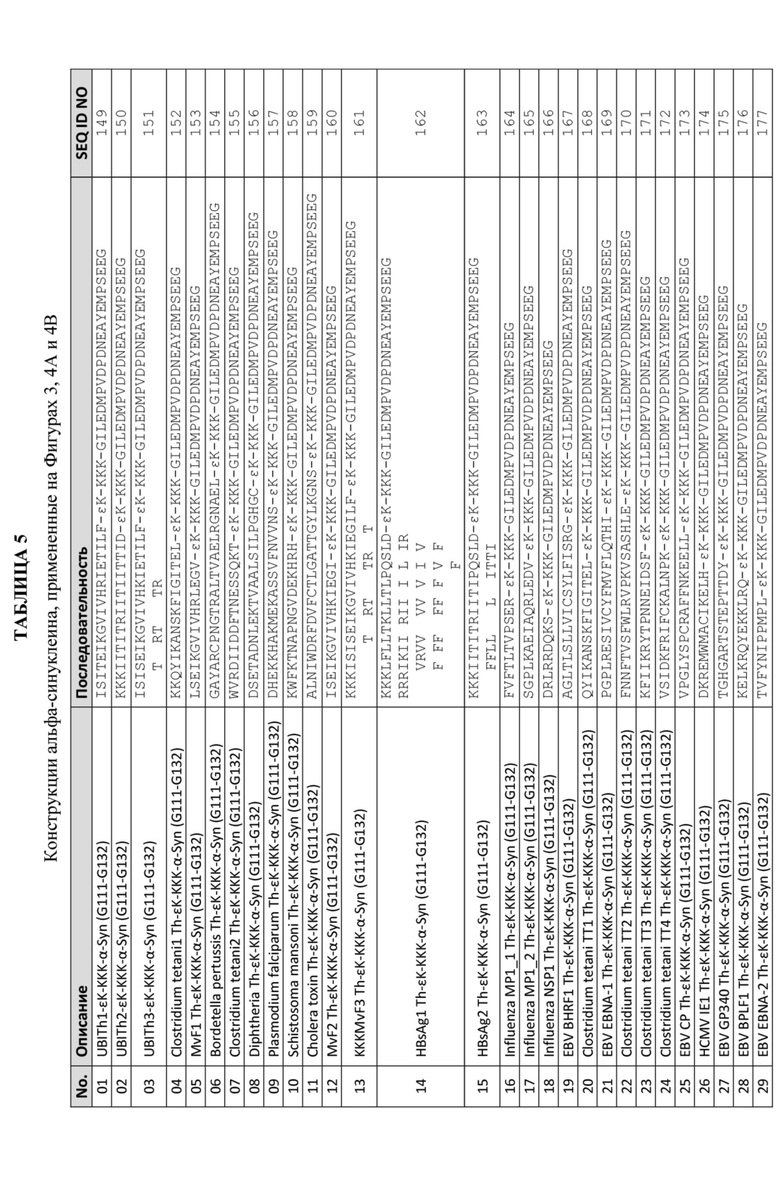
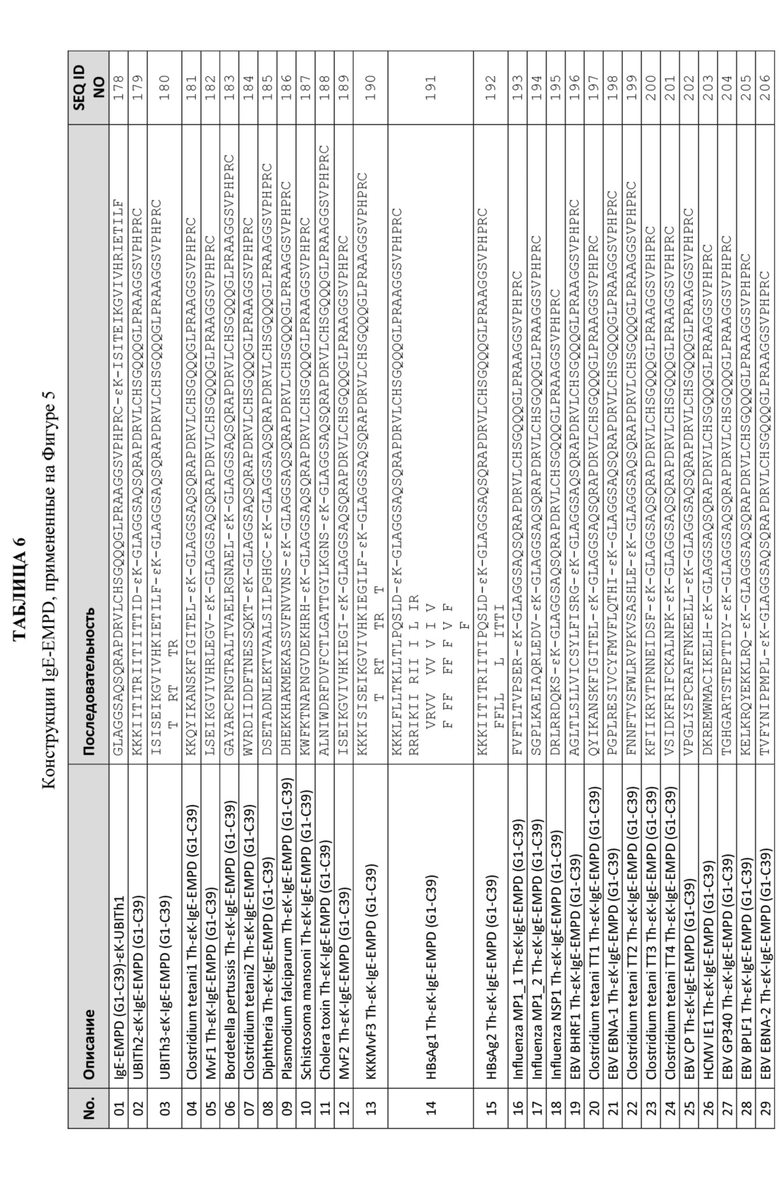
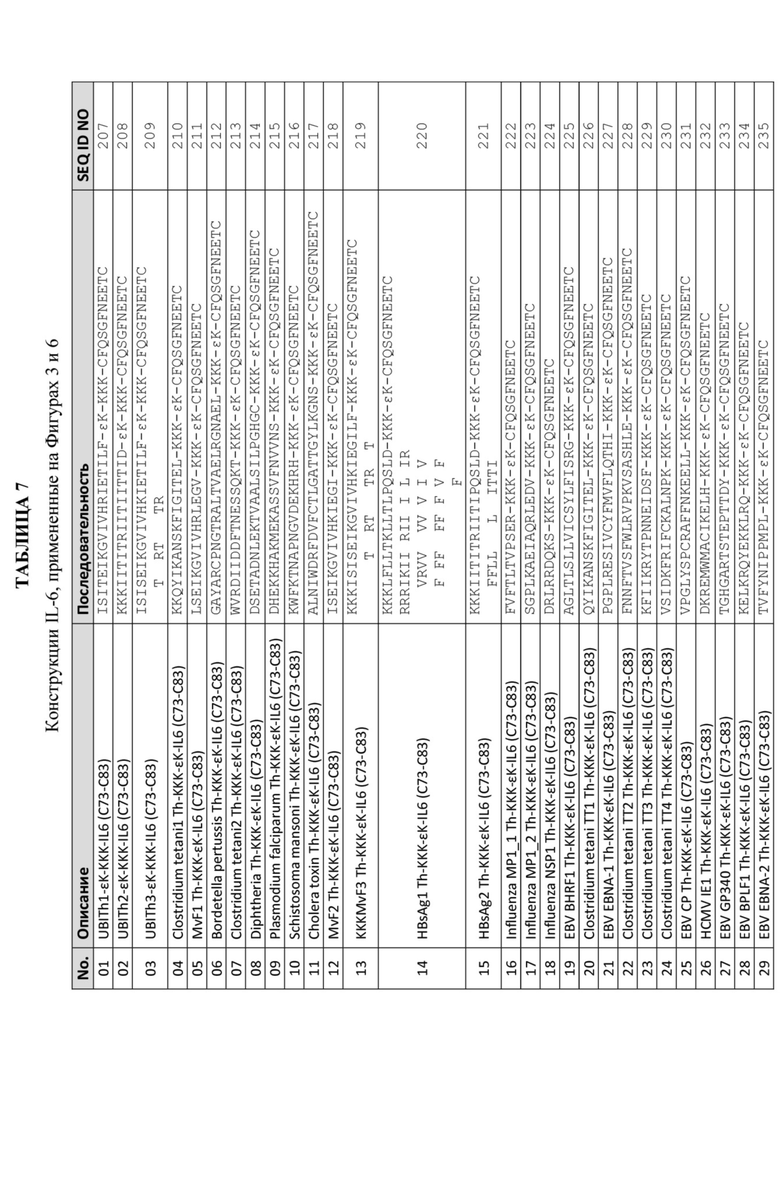
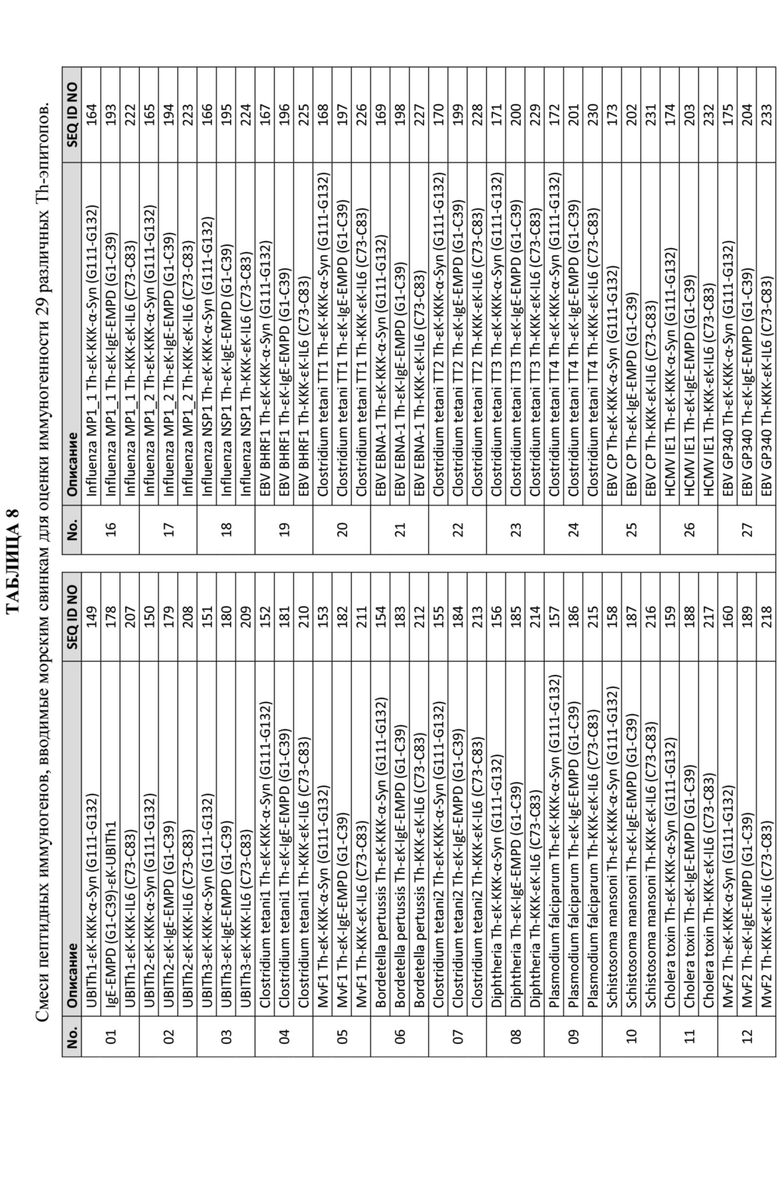
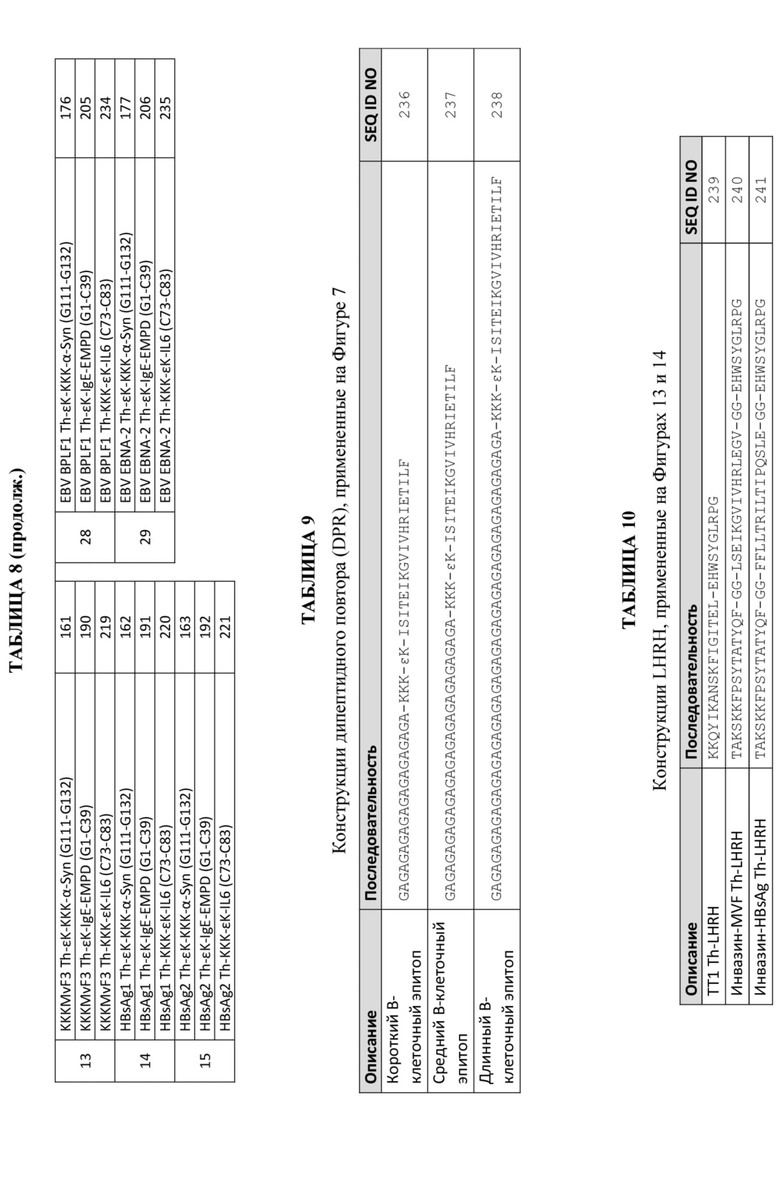
ТАБЛИЦА 11
Исключительная иммуногенность иммуногенов Aβ1-14, которые нацелены на пептиды Aβ, но не на Th-эпитопы, у морских свинок
№
(SEQ ID NO: 60)
(SEQ ID NO: 17)
(SEQ ID NO: 28)
(SEQ ID NO: 67)
+
Aβ1-14-εK-HBsAg3 Th
(SEQ ID NO: 68)
(SD)
(0,48)
(0,12)
(0,09)
ТАБЛИЦА 12
Измерение концентрации цитокинов в мононуклеарных клетках периферической крови (PBMC) вакцинированных UB311 или здоровых макаков Cynomolgus, собранных в моменты времени 15, 21 и 25,5 wpi при стимуляции пептидами Aβ1-14, Aβ1-42 или митогеном PHA (фитогемагглютином)
(SEQ ID NO: 56)
(SEQ ID NO: 58)
a Результат был приведен как среднее значение ± стандартное отклонение
b BDL, ниже уровня обнаружения
ТАБЛИЦА 13
Индекс стимуляции PBMC, оцененный у 19 пациентов с болезнью Альцгеймера
(SEQ ID NO: 56)
(SEQ ID NO: 57)
(SEQ ID NO: 59)
(SEQ ID NO: 58)
(SEQ ID NO: 60)
(нерелевантный пептид)
ТАБЛИЦА 14
Концентрации цитокинов в мононуклеарных клетках периферической крови (PBMC), оцененные у 19 пациентов при стимуляции пептидами Aβ или митогеном PHA1
(SEQ ID NO: 56)
(32,5)
(24,3)
(16,9)
(12,9)
(29,7)
(52,0)
(1,6)
(1,6)
(62,8)
(51,7)
(SEQ ID NO: 57)
(31,4)
(23,9)
(16,1)
(14,2)
(31,7)
(42,6)
(1,6)
(1,8)
(72,2)
(69,7)
(SEQ ID NO: 59)
(34,3)
(28,0)
(23,6)
(24,4)
(25,4)
(41,8)
(1,5)
(2,5)
(66,5)
(67,8)
(SEQ ID NO: 58)
(25,7)
(21,2)
(9,7)
(15,6)
(70,9)3
(51,3)
(0,86)
(1,5)
(18,4)
(39,3)
(SEQ ID NO: 60)
(17,7)
(16,9)
(16,1)
(77,3)
(130)4
(121)5
(22,7)
(50,2)
(71,5)3
(133)6
(27,4)
(26,0)
(18,4)
(30,0)
(52,1)
(95,8)
(0,64)
(0,53)
(23,5)
(29,3)
(11,3)
(6,5)
(0,00)2
(4,8)2
(0,00)2
(0,00)2
(84,8)
(99,7)
(30,5)2
(46,5)2
(24,9)
(33,1)
(12,3)
(18,2)
(41,9)
(76,5)
(2,5)
(1,6)
(70,9)
(67,8)
1 Поддающийся количественной оценке диапазон анализа составляет от 5 до 320 пг/мл
2 Концентрация у > 90% субъектов была выше верхнего предела количественного определения (AQL> 320 пг/мл)
3 У одного пациента было значение AQL 4 У шести пациентов были значения AQL
5 У восьми пациентов были значения AQL 6 У четырех пациентов были значения AQL
7 Отсутствие продукции IL-2, наблюдаемое в ответ на митоген PHA, соответствовало данным, полученным в аналогичных экспериментальных условиях.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UBI US Holdings, LLC
UBI IP Holdings
Wang, Chang Yi
<120> ИСКУССТВЕННЫЕ НЕИЗБИРАТЕЛЬНЫЕ ЭПИТОПЫ КЛЕТОК Т-ХЕЛПЕРОВ КАК
ИММУННЫЕ СТИМУЛЯТОРЫ ДЛЯ СИНТЕТИЧЕСКИХ ПЕПТИДНЫХ ИММУНОГЕНОВ
<130> 2039-WO
<140> TBD
<141> 2019-12-19
<150> US 62/782,253
<151> 2018-12-19
<160> 243
<170> PatentIn версия 3.5
<210> 1
<211> 16
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> MvF Th
<400> 1
Asp Leu Ser Asp Leu Lys Gly Leu Leu Leu His Lys Leu Asp Gly Leu
1 5 10 15
<210> 2
<211> 16
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> MvF Th
<400> 2
Glu Ile Ser Asp Glu Ile Arg Leu Ile Ile Ile Lys Arg Ile Glu Ile
1 5 10 15
<210> 3
<211> 16
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> MvF Th
<400> 3
Asp Val Ser Asp Val Lys Gly Val Val Val His Lys Val Asp Gly Val
1 5 10 15
<210> 4
<211> 16
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> MvF Th
<400> 4
Asp Phe Ser Asp Phe Lys Gly Phe Phe Phe His Lys Phe Asp Gly Phe
1 5 10 15
<210> 5
<211> 16
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> MvF Th
<220>
<221> САЙТ
<222> (1)..(1)
<223> D или E
<220>
<221> САЙТ
<222> (2)..(2)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (4)..(4)
<223> D или E
<220>
<221> САЙТ
<222> (5)..(5)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (6)..(6)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (9)..(9)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (12)..(12)
<223> K или R
<220>
<221> САЙТ
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (14)..(14)
<223> D или E
<220>
<221> САЙТ
<222> (16)..(16)
<223> L или I или V или F
<400> 5
Xaa Xaa Ser Xaa Xaa Xaa Gly Xaa Xaa Xaa His Xaa Xaa Xaa Gly Xaa
1 5 10 15
<210> 6
<211> 15
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF1 Th
<400> 6
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val
1 5 10 15
<210> 7
<211> 15
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF 2 Th
<400> 7
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile
1 5 10 15
<210> 8
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF3 Th
<400> 8
Ile Ser Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly
1 5 10 15
Ile Leu Phe
<210> 9
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 3Th
<400> 9
Ile Ser Ile Thr Glu Ile Arg Thr Val Ile Val Thr Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 10
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF3 Th
<220>
<221> САЙТ
<222> (4)..(4)
<223> S или T
<220>
<221> САЙТ
<222> (7)..(7)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> G или T
<220>
<221> САЙТ
<222> (12)..(12)
<223> H или T
<220>
<221> САЙТ
<222> (13)..(13)
<223> K или R
<220>
<221> САЙТ
<222> (16)..(16)
<223> G или T
<400> 10
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 11
<211> 22
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF3 Th
<400> 11
Lys Lys Lys Ile Ser Ile Ser Glu Ile Lys Gly Val Ile Val His Lys
1 5 10 15
Ile Glu Gly Ile Leu Phe
20
<210> 12
<211> 22
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF3 Th
<400> 12
Lys Lys Lys Ile Ser Ile Thr Glu Ile Arg Thr Val Ile Val Thr Arg
1 5 10 15
Ile Glu Thr Ile Leu Phe
20
<210> 13
<211> 22
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF 3 Th
<220>
<221> САЙТ
<222> (7)..(7)
<223> S или T
<220>
<221> САЙТ
<222> (10)..(10)
<223> K или R
<220>
<221> САЙТ
<222> (11)..(11)
<223> G или T
<220>
<221> САЙТ
<222> (15)..(15)
<223> H или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> K или R
<220>
<221> САЙТ
<222> (19)..(19)
<223> G или T
<400> 13
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe
20
<210> 14
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF4 Th
<400> 14
Ile Ser Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 15
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF4 Th
<400> 15
Ile Ser Ile Thr Glu Ile Arg Thr Val Ile Val Thr Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 16
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF4 Th (UBITh3)
<220>
<221> САЙТ
<222> (4)..(4)
<223> S или T
<220>
<221> САЙТ
<222> (7)..(7)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> G или T
<220>
<221> САЙТ
<222> (12)..(12)
<223> H или T
<220>
<221> САЙТ
<222> (13)..(13)
<223> K или R
<400> 16
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 17
<211> 19
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF5 Th (UBITh1)
<400> 17
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 18
<211> 22
<212> PRT
<213> Вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF5 Th (UBITh1a)
<400> 18
Lys Lys Lys Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg
1 5 10 15
Ile Glu Thr Ile Leu Phe
20
<210> 19
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg1 Th
<400> 19
Lys Lys Lys Leu Phe Leu Leu Thr Lys Leu Leu Thr Leu Pro Gln Ser
1 5 10 15
Leu Asp
<210> 20
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg1 Th
<400> 20
Arg Arg Arg Ile Lys Ile Ile Thr Arg Ile Ile Thr Ile Pro Leu Ser
1 5 10 15
Ile Arg
<210> 21
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg1 Th
<400> 21
Lys Lys Lys Val Arg Val Val Thr Lys Val Val Thr Val Pro Ile Ser
1 5 10 15
Val Asp
<210> 22
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg1 Th
<400> 22
Lys Lys Lys Phe Phe Phe Phe Thr Lys Phe Phe Thr Phe Pro Val Ser
1 5 10 15
Phe Asp
<210> 23
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg1 Th
<400> 23
Lys Lys Lys Leu Phe Leu Leu Thr Lys Leu Leu Thr Leu Pro Phe Ser
1 5 10 15
Leu Asp
<210> 24
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 1 Th
<220>
<221> САЙТ
<222> (1)..(1)
<223> K или R
<220>
<221> САЙТ
<222> (2)..(2)
<223> K или R
<220>
<221> САЙТ
<222> (3)..(3)
<223> K или R
<220>
<221> САЙТ
<222> (4)..(4)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> F или K или R
<220>
<221> САЙТ
<222> (6)..(6)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (7)..(7)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (9)..(9)
<223> K или R
<220>
<221> САЙТ
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (11)..(11)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или L или I или V или F
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (18)..(18)
<223> D или R
<400> 24
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa
<210> 25
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg2 Th
<400> 25
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Pro Gln Ser
1 5 10 15
Leu Asp
<210> 26
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg2 Th
<400> 26
Lys Lys Lys Phe Phe Leu Leu Thr Arg Ile Leu Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp
<210> 27
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 2 Th
<220>
<221> САЙТ
<222> (4)..(4)
<223> I или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> I или F
<220>
<221> САЙТ
<222> (6)..(6)
<223> T или L
<220>
<221> САЙТ
<222> (7)..(7)
<223> I или L
<220>
<221> САЙТ
<222> (11)..(11)
<223> I или L
<220>
<221> САЙТ
<222> (14)..(14)
<223> P или I
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> S или T
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I
<400> 27
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp
<210> 28
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 3 Th
<400> 28
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp
<210> 29
<211> 15
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> HBsAg4 Th (UBITh4)
<400> 29
Phe Phe Leu Leu Thr Arg Ile Leu Thr Ile Pro Gln Ser Leu Asp
1 5 10 15
<210> 30
<211> 18
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> KKK-HBsAg Th
<400> 30
Lys Lys Lys Phe Phe Leu Leu Thr Arg Ile Leu Thr Ile Pro Gln Ser
1 5 10 15
Leu Asp
<210> 31
<211> 14
<212> PRT
<213> Вирус гепатита В
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HBsAg Th
<400> 31
Phe Phe Leu Leu Thr Arg Ile Leu Thr Ile Pro Gln Ser Leu
1 5 10
<210> 32
<211> 24
<212> PRT
<213> Bordetella pertussis
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> Bordetella pertussis Th
<400> 32
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu
20
<210> 33
<211> 25
<212> PRT
<213> Cholera Toxin
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> Cholera Toxin Th
<400> 33
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser
20 25
<210> 34
<211> 15
<212> PRT
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Clostridium tetani TT1 Th
<400> 34
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu
1 5 10 15
<210> 35
<211> 17
<212> PRT
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani 1 Th
<400> 35
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu
<210> 36
<211> 21
<212> PRT
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Clostridium tetani TT2 Th
<400> 36
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu
20
<210> 37
<211> 16
<212> PRT
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT3 Th
<400> 37
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
<210> 38
<211> 16
<212> PRT
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT4 Th
<400> 38
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
<210> 39
<211> 17
<212> PRT
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani 2 Th
<400> 39
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr
<210> 40
<211> 23
<212> PRT
<213> дифтерийная палочка
<220>
<221> ПЕПТИД
<222> (1)..(23)
<223> Diphtheria Th
<400> 40
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys
20
<210> 41
<211> 19
<212> PRT
<213> Вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> EBV BHRF1 Th
<400> 41
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly
<210> 42
<211> 20
<212> PRT
<213> Вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> EBV EBNA-1 Th
<400> 42
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile
20
<210> 43
<211> 18
<212> PRT
<213> Вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> EBV CP Th
<400> 43
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu
<210> 44
<211> 15
<212> PRT
<213> Вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> EBV GP340 Th
<400> 44
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr
1 5 10 15
<210> 45
<211> 13
<212> PRT
<213> Вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> EBV BPLF1 Th
<400> 45
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln
1 5 10
<210> 46
<211> 11
<212> PRT
<213> Вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> EBV EBNA-2 Th
<400> 46
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu
1 5 10
<210> 47
<211> 14
<212> PRT
<213> Цитомегаловирус человека
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HCMV IE1 Th
<400> 47
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His
1 5 10
<210> 48
<211> 11
<212> PRT
<213> Вирус гриппа
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> Матриксный протеин вируса гриппа 1_1 Th
<400> 48
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg
1 5 10
<210> 49
<211> 15
<212> PRT
<213> Вирус гриппа
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Матриксный протеин вируса гриппа 1_2 Th
<400> 49
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val
1 5 10 15
<210> 50
<211> 9
<212> PRT
<213> Вирус гриппа
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> Неструктурный протеин вируса гриппа 1 Th
<400> 50
Asp Arg Leu Arg Arg Asp Gln Lys Ser
1 5
<210> 51
<211> 21
<212> PRT
<213> Plasmodium falciparum
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Plasmodium falciparum Th
<400> 51
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser
20
<210> 52
<211> 17
<212> PRT
<213> Schistosoma mansoni
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Schistosoma mansoni Th
<400> 52
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His
<210> 53
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> САЙТ
<222> (1)..(1)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (1)..(4)
<223> эпсилон-K-KKK как спейсер
<400> 53
Lys Lys Lys Lys
1
<210> 54
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(4)
<223> KKK-эпсилон-K как спейсер
<220>
<221> САЙТ
<222> (4)..(4)
<223> эпсилон-K
<400> 54
Lys Lys Lys Lys
1
<210> 55
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(6)
<223> Гибкий шарнирный спейсер
<400> 55
Pro Pro Xaa Pro Xaa Pro
1 5
<210> 56
<211> 14
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> Abeta1-14
<400> 56
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His
1 5 10
<210> 57
<211> 16
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Abeta1-16
<400> 57
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Gln Lys
1 5 10 15
<210> 58
<211> 26
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(26)
<223> Abeta17-42
<400> 58
Leu Val Phe Phe Ala Glu Asp Val Gly Ser Asn Lys Gly Ala Ile Ile
1 5 10 15
Gly Leu Met Val Gly Gly Val Val Ile Ala
20 25
<210> 59
<211> 28
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(28)
<223> Abeta1-28
<400> 59
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Gln Lys
1 5 10 15
Leu Val Phe Phe Ala Glu Asp Val Gly Ser Asn Lys
20 25
<210> 60
<211> 42
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(42)
<223> Abeta1-42
<400> 60
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Gln Lys
1 5 10 15
Leu Val Phe Phe Ala Glu Asp Val Gly Ser Asn Lys Gly Ala Ile Ile
20 25 30
Gly Leu Met Val Gly Gly Val Val Ile Ala
35 40
<210> 61
<211> 22
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> alpha-Syn111-132 (Получено из GenBank: NP_000336)
<400> 61
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
1 5 10 15
Met Pro Ser Glu Glu Gly
20
<210> 62
<211> 39
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(39)
<223> IgE-EMPD1-39
<400> 62
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
20 25 30
Ser Val Pro His Pro Arg Cys
35
<210> 63
<211> 30
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(30)
<223> Tau379-408 (Получено из GenBank: AGF19246.1)
<400> 63
Arg Glu Asn Ala Lys Ala Lys Thr Asp His Gly Ala Glu Ile Val Tyr
1 5 10 15
Lys Ser Pro Val Val Ser Gly Asp Thr Ser Pro Arg His Leu
20 25 30
<210> 64
<211> 48
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(48)
<223> IL-3197-144 (Получено из Uniprot C7G0W1-1; GenBank: BAH97742.1)
<400> 64
Leu Ser Asp Lys Asn Ile Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
1 5 10 15
Leu Lys Phe Gln His Glu Pro Glu Thr Glu Ile Ser Val Pro Ala Asp
20 25 30
Thr Phe Glu Cys Lys Ser Phe Ile Leu Thr Ile Leu Gln Gln Phe Ser
35 40 45
<210> 65
<211> 37
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> Abeta 1-14
<220>
<221> ПЕПТИД
<222> (15)..(18)
<223> эпсилон-K-KKK как спейсер
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(37)
<223> MvF 4 Th
<220>
<221> САЙТ
<222> (22)..(22)
<223> S или T
<220>
<221> САЙТ
<222> (25)..(25)
<223> K или R
<220>
<221> САЙТ
<222> (26)..(26)
<223> G или T
<220>
<221> САЙТ
<222> (30)..(30)
<223> H или T
<220>
<221> САЙТ
<222> (31)..(31)
<223> K или R
<400> 65
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Lys Lys
1 5 10 15
Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile
20 25 30
Glu Thr Ile Leu Phe
35
<210> 66
<211> 33
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> Abeta 1-14
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (16)..(33)
<223> HBsAg 2 Th
<220>
<221> САЙТ
<222> (19)..(19)
<223> I или F
<220>
<221> САЙТ
<222> (20)..(20)
<223> I или F
<220>
<221> САЙТ
<222> (21)..(21)
<223> T или L
<220>
<221> САЙТ
<222> (22)..(22)
<223> I или L
<220>
<221> САЙТ
<222> (26)..(26)
<223> I или L
<220>
<221> САЙТ
<222> (29)..(29)
<223> P или I
<220>
<221> САЙТ
<222> (30)..(30)
<223> Q или T
<220>
<221> САЙТ
<222> (31)..(31)
<223> S или T
<220>
<221> САЙТ
<222> (32)..(32)
<223> L или I
<400> 66
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Lys Lys
1 5 10 15
Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa Xaa
20 25 30
Asp
<210> 67
<211> 37
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> Abeta 1-14
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон K
<220>
<221> ПЕПТИД
<222> (15)..(18)
<223> эпсилон K-KKK как спейсер
<220>
<221> ПЕПТИД
<222> (19)..(37)
<223> MvF5 Th (UBITh1)
<400> 67
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Lys Lys
1 5 10 15
Lys Lys Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile
20 25 30
Glu Thr Ile Leu Phe
35
<210> 68
<211> 33
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> Abeta 1-14
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон K
<220>
<221> ПЕПТИД
<222> (16)..(33)
<223> HBsAg3 Th
<400> 68
Asp Ala Glu Phe Arg His Asp Ser Gly Tyr Glu Val His His Lys Lys
1 5 10 15
Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr Ile
20 25 30
Asp
<210> 69
<211> 16
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Tau145-160
<400> 69
Ala Asp Gly Lys Thr Lys Ile Ala Thr Pro Arg Gly Ala Ala Pro Pro
1 5 10 15
<210> 70
<211> 50
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(50)
<223> Tau1-50
<400> 70
Met Ala Glu Pro Arg Gln Glu Phe Glu Val Met Glu Asp His Ala Gly
1 5 10 15
Thr Tyr Gly Leu Gly Asp Arg Lys Asp Gln Gly Gly Tyr Thr Met His
20 25 30
Gln Asp Gln Glu Gly Asp Thr Asp Ala Gly Leu Lys Glu Ser Pro Leu
35 40 45
Gln Thr
50
<210> 71
<211> 15
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Tau297-311
<400> 71
Ile Lys His Val Pro Gly Gly Gly Ser Val Gln Ile Val Tyr Lys
1 5 10 15
<210> 72
<211> 31
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(31)
<223> IL-31 85-115 (S85 to C85, I115 to C115)
<400> 72
Cys Pro Ala Ile Arg Ala Tyr Leu Lys Thr Ile Arg Gln Leu Asp Asn
1 5 10 15
Lys Ser Val Ile Asp Glu Ile Ile Glu His Leu Asp Lys Leu Cys
20 25 30
<210> 73
<211> 25
<212> PRT
<213> Melanomma radicans
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> Неоантиген меланомы
<400> 73
Ser Gly Ser Pro Pro Leu Arg Val Ser Val Gly Asp Phe Ser Gln Glu
1 5 10 15
Phe Ser Pro Gln Glu Ala Gln Gln Asp
20 25
<210> 74
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Глиома
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> Гистон 3 вариант H3.3K27M для глиомы
<400> 74
Arg Met Ser Ala Pro Ser Thr Gly Gly Val
1 5 10
<210> 75
<211> 24
<212> PRT
<213> Искусственная последовательность
<220>
<223> KRAS Метастатический колоректальный рак
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> KRAS Метастатический колоректальный рак (МУТАНТ G-to-D)
<400> 75
Met Thr Glu Tyr Lys Leu Val Val Val Gly Ala Asp Gly Val Gly Lys
1 5 10 15
Ser Ala Leu Thr Ile Gln Leu Ile
20
<210> 76
<211> 32
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(32)
<223> HIV CTL Gag P24
<400> 76
Ser Ile Leu Asp Ile Lys Gln Gly Pro Lys Glu Pro Phe Arg Asp Tyr
1 5 10 15
Val Asp Arg Phe Tyr Lys Thr Leu Arg Ala Glu Gln Ala Ser Gln Glu
20 25 30
<210> 77
<211> 22
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> HIV CTL Gag P17
<400> 77
Ala Ser Arg Glu Leu Glu Arg Phe Ala Val Asn Pro Gly Leu Leu Glu
1 5 10 15
Thr Ser Glu Gly Cys Arg
20
<210> 78
<211> 20
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> HIV CTL Gag 293-312
<400> 78
Phe Arg Asp Tyr Val Asp Arg Phe Tyr Lys Thr Leu Arg Ala Glu Gln
1 5 10 15
Ala Ser Gln Glu
20
<210> 79
<211> 35
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(35)
<223> HIV CTL POL PRT
<400> 79
Pro Arg Thr Lys Met Ile Gly Gly Ile Gly Gly Phe Ile Lys Val Arg
1 5 10 15
Gln Tyr Asp Gln Ile Leu Ile Glu Cys Gly His Lys Ala Ile Gly Thr
20 25 30
Val Leu Val
35
<210> 80
<211> 26
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(26)
<223> HIV CTL POL RT
<400> 80
Leu Arg Trp Gly Phe Thr Thr Pro Asp Lys Lys His Gln Lys Glu Pro
1 5 10 15
Pro Phe Leu Trp Met Gly Tyr Glu Leu His
20 25
<210> 81
<211> 17
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> HIV CTL POL INT
<400> 81
Thr Lys Glu Leu Gln Lys Gln Ile Thr Lys Ile Gln Asn Phe Arg Val
1 5 10 15
Tyr
<210> 82
<211> 16
<212> PRT
<213> Вирус иммунодефицита человека
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> HIV CTL VIF
<400> 82
Cys Phe Ala Asp Ser Ala Ile Arg Lys Ala Ile Leu Gly His Ile Val
1 5 10 15
<210> 83
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL VP11/12 (66-74)
<400> 83
Phe Leu Thr Cys Thr Asp Arg Ser Val
1 5
<210> 84
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL VP11/12 (197-205)
<400> 84
Arg Ile Gln Gln Tyr Met Phe Phe Met
1 5
<210> 85
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL VP11/12 (220-228)
<400> 85
Arg Leu Asn Glu Leu Leu Ala Tyr Val
1 5
<210> 86
<211> 17
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> HSV CTL VP11/12 (230-246)
<400> 86
Val Leu Tyr Arg Trp Ala Ser Trp Met Leu Trp Thr Thr Asp Lys His
1 5 10 15
Val
<210> 87
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL VP11/12 (702-710)
<400> 87
Ala Leu Ser Ala Leu Leu Thr Lys Leu
1 5
<210> 88
<211> 8
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> HSV CTL gamma1 gB498
<400> 88
Ser Ser Ile Glu Phe Ala Arg Leu
1 5
<210> 89
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL gamma1 gB452
<400> 89
Tyr Gln Pro Leu Leu Ser Asn Thr Leu
1 5
<210> 90
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL gamma1 gB560
<400> 90
Ser Ala Arg Met Leu Gly Asp Val Met
1 5
<210> 91
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL gamma1 gG4
<400> 91
Gly Ala Met Arg Ala Val Val Pro Ile
1 5
<210> 92
<211> 8
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> HSV CTL beta PR1982
<400> 92
Phe Ala Pro Leu Phe Thr Asn Leu
1 5
<210> 93
<211> 8
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> HSV CTL beta PR1_822
<400> 93
Gln Thr Phe Asp Phe Gly Arg Leu
1 5
<210> 94
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL beta PR1_372
<400> 94
Phe Gly Leu Leu Asn Tyr Ala Leu Val
1 5
<210> 95
<211> 8
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> HSV CTL beta ICP_876
<400> 95
Gly Ala Ile Asn Phe Ile Asn Leu
1 5
<210> 96
<211> 11
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> HSV CTL beta ICP_168
<400> 96
Ala Val Cys Ile Asn Asn Thr Phe Leu His Leu
1 5 10
<210> 97
<211> 10
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> HSV CTL beta PR2_81
<400> 97
Ser Phe Tyr Arg Phe Leu Phe Ala Phe Leu
1 5 10
<210> 98
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL beta PR2_279
<400> 98
Ala Ala Ile Glu Asn Tyr Val Arg Phe
1 5
<210> 99
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL beta UL9_251
<400> 99
Phe Leu Pro Arg Leu Gly Thr Glu Leu
1 5
<210> 100
<211> 8
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> HSV CTL beta UL41_181
<400> 100
Leu Gly Tyr Ala Tyr Ile Asn Ser
1 5
<210> 101
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL gamma2 gC8
<400> 101
Leu Ala Val Val Leu Trp Ser Leu Leu
1 5
<210> 102
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL beta UL28_629
<400> 102
Tyr Ser Val Glu Asn Val Gly Leu Leu
1 5
<210> 103
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL gamma2 gH391
<400> 103
Phe Ala Phe Val Asn Ala Ala His Ala
1 5
<210> 104
<211> 9
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> HSV CTL gamma2 gK54
<400> 104
Trp Met Lys Met Asn Gln Thr Leu Leu
1 5
<210> 105
<211> 20
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> HSV CTL gB161-180
<400> 105
Ala Thr Met Tyr Tyr Lys Asp Val Thr Val Ser Gln Val Trp Phe Gly
1 5 10 15
His Arg Tyr Ser
20
<210> 106
<211> 15
<212> PRT
<213> вирус герпеса человека 1
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> HSV CTL gB 666-680
<400> 106
Thr Val Ser Thr Phe Ile Asp Leu Asn Ile Thr Met Leu Glu Asp
1 5 10 15
<210> 107
<211> 8
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> FMDV CTL L_65
<400> 107
Glu Pro Phe Phe Asp Trp Val Tyr
1 5
<210> 108
<211> 9
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> FMDV CTL VP4_227
<400> 108
Tyr Met Gln Gln Tyr Gln Asn Ser Met
1 5
<210> 109
<211> 9
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> FMDV CTL VP2_314
<400> 109
Ser Ser Val Gly Val Thr Tyr Gly Tyr
1 5
<210> 110
<211> 8
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> FMDV CTL VP2_346
<400> 110
Arg Phe Phe Lys Thr His Leu Phe
1 5
<210> 111
<211> 11
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> FMDV CTL VP2_385
<400> 111
Ala Tyr Met Arg Asn Gly Trp Asp Val Glu Val
1 5 10
<210> 112
<211> 13
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> FMDV CTL VP2_421
<400> 112
Arg Glu Leu Tyr Gln Leu Thr Leu Phe Pro His Gln Phe
1 5 10
<210> 113
<211> 8
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> FMDV CTL VP3_622
<400> 113
Lys Ala Arg Tyr Met Ile Ala Tyr
1 5
<210> 114
<211> 8
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(8)
<223> FMDV CTL 2C1309
<400> 114
Ile Ile Ala Thr Thr Asn Leu Tyr
1 5
<210> 115
<211> 11
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> FMDV CTL 2C1369
<400> 115
Phe Gln Tyr Asp Cys Ala Leu Leu Asn Gly Met
1 5 10
<210> 116
<211> 10
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> FMDV CTL 3C1720
<400> 116
Met Leu Ser Asp Ala Ala Leu Met Val Leu
1 5 10
<210> 117
<211> 19
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> FMDV CTL 3D2083
<400> 117
Trp Gln Arg Phe Gly Thr His Phe Ala Gln Tyr Arg Asn Val Trp Asp
1 5 10 15
Val Asp Tyr
<210> 118
<211> 11
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> FMDV CTL 3D2159
<400> 118
Asn Thr Ile Leu Asn Asn Ile Tyr Val Leu Tyr
1 5 10
<210> 119
<211> 31
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(31)
<223> FMDV CTL 3D2231/43/58
<400> 119
Ser Ile Thr Asp Val Thr Phe Leu Lys Lys Lys His Met Asp Tyr Gly
1 5 10 15
Thr Gly Phe Tyr Lys Lys Lys Thr Leu Glu Ala Ile Leu Ser Phe
20 25 30
<210> 120
<211> 17
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> FMDV CTL 3D2295
<400> 120
Phe Glu Pro Phe Gln Gly Leu Phe Glu Ile Pro Ser Tyr Arg Ser Leu
1 5 10 15
Tyr
<210> 121
<211> 10
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> FMDV CTL 3D2302
<400> 121
Phe Glu Ile Pro Ser Tyr Arg Ser Leu Tyr
1 5 10
<210> 122
<211> 9
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> FMDV CTL Тип O 750
<400> 122
Arg Arg Gln His Thr Asp Val Ser Phe
1 5
<210> 123
<211> 9
<212> PRT
<213> Вирус ящура
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> FMDV CTL Тип O 881
<400> 123
Arg Thr Leu Pro Thr Ser Phe Asn Tyr
1 5
<210> 124
<211> 14
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> PRRSV CTL ORF1a
<400> 124
Arg Leu Gly Lys Ile Ile Ser Leu Cys Gln Val Ile Glu Asp
1 5 10
<210> 125
<211> 15
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> PRRSV CTL ORF1a
<400> 125
Val Pro Val Ile Thr Cys Gly Val Val His Leu Leu Ala Ile Ile
1 5 10 15
<210> 126
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL ORF7
<400> 126
Leu Ser Asp Ser Gly Arg Ile Ser Tyr
1 5
<210> 127
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL ORF2
<400> 127
Arg Thr Ala Pro Asn Glu Ile Ala Phe
1 5
<210> 128
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL ORF2a
<400> 128
Ala Ser Asp Trp Phe Ala Pro Arg Tyr
1 5
<210> 129
<211> 10
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> PRRSV CTL ORF5
<400> 129
Met Ser Trp Arg Tyr Ser Cys Thr Arg Tyr
1 5 10
<210> 130
<211> 10
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> PRRSV CTL NSP9
<400> 130
Thr Thr Met Pro Ser Gly Phe Glu Leu Tyr
1 5 10
<210> 131
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL NSP10
<400> 131
Asn Ser Phe Leu Asp Glu Ala Ala Tyr
1 5
<210> 132
<211> 10
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> PRRSV CTL ORF6
<400> 132
Arg Gly Arg Leu Leu Gly Leu Leu His Leu
1 5 10
<210> 133
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL ORF5
<400> 133
Leu Tyr Arg Trp Arg Ser Pro Val Ile
1 5
<210> 134
<211> 10
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> PRRSV CTL NSP9
<400> 134
Met Pro Asn Tyr His Trp Trp Val Glu His
1 5 10
<210> 135
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL NSP9
<400> 135
Glu Val Ala Leu Ser Ala Gln Ile Ile
1 5
<210> 136
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL Тип 1_4596
<400> 136
Cys Leu Phe Ala Ile Leu Leu Ala Thr
1 5
<210> 137
<211> 12
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(12)
<223> PRRSV CTL Тип 1_4721
<400> 137
Cys Ala Phe Ala Ala Phe Val Cys Phe Val Ile Arg
1 5 10
<210> 138
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL Тип 1_5028
<400> 138
Lys Pro Glu Lys Pro His Phe Pro Leu
1 5
<210> 139
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL Тип 1_5083
<400> 139
Phe Met Leu Pro Val Ala His Thr Val
1 5
<210> 140
<211> 9
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> PRRSV CTL Тип 2_2702
<400> 140
Thr Met Pro Pro Gly Phe Glu Leu Tyr
1 5
<210> 141
<211> 15
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> PRRSV CTL Тип 2_4765
<400> 141
Leu Ala Ala Leu Ile Cys Phe Val Ile Arg Leu Ala Lys Asn Cys
1 5 10 15
<210> 142
<211> 16
<212> PRT
<213> Вирус репродуктивного и респираторного синдрома свиней
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> PRRSV CTL Тип 2_4797
<400> 142
Lys Gly Arg Leu Tyr Arg Trp Arg Ser Pro Val Ile Ile Val Glu Lys
1 5 10 15
<210> 143
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вирус классической чумы свиней
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> CSFV CTL Тип 1_1446
<400> 143
Lys His Lys Val Arg Asn Glu Val Met Val His Trp Phe Asp Asp
1 5 10 15
<210> 144
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вирус классической чумы свиней
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> CSFV CTL Тип 1_2276
<400> 144
Glu Asn Ala Leu Leu Val Ala Leu Phe
1 5
<210> 145
<211> 11
<212> PRT
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> IL-6 73-83
<400> 145
Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
1 5 10
<210> 146
<211> 32
<212> DNA
<213> Искусственная последовательность
<220>
<223> CpG1 олигонуклеотид ODN
<400> 146
tcgtcgtttt gtcgttttgt cgttttgtcg tt 32
<210> 147
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> CpG2 олигонуклеотид ODN
<220>
<221> misc_feature
<222> (1)..(1)
<223> фосфоротиоатная группа
<400> 147
tcgtcgtttt gtcgttttgt cgtt 24
<210> 148
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> CpG3 олигонуклеотид ODN
<400> 148
tcgtcgtttt gtcgttttgt cgtt 24
<210> 149
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF5 Th (UBITh1)
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (24)..(45)
<223> альфа-синуклеин (G111-G132)
<400> 149
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 150
<211> 44
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg3 Th (UBITh2)
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (23)..(44)
<223> альфа-синуклеин (G111-G132)
<400> 150
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro
20 25 30
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 151
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF4 Th (UBITh3)
<220>
<221> САЙТ
<222> (4)..(4)
<223> S или T
<220>
<221> САЙТ
<222> (7)..(7)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> G или T
<220>
<221> САЙТ
<222> (12)..(12)
<223> H или T
<220>
<221> САЙТ
<222> (13)..(13)
<223> K или R
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (24)..(45)
<223> альфа-синуклеин (G111-G132)
<400> 151
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 152
<211> 43
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani1 Th
<220>
<221> ПЕПТИД
<222> (18)..(21)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(43)
<223> альфа-синуклеин (G111-G132)
<400> 152
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp
20 25 30
Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 153
<211> 41
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин (G111-G132)
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF1 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(41)
<223> альфа-синуклеин (G111-G132)
<400> 153
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val Lys
1 5 10 15
Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 154
<211> 50
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> Bordetella pertussis Th
<220>
<221> ПЕПТИД
<222> (25)..(28)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (25)..(25)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (29)..(50)
<223> альфа-синуклеин (G111-G132)
<400> 154
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu Lys Lys Lys Lys Gly Ile Leu Glu
20 25 30
Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu
35 40 45
Glu Gly
50
<210> 155
<211> 43
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani2 Th
<220>
<221> ПЕПТИД
<222> (18)..(21)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(43)
<223> альфа-синуклеин (G111-G132)
<400> 155
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp
20 25 30
Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 156
<211> 49
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(23)
<223> Diphtheria Th
<220>
<221> ПЕПТИД
<222> (24)..(27)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (24)..(24)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (28)..(49)
<223> альфа-синуклеин (G111-G132)
<400> 156
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys Lys Lys Lys Lys Gly Ile Leu Glu Asp
20 25 30
Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu
35 40 45
Gly
<210> 157
<211> 47
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Plasmodium falciparum Th
<220>
<221> ПЕПТИД
<222> (22)..(25)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (26)..(47)
<223> альфа-синуклеин (G111-G132)
<400> 157
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro
20 25 30
Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 158
<211> 43
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Schistosoma mansoni Th
<220>
<221> ПЕПТИД
<222> (18)..(21)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(43)
<223> альфа-синуклеин (G111-G132)
<400> 158
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp
20 25 30
Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 159
<211> 51
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> Cholera Toxin Th
<220>
<221> ПЕПТИД
<222> (26)..(29)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (26)..(26)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (30)..(51)
<223> альфа-синуклеин (G111-G132)
<400> 159
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser Lys Lys Lys Lys Gly Ile Leu
20 25 30
Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser
35 40 45
Glu Glu Gly
50
<210> 160
<211> 41
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF2 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(41)
<223> альфа-синуклеин (G111-G132)
<400> 160
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile Lys
1 5 10 15
Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 161
<211> 48
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF3 Th
<220>
<221> САЙТ
<222> (7)..(7)
<223> S или T
<220>
<221> САЙТ
<222> (10)..(10)
<223> K или R
<220>
<221> САЙТ
<222> (11)..(11)
<223> G или T
<220>
<221> САЙТ
<222> (15)..(15)
<223> H или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> K или R
<220>
<221> САЙТ
<222> (19)..(19)
<223> G или T
<220>
<221> САЙТ
<222> (23)..(23)
<223> эпсилон K как спейсер
<220>
<221> ПЕПТИД
<222> (23)..(26)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (27)..(48)
<223> альфа-синуклеин 111-132
<220>
<221> ПЕПТИД
<222> (27)..(48)
<223> альфа-синуклеин (G111-G132)
<400> 161
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met
20 25 30
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 162
<211> 44
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 1 Th
<220>
<221> САЙТ
<222> (1)..(1)
<223> K или R
<220>
<221> САЙТ
<222> (2)..(2)
<223> K или R
<220>
<221> САЙТ
<222> (3)..(3)
<223> K или R
<220>
<221> САЙТ
<222> (4)..(4)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> F или K или R
<220>
<221> САЙТ
<222> (6)..(6)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (7)..(7)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (9)..(9)
<223> K или R
<220>
<221> САЙТ
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (11)..(11)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или L или I или V или F
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (18)..(18)
<223> D или R
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон K как спейсер
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (23)..(44)
<223> альфа-синуклеин 111-132
<220>
<221> ПЕПТИД
<222> (23)..(44)
<223> альфа-синуклеин (G111-G132)
<400> 162
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro
20 25 30
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 163
<211> 44
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 2 Th
<220>
<221> САЙТ
<222> (4)..(4)
<223> I или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> I или F
<220>
<221> САЙТ
<222> (6)..(6)
<223> T или L
<220>
<221> САЙТ
<222> (7)..(7)
<223> I или L
<220>
<221> САЙТ
<222> (11)..(11)
<223> I или L
<220>
<221> САЙТ
<222> (14)..(14)
<223> P или I
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> S или T
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон K как спейсер
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (23)..(44)
<223> альфа-синуклеин (G111-G132)
<400> 163
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro
20 25 30
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 164
<211> 37
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> Influenza MP1_1 Th
<220>
<221> ПЕПТИД
<222> (12)..(15)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (12)..(12)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (16)..(37)
<223> альфа-синуклеин (G111-G132)
<400> 164
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg Lys Lys Lys Lys Gly
1 5 10 15
Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met
20 25 30
Pro Ser Glu Glu Gly
35
<210> 165
<211> 41
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Influenza MP1_2 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(41)
<223> альфа-синуклеин (G111-G132)
<400> 165
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val Lys
1 5 10 15
Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 166
<211> 35
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> Influenza NSP1 Th
<220>
<221> ПЕПТИД
<222> (10)..(13)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (10)..(10)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (14)..(35)
<223> альфа-синуклеин (G111-G132)
<400> 166
Asp Arg Leu Arg Arg Asp Gln Lys Ser Lys Lys Lys Lys Gly Ile Leu
1 5 10 15
Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser
20 25 30
Glu Glu Gly
35
<210> 167
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> EBV BHRF1 Th
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(45)
<223> альфа-синуклеин (G111-G132)
<400> 167
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 168
<211> 41
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Clostridium tetani TT1 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(41)
<223> альфа-синуклеин (G111-G132)
<400> 168
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu Lys
1 5 10 15
Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 169
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> EBV EBNA-1 Th
<220>
<221> ПЕПТИД
<222> (21)..(24)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (21)..(21)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (25)..(46)
<223> альфа-синуклеин (G111-G132)
<400> 169
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val
20 25 30
Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 170
<211> 47
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Clostridium tetani TT2 Th
<220>
<221> ПЕПТИД
<222> (22)..(25)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (26)..(47)
<223> альфа-синуклеин (G111-G132)
<400> 170
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro
20 25 30
Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210> 171
<211> 42
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT3 Th
<220>
<221> ПЕПТИД
<222> (17)..(20)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (17)..(17)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (21)..(42)
<223> альфа-синуклеин (G111-G132)
<400> 171
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn
20 25 30
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 172
<211> 42
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT4 Th
<220>
<221> ПЕПТИД
<222> (17)..(20)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (17)..(17)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (21)..(42)
<223> альфа-синуклеин (G111-G132)
<400> 172
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn
20 25 30
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 173
<211> 44
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> EBV CP Th
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(44)
<223> альфа-синуклеин (G111-G132)
<400> 173
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro
20 25 30
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 174
<211> 40
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HCMV IE1 Th
<220>
<221> ПЕПТИД
<222> (15)..(18)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(40)
<223> альфа-синуклеин (G111-G132)
<400> 174
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His Lys Lys
1 5 10 15
Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala
20 25 30
Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 175
<211> 41
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> EBV GP340 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(41)
<223> альфа-синуклеин (G111-G132)
<400> 175
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr Lys
1 5 10 15
Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210> 176
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> EBV BPLF1 Th
<220>
<221> ПЕПТИД
<222> (14)..(17)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (14)..(14)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (18)..(39)
<223> альфа-синуклеин (G111-G132)
<400> 176
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln Lys Lys Lys
1 5 10 15
Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
20 25 30
Glu Met Pro Ser Glu Glu Gly
35
<210> 177
<211> 37
<212> PRT
<213> Искусственная последовательность
<220>
<223> альфа-синуклеин Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> EBV EBNA-2 Th
<220>
<221> ПЕПТИД
<222> (12)..(15)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (12)..(12)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (16)..(37)
<223> альфа-синуклеин (G111-G132)
<400> 177
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu Lys Lys Lys Lys Gly
1 5 10 15
Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met
20 25 30
Pro Ser Glu Glu Gly
35
<210> 178
<211> 59
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(39)
<223> IgE-EMPD (G1-C39)
<220>
<221> САЙТ
<222> (40)..(40)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (41)..(59)
<223> MvF5 Th (UBITh1)
<400> 178
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
20 25 30
Ser Val Pro His Pro Arg Cys Lys Ile Ser Ile Thr Glu Ile Lys Gly
35 40 45
Val Ile Val His Arg Ile Glu Thr Ile Leu Phe
50 55
<210> 179
<211> 58
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg3 Th (UBITh2)
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD (G1-C39)
<400> 179
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 180
<211> 59
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 4 Th (UBITh3)
<220>
<221> САЙТ
<222> (4)..(4)
<223> S или T
<220>
<221> САЙТ
<222> (7)..(7)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> G или T
<220>
<221> САЙТ
<222> (12)..(12)
<223> H или T
<220>
<221> САЙТ
<222> (13)..(13)
<223> K или R
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (21)..(59)
<223> IgE-EMPD (G1-C39)
<400> 180
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
20 25 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg
35 40 45
Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 181
<211> 57
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani1 Th
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(57)
<223> IgE-EMPD (G1-C39)
<400> 181
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
20 25 30
Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
35 40 45
Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 182
<211> 55
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF1 Th
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD (G1-C39)
<400> 182
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 183
<211> 64
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> Bordetella pertussis Th
<220>
<221> САЙТ
<222> (25)..(25)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (26)..(64)
<223> IgE-EMPD (G1-C39)
<400> 183
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu Lys Gly Leu Ala Gly Gly Ser Ala
20 25 30
Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln
35 40 45
Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 184
<211> 57
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani2 Th
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(57)
<223> IgE-EMPD (G1-C39)
<400> 184
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
20 25 30
Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
35 40 45
Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 185
<211> 63
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(23)
<223> Diphtheria Th
<220>
<221> САЙТ
<222> (24)..(24)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (25)..(63)
<223> IgE-EMPD (G1-C39)
<400> 185
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys Lys Gly Leu Ala Gly Gly Ser Ala Gln
20 25 30
Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln
35 40 45
Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 186
<211> 61
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(21)
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(61)
<223> IgE-EMPD (G1-C39)
<400> 186
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln
20 25 30
Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu
35 40 45
Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 187
<211> 57
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Schistosoma mansoni Th
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(57)
<223> IgE-EMPD (G1-C39)
<400> 187
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
20 25 30
Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
35 40 45
Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 188
<211> 65
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> Cholera Toxin Th
<220>
<221> САЙТ
<222> (26)..(26)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (27)..(65)
<223> IgE-EMPD (G1-C39)
<400> 188
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser Lys Gly Leu Ala Gly Gly Ser
20 25 30
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
35 40 45
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
50 55 60
Cys
65
<210> 189
<211> 55
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF2 Th
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD (G1-C39)
<400> 189
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 190
<211> 62
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF3 Th
<220>
<221> САЙТ
<222> (7)..(7)
<223> S или T
<220>
<221> САЙТ
<222> (10)..(10)
<223> K или R
<220>
<221> САЙТ
<222> (11)..(11)
<223> G или T
<220>
<221> САЙТ
<222> (15)..(15)
<223> H или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> K или R
<220>
<221> САЙТ
<222> (19)..(19)
<223> G или T
<220>
<221> САЙТ
<222> (23)..(23)
<223> эпсилон K как спейсер
<220>
<221> ПЕПТИД
<222> (24)..(62)
<223> IgE-EMPD (G1-C39)
<400> 190
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser
20 25 30
Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly
35 40 45
Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 191
<211> 58
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 1 Th
<220>
<221> САЙТ
<222> (1)..(1)
<223> K или R
<220>
<221> САЙТ
<222> (2)..(2)
<223> K или R
<220>
<221> САЙТ
<222> (3)..(3)
<223> K или R
<220>
<221> САЙТ
<222> (4)..(4)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> F или K или R
<220>
<221> САЙТ
<222> (6)..(6)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (7)..(7)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (9)..(9)
<223> K или R
<220>
<221> САЙТ
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (11)..(11)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или L или I или V или F
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (18)..(18)
<223> D или R
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон K как спейсер
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD (G1-C39)
<400> 191
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 192
<211> 58
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 2 Th
<220>
<221> САЙТ
<222> (4)..(4)
<223> I или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> I или F
<220>
<221> САЙТ
<222> (6)..(6)
<223> T или L
<220>
<221> САЙТ
<222> (7)..(7)
<223> I или L
<220>
<221> САЙТ
<222> (11)..(11)
<223> I или L
<220>
<221> САЙТ
<222> (14)..(14)
<223> P или I
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> S или T
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон K как спейсер
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD (G1-C39)
<400> 192
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 193
<211> 51
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> Influenza MP1_1 Th
<220>
<221> САЙТ
<222> (12)..(12)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (13)..(51)
<223> IgE-EMPD (G1-C39)
<400> 193
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg Lys Gly Leu Ala Gly
1 5 10 15
Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser
20 25 30
Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His
35 40 45
Pro Arg Cys
50
<210> 194
<211> 55
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Influenza MP1_2 Th
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD (G1-C39)
<400> 194
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 195
<211> 49
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> Influenza NSP1 Th
<220>
<221> САЙТ
<222> (10)..(10)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (11)..(49)
<223> IgE-EMPD (G1-C39)
<400> 195
Asp Arg Leu Arg Arg Asp Gln Lys Ser Lys Gly Leu Ala Gly Gly Ser
1 5 10 15
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
20 25 30
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
35 40 45
Cys
<210> 196
<211> 59
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> EBV BHRF1 Th
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (21)..(59)
<223> IgE-EMPD (G1-C39)
<400> 196
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
20 25 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg
35 40 45
Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 197
<211> 55
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Clostridium tetani TT1 Th
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD (G1-C39)
<400> 197
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 198
<211> 60
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> EBV EBNA-1 Th
<220>
<221> САЙТ
<222> (21)..(21)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(60)
<223> IgE-EMPD (G1-C39)
<400> 198
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg
20 25 30
Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro
35 40 45
Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 199
<211> 61
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Clostridium tetani TT2 Th
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(61)
<223> IgE-EMPD (G1-C39)
<400> 199
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln
20 25 30
Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu
35 40 45
Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 200
<211> 56
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT3 Th
<220>
<221> САЙТ
<222> (17)..(17)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (18)..(56)
<223> IgE-EMPD (G1-C39)
<400> 200
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg
20 25 30
Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly
35 40 45
Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 201
<211> 56
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT4 Th
<220>
<221> САЙТ
<222> (17)..(17)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (18)..(56)
<223> IgE-EMPD (G1-C39)
<400> 201
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg
20 25 30
Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly
35 40 45
Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 202
<211> 58
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> EBV CP Th
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD (G1-C39)
<400> 202
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 203
<211> 54
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HCMV IE1 Th
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (16)..(54)
<223> IgE-EMPD (G1-C39)
<400> 203
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His Lys Gly
1 5 10 15
Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu
20 25 30
Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser
35 40 45
Val Pro His Pro Arg Cys
50
<210> 204
<211> 55
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> EBV GP340 Th
<220>
<221> САЙТ
<222> (16)..(16)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD (G1-C39)
<400> 204
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 205
<211> 53
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> EBV BPLF1 Th
<220>
<221> САЙТ
<222> (14)..(14)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (15)..(53)
<223> IgE-EMPD (G1-C39)
<400> 205
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln Lys Gly Leu
1 5 10 15
Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys
20 25 30
His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val
35 40 45
Pro His Pro Arg Cys
50
<210> 206
<211> 51
<212> PRT
<213> Искусственная последовательность
<220>
<223> IgE-EMPD Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> EBV EBNA-2 Th
<220>
<221> САЙТ
<222> (12)..(12)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (13)..(51)
<223> IgE-EMPD (G1-C39)
<400> 206
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu Lys Gly Leu Ala Gly
1 5 10 15
Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser
20 25 30
Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His
35 40 45
Pro Arg Cys
50
<210> 207
<211> 34
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF5 Th (UBITh1)
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (24)..(34)
<223> IL6 (C73-C83)
<400> 207
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu
20 25 30
Thr Cys
<210> 208
<211> 33
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg3 Th (UBITh2)
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> эпсилон-K-KKK спейсер
<220>
<221> ПЕПТИД
<222> (23)..(33)
<223> IL6 (C73-C83)
<400> 208
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr
20 25 30
Cys
<210> 209
<211> 34
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 4 Th (UBITh3)
<220>
<221> САЙТ
<222> (4)..(4)
<223> S или T
<220>
<221> САЙТ
<222> (7)..(7)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> G или T
<220>
<221> САЙТ
<222> (12)..(12)
<223> H или T
<220>
<221> САЙТ
<222> (13)..(13)
<223> K или R
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон-K-KKK спейсер
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(34)
<223> IL6 (C73-C83)
<400> 209
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu
20 25 30
Thr Cys
<210> 210
<211> 32
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani1 Th
<220>
<221> ПЕПТИД
<222> (18)..(21)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (21)..(21)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(32)
<223> IL6 (C73-C83)
<400> 210
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 211
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF1 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(30)
<223> IL6 (C73-C83)
<400> 211
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val Lys
1 5 10 15
Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 212
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> Bordetella pertussis Th
<220>
<221> ПЕПТИД
<222> (25)..(28)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (28)..(28)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (29)..(39)
<223> IL6 (C73-C83)
<400> 212
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu Lys Lys Lys Lys Cys Phe Gln Ser
20 25 30
Gly Phe Asn Glu Glu Thr Cys
35
<210> 213
<211> 32
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani2 Th
<220>
<221> ПЕПТИД
<222> (18)..(21)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (21)..(21)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(32)
<223> IL6 (C73-C83)
<400> 213
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 214
<211> 38
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(23)
<223> Diphtheria Th
<220>
<221> ПЕПТИД
<222> (24)..(27)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (27)..(27)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (28)..(38)
<223> IL6 (C73-C83)
<400> 214
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys Lys Lys Lys Lys Cys Phe Gln Ser Gly
20 25 30
Phe Asn Glu Glu Thr Cys
35
<210> 215
<211> 36
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Plasmodium falciparum Th
<220>
<221> ПЕПТИД
<222> (22)..(25)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (25)..(25)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (26)..(36)
<223> IL6 (C73-C83)
<400> 215
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn
20 25 30
Glu Glu Thr Cys
35
<210> 216
<211> 32
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Schistosoma mansoni Th
<220>
<221> ПЕПТИД
<222> (18)..(21)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (21)..(21)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (22)..(32)
<223> IL6 (C73-C83)
<400> 216
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 217
<211> 40
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> Cholera Toxin Th
<220>
<221> ПЕПТИД
<222> (26)..(29)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (29)..(29)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (30)..(40)
<223> IL6 (C73-C83)
<400> 217
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser Lys Lys Lys Lys Cys Phe Gln
20 25 30
Ser Gly Phe Asn Glu Glu Thr Cys
35 40
<210> 218
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF2 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(30)
<223> IL6 (C73-C83)
<400> 218
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile Lys
1 5 10 15
Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 219
<211> 37
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF3 Th
<220>
<221> САЙТ
<222> (7)..(7)
<223> S или T
<220>
<221> САЙТ
<222> (10)..(10)
<223> K или R
<220>
<221> САЙТ
<222> (11)..(11)
<223> G или T
<220>
<221> САЙТ
<222> (15)..(15)
<223> H или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> K или R
<220>
<221> САЙТ
<222> (19)..(19)
<223> G или T
<220>
<221> ПЕПТИД
<222> (23)..(26)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (26)..(26)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (27)..(37)
<223> IL6 (C73-C83)
<400> 219
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe
20 25 30
Asn Glu Glu Thr Cys
35
<210> 220
<211> 33
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 1 Th
<220>
<221> САЙТ
<222> (1)..(1)
<223> K или R
<220>
<221> САЙТ
<222> (2)..(2)
<223> K или R
<220>
<221> САЙТ
<222> (3)..(3)
<223> K или R
<220>
<221> САЙТ
<222> (4)..(4)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> F или K или R
<220>
<221> САЙТ
<222> (6)..(6)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (7)..(7)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (9)..(9)
<223> K или R
<220>
<221> САЙТ
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (11)..(11)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или L или I или V или F
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I или V или F
<220>
<221> САЙТ
<222> (18)..(18)
<223> D или R
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(33)
<223> IL6 (C73-C83)
<400> 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr
20 25 30
Cys
<210> 221
<211> 33
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 2 Th
<220>
<221> САЙТ
<222> (4)..(4)
<223> I или F
<220>
<221> САЙТ
<222> (5)..(5)
<223> I или F
<220>
<221> САЙТ
<222> (6)..(6)
<223> T или L
<220>
<221> САЙТ
<222> (7)..(7)
<223> I или L
<220>
<221> САЙТ
<222> (11)..(11)
<223> I или L
<220>
<221> САЙТ
<222> (14)..(14)
<223> P или I
<220>
<221> САЙТ
<222> (15)..(15)
<223> Q или T
<220>
<221> САЙТ
<222> (16)..(16)
<223> S или T
<220>
<221> САЙТ
<222> (17)..(17)
<223> L или I
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(33)
<223> IL6 (C73-C83)
<400> 221
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr
20 25 30
Cys
<210> 222
<211> 26
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> Influenza MP1_1 Th
<220>
<221> ПЕПТИД
<222> (12)..(15)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (16)..(26)
<223> IL6 (C73-C83)
<400> 222
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg Lys Lys Lys Lys Cys
1 5 10 15
Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25
<210> 223
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Influenza MP1_2 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(30)
<223> IL6 (C73-C83)
<400> 223
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val Lys
1 5 10 15
Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 224
<211> 24
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> Influenza NSP1 Th
<220>
<221> ПЕПТИД
<222> (10)..(13)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (13)..(13)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (14)..(24)
<223> IL6 (C73-C83)
<400> 224
Asp Arg Leu Arg Arg Asp Gln Lys Ser Lys Lys Lys Lys Cys Phe Gln
1 5 10 15
Ser Gly Phe Asn Glu Glu Thr Cys
20
<210> 225
<211> 34
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> EBV BHRF1 Th
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (23)..(23)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(34)
<223> IL6 (C73-C83)
<400> 225
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu
20 25 30
Thr Cys
<210> 226
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Clostridium tetani TT1 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(30)
<223> IL6 (C73-C83)
<400> 226
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu Lys
1 5 10 15
Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 227
<211> 35
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> EBV EBNA-1 Th
<220>
<221> ПЕПТИД
<222> (21)..(24)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (24)..(24)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (25)..(35)
<223> IL6 (C73-C83)
<400> 227
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu
20 25 30
Glu Thr Cys
35
<210> 228
<211> 36
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Clostridium tetani TT2 Th
<220>
<221> ПЕПТИД
<222> (22)..(25)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (25)..(25)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (26)..(36)
<223> IL6 (C73-C83)
<400> 228
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn
20 25 30
Glu Glu Thr Cys
35
<210> 229
<211> 31
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT3 Th
<220>
<221> ПЕПТИД
<222> (17)..(20)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (21)..(31)
<223> IL6 (C73-C83)
<400> 229
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 230
<211> 31
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT4 Th
<220>
<221> ПЕПТИД
<222> (17)..(20)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (21)..(31)
<223> IL6 (C73-C83)
<400> 230
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 231
<211> 33
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> EBV CP Th
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (22)..(22)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(33)
<223> IL6 (C73-C83)
<400> 231
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu Lys Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr
20 25 30
Cys
<210> 232
<211> 29
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HCMV IE1 Th
<220>
<221> ПЕПТИД
<222> (15)..(18)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (18)..(18)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (19)..(29)
<223> IL6 (C73-C83)
<400> 232
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His Lys Lys
1 5 10 15
Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25
<210> 233
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> EBV GP340 Th
<220>
<221> ПЕПТИД
<222> (16)..(19)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (20)..(30)
<223> IL6 (C73-C83)
<400> 233
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr Lys
1 5 10 15
Lys Lys Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25 30
<210> 234
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> EBV BPLF1 Th
<220>
<221> ПЕПТИД
<222> (14)..(17)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (17)..(17)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (18)..(28)
<223> IL6 (C73-C83)
<400> 234
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln Lys Lys Lys
1 5 10 15
Lys Cys Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25
<210> 235
<211> 26
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> EBV EBNA-2 Th
<220>
<221> ПЕПТИД
<222> (12)..(15)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (15)..(15)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (16)..(26)
<223> IL6 (C73-C83)
<400> 235
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu Lys Lys Lys Lys Cys
1 5 10 15
Phe Gln Ser Gly Phe Asn Glu Glu Thr Cys
20 25
<210> 236
<211> 43
<212> PRT
<213> Искусственная последовательность
<220>
<223> DPR Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> (GA) 10 repeats
<220>
<221> ПЕПТИД
<222> (21)..(24)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (24)..(24)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (25)..(43)
<223> UBITh1
<400> 236
Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala
1 5 10 15
Gly Ala Gly Ala Lys Lys Lys Lys Ile Ser Ile Thr Glu Ile Lys Gly
20 25 30
Val Ile Val His Arg Ile Glu Thr Ile Leu Phe
35 40
<210> 237
<211> 53
<212> PRT
<213> Искусственная последовательность
<220>
<223> DPR Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(30)
<223> (GA) 15 повторов
<220>
<221> ПЕПТИД
<222> (31)..(34)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (34)..(34)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (35)..(53)
<223> UBITh1
<400> 237
Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala
1 5 10 15
Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Lys Lys
20 25 30
Lys Lys Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile
35 40 45
Glu Thr Ile Leu Phe
50
<210> 238
<211> 73
<212> PRT
<213> Искусственная последовательность
<220>
<223> DPR Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(50)
<223> (GA) 25 повторов
<220>
<221> ПЕПТИД
<222> (51)..(54)
<223> KKK-эпсилон-K спейсер
<220>
<221> САЙТ
<222> (54)..(54)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (55)..(73)
<223> UBITh1
<400> 238
Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala
1 5 10 15
Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala
20 25 30
Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala Gly Ala
35 40 45
Gly Ala Lys Lys Lys Lys Ile Ser Ile Thr Glu Ile Lys Gly Val Ile
50 55 60
Val His Arg Ile Glu Thr Ile Leu Phe
65 70
<210> 239
<211> 27
<212> PRT
<213> Искусственная последовательность
<220>
<223> LHRH Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> TT1 Th
<220>
<221> ПЕПТИД
<222> (18)..(27)
<223> LHRH
<400> 239
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu Glu His Trp Ser Tyr Gly Leu Arg Pro Gly
20 25
<210> 240
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> LHRH Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Инвазин
<220>
<221> САЙТ
<222> (17)..(18)
<223> GG спейсер
<220>
<221> ПЕПТИД
<222> (19)..(33)
<223> MVF Th
<220>
<221> САЙТ
<222> (34)..(35)
<223> GG спейсер
<220>
<221> ПЕПТИД
<222> (36)..(45)
<223> LHRH
<400> 240
Thr Ala Lys Ser Lys Lys Phe Pro Ser Tyr Thr Ala Thr Tyr Gln Phe
1 5 10 15
Gly Gly Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly
20 25 30
Val Gly Gly Glu His Trp Ser Tyr Gly Leu Arg Pro Gly
35 40 45
<210> 241
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> LHRH Пептидная иммуногенная конструкция
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Инвазин
<220>
<221> САЙТ
<222> (17)..(18)
<223> GG спейсер
<220>
<221> ПЕПТИД
<222> (19)..(33)
<223> HBsAg Th
<220>
<221> САЙТ
<222> (34)..(35)
<223> GG спейсер
<220>
<221> ПЕПТИД
<222> (36)..(45)
<223> LHRH
<400> 241
Thr Ala Lys Ser Lys Lys Phe Pro Ser Tyr Thr Ala Thr Tyr Gln Phe
1 5 10 15
Gly Gly Phe Phe Leu Leu Thr Arg Ile Leu Thr Ile Pro Gln Ser Leu
20 25 30
Glu Gly Gly Glu His Trp Ser Tyr Gly Leu Arg Pro Gly
35 40 45
<210> 242
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> IL-6 (72-82) (аналог крысы)
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> IL-6 (72-82) аналог крысы
<400> 242
Cys Phe Gln Thr Gly Tyr Asn Gln Glu Ile Cys
1 5 10
<210> 243
<211> 34
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 4 Th (UBITh3)
<220>
<221> САЙТ
<222> (4)..(4)
<223> S или T
<220>
<221> САЙТ
<222> (7)..(7)
<223> K или R
<220>
<221> САЙТ
<222> (8)..(8)
<223> G или T
<220>
<221> САЙТ
<222> (12)..(12)
<223> H или T
<220>
<221> САЙТ
<222> (13)..(13)
<223> K или R
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон K-KKK спейсер
<220>
<221> САЙТ
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(34)
<223> аналог крысы IL-6 (72-82)
<400> 243
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Cys Phe Gln Thr Gly Tyr Asn Gln Glu
20 25 30
Ile Cys
<---

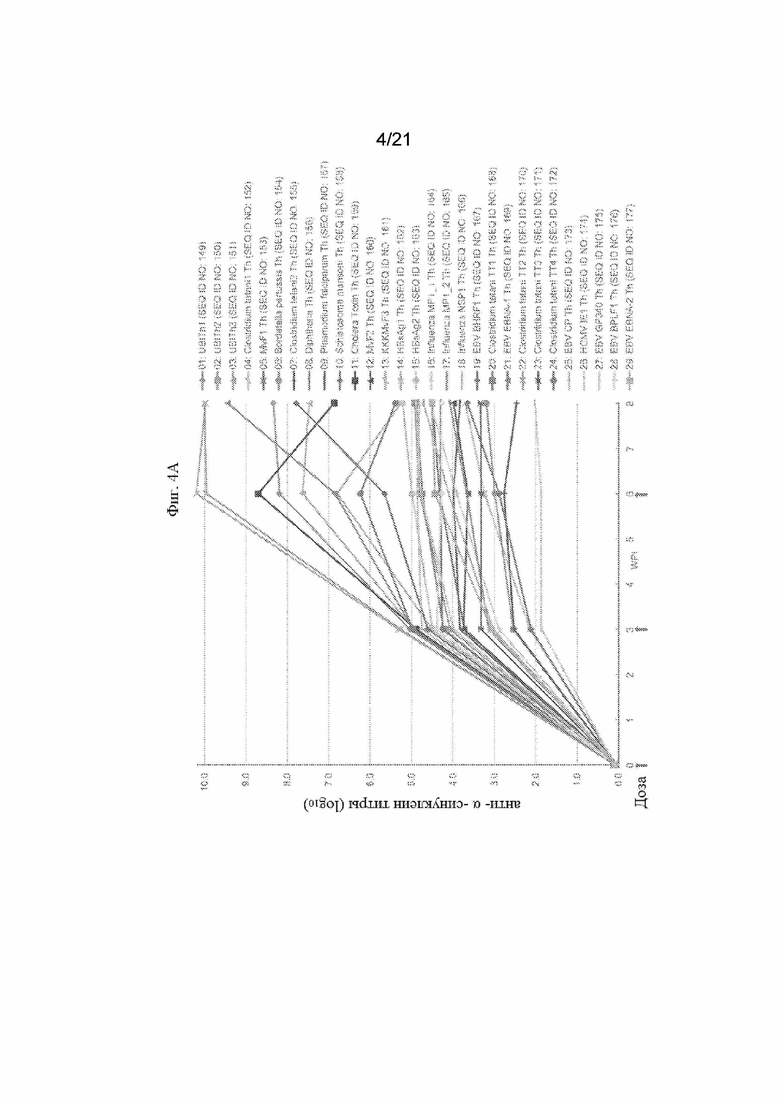
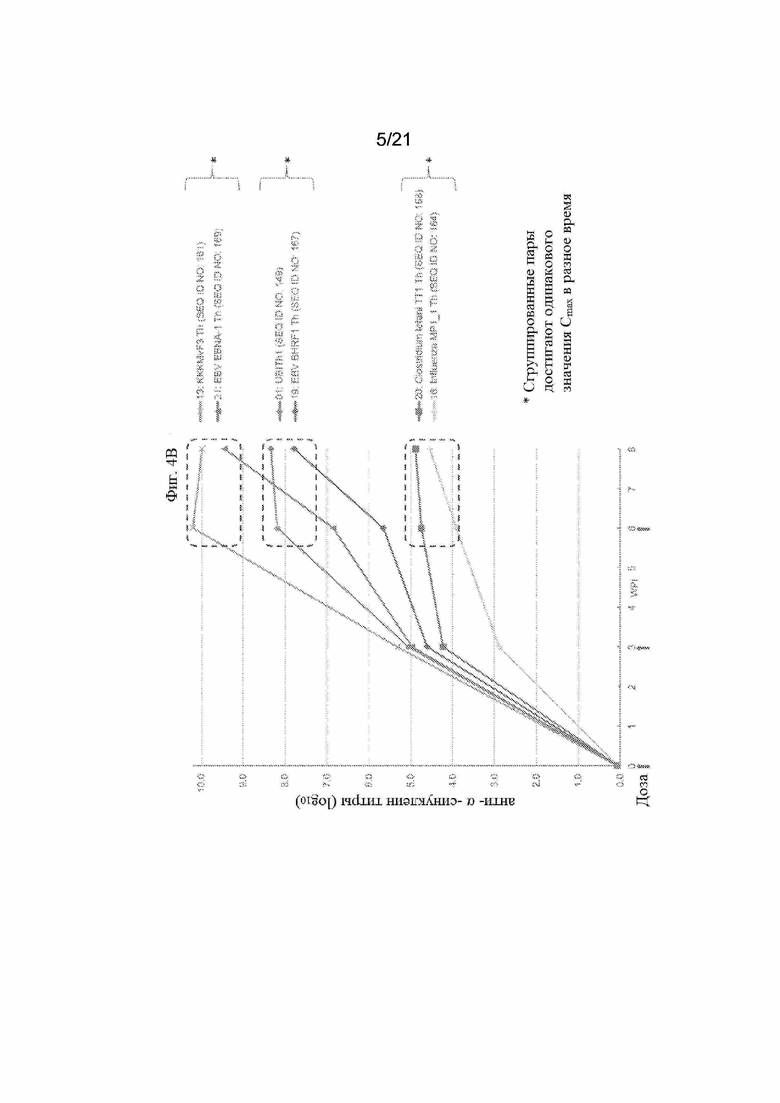
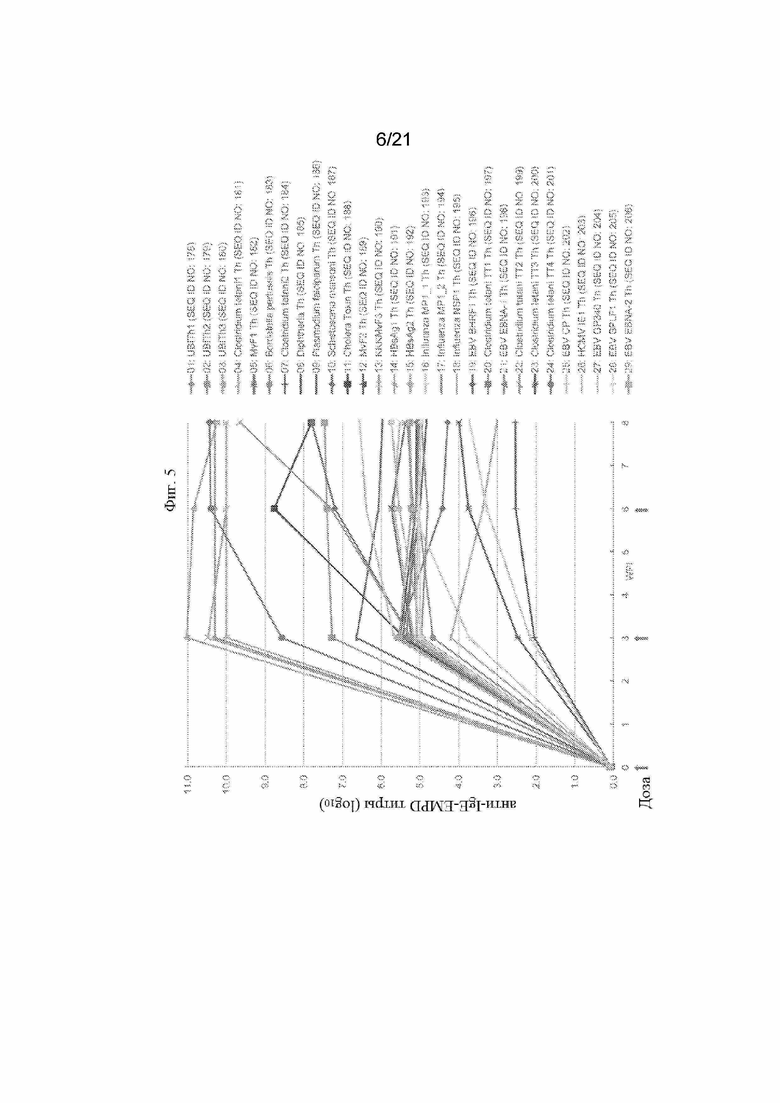
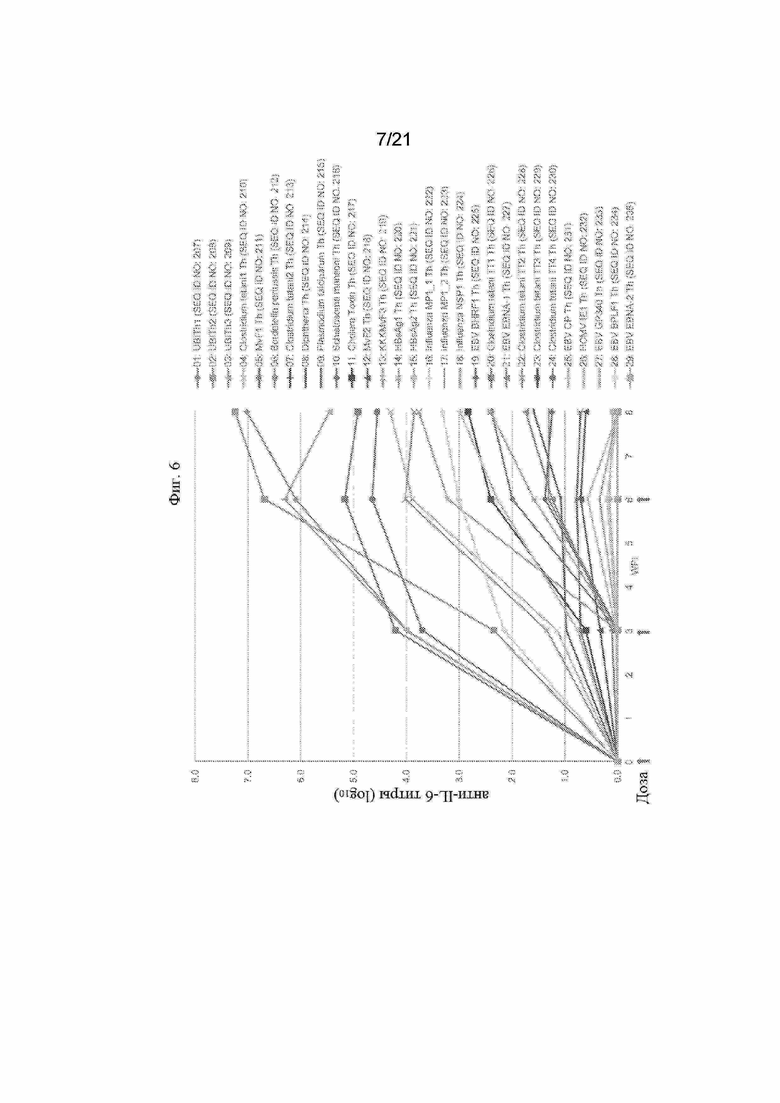
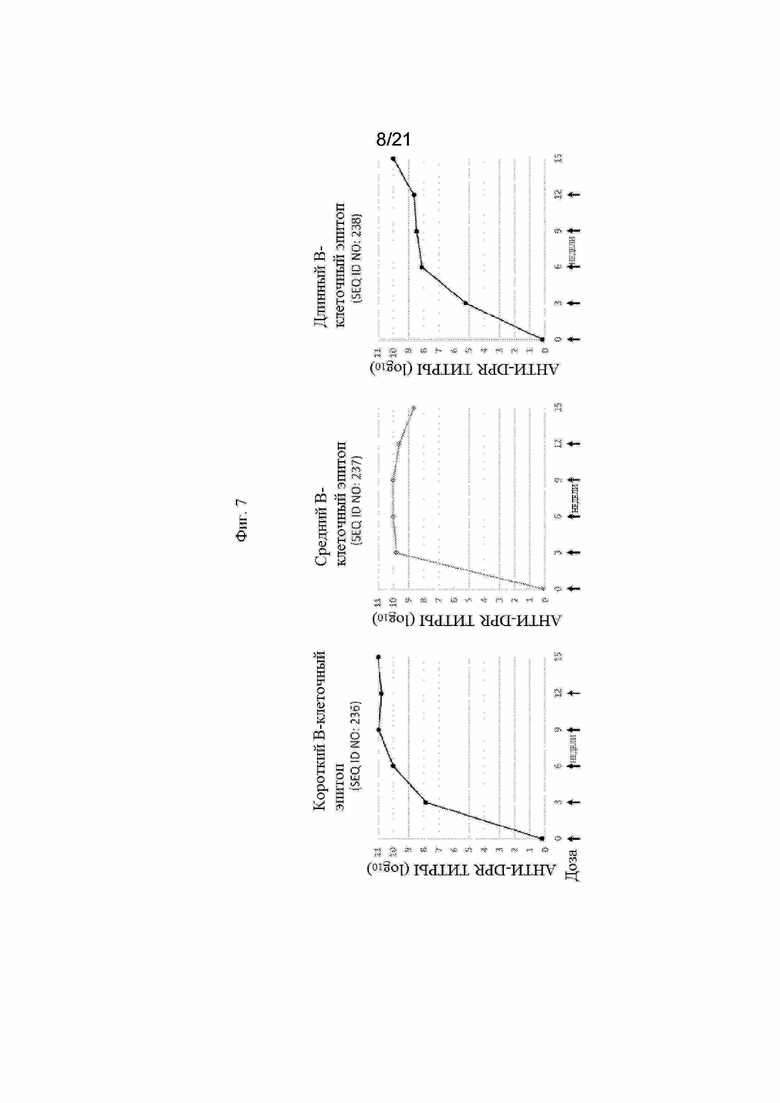
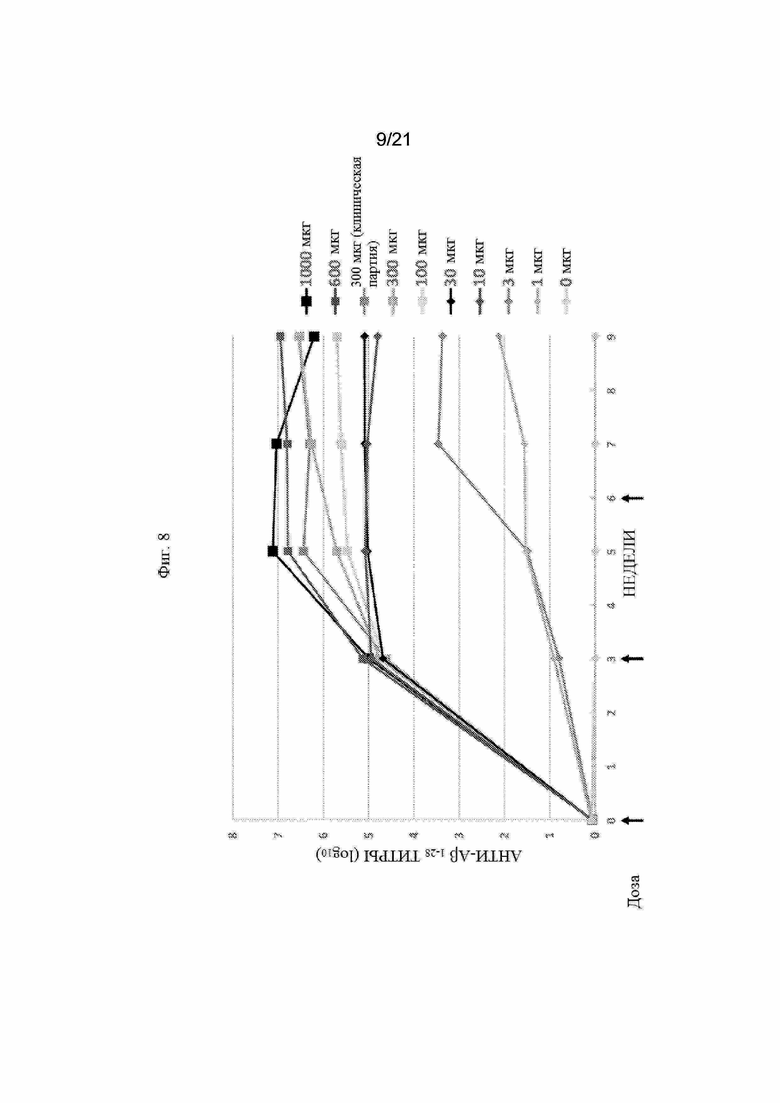
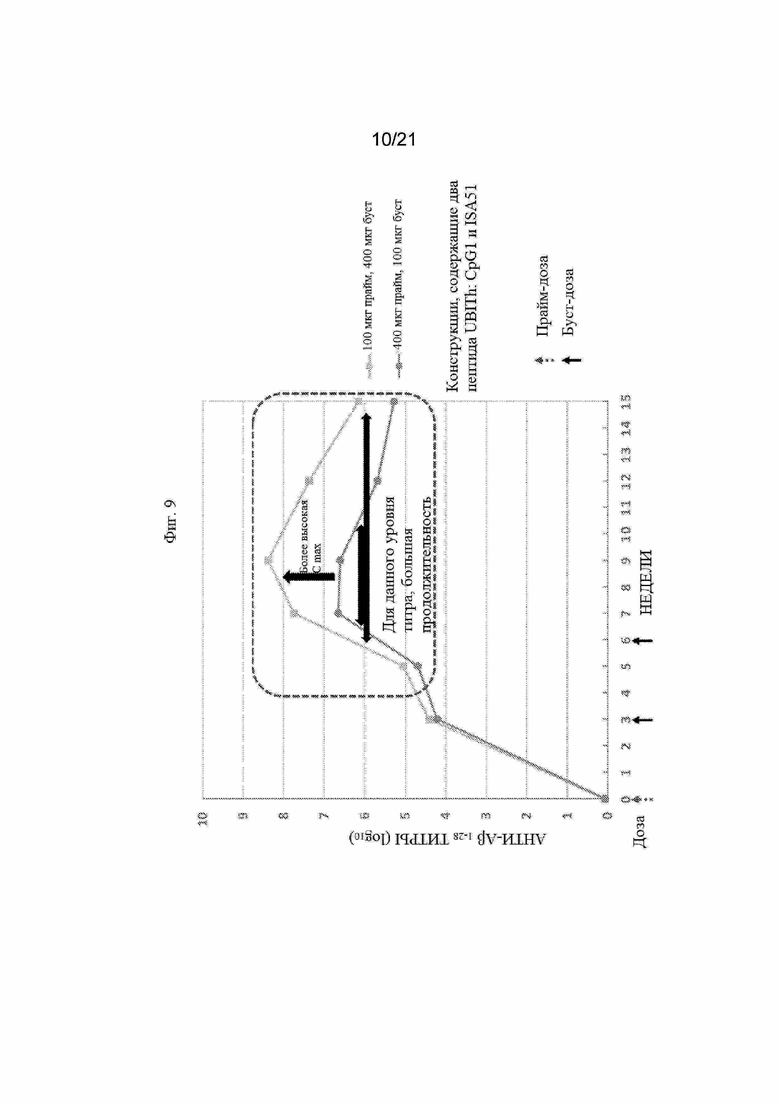
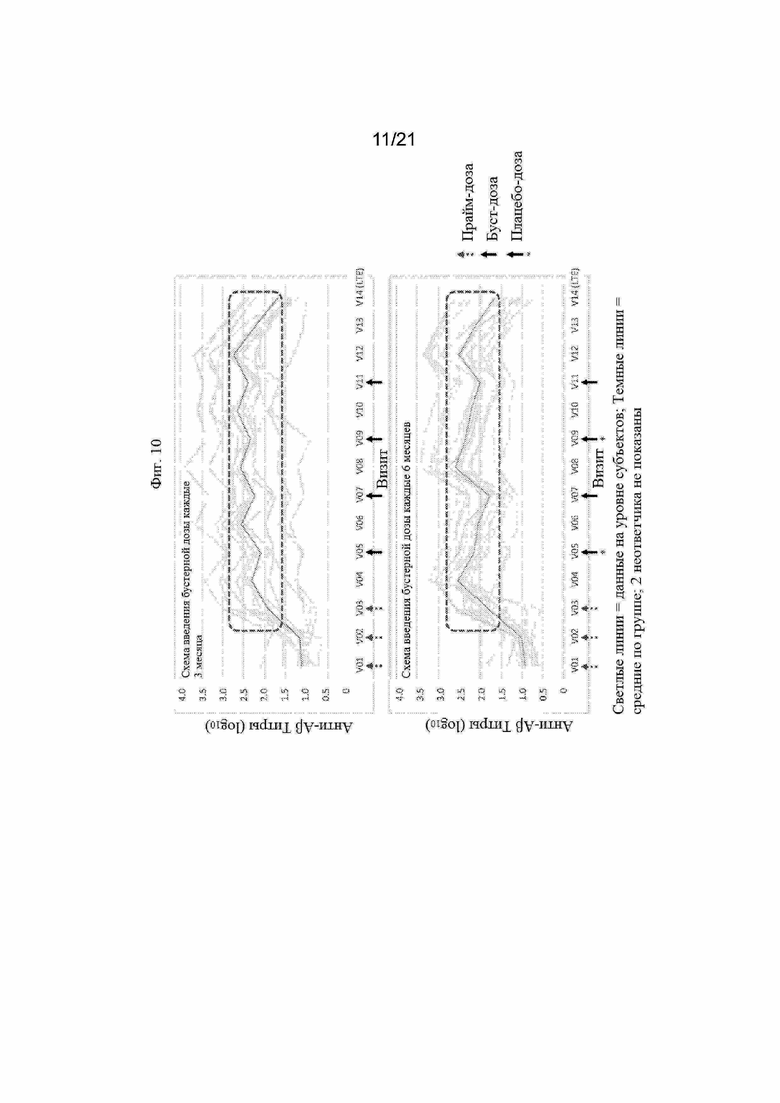
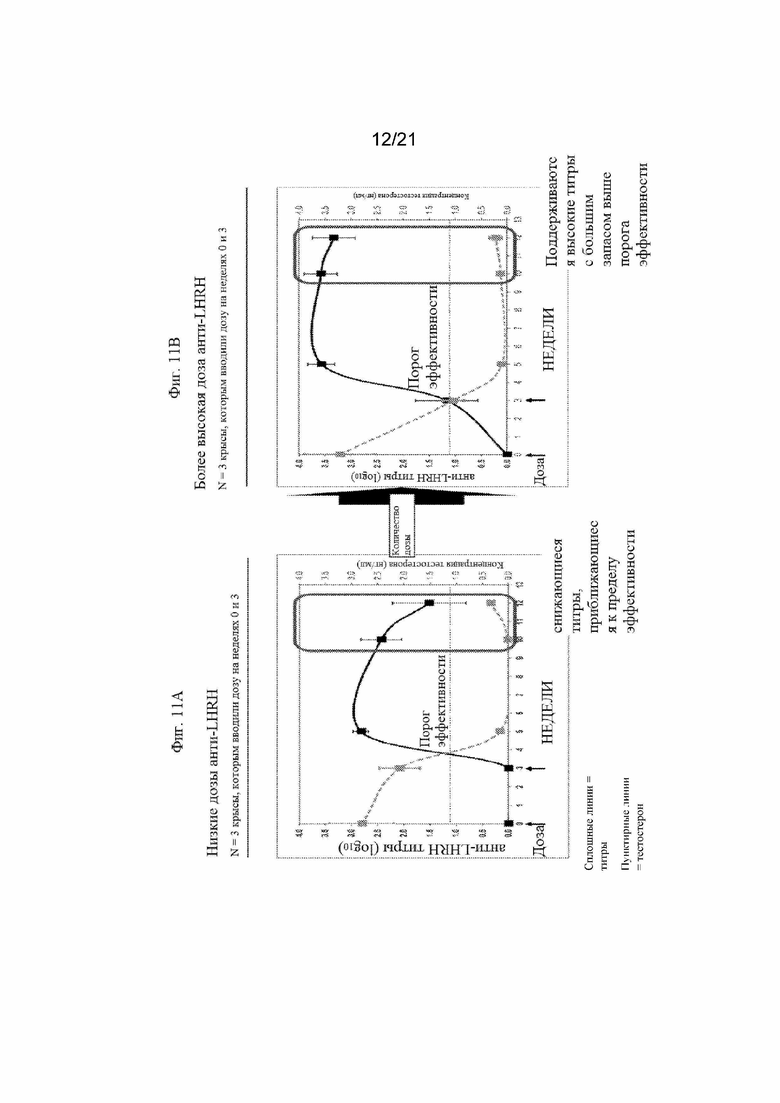
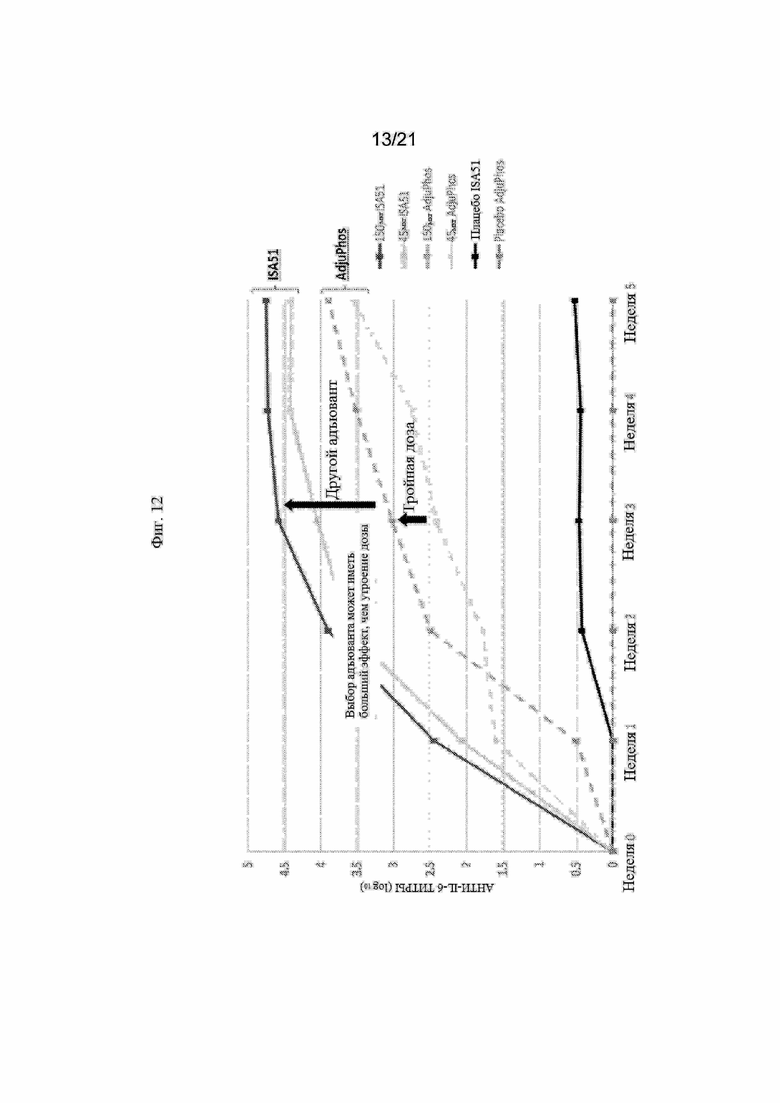
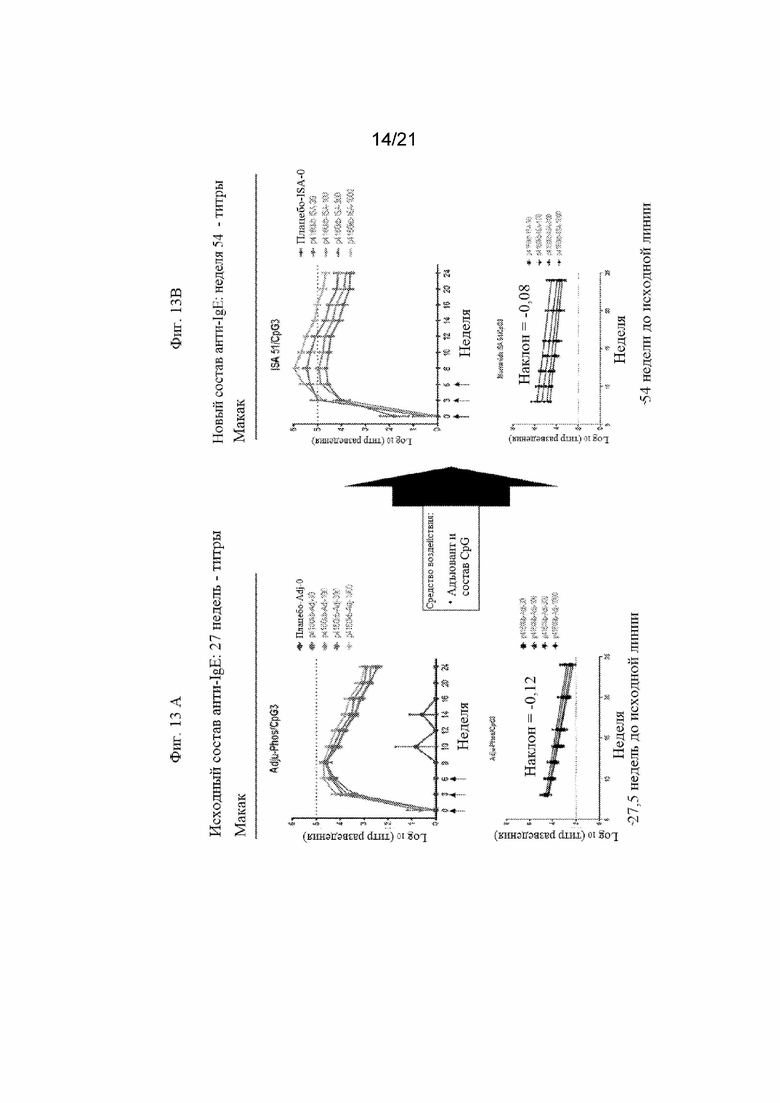
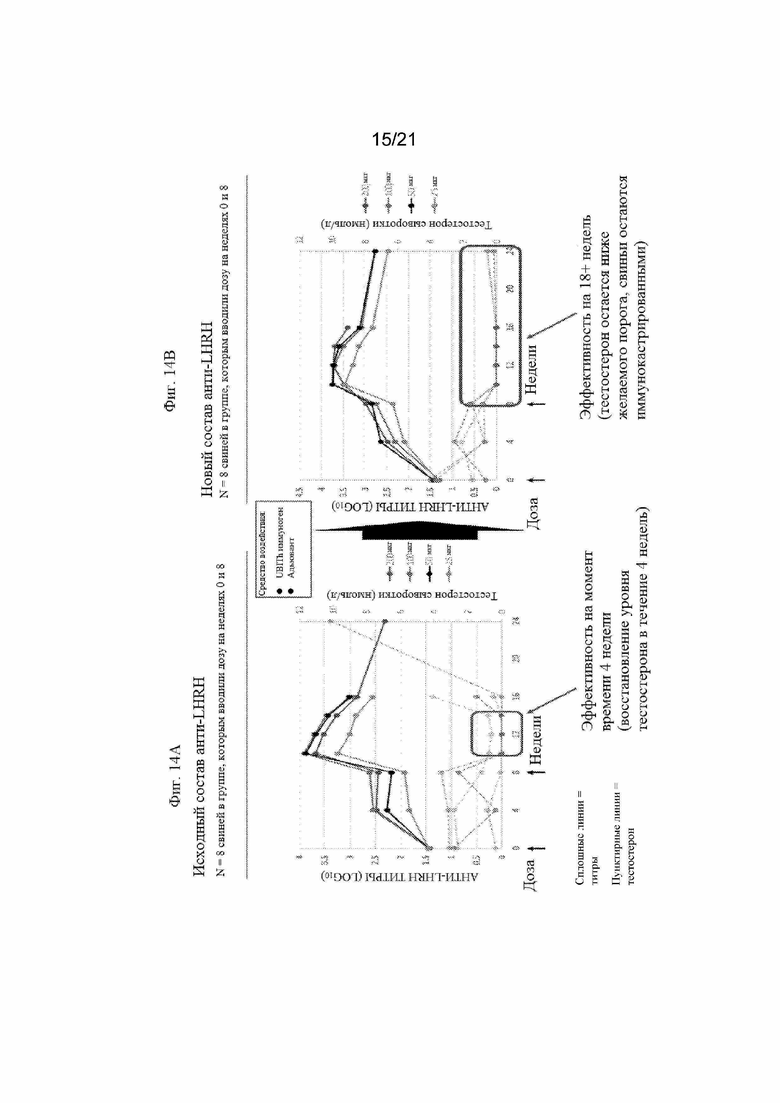
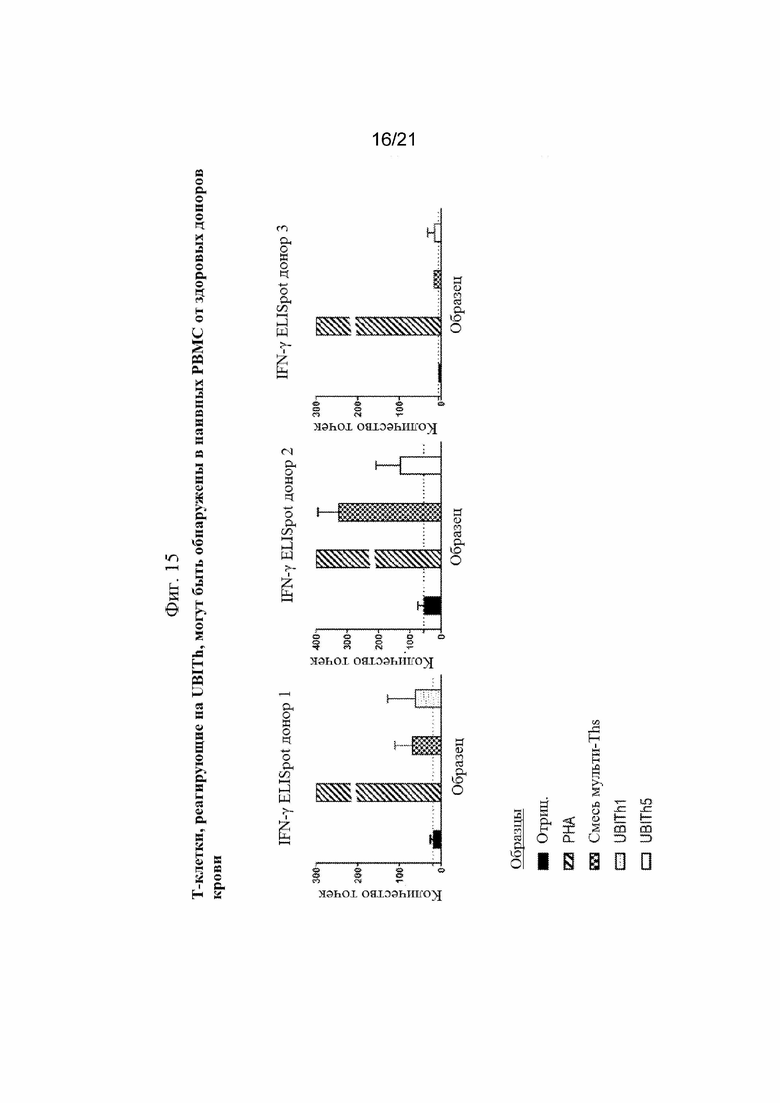

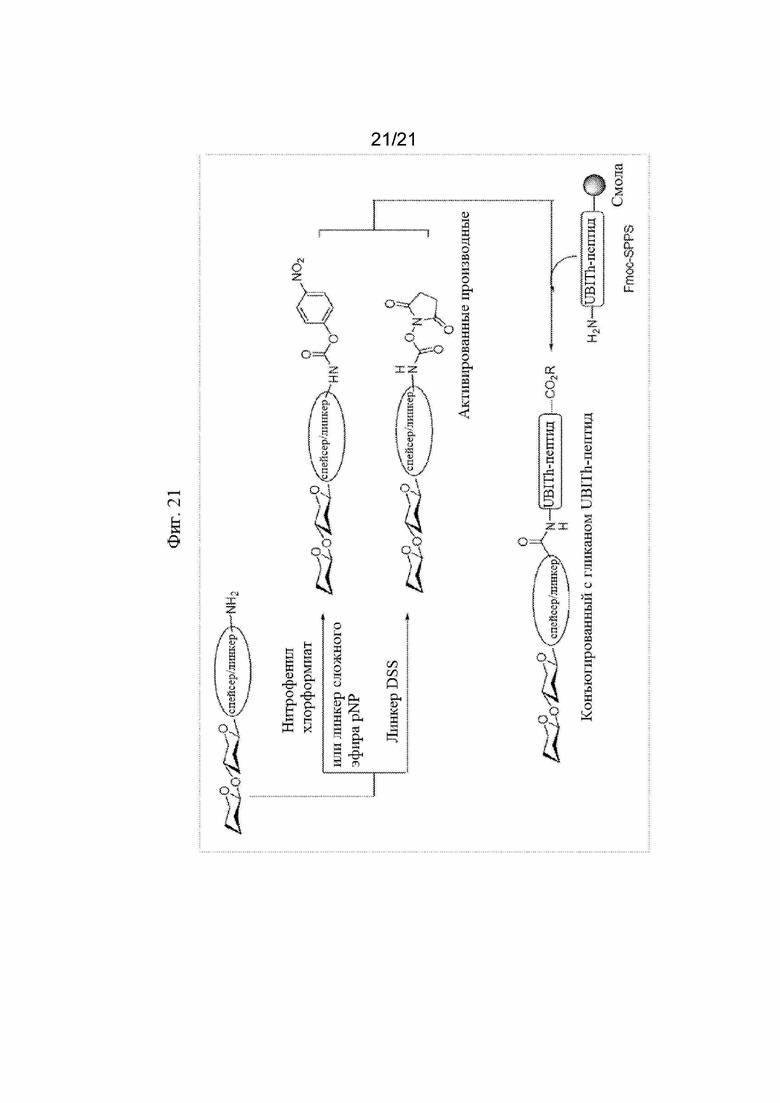
Изобретение относится к биотехнологии, иммунологии и медицине, в частности к новым неизбирательным и искусственным эпитопам Т-хелперных клеток (Th-эпитопам), предназначенных для обеспечения оптимальной иммуногенности целевого антигенного сайта. Целевой антигенный сайт может включать В-клеточный эпитоп, эпитоп CTL, пептидный гаптен, непептидный гаптен или их любой иммунологически реактивный аналог. Описанные Th-эпитопы, когда они ковалентно связаны с целевым антигенным сайтом в пептидной иммуногенной конструкции, вызывают сильный В-клеточный иммунный ответ или ответ эффекторных Т-клеток на целевой антигенный сайт. Изобретение обеспечивает эффективные и безопасные пептидные иммуногены, которые после введения не вызывают воспалительных клеточно-опосредованных иммунных ответов против аутоантигенов. 9 н. и 2 з.п. ф-лы, 21 ил., 15 табл., 12 пр.
1. Пептидная иммуногенная конструкция для индукции опосредованного клетками Т-хелперами иммунного ответа против целевого антигенного сайта, представленная следующими формулами:
(A)n-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X,
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(A)n-X,
или
(A)n-(Th)m-(B)o-(целевой антигенный сайт)-(B)o-(Th)m-(A)n-X,
или
{(A)n-(Th)p-(B)o-(целевой антигенный сайт)-(B)o-(Th)p-(A)n-X}m,
где каждый A независимо представляет собой аминокислоту;
каждый B независимо представляет собой гетерологичный спейсер;
каждый Th независимо представляет собой неизбирательный искусственный Th-эпитоп, выбранный из группы, состоящей из SEQ ID NO: 1-52;
целевой антигенный сайт представляет собой белок аутоантигена, выбранный из группы, состоящей из альфа-синуклеина, имеющего аминокислотную последовательность SEQ ID NO: 61, IgE EMPD, имеющего аминокислотную последовательность SEQ ID NO: 62, IL-6, имеющего аминокислотную последовательность SEQ ID NO: 145 или 242, Aβ1-14 c аминокислотной последовательностью SEQ ID NO: 56-60, пептида IL-31, имеющего аминокислотную последовательность SEQ ID NO: 64 или 72, или Tau пептида, имеющего аминокислотную последовательность SEQ ID NO: 63, 69, 70 или 71;
X представляет собой аминокислоту, α-COOH или α-CONH2;
n равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10;
m равно 1, 2, 3 или 4; а также
o равно 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10;
p равно 0, 1, 2, 3 или 4.
2. Пептидная иммуногенная конструкция по п. 1, где гетерологичный спейсер компонента B выбирают из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N) Lys, ε-N-Lys-Lys-Lys-Lys, представленного в SEQ ID NO: 53, Lys-Lys-Lys-εNLys, представленного в SEQ ID NO: 54, Gly-Gly, Pro-Pro-Xaa-Pro-Xaa-Pro, представленного в SEQ ID NO: 55, и любой их комбинации.
3. Пептидная иммуногенная конструкция по п. 1, где гетерологичный спейсер выбирают из группы, состоящей из (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys, представленного в SEQ ID NO: 53, и Lys-Lys-Lys-εNLys, представленного в SEQ ID NO: 54.
4. Фармацевтическая композиция для лечения болезни Паркинсона, содержащая пептидную иммуногенную конструкцию по п. 1, где целевой антигенный сайт представляет собой альфа-синуклеин, имеющий аминокислотную последовательность SEQ ID NO: 61.
5. Способ предотвращения или лечения заболевания, патологического состояния или болезни у субъекта, включающий введение указанному субъекту фармацевтически эффективного количества фармацевтической композиции по п. 4, где указанное заболевание, патологическое состояние или болезнь включает болезнь Паркинсона.
6. Фармацевтическая композиция для лечения болезни Альцгеймера, содержащая пептидную иммуногенную конструкцию по п. 1, где целевой антигенный сайт выбран из группы, состоящей из Aβ1-14 c аминокислотной последовательностью SEQ ID NO: 56-60 или Tau пептида, имеющего аминокислотную последовательность SEQ ID NO: 63, 69, 70 или 71.
7. Способ предотвращения или лечения заболевания, патологического состояния или болезни у субъекта, включающий введение указанному субъекту фармацевтически эффективного количества фармацевтической композиции по п. 6, где указанное заболевание, патологическое состояние или болезнь включает болезнь Альцгеймера.
8. Фармацевтическая композиция для лечения аллергических заболеваний, содержащая пептидную иммуногенную конструкцию по п. 1, где целевой антигенный сайт выбран из группы, состоящей из IgE EMPD, имеющего аминокислотную последовательность SEQ ID NO: 62, или пептида IL-31, имеющего аминокислотную последовательность SEQ ID NO: 64 или 72.
9. Способ предотвращения или лечения заболевания, патологического состояния или болезни у субъекта, включающий введение указанному субъекту фармацевтически эффективного количества фармацевтической композиции по п. 8, где указанное заболевание, патологическое состояние или болезнь включает аллергические заболевания.
10. Фармацевтическая композиция для лечения связанных с цитокинами заболеваний, содержащая пептидную иммуногенную конструкцию по п. 1, где целевой антигенный сайт представляет собой IL-6, имеющий аминокислотную последовательность SEQ ID NO: 145 или 242.
11. Способ предотвращения или лечения заболевания, патологического состояния или болезни у субъекта, включающий введение указанному субъекту фармацевтически эффективного количества фармацевтической композиции по п. 10, где указанное заболевание, патологическое состояние или болезнь включает связанные с цитокинами заболевания.
| US 2018244739 A1, 30.08.2018 | |||
| СПОСОБ ПОЛУЧЕНИЯ 1,3-ПРОПАНДИОЛА ПУТЕМ КАТАЛИТИЧЕСКОГО ГИДРИРОВАНИЯ 3-ГИДРОКСИПРОПАНАЛЯ В ПРИСУТСТВИИ СОКАТАЛИЗАТОРА ГИДРАТАЦИИ | 2004 |
|
RU2351581C2 |
| КОРОЕВ Д | |||
| О | |||
| и др., Предсказание структуры пептидов, способных индуцировать образование антител у мышей, Биоорганическая химия, 2002, т | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ANEZ G | |||
| et al., Passage of dengue virus type 4 vaccine candidates in fetal rhesus lung cells selects heparin-sensitive variants that | |||

